随着频繁的人类活动和异常自然干扰的发生, 森林生态系统结构与功能正受到严重的威胁(Vitousek et al ., 1997 )。干扰生态学已成为生态学研究最活跃的领域之一(朱教君和刘世荣, 2007 ; Bond-Lamberty et al ., 2014 ), 其重点是从森林群落的干扰历史和林窗(隙)动态两个方面对中小规模干扰的特征及影响开展研究, 且有关林窗动态的研究相对较多(臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 )。重建干扰历史是植物群落演替和更新研究的基础, 越来越受到重视。森林群落的干扰动态可以通过固定样地的长期观测获得(Cooke & Lorenzetti, 2006 ), 但对于缺乏长期观测的森林群落, 人们无法得知其干扰历史。树木年轮是树木生长的历史记录(Smith, 2008 ), 其宽度、密度、同位素含量以及解剖结构等记载了树木逐年的生长信息及内、外部环境变化的影响(Cook & Kairiuktis, 1990 )。树轮具有生长时间长、定年准确、材料广泛、容易获取等优点, 是研究树木生长环境变化的重要手段, 也是重建森林群落动态的重要材料。
国外已有一些研究利用树木年轮重建森林干扰历史(Nowacki & Abrams, 1997 ; Ishikawa et al ., 1999 ; Rubino & McCarthyz, 2004 ; Brown & Giesecke, 2014 ), 而国内这方面研究起步较晚, 且相对较少。20世纪90年代初, 阳含熙和谢海生(1994)最先利用树木生长释放和抑制对长白山红松(Pinus koraiensis )混交林干扰历史进行了重建, 并指出该方法是以风作为主要干扰类型, 重建干扰历史最准确、最可靠、提供信息量最大的方法; 侯向阳等(2000)研究长白山红松林干扰节律时, 也得到了类似的结论。乔佃锋(2005)利用该方法(阳含熙和谢海生, 1994 )重建了长白山敦化地区阔叶红松林的干扰历史, 发现该区红松林干扰以小规模的风干扰为主, 1980年出现了明显的干扰峰, 干扰峰之间60年的时间间隔正好与红松的更替世代相吻合。王晓春和赵玉芳(2011)发现前一年径向生长对生长释放事件的判定很重要, 小兴安岭北端黑河地区的红松和红皮云杉(Picea koraiensis )的生长释放可能受林窗产生、微环境变化以及温度和降水等气候因素影响。夏冰等(1995)在天目山建立了生长释放和抑制的判定标准, 并重建了黄山松(Pinus taiwanensis )阔叶林树冠的干扰历史; 同时, 还对不同树种年轮宽度的生长释放判别方法进行了研究, 并做了重建干扰强度的尝试(夏冰等, 1997 , 2001 )。此外, 不同学者还利用树轮生态学方法对火灾(张磊和王晓春, 2013 )、泥石流(铁永波等, 2014 )、山体滑坡(洪婷等, 2012 )和冰川活动(Zhu et al ., 2013 )等干扰进行了重建。
阔叶红松林是我国东北东部山地的地带性顶极植被, 分布于长白山、完达山、小兴安岭一带的中低山区, 是世界上为数不多的大面积原生针阔混交林, 以其结构复杂、组成独特、生物多样性丰富而著称(李景文, 1997 )。然而, 由于过去不合理的采伐利用, 天然阔叶红松林资源逐年锐减, 原始阔叶红松林几乎荡然无存。开展阔叶红松林主要树种生长释放及干扰历史研究, 对探讨现存阔叶红松林的稳定性、碳源汇功能等具有重要意义。目前已有的阔叶红松林干扰重建研究主要集中在长白山一带, 且多停留在干扰年代和干扰频率的确定上, 并未对干扰强度以及可能存在的干扰机制进行探讨。小兴安岭是阔叶红松林在我国分布最为集中, 且保存完好的核心地区。红松和臭冷杉(Abies nephrolepis )是阔叶红松林内的主要针叶树种。因此, 本文利用其生长抑制和释放规律, 确定小兴安岭典型阔叶红松林林窗干扰的发生时间和频率, 探讨过去的干扰历史及干扰形成机制, 为阔叶红松林的保护与可持续利用提供科学依据。
1 材料和方法
1.1 研究地区概况
本研究实验地设在黑龙江省凉水国家级自然保护区(47.18° N, 128.96° E)。该保护区位于黑龙江省伊春市带岭区, 属小兴安岭南部达里带岭支脉的东坡。保护区海拔280-707 m, 为典型的低山丘陵地貌。森林覆被率达96%, 是我国现有的较大片原始红松林之一。该区地处中纬度大陆东岸, 属温带大陆性季风气候, 冬长夏短。冬季气候严寒、干燥; 夏季降水集中, 气温较高; 春秋两季气候多变。该地区常刮起偏南大风, 春、夏多西南风; 秋、冬多西北风。年平均气温-0.3 ℃, 年降水量676 mm, 年平均相对湿度78%-96%, 年蒸发量805.4 mm。年日照时间1850 h, 无霜期100-120天。地带性土壤为暗 棕壤。
本研究在已有的9 hm2 典型阔叶红松林森林动态监测样地内进行。该样地位于保护区的中段, 从凉水沟谷地到海拔600 m的中山山脊。样地以红松为优势种, 伴生有臭冷杉、水曲柳(Fraxinus mandshurica )、紫椴(Tilia amurensis )、枫桦(Betula costata )、大青杨(Populus ussuriensis )、裂叶榆(Ulmus laciniata )、色木槭(Acer mono )和花楷槭(Acer ukurunduense )等20余种针阔叶树种; 主要下木有刺五加(Acanthopanax senticosus )、毛榛子(Corylus mand- shurica )、东北山梅花(Philadelphus schrenkii )等。草本植物主要有薹草(Carex spp.)、东北蹄盖蕨(Athyrium brevifrons )和东北羊角芹(Aegopodium alpestre )等, 藤本植物有五味子(Schisandra chinensis )、狗枣猕猴桃(Actinidia kolomikta )等(徐丽娜和金光泽, 2012 )。
1.2 研究方法
2013年6月, 在对9 hm2 固定样地大面积踏查的基础上, 选择比较典型的300 m × 200 m地块作为实验样地。采用样方调查法(臧润国等, 1999 ), 以林内东北点作为调查起点, 沿罗盘仪所指方向, 由北向南穿行, 调查每一个林窗的基本特征(本文定义树冠空隙面积10 m2 以上的林冠空隙为林窗)。对样地内每一株林窗边缘木(gap tree)(距林窗边缘3 m以内立木)进行生长锥取样。取样位置在树高约1.2 m处, 取样方向均朝向林窗中心点处, 所取样芯尽量达到髓心。在符合要求的44个林窗中共获得606个边缘木样芯, 其中红松461个(平均年龄约160年), 臭冷杉145个(平均年龄约80年)。因红松与臭冷杉均存在腐心现象, 部分样芯未取到髓心, 因此实际树龄比样芯年龄更长。此外, 还在固定样地外随机钻取非林窗木(non-gap tree)样芯(每个树种≥40个样芯)作为对照。将钻取的树芯装入孔径相当的塑料管并进行编号, 同时记录每一个样本的特征及生境条件。
将野外采集的所有样芯带回实验室, 进行固定、干燥、打磨等预处理(Stokes & Smiley, 1996 )。在显微镜下用骨架图法进行交叉定年, 然后用年轮分析仪进行宽度测量, 精度为0.001 mm。最后, 用COFECHA程序来检验定年结果(Holmes, 1983 ), 根据软件提示在显微镜下检查并修订定年与测量中出现的误差, 以确保年轮的日历年代准确。
1.3 数据处理与统计分析
本文采用王晓春和赵玉芳(2011)的方法计算当年年轮宽度序列的生长变化百分率(Percentage Growth Change, GC)。该方法在Nowacki和Abrams (1997)的方法基础上, 将滑动平均值法修正为中值法, 并将滑动窗口由10 a调整为5 a, 以凸显非正常干扰事件的影响(Rubino & McCarthyz, 2004 ; 王晓春和赵玉芳, 2011 )。研究中以林窗为单位计算臭冷杉和红松的生长释放标准曲线, 并利用该标准曲线判断每一边缘木的生长释放情况。将GC介于25%-50%之间且持续5 a以上的释放记为“中等释放”, GC > 50%且持续5 a以上的释放记为“主要释放”, GC < 25%则不考虑为真正释放。林窗木与非林窗木生长变化百分率的差值可以在很大程度上反映林窗的干扰情况。首先, 求出样地内每一个林窗所有边缘木生长变化百分率的平均值; 其次, 将各平均值逐一与非林窗木生长变化百分率的平均值作差, 得出各个林窗干扰情况随时间的变化序列; 最后, 再将得到的44个时间序列每一年的值, 按正、负进行求和后便得出柱状累计图(图1 )。本文以平均每株树木经历生长释放次数的倒数乘以生长释放频率, 推测过去树木的死亡率即干扰强度(夏冰等, 1997 , 2001 )。例如, 臭冷杉经历的平均主要生长释放次数为1.05次, 其平均生长释放频率为每10 a 8.79%, 则其平均干扰强度是每10 a 8.37% (表1 )。
图1 臭冷杉(A, B)和红松(C, D)林窗木与非林窗木生长变化百分率比较及其差值变化。
Fig. 1 Comparison of percentage growth changes between gap trees and non-gap trees for Abies nephrolepis (A, B) and Pinus koraiensis (C, D) and differences in percentage growth changes between trees in gap trees and non-gap trees.
气候资料由中国气象科学数据共享服务网(http://cdc.cma.gov.cn/)提供。选择与采样点距离较近且位于同一气候区的伊春气象站点(47.73° N, 128.92° E, 海拔240.9 m) 1956-2013年的月平均最高气温、月平均最低气温、风速等气候变量用于计算气候指标异常值。气候指标异常值的计算方法以气温异常值为例, 气温异常值用公式T diff = T 2 - T 1 计算, 式中T diff 为气温异常值, T 1 和T 2 分别为前5 a和随后5 a气温的平均值, 用此公式在每个气温序列上滑动得到气温异常值序列。太阳黑子数从太阳黑子指数和太阳活动长期观测网(http://www.sidc.be/ silso/)上免费下载, 其异常值计算方法同气温异常值计算方法。本文太阳黑子爆发年是指年太阳黑子数大于1800个的年份; 风速异常值年指最大风速大于13.9 m·s-1 的年份; 极端高、低温年分别指9-10月极端高温平均值大于25 ℃、7-8月极端低温平均值小于7 ℃的年份。
用SPSS 19.5软件进行Pearson相关性分析; 用MTM程序对生长释放变化进行多带谱分析, 以检验其干扰的周期及显著性; 用EVENT程序进行叠加点事件分析(superposed epoch analysis, SEA), 以确定生长抑制、释放的干扰机制; 用SigmaPlot 12.5和Excel 2010软件完成图表制作。
2 结果
2.1 干扰特征分析
2.1.1 生长变化百分率
在树木生长初期, 林窗与非林窗树木生长变化百分率差异较大, 而树木生长后期的变化规律基本一致, 这可能与前期样本量较少有关(图1 )。林窗与非林窗臭冷杉均在1935-1940、1970-1975年等时间段内发生了明显的生长抑制, 1925-1930、1945-1950和1980-1985年等时间段内发生了明显的生长释放。林窗与非林窗红松生长释放和抑制遍布树木生长的全过程, 生长释放和抑制现象交替出现且具有一定的周期性, 在1815-1820、1910-1915、1925-1930和1995-2000年等时间段内有明显的生长抑制, 1795- 1800、1850-1855、1920-1925和1930-1935年内均有明显的生长释放(图1 )。臭冷杉生长释放的最大峰值约为300%, 而红松生长释放的最大峰值仅为80%左右, 说明臭冷杉比红松对外界干扰的反应更为敏感, 臭冷杉更适用于干扰历史重建, 但由于臭冷杉寿命较短, 重建长时间尺度干扰时往往需借助于红松等寿命较长的树种。
林窗与非林窗树木生长变化百分率差值表明: 以林窗为统计单位, 研究区内林窗与非林窗臭冷杉生长变化百分率差异较大, 臭冷杉对干扰的响应比红松更敏感。臭冷杉在1894、1934、1951、1968和1990年出现了明显的干扰峰, 1885-1890、1930- 1935、1945-1955、1965-1975和1980-1990年间出现了明显的干扰谷(图1B ), 表明臭冷杉在1885、1930、1945、1965和1980年左右发生了较明显的生长抑制, 1890、1975和1985年左右发生了较明显的生长释放。红松干扰峰和干扰谷交替出现, 峰谷之间的时间间隔约11年, 在1785、1800、1830、1870和1890年左右生长抑制较为明显, 1780、1795、1825、1880、1895、1930、1945和2000年左右生长释放较为明显(图1D )。
2.1.2 林窗木生长释放特征统计
臭冷杉发生主要释放、中等以上释放占其所有释放的比例均高于红松(表1 ), 这与树种对干扰的敏感度及所取样芯的年龄有关, 树种对干扰的敏感度越低或低龄样本比例越大, 生长释放林木所占的比例就越小。单株臭冷杉发生中等以上生长释放的次数低于红松, 但发生主要释放的次数却高于红松。臭冷杉发生中等以上释放与主要释放的释放期平均长度均比红松长约1年, 初次释放年龄和平均间隔期均小于红松10 a以上, 且这些指标均存在较大的变异性。就单个林窗而言, 因林窗边缘木中红松的比例较多且年龄相对较长, 故臭冷杉发生中等以上释放与主要释放的次数均明显少于红松。
臭冷杉在过去的150 a中所经历的中等以上释放和主要释放的平均释放频率明显高于红松过去280 a中所经历的平均生长释放频率(表1 ), 臭冷杉中等以上释放和主要释放的干扰强度亦高于红松。臭冷杉发生中等以上释放、主要释放的干扰间隔期均小于红松, 且两树种发生两种类型释放的干扰间隔期均与树木的寿命接近。上述生长释放特征表明干扰对臭冷杉的影响大于红松, 臭冷杉对干扰更为敏感。
2.1.3 干扰年表的建立
将44个林窗的所有边缘木发生生长释放事件的数量和频率, 以年为单位进行统计, 得出了红松和臭冷杉的干扰年表(图2 , 图3 )。
图2 小兴安岭红松干扰年表。A, 生长释放树木个数随时间的变化。B, 生长释放树木数量占总采样树木数量的比例随时间的变化。
Fig. 2 Disturbance chronology of Pinus koraiensis in Xiaoxing’an Mountain. A, Number of trees with release changes over time. B, Percentage of trees with release changes over time.
图3 小兴安岭臭冷杉干扰年表。A, 生长释放树木个数随时间的变化。B, 生长释放树木数量占总采样树木的比例随时间的变化。
Fig. 3 Disturbance chronology of Abies nephrolepis in Xiaoxing’an Mountain. A, number of trees with release changes over time. B, percentage of trees with release changes over time.
红松的干扰年表从1721-2008年, 共288 a (图2 )。发生中等释放的数量和频率均明显大于主要释放, 在1751、1770、1800、1830、1842和1937年等年份出现了明显的释放高峰。随着样本量的增加, 发生生长释放的样芯数量急剧增加并逐渐稳定。但释放样芯所占比例变化相对稳定, 在15%左右上下波动, 并未因样本量增加而增加, 且呈现出一定的周期性(图2 B)。
臭冷杉的干扰年表从1855-2008年, 共154 a (图3 )。发生中等释放的数量和频率均明显多于主要释放。随着样本量的增加, 释放样芯的数量急剧增加并逐渐稳定。但释放样芯所占比例随着样本量的增加, 变化相对稳定, 在25%左右上下波动。在1894、1951和1990年等年份出现了明显的释放高峰, 与红松一样, 臭冷杉生长释放亦呈现出一定的周期性。
2.1.4 干扰周期
红松干扰年表(生长释放树木数量占总采样树木的比例随时间的变化序列)存在9.1-18.2 a、7.3-7.9 a、3.8 a、3.5 a和2.0 a的极显著(p < 0.01)周期, 臭冷杉干扰年表(生长释放树木数量占总采样树木的比例随时间的变化序列)存在65-85 a、7.5-48.8 a和3.5-3.6 a的极显著周期(图4 )。通常气象因子波动和植物生长具有2-3 a的周期, 红松与臭冷杉干扰年表也记录了该周期; 2-7 a的干扰周期也可能与厄尔尼诺-南方涛动(El Niño-Southern Oscillation, ENSO)事件有关; 9.1-18.2 a的干扰周期可能受太阳活动的影响。而臭冷杉干扰年表中65-85 a的干扰周期可能受北大西洋年代际振荡(Atlantic Multidecadal Oscillation, AMO)的影响, 已有研究表明AMO会影响东北亚树木生长(Wang et al ., 2011 ; 姚启超和王晓春, 2013 )。
图4 红松(A)和臭冷杉(B)干扰年表的多带谱分析。图中数值代表该时段周期显著(p < 0.01)。
Fig. 4 Multi-taper method spectrums analysis for Pinus koraiensis (A) and Abies nephrolepis (B) disturbance chronologies. Values shown in the figure represent significant periods (p < 0.01).
