高山恶劣的自然环境条件如低温、大风、多雨以及生长季短等不利因素限制了传粉昆虫的种类及其活动能力和访花频率(Arroyo et al ., 1982 , 1985 ; Bingham & Orthner, 1998 ), 导致到达柱头的异花花粉数量减少, 继而影响植物的生殖成功率(张大勇和姜新华, 2001 )。但已有的研究表明, 具有异花传粉繁育系统的植物在高山植物群落中普遍存在, 它们往往采取延长花寿命(Arroyo et al ., 1985 ; Bingham & Orthner, 1998 ; Blionis & Vokou, 2002 ; Steinacher & Wagner, 2010 )、增加对繁殖结构的资源分配(Fabbro & Körner, 2004 )等策略来适应传粉昆虫的缺乏, 以保证种群的延续。同时, 一些高山植物会选择自花授粉这种以降低后代适合度为代价的方式保证种群的延续(Schoen et al ., 1996 ; 段元文等, 2007b )。
海拔带来的环境差异会使各类昆虫的活动受到限制或促进, 导致不同海拔地区访花者种类和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b )。在高山植物群落中, 鸟类和蝴蝶的传粉作用逐渐减弱, 熊蜂和蝇类成为主要的传粉昆虫(何亚平和刘建全, 2004 ; Duan et al ., 2007 )。对毛茛科植物Ranunculus acris 的研究发现, 低海拔地区昆虫的访花频率是高海拔地区的3倍(Totland, 2001 )。对凤仙花属(Impatiens )植物的研究发现, 高海拔地区和低海拔地区的传粉昆虫种类存在差异, 熊蜂在高海拔地区的花粉传递中扮演着重要的角色(Tian et al ., 2004 ; 肖乐希和刘克明, 2009 ; 肖乐希, 2009 ; 毛志斌等, 2011 )。对露蕊乌头(Aconitum gymnandrum )的研究表明, 低海拔种群的熊蜂种类(3种)多于高海拔种群(2种), 且熊蜂在低海拔种群的总访花频率远远高于高海拔种群(张挺峰等, 2006 )。对横断山区大钟花(Megacodon stylophorus )的研究亦得到相似结果(Meng et al ., 2012 )。对海南凤仙花(Impatiens hainanensis )的传粉生物学研究亦发现, 高海拔种群的传粉者以绿条无垫蜂(Amegilla zonata )为主, 低、中海拔种群以黄黑无垫蜂(Amegilla leptocoma )为主, 且黄黑无垫蜂在低、中海拔种群的访问频率远高于高海拔种群(钟云芳等, 2014 )。
高山环境下, 由于传粉者的不足使得大多数植物存在花粉限制(段元文等, 2007b ; 司庆文等, 2010 ; 钟云芳等, 2014 )。面对传粉者缺乏的选择压力, 植物会采取多样的策略提高对传粉昆虫的吸引力, 如提高花展示(邵剑文等, 2008 ; 彭德力等, 2012 )、提供生殖场所(Fishman & Hadany, 2013 )等, 随着海拔升高, 一些植物通过延长花寿命获得交配的机会(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 ); 而低温环境下, 一些植物则通过提高花序内或花内温度来达到吸引传粉昆虫的目的(Stanton & Galen, 1989 )。如水母雪兔子(Saussurea medusa )通过花序温度的升高使熊蜂在花序上的停留时间超过1 min, 这在一定程度上对传粉者起到了吸引作用(Tsukaya, 2002 )。对冬季开花的毛茛科铁筷子属植物Helleborus foetidus 和Helleborus bocconei 的研究亦发现, 花内温度的升高使传粉者更加活跃(Vesprini & Pacini, 2010 )。同时, 一些植物往往会进化出各异的自交机制, 如通过雌雄蕊位置的变化促进花粉传递以保障成功繁殖。总结已有的研究发现, 植物雌雄蕊位置关系的改变方式呈现出多样化的特征, 如花丝伸长(段元文等, 2007a , 2007b )、花柱伸长(侯勤正等, 2009 )、花柱卷曲(孙杉等, 2010 )等。
绿绒蒿属(Meconopsis )植物绝大多数分布在以横断山脉-喜马拉雅为中心的高山环境中(庄璇, 1981 ), 作为传统藏药, 目前的研究主要集中在其化学成分及药理活性方面(尚小雅等, 2006 ; Shang et al ., 2006 ; Zhou et al ., 2009 ), 一些学者对本属植物进行了系统分类方面的探讨(Meng et al ., 2006 ; Singhal & Kumar, 2008 ; Yang et al ., 2012 )。作为一类长期适应高山环境的植物, 目前对绿绒蒿属植物的繁殖特征、适应进化等方面尚缺乏系统研究(Dar et al ., 2010 )。
与本属其他种以较小的种群分布于狭小的范围内相比, 全缘叶绿绒蒿(Meconopsis integrifolia )有着较广的分布范围(Sulaiman & Babu, 1996 )。作为一年生或多年生草本, 它主要分布于青藏高原海拔2500-5000 m的范围内, 一般在5月底到6月初进入花期, 此时属于高山植物的生长季早期, 传粉昆虫数量少, 异花传粉困难。全缘叶绿绒蒿会采取何种繁殖策略以适应高山环境, 这种繁殖策略是否随着环境的变化而发生改变? 为此, 本研究通过在四川省阿坝州巴郎山地区对不同海拔样地中的全缘叶绿绒蒿2年的连续观察和人工授粉试验, 重点回答以下问题: (1)全缘叶绿绒蒿的主要传粉昆虫是什么?异花传粉者的访花频率在不同环境中是否有差异?(2)它的繁育系统特点是什么?(3)其繁殖策略是否表现出随环境变化的特征?
1 材料和方法
1.1 研究地点概况
观察和试验于2013年、2014年5月下旬至9月下旬在四川省阿坝藏族羌族自治州汶川县卧龙自然保护区巴郎山(102.90°-102.95° E, 30.88°-30.91° N, 海拔4484 m)进行, 巴郎山年平均气温8.6 ℃ (1月平均气温-1.7 ℃, 7月平均气温17.0 ℃), 年降水量700-900 mm, 降水集中在5-9月。在充分踏查的基础上, 共选择了5个全缘叶绿绒蒿研究样地(海拔高度分别为4452、4215、4081、3841和3681 m)。
1.2 研究材料
全缘叶绿绒蒿为罂粟科绿绒蒿属一年生或多年生草本, 主要分布于海拔2500 m的林缘到海拔5000 m的雪线区间, 生境多为高山草甸(图1A )。植株高15-150 cm (图1C ), 花黄色(图1B ), 通常3-5朵生于茎上部叶腋内; 雄蕊多数; 子房卵形; 蒴果。花期5-8月。
图1 全缘叶绿绒蒿生境及各项特征。A, B, C, D和E分别为全缘叶绿绒蒿的生境、花形、植株形态、开花早期和开花末期的雌雄蕊位置。图1B中a、b分别表示柱头高度和最高雄蕊高度。
Fig. 1 Habitat and floral traits of Meconopsis integrifolia in this study. A, Habitat of M . integrifolia; B, Flower; C, Whole plant; D and E, Showing the relative position between stamen and stigma at the early and end of blossom, respectively. a and b in Fig. 1B show the height of stigma and stamen, respectively.
1.3 研究方法
1.3.1 花部综合特征
1.3.1.1 开花过程 各样地分别随机选取并标记20个即将开放的花蕾, 从2013年6月11日起同时对其进行开放期间的全程观察(每天一次), 每天于9:00-10:00记录标记花的状态, 直至标记的20朵花花瓣凋落柱头萎蔫后结束观察。每次观察记录并测定花是否开放(以花萼片脱落为花开放的标志)、柱头高度(图1B )及柱头表面有无花粉、最高雄蕊高度(图1B )、花药散粉时间(以单花某一花药开始散粉的时间为准)、散粉结束时间(以单花所有花药散粉结束的时间为准)、花萎蔫时间(以花瓣凋落柱头萎蔫为准), 以用于计算花寿命、花药散粉持续时间及雌雄蕊空间位置变化。花寿命即从标记的花蕾花萼片脱落时至花瓣凋落柱头萎蔫的时间; 花药散粉持续时间即花药开始散粉时至所有花药散粉结束的 时间。
1.3.1.2 花粉胚珠比例(P/O) 各样地分别随机选取即将开放的花蕾20个固定于FAA固定液中, 并带回实验室。室内分别将每朵花的花药全部取出, 置于盛有蒸馏水的5 mL离心管中, 充分挤碎, 使花粉散布于离心管内, 加水定容至5 mL, 充分摇匀后用移液枪取出1 μL, 置于显微镜下计数花粉数, 重复10次并计算平均值, 得出每花的平均花粉数目; 剖开子房, 光学显微镜(Olympus CX21, Olympus Corporation, Tokyo, Japan)下统计胚珠数目。计算花粉胚珠比例(P/O), 以初步判断全缘叶绿绒蒿的繁育系统类型。
1.3.2 柱头可授性与花粉活力检测
在2013年盛花期进行柱头可授性与花粉活力检测。采用联苯胺-过氧化氢法(叶要妹等, 2007 )检测柱头可授性。分别于各样地随机选取5朵当天开放的花并标记, 此后每天均随机选取当日开放的5朵花直至最早标记的花柱头完全变黑, 收集各天标记的花, 将其柱头浸入凹面载玻片中, 若柱头具有可授性则柱头周围反应液呈蓝色并有大量气泡产生。用-和+表示观察到的情况。-表示柱头不具有可授性, 即无气泡与变蓝情况; +表示柱头具有可授性, 即有轻微的变蓝和气泡产生; ++表示柱头具有较强可授性, 即有较多的气泡和比较明显的变蓝情况; +++表示柱头具有极强可授性, 即有大量气泡和非常明显的变蓝情况。
用TTC法(潘成臣等, 2012 )测定花粉活力。各样地分别随机选取20个植株上的20个即将开放的花蕾并标记, 于散粉当天采集标记的花朵并带回, 放于4 ℃冰箱冷藏。采集当日每隔2小时取出一批进行染色, 显微镜下统计3-5个视域内被染色花粉数和未被染色花粉数, 计算花粉活力。花粉活力=被染色花粉数/(被染色花粉数+未被染色花粉数) × 100%。此后每天取出一批进行染色并统计直至花粉没有活性。
1.3.3 人工授粉试验
为检测自交亲和性及自花传粉及异花传粉对结实的贡献, 参考段元文等(2007b) 的方法, 分别于2013年和2014年对5个样地进行了5种人工授粉试验: (1)去雄套袋: 随机选择并标记20个植株上的20个将要开放的花蕾, 去除雄蕊后套袋以检测是否存在无融合生殖现象; (2)去雄不套网: 于开花初期, 随机选择并标记20个植株上的20个将要开放的花蕾, 去掉雄蕊后不做任何处理, 拟检查异花花粉对全缘叶绿绒蒿结实的贡献; (3)套网: 随机选取并标记20个植株上的20个将要开放的花蕾并套网, 拟检测全缘叶绿绒蒿自花传粉的能力以及是否产生种子; (4)人工自交; (5)人工异交: 开花初期, 随机选取20对位于不同植株上将要开放的花蕾并去雄套袋(套袋隔离的花中未发现任何昆虫), 根据同一样地中其他花的开放时间(约2天), 将每对中的一朵花授以同一植株上刚散出的花粉, 另一朵花授以10 m以外(保证异交)的植株上刚散出的花粉, 授粉后仍套袋(防止雨水冲刷柱头上的花粉和隔离访花昆虫)。另外每样地随机选取20朵花不做任何处理作为上述所有处理的对照。值得注意的是, 同种处理均选取不同植株上的花蕾。于授粉后40天(果实成熟但未开裂)收集所标记的花, 统计坐果率(fruit set rate)和每个果实的成熟种子数、不成熟种子数和败育胚珠数, 计算结实率。结实率(seed set rate)=成熟种子数/ (成熟种子数+不成熟种子数+败育种子数) × 100%。
1.3.4 访花昆虫的观察
2013年和2014年进行访花昆虫的观察。各样地访花昆虫的观察同时进行, 在样地的花开放盛期, 每天随机选取10朵正在开放的花, 在7:00-19:30之间(由于低温的缘故, 各样地7:00以前、20:00以后基本未见新的昆虫访花)连续观察并记录所有访花的昆虫, 单位时间1 h (如7:00-8:00、18:00-19:00, 以19:00-19:30的观察代表19:00-20:00), 记录访花昆虫的种类、数目、访花次数和访花行为。以一个单位时间前30 min的观察记录代表该单位时间的观察记录(如以12:00-12:30、14:00-14:30的观察记录分别代表12:00-13:00、14:00-15:00的观察), 每个样地访花昆虫的观察时间总计不少于6天和39 h (2013年和2014年各3天、19.5 h)。昆虫的访花频率用单花每小时的访问次数表示。观察结束后, 捕捉访花的昆虫并制作成标本, 带回实验室用于鉴定。访花昆虫的观察时间分别为2013年6月7日-9日和2014年6月10日-12日。
1.3.5 花内温度与环境温度测定
全缘叶绿绒蒿不分泌花蜜, 访花昆虫亦不取食花粉, 为验证其是否通过花内温度的提高吸引传粉者, 于2013年和2014年的盛花期在观察访花昆虫的同时对花内温度、环境温度进行了测定。
于各样地内每天随机选取并标记10朵正在开放的花(与访花昆虫观察同为10朵花), 在7:00-19:30 (由于环境温度大大低于昆虫的活动温度, 各样地7:00以前、20:00以后基本未见新的昆虫访花)同时测定并记录标记花的花内温度、环境温度, 单位时间1 h (如7:00-8:00、18:00-19:00, 其中以19:00-19:30的观察代表19 :00-20:00)。单位时间内每5 min测定一次花内温度、环境温度, 并以前30 min (如11:00-11:30)测定的温度代表该单位时间的温度。花内温度、环境温度用手持红外测温仪(Hcjet HT-835, 宏诚科技, 深圳)测定, 花内温度以花丝基部的温度为准, 环境温度以植株周围2 m范围内随机测定的5个点的平均温度为准。花内温度与环境温度的测定时间分别为2013年6月7-9日和2014年6月10-12日。
1.4 统计分析
采用SPSS 19.0统计分析软件进行数据分析。在比较前, 先检验数据是否符合正态分布。对于服从正态分布的数据, 采用独立样本t 检验(independent- samples t test)比较2组数据间的差异, 用单因素方差分析(one-way ANOVA)比较3组及其以上数据间的差异; 而对于不服从正态分布的数据, 采用非参数检验中2 Independent Samples比较2组数据间的差异, 用K Independent Samples比较3组及其以上数据间的差异。统计数据用平均值±标准误差表示, n 表示样本量。
2 研究结果
2.1 花部综合特征
全缘叶绿绒蒿花寿命表现出随海拔升高而延长的趋势, 其中样地1和2单花寿命((15.81 ± 0.37)、(14.11 ± 0.46)天)显著(p < 0.05)长于样地3、4和5 (分别为(13.03 ± 0.40)、(11.64 ± 0.26)、(12.61 ± 0.39)天)(表1 )。开花5天后, 花药开始散粉, 直至花冠萎蔫。花开放时首先将柱头展现于环境中, 雄蕊包围花柱, 并位于柱头下方(高度差约4.88 mm), 随着开花时间的延长, 最高花药与柱头的高度差逐渐减小(图1D , 1E; 图2 ), 开花中后期(第7-9天)花药与柱头相互接触, 造成自花花粉直接释放到柱头上(开花中后期, 约65%观察个体的花药与柱头接触; 样地1的花药没有达到柱头高度, 两者高度差约0.04 mm)。这些结果表明: 全缘叶绿绒蒿具有不完全雌雄异位的花部特征, 具有在花末期自交的可能性。
图2 各海拔样地全缘叶绿绒蒿柱头与最高花药高度差随花开放时间的变化(平均值±标准误差, n = 20)。
Fig. 2 Changes in the height difference between stigma and the highest stamen of Meconopsis integrifolia with time (mean ± SE, n = 20).
全缘叶绿绒蒿的花粉数目为(876027 ± 53243) (n = 100), 胚珠数目为(985 ± 42)(n = 100), P/O值为(901.24 ± 43.01) (n = 100)。
2.2 柱头可授性与花粉活力检测
全缘叶绿绒蒿的柱头在开花后1天有微弱的活性; 第2天到第6天过氧化物酶活性明显升高, 其中第3-5天时段内柱头活性最高; 第7-8天柱头活性有所下降; 第9天开始柱头活性明显下降, 部分样地中柱头上已检测不到过氧化物酶活性; 第10-11天柱头完全变黑, 各样地检测不到过氧化物酶活性。由表2 可知, 随着海拔的升高, 样地3和4柱头可授持续时间比样地5略有延长, 样地1和2又比样地3和4略有延长。
图3 各样地全缘叶绿绒蒿花粉活力随时间的变化(平均值±标准误差)。
Fig. 3 Changes in pollen viability of Meconopsis integrifolia with time in plots of different elevations (mean ± SE ).
各样地花粉活力随时间的变化基本一致, 即在散粉之初较高(60%以上); 之后花粉活力明显下降, 到散粉后12 h各样地花粉活力降至20%左右; 散粉后48 h花粉活力降至15%左右; 散粉后72 h花粉活力降至2%左右; 散粉后96 h花粉没有活力(图3 )。由此可知, 自然状态下花粉寿命约为2天, 观察发现, 花药散粉开始于花开放的第5天, 这就使得单花有活力的自花花粉与有可授性的柱头存在约2天(开花第5-6天)的重叠期。这些结果表明: 全缘叶绿绒蒿具有雌雄异熟的花特点, 但存在一定的重叠期。
2.3 人工授粉试验检测
不同海拔样地中人工自交的坐果率和结实率分别在60%和40%以上, 说明全缘叶绿绒蒿具有自交部分亲和的特性。在不同海拔样地中人工自交的坐果率和结实率均低于人工异交, 且海拔越低, 差异越显著(样地1和2人工自交结实率低于人工异交约15%, 样地3、4和5的差异达到了20%)(表3 )。
各样地中去雄套袋处理的花没有产生果实和种子, 说明全缘叶绿绒蒿不存在无融合生殖现象。不去雄套网处理的坐果率(30%-45%)和结实率(15%- 35%)显著低于自然对照花的坐果率(60%-75%)和结实率(35%-50%)(表3 ), 说明各样地全缘叶绿绒蒿具有自交能力, 但其通过自花花粉产生的种子数目较少。样地1和样地2, 去雄不套网处理的结实率(27.8%、31.3%)与对照(34.5%、37.8%)没有显著差异, 而样地3、4和5此处理的结实率(28.0%、32.1%、30.7%)显著低于对照(50.4%、52.4%、49.8%)(p < 0.05)(表3 ), 表明样地1和样地2中异花花粉可能是全缘叶绿绒蒿产生种子的主要花粉来源, 而样地3、4和5中自然对照花可能由于获得更多的自花花粉而产生了更多的种子。各样地中自然对照花的结实率显著低于(p < 0.05)人工异交处理的结实率(表3 ), 说明自然状态下全缘叶绿绒蒿存在一定程度的传粉限制。
2.4 访花昆虫与花内温度检测
各样地两年共计39 h的观察表明, 蝇类是全缘叶绿绒蒿最重要的异花传粉昆虫。它们不取食花粉, 但在花内的停留时间较长(>10 min), 能够携带较多花粉。访花时, 蝇类背部先接触柱头从而实现了花粉输入; 之后会在花内来回爬动, 在进入花冠底部的过程中, 其背部与正在散粉的花药接触, 离开时将这些花粉带出。蝇类在各样地的访花频率存在显著差异, 样地1访花频率最低((0.15 ± 0.09)次·花-1 ·h-1 ), 样地2访花频率最高((0.59 ± 0.04)次·花-1 ·h-1 ) (图4 )。
图4 不同海拔样地全缘叶绿绒蒿的昆虫访花频率(平均值±标准误差)。不同字母表示差异显著(p < 0.05)。
Fig. 4 Flower visiting frequency of Meconopsis integrifolia in plots of different elevations (mean ± SE ). Different letters indicate significant differences (p < 0.05).
样地1和样地2的传粉昆虫主要是蝇类, 它们在不同植株间的活动能够保证异花传粉结实, 同时, 它们在花内的活动会引起“协助自交”; 样地3、样地4和样地5传粉昆虫是蝇类和蓟马(Thripidae spp.), 蓟马等小型昆虫在花内停留时间较长(>5 min)并在柱头与花药间频繁活动, 导致花粉在同一朵花内的传递。
环境温度的测定发现, 各海拔样地7:00环境温度均低于5 ℃, 19:00环境温度均低于3.5 ℃, 这大大低于昆虫的活动温度(贺海霞, 2006 ; 朱建华等, 2006 , 2010 ); 且7:00以前、20:00以后既未见昆虫飞行亦未见新的昆虫进入花内, 此时间段花冠不同程度的闭合, 昆虫蛰伏于花内。由此初步推测, 每日7:00以前和20:00以后没有昆虫访花。
花内温度、环境温度测定发现, 每日7:00-9:00花内温度显著高于环境温度(p < 0.05), 7:00高出2.57-5.83 ℃; 之后环境温度迅速升高, 12:00-15:00环境温度显著高于花内温度(p < 0.05); 17:00后环境温度迅速下降, 花内温度(17:00各样地花内温度平均为5.48 ℃)逐渐升高, 18:00花内温度基本高于环境温度, 到19:00花内温度(各样地花内温度平均为6.84 ℃)显著高于环境温度(p < 0.05), 高出2.5-4.9 ℃ (图5 )。一天中访花的蝇类数量随花内温度与环境温度间温差的变化呈现出规律性的变化, 即当花内温度高于环境温度时, 蝇类数量较多, 当花内温度低于环境温度时, 蝇类数量较少, 甚至没有。各海拔样地中, 花内温度与环境温度的差值与访花蝇类数量存在极显著(p < 0.01)的相关关系(样地5为显著, p = 0.018), 且两者呈现较好的线性关系, 即样地1: y = -0.033x 2 + 0.164x + 2.01 (R 2 = 0.762, F = 15.102, p < 0.01)、样地2: y = -0.005x 2 + 0.233x +1.571 (R 2 = 0.894, F = 42.507, p < 0.01)、样地3: y = -0.041x 2 + 0.332x + 1.012 (R 2 = 0.724, F = 9.129, p < 0.01)、样地4: y = -0.03x 2 + 0.292x + 1.041 (R 2 = 0.851, F = 11.172, p < 0.01)、样地5: y = -0.151x 2 + 0.477x + 1.124 (R 2 = 0.587, F = 6.997, p < 0.05)。式中, y 为访花蝇类的数量, x 为温差。
图5 一天中花内温度与环境温度的温差变化(平均值±标准误差)。
Fig. 5 Changes in temperature difference between flowers and the environment during a day (mean ± SE ).
3 讨论
3.1 花寿命与柱头可授性
花寿命具有可塑性, 环境的变化往往造成花寿命的延长或缩短(Clark & Husband, 2007 ; 张志强和李庆军, 2009 ; Torres-Díaz et al ., 2011 ), 而花寿命与植物繁殖成功率相关(Torres-Díaz et al ., 2011 )。Vesprini和Pacini (2005) 亦发现, 低温条件下, 铁筷子属植物Helleborus bocconei 和H . foetidus 采取长花寿命, 每天花粉囊以依次少量开裂的方式来适应传粉昆虫不足的情况。Steinacher和Wagner (2010) 对7种高山植物的研究发现, 当花朵被拜访的频率很低时, 植物会通过延长柱头可授性和增加花冠保鲜时间来延长花寿命。本研究发现, 海拔越高全缘叶绿绒蒿的花寿命与柱头可授性持续时间越长, 这可能与高海拔的低温条件以及由此造成的传粉者访花频率低(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 )有关。对于需要传粉者才能完成授粉的植物而言, 花寿命的延长不仅可以增加花被传粉者拜访的概率(张志强和李庆军, 2009 ), 也会增加自身花粉的移出和外来花粉落置的机会(彭德力等, 2012 ), 同时有利于获得更多的交配机会, 保证繁殖成功(Teixido et al ., 2011 )。可见, 全缘叶绿绒蒿花寿命的延长可能是其应对高山环境较低昆虫访花频率的一种“机会主义”有性生殖策略(侯勤正等, 2009 ), 是植物对高山环境有效适应的结果。
3.2 传粉昆虫
以往对高山植物传粉昆虫的研究发现, 熊蜂以其稳定的热调节系统、较强的活动能力和较高的传粉效率, 使其成为高山环境下最重要的传粉者(Bergman et al ., 1996 ; Blionis & Vokou, 2001 ; 侯勤正等, 2009 ; Rader et al ., 2013 )。但在高山植物的传粉过程中, 蝇类往往也扮演着重要的角色。发现厕蝇(Fannia sp.)是茄科植物山莨菪(Anisodus tanguticus )最重要的有效异花传粉昆虫; Pickering和Stock (2004) 在对澳大利亚高山群落中传粉昆虫的调查也发现, 蝇类比重高达62.3%, 且种类丰富、访花频繁; Sieber等(2011) 对欧洲高山植物紫草科齿缘草属植物Eritrichium nanum 的传粉昆虫调查发现, 蝇类比重达到97.5%。本研究亦发现, 尽管熊蜂在高山环境中有分布, 但其更倾向于拜访分泌花蜜的报春类植物, 而蝇类才是全缘叶绿绒蒿最重要的异花传粉昆虫。这可能与巴郎山环境中牲畜(牦牛、马)的粪便较多, 为蝇类提供了食物来源有关; 此外, 全缘叶绿绒蒿花冠的保温效果能够保障在环境条件恶劣时(如低温、降雨等)为蝇类等传粉昆虫提供庇护场所。观察中没有发现全缘叶绿绒蒿分泌花蜜, 亦没有发现蝇类取食花粉, 而蝇类数量与花内温度、环境温度的温差间的极显著相关关系(p < 0.01)表明全缘叶绿绒蒿可以通过向蝇类等提供保温庇护场所达到吸引昆虫为之传粉的目的。与生活在喜马拉雅山和横断山冰缘带的“温室植物” (greenhouse plant)和“棉毛植物”(cotton plant)以较大苞片调节花内温度不同, 全缘叶绿绒蒿的花瓣在调节花内温度的过程中起着重要作用。全缘叶绿绒蒿随环境温度改变而调节自身花内温度的机制是否对雌雄配子体的发育产生影响有待于进一步证实。
有研究表明, 海拔较高或积温较低的地区, 某些昆虫会受到温度的限制(Heinrich, 1972 ; Totland, 2001 ; Fabbro & Körner, 2004 ), 导致不同海拔地区访问者和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b ; Sieber et al ., 2011 )。全缘叶绿绒蒿最高海拔样地1 (4452 m)蝇类的访花频率最低((0.15 ± 0.09)次·花-1 ·h-1 ), 这可能与高海拔地区较低的温度有关; 但最高的蝇类访花频率((0.59 ± 0.04)次·花-1 ·h-1 )没有出现在较低海拔的样地中, 而是出现在样地2 (4215 m), 野外观察研究发现, 样地2中全缘叶绿绒蒿的个体数量最多、开花最为繁盛, 由此初步推测, 样地2中最高的蝇类访花频率可能与其较高的花展示度有关, 同时, 观察时间和研究样地缺乏重复试验的限制亦可能造成访花频率在各海拔间的差异。
3.3 繁育系统
一般而言, 自交不亲和、雌雄异位和雌雄异熟被认为是促进异交、避免自交的重要机制(Ishii & Sakai, 2002 ; Anderson et al ., 2003 ; 侯勤正等, 2009 )。在异花传粉者不足等选择压力下, 一些物种能够演化出自交亲和或依赖自交的种群和个体(Busch, 2005 ; Herlihy & Eckert, 2005 ; 彭德力等, 2012 )。全缘叶绿绒蒿具有不完全雌雄异位和不完全雌雄异熟的特点, 属于自交亲和物种(self-compatible species)。花部综合特征观察及人工授粉试验的研究表明, 生长于高山环境中的全缘叶绿绒蒿具有花药与柱头间的距离缩小、雌雄性功能展示时间的重叠等特征的变化, 且面临着传粉者不足造成的传粉限制。全缘叶绿绒蒿自交亲和及雌雄不完全异熟为单花的自动自交提供了前提条件, 同时不完全雌雄异位直接导致了单花自动自交现象的发生。具体而言, 在研究的5个样地中, 均发现花药于开花的第5天开始散粉, 此时柱头还具有较强的可授性, 这为自交提供了前提; 其次, 大多数观察个体(约65%)的花药和柱头在开花发育的中后期完全接触, 这为自动自交提供了花粉来源; 再者, 全缘叶绿绒蒿个体自交部分亲和, 这为自动自交奠定了生理基础。同时, 花药在花开放过程中始终紧贴花柱, 开花中后期花药与柱头的接触, 可能会由于昆虫在花内的活动而引起协助自交。各样地的结实均由于异花传粉者不足而受到传粉限制。两种不同类型的自交机制恰恰为该植物异花传粉者不足提供了一定程度上的繁殖补偿(Zhang & Li, 2008 )。
人工授粉试验表明, 海拔较高的样地1和样地2异花花粉可能是全缘叶绿绒蒿产生种子的主要花粉来源, 这与样地2最高的蝇类访花频率相对应, 而样地1异花花粉对结实率的贡献大可能与异花传粉昆虫传粉的有效性有关。而海拔稍低的样地3、4和样地5蝇类访花频率低于样地2, 同时, 样地3、4和5中除由蝇类引起的“协助自交”外, 蓟马等小型昆虫亦导致自身花粉在同一花内的传递, 这些因素共同作用可能造成样地3、4和5的花获得更多的自花花粉而产生更多的种子。
全缘叶绿绒蒿P/O值为901.24 ± 43.01, 这与Dafni (1992) 指出的兼性异交的物种相似, 这也说明全缘叶绿绒蒿同时具有异交和自交的混合交配系统, 正如曾经报道过的菊科植物Chaetanthera euphrasioides (Arroyo et al ., 2006 )、茄科植物山莨菪(段元文等, 2007b )、龙胆科植物线叶龙胆(Gentiana lawrencei var. farreri )(侯勤正等, 2009 )、茄参(Mand- ragora caulescens ) (万金鹏等, 2011 )、锦葵科植物Kosteletzkya virginica (Ruan et al ., 2011 )以及With- ania ashwagandha (Mir et al ., 2013 )等植物一样, 导致植物异交的隔离机制处于非完全状态, 使得全缘叶绿绒蒿能够通过自交方式产生种子以保证结实。
致谢 感谢四川农业大学风景园林学院张倩、廖捷敏、侯舒婷、曾丽云、刘思岑、刘梦颖在实验室统计分析过程中给予的帮助, 感谢四川农业大学风景园林学院宋会兴教授对论文提出的宝贵意见和建议!
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Anderson B Midgley JJ Stewart BA 2003 ). Facilitated selfing offers reproductive assurance: A mutualism between a hemipteran and carnivorous plant
. American Journal of Botany , 90 , 1009 -1015 .
[本文引用: 1]
[2]
Arroyo MTK Armesto JJ Primack RB 1985 ). Community studies in pollination ecology in the high temperate Andes of central Chile. II. Effect of temperature on visitation rates and pollination possibilities
. Plant Systematics and Evolution , 149 , 187 -203 .
[本文引用: 2]
[3]
Arroyo MTK Muñoz MS Henríquez C Till-Bottraud I Pérez F 2006 ). Erratic pollination, high selfing levels and their correlates and consequences in an attitudinally widespread above-tree-line species in the high Andes of Chile
. Acta Oecologica , 30 , 248 -257 .
[本文引用: 1]
[4]
Arroyo MTK Primack R Armesto J 1982 ). Community studies in pollination ecology in the high temperate Andes of central Chile. I. Pollination mechanisms and altitudinal variation
. American Journal of Botany , 69 , 82 -97 .
[本文引用: 1]
[5]
Bergman P Molar U Holmgren B 1996 ). Micrometeorological impacts on insect activity and plant reproductive success in an alpine environment, Swedish Lapland
. Arctic and Alpine Research , 28 , 196 -202 .
[本文引用: 1]
[6]
Bingham RA Orthner AR 1998 ). Efficient pollination of alpine plants
. Nature , 391 , 238 -239 .
[本文引用: 2]
[7]
Blionis GJ Vokou D 2001 ). Pollination ecology of Campanula species on Mt Olympos, Greece
. Ecography , 24 , 287 -297 .
[本文引用: 1]
[8]
Blionis GJ Vokou D 2002 ). Structural and functional divergence of Campanula spatulata subspecies on Mt Olympos (Greece)
. Plant Systematics and Evolution , 232 , 89 -105 .
[本文引用: 3]
[9]
Busch JW 2005 ). The evolution of self-compatibility in geographically peripheral populations of Leavenworthia alabamica (Brassicaceae)
. American Journal of Botany , 92 , 1503 -1512 .
[本文引用: 1]
[10]
Chuang H 1981 ). The systematic evolution and the geographical distribution of Meconopsis vig
. Acta Botanica Yunnanica , 3 , 139 -146 .
[本文引用: 1]
(in Chinese with English abstract) [庄璇 (1981 ). 绿绒蒿属的系统演化及地理分布
. 云南植物研究 ,3 , 139 -146 .]
[本文引用: 1]
[11]
Clark MJ Husband BC 2007 ). Plasticity and timing of flower closure in response to pollination in Chamerion angustifolium (Onagraceae)
. International Journal of Plant Science , 168 , 619 -625 .
[本文引用: 1]
[12]
Dafni A 1992). Pollination Ecology : A Practical Approach. Oxford University Press , Oxford . 1 -57 .
[本文引用: 1]
[13]
Dar AR Zafar R Dar GH 2010 ). Reproductive ecology of an endemic angiosperm, Meconopsis latifolia Prain (Papaveraceae), in the Kashmir Himalaya, India
. Journal of Biological Science , 10 , 490 -498 .
[本文引用: 1]
[14]
Devoto MD Medan D Montaldo NH 2005 ). Patterns of interaction between plants and pollinators along an environmental gradient
. Oikos , 109 , 461 -472 .
[本文引用: 2]
[15]
Duan YW He YP Zhang TF Liu JQ 2007 a). Delayed selfing in an alpine species Gentianopsis barbata. Journal of Plant Ecology (Chinese Version)
, 31 , 110 -117 .
[本文引用: 1]
(in Chinese with English abstract) [段元文 , 何亚平 , 张挺峰 , 刘建全 (2007 a). 高山植物扁蕾的延迟自交机制
. 植物生态学报 , 31 , 110 -117 .]
[本文引用: 1]
[16]
Duan YW Zhang TF Liu JQ 2007 ). Interannual fluctuations in floral longevity, pollinator visitation and pollination limitation of an alpine plant (Gentiana straminea Maxim., Gentianaceae) at two altitudes in the Qinghai-Tibetan Plateau
. Plant Systematics and Evolution , 267 , 255 -265 .
[本文引用: 3]
[17]
Duan YW Zhang TF Liu JQ 2007 b). Pollination biology of Anisodus tanguticus (Solanaceae)
. Biodiversity Science , 15 , 584 -591 .
[本文引用: 5]
(in Chinese with English abstract) [段元文 , 张挺峰 , 刘建全 (2007 b). 山莨菪(茄科)的传粉生物学
. 生物多样性 , 15 , 584 -591 .]
[本文引用: 5]
[18]
Fabbro T Körner C 2004 ). Altitudinal differences in flower traits and reproductive allocation
. Flora , 199 , 70 -81 .
[本文引用: 2]
[19]
Fishman MA Hadany L 2013 ). Pollinators’ mating rendezvous and the evolution of floral advertisement
. Journal of Theoretical Biology , 316 , 99 -106 .
[本文引用: 1]
[20]
He HX 2006 ). Pollination Biology and Breeding System of the Endangered Chinese Endemic Kingdonia uniflora (Ranunculaceae). Master degree dissertation, Shaanxi Normal University
, Xi’an. 12 .
[本文引用: 1]
(in Chinese with English abstract). [贺海霞 (2006 ). 濒危植物独叶草(毛茛科)的传粉生物学及繁育系统研究
. 硕士论文, 陕西师范大学, 西安 . 12 .]
[本文引用: 1]
[21]
He YP Liu JQ 2004 ). Pollination ecology of Gentiana straminea Maxim. (Gentianaceae), an alpine perennial in the Qinghai-Tibet Plateau
. Acta Ecologica Sinica , 24 , 215 -220 .
[本文引用: 1]
(in Chinese with English abstract) [何亚平 , 刘建全 (2004 ). 青藏高原高山植物麻花艽的传粉生态学研究
. 生态学报 , 24 , 215 -220 .]
[本文引用: 1]
[22]
Heinrich B 1972 ). Energetics of temperature regulation and foraging in a bumblebee, Bombus terricola Kirby
. Journal of Comparative Physiology , 77 , 49 -64 .
[本文引用: 1]
[23]
Herlihy CR Eckert CG 2005 ). Evolution of self-fertilization at geographical range margins? A comparison of demographic, floral, and mating system variables in central vs. peripheral populations of Aquilegia canadensis (Ranunculaceae)
. American Journal of Botany , 92 , 744 -751 .
[本文引用: 1]
[24]
Hou QZ Duan YW Si QW Yang HL 2009 ). Pollination ecology of Gentiana lawrencei var. Farreri, a late-flowering Qinghai-Tibet Plateau species
. Chinese Journal of Plant Ecology , 33 , 1156 -1164 .
[本文引用: 5]
(in Chinese with English abstract) [侯勤正 , 段元文 , 司庆文 , 杨慧玲 (2009 ). 青藏高原晚期开花植物线叶龙胆的传粉生态学
. 植物生态学报 , 33 , 1156 -1164 .]
[本文引用: 5]
[25]
Ishii HS Sakai S 2002 ). Temporal variation in floral display size and individual floral sex allocation in racemes of Narthecium asiaticum (Liliaceae)
. American Journal of Botany , 89 , 441 -446 .
[本文引用: 1]
[26]
Makrodimos N Blionis GJ Krigas N Vokou D 2008 ). Flower morphology, phenology and visitor patterns in an alpine community on Mt Olympos, Greece
. Flora—Morphology, Distribution, Functional Ecology of Plants , 203 , 449 -468 .
[本文引用: 2]
[27]
Mao ZB Boehler C Ge XJ 2011 ). Pollination ecology and breeding system of Impatiens lateristachys (Balsaminaceae) endemic to China
. Guihaia , 31 , 160 -166 .
[本文引用: 1]
(in Chinese with abstract) [毛志斌 , Boehler C 葛学军 (2011 ). 侧穗凤仙花的传粉生态和繁育系统
. 广西植物 , 31 , 160 -166 .]
[本文引用: 1]
[28]
Meng LH Wang Y Luo J Liu CY Yang YP Duan YW 2012 ). Pollination ecology and its implication for conservation of an endangered perennial herb native to the East-Himalaya, Megacodon stylophorus (Gentianaceae)
. Plant Ecology and Evolution , 145 , 356 -362 .
[本文引用: 1]
[29]
Meng Y Xie HY Nie ZL Gu ZJ Yang YP 2006 ). A karyomorphological study on four species of Meconopsis Vig. (Papaveraceae) from the Hengduan Mountains, SW China
. Garyologia , 59 , 1 -6 .
[本文引用: 1]
[30]
Mir BA Koul S Soodan AS 2013 ). Reproductive biology of Withania ashwagandha sp. Novo (Solanaceae)
. Industrial Crops and Products , 45 , 442 -446 .
[本文引用: 1]
[31]
Pan CC Liu LD Zhao HL Hou YL Zhang L Wang LJ 2012 ). Pollination biology of Elaeagnus angustifolia in the middle reaches of the Heihe River in Northwest China
. Journal of Desert Research , 32 , 780 -783 .
[本文引用: 1]
(in Chinese with English abstract) [潘成臣 , 刘林德 , 赵哈林 , 侯月利 , 张莉 , 王丽娟 (2012 ). 黑河中游沙枣的传粉生物学研究
. 中国沙漠 , 32 , 780 -783 .]
[本文引用: 1]
[32]
Peng DL Zhang ZQ Niu Y Yang Y Song B Sun H Li ZM 2012 ). Advances in the studies of reproductive strategies of alpine plants
. Biodiversity Science , 20 , 286 -299 .
[本文引用: 3]
(in Chinese with English abstract) [彭德力 , 张志强 , 牛洋 , 杨扬 , 宋波 , 孙航 , 李志敏 (2012 ). 高山植物繁殖策略的研究进展
. 生物多样性 , 20 , 286 -290 .]
[本文引用: 3]
[33]
Pickering CM Stock M 2004 ). Insect colour preference compared to flower colours in the Australian Alps
. Nordic Journal of Botany , 23 , 217 -223 .
[本文引用: 1]
[34]
Rader R Edwards W Westcott DA Cunningham SA Howlett BG 2013 ). Diurnal effectiveness of pollination by bees and flies in agricultural Brassica rapa: Implications for ecosystem resilience
. Basic and Applied Ecology , 14 , 20 -27 .
[本文引用: 1]
[35]
Ruan CJ Qin P Teixeira da Silva JA 2011 ). Relationship between reproductive assurance and mixed mating in perennial Kosteletzkya virginica
. South African Journal of Botany , 77 , 280 -291 .
[本文引用: 1]
[36]
Schoen DJ Morgan MT Bataillon T 1996 ). How does self-pollination evolve? Inferences from floral ecology and molecular genetic variation. Philosophical Transactions of the Royal Society of London (
Series B ), 351 , 1281 -1290 .
[本文引用: 1]
[37]
Shang XY Li C Zhang CZ Yang YC Shi JG 2006 ). Non-alkaloid constituents from a Tibetan medicine Meconopsis quintuplinervia
. China Journal of Chinese Materia Medica , 31 , 468 -471 .
[本文引用: 1]
(in Chinese with English abstract) [尚小雅 , 李冲 , 张承忠 , 杨永春 , 石建功 (2006 ). 藏药五脉绿绒蒿中非生物碱成分
. 中国中药杂志 , 31 , 468 -471 .]
[本文引用: 1]
[38]
Shang XY Wang YH Li C Zhang CZ Yang YC Shi JG 2006 ). Acetylated flavonol diglucosides from Meconopsis quintuplinervia
. Phytochemistry , 67 , 511 -515 .
[本文引用: 1]
[39]
Shao JW Zhang XP Zhang ZX Zhu GP 2008 ). Identification of effective pollinators of Primula merrilliana and effects of flower density and population size on pollination efficiency
. Journal of Systematics and Evolution , 46 , 537 -544 .
[本文引用: 1]
(in Chinese with English abstract) [邵剑文 , 张小平 , 张中信 , 朱国萍 (2008 ). 安徽羽叶报春的有效传粉昆虫及花朵密度和种群大小对传粉效果的影响
. 植物分类学报 , 46 , 537 -544 .]
[本文引用: 1]
[40]
Si QW Hou QZ Zhu XF Zhou DW Yang HL 2010 ). Pollination biology of Gentiana dahurica (Gentianaceae)
. Acta Botanica Boreali-Occidentalia Sinica , 30 , 2433 -2436 .
[本文引用: 1]
(in Chinese with English abstract) [司庆文 , 侯勤正 , 朱兴福 , 周党卫 , 杨慧玲 (2010 ). 达乌里秦艽的传粉生物学研究
. 西北植物学报 , 30 , 2433 -2436 .]
[本文引用: 1]
[41]
Sieber Y Holderegger R Waser NM Thomas VFD Braun S Erhardt A Reyer HU Wirth LR 2011 ). Do alpine plants facilitate each other’s pollination? Experiments at a small spatial scale
. Acta Oecologica , 37 , 369 -374 .
[本文引用: 2]
[42]
Singhal VK Kumar P 2008 ). Impact of cytomixis on meiosis, pollen viability and pollen size in wild populations of Himalayan poppy (Meconopsis aculeate Royle)
. Journal of Bioscience , 33 , 371 -380 .
[本文引用: 1]
[43]
Stanton ML Galen C 1989 ). Consequences of flower heliotropism for reproduction in an alpine buttercup (Ranunculus adoneus)
. Oecologia , 78 , 477 -485 .
[本文引用: 1]
[44]
Steinacher G Wagner J 2010 ). Flower longevity and duration of pistil receptivity in high mountain plants
. Flora— Morphology, Distribution, Functional Ecology of Plants , 205 , 376 -387 .
[本文引用: 2]
[45]
Sulaiman IM Babu CR 1996 ). Enzyme polymorphism analyses in three endangered species of Himalayan poppy, Meconopsis (Papaveraceae)
. Genetic Resources and Crop Evolution , 43 , 351 -356
[本文引用: 1]
[46]
Sun S Cao GX Luo YJ Li QJ 2010 ). Maintenance and functional gender specialization of flexistyly
. Chinese Journal of Plant Ecology , 34 , 827 -838 .
[本文引用: 1]
(in Chinese with English abstract) [孙杉 , 操国兴 , 罗燕江 , 李庆军 (2010 ). 花柱卷曲性的维持及功能性别特化
. 植物生态学报 , 34 , 827 -838 .]
[本文引用: 1]
[47]
Teixido AL Méndez M Valladares F 2011 ). Flower size and longevity influence florivory in the large-flowered shrub Cistus ladanifer
. Acta Oecologica , 37 , 418 -421 .
[本文引用: 1]
[48]
Tian JP Liu KM Hu GW 2004 ). Pollination ecology and pollination system of Impatiens reptans (Balsaminaceae) endemic to China
. Annals of Botany , 93 , 167 -175 .
[本文引用: 1]
[49]
Torres-Díaz C Gómez-González S Stotz GC Torres-Morales P Paredes B Pérez-Millaqueo M Gianoli E 2011 ). Extremely long-lived stigmas allow extended cross- pollination opportunities in a high Andean plant
. PLoS ONE , 6 , e19497 .
[本文引用: 2]
[50]
Totland Ø 2001 ). Environment-dependent pollen limitation and selection on floral traits in an alpine species
. Ecology , 82 , 2233 -2244 .
[本文引用: 2]
[51]
Tsukaya H 2002 ). Optical and anatomical characteristics of bracts from the Chinese “glasshouse” plant, Rheum alexandrae Batalin (Ploygonaceae), in Yunnan, China
. Journal of Plant Research , 115 , 59 -63 .
[本文引用: 1]
[52]
Vesprini JL Pacini E 2005 ). Temperature-dependent floral longevity in two Helleborus species
. Plant Systematics and Evolution , 252 , 63 -70 .
[本文引用: 1]
[53]
Vesprini JL Pacini E 2010 ). Pollination ecology in sympatric winter flowering Helleborus (Ranunculaceae)
. Flora— Morphology, Distribution, Functional Ecology of Plants , 205 , 627 -632 .
[本文引用: 1]
[54]
Wan JP Zhu XF Li QJ 2011 ). Breeding system of protogynous Mandragora caulescens (Solanaceae)
. Plant Diversity and Resources , 33 , 565 -570 .
[本文引用: 1]
(in Chinese with English abstract) [万金鹏 , 朱兴福 , 李庆军 (2011 ). 雌先熟植物茄参(茄科)的繁育系统
. 植物分类与资源学报 , 33 , 565 -570 .]
[本文引用: 1]
[55]
Xiao LX 2009 ). The Pollination Biology of Four Species of Impatiens L. Master degree dissertation, Hunan Normal University
, Changsha. 4 .
[本文引用: 1]
(in Chinese with English abstract). [肖乐希 (2009 ). 四种凤仙花属(Impatiens L.) 植物的传粉生物学研究
. 硕士论文, 湖南师范大学, 长沙 . 4 .]
[本文引用: 1]
[56]
Xiao LX Liu KM 2009 ). Floral traits and pollination system of Impatiens chinensis (Balsaminaceae)
. Bulletin of Botanical Research , 29 , 164 -168 .
[本文引用: 1]
(in Chinese with English abstract) [肖乐希 , 刘克明 (2009 ). 华凤仙花部特征和传粉系统研究
. 植物研究 , 29 , 164 -168 .]
[本文引用: 1]
[57]
Yang FS Qin AL Li YF Wang XQ 2012 ). Great genetic differentiation among populations of Meconopsis integrifolia and its implication for plant speciation in the Qinghai- Tibetan Plateau
. PLoS ONE , 7 , e37196 .
[本文引用: 1]
[58]
Ye YM Zhang JW Qi YC Chen TH Bao MZ 2007 ). Studies on stigma receptivity and pollen viability of Zinnia elegans
. Scientia Agricultura Sincia , 40 , 2376 -2381 .
[本文引用: 1]
(in Chinese with English abstract) [叶要妹 , 张俊卫 , 齐迎春 , 陈天花 , 包满珠 (2007 ). 百日草柱头可授性和花粉生活力的研究
. 中国农业科学 , 40 , 2376 -2381 .]
[本文引用: 1]
[59]
Zhang DY Jiang XH 2001 ). Mating system evolution, resource allocation, and genetic diversity in plants
. Acta Phytoecologica Sinica , 25 , 130 -143 .
[本文引用: 1]
(in Chinese with English abstract) [张大勇 , 姜新华 (2001 ). 植物交配系统的进化、资源分配对策与遗传多样性
. 植物生态学报 , 25 , 130 -143 .]
[本文引用: 1]
[60]
Zhang TF Duan YW Liu JQ 2006 ). Pollination ecology of Aconitum gymnandrum (Ranunculaceae) at two sites with different altitudes
. Acta Phytotaxonomica Sinica , 44 , 362 -370 .
[本文引用: 1]
(in Chinese with English abstract) [张挺峰 , 段元文 , 刘建全 (2006 ). 露蕊乌头(毛茛科)不同海拔居群的传粉生态学
. 植物分类学报 , 44 , 362 -370 .]
[本文引用: 1]
[61]
Zhang ZQ Li QJ 2008 ). Autonomous selfing provides reproductive assurance in an alpine ginger Roscoea schneideriana (Zingiberaceae)
. Annals of Botany , 102 , 531 -538 .
[本文引用: 1]
[62]
Zhang ZQ Li QJ 2009 ). Review of evolutionary ecology of floral longevity
. Chinese Journal of Plant Ecology , 33 , 598 -606 .
[本文引用: 2]
(in Chinese with English abstract) [张志强 , 李庆军 (2009 ). 花寿命的进化生态学意义
. 植物生态学报 , 33 , 598 -606 .]
[本文引用: 2]
[63]
Zhong YF Zhang Z Song XQ Zhou ZD 2014 ). Pollination biology of Impatiens hainanensis (Balsaminaceae) populations at different altitudes
. Biodiversity Science , 22 , 467 -475 .
[本文引用: 2]
(in Chinese with English abstract) [钟云芳 , 张哲 , 宋希强 , 周兆德 (2014 ). 海南凤仙花不同海拔种群的传粉生物学
. 生物多样性 , 22 , 467 -475 .]
[本文引用: 2]
[64]
Zhou Y Song JZ Choi FFK Wu HF Qiao CF Ding LS Gesang SL Xu HX 2009 ). An experimental design approach using response surface techniques to obtain optimal liquid chromatography and mass spectrometry conditions to determine the alkaloids in Meconops species
. Journal of Chromatography A , 1216 , 7013 -7023 .
[本文引用: 1]
[65]
Zhu JH Ou SJ Mai FZ He QG Xu N Peng HX 2006 ). Effect of flies on mango pollination and the relationship between their pollination activity and the temperature
. Chinese Journal of Tropical Crops , 27 (4 ), 5 -8 .
[本文引用: 1]
(in Chinese with English abstract) [朱建华 , 欧世金 , 麦福珍 , 何全光 , 徐宁 , 彭宏祥 (2006 ). 苍蝇对杧果的传粉作用及其与温度的关系
. 热带作物学报 , 27 (4 , 5 -8 .]
[本文引用: 1]
[66]
Zhu JH Xu N Wang ZY Li HL Lu GF Li DB Li GW Huang FZ Peng HX 2010 ). Study on the species of pollination insects of longan and the relationship between their pollination activity and temperature
. Chinese Journal of Tropical Crops , 31 , 646 -650 .
[本文引用: 1]
(in Chinese with English abstract) [朱建华 , 徐宁 , 王助引 , 李鸿莉 , 陆贵锋 , 李冬波 , 黎光旺 , 黄凤珠 , 彭宏祥 (2010 ). 龙眼传粉昆虫种类及其传粉活动与温度的关系
. 热带作物学报 , 31 , 646 -650 .]
[本文引用: 1]
Facilitated selfing offers reproductive assurance: A mutualism between a hemipteran and carnivorous plant
1
2003
... 一般而言, 自交不亲和、雌雄异位和雌雄异熟被认为是促进异交、避免自交的重要机制(Ishii & Sakai, 2002 ; Anderson et al ., 2003 ; 侯勤正等, 2009 ).在异花传粉者不足等选择压力下, 一些物种能够演化出自交亲和或依赖自交的种群和个体(Busch, 2005 ; Herlihy & Eckert, 2005 ; 彭德力等, 2012 ).全缘叶绿绒蒿具有不完全雌雄异位和不完全雌雄异熟的特点, 属于自交亲和物种(self-compatible species).花部综合特征观察及人工授粉试验的研究表明, 生长于高山环境中的全缘叶绿绒蒿具有花药与柱头间的距离缩小、雌雄性功能展示时间的重叠等特征的变化, 且面临着传粉者不足造成的传粉限制.全缘叶绿绒蒿自交亲和及雌雄不完全异熟为单花的自动自交提供了前提条件, 同时不完全雌雄异位直接导致了单花自动自交现象的发生.具体而言, 在研究的5个样地中, 均发现花药于开花的第5天开始散粉, 此时柱头还具有较强的可授性, 这为自交提供了前提; 其次, 大多数观察个体(约65%)的花药和柱头在开花发育的中后期完全接触, 这为自动自交提供了花粉来源; 再者, 全缘叶绿绒蒿个体自交部分亲和, 这为自动自交奠定了生理基础.同时, 花药在花开放过程中始终紧贴花柱, 开花中后期花药与柱头的接触, 可能会由于昆虫在花内的活动而引起协助自交.各样地的结实均由于异花传粉者不足而受到传粉限制.两种不同类型的自交机制恰恰为该植物异花传粉者不足提供了一定程度上的繁殖补偿(Zhang & Li, 2008 ). ...
