倒木是影响陆地生态系统物质循环与能量流动的重要组分之一(Harmon et al ., 1986 ), 在调控碳(C)库与养分循环等方面发挥着重要的作用(Tinker & Knight, 2000 )。然而, 倒木C和养分的释放受其他关键养分元素(如氮(N)、磷(P)等)含量及其有效性的控制。C:N:P化学计量比以及木质素和纤维素含量特征, 不仅可以表征倒木分解过程中基质的质量变化和养分限制状况(Wardle et al ., 2004 ; Wassen et al ., 2005 ), 而且可以指示物质循环和能量流动的方向。由于树皮、边材和心材基质质量(Schwarze, 2007 ; Bebber et al ., 2011 )和分解时间(Shorohova & Kapitsa, 2014 )的差异, 树皮、边材和心材在不同的分解阶段可能具有不同的质量变化特征。相对于边材和心材, 树皮最先受到环境的影响(Mukhortova, 2012 ; Stanton et al ., 2014 )和外来分解者的侵入(Zhou & Dai, 2012 ), 可能在初期具有分解速率较高和质量变化迅速的特征。 相对于心材, 边材受到物理化学构造的影响, 初期养分含量低(Schwarze, 2007 ), 随着活木保护机制的消失和树皮保护功能的减弱, 分解者和外界环境的影响加剧, 可能在分解中期或后期表现出较快的质量变化特征。心材由于储存了大量能源物质和难分解组分(Schwarze, 2007 ), 可能有较高的C含量和相对较低的养分含量, 但其结构疏松, 易于分解者活动和居住, 其分解过程和质量变化特征更为复杂。已有的研究更加关注于不同林型、不同管理条件及不同演替类型等林分内粗木质残体(CWD)储量和分解速率, 而缺乏关于高寒森林不同分解阶段心材、边材和树皮质量特征的研究, 极大地限制了人们对倒木分解过程的 认识。
川西高寒森林是我国西南林区的主体, 在调节局域气候、涵养水源、保持水土等方面发挥着重要作用(Yang et al ., 2005 )。受频繁的自然灾害和林分自然更新影响, 森林倒木储量巨大(Meyer & Schmidt, 2011 ), 往往受低温限制, 倒木分解缓慢(Kueppers et al ., 2004 ), 各分解阶段的倒木普遍存在(Kueppers et al ., 2004 )。地表倒木作为一个长期存在于森林生态系统的稳定成分, 在地区水源涵养、生物多样性保育、物质循环及水土保持等方面具有不可替代的作用。基于我们研究团队已建立的研究平台和较为成熟的研究方法, 以不同分解阶段的岷江冷杉(Abies faxoniana )倒木为研究对象, 我们研究了倒木树皮、边材和心材等组分在不同分解阶段的质量变化特征, 以期为深入认识倒木分解及其相关的物质循环过程提供一定的理论依据。
1 材料和方法
1.1 研究区概况
研究区位于四川省阿坝州理县毕棚沟自然保护区(102.88°-102.95° E, 31.23°-31.32° N, 海拔2458- 4619 m), 地处青藏高原东缘到四川盆地过渡地带, 四姑娘山北麓, 总面积180 km2 。属丹巴-松潘半湿润气候, 年平均气温2-4 ℃, 最高气温23.7 ℃, 最低气温-18.1℃, 年降水量850 mm, 降水主要分布在生长季节。受季风的影响, 区域内干湿季节差异显著: 干季日照强、降水少、气候寒冷、空气干燥; 湿季日照少、降水多、气候温暖、多云雾。主要植被类型为针阔混交林和针叶林, 乔木主要有岷江冷杉、红桦(Betula albosinensis )、四川红杉(Larix mastersiana )、方枝柏(Sabina saltuaria )等, 林下灌木主要有康定柳(Salix paraplesia )、高山杜鹃(Rhododendron lapponicum )、刺黑珠(别名三颗针) (Berberis sargentiana )、红毛花楸(Sorbus rufopilosa )、沙棘(Hippophae rhamnoides )、扁刺蔷薇(Rosa sweginzowii )等, 草本植物主要有蟹甲草(Cacalia spp.)、冷蕨(Cystopteris fragilis )、薹草属(Carex spp.)和莎草属(Cyperus spp.)等。
1.2 实验方法
1.2.1 样品采集
为避免其他因素的影响, 首先对研究区域内活立木物种组成与储量, 倒木储量及分布特征, 坡度、坡向和土壤性质等进行了调查研究(表1 )。倒木分解阶段的划分按照Rouvinen等(2002) 和闫恩荣等(2005) 提出的CWD 5级腐烂系划分系统(I级: 新鲜, 树木死亡不足1年; II级: 开始腐解, 小刀可刺进几mm; III级: 小刀可刺进2 cm; IV级: 小刀可刺进2-5 cm; V级: 小刀可任意刺穿木质体)。
基于前期调查, 2013年8月在高寒森林海拔3579 m (31.23° N, 102.88° E)处设置了3个100 m × 100 m的典型样地, 每个样地至少间隔500 m, 在样地选取I-V 5个分解阶段的岷江冷杉倒木, 每个分解阶段3株, 统一直径为(30 ± 5) cm, 避免径级差异造成的影响。对于I-III分解阶段的倒木, 截取代表性圆盘, 厚度5 cm, 对于高度分解的倒木则直接取样。采用de Aza等(2011) 的方法将各分解阶段的倒木样品分为心材、边材和树皮, 分别采集各个分解阶段的倒木样品, 去除泥土、石块、根系、苔藓等附生物质, 做好标记后, 装于密封袋内保存, 带回实验室分析, 共计45个样品。
1.2.2 室内分析测定
将采集的样品粉碎, 过60目筛, 于65 ℃条件下烘干至恒重, 存放于干燥环境中待测。测定指标有: 总有机碳含量(重铬酸钾-外加热法(鲁如坤, 1999 )测定)、全氮含量(半微量凯氏定氮法(鲁如坤, 1999 )测定)、全磷含量(钼锑钪比色法(鲁如坤, 1999 )测定)、木质素和纤维素含量(改进的Van Soest中性洗涤纤维(NDF)及酸性洗涤纤维(ADF)方法测定(Rowland & Roberts, 1994 ; Du et al ., 2004 ))。倒木样品按采样份数分别测定, 即每个分解阶段3个重复, 结果取平均值。
1.2.3 数据处理与统计方法
CENTURY模型(Parton et al ., 1987 ): 植物残体易分解比例F m = 0.99 - 0.018L/N, 式中L 为初始木质素含量, N 为初始氮含量。
数据统计与分析采用SPSS 17.0和Excel完成。用单因素方差分析(one-way ANOVA)和最小显著差异法(LSD )检验倒木不同结构组分在不同分解阶段质量变化的差异。用Pearson相关性分析检验倒木的碳格局与养分含量的关系。显著性水平设定为p = 0.05。
2 结果和分析
2.1 C、N、P含量及其生态化学计量比
随着分解程度加深, 倒木各组分C含量显著降低, 下降程度: 树皮>边材>心材(图1 )。其中心材和边材的C含量在I到II分解阶段时随着分解程度加深而缓慢降低, 心材C含量从III分解阶段开始显著降低, 边材C含量从IV分解阶段开始显著降低, 树皮在I到III分解阶段时C含量缓慢增加, IV分解阶段开始显著降低。倒木各组分N含量在各分解阶段的差异不显著, 除III分解阶段的心材外, 均总体表现为随着分解程度加深而增加的趋势, 以边材增加最为显著, V分解阶段时显著升高(p < 0.05), 树皮N含量一直处于最高水平, 相对I到III分解阶段, IV和V分解阶段N含量有较大幅度提升, 但未达到显著性水平。倒木各组分P含量在各分解阶段的表现不同, 树皮初始P含量最高, 且在各分解阶段均处于较高水平, 随着分解程度加深, 大体呈先降后升趋势; 心材初始P含量仅次于树皮, III分解阶段时P含量显著降低(p < 0.05), 而后显著提升(p < 0.05), 与树皮呈现相同的变化趋势; 边材初始P含量最低, III分解阶段时P含量显著升高(p < 0.05), IV和V分解阶段时P含量一直处于较高水平。
图1 岷江冷杉倒木在不同分解阶段(I-V)的C、N、P含量(平均值±标准偏差, n = 3)。不同小写字母表示倒木3个组分(心材、边材、树皮)间差异显著(p < 0.05)。I, 新鲜, 树木死亡不足1年; II, 开始腐解, 小刀可刺进几mm; III, 小刀可刺进2 cm; IV, 小刀可刺进2-5 cm; V, 小刀可任意刺穿木质体。
Fig. 1 Contents of C, N, P in logs of Abies faxoniana at different decay stages (I-V) (mean ± SD, n = 3). Different lowercase letters indicate significant differences among the three log components (i.e. heartwood, sapwood, and bark) (p < 0.05). I, died less than one year prior to sampling, cambium still fresh; II, cambium decayed, a knife blade penetrates a few millimetres; III, a knife blade penetrates less than 2 cm; IV, a knife blade penetrates 2-5 cm; V, a knife blade penetrates all the way.
总体来看, 各分解阶段树皮C:N:P质量比一直低于其他两个组分, 边材在I和II分解阶段时C:N:P质量比值最高, 心材在III到V级分解阶段时C:N:P质量比最高(表2 )。C:N、C:P和N:P值有各自不同的变化特征(表3 ), 具体表现为: 心材初始C:N值最高, 随着分解程度加深表现为先降后升的趋势, III分解阶段时, 心材初始C:N值最小; 树皮初始C:N值显著低于其他组分, 随着分解程度加深呈先升后降的趋势, 各分解阶段C:N值差异显著(p < 0.05); 边材和树皮呈相同的变化趋势, 分别在II和III分解阶段时, 初始C:N值最大, 且初始N含量越高临界值越小。C:P值在各分解阶段间差异均达到显著性水平(p < 0.05), 随着分解程度加深均呈先升后降的趋势, III分解阶段的心材、II分解阶段的边材和树皮C:P值最大, 且初始P含量越高临界值越小; 树皮最大的C:P值为3022, 仍低于其他组分的最小值3453。N:P值在心材和边材各分解阶段间差异显著(p < 0.01), 边材初始N:P值116, 显著高于心材和树皮, 随着分解程度加深呈逐渐降低趋势, 但其值保持在20-30之间; 心材初始N:P值略低于20, 随着分解程度加深表现为先升后降趋势, III分解阶段时N:P值最高; 树皮初始N:P略高于20, 随着分解程度加深表现为逐渐升高趋势。
2.2 木质素和纤维素的特征
随着分解程度加深, 木质素含量显著增加(p < 0.05), V分解阶段时木质素含量最高。心材初始木质素含量显著低于其他组分(p < 0.05), V分解阶段初始木质素含量增幅最大, 其次是边材; 树皮在II分解阶段时木质素含量显著升高, 后期基本保持不变(图2 )。心材和边材纤维素含量随着分解程度加深显著降低(p < 0.05), I分解阶段最高, V分解阶段最低; 树皮纤维素含量大致呈降低趋势, 但降低不显著, 至V分解阶段时纤维素含量降低程度: 心材>边材>树皮, 即初始含量越高, 降低程度越大, 且心材最先表现出显著性降低。I分解阶段时3个组分木质素:纤维素的差异极显著(p < 0.05), V分解阶段时倒木3个组分间木质素:纤维素的差异不显著(p = 0.696); 心材和边材的木质素:纤维素比值随着分解程度加深呈递增趋势, 其中边材增加趋势较为缓慢, III分解阶段开始木质素:纤维素比值大于1; 树皮木质素:纤维素比值一直处于较高水平; 木材结构性组分格局以初始值最小的心材增加幅度最大, 初始值最大的树皮变化幅度最小。
2.3 木质素和纤维素与N和P的相对比例
单因素方差分析(表4 )显示: 仅心材木质素:N、木质素:P、纤维素:N、纤维素:P在不同分解阶段间有显著性差异, 随着分解程度加深, 木质素:N和纤维素:N先降低后升高, 木质素:P和纤维素:P呈相反的变化趋势; 边材木质素:P和纤维素:P随着分解程度加深大致呈逐渐降低的变化趋势, 木质素:N和纤维素:N表现为先降低后升高; 树皮变化较为平稳。不同分解阶段倒木的木质素、纤维素与N和P的相关性分析(表5 )表明, N含量显著影响心材和边材木质纤维素分配比例, 但对木质素和纤维素含量影响不大, P含量对木质素、纤维素分配比例和木质素、纤维素含量的影响不大。
图2 岷江冷杉倒木在不同分解阶段(I-V)木质素、纤维素含量及木质素纤维素变化特征(平均值±标准偏差, n = 3)。不同小写字母表示倒木3个组分(心材、边材、树皮)间差异显著(p < 0.05)。I, 新鲜, 树木死亡不足1年; II, 开始腐解, 小刀可刺进几mm; III, 小刀可刺进2 cm; IV, 小刀可刺进2-5 cm; V, 小刀可任意刺穿木质体。
Fig. 2 Characteristics of lignin, cellulose content and the ratio of lignin to cellulose for various decay stages (I-V) (mean ± SD, n = 3). Different lowercase letters indicate significant differences among the three log components (i.e. heartwood, sapwood, and bark) (p < 0.05). I, died less than one year prior to sampling, cambium still fresh; II, cambium decayed, a knife blade penetrates a few millimetres; III, a knife blade penetrates less than 2 cm; IV, a knife blade penetrates 2-5 cm; V, a knife blade penetrates all the way.
心材、边材和树皮的易分解比例F m 依次为0.66 ± 0.09、0.60 ± 0.10和0.88 ± 0.03, 其中树皮与其他两个组分的差异显著(p < 0.05), 且树皮的易分解组分比例最高, 这与树皮具有最小的C:N:P化学计量比结果一致。
3 讨论
高寒森林生态系统普遍存在各个分解阶段的倒木(Yoon et al ., 2011 )。倒木化学成分和物理结构的差异显著影响其不同解剖结构的质量特征及分解过程。以往研究多将倒木看作是一个整体(Romero et al ., 2005 ; 张利敏和王传宽, 2010 ), 忽略了倒木不同解剖结构的生理代谢途径各异可能会形成不同养分格局, 进而导致不同的养分释放模式。本研究结果表明: 随着分解程度加深, I到III分解阶段的树皮C含量略升高后显著降低, 其余各组分C含量降低, IV分解阶段开始降低程度尤为显著; 倒木各组分N含量在各分解阶段差异不显著, 除III分解阶段的心材外, 总体表现为随着分解程度加深而增加; P含量因结构组分不同表现出“富集—释放”和“释放”模式, 树皮C:N:P化学计量比一直处于较低水平, 易分解组分比例最高; C:N和C:P临界值出现在II和III分解阶段, 且与初始N、P含量成反比。这不仅表明倒木结构和化学组成在不同分解阶段具有显著的差异, 而且在一定程度上表征了N和P对不同分解阶段倒木C释放的限制特征。
林分养分水平因树种、生长情况、环境条件、立地特征和管理方式的不同而表现出一定的差异(Elia & Potvin, 2003 ; Feng et al ., 2013 )。木材不同解剖结构的物理构造和化学组成不同(Schwarze, 2007 ), 化合物和元素含量的差异也较大(Meerts, 2002 ; Augusto et al ., 2008 )。de Aza等(2011) 对3种针叶树种的研究发现, 其心材的C含量显著高于边材, 受木质素含量影响较大。Cowling和Merrill (1966)对裸子植物的研究发现: 同一横截面树皮N含量占比例最高, 边材和内皮的N含量高于心材和外皮, 边材N含量是心材的1.5-2.0倍, 与本项研究结果基本一致。本研究中, 初始C含量由高到低依次为心材、边材和树皮, 变化范围37%-58%, 随着分解程度加深C含量显著降低, 主要是由于纤维素分解, 本研究中纤维素与C含量变化趋势一致, 呈极显著负相关(p < 0.01)。环境中的无脊椎动物在倒木不同分解阶段和不同组分中发挥着重要作用(Harmon et al ., 1986 ), 心材纤维素含量下降稍快于边材可能是由于无脊椎动物在咀嚼或消化木材的同时引入了更多的微生物和捕食者, 而边材分解初期不利于无脊椎动物入侵。随着树皮和心材分解的加深, 边材与环境接触面积增大, 水热条件及通气量得以改善, 促进了微生物在边材的入侵及生长。倒木分解是复杂化合物转化成简单小分子物质、有机物矿化成无机物的过程, 养分释放过程受控于生物和物理因素, 矿化过程主要由分解者的活性调控(Manzoni et al ., 2010 ), 基质养分条件和物理性质影响着微生物的活性(Harmon et al ., 1986 ; Gonzalez-Polo et al ., 2013 )。分解者将环境有机基质中有机化合物和固存的C释放, 以满足自身生命活动需求, 同时将无机态的养分释放到环境中以保持自身化学计量特征的稳定(Cleveland & Liptzin, 2007 )。木材的初始N、P水平较低, 高质量基质(低C:N、C:P)有利于分解者入侵和附生植物定植生长(Benner & Vitousek, 2007 ), 加快倒木分解进程, 但当N、P含量达到一定水平后, 微生物的保护机制将多余的养分释放到环境中, 因此N、P表现出不同释放特征。此外, N、P含量显著影响C:N和C:P值, N、P初始含量分别反作用于C:N和C:P临界值大小。生态化学计量学理论认为大多数情况下C:N > 27或C:P > 186时微生物生长受限制(Sinsabaugh et al ., 2009 ), 本研究中, 心材和边材在整个分解阶段均易受到N、P限制, 树皮分解前期易受到N限制, 整个分解阶段均易受到P限制。认为初始N:P值也能很好地预测N、P动态, 他们的针叶树种分解过程养分释放研究发现临界N:P值约为20。本研究中, 心材和树皮N:P初始值分别为16和27, 表现为N相对含量从流失到富集的模式, 边材N:P初始值为116, 表现为相对含量的富集模式, 本研究结果与Laiho和Prescott (2004) 的研究发现不矛盾, 但仍需进一步验证。此外, 心材中的酚类物质含量高, 微生物底物有效性较低; 边材在树木死亡后保护机制迅速消失, 抑菌物质含量较低(Schwarze, 2007 ), 易于微生物入侵, 养分状况更易受微生物调控。以上结果表明: 不同分解阶段的倒木养分格局差异显著, 基质初始化学计量特征调控并影响后续分解阶段的养分格局; 倒木不同解剖结构的质量特征各异, 进而呈现不同的养分释放模式。
森林中的植物残体主要分为凋落叶和木质残体(Bebber et al ., 2011 )。已有大量研究(Mukhortova, 2012 ; 郭平等, 2013 )证实, 基质N含量显著影响森林凋落物分解过程, 主要通过改变分解速率方式影响C固存。易分解指数F m 通过植物体的初始含量木质素和N在一定程度上可反映植物残体的易分解能力。我们的研究表明树皮的F m 指数显著高于其他组分(如心材、边材), 而且在整个分解过程中C:N:P化学计量比最低, 更易于微生物分解, 上述两组数据得出的结论一致。本研究中, N含量显著影响不同分解阶段倒木心材和边材的C格局(木质素:纤维素)(p < 0.05), 这是因为倒木心材和边材有高C:N初始值, 随着分解阶段加深逐渐增加的N浓度促进了微生物(主要是担子菌类)对木质素的分解(Watkinson et al ., 2006 ), 间接地影响了对纤维素的分解。而富N环境极大地促进了细菌类微生物生长, 分解木质素的微生物失去竞争优势, 故木质素分解受限, 在分解后期木质素:纤维素显著升高, 这表明可能存在一个木质素:N的最优区间, 当环境中木质素:N在此区间内会有较高的木质素分解效率, 过高过低均会抑制木质素的分解。高海拔地区N沉降现象越来越严重, 这可能会抑制倒木分解, 影响C释放过程。Bebber等(2011)研究发现在低N环境中增加可利用N含量可加速倒木分解, 与本研究结果一致。
综上所述, 高寒森林倒木不同组分在不同分解阶段的质量特征各异。在整个分解过程中, 树皮最小的C:N:P化学计量比和最大的易分解比例F m 指示着较高的分解速率(黄耀等, 2003 )。心材和边材的C:N:P化学计量比随着分解程度加深而逐渐降低, 且边材下降速度更快。C:N和C:P的变化特征也表明: 除心材C:N外, 其他组分C:N和C:P均在II或III级分解阶段达到最大值, 受N、P限制最强。N含量显著影响木质素和纤维素的相对分解速率。这些结果表明高寒森林倒木不同组分在同一分解阶段的质量特征各异, 并有不同的分解模式。本研究可为深入探索高寒森林倒木分解及其相关的物质循环提供重要的理论依据。
致谢 感谢四川农业大学生态林业研究所熊莉女士在野外采样工作中给予的帮助。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Augusto L Meredieu C Bert D Trichet P Porté A Bosc A Lagane F Loustau D Pellerin S Danjon F Ranger J Gelpe J 2008 ). Improving models of forest nutrient export with equations that predict the nutrient concentration of tree compartments
. Annals of Forest Science , 65 , 808 -821 .
[本文引用: 1]
[2]
Bebber DP Watkinson SC Boddy L Darrah PR 2011 ). Simulated nitrogen deposition affects wood decomposition by cord-forming fungi
. Oecologia , 167 , 1177 -1184 .
[本文引用: 2]
[3]
Benner JW Vitousek PM 2007 ). Development of a diverse epiphyte community in response to phosphorus fertilization
. Ecology Letters , 10 , 628 -636 .
[本文引用: 1]
[4]
Cleveland CC Liptzin D 2007 ). C:N:P stoichiometry in soil: Is there a ‘‘Redfield ratio’’ for the microbial biomass?
Biogeochemistry , 85 , 235 -252 .
[本文引用: 1]
[5]
Cowling EB Merrill W 1966 ). Nitrogen in wood and its role in wood deterioration
. Canadian Journal of Botany , 44 , 1539 -1554 .
[6]
de Aza CH Turrión MB Pando V Bravo F 2011 ). Carbon in heartwood, sapwood and bark along the stem profile in three Mediterranean Pinus species
. Annals of Forest Science , 68 , 1067 -1076 .
[本文引用: 2]
[7]
Du FY Zhang XY Wang HX 2004 ). Studies on quantitative assay and degradation law of lignocelluloses
. Biotechnology , 14 , 46 -48 .
[本文引用: 1]
[8]
Elia M Potvin C 2003 ). Assessing inter-and intra-specific variation in trunk carbon concentration for 32 neotropical tree species
. Canadian Journal of Forest Research , 33 , 1039 -1045 .
[本文引用: 1]
[9]
Feng SH Cheng SN Yuan ZS Leitch M Xu CB 2013 ). Valorization of bark for chemicals and materials: A review
. Renewable and Sustainable Energy Reviews , 26 , 560 -578 .
[本文引用: 1]
[10]
Gonzalez-Polo M Fernández-Souto A Austin AT 2013 ). Coarse woody debris stimulates soil enzymatic activity and litter decomposition in an old-growth temperate forest of Patagonia, Argentina
. Ecosystems , 16 , 1025 -1038 .
[本文引用: 1]
[11]
Guo P Wang YQ Wang YJ Zhang HL Wang R Liu CX 2013 ). Litterfall mass, nutrient contents, and nutrient release characteristics of typical forests in Jinyun Mountains of China under the background of acid rain
. Chinese Journal of Ecology , 32 , 2339 -2346 .
[本文引用: 1]
(in Chinese with English abstract) [郭平 , 王云琦 , 王玉杰 , 张会兰 , 王冉 , 刘春霞 (2013 ). 酸雨背景下缙云山典型林分凋落物量和营养元素含量及其释放特征
. 生态学杂志 , 32 , 2339 -2346 .]
[本文引用: 1]
[12]
Harmon ME Anderson NH Franklin JF Cline SP Swanson JF Aumen NG Solins P Sedell JR Gregpry SV Lienkaemper GW Lattin JD Cromack K Cummins KW 1986 ). Ecology of coarse woody debris in temperate ecosystem
. Advances in Ecological Research , 15 , 133 -302 .
[本文引用: 3]
[13]
Huang Y Shen Y Zhou M Ma RS 2003 ). Decomposition of plant residue as influenced by its lignin and nitrogen
. Chinese Journal of Plant Ecology , 27 , 183 -188 .
[本文引用: 1]
(in Chinese with English abstract) [黄耀 , 沈雨 , 周密 , 马瑞升 (2003 ). 木质素和氮含量对植物残体分解的影响
. 植物生态学报 , 27 , 183 -188 .]
[本文引用: 1]
[14]
Kueppers LM Southon J Baer P Harte J 2004 ). Dead wood biomass and turnover time, measured by radiocarbon, along a subalpine elevation gradient
. Oecologia , 141 , 641 -651 .
[本文引用: 2]
[15]
Laiho R Prescott CE 2004 ). Decay and nutrient dynamics of coarse woody debris in northern coniferous forests: A synthesis
. Canadian Journal of Forest Research , 34 , 763 -777 .
[本文引用: 1]
[16]
Lu RK 1999). Soil and Agrochemical Analytical Methods . China Agricultural Science and Technology Press , Beijing . 296 -338 . (in Chinese)
[本文引用: 3]
[鲁如坤 (1999). 土壤农化分析方法. 中国农业科技出版社 , 北京 . 296 -338 .]
[本文引用: 3]
[17]
Manzoni S Trofymow JA Jackson RB Porporato A 2010 ). Stoichiometric controls on carbon, nitrogen, and phosphorus dynamics in decomposing litter
. Ecological Monographs , 80 , 89 -106 .
[本文引用: 1]
[18]
Meerts P 2002 ). Mineral nutrient concentrations in sapwood and heartwood: A literature review
. Annals of Forest Science , 59 , 713 -722 .
[本文引用: 1]
[19]
Meyer P Schmidt M 2011 ). Accumulation of dead wood in abandoned beech (Fagus sylvatica L.) forests in northwestern Germany
. Forest Ecology and Management , 261 , 342 -352 .
[本文引用: 1]
[20]
Mukhortova LV 2012 ). Carbon and nutrient release during decomposition of coarse woody debris in forest ecosystems of Central Siberia
. Folia Forestalia Polonica , 52 , 71 -83 .
[本文引用: 2]
[21]
Parton WJ Schimel DS Cole CV Ojima DS 1987 ). Analysis of factors controlling soil organic matter levels in Great Plains grasslands
. Soil Science Society of America Journal , 51 , 1173 -1179 .
[本文引用: 1]
[22]
Romero LM Smith TJ Fourqurean JW 2005 ). Changes in mass and nutrient content of wood during decomposition in a south Florida mangrove forest
. Journal of Ecology , 93 , 618 -631 .
[本文引用: 1]
[23]
Rouvinen S Kuuluvainen T Karjalainen L 2002 ). Coarse woody debris in old Pinus sylvestris dominated forests along a geographic and human impact gradient in boreal Fennoscandia
. Canadian Journal of Forest Research , 32 , 2184 –2200 .
[本文引用: 1]
[24]
Rowland AP Roberts JD 1994 ). Lignin and cellulose fractionation in decomposition studies using Acid-Detergent Fibre methods
. Communications in Soil Science & Plant Analysis , 25 , 269 -277 .
[本文引用: 1]
[25]
Schwarze FWMR 2007 ). Wood decay under the microscope
. Fungal Biology Reviews , 21 , 133 -170 .
[本文引用: 5]
[26]
Shorohova E Kapitsa E 2014 ). Mineralization and fragmentation rates of bark attached to logs in a northern boreal forest
. Forest Ecology and Management , 315 , 185 -190 .
[本文引用: 1]
[27]
Sinsabaugh RL Hill BH Follstad Shah JJ 2009 ). Ecoenzymatic stoichiometry of microbial organic nutrient acquisition in soil and sediment
. Nature , 462 , 795 -799 .
[本文引用: 1]
[28]
Stanton DE Huallpa Chávez J Villegas L Villasante F Armesto J Hedin LO Horn H 2014 ). Epiphytes improve host plant water use by microenvironment modification
. Functional Ecology , 28 , 1274 -1283 .
[本文引用: 1]
[29]
Tinker DB Knight DH 2000 ). Coarse woody debris following fire and logging in Wyoming lodgepole pine forests
. Ecosystems , 3 , 472 -483 .
[本文引用: 1]
[30]
Wardle DA Walker LR Bardgett RD 2004 ). Ecosystem properties and forest decline in contrasting long-term chronosequences
. Science , 305 , 509 -513 .
[本文引用: 1]
[31]
Wassen MJ Venterink HO Lapshina ED Tanneberger F 2005 ). Endangered plants persist under phosphorus limitation
. Nature , 437 , 547 -550 .
[本文引用: 1]
[32]
Watkinson S Bebber DP Darrah P Fricker M Tlalka M Boddy L 2006). The role of wood decay fungi in the carbon and nitrogen dynamics of the forest floor. In: Gadd GM ed. Fungi in Biogeochemical Cycles. Cambridge University Press, Cambridge , UK . 151 -158 .
[本文引用: 1]
[33]
Yan ER Wang XH Huang JJ 2005 ). Concept and classification of coarse woody debris in forest ecosystems
. Acta Ecologica Sinica , 25 , 158 -167 .
[本文引用: 1]
(in Chinese with English abstract) [闫恩荣 , 王希华 , 黄建军 (2005 ). 森林粗死木质残体的概念及其分类
. 生态学报 , 25 , 158 -167 .]
[本文引用: 1]
[34]
Yang WQ Wang KY Kellomäki S Gong HD 2005 ). Litter dynamics of three subalpine forests in western Sichuan
. Pedosphere , 15 , 653 -659 .
[本文引用: 1]
[35]
Yoon TK Noh NJ Kim RH Seo KW Lee SK Yi K Lee IK Lim JH Son Y 2011 ). Mass dynamics of coarse woody debris in an old-growth deciduous forest of Gwangneung, Korea
. Forest Science and Technology , 7 , 145 -150 .
[本文引用: 1]
[36]
Zhang LM Wang CK 2010 ). Carbon and nitrogen release during decomposition of coarse woody debris for eleven temperate tree species in the eastern mountain region of northeast China
. Chinese Journal of Plant Ecology , 34 , 368 -374 .
[本文引用: 1]
(in Chinese with English abstract) [张利敏 , 王传宽 (2010 ). 东北东部山区11种温带树种粗木质残体分解与碳氮释放
. 植物生态学报 , 34 , 368 -374 .]
[本文引用: 1]
[37]
Zhou LW Dai YC 2012 ). Recognizing ecological patterns of wood-decaying polypores on gymnosperm and angiosperm trees in northeast China
. Fungal Ecology , 5 , 230 -235 .
[本文引用: 1]
Improving models of forest nutrient export with equations that predict the nutrient concentration of tree compartments
1
2008
... 林分养分水平因树种、生长情况、环境条件、立地特征和管理方式的不同而表现出一定的差异(Elia & Potvin, 2003 ; Feng et al ., 2013 ).木材不同解剖结构的物理构造和化学组成不同(Schwarze, 2007 ), 化合物和元素含量的差异也较大(Meerts, 2002 ; Augusto et al ., 2008 ).de Aza等(2011) 对3种针叶树种的研究发现, 其心材的C含量显著高于边材, 受木质素含量影响较大.Cowling和Merrill (1966)对裸子植物的研究发现: 同一横截面树皮N含量占比例最高, 边材和内皮的N含量高于心材和外皮, 边材N含量是心材的1.5-2.0倍, 与本项研究结果基本一致.本研究中, 初始C含量由高到低依次为心材、边材和树皮, 变化范围37%-58%, 随着分解程度加深C含量显著降低, 主要是由于纤维素分解, 本研究中纤维素与C含量变化趋势一致, 呈极显著负相关(p < 0.01).环境中的无脊椎动物在倒木不同分解阶段和不同组分中发挥着重要作用(Harmon et al ., 1986 ), 心材纤维素含量下降稍快于边材可能是由于无脊椎动物在咀嚼或消化木材的同时引入了更多的微生物和捕食者, 而边材分解初期不利于无脊椎动物入侵.随着树皮和心材分解的加深, 边材与环境接触面积增大, 水热条件及通气量得以改善, 促进了微生物在边材的入侵及生长.倒木分解是复杂化合物转化成简单小分子物质、有机物矿化成无机物的过程, 养分释放过程受控于生物和物理因素, 矿化过程主要由分解者的活性调控(Manzoni et al ., 2010 ), 基质养分条件和物理性质影响着微生物的活性(Harmon et al ., 1986 ; Gonzalez-Polo et al ., 2013 ).分解者将环境有机基质中有机化合物和固存的C释放, 以满足自身生命活动需求, 同时将无机态的养分释放到环境中以保持自身化学计量特征的稳定(Cleveland & Liptzin, 2007 ).木材的初始N、P水平较低, 高质量基质(低C:N、C:P)有利于分解者入侵和附生植物定植生长(Benner & Vitousek, 2007 ), 加快倒木分解进程, 但当N、P含量达到一定水平后, 微生物的保护机制将多余的养分释放到环境中, 因此N、P表现出不同释放特征.此外, N、P含量显著影响C:N和C:P值, N、P初始含量分别反作用于C:N和C:P临界值大小.生态化学计量学理论认为大多数情况下C:N > 27或C:P > 186时微生物生长受限制(Sinsabaugh et al ., 2009 ), 本研究中, 心材和边材在整个分解阶段均易受到N、P限制, 树皮分解前期易受到N限制, 整个分解阶段均易受到P限制.认为初始N:P值也能很好地预测N、P动态, 他们的针叶树种分解过程养分释放研究发现临界N:P值约为20.本研究中, 心材和树皮N:P初始值分别为16和27, 表现为N相对含量从流失到富集的模式, 边材N:P初始值为116, 表现为相对含量的富集模式, 本研究结果与Laiho和Prescott (2004) 的研究发现不矛盾, 但仍需进一步验证.此外, 心材中的酚类物质含量高, 微生物底物有效性较低; 边材在树木死亡后保护机制迅速消失, 抑菌物质含量较低(Schwarze, 2007 ), 易于微生物入侵, 养分状况更易受微生物调控.以上结果表明: 不同分解阶段的倒木养分格局差异显著, 基质初始化学计量特征调控并影响后续分解阶段的养分格局; 倒木不同解剖结构的质量特征各异, 进而呈现不同的养分释放模式. ...
Simulated nitrogen deposition affects wood decomposition by cord-forming fungi
2
2011
... 倒木是影响陆地生态系统物质循环与能量流动的重要组分之一(Harmon et al ., 1986 ), 在调控碳(C)库与养分循环等方面发挥着重要的作用(Tinker & Knight, 2000 ).然而, 倒木C和养分的释放受其他关键养分元素(如氮(N)、磷(P)等)含量及其有效性的控制.C:N:P化学计量比以及木质素和纤维素含量特征, 不仅可以表征倒木分解过程中基质的质量变化和养分限制状况(Wardle et al ., 2004 ; Wassen et al ., 2005 ), 而且可以指示物质循环和能量流动的方向.由于树皮、边材和心材基质质量(Schwarze, 2007 ; Bebber et al ., 2011 )和分解时间(Shorohova & Kapitsa, 2014 )的差异, 树皮、边材和心材在不同的分解阶段可能具有不同的质量变化特征.相对于边材和心材, 树皮最先受到环境的影响(Mukhortova, 2012 ; Stanton et al ., 2014 )和外来分解者的侵入(Zhou & Dai, 2012 ), 可能在初期具有分解速率较高和质量变化迅速的特征. 相对于心材, 边材受到物理化学构造的影响, 初期养分含量低(Schwarze, 2007 ), 随着活木保护机制的消失和树皮保护功能的减弱, 分解者和外界环境的影响加剧, 可能在分解中期或后期表现出较快的质量变化特征.心材由于储存了大量能源物质和难分解组分(Schwarze, 2007 ), 可能有较高的C含量和相对较低的养分含量, 但其结构疏松, 易于分解者活动和居住, 其分解过程和质量变化特征更为复杂.已有的研究更加关注于不同林型、不同管理条件及不同演替类型等林分内粗木质残体(CWD)储量和分解速率, 而缺乏关于高寒森林不同分解阶段心材、边材和树皮质量特征的研究, 极大地限制了人们对倒木分解过程的 认识. ...
... 森林中的植物残体主要分为凋落叶和木质残体(Bebber et al ., 2011 ).已有大量研究(Mukhortova, 2012 ; 郭平等, 2013 )证实, 基质N含量显著影响森林凋落物分解过程, 主要通过改变分解速率方式影响C固存.易分解指数F m 通过植物体的初始含量木质素和N在一定程度上可反映植物残体的易分解能力.我们的研究表明树皮的F m 指数显著高于其他组分(如心材、边材), 而且在整个分解过程中C:N:P化学计量比最低, 更易于微生物分解, 上述两组数据得出的结论一致.本研究中, N含量显著影响不同分解阶段倒木心材和边材的C格局(木质素:纤维素)(p < 0.05), 这是因为倒木心材和边材有高C:N初始值, 随着分解阶段加深逐渐增加的N浓度促进了微生物(主要是担子菌类)对木质素的分解(Watkinson et al ., 2006 ), 间接地影响了对纤维素的分解.而富N环境极大地促进了细菌类微生物生长, 分解木质素的微生物失去竞争优势, 故木质素分解受限, 在分解后期木质素:纤维素显著升高, 这表明可能存在一个木质素:N的最优区间, 当环境中木质素:N在此区间内会有较高的木质素分解效率, 过高过低均会抑制木质素的分解.高海拔地区N沉降现象越来越严重, 这可能会抑制倒木分解, 影响C释放过程.Bebber等(2011)研究发现在低N环境中增加可利用N含量可加速倒木分解, 与本研究结果一致. ...
Development of a diverse epiphyte community in response to phosphorus fertilization
1
2007
... 林分养分水平因树种、生长情况、环境条件、立地特征和管理方式的不同而表现出一定的差异(Elia & Potvin, 2003 ; Feng et al ., 2013 ).木材不同解剖结构的物理构造和化学组成不同(Schwarze, 2007 ), 化合物和元素含量的差异也较大(Meerts, 2002 ; Augusto et al ., 2008 ).de Aza等(2011) 对3种针叶树种的研究发现, 其心材的C含量显著高于边材, 受木质素含量影响较大.Cowling和Merrill (1966)对裸子植物的研究发现: 同一横截面树皮N含量占比例最高, 边材和内皮的N含量高于心材和外皮, 边材N含量是心材的1.5-2.0倍, 与本项研究结果基本一致.本研究中, 初始C含量由高到低依次为心材、边材和树皮, 变化范围37%-58%, 随着分解程度加深C含量显著降低, 主要是由于纤维素分解, 本研究中纤维素与C含量变化趋势一致, 呈极显著负相关(p < 0.01).环境中的无脊椎动物在倒木不同分解阶段和不同组分中发挥着重要作用(Harmon et al ., 1986 ), 心材纤维素含量下降稍快于边材可能是由于无脊椎动物在咀嚼或消化木材的同时引入了更多的微生物和捕食者, 而边材分解初期不利于无脊椎动物入侵.随着树皮和心材分解的加深, 边材与环境接触面积增大, 水热条件及通气量得以改善, 促进了微生物在边材的入侵及生长.倒木分解是复杂化合物转化成简单小分子物质、有机物矿化成无机物的过程, 养分释放过程受控于生物和物理因素, 矿化过程主要由分解者的活性调控(Manzoni et al ., 2010 ), 基质养分条件和物理性质影响着微生物的活性(Harmon et al ., 1986 ; Gonzalez-Polo et al ., 2013 ).分解者将环境有机基质中有机化合物和固存的C释放, 以满足自身生命活动需求, 同时将无机态的养分释放到环境中以保持自身化学计量特征的稳定(Cleveland & Liptzin, 2007 ).木材的初始N、P水平较低, 高质量基质(低C:N、C:P)有利于分解者入侵和附生植物定植生长(Benner & Vitousek, 2007 ), 加快倒木分解进程, 但当N、P含量达到一定水平后, 微生物的保护机制将多余的养分释放到环境中, 因此N、P表现出不同释放特征.此外, N、P含量显著影响C:N和C:P值, N、P初始含量分别反作用于C:N和C:P临界值大小.生态化学计量学理论认为大多数情况下C:N > 27或C:P > 186时微生物生长受限制(Sinsabaugh et al ., 2009 ), 本研究中, 心材和边材在整个分解阶段均易受到N、P限制, 树皮分解前期易受到N限制, 整个分解阶段均易受到P限制.认为初始N:P值也能很好地预测N、P动态, 他们的针叶树种分解过程养分释放研究发现临界N:P值约为20.本研究中, 心材和树皮N:P初始值分别为16和27, 表现为N相对含量从流失到富集的模式, 边材N:P初始值为116, 表现为相对含量的富集模式, 本研究结果与Laiho和Prescott (2004) 的研究发现不矛盾, 但仍需进一步验证.此外, 心材中的酚类物质含量高, 微生物底物有效性较低; 边材在树木死亡后保护机制迅速消失, 抑菌物质含量较低(Schwarze, 2007 ), 易于微生物入侵, 养分状况更易受微生物调控.以上结果表明: 不同分解阶段的倒木养分格局差异显著, 基质初始化学计量特征调控并影响后续分解阶段的养分格局; 倒木不同解剖结构的质量特征各异, 进而呈现不同的养分释放模式. ...
C:N:P stoichiometry in soil: Is there a ‘‘Redfield ratio’’ for the microbial biomass?
1
2007
... 林分养分水平因树种、生长情况、环境条件、立地特征和管理方式的不同而表现出一定的差异(Elia & Potvin, 2003 ; Feng et al ., 2013 ).木材不同解剖结构的物理构造和化学组成不同(Schwarze, 2007 ), 化合物和元素含量的差异也较大(Meerts, 2002 ; Augusto et al ., 2008 ).de Aza等(2011) 对3种针叶树种的研究发现, 其心材的C含量显著高于边材, 受木质素含量影响较大.Cowling和Merrill (1966)对裸子植物的研究发现: 同一横截面树皮N含量占比例最高, 边材和内皮的N含量高于心材和外皮, 边材N含量是心材的1.5-2.0倍, 与本项研究结果基本一致.本研究中, 初始C含量由高到低依次为心材、边材和树皮, 变化范围37%-58%, 随着分解程度加深C含量显著降低, 主要是由于纤维素分解, 本研究中纤维素与C含量变化趋势一致, 呈极显著负相关(p < 0.01).环境中的无脊椎动物在倒木不同分解阶段和不同组分中发挥着重要作用(Harmon et al ., 1986 ), 心材纤维素含量下降稍快于边材可能是由于无脊椎动物在咀嚼或消化木材的同时引入了更多的微生物和捕食者, 而边材分解初期不利于无脊椎动物入侵.随着树皮和心材分解的加深, 边材与环境接触面积增大, 水热条件及通气量得以改善, 促进了微生物在边材的入侵及生长.倒木分解是复杂化合物转化成简单小分子物质、有机物矿化成无机物的过程, 养分释放过程受控于生物和物理因素, 矿化过程主要由分解者的活性调控(Manzoni et al ., 2010 ), 基质养分条件和物理性质影响着微生物的活性(Harmon et al ., 1986 ; Gonzalez-Polo et al ., 2013 ).分解者将环境有机基质中有机化合物和固存的C释放, 以满足自身生命活动需求, 同时将无机态的养分释放到环境中以保持自身化学计量特征的稳定(Cleveland & Liptzin, 2007 ).木材的初始N、P水平较低, 高质量基质(低C:N、C:P)有利于分解者入侵和附生植物定植生长(Benner & Vitousek, 2007 ), 加快倒木分解进程, 但当N、P含量达到一定水平后, 微生物的保护机制将多余的养分释放到环境中, 因此N、P表现出不同释放特征.此外, N、P含量显著影响C:N和C:P值, N、P初始含量分别反作用于C:N和C:P临界值大小.生态化学计量学理论认为大多数情况下C:N > 27或C:P > 186时微生物生长受限制(Sinsabaugh et al ., 2009 ), 本研究中, 心材和边材在整个分解阶段均易受到N、P限制, 树皮分解前期易受到N限制, 整个分解阶段均易受到P限制.认为初始N:P值也能很好地预测N、P动态, 他们的针叶树种分解过程养分释放研究发现临界N:P值约为20.本研究中, 心材和树皮N:P初始值分别为16和27, 表现为N相对含量从流失到富集的模式, 边材N:P初始值为116, 表现为相对含量的富集模式, 本研究结果与Laiho和Prescott (2004) 的研究发现不矛盾, 但仍需进一步验证.此外, 心材中的酚类物质含量高, 微生物底物有效性较低; 边材在树木死亡后保护机制迅速消失, 抑菌物质含量较低(Schwarze, 2007 ), 易于微生物入侵, 养分状况更易受微生物调控.以上结果表明: 不同分解阶段的倒木养分格局差异显著, 基质初始化学计量特征调控并影响后续分解阶段的养分格局; 倒木不同解剖结构的质量特征各异, 进而呈现不同的养分释放模式. ...
Nitrogen in wood and its role in wood deterioration
0
1966
Carbon in heartwood, sapwood and bark along the stem profile in three Mediterranean Pinus species
2
2011
... 基于前期调查, 2013年8月在高寒森林海拔3579 m (31.23° N, 102.88° E)处设置了3个100 m × 100 m的典型样地, 每个样地至少间隔500 m, 在样地选取I-V 5个分解阶段的岷江冷杉倒木, 每个分解阶段3株, 统一直径为(30 ± 5) cm, 避免径级差异造成的影响.对于I-III分解阶段的倒木, 截取代表性圆盘, 厚度5 cm, 对于高度分解的倒木则直接取样.采用de Aza等(2011) 的方法将各分解阶段的倒木样品分为心材、边材和树皮, 分别采集各个分解阶段的倒木样品, 去除泥土、石块、根系、苔藓等附生物质, 做好标记后, 装于密封袋内保存, 带回实验室分析, 共计45个样品. ...
... 林分养分水平因树种、生长情况、环境条件、立地特征和管理方式的不同而表现出一定的差异(Elia & Potvin, 2003 ; Feng et al ., 2013 ).木材不同解剖结构的物理构造和化学组成不同(Schwarze, 2007 ), 化合物和元素含量的差异也较大(Meerts, 2002 ; Augusto et al ., 2008 ).de Aza等(2011) 对3种针叶树种的研究发现, 其心材的C含量显著高于边材, 受木质素含量影响较大.Cowling和Merrill (1966)对裸子植物的研究发现: 同一横截面树皮N含量占比例最高, 边材和内皮的N含量高于心材和外皮, 边材N含量是心材的1.5-2.0倍, 与本项研究结果基本一致.本研究中, 初始C含量由高到低依次为心材、边材和树皮, 变化范围37%-58%, 随着分解程度加深C含量显著降低, 主要是由于纤维素分解, 本研究中纤维素与C含量变化趋势一致, 呈极显著负相关(p < 0.01).环境中的无脊椎动物在倒木不同分解阶段和不同组分中发挥着重要作用(Harmon et al ., 1986 ), 心材纤维素含量下降稍快于边材可能是由于无脊椎动物在咀嚼或消化木材的同时引入了更多的微生物和捕食者, 而边材分解初期不利于无脊椎动物入侵.随着树皮和心材分解的加深, 边材与环境接触面积增大, 水热条件及通气量得以改善, 促进了微生物在边材的入侵及生长.倒木分解是复杂化合物转化成简单小分子物质、有机物矿化成无机物的过程, 养分释放过程受控于生物和物理因素, 矿化过程主要由分解者的活性调控(Manzoni et al ., 2010 ), 基质养分条件和物理性质影响着微生物的活性(Harmon et al ., 1986 ; Gonzalez-Polo et al ., 2013 ).分解者将环境有机基质中有机化合物和固存的C释放, 以满足自身生命活动需求, 同时将无机态的养分释放到环境中以保持自身化学计量特征的稳定(Cleveland & Liptzin, 2007 ).木材的初始N、P水平较低, 高质量基质(低C:N、C:P)有利于分解者入侵和附生植物定植生长(Benner & Vitousek, 2007 ), 加快倒木分解进程, 但当N、P含量达到一定水平后, 微生物的保护机制将多余的养分释放到环境中, 因此N、P表现出不同释放特征.此外, N、P含量显著影响C:N和C:P值, N、P初始含量分别反作用于C:N和C:P临界值大小.生态化学计量学理论认为大多数情况下C:N > 27或C:P > 186时微生物生长受限制(Sinsabaugh et al ., 2009 ), 本研究中, 心材和边材在整个分解阶段均易受到N、P限制, 树皮分解前期易受到N限制, 整个分解阶段均易受到P限制.认为初始N:P值也能很好地预测N、P动态, 他们的针叶树种分解过程养分释放研究发现临界N:P值约为20.本研究中, 心材和树皮N:P初始值分别为16和27, 表现为N相对含量从流失到富集的模式, 边材N:P初始值为116, 表现为相对含量的富集模式, 本研究结果与Laiho和Prescott (2004) 的研究发现不矛盾, 但仍需进一步验证.此外, 心材中的酚类物质含量高, 微生物底物有效性较低; 边材在树木死亡后保护机制迅速消失, 抑菌物质含量较低(Schwarze, 2007 ), 易于微生物入侵, 养分状况更易受微生物调控.以上结果表明: 不同分解阶段的倒木养分格局差异显著, 基质初始化学计量特征调控并影响后续分解阶段的养分格局; 倒木不同解剖结构的质量特征各异, 进而呈现不同的养分释放模式. ...
Studies on quantitative assay and degradation law of lignocelluloses
1
2004
... 将采集的样品粉碎, 过60目筛, 于65 ℃条件下烘干至恒重, 存放于干燥环境中待测.测定指标有: 总有机碳含量(重铬酸钾-外加热法(鲁如坤, 1999 )测定)、全氮含量(半微量凯氏定氮法(鲁如坤, 1999 )测定)、全磷含量(钼锑钪比色法(鲁如坤, 1999 )测定)、木质素和纤维素含量(改进的Van Soest中性洗涤纤维(NDF)及酸性洗涤纤维(ADF)方法测定(Rowland & Roberts, 1994 ; Du et al ., 2004 )).倒木样品按采样份数分别测定, 即每个分解阶段3个重复, 结果取平均值. ...
Assessing inter-and intra-specific variation in trunk carbon concentration for 32 neotropical tree species
1
2003
... 林分养分水平因树种、生长情况、环境条件、立地特征和管理方式的不同而表现出一定的差异(Elia & Potvin, 2003 ; Feng et al ., 2013 ).木材不同解剖结构的物理构造和化学组成不同(Schwarze, 2007 ), 化合物和元素含量的差异也较大(Meerts, 2002 ; Augusto et al ., 2008 ).de Aza等(2011) 对3种针叶树种的研究发现, 其心材的C含量显著高于边材, 受木质素含量影响较大.Cowling和Merrill (1966)对裸子植物的研究发现: 同一横截面树皮N含量占比例最高, 边材和内皮的N含量高于心材和外皮, 边材N含量是心材的1.5-2.0倍, 与本项研究结果基本一致.本研究中, 初始C含量由高到低依次为心材、边材和树皮, 变化范围37%-58%, 随着分解程度加深C含量显著降低, 主要是由于纤维素分解, 本研究中纤维素与C含量变化趋势一致, 呈极显著负相关(p < 0.01).环境中的无脊椎动物在倒木不同分解阶段和不同组分中发挥着重要作用(Harmon et al ., 1986 ), 心材纤维素含量下降稍快于边材可能是由于无脊椎动物在咀嚼或消化木材的同时引入了更多的微生物和捕食者, 而边材分解初期不利于无脊椎动物入侵.随着树皮和心材分解的加深, 边材与环境接触面积增大, 水热条件及通气量得以改善, 促进了微生物在边材的入侵及生长.倒木分解是复杂化合物转化成简单小分子物质、有机物矿化成无机物的过程, 养分释放过程受控于生物和物理因素, 矿化过程主要由分解者的活性调控(Manzoni et al ., 2010 ), 基质养分条件和物理性质影响着微生物的活性(Harmon et al ., 1986 ; Gonzalez-Polo et al ., 2013 ).分解者将环境有机基质中有机化合物和固存的C释放, 以满足自身生命活动需求, 同时将无机态的养分释放到环境中以保持自身化学计量特征的稳定(Cleveland & Liptzin, 2007 ).木材的初始N、P水平较低, 高质量基质(低C:N、C:P)有利于分解者入侵和附生植物定植生长(Benner & Vitousek, 2007 ), 加快倒木分解进程, 但当N、P含量达到一定水平后, 微生物的保护机制将多余的养分释放到环境中, 因此N、P表现出不同释放特征.此外, N、P含量显著影响C:N和C:P值, N、P初始含量分别反作用于C:N和C:P临界值大小.生态化学计量学理论认为大多数情况下C:N > 27或C:P > 186时微生物生长受限制(Sinsabaugh et al ., 2009 ), 本研究中, 心材和边材在整个分解阶段均易受到N、P限制, 树皮分解前期易受到N限制, 整个分解阶段均易受到P限制.认为初始N:P值也能很好地预测N、P动态, 他们的针叶树种分解过程养分释放研究发现临界N:P值约为20.本研究中, 心材和树皮N:P初始值分别为16和27, 表现为N相对含量从流失到富集的模式, 边材N:P初始值为116, 表现为相对含量的富集模式, 本研究结果与Laiho和Prescott (2004) 的研究发现不矛盾, 但仍需进一步验证.此外, 心材中的酚类物质含量高, 微生物底物有效性较低; 边材在树木死亡后保护机制迅速消失, 抑菌物质含量较低(Schwarze, 2007 ), 易于微生物入侵, 养分状况更易受微生物调控.以上结果表明: 不同分解阶段的倒木养分格局差异显著, 基质初始化学计量特征调控并影响后续分解阶段的养分格局; 倒木不同解剖结构的质量特征各异, 进而呈现不同的养分释放模式. ...
Valorization of bark for chemicals and materials: A review
1
2013
... 林分养分水平因树种、生长情况、环境条件、立地特征和管理方式的不同而表现出一定的差异(Elia & Potvin, 2003 ; Feng et al ., 2013 ).木材不同解剖结构的物理构造和化学组成不同(Schwarze, 2007 ), 化合物和元素含量的差异也较大(Meerts, 2002 ; Augusto et al ., 2008 ).de Aza等(2011) 对3种针叶树种的研究发现, 其心材的C含量显著高于边材, 受木质素含量影响较大.Cowling和Merrill (1966)对裸子植物的研究发现: 同一横截面树皮N含量占比例最高, 边材和内皮的N含量高于心材和外皮, 边材N含量是心材的1.5-2.0倍, 与本项研究结果基本一致.本研究中, 初始C含量由高到低依次为心材、边材和树皮, 变化范围37%-58%, 随着分解程度加深C含量显著降低, 主要是由于纤维素分解, 本研究中纤维素与C含量变化趋势一致, 呈极显著负相关(p < 0.01).环境中的无脊椎动物在倒木不同分解阶段和不同组分中发挥着重要作用(Harmon et al ., 1986 ), 心材纤维素含量下降稍快于边材可能是由于无脊椎动物在咀嚼或消化木材的同时引入了更多的微生物和捕食者, 而边材分解初期不利于无脊椎动物入侵.随着树皮和心材分解的加深, 边材与环境接触面积增大, 水热条件及通气量得以改善, 促进了微生物在边材的入侵及生长.倒木分解是复杂化合物转化成简单小分子物质、有机物矿化成无机物的过程, 养分释放过程受控于生物和物理因素, 矿化过程主要由分解者的活性调控(Manzoni et al ., 2010 ), 基质养分条件和物理性质影响着微生物的活性(Harmon et al ., 1986 ; Gonzalez-Polo et al ., 2013 ).分解者将环境有机基质中有机化合物和固存的C释放, 以满足自身生命活动需求, 同时将无机态的养分释放到环境中以保持自身化学计量特征的稳定(Cleveland & Liptzin, 2007 ).木材的初始N、P水平较低, 高质量基质(低C:N、C:P)有利于分解者入侵和附生植物定植生长(Benner & Vitousek, 2007 ), 加快倒木分解进程, 但当N、P含量达到一定水平后, 微生物的保护机制将多余的养分释放到环境中, 因此N、P表现出不同释放特征.此外, N、P含量显著影响C:N和C:P值, N、P初始含量分别反作用于C:N和C:P临界值大小.生态化学计量学理论认为大多数情况下C:N > 27或C:P > 186时微生物生长受限制(Sinsabaugh et al ., 2009 ), 本研究中, 心材和边材在整个分解阶段均易受到N、P限制, 树皮分解前期易受到N限制, 整个分解阶段均易受到P限制.认为初始N:P值也能很好地预测N、P动态, 他们的针叶树种分解过程养分释放研究发现临界N:P值约为20.本研究中, 心材和树皮N:P初始值分别为16和27, 表现为N相对含量从流失到富集的模式, 边材N:P初始值为116, 表现为相对含量的富集模式, 本研究结果与Laiho和Prescott (2004) 的研究发现不矛盾, 但仍需进一步验证.此外, 心材中的酚类物质含量高, 微生物底物有效性较低; 边材在树木死亡后保护机制迅速消失, 抑菌物质含量较低(Schwarze, 2007 ), 易于微生物入侵, 养分状况更易受微生物调控.以上结果表明: 不同分解阶段的倒木养分格局差异显著, 基质初始化学计量特征调控并影响后续分解阶段的养分格局; 倒木不同解剖结构的质量特征各异, 进而呈现不同的养分释放模式. ...
Coarse woody debris stimulates soil enzymatic activity and litter decomposition in an old-growth temperate forest of Patagonia, Argentina
1
2013
... 林分养分水平因树种、生长情况、环境条件、立地特征和管理方式的不同而表现出一定的差异(Elia & Potvin, 2003 ; Feng et al ., 2013 ).木材不同解剖结构的物理构造和化学组成不同(Schwarze, 2007 ), 化合物和元素含量的差异也较大(Meerts, 2002 ; Augusto et al ., 2008 ).de Aza等(2011) 对3种针叶树种的研究发现, 其心材的C含量显著高于边材, 受木质素含量影响较大.Cowling和Merrill (1966)对裸子植物的研究发现: 同一横截面树皮N含量占比例最高, 边材和内皮的N含量高于心材和外皮, 边材N含量是心材的1.5-2.0倍, 与本项研究结果基本一致.本研究中, 初始C含量由高到低依次为心材、边材和树皮, 变化范围37%-58%, 随着分解程度加深C含量显著降低, 主要是由于纤维素分解, 本研究中纤维素与C含量变化趋势一致, 呈极显著负相关(p < 0.01).环境中的无脊椎动物在倒木不同分解阶段和不同组分中发挥着重要作用(Harmon et al ., 1986 ), 心材纤维素含量下降稍快于边材可能是由于无脊椎动物在咀嚼或消化木材的同时引入了更多的微生物和捕食者, 而边材分解初期不利于无脊椎动物入侵.随着树皮和心材分解的加深, 边材与环境接触面积增大, 水热条件及通气量得以改善, 促进了微生物在边材的入侵及生长.倒木分解是复杂化合物转化成简单小分子物质、有机物矿化成无机物的过程, 养分释放过程受控于生物和物理因素, 矿化过程主要由分解者的活性调控(Manzoni et al ., 2010 ), 基质养分条件和物理性质影响着微生物的活性(Harmon et al ., 1986 ; Gonzalez-Polo et al ., 2013 ).分解者将环境有机基质中有机化合物和固存的C释放, 以满足自身生命活动需求, 同时将无机态的养分释放到环境中以保持自身化学计量特征的稳定(Cleveland & Liptzin, 2007 ).木材的初始N、P水平较低, 高质量基质(低C:N、C:P)有利于分解者入侵和附生植物定植生长(Benner & Vitousek, 2007 ), 加快倒木分解进程, 但当N、P含量达到一定水平后, 微生物的保护机制将多余的养分释放到环境中, 因此N、P表现出不同释放特征.此外, N、P含量显著影响C:N和C:P值, N、P初始含量分别反作用于C:N和C:P临界值大小.生态化学计量学理论认为大多数情况下C:N > 27或C:P > 186时微生物生长受限制(Sinsabaugh et al ., 2009 ), 本研究中, 心材和边材在整个分解阶段均易受到N、P限制, 树皮分解前期易受到N限制, 整个分解阶段均易受到P限制.认为初始N:P值也能很好地预测N、P动态, 他们的针叶树种分解过程养分释放研究发现临界N:P值约为20.本研究中, 心材和树皮N:P初始值分别为16和27, 表现为N相对含量从流失到富集的模式, 边材N:P初始值为116, 表现为相对含量的富集模式, 本研究结果与Laiho和Prescott (2004) 的研究发现不矛盾, 但仍需进一步验证.此外, 心材中的酚类物质含量高, 微生物底物有效性较低; 边材在树木死亡后保护机制迅速消失, 抑菌物质含量较低(Schwarze, 2007 ), 易于微生物入侵, 养分状况更易受微生物调控.以上结果表明: 不同分解阶段的倒木养分格局差异显著, 基质初始化学计量特征调控并影响后续分解阶段的养分格局; 倒木不同解剖结构的质量特征各异, 进而呈现不同的养分释放模式. ...
酸雨背景下缙云山典型林分凋落物量和营养元素含量及其释放特征
1
2013
... 森林中的植物残体主要分为凋落叶和木质残体(Bebber et al ., 2011 ).已有大量研究(Mukhortova, 2012 ; 郭平等, 2013 )证实, 基质N含量显著影响森林凋落物分解过程, 主要通过改变分解速率方式影响C固存.易分解指数F m 通过植物体的初始含量木质素和N在一定程度上可反映植物残体的易分解能力.我们的研究表明树皮的F m 指数显著高于其他组分(如心材、边材), 而且在整个分解过程中C:N:P化学计量比最低, 更易于微生物分解, 上述两组数据得出的结论一致.本研究中, N含量显著影响不同分解阶段倒木心材和边材的C格局(木质素:纤维素)(p < 0.05), 这是因为倒木心材和边材有高C:N初始值, 随着分解阶段加深逐渐增加的N浓度促进了微生物(主要是担子菌类)对木质素的分解(Watkinson et al ., 2006 ), 间接地影响了对纤维素的分解.而富N环境极大地促进了细菌类微生物生长, 分解木质素的微生物失去竞争优势, 故木质素分解受限, 在分解后期木质素:纤维素显著升高, 这表明可能存在一个木质素:N的最优区间, 当环境中木质素:N在此区间内会有较高的木质素分解效率, 过高过低均会抑制木质素的分解.高海拔地区N沉降现象越来越严重, 这可能会抑制倒木分解, 影响C释放过程.Bebber等(2011)研究发现在低N环境中增加可利用N含量可加速倒木分解, 与本研究结果一致. ...
酸雨背景下缙云山典型林分凋落物量和营养元素含量及其释放特征
1
2013
... 森林中的植物残体主要分为凋落叶和木质残体(Bebber et al ., 2011 ).已有大量研究(Mukhortova, 2012 ; 郭平等, 2013 )证实, 基质N含量显著影响森林凋落物分解过程, 主要通过改变分解速率方式影响C固存.易分解指数F m 通过植物体的初始含量木质素和N在一定程度上可反映植物残体的易分解能力.我们的研究表明树皮的F m 指数显著高于其他组分(如心材、边材), 而且在整个分解过程中C:N:P化学计量比最低, 更易于微生物分解, 上述两组数据得出的结论一致.本研究中, N含量显著影响不同分解阶段倒木心材和边材的C格局(木质素:纤维素)(p < 0.05), 这是因为倒木心材和边材有高C:N初始值, 随着分解阶段加深逐渐增加的N浓度促进了微生物(主要是担子菌类)对木质素的分解(Watkinson et al ., 2006 ), 间接地影响了对纤维素的分解.而富N环境极大地促进了细菌类微生物生长, 分解木质素的微生物失去竞争优势, 故木质素分解受限, 在分解后期木质素:纤维素显著升高, 这表明可能存在一个木质素:N的最优区间, 当环境中木质素:N在此区间内会有较高的木质素分解效率, 过高过低均会抑制木质素的分解.高海拔地区N沉降现象越来越严重, 这可能会抑制倒木分解, 影响C释放过程.Bebber等(2011)研究发现在低N环境中增加可利用N含量可加速倒木分解, 与本研究结果一致. ...
Ecology of coarse woody debris in temperate ecosystem
3
1986
... 倒木是影响陆地生态系统物质循环与能量流动的重要组分之一(Harmon et al ., 1986 ), 在调控碳(C)库与养分循环等方面发挥着重要的作用(Tinker & Knight, 2000 ).然而, 倒木C和养分的释放受其他关键养分元素(如氮(N)、磷(P)等)含量及其有效性的控制.C:N:P化学计量比以及木质素和纤维素含量特征, 不仅可以表征倒木分解过程中基质的质量变化和养分限制状况(Wardle et al ., 2004 ; Wassen et al ., 2005 ), 而且可以指示物质循环和能量流动的方向.由于树皮、边材和心材基质质量(Schwarze, 2007 ; Bebber et al ., 2011 )和分解时间(Shorohova & Kapitsa, 2014 )的差异, 树皮、边材和心材在不同的分解阶段可能具有不同的质量变化特征.相对于边材和心材, 树皮最先受到环境的影响(Mukhortova, 2012 ; Stanton et al ., 2014 )和外来分解者的侵入(Zhou & Dai, 2012 ), 可能在初期具有分解速率较高和质量变化迅速的特征. 相对于心材, 边材受到物理化学构造的影响, 初期养分含量低(Schwarze, 2007 ), 随着活木保护机制的消失和树皮保护功能的减弱, 分解者和外界环境的影响加剧, 可能在分解中期或后期表现出较快的质量变化特征.心材由于储存了大量能源物质和难分解组分(Schwarze, 2007 ), 可能有较高的C含量和相对较低的养分含量, 但其结构疏松, 易于分解者活动和居住, 其分解过程和质量变化特征更为复杂.已有的研究更加关注于不同林型、不同管理条件及不同演替类型等林分内粗木质残体(CWD)储量和分解速率, 而缺乏关于高寒森林不同分解阶段心材、边材和树皮质量特征的研究, 极大地限制了人们对倒木分解过程的 认识. ...
... 林分养分水平因树种、生长情况、环境条件、立地特征和管理方式的不同而表现出一定的差异(Elia & Potvin, 2003 ; Feng et al ., 2013 ).木材不同解剖结构的物理构造和化学组成不同(Schwarze, 2007 ), 化合物和元素含量的差异也较大(Meerts, 2002 ; Augusto et al ., 2008 ).de Aza等(2011) 对3种针叶树种的研究发现, 其心材的C含量显著高于边材, 受木质素含量影响较大.Cowling和Merrill (1966)对裸子植物的研究发现: 同一横截面树皮N含量占比例最高, 边材和内皮的N含量高于心材和外皮, 边材N含量是心材的1.5-2.0倍, 与本项研究结果基本一致.本研究中, 初始C含量由高到低依次为心材、边材和树皮, 变化范围37%-58%, 随着分解程度加深C含量显著降低, 主要是由于纤维素分解, 本研究中纤维素与C含量变化趋势一致, 呈极显著负相关(p < 0.01).环境中的无脊椎动物在倒木不同分解阶段和不同组分中发挥着重要作用(Harmon et al ., 1986 ), 心材纤维素含量下降稍快于边材可能是由于无脊椎动物在咀嚼或消化木材的同时引入了更多的微生物和捕食者, 而边材分解初期不利于无脊椎动物入侵.随着树皮和心材分解的加深, 边材与环境接触面积增大, 水热条件及通气量得以改善, 促进了微生物在边材的入侵及生长.倒木分解是复杂化合物转化成简单小分子物质、有机物矿化成无机物的过程, 养分释放过程受控于生物和物理因素, 矿化过程主要由分解者的活性调控(Manzoni et al ., 2010 ), 基质养分条件和物理性质影响着微生物的活性(Harmon et al ., 1986 ; Gonzalez-Polo et al ., 2013 ).分解者将环境有机基质中有机化合物和固存的C释放, 以满足自身生命活动需求, 同时将无机态的养分释放到环境中以保持自身化学计量特征的稳定(Cleveland & Liptzin, 2007 ).木材的初始N、P水平较低, 高质量基质(低C:N、C:P)有利于分解者入侵和附生植物定植生长(Benner & Vitousek, 2007 ), 加快倒木分解进程, 但当N、P含量达到一定水平后, 微生物的保护机制将多余的养分释放到环境中, 因此N、P表现出不同释放特征.此外, N、P含量显著影响C:N和C:P值, N、P初始含量分别反作用于C:N和C:P临界值大小.生态化学计量学理论认为大多数情况下C:N > 27或C:P > 186时微生物生长受限制(Sinsabaugh et al ., 2009 ), 本研究中, 心材和边材在整个分解阶段均易受到N、P限制, 树皮分解前期易受到N限制, 整个分解阶段均易受到P限制.认为初始N:P值也能很好地预测N、P动态, 他们的针叶树种分解过程养分释放研究发现临界N:P值约为20.本研究中, 心材和树皮N:P初始值分别为16和27, 表现为N相对含量从流失到富集的模式, 边材N:P初始值为116, 表现为相对含量的富集模式, 本研究结果与Laiho和Prescott (2004) 的研究发现不矛盾, 但仍需进一步验证.此外, 心材中的酚类物质含量高, 微生物底物有效性较低; 边材在树木死亡后保护机制迅速消失, 抑菌物质含量较低(Schwarze, 2007 ), 易于微生物入侵, 养分状况更易受微生物调控.以上结果表明: 不同分解阶段的倒木养分格局差异显著, 基质初始化学计量特征调控并影响后续分解阶段的养分格局; 倒木不同解剖结构的质量特征各异, 进而呈现不同的养分释放模式. ...
... ), 基质养分条件和物理性质影响着微生物的活性(Harmon et al ., 1986 ; Gonzalez-Polo et al ., 2013 ).分解者将环境有机基质中有机化合物和固存的C释放, 以满足自身生命活动需求, 同时将无机态的养分释放到环境中以保持自身化学计量特征的稳定(Cleveland & Liptzin, 2007 ).木材的初始N、P水平较低, 高质量基质(低C:N、C:P)有利于分解者入侵和附生植物定植生长(Benner & Vitousek, 2007 ), 加快倒木分解进程, 但当N、P含量达到一定水平后, 微生物的保护机制将多余的养分释放到环境中, 因此N、P表现出不同释放特征.此外, N、P含量显著影响C:N和C:P值, N、P初始含量分别反作用于C:N和C:P临界值大小.生态化学计量学理论认为大多数情况下C:N > 27或C:P > 186时微生物生长受限制(Sinsabaugh et al ., 2009 ), 本研究中, 心材和边材在整个分解阶段均易受到N、P限制, 树皮分解前期易受到N限制, 整个分解阶段均易受到P限制.认为初始N:P值也能很好地预测N、P动态, 他们的针叶树种分解过程养分释放研究发现临界N:P值约为20.本研究中, 心材和树皮N:P初始值分别为16和27, 表现为N相对含量从流失到富集的模式, 边材N:P初始值为116, 表现为相对含量的富集模式, 本研究结果与Laiho和Prescott (2004) 的研究发现不矛盾, 但仍需进一步验证.此外, 心材中的酚类物质含量高, 微生物底物有效性较低; 边材在树木死亡后保护机制迅速消失, 抑菌物质含量较低(Schwarze, 2007 ), 易于微生物入侵, 养分状况更易受微生物调控.以上结果表明: 不同分解阶段的倒木养分格局差异显著, 基质初始化学计量特征调控并影响后续分解阶段的养分格局; 倒木不同解剖结构的质量特征各异, 进而呈现不同的养分释放模式. ...
木质素和氮含量对植物残体分解的影响
1
2003
... 综上所述, 高寒森林倒木不同组分在不同分解阶段的质量特征各异.在整个分解过程中, 树皮最小的C:N:P化学计量比和最大的易分解比例F m 指示着较高的分解速率(黄耀等, 2003 ).心材和边材的C:N:P化学计量比随着分解程度加深而逐渐降低, 且边材下降速度更快.C:N和C:P的变化特征也表明: 除心材C:N外, 其他组分C:N和C:P均在II或III级分解阶段达到最大值, 受N、P限制最强.N含量显著影响木质素和纤维素的相对分解速率.这些结果表明高寒森林倒木不同组分在同一分解阶段的质量特征各异, 并有不同的分解模式.本研究可为深入探索高寒森林倒木分解及其相关的物质循环提供重要的理论依据. ...
木质素和氮含量对植物残体分解的影响
1
2003
... 综上所述, 高寒森林倒木不同组分在不同分解阶段的质量特征各异.在整个分解过程中, 树皮最小的C:N:P化学计量比和最大的易分解比例F m 指示着较高的分解速率(黄耀等, 2003 ).心材和边材的C:N:P化学计量比随着分解程度加深而逐渐降低, 且边材下降速度更快.C:N和C:P的变化特征也表明: 除心材C:N外, 其他组分C:N和C:P均在II或III级分解阶段达到最大值, 受N、P限制最强.N含量显著影响木质素和纤维素的相对分解速率.这些结果表明高寒森林倒木不同组分在同一分解阶段的质量特征各异, 并有不同的分解模式.本研究可为深入探索高寒森林倒木分解及其相关的物质循环提供重要的理论依据. ...
Dead wood biomass and turnover time, measured by radiocarbon, along a subalpine elevation gradient
2
2004
... 川西高寒森林是我国西南林区的主体, 在调节局域气候、涵养水源、保持水土等方面发挥着重要作用(Yang et al ., 2005 ).受频繁的自然灾害和林分自然更新影响, 森林倒木储量巨大(Meyer & Schmidt, 2011 ), 往往受低温限制, 倒木分解缓慢(Kueppers et al ., 2004 ), 各分解阶段的倒木普遍存在(Kueppers et al ., 2004 ).地表倒木作为一个长期存在于森林生态系统的稳定成分, 在地区水源涵养、生物多样性保育、物质循环及水土保持等方面具有不可替代的作用.基于我们研究团队已建立的研究平台和较为成熟的研究方法, 以不同分解阶段的岷江冷杉(Abies faxoniana )倒木为研究对象, 我们研究了倒木树皮、边材和心材等组分在不同分解阶段的质量变化特征, 以期为深入认识倒木分解及其相关的物质循环过程提供一定的理论依据. ...
... ), 各分解阶段的倒木普遍存在(Kueppers et al ., 2004 ).地表倒木作为一个长期存在于森林生态系统的稳定成分, 在地区水源涵养、生物多样性保育、物质循环及水土保持等方面具有不可替代的作用.基于我们研究团队已建立的研究平台和较为成熟的研究方法, 以不同分解阶段的岷江冷杉(Abies faxoniana )倒木为研究对象, 我们研究了倒木树皮、边材和心材等组分在不同分解阶段的质量变化特征, 以期为深入认识倒木分解及其相关的物质循环过程提供一定的理论依据. ...
Decay and nutrient dynamics of coarse woody debris in northern coniferous forests: A synthesis
1
2004
... 林分养分水平因树种、生长情况、环境条件、立地特征和管理方式的不同而表现出一定的差异(Elia & Potvin, 2003 ; Feng et al ., 2013 ).木材不同解剖结构的物理构造和化学组成不同(Schwarze, 2007 ), 化合物和元素含量的差异也较大(Meerts, 2002 ; Augusto et al ., 2008 ).de Aza等(2011) 对3种针叶树种的研究发现, 其心材的C含量显著高于边材, 受木质素含量影响较大.Cowling和Merrill (1966)对裸子植物的研究发现: 同一横截面树皮N含量占比例最高, 边材和内皮的N含量高于心材和外皮, 边材N含量是心材的1.5-2.0倍, 与本项研究结果基本一致.本研究中, 初始C含量由高到低依次为心材、边材和树皮, 变化范围37%-58%, 随着分解程度加深C含量显著降低, 主要是由于纤维素分解, 本研究中纤维素与C含量变化趋势一致, 呈极显著负相关(p < 0.01).环境中的无脊椎动物在倒木不同分解阶段和不同组分中发挥着重要作用(Harmon et al ., 1986 ), 心材纤维素含量下降稍快于边材可能是由于无脊椎动物在咀嚼或消化木材的同时引入了更多的微生物和捕食者, 而边材分解初期不利于无脊椎动物入侵.随着树皮和心材分解的加深, 边材与环境接触面积增大, 水热条件及通气量得以改善, 促进了微生物在边材的入侵及生长.倒木分解是复杂化合物转化成简单小分子物质、有机物矿化成无机物的过程, 养分释放过程受控于生物和物理因素, 矿化过程主要由分解者的活性调控(Manzoni et al ., 2010 ), 基质养分条件和物理性质影响着微生物的活性(Harmon et al ., 1986 ; Gonzalez-Polo et al ., 2013 ).分解者将环境有机基质中有机化合物和固存的C释放, 以满足自身生命活动需求, 同时将无机态的养分释放到环境中以保持自身化学计量特征的稳定(Cleveland & Liptzin, 2007 ).木材的初始N、P水平较低, 高质量基质(低C:N、C:P)有利于分解者入侵和附生植物定植生长(Benner & Vitousek, 2007 ), 加快倒木分解进程, 但当N、P含量达到一定水平后, 微生物的保护机制将多余的养分释放到环境中, 因此N、P表现出不同释放特征.此外, N、P含量显著影响C:N和C:P值, N、P初始含量分别反作用于C:N和C:P临界值大小.生态化学计量学理论认为大多数情况下C:N > 27或C:P > 186时微生物生长受限制(Sinsabaugh et al ., 2009 ), 本研究中, 心材和边材在整个分解阶段均易受到N、P限制, 树皮分解前期易受到N限制, 整个分解阶段均易受到P限制.认为初始N:P值也能很好地预测N、P动态, 他们的针叶树种分解过程养分释放研究发现临界N:P值约为20.本研究中, 心材和树皮N:P初始值分别为16和27, 表现为N相对含量从流失到富集的模式, 边材N:P初始值为116, 表现为相对含量的富集模式, 本研究结果与Laiho和Prescott (2004) 的研究发现不矛盾, 但仍需进一步验证.此外, 心材中的酚类物质含量高, 微生物底物有效性较低; 边材在树木死亡后保护机制迅速消失, 抑菌物质含量较低(Schwarze, 2007 ), 易于微生物入侵, 养分状况更易受微生物调控.以上结果表明: 不同分解阶段的倒木养分格局差异显著, 基质初始化学计量特征调控并影响后续分解阶段的养分格局; 倒木不同解剖结构的质量特征各异, 进而呈现不同的养分释放模式. ...
3
... 将采集的样品粉碎, 过60目筛, 于65 ℃条件下烘干至恒重, 存放于干燥环境中待测.测定指标有: 总有机碳含量(重铬酸钾-外加热法(鲁如坤, 1999 )测定)、全氮含量(半微量凯氏定氮法(鲁如坤, 1999 )测定)、全磷含量(钼锑钪比色法(鲁如坤, 1999 )测定)、木质素和纤维素含量(改进的Van Soest中性洗涤纤维(NDF)及酸性洗涤纤维(ADF)方法测定(Rowland & Roberts, 1994 ; Du et al ., 2004 )).倒木样品按采样份数分别测定, 即每个分解阶段3个重复, 结果取平均值. ...
... )测定)、全氮含量(半微量凯氏定氮法(鲁如坤, 1999 )测定)、全磷含量(钼锑钪比色法(鲁如坤, 1999 )测定)、木质素和纤维素含量(改进的Van Soest中性洗涤纤维(NDF)及酸性洗涤纤维(ADF)方法测定(Rowland & Roberts, 1994 ; Du et al ., 2004 )).倒木样品按采样份数分别测定, 即每个分解阶段3个重复, 结果取平均值. ...
... )测定)、全磷含量(钼锑钪比色法(鲁如坤, 1999 )测定)、木质素和纤维素含量(改进的Van Soest中性洗涤纤维(NDF)及酸性洗涤纤维(ADF)方法测定(Rowland & Roberts, 1994 ; Du et al ., 2004 )).倒木样品按采样份数分别测定, 即每个分解阶段3个重复, 结果取平均值. ...
3
... 将采集的样品粉碎, 过60目筛, 于65 ℃条件下烘干至恒重, 存放于干燥环境中待测.测定指标有: 总有机碳含量(重铬酸钾-外加热法(鲁如坤, 1999 )测定)、全氮含量(半微量凯氏定氮法(鲁如坤, 1999 )测定)、全磷含量(钼锑钪比色法(鲁如坤, 1999 )测定)、木质素和纤维素含量(改进的Van Soest中性洗涤纤维(NDF)及酸性洗涤纤维(ADF)方法测定(Rowland & Roberts, 1994 ; Du et al ., 2004 )).倒木样品按采样份数分别测定, 即每个分解阶段3个重复, 结果取平均值. ...
... )测定)、全氮含量(半微量凯氏定氮法(鲁如坤, 1999 )测定)、全磷含量(钼锑钪比色法(鲁如坤, 1999 )测定)、木质素和纤维素含量(改进的Van Soest中性洗涤纤维(NDF)及酸性洗涤纤维(ADF)方法测定(Rowland & Roberts, 1994 ; Du et al ., 2004 )).倒木样品按采样份数分别测定, 即每个分解阶段3个重复, 结果取平均值. ...
... )测定)、全磷含量(钼锑钪比色法(鲁如坤, 1999 )测定)、木质素和纤维素含量(改进的Van Soest中性洗涤纤维(NDF)及酸性洗涤纤维(ADF)方法测定(Rowland & Roberts, 1994 ; Du et al ., 2004 )).倒木样品按采样份数分别测定, 即每个分解阶段3个重复, 结果取平均值. ...
Stoichiometric controls on carbon, nitrogen, and phosphorus dynamics in decomposing litter
1
2010
... 林分养分水平因树种、生长情况、环境条件、立地特征和管理方式的不同而表现出一定的差异(Elia & Potvin, 2003 ; Feng et al ., 2013 ).木材不同解剖结构的物理构造和化学组成不同(Schwarze, 2007 ), 化合物和元素含量的差异也较大(Meerts, 2002 ; Augusto et al ., 2008 ).de Aza等(2011) 对3种针叶树种的研究发现, 其心材的C含量显著高于边材, 受木质素含量影响较大.Cowling和Merrill (1966)对裸子植物的研究发现: 同一横截面树皮N含量占比例最高, 边材和内皮的N含量高于心材和外皮, 边材N含量是心材的1.5-2.0倍, 与本项研究结果基本一致.本研究中, 初始C含量由高到低依次为心材、边材和树皮, 变化范围37%-58%, 随着分解程度加深C含量显著降低, 主要是由于纤维素分解, 本研究中纤维素与C含量变化趋势一致, 呈极显著负相关(p < 0.01).环境中的无脊椎动物在倒木不同分解阶段和不同组分中发挥着重要作用(Harmon et al ., 1986 ), 心材纤维素含量下降稍快于边材可能是由于无脊椎动物在咀嚼或消化木材的同时引入了更多的微生物和捕食者, 而边材分解初期不利于无脊椎动物入侵.随着树皮和心材分解的加深, 边材与环境接触面积增大, 水热条件及通气量得以改善, 促进了微生物在边材的入侵及生长.倒木分解是复杂化合物转化成简单小分子物质、有机物矿化成无机物的过程, 养分释放过程受控于生物和物理因素, 矿化过程主要由分解者的活性调控(Manzoni et al ., 2010 ), 基质养分条件和物理性质影响着微生物的活性(Harmon et al ., 1986 ; Gonzalez-Polo et al ., 2013 ).分解者将环境有机基质中有机化合物和固存的C释放, 以满足自身生命活动需求, 同时将无机态的养分释放到环境中以保持自身化学计量特征的稳定(Cleveland & Liptzin, 2007 ).木材的初始N、P水平较低, 高质量基质(低C:N、C:P)有利于分解者入侵和附生植物定植生长(Benner & Vitousek, 2007 ), 加快倒木分解进程, 但当N、P含量达到一定水平后, 微生物的保护机制将多余的养分释放到环境中, 因此N、P表现出不同释放特征.此外, N、P含量显著影响C:N和C:P值, N、P初始含量分别反作用于C:N和C:P临界值大小.生态化学计量学理论认为大多数情况下C:N > 27或C:P > 186时微生物生长受限制(Sinsabaugh et al ., 2009 ), 本研究中, 心材和边材在整个分解阶段均易受到N、P限制, 树皮分解前期易受到N限制, 整个分解阶段均易受到P限制.认为初始N:P值也能很好地预测N、P动态, 他们的针叶树种分解过程养分释放研究发现临界N:P值约为20.本研究中, 心材和树皮N:P初始值分别为16和27, 表现为N相对含量从流失到富集的模式, 边材N:P初始值为116, 表现为相对含量的富集模式, 本研究结果与Laiho和Prescott (2004) 的研究发现不矛盾, 但仍需进一步验证.此外, 心材中的酚类物质含量高, 微生物底物有效性较低; 边材在树木死亡后保护机制迅速消失, 抑菌物质含量较低(Schwarze, 2007 ), 易于微生物入侵, 养分状况更易受微生物调控.以上结果表明: 不同分解阶段的倒木养分格局差异显著, 基质初始化学计量特征调控并影响后续分解阶段的养分格局; 倒木不同解剖结构的质量特征各异, 进而呈现不同的养分释放模式. ...
Mineral nutrient concentrations in sapwood and heartwood: A literature review
1
2002
... 林分养分水平因树种、生长情况、环境条件、立地特征和管理方式的不同而表现出一定的差异(Elia & Potvin, 2003 ; Feng et al ., 2013 ).木材不同解剖结构的物理构造和化学组成不同(Schwarze, 2007 ), 化合物和元素含量的差异也较大(Meerts, 2002 ; Augusto et al ., 2008 ).de Aza等(2011) 对3种针叶树种的研究发现, 其心材的C含量显著高于边材, 受木质素含量影响较大.Cowling和Merrill (1966)对裸子植物的研究发现: 同一横截面树皮N含量占比例最高, 边材和内皮的N含量高于心材和外皮, 边材N含量是心材的1.5-2.0倍, 与本项研究结果基本一致.本研究中, 初始C含量由高到低依次为心材、边材和树皮, 变化范围37%-58%, 随着分解程度加深C含量显著降低, 主要是由于纤维素分解, 本研究中纤维素与C含量变化趋势一致, 呈极显著负相关(p < 0.01).环境中的无脊椎动物在倒木不同分解阶段和不同组分中发挥着重要作用(Harmon et al ., 1986 ), 心材纤维素含量下降稍快于边材可能是由于无脊椎动物在咀嚼或消化木材的同时引入了更多的微生物和捕食者, 而边材分解初期不利于无脊椎动物入侵.随着树皮和心材分解的加深, 边材与环境接触面积增大, 水热条件及通气量得以改善, 促进了微生物在边材的入侵及生长.倒木分解是复杂化合物转化成简单小分子物质、有机物矿化成无机物的过程, 养分释放过程受控于生物和物理因素, 矿化过程主要由分解者的活性调控(Manzoni et al ., 2010 ), 基质养分条件和物理性质影响着微生物的活性(Harmon et al ., 1986 ; Gonzalez-Polo et al ., 2013 ).分解者将环境有机基质中有机化合物和固存的C释放, 以满足自身生命活动需求, 同时将无机态的养分释放到环境中以保持自身化学计量特征的稳定(Cleveland & Liptzin, 2007 ).木材的初始N、P水平较低, 高质量基质(低C:N、C:P)有利于分解者入侵和附生植物定植生长(Benner & Vitousek, 2007 ), 加快倒木分解进程, 但当N、P含量达到一定水平后, 微生物的保护机制将多余的养分释放到环境中, 因此N、P表现出不同释放特征.此外, N、P含量显著影响C:N和C:P值, N、P初始含量分别反作用于C:N和C:P临界值大小.生态化学计量学理论认为大多数情况下C:N > 27或C:P > 186时微生物生长受限制(Sinsabaugh et al ., 2009 ), 本研究中, 心材和边材在整个分解阶段均易受到N、P限制, 树皮分解前期易受到N限制, 整个分解阶段均易受到P限制.认为初始N:P值也能很好地预测N、P动态, 他们的针叶树种分解过程养分释放研究发现临界N:P值约为20.本研究中, 心材和树皮N:P初始值分别为16和27, 表现为N相对含量从流失到富集的模式, 边材N:P初始值为116, 表现为相对含量的富集模式, 本研究结果与Laiho和Prescott (2004) 的研究发现不矛盾, 但仍需进一步验证.此外, 心材中的酚类物质含量高, 微生物底物有效性较低; 边材在树木死亡后保护机制迅速消失, 抑菌物质含量较低(Schwarze, 2007 ), 易于微生物入侵, 养分状况更易受微生物调控.以上结果表明: 不同分解阶段的倒木养分格局差异显著, 基质初始化学计量特征调控并影响后续分解阶段的养分格局; 倒木不同解剖结构的质量特征各异, 进而呈现不同的养分释放模式. ...
Accumulation of dead wood in abandoned beech (Fagus sylvatica L.) forests in northwestern Germany
1
2011
... 川西高寒森林是我国西南林区的主体, 在调节局域气候、涵养水源、保持水土等方面发挥着重要作用(Yang et al ., 2005 ).受频繁的自然灾害和林分自然更新影响, 森林倒木储量巨大(Meyer & Schmidt, 2011 ), 往往受低温限制, 倒木分解缓慢(Kueppers et al ., 2004 ), 各分解阶段的倒木普遍存在(Kueppers et al ., 2004 ).地表倒木作为一个长期存在于森林生态系统的稳定成分, 在地区水源涵养、生物多样性保育、物质循环及水土保持等方面具有不可替代的作用.基于我们研究团队已建立的研究平台和较为成熟的研究方法, 以不同分解阶段的岷江冷杉(Abies faxoniana )倒木为研究对象, 我们研究了倒木树皮、边材和心材等组分在不同分解阶段的质量变化特征, 以期为深入认识倒木分解及其相关的物质循环过程提供一定的理论依据. ...
Carbon and nutrient release during decomposition of coarse woody debris in forest ecosystems of Central Siberia
2
2012
... 倒木是影响陆地生态系统物质循环与能量流动的重要组分之一(Harmon et al ., 1986 ), 在调控碳(C)库与养分循环等方面发挥着重要的作用(Tinker & Knight, 2000 ).然而, 倒木C和养分的释放受其他关键养分元素(如氮(N)、磷(P)等)含量及其有效性的控制.C:N:P化学计量比以及木质素和纤维素含量特征, 不仅可以表征倒木分解过程中基质的质量变化和养分限制状况(Wardle et al ., 2004 ; Wassen et al ., 2005 ), 而且可以指示物质循环和能量流动的方向.由于树皮、边材和心材基质质量(Schwarze, 2007 ; Bebber et al ., 2011 )和分解时间(Shorohova & Kapitsa, 2014 )的差异, 树皮、边材和心材在不同的分解阶段可能具有不同的质量变化特征.相对于边材和心材, 树皮最先受到环境的影响(Mukhortova, 2012 ; Stanton et al ., 2014 )和外来分解者的侵入(Zhou & Dai, 2012 ), 可能在初期具有分解速率较高和质量变化迅速的特征. 相对于心材, 边材受到物理化学构造的影响, 初期养分含量低(Schwarze, 2007 ), 随着活木保护机制的消失和树皮保护功能的减弱, 分解者和外界环境的影响加剧, 可能在分解中期或后期表现出较快的质量变化特征.心材由于储存了大量能源物质和难分解组分(Schwarze, 2007 ), 可能有较高的C含量和相对较低的养分含量, 但其结构疏松, 易于分解者活动和居住, 其分解过程和质量变化特征更为复杂.已有的研究更加关注于不同林型、不同管理条件及不同演替类型等林分内粗木质残体(CWD)储量和分解速率, 而缺乏关于高寒森林不同分解阶段心材、边材和树皮质量特征的研究, 极大地限制了人们对倒木分解过程的 认识. ...
... 森林中的植物残体主要分为凋落叶和木质残体(Bebber et al ., 2011 ).已有大量研究(Mukhortova, 2012 ; 郭平等, 2013 )证实, 基质N含量显著影响森林凋落物分解过程, 主要通过改变分解速率方式影响C固存.易分解指数F m 通过植物体的初始含量木质素和N在一定程度上可反映植物残体的易分解能力.我们的研究表明树皮的F m 指数显著高于其他组分(如心材、边材), 而且在整个分解过程中C:N:P化学计量比最低, 更易于微生物分解, 上述两组数据得出的结论一致.本研究中, N含量显著影响不同分解阶段倒木心材和边材的C格局(木质素:纤维素)(p < 0.05), 这是因为倒木心材和边材有高C:N初始值, 随着分解阶段加深逐渐增加的N浓度促进了微生物(主要是担子菌类)对木质素的分解(Watkinson et al ., 2006 ), 间接地影响了对纤维素的分解.而富N环境极大地促进了细菌类微生物生长, 分解木质素的微生物失去竞争优势, 故木质素分解受限, 在分解后期木质素:纤维素显著升高, 这表明可能存在一个木质素:N的最优区间, 当环境中木质素:N在此区间内会有较高的木质素分解效率, 过高过低均会抑制木质素的分解.高海拔地区N沉降现象越来越严重, 这可能会抑制倒木分解, 影响C释放过程.Bebber等(2011)研究发现在低N环境中增加可利用N含量可加速倒木分解, 与本研究结果一致. ...
Analysis of factors controlling soil organic matter levels in Great Plains grasslands
1
1987
... CENTURY模型(Parton et al ., 1987 ): 植物残体易分解比例F m = 0.99 - 0.018L/N, 式中L 为初始木质素含量, N 为初始氮含量. ...
Changes in mass and nutrient content of wood during decomposition in a south Florida mangrove forest
1
2005
... 高寒森林生态系统普遍存在各个分解阶段的倒木(Yoon et al ., 2011 ).倒木化学成分和物理结构的差异显著影响其不同解剖结构的质量特征及分解过程.以往研究多将倒木看作是一个整体(Romero et al ., 2005 ; 张利敏和王传宽, 2010 ), 忽略了倒木不同解剖结构的生理代谢途径各异可能会形成不同养分格局, 进而导致不同的养分释放模式.本研究结果表明: 随着分解程度加深, I到III分解阶段的树皮C含量略升高后显著降低, 其余各组分C含量降低, IV分解阶段开始降低程度尤为显著; 倒木各组分N含量在各分解阶段差异不显著, 除III分解阶段的心材外, 总体表现为随着分解程度加深而增加; P含量因结构组分不同表现出“富集—释放”和“释放”模式, 树皮C:N:P化学计量比一直处于较低水平, 易分解组分比例最高; C:N和C:P临界值出现在II和III分解阶段, 且与初始N、P含量成反比.这不仅表明倒木结构和化学组成在不同分解阶段具有显著的差异, 而且在一定程度上表征了N和P对不同分解阶段倒木C释放的限制特征. ...
Coarse woody debris in old Pinus sylvestris dominated forests along a geographic and human impact gradient in boreal Fennoscandia
1
2002
... 为避免其他因素的影响, 首先对研究区域内活立木物种组成与储量, 倒木储量及分布特征, 坡度、坡向和土壤性质等进行了调查研究(表1 ).倒木分解阶段的划分按照Rouvinen等(2002) 和闫恩荣等(2005) 提出的CWD 5级腐烂系划分系统(I级: 新鲜, 树木死亡不足1年; II级: 开始腐解, 小刀可刺进几mm; III级: 小刀可刺进2 cm; IV级: 小刀可刺进2-5 cm; V级: 小刀可任意刺穿木质体). ...
Lignin and cellulose fractionation in decomposition studies using Acid-Detergent Fibre methods
1
1994
... 将采集的样品粉碎, 过60目筛, 于65 ℃条件下烘干至恒重, 存放于干燥环境中待测.测定指标有: 总有机碳含量(重铬酸钾-外加热法(鲁如坤, 1999 )测定)、全氮含量(半微量凯氏定氮法(鲁如坤, 1999 )测定)、全磷含量(钼锑钪比色法(鲁如坤, 1999 )测定)、木质素和纤维素含量(改进的Van Soest中性洗涤纤维(NDF)及酸性洗涤纤维(ADF)方法测定(Rowland & Roberts, 1994 ; Du et al ., 2004 )).倒木样品按采样份数分别测定, 即每个分解阶段3个重复, 结果取平均值. ...
Wood decay under the microscope
5
2007
... 倒木是影响陆地生态系统物质循环与能量流动的重要组分之一(Harmon et al ., 1986 ), 在调控碳(C)库与养分循环等方面发挥着重要的作用(Tinker & Knight, 2000 ).然而, 倒木C和养分的释放受其他关键养分元素(如氮(N)、磷(P)等)含量及其有效性的控制.C:N:P化学计量比以及木质素和纤维素含量特征, 不仅可以表征倒木分解过程中基质的质量变化和养分限制状况(Wardle et al ., 2004 ; Wassen et al ., 2005 ), 而且可以指示物质循环和能量流动的方向.由于树皮、边材和心材基质质量(Schwarze, 2007 ; Bebber et al ., 2011 )和分解时间(Shorohova & Kapitsa, 2014 )的差异, 树皮、边材和心材在不同的分解阶段可能具有不同的质量变化特征.相对于边材和心材, 树皮最先受到环境的影响(Mukhortova, 2012 ; Stanton et al ., 2014 )和外来分解者的侵入(Zhou & Dai, 2012 ), 可能在初期具有分解速率较高和质量变化迅速的特征. 相对于心材, 边材受到物理化学构造的影响, 初期养分含量低(Schwarze, 2007 ), 随着活木保护机制的消失和树皮保护功能的减弱, 分解者和外界环境的影响加剧, 可能在分解中期或后期表现出较快的质量变化特征.心材由于储存了大量能源物质和难分解组分(Schwarze, 2007 ), 可能有较高的C含量和相对较低的养分含量, 但其结构疏松, 易于分解者活动和居住, 其分解过程和质量变化特征更为复杂.已有的研究更加关注于不同林型、不同管理条件及不同演替类型等林分内粗木质残体(CWD)储量和分解速率, 而缺乏关于高寒森林不同分解阶段心材、边材和树皮质量特征的研究, 极大地限制了人们对倒木分解过程的 认识. ...
... ), 可能在初期具有分解速率较高和质量变化迅速的特征. 相对于心材, 边材受到物理化学构造的影响, 初期养分含量低(Schwarze, 2007 ), 随着活木保护机制的消失和树皮保护功能的减弱, 分解者和外界环境的影响加剧, 可能在分解中期或后期表现出较快的质量变化特征.心材由于储存了大量能源物质和难分解组分(Schwarze, 2007 ), 可能有较高的C含量和相对较低的养分含量, 但其结构疏松, 易于分解者活动和居住, 其分解过程和质量变化特征更为复杂.已有的研究更加关注于不同林型、不同管理条件及不同演替类型等林分内粗木质残体(CWD)储量和分解速率, 而缺乏关于高寒森林不同分解阶段心材、边材和树皮质量特征的研究, 极大地限制了人们对倒木分解过程的 认识. ...
... ), 随着活木保护机制的消失和树皮保护功能的减弱, 分解者和外界环境的影响加剧, 可能在分解中期或后期表现出较快的质量变化特征.心材由于储存了大量能源物质和难分解组分(Schwarze, 2007 ), 可能有较高的C含量和相对较低的养分含量, 但其结构疏松, 易于分解者活动和居住, 其分解过程和质量变化特征更为复杂.已有的研究更加关注于不同林型、不同管理条件及不同演替类型等林分内粗木质残体(CWD)储量和分解速率, 而缺乏关于高寒森林不同分解阶段心材、边材和树皮质量特征的研究, 极大地限制了人们对倒木分解过程的 认识. ...
... 林分养分水平因树种、生长情况、环境条件、立地特征和管理方式的不同而表现出一定的差异(Elia & Potvin, 2003 ; Feng et al ., 2013 ).木材不同解剖结构的物理构造和化学组成不同(Schwarze, 2007 ), 化合物和元素含量的差异也较大(Meerts, 2002 ; Augusto et al ., 2008 ).de Aza等(2011) 对3种针叶树种的研究发现, 其心材的C含量显著高于边材, 受木质素含量影响较大.Cowling和Merrill (1966)对裸子植物的研究发现: 同一横截面树皮N含量占比例最高, 边材和内皮的N含量高于心材和外皮, 边材N含量是心材的1.5-2.0倍, 与本项研究结果基本一致.本研究中, 初始C含量由高到低依次为心材、边材和树皮, 变化范围37%-58%, 随着分解程度加深C含量显著降低, 主要是由于纤维素分解, 本研究中纤维素与C含量变化趋势一致, 呈极显著负相关(p < 0.01).环境中的无脊椎动物在倒木不同分解阶段和不同组分中发挥着重要作用(Harmon et al ., 1986 ), 心材纤维素含量下降稍快于边材可能是由于无脊椎动物在咀嚼或消化木材的同时引入了更多的微生物和捕食者, 而边材分解初期不利于无脊椎动物入侵.随着树皮和心材分解的加深, 边材与环境接触面积增大, 水热条件及通气量得以改善, 促进了微生物在边材的入侵及生长.倒木分解是复杂化合物转化成简单小分子物质、有机物矿化成无机物的过程, 养分释放过程受控于生物和物理因素, 矿化过程主要由分解者的活性调控(Manzoni et al ., 2010 ), 基质养分条件和物理性质影响着微生物的活性(Harmon et al ., 1986 ; Gonzalez-Polo et al ., 2013 ).分解者将环境有机基质中有机化合物和固存的C释放, 以满足自身生命活动需求, 同时将无机态的养分释放到环境中以保持自身化学计量特征的稳定(Cleveland & Liptzin, 2007 ).木材的初始N、P水平较低, 高质量基质(低C:N、C:P)有利于分解者入侵和附生植物定植生长(Benner & Vitousek, 2007 ), 加快倒木分解进程, 但当N、P含量达到一定水平后, 微生物的保护机制将多余的养分释放到环境中, 因此N、P表现出不同释放特征.此外, N、P含量显著影响C:N和C:P值, N、P初始含量分别反作用于C:N和C:P临界值大小.生态化学计量学理论认为大多数情况下C:N > 27或C:P > 186时微生物生长受限制(Sinsabaugh et al ., 2009 ), 本研究中, 心材和边材在整个分解阶段均易受到N、P限制, 树皮分解前期易受到N限制, 整个分解阶段均易受到P限制.认为初始N:P值也能很好地预测N、P动态, 他们的针叶树种分解过程养分释放研究发现临界N:P值约为20.本研究中, 心材和树皮N:P初始值分别为16和27, 表现为N相对含量从流失到富集的模式, 边材N:P初始值为116, 表现为相对含量的富集模式, 本研究结果与Laiho和Prescott (2004) 的研究发现不矛盾, 但仍需进一步验证.此外, 心材中的酚类物质含量高, 微生物底物有效性较低; 边材在树木死亡后保护机制迅速消失, 抑菌物质含量较低(Schwarze, 2007 ), 易于微生物入侵, 养分状况更易受微生物调控.以上结果表明: 不同分解阶段的倒木养分格局差异显著, 基质初始化学计量特征调控并影响后续分解阶段的养分格局; 倒木不同解剖结构的质量特征各异, 进而呈现不同的养分释放模式. ...
... 的研究发现不矛盾, 但仍需进一步验证.此外, 心材中的酚类物质含量高, 微生物底物有效性较低; 边材在树木死亡后保护机制迅速消失, 抑菌物质含量较低(Schwarze, 2007 ), 易于微生物入侵, 养分状况更易受微生物调控.以上结果表明: 不同分解阶段的倒木养分格局差异显著, 基质初始化学计量特征调控并影响后续分解阶段的养分格局; 倒木不同解剖结构的质量特征各异, 进而呈现不同的养分释放模式. ...
Mineralization and fragmentation rates of bark attached to logs in a northern boreal forest
1
2014
... 倒木是影响陆地生态系统物质循环与能量流动的重要组分之一(Harmon et al ., 1986 ), 在调控碳(C)库与养分循环等方面发挥着重要的作用(Tinker & Knight, 2000 ).然而, 倒木C和养分的释放受其他关键养分元素(如氮(N)、磷(P)等)含量及其有效性的控制.C:N:P化学计量比以及木质素和纤维素含量特征, 不仅可以表征倒木分解过程中基质的质量变化和养分限制状况(Wardle et al ., 2004 ; Wassen et al ., 2005 ), 而且可以指示物质循环和能量流动的方向.由于树皮、边材和心材基质质量(Schwarze, 2007 ; Bebber et al ., 2011 )和分解时间(Shorohova & Kapitsa, 2014 )的差异, 树皮、边材和心材在不同的分解阶段可能具有不同的质量变化特征.相对于边材和心材, 树皮最先受到环境的影响(Mukhortova, 2012 ; Stanton et al ., 2014 )和外来分解者的侵入(Zhou & Dai, 2012 ), 可能在初期具有分解速率较高和质量变化迅速的特征. 相对于心材, 边材受到物理化学构造的影响, 初期养分含量低(Schwarze, 2007 ), 随着活木保护机制的消失和树皮保护功能的减弱, 分解者和外界环境的影响加剧, 可能在分解中期或后期表现出较快的质量变化特征.心材由于储存了大量能源物质和难分解组分(Schwarze, 2007 ), 可能有较高的C含量和相对较低的养分含量, 但其结构疏松, 易于分解者活动和居住, 其分解过程和质量变化特征更为复杂.已有的研究更加关注于不同林型、不同管理条件及不同演替类型等林分内粗木质残体(CWD)储量和分解速率, 而缺乏关于高寒森林不同分解阶段心材、边材和树皮质量特征的研究, 极大地限制了人们对倒木分解过程的 认识. ...
Ecoenzymatic stoichiometry of microbial organic nutrient acquisition in soil and sediment
1
2009
... 林分养分水平因树种、生长情况、环境条件、立地特征和管理方式的不同而表现出一定的差异(Elia & Potvin, 2003 ; Feng et al ., 2013 ).木材不同解剖结构的物理构造和化学组成不同(Schwarze, 2007 ), 化合物和元素含量的差异也较大(Meerts, 2002 ; Augusto et al ., 2008 ).de Aza等(2011) 对3种针叶树种的研究发现, 其心材的C含量显著高于边材, 受木质素含量影响较大.Cowling和Merrill (1966)对裸子植物的研究发现: 同一横截面树皮N含量占比例最高, 边材和内皮的N含量高于心材和外皮, 边材N含量是心材的1.5-2.0倍, 与本项研究结果基本一致.本研究中, 初始C含量由高到低依次为心材、边材和树皮, 变化范围37%-58%, 随着分解程度加深C含量显著降低, 主要是由于纤维素分解, 本研究中纤维素与C含量变化趋势一致, 呈极显著负相关(p < 0.01).环境中的无脊椎动物在倒木不同分解阶段和不同组分中发挥着重要作用(Harmon et al ., 1986 ), 心材纤维素含量下降稍快于边材可能是由于无脊椎动物在咀嚼或消化木材的同时引入了更多的微生物和捕食者, 而边材分解初期不利于无脊椎动物入侵.随着树皮和心材分解的加深, 边材与环境接触面积增大, 水热条件及通气量得以改善, 促进了微生物在边材的入侵及生长.倒木分解是复杂化合物转化成简单小分子物质、有机物矿化成无机物的过程, 养分释放过程受控于生物和物理因素, 矿化过程主要由分解者的活性调控(Manzoni et al ., 2010 ), 基质养分条件和物理性质影响着微生物的活性(Harmon et al ., 1986 ; Gonzalez-Polo et al ., 2013 ).分解者将环境有机基质中有机化合物和固存的C释放, 以满足自身生命活动需求, 同时将无机态的养分释放到环境中以保持自身化学计量特征的稳定(Cleveland & Liptzin, 2007 ).木材的初始N、P水平较低, 高质量基质(低C:N、C:P)有利于分解者入侵和附生植物定植生长(Benner & Vitousek, 2007 ), 加快倒木分解进程, 但当N、P含量达到一定水平后, 微生物的保护机制将多余的养分释放到环境中, 因此N、P表现出不同释放特征.此外, N、P含量显著影响C:N和C:P值, N、P初始含量分别反作用于C:N和C:P临界值大小.生态化学计量学理论认为大多数情况下C:N > 27或C:P > 186时微生物生长受限制(Sinsabaugh et al ., 2009 ), 本研究中, 心材和边材在整个分解阶段均易受到N、P限制, 树皮分解前期易受到N限制, 整个分解阶段均易受到P限制.认为初始N:P值也能很好地预测N、P动态, 他们的针叶树种分解过程养分释放研究发现临界N:P值约为20.本研究中, 心材和树皮N:P初始值分别为16和27, 表现为N相对含量从流失到富集的模式, 边材N:P初始值为116, 表现为相对含量的富集模式, 本研究结果与Laiho和Prescott (2004) 的研究发现不矛盾, 但仍需进一步验证.此外, 心材中的酚类物质含量高, 微生物底物有效性较低; 边材在树木死亡后保护机制迅速消失, 抑菌物质含量较低(Schwarze, 2007 ), 易于微生物入侵, 养分状况更易受微生物调控.以上结果表明: 不同分解阶段的倒木养分格局差异显著, 基质初始化学计量特征调控并影响后续分解阶段的养分格局; 倒木不同解剖结构的质量特征各异, 进而呈现不同的养分释放模式. ...
Epiphytes improve host plant water use by microenvironment modification
1
2014
... 倒木是影响陆地生态系统物质循环与能量流动的重要组分之一(Harmon et al ., 1986 ), 在调控碳(C)库与养分循环等方面发挥着重要的作用(Tinker & Knight, 2000 ).然而, 倒木C和养分的释放受其他关键养分元素(如氮(N)、磷(P)等)含量及其有效性的控制.C:N:P化学计量比以及木质素和纤维素含量特征, 不仅可以表征倒木分解过程中基质的质量变化和养分限制状况(Wardle et al ., 2004 ; Wassen et al ., 2005 ), 而且可以指示物质循环和能量流动的方向.由于树皮、边材和心材基质质量(Schwarze, 2007 ; Bebber et al ., 2011 )和分解时间(Shorohova & Kapitsa, 2014 )的差异, 树皮、边材和心材在不同的分解阶段可能具有不同的质量变化特征.相对于边材和心材, 树皮最先受到环境的影响(Mukhortova, 2012 ; Stanton et al ., 2014 )和外来分解者的侵入(Zhou & Dai, 2012 ), 可能在初期具有分解速率较高和质量变化迅速的特征. 相对于心材, 边材受到物理化学构造的影响, 初期养分含量低(Schwarze, 2007 ), 随着活木保护机制的消失和树皮保护功能的减弱, 分解者和外界环境的影响加剧, 可能在分解中期或后期表现出较快的质量变化特征.心材由于储存了大量能源物质和难分解组分(Schwarze, 2007 ), 可能有较高的C含量和相对较低的养分含量, 但其结构疏松, 易于分解者活动和居住, 其分解过程和质量变化特征更为复杂.已有的研究更加关注于不同林型、不同管理条件及不同演替类型等林分内粗木质残体(CWD)储量和分解速率, 而缺乏关于高寒森林不同分解阶段心材、边材和树皮质量特征的研究, 极大地限制了人们对倒木分解过程的 认识. ...
Coarse woody debris following fire and logging in Wyoming lodgepole pine forests
1
2000
... 倒木是影响陆地生态系统物质循环与能量流动的重要组分之一(Harmon et al ., 1986 ), 在调控碳(C)库与养分循环等方面发挥着重要的作用(Tinker & Knight, 2000 ).然而, 倒木C和养分的释放受其他关键养分元素(如氮(N)、磷(P)等)含量及其有效性的控制.C:N:P化学计量比以及木质素和纤维素含量特征, 不仅可以表征倒木分解过程中基质的质量变化和养分限制状况(Wardle et al ., 2004 ; Wassen et al ., 2005 ), 而且可以指示物质循环和能量流动的方向.由于树皮、边材和心材基质质量(Schwarze, 2007 ; Bebber et al ., 2011 )和分解时间(Shorohova & Kapitsa, 2014 )的差异, 树皮、边材和心材在不同的分解阶段可能具有不同的质量变化特征.相对于边材和心材, 树皮最先受到环境的影响(Mukhortova, 2012 ; Stanton et al ., 2014 )和外来分解者的侵入(Zhou & Dai, 2012 ), 可能在初期具有分解速率较高和质量变化迅速的特征. 相对于心材, 边材受到物理化学构造的影响, 初期养分含量低(Schwarze, 2007 ), 随着活木保护机制的消失和树皮保护功能的减弱, 分解者和外界环境的影响加剧, 可能在分解中期或后期表现出较快的质量变化特征.心材由于储存了大量能源物质和难分解组分(Schwarze, 2007 ), 可能有较高的C含量和相对较低的养分含量, 但其结构疏松, 易于分解者活动和居住, 其分解过程和质量变化特征更为复杂.已有的研究更加关注于不同林型、不同管理条件及不同演替类型等林分内粗木质残体(CWD)储量和分解速率, 而缺乏关于高寒森林不同分解阶段心材、边材和树皮质量特征的研究, 极大地限制了人们对倒木分解过程的 认识. ...
Ecosystem properties and forest decline in contrasting long-term chronosequences
1
2004
... 倒木是影响陆地生态系统物质循环与能量流动的重要组分之一(Harmon et al ., 1986 ), 在调控碳(C)库与养分循环等方面发挥着重要的作用(Tinker & Knight, 2000 ).然而, 倒木C和养分的释放受其他关键养分元素(如氮(N)、磷(P)等)含量及其有效性的控制.C:N:P化学计量比以及木质素和纤维素含量特征, 不仅可以表征倒木分解过程中基质的质量变化和养分限制状况(Wardle et al ., 2004 ; Wassen et al ., 2005 ), 而且可以指示物质循环和能量流动的方向.由于树皮、边材和心材基质质量(Schwarze, 2007 ; Bebber et al ., 2011 )和分解时间(Shorohova & Kapitsa, 2014 )的差异, 树皮、边材和心材在不同的分解阶段可能具有不同的质量变化特征.相对于边材和心材, 树皮最先受到环境的影响(Mukhortova, 2012 ; Stanton et al ., 2014 )和外来分解者的侵入(Zhou & Dai, 2012 ), 可能在初期具有分解速率较高和质量变化迅速的特征. 相对于心材, 边材受到物理化学构造的影响, 初期养分含量低(Schwarze, 2007 ), 随着活木保护机制的消失和树皮保护功能的减弱, 分解者和外界环境的影响加剧, 可能在分解中期或后期表现出较快的质量变化特征.心材由于储存了大量能源物质和难分解组分(Schwarze, 2007 ), 可能有较高的C含量和相对较低的养分含量, 但其结构疏松, 易于分解者活动和居住, 其分解过程和质量变化特征更为复杂.已有的研究更加关注于不同林型、不同管理条件及不同演替类型等林分内粗木质残体(CWD)储量和分解速率, 而缺乏关于高寒森林不同分解阶段心材、边材和树皮质量特征的研究, 极大地限制了人们对倒木分解过程的 认识. ...
Endangered plants persist under phosphorus limitation
1
2005
... 倒木是影响陆地生态系统物质循环与能量流动的重要组分之一(Harmon et al ., 1986 ), 在调控碳(C)库与养分循环等方面发挥着重要的作用(Tinker & Knight, 2000 ).然而, 倒木C和养分的释放受其他关键养分元素(如氮(N)、磷(P)等)含量及其有效性的控制.C:N:P化学计量比以及木质素和纤维素含量特征, 不仅可以表征倒木分解过程中基质的质量变化和养分限制状况(Wardle et al ., 2004 ; Wassen et al ., 2005 ), 而且可以指示物质循环和能量流动的方向.由于树皮、边材和心材基质质量(Schwarze, 2007 ; Bebber et al ., 2011 )和分解时间(Shorohova & Kapitsa, 2014 )的差异, 树皮、边材和心材在不同的分解阶段可能具有不同的质量变化特征.相对于边材和心材, 树皮最先受到环境的影响(Mukhortova, 2012 ; Stanton et al ., 2014 )和外来分解者的侵入(Zhou & Dai, 2012 ), 可能在初期具有分解速率较高和质量变化迅速的特征. 相对于心材, 边材受到物理化学构造的影响, 初期养分含量低(Schwarze, 2007 ), 随着活木保护机制的消失和树皮保护功能的减弱, 分解者和外界环境的影响加剧, 可能在分解中期或后期表现出较快的质量变化特征.心材由于储存了大量能源物质和难分解组分(Schwarze, 2007 ), 可能有较高的C含量和相对较低的养分含量, 但其结构疏松, 易于分解者活动和居住, 其分解过程和质量变化特征更为复杂.已有的研究更加关注于不同林型、不同管理条件及不同演替类型等林分内粗木质残体(CWD)储量和分解速率, 而缺乏关于高寒森林不同分解阶段心材、边材和树皮质量特征的研究, 极大地限制了人们对倒木分解过程的 认识. ...
1
... 森林中的植物残体主要分为凋落叶和木质残体(Bebber et al ., 2011 ).已有大量研究(Mukhortova, 2012 ; 郭平等, 2013 )证实, 基质N含量显著影响森林凋落物分解过程, 主要通过改变分解速率方式影响C固存.易分解指数F m 通过植物体的初始含量木质素和N在一定程度上可反映植物残体的易分解能力.我们的研究表明树皮的F m 指数显著高于其他组分(如心材、边材), 而且在整个分解过程中C:N:P化学计量比最低, 更易于微生物分解, 上述两组数据得出的结论一致.本研究中, N含量显著影响不同分解阶段倒木心材和边材的C格局(木质素:纤维素)(p < 0.05), 这是因为倒木心材和边材有高C:N初始值, 随着分解阶段加深逐渐增加的N浓度促进了微生物(主要是担子菌类)对木质素的分解(Watkinson et al ., 2006 ), 间接地影响了对纤维素的分解.而富N环境极大地促进了细菌类微生物生长, 分解木质素的微生物失去竞争优势, 故木质素分解受限, 在分解后期木质素:纤维素显著升高, 这表明可能存在一个木质素:N的最优区间, 当环境中木质素:N在此区间内会有较高的木质素分解效率, 过高过低均会抑制木质素的分解.高海拔地区N沉降现象越来越严重, 这可能会抑制倒木分解, 影响C释放过程.Bebber等(2011)研究发现在低N环境中增加可利用N含量可加速倒木分解, 与本研究结果一致. ...
森林粗死木质残体的概念及其分类
1
2005
... 为避免其他因素的影响, 首先对研究区域内活立木物种组成与储量, 倒木储量及分布特征, 坡度、坡向和土壤性质等进行了调查研究(表1 ).倒木分解阶段的划分按照Rouvinen等(2002) 和闫恩荣等(2005) 提出的CWD 5级腐烂系划分系统(I级: 新鲜, 树木死亡不足1年; II级: 开始腐解, 小刀可刺进几mm; III级: 小刀可刺进2 cm; IV级: 小刀可刺进2-5 cm; V级: 小刀可任意刺穿木质体). ...
森林粗死木质残体的概念及其分类
1
2005
... 为避免其他因素的影响, 首先对研究区域内活立木物种组成与储量, 倒木储量及分布特征, 坡度、坡向和土壤性质等进行了调查研究(表1 ).倒木分解阶段的划分按照Rouvinen等(2002) 和闫恩荣等(2005) 提出的CWD 5级腐烂系划分系统(I级: 新鲜, 树木死亡不足1年; II级: 开始腐解, 小刀可刺进几mm; III级: 小刀可刺进2 cm; IV级: 小刀可刺进2-5 cm; V级: 小刀可任意刺穿木质体). ...
Litter dynamics of three subalpine forests in western Sichuan
1
2005
... 川西高寒森林是我国西南林区的主体, 在调节局域气候、涵养水源、保持水土等方面发挥着重要作用(Yang et al ., 2005 ).受频繁的自然灾害和林分自然更新影响, 森林倒木储量巨大(Meyer & Schmidt, 2011 ), 往往受低温限制, 倒木分解缓慢(Kueppers et al ., 2004 ), 各分解阶段的倒木普遍存在(Kueppers et al ., 2004 ).地表倒木作为一个长期存在于森林生态系统的稳定成分, 在地区水源涵养、生物多样性保育、物质循环及水土保持等方面具有不可替代的作用.基于我们研究团队已建立的研究平台和较为成熟的研究方法, 以不同分解阶段的岷江冷杉(Abies faxoniana )倒木为研究对象, 我们研究了倒木树皮、边材和心材等组分在不同分解阶段的质量变化特征, 以期为深入认识倒木分解及其相关的物质循环过程提供一定的理论依据. ...
Mass dynamics of coarse woody debris in an old-growth deciduous forest of Gwangneung, Korea
1
2011
... 高寒森林生态系统普遍存在各个分解阶段的倒木(Yoon et al ., 2011 ).倒木化学成分和物理结构的差异显著影响其不同解剖结构的质量特征及分解过程.以往研究多将倒木看作是一个整体(Romero et al ., 2005 ; 张利敏和王传宽, 2010 ), 忽略了倒木不同解剖结构的生理代谢途径各异可能会形成不同养分格局, 进而导致不同的养分释放模式.本研究结果表明: 随着分解程度加深, I到III分解阶段的树皮C含量略升高后显著降低, 其余各组分C含量降低, IV分解阶段开始降低程度尤为显著; 倒木各组分N含量在各分解阶段差异不显著, 除III分解阶段的心材外, 总体表现为随着分解程度加深而增加; P含量因结构组分不同表现出“富集—释放”和“释放”模式, 树皮C:N:P化学计量比一直处于较低水平, 易分解组分比例最高; C:N和C:P临界值出现在II和III分解阶段, 且与初始N、P含量成反比.这不仅表明倒木结构和化学组成在不同分解阶段具有显著的差异, 而且在一定程度上表征了N和P对不同分解阶段倒木C释放的限制特征. ...
东北东部山区11种温带树种粗木质残体分解与碳氮释放
1
2010
... 高寒森林生态系统普遍存在各个分解阶段的倒木(Yoon et al ., 2011 ).倒木化学成分和物理结构的差异显著影响其不同解剖结构的质量特征及分解过程.以往研究多将倒木看作是一个整体(Romero et al ., 2005 ; 张利敏和王传宽, 2010 ), 忽略了倒木不同解剖结构的生理代谢途径各异可能会形成不同养分格局, 进而导致不同的养分释放模式.本研究结果表明: 随着分解程度加深, I到III分解阶段的树皮C含量略升高后显著降低, 其余各组分C含量降低, IV分解阶段开始降低程度尤为显著; 倒木各组分N含量在各分解阶段差异不显著, 除III分解阶段的心材外, 总体表现为随着分解程度加深而增加; P含量因结构组分不同表现出“富集—释放”和“释放”模式, 树皮C:N:P化学计量比一直处于较低水平, 易分解组分比例最高; C:N和C:P临界值出现在II和III分解阶段, 且与初始N、P含量成反比.这不仅表明倒木结构和化学组成在不同分解阶段具有显著的差异, 而且在一定程度上表征了N和P对不同分解阶段倒木C释放的限制特征. ...
东北东部山区11种温带树种粗木质残体分解与碳氮释放
1
2010
... 高寒森林生态系统普遍存在各个分解阶段的倒木(Yoon et al ., 2011 ).倒木化学成分和物理结构的差异显著影响其不同解剖结构的质量特征及分解过程.以往研究多将倒木看作是一个整体(Romero et al ., 2005 ; 张利敏和王传宽, 2010 ), 忽略了倒木不同解剖结构的生理代谢途径各异可能会形成不同养分格局, 进而导致不同的养分释放模式.本研究结果表明: 随着分解程度加深, I到III分解阶段的树皮C含量略升高后显著降低, 其余各组分C含量降低, IV分解阶段开始降低程度尤为显著; 倒木各组分N含量在各分解阶段差异不显著, 除III分解阶段的心材外, 总体表现为随着分解程度加深而增加; P含量因结构组分不同表现出“富集—释放”和“释放”模式, 树皮C:N:P化学计量比一直处于较低水平, 易分解组分比例最高; C:N和C:P临界值出现在II和III分解阶段, 且与初始N、P含量成反比.这不仅表明倒木结构和化学组成在不同分解阶段具有显著的差异, 而且在一定程度上表征了N和P对不同分解阶段倒木C释放的限制特征. ...
Recognizing ecological patterns of wood-decaying polypores on gymnosperm and angiosperm trees in northeast China
1
2012
... 倒木是影响陆地生态系统物质循环与能量流动的重要组分之一(Harmon et al ., 1986 ), 在调控碳(C)库与养分循环等方面发挥着重要的作用(Tinker & Knight, 2000 ).然而, 倒木C和养分的释放受其他关键养分元素(如氮(N)、磷(P)等)含量及其有效性的控制.C:N:P化学计量比以及木质素和纤维素含量特征, 不仅可以表征倒木分解过程中基质的质量变化和养分限制状况(Wardle et al ., 2004 ; Wassen et al ., 2005 ), 而且可以指示物质循环和能量流动的方向.由于树皮、边材和心材基质质量(Schwarze, 2007 ; Bebber et al ., 2011 )和分解时间(Shorohova & Kapitsa, 2014 )的差异, 树皮、边材和心材在不同的分解阶段可能具有不同的质量变化特征.相对于边材和心材, 树皮最先受到环境的影响(Mukhortova, 2012 ; Stanton et al ., 2014 )和外来分解者的侵入(Zhou & Dai, 2012 ), 可能在初期具有分解速率较高和质量变化迅速的特征. 相对于心材, 边材受到物理化学构造的影响, 初期养分含量低(Schwarze, 2007 ), 随着活木保护机制的消失和树皮保护功能的减弱, 分解者和外界环境的影响加剧, 可能在分解中期或后期表现出较快的质量变化特征.心材由于储存了大量能源物质和难分解组分(Schwarze, 2007 ), 可能有较高的C含量和相对较低的养分含量, 但其结构疏松, 易于分解者活动和居住, 其分解过程和质量变化特征更为复杂.已有的研究更加关注于不同林型、不同管理条件及不同演替类型等林分内粗木质残体(CWD)储量和分解速率, 而缺乏关于高寒森林不同分解阶段心材、边材和树皮质量特征的研究, 极大地限制了人们对倒木分解过程的 认识. ...
 , 谭波, 肖洒, 李俊, 苟小林
, 谭波, 肖洒, 李俊, 苟小林


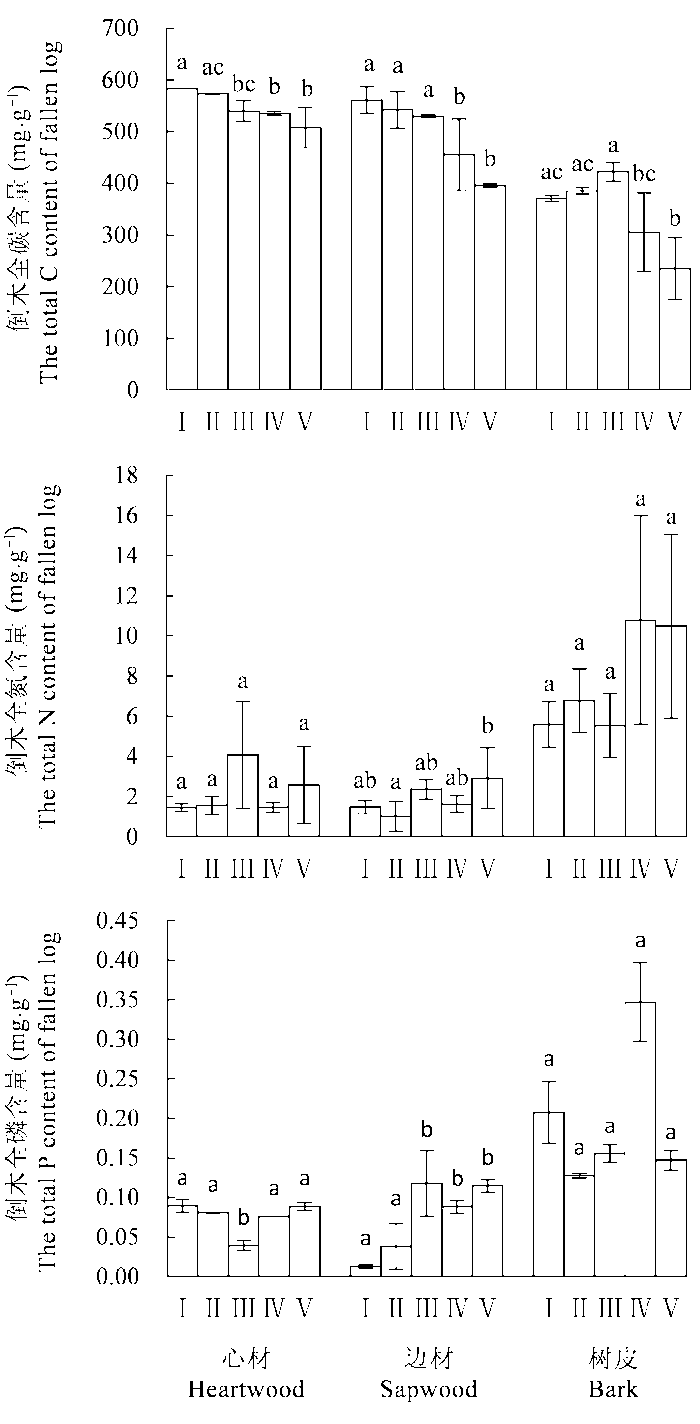
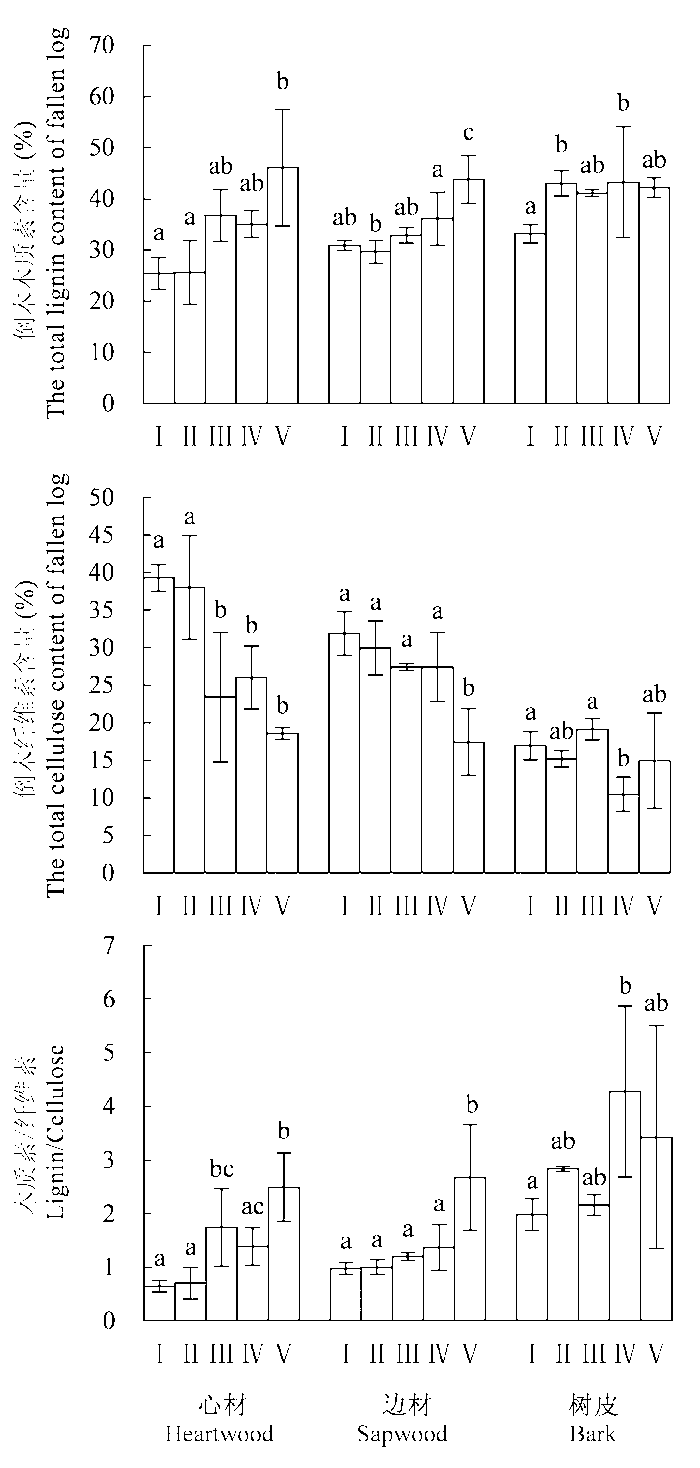
{kind=link}
{kind=link}
{kind=link}
{kind=link}