


植物功能性状反映了植物对环境的适应性和资源利用效率。目前, 植物功能性状被用于探讨群落的组装机制、植物分布、生态位分化, 以及生态系统服务功能的研究。植物功能性状发育受自身结构、遗传和环境资源条件(如水分条件)的限制, 并且不同结构特征或功能性状之间相互制约, 所以植物在适应环境的过程中往往表现出一些功能性状之间的相关性和权衡关系(Meinzer, 2003; Reich et al., 2003; Bucci et al., 2004; Swenson & Enquist, 2007)。比如比叶重较高的叶片, 叶片较致密且厚, 一般具有更强的抗干旱能力(Poorter et al., 2009; Markesteijn et al., 2011; Fu et al., 2012), 但是这种叶片不利于内部气体的扩散, 同时也会稀释单位质量光合组织的浓度, 从而降低其光合作用能力。因此叶片抗干旱能力和光合能力之间可能存在一定的权衡关系, 并受到叶片密度和厚度的制约。
叶片解剖结构特征对植物的生理功能和在特定环境中的适应性有重要影响(Tomas et al., 2013), 能够直接影响植物叶片的光合潜力(Niinemets et al., 2009; Scafaro et al., 2011; Terashima et al., 2011)。叶片的上表皮影响进入叶肉的光量子总量(Brodersen & Vogelmann, 2007), 且多数上表皮细胞具有凸面, 能够聚集通过它们的光(Poulson & Vogelmann, 1990; Vogelmann, 1993; Vogelmann et al., 1996)。有些植物叶片中维管束向叶表皮方向的扩展可以将光传递到叶片的深层(Nikolopoulos et al., 2002), 地中海地区的硬叶植物叶片上表皮细胞中的石细胞也可以将光传递到叶片的内部, 起到光纤维的作用(Karabourniotis et al., 1994; Karabourniotis, 1998)。而柱状的栅栏组织细胞具有光线通道的作用, 将光线传播到叶肉深层(Vogelmann & Martin, 1993)。球状的海绵组织细胞排列不规则, 使得光线散射多次, 在叶肉中的传播路径延长, 使光量子得到充分吸收(DeLucia et al., 1996; Evans et al., 2004)。
生长地点的水分条件可以导致植物的解剖结构发生改变(Chartzoulakis et al., 2002)。旱生植物叶片一般都具有较低的面积/体积比(Karabourniotis & Bornman, 1999; Richardson & Berlyn, 2002), 较厚的角质层(Richardson & Berlyn, 2002)和上下表皮(Bacelar et al., 2004)以及较小的叶肉细胞和细胞间隙(Mediavilla et al., 2001)。干旱生境中, 植物叶片表面的毛状体可以减少水分蒸腾(Bacelar et al., 2004), 增加边界层阻力(Savé et al., 2000), 从而提高水分利用效率。同时还可以增加植物对小降水量和冷凝水的利用(Savé et al., 2000)。植物叶片忍耐低水势的能力决定了植物在旱季的存活能力以及植物的分布模式(Kursar et al., 2009)。植物压力-容积曲线参数是植物叶片耐失水能力的指标。膨压丧失点时的叶片水势(πtlp)与植物生长地点的降水量有很强的相关性, 是代表植物耐旱性的一个很有效的功能性状(Bartlett et al., 2012b)。
热带雨林是生物多样性最丰富的陆地生态系统, 其物种共存机制是研究的热点。近年来, 我国学者对热带雨林的生物多样性维持机制开展了大量研究, 主要集中在利用模型探讨中性理论、生态位分化、密度依赖等解释物种分布和共存的机制(Lan et al., 2012; Lin et al., 2012; Hu et al., 2013)。但是, 把植物功能性状特别是植物抗逆性性状用于物种共存机制的研究还很少, 一个原因是因为测定这些抗逆性性状很费工费时。本研究选取了西双版纳热带季雨林在水分条件不同的几个生境(山脊、山坡、沟谷)中分布的18种大戟科植物。测量了其幼树叶片的叶片解剖结构、水分关系特征、最大光合能力和暗呼吸。大戟科是热带雨林很重要的一个科, 其重要值在西双版纳热带雨林20 hm2样地排第三(兰国玉等, 2008)。此外, 同一个科内的物种进化历史相近, 在探讨性状的关联进化时可以排除进化历史差异较大带来的影响。本研究主要探讨: 1)热带季雨林不同生境分布的植物功能性状之间的适应性差异; 2)植物叶片解剖结构和植物光合以及水分关系特征之间的关系。从而为解释植物分布和共存机制提供依据。因为亲缘关系较近的物种之间存在进化上的自相似性, 从而导致用传统的统计方法检验性状之间的相关关系时会产生偏差(Felsenstein, 1985), 所以本研究利用系统发育独立对照差的方法, 考虑了系统发育的影响, 分析亲缘关系相近的植物类群性状的关联进化(Felsenstein, 1985; Garland et al., 1992)。
1 材料和方法
1.1 实验样地与材料
本研究在西双版纳热带季雨林20 hm2动态监测样地内进行, 样地位于云南省西双版纳傣族自治州勐腊县补蚌村南贡山东部斑马山脚, 地理位置为101.57°-101.58° E, 21.61°-21.62° N (兰国玉等, 2008)。该地区属于热带季风气候, 5到10月为雨季, 11月到翌年4月为旱季(Cao et al., 2006)。平均年降水量为1493 mm, 其中雨季占84%, 旱季降水稀少(Cao et al., 2006)。物种分布和物种的生境类型划分参照对该样地地形特征和物种分布的基础调查(Lan et al., 2012), 选取了样地内不同生境(山脊、山坡、沟谷)的18种大戟科植物的幼树(1.0 cm <胸径(DBH) < 5.0 cm), 树高介于1.0-2.5 m之间(种名、丰度(每hm2样地内的个体数)及分布见表1), 其中广布种是在3个生境内均匀分布的种。林下小树因为根系相对较浅, 所以对生境水分条件变化更为敏感(Becker & Wong, 1994)。实验选取的3个生境存在显著的水分差异。沟谷常年有小溪存在, 而山脊和山坡无地表水积存。这种水分差异在旱季尤为明显。3个生境的光环境也有所不同, 沟谷两侧的林下郁闭度高, 而山脊比较空旷, 有明显的林窗存在。山坡林下的光环境介于二者之间。
表1 样地内18种大戟科植物物种名、分布及丰度
Table 1
| 种 Species | 分布 Distribution | 丰度 Abundance (Ind.·hm-2) |
|---|---|---|
| 长梗三宝木 Trigonostemon thyrsoideus | 沟谷 Valley | 40.45 |
| 粉绿野桐 Mallotus garrettii | 沟谷 Valley | 34.15 |
| 勐腊核果木 Drypetes hoaensis | 沟谷 Valley | 28.35 |
| 棒柄花 Cleidion brevipetiolatum | 沟谷 Valley | 48.55 |
| 秋枫 Bischofia javanica | 沟谷 Valley | 1.65 |
| 缅桐 Sumbaviopsis albicans | 山坡 Slope | 23.00 |
| 轮叶戟 Lasiococca comberi | 山坡 Slope | 9.20 |
| 风轮桐 Epiprinus siletianus | 山坡 Slope | 6.20 |
| 网脉核果木 Drypetes perreticulata | 山坡 Slope | 1.50 |
| 土蜜树 Bridelia tomentosa | 山坡 Slope | 1.05 |
| 木奶果 Baccaurea ramilflora | 遍及整个样地 Throughout the entire sample plot | 160.60 |
| 山地五月茶 Antidesma montanum | 遍及整个样地 Throughout the entire sample plot | 22.75 |
| 日本五月茶 Antidesma japonicum | 遍及整个样地 Throughout the entire sample plot | 13.75 |
| 尾叶血桐 Macaranga kurzii | 山脊 Ridge | 0.40 |
| 越南巴豆 Croton kongensis | 山脊 Ridge | 7.35 |
| 银背巴豆 Croton argyratus | 山脊 Ridge | 3.00 |
| 椴叶山麻杆 Alchornea tiliifolia | 山脊 Ridge | 18.75 |
| 云南银柴 Aporusa yunnanensis | 山脊 Ridge | 26.40 |
1.2 压力-容积曲线的测定
压力-容积曲线的测定在旱季进行。每个种分别随机抽取6个个体, 广布种在各个生境内各选取4个个体。每个个体在傍晚时采集一根枝条, 用湿润的纸巾包裹住切口, 装入黑色采样袋内, 带回室内, 复水一夜。第二天, 用刀片将带叶柄的叶片从枝条上切下, 放入Whirl-Pak (Nasco, Fort Atkinson, Wisconsin, USA)袋中, 使用Model 1000压力势仪(PMS Instrument Company, Oregon, USA)进行叶片水势的测定。通过压力-容积曲线, 使用标准方法(Bartlett et al., 2012a), 计算得到饱和含水量(SWC)、饱和渗透势(πo)、膨压丧失点时的水势(πtlp)、膨压丧失点时的相对含水量(RWCtlp)、弹性模数(ε)。在计算过程中, 表示过饱和的共质体含水量的数据点被剔除(Kubiske & Abrams, 1990; Sack & Pasquet-Kok, 2011)。
1.3 气体交换的测定
使用LI-6400便携式光合仪(LI-COR, Lincoln, USA)配合红/蓝光源, 从每个种的幼树中随机抽取的6个个体(广布种幼树的选取同1.2), 测定其相对阳生且完全展开的叶片的气体交换状况。初始得到的值为单位面积的值, 随后乘以比叶面积(SLA)转化为单位质量的值。每天9:30-11:30进行最大光合速率的测定, 光强设定为1200 μmol·m-2·s-1。使用外接CO2小钢瓶来减少林下环境波动对测定的影响, CO2浓度设定为500 μmol·mol-1。暗呼吸的测定在20:30-22:30进行。最大光合速率和暗呼吸速率(R)的测定均采用原地活体测量。由于暗呼吸速率的值为负值, 为表述方便取其绝对值将其转化为正值。
1.4 叶片解剖结构和功能性状的测定
每个个体选取3片健康、相对阳生且无明显缺损的叶片, 使用显微镜(MD 2500; Leica Microsystems Ltd., Wetzlar, Germany)对叶片顶端、中部和底部的横切切片进行叶片各结构成分(上表皮厚度(UET)、栅栏组织厚度(PT)、海绵组织厚度(ST)、下表皮厚度(LET)和叶片厚度(LT))的测定。使用便携式叶面积仪(LI-3000A; Lincoln, USA)测量叶片面积, 随后将叶片装入信封中, 放入烘箱中80 ℃烘干48 h。用分析天平(AL204; Mettler Toledo Group, 上海)测定叶片干质量。SLA =叶片面积/叶片干质量。比叶质量(LMA) =叶片干质量/叶片面积。叶片密度(LD) = LMA/LT。栅栏组织厚度/海绵组织厚度(P/S) = PT/ST。
1.5 系统发育树的构建
我们在属水平上利用rbcL和matK DNA序列构建系统发育树。山地五月茶(Antidesma montanum)、越南巴豆(Croton kongensis)、银背巴豆(Croton argyratus)、网脉核果木(Drypetes perreticulata)、风轮桐(Epiprinus siletianus)和轮叶戟(Lasiococca comberi)的rbcL和matK DNA序列来自西双版纳热带植物园DNA Barcoding项目。棒柄花(Cleidion brevipetiolatum)的matK序列和勐腊核果木(Drypetes hoaensis)、酸味子(Antidesma japonicum)的rbcL序列也来自这个项目。白叶桐(Sumbaviopsis albicans)、秋枫(Bischofia javanica)、土蜜树(Bridelia tomentosa)、木奶果(Baccaurea ramiflora)、云南银柴(Aporosa yunnanensis)、长梗三宝木(Trigonostemon thyrsoideus)、粉绿野桐(Mallotus garrettii)、尾叶血桐(Macaranga Kurzii)和椴叶山麻杆(Alchornea tiliifolia)的rbcL和matK DNA序列从GenBank获得。由于木奶果、云南银柴、长梗三宝木、粉绿野桐、尾叶血桐和椴叶山麻杆其序列无法获得, 我们使用同属种的基因序列代替。利用Mega 5.2 (Tamura et al., 2011)进行系统发育树的构建, 算法采用最大似然法(maximum likelihood method)。
1.6 统计方法
分别采用Pearson相关分析和系统发育独立对照差相关分析, 对各性状间的相关关系进行了研究。各类群之间的差异采用单因素方差分析(one- way ANOVA)进行检验, 对差异显著的性状用Tukey HSD法进行多重比较。使用R 3.0.2进行上述统计分析。
2 实验结果
2.1 各类群植物间的性状差异
从表2可见, 广布种具有最大的SWC。山脊和山坡分布种的πo和πtlp显著低于沟谷分布种和广布种。山脊分布种的RWCtlp最小, 沟谷分布种的RWCtlp最大。广布种具有最大的UET。山坡分布种的LD显著高于沟谷分布种。山脊分布种的R最大, 山坡分布种的R最小。其余的性状各类群间无显著性差异。其中, 广布种具有最大的LT、PT、ST和最小的ε和单位叶片干质量的最大净光合速率(Am), 而山坡分布种具有最小的LT、PT、ST和最大的ε、LMA。山脊分布种具有最大的P/S和单位叶面积的最大光合速率(Aa)、Am。沟谷分布种则具有最小的LMA、P/S和Aa。山脊分布种和山坡分布种的LET比沟谷分布种和广布种小。山脊分布种和沟谷分布种的Rm比广布种和山坡分布种大。
表2 大戟科植物不同小生境分布类群的叶片性状及方差分析结果(平均值±标准误差)
Table 2
| 性状 Trait | 山脊 Ridge | 山坡 Slope | 沟谷 Valley | 广布种 Cosmopolitan species |
|---|---|---|---|---|
| SWC (%) | 2.660 ± 0.840ab | 1.860 ± 0.330a | 3.280 ± 1.110ab | 4.260 ± 0.770b |
| πo (MPa) | -1.670 ± 0.180a | -1.740 ± 0.250a | -1.190 ± 0.220b | -1.190 ± 0.040b |
| πtlp (MPa) | -1.950 ± 0.180a | -1.990 ± 0.220a | -1.400 ± 0.270b | -1.400 ± 0.020b |
| ε (MPa) | 15.21 ± 6.540a | 22.32 ± 7.480a | 18.70 ± 6.300a | 11.88 ± 2.190a |
| RWCtlp (%) | 84.62 ± 5.280a | 90.70 ± 4.600ab | 92.61 ± 3.560b | 86.99 ± 0.810ab |
| LT (mm) | 0.670 ± 0.380a | 0.540 ± 0.140a | 0.900 ± 0.300a | 1.100 ± 0.460a |
| UET (mm) | 0.069 ± 0.036a | 0.061 ± 0.010a | 0.083 ± 0.017a | 0.164 ± 0.053b |
| PT (mm) | 0.240 ± 0.130a | 0.150 ± 0.060a | 0.190 ± 0.090a | 0.280 ± 0.060a |
| ST (mm) | 0.300 ± 0.230a | 0.260 ± 0.100a | 0.540 ± 0.220a | 0.550 ± 0.340a |
| LET (mm) | 0.063 ± 0.022a | 0.063 ± 0.015a | 0.080 ± 0.018a | 0.101 ± 0.023a |
| P/S (%) | 1.214 ± 0.611a | 0.670 ± 0.345a | 0.432 ± 0.270a | 0.618 ± 0.208a |
| LD (g·cm-3) | 940.4 ± 391.2ab | 1 175.0 ± 385.5a | 514.2 ± 94.2b | 529.5 ± 171.1ab |
| LMA (g·cm-2) | 51.36 ± 12.810a | 59.56 ± 14.080a | 44.91 ± 14.750a | 53.49 ± 6.640a |
| Aa (μmol·m-2·s-1) | 10.380 ± 2.310a | 8.400 ± 3.410a | 7.690 ± 1.520a | 7.710 ± 0.750a |
| Am (nmol·g-1·s-1) | 0.210 ± 0.150a | 0.160 ± 0.110a | 0.190 ± 0.110a | 0.150 ± 0.030a |
| R (μmol·m-2·s-1) | 0.600 ± 0.074a | 0.470 ± 0.074b | 0.500 ± 0.064ab | 0.480 ± 0.046ab |
| Rm (nmol·g-1·s-1) | 0.012 ± 0.003a | 0.008 ± 0.003a | 0.012 ± 0.006a | 0.009 ± 0.001a |
ε, 弹性模数; πo, 饱和渗透势; πtlp, 膨压丧失点时的水势; Aa, 单位叶面积最大光合速率; Am, 单位叶片干质量最大光合速率; LD, 叶片密度; LET, 下表皮厚度; LMA, 比叶质量; LT, 叶片厚度; P/S, 栅栏组织厚度/海绵组织厚度; PT, 栅栏组织厚度; R, 单位叶面积暗呼吸速率; Rm, 单位叶干质量暗呼吸速率; RWCtlp, 膨压丧失点时的相对含水量; ST, 海绵组织厚度; SWC, 饱和含水量; UET, 上表皮厚度。不同小写字母表示差异显著(p < 0.05)。
ε, modulus of elasticity at full turgor; πo, osmotic potential at full turgor; πtlp, water potential at turgor loss point; Aa, maximum photosynthesis rate per leaf area; Am, maximum photosynthesis rate per leaf dry mass; LD, leaf density; LET, lower epidermis thickness; LMA, leaf mass per area; LT, leaf thickness; P/S, palisade thickness/spongy thickness; PT, palisade tissue thickness; R, dark respiration rate per leaf area; Rm, dark respiration rate per leaf dry mass; RWCtlp, relative water content at turgor loss point; ST, spongy tissue thickness; SWC, saturated water content; UET, upper epidermis thickness. Different lowercase letters indicate significant differences (p < 0.05).
2.2 LT、LD和LMA之间的相关关系
表3 大戟科植物叶片各性状间的相关关系。左下为传统Pearson相关,右上为系统发育独立性比较后的相关
Table 3
| SWC | πo | πtlp | ε | RWCtlp | LT | UET | PT | ST | LET | P/S | LD | LMA | Aa | Am | R | Rm | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SWC | 1 | 0.81*** | 0.82*** | -0.25 | 0.07 | 0.62** | 0.61** | -0.05 | 0.70** | 0.60* | -0.81*** | -0.71*** | -0.44 | 0.25 | 0.51* | 0.19 | 0.57* |
| πo | 0.70** | 1 | 0.98*** | -0.25 | 0.20 | 0.59* | 0.52* | -0.23 | 0.74*** | 0.37 | -0.76*** | -0.71*** | -0.86*** | 0.27 | 0.63** | -0.11 | 0.66** |
| πtlp | 0.77*** | 0.98*** | 1 | -0.20 | 0.28 | 0.55* | 0.53* | -0.24 | 0.69** | 0.41 | -0.82*** | -0.70** | -0.79*** | 0.18 | 0.57** | -0.17 | 0.61** |
| ε | -0.47 | -0.68** | -0.57* | 1 | 0.81*** | -0.65** | -0.45 | 0.01 | -0.74*** | -0.44 | 0.43 | 0.24 | 0.66** | -0.65** | -0.74*** | -0.37 | -0.69** |
| RWCtlp | -0.05 | -0.26 | -0.10 | 0.77*** | 1 | -0.31 | -0.18 | -0.05 | -0.34 | -0.06 | -0.12 | -0.13 | 0.47 | -0.56* | -0.54* | -0.44 | -0.52* |
| LT | 0.64** | 0.60** | 0.60** | -0.13 | 0.25 | 1 | 0.84*** | 0.74*** | 0.95*** | 0.91*** | -0.56* | -0.78*** | 0.33 | 0.01 | -0.25 | -0.03 | -0.33 |
| UET | 0.66** | 0.55* | 0.56* | -0.31 | 0 | 0.71** | 1 | 0.71*** | 0.69** | 0.85*** | -0.45 | -0.62** | 0.23 | -0.06 | -0.23 | -0.10 | -0.28 |
| PT | 0.65** | 0.42 | 0.43 | -0.23 | -0.06 | 0.51* | 0.27 | 1 | 0.49* | 0.69** | 0.20 | -0.61** | 0.23 | 0.40 | 0.05 | 0.36 | -0.08 |
| ST | 0.50* | 0.56* | 0.55* | -0.03 | 0.37 | 0.95*** | 0.66** | 0.23 | 1 | 0.82*** | -0.70** | -0.73*** | 0.32 | -0.13 | -0.33 | -0.17 | -0.37 |
| LET | 0.54* | 0.52* | 0.54* | -0.20 | 0.20 | 0.83*** | 0.63** | 0.60* | 0.71** | 1 | -0.50* | -0.76*** | 0.34 | -0.03 | -0.25 | -0.16 | -0.38 |
| P/S | -0.21 | -0.37 | -0.37 | -0.37 | -0.68** | -0.54* | -0.30 | -0.00 | -0.70** | -0.57* | 1 | 0.69** | 0.44 | -0.21 | -0.45 | 0.06 | -0.41 |
| LD | -0.70** | -0.89*** | -0.87*** | 0.75*** | 0.36 | -0.75** | -0.56* | -0.10 | -0.81*** | -0.60* | 0.48* | 1 | 0.76*** | -0.53* | -0.81*** | -0.23 | -0.82*** |
| LMA | -0.16 | -0.29 | -0.27 | 0.31 | 0.33 | -0.29 | -0.24 | 0.40 | -0.47 | 0 | -0.37 | 0.14 | 1 | -0.22 | -0.77*** | 0.16 | -0.75** |
| Aa | 0.16 | 0 | -0.04 | -0.39 | -0.42 | 0.69** | 0.22 | 0.58* | 0.61** | 0.54* | 0.35 | -0.20 | -0.28 | 1 | 0.77*** | 0.76*** | 0.44 |
| Am | 0.14 | 0.10 | 0.10 | -0.37 | -0.37 | 0.60* | 0.27 | 0.18 | 0.62** | 0.40 | 0.44 | -0.18 | -0.68** | 0.80*** | 1 | 0.49* | 0.87*** |
| R | 0.20 | -0.08 | -0.14 | -0.41 | -0.56* | 0.53* | 0.10 | 0.70** | 0.38 | 0.52* | 0.48* | -0.18 | -0.24 | 0.59** | 0.52* | 1 | -0.62** |
| Rm | 0.16 | 0.12 | 0.12 | -0.36 | -0.40 | 0.50* | 0.25 | 0.04 | 0.56* | 0.30 | 0.49* | -0.15 | -0.86*** | 0.58* | 0.92*** | -0.45 | 1 |
ε, 弹性模数; πo, 饱和渗透势; πtlp, 膨压丧失点时的水势; Aa, 单位叶面积最大光合速率; Am, 单位叶干质量最大光合速率; LD, 叶片密度; LET, 下表皮厚度; LMA, 比叶质量; LT, 叶片厚度; P/S, 栅栏组织厚度/海绵组织厚度; PT, 栅栏组织厚度; R, 单位叶面积暗呼吸速率; Rm, 单位叶干质量暗呼吸速率; RWCtlp, 膨压丧失点时的相对含水量; ST, 海绵组织厚度; SWC, 饱和含水量; UET, 上表皮 厚度。*, 0.01 < p < 0.05; **, 0.001 < p < 0.01; ***, p < 0.001。
ε, modulus of elasticity at full turgor; πo, osmotic potential at full turgor; πtlp, water potential at turgor loss point; Aa, maximum photosynthesis per leaf area; Am, maximum photosynthesis per leaf dry mass; LD, leaf density; LET, lower epidermis thickness; LMA, leaf mass per area; LT, leaf thickness; P/S, palisade thickness/spongy thickness; PT, palisade tissue thickness; R, dark respiration per leaf area; Rm, dark respiration per leaf dry mass; RWCtlp, relative water content at turgor loss point; ST, spongy tissue thickness; SWC, saturated water content; UET, upper epidermis thickness. *, 0.01 < p < 0.05; **, 0.001 < p < 0.01; ***, p < 0.001.
2.3 叶片解剖和水分关系特征之间的相关关系
图1
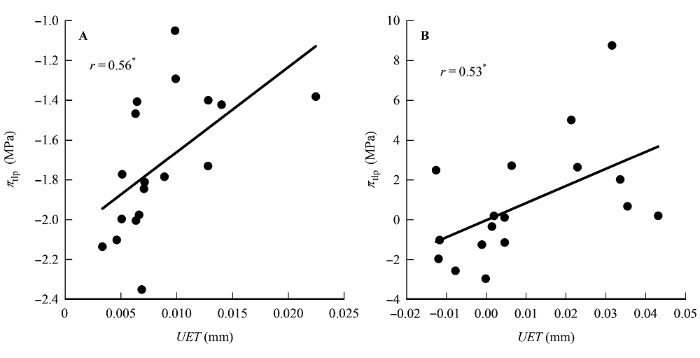
图1
上表皮厚度(UET)与膨压丧失点时的水势(πtlp)的相关关系。
A, 传统Pearson相关。B, 系统发育独立性比较。*, 0.01 < p < 0.05。
Fig. 1
Correlations between upper epidermis thickness (UET) and water potential at turgor loss point (πtlp).
A, Traditional Pearson correlation. B, Correlation given by the phylogenetic independent contrasts analysis. *, 0.01 < p < 0.05.
图2
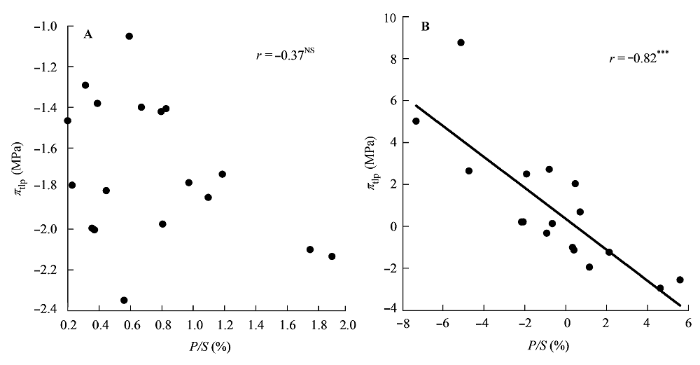
图2
栅栏组织厚度/海绵组织厚度(P/S)与膨压丧失点时的水势(πtlp)的相关关系。
A, 传统Pearson相关。B, 系统发育独立性比较。***, p < 0.001; NS, p > 0.05。
Fig. 2
Correlations between the palisade thickness/spongy thickness (P/S) and water potential at turgor loss point (πtlp).
A, Traditional Pearson correlation. B, Correlation given by the phylogenetic independent contrasts analysis. ***, p < 0.001; NS, p > 0.05.
图3
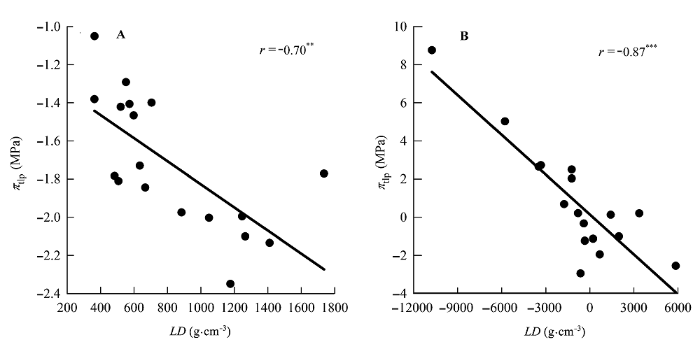
图3
叶片密度(LD)与膨压丧失点时的水势(πtlp)的相关关系。
A, 传统Pearson相关。B, 系统发育独立性比较。**, 0.001 < p < 0.01; ***, p < 0.001。
Fig. 3
Correlations between leaf density (LD) and water potential at turgor loss point (πtlp).
A, Traditional Pearson correlation. B, Correlation given by the phylogenetic independent contrasts analysis. **, 0.001 < p < 0.01; ***, p < 0.001.
图4
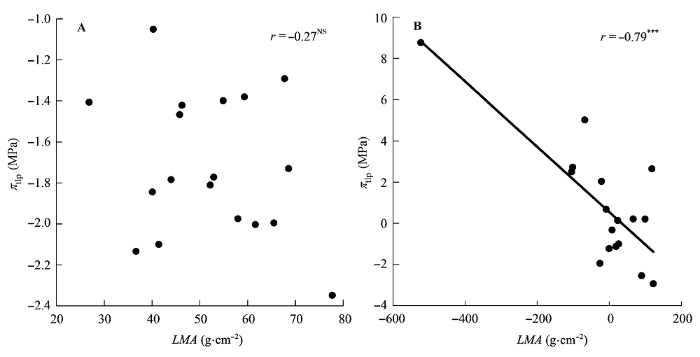
图4
比叶质量(LMA)与膨压丧失点时的水势(πtlp)的相关关系。
A, 传统Pearson相关。B, 系统发育独立性比较。***, p < 0.001; NS, p > 0.05。
Fig. 4
Correlations between leaf mass per area (LMA) and water potential at turgor loss point (πtlp).
A, Traditional Pearson correlation. B, Correlation given by the phylogenetic independent contrasts analysis. ***, p < 0.001; NS, p > 0.05.
2.4 叶片解剖结构和气体交换特征之间的相关 关系
图5
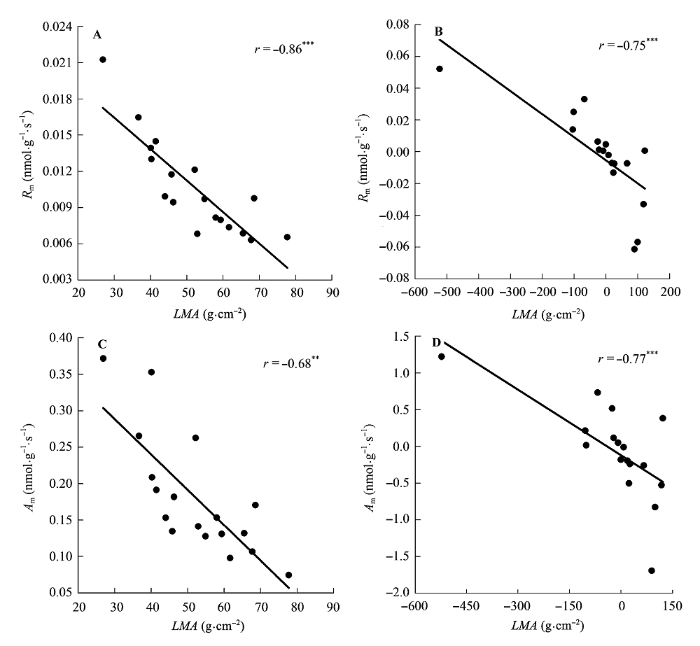
图5
比叶质量(LMA)与单位叶干质量最大光合速率(Am)和单位叶干质量暗呼吸速率(Rm)的相关关系。
A, B, 传统Pearson相关。C, D, 系统发育独立性比较。**, 0.001 < p < 0.01; ***, p < 0.001。
Fig. 5
Correlations of leaf mass per area (LMA) with maximum photosynthesis per leaf dry mass (Am) and dark respiration per leaf dry mass (Rm).
A, B, Traditional Pearson correlation. C, D, Correlation given by the phylogenetic independent contrasts analysis. **, 0.001 < p < 0.01; ***, p < 0.001.
图6
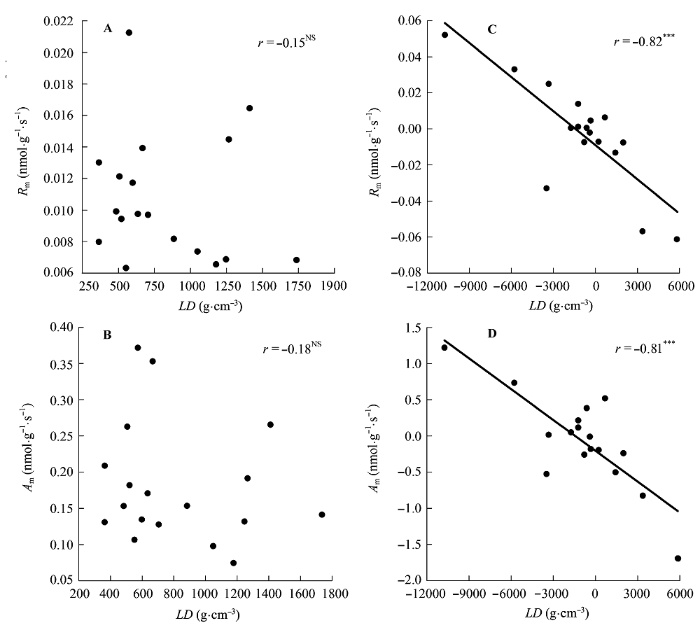
图6
叶片密度(LD)与单位叶干质量最大光合速率(Am)和单位叶干质量暗呼吸速率(Rm)的相关关系。
A, B, 传统Pearson相关。C, D, 系统发育独立性比较。***, p < 0.001; NS, p > 0.05。
Fig. 6
Correlations of leaf density (LD) with maximum photosynthesis per leaf dry mass (Am) and dark respiration per leaf dry mass (Rm).
A, B, Traditional Pearson correlation. C, D, Correlation given by the phylogenetic independent contrasts analysis. ***, p < 0.001; NS, p > 0.05.
2.5 叶片水分关系特征与气体交换之间的相关关系
图7
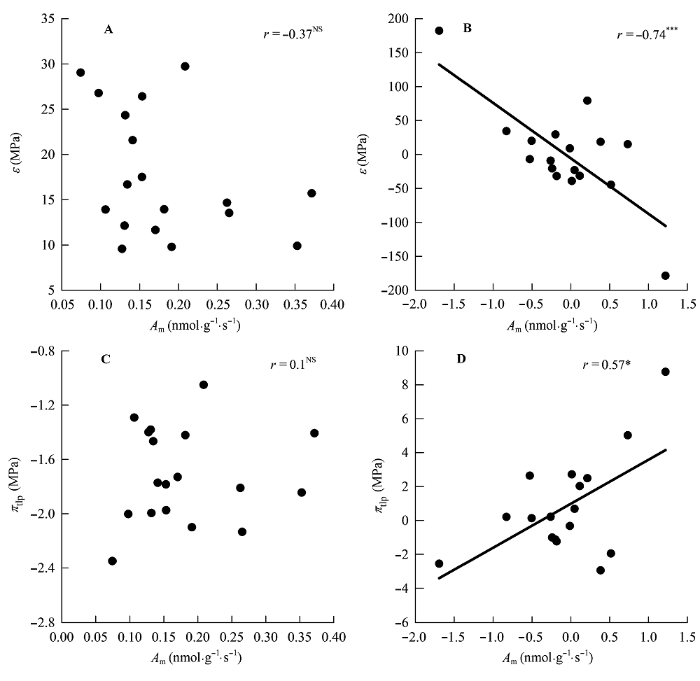
图7
单位叶干质量最大光合速率(Am)与膨压丧失点时的水势(πtlp)和叶片弹性模数(ε)的相关关系。
A、B, 传统Pearson相关。C、D, 系统发育独立性比较。*, 0.01 < p < 0.05; ***, p < 0.001; NS, p > 0.05。
Fig. 7
Correlations of maximum photosynthesis per leaf dry mass (Am) with water potential at turgor loss point (πtlp) and modulus of elasticity at full turgor (ε).
A, B, Traditional Pearson correlation. C, D, Correlation given by the phylogenetic independent contrasts analysis. *, 0.01 < p < 0.05; ***, p < 0.001; NS, p > 0.05.
3 讨论
本研究发现, 生境内大戟科植物幼树表现出了一定的结构和功能的趋同性, 分布在山脊和山坡的种与分布在沟谷的种和广布种相比具有更低的膨压丧失点和饱和渗透势, 以及更高的光合能力和P/S (表2); 植物的耐失水能力与植物的光合能力之间存在权衡关系(图7)。因此, 植物在耐失水能力和光合能力方向上的分化和二者之间的权衡关系可能为大戟科植物的共存机制和分布格局提供了解释。有研究表明, 植物叶片耐失水能力是植物耐旱性的一个重要指标(Bartlett et al., 2012b; Fu et al., 2012)。分布在山脊和山坡的种具有较低的膨压丧失点和饱和渗透势, 表明它们具有更强的耐失水能力, 能够避免日间和干旱季节膨压丧失造成的气孔关闭所引起的光合气体交换受限, 从而有利于它们占据相对干旱的生境。与山脊和山坡相比, 沟谷生境常年有地表水流, 水分条件较好, 很少受旱季缺水的影响。但是沟谷林下光环境较差, 植物叶片的P/S较小(表2), 即海绵组织比例较大, 使得光线多次反射, 光路径延长(Vogelman et al., 1996; Smith et al., 1997), 增加了林下光谱的主要成分—绿光的吸收(Terashima et al., 2011)。同时, 较低的R (表2)减少了植物维持的消耗, 增加了光合产物的积累, 提高了植物的耐阴性, 从而能够将耐旱的植物从郁闭生境排挤出去。相关研究表明, 植物的抗逆性和光合、生长之间总是存在权衡关系。例如, 叶片N向防御方面的投资与光合方面的投资存在权衡关系(Feng et al., 2009); 木质部抗冰冻-解冻引起的栓塞化的能力和光合能力之间存在权衡关系(Choat et al., 2011); 植物的耐阴性和光合能力之间存在权衡关系(Walters & Reich, 2000)。本研究结果支持了植物耐旱性和光合能力之间的权衡关系(Aranda et al., 2005)。尽管Lan等(2012)、Lin等(2012)发现中性过程自身或者与生态位分化相结合影响了西双版纳地区热带雨林物种的分布和共存, 但是我们的结果显示植物的叶片结构和功能表现出对不同小生境光和水分条件的适应, 植物的耐失水能力与光合能力之间存在权衡关系, 表明物种在环境水分梯度和光梯度上产生了生态位的分化。因此更加支持生态位分化对该热带雨林高的生物多样性维持和大量物种的长期共存所起的重要作用(Brown et al., 2013)。
我们发现植物的耐失水能力和光合能力之间的权衡关系是由叶片自身结构的限制所决定的。一方面叶片结构影响植物的耐失水能力。SWC随LT、UET、PT、ST以及LET的增加而增加(表3)。而由于表皮细胞体积的99%由液泡所占据(Winter et al., 1993), 同时液泡中的氨基酸浓度比细胞基质和叶绿体中的低一个数量级(Winter et al., 1993)。因此表皮细胞越小, 植物叶片的πo越低(表3), 植物的πtlp也就越低(图1)。这一结果与之前的研究一致(Cutler et al., 1977; Moore et al., 2008), 表明植物细胞的减小是植物耐旱的一个重要机制(Cutler et al., 1977), 植物通过对叶片细胞大小的调整来影响植物的πo, 从而适应不同的水分环境。P/S是植物耐旱性的一个特征(Cao, 2000)。它与SWC、πo和πtlp有很强的负相关关系(图2; 表3)。我们推测P/S对植物叶片水分关系的影响, 可能是由P/S与叶脉密度的正相关关系所决定的(Sack & Frole, 2006)。较小的LMA意味着植物叶片的蒸腾表面积小, 减少了植物在干旱条件下对水分的需求(Poorter et al., 2009)。LMA与SWC、πo和πtlp存在负相关关系(图4; 表3)。这可能是由LD引起的(图3; 表3), 因为具有较高LD的叶片, 细胞较小, 排列紧密, 细胞壁厚(Niinemets, 2001), 能够有效地积累渗透物质(Morgan, 1984)。因此, 在跨生境的尺度上, 叶片结构特征差异特别是致密程度很好地解释了水分特征的差异, 也就是说, LD是植物耐旱性的一个重要指标。尽管在传统的Pearson相关分析中我们并没有发现LMA与ε有相关关系, 但是在排除了系统发育亲缘关系的影响后, LMA与ε存在正相关关系(表3)。说明二者之间的关系受系统发育影响, 存在关联进化。具有较高的ε的叶片在组织含水量变化一定的情况下水势改变较大。因此, 虽然叶片损失的水分较少, 但是叶片和土壤之间的水势梯度却较大(Niinemets, 2001; Bartlett et al., 2012b), 从而使植物能够从干旱的土壤中吸收水分。
另一方面, 叶片结构能够影响植物的光合能力, 不同的叶片结构对光合能力的影响不同。表皮细胞可以聚焦光线(Vogelman et al., 1996a; Smith et al., 1997; Brodersen & Vogelmann, 2007), 栅栏组织将光线传递到叶片深层(Vogelman et al., 1996b; Smith et al., 1997)、吸收红蓝光(Terashima et al., 2011), 海绵组织则多次反射光线(Vogelman et al., 1996b; Smith et al., 1997), 使得光线特别是绿光得到充分吸收(Terashima et al., 2011)。叶片厚度与Aa存在正相关关系, 与R存在负相关关系(表3)。叶片厚度的增加使得CO2的扩散阻力变大(Hanba et al., 1999), 因此需要较大的单位叶面积暴露在细胞间隙的叶肉面积(Smes)来增加CO2的溶解面积和扩散途径(Terashima et al., 2011)。而这主要是通过栅栏组织的增加来实现的(Parkhurst, 1994)。此外, 叶片栅栏组织厚度增加, 植物的捕光能力和光合酶的单位面积浓度也增加。因此, 植物很可能是通过调节栅栏组织结构来影响Aa。有研究表明, LD越大, CO2气相阶段的扩散阻力也越大(Niinemets, 1999; Fu et al., 2012)。LD的增加是由细胞壁和其他木质化的结构成分的增加引起的, 因此LD与单位质量的氮含量(Nm)负相关(Niinemets, 1999), LD越大, 植物叶片的光合组织越少。这些都导致了LD与Am的负相关关系, 与Rm正相关关系的存在(图6)。同时与Wright等(2004)和Hassiotou等(2010)结果类似, LMA与Am存在负相关关系, 与Rm存在正相关关系(图5), 这些关系受系统发育的影响, 存在关联进化。高LMA的植物叶片, CO2从气孔到叶绿体的扩散路径长, 低层的叶绿体受到内部的遮阴严重, 使得叶片在相同蛋白质含量的情况下Am值较低(Parkhurst, 1994)。而且, 高LMA的植物可能会将N更多地投向非光合的叶片组织(Poorter & Evans, 1998; Green & Kruger, 2001)。
本研究表明植物在耐失水能力上的投入可能同时在结构和养分投入上限制了植物的光合能力。尽管Aa和R与πo和πtlp不存在相关关系, 但是在去除了系统发育的影响后, Am与πo和πtlp存在正相关关系, Rm与πo和πtlp存在正相关关系(表3; 图7)而ε与Am存在负相关关系(图7), 这表明植物的耐失水能力与植物的光合能力在进化上存在权衡关系。πtlp主要是由πo所决定的(表3) (Bartlett et al., 2012b), 低的πo需要高浓度的渗透物质, 这些物质在细胞质内一方面促进水分的吸收和保留, 另一方面保护大分子和膜系统免受干旱引起的损伤(Hessini et al., 2009)。然而这些兼容性的渗透物质的生产成本很高(Patakas et al., 2002)。与此同时, 高的ε需要厚的细胞壁, 导致N向光合组织的投资减少(Onoda et al., 2004; Feng et al., 2009)。这使得LD和LMA值增加(图3, 图4), CO2扩散阻力增加(Tomas et al., 2013)从而导致了植物光合能力的下降(表3; 图7)和维持成本的增加。因此, 我们推断由于叶片自身结构的限制使得植物在提高耐失水能力(高的P/S、LMA、LD)的同时限制了植物光合能力的增加(低的LD、LMA), 植物的这种沿水分梯度和光梯度的结构和功能的分化可能是大戟科植物在热带雨林中分布格局的形成和物种共存的重要机制(Smith & Huston, 1989)。
研究中我们还发现, 有些性状之间的相关关系无论是用传统的Pearson相关分析还是用系统发育独立对照差相关分析, 都存在一致的相关关系, 如UET与πtlp之间的正相关关系(图1), LD与πtlp之间的负相关关系(图2)。对于这类性状, 这两种相关分析方法都能够很好地揭示它们之间的关系。有些性状, Pearson相关分析并不显示它们之间有相关关系存在, 而用系统发育独立对照差相关分析则显示了相关关系的存在, 如P/S与πtlp (图2)、LD与Rm (图6A、6C)、LD与Am (图6B、6D)等。对于这类性状如果用Pearson相关分析则犯了I类错误(type I error)。还有些性状用Pearson相关分析发现它们之间存在相关关系, 用系统发育独立对照差相关分析发现这种相关关系消失, 如PT与R和Aa等(表3)。对于这类性状用Pearson相关分析则犯了II类错误(type II error)。因此, 这些结果表明对于亲缘关系较近的植物类群, 必须要使用系统发育独立对照差相关分析(Felsen- stein, 1985)。否则, 其结果可信度会受到质疑。
综上所述, 小生境内大戟科植物幼树表现出了一定结构和功能的趋同性, 且植物的耐失水能力与植物的光合能力之间存在权衡关系。由于植物叶片结构的限制, 植物在水分梯度和光梯度上发生生态位分化, 从而使物种能够共存、热带雨林能够维持较高的生物多样性。此外, 本研究还表明, 在研究亲缘关系较近的植物类群的性状相关关系时, 使用系统发育独立对照差相关分析是十分必要的。
致谢
感谢中国科学院西双版纳热带雨林生态系统研究站提供的帮助和数据支持。
参考文献
Effects of the interaction between drought and shade on water relations, gas exchange and morphological traits in cork oak (Quercus suber L.) seedlings
Sclerophylly and leaf anatomical traits of five field-grown olive cultivars growing under drought conditions
Rapid determination of comparative drought tolerance traits: using an osmometer to predict turgor loss point
The determinants of leaf turgor loss point and prediction of drought tolerance of species and biomes: a global meta-analysis
Drought induced mortality in tropical heath forest
Do epidermal lens cells facilitate the absorptance of diffuse light?
Multispecies coexistence of trees in tropical forests: spatial signals of topographic niche differentiation increase with environmental heterogeneity

Neutral and niche theories give contrasting explanations for the maintenance of tropical tree species diversity. Both have some empirical support, but methods to disentangle their effects have not yet been developed. We applied a statistical measure of spatial structure to data from 14 large tropical forest plots to test a prediction of niche theory that is incompatible with neutral theory: that species in heterogeneous environments should separate out in space according to their niche preferences. We chose plots across a range of topographic heterogeneity, and tested whether pairwise spatial associations among species were more variable in more heterogeneous sites. We found strong support for this prediction, based on a strong positive relationship between variance in the spatial structure of species pairs and topographic heterogeneity across sites. We interpret this pattern as evidence of pervasive niche differentiation, which increases in importance with increasing environmental heterogeneity.
Functional convergence in hydraulic architecture and water relations of tropical savanna trees: from leaf to whole plant
Leaf anatomy and chlorophyll content of 12 woody species in contrasting light conditions in a bornean heath forest
Tropical forests of Xishuangbanna, China
Water stress affects leaf anatomy, gas exchange, water relations and growth of two avocado cultivars
Xylem traits mediate a trade-off between resistance to freeze-thaw-induced embolism and photosynthetic capacity in overwintering evergreens
The importance of cell size in the water relations of plants
Contribution of intercellular reflectance to photosynthesis in shade leaves
Chloroplast to leaf. In: Smith W, Vogelmann T, Critchley C eds
Phylogenies and the comparative method
Evolutionary tradeoffs for nitrogen allocation to photosynthesis versus cell walls in an invasive plant
Stem hydraulic traits and leaf water-stress tolerance are co-ordinated with the leaf phenology of angiosperm trees in an asian tropical dry karst forest
BACKGROUND AND AIMS: The co-occurring of evergreen and deciduous angiosperm trees in Asian tropical dry forests on karst substrates suggests the existence of different water-use strategies among species. In this study it is hypothesized that the co-occurring evergreen and deciduous trees differ in stem hydraulic traits and leaf water relationships, and there will be correlated evolution in drought tolerance between leaves and stems. METHODS: A comparison was made of stem hydraulic conductivity, vulnerability curves, wood anatomy, leaf life span, leaf pressure-volume characteristics and photosynthetic capacity of six evergreen and six deciduous tree species co-occurring in a tropical dry karst forest in south-west China. The correlated evolution of leaf and stem traits was examined using both traditional and phylogenetic independent contrasts correlations. KEY RESULTS: It was found that the deciduous trees had higher stem hydraulic efficiency, greater hydraulically weighted vessel diameter (D(h)) and higher mass-based photosynthetic rate (A(m)); while the evergreen species had greater xylem-cavitation resistance, lower leaf turgor-loss point water potential (pi(0)) and higher bulk modulus of elasticity. There were evolutionary correlations between leaf life span and stem hydraulic efficiency, A(m), and dry season pi(0). Xylem-cavitation resistance was evolutionarily correlated with stem hydraulic efficiency, D(h), as well as dry season pi(0). Both wood density and leaf density were closely correlated with leaf water-stress tolerance and A(m). CONCLUSIONS: The results reveal the clear distinctions in stem hydraulic traits and leaf water-stress tolerance between the co-occurring evergreen and deciduous angiosperm trees in an Asian dry karst forest. A novel pattern was demonstrated linking leaf longevity with stem hydraulic efficiency and leaf water-stress tolerance. The results show the correlated evolution in drought tolerance between stems and leaves.
Procedures for the analysis of comparative data using phylogenetically independent contrasts
Light-mediated constraints on leaf function correlate with leaf structure among deciduous and evergreen tree species
The influence of leaf thickness on the CO2 transfer conductance and leaf stable carbon isotope ratio for some evergreen tree species in Japanese warm-temperate forests
Photosynthesis at an extreme end of the leaf trait spectrum: How does it relate to high leaf dry mass per area and associated structural parameters?
Leaf dry mass per area (LMA) is a composite parameter relating to a suite of structural traits that have the potential to influence photosynthesis. However, the extent to which each of these traits contributes to variation in LMA and photosynthetic rates is not well understood, especially at the high end of the LMA spectrum. In this study, the genus Banksia (Proteaceae) was chosen as a model group, and key structural traits such as LMA, leaf thickness, and density were measured in 49 species. Based on the leaf trait variation obtained, a subset of 18 species displaying a wide range in LMA of 134-507 g m(-2) was selected for analyses of relationships between leaf structural and photosynthetic characteristics. High LMA was associated with more structural tissue, lower mass-based chlorophyll and nitrogen concentrations, and therefore lower mass-based photosynthesis. In contrast, area-based photosynthesis did not correlate with LMA, despite mesophyll volume per area increasing with increases in LMA. Photosynthetic rate per unit mesophyll volume declined with increasing LMA, which is possibly associated with structural limitations and, to a lesser extent, with lower nitrogen allocation. Mesophyll cell wall thickness significantly increased with LMA, which would contribute to lower mesophyll conductance at high LMA. Photosynthetic nitrogen use efficiency and the nitrogen allocation to Rubisco and thylakoids tended to decrease at high LMA. The interplay between anatomy and physiology renders area-based photosynthesis independent of LMA in Banksia species.
Effect of water stress on growth, osmo- tic adjustment, cell wall elasticity and water-use efficiency in Spartina alterniflora
The environment, not space, dominantly structures the landscape patterns of the richness and composition of the tropical understory vegetation
Light-guiding function of foliar sclereids in the evergreen sclerophyll phillyrea latifolia: a quantitative approach
Penetration of UV-A, UV-B and blue light through the leaf trichome layers of two xeromorphic plants, olive and oak, measured by optical fibre microprobes
Foliar sclereids of Olea europaea may function as optical fibres
Pressure-volume relation- ships in non-rehydrated tissue at various water deficits
Tolerance to low leaf water status of tropical tree seedlings is related to drought performance and distribution
Spatial distribution and interspecific associations of tree species in a tropical seasonal rain forest of China
Studying the spatial pattern and interspecific associations of plant species may provide valuable insights into processes and mechanisms that maintain species coexistence. Point pattern analysis was used to analyze the spatial distribution patterns of twenty dominant tree species, their interspecific spatial associations and changes across life stages in a 20-ha permanent plot of seasonal tropical rainforest in Xishuangbanna, China, to test mechanisms maintaining species coexistence. Torus-translation tests were used to quantify positive or negative associations of the species to topographic habitats. The results showed: (1) fourteen of the twenty tree species were negatively (or positively) associated with one or two of the topographic variables, which evidences that the niche contributes to the spatial pattern of these species. (2) Most saplings of the study species showed a significantly clumped distribution at small scales (0-10 m) which was lost at larger scales (10-30 m). (3) The degree of spatial clumping deceases from saplings, to poles, to adults indicates that density-dependent mortality of the offspring is ubiquitous in species. (4) It is notable that a high number of positive small-scale interactions were found among the twenty species. For saplings, 42.6% of all combinations of species pairs showed positive associations at neighborhood scales up to five meters, but only 38.4% were negative. For poles and adults, positive associations at these distances still made up 45.5% and 29.5%, respectively. In conclusion, there is considerable evidence for the presence of positive interactions among the tree species, which suggests that species herd protection may occur in our plot. In addition, niche assembly and limited dispersal (likely) contribute to the spatial patterns of tree species in the tropical seasonal rain forest in Xishuangbanna, China.
Establishment of Xishuangbanna tropical forest dynamics plot: species compositions and spatial distribution patterns
西双版纳热带森林动态监测样地——树种组成与空间分布格局
Seasonal differentiation in density-dependent seedling survival in a tropical rain forest
Ecological differentiation in xylem cavitation resistance is associated with stem and leaf structural traits
Internal leaf anatomy and photosynthetic resource-use efficiency: inter- specific and intraspecific comparisons
Leaf mass per unit area (LMA) and internal leaf anatomy often affect net gas exchange because of their effects on internal CO2 conductance to the site of carboxylation, internal shading, competition for CO2 among carboxylation sites, nitrogen concentration and its partitioning. To evaluate effects of LMA and leaf anatomy on CO2 assimilation, water-use efficiency (WUE) and nitrogen-use efficiency (NUE), we measured LMA, leaf thickness, the thickness of mesophyll components, and gas exchange rates at ambient CO2 concentration in leaves of six woody deciduous and evergreen species with different leaf life spans. In two species, CO2 assimilation was also estimated at saturating CO2 concentrations. There were interspecific differences in all morphological variables studied. Long-lived leaves had higher LMA and were thicker than short-lived leaves. Species with high LMA had low assimilation rates and NUE, both in ambient and saturating CO2 concentrations. Thus, in species with high LMA, assimilation was reduced by non-stomatal limitations, possibly because of a lower allocation of N to the photosynthetic machinery than in species with low LMA. Within a species, thicker leaves tended to have a lower tissue density. In intraspecific comparisons under field conditions, increasing internal air volume had positive effects on WUE, probably because of enhanced internal CO2 conductance to the site of carboxylation. We conclude that, in interspecific comparisons, different patterns of N partitioning strongly influence NUE, whereas in intraspecific comparisons, internal leaf anatomy is a key factor regulating resource-use efficiency.
Functional convergence in plant responses to the environment
Much comparative ecophysiological research has focused on contrasting species-specific behavior or ecological strategies with regard to regulation of basic physiological processes such as transpiration, photosynthesis and growth, leading to an emphasis on divergence rather than convergence in plant functioning. This review highlights selected examples in which substantial functional convergence among taxonomically, phylogenetically and architecturally diverse species has been revealed by applying appropriate scaling factors and identifying universal constraints or trade-offs. Recent empirical and theoretical scaling models emphasize the strong role that plant size, architecture, allometry and chemistry play in constraining functional traits related to water and carbon economy and growth. Taken together, the findings summarized here strongly suggest that there are a limited number of physiological solutions to a given problem of plant adaptation to the environment. Comparative ecophysiological studies will therefore benefit from consideration of the constraints that plant anatomical, structural and chemical attributes place on physiological functioning.
Adaptations of higher plant cell walls to water loss: drought vs desiccation
Osmoregulation and water stress in higher plants
Research review. Components of leaf dry mass per area-thickness and density-alter leaf photo- synthetic capacity in reverse directions in woody plants
Global-scale climatic controls of leaf dry mass per area, density, and thickness in trees and shrubs
Role of mesophyll diffusion conductance in constraining potential photosynthetic productivity in the field
Limited mesophyll diffusion conductance to CO(2) (g(m)) can significantly constrain plant photosynthesis, but the extent of g(m)-limitation is still imperfectly known. As g(m) scales positively with foliage photosynthetic capacity (A), the CO(2) drawdown from substomatal cavities (C(i)) to chloroplasts (C(C), C(i)-C(C)=A/g(m)) rather than g(m) alone characterizes the mesophyll diffusion limitations of photosynthesis. The dependencies of g(m) on A, foliage structure (leaf dry mass per unit area, M(A)), and the resulting drawdowns across a dataset of 81 species of contrasting foliage structure and photosynthetic potentials measured under non-stressed conditions were analysed to describe the structure-driven potential photosynthetic limitations due to g(m). Further the effects of key environmental stress factors and leaf and plant developmental alterations on g(m) and CO(2) drawdown were evaluated and the implications of varying g(m) on foliage photosynthesis in the field were simulated. The meta-analysis demonstrated that g(m) of non-stressed leaves was negatively correlated with M(A), and despite the positive relationship between g(m) and A, the CO(2) drawdown was larger in leaves with more robust structure. The correlations were stronger with mass-based g(m) and A, probably reflecting the circumstance that mesophyll diffusion is a complex three-dimensional process that scales better with mesophyll volume-weighted than with leaf area-weighted traits. The analysis of key environmental stress effects on g(m) and CO(2) drawdowns demonstrated that the effect of individual stresses on CO(2) drawdowns varies depending on the stress effects on foliage structure and assimilation rates. Leaf diffusion limitations are larger in non-senescent older leaves and also in senescent leaves, again reflecting more robust leaf structure and/or non-co-ordinated alterations in leaf photosynthesis and g(m). According to simulation analyses, in plants with a larger part of the overall diffusion conductance from the ambient atmosphere to the chloroplasts in the mesophyll, photosynthesis is less sensitive to changes in stomatal conductance. Accordingly, in harsher environments that support vegetation with tougher long-living stress-tolerant leaves with lower g(m), reductions in stomatal conductance that are common during stress periods are expected to alter photosynthesis less than in species where a larger part of the total diffusion limitation is determined by stomata. While structural robustness improves plant performance under environmental stress, low g(m) and inherently large CO(2) drawdown in robust leaves limits the photosynthesis of these plants more severely under favourable conditions when stomatal conductance is high. The differences in overall responsiveness to environmental modifications of plants with varying g(m) need consideration in current large-scale ecosystem productivity models.
The relationship between anatomy and photosynthetic performance of heterobaric leaves
Heterobaric leaves show heterogeneous pigmentation due to the occurrence of a network of transparent areas that are created from the bundle sheaths extensions (BSEs). Image analysis showed that the percentage of photosynthetically active leaf area (Ap) of the heterobaric leaves of 31 plant species was species dependent, ranging from 91% in Malva sylvestris to only 48% in Gynerium sp. Although a significant portion of the leaf surface does not correspond to photosynthetic tissue, the photosynthetic capacity of these leaves, expressed per unit of projected area (Pmax), was not considerably affected by the size of their transparent leaf area (At). This means that the photosynthetic capacity expressed per Ap (P*max) should increase with At. Moreover, the expression of P*max could be allowing the interpretation of the photosynthetic performance in relation to some critical anatomical traits. The P*max, irrespective of plant species, correlated with the specific leaf transparent volume (lambda(t)), as well as with the transparent leaf area complexity factor ((CF)A(t)), parameters indicating the volume per unit leaf area and length/density of the transparent tissues, respectively. Moreover, both parameters increased exponentially with leaf thickness, suggesting an essential functional role of BSEs mainly in thick leaves. The results of the present study suggest that although the Ap of an heterobaric leaf is reduced, the photosynthetic performance of each areole is increased, possibly due to the light transferring capacity of BSEs. This mechanism may allow a significant increase in leaf thickness and a consequent increase of the photosynthetic capacity per unit (projected) area, offering adaptive advantages in xerothermic environments.
Allocation of nitrogen to cell walls decreases photosynthetic nitrogen-use efficiency
Diffusion of CO2 and other gases inside leaves
The role of organic solute and ion accumulation in osmotic adjustment in drought-stressed grapevines
Photosynthetic nitrogen-use efficiency of species that differ inherently in specific leaf area
Causes and consequences of variation in leaf mass per area (LMA): a meta-analysis
Epidermal focussing and effects upon photosynthetic light-harvesting in leaves of oxalis
The evolution of plant functional variation: traits, spectra, and strategies
Changes in foliar spectral reflectance and chlorophyll fluorescence of four temperate species following branch cutting
Spectral reflectance and chlorophyll fluorescence are rapid non-invasive methods that can be used to quantify plant stress. Because variation in ambient light (e.g., diurnal patterns of solar radiation) may have a confounding effect on these measurements, branches are often excised in the field and then measured under controlled conditions in the laboratory. We studied four temperate tree species (Abies balsamea (L.) Mill. (balsam fir), Betula papyrifera var. cordifolia (Regel) Fern. (paper birch), Picea rubens Sarg. (red spruce) and Sorbus americana Marsh. (mountain-ash)) to determine how quickly reflectance and fluorescence change following branch cutting. We hypothesized that conifer species, which have tough xeromorphic foliage, would exhibit changes more slowly than broadleaf species. Furthermore, we hypothesized that keeping broadleaf samples cool and moist would delay the onset of reflectance and fluorescence changes. In one set of experiments, we did not use any treatments to maintain the freshness of cut branches. During the first 12 h following cutting, changes in reflectance and fluorescence were slight for all species. Two or 3 days after branch cutting, the two conifers still showed only small changes in the ratio of variable to maximum fluorescence (Fv/Fm) and most reflectance indices, whereas paper birch and mountain-ash showed larger and more rapid declines in Fv/Fm and most reflectance indices. We attribute these declines to loss of water. As a consequence of xeromorphic leaf structure, the conifers were better able to minimize water loss than the two broadleaf species. In another experiment, paper birch that had been kept cool and moist after cutting showed only slight changes in fluorescence and reflectance, even after 3 days, indicating that with careful handling the time interval between collection and measurement of reflectance and fluorescence of many broadleaf specimens can be extended to several days. We conclude that measurements of reflectance and fluorescence need not be made in situ to be accurate and reliable.
Leaf structural diversity is related to hydraulic capacity in tropical rain forest trees
The hydraulic resistance of the leaf (R1) is a major bottleneck in the whole plant water transport pathway and may thus be linked with the enormous variation in leaf structure and function among tropical rain forest trees. A previous study found that R1 varied by an order of magnitude across 10 tree species of Panamanian tropical lowland rain forest. Here, correlations were tested between R1 and 24 traits relating to leaf venation and mesophyll structure, and to gross leaf form. Across species, R1 was related to both venation architecture and mesophyll structure. R1 was positively related to the theoretical axial resistivity of the midrib, determined from xylem conduit numbers and dimensions, and R1 was negatively related to venation density in nine of 10 species. R1 was also negatively related to both palisade mesophyll thickness and to the ratio of palisade to spongy mesophyll. By contrast, numerous leaf traits were independent of R1, including area, shape, thickness, and density, demonstrating that leaves can be diverse in gross structure without intrinsic trade-offs in hydraulic capacity. Variation in both R1-linked and R1-independent traits related strongly to regeneration irradiance, indicating the potential importance of both types of traits in establishment ecology.
Leaf pubescence, water relations and chlorophyll fluorescence in two subspecies of Lotus creticus L
The objective of the present study was to compare the plant morphology, water relations and photochemical efficiency of photosystem 2 in two wild. Mediterranean species Lotus creticus creticus and Lotus creticus cytisoides. L. creticus creticus showed higher density of trichomes and stomatal density on the adaxial leaf surface than L. creticus cytisoides, whereas L. creticus cytisoides showed higher stomatal density in abaxial surface than L. creticus creticus. These morphological traits promoted clear differences in leaf surface water retention and leaf reflectance. Leaf water potential and photochemical efficiency were lower in L. creticus cytisoides than in L. creticus creticus.]]>
Temperature response of mesophyll conductance in cultiv- ated and wild oryza species with contrasting mesophyll cell wall thickness
A theory of the spatial and temporal dynamics of plant communities
Leaf form and photosynthesis
Ecological and evolutionary determinants of a key plant functional trait: wood density and its community-wide variation across latitude and elevation
Wood density is an important trait in trees indicative of their life history and mechanical and physiological strategies. The following examines the evolutionary ecology of wood density using a large database for seed plants. In particular, we focused on the geographic and phylogenetic variation in wood density for both gymnosperms and angiosperms. A phylogenetic supertree was constructed for over 4600 taxa, allowing for comprehensive analyses of divergences across the seed plant phylogeny. Community-wide means and variances for wood densities were quantified for 171 standardized forest communities. Wood density was generally highly conserved across the entire seed plant phylogeny, yet large divergences were found within the rosid clade. Geographic and community variation in wood density, however, was significantly lower in temperate and high elevation communities, dominated by gymnosperms, than in tropical lowland communities, dominated by angiosperms, suggesting an increase in trait and, to some extent, clade filtering with latitude and elevation. Together, our results support the notion that both biotic and abiotic forces have been important in the evolution of wood density as well as in controlling the observed trait mean and variance across geographic gradients.
MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods
Leaf functional anatomy in relation to photosynthesis
Importance of leaf anatomy in determining mesophyll diffusion conductance to CO2 across species: quantitative limitations and scaling up by models
Plant tissue optics
Focusing of light by leaf epidermal cells
The functional-significance of palisade tissue—Penetration of directional versus diffuse light
Leaves and light capture: light propagation and gradients of carbon fixation within leaves
Trade-offs in low-light CO2 exchange: a component of variation in shade tolerance among cold temperate tree seedlings
Subcellular volumes and metabolite concentrations in barley leaves
The worldwide leaf economics spectrum
Bringing together leaf trait data spanning 2,548 species and 175 sites we describe, for the first time at global scale, a universal spectrum of leaf economics consisting of key chemical, structural and physiological properties. The spectrum runs from quick to slow return on investments of nutrients and dry mass in leaves, and operates largely independently of growth form, plant functional type or biome. Categories along the spectrum would, in general, describe leaf economic variation at the global scale better than plant functional types, because functional types overlap substantially in their leaf traits. Overall, modulation of leaf traits and trait relationships by climate is surprisingly modest, although some striking and significant patterns can be seen. Reliable quantification of the leaf economics spectrum and its interaction with climate will prove valuable for modelling nutrient fluxes and vegetation boundaries under changing land-use and climate.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}