Community studies in pollination ecology in the high temperate Andes of central Chile. II. Effect of temperature on visitation rates and pollination possibilities
2
1985
... 高山恶劣的自然环境条件如低温、大风、多雨以及生长季短等不利因素限制了传粉昆虫的种类及其活动能力和访花频率(Arroyo et al ., 1982 , 1985 ; Bingham & Orthner, 1998 ), 导致到达柱头的异花花粉数量减少, 继而影响植物的生殖成功率(张大勇和姜新华, 2001 ).但已有的研究表明, 具有异花传粉繁育系统的植物在高山植物群落中普遍存在, 它们往往采取延长花寿命(Arroyo et al ., 1985 ; Bingham & Orthner, 1998 ; Blionis & Vokou, 2002 ; Steinacher & Wagner, 2010 )、增加对繁殖结构的资源分配(Fabbro & Körner, 2004 )等策略来适应传粉昆虫的缺乏, 以保证种群的延续.同时, 一些高山植物会选择自花授粉这种以降低后代适合度为代价的方式保证种群的延续(Schoen et al ., 1996 ; 段元文等, 2007b ). ...
... ).但已有的研究表明, 具有异花传粉繁育系统的植物在高山植物群落中普遍存在, 它们往往采取延长花寿命(Arroyo et al ., 1985 ; Bingham & Orthner, 1998 ; Blionis & Vokou, 2002 ; Steinacher & Wagner, 2010 )、增加对繁殖结构的资源分配(Fabbro & Körner, 2004 )等策略来适应传粉昆虫的缺乏, 以保证种群的延续.同时, 一些高山植物会选择自花授粉这种以降低后代适合度为代价的方式保证种群的延续(Schoen et al ., 1996 ; 段元文等, 2007b ). ...
Erratic pollination, high selfing levels and their correlates and consequences in an attitudinally widespread above-tree-line species in the high Andes of Chile
1
2006
... 全缘叶绿绒蒿P/O值为901.24 ± 43.01, 这与Dafni (1992) 指出的兼性异交的物种相似, 这也说明全缘叶绿绒蒿同时具有异交和自交的混合交配系统, 正如曾经报道过的菊科植物Chaetanthera euphrasioides (Arroyo et al ., 2006 )、茄科植物山莨菪(段元文等, 2007b )、龙胆科植物线叶龙胆(Gentiana lawrencei var. farreri )(侯勤正等, 2009 )、茄参(Mand- ragora caulescens ) (万金鹏等, 2011 )、锦葵科植物Kosteletzkya virginica (Ruan et al ., 2011 )以及With- ania ashwagandha (Mir et al ., 2013 )等植物一样, 导致植物异交的隔离机制处于非完全状态, 使得全缘叶绿绒蒿能够通过自交方式产生种子以保证结实. ...
Community studies in pollination ecology in the high temperate Andes of central Chile. I. Pollination mechanisms and altitudinal variation
1
1982
... 高山恶劣的自然环境条件如低温、大风、多雨以及生长季短等不利因素限制了传粉昆虫的种类及其活动能力和访花频率(Arroyo et al ., 1982 , 1985 ; Bingham & Orthner, 1998 ), 导致到达柱头的异花花粉数量减少, 继而影响植物的生殖成功率(张大勇和姜新华, 2001 ).但已有的研究表明, 具有异花传粉繁育系统的植物在高山植物群落中普遍存在, 它们往往采取延长花寿命(Arroyo et al ., 1985 ; Bingham & Orthner, 1998 ; Blionis & Vokou, 2002 ; Steinacher & Wagner, 2010 )、增加对繁殖结构的资源分配(Fabbro & Körner, 2004 )等策略来适应传粉昆虫的缺乏, 以保证种群的延续.同时, 一些高山植物会选择自花授粉这种以降低后代适合度为代价的方式保证种群的延续(Schoen et al ., 1996 ; 段元文等, 2007b ). ...
Micrometeorological impacts on insect activity and plant reproductive success in an alpine environment, Swedish Lapland
1
1996
... 以往对高山植物传粉昆虫的研究发现, 熊蜂以其稳定的热调节系统、较强的活动能力和较高的传粉效率, 使其成为高山环境下最重要的传粉者(Bergman et al ., 1996 ; Blionis & Vokou, 2001 ; 侯勤正等, 2009 ; Rader et al ., 2013 ).但在高山植物的传粉过程中, 蝇类往往也扮演着重要的角色.发现厕蝇(Fannia sp.)是茄科植物山莨菪(Anisodus tanguticus )最重要的有效异花传粉昆虫; Pickering和Stock (2004) 在对澳大利亚高山群落中传粉昆虫的调查也发现, 蝇类比重高达62.3%, 且种类丰富、访花频繁; Sieber等(2011) 对欧洲高山植物紫草科齿缘草属植物Eritrichium nanum 的传粉昆虫调查发现, 蝇类比重达到97.5%.本研究亦发现, 尽管熊蜂在高山环境中有分布, 但其更倾向于拜访分泌花蜜的报春类植物, 而蝇类才是全缘叶绿绒蒿最重要的异花传粉昆虫.这可能与巴郎山环境中牲畜(牦牛、马)的粪便较多, 为蝇类提供了食物来源有关; 此外, 全缘叶绿绒蒿花冠的保温效果能够保障在环境条件恶劣时(如低温、降雨等)为蝇类等传粉昆虫提供庇护场所.观察中没有发现全缘叶绿绒蒿分泌花蜜, 亦没有发现蝇类取食花粉, 而蝇类数量与花内温度、环境温度的温差间的极显著相关关系(p < 0.01)表明全缘叶绿绒蒿可以通过向蝇类等提供保温庇护场所达到吸引昆虫为之传粉的目的.与生活在喜马拉雅山和横断山冰缘带的“温室植物” (greenhouse plant)和“棉毛植物”(cotton plant)以较大苞片调节花内温度不同, 全缘叶绿绒蒿的花瓣在调节花内温度的过程中起着重要作用.全缘叶绿绒蒿随环境温度改变而调节自身花内温度的机制是否对雌雄配子体的发育产生影响有待于进一步证实. ...
Efficient pollination of alpine plants
2
1998
... 高山恶劣的自然环境条件如低温、大风、多雨以及生长季短等不利因素限制了传粉昆虫的种类及其活动能力和访花频率(Arroyo et al ., 1982 , 1985 ; Bingham & Orthner, 1998 ), 导致到达柱头的异花花粉数量减少, 继而影响植物的生殖成功率(张大勇和姜新华, 2001 ).但已有的研究表明, 具有异花传粉繁育系统的植物在高山植物群落中普遍存在, 它们往往采取延长花寿命(Arroyo et al ., 1985 ; Bingham & Orthner, 1998 ; Blionis & Vokou, 2002 ; Steinacher & Wagner, 2010 )、增加对繁殖结构的资源分配(Fabbro & Körner, 2004 )等策略来适应传粉昆虫的缺乏, 以保证种群的延续.同时, 一些高山植物会选择自花授粉这种以降低后代适合度为代价的方式保证种群的延续(Schoen et al ., 1996 ; 段元文等, 2007b ). ...
... ; Bingham & Orthner, 1998 ; Blionis & Vokou, 2002 ; Steinacher & Wagner, 2010 )、增加对繁殖结构的资源分配(Fabbro & Körner, 2004 )等策略来适应传粉昆虫的缺乏, 以保证种群的延续.同时, 一些高山植物会选择自花授粉这种以降低后代适合度为代价的方式保证种群的延续(Schoen et al ., 1996 ; 段元文等, 2007b ). ...
Pollination ecology of Campanula species on Mt Olympos, Greece
1
2001
... 以往对高山植物传粉昆虫的研究发现, 熊蜂以其稳定的热调节系统、较强的活动能力和较高的传粉效率, 使其成为高山环境下最重要的传粉者(Bergman et al ., 1996 ; Blionis & Vokou, 2001 ; 侯勤正等, 2009 ; Rader et al ., 2013 ).但在高山植物的传粉过程中, 蝇类往往也扮演着重要的角色.发现厕蝇(Fannia sp.)是茄科植物山莨菪(Anisodus tanguticus )最重要的有效异花传粉昆虫; Pickering和Stock (2004) 在对澳大利亚高山群落中传粉昆虫的调查也发现, 蝇类比重高达62.3%, 且种类丰富、访花频繁; Sieber等(2011) 对欧洲高山植物紫草科齿缘草属植物Eritrichium nanum 的传粉昆虫调查发现, 蝇类比重达到97.5%.本研究亦发现, 尽管熊蜂在高山环境中有分布, 但其更倾向于拜访分泌花蜜的报春类植物, 而蝇类才是全缘叶绿绒蒿最重要的异花传粉昆虫.这可能与巴郎山环境中牲畜(牦牛、马)的粪便较多, 为蝇类提供了食物来源有关; 此外, 全缘叶绿绒蒿花冠的保温效果能够保障在环境条件恶劣时(如低温、降雨等)为蝇类等传粉昆虫提供庇护场所.观察中没有发现全缘叶绿绒蒿分泌花蜜, 亦没有发现蝇类取食花粉, 而蝇类数量与花内温度、环境温度的温差间的极显著相关关系(p < 0.01)表明全缘叶绿绒蒿可以通过向蝇类等提供保温庇护场所达到吸引昆虫为之传粉的目的.与生活在喜马拉雅山和横断山冰缘带的“温室植物” (greenhouse plant)和“棉毛植物”(cotton plant)以较大苞片调节花内温度不同, 全缘叶绿绒蒿的花瓣在调节花内温度的过程中起着重要作用.全缘叶绿绒蒿随环境温度改变而调节自身花内温度的机制是否对雌雄配子体的发育产生影响有待于进一步证实. ...
Structural and functional divergence of Campanula spatulata subspecies on Mt Olympos (Greece)
3
2002
... 高山恶劣的自然环境条件如低温、大风、多雨以及生长季短等不利因素限制了传粉昆虫的种类及其活动能力和访花频率(Arroyo et al ., 1982 , 1985 ; Bingham & Orthner, 1998 ), 导致到达柱头的异花花粉数量减少, 继而影响植物的生殖成功率(张大勇和姜新华, 2001 ).但已有的研究表明, 具有异花传粉繁育系统的植物在高山植物群落中普遍存在, 它们往往采取延长花寿命(Arroyo et al ., 1985 ; Bingham & Orthner, 1998 ; Blionis & Vokou, 2002 ; Steinacher & Wagner, 2010 )、增加对繁殖结构的资源分配(Fabbro & Körner, 2004 )等策略来适应传粉昆虫的缺乏, 以保证种群的延续.同时, 一些高山植物会选择自花授粉这种以降低后代适合度为代价的方式保证种群的延续(Schoen et al ., 1996 ; 段元文等, 2007b ). ...
... 高山环境下, 由于传粉者的不足使得大多数植物存在花粉限制(段元文等, 2007b ; 司庆文等, 2010 ; 钟云芳等, 2014 ).面对传粉者缺乏的选择压力, 植物会采取多样的策略提高对传粉昆虫的吸引力, 如提高花展示(邵剑文等, 2008 ; 彭德力等, 2012 )、提供生殖场所(Fishman & Hadany, 2013 )等, 随着海拔升高, 一些植物通过延长花寿命获得交配的机会(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 ); 而低温环境下, 一些植物则通过提高花序内或花内温度来达到吸引传粉昆虫的目的(Stanton & Galen, 1989 ).如水母雪兔子(Saussurea medusa )通过花序温度的升高使熊蜂在花序上的停留时间超过1 min, 这在一定程度上对传粉者起到了吸引作用(Tsukaya, 2002 ).对冬季开花的毛茛科铁筷子属植物Helleborus foetidus 和Helleborus bocconei 的研究亦发现, 花内温度的升高使传粉者更加活跃(Vesprini & Pacini, 2010 ).同时, 一些植物往往会进化出各异的自交机制, 如通过雌雄蕊位置的变化促进花粉传递以保障成功繁殖.总结已有的研究发现, 植物雌雄蕊位置关系的改变方式呈现出多样化的特征, 如花丝伸长(段元文等, 2007a , 2007b )、花柱伸长(侯勤正等, 2009 )、花柱卷曲(孙杉等, 2010 )等. ...
... 花寿命具有可塑性, 环境的变化往往造成花寿命的延长或缩短(Clark & Husband, 2007 ; 张志强和李庆军, 2009 ; Torres-Díaz et al ., 2011 ), 而花寿命与植物繁殖成功率相关(Torres-Díaz et al ., 2011 ).Vesprini和Pacini (2005) 亦发现, 低温条件下, 铁筷子属植物Helleborus bocconei 和H . foetidus 采取长花寿命, 每天花粉囊以依次少量开裂的方式来适应传粉昆虫不足的情况.Steinacher和Wagner (2010) 对7种高山植物的研究发现, 当花朵被拜访的频率很低时, 植物会通过延长柱头可授性和增加花冠保鲜时间来延长花寿命.本研究发现, 海拔越高全缘叶绿绒蒿的花寿命与柱头可授性持续时间越长, 这可能与高海拔的低温条件以及由此造成的传粉者访花频率低(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 )有关.对于需要传粉者才能完成授粉的植物而言, 花寿命的延长不仅可以增加花被传粉者拜访的概率(张志强和李庆军, 2009 ), 也会增加自身花粉的移出和外来花粉落置的机会(彭德力等, 2012 ), 同时有利于获得更多的交配机会, 保证繁殖成功(Teixido et al ., 2011 ).可见, 全缘叶绿绒蒿花寿命的延长可能是其应对高山环境较低昆虫访花频率的一种“机会主义”有性生殖策略(侯勤正等, 2009 ), 是植物对高山环境有效适应的结果. ...
The evolution of self-compatibility in geographically peripheral populations of Leavenworthia alabamica (Brassicaceae)
1
2005
... 一般而言, 自交不亲和、雌雄异位和雌雄异熟被认为是促进异交、避免自交的重要机制(Ishii & Sakai, 2002 ; Anderson et al ., 2003 ; 侯勤正等, 2009 ).在异花传粉者不足等选择压力下, 一些物种能够演化出自交亲和或依赖自交的种群和个体(Busch, 2005 ; Herlihy & Eckert, 2005 ; 彭德力等, 2012 ).全缘叶绿绒蒿具有不完全雌雄异位和不完全雌雄异熟的特点, 属于自交亲和物种(self-compatible species).花部综合特征观察及人工授粉试验的研究表明, 生长于高山环境中的全缘叶绿绒蒿具有花药与柱头间的距离缩小、雌雄性功能展示时间的重叠等特征的变化, 且面临着传粉者不足造成的传粉限制.全缘叶绿绒蒿自交亲和及雌雄不完全异熟为单花的自动自交提供了前提条件, 同时不完全雌雄异位直接导致了单花自动自交现象的发生.具体而言, 在研究的5个样地中, 均发现花药于开花的第5天开始散粉, 此时柱头还具有较强的可授性, 这为自交提供了前提; 其次, 大多数观察个体(约65%)的花药和柱头在开花发育的中后期完全接触, 这为自动自交提供了花粉来源; 再者, 全缘叶绿绒蒿个体自交部分亲和, 这为自动自交奠定了生理基础.同时, 花药在花开放过程中始终紧贴花柱, 开花中后期花药与柱头的接触, 可能会由于昆虫在花内的活动而引起协助自交.各样地的结实均由于异花传粉者不足而受到传粉限制.两种不同类型的自交机制恰恰为该植物异花传粉者不足提供了一定程度上的繁殖补偿(Zhang & Li, 2008 ). ...
绿绒蒿属的系统演化及地理分布
1
1981
... 绿绒蒿属(Meconopsis )植物绝大多数分布在以横断山脉-喜马拉雅为中心的高山环境中(庄璇, 1981 ), 作为传统藏药, 目前的研究主要集中在其化学成分及药理活性方面(尚小雅等, 2006 ; Shang et al ., 2006 ; Zhou et al ., 2009 ), 一些学者对本属植物进行了系统分类方面的探讨(Meng et al ., 2006 ; Singhal & Kumar, 2008 ; Yang et al ., 2012 ).作为一类长期适应高山环境的植物, 目前对绿绒蒿属植物的繁殖特征、适应进化等方面尚缺乏系统研究(Dar et al ., 2010 ). ...
绿绒蒿属的系统演化及地理分布
1
1981
... 绿绒蒿属(Meconopsis )植物绝大多数分布在以横断山脉-喜马拉雅为中心的高山环境中(庄璇, 1981 ), 作为传统藏药, 目前的研究主要集中在其化学成分及药理活性方面(尚小雅等, 2006 ; Shang et al ., 2006 ; Zhou et al ., 2009 ), 一些学者对本属植物进行了系统分类方面的探讨(Meng et al ., 2006 ; Singhal & Kumar, 2008 ; Yang et al ., 2012 ).作为一类长期适应高山环境的植物, 目前对绿绒蒿属植物的繁殖特征、适应进化等方面尚缺乏系统研究(Dar et al ., 2010 ). ...
Plasticity and timing of flower closure in response to pollination in Chamerion angustifolium (Onagraceae)
1
2007
... 花寿命具有可塑性, 环境的变化往往造成花寿命的延长或缩短(Clark & Husband, 2007 ; 张志强和李庆军, 2009 ; Torres-Díaz et al ., 2011 ), 而花寿命与植物繁殖成功率相关(Torres-Díaz et al ., 2011 ).Vesprini和Pacini (2005) 亦发现, 低温条件下, 铁筷子属植物Helleborus bocconei 和H . foetidus 采取长花寿命, 每天花粉囊以依次少量开裂的方式来适应传粉昆虫不足的情况.Steinacher和Wagner (2010) 对7种高山植物的研究发现, 当花朵被拜访的频率很低时, 植物会通过延长柱头可授性和增加花冠保鲜时间来延长花寿命.本研究发现, 海拔越高全缘叶绿绒蒿的花寿命与柱头可授性持续时间越长, 这可能与高海拔的低温条件以及由此造成的传粉者访花频率低(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 )有关.对于需要传粉者才能完成授粉的植物而言, 花寿命的延长不仅可以增加花被传粉者拜访的概率(张志强和李庆军, 2009 ), 也会增加自身花粉的移出和外来花粉落置的机会(彭德力等, 2012 ), 同时有利于获得更多的交配机会, 保证繁殖成功(Teixido et al ., 2011 ).可见, 全缘叶绿绒蒿花寿命的延长可能是其应对高山环境较低昆虫访花频率的一种“机会主义”有性生殖策略(侯勤正等, 2009 ), 是植物对高山环境有效适应的结果. ...
1
... 全缘叶绿绒蒿P/O值为901.24 ± 43.01, 这与Dafni (1992) 指出的兼性异交的物种相似, 这也说明全缘叶绿绒蒿同时具有异交和自交的混合交配系统, 正如曾经报道过的菊科植物Chaetanthera euphrasioides (Arroyo et al ., 2006 )、茄科植物山莨菪(段元文等, 2007b )、龙胆科植物线叶龙胆(Gentiana lawrencei var. farreri )(侯勤正等, 2009 )、茄参(Mand- ragora caulescens ) (万金鹏等, 2011 )、锦葵科植物Kosteletzkya virginica (Ruan et al ., 2011 )以及With- ania ashwagandha (Mir et al ., 2013 )等植物一样, 导致植物异交的隔离机制处于非完全状态, 使得全缘叶绿绒蒿能够通过自交方式产生种子以保证结实. ...
Reproductive ecology of an endemic angiosperm, Meconopsis latifolia Prain (Papaveraceae), in the Kashmir Himalaya, India
1
2010
... 绿绒蒿属(Meconopsis )植物绝大多数分布在以横断山脉-喜马拉雅为中心的高山环境中(庄璇, 1981 ), 作为传统藏药, 目前的研究主要集中在其化学成分及药理活性方面(尚小雅等, 2006 ; Shang et al ., 2006 ; Zhou et al ., 2009 ), 一些学者对本属植物进行了系统分类方面的探讨(Meng et al ., 2006 ; Singhal & Kumar, 2008 ; Yang et al ., 2012 ).作为一类长期适应高山环境的植物, 目前对绿绒蒿属植物的繁殖特征、适应进化等方面尚缺乏系统研究(Dar et al ., 2010 ). ...
Patterns of interaction between plants and pollinators along an environmental gradient
2
2005
... 海拔带来的环境差异会使各类昆虫的活动受到限制或促进, 导致不同海拔地区访花者种类和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b ).在高山植物群落中, 鸟类和蝴蝶的传粉作用逐渐减弱, 熊蜂和蝇类成为主要的传粉昆虫(何亚平和刘建全, 2004 ; Duan et al ., 2007 ).对毛茛科植物Ranunculus acris 的研究发现, 低海拔地区昆虫的访花频率是高海拔地区的3倍(Totland, 2001 ).对凤仙花属(Impatiens )植物的研究发现, 高海拔地区和低海拔地区的传粉昆虫种类存在差异, 熊蜂在高海拔地区的花粉传递中扮演着重要的角色(Tian et al ., 2004 ; 肖乐希和刘克明, 2009 ; 肖乐希, 2009 ; 毛志斌等, 2011 ).对露蕊乌头(Aconitum gymnandrum )的研究表明, 低海拔种群的熊蜂种类(3种)多于高海拔种群(2种), 且熊蜂在低海拔种群的总访花频率远远高于高海拔种群(张挺峰等, 2006 ).对横断山区大钟花(Megacodon stylophorus )的研究亦得到相似结果(Meng et al ., 2012 ).对海南凤仙花(Impatiens hainanensis )的传粉生物学研究亦发现, 高海拔种群的传粉者以绿条无垫蜂(Amegilla zonata )为主, 低、中海拔种群以黄黑无垫蜂(Amegilla leptocoma )为主, 且黄黑无垫蜂在低、中海拔种群的访问频率远高于高海拔种群(钟云芳等, 2014 ). ...
... 有研究表明, 海拔较高或积温较低的地区, 某些昆虫会受到温度的限制(Heinrich, 1972 ; Totland, 2001 ; Fabbro & Körner, 2004 ), 导致不同海拔地区访问者和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b ; Sieber et al ., 2011 ).全缘叶绿绒蒿最高海拔样地1 (4452 m)蝇类的访花频率最低((0.15 ± 0.09)次·花-1 ·h-1 ), 这可能与高海拔地区较低的温度有关; 但最高的蝇类访花频率((0.59 ± 0.04)次·花-1 ·h-1 )没有出现在较低海拔的样地中, 而是出现在样地2 (4215 m), 野外观察研究发现, 样地2中全缘叶绿绒蒿的个体数量最多、开花最为繁盛, 由此初步推测, 样地2中最高的蝇类访花频率可能与其较高的花展示度有关, 同时, 观察时间和研究样地缺乏重复试验的限制亦可能造成访花频率在各海拔间的差异. ...
高山植物扁蕾的延迟自交机制
1
2007
... 高山恶劣的自然环境条件如低温、大风、多雨以及生长季短等不利因素限制了传粉昆虫的种类及其活动能力和访花频率(Arroyo et al ., 1982 , 1985 ; Bingham & Orthner, 1998 ), 导致到达柱头的异花花粉数量减少, 继而影响植物的生殖成功率(张大勇和姜新华, 2001 ).但已有的研究表明, 具有异花传粉繁育系统的植物在高山植物群落中普遍存在, 它们往往采取延长花寿命(Arroyo et al ., 1985 ; Bingham & Orthner, 1998 ; Blionis & Vokou, 2002 ; Steinacher & Wagner, 2010 )、增加对繁殖结构的资源分配(Fabbro & Körner, 2004 )等策略来适应传粉昆虫的缺乏, 以保证种群的延续.同时, 一些高山植物会选择自花授粉这种以降低后代适合度为代价的方式保证种群的延续(Schoen et al ., 1996 ; 段元文等, 2007b ). ...
高山植物扁蕾的延迟自交机制
1
2007
... 高山恶劣的自然环境条件如低温、大风、多雨以及生长季短等不利因素限制了传粉昆虫的种类及其活动能力和访花频率(Arroyo et al ., 1982 , 1985 ; Bingham & Orthner, 1998 ), 导致到达柱头的异花花粉数量减少, 继而影响植物的生殖成功率(张大勇和姜新华, 2001 ).但已有的研究表明, 具有异花传粉繁育系统的植物在高山植物群落中普遍存在, 它们往往采取延长花寿命(Arroyo et al ., 1985 ; Bingham & Orthner, 1998 ; Blionis & Vokou, 2002 ; Steinacher & Wagner, 2010 )、增加对繁殖结构的资源分配(Fabbro & Körner, 2004 )等策略来适应传粉昆虫的缺乏, 以保证种群的延续.同时, 一些高山植物会选择自花授粉这种以降低后代适合度为代价的方式保证种群的延续(Schoen et al ., 1996 ; 段元文等, 2007b ). ...
Interannual fluctuations in floral longevity, pollinator visitation and pollination limitation of an alpine plant (Gentiana straminea Maxim., Gentianaceae) at two altitudes in the Qinghai-Tibetan Plateau
3
2007
... 海拔带来的环境差异会使各类昆虫的活动受到限制或促进, 导致不同海拔地区访花者种类和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b ).在高山植物群落中, 鸟类和蝴蝶的传粉作用逐渐减弱, 熊蜂和蝇类成为主要的传粉昆虫(何亚平和刘建全, 2004 ; Duan et al ., 2007 ).对毛茛科植物Ranunculus acris 的研究发现, 低海拔地区昆虫的访花频率是高海拔地区的3倍(Totland, 2001 ).对凤仙花属(Impatiens )植物的研究发现, 高海拔地区和低海拔地区的传粉昆虫种类存在差异, 熊蜂在高海拔地区的花粉传递中扮演着重要的角色(Tian et al ., 2004 ; 肖乐希和刘克明, 2009 ; 肖乐希, 2009 ; 毛志斌等, 2011 ).对露蕊乌头(Aconitum gymnandrum )的研究表明, 低海拔种群的熊蜂种类(3种)多于高海拔种群(2种), 且熊蜂在低海拔种群的总访花频率远远高于高海拔种群(张挺峰等, 2006 ).对横断山区大钟花(Megacodon stylophorus )的研究亦得到相似结果(Meng et al ., 2012 ).对海南凤仙花(Impatiens hainanensis )的传粉生物学研究亦发现, 高海拔种群的传粉者以绿条无垫蜂(Amegilla zonata )为主, 低、中海拔种群以黄黑无垫蜂(Amegilla leptocoma )为主, 且黄黑无垫蜂在低、中海拔种群的访问频率远高于高海拔种群(钟云芳等, 2014 ). ...
... 高山环境下, 由于传粉者的不足使得大多数植物存在花粉限制(段元文等, 2007b ; 司庆文等, 2010 ; 钟云芳等, 2014 ).面对传粉者缺乏的选择压力, 植物会采取多样的策略提高对传粉昆虫的吸引力, 如提高花展示(邵剑文等, 2008 ; 彭德力等, 2012 )、提供生殖场所(Fishman & Hadany, 2013 )等, 随着海拔升高, 一些植物通过延长花寿命获得交配的机会(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 ); 而低温环境下, 一些植物则通过提高花序内或花内温度来达到吸引传粉昆虫的目的(Stanton & Galen, 1989 ).如水母雪兔子(Saussurea medusa )通过花序温度的升高使熊蜂在花序上的停留时间超过1 min, 这在一定程度上对传粉者起到了吸引作用(Tsukaya, 2002 ).对冬季开花的毛茛科铁筷子属植物Helleborus foetidus 和Helleborus bocconei 的研究亦发现, 花内温度的升高使传粉者更加活跃(Vesprini & Pacini, 2010 ).同时, 一些植物往往会进化出各异的自交机制, 如通过雌雄蕊位置的变化促进花粉传递以保障成功繁殖.总结已有的研究发现, 植物雌雄蕊位置关系的改变方式呈现出多样化的特征, 如花丝伸长(段元文等, 2007a , 2007b )、花柱伸长(侯勤正等, 2009 )、花柱卷曲(孙杉等, 2010 )等. ...
... , 2007b )、花柱伸长(侯勤正等, 2009 )、花柱卷曲(孙杉等, 2010 )等. ...
山莨菪(茄科)的传粉生物学
5
2007
... 海拔带来的环境差异会使各类昆虫的活动受到限制或促进, 导致不同海拔地区访花者种类和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b ).在高山植物群落中, 鸟类和蝴蝶的传粉作用逐渐减弱, 熊蜂和蝇类成为主要的传粉昆虫(何亚平和刘建全, 2004 ; Duan et al ., 2007 ).对毛茛科植物Ranunculus acris 的研究发现, 低海拔地区昆虫的访花频率是高海拔地区的3倍(Totland, 2001 ).对凤仙花属(Impatiens )植物的研究发现, 高海拔地区和低海拔地区的传粉昆虫种类存在差异, 熊蜂在高海拔地区的花粉传递中扮演着重要的角色(Tian et al ., 2004 ; 肖乐希和刘克明, 2009 ; 肖乐希, 2009 ; 毛志斌等, 2011 ).对露蕊乌头(Aconitum gymnandrum )的研究表明, 低海拔种群的熊蜂种类(3种)多于高海拔种群(2种), 且熊蜂在低海拔种群的总访花频率远远高于高海拔种群(张挺峰等, 2006 ).对横断山区大钟花(Megacodon stylophorus )的研究亦得到相似结果(Meng et al ., 2012 ).对海南凤仙花(Impatiens hainanensis )的传粉生物学研究亦发现, 高海拔种群的传粉者以绿条无垫蜂(Amegilla zonata )为主, 低、中海拔种群以黄黑无垫蜂(Amegilla leptocoma )为主, 且黄黑无垫蜂在低、中海拔种群的访问频率远高于高海拔种群(钟云芳等, 2014 ). ...
... 高山环境下, 由于传粉者的不足使得大多数植物存在花粉限制(段元文等, 2007b ; 司庆文等, 2010 ; 钟云芳等, 2014 ).面对传粉者缺乏的选择压力, 植物会采取多样的策略提高对传粉昆虫的吸引力, 如提高花展示(邵剑文等, 2008 ; 彭德力等, 2012 )、提供生殖场所(Fishman & Hadany, 2013 )等, 随着海拔升高, 一些植物通过延长花寿命获得交配的机会(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 ); 而低温环境下, 一些植物则通过提高花序内或花内温度来达到吸引传粉昆虫的目的(Stanton & Galen, 1989 ).如水母雪兔子(Saussurea medusa )通过花序温度的升高使熊蜂在花序上的停留时间超过1 min, 这在一定程度上对传粉者起到了吸引作用(Tsukaya, 2002 ).对冬季开花的毛茛科铁筷子属植物Helleborus foetidus 和Helleborus bocconei 的研究亦发现, 花内温度的升高使传粉者更加活跃(Vesprini & Pacini, 2010 ).同时, 一些植物往往会进化出各异的自交机制, 如通过雌雄蕊位置的变化促进花粉传递以保障成功繁殖.总结已有的研究发现, 植物雌雄蕊位置关系的改变方式呈现出多样化的特征, 如花丝伸长(段元文等, 2007a , 2007b )、花柱伸长(侯勤正等, 2009 )、花柱卷曲(孙杉等, 2010 )等. ...
... 为检测自交亲和性及自花传粉及异花传粉对结实的贡献, 参考段元文等(2007b) 的方法, 分别于2013年和2014年对5个样地进行了5种人工授粉试验: (1)去雄套袋: 随机选择并标记20个植株上的20个将要开放的花蕾, 去除雄蕊后套袋以检测是否存在无融合生殖现象; (2)去雄不套网: 于开花初期, 随机选择并标记20个植株上的20个将要开放的花蕾, 去掉雄蕊后不做任何处理, 拟检查异花花粉对全缘叶绿绒蒿结实的贡献; (3)套网: 随机选取并标记20个植株上的20个将要开放的花蕾并套网, 拟检测全缘叶绿绒蒿自花传粉的能力以及是否产生种子; (4)人工自交; (5)人工异交: 开花初期, 随机选取20对位于不同植株上将要开放的花蕾并去雄套袋(套袋隔离的花中未发现任何昆虫), 根据同一样地中其他花的开放时间(约2天), 将每对中的一朵花授以同一植株上刚散出的花粉, 另一朵花授以10 m以外(保证异交)的植株上刚散出的花粉, 授粉后仍套袋(防止雨水冲刷柱头上的花粉和隔离访花昆虫).另外每样地随机选取20朵花不做任何处理作为上述所有处理的对照.值得注意的是, 同种处理均选取不同植株上的花蕾.于授粉后40天(果实成熟但未开裂)收集所标记的花, 统计坐果率(fruit set rate)和每个果实的成熟种子数、不成熟种子数和败育胚珠数, 计算结实率.结实率(seed set rate)=成熟种子数/ (成熟种子数+不成熟种子数+败育种子数) × 100%. ...
... 有研究表明, 海拔较高或积温较低的地区, 某些昆虫会受到温度的限制(Heinrich, 1972 ; Totland, 2001 ; Fabbro & Körner, 2004 ), 导致不同海拔地区访问者和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b ; Sieber et al ., 2011 ).全缘叶绿绒蒿最高海拔样地1 (4452 m)蝇类的访花频率最低((0.15 ± 0.09)次·花-1 ·h-1 ), 这可能与高海拔地区较低的温度有关; 但最高的蝇类访花频率((0.59 ± 0.04)次·花-1 ·h-1 )没有出现在较低海拔的样地中, 而是出现在样地2 (4215 m), 野外观察研究发现, 样地2中全缘叶绿绒蒿的个体数量最多、开花最为繁盛, 由此初步推测, 样地2中最高的蝇类访花频率可能与其较高的花展示度有关, 同时, 观察时间和研究样地缺乏重复试验的限制亦可能造成访花频率在各海拔间的差异. ...
... 全缘叶绿绒蒿P/O值为901.24 ± 43.01, 这与Dafni (1992) 指出的兼性异交的物种相似, 这也说明全缘叶绿绒蒿同时具有异交和自交的混合交配系统, 正如曾经报道过的菊科植物Chaetanthera euphrasioides (Arroyo et al ., 2006 )、茄科植物山莨菪(段元文等, 2007b )、龙胆科植物线叶龙胆(Gentiana lawrencei var. farreri )(侯勤正等, 2009 )、茄参(Mand- ragora caulescens ) (万金鹏等, 2011 )、锦葵科植物Kosteletzkya virginica (Ruan et al ., 2011 )以及With- ania ashwagandha (Mir et al ., 2013 )等植物一样, 导致植物异交的隔离机制处于非完全状态, 使得全缘叶绿绒蒿能够通过自交方式产生种子以保证结实. ...
山莨菪(茄科)的传粉生物学
5
2007
... 海拔带来的环境差异会使各类昆虫的活动受到限制或促进, 导致不同海拔地区访花者种类和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b ).在高山植物群落中, 鸟类和蝴蝶的传粉作用逐渐减弱, 熊蜂和蝇类成为主要的传粉昆虫(何亚平和刘建全, 2004 ; Duan et al ., 2007 ).对毛茛科植物Ranunculus acris 的研究发现, 低海拔地区昆虫的访花频率是高海拔地区的3倍(Totland, 2001 ).对凤仙花属(Impatiens )植物的研究发现, 高海拔地区和低海拔地区的传粉昆虫种类存在差异, 熊蜂在高海拔地区的花粉传递中扮演着重要的角色(Tian et al ., 2004 ; 肖乐希和刘克明, 2009 ; 肖乐希, 2009 ; 毛志斌等, 2011 ).对露蕊乌头(Aconitum gymnandrum )的研究表明, 低海拔种群的熊蜂种类(3种)多于高海拔种群(2种), 且熊蜂在低海拔种群的总访花频率远远高于高海拔种群(张挺峰等, 2006 ).对横断山区大钟花(Megacodon stylophorus )的研究亦得到相似结果(Meng et al ., 2012 ).对海南凤仙花(Impatiens hainanensis )的传粉生物学研究亦发现, 高海拔种群的传粉者以绿条无垫蜂(Amegilla zonata )为主, 低、中海拔种群以黄黑无垫蜂(Amegilla leptocoma )为主, 且黄黑无垫蜂在低、中海拔种群的访问频率远高于高海拔种群(钟云芳等, 2014 ). ...
... 高山环境下, 由于传粉者的不足使得大多数植物存在花粉限制(段元文等, 2007b ; 司庆文等, 2010 ; 钟云芳等, 2014 ).面对传粉者缺乏的选择压力, 植物会采取多样的策略提高对传粉昆虫的吸引力, 如提高花展示(邵剑文等, 2008 ; 彭德力等, 2012 )、提供生殖场所(Fishman & Hadany, 2013 )等, 随着海拔升高, 一些植物通过延长花寿命获得交配的机会(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 ); 而低温环境下, 一些植物则通过提高花序内或花内温度来达到吸引传粉昆虫的目的(Stanton & Galen, 1989 ).如水母雪兔子(Saussurea medusa )通过花序温度的升高使熊蜂在花序上的停留时间超过1 min, 这在一定程度上对传粉者起到了吸引作用(Tsukaya, 2002 ).对冬季开花的毛茛科铁筷子属植物Helleborus foetidus 和Helleborus bocconei 的研究亦发现, 花内温度的升高使传粉者更加活跃(Vesprini & Pacini, 2010 ).同时, 一些植物往往会进化出各异的自交机制, 如通过雌雄蕊位置的变化促进花粉传递以保障成功繁殖.总结已有的研究发现, 植物雌雄蕊位置关系的改变方式呈现出多样化的特征, 如花丝伸长(段元文等, 2007a , 2007b )、花柱伸长(侯勤正等, 2009 )、花柱卷曲(孙杉等, 2010 )等. ...
... 为检测自交亲和性及自花传粉及异花传粉对结实的贡献, 参考段元文等(2007b) 的方法, 分别于2013年和2014年对5个样地进行了5种人工授粉试验: (1)去雄套袋: 随机选择并标记20个植株上的20个将要开放的花蕾, 去除雄蕊后套袋以检测是否存在无融合生殖现象; (2)去雄不套网: 于开花初期, 随机选择并标记20个植株上的20个将要开放的花蕾, 去掉雄蕊后不做任何处理, 拟检查异花花粉对全缘叶绿绒蒿结实的贡献; (3)套网: 随机选取并标记20个植株上的20个将要开放的花蕾并套网, 拟检测全缘叶绿绒蒿自花传粉的能力以及是否产生种子; (4)人工自交; (5)人工异交: 开花初期, 随机选取20对位于不同植株上将要开放的花蕾并去雄套袋(套袋隔离的花中未发现任何昆虫), 根据同一样地中其他花的开放时间(约2天), 将每对中的一朵花授以同一植株上刚散出的花粉, 另一朵花授以10 m以外(保证异交)的植株上刚散出的花粉, 授粉后仍套袋(防止雨水冲刷柱头上的花粉和隔离访花昆虫).另外每样地随机选取20朵花不做任何处理作为上述所有处理的对照.值得注意的是, 同种处理均选取不同植株上的花蕾.于授粉后40天(果实成熟但未开裂)收集所标记的花, 统计坐果率(fruit set rate)和每个果实的成熟种子数、不成熟种子数和败育胚珠数, 计算结实率.结实率(seed set rate)=成熟种子数/ (成熟种子数+不成熟种子数+败育种子数) × 100%. ...
... 有研究表明, 海拔较高或积温较低的地区, 某些昆虫会受到温度的限制(Heinrich, 1972 ; Totland, 2001 ; Fabbro & Körner, 2004 ), 导致不同海拔地区访问者和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b ; Sieber et al ., 2011 ).全缘叶绿绒蒿最高海拔样地1 (4452 m)蝇类的访花频率最低((0.15 ± 0.09)次·花-1 ·h-1 ), 这可能与高海拔地区较低的温度有关; 但最高的蝇类访花频率((0.59 ± 0.04)次·花-1 ·h-1 )没有出现在较低海拔的样地中, 而是出现在样地2 (4215 m), 野外观察研究发现, 样地2中全缘叶绿绒蒿的个体数量最多、开花最为繁盛, 由此初步推测, 样地2中最高的蝇类访花频率可能与其较高的花展示度有关, 同时, 观察时间和研究样地缺乏重复试验的限制亦可能造成访花频率在各海拔间的差异. ...
... 全缘叶绿绒蒿P/O值为901.24 ± 43.01, 这与Dafni (1992) 指出的兼性异交的物种相似, 这也说明全缘叶绿绒蒿同时具有异交和自交的混合交配系统, 正如曾经报道过的菊科植物Chaetanthera euphrasioides (Arroyo et al ., 2006 )、茄科植物山莨菪(段元文等, 2007b )、龙胆科植物线叶龙胆(Gentiana lawrencei var. farreri )(侯勤正等, 2009 )、茄参(Mand- ragora caulescens ) (万金鹏等, 2011 )、锦葵科植物Kosteletzkya virginica (Ruan et al ., 2011 )以及With- ania ashwagandha (Mir et al ., 2013 )等植物一样, 导致植物异交的隔离机制处于非完全状态, 使得全缘叶绿绒蒿能够通过自交方式产生种子以保证结实. ...
Altitudinal differences in flower traits and reproductive allocation
2
2004
... 高山恶劣的自然环境条件如低温、大风、多雨以及生长季短等不利因素限制了传粉昆虫的种类及其活动能力和访花频率(Arroyo et al ., 1982 , 1985 ; Bingham & Orthner, 1998 ), 导致到达柱头的异花花粉数量减少, 继而影响植物的生殖成功率(张大勇和姜新华, 2001 ).但已有的研究表明, 具有异花传粉繁育系统的植物在高山植物群落中普遍存在, 它们往往采取延长花寿命(Arroyo et al ., 1985 ; Bingham & Orthner, 1998 ; Blionis & Vokou, 2002 ; Steinacher & Wagner, 2010 )、增加对繁殖结构的资源分配(Fabbro & Körner, 2004 )等策略来适应传粉昆虫的缺乏, 以保证种群的延续.同时, 一些高山植物会选择自花授粉这种以降低后代适合度为代价的方式保证种群的延续(Schoen et al ., 1996 ; 段元文等, 2007b ). ...
... 有研究表明, 海拔较高或积温较低的地区, 某些昆虫会受到温度的限制(Heinrich, 1972 ; Totland, 2001 ; Fabbro & Körner, 2004 ), 导致不同海拔地区访问者和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b ; Sieber et al ., 2011 ).全缘叶绿绒蒿最高海拔样地1 (4452 m)蝇类的访花频率最低((0.15 ± 0.09)次·花-1 ·h-1 ), 这可能与高海拔地区较低的温度有关; 但最高的蝇类访花频率((0.59 ± 0.04)次·花-1 ·h-1 )没有出现在较低海拔的样地中, 而是出现在样地2 (4215 m), 野外观察研究发现, 样地2中全缘叶绿绒蒿的个体数量最多、开花最为繁盛, 由此初步推测, 样地2中最高的蝇类访花频率可能与其较高的花展示度有关, 同时, 观察时间和研究样地缺乏重复试验的限制亦可能造成访花频率在各海拔间的差异. ...
Pollinators’ mating rendezvous and the evolution of floral advertisement
1
2013
... 高山环境下, 由于传粉者的不足使得大多数植物存在花粉限制(段元文等, 2007b ; 司庆文等, 2010 ; 钟云芳等, 2014 ).面对传粉者缺乏的选择压力, 植物会采取多样的策略提高对传粉昆虫的吸引力, 如提高花展示(邵剑文等, 2008 ; 彭德力等, 2012 )、提供生殖场所(Fishman & Hadany, 2013 )等, 随着海拔升高, 一些植物通过延长花寿命获得交配的机会(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 ); 而低温环境下, 一些植物则通过提高花序内或花内温度来达到吸引传粉昆虫的目的(Stanton & Galen, 1989 ).如水母雪兔子(Saussurea medusa )通过花序温度的升高使熊蜂在花序上的停留时间超过1 min, 这在一定程度上对传粉者起到了吸引作用(Tsukaya, 2002 ).对冬季开花的毛茛科铁筷子属植物Helleborus foetidus 和Helleborus bocconei 的研究亦发现, 花内温度的升高使传粉者更加活跃(Vesprini & Pacini, 2010 ).同时, 一些植物往往会进化出各异的自交机制, 如通过雌雄蕊位置的变化促进花粉传递以保障成功繁殖.总结已有的研究发现, 植物雌雄蕊位置关系的改变方式呈现出多样化的特征, 如花丝伸长(段元文等, 2007a , 2007b )、花柱伸长(侯勤正等, 2009 )、花柱卷曲(孙杉等, 2010 )等. ...
濒危植物独叶草(毛茛科)的传粉生物学及繁育系统研究
1
2006
... 环境温度的测定发现, 各海拔样地7:00环境温度均低于5 ℃, 19:00环境温度均低于3.5 ℃, 这大大低于昆虫的活动温度(贺海霞, 2006 ; 朱建华等, 2006 , 2010 ); 且7:00以前、20:00以后既未见昆虫飞行亦未见新的昆虫进入花内, 此时间段花冠不同程度的闭合, 昆虫蛰伏于花内.由此初步推测, 每日7:00以前和20:00以后没有昆虫访花. ...
濒危植物独叶草(毛茛科)的传粉生物学及繁育系统研究
1
2006
... 环境温度的测定发现, 各海拔样地7:00环境温度均低于5 ℃, 19:00环境温度均低于3.5 ℃, 这大大低于昆虫的活动温度(贺海霞, 2006 ; 朱建华等, 2006 , 2010 ); 且7:00以前、20:00以后既未见昆虫飞行亦未见新的昆虫进入花内, 此时间段花冠不同程度的闭合, 昆虫蛰伏于花内.由此初步推测, 每日7:00以前和20:00以后没有昆虫访花. ...
青藏高原高山植物麻花艽的传粉生态学研究
1
2004
... 海拔带来的环境差异会使各类昆虫的活动受到限制或促进, 导致不同海拔地区访花者种类和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b ).在高山植物群落中, 鸟类和蝴蝶的传粉作用逐渐减弱, 熊蜂和蝇类成为主要的传粉昆虫(何亚平和刘建全, 2004 ; Duan et al ., 2007 ).对毛茛科植物Ranunculus acris 的研究发现, 低海拔地区昆虫的访花频率是高海拔地区的3倍(Totland, 2001 ).对凤仙花属(Impatiens )植物的研究发现, 高海拔地区和低海拔地区的传粉昆虫种类存在差异, 熊蜂在高海拔地区的花粉传递中扮演着重要的角色(Tian et al ., 2004 ; 肖乐希和刘克明, 2009 ; 肖乐希, 2009 ; 毛志斌等, 2011 ).对露蕊乌头(Aconitum gymnandrum )的研究表明, 低海拔种群的熊蜂种类(3种)多于高海拔种群(2种), 且熊蜂在低海拔种群的总访花频率远远高于高海拔种群(张挺峰等, 2006 ).对横断山区大钟花(Megacodon stylophorus )的研究亦得到相似结果(Meng et al ., 2012 ).对海南凤仙花(Impatiens hainanensis )的传粉生物学研究亦发现, 高海拔种群的传粉者以绿条无垫蜂(Amegilla zonata )为主, 低、中海拔种群以黄黑无垫蜂(Amegilla leptocoma )为主, 且黄黑无垫蜂在低、中海拔种群的访问频率远高于高海拔种群(钟云芳等, 2014 ). ...
青藏高原高山植物麻花艽的传粉生态学研究
1
2004
... 海拔带来的环境差异会使各类昆虫的活动受到限制或促进, 导致不同海拔地区访花者种类和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b ).在高山植物群落中, 鸟类和蝴蝶的传粉作用逐渐减弱, 熊蜂和蝇类成为主要的传粉昆虫(何亚平和刘建全, 2004 ; Duan et al ., 2007 ).对毛茛科植物Ranunculus acris 的研究发现, 低海拔地区昆虫的访花频率是高海拔地区的3倍(Totland, 2001 ).对凤仙花属(Impatiens )植物的研究发现, 高海拔地区和低海拔地区的传粉昆虫种类存在差异, 熊蜂在高海拔地区的花粉传递中扮演着重要的角色(Tian et al ., 2004 ; 肖乐希和刘克明, 2009 ; 肖乐希, 2009 ; 毛志斌等, 2011 ).对露蕊乌头(Aconitum gymnandrum )的研究表明, 低海拔种群的熊蜂种类(3种)多于高海拔种群(2种), 且熊蜂在低海拔种群的总访花频率远远高于高海拔种群(张挺峰等, 2006 ).对横断山区大钟花(Megacodon stylophorus )的研究亦得到相似结果(Meng et al ., 2012 ).对海南凤仙花(Impatiens hainanensis )的传粉生物学研究亦发现, 高海拔种群的传粉者以绿条无垫蜂(Amegilla zonata )为主, 低、中海拔种群以黄黑无垫蜂(Amegilla leptocoma )为主, 且黄黑无垫蜂在低、中海拔种群的访问频率远高于高海拔种群(钟云芳等, 2014 ). ...
Energetics of temperature regulation and foraging in a bumblebee, Bombus terricola Kirby
1
1972
... 有研究表明, 海拔较高或积温较低的地区, 某些昆虫会受到温度的限制(Heinrich, 1972 ; Totland, 2001 ; Fabbro & Körner, 2004 ), 导致不同海拔地区访问者和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b ; Sieber et al ., 2011 ).全缘叶绿绒蒿最高海拔样地1 (4452 m)蝇类的访花频率最低((0.15 ± 0.09)次·花-1 ·h-1 ), 这可能与高海拔地区较低的温度有关; 但最高的蝇类访花频率((0.59 ± 0.04)次·花-1 ·h-1 )没有出现在较低海拔的样地中, 而是出现在样地2 (4215 m), 野外观察研究发现, 样地2中全缘叶绿绒蒿的个体数量最多、开花最为繁盛, 由此初步推测, 样地2中最高的蝇类访花频率可能与其较高的花展示度有关, 同时, 观察时间和研究样地缺乏重复试验的限制亦可能造成访花频率在各海拔间的差异. ...
Evolution of self-fertilization at geographical range margins? A comparison of demographic, floral, and mating system variables in central vs. peripheral populations of Aquilegia canadensis (Ranunculaceae)
1
2005
... 一般而言, 自交不亲和、雌雄异位和雌雄异熟被认为是促进异交、避免自交的重要机制(Ishii & Sakai, 2002 ; Anderson et al ., 2003 ; 侯勤正等, 2009 ).在异花传粉者不足等选择压力下, 一些物种能够演化出自交亲和或依赖自交的种群和个体(Busch, 2005 ; Herlihy & Eckert, 2005 ; 彭德力等, 2012 ).全缘叶绿绒蒿具有不完全雌雄异位和不完全雌雄异熟的特点, 属于自交亲和物种(self-compatible species).花部综合特征观察及人工授粉试验的研究表明, 生长于高山环境中的全缘叶绿绒蒿具有花药与柱头间的距离缩小、雌雄性功能展示时间的重叠等特征的变化, 且面临着传粉者不足造成的传粉限制.全缘叶绿绒蒿自交亲和及雌雄不完全异熟为单花的自动自交提供了前提条件, 同时不完全雌雄异位直接导致了单花自动自交现象的发生.具体而言, 在研究的5个样地中, 均发现花药于开花的第5天开始散粉, 此时柱头还具有较强的可授性, 这为自交提供了前提; 其次, 大多数观察个体(约65%)的花药和柱头在开花发育的中后期完全接触, 这为自动自交提供了花粉来源; 再者, 全缘叶绿绒蒿个体自交部分亲和, 这为自动自交奠定了生理基础.同时, 花药在花开放过程中始终紧贴花柱, 开花中后期花药与柱头的接触, 可能会由于昆虫在花内的活动而引起协助自交.各样地的结实均由于异花传粉者不足而受到传粉限制.两种不同类型的自交机制恰恰为该植物异花传粉者不足提供了一定程度上的繁殖补偿(Zhang & Li, 2008 ). ...
青藏高原晚期开花植物线叶龙胆的传粉生态学
5
2009
... 高山环境下, 由于传粉者的不足使得大多数植物存在花粉限制(段元文等, 2007b ; 司庆文等, 2010 ; 钟云芳等, 2014 ).面对传粉者缺乏的选择压力, 植物会采取多样的策略提高对传粉昆虫的吸引力, 如提高花展示(邵剑文等, 2008 ; 彭德力等, 2012 )、提供生殖场所(Fishman & Hadany, 2013 )等, 随着海拔升高, 一些植物通过延长花寿命获得交配的机会(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 ); 而低温环境下, 一些植物则通过提高花序内或花内温度来达到吸引传粉昆虫的目的(Stanton & Galen, 1989 ).如水母雪兔子(Saussurea medusa )通过花序温度的升高使熊蜂在花序上的停留时间超过1 min, 这在一定程度上对传粉者起到了吸引作用(Tsukaya, 2002 ).对冬季开花的毛茛科铁筷子属植物Helleborus foetidus 和Helleborus bocconei 的研究亦发现, 花内温度的升高使传粉者更加活跃(Vesprini & Pacini, 2010 ).同时, 一些植物往往会进化出各异的自交机制, 如通过雌雄蕊位置的变化促进花粉传递以保障成功繁殖.总结已有的研究发现, 植物雌雄蕊位置关系的改变方式呈现出多样化的特征, 如花丝伸长(段元文等, 2007a , 2007b )、花柱伸长(侯勤正等, 2009 )、花柱卷曲(孙杉等, 2010 )等. ...
... 花寿命具有可塑性, 环境的变化往往造成花寿命的延长或缩短(Clark & Husband, 2007 ; 张志强和李庆军, 2009 ; Torres-Díaz et al ., 2011 ), 而花寿命与植物繁殖成功率相关(Torres-Díaz et al ., 2011 ).Vesprini和Pacini (2005) 亦发现, 低温条件下, 铁筷子属植物Helleborus bocconei 和H . foetidus 采取长花寿命, 每天花粉囊以依次少量开裂的方式来适应传粉昆虫不足的情况.Steinacher和Wagner (2010) 对7种高山植物的研究发现, 当花朵被拜访的频率很低时, 植物会通过延长柱头可授性和增加花冠保鲜时间来延长花寿命.本研究发现, 海拔越高全缘叶绿绒蒿的花寿命与柱头可授性持续时间越长, 这可能与高海拔的低温条件以及由此造成的传粉者访花频率低(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 )有关.对于需要传粉者才能完成授粉的植物而言, 花寿命的延长不仅可以增加花被传粉者拜访的概率(张志强和李庆军, 2009 ), 也会增加自身花粉的移出和外来花粉落置的机会(彭德力等, 2012 ), 同时有利于获得更多的交配机会, 保证繁殖成功(Teixido et al ., 2011 ).可见, 全缘叶绿绒蒿花寿命的延长可能是其应对高山环境较低昆虫访花频率的一种“机会主义”有性生殖策略(侯勤正等, 2009 ), 是植物对高山环境有效适应的结果. ...
... 以往对高山植物传粉昆虫的研究发现, 熊蜂以其稳定的热调节系统、较强的活动能力和较高的传粉效率, 使其成为高山环境下最重要的传粉者(Bergman et al ., 1996 ; Blionis & Vokou, 2001 ; 侯勤正等, 2009 ; Rader et al ., 2013 ).但在高山植物的传粉过程中, 蝇类往往也扮演着重要的角色.发现厕蝇(Fannia sp.)是茄科植物山莨菪(Anisodus tanguticus )最重要的有效异花传粉昆虫; Pickering和Stock (2004) 在对澳大利亚高山群落中传粉昆虫的调查也发现, 蝇类比重高达62.3%, 且种类丰富、访花频繁; Sieber等(2011) 对欧洲高山植物紫草科齿缘草属植物Eritrichium nanum 的传粉昆虫调查发现, 蝇类比重达到97.5%.本研究亦发现, 尽管熊蜂在高山环境中有分布, 但其更倾向于拜访分泌花蜜的报春类植物, 而蝇类才是全缘叶绿绒蒿最重要的异花传粉昆虫.这可能与巴郎山环境中牲畜(牦牛、马)的粪便较多, 为蝇类提供了食物来源有关; 此外, 全缘叶绿绒蒿花冠的保温效果能够保障在环境条件恶劣时(如低温、降雨等)为蝇类等传粉昆虫提供庇护场所.观察中没有发现全缘叶绿绒蒿分泌花蜜, 亦没有发现蝇类取食花粉, 而蝇类数量与花内温度、环境温度的温差间的极显著相关关系(p < 0.01)表明全缘叶绿绒蒿可以通过向蝇类等提供保温庇护场所达到吸引昆虫为之传粉的目的.与生活在喜马拉雅山和横断山冰缘带的“温室植物” (greenhouse plant)和“棉毛植物”(cotton plant)以较大苞片调节花内温度不同, 全缘叶绿绒蒿的花瓣在调节花内温度的过程中起着重要作用.全缘叶绿绒蒿随环境温度改变而调节自身花内温度的机制是否对雌雄配子体的发育产生影响有待于进一步证实. ...
... 一般而言, 自交不亲和、雌雄异位和雌雄异熟被认为是促进异交、避免自交的重要机制(Ishii & Sakai, 2002 ; Anderson et al ., 2003 ; 侯勤正等, 2009 ).在异花传粉者不足等选择压力下, 一些物种能够演化出自交亲和或依赖自交的种群和个体(Busch, 2005 ; Herlihy & Eckert, 2005 ; 彭德力等, 2012 ).全缘叶绿绒蒿具有不完全雌雄异位和不完全雌雄异熟的特点, 属于自交亲和物种(self-compatible species).花部综合特征观察及人工授粉试验的研究表明, 生长于高山环境中的全缘叶绿绒蒿具有花药与柱头间的距离缩小、雌雄性功能展示时间的重叠等特征的变化, 且面临着传粉者不足造成的传粉限制.全缘叶绿绒蒿自交亲和及雌雄不完全异熟为单花的自动自交提供了前提条件, 同时不完全雌雄异位直接导致了单花自动自交现象的发生.具体而言, 在研究的5个样地中, 均发现花药于开花的第5天开始散粉, 此时柱头还具有较强的可授性, 这为自交提供了前提; 其次, 大多数观察个体(约65%)的花药和柱头在开花发育的中后期完全接触, 这为自动自交提供了花粉来源; 再者, 全缘叶绿绒蒿个体自交部分亲和, 这为自动自交奠定了生理基础.同时, 花药在花开放过程中始终紧贴花柱, 开花中后期花药与柱头的接触, 可能会由于昆虫在花内的活动而引起协助自交.各样地的结实均由于异花传粉者不足而受到传粉限制.两种不同类型的自交机制恰恰为该植物异花传粉者不足提供了一定程度上的繁殖补偿(Zhang & Li, 2008 ). ...
... 全缘叶绿绒蒿P/O值为901.24 ± 43.01, 这与Dafni (1992) 指出的兼性异交的物种相似, 这也说明全缘叶绿绒蒿同时具有异交和自交的混合交配系统, 正如曾经报道过的菊科植物Chaetanthera euphrasioides (Arroyo et al ., 2006 )、茄科植物山莨菪(段元文等, 2007b )、龙胆科植物线叶龙胆(Gentiana lawrencei var. farreri )(侯勤正等, 2009 )、茄参(Mand- ragora caulescens ) (万金鹏等, 2011 )、锦葵科植物Kosteletzkya virginica (Ruan et al ., 2011 )以及With- ania ashwagandha (Mir et al ., 2013 )等植物一样, 导致植物异交的隔离机制处于非完全状态, 使得全缘叶绿绒蒿能够通过自交方式产生种子以保证结实. ...
青藏高原晚期开花植物线叶龙胆的传粉生态学
5
2009
... 高山环境下, 由于传粉者的不足使得大多数植物存在花粉限制(段元文等, 2007b ; 司庆文等, 2010 ; 钟云芳等, 2014 ).面对传粉者缺乏的选择压力, 植物会采取多样的策略提高对传粉昆虫的吸引力, 如提高花展示(邵剑文等, 2008 ; 彭德力等, 2012 )、提供生殖场所(Fishman & Hadany, 2013 )等, 随着海拔升高, 一些植物通过延长花寿命获得交配的机会(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 ); 而低温环境下, 一些植物则通过提高花序内或花内温度来达到吸引传粉昆虫的目的(Stanton & Galen, 1989 ).如水母雪兔子(Saussurea medusa )通过花序温度的升高使熊蜂在花序上的停留时间超过1 min, 这在一定程度上对传粉者起到了吸引作用(Tsukaya, 2002 ).对冬季开花的毛茛科铁筷子属植物Helleborus foetidus 和Helleborus bocconei 的研究亦发现, 花内温度的升高使传粉者更加活跃(Vesprini & Pacini, 2010 ).同时, 一些植物往往会进化出各异的自交机制, 如通过雌雄蕊位置的变化促进花粉传递以保障成功繁殖.总结已有的研究发现, 植物雌雄蕊位置关系的改变方式呈现出多样化的特征, 如花丝伸长(段元文等, 2007a , 2007b )、花柱伸长(侯勤正等, 2009 )、花柱卷曲(孙杉等, 2010 )等. ...
... 花寿命具有可塑性, 环境的变化往往造成花寿命的延长或缩短(Clark & Husband, 2007 ; 张志强和李庆军, 2009 ; Torres-Díaz et al ., 2011 ), 而花寿命与植物繁殖成功率相关(Torres-Díaz et al ., 2011 ).Vesprini和Pacini (2005) 亦发现, 低温条件下, 铁筷子属植物Helleborus bocconei 和H . foetidus 采取长花寿命, 每天花粉囊以依次少量开裂的方式来适应传粉昆虫不足的情况.Steinacher和Wagner (2010) 对7种高山植物的研究发现, 当花朵被拜访的频率很低时, 植物会通过延长柱头可授性和增加花冠保鲜时间来延长花寿命.本研究发现, 海拔越高全缘叶绿绒蒿的花寿命与柱头可授性持续时间越长, 这可能与高海拔的低温条件以及由此造成的传粉者访花频率低(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 )有关.对于需要传粉者才能完成授粉的植物而言, 花寿命的延长不仅可以增加花被传粉者拜访的概率(张志强和李庆军, 2009 ), 也会增加自身花粉的移出和外来花粉落置的机会(彭德力等, 2012 ), 同时有利于获得更多的交配机会, 保证繁殖成功(Teixido et al ., 2011 ).可见, 全缘叶绿绒蒿花寿命的延长可能是其应对高山环境较低昆虫访花频率的一种“机会主义”有性生殖策略(侯勤正等, 2009 ), 是植物对高山环境有效适应的结果. ...
... 以往对高山植物传粉昆虫的研究发现, 熊蜂以其稳定的热调节系统、较强的活动能力和较高的传粉效率, 使其成为高山环境下最重要的传粉者(Bergman et al ., 1996 ; Blionis & Vokou, 2001 ; 侯勤正等, 2009 ; Rader et al ., 2013 ).但在高山植物的传粉过程中, 蝇类往往也扮演着重要的角色.发现厕蝇(Fannia sp.)是茄科植物山莨菪(Anisodus tanguticus )最重要的有效异花传粉昆虫; Pickering和Stock (2004) 在对澳大利亚高山群落中传粉昆虫的调查也发现, 蝇类比重高达62.3%, 且种类丰富、访花频繁; Sieber等(2011) 对欧洲高山植物紫草科齿缘草属植物Eritrichium nanum 的传粉昆虫调查发现, 蝇类比重达到97.5%.本研究亦发现, 尽管熊蜂在高山环境中有分布, 但其更倾向于拜访分泌花蜜的报春类植物, 而蝇类才是全缘叶绿绒蒿最重要的异花传粉昆虫.这可能与巴郎山环境中牲畜(牦牛、马)的粪便较多, 为蝇类提供了食物来源有关; 此外, 全缘叶绿绒蒿花冠的保温效果能够保障在环境条件恶劣时(如低温、降雨等)为蝇类等传粉昆虫提供庇护场所.观察中没有发现全缘叶绿绒蒿分泌花蜜, 亦没有发现蝇类取食花粉, 而蝇类数量与花内温度、环境温度的温差间的极显著相关关系(p < 0.01)表明全缘叶绿绒蒿可以通过向蝇类等提供保温庇护场所达到吸引昆虫为之传粉的目的.与生活在喜马拉雅山和横断山冰缘带的“温室植物” (greenhouse plant)和“棉毛植物”(cotton plant)以较大苞片调节花内温度不同, 全缘叶绿绒蒿的花瓣在调节花内温度的过程中起着重要作用.全缘叶绿绒蒿随环境温度改变而调节自身花内温度的机制是否对雌雄配子体的发育产生影响有待于进一步证实. ...
... 一般而言, 自交不亲和、雌雄异位和雌雄异熟被认为是促进异交、避免自交的重要机制(Ishii & Sakai, 2002 ; Anderson et al ., 2003 ; 侯勤正等, 2009 ).在异花传粉者不足等选择压力下, 一些物种能够演化出自交亲和或依赖自交的种群和个体(Busch, 2005 ; Herlihy & Eckert, 2005 ; 彭德力等, 2012 ).全缘叶绿绒蒿具有不完全雌雄异位和不完全雌雄异熟的特点, 属于自交亲和物种(self-compatible species).花部综合特征观察及人工授粉试验的研究表明, 生长于高山环境中的全缘叶绿绒蒿具有花药与柱头间的距离缩小、雌雄性功能展示时间的重叠等特征的变化, 且面临着传粉者不足造成的传粉限制.全缘叶绿绒蒿自交亲和及雌雄不完全异熟为单花的自动自交提供了前提条件, 同时不完全雌雄异位直接导致了单花自动自交现象的发生.具体而言, 在研究的5个样地中, 均发现花药于开花的第5天开始散粉, 此时柱头还具有较强的可授性, 这为自交提供了前提; 其次, 大多数观察个体(约65%)的花药和柱头在开花发育的中后期完全接触, 这为自动自交提供了花粉来源; 再者, 全缘叶绿绒蒿个体自交部分亲和, 这为自动自交奠定了生理基础.同时, 花药在花开放过程中始终紧贴花柱, 开花中后期花药与柱头的接触, 可能会由于昆虫在花内的活动而引起协助自交.各样地的结实均由于异花传粉者不足而受到传粉限制.两种不同类型的自交机制恰恰为该植物异花传粉者不足提供了一定程度上的繁殖补偿(Zhang & Li, 2008 ). ...
... 全缘叶绿绒蒿P/O值为901.24 ± 43.01, 这与Dafni (1992) 指出的兼性异交的物种相似, 这也说明全缘叶绿绒蒿同时具有异交和自交的混合交配系统, 正如曾经报道过的菊科植物Chaetanthera euphrasioides (Arroyo et al ., 2006 )、茄科植物山莨菪(段元文等, 2007b )、龙胆科植物线叶龙胆(Gentiana lawrencei var. farreri )(侯勤正等, 2009 )、茄参(Mand- ragora caulescens ) (万金鹏等, 2011 )、锦葵科植物Kosteletzkya virginica (Ruan et al ., 2011 )以及With- ania ashwagandha (Mir et al ., 2013 )等植物一样, 导致植物异交的隔离机制处于非完全状态, 使得全缘叶绿绒蒿能够通过自交方式产生种子以保证结实. ...
Temporal variation in floral display size and individual floral sex allocation in racemes of Narthecium asiaticum (Liliaceae)
1
2002
... 一般而言, 自交不亲和、雌雄异位和雌雄异熟被认为是促进异交、避免自交的重要机制(Ishii & Sakai, 2002 ; Anderson et al ., 2003 ; 侯勤正等, 2009 ).在异花传粉者不足等选择压力下, 一些物种能够演化出自交亲和或依赖自交的种群和个体(Busch, 2005 ; Herlihy & Eckert, 2005 ; 彭德力等, 2012 ).全缘叶绿绒蒿具有不完全雌雄异位和不完全雌雄异熟的特点, 属于自交亲和物种(self-compatible species).花部综合特征观察及人工授粉试验的研究表明, 生长于高山环境中的全缘叶绿绒蒿具有花药与柱头间的距离缩小、雌雄性功能展示时间的重叠等特征的变化, 且面临着传粉者不足造成的传粉限制.全缘叶绿绒蒿自交亲和及雌雄不完全异熟为单花的自动自交提供了前提条件, 同时不完全雌雄异位直接导致了单花自动自交现象的发生.具体而言, 在研究的5个样地中, 均发现花药于开花的第5天开始散粉, 此时柱头还具有较强的可授性, 这为自交提供了前提; 其次, 大多数观察个体(约65%)的花药和柱头在开花发育的中后期完全接触, 这为自动自交提供了花粉来源; 再者, 全缘叶绿绒蒿个体自交部分亲和, 这为自动自交奠定了生理基础.同时, 花药在花开放过程中始终紧贴花柱, 开花中后期花药与柱头的接触, 可能会由于昆虫在花内的活动而引起协助自交.各样地的结实均由于异花传粉者不足而受到传粉限制.两种不同类型的自交机制恰恰为该植物异花传粉者不足提供了一定程度上的繁殖补偿(Zhang & Li, 2008 ). ...
Flower morphology, phenology and visitor patterns in an alpine community on Mt Olympos, Greece
2
2008
... 高山环境下, 由于传粉者的不足使得大多数植物存在花粉限制(段元文等, 2007b ; 司庆文等, 2010 ; 钟云芳等, 2014 ).面对传粉者缺乏的选择压力, 植物会采取多样的策略提高对传粉昆虫的吸引力, 如提高花展示(邵剑文等, 2008 ; 彭德力等, 2012 )、提供生殖场所(Fishman & Hadany, 2013 )等, 随着海拔升高, 一些植物通过延长花寿命获得交配的机会(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 ); 而低温环境下, 一些植物则通过提高花序内或花内温度来达到吸引传粉昆虫的目的(Stanton & Galen, 1989 ).如水母雪兔子(Saussurea medusa )通过花序温度的升高使熊蜂在花序上的停留时间超过1 min, 这在一定程度上对传粉者起到了吸引作用(Tsukaya, 2002 ).对冬季开花的毛茛科铁筷子属植物Helleborus foetidus 和Helleborus bocconei 的研究亦发现, 花内温度的升高使传粉者更加活跃(Vesprini & Pacini, 2010 ).同时, 一些植物往往会进化出各异的自交机制, 如通过雌雄蕊位置的变化促进花粉传递以保障成功繁殖.总结已有的研究发现, 植物雌雄蕊位置关系的改变方式呈现出多样化的特征, 如花丝伸长(段元文等, 2007a , 2007b )、花柱伸长(侯勤正等, 2009 )、花柱卷曲(孙杉等, 2010 )等. ...
... 花寿命具有可塑性, 环境的变化往往造成花寿命的延长或缩短(Clark & Husband, 2007 ; 张志强和李庆军, 2009 ; Torres-Díaz et al ., 2011 ), 而花寿命与植物繁殖成功率相关(Torres-Díaz et al ., 2011 ).Vesprini和Pacini (2005) 亦发现, 低温条件下, 铁筷子属植物Helleborus bocconei 和H . foetidus 采取长花寿命, 每天花粉囊以依次少量开裂的方式来适应传粉昆虫不足的情况.Steinacher和Wagner (2010) 对7种高山植物的研究发现, 当花朵被拜访的频率很低时, 植物会通过延长柱头可授性和增加花冠保鲜时间来延长花寿命.本研究发现, 海拔越高全缘叶绿绒蒿的花寿命与柱头可授性持续时间越长, 这可能与高海拔的低温条件以及由此造成的传粉者访花频率低(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 )有关.对于需要传粉者才能完成授粉的植物而言, 花寿命的延长不仅可以增加花被传粉者拜访的概率(张志强和李庆军, 2009 ), 也会增加自身花粉的移出和外来花粉落置的机会(彭德力等, 2012 ), 同时有利于获得更多的交配机会, 保证繁殖成功(Teixido et al ., 2011 ).可见, 全缘叶绿绒蒿花寿命的延长可能是其应对高山环境较低昆虫访花频率的一种“机会主义”有性生殖策略(侯勤正等, 2009 ), 是植物对高山环境有效适应的结果. ...
侧穗凤仙花的传粉生态和繁育系统
1
2011
... 海拔带来的环境差异会使各类昆虫的活动受到限制或促进, 导致不同海拔地区访花者种类和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b ).在高山植物群落中, 鸟类和蝴蝶的传粉作用逐渐减弱, 熊蜂和蝇类成为主要的传粉昆虫(何亚平和刘建全, 2004 ; Duan et al ., 2007 ).对毛茛科植物Ranunculus acris 的研究发现, 低海拔地区昆虫的访花频率是高海拔地区的3倍(Totland, 2001 ).对凤仙花属(Impatiens )植物的研究发现, 高海拔地区和低海拔地区的传粉昆虫种类存在差异, 熊蜂在高海拔地区的花粉传递中扮演着重要的角色(Tian et al ., 2004 ; 肖乐希和刘克明, 2009 ; 肖乐希, 2009 ; 毛志斌等, 2011 ).对露蕊乌头(Aconitum gymnandrum )的研究表明, 低海拔种群的熊蜂种类(3种)多于高海拔种群(2种), 且熊蜂在低海拔种群的总访花频率远远高于高海拔种群(张挺峰等, 2006 ).对横断山区大钟花(Megacodon stylophorus )的研究亦得到相似结果(Meng et al ., 2012 ).对海南凤仙花(Impatiens hainanensis )的传粉生物学研究亦发现, 高海拔种群的传粉者以绿条无垫蜂(Amegilla zonata )为主, 低、中海拔种群以黄黑无垫蜂(Amegilla leptocoma )为主, 且黄黑无垫蜂在低、中海拔种群的访问频率远高于高海拔种群(钟云芳等, 2014 ). ...
侧穗凤仙花的传粉生态和繁育系统
1
2011
... 海拔带来的环境差异会使各类昆虫的活动受到限制或促进, 导致不同海拔地区访花者种类和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b ).在高山植物群落中, 鸟类和蝴蝶的传粉作用逐渐减弱, 熊蜂和蝇类成为主要的传粉昆虫(何亚平和刘建全, 2004 ; Duan et al ., 2007 ).对毛茛科植物Ranunculus acris 的研究发现, 低海拔地区昆虫的访花频率是高海拔地区的3倍(Totland, 2001 ).对凤仙花属(Impatiens )植物的研究发现, 高海拔地区和低海拔地区的传粉昆虫种类存在差异, 熊蜂在高海拔地区的花粉传递中扮演着重要的角色(Tian et al ., 2004 ; 肖乐希和刘克明, 2009 ; 肖乐希, 2009 ; 毛志斌等, 2011 ).对露蕊乌头(Aconitum gymnandrum )的研究表明, 低海拔种群的熊蜂种类(3种)多于高海拔种群(2种), 且熊蜂在低海拔种群的总访花频率远远高于高海拔种群(张挺峰等, 2006 ).对横断山区大钟花(Megacodon stylophorus )的研究亦得到相似结果(Meng et al ., 2012 ).对海南凤仙花(Impatiens hainanensis )的传粉生物学研究亦发现, 高海拔种群的传粉者以绿条无垫蜂(Amegilla zonata )为主, 低、中海拔种群以黄黑无垫蜂(Amegilla leptocoma )为主, 且黄黑无垫蜂在低、中海拔种群的访问频率远高于高海拔种群(钟云芳等, 2014 ). ...
Pollination ecology and its implication for conservation of an endangered perennial herb native to the East-Himalaya, Megacodon stylophorus (Gentianaceae)
1
2012
... 海拔带来的环境差异会使各类昆虫的活动受到限制或促进, 导致不同海拔地区访花者种类和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b ).在高山植物群落中, 鸟类和蝴蝶的传粉作用逐渐减弱, 熊蜂和蝇类成为主要的传粉昆虫(何亚平和刘建全, 2004 ; Duan et al ., 2007 ).对毛茛科植物Ranunculus acris 的研究发现, 低海拔地区昆虫的访花频率是高海拔地区的3倍(Totland, 2001 ).对凤仙花属(Impatiens )植物的研究发现, 高海拔地区和低海拔地区的传粉昆虫种类存在差异, 熊蜂在高海拔地区的花粉传递中扮演着重要的角色(Tian et al ., 2004 ; 肖乐希和刘克明, 2009 ; 肖乐希, 2009 ; 毛志斌等, 2011 ).对露蕊乌头(Aconitum gymnandrum )的研究表明, 低海拔种群的熊蜂种类(3种)多于高海拔种群(2种), 且熊蜂在低海拔种群的总访花频率远远高于高海拔种群(张挺峰等, 2006 ).对横断山区大钟花(Megacodon stylophorus )的研究亦得到相似结果(Meng et al ., 2012 ).对海南凤仙花(Impatiens hainanensis )的传粉生物学研究亦发现, 高海拔种群的传粉者以绿条无垫蜂(Amegilla zonata )为主, 低、中海拔种群以黄黑无垫蜂(Amegilla leptocoma )为主, 且黄黑无垫蜂在低、中海拔种群的访问频率远高于高海拔种群(钟云芳等, 2014 ). ...
A karyomorphological study on four species of Meconopsis Vig. (Papaveraceae) from the Hengduan Mountains, SW China
1
2006
... 绿绒蒿属(Meconopsis )植物绝大多数分布在以横断山脉-喜马拉雅为中心的高山环境中(庄璇, 1981 ), 作为传统藏药, 目前的研究主要集中在其化学成分及药理活性方面(尚小雅等, 2006 ; Shang et al ., 2006 ; Zhou et al ., 2009 ), 一些学者对本属植物进行了系统分类方面的探讨(Meng et al ., 2006 ; Singhal & Kumar, 2008 ; Yang et al ., 2012 ).作为一类长期适应高山环境的植物, 目前对绿绒蒿属植物的繁殖特征、适应进化等方面尚缺乏系统研究(Dar et al ., 2010 ). ...
Reproductive biology of Withania ashwagandha sp. Novo (Solanaceae)
1
2013
... 全缘叶绿绒蒿P/O值为901.24 ± 43.01, 这与Dafni (1992) 指出的兼性异交的物种相似, 这也说明全缘叶绿绒蒿同时具有异交和自交的混合交配系统, 正如曾经报道过的菊科植物Chaetanthera euphrasioides (Arroyo et al ., 2006 )、茄科植物山莨菪(段元文等, 2007b )、龙胆科植物线叶龙胆(Gentiana lawrencei var. farreri )(侯勤正等, 2009 )、茄参(Mand- ragora caulescens ) (万金鹏等, 2011 )、锦葵科植物Kosteletzkya virginica (Ruan et al ., 2011 )以及With- ania ashwagandha (Mir et al ., 2013 )等植物一样, 导致植物异交的隔离机制处于非完全状态, 使得全缘叶绿绒蒿能够通过自交方式产生种子以保证结实. ...
黑河中游沙枣的传粉生物学研究
1
2012
... 用TTC法(潘成臣等, 2012 )测定花粉活力.各样地分别随机选取20个植株上的20个即将开放的花蕾并标记, 于散粉当天采集标记的花朵并带回, 放于4 ℃冰箱冷藏.采集当日每隔2小时取出一批进行染色, 显微镜下统计3-5个视域内被染色花粉数和未被染色花粉数, 计算花粉活力.花粉活力=被染色花粉数/(被染色花粉数+未被染色花粉数) × 100%.此后每天取出一批进行染色并统计直至花粉没有活性. ...
黑河中游沙枣的传粉生物学研究
1
2012
... 用TTC法(潘成臣等, 2012 )测定花粉活力.各样地分别随机选取20个植株上的20个即将开放的花蕾并标记, 于散粉当天采集标记的花朵并带回, 放于4 ℃冰箱冷藏.采集当日每隔2小时取出一批进行染色, 显微镜下统计3-5个视域内被染色花粉数和未被染色花粉数, 计算花粉活力.花粉活力=被染色花粉数/(被染色花粉数+未被染色花粉数) × 100%.此后每天取出一批进行染色并统计直至花粉没有活性. ...
高山植物繁殖策略的研究进展
3
2012
... 高山环境下, 由于传粉者的不足使得大多数植物存在花粉限制(段元文等, 2007b ; 司庆文等, 2010 ; 钟云芳等, 2014 ).面对传粉者缺乏的选择压力, 植物会采取多样的策略提高对传粉昆虫的吸引力, 如提高花展示(邵剑文等, 2008 ; 彭德力等, 2012 )、提供生殖场所(Fishman & Hadany, 2013 )等, 随着海拔升高, 一些植物通过延长花寿命获得交配的机会(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 ); 而低温环境下, 一些植物则通过提高花序内或花内温度来达到吸引传粉昆虫的目的(Stanton & Galen, 1989 ).如水母雪兔子(Saussurea medusa )通过花序温度的升高使熊蜂在花序上的停留时间超过1 min, 这在一定程度上对传粉者起到了吸引作用(Tsukaya, 2002 ).对冬季开花的毛茛科铁筷子属植物Helleborus foetidus 和Helleborus bocconei 的研究亦发现, 花内温度的升高使传粉者更加活跃(Vesprini & Pacini, 2010 ).同时, 一些植物往往会进化出各异的自交机制, 如通过雌雄蕊位置的变化促进花粉传递以保障成功繁殖.总结已有的研究发现, 植物雌雄蕊位置关系的改变方式呈现出多样化的特征, 如花丝伸长(段元文等, 2007a , 2007b )、花柱伸长(侯勤正等, 2009 )、花柱卷曲(孙杉等, 2010 )等. ...
... 花寿命具有可塑性, 环境的变化往往造成花寿命的延长或缩短(Clark & Husband, 2007 ; 张志强和李庆军, 2009 ; Torres-Díaz et al ., 2011 ), 而花寿命与植物繁殖成功率相关(Torres-Díaz et al ., 2011 ).Vesprini和Pacini (2005) 亦发现, 低温条件下, 铁筷子属植物Helleborus bocconei 和H . foetidus 采取长花寿命, 每天花粉囊以依次少量开裂的方式来适应传粉昆虫不足的情况.Steinacher和Wagner (2010) 对7种高山植物的研究发现, 当花朵被拜访的频率很低时, 植物会通过延长柱头可授性和增加花冠保鲜时间来延长花寿命.本研究发现, 海拔越高全缘叶绿绒蒿的花寿命与柱头可授性持续时间越长, 这可能与高海拔的低温条件以及由此造成的传粉者访花频率低(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 )有关.对于需要传粉者才能完成授粉的植物而言, 花寿命的延长不仅可以增加花被传粉者拜访的概率(张志强和李庆军, 2009 ), 也会增加自身花粉的移出和外来花粉落置的机会(彭德力等, 2012 ), 同时有利于获得更多的交配机会, 保证繁殖成功(Teixido et al ., 2011 ).可见, 全缘叶绿绒蒿花寿命的延长可能是其应对高山环境较低昆虫访花频率的一种“机会主义”有性生殖策略(侯勤正等, 2009 ), 是植物对高山环境有效适应的结果. ...
... 一般而言, 自交不亲和、雌雄异位和雌雄异熟被认为是促进异交、避免自交的重要机制(Ishii & Sakai, 2002 ; Anderson et al ., 2003 ; 侯勤正等, 2009 ).在异花传粉者不足等选择压力下, 一些物种能够演化出自交亲和或依赖自交的种群和个体(Busch, 2005 ; Herlihy & Eckert, 2005 ; 彭德力等, 2012 ).全缘叶绿绒蒿具有不完全雌雄异位和不完全雌雄异熟的特点, 属于自交亲和物种(self-compatible species).花部综合特征观察及人工授粉试验的研究表明, 生长于高山环境中的全缘叶绿绒蒿具有花药与柱头间的距离缩小、雌雄性功能展示时间的重叠等特征的变化, 且面临着传粉者不足造成的传粉限制.全缘叶绿绒蒿自交亲和及雌雄不完全异熟为单花的自动自交提供了前提条件, 同时不完全雌雄异位直接导致了单花自动自交现象的发生.具体而言, 在研究的5个样地中, 均发现花药于开花的第5天开始散粉, 此时柱头还具有较强的可授性, 这为自交提供了前提; 其次, 大多数观察个体(约65%)的花药和柱头在开花发育的中后期完全接触, 这为自动自交提供了花粉来源; 再者, 全缘叶绿绒蒿个体自交部分亲和, 这为自动自交奠定了生理基础.同时, 花药在花开放过程中始终紧贴花柱, 开花中后期花药与柱头的接触, 可能会由于昆虫在花内的活动而引起协助自交.各样地的结实均由于异花传粉者不足而受到传粉限制.两种不同类型的自交机制恰恰为该植物异花传粉者不足提供了一定程度上的繁殖补偿(Zhang & Li, 2008 ). ...
高山植物繁殖策略的研究进展
3
2012
... 高山环境下, 由于传粉者的不足使得大多数植物存在花粉限制(段元文等, 2007b ; 司庆文等, 2010 ; 钟云芳等, 2014 ).面对传粉者缺乏的选择压力, 植物会采取多样的策略提高对传粉昆虫的吸引力, 如提高花展示(邵剑文等, 2008 ; 彭德力等, 2012 )、提供生殖场所(Fishman & Hadany, 2013 )等, 随着海拔升高, 一些植物通过延长花寿命获得交配的机会(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 ); 而低温环境下, 一些植物则通过提高花序内或花内温度来达到吸引传粉昆虫的目的(Stanton & Galen, 1989 ).如水母雪兔子(Saussurea medusa )通过花序温度的升高使熊蜂在花序上的停留时间超过1 min, 这在一定程度上对传粉者起到了吸引作用(Tsukaya, 2002 ).对冬季开花的毛茛科铁筷子属植物Helleborus foetidus 和Helleborus bocconei 的研究亦发现, 花内温度的升高使传粉者更加活跃(Vesprini & Pacini, 2010 ).同时, 一些植物往往会进化出各异的自交机制, 如通过雌雄蕊位置的变化促进花粉传递以保障成功繁殖.总结已有的研究发现, 植物雌雄蕊位置关系的改变方式呈现出多样化的特征, 如花丝伸长(段元文等, 2007a , 2007b )、花柱伸长(侯勤正等, 2009 )、花柱卷曲(孙杉等, 2010 )等. ...
... 花寿命具有可塑性, 环境的变化往往造成花寿命的延长或缩短(Clark & Husband, 2007 ; 张志强和李庆军, 2009 ; Torres-Díaz et al ., 2011 ), 而花寿命与植物繁殖成功率相关(Torres-Díaz et al ., 2011 ).Vesprini和Pacini (2005) 亦发现, 低温条件下, 铁筷子属植物Helleborus bocconei 和H . foetidus 采取长花寿命, 每天花粉囊以依次少量开裂的方式来适应传粉昆虫不足的情况.Steinacher和Wagner (2010) 对7种高山植物的研究发现, 当花朵被拜访的频率很低时, 植物会通过延长柱头可授性和增加花冠保鲜时间来延长花寿命.本研究发现, 海拔越高全缘叶绿绒蒿的花寿命与柱头可授性持续时间越长, 这可能与高海拔的低温条件以及由此造成的传粉者访花频率低(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 )有关.对于需要传粉者才能完成授粉的植物而言, 花寿命的延长不仅可以增加花被传粉者拜访的概率(张志强和李庆军, 2009 ), 也会增加自身花粉的移出和外来花粉落置的机会(彭德力等, 2012 ), 同时有利于获得更多的交配机会, 保证繁殖成功(Teixido et al ., 2011 ).可见, 全缘叶绿绒蒿花寿命的延长可能是其应对高山环境较低昆虫访花频率的一种“机会主义”有性生殖策略(侯勤正等, 2009 ), 是植物对高山环境有效适应的结果. ...
... 一般而言, 自交不亲和、雌雄异位和雌雄异熟被认为是促进异交、避免自交的重要机制(Ishii & Sakai, 2002 ; Anderson et al ., 2003 ; 侯勤正等, 2009 ).在异花传粉者不足等选择压力下, 一些物种能够演化出自交亲和或依赖自交的种群和个体(Busch, 2005 ; Herlihy & Eckert, 2005 ; 彭德力等, 2012 ).全缘叶绿绒蒿具有不完全雌雄异位和不完全雌雄异熟的特点, 属于自交亲和物种(self-compatible species).花部综合特征观察及人工授粉试验的研究表明, 生长于高山环境中的全缘叶绿绒蒿具有花药与柱头间的距离缩小、雌雄性功能展示时间的重叠等特征的变化, 且面临着传粉者不足造成的传粉限制.全缘叶绿绒蒿自交亲和及雌雄不完全异熟为单花的自动自交提供了前提条件, 同时不完全雌雄异位直接导致了单花自动自交现象的发生.具体而言, 在研究的5个样地中, 均发现花药于开花的第5天开始散粉, 此时柱头还具有较强的可授性, 这为自交提供了前提; 其次, 大多数观察个体(约65%)的花药和柱头在开花发育的中后期完全接触, 这为自动自交提供了花粉来源; 再者, 全缘叶绿绒蒿个体自交部分亲和, 这为自动自交奠定了生理基础.同时, 花药在花开放过程中始终紧贴花柱, 开花中后期花药与柱头的接触, 可能会由于昆虫在花内的活动而引起协助自交.各样地的结实均由于异花传粉者不足而受到传粉限制.两种不同类型的自交机制恰恰为该植物异花传粉者不足提供了一定程度上的繁殖补偿(Zhang & Li, 2008 ). ...
Insect colour preference compared to flower colours in the Australian Alps
1
2004
... 以往对高山植物传粉昆虫的研究发现, 熊蜂以其稳定的热调节系统、较强的活动能力和较高的传粉效率, 使其成为高山环境下最重要的传粉者(Bergman et al ., 1996 ; Blionis & Vokou, 2001 ; 侯勤正等, 2009 ; Rader et al ., 2013 ).但在高山植物的传粉过程中, 蝇类往往也扮演着重要的角色.发现厕蝇(Fannia sp.)是茄科植物山莨菪(Anisodus tanguticus )最重要的有效异花传粉昆虫; Pickering和Stock (2004) 在对澳大利亚高山群落中传粉昆虫的调查也发现, 蝇类比重高达62.3%, 且种类丰富、访花频繁; Sieber等(2011) 对欧洲高山植物紫草科齿缘草属植物Eritrichium nanum 的传粉昆虫调查发现, 蝇类比重达到97.5%.本研究亦发现, 尽管熊蜂在高山环境中有分布, 但其更倾向于拜访分泌花蜜的报春类植物, 而蝇类才是全缘叶绿绒蒿最重要的异花传粉昆虫.这可能与巴郎山环境中牲畜(牦牛、马)的粪便较多, 为蝇类提供了食物来源有关; 此外, 全缘叶绿绒蒿花冠的保温效果能够保障在环境条件恶劣时(如低温、降雨等)为蝇类等传粉昆虫提供庇护场所.观察中没有发现全缘叶绿绒蒿分泌花蜜, 亦没有发现蝇类取食花粉, 而蝇类数量与花内温度、环境温度的温差间的极显著相关关系(p < 0.01)表明全缘叶绿绒蒿可以通过向蝇类等提供保温庇护场所达到吸引昆虫为之传粉的目的.与生活在喜马拉雅山和横断山冰缘带的“温室植物” (greenhouse plant)和“棉毛植物”(cotton plant)以较大苞片调节花内温度不同, 全缘叶绿绒蒿的花瓣在调节花内温度的过程中起着重要作用.全缘叶绿绒蒿随环境温度改变而调节自身花内温度的机制是否对雌雄配子体的发育产生影响有待于进一步证实. ...
Diurnal effectiveness of pollination by bees and flies in agricultural Brassica rapa: Implications for ecosystem resilience
1
2013
... 以往对高山植物传粉昆虫的研究发现, 熊蜂以其稳定的热调节系统、较强的活动能力和较高的传粉效率, 使其成为高山环境下最重要的传粉者(Bergman et al ., 1996 ; Blionis & Vokou, 2001 ; 侯勤正等, 2009 ; Rader et al ., 2013 ).但在高山植物的传粉过程中, 蝇类往往也扮演着重要的角色.发现厕蝇(Fannia sp.)是茄科植物山莨菪(Anisodus tanguticus )最重要的有效异花传粉昆虫; Pickering和Stock (2004) 在对澳大利亚高山群落中传粉昆虫的调查也发现, 蝇类比重高达62.3%, 且种类丰富、访花频繁; Sieber等(2011) 对欧洲高山植物紫草科齿缘草属植物Eritrichium nanum 的传粉昆虫调查发现, 蝇类比重达到97.5%.本研究亦发现, 尽管熊蜂在高山环境中有分布, 但其更倾向于拜访分泌花蜜的报春类植物, 而蝇类才是全缘叶绿绒蒿最重要的异花传粉昆虫.这可能与巴郎山环境中牲畜(牦牛、马)的粪便较多, 为蝇类提供了食物来源有关; 此外, 全缘叶绿绒蒿花冠的保温效果能够保障在环境条件恶劣时(如低温、降雨等)为蝇类等传粉昆虫提供庇护场所.观察中没有发现全缘叶绿绒蒿分泌花蜜, 亦没有发现蝇类取食花粉, 而蝇类数量与花内温度、环境温度的温差间的极显著相关关系(p < 0.01)表明全缘叶绿绒蒿可以通过向蝇类等提供保温庇护场所达到吸引昆虫为之传粉的目的.与生活在喜马拉雅山和横断山冰缘带的“温室植物” (greenhouse plant)和“棉毛植物”(cotton plant)以较大苞片调节花内温度不同, 全缘叶绿绒蒿的花瓣在调节花内温度的过程中起着重要作用.全缘叶绿绒蒿随环境温度改变而调节自身花内温度的机制是否对雌雄配子体的发育产生影响有待于进一步证实. ...
Relationship between reproductive assurance and mixed mating in perennial Kosteletzkya virginica
1
2011
... 全缘叶绿绒蒿P/O值为901.24 ± 43.01, 这与Dafni (1992) 指出的兼性异交的物种相似, 这也说明全缘叶绿绒蒿同时具有异交和自交的混合交配系统, 正如曾经报道过的菊科植物Chaetanthera euphrasioides (Arroyo et al ., 2006 )、茄科植物山莨菪(段元文等, 2007b )、龙胆科植物线叶龙胆(Gentiana lawrencei var. farreri )(侯勤正等, 2009 )、茄参(Mand- ragora caulescens ) (万金鹏等, 2011 )、锦葵科植物Kosteletzkya virginica (Ruan et al ., 2011 )以及With- ania ashwagandha (Mir et al ., 2013 )等植物一样, 导致植物异交的隔离机制处于非完全状态, 使得全缘叶绿绒蒿能够通过自交方式产生种子以保证结实. ...
How does self-pollination evolve? Inferences from floral ecology and molecular genetic variation. Philosophical Transactions of the Royal Society of London (
1
1996
... 高山恶劣的自然环境条件如低温、大风、多雨以及生长季短等不利因素限制了传粉昆虫的种类及其活动能力和访花频率(Arroyo et al ., 1982 , 1985 ; Bingham & Orthner, 1998 ), 导致到达柱头的异花花粉数量减少, 继而影响植物的生殖成功率(张大勇和姜新华, 2001 ).但已有的研究表明, 具有异花传粉繁育系统的植物在高山植物群落中普遍存在, 它们往往采取延长花寿命(Arroyo et al ., 1985 ; Bingham & Orthner, 1998 ; Blionis & Vokou, 2002 ; Steinacher & Wagner, 2010 )、增加对繁殖结构的资源分配(Fabbro & Körner, 2004 )等策略来适应传粉昆虫的缺乏, 以保证种群的延续.同时, 一些高山植物会选择自花授粉这种以降低后代适合度为代价的方式保证种群的延续(Schoen et al ., 1996 ; 段元文等, 2007b ). ...
藏药五脉绿绒蒿中非生物碱成分
1
2006
... 绿绒蒿属(Meconopsis )植物绝大多数分布在以横断山脉-喜马拉雅为中心的高山环境中(庄璇, 1981 ), 作为传统藏药, 目前的研究主要集中在其化学成分及药理活性方面(尚小雅等, 2006 ; Shang et al ., 2006 ; Zhou et al ., 2009 ), 一些学者对本属植物进行了系统分类方面的探讨(Meng et al ., 2006 ; Singhal & Kumar, 2008 ; Yang et al ., 2012 ).作为一类长期适应高山环境的植物, 目前对绿绒蒿属植物的繁殖特征、适应进化等方面尚缺乏系统研究(Dar et al ., 2010 ). ...
藏药五脉绿绒蒿中非生物碱成分
1
2006
... 绿绒蒿属(Meconopsis )植物绝大多数分布在以横断山脉-喜马拉雅为中心的高山环境中(庄璇, 1981 ), 作为传统藏药, 目前的研究主要集中在其化学成分及药理活性方面(尚小雅等, 2006 ; Shang et al ., 2006 ; Zhou et al ., 2009 ), 一些学者对本属植物进行了系统分类方面的探讨(Meng et al ., 2006 ; Singhal & Kumar, 2008 ; Yang et al ., 2012 ).作为一类长期适应高山环境的植物, 目前对绿绒蒿属植物的繁殖特征、适应进化等方面尚缺乏系统研究(Dar et al ., 2010 ). ...
Acetylated flavonol diglucosides from Meconopsis quintuplinervia
1
2006
... 绿绒蒿属(Meconopsis )植物绝大多数分布在以横断山脉-喜马拉雅为中心的高山环境中(庄璇, 1981 ), 作为传统藏药, 目前的研究主要集中在其化学成分及药理活性方面(尚小雅等, 2006 ; Shang et al ., 2006 ; Zhou et al ., 2009 ), 一些学者对本属植物进行了系统分类方面的探讨(Meng et al ., 2006 ; Singhal & Kumar, 2008 ; Yang et al ., 2012 ).作为一类长期适应高山环境的植物, 目前对绿绒蒿属植物的繁殖特征、适应进化等方面尚缺乏系统研究(Dar et al ., 2010 ). ...
安徽羽叶报春的有效传粉昆虫及花朵密度和种群大小对传粉效果的影响
1
2008
... 高山环境下, 由于传粉者的不足使得大多数植物存在花粉限制(段元文等, 2007b ; 司庆文等, 2010 ; 钟云芳等, 2014 ).面对传粉者缺乏的选择压力, 植物会采取多样的策略提高对传粉昆虫的吸引力, 如提高花展示(邵剑文等, 2008 ; 彭德力等, 2012 )、提供生殖场所(Fishman & Hadany, 2013 )等, 随着海拔升高, 一些植物通过延长花寿命获得交配的机会(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 ); 而低温环境下, 一些植物则通过提高花序内或花内温度来达到吸引传粉昆虫的目的(Stanton & Galen, 1989 ).如水母雪兔子(Saussurea medusa )通过花序温度的升高使熊蜂在花序上的停留时间超过1 min, 这在一定程度上对传粉者起到了吸引作用(Tsukaya, 2002 ).对冬季开花的毛茛科铁筷子属植物Helleborus foetidus 和Helleborus bocconei 的研究亦发现, 花内温度的升高使传粉者更加活跃(Vesprini & Pacini, 2010 ).同时, 一些植物往往会进化出各异的自交机制, 如通过雌雄蕊位置的变化促进花粉传递以保障成功繁殖.总结已有的研究发现, 植物雌雄蕊位置关系的改变方式呈现出多样化的特征, 如花丝伸长(段元文等, 2007a , 2007b )、花柱伸长(侯勤正等, 2009 )、花柱卷曲(孙杉等, 2010 )等. ...
安徽羽叶报春的有效传粉昆虫及花朵密度和种群大小对传粉效果的影响
1
2008
... 高山环境下, 由于传粉者的不足使得大多数植物存在花粉限制(段元文等, 2007b ; 司庆文等, 2010 ; 钟云芳等, 2014 ).面对传粉者缺乏的选择压力, 植物会采取多样的策略提高对传粉昆虫的吸引力, 如提高花展示(邵剑文等, 2008 ; 彭德力等, 2012 )、提供生殖场所(Fishman & Hadany, 2013 )等, 随着海拔升高, 一些植物通过延长花寿命获得交配的机会(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 ); 而低温环境下, 一些植物则通过提高花序内或花内温度来达到吸引传粉昆虫的目的(Stanton & Galen, 1989 ).如水母雪兔子(Saussurea medusa )通过花序温度的升高使熊蜂在花序上的停留时间超过1 min, 这在一定程度上对传粉者起到了吸引作用(Tsukaya, 2002 ).对冬季开花的毛茛科铁筷子属植物Helleborus foetidus 和Helleborus bocconei 的研究亦发现, 花内温度的升高使传粉者更加活跃(Vesprini & Pacini, 2010 ).同时, 一些植物往往会进化出各异的自交机制, 如通过雌雄蕊位置的变化促进花粉传递以保障成功繁殖.总结已有的研究发现, 植物雌雄蕊位置关系的改变方式呈现出多样化的特征, 如花丝伸长(段元文等, 2007a , 2007b )、花柱伸长(侯勤正等, 2009 )、花柱卷曲(孙杉等, 2010 )等. ...
达乌里秦艽的传粉生物学研究
1
2010
... 高山环境下, 由于传粉者的不足使得大多数植物存在花粉限制(段元文等, 2007b ; 司庆文等, 2010 ; 钟云芳等, 2014 ).面对传粉者缺乏的选择压力, 植物会采取多样的策略提高对传粉昆虫的吸引力, 如提高花展示(邵剑文等, 2008 ; 彭德力等, 2012 )、提供生殖场所(Fishman & Hadany, 2013 )等, 随着海拔升高, 一些植物通过延长花寿命获得交配的机会(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 ); 而低温环境下, 一些植物则通过提高花序内或花内温度来达到吸引传粉昆虫的目的(Stanton & Galen, 1989 ).如水母雪兔子(Saussurea medusa )通过花序温度的升高使熊蜂在花序上的停留时间超过1 min, 这在一定程度上对传粉者起到了吸引作用(Tsukaya, 2002 ).对冬季开花的毛茛科铁筷子属植物Helleborus foetidus 和Helleborus bocconei 的研究亦发现, 花内温度的升高使传粉者更加活跃(Vesprini & Pacini, 2010 ).同时, 一些植物往往会进化出各异的自交机制, 如通过雌雄蕊位置的变化促进花粉传递以保障成功繁殖.总结已有的研究发现, 植物雌雄蕊位置关系的改变方式呈现出多样化的特征, 如花丝伸长(段元文等, 2007a , 2007b )、花柱伸长(侯勤正等, 2009 )、花柱卷曲(孙杉等, 2010 )等. ...
达乌里秦艽的传粉生物学研究
1
2010
... 高山环境下, 由于传粉者的不足使得大多数植物存在花粉限制(段元文等, 2007b ; 司庆文等, 2010 ; 钟云芳等, 2014 ).面对传粉者缺乏的选择压力, 植物会采取多样的策略提高对传粉昆虫的吸引力, 如提高花展示(邵剑文等, 2008 ; 彭德力等, 2012 )、提供生殖场所(Fishman & Hadany, 2013 )等, 随着海拔升高, 一些植物通过延长花寿命获得交配的机会(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 ); 而低温环境下, 一些植物则通过提高花序内或花内温度来达到吸引传粉昆虫的目的(Stanton & Galen, 1989 ).如水母雪兔子(Saussurea medusa )通过花序温度的升高使熊蜂在花序上的停留时间超过1 min, 这在一定程度上对传粉者起到了吸引作用(Tsukaya, 2002 ).对冬季开花的毛茛科铁筷子属植物Helleborus foetidus 和Helleborus bocconei 的研究亦发现, 花内温度的升高使传粉者更加活跃(Vesprini & Pacini, 2010 ).同时, 一些植物往往会进化出各异的自交机制, 如通过雌雄蕊位置的变化促进花粉传递以保障成功繁殖.总结已有的研究发现, 植物雌雄蕊位置关系的改变方式呈现出多样化的特征, 如花丝伸长(段元文等, 2007a , 2007b )、花柱伸长(侯勤正等, 2009 )、花柱卷曲(孙杉等, 2010 )等. ...
Do alpine plants facilitate each other’s pollination? Experiments at a small spatial scale
2
2011
... 以往对高山植物传粉昆虫的研究发现, 熊蜂以其稳定的热调节系统、较强的活动能力和较高的传粉效率, 使其成为高山环境下最重要的传粉者(Bergman et al ., 1996 ; Blionis & Vokou, 2001 ; 侯勤正等, 2009 ; Rader et al ., 2013 ).但在高山植物的传粉过程中, 蝇类往往也扮演着重要的角色.发现厕蝇(Fannia sp.)是茄科植物山莨菪(Anisodus tanguticus )最重要的有效异花传粉昆虫; Pickering和Stock (2004) 在对澳大利亚高山群落中传粉昆虫的调查也发现, 蝇类比重高达62.3%, 且种类丰富、访花频繁; Sieber等(2011) 对欧洲高山植物紫草科齿缘草属植物Eritrichium nanum 的传粉昆虫调查发现, 蝇类比重达到97.5%.本研究亦发现, 尽管熊蜂在高山环境中有分布, 但其更倾向于拜访分泌花蜜的报春类植物, 而蝇类才是全缘叶绿绒蒿最重要的异花传粉昆虫.这可能与巴郎山环境中牲畜(牦牛、马)的粪便较多, 为蝇类提供了食物来源有关; 此外, 全缘叶绿绒蒿花冠的保温效果能够保障在环境条件恶劣时(如低温、降雨等)为蝇类等传粉昆虫提供庇护场所.观察中没有发现全缘叶绿绒蒿分泌花蜜, 亦没有发现蝇类取食花粉, 而蝇类数量与花内温度、环境温度的温差间的极显著相关关系(p < 0.01)表明全缘叶绿绒蒿可以通过向蝇类等提供保温庇护场所达到吸引昆虫为之传粉的目的.与生活在喜马拉雅山和横断山冰缘带的“温室植物” (greenhouse plant)和“棉毛植物”(cotton plant)以较大苞片调节花内温度不同, 全缘叶绿绒蒿的花瓣在调节花内温度的过程中起着重要作用.全缘叶绿绒蒿随环境温度改变而调节自身花内温度的机制是否对雌雄配子体的发育产生影响有待于进一步证实. ...
... 有研究表明, 海拔较高或积温较低的地区, 某些昆虫会受到温度的限制(Heinrich, 1972 ; Totland, 2001 ; Fabbro & Körner, 2004 ), 导致不同海拔地区访问者和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b ; Sieber et al ., 2011 ).全缘叶绿绒蒿最高海拔样地1 (4452 m)蝇类的访花频率最低((0.15 ± 0.09)次·花-1 ·h-1 ), 这可能与高海拔地区较低的温度有关; 但最高的蝇类访花频率((0.59 ± 0.04)次·花-1 ·h-1 )没有出现在较低海拔的样地中, 而是出现在样地2 (4215 m), 野外观察研究发现, 样地2中全缘叶绿绒蒿的个体数量最多、开花最为繁盛, 由此初步推测, 样地2中最高的蝇类访花频率可能与其较高的花展示度有关, 同时, 观察时间和研究样地缺乏重复试验的限制亦可能造成访花频率在各海拔间的差异. ...
Impact of cytomixis on meiosis, pollen viability and pollen size in wild populations of Himalayan poppy (Meconopsis aculeate Royle)
1
2008
... 绿绒蒿属(Meconopsis )植物绝大多数分布在以横断山脉-喜马拉雅为中心的高山环境中(庄璇, 1981 ), 作为传统藏药, 目前的研究主要集中在其化学成分及药理活性方面(尚小雅等, 2006 ; Shang et al ., 2006 ; Zhou et al ., 2009 ), 一些学者对本属植物进行了系统分类方面的探讨(Meng et al ., 2006 ; Singhal & Kumar, 2008 ; Yang et al ., 2012 ).作为一类长期适应高山环境的植物, 目前对绿绒蒿属植物的繁殖特征、适应进化等方面尚缺乏系统研究(Dar et al ., 2010 ). ...
Consequences of flower heliotropism for reproduction in an alpine buttercup (Ranunculus adoneus)
1
1989
... 高山环境下, 由于传粉者的不足使得大多数植物存在花粉限制(段元文等, 2007b ; 司庆文等, 2010 ; 钟云芳等, 2014 ).面对传粉者缺乏的选择压力, 植物会采取多样的策略提高对传粉昆虫的吸引力, 如提高花展示(邵剑文等, 2008 ; 彭德力等, 2012 )、提供生殖场所(Fishman & Hadany, 2013 )等, 随着海拔升高, 一些植物通过延长花寿命获得交配的机会(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 ); 而低温环境下, 一些植物则通过提高花序内或花内温度来达到吸引传粉昆虫的目的(Stanton & Galen, 1989 ).如水母雪兔子(Saussurea medusa )通过花序温度的升高使熊蜂在花序上的停留时间超过1 min, 这在一定程度上对传粉者起到了吸引作用(Tsukaya, 2002 ).对冬季开花的毛茛科铁筷子属植物Helleborus foetidus 和Helleborus bocconei 的研究亦发现, 花内温度的升高使传粉者更加活跃(Vesprini & Pacini, 2010 ).同时, 一些植物往往会进化出各异的自交机制, 如通过雌雄蕊位置的变化促进花粉传递以保障成功繁殖.总结已有的研究发现, 植物雌雄蕊位置关系的改变方式呈现出多样化的特征, 如花丝伸长(段元文等, 2007a , 2007b )、花柱伸长(侯勤正等, 2009 )、花柱卷曲(孙杉等, 2010 )等. ...
Flower longevity and duration of pistil receptivity in high mountain plants
2
2010
... 高山恶劣的自然环境条件如低温、大风、多雨以及生长季短等不利因素限制了传粉昆虫的种类及其活动能力和访花频率(Arroyo et al ., 1982 , 1985 ; Bingham & Orthner, 1998 ), 导致到达柱头的异花花粉数量减少, 继而影响植物的生殖成功率(张大勇和姜新华, 2001 ).但已有的研究表明, 具有异花传粉繁育系统的植物在高山植物群落中普遍存在, 它们往往采取延长花寿命(Arroyo et al ., 1985 ; Bingham & Orthner, 1998 ; Blionis & Vokou, 2002 ; Steinacher & Wagner, 2010 )、增加对繁殖结构的资源分配(Fabbro & Körner, 2004 )等策略来适应传粉昆虫的缺乏, 以保证种群的延续.同时, 一些高山植物会选择自花授粉这种以降低后代适合度为代价的方式保证种群的延续(Schoen et al ., 1996 ; 段元文等, 2007b ). ...
... 花寿命具有可塑性, 环境的变化往往造成花寿命的延长或缩短(Clark & Husband, 2007 ; 张志强和李庆军, 2009 ; Torres-Díaz et al ., 2011 ), 而花寿命与植物繁殖成功率相关(Torres-Díaz et al ., 2011 ).Vesprini和Pacini (2005) 亦发现, 低温条件下, 铁筷子属植物Helleborus bocconei 和H . foetidus 采取长花寿命, 每天花粉囊以依次少量开裂的方式来适应传粉昆虫不足的情况.Steinacher和Wagner (2010) 对7种高山植物的研究发现, 当花朵被拜访的频率很低时, 植物会通过延长柱头可授性和增加花冠保鲜时间来延长花寿命.本研究发现, 海拔越高全缘叶绿绒蒿的花寿命与柱头可授性持续时间越长, 这可能与高海拔的低温条件以及由此造成的传粉者访花频率低(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 )有关.对于需要传粉者才能完成授粉的植物而言, 花寿命的延长不仅可以增加花被传粉者拜访的概率(张志强和李庆军, 2009 ), 也会增加自身花粉的移出和外来花粉落置的机会(彭德力等, 2012 ), 同时有利于获得更多的交配机会, 保证繁殖成功(Teixido et al ., 2011 ).可见, 全缘叶绿绒蒿花寿命的延长可能是其应对高山环境较低昆虫访花频率的一种“机会主义”有性生殖策略(侯勤正等, 2009 ), 是植物对高山环境有效适应的结果. ...
Enzyme polymorphism analyses in three endangered species of Himalayan poppy, Meconopsis (Papaveraceae)
1
1996
... 与本属其他种以较小的种群分布于狭小的范围内相比, 全缘叶绿绒蒿(Meconopsis integrifolia )有着较广的分布范围(Sulaiman & Babu, 1996 ).作为一年生或多年生草本, 它主要分布于青藏高原海拔2500-5000 m的范围内, 一般在5月底到6月初进入花期, 此时属于高山植物的生长季早期, 传粉昆虫数量少, 异花传粉困难.全缘叶绿绒蒿会采取何种繁殖策略以适应高山环境, 这种繁殖策略是否随着环境的变化而发生改变? 为此, 本研究通过在四川省阿坝州巴郎山地区对不同海拔样地中的全缘叶绿绒蒿2年的连续观察和人工授粉试验, 重点回答以下问题: (1)全缘叶绿绒蒿的主要传粉昆虫是什么?异花传粉者的访花频率在不同环境中是否有差异?(2)它的繁育系统特点是什么?(3)其繁殖策略是否表现出随环境变化的特征? ...
花柱卷曲性的维持及功能性别特化
1
2010
... 高山环境下, 由于传粉者的不足使得大多数植物存在花粉限制(段元文等, 2007b ; 司庆文等, 2010 ; 钟云芳等, 2014 ).面对传粉者缺乏的选择压力, 植物会采取多样的策略提高对传粉昆虫的吸引力, 如提高花展示(邵剑文等, 2008 ; 彭德力等, 2012 )、提供生殖场所(Fishman & Hadany, 2013 )等, 随着海拔升高, 一些植物通过延长花寿命获得交配的机会(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 ); 而低温环境下, 一些植物则通过提高花序内或花内温度来达到吸引传粉昆虫的目的(Stanton & Galen, 1989 ).如水母雪兔子(Saussurea medusa )通过花序温度的升高使熊蜂在花序上的停留时间超过1 min, 这在一定程度上对传粉者起到了吸引作用(Tsukaya, 2002 ).对冬季开花的毛茛科铁筷子属植物Helleborus foetidus 和Helleborus bocconei 的研究亦发现, 花内温度的升高使传粉者更加活跃(Vesprini & Pacini, 2010 ).同时, 一些植物往往会进化出各异的自交机制, 如通过雌雄蕊位置的变化促进花粉传递以保障成功繁殖.总结已有的研究发现, 植物雌雄蕊位置关系的改变方式呈现出多样化的特征, 如花丝伸长(段元文等, 2007a , 2007b )、花柱伸长(侯勤正等, 2009 )、花柱卷曲(孙杉等, 2010 )等. ...
花柱卷曲性的维持及功能性别特化
1
2010
... 高山环境下, 由于传粉者的不足使得大多数植物存在花粉限制(段元文等, 2007b ; 司庆文等, 2010 ; 钟云芳等, 2014 ).面对传粉者缺乏的选择压力, 植物会采取多样的策略提高对传粉昆虫的吸引力, 如提高花展示(邵剑文等, 2008 ; 彭德力等, 2012 )、提供生殖场所(Fishman & Hadany, 2013 )等, 随着海拔升高, 一些植物通过延长花寿命获得交配的机会(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 ); 而低温环境下, 一些植物则通过提高花序内或花内温度来达到吸引传粉昆虫的目的(Stanton & Galen, 1989 ).如水母雪兔子(Saussurea medusa )通过花序温度的升高使熊蜂在花序上的停留时间超过1 min, 这在一定程度上对传粉者起到了吸引作用(Tsukaya, 2002 ).对冬季开花的毛茛科铁筷子属植物Helleborus foetidus 和Helleborus bocconei 的研究亦发现, 花内温度的升高使传粉者更加活跃(Vesprini & Pacini, 2010 ).同时, 一些植物往往会进化出各异的自交机制, 如通过雌雄蕊位置的变化促进花粉传递以保障成功繁殖.总结已有的研究发现, 植物雌雄蕊位置关系的改变方式呈现出多样化的特征, 如花丝伸长(段元文等, 2007a , 2007b )、花柱伸长(侯勤正等, 2009 )、花柱卷曲(孙杉等, 2010 )等. ...
Flower size and longevity influence florivory in the large-flowered shrub Cistus ladanifer
1
2011
... 花寿命具有可塑性, 环境的变化往往造成花寿命的延长或缩短(Clark & Husband, 2007 ; 张志强和李庆军, 2009 ; Torres-Díaz et al ., 2011 ), 而花寿命与植物繁殖成功率相关(Torres-Díaz et al ., 2011 ).Vesprini和Pacini (2005) 亦发现, 低温条件下, 铁筷子属植物Helleborus bocconei 和H . foetidus 采取长花寿命, 每天花粉囊以依次少量开裂的方式来适应传粉昆虫不足的情况.Steinacher和Wagner (2010) 对7种高山植物的研究发现, 当花朵被拜访的频率很低时, 植物会通过延长柱头可授性和增加花冠保鲜时间来延长花寿命.本研究发现, 海拔越高全缘叶绿绒蒿的花寿命与柱头可授性持续时间越长, 这可能与高海拔的低温条件以及由此造成的传粉者访花频率低(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 )有关.对于需要传粉者才能完成授粉的植物而言, 花寿命的延长不仅可以增加花被传粉者拜访的概率(张志强和李庆军, 2009 ), 也会增加自身花粉的移出和外来花粉落置的机会(彭德力等, 2012 ), 同时有利于获得更多的交配机会, 保证繁殖成功(Teixido et al ., 2011 ).可见, 全缘叶绿绒蒿花寿命的延长可能是其应对高山环境较低昆虫访花频率的一种“机会主义”有性生殖策略(侯勤正等, 2009 ), 是植物对高山环境有效适应的结果. ...
Pollination ecology and pollination system of Impatiens reptans (Balsaminaceae) endemic to China
1
2004
... 海拔带来的环境差异会使各类昆虫的活动受到限制或促进, 导致不同海拔地区访花者种类和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b ).在高山植物群落中, 鸟类和蝴蝶的传粉作用逐渐减弱, 熊蜂和蝇类成为主要的传粉昆虫(何亚平和刘建全, 2004 ; Duan et al ., 2007 ).对毛茛科植物Ranunculus acris 的研究发现, 低海拔地区昆虫的访花频率是高海拔地区的3倍(Totland, 2001 ).对凤仙花属(Impatiens )植物的研究发现, 高海拔地区和低海拔地区的传粉昆虫种类存在差异, 熊蜂在高海拔地区的花粉传递中扮演着重要的角色(Tian et al ., 2004 ; 肖乐希和刘克明, 2009 ; 肖乐希, 2009 ; 毛志斌等, 2011 ).对露蕊乌头(Aconitum gymnandrum )的研究表明, 低海拔种群的熊蜂种类(3种)多于高海拔种群(2种), 且熊蜂在低海拔种群的总访花频率远远高于高海拔种群(张挺峰等, 2006 ).对横断山区大钟花(Megacodon stylophorus )的研究亦得到相似结果(Meng et al ., 2012 ).对海南凤仙花(Impatiens hainanensis )的传粉生物学研究亦发现, 高海拔种群的传粉者以绿条无垫蜂(Amegilla zonata )为主, 低、中海拔种群以黄黑无垫蜂(Amegilla leptocoma )为主, 且黄黑无垫蜂在低、中海拔种群的访问频率远高于高海拔种群(钟云芳等, 2014 ). ...
Extremely long-lived stigmas allow extended cross- pollination opportunities in a high Andean plant
2
2011
... 花寿命具有可塑性, 环境的变化往往造成花寿命的延长或缩短(Clark & Husband, 2007 ; 张志强和李庆军, 2009 ; Torres-Díaz et al ., 2011 ), 而花寿命与植物繁殖成功率相关(Torres-Díaz et al ., 2011 ).Vesprini和Pacini (2005) 亦发现, 低温条件下, 铁筷子属植物Helleborus bocconei 和H . foetidus 采取长花寿命, 每天花粉囊以依次少量开裂的方式来适应传粉昆虫不足的情况.Steinacher和Wagner (2010) 对7种高山植物的研究发现, 当花朵被拜访的频率很低时, 植物会通过延长柱头可授性和增加花冠保鲜时间来延长花寿命.本研究发现, 海拔越高全缘叶绿绒蒿的花寿命与柱头可授性持续时间越长, 这可能与高海拔的低温条件以及由此造成的传粉者访花频率低(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 )有关.对于需要传粉者才能完成授粉的植物而言, 花寿命的延长不仅可以增加花被传粉者拜访的概率(张志强和李庆军, 2009 ), 也会增加自身花粉的移出和外来花粉落置的机会(彭德力等, 2012 ), 同时有利于获得更多的交配机会, 保证繁殖成功(Teixido et al ., 2011 ).可见, 全缘叶绿绒蒿花寿命的延长可能是其应对高山环境较低昆虫访花频率的一种“机会主义”有性生殖策略(侯勤正等, 2009 ), 是植物对高山环境有效适应的结果. ...
... ), 而花寿命与植物繁殖成功率相关(Torres-Díaz et al ., 2011 ).Vesprini和Pacini (2005) 亦发现, 低温条件下, 铁筷子属植物Helleborus bocconei 和H . foetidus 采取长花寿命, 每天花粉囊以依次少量开裂的方式来适应传粉昆虫不足的情况.Steinacher和Wagner (2010) 对7种高山植物的研究发现, 当花朵被拜访的频率很低时, 植物会通过延长柱头可授性和增加花冠保鲜时间来延长花寿命.本研究发现, 海拔越高全缘叶绿绒蒿的花寿命与柱头可授性持续时间越长, 这可能与高海拔的低温条件以及由此造成的传粉者访花频率低(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 )有关.对于需要传粉者才能完成授粉的植物而言, 花寿命的延长不仅可以增加花被传粉者拜访的概率(张志强和李庆军, 2009 ), 也会增加自身花粉的移出和外来花粉落置的机会(彭德力等, 2012 ), 同时有利于获得更多的交配机会, 保证繁殖成功(Teixido et al ., 2011 ).可见, 全缘叶绿绒蒿花寿命的延长可能是其应对高山环境较低昆虫访花频率的一种“机会主义”有性生殖策略(侯勤正等, 2009 ), 是植物对高山环境有效适应的结果. ...
Environment-dependent pollen limitation and selection on floral traits in an alpine species
2
2001
... 海拔带来的环境差异会使各类昆虫的活动受到限制或促进, 导致不同海拔地区访花者种类和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b ).在高山植物群落中, 鸟类和蝴蝶的传粉作用逐渐减弱, 熊蜂和蝇类成为主要的传粉昆虫(何亚平和刘建全, 2004 ; Duan et al ., 2007 ).对毛茛科植物Ranunculus acris 的研究发现, 低海拔地区昆虫的访花频率是高海拔地区的3倍(Totland, 2001 ).对凤仙花属(Impatiens )植物的研究发现, 高海拔地区和低海拔地区的传粉昆虫种类存在差异, 熊蜂在高海拔地区的花粉传递中扮演着重要的角色(Tian et al ., 2004 ; 肖乐希和刘克明, 2009 ; 肖乐希, 2009 ; 毛志斌等, 2011 ).对露蕊乌头(Aconitum gymnandrum )的研究表明, 低海拔种群的熊蜂种类(3种)多于高海拔种群(2种), 且熊蜂在低海拔种群的总访花频率远远高于高海拔种群(张挺峰等, 2006 ).对横断山区大钟花(Megacodon stylophorus )的研究亦得到相似结果(Meng et al ., 2012 ).对海南凤仙花(Impatiens hainanensis )的传粉生物学研究亦发现, 高海拔种群的传粉者以绿条无垫蜂(Amegilla zonata )为主, 低、中海拔种群以黄黑无垫蜂(Amegilla leptocoma )为主, 且黄黑无垫蜂在低、中海拔种群的访问频率远高于高海拔种群(钟云芳等, 2014 ). ...
... 有研究表明, 海拔较高或积温较低的地区, 某些昆虫会受到温度的限制(Heinrich, 1972 ; Totland, 2001 ; Fabbro & Körner, 2004 ), 导致不同海拔地区访问者和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b ; Sieber et al ., 2011 ).全缘叶绿绒蒿最高海拔样地1 (4452 m)蝇类的访花频率最低((0.15 ± 0.09)次·花-1 ·h-1 ), 这可能与高海拔地区较低的温度有关; 但最高的蝇类访花频率((0.59 ± 0.04)次·花-1 ·h-1 )没有出现在较低海拔的样地中, 而是出现在样地2 (4215 m), 野外观察研究发现, 样地2中全缘叶绿绒蒿的个体数量最多、开花最为繁盛, 由此初步推测, 样地2中最高的蝇类访花频率可能与其较高的花展示度有关, 同时, 观察时间和研究样地缺乏重复试验的限制亦可能造成访花频率在各海拔间的差异. ...
Optical and anatomical characteristics of bracts from the Chinese “glasshouse” plant, Rheum alexandrae Batalin (Ploygonaceae), in Yunnan, China
1
2002
... 高山环境下, 由于传粉者的不足使得大多数植物存在花粉限制(段元文等, 2007b ; 司庆文等, 2010 ; 钟云芳等, 2014 ).面对传粉者缺乏的选择压力, 植物会采取多样的策略提高对传粉昆虫的吸引力, 如提高花展示(邵剑文等, 2008 ; 彭德力等, 2012 )、提供生殖场所(Fishman & Hadany, 2013 )等, 随着海拔升高, 一些植物通过延长花寿命获得交配的机会(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 ); 而低温环境下, 一些植物则通过提高花序内或花内温度来达到吸引传粉昆虫的目的(Stanton & Galen, 1989 ).如水母雪兔子(Saussurea medusa )通过花序温度的升高使熊蜂在花序上的停留时间超过1 min, 这在一定程度上对传粉者起到了吸引作用(Tsukaya, 2002 ).对冬季开花的毛茛科铁筷子属植物Helleborus foetidus 和Helleborus bocconei 的研究亦发现, 花内温度的升高使传粉者更加活跃(Vesprini & Pacini, 2010 ).同时, 一些植物往往会进化出各异的自交机制, 如通过雌雄蕊位置的变化促进花粉传递以保障成功繁殖.总结已有的研究发现, 植物雌雄蕊位置关系的改变方式呈现出多样化的特征, 如花丝伸长(段元文等, 2007a , 2007b )、花柱伸长(侯勤正等, 2009 )、花柱卷曲(孙杉等, 2010 )等. ...
Temperature-dependent floral longevity in two Helleborus species
1
2005
... 花寿命具有可塑性, 环境的变化往往造成花寿命的延长或缩短(Clark & Husband, 2007 ; 张志强和李庆军, 2009 ; Torres-Díaz et al ., 2011 ), 而花寿命与植物繁殖成功率相关(Torres-Díaz et al ., 2011 ).Vesprini和Pacini (2005) 亦发现, 低温条件下, 铁筷子属植物Helleborus bocconei 和H . foetidus 采取长花寿命, 每天花粉囊以依次少量开裂的方式来适应传粉昆虫不足的情况.Steinacher和Wagner (2010) 对7种高山植物的研究发现, 当花朵被拜访的频率很低时, 植物会通过延长柱头可授性和增加花冠保鲜时间来延长花寿命.本研究发现, 海拔越高全缘叶绿绒蒿的花寿命与柱头可授性持续时间越长, 这可能与高海拔的低温条件以及由此造成的传粉者访花频率低(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 )有关.对于需要传粉者才能完成授粉的植物而言, 花寿命的延长不仅可以增加花被传粉者拜访的概率(张志强和李庆军, 2009 ), 也会增加自身花粉的移出和外来花粉落置的机会(彭德力等, 2012 ), 同时有利于获得更多的交配机会, 保证繁殖成功(Teixido et al ., 2011 ).可见, 全缘叶绿绒蒿花寿命的延长可能是其应对高山环境较低昆虫访花频率的一种“机会主义”有性生殖策略(侯勤正等, 2009 ), 是植物对高山环境有效适应的结果. ...
Pollination ecology in sympatric winter flowering Helleborus (Ranunculaceae)
1
2010
... 高山环境下, 由于传粉者的不足使得大多数植物存在花粉限制(段元文等, 2007b ; 司庆文等, 2010 ; 钟云芳等, 2014 ).面对传粉者缺乏的选择压力, 植物会采取多样的策略提高对传粉昆虫的吸引力, 如提高花展示(邵剑文等, 2008 ; 彭德力等, 2012 )、提供生殖场所(Fishman & Hadany, 2013 )等, 随着海拔升高, 一些植物通过延长花寿命获得交配的机会(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 ); 而低温环境下, 一些植物则通过提高花序内或花内温度来达到吸引传粉昆虫的目的(Stanton & Galen, 1989 ).如水母雪兔子(Saussurea medusa )通过花序温度的升高使熊蜂在花序上的停留时间超过1 min, 这在一定程度上对传粉者起到了吸引作用(Tsukaya, 2002 ).对冬季开花的毛茛科铁筷子属植物Helleborus foetidus 和Helleborus bocconei 的研究亦发现, 花内温度的升高使传粉者更加活跃(Vesprini & Pacini, 2010 ).同时, 一些植物往往会进化出各异的自交机制, 如通过雌雄蕊位置的变化促进花粉传递以保障成功繁殖.总结已有的研究发现, 植物雌雄蕊位置关系的改变方式呈现出多样化的特征, 如花丝伸长(段元文等, 2007a , 2007b )、花柱伸长(侯勤正等, 2009 )、花柱卷曲(孙杉等, 2010 )等. ...
雌先熟植物茄参(茄科)的繁育系统
1
2011
... 全缘叶绿绒蒿P/O值为901.24 ± 43.01, 这与Dafni (1992) 指出的兼性异交的物种相似, 这也说明全缘叶绿绒蒿同时具有异交和自交的混合交配系统, 正如曾经报道过的菊科植物Chaetanthera euphrasioides (Arroyo et al ., 2006 )、茄科植物山莨菪(段元文等, 2007b )、龙胆科植物线叶龙胆(Gentiana lawrencei var. farreri )(侯勤正等, 2009 )、茄参(Mand- ragora caulescens ) (万金鹏等, 2011 )、锦葵科植物Kosteletzkya virginica (Ruan et al ., 2011 )以及With- ania ashwagandha (Mir et al ., 2013 )等植物一样, 导致植物异交的隔离机制处于非完全状态, 使得全缘叶绿绒蒿能够通过自交方式产生种子以保证结实. ...
雌先熟植物茄参(茄科)的繁育系统
1
2011
... 全缘叶绿绒蒿P/O值为901.24 ± 43.01, 这与Dafni (1992) 指出的兼性异交的物种相似, 这也说明全缘叶绿绒蒿同时具有异交和自交的混合交配系统, 正如曾经报道过的菊科植物Chaetanthera euphrasioides (Arroyo et al ., 2006 )、茄科植物山莨菪(段元文等, 2007b )、龙胆科植物线叶龙胆(Gentiana lawrencei var. farreri )(侯勤正等, 2009 )、茄参(Mand- ragora caulescens ) (万金鹏等, 2011 )、锦葵科植物Kosteletzkya virginica (Ruan et al ., 2011 )以及With- ania ashwagandha (Mir et al ., 2013 )等植物一样, 导致植物异交的隔离机制处于非完全状态, 使得全缘叶绿绒蒿能够通过自交方式产生种子以保证结实. ...
四种凤仙花属(Impatiens L.) 植物的传粉生物学研究
1
2009
... 海拔带来的环境差异会使各类昆虫的活动受到限制或促进, 导致不同海拔地区访花者种类和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b ).在高山植物群落中, 鸟类和蝴蝶的传粉作用逐渐减弱, 熊蜂和蝇类成为主要的传粉昆虫(何亚平和刘建全, 2004 ; Duan et al ., 2007 ).对毛茛科植物Ranunculus acris 的研究发现, 低海拔地区昆虫的访花频率是高海拔地区的3倍(Totland, 2001 ).对凤仙花属(Impatiens )植物的研究发现, 高海拔地区和低海拔地区的传粉昆虫种类存在差异, 熊蜂在高海拔地区的花粉传递中扮演着重要的角色(Tian et al ., 2004 ; 肖乐希和刘克明, 2009 ; 肖乐希, 2009 ; 毛志斌等, 2011 ).对露蕊乌头(Aconitum gymnandrum )的研究表明, 低海拔种群的熊蜂种类(3种)多于高海拔种群(2种), 且熊蜂在低海拔种群的总访花频率远远高于高海拔种群(张挺峰等, 2006 ).对横断山区大钟花(Megacodon stylophorus )的研究亦得到相似结果(Meng et al ., 2012 ).对海南凤仙花(Impatiens hainanensis )的传粉生物学研究亦发现, 高海拔种群的传粉者以绿条无垫蜂(Amegilla zonata )为主, 低、中海拔种群以黄黑无垫蜂(Amegilla leptocoma )为主, 且黄黑无垫蜂在低、中海拔种群的访问频率远高于高海拔种群(钟云芳等, 2014 ). ...
四种凤仙花属(Impatiens L.) 植物的传粉生物学研究
1
2009
... 海拔带来的环境差异会使各类昆虫的活动受到限制或促进, 导致不同海拔地区访花者种类和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b ).在高山植物群落中, 鸟类和蝴蝶的传粉作用逐渐减弱, 熊蜂和蝇类成为主要的传粉昆虫(何亚平和刘建全, 2004 ; Duan et al ., 2007 ).对毛茛科植物Ranunculus acris 的研究发现, 低海拔地区昆虫的访花频率是高海拔地区的3倍(Totland, 2001 ).对凤仙花属(Impatiens )植物的研究发现, 高海拔地区和低海拔地区的传粉昆虫种类存在差异, 熊蜂在高海拔地区的花粉传递中扮演着重要的角色(Tian et al ., 2004 ; 肖乐希和刘克明, 2009 ; 肖乐希, 2009 ; 毛志斌等, 2011 ).对露蕊乌头(Aconitum gymnandrum )的研究表明, 低海拔种群的熊蜂种类(3种)多于高海拔种群(2种), 且熊蜂在低海拔种群的总访花频率远远高于高海拔种群(张挺峰等, 2006 ).对横断山区大钟花(Megacodon stylophorus )的研究亦得到相似结果(Meng et al ., 2012 ).对海南凤仙花(Impatiens hainanensis )的传粉生物学研究亦发现, 高海拔种群的传粉者以绿条无垫蜂(Amegilla zonata )为主, 低、中海拔种群以黄黑无垫蜂(Amegilla leptocoma )为主, 且黄黑无垫蜂在低、中海拔种群的访问频率远高于高海拔种群(钟云芳等, 2014 ). ...
华凤仙花部特征和传粉系统研究
1
2009
... 海拔带来的环境差异会使各类昆虫的活动受到限制或促进, 导致不同海拔地区访花者种类和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b ).在高山植物群落中, 鸟类和蝴蝶的传粉作用逐渐减弱, 熊蜂和蝇类成为主要的传粉昆虫(何亚平和刘建全, 2004 ; Duan et al ., 2007 ).对毛茛科植物Ranunculus acris 的研究发现, 低海拔地区昆虫的访花频率是高海拔地区的3倍(Totland, 2001 ).对凤仙花属(Impatiens )植物的研究发现, 高海拔地区和低海拔地区的传粉昆虫种类存在差异, 熊蜂在高海拔地区的花粉传递中扮演着重要的角色(Tian et al ., 2004 ; 肖乐希和刘克明, 2009 ; 肖乐希, 2009 ; 毛志斌等, 2011 ).对露蕊乌头(Aconitum gymnandrum )的研究表明, 低海拔种群的熊蜂种类(3种)多于高海拔种群(2种), 且熊蜂在低海拔种群的总访花频率远远高于高海拔种群(张挺峰等, 2006 ).对横断山区大钟花(Megacodon stylophorus )的研究亦得到相似结果(Meng et al ., 2012 ).对海南凤仙花(Impatiens hainanensis )的传粉生物学研究亦发现, 高海拔种群的传粉者以绿条无垫蜂(Amegilla zonata )为主, 低、中海拔种群以黄黑无垫蜂(Amegilla leptocoma )为主, 且黄黑无垫蜂在低、中海拔种群的访问频率远高于高海拔种群(钟云芳等, 2014 ). ...
华凤仙花部特征和传粉系统研究
1
2009
... 海拔带来的环境差异会使各类昆虫的活动受到限制或促进, 导致不同海拔地区访花者种类和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b ).在高山植物群落中, 鸟类和蝴蝶的传粉作用逐渐减弱, 熊蜂和蝇类成为主要的传粉昆虫(何亚平和刘建全, 2004 ; Duan et al ., 2007 ).对毛茛科植物Ranunculus acris 的研究发现, 低海拔地区昆虫的访花频率是高海拔地区的3倍(Totland, 2001 ).对凤仙花属(Impatiens )植物的研究发现, 高海拔地区和低海拔地区的传粉昆虫种类存在差异, 熊蜂在高海拔地区的花粉传递中扮演着重要的角色(Tian et al ., 2004 ; 肖乐希和刘克明, 2009 ; 肖乐希, 2009 ; 毛志斌等, 2011 ).对露蕊乌头(Aconitum gymnandrum )的研究表明, 低海拔种群的熊蜂种类(3种)多于高海拔种群(2种), 且熊蜂在低海拔种群的总访花频率远远高于高海拔种群(张挺峰等, 2006 ).对横断山区大钟花(Megacodon stylophorus )的研究亦得到相似结果(Meng et al ., 2012 ).对海南凤仙花(Impatiens hainanensis )的传粉生物学研究亦发现, 高海拔种群的传粉者以绿条无垫蜂(Amegilla zonata )为主, 低、中海拔种群以黄黑无垫蜂(Amegilla leptocoma )为主, 且黄黑无垫蜂在低、中海拔种群的访问频率远高于高海拔种群(钟云芳等, 2014 ). ...
Great genetic differentiation among populations of Meconopsis integrifolia and its implication for plant speciation in the Qinghai- Tibetan Plateau
1
2012
... 绿绒蒿属(Meconopsis )植物绝大多数分布在以横断山脉-喜马拉雅为中心的高山环境中(庄璇, 1981 ), 作为传统藏药, 目前的研究主要集中在其化学成分及药理活性方面(尚小雅等, 2006 ; Shang et al ., 2006 ; Zhou et al ., 2009 ), 一些学者对本属植物进行了系统分类方面的探讨(Meng et al ., 2006 ; Singhal & Kumar, 2008 ; Yang et al ., 2012 ).作为一类长期适应高山环境的植物, 目前对绿绒蒿属植物的繁殖特征、适应进化等方面尚缺乏系统研究(Dar et al ., 2010 ). ...
百日草柱头可授性和花粉生活力的研究
1
2007
... 在2013年盛花期进行柱头可授性与花粉活力检测.采用联苯胺-过氧化氢法(叶要妹等, 2007 )检测柱头可授性.分别于各样地随机选取5朵当天开放的花并标记, 此后每天均随机选取当日开放的5朵花直至最早标记的花柱头完全变黑, 收集各天标记的花, 将其柱头浸入凹面载玻片中, 若柱头具有可授性则柱头周围反应液呈蓝色并有大量气泡产生.用-和+表示观察到的情况.-表示柱头不具有可授性, 即无气泡与变蓝情况; +表示柱头具有可授性, 即有轻微的变蓝和气泡产生; ++表示柱头具有较强可授性, 即有较多的气泡和比较明显的变蓝情况; +++表示柱头具有极强可授性, 即有大量气泡和非常明显的变蓝情况. ...
百日草柱头可授性和花粉生活力的研究
1
2007
... 在2013年盛花期进行柱头可授性与花粉活力检测.采用联苯胺-过氧化氢法(叶要妹等, 2007 )检测柱头可授性.分别于各样地随机选取5朵当天开放的花并标记, 此后每天均随机选取当日开放的5朵花直至最早标记的花柱头完全变黑, 收集各天标记的花, 将其柱头浸入凹面载玻片中, 若柱头具有可授性则柱头周围反应液呈蓝色并有大量气泡产生.用-和+表示观察到的情况.-表示柱头不具有可授性, 即无气泡与变蓝情况; +表示柱头具有可授性, 即有轻微的变蓝和气泡产生; ++表示柱头具有较强可授性, 即有较多的气泡和比较明显的变蓝情况; +++表示柱头具有极强可授性, 即有大量气泡和非常明显的变蓝情况. ...
植物交配系统的进化、资源分配对策与遗传多样性
1
2001
... 高山恶劣的自然环境条件如低温、大风、多雨以及生长季短等不利因素限制了传粉昆虫的种类及其活动能力和访花频率(Arroyo et al ., 1982 , 1985 ; Bingham & Orthner, 1998 ), 导致到达柱头的异花花粉数量减少, 继而影响植物的生殖成功率(张大勇和姜新华, 2001 ).但已有的研究表明, 具有异花传粉繁育系统的植物在高山植物群落中普遍存在, 它们往往采取延长花寿命(Arroyo et al ., 1985 ; Bingham & Orthner, 1998 ; Blionis & Vokou, 2002 ; Steinacher & Wagner, 2010 )、增加对繁殖结构的资源分配(Fabbro & Körner, 2004 )等策略来适应传粉昆虫的缺乏, 以保证种群的延续.同时, 一些高山植物会选择自花授粉这种以降低后代适合度为代价的方式保证种群的延续(Schoen et al ., 1996 ; 段元文等, 2007b ). ...
植物交配系统的进化、资源分配对策与遗传多样性
1
2001
... 高山恶劣的自然环境条件如低温、大风、多雨以及生长季短等不利因素限制了传粉昆虫的种类及其活动能力和访花频率(Arroyo et al ., 1982 , 1985 ; Bingham & Orthner, 1998 ), 导致到达柱头的异花花粉数量减少, 继而影响植物的生殖成功率(张大勇和姜新华, 2001 ).但已有的研究表明, 具有异花传粉繁育系统的植物在高山植物群落中普遍存在, 它们往往采取延长花寿命(Arroyo et al ., 1985 ; Bingham & Orthner, 1998 ; Blionis & Vokou, 2002 ; Steinacher & Wagner, 2010 )、增加对繁殖结构的资源分配(Fabbro & Körner, 2004 )等策略来适应传粉昆虫的缺乏, 以保证种群的延续.同时, 一些高山植物会选择自花授粉这种以降低后代适合度为代价的方式保证种群的延续(Schoen et al ., 1996 ; 段元文等, 2007b ). ...
露蕊乌头(毛茛科)不同海拔居群的传粉生态学
1
2006
... 海拔带来的环境差异会使各类昆虫的活动受到限制或促进, 导致不同海拔地区访花者种类和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b ).在高山植物群落中, 鸟类和蝴蝶的传粉作用逐渐减弱, 熊蜂和蝇类成为主要的传粉昆虫(何亚平和刘建全, 2004 ; Duan et al ., 2007 ).对毛茛科植物Ranunculus acris 的研究发现, 低海拔地区昆虫的访花频率是高海拔地区的3倍(Totland, 2001 ).对凤仙花属(Impatiens )植物的研究发现, 高海拔地区和低海拔地区的传粉昆虫种类存在差异, 熊蜂在高海拔地区的花粉传递中扮演着重要的角色(Tian et al ., 2004 ; 肖乐希和刘克明, 2009 ; 肖乐希, 2009 ; 毛志斌等, 2011 ).对露蕊乌头(Aconitum gymnandrum )的研究表明, 低海拔种群的熊蜂种类(3种)多于高海拔种群(2种), 且熊蜂在低海拔种群的总访花频率远远高于高海拔种群(张挺峰等, 2006 ).对横断山区大钟花(Megacodon stylophorus )的研究亦得到相似结果(Meng et al ., 2012 ).对海南凤仙花(Impatiens hainanensis )的传粉生物学研究亦发现, 高海拔种群的传粉者以绿条无垫蜂(Amegilla zonata )为主, 低、中海拔种群以黄黑无垫蜂(Amegilla leptocoma )为主, 且黄黑无垫蜂在低、中海拔种群的访问频率远高于高海拔种群(钟云芳等, 2014 ). ...
露蕊乌头(毛茛科)不同海拔居群的传粉生态学
1
2006
... 海拔带来的环境差异会使各类昆虫的活动受到限制或促进, 导致不同海拔地区访花者种类和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b ).在高山植物群落中, 鸟类和蝴蝶的传粉作用逐渐减弱, 熊蜂和蝇类成为主要的传粉昆虫(何亚平和刘建全, 2004 ; Duan et al ., 2007 ).对毛茛科植物Ranunculus acris 的研究发现, 低海拔地区昆虫的访花频率是高海拔地区的3倍(Totland, 2001 ).对凤仙花属(Impatiens )植物的研究发现, 高海拔地区和低海拔地区的传粉昆虫种类存在差异, 熊蜂在高海拔地区的花粉传递中扮演着重要的角色(Tian et al ., 2004 ; 肖乐希和刘克明, 2009 ; 肖乐希, 2009 ; 毛志斌等, 2011 ).对露蕊乌头(Aconitum gymnandrum )的研究表明, 低海拔种群的熊蜂种类(3种)多于高海拔种群(2种), 且熊蜂在低海拔种群的总访花频率远远高于高海拔种群(张挺峰等, 2006 ).对横断山区大钟花(Megacodon stylophorus )的研究亦得到相似结果(Meng et al ., 2012 ).对海南凤仙花(Impatiens hainanensis )的传粉生物学研究亦发现, 高海拔种群的传粉者以绿条无垫蜂(Amegilla zonata )为主, 低、中海拔种群以黄黑无垫蜂(Amegilla leptocoma )为主, 且黄黑无垫蜂在低、中海拔种群的访问频率远高于高海拔种群(钟云芳等, 2014 ). ...
Autonomous selfing provides reproductive assurance in an alpine ginger Roscoea schneideriana (Zingiberaceae)
1
2008
... 一般而言, 自交不亲和、雌雄异位和雌雄异熟被认为是促进异交、避免自交的重要机制(Ishii & Sakai, 2002 ; Anderson et al ., 2003 ; 侯勤正等, 2009 ).在异花传粉者不足等选择压力下, 一些物种能够演化出自交亲和或依赖自交的种群和个体(Busch, 2005 ; Herlihy & Eckert, 2005 ; 彭德力等, 2012 ).全缘叶绿绒蒿具有不完全雌雄异位和不完全雌雄异熟的特点, 属于自交亲和物种(self-compatible species).花部综合特征观察及人工授粉试验的研究表明, 生长于高山环境中的全缘叶绿绒蒿具有花药与柱头间的距离缩小、雌雄性功能展示时间的重叠等特征的变化, 且面临着传粉者不足造成的传粉限制.全缘叶绿绒蒿自交亲和及雌雄不完全异熟为单花的自动自交提供了前提条件, 同时不完全雌雄异位直接导致了单花自动自交现象的发生.具体而言, 在研究的5个样地中, 均发现花药于开花的第5天开始散粉, 此时柱头还具有较强的可授性, 这为自交提供了前提; 其次, 大多数观察个体(约65%)的花药和柱头在开花发育的中后期完全接触, 这为自动自交提供了花粉来源; 再者, 全缘叶绿绒蒿个体自交部分亲和, 这为自动自交奠定了生理基础.同时, 花药在花开放过程中始终紧贴花柱, 开花中后期花药与柱头的接触, 可能会由于昆虫在花内的活动而引起协助自交.各样地的结实均由于异花传粉者不足而受到传粉限制.两种不同类型的自交机制恰恰为该植物异花传粉者不足提供了一定程度上的繁殖补偿(Zhang & Li, 2008 ). ...
花寿命的进化生态学意义
2
2009
... 花寿命具有可塑性, 环境的变化往往造成花寿命的延长或缩短(Clark & Husband, 2007 ; 张志强和李庆军, 2009 ; Torres-Díaz et al ., 2011 ), 而花寿命与植物繁殖成功率相关(Torres-Díaz et al ., 2011 ).Vesprini和Pacini (2005) 亦发现, 低温条件下, 铁筷子属植物Helleborus bocconei 和H . foetidus 采取长花寿命, 每天花粉囊以依次少量开裂的方式来适应传粉昆虫不足的情况.Steinacher和Wagner (2010) 对7种高山植物的研究发现, 当花朵被拜访的频率很低时, 植物会通过延长柱头可授性和增加花冠保鲜时间来延长花寿命.本研究发现, 海拔越高全缘叶绿绒蒿的花寿命与柱头可授性持续时间越长, 这可能与高海拔的低温条件以及由此造成的传粉者访花频率低(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 )有关.对于需要传粉者才能完成授粉的植物而言, 花寿命的延长不仅可以增加花被传粉者拜访的概率(张志强和李庆军, 2009 ), 也会增加自身花粉的移出和外来花粉落置的机会(彭德力等, 2012 ), 同时有利于获得更多的交配机会, 保证繁殖成功(Teixido et al ., 2011 ).可见, 全缘叶绿绒蒿花寿命的延长可能是其应对高山环境较低昆虫访花频率的一种“机会主义”有性生殖策略(侯勤正等, 2009 ), 是植物对高山环境有效适应的结果. ...
... )有关.对于需要传粉者才能完成授粉的植物而言, 花寿命的延长不仅可以增加花被传粉者拜访的概率(张志强和李庆军, 2009 ), 也会增加自身花粉的移出和外来花粉落置的机会(彭德力等, 2012 ), 同时有利于获得更多的交配机会, 保证繁殖成功(Teixido et al ., 2011 ).可见, 全缘叶绿绒蒿花寿命的延长可能是其应对高山环境较低昆虫访花频率的一种“机会主义”有性生殖策略(侯勤正等, 2009 ), 是植物对高山环境有效适应的结果. ...
花寿命的进化生态学意义
2
2009
... 花寿命具有可塑性, 环境的变化往往造成花寿命的延长或缩短(Clark & Husband, 2007 ; 张志强和李庆军, 2009 ; Torres-Díaz et al ., 2011 ), 而花寿命与植物繁殖成功率相关(Torres-Díaz et al ., 2011 ).Vesprini和Pacini (2005) 亦发现, 低温条件下, 铁筷子属植物Helleborus bocconei 和H . foetidus 采取长花寿命, 每天花粉囊以依次少量开裂的方式来适应传粉昆虫不足的情况.Steinacher和Wagner (2010) 对7种高山植物的研究发现, 当花朵被拜访的频率很低时, 植物会通过延长柱头可授性和增加花冠保鲜时间来延长花寿命.本研究发现, 海拔越高全缘叶绿绒蒿的花寿命与柱头可授性持续时间越长, 这可能与高海拔的低温条件以及由此造成的传粉者访花频率低(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 )有关.对于需要传粉者才能完成授粉的植物而言, 花寿命的延长不仅可以增加花被传粉者拜访的概率(张志强和李庆军, 2009 ), 也会增加自身花粉的移出和外来花粉落置的机会(彭德力等, 2012 ), 同时有利于获得更多的交配机会, 保证繁殖成功(Teixido et al ., 2011 ).可见, 全缘叶绿绒蒿花寿命的延长可能是其应对高山环境较低昆虫访花频率的一种“机会主义”有性生殖策略(侯勤正等, 2009 ), 是植物对高山环境有效适应的结果. ...
... )有关.对于需要传粉者才能完成授粉的植物而言, 花寿命的延长不仅可以增加花被传粉者拜访的概率(张志强和李庆军, 2009 ), 也会增加自身花粉的移出和外来花粉落置的机会(彭德力等, 2012 ), 同时有利于获得更多的交配机会, 保证繁殖成功(Teixido et al ., 2011 ).可见, 全缘叶绿绒蒿花寿命的延长可能是其应对高山环境较低昆虫访花频率的一种“机会主义”有性生殖策略(侯勤正等, 2009 ), 是植物对高山环境有效适应的结果. ...
海南凤仙花不同海拔种群的传粉生物学
2
2014
... 海拔带来的环境差异会使各类昆虫的活动受到限制或促进, 导致不同海拔地区访花者种类和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b ).在高山植物群落中, 鸟类和蝴蝶的传粉作用逐渐减弱, 熊蜂和蝇类成为主要的传粉昆虫(何亚平和刘建全, 2004 ; Duan et al ., 2007 ).对毛茛科植物Ranunculus acris 的研究发现, 低海拔地区昆虫的访花频率是高海拔地区的3倍(Totland, 2001 ).对凤仙花属(Impatiens )植物的研究发现, 高海拔地区和低海拔地区的传粉昆虫种类存在差异, 熊蜂在高海拔地区的花粉传递中扮演着重要的角色(Tian et al ., 2004 ; 肖乐希和刘克明, 2009 ; 肖乐希, 2009 ; 毛志斌等, 2011 ).对露蕊乌头(Aconitum gymnandrum )的研究表明, 低海拔种群的熊蜂种类(3种)多于高海拔种群(2种), 且熊蜂在低海拔种群的总访花频率远远高于高海拔种群(张挺峰等, 2006 ).对横断山区大钟花(Megacodon stylophorus )的研究亦得到相似结果(Meng et al ., 2012 ).对海南凤仙花(Impatiens hainanensis )的传粉生物学研究亦发现, 高海拔种群的传粉者以绿条无垫蜂(Amegilla zonata )为主, 低、中海拔种群以黄黑无垫蜂(Amegilla leptocoma )为主, 且黄黑无垫蜂在低、中海拔种群的访问频率远高于高海拔种群(钟云芳等, 2014 ). ...
... 高山环境下, 由于传粉者的不足使得大多数植物存在花粉限制(段元文等, 2007b ; 司庆文等, 2010 ; 钟云芳等, 2014 ).面对传粉者缺乏的选择压力, 植物会采取多样的策略提高对传粉昆虫的吸引力, 如提高花展示(邵剑文等, 2008 ; 彭德力等, 2012 )、提供生殖场所(Fishman & Hadany, 2013 )等, 随着海拔升高, 一些植物通过延长花寿命获得交配的机会(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 ); 而低温环境下, 一些植物则通过提高花序内或花内温度来达到吸引传粉昆虫的目的(Stanton & Galen, 1989 ).如水母雪兔子(Saussurea medusa )通过花序温度的升高使熊蜂在花序上的停留时间超过1 min, 这在一定程度上对传粉者起到了吸引作用(Tsukaya, 2002 ).对冬季开花的毛茛科铁筷子属植物Helleborus foetidus 和Helleborus bocconei 的研究亦发现, 花内温度的升高使传粉者更加活跃(Vesprini & Pacini, 2010 ).同时, 一些植物往往会进化出各异的自交机制, 如通过雌雄蕊位置的变化促进花粉传递以保障成功繁殖.总结已有的研究发现, 植物雌雄蕊位置关系的改变方式呈现出多样化的特征, 如花丝伸长(段元文等, 2007a , 2007b )、花柱伸长(侯勤正等, 2009 )、花柱卷曲(孙杉等, 2010 )等. ...
海南凤仙花不同海拔种群的传粉生物学
2
2014
... 海拔带来的环境差异会使各类昆虫的活动受到限制或促进, 导致不同海拔地区访花者种类和访问比例的差异(Devoto et al ., 2005 ; 段元文等, 2007b ).在高山植物群落中, 鸟类和蝴蝶的传粉作用逐渐减弱, 熊蜂和蝇类成为主要的传粉昆虫(何亚平和刘建全, 2004 ; Duan et al ., 2007 ).对毛茛科植物Ranunculus acris 的研究发现, 低海拔地区昆虫的访花频率是高海拔地区的3倍(Totland, 2001 ).对凤仙花属(Impatiens )植物的研究发现, 高海拔地区和低海拔地区的传粉昆虫种类存在差异, 熊蜂在高海拔地区的花粉传递中扮演着重要的角色(Tian et al ., 2004 ; 肖乐希和刘克明, 2009 ; 肖乐希, 2009 ; 毛志斌等, 2011 ).对露蕊乌头(Aconitum gymnandrum )的研究表明, 低海拔种群的熊蜂种类(3种)多于高海拔种群(2种), 且熊蜂在低海拔种群的总访花频率远远高于高海拔种群(张挺峰等, 2006 ).对横断山区大钟花(Megacodon stylophorus )的研究亦得到相似结果(Meng et al ., 2012 ).对海南凤仙花(Impatiens hainanensis )的传粉生物学研究亦发现, 高海拔种群的传粉者以绿条无垫蜂(Amegilla zonata )为主, 低、中海拔种群以黄黑无垫蜂(Amegilla leptocoma )为主, 且黄黑无垫蜂在低、中海拔种群的访问频率远高于高海拔种群(钟云芳等, 2014 ). ...
... 高山环境下, 由于传粉者的不足使得大多数植物存在花粉限制(段元文等, 2007b ; 司庆文等, 2010 ; 钟云芳等, 2014 ).面对传粉者缺乏的选择压力, 植物会采取多样的策略提高对传粉昆虫的吸引力, 如提高花展示(邵剑文等, 2008 ; 彭德力等, 2012 )、提供生殖场所(Fishman & Hadany, 2013 )等, 随着海拔升高, 一些植物通过延长花寿命获得交配的机会(Blionis & Vokou, 2002 ; Makrodimos et al ., 2008 ); 而低温环境下, 一些植物则通过提高花序内或花内温度来达到吸引传粉昆虫的目的(Stanton & Galen, 1989 ).如水母雪兔子(Saussurea medusa )通过花序温度的升高使熊蜂在花序上的停留时间超过1 min, 这在一定程度上对传粉者起到了吸引作用(Tsukaya, 2002 ).对冬季开花的毛茛科铁筷子属植物Helleborus foetidus 和Helleborus bocconei 的研究亦发现, 花内温度的升高使传粉者更加活跃(Vesprini & Pacini, 2010 ).同时, 一些植物往往会进化出各异的自交机制, 如通过雌雄蕊位置的变化促进花粉传递以保障成功繁殖.总结已有的研究发现, 植物雌雄蕊位置关系的改变方式呈现出多样化的特征, 如花丝伸长(段元文等, 2007a , 2007b )、花柱伸长(侯勤正等, 2009 )、花柱卷曲(孙杉等, 2010 )等. ...
An experimental design approach using response surface techniques to obtain optimal liquid chromatography and mass spectrometry conditions to determine the alkaloids in Meconops species
1
2009
... 绿绒蒿属(Meconopsis )植物绝大多数分布在以横断山脉-喜马拉雅为中心的高山环境中(庄璇, 1981 ), 作为传统藏药, 目前的研究主要集中在其化学成分及药理活性方面(尚小雅等, 2006 ; Shang et al ., 2006 ; Zhou et al ., 2009 ), 一些学者对本属植物进行了系统分类方面的探讨(Meng et al ., 2006 ; Singhal & Kumar, 2008 ; Yang et al ., 2012 ).作为一类长期适应高山环境的植物, 目前对绿绒蒿属植物的繁殖特征、适应进化等方面尚缺乏系统研究(Dar et al ., 2010 ). ...
苍蝇对杧果的传粉作用及其与温度的关系
1
2006
... 环境温度的测定发现, 各海拔样地7:00环境温度均低于5 ℃, 19:00环境温度均低于3.5 ℃, 这大大低于昆虫的活动温度(贺海霞, 2006 ; 朱建华等, 2006 , 2010 ); 且7:00以前、20:00以后既未见昆虫飞行亦未见新的昆虫进入花内, 此时间段花冠不同程度的闭合, 昆虫蛰伏于花内.由此初步推测, 每日7:00以前和20:00以后没有昆虫访花. ...
苍蝇对杧果的传粉作用及其与温度的关系
1
2006
... 环境温度的测定发现, 各海拔样地7:00环境温度均低于5 ℃, 19:00环境温度均低于3.5 ℃, 这大大低于昆虫的活动温度(贺海霞, 2006 ; 朱建华等, 2006 , 2010 ); 且7:00以前、20:00以后既未见昆虫飞行亦未见新的昆虫进入花内, 此时间段花冠不同程度的闭合, 昆虫蛰伏于花内.由此初步推测, 每日7:00以前和20:00以后没有昆虫访花. ...
龙眼传粉昆虫种类及其传粉活动与温度的关系
1
2010
... 环境温度的测定发现, 各海拔样地7:00环境温度均低于5 ℃, 19:00环境温度均低于3.5 ℃, 这大大低于昆虫的活动温度(贺海霞, 2006 ; 朱建华等, 2006 , 2010 ); 且7:00以前、20:00以后既未见昆虫飞行亦未见新的昆虫进入花内, 此时间段花冠不同程度的闭合, 昆虫蛰伏于花内.由此初步推测, 每日7:00以前和20:00以后没有昆虫访花. ...
龙眼传粉昆虫种类及其传粉活动与温度的关系
1
2010
... 环境温度的测定发现, 各海拔样地7:00环境温度均低于5 ℃, 19:00环境温度均低于3.5 ℃, 这大大低于昆虫的活动温度(贺海霞, 2006 ; 朱建华等, 2006 , 2010 ); 且7:00以前、20:00以后既未见昆虫飞行亦未见新的昆虫进入花内, 此时间段花冠不同程度的闭合, 昆虫蛰伏于花内.由此初步推测, 每日7:00以前和20:00以后没有昆虫访花. ...
 , 操国兴
, 操国兴


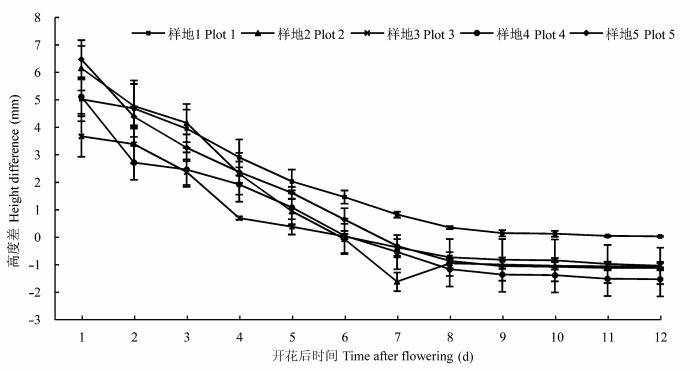
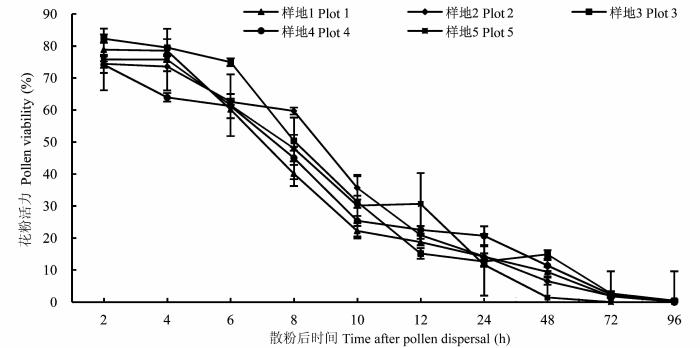
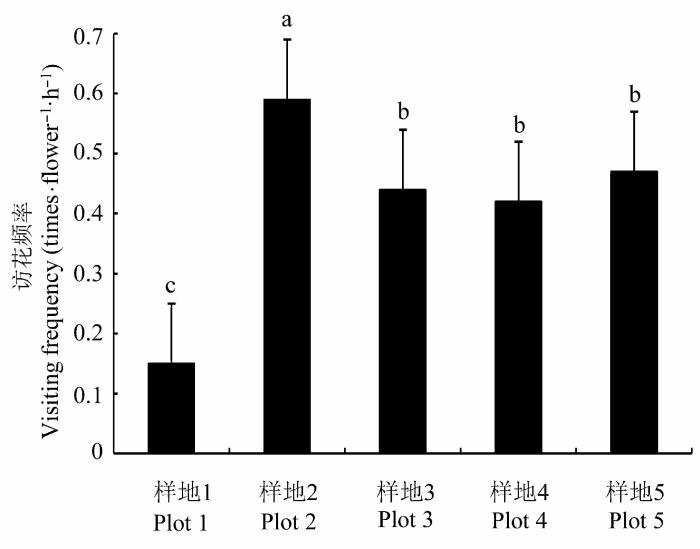
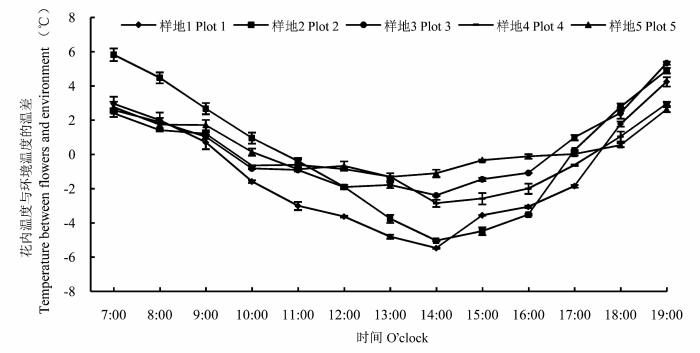
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}