2.2 干扰机制分析
2.2.1 异常风速对生长释放的影响
风速异常值与树木生长变化百分率相关性分析表明(图5 ), 异常风速对林窗木的生长影响很大, 风速异常值与红松生长变化百分率呈负相关, 与臭冷杉生长变化百分率呈正相关。红松生长变化百分率与前一年8月、11月以及当年8月的平均风速异常值呈正相关; 与前一年6-7月、9-10月、前一年12月至当年4月、当年6-7月以及9月的平均风速异常值显著(p < 0.05)负相关; 而臭冷杉生长变化百分率仅与前一年10月和当年1月的平均风速异常值呈负相关, 与前一年4-6月、8-9月、当年2-3月以及5月的平均风速异常值显著正相关。两树种生长变化百分率和最大风速异常值相关性与平均风速异常值类似。树木自身的遗传差异或所处的空间环境不同, 可能是导致红松和臭冷杉生长变化百分率与异常风速的响应存在明显差异的原因。
图5 红松和臭冷杉生长变化百分率与月平均风速(A)、最大风速(B)异常值的相关性。灰色柱表示显著相关(p < 0.05)。P表示前一年, 如”P8”指前一年8月。
Fig. 5 Correlation coefficients between percentage growth changes in Pinus koraiensis (black bar) and Abies nephrolepis (blank bar) and monthly mean wind speed (A) and maximum anomalous wind speed (B). The gray columns indicate significant correlations (p < 0.05). The letter P represents the previous year. For instance, “P8” represents the previous August.
由最大风速与红松和臭冷杉生长变化百分率的SEA分析结果可知(图6 ), 小兴安岭地区林窗红松树轮记录的生长变化百分率, 由最大风速发生前1-4 a的负相关转变为发生后1-3 a的正相关(图6 A)。这表明林窗红松生长变化百分率受风速影响很大, 风可能是导致林窗红松生长释放的主要原因。臭冷杉可能因自身遗传原因, 生长变化百分率受最大风速影响不大(图6 B)。上述结果表明, 风对该地区树木的生长释放有很大影响, 这种影响可能因树种不同而存在差异。
图6 最大异常风速与红松(A)和臭冷杉(B)生长变化百分率的叠加事件分析。
Fig. 6 Superimposed events analysis between maximum anomalous wind speed and percentage growth changes in Pinus koraiensis (A) and Abies nephrolepis (B).
2.2.2 异常气温对生长释放的影响
高温异常值对本地区红松生长释放限制较强, 低温异常值对本地区红松生长释放限制较弱, 臭冷杉则相反, 总体来看, 异常气温对臭冷杉的影响比对红松的影响大(图7 )。红松生长变化百分率与前一年6月和11月、当年2月和6月的低温异常值显著正相关, 与前一年和当年5月高温异常值显著负相关(图7 A)。另外, 生长季(5-6月)和冬半年(前一年10月-3月)异常气温对红松生长变化百分率的影响也较大。臭冷杉生长变化百分率与前一年和当年4-6月、8-9月低温异常值显著负相关, 与前一年10月低温异常值显著正相关; 臭冷杉生长变化百分率与前一年和当年6月高温异常值显著负相关, 与前一年10-11月及当年2-3月显著正相关(图7 B)。夏半年(4-9月)异常气温对臭冷杉生长释放有明显的抑制作用, 而冬半年(前一年10月-3月)异常气温有明显的促进作用。
图7 红松(A)和臭冷杉(B)生长变化百分率与月最低气温、最高气温异常值的相关性。灰色柱表示显著相关(p < 0.05)。P表示前一年, 如”P8”指前一年8月。
Fig. 7 Correlation coefficients between percentage growth changes in Pinus koraiensis (A) and Abies nephrolepis (B) and minimum anomalous air temperature and maximum anomalous air temperature. The gray columns indicate significant correlations (p < 0.05). The letter P represents the previous year. For instance, “P8” represents the previous August.
小兴安岭地区林窗红松树轮记录的生长变化百分率, 由极端高温年发生前1-3 a的显著负相关转变为发生后3 a的显著正相关(图8A ), 这表明异常高温对林窗红松生长释放影响较大, 异常高温也可能是导致林窗红松生长释放的原因之一。臭冷杉树轮记录的生长变化百分率, 由极端低温年发生前3-5 a的显著正相关转变为发生后的不显著负相关(图8B ), 说明异常低温对该地区臭冷杉生长释放有重要影响。上述结果表明, 异常气温对林窗边缘树木的生长释放有重要影响, 且该影响可能因树种及作用时间差异而不同。
图8 极端气温与红松(A)和臭冷杉(B)生长变化百分率的叠加事件分析。灰色柱表示显著相关(p < 0.05)
Fig. 8 Superimposed events analysis between extreme air temperatures and percentage growth changes in Pinus koraiensis (A) and Abies nephrolepis (B). The gray columns indicate significant correlations (p < 0.05).
3 讨论
3.1 生长释放与林窗干扰
林窗干扰是自然界中最常见的一种森林干扰(臧润国和徐化成, 1998 ; 臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 )。林窗干扰发生后, 周围树木由于竞争去除而产生生长释放(王晓春和赵玉芳, 2011 ), 并且这种释放可以持续多年。通常将大于一定阈值且持续5 a或10 a以上的生长释放视为“真正释放”, 并将生长释放开始的时间视为干扰的发生时间(Black & Abrams, 2004 ; Rubino & McCarthyz, 2004 ; 王晓春和赵玉芳, 2011 )。确定生长释放的发生时间是重建森林干扰历史的最基本过程(Black & Abrams, 2004 ), 林窗形成时间常以林窗内幼苗更新时间(刘少冲等, 2013 )、倒木倾倒时间(用腐烂等级确定, 韩路等, 2011 )以及林窗周边树木生长释放时间(阳含熙和谢海生, 1994 ; 夏冰等, 1995 )等确定。本研究林窗干扰时间的确定主要用树木生长释放来判断, 同时还参考了林窗内的幼苗更新及倒木腐烂等级, 但有些林窗更新的非均一性以及林窗内倒木腐烂情况比较复杂也影响林窗形成时间的确定。
生长释放判定方法的选择及判定标准的确定是树轮干扰历史重建的关键。判定方法和标准不同, 得出的生长释放结果亦不同(Rubino & McCarthyz, 2004 ), 每一种方法都有各自的优缺点及使用范围(封晓辉等, 2011 )。在实践中, 通过主观判断去选择生长释放判定方法及判定标准, 往往会因主观原因放大或弱化干扰事件。因此, 在判定方法及判定标准选择时, 在充分考虑研究需要的同时, 还需结合研究地点及环境条件等合理选择(阳含熙和谢海生, 1994 ; 夏冰等, 1997 ; 王晓春和赵玉芳, 2011 )。为了保留更多的干扰信号, 本文选用生长释放标准曲线法(王晓春和赵玉芳, 2011 ), 即生长变化率超过一定的标准便可视为生长释放发生, 该方法是目前应用最多的方法(封晓辉等, 2011 )。本研究中林窗木与非林窗木的生长变化百分率有很强的一致性(仅局部存在部分差异), 说明林窗木生长变化百分率序列中包含了部分非干扰信号。通过生长释放判定得到的生长释放中必定会有部分噪音即非干扰信号, 即并不是所有的释放(包括中等释放和主要释放)都能代表林窗干扰(王晓春和赵玉芳, 2011 ), 因此主要释放或部分主要释放才能代表真正意义上的干扰。 生长变化百分率差值图(图1 )与干扰年表(图2 , 图3 )谷、峰值具有很强的一致性, 且干扰年表在一定程度上对生长变化百分率差值图中不太明显的峰值具有局部放大作用, 经局部放大的部分信号可能是噪音的主要来源。因此, 通过林窗边缘木生长释放判定得出的干扰年表, 能够很好地反映该区阔叶红松林的干扰历史, 且能将一些不太明显的微小干扰包括其中。
3.2 干扰的变异性和区域性
本研究发现小兴安岭地区林窗木与非林窗木的年生长变化规律(平均生长变化百分率)基本一致, 而以林窗为单位得到的林窗木生长变化百分率与非林窗木之间的差异却很明显, 可能是不同林窗或同一林窗不同树木对同一干扰事件的响应存在差异的结果。自然干扰及其所产生的影响存在巨大的变异性(魏晓华, 2010 ), 可能因海拔、坡度和坡位等因素而出现不同的格局(沈泽昊等, 2001 )。刘妍妍和金光泽(2009)在分析本研究样地内粗木质残体与地形因子的关系时, 也得到了类似的结论。林窗木生长释放特征也具有极大的变异性, 如臭冷杉发生中等以上释放每株2.16次,介于每株0-6次之间; 初次释放平均年龄为32.14 a, 介于15-116 a; 平均间隔期为16.56 a, 介于2-67 a, 从另一方面印证了小兴安岭地区典型阔叶红松林干扰及其所产生的影响存在巨大的变异性。夏冰等(1997)在亚高山云冷杉混交林得到了类似的研究结果。此外, 本研究还发现了一些连续多年的生长释放, 如1889-1904、1932-1938、1947-1973年等, 这可能是连续发生释放或者间隔释放相叠加的结果, 也可能是区域尺度上大范围干扰事件造成的。
小兴安岭地区阔叶红松林内红松干扰事件主要发生在18世纪30年代、19世纪10年代、19世纪30-40年代、20世纪20-30年代, 臭冷杉主要发生在19世纪90年代、20世纪30年代、20世纪50-60年代、20世纪90年代。该结果与其他学者在长白山地区(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭地区(赵玉芳, 2011 )以及俄罗斯远东地区(Ishikawa et al ., 1999 )等阔叶红松林主要分布区的研究结果相一致, 说明阔叶红松林干扰事件不只是局限在研究区域内的小范围事件, 而且可能为区域尺度上(整个阔叶红松林分布区)的大范围干扰事件。红松和臭冷杉干扰事件可能受区域性气候模态周期影响(图4 ), 进一步说明阔叶红松林干扰事件是区域尺度上的大范围干扰事件。小兴安岭丰林自然保护区臭冷杉径向生长与AMO和太平洋年代际振荡(Pacific Decadal Oscillation, PDO)有直接关系, AMO、PDO的相位转变可能是谷地臭冷杉衰退的原因(姚启超和王晓春, 2013 )。长白山安图(徐海等, 2002 )和小兴安岭五营(尹红等, 2009 )红松树轮重建的降水或气温序列中也发现了与ENSO等区域性气候模态对应的周期, 也说明红松径向生长受区域性气候模态的影响。大尺度气候波动虽然不会直接影响某一区域树木的生长, 但能通过影响局地气候间接影响树木生长(张先亮等, 2010 )。当这种影响超过一定的阈值时, 势必会导致树木出现生长衰退或死亡。因此, 外力作用下更易形成林窗, 造成周围树体生长释放。另外, 区域气候模态(尤其是ENSO)与东亚季风关系密切(徐袁和钱维宏, 2003 ), 由此形成的风暴更是触发林窗形成的关键原因。
3.3 影响干扰的因素
本研究发现风干扰是小兴安岭原始阔叶红松林树木生长释放的一个主要因素, 风干扰是该区阔叶红松林干扰的主要类型, 这与其他学者在长白山(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭(王晓春和赵玉芳, 2011 ; 赵玉芳, 2011 )及俄罗斯远东(Ishikawa et al ., 1999 )等地阔叶红松林干扰研究的结果一致。风干扰造成树体倾倒(尤其是林冠层乔木, 如红松)形成林窗(图9 ), 林窗的出现解除了周围树木生长的竞争, 林窗边缘木将出现明显的生长释放(王晓春和赵玉芳, 2011 )。例如, 研究区春季(4-5月)多风且风速较大, 春季最大风速平均值前10个最大年份分别为1971、1976、1987、1989-1990、1994-1996和1998-1999年, 这些年份当年或后几年红松生长变化百分率均出现明显增加的趋势。臭冷杉由于是喜阴树种且多处于林冠下层, 生长释放不太明显。此外, 林窗内形成木数量、腐烂等级以及林窗边缘木生长释放特征, 很好地证明了森林遭大风袭击后摧毁了部分林冠层树木, 造成林冠参差不齐。林窗形成后, 该区对风的抵抗力便会大大削弱, 林分很容易发生持续的风倒, 产生多林窗潮(multiple-gap episodes)(阳含熙和谢海生, 1994 ; 臧润国和徐化成, 1998 )。
图9 风倒木照片。A, 水曲柳掘根风倒。B, 红松干中折断。
Fig. 9 Photos of windthrow. A, Fraxinus mandschurica tree uprooted by the wind. B, Pinus koraiensis tree broken on trunk.
风干扰是阔叶红松林干扰的主要因素之一(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 ), 然而并不意味着风速越大, 形成林窗的数量就越多, 生长释放就越明显。风只是林窗干扰发生的一个外因, 林窗干扰是否发生以及干扰的强弱还取决于树体自身的生长情况及其生存的环境条件。此外, 异常气温也在很大程度上影响着该区树木的生长释放, 这一结果与王晓春和赵玉芳(2011)的研究结果一致。异常气温影响研究区的干旱化现象(贺伟等, 2013 ), 导致该区树木生长衰退或死亡, 对外界干扰的抵抗能力降低或丧失, 在外力作用下更易形成倒木, 进而造成林窗边缘树木发生释放。
中等释放与主要释放的主要差别在于干扰强度不同, 仅从生长释放强度上无法判断这些释放的具体干扰因子。但释放强度不同可能有两个原因: 其一是树木的竞争程度, 如果释放前树木竞争较为激烈, 那么一旦竞争解除, 生长释放就会较大; 其二是干扰强度的大小, 干扰较大会使释放后的树木获得更多资源, 生长释放也就较大。如不考虑竞争因素, 主要释放可能是由异常气候(如台风等)产生的较大干扰造成的, 而中等释放除干扰外(干扰产生一定程度的林窗开放), 还可能与限制树木生长因素的较大波动有关。
红松和臭冷杉干扰事件的周期性中, 存在11 a的极显著(p < 0.01)太阳活动周期, 而且红松生长变化百分率与太阳黑子异常值呈显著负相关(p < 0.05) (图10 ), 也表明太阳活动很大程度上影响着该区树木的生长释放。太阳黑子爆发年与红松、臭冷杉生长变化百分率的SEA分析(图11 )表明: 小兴安岭地区红松树轮记录的生长变化百分率由太阳黑子爆发前2-4 a的显著(p < 0.05)负相关, 转变为爆发后1-4 a的正相关; 臭冷杉叠加事件分析的结果与红松基本一致, 但未达到显著水平。太阳活动可通过影响局地气候间接影响树木的生长(徐海等, 2002 ; 尹红等, 2009 )。太阳活动剧烈时, 气温异常值随之增大或减小, 风速异常值也随之增大或减小, 太阳活动通过影响局地的气温和风速变化(表2 )间接影响该区树木生长, 具体表现为造成树木衰退或死亡, 抗干扰能力降低, 最终导致树体间竞争解除或减弱, 产生生长释放。太阳活动不仅直接影响着区域尺度下的气温、降水和风速变化, 还在很大程度上影响着区域性气候模态(赵亮等, 2011 )。因此, 太阳活动通过影响气温、降水和风速等局地气候以及其他大尺度气候模态干扰该地区森林动态, 是小兴安岭典型阔叶红松林可能存在的干扰机制。
图10 红松和臭冷杉生长变化百分率与月太阳黑子数异常值的相关性。柱体的灰色表示显著相关(p < 0.05)。P表示前一年, 如”P8”指前一年8月。
Fig. 10 Correlation coefficients between percentage growth changes in Pinus koraiensis and Abies nephrolepis and monthly sunspot number anomalous value. The gray color of columns indicate significant correlations (p < 0.05). The letter P in the horizontal axis represents the previous year. For instance, “P8” represents the previous August.
图11 太阳黑子爆发与红松(A)和臭冷杉(B)生长变化百分率的叠加事件分析。灰色柱表示显著相关(p < 0.05)。
Fig. 11 Superimposed events analysis between sunspot eruption and percentage growth changes in Pinus koraiensis (A) and Abies nephrolepis (B). The gray columns indicate significant correlations (p < 0.05).
4 结论
本文用红松和臭冷杉树轮重建了小兴安岭典型阔叶红松林的干扰历史, 红松和臭冷杉林的干扰事件有差别但也有重叠。红松林窗干扰主要出现在1733-1738、1748-1752、1769-1771、1798-1801、1827-1833、1841-1844、1935-1939和1968-1973年间, 臭冷杉林窗主要干扰在1889-1904、1932-1938、1947-1973和1986-2005年间。红松林窗干扰存在2.0 a、3.5 a、3.8 a、7.3-7.9 a和9.1-18.2 a的显著周期, 而臭冷杉林窗干扰存在3.5-3.6 a、7.5-48.8 a和65-85 a的显著周期。风干扰是小兴安岭典型阔叶红松林干扰释放的主要因子, 异常气温也影响该地区树木生长释放。太阳活动通过改变局地风速、气温、降水等气候因子以及其他大尺度气候模态影响林窗动态可能是阔叶红松林的干扰机制之一。阔叶红松林的干扰历史既存在小尺度的局域事件, 也有区域尺度上的大范围干扰事件。因此, 林窗动态存在较大的变异性。
致谢 感谢黑龙江凉水国家级自然保护区和东北林业大学凉水实验林场提供的野外基础支持。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Black BA Abrams MD 2004 ). Development and application of boundary-line release criteria.
Dendrochronologia , 22 , 31 -42 .
[本文引用: 2]
[2]
Bond-Lamberty B Rocha AV Calvin K Holmes B Wang CK Goulden ML 2014 ). Disturbance legacies and climate jointly drive tree growth and mortality in an intensively studied boreal forest.
Global Change Biology , 20 , 216 -227 .
[本文引用: 1]
[3]
Brown KJ Giesecke T 2014 ). Holocene fire disturbance in the boreal forest of central Sweden.
Boreas , 43 , 639 -651 .
[本文引用: 1]
[4]
Cook ER Kairiuktis LA 1990 ). Methods of Dendrochronology: Applications in the Environmental Sciences. Kluwer Academic Publishers
, London.
[本文引用: 1]
[5]
Cooke BJ Lorenzetti F 2006 ). The dynamics of forest tent caterpillar outbreaks in Québec, Canada.
Forest Ecology and Management , 226 , 110 -121 .
[本文引用: 1]
[6]
Feng XH Cheng RM Xiao WF Wang XR Wang RL 2011 ). The application of tree-ring on forest disturbance history reconstruction.
Acta Ecologica Sinica , 31 , 3215 -3222 .(in Chinese with English abstract)
[本文引用: 2]
[封晓辉 , 程瑞梅 , 肖文发 , 王晓荣 , 王瑞丽 (2011 ). 树木年轮在干扰历史重建中的应用
. 生态学报 , 31 , 3215 -3222 .]
[本文引用: 2]
[7]
Han L Wang HZ Chen JL Yu J 2011 ). Disturbance regimes and gaps characteristics of the desert riparian forest at the middle reaches of Tarim River.
Acta Ecologica Sinica , 31 , 4699 -4708 .(in Chinese with English abstract)
[本文引用: 3]
[韩路 , 王海珍 , 陈加利 , 于军 (2011 ). 塔里木荒漠河岸林干扰状况与林隙特征
. 生态学报 , 31 , 4699 -4708 .]
[本文引用: 3]
[8]
He W Bu RC Xiong ZP Hu YM 2013 ). Characteristics of temperature and precipitation in Northeastern China from 1961 to 2005.
Acta Ecologica Sinica , 33 , 519 -531 .(in Chinese with English abstract)
[本文引用: 1]
[贺伟 , 布仁仓 , 熊在平 , 胡远满 (2013 ). 1961-2005年东北地区气温和降水变化趋势
. 生态学报 , 33 , 519 -531 .]
[本文引用: 1]
[9]
Holmes RL 1983 ). Computer-assisted quality control in tree-ring dating and measurement.
Tree-Ring Bulletin , 43 , 69 -78 .
[本文引用: 1]
[10]
Hong T Bai SB Wang J Zhang ZG 2012 ). Reconstruct the activity years of Jiufangshan landslide by means of tree-rings.
Journal of Mountain Science , 30 , 57 -64 .(in Chinese with English abstract)
[本文引用: 1]
[洪婷 , 白世彪 , 王建 , 张志刚 (2012 ). 利用树轮重建九房山滑坡活动年份
. 山地学报 , 30 , 57 -64 .]
[本文引用: 1]
[11]
Hou XY Han JX Xie HS 2000 ). Disturbance regimes in the Korean-pine forests in Changbai Mountain.
Acta Ecologica Sinica , 20 , 409 -411 .(in Chinese with English abstract)
[本文引用: 3]
[侯向阳 , 韩进轩 , 谢海生 (2000 ). 长白山红松林干扰节律研究
. 生态学报 , 20 , 409 -411 .]
[本文引用: 3]
[12]
Ishikawa Y Krestov PV Namikawa K 1999 ). Disturbance history and tree establishment in old-growth Pinus koraiensis-hardwood forests in the Russian Far East.
Journal of Vegetation Science , 10 , 439 -448 .
[本文引用: 3]
[13]
Li JW 1997 ). Korean Pine Mixed Forest Ecology and Management . Northeast Forestry University Press , Harbin .(in Chinese)
[本文引用: 1]
[李景文 (1997 ). 红松混交林生态与经营 . 东北林业大学出版社 , 哈尔滨 .]
[本文引用: 1]
[14]
Liu SC Wang JH Duan WB Chen LX Wang LX Du S Zhao JH 2013 ). Gap characteristics in the mixed broad-leaved Korean pine forest in Xiaoxing’an Mountains.
Acta Ecologica Sinica , 33 , 5234 -5244 .(in Chinese with English abstract)
[本文引用: 3]
[刘少冲 , 王敬华 , 段文标 , 陈立新 , 王丽霞 , 杜珊 , 赵健慧 (2013 ). 小兴安岭阔叶红松混交林林隙特征
. 生态学报 , 33 , 5234 -5244 .]
[本文引用: 3]
[15]
Liu YY Jin GZ 2009 ). Influence of topography on coarse woody debris in a mixed broadleaved-Korean pine forest in Xiaoxing’an Mountains, China.
Acta Ecologica Sinica , 29 , 1398 -1407 .(in Chinese with English abstract)
[刘妍妍 , 金光泽 (2009 ). 地形对小兴安岭阔叶红松(Pinus koraiensis)林粗木质残体分布的影响
. 生态学报 , 29 , 1398 -1407 .]
[16]
Nowacki GJ Abrams MD 1997 ). Radial-growth averaging criteria for reconstructing disturbance histories from resettlement-origin oaks.
Ecological Monographs , 67 , 225 -249 .
[本文引用: 1]
[17]
Qiao DF 2005 ). Study on Disturbance History of Korean Pine Forest in Dunhua Region
. PhD dissertation, Beijing Normal University, Beijing. (in Chinese with English abstract)
[本文引用: 3]
[乔佃锋 (2005 ). 敦化地区红松林干扰历史研究
. 博士学位论文, 北京师范大学, 北京 .]
[本文引用: 3]
[18]
Rubino DL McCarthyz BC 2004 ). Comparative analysis of dendroecological methods used to assess disturbance events.
Dendrochronologia , 21 , 97 -115 .
[本文引用: 4]
[19]
Shen ZH Wang GF Li XD 2001 ). Gap related disturbance in mixed mountain forests at Mt. Dalaoling in the Three Gorges II. Topographic patterns.
Acta Phytoecologica Sinica , 26 , 149 -156 .(in Chinese with English abstract)
[本文引用: 1]
[沈泽昊 , 王功芳 , 李道兴 (2001 ). 三峡大老岭山地常绿落叶阔叶混交林林隙干扰研究II
. 林隙干扰的地形格局. 植物生态学报 , 26 , 149 -156 .]
[本文引用: 1]
[20]
Smith KT 2008 ). An organismal view of dendrochronology.
Dendrochronologia , 26 , 185 -193 .
[本文引用: 1]
[21]
Stokes MA Smiley TL 1996 ). An Introduction to Tree-ring Dating . The University of Arizona Press , Tucson, USA .
[本文引用: 1]
[22]
Tie YB Malik I Owczarek P 2014 ). Dendrochronological dating of debris flow historical events in high mountain area—Take Daozao debris flow as an example.
Mountain Research , 32 , 226 -232 .(in Chinese with English abstract)
[本文引用: 1]
[铁永波 , Malik I Owczarek P 2014 ). 树木年代学在高寒山区泥石流历史事件重建中的应用——以磨西河流域倒灶沟为例
. 山地学报 , 32 , 226 -232 .]
[本文引用: 1]
[23]
Vitousek PM Mooney HA Lubchenco J Melillo JM 1997 ). Human domination of Earth’s ecosystems.
Science , 277 , 494 -499 .
[本文引用: 1]
[24]
Wang XC Brown PM Zhang YN Song LP 2011 ). Imprint of the Atlantic multidecadal oscillation on tree-ring widths in Northeastern Asia since 1568.
PLoS ONE , 6 , e22740 .
[本文引用: 1]
[25]
Wang XC Zhao YF 2011 ). Growth release determination and interpretation of Korean pine and Koyama spruce in Shengshan National Nature Reserve, Heilongjiang Province, China.
Acta Ecologica Sinica , 31 , 1230 -1239 .(in Chinese with English abstract)
[本文引用: 8]
[王晓春 , 赵玉芳 (2011 ). 黑河胜山国家自然保护区红松和红皮云杉生长释放判定及解释
. 生态学报 , 31 , 1230 -1239 .]
[本文引用: 8]
[26]
Wei XH 2010 ). Disturbance ecology: A science needing attention.
Acta Agriculturae Universitatis Jiangxiensis , 32 , 1032 -1039 .(in Chinese with English abstract)
[本文引用: 1]
[魏晓华 (2010 ). 干扰生态学: 一门必须重视的学科
. 江西农业大学学报 , 32 , 1032 -1039 .]
[本文引用: 1]
[27]
Xia B Deng F Zhou K Wang Q Dong X 2001 ). A case study on the verification of density of forest disturbance through tree-ring analysis.
Journal of Plant Resources and Environment , 10 (1 ), 1 -6 .(in Chinese with English abstract)
[本文引用: 2]
[夏冰 , 邓飞 , 周康 , 王谦 , 董旭 (2001 ). 树轮分析用于森林干扰强度推测的实例研究
. 植物资源与环境学报 , 10 (1 ), 1 -6 .]
[本文引用: 2]
[28]
Xia B He SA Lan T Deng F Yao G 1997 ). Growth release and disturbance of conifer trees in subalpine spruce-fir mixed forest.
Journal of Plant Resources and Environment , 6 (1 ), 1 -8 .(in Chinese with English abstract)
[本文引用: 3]
[夏冰 , 贺善安 , 兰涛 , 邓飞 , 姚淦 (1997 ). 亚高山云冷杉混交林树木生长释放与干扰分析
. 植物资源与环境 , 6 (1 ), 1 -8 .]
[本文引用: 3]
[29]
Xia B Lan T He SA Deng F Yang FC 1995 ). Estimation of canopy disturbance and dynamics of a stand of Pinus taiwanensis and broadleaf trees in the west Tianmu Mountain.
Journal of Plant Resources and Environment , 4 (3 ), 15 -20 .(in Chinese with English abstract)
[本文引用: 1]
[夏冰 , 兰涛 , 贺善安 , 邓飞 , 杨逢春 (1995 ). 西天目山黄山松阔叶林的冠层干扰与动态推测
. 植物资源与环境 , 4 (3 ), 15 -20 .]
[本文引用: 1]
[30]
Xu H Hong YT Zhu YX Liu GS 2002 ). Information on climate change recorded in δ13 C and δ18 O series of Pinus koraiensis tree ring cellulose in Antu area.
Geology- Geochemistry , 30 (2 ), 59 -65 .(in Chinese with English abstract)
[本文引用: 2]
[徐海 , 洪业汤 , 朱咏煊 , 刘广深 (2002 ). 安图红松树轮稳定δ13 C、δ18 O序列记录的气候变化信息
. 地质地球化学 , 30 (2 ), 59 -65 .]
[本文引用: 2]
[31]
Xu LN Jin GZ 2012 ). Species composition and community structure of a typical mixed broadleaved-Korean pine (Pinus koraiensis) forest plot in Liangshui Nature Reserve, Northeast China.
Biodiversity Science , 20 , 470 -481 .(in Chinese with English abstract)
[本文引用: 1]
[徐丽娜 , 金光泽 (2012 ). 小兴安岭凉水典型阔叶红松林动态监测样地: 物种组成与群落结构
. 生物多样性 , 20 , 470 -481 .]
[本文引用: 1]
[32]
Xu Y Qian WH 2003 ). Research on East Asian summer monsoon: A Review.
Acta Geographica Sinica , 58 , 138 -146 .(in Chinese with English abstract)
[本文引用: 1]
[徐袁 , 钱维宏 (2003 ). 东亚季风边缘活动带研究综述
. 地理学报 , 58 , 138 -146 .]
[本文引用: 1]
[33]
Yang HX Xie HS 1994 ). Study on the reconstruction of disturbance history of Pinus koraiensis mixed forest in Changbai Mountain.
Acta Phytoecologica Sinica , 18 , 201 -208 .(in Chinese with English abstract)
[本文引用: 7]
[阳含熙 , 谢海生 (1994 ). 长白山红松混交林干扰历史的重构研究
. 植物生态学报 , 18 , 201 -208 .]
[本文引用: 7]
[34]
Yao QC Wang XC 2013 ). Climate-growth relationships of Abies nephrolepis and its connection with large-scale climate change in Xiaoxing’an Mountains, northeastern China.
Journal of Beijing Forestry University , 35 (2 ), 30 -38 .(in Chinese with English abstract)
[本文引用: 2]
[姚启超 , 王晓春 (2013 ). 小兴安岭不同海拔臭冷杉年轮-气候关系及大尺度气候影响
. 北京林业大学学报 , 35 (2 ), 30 -38 .]
[本文引用: 2]
[35]
Yin H Guo PW Liu HB Huang L Yu HM Guo SY Wang F 2009 ). Reconstruction of the October mean temperature since 1796 at Wuying from tree ring data.
Advances in Climate Change Research , 5 , 18 -23 .(in Chinese with English abstract)
[本文引用: 2]
[尹红 , 郭品文 , 刘洪滨 , 黄磊 , 于宏敏 , 国世友 , 王芳 (2009 ). 利用树轮重建小兴安岭五营1796年以来的温度变化
. 气候变化研究进展 , 5 , 18 -23 .]
[本文引用: 2]
[36]
Zang RG Xu HC 1998 ). Advances in forest gap disturbance research.
Scientia Silvae Sinicae , 34 (1 ), 90 -98 .(in Chinese with English abstract)
[本文引用: 2]
[臧润国 , 徐化成 (1998 ). 林隙(GAP)干扰研究进展
. 林业科学 , 34 (1 ), 90 -98 .]
[本文引用: 2]
[37]
Zang RG Yang YC Liu JY Yu SX Yang XS 1999 ). Gaps and their natural disturbance regimes in the tropical montane rain forest of Hainan Island.
Scientia Silvae Sinicae , 35 (1 ), 2 -8 .(in Chinese with English abstract)
[本文引用: 3]
[臧润国 , 杨彦承 , 刘静艳 , 余世孝 , 杨秀森 (1999 ). 海南岛热带山地雨林林隙及其自然干扰特征
. 林业科学 , 35 (1 ), 2 -8 .]
[本文引用: 3]
[38]
Zhang L Wang XC 2013 ). Fire history reconstruction of a larch forest in Pangu of Daxing’an Mountains, China. Journal of Northeast Normal University (Natural Sciences Edition)
, 45 (1 ), 117 -122 .(in Chinese with English abstract)
[本文引用: 1]
[张磊 , 王晓春 (2013 ). 基于树轮火疤重建大兴安岭盘古兴安落叶松林近200年火灾历史
. 东北师范大学学报(自然科学版) , 45 (1 ), 117 -122 .]
[本文引用: 1]
[39]
Zhang XL Cui MX Ma YJ Wu T Chen ZJ Ding WH 2010 ). Larix gmelinii tree-ring width chronology and its responses to climate change in Kuduer, Daxing’an Mountains.
Chinese Journal of Applied Ecology , 21 , 2501 -2507 .(in Chinese with English abstract)
[本文引用: 1]
[张先亮 , 崔明星 , 马艳军 , 吴涛 , 陈振举 , 丁玮航 (2010 ). 大兴安岭库都尔地区兴安落叶松年轮宽度年表及其与气候变化的关系
. 应用生态学报 , 21 , 2501 -2507 .]
[本文引用: 1]
[40]
Zhang Y Drobyshev I Gao LS Zhao XH Bergeron Y 2014 ). Disturbance and regeneration dynamics of a mixed Korean pine dominated forest on Changbai Mountain, North- Eastern China.
Dendrochronologia , 32 , 21 -31 .
[本文引用: 3]
[41]
Zhao L Xu Y Wang JS Ding YH Xiao ZN 2011 ). Progress in studies on the influence of solar activity on climate change during the last 100 years.
Advances in Meteorological Science and Technology , 1 (4 ), 37 -48 .(in Chinese with English abstract)
[本文引用: 1]
[赵亮 , 徐影 , 王劲松 , 丁一汇 , 肖子牛 (2011 ). 太阳活动对近百年气候变化的影响研究进展
. 气象科技进展 , 1 (4 ), 37 -48 .]
[本文引用: 1]
[42]
Zhao YF 2011 ). Growth Release Determination and Interpretation of Korean Pine in Heilongjiang Province, China. Master degree dissertation, Northeast Forestry University
, Harbin. (in Chinese with English abstract)
[本文引用: 2]
[赵玉芳 (2011 ). 黑龙江省红松生长释放判定及解释
. 硕士学位论文, 东北林业大学, 哈尔滨.]
[本文引用: 2]
[43]
Zhu HF Xu P Shao XM Luo HJ 2013 ). Little Ice Age glacier fluctuations reconstructed for the southeastern Tibetan Plateau using tree rings.
Quaternary International , 283 , 134 -138 .
[本文引用: 1]
[44]
Zhu JJ Liu SR 2007 ). A review on Disturbance Ecology of Forest . China Forestry Publishing House , Beijing .(in Chinese)
[本文引用: 1]
[朱教君 , 刘世荣 (2007 ). 森林干扰生态研究 . 中国林业出版社 , 北京 .]
[本文引用: 1]
Development and application of boundary-line release criteria.
2
2004
... 林窗干扰是自然界中最常见的一种森林干扰(臧润国和徐化成, 1998 ; 臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).林窗干扰发生后, 周围树木由于竞争去除而产生生长释放(王晓春和赵玉芳, 2011 ), 并且这种释放可以持续多年.通常将大于一定阈值且持续5 a或10 a以上的生长释放视为“真正释放”, 并将生长释放开始的时间视为干扰的发生时间(Black & Abrams, 2004 ; Rubino & McCarthyz, 2004 ; 王晓春和赵玉芳, 2011 ).确定生长释放的发生时间是重建森林干扰历史的最基本过程(Black & Abrams, 2004 ), 林窗形成时间常以林窗内幼苗更新时间(刘少冲等, 2013 )、倒木倾倒时间(用腐烂等级确定, 韩路等, 2011 )以及林窗周边树木生长释放时间(阳含熙和谢海生, 1994 ; 夏冰等, 1995 )等确定.本研究林窗干扰时间的确定主要用树木生长释放来判断, 同时还参考了林窗内的幼苗更新及倒木腐烂等级, 但有些林窗更新的非均一性以及林窗内倒木腐烂情况比较复杂也影响林窗形成时间的确定. ...
... ).确定生长释放的发生时间是重建森林干扰历史的最基本过程(Black & Abrams, 2004 ), 林窗形成时间常以林窗内幼苗更新时间(刘少冲等, 2013 )、倒木倾倒时间(用腐烂等级确定, 韩路等, 2011 )以及林窗周边树木生长释放时间(阳含熙和谢海生, 1994 ; 夏冰等, 1995 )等确定.本研究林窗干扰时间的确定主要用树木生长释放来判断, 同时还参考了林窗内的幼苗更新及倒木腐烂等级, 但有些林窗更新的非均一性以及林窗内倒木腐烂情况比较复杂也影响林窗形成时间的确定. ...
Disturbance legacies and climate jointly drive tree growth and mortality in an intensively studied boreal forest.
1
2014
... 随着频繁的人类活动和异常自然干扰的发生, 森林生态系统结构与功能正受到严重的威胁(Vitousek et al ., 1997 ).干扰生态学已成为生态学研究最活跃的领域之一(朱教君和刘世荣, 2007 ; Bond-Lamberty et al ., 2014 ), 其重点是从森林群落的干扰历史和林窗(隙)动态两个方面对中小规模干扰的特征及影响开展研究, 且有关林窗动态的研究相对较多(臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).重建干扰历史是植物群落演替和更新研究的基础, 越来越受到重视.森林群落的干扰动态可以通过固定样地的长期观测获得(Cooke & Lorenzetti, 2006 ), 但对于缺乏长期观测的森林群落, 人们无法得知其干扰历史.树木年轮是树木生长的历史记录(Smith, 2008 ), 其宽度、密度、同位素含量以及解剖结构等记载了树木逐年的生长信息及内、外部环境变化的影响(Cook & Kairiuktis, 1990 ).树轮具有生长时间长、定年准确、材料广泛、容易获取等优点, 是研究树木生长环境变化的重要手段, 也是重建森林群落动态的重要材料. ...
Holocene fire disturbance in the boreal forest of central Sweden.
1
2014
... 国外已有一些研究利用树木年轮重建森林干扰历史(Nowacki & Abrams, 1997 ; Ishikawa et al ., 1999 ; Rubino & McCarthyz, 2004 ; Brown & Giesecke, 2014 ), 而国内这方面研究起步较晚, 且相对较少.20世纪90年代初, 阳含熙和谢海生(1994)最先利用树木生长释放和抑制对长白山红松(Pinus koraiensis )混交林干扰历史进行了重建, 并指出该方法是以风作为主要干扰类型, 重建干扰历史最准确、最可靠、提供信息量最大的方法; 侯向阳等(2000)研究长白山红松林干扰节律时, 也得到了类似的结论.乔佃锋(2005)利用该方法(阳含熙和谢海生, 1994 )重建了长白山敦化地区阔叶红松林的干扰历史, 发现该区红松林干扰以小规模的风干扰为主, 1980年出现了明显的干扰峰, 干扰峰之间60年的时间间隔正好与红松的更替世代相吻合.王晓春和赵玉芳(2011)发现前一年径向生长对生长释放事件的判定很重要, 小兴安岭北端黑河地区的红松和红皮云杉(Picea koraiensis )的生长释放可能受林窗产生、微环境变化以及温度和降水等气候因素影响.夏冰等(1995)在天目山建立了生长释放和抑制的判定标准, 并重建了黄山松(Pinus taiwanensis )阔叶林树冠的干扰历史; 同时, 还对不同树种年轮宽度的生长释放判别方法进行了研究, 并做了重建干扰强度的尝试(夏冰等, 1997 , 2001 ).此外, 不同学者还利用树轮生态学方法对火灾(张磊和王晓春, 2013 )、泥石流(铁永波等, 2014 )、山体滑坡(洪婷等, 2012 )和冰川活动(Zhu et al ., 2013 )等干扰进行了重建. ...
Methods of Dendrochronology: Applications in the Environmental Sciences. Kluwer Academic Publishers
1
1990
... 随着频繁的人类活动和异常自然干扰的发生, 森林生态系统结构与功能正受到严重的威胁(Vitousek et al ., 1997 ).干扰生态学已成为生态学研究最活跃的领域之一(朱教君和刘世荣, 2007 ; Bond-Lamberty et al ., 2014 ), 其重点是从森林群落的干扰历史和林窗(隙)动态两个方面对中小规模干扰的特征及影响开展研究, 且有关林窗动态的研究相对较多(臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).重建干扰历史是植物群落演替和更新研究的基础, 越来越受到重视.森林群落的干扰动态可以通过固定样地的长期观测获得(Cooke & Lorenzetti, 2006 ), 但对于缺乏长期观测的森林群落, 人们无法得知其干扰历史.树木年轮是树木生长的历史记录(Smith, 2008 ), 其宽度、密度、同位素含量以及解剖结构等记载了树木逐年的生长信息及内、外部环境变化的影响(Cook & Kairiuktis, 1990 ).树轮具有生长时间长、定年准确、材料广泛、容易获取等优点, 是研究树木生长环境变化的重要手段, 也是重建森林群落动态的重要材料. ...
The dynamics of forest tent caterpillar outbreaks in Québec, Canada.
1
2006
... 随着频繁的人类活动和异常自然干扰的发生, 森林生态系统结构与功能正受到严重的威胁(Vitousek et al ., 1997 ).干扰生态学已成为生态学研究最活跃的领域之一(朱教君和刘世荣, 2007 ; Bond-Lamberty et al ., 2014 ), 其重点是从森林群落的干扰历史和林窗(隙)动态两个方面对中小规模干扰的特征及影响开展研究, 且有关林窗动态的研究相对较多(臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).重建干扰历史是植物群落演替和更新研究的基础, 越来越受到重视.森林群落的干扰动态可以通过固定样地的长期观测获得(Cooke & Lorenzetti, 2006 ), 但对于缺乏长期观测的森林群落, 人们无法得知其干扰历史.树木年轮是树木生长的历史记录(Smith, 2008 ), 其宽度、密度、同位素含量以及解剖结构等记载了树木逐年的生长信息及内、外部环境变化的影响(Cook & Kairiuktis, 1990 ).树轮具有生长时间长、定年准确、材料广泛、容易获取等优点, 是研究树木生长环境变化的重要手段, 也是重建森林群落动态的重要材料. ...
树木年轮在干扰历史重建中的应用
2
2011
... 生长释放判定方法的选择及判定标准的确定是树轮干扰历史重建的关键.判定方法和标准不同, 得出的生长释放结果亦不同(Rubino & McCarthyz, 2004 ), 每一种方法都有各自的优缺点及使用范围(封晓辉等, 2011 ).在实践中, 通过主观判断去选择生长释放判定方法及判定标准, 往往会因主观原因放大或弱化干扰事件.因此, 在判定方法及判定标准选择时, 在充分考虑研究需要的同时, 还需结合研究地点及环境条件等合理选择(阳含熙和谢海生, 1994 ; 夏冰等, 1997 ; 王晓春和赵玉芳, 2011 ).为了保留更多的干扰信号, 本文选用生长释放标准曲线法(王晓春和赵玉芳, 2011 ), 即生长变化率超过一定的标准便可视为生长释放发生, 该方法是目前应用最多的方法(封晓辉等, 2011 ).本研究中林窗木与非林窗木的生长变化百分率有很强的一致性(仅局部存在部分差异), 说明林窗木生长变化百分率序列中包含了部分非干扰信号.通过生长释放判定得到的生长释放中必定会有部分噪音即非干扰信号, 即并不是所有的释放(包括中等释放和主要释放)都能代表林窗干扰(王晓春和赵玉芳, 2011 ), 因此主要释放或部分主要释放才能代表真正意义上的干扰. 生长变化百分率差值图(图1 )与干扰年表(图2 , 图3 )谷、峰值具有很强的一致性, 且干扰年表在一定程度上对生长变化百分率差值图中不太明显的峰值具有局部放大作用, 经局部放大的部分信号可能是噪音的主要来源.因此, 通过林窗边缘木生长释放判定得出的干扰年表, 能够很好地反映该区阔叶红松林的干扰历史, 且能将一些不太明显的微小干扰包括其中. ...
... ), 即生长变化率超过一定的标准便可视为生长释放发生, 该方法是目前应用最多的方法(封晓辉等, 2011 ).本研究中林窗木与非林窗木的生长变化百分率有很强的一致性(仅局部存在部分差异), 说明林窗木生长变化百分率序列中包含了部分非干扰信号.通过生长释放判定得到的生长释放中必定会有部分噪音即非干扰信号, 即并不是所有的释放(包括中等释放和主要释放)都能代表林窗干扰(王晓春和赵玉芳, 2011 ), 因此主要释放或部分主要释放才能代表真正意义上的干扰. 生长变化百分率差值图(图1 )与干扰年表(图2 , 图3 )谷、峰值具有很强的一致性, 且干扰年表在一定程度上对生长变化百分率差值图中不太明显的峰值具有局部放大作用, 经局部放大的部分信号可能是噪音的主要来源.因此, 通过林窗边缘木生长释放判定得出的干扰年表, 能够很好地反映该区阔叶红松林的干扰历史, 且能将一些不太明显的微小干扰包括其中. ...
树木年轮在干扰历史重建中的应用
2
2011
... 生长释放判定方法的选择及判定标准的确定是树轮干扰历史重建的关键.判定方法和标准不同, 得出的生长释放结果亦不同(Rubino & McCarthyz, 2004 ), 每一种方法都有各自的优缺点及使用范围(封晓辉等, 2011 ).在实践中, 通过主观判断去选择生长释放判定方法及判定标准, 往往会因主观原因放大或弱化干扰事件.因此, 在判定方法及判定标准选择时, 在充分考虑研究需要的同时, 还需结合研究地点及环境条件等合理选择(阳含熙和谢海生, 1994 ; 夏冰等, 1997 ; 王晓春和赵玉芳, 2011 ).为了保留更多的干扰信号, 本文选用生长释放标准曲线法(王晓春和赵玉芳, 2011 ), 即生长变化率超过一定的标准便可视为生长释放发生, 该方法是目前应用最多的方法(封晓辉等, 2011 ).本研究中林窗木与非林窗木的生长变化百分率有很强的一致性(仅局部存在部分差异), 说明林窗木生长变化百分率序列中包含了部分非干扰信号.通过生长释放判定得到的生长释放中必定会有部分噪音即非干扰信号, 即并不是所有的释放(包括中等释放和主要释放)都能代表林窗干扰(王晓春和赵玉芳, 2011 ), 因此主要释放或部分主要释放才能代表真正意义上的干扰. 生长变化百分率差值图(图1 )与干扰年表(图2 , 图3 )谷、峰值具有很强的一致性, 且干扰年表在一定程度上对生长变化百分率差值图中不太明显的峰值具有局部放大作用, 经局部放大的部分信号可能是噪音的主要来源.因此, 通过林窗边缘木生长释放判定得出的干扰年表, 能够很好地反映该区阔叶红松林的干扰历史, 且能将一些不太明显的微小干扰包括其中. ...
... ), 即生长变化率超过一定的标准便可视为生长释放发生, 该方法是目前应用最多的方法(封晓辉等, 2011 ).本研究中林窗木与非林窗木的生长变化百分率有很强的一致性(仅局部存在部分差异), 说明林窗木生长变化百分率序列中包含了部分非干扰信号.通过生长释放判定得到的生长释放中必定会有部分噪音即非干扰信号, 即并不是所有的释放(包括中等释放和主要释放)都能代表林窗干扰(王晓春和赵玉芳, 2011 ), 因此主要释放或部分主要释放才能代表真正意义上的干扰. 生长变化百分率差值图(图1 )与干扰年表(图2 , 图3 )谷、峰值具有很强的一致性, 且干扰年表在一定程度上对生长变化百分率差值图中不太明显的峰值具有局部放大作用, 经局部放大的部分信号可能是噪音的主要来源.因此, 通过林窗边缘木生长释放判定得出的干扰年表, 能够很好地反映该区阔叶红松林的干扰历史, 且能将一些不太明显的微小干扰包括其中. ...
塔里木荒漠河岸林干扰状况与林隙特征
3
2011
... 随着频繁的人类活动和异常自然干扰的发生, 森林生态系统结构与功能正受到严重的威胁(Vitousek et al ., 1997 ).干扰生态学已成为生态学研究最活跃的领域之一(朱教君和刘世荣, 2007 ; Bond-Lamberty et al ., 2014 ), 其重点是从森林群落的干扰历史和林窗(隙)动态两个方面对中小规模干扰的特征及影响开展研究, 且有关林窗动态的研究相对较多(臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).重建干扰历史是植物群落演替和更新研究的基础, 越来越受到重视.森林群落的干扰动态可以通过固定样地的长期观测获得(Cooke & Lorenzetti, 2006 ), 但对于缺乏长期观测的森林群落, 人们无法得知其干扰历史.树木年轮是树木生长的历史记录(Smith, 2008 ), 其宽度、密度、同位素含量以及解剖结构等记载了树木逐年的生长信息及内、外部环境变化的影响(Cook & Kairiuktis, 1990 ).树轮具有生长时间长、定年准确、材料广泛、容易获取等优点, 是研究树木生长环境变化的重要手段, 也是重建森林群落动态的重要材料. ...
... 林窗干扰是自然界中最常见的一种森林干扰(臧润国和徐化成, 1998 ; 臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).林窗干扰发生后, 周围树木由于竞争去除而产生生长释放(王晓春和赵玉芳, 2011 ), 并且这种释放可以持续多年.通常将大于一定阈值且持续5 a或10 a以上的生长释放视为“真正释放”, 并将生长释放开始的时间视为干扰的发生时间(Black & Abrams, 2004 ; Rubino & McCarthyz, 2004 ; 王晓春和赵玉芳, 2011 ).确定生长释放的发生时间是重建森林干扰历史的最基本过程(Black & Abrams, 2004 ), 林窗形成时间常以林窗内幼苗更新时间(刘少冲等, 2013 )、倒木倾倒时间(用腐烂等级确定, 韩路等, 2011 )以及林窗周边树木生长释放时间(阳含熙和谢海生, 1994 ; 夏冰等, 1995 )等确定.本研究林窗干扰时间的确定主要用树木生长释放来判断, 同时还参考了林窗内的幼苗更新及倒木腐烂等级, 但有些林窗更新的非均一性以及林窗内倒木腐烂情况比较复杂也影响林窗形成时间的确定. ...
... )、倒木倾倒时间(用腐烂等级确定, 韩路等, 2011 )以及林窗周边树木生长释放时间(阳含熙和谢海生, 1994 ; 夏冰等, 1995 )等确定.本研究林窗干扰时间的确定主要用树木生长释放来判断, 同时还参考了林窗内的幼苗更新及倒木腐烂等级, 但有些林窗更新的非均一性以及林窗内倒木腐烂情况比较复杂也影响林窗形成时间的确定. ...
塔里木荒漠河岸林干扰状况与林隙特征
3
2011
... 随着频繁的人类活动和异常自然干扰的发生, 森林生态系统结构与功能正受到严重的威胁(Vitousek et al ., 1997 ).干扰生态学已成为生态学研究最活跃的领域之一(朱教君和刘世荣, 2007 ; Bond-Lamberty et al ., 2014 ), 其重点是从森林群落的干扰历史和林窗(隙)动态两个方面对中小规模干扰的特征及影响开展研究, 且有关林窗动态的研究相对较多(臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).重建干扰历史是植物群落演替和更新研究的基础, 越来越受到重视.森林群落的干扰动态可以通过固定样地的长期观测获得(Cooke & Lorenzetti, 2006 ), 但对于缺乏长期观测的森林群落, 人们无法得知其干扰历史.树木年轮是树木生长的历史记录(Smith, 2008 ), 其宽度、密度、同位素含量以及解剖结构等记载了树木逐年的生长信息及内、外部环境变化的影响(Cook & Kairiuktis, 1990 ).树轮具有生长时间长、定年准确、材料广泛、容易获取等优点, 是研究树木生长环境变化的重要手段, 也是重建森林群落动态的重要材料. ...
... 林窗干扰是自然界中最常见的一种森林干扰(臧润国和徐化成, 1998 ; 臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).林窗干扰发生后, 周围树木由于竞争去除而产生生长释放(王晓春和赵玉芳, 2011 ), 并且这种释放可以持续多年.通常将大于一定阈值且持续5 a或10 a以上的生长释放视为“真正释放”, 并将生长释放开始的时间视为干扰的发生时间(Black & Abrams, 2004 ; Rubino & McCarthyz, 2004 ; 王晓春和赵玉芳, 2011 ).确定生长释放的发生时间是重建森林干扰历史的最基本过程(Black & Abrams, 2004 ), 林窗形成时间常以林窗内幼苗更新时间(刘少冲等, 2013 )、倒木倾倒时间(用腐烂等级确定, 韩路等, 2011 )以及林窗周边树木生长释放时间(阳含熙和谢海生, 1994 ; 夏冰等, 1995 )等确定.本研究林窗干扰时间的确定主要用树木生长释放来判断, 同时还参考了林窗内的幼苗更新及倒木腐烂等级, 但有些林窗更新的非均一性以及林窗内倒木腐烂情况比较复杂也影响林窗形成时间的确定. ...
... )、倒木倾倒时间(用腐烂等级确定, 韩路等, 2011 )以及林窗周边树木生长释放时间(阳含熙和谢海生, 1994 ; 夏冰等, 1995 )等确定.本研究林窗干扰时间的确定主要用树木生长释放来判断, 同时还参考了林窗内的幼苗更新及倒木腐烂等级, 但有些林窗更新的非均一性以及林窗内倒木腐烂情况比较复杂也影响林窗形成时间的确定. ...
1961-2005年东北地区气温和降水变化趋势
1
2013
... 风干扰是阔叶红松林干扰的主要因素之一(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 ), 然而并不意味着风速越大, 形成林窗的数量就越多, 生长释放就越明显.风只是林窗干扰发生的一个外因, 林窗干扰是否发生以及干扰的强弱还取决于树体自身的生长情况及其生存的环境条件.此外, 异常气温也在很大程度上影响着该区树木的生长释放, 这一结果与王晓春和赵玉芳(2011)的研究结果一致.异常气温影响研究区的干旱化现象(贺伟等, 2013 ), 导致该区树木生长衰退或死亡, 对外界干扰的抵抗能力降低或丧失, 在外力作用下更易形成倒木, 进而造成林窗边缘树木发生释放. ...
1961-2005年东北地区气温和降水变化趋势
1
2013
... 风干扰是阔叶红松林干扰的主要因素之一(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 ), 然而并不意味着风速越大, 形成林窗的数量就越多, 生长释放就越明显.风只是林窗干扰发生的一个外因, 林窗干扰是否发生以及干扰的强弱还取决于树体自身的生长情况及其生存的环境条件.此外, 异常气温也在很大程度上影响着该区树木的生长释放, 这一结果与王晓春和赵玉芳(2011)的研究结果一致.异常气温影响研究区的干旱化现象(贺伟等, 2013 ), 导致该区树木生长衰退或死亡, 对外界干扰的抵抗能力降低或丧失, 在外力作用下更易形成倒木, 进而造成林窗边缘树木发生释放. ...
Computer-assisted quality control in tree-ring dating and measurement.
1
1983
... 将野外采集的所有样芯带回实验室, 进行固定、干燥、打磨等预处理(Stokes & Smiley, 1996 ).在显微镜下用骨架图法进行交叉定年, 然后用年轮分析仪进行宽度测量, 精度为0.001 mm.最后, 用COFECHA程序来检验定年结果(Holmes, 1983 ), 根据软件提示在显微镜下检查并修订定年与测量中出现的误差, 以确保年轮的日历年代准确. ...
利用树轮重建九房山滑坡活动年份
1
2012
... 国外已有一些研究利用树木年轮重建森林干扰历史(Nowacki & Abrams, 1997 ; Ishikawa et al ., 1999 ; Rubino & McCarthyz, 2004 ; Brown & Giesecke, 2014 ), 而国内这方面研究起步较晚, 且相对较少.20世纪90年代初, 阳含熙和谢海生(1994)最先利用树木生长释放和抑制对长白山红松(Pinus koraiensis )混交林干扰历史进行了重建, 并指出该方法是以风作为主要干扰类型, 重建干扰历史最准确、最可靠、提供信息量最大的方法; 侯向阳等(2000)研究长白山红松林干扰节律时, 也得到了类似的结论.乔佃锋(2005)利用该方法(阳含熙和谢海生, 1994 )重建了长白山敦化地区阔叶红松林的干扰历史, 发现该区红松林干扰以小规模的风干扰为主, 1980年出现了明显的干扰峰, 干扰峰之间60年的时间间隔正好与红松的更替世代相吻合.王晓春和赵玉芳(2011)发现前一年径向生长对生长释放事件的判定很重要, 小兴安岭北端黑河地区的红松和红皮云杉(Picea koraiensis )的生长释放可能受林窗产生、微环境变化以及温度和降水等气候因素影响.夏冰等(1995)在天目山建立了生长释放和抑制的判定标准, 并重建了黄山松(Pinus taiwanensis )阔叶林树冠的干扰历史; 同时, 还对不同树种年轮宽度的生长释放判别方法进行了研究, 并做了重建干扰强度的尝试(夏冰等, 1997 , 2001 ).此外, 不同学者还利用树轮生态学方法对火灾(张磊和王晓春, 2013 )、泥石流(铁永波等, 2014 )、山体滑坡(洪婷等, 2012 )和冰川活动(Zhu et al ., 2013 )等干扰进行了重建. ...
利用树轮重建九房山滑坡活动年份
1
2012
... 国外已有一些研究利用树木年轮重建森林干扰历史(Nowacki & Abrams, 1997 ; Ishikawa et al ., 1999 ; Rubino & McCarthyz, 2004 ; Brown & Giesecke, 2014 ), 而国内这方面研究起步较晚, 且相对较少.20世纪90年代初, 阳含熙和谢海生(1994)最先利用树木生长释放和抑制对长白山红松(Pinus koraiensis )混交林干扰历史进行了重建, 并指出该方法是以风作为主要干扰类型, 重建干扰历史最准确、最可靠、提供信息量最大的方法; 侯向阳等(2000)研究长白山红松林干扰节律时, 也得到了类似的结论.乔佃锋(2005)利用该方法(阳含熙和谢海生, 1994 )重建了长白山敦化地区阔叶红松林的干扰历史, 发现该区红松林干扰以小规模的风干扰为主, 1980年出现了明显的干扰峰, 干扰峰之间60年的时间间隔正好与红松的更替世代相吻合.王晓春和赵玉芳(2011)发现前一年径向生长对生长释放事件的判定很重要, 小兴安岭北端黑河地区的红松和红皮云杉(Picea koraiensis )的生长释放可能受林窗产生、微环境变化以及温度和降水等气候因素影响.夏冰等(1995)在天目山建立了生长释放和抑制的判定标准, 并重建了黄山松(Pinus taiwanensis )阔叶林树冠的干扰历史; 同时, 还对不同树种年轮宽度的生长释放判别方法进行了研究, 并做了重建干扰强度的尝试(夏冰等, 1997 , 2001 ).此外, 不同学者还利用树轮生态学方法对火灾(张磊和王晓春, 2013 )、泥石流(铁永波等, 2014 )、山体滑坡(洪婷等, 2012 )和冰川活动(Zhu et al ., 2013 )等干扰进行了重建. ...
长白山红松林干扰节律研究
3
2000
... 小兴安岭地区阔叶红松林内红松干扰事件主要发生在18世纪30年代、19世纪10年代、19世纪30-40年代、20世纪20-30年代, 臭冷杉主要发生在19世纪90年代、20世纪30年代、20世纪50-60年代、20世纪90年代.该结果与其他学者在长白山地区(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭地区(赵玉芳, 2011 )以及俄罗斯远东地区(Ishikawa et al ., 1999 )等阔叶红松林主要分布区的研究结果相一致, 说明阔叶红松林干扰事件不只是局限在研究区域内的小范围事件, 而且可能为区域尺度上(整个阔叶红松林分布区)的大范围干扰事件.红松和臭冷杉干扰事件可能受区域性气候模态周期影响(图4 ), 进一步说明阔叶红松林干扰事件是区域尺度上的大范围干扰事件.小兴安岭丰林自然保护区臭冷杉径向生长与AMO和太平洋年代际振荡(Pacific Decadal Oscillation, PDO)有直接关系, AMO、PDO的相位转变可能是谷地臭冷杉衰退的原因(姚启超和王晓春, 2013 ).长白山安图(徐海等, 2002 )和小兴安岭五营(尹红等, 2009 )红松树轮重建的降水或气温序列中也发现了与ENSO等区域性气候模态对应的周期, 也说明红松径向生长受区域性气候模态的影响.大尺度气候波动虽然不会直接影响某一区域树木的生长, 但能通过影响局地气候间接影响树木生长(张先亮等, 2010 ).当这种影响超过一定的阈值时, 势必会导致树木出现生长衰退或死亡.因此, 外力作用下更易形成林窗, 造成周围树体生长释放.另外, 区域气候模态(尤其是ENSO)与东亚季风关系密切(徐袁和钱维宏, 2003 ), 由此形成的风暴更是触发林窗形成的关键原因. ...
... 本研究发现风干扰是小兴安岭原始阔叶红松林树木生长释放的一个主要因素, 风干扰是该区阔叶红松林干扰的主要类型, 这与其他学者在长白山(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭(王晓春和赵玉芳, 2011 ; 赵玉芳, 2011 )及俄罗斯远东(Ishikawa et al ., 1999 )等地阔叶红松林干扰研究的结果一致.风干扰造成树体倾倒(尤其是林冠层乔木, 如红松)形成林窗(图9 ), 林窗的出现解除了周围树木生长的竞争, 林窗边缘木将出现明显的生长释放(王晓春和赵玉芳, 2011 ).例如, 研究区春季(4-5月)多风且风速较大, 春季最大风速平均值前10个最大年份分别为1971、1976、1987、1989-1990、1994-1996和1998-1999年, 这些年份当年或后几年红松生长变化百分率均出现明显增加的趋势.臭冷杉由于是喜阴树种且多处于林冠下层, 生长释放不太明显.此外, 林窗内形成木数量、腐烂等级以及林窗边缘木生长释放特征, 很好地证明了森林遭大风袭击后摧毁了部分林冠层树木, 造成林冠参差不齐.林窗形成后, 该区对风的抵抗力便会大大削弱, 林分很容易发生持续的风倒, 产生多林窗潮(multiple-gap episodes)(阳含熙和谢海生, 1994 ; 臧润国和徐化成, 1998 ). ...
... 风干扰是阔叶红松林干扰的主要因素之一(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 ), 然而并不意味着风速越大, 形成林窗的数量就越多, 生长释放就越明显.风只是林窗干扰发生的一个外因, 林窗干扰是否发生以及干扰的强弱还取决于树体自身的生长情况及其生存的环境条件.此外, 异常气温也在很大程度上影响着该区树木的生长释放, 这一结果与王晓春和赵玉芳(2011)的研究结果一致.异常气温影响研究区的干旱化现象(贺伟等, 2013 ), 导致该区树木生长衰退或死亡, 对外界干扰的抵抗能力降低或丧失, 在外力作用下更易形成倒木, 进而造成林窗边缘树木发生释放. ...
长白山红松林干扰节律研究
3
2000
... 小兴安岭地区阔叶红松林内红松干扰事件主要发生在18世纪30年代、19世纪10年代、19世纪30-40年代、20世纪20-30年代, 臭冷杉主要发生在19世纪90年代、20世纪30年代、20世纪50-60年代、20世纪90年代.该结果与其他学者在长白山地区(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭地区(赵玉芳, 2011 )以及俄罗斯远东地区(Ishikawa et al ., 1999 )等阔叶红松林主要分布区的研究结果相一致, 说明阔叶红松林干扰事件不只是局限在研究区域内的小范围事件, 而且可能为区域尺度上(整个阔叶红松林分布区)的大范围干扰事件.红松和臭冷杉干扰事件可能受区域性气候模态周期影响(图4 ), 进一步说明阔叶红松林干扰事件是区域尺度上的大范围干扰事件.小兴安岭丰林自然保护区臭冷杉径向生长与AMO和太平洋年代际振荡(Pacific Decadal Oscillation, PDO)有直接关系, AMO、PDO的相位转变可能是谷地臭冷杉衰退的原因(姚启超和王晓春, 2013 ).长白山安图(徐海等, 2002 )和小兴安岭五营(尹红等, 2009 )红松树轮重建的降水或气温序列中也发现了与ENSO等区域性气候模态对应的周期, 也说明红松径向生长受区域性气候模态的影响.大尺度气候波动虽然不会直接影响某一区域树木的生长, 但能通过影响局地气候间接影响树木生长(张先亮等, 2010 ).当这种影响超过一定的阈值时, 势必会导致树木出现生长衰退或死亡.因此, 外力作用下更易形成林窗, 造成周围树体生长释放.另外, 区域气候模态(尤其是ENSO)与东亚季风关系密切(徐袁和钱维宏, 2003 ), 由此形成的风暴更是触发林窗形成的关键原因. ...
... 本研究发现风干扰是小兴安岭原始阔叶红松林树木生长释放的一个主要因素, 风干扰是该区阔叶红松林干扰的主要类型, 这与其他学者在长白山(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭(王晓春和赵玉芳, 2011 ; 赵玉芳, 2011 )及俄罗斯远东(Ishikawa et al ., 1999 )等地阔叶红松林干扰研究的结果一致.风干扰造成树体倾倒(尤其是林冠层乔木, 如红松)形成林窗(图9 ), 林窗的出现解除了周围树木生长的竞争, 林窗边缘木将出现明显的生长释放(王晓春和赵玉芳, 2011 ).例如, 研究区春季(4-5月)多风且风速较大, 春季最大风速平均值前10个最大年份分别为1971、1976、1987、1989-1990、1994-1996和1998-1999年, 这些年份当年或后几年红松生长变化百分率均出现明显增加的趋势.臭冷杉由于是喜阴树种且多处于林冠下层, 生长释放不太明显.此外, 林窗内形成木数量、腐烂等级以及林窗边缘木生长释放特征, 很好地证明了森林遭大风袭击后摧毁了部分林冠层树木, 造成林冠参差不齐.林窗形成后, 该区对风的抵抗力便会大大削弱, 林分很容易发生持续的风倒, 产生多林窗潮(multiple-gap episodes)(阳含熙和谢海生, 1994 ; 臧润国和徐化成, 1998 ). ...
... 风干扰是阔叶红松林干扰的主要因素之一(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 ), 然而并不意味着风速越大, 形成林窗的数量就越多, 生长释放就越明显.风只是林窗干扰发生的一个外因, 林窗干扰是否发生以及干扰的强弱还取决于树体自身的生长情况及其生存的环境条件.此外, 异常气温也在很大程度上影响着该区树木的生长释放, 这一结果与王晓春和赵玉芳(2011)的研究结果一致.异常气温影响研究区的干旱化现象(贺伟等, 2013 ), 导致该区树木生长衰退或死亡, 对外界干扰的抵抗能力降低或丧失, 在外力作用下更易形成倒木, 进而造成林窗边缘树木发生释放. ...
Disturbance history and tree establishment in old-growth Pinus koraiensis-hardwood forests in the Russian Far East.
3
1999
... 国外已有一些研究利用树木年轮重建森林干扰历史(Nowacki & Abrams, 1997 ; Ishikawa et al ., 1999 ; Rubino & McCarthyz, 2004 ; Brown & Giesecke, 2014 ), 而国内这方面研究起步较晚, 且相对较少.20世纪90年代初, 阳含熙和谢海生(1994)最先利用树木生长释放和抑制对长白山红松(Pinus koraiensis )混交林干扰历史进行了重建, 并指出该方法是以风作为主要干扰类型, 重建干扰历史最准确、最可靠、提供信息量最大的方法; 侯向阳等(2000)研究长白山红松林干扰节律时, 也得到了类似的结论.乔佃锋(2005)利用该方法(阳含熙和谢海生, 1994 )重建了长白山敦化地区阔叶红松林的干扰历史, 发现该区红松林干扰以小规模的风干扰为主, 1980年出现了明显的干扰峰, 干扰峰之间60年的时间间隔正好与红松的更替世代相吻合.王晓春和赵玉芳(2011)发现前一年径向生长对生长释放事件的判定很重要, 小兴安岭北端黑河地区的红松和红皮云杉(Picea koraiensis )的生长释放可能受林窗产生、微环境变化以及温度和降水等气候因素影响.夏冰等(1995)在天目山建立了生长释放和抑制的判定标准, 并重建了黄山松(Pinus taiwanensis )阔叶林树冠的干扰历史; 同时, 还对不同树种年轮宽度的生长释放判别方法进行了研究, 并做了重建干扰强度的尝试(夏冰等, 1997 , 2001 ).此外, 不同学者还利用树轮生态学方法对火灾(张磊和王晓春, 2013 )、泥石流(铁永波等, 2014 )、山体滑坡(洪婷等, 2012 )和冰川活动(Zhu et al ., 2013 )等干扰进行了重建. ...
... 小兴安岭地区阔叶红松林内红松干扰事件主要发生在18世纪30年代、19世纪10年代、19世纪30-40年代、20世纪20-30年代, 臭冷杉主要发生在19世纪90年代、20世纪30年代、20世纪50-60年代、20世纪90年代.该结果与其他学者在长白山地区(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭地区(赵玉芳, 2011 )以及俄罗斯远东地区(Ishikawa et al ., 1999 )等阔叶红松林主要分布区的研究结果相一致, 说明阔叶红松林干扰事件不只是局限在研究区域内的小范围事件, 而且可能为区域尺度上(整个阔叶红松林分布区)的大范围干扰事件.红松和臭冷杉干扰事件可能受区域性气候模态周期影响(图4 ), 进一步说明阔叶红松林干扰事件是区域尺度上的大范围干扰事件.小兴安岭丰林自然保护区臭冷杉径向生长与AMO和太平洋年代际振荡(Pacific Decadal Oscillation, PDO)有直接关系, AMO、PDO的相位转变可能是谷地臭冷杉衰退的原因(姚启超和王晓春, 2013 ).长白山安图(徐海等, 2002 )和小兴安岭五营(尹红等, 2009 )红松树轮重建的降水或气温序列中也发现了与ENSO等区域性气候模态对应的周期, 也说明红松径向生长受区域性气候模态的影响.大尺度气候波动虽然不会直接影响某一区域树木的生长, 但能通过影响局地气候间接影响树木生长(张先亮等, 2010 ).当这种影响超过一定的阈值时, 势必会导致树木出现生长衰退或死亡.因此, 外力作用下更易形成林窗, 造成周围树体生长释放.另外, 区域气候模态(尤其是ENSO)与东亚季风关系密切(徐袁和钱维宏, 2003 ), 由此形成的风暴更是触发林窗形成的关键原因. ...
... 本研究发现风干扰是小兴安岭原始阔叶红松林树木生长释放的一个主要因素, 风干扰是该区阔叶红松林干扰的主要类型, 这与其他学者在长白山(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭(王晓春和赵玉芳, 2011 ; 赵玉芳, 2011 )及俄罗斯远东(Ishikawa et al ., 1999 )等地阔叶红松林干扰研究的结果一致.风干扰造成树体倾倒(尤其是林冠层乔木, 如红松)形成林窗(图9 ), 林窗的出现解除了周围树木生长的竞争, 林窗边缘木将出现明显的生长释放(王晓春和赵玉芳, 2011 ).例如, 研究区春季(4-5月)多风且风速较大, 春季最大风速平均值前10个最大年份分别为1971、1976、1987、1989-1990、1994-1996和1998-1999年, 这些年份当年或后几年红松生长变化百分率均出现明显增加的趋势.臭冷杉由于是喜阴树种且多处于林冠下层, 生长释放不太明显.此外, 林窗内形成木数量、腐烂等级以及林窗边缘木生长释放特征, 很好地证明了森林遭大风袭击后摧毁了部分林冠层树木, 造成林冠参差不齐.林窗形成后, 该区对风的抵抗力便会大大削弱, 林分很容易发生持续的风倒, 产生多林窗潮(multiple-gap episodes)(阳含熙和谢海生, 1994 ; 臧润国和徐化成, 1998 ). ...
1
1997
... 阔叶红松林是我国东北东部山地的地带性顶极植被, 分布于长白山、完达山、小兴安岭一带的中低山区, 是世界上为数不多的大面积原生针阔混交林, 以其结构复杂、组成独特、生物多样性丰富而著称(李景文, 1997 ).然而, 由于过去不合理的采伐利用, 天然阔叶红松林资源逐年锐减, 原始阔叶红松林几乎荡然无存.开展阔叶红松林主要树种生长释放及干扰历史研究, 对探讨现存阔叶红松林的稳定性、碳源汇功能等具有重要意义.目前已有的阔叶红松林干扰重建研究主要集中在长白山一带, 且多停留在干扰年代和干扰频率的确定上, 并未对干扰强度以及可能存在的干扰机制进行探讨.小兴安岭是阔叶红松林在我国分布最为集中, 且保存完好的核心地区.红松和臭冷杉(Abies nephrolepis )是阔叶红松林内的主要针叶树种.因此, 本文利用其生长抑制和释放规律, 确定小兴安岭典型阔叶红松林林窗干扰的发生时间和频率, 探讨过去的干扰历史及干扰形成机制, 为阔叶红松林的保护与可持续利用提供科学依据. ...
1
1997
... 阔叶红松林是我国东北东部山地的地带性顶极植被, 分布于长白山、完达山、小兴安岭一带的中低山区, 是世界上为数不多的大面积原生针阔混交林, 以其结构复杂、组成独特、生物多样性丰富而著称(李景文, 1997 ).然而, 由于过去不合理的采伐利用, 天然阔叶红松林资源逐年锐减, 原始阔叶红松林几乎荡然无存.开展阔叶红松林主要树种生长释放及干扰历史研究, 对探讨现存阔叶红松林的稳定性、碳源汇功能等具有重要意义.目前已有的阔叶红松林干扰重建研究主要集中在长白山一带, 且多停留在干扰年代和干扰频率的确定上, 并未对干扰强度以及可能存在的干扰机制进行探讨.小兴安岭是阔叶红松林在我国分布最为集中, 且保存完好的核心地区.红松和臭冷杉(Abies nephrolepis )是阔叶红松林内的主要针叶树种.因此, 本文利用其生长抑制和释放规律, 确定小兴安岭典型阔叶红松林林窗干扰的发生时间和频率, 探讨过去的干扰历史及干扰形成机制, 为阔叶红松林的保护与可持续利用提供科学依据. ...
小兴安岭阔叶红松混交林林隙特征
3
2013
... 随着频繁的人类活动和异常自然干扰的发生, 森林生态系统结构与功能正受到严重的威胁(Vitousek et al ., 1997 ).干扰生态学已成为生态学研究最活跃的领域之一(朱教君和刘世荣, 2007 ; Bond-Lamberty et al ., 2014 ), 其重点是从森林群落的干扰历史和林窗(隙)动态两个方面对中小规模干扰的特征及影响开展研究, 且有关林窗动态的研究相对较多(臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).重建干扰历史是植物群落演替和更新研究的基础, 越来越受到重视.森林群落的干扰动态可以通过固定样地的长期观测获得(Cooke & Lorenzetti, 2006 ), 但对于缺乏长期观测的森林群落, 人们无法得知其干扰历史.树木年轮是树木生长的历史记录(Smith, 2008 ), 其宽度、密度、同位素含量以及解剖结构等记载了树木逐年的生长信息及内、外部环境变化的影响(Cook & Kairiuktis, 1990 ).树轮具有生长时间长、定年准确、材料广泛、容易获取等优点, 是研究树木生长环境变化的重要手段, 也是重建森林群落动态的重要材料. ...
... 林窗干扰是自然界中最常见的一种森林干扰(臧润国和徐化成, 1998 ; 臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).林窗干扰发生后, 周围树木由于竞争去除而产生生长释放(王晓春和赵玉芳, 2011 ), 并且这种释放可以持续多年.通常将大于一定阈值且持续5 a或10 a以上的生长释放视为“真正释放”, 并将生长释放开始的时间视为干扰的发生时间(Black & Abrams, 2004 ; Rubino & McCarthyz, 2004 ; 王晓春和赵玉芳, 2011 ).确定生长释放的发生时间是重建森林干扰历史的最基本过程(Black & Abrams, 2004 ), 林窗形成时间常以林窗内幼苗更新时间(刘少冲等, 2013 )、倒木倾倒时间(用腐烂等级确定, 韩路等, 2011 )以及林窗周边树木生长释放时间(阳含熙和谢海生, 1994 ; 夏冰等, 1995 )等确定.本研究林窗干扰时间的确定主要用树木生长释放来判断, 同时还参考了林窗内的幼苗更新及倒木腐烂等级, 但有些林窗更新的非均一性以及林窗内倒木腐烂情况比较复杂也影响林窗形成时间的确定. ...
... ), 林窗形成时间常以林窗内幼苗更新时间(刘少冲等, 2013 )、倒木倾倒时间(用腐烂等级确定, 韩路等, 2011 )以及林窗周边树木生长释放时间(阳含熙和谢海生, 1994 ; 夏冰等, 1995 )等确定.本研究林窗干扰时间的确定主要用树木生长释放来判断, 同时还参考了林窗内的幼苗更新及倒木腐烂等级, 但有些林窗更新的非均一性以及林窗内倒木腐烂情况比较复杂也影响林窗形成时间的确定. ...
小兴安岭阔叶红松混交林林隙特征
3
2013
... 随着频繁的人类活动和异常自然干扰的发生, 森林生态系统结构与功能正受到严重的威胁(Vitousek et al ., 1997 ).干扰生态学已成为生态学研究最活跃的领域之一(朱教君和刘世荣, 2007 ; Bond-Lamberty et al ., 2014 ), 其重点是从森林群落的干扰历史和林窗(隙)动态两个方面对中小规模干扰的特征及影响开展研究, 且有关林窗动态的研究相对较多(臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).重建干扰历史是植物群落演替和更新研究的基础, 越来越受到重视.森林群落的干扰动态可以通过固定样地的长期观测获得(Cooke & Lorenzetti, 2006 ), 但对于缺乏长期观测的森林群落, 人们无法得知其干扰历史.树木年轮是树木生长的历史记录(Smith, 2008 ), 其宽度、密度、同位素含量以及解剖结构等记载了树木逐年的生长信息及内、外部环境变化的影响(Cook & Kairiuktis, 1990 ).树轮具有生长时间长、定年准确、材料广泛、容易获取等优点, 是研究树木生长环境变化的重要手段, 也是重建森林群落动态的重要材料. ...
... 林窗干扰是自然界中最常见的一种森林干扰(臧润国和徐化成, 1998 ; 臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).林窗干扰发生后, 周围树木由于竞争去除而产生生长释放(王晓春和赵玉芳, 2011 ), 并且这种释放可以持续多年.通常将大于一定阈值且持续5 a或10 a以上的生长释放视为“真正释放”, 并将生长释放开始的时间视为干扰的发生时间(Black & Abrams, 2004 ; Rubino & McCarthyz, 2004 ; 王晓春和赵玉芳, 2011 ).确定生长释放的发生时间是重建森林干扰历史的最基本过程(Black & Abrams, 2004 ), 林窗形成时间常以林窗内幼苗更新时间(刘少冲等, 2013 )、倒木倾倒时间(用腐烂等级确定, 韩路等, 2011 )以及林窗周边树木生长释放时间(阳含熙和谢海生, 1994 ; 夏冰等, 1995 )等确定.本研究林窗干扰时间的确定主要用树木生长释放来判断, 同时还参考了林窗内的幼苗更新及倒木腐烂等级, 但有些林窗更新的非均一性以及林窗内倒木腐烂情况比较复杂也影响林窗形成时间的确定. ...
... ), 林窗形成时间常以林窗内幼苗更新时间(刘少冲等, 2013 )、倒木倾倒时间(用腐烂等级确定, 韩路等, 2011 )以及林窗周边树木生长释放时间(阳含熙和谢海生, 1994 ; 夏冰等, 1995 )等确定.本研究林窗干扰时间的确定主要用树木生长释放来判断, 同时还参考了林窗内的幼苗更新及倒木腐烂等级, 但有些林窗更新的非均一性以及林窗内倒木腐烂情况比较复杂也影响林窗形成时间的确定. ...
地形对小兴安岭阔叶红松(Pinus koraiensis)林粗木质残体分布的影响
2009
地形对小兴安岭阔叶红松(Pinus koraiensis)林粗木质残体分布的影响
2009
Radial-growth averaging criteria for reconstructing disturbance histories from resettlement-origin oaks.
1
1997
... 国外已有一些研究利用树木年轮重建森林干扰历史(Nowacki & Abrams, 1997 ; Ishikawa et al ., 1999 ; Rubino & McCarthyz, 2004 ; Brown & Giesecke, 2014 ), 而国内这方面研究起步较晚, 且相对较少.20世纪90年代初, 阳含熙和谢海生(1994)最先利用树木生长释放和抑制对长白山红松(Pinus koraiensis )混交林干扰历史进行了重建, 并指出该方法是以风作为主要干扰类型, 重建干扰历史最准确、最可靠、提供信息量最大的方法; 侯向阳等(2000)研究长白山红松林干扰节律时, 也得到了类似的结论.乔佃锋(2005)利用该方法(阳含熙和谢海生, 1994 )重建了长白山敦化地区阔叶红松林的干扰历史, 发现该区红松林干扰以小规模的风干扰为主, 1980年出现了明显的干扰峰, 干扰峰之间60年的时间间隔正好与红松的更替世代相吻合.王晓春和赵玉芳(2011)发现前一年径向生长对生长释放事件的判定很重要, 小兴安岭北端黑河地区的红松和红皮云杉(Picea koraiensis )的生长释放可能受林窗产生、微环境变化以及温度和降水等气候因素影响.夏冰等(1995)在天目山建立了生长释放和抑制的判定标准, 并重建了黄山松(Pinus taiwanensis )阔叶林树冠的干扰历史; 同时, 还对不同树种年轮宽度的生长释放判别方法进行了研究, 并做了重建干扰强度的尝试(夏冰等, 1997 , 2001 ).此外, 不同学者还利用树轮生态学方法对火灾(张磊和王晓春, 2013 )、泥石流(铁永波等, 2014 )、山体滑坡(洪婷等, 2012 )和冰川活动(Zhu et al ., 2013 )等干扰进行了重建. ...
敦化地区红松林干扰历史研究
3
2005
... 小兴安岭地区阔叶红松林内红松干扰事件主要发生在18世纪30年代、19世纪10年代、19世纪30-40年代、20世纪20-30年代, 臭冷杉主要发生在19世纪90年代、20世纪30年代、20世纪50-60年代、20世纪90年代.该结果与其他学者在长白山地区(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭地区(赵玉芳, 2011 )以及俄罗斯远东地区(Ishikawa et al ., 1999 )等阔叶红松林主要分布区的研究结果相一致, 说明阔叶红松林干扰事件不只是局限在研究区域内的小范围事件, 而且可能为区域尺度上(整个阔叶红松林分布区)的大范围干扰事件.红松和臭冷杉干扰事件可能受区域性气候模态周期影响(图4 ), 进一步说明阔叶红松林干扰事件是区域尺度上的大范围干扰事件.小兴安岭丰林自然保护区臭冷杉径向生长与AMO和太平洋年代际振荡(Pacific Decadal Oscillation, PDO)有直接关系, AMO、PDO的相位转变可能是谷地臭冷杉衰退的原因(姚启超和王晓春, 2013 ).长白山安图(徐海等, 2002 )和小兴安岭五营(尹红等, 2009 )红松树轮重建的降水或气温序列中也发现了与ENSO等区域性气候模态对应的周期, 也说明红松径向生长受区域性气候模态的影响.大尺度气候波动虽然不会直接影响某一区域树木的生长, 但能通过影响局地气候间接影响树木生长(张先亮等, 2010 ).当这种影响超过一定的阈值时, 势必会导致树木出现生长衰退或死亡.因此, 外力作用下更易形成林窗, 造成周围树体生长释放.另外, 区域气候模态(尤其是ENSO)与东亚季风关系密切(徐袁和钱维宏, 2003 ), 由此形成的风暴更是触发林窗形成的关键原因. ...
... 本研究发现风干扰是小兴安岭原始阔叶红松林树木生长释放的一个主要因素, 风干扰是该区阔叶红松林干扰的主要类型, 这与其他学者在长白山(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭(王晓春和赵玉芳, 2011 ; 赵玉芳, 2011 )及俄罗斯远东(Ishikawa et al ., 1999 )等地阔叶红松林干扰研究的结果一致.风干扰造成树体倾倒(尤其是林冠层乔木, 如红松)形成林窗(图9 ), 林窗的出现解除了周围树木生长的竞争, 林窗边缘木将出现明显的生长释放(王晓春和赵玉芳, 2011 ).例如, 研究区春季(4-5月)多风且风速较大, 春季最大风速平均值前10个最大年份分别为1971、1976、1987、1989-1990、1994-1996和1998-1999年, 这些年份当年或后几年红松生长变化百分率均出现明显增加的趋势.臭冷杉由于是喜阴树种且多处于林冠下层, 生长释放不太明显.此外, 林窗内形成木数量、腐烂等级以及林窗边缘木生长释放特征, 很好地证明了森林遭大风袭击后摧毁了部分林冠层树木, 造成林冠参差不齐.林窗形成后, 该区对风的抵抗力便会大大削弱, 林分很容易发生持续的风倒, 产生多林窗潮(multiple-gap episodes)(阳含熙和谢海生, 1994 ; 臧润国和徐化成, 1998 ). ...
... 风干扰是阔叶红松林干扰的主要因素之一(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 ), 然而并不意味着风速越大, 形成林窗的数量就越多, 生长释放就越明显.风只是林窗干扰发生的一个外因, 林窗干扰是否发生以及干扰的强弱还取决于树体自身的生长情况及其生存的环境条件.此外, 异常气温也在很大程度上影响着该区树木的生长释放, 这一结果与王晓春和赵玉芳(2011)的研究结果一致.异常气温影响研究区的干旱化现象(贺伟等, 2013 ), 导致该区树木生长衰退或死亡, 对外界干扰的抵抗能力降低或丧失, 在外力作用下更易形成倒木, 进而造成林窗边缘树木发生释放. ...
敦化地区红松林干扰历史研究
3
2005
... 小兴安岭地区阔叶红松林内红松干扰事件主要发生在18世纪30年代、19世纪10年代、19世纪30-40年代、20世纪20-30年代, 臭冷杉主要发生在19世纪90年代、20世纪30年代、20世纪50-60年代、20世纪90年代.该结果与其他学者在长白山地区(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭地区(赵玉芳, 2011 )以及俄罗斯远东地区(Ishikawa et al ., 1999 )等阔叶红松林主要分布区的研究结果相一致, 说明阔叶红松林干扰事件不只是局限在研究区域内的小范围事件, 而且可能为区域尺度上(整个阔叶红松林分布区)的大范围干扰事件.红松和臭冷杉干扰事件可能受区域性气候模态周期影响(图4 ), 进一步说明阔叶红松林干扰事件是区域尺度上的大范围干扰事件.小兴安岭丰林自然保护区臭冷杉径向生长与AMO和太平洋年代际振荡(Pacific Decadal Oscillation, PDO)有直接关系, AMO、PDO的相位转变可能是谷地臭冷杉衰退的原因(姚启超和王晓春, 2013 ).长白山安图(徐海等, 2002 )和小兴安岭五营(尹红等, 2009 )红松树轮重建的降水或气温序列中也发现了与ENSO等区域性气候模态对应的周期, 也说明红松径向生长受区域性气候模态的影响.大尺度气候波动虽然不会直接影响某一区域树木的生长, 但能通过影响局地气候间接影响树木生长(张先亮等, 2010 ).当这种影响超过一定的阈值时, 势必会导致树木出现生长衰退或死亡.因此, 外力作用下更易形成林窗, 造成周围树体生长释放.另外, 区域气候模态(尤其是ENSO)与东亚季风关系密切(徐袁和钱维宏, 2003 ), 由此形成的风暴更是触发林窗形成的关键原因. ...
... 本研究发现风干扰是小兴安岭原始阔叶红松林树木生长释放的一个主要因素, 风干扰是该区阔叶红松林干扰的主要类型, 这与其他学者在长白山(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭(王晓春和赵玉芳, 2011 ; 赵玉芳, 2011 )及俄罗斯远东(Ishikawa et al ., 1999 )等地阔叶红松林干扰研究的结果一致.风干扰造成树体倾倒(尤其是林冠层乔木, 如红松)形成林窗(图9 ), 林窗的出现解除了周围树木生长的竞争, 林窗边缘木将出现明显的生长释放(王晓春和赵玉芳, 2011 ).例如, 研究区春季(4-5月)多风且风速较大, 春季最大风速平均值前10个最大年份分别为1971、1976、1987、1989-1990、1994-1996和1998-1999年, 这些年份当年或后几年红松生长变化百分率均出现明显增加的趋势.臭冷杉由于是喜阴树种且多处于林冠下层, 生长释放不太明显.此外, 林窗内形成木数量、腐烂等级以及林窗边缘木生长释放特征, 很好地证明了森林遭大风袭击后摧毁了部分林冠层树木, 造成林冠参差不齐.林窗形成后, 该区对风的抵抗力便会大大削弱, 林分很容易发生持续的风倒, 产生多林窗潮(multiple-gap episodes)(阳含熙和谢海生, 1994 ; 臧润国和徐化成, 1998 ). ...
... 风干扰是阔叶红松林干扰的主要因素之一(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 ), 然而并不意味着风速越大, 形成林窗的数量就越多, 生长释放就越明显.风只是林窗干扰发生的一个外因, 林窗干扰是否发生以及干扰的强弱还取决于树体自身的生长情况及其生存的环境条件.此外, 异常气温也在很大程度上影响着该区树木的生长释放, 这一结果与王晓春和赵玉芳(2011)的研究结果一致.异常气温影响研究区的干旱化现象(贺伟等, 2013 ), 导致该区树木生长衰退或死亡, 对外界干扰的抵抗能力降低或丧失, 在外力作用下更易形成倒木, 进而造成林窗边缘树木发生释放. ...
Comparative analysis of dendroecological methods used to assess disturbance events.
4
2004
... 国外已有一些研究利用树木年轮重建森林干扰历史(Nowacki & Abrams, 1997 ; Ishikawa et al ., 1999 ; Rubino & McCarthyz, 2004 ; Brown & Giesecke, 2014 ), 而国内这方面研究起步较晚, 且相对较少.20世纪90年代初, 阳含熙和谢海生(1994)最先利用树木生长释放和抑制对长白山红松(Pinus koraiensis )混交林干扰历史进行了重建, 并指出该方法是以风作为主要干扰类型, 重建干扰历史最准确、最可靠、提供信息量最大的方法; 侯向阳等(2000)研究长白山红松林干扰节律时, 也得到了类似的结论.乔佃锋(2005)利用该方法(阳含熙和谢海生, 1994 )重建了长白山敦化地区阔叶红松林的干扰历史, 发现该区红松林干扰以小规模的风干扰为主, 1980年出现了明显的干扰峰, 干扰峰之间60年的时间间隔正好与红松的更替世代相吻合.王晓春和赵玉芳(2011)发现前一年径向生长对生长释放事件的判定很重要, 小兴安岭北端黑河地区的红松和红皮云杉(Picea koraiensis )的生长释放可能受林窗产生、微环境变化以及温度和降水等气候因素影响.夏冰等(1995)在天目山建立了生长释放和抑制的判定标准, 并重建了黄山松(Pinus taiwanensis )阔叶林树冠的干扰历史; 同时, 还对不同树种年轮宽度的生长释放判别方法进行了研究, 并做了重建干扰强度的尝试(夏冰等, 1997 , 2001 ).此外, 不同学者还利用树轮生态学方法对火灾(张磊和王晓春, 2013 )、泥石流(铁永波等, 2014 )、山体滑坡(洪婷等, 2012 )和冰川活动(Zhu et al ., 2013 )等干扰进行了重建. ...
... 本文采用王晓春和赵玉芳(2011)的方法计算当年年轮宽度序列的生长变化百分率(Percentage Growth Change, GC).该方法在Nowacki和Abrams (1997)的方法基础上, 将滑动平均值法修正为中值法, 并将滑动窗口由10 a调整为5 a, 以凸显非正常干扰事件的影响(Rubino & McCarthyz, 2004 ; 王晓春和赵玉芳, 2011 ).研究中以林窗为单位计算臭冷杉和红松的生长释放标准曲线, 并利用该标准曲线判断每一边缘木的生长释放情况.将GC介于25%-50%之间且持续5 a以上的释放记为“中等释放”, GC > 50%且持续5 a以上的释放记为“主要释放”, GC < 25%则不考虑为真正释放.林窗木与非林窗木生长变化百分率的差值可以在很大程度上反映林窗的干扰情况.首先, 求出样地内每一个林窗所有边缘木生长变化百分率的平均值; 其次, 将各平均值逐一与非林窗木生长变化百分率的平均值作差, 得出各个林窗干扰情况随时间的变化序列; 最后, 再将得到的44个时间序列每一年的值, 按正、负进行求和后便得出柱状累计图(图1 ).本文以平均每株树木经历生长释放次数的倒数乘以生长释放频率, 推测过去树木的死亡率即干扰强度(夏冰等, 1997 , 2001 ).例如, 臭冷杉经历的平均主要生长释放次数为1.05次, 其平均生长释放频率为每10 a 8.79%, 则其平均干扰强度是每10 a 8.37% (表1 ). ...
... 林窗干扰是自然界中最常见的一种森林干扰(臧润国和徐化成, 1998 ; 臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).林窗干扰发生后, 周围树木由于竞争去除而产生生长释放(王晓春和赵玉芳, 2011 ), 并且这种释放可以持续多年.通常将大于一定阈值且持续5 a或10 a以上的生长释放视为“真正释放”, 并将生长释放开始的时间视为干扰的发生时间(Black & Abrams, 2004 ; Rubino & McCarthyz, 2004 ; 王晓春和赵玉芳, 2011 ).确定生长释放的发生时间是重建森林干扰历史的最基本过程(Black & Abrams, 2004 ), 林窗形成时间常以林窗内幼苗更新时间(刘少冲等, 2013 )、倒木倾倒时间(用腐烂等级确定, 韩路等, 2011 )以及林窗周边树木生长释放时间(阳含熙和谢海生, 1994 ; 夏冰等, 1995 )等确定.本研究林窗干扰时间的确定主要用树木生长释放来判断, 同时还参考了林窗内的幼苗更新及倒木腐烂等级, 但有些林窗更新的非均一性以及林窗内倒木腐烂情况比较复杂也影响林窗形成时间的确定. ...
... 生长释放判定方法的选择及判定标准的确定是树轮干扰历史重建的关键.判定方法和标准不同, 得出的生长释放结果亦不同(Rubino & McCarthyz, 2004 ), 每一种方法都有各自的优缺点及使用范围(封晓辉等, 2011 ).在实践中, 通过主观判断去选择生长释放判定方法及判定标准, 往往会因主观原因放大或弱化干扰事件.因此, 在判定方法及判定标准选择时, 在充分考虑研究需要的同时, 还需结合研究地点及环境条件等合理选择(阳含熙和谢海生, 1994 ; 夏冰等, 1997 ; 王晓春和赵玉芳, 2011 ).为了保留更多的干扰信号, 本文选用生长释放标准曲线法(王晓春和赵玉芳, 2011 ), 即生长变化率超过一定的标准便可视为生长释放发生, 该方法是目前应用最多的方法(封晓辉等, 2011 ).本研究中林窗木与非林窗木的生长变化百分率有很强的一致性(仅局部存在部分差异), 说明林窗木生长变化百分率序列中包含了部分非干扰信号.通过生长释放判定得到的生长释放中必定会有部分噪音即非干扰信号, 即并不是所有的释放(包括中等释放和主要释放)都能代表林窗干扰(王晓春和赵玉芳, 2011 ), 因此主要释放或部分主要释放才能代表真正意义上的干扰. 生长变化百分率差值图(图1 )与干扰年表(图2 , 图3 )谷、峰值具有很强的一致性, 且干扰年表在一定程度上对生长变化百分率差值图中不太明显的峰值具有局部放大作用, 经局部放大的部分信号可能是噪音的主要来源.因此, 通过林窗边缘木生长释放判定得出的干扰年表, 能够很好地反映该区阔叶红松林的干扰历史, 且能将一些不太明显的微小干扰包括其中. ...
三峡大老岭山地常绿落叶阔叶混交林林隙干扰研究II
1
2001
... 本研究发现小兴安岭地区林窗木与非林窗木的年生长变化规律(平均生长变化百分率)基本一致, 而以林窗为单位得到的林窗木生长变化百分率与非林窗木之间的差异却很明显, 可能是不同林窗或同一林窗不同树木对同一干扰事件的响应存在差异的结果.自然干扰及其所产生的影响存在巨大的变异性(魏晓华, 2010 ), 可能因海拔、坡度和坡位等因素而出现不同的格局(沈泽昊等, 2001 ).刘妍妍和金光泽(2009)在分析本研究样地内粗木质残体与地形因子的关系时, 也得到了类似的结论.林窗木生长释放特征也具有极大的变异性, 如臭冷杉发生中等以上释放每株2.16次,介于每株0-6次之间; 初次释放平均年龄为32.14 a, 介于15-116 a; 平均间隔期为16.56 a, 介于2-67 a, 从另一方面印证了小兴安岭地区典型阔叶红松林干扰及其所产生的影响存在巨大的变异性.夏冰等(1997)在亚高山云冷杉混交林得到了类似的研究结果.此外, 本研究还发现了一些连续多年的生长释放, 如1889-1904、1932-1938、1947-1973年等, 这可能是连续发生释放或者间隔释放相叠加的结果, 也可能是区域尺度上大范围干扰事件造成的. ...
三峡大老岭山地常绿落叶阔叶混交林林隙干扰研究II
1
2001
... 本研究发现小兴安岭地区林窗木与非林窗木的年生长变化规律(平均生长变化百分率)基本一致, 而以林窗为单位得到的林窗木生长变化百分率与非林窗木之间的差异却很明显, 可能是不同林窗或同一林窗不同树木对同一干扰事件的响应存在差异的结果.自然干扰及其所产生的影响存在巨大的变异性(魏晓华, 2010 ), 可能因海拔、坡度和坡位等因素而出现不同的格局(沈泽昊等, 2001 ).刘妍妍和金光泽(2009)在分析本研究样地内粗木质残体与地形因子的关系时, 也得到了类似的结论.林窗木生长释放特征也具有极大的变异性, 如臭冷杉发生中等以上释放每株2.16次,介于每株0-6次之间; 初次释放平均年龄为32.14 a, 介于15-116 a; 平均间隔期为16.56 a, 介于2-67 a, 从另一方面印证了小兴安岭地区典型阔叶红松林干扰及其所产生的影响存在巨大的变异性.夏冰等(1997)在亚高山云冷杉混交林得到了类似的研究结果.此外, 本研究还发现了一些连续多年的生长释放, 如1889-1904、1932-1938、1947-1973年等, 这可能是连续发生释放或者间隔释放相叠加的结果, 也可能是区域尺度上大范围干扰事件造成的. ...
An organismal view of dendrochronology.
1
2008
... 随着频繁的人类活动和异常自然干扰的发生, 森林生态系统结构与功能正受到严重的威胁(Vitousek et al ., 1997 ).干扰生态学已成为生态学研究最活跃的领域之一(朱教君和刘世荣, 2007 ; Bond-Lamberty et al ., 2014 ), 其重点是从森林群落的干扰历史和林窗(隙)动态两个方面对中小规模干扰的特征及影响开展研究, 且有关林窗动态的研究相对较多(臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).重建干扰历史是植物群落演替和更新研究的基础, 越来越受到重视.森林群落的干扰动态可以通过固定样地的长期观测获得(Cooke & Lorenzetti, 2006 ), 但对于缺乏长期观测的森林群落, 人们无法得知其干扰历史.树木年轮是树木生长的历史记录(Smith, 2008 ), 其宽度、密度、同位素含量以及解剖结构等记载了树木逐年的生长信息及内、外部环境变化的影响(Cook & Kairiuktis, 1990 ).树轮具有生长时间长、定年准确、材料广泛、容易获取等优点, 是研究树木生长环境变化的重要手段, 也是重建森林群落动态的重要材料. ...
1
1996
... 将野外采集的所有样芯带回实验室, 进行固定、干燥、打磨等预处理(Stokes & Smiley, 1996 ).在显微镜下用骨架图法进行交叉定年, 然后用年轮分析仪进行宽度测量, 精度为0.001 mm.最后, 用COFECHA程序来检验定年结果(Holmes, 1983 ), 根据软件提示在显微镜下检查并修订定年与测量中出现的误差, 以确保年轮的日历年代准确. ...
树木年代学在高寒山区泥石流历史事件重建中的应用——以磨西河流域倒灶沟为例
1
2014
... 国外已有一些研究利用树木年轮重建森林干扰历史(Nowacki & Abrams, 1997 ; Ishikawa et al ., 1999 ; Rubino & McCarthyz, 2004 ; Brown & Giesecke, 2014 ), 而国内这方面研究起步较晚, 且相对较少.20世纪90年代初, 阳含熙和谢海生(1994)最先利用树木生长释放和抑制对长白山红松(Pinus koraiensis )混交林干扰历史进行了重建, 并指出该方法是以风作为主要干扰类型, 重建干扰历史最准确、最可靠、提供信息量最大的方法; 侯向阳等(2000)研究长白山红松林干扰节律时, 也得到了类似的结论.乔佃锋(2005)利用该方法(阳含熙和谢海生, 1994 )重建了长白山敦化地区阔叶红松林的干扰历史, 发现该区红松林干扰以小规模的风干扰为主, 1980年出现了明显的干扰峰, 干扰峰之间60年的时间间隔正好与红松的更替世代相吻合.王晓春和赵玉芳(2011)发现前一年径向生长对生长释放事件的判定很重要, 小兴安岭北端黑河地区的红松和红皮云杉(Picea koraiensis )的生长释放可能受林窗产生、微环境变化以及温度和降水等气候因素影响.夏冰等(1995)在天目山建立了生长释放和抑制的判定标准, 并重建了黄山松(Pinus taiwanensis )阔叶林树冠的干扰历史; 同时, 还对不同树种年轮宽度的生长释放判别方法进行了研究, 并做了重建干扰强度的尝试(夏冰等, 1997 , 2001 ).此外, 不同学者还利用树轮生态学方法对火灾(张磊和王晓春, 2013 )、泥石流(铁永波等, 2014 )、山体滑坡(洪婷等, 2012 )和冰川活动(Zhu et al ., 2013 )等干扰进行了重建. ...
树木年代学在高寒山区泥石流历史事件重建中的应用——以磨西河流域倒灶沟为例
1
2014
... 国外已有一些研究利用树木年轮重建森林干扰历史(Nowacki & Abrams, 1997 ; Ishikawa et al ., 1999 ; Rubino & McCarthyz, 2004 ; Brown & Giesecke, 2014 ), 而国内这方面研究起步较晚, 且相对较少.20世纪90年代初, 阳含熙和谢海生(1994)最先利用树木生长释放和抑制对长白山红松(Pinus koraiensis )混交林干扰历史进行了重建, 并指出该方法是以风作为主要干扰类型, 重建干扰历史最准确、最可靠、提供信息量最大的方法; 侯向阳等(2000)研究长白山红松林干扰节律时, 也得到了类似的结论.乔佃锋(2005)利用该方法(阳含熙和谢海生, 1994 )重建了长白山敦化地区阔叶红松林的干扰历史, 发现该区红松林干扰以小规模的风干扰为主, 1980年出现了明显的干扰峰, 干扰峰之间60年的时间间隔正好与红松的更替世代相吻合.王晓春和赵玉芳(2011)发现前一年径向生长对生长释放事件的判定很重要, 小兴安岭北端黑河地区的红松和红皮云杉(Picea koraiensis )的生长释放可能受林窗产生、微环境变化以及温度和降水等气候因素影响.夏冰等(1995)在天目山建立了生长释放和抑制的判定标准, 并重建了黄山松(Pinus taiwanensis )阔叶林树冠的干扰历史; 同时, 还对不同树种年轮宽度的生长释放判别方法进行了研究, 并做了重建干扰强度的尝试(夏冰等, 1997 , 2001 ).此外, 不同学者还利用树轮生态学方法对火灾(张磊和王晓春, 2013 )、泥石流(铁永波等, 2014 )、山体滑坡(洪婷等, 2012 )和冰川活动(Zhu et al ., 2013 )等干扰进行了重建. ...
Human domination of Earth’s ecosystems.
1
1997
... 随着频繁的人类活动和异常自然干扰的发生, 森林生态系统结构与功能正受到严重的威胁(Vitousek et al ., 1997 ).干扰生态学已成为生态学研究最活跃的领域之一(朱教君和刘世荣, 2007 ; Bond-Lamberty et al ., 2014 ), 其重点是从森林群落的干扰历史和林窗(隙)动态两个方面对中小规模干扰的特征及影响开展研究, 且有关林窗动态的研究相对较多(臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).重建干扰历史是植物群落演替和更新研究的基础, 越来越受到重视.森林群落的干扰动态可以通过固定样地的长期观测获得(Cooke & Lorenzetti, 2006 ), 但对于缺乏长期观测的森林群落, 人们无法得知其干扰历史.树木年轮是树木生长的历史记录(Smith, 2008 ), 其宽度、密度、同位素含量以及解剖结构等记载了树木逐年的生长信息及内、外部环境变化的影响(Cook & Kairiuktis, 1990 ).树轮具有生长时间长、定年准确、材料广泛、容易获取等优点, 是研究树木生长环境变化的重要手段, 也是重建森林群落动态的重要材料. ...
Imprint of the Atlantic multidecadal oscillation on tree-ring widths in Northeastern Asia since 1568.
1
2011
... 红松干扰年表(生长释放树木数量占总采样树木的比例随时间的变化序列)存在9.1-18.2 a、7.3-7.9 a、3.8 a、3.5 a和2.0 a的极显著(p < 0.01)周期, 臭冷杉干扰年表(生长释放树木数量占总采样树木的比例随时间的变化序列)存在65-85 a、7.5-48.8 a和3.5-3.6 a的极显著周期(图4 ).通常气象因子波动和植物生长具有2-3 a的周期, 红松与臭冷杉干扰年表也记录了该周期; 2-7 a的干扰周期也可能与厄尔尼诺-南方涛动(El Niño-Southern Oscillation, ENSO)事件有关; 9.1-18.2 a的干扰周期可能受太阳活动的影响.而臭冷杉干扰年表中65-85 a的干扰周期可能受北大西洋年代际振荡(Atlantic Multidecadal Oscillation, AMO)的影响, 已有研究表明AMO会影响东北亚树木生长(Wang et al ., 2011 ; 姚启超和王晓春, 2013 ). ...
黑河胜山国家自然保护区红松和红皮云杉生长释放判定及解释
8
2011
... 本文采用王晓春和赵玉芳(2011)的方法计算当年年轮宽度序列的生长变化百分率(Percentage Growth Change, GC).该方法在Nowacki和Abrams (1997)的方法基础上, 将滑动平均值法修正为中值法, 并将滑动窗口由10 a调整为5 a, 以凸显非正常干扰事件的影响(Rubino & McCarthyz, 2004 ; 王晓春和赵玉芳, 2011 ).研究中以林窗为单位计算臭冷杉和红松的生长释放标准曲线, 并利用该标准曲线判断每一边缘木的生长释放情况.将GC介于25%-50%之间且持续5 a以上的释放记为“中等释放”, GC > 50%且持续5 a以上的释放记为“主要释放”, GC < 25%则不考虑为真正释放.林窗木与非林窗木生长变化百分率的差值可以在很大程度上反映林窗的干扰情况.首先, 求出样地内每一个林窗所有边缘木生长变化百分率的平均值; 其次, 将各平均值逐一与非林窗木生长变化百分率的平均值作差, 得出各个林窗干扰情况随时间的变化序列; 最后, 再将得到的44个时间序列每一年的值, 按正、负进行求和后便得出柱状累计图(图1 ).本文以平均每株树木经历生长释放次数的倒数乘以生长释放频率, 推测过去树木的死亡率即干扰强度(夏冰等, 1997 , 2001 ).例如, 臭冷杉经历的平均主要生长释放次数为1.05次, 其平均生长释放频率为每10 a 8.79%, 则其平均干扰强度是每10 a 8.37% (表1 ). ...
... 林窗干扰是自然界中最常见的一种森林干扰(臧润国和徐化成, 1998 ; 臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).林窗干扰发生后, 周围树木由于竞争去除而产生生长释放(王晓春和赵玉芳, 2011 ), 并且这种释放可以持续多年.通常将大于一定阈值且持续5 a或10 a以上的生长释放视为“真正释放”, 并将生长释放开始的时间视为干扰的发生时间(Black & Abrams, 2004 ; Rubino & McCarthyz, 2004 ; 王晓春和赵玉芳, 2011 ).确定生长释放的发生时间是重建森林干扰历史的最基本过程(Black & Abrams, 2004 ), 林窗形成时间常以林窗内幼苗更新时间(刘少冲等, 2013 )、倒木倾倒时间(用腐烂等级确定, 韩路等, 2011 )以及林窗周边树木生长释放时间(阳含熙和谢海生, 1994 ; 夏冰等, 1995 )等确定.本研究林窗干扰时间的确定主要用树木生长释放来判断, 同时还参考了林窗内的幼苗更新及倒木腐烂等级, 但有些林窗更新的非均一性以及林窗内倒木腐烂情况比较复杂也影响林窗形成时间的确定. ...
... ; 王晓春和赵玉芳, 2011 ).确定生长释放的发生时间是重建森林干扰历史的最基本过程(Black & Abrams, 2004 ), 林窗形成时间常以林窗内幼苗更新时间(刘少冲等, 2013 )、倒木倾倒时间(用腐烂等级确定, 韩路等, 2011 )以及林窗周边树木生长释放时间(阳含熙和谢海生, 1994 ; 夏冰等, 1995 )等确定.本研究林窗干扰时间的确定主要用树木生长释放来判断, 同时还参考了林窗内的幼苗更新及倒木腐烂等级, 但有些林窗更新的非均一性以及林窗内倒木腐烂情况比较复杂也影响林窗形成时间的确定. ...
... 生长释放判定方法的选择及判定标准的确定是树轮干扰历史重建的关键.判定方法和标准不同, 得出的生长释放结果亦不同(Rubino & McCarthyz, 2004 ), 每一种方法都有各自的优缺点及使用范围(封晓辉等, 2011 ).在实践中, 通过主观判断去选择生长释放判定方法及判定标准, 往往会因主观原因放大或弱化干扰事件.因此, 在判定方法及判定标准选择时, 在充分考虑研究需要的同时, 还需结合研究地点及环境条件等合理选择(阳含熙和谢海生, 1994 ; 夏冰等, 1997 ; 王晓春和赵玉芳, 2011 ).为了保留更多的干扰信号, 本文选用生长释放标准曲线法(王晓春和赵玉芳, 2011 ), 即生长变化率超过一定的标准便可视为生长释放发生, 该方法是目前应用最多的方法(封晓辉等, 2011 ).本研究中林窗木与非林窗木的生长变化百分率有很强的一致性(仅局部存在部分差异), 说明林窗木生长变化百分率序列中包含了部分非干扰信号.通过生长释放判定得到的生长释放中必定会有部分噪音即非干扰信号, 即并不是所有的释放(包括中等释放和主要释放)都能代表林窗干扰(王晓春和赵玉芳, 2011 ), 因此主要释放或部分主要释放才能代表真正意义上的干扰. 生长变化百分率差值图(图1 )与干扰年表(图2 , 图3 )谷、峰值具有很强的一致性, 且干扰年表在一定程度上对生长变化百分率差值图中不太明显的峰值具有局部放大作用, 经局部放大的部分信号可能是噪音的主要来源.因此, 通过林窗边缘木生长释放判定得出的干扰年表, 能够很好地反映该区阔叶红松林的干扰历史, 且能将一些不太明显的微小干扰包括其中. ...
... ).为了保留更多的干扰信号, 本文选用生长释放标准曲线法(王晓春和赵玉芳, 2011 ), 即生长变化率超过一定的标准便可视为生长释放发生, 该方法是目前应用最多的方法(封晓辉等, 2011 ).本研究中林窗木与非林窗木的生长变化百分率有很强的一致性(仅局部存在部分差异), 说明林窗木生长变化百分率序列中包含了部分非干扰信号.通过生长释放判定得到的生长释放中必定会有部分噪音即非干扰信号, 即并不是所有的释放(包括中等释放和主要释放)都能代表林窗干扰(王晓春和赵玉芳, 2011 ), 因此主要释放或部分主要释放才能代表真正意义上的干扰. 生长变化百分率差值图(图1 )与干扰年表(图2 , 图3 )谷、峰值具有很强的一致性, 且干扰年表在一定程度上对生长变化百分率差值图中不太明显的峰值具有局部放大作用, 经局部放大的部分信号可能是噪音的主要来源.因此, 通过林窗边缘木生长释放判定得出的干扰年表, 能够很好地反映该区阔叶红松林的干扰历史, 且能将一些不太明显的微小干扰包括其中. ...
... ).本研究中林窗木与非林窗木的生长变化百分率有很强的一致性(仅局部存在部分差异), 说明林窗木生长变化百分率序列中包含了部分非干扰信号.通过生长释放判定得到的生长释放中必定会有部分噪音即非干扰信号, 即并不是所有的释放(包括中等释放和主要释放)都能代表林窗干扰(王晓春和赵玉芳, 2011 ), 因此主要释放或部分主要释放才能代表真正意义上的干扰. 生长变化百分率差值图(图1 )与干扰年表(图2 , 图3 )谷、峰值具有很强的一致性, 且干扰年表在一定程度上对生长变化百分率差值图中不太明显的峰值具有局部放大作用, 经局部放大的部分信号可能是噪音的主要来源.因此, 通过林窗边缘木生长释放判定得出的干扰年表, 能够很好地反映该区阔叶红松林的干扰历史, 且能将一些不太明显的微小干扰包括其中. ...
... 本研究发现风干扰是小兴安岭原始阔叶红松林树木生长释放的一个主要因素, 风干扰是该区阔叶红松林干扰的主要类型, 这与其他学者在长白山(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭(王晓春和赵玉芳, 2011 ; 赵玉芳, 2011 )及俄罗斯远东(Ishikawa et al ., 1999 )等地阔叶红松林干扰研究的结果一致.风干扰造成树体倾倒(尤其是林冠层乔木, 如红松)形成林窗(图9 ), 林窗的出现解除了周围树木生长的竞争, 林窗边缘木将出现明显的生长释放(王晓春和赵玉芳, 2011 ).例如, 研究区春季(4-5月)多风且风速较大, 春季最大风速平均值前10个最大年份分别为1971、1976、1987、1989-1990、1994-1996和1998-1999年, 这些年份当年或后几年红松生长变化百分率均出现明显增加的趋势.臭冷杉由于是喜阴树种且多处于林冠下层, 生长释放不太明显.此外, 林窗内形成木数量、腐烂等级以及林窗边缘木生长释放特征, 很好地证明了森林遭大风袭击后摧毁了部分林冠层树木, 造成林冠参差不齐.林窗形成后, 该区对风的抵抗力便会大大削弱, 林分很容易发生持续的风倒, 产生多林窗潮(multiple-gap episodes)(阳含熙和谢海生, 1994 ; 臧润国和徐化成, 1998 ). ...
... ), 林窗的出现解除了周围树木生长的竞争, 林窗边缘木将出现明显的生长释放(王晓春和赵玉芳, 2011 ).例如, 研究区春季(4-5月)多风且风速较大, 春季最大风速平均值前10个最大年份分别为1971、1976、1987、1989-1990、1994-1996和1998-1999年, 这些年份当年或后几年红松生长变化百分率均出现明显增加的趋势.臭冷杉由于是喜阴树种且多处于林冠下层, 生长释放不太明显.此外, 林窗内形成木数量、腐烂等级以及林窗边缘木生长释放特征, 很好地证明了森林遭大风袭击后摧毁了部分林冠层树木, 造成林冠参差不齐.林窗形成后, 该区对风的抵抗力便会大大削弱, 林分很容易发生持续的风倒, 产生多林窗潮(multiple-gap episodes)(阳含熙和谢海生, 1994 ; 臧润国和徐化成, 1998 ). ...
黑河胜山国家自然保护区红松和红皮云杉生长释放判定及解释
8
2011
... 本文采用王晓春和赵玉芳(2011)的方法计算当年年轮宽度序列的生长变化百分率(Percentage Growth Change, GC).该方法在Nowacki和Abrams (1997)的方法基础上, 将滑动平均值法修正为中值法, 并将滑动窗口由10 a调整为5 a, 以凸显非正常干扰事件的影响(Rubino & McCarthyz, 2004 ; 王晓春和赵玉芳, 2011 ).研究中以林窗为单位计算臭冷杉和红松的生长释放标准曲线, 并利用该标准曲线判断每一边缘木的生长释放情况.将GC介于25%-50%之间且持续5 a以上的释放记为“中等释放”, GC > 50%且持续5 a以上的释放记为“主要释放”, GC < 25%则不考虑为真正释放.林窗木与非林窗木生长变化百分率的差值可以在很大程度上反映林窗的干扰情况.首先, 求出样地内每一个林窗所有边缘木生长变化百分率的平均值; 其次, 将各平均值逐一与非林窗木生长变化百分率的平均值作差, 得出各个林窗干扰情况随时间的变化序列; 最后, 再将得到的44个时间序列每一年的值, 按正、负进行求和后便得出柱状累计图(图1 ).本文以平均每株树木经历生长释放次数的倒数乘以生长释放频率, 推测过去树木的死亡率即干扰强度(夏冰等, 1997 , 2001 ).例如, 臭冷杉经历的平均主要生长释放次数为1.05次, 其平均生长释放频率为每10 a 8.79%, 则其平均干扰强度是每10 a 8.37% (表1 ). ...
... 林窗干扰是自然界中最常见的一种森林干扰(臧润国和徐化成, 1998 ; 臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).林窗干扰发生后, 周围树木由于竞争去除而产生生长释放(王晓春和赵玉芳, 2011 ), 并且这种释放可以持续多年.通常将大于一定阈值且持续5 a或10 a以上的生长释放视为“真正释放”, 并将生长释放开始的时间视为干扰的发生时间(Black & Abrams, 2004 ; Rubino & McCarthyz, 2004 ; 王晓春和赵玉芳, 2011 ).确定生长释放的发生时间是重建森林干扰历史的最基本过程(Black & Abrams, 2004 ), 林窗形成时间常以林窗内幼苗更新时间(刘少冲等, 2013 )、倒木倾倒时间(用腐烂等级确定, 韩路等, 2011 )以及林窗周边树木生长释放时间(阳含熙和谢海生, 1994 ; 夏冰等, 1995 )等确定.本研究林窗干扰时间的确定主要用树木生长释放来判断, 同时还参考了林窗内的幼苗更新及倒木腐烂等级, 但有些林窗更新的非均一性以及林窗内倒木腐烂情况比较复杂也影响林窗形成时间的确定. ...
... ; 王晓春和赵玉芳, 2011 ).确定生长释放的发生时间是重建森林干扰历史的最基本过程(Black & Abrams, 2004 ), 林窗形成时间常以林窗内幼苗更新时间(刘少冲等, 2013 )、倒木倾倒时间(用腐烂等级确定, 韩路等, 2011 )以及林窗周边树木生长释放时间(阳含熙和谢海生, 1994 ; 夏冰等, 1995 )等确定.本研究林窗干扰时间的确定主要用树木生长释放来判断, 同时还参考了林窗内的幼苗更新及倒木腐烂等级, 但有些林窗更新的非均一性以及林窗内倒木腐烂情况比较复杂也影响林窗形成时间的确定. ...
... 生长释放判定方法的选择及判定标准的确定是树轮干扰历史重建的关键.判定方法和标准不同, 得出的生长释放结果亦不同(Rubino & McCarthyz, 2004 ), 每一种方法都有各自的优缺点及使用范围(封晓辉等, 2011 ).在实践中, 通过主观判断去选择生长释放判定方法及判定标准, 往往会因主观原因放大或弱化干扰事件.因此, 在判定方法及判定标准选择时, 在充分考虑研究需要的同时, 还需结合研究地点及环境条件等合理选择(阳含熙和谢海生, 1994 ; 夏冰等, 1997 ; 王晓春和赵玉芳, 2011 ).为了保留更多的干扰信号, 本文选用生长释放标准曲线法(王晓春和赵玉芳, 2011 ), 即生长变化率超过一定的标准便可视为生长释放发生, 该方法是目前应用最多的方法(封晓辉等, 2011 ).本研究中林窗木与非林窗木的生长变化百分率有很强的一致性(仅局部存在部分差异), 说明林窗木生长变化百分率序列中包含了部分非干扰信号.通过生长释放判定得到的生长释放中必定会有部分噪音即非干扰信号, 即并不是所有的释放(包括中等释放和主要释放)都能代表林窗干扰(王晓春和赵玉芳, 2011 ), 因此主要释放或部分主要释放才能代表真正意义上的干扰. 生长变化百分率差值图(图1 )与干扰年表(图2 , 图3 )谷、峰值具有很强的一致性, 且干扰年表在一定程度上对生长变化百分率差值图中不太明显的峰值具有局部放大作用, 经局部放大的部分信号可能是噪音的主要来源.因此, 通过林窗边缘木生长释放判定得出的干扰年表, 能够很好地反映该区阔叶红松林的干扰历史, 且能将一些不太明显的微小干扰包括其中. ...
... ).为了保留更多的干扰信号, 本文选用生长释放标准曲线法(王晓春和赵玉芳, 2011 ), 即生长变化率超过一定的标准便可视为生长释放发生, 该方法是目前应用最多的方法(封晓辉等, 2011 ).本研究中林窗木与非林窗木的生长变化百分率有很强的一致性(仅局部存在部分差异), 说明林窗木生长变化百分率序列中包含了部分非干扰信号.通过生长释放判定得到的生长释放中必定会有部分噪音即非干扰信号, 即并不是所有的释放(包括中等释放和主要释放)都能代表林窗干扰(王晓春和赵玉芳, 2011 ), 因此主要释放或部分主要释放才能代表真正意义上的干扰. 生长变化百分率差值图(图1 )与干扰年表(图2 , 图3 )谷、峰值具有很强的一致性, 且干扰年表在一定程度上对生长变化百分率差值图中不太明显的峰值具有局部放大作用, 经局部放大的部分信号可能是噪音的主要来源.因此, 通过林窗边缘木生长释放判定得出的干扰年表, 能够很好地反映该区阔叶红松林的干扰历史, 且能将一些不太明显的微小干扰包括其中. ...
... ).本研究中林窗木与非林窗木的生长变化百分率有很强的一致性(仅局部存在部分差异), 说明林窗木生长变化百分率序列中包含了部分非干扰信号.通过生长释放判定得到的生长释放中必定会有部分噪音即非干扰信号, 即并不是所有的释放(包括中等释放和主要释放)都能代表林窗干扰(王晓春和赵玉芳, 2011 ), 因此主要释放或部分主要释放才能代表真正意义上的干扰. 生长变化百分率差值图(图1 )与干扰年表(图2 , 图3 )谷、峰值具有很强的一致性, 且干扰年表在一定程度上对生长变化百分率差值图中不太明显的峰值具有局部放大作用, 经局部放大的部分信号可能是噪音的主要来源.因此, 通过林窗边缘木生长释放判定得出的干扰年表, 能够很好地反映该区阔叶红松林的干扰历史, 且能将一些不太明显的微小干扰包括其中. ...
... 本研究发现风干扰是小兴安岭原始阔叶红松林树木生长释放的一个主要因素, 风干扰是该区阔叶红松林干扰的主要类型, 这与其他学者在长白山(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭(王晓春和赵玉芳, 2011 ; 赵玉芳, 2011 )及俄罗斯远东(Ishikawa et al ., 1999 )等地阔叶红松林干扰研究的结果一致.风干扰造成树体倾倒(尤其是林冠层乔木, 如红松)形成林窗(图9 ), 林窗的出现解除了周围树木生长的竞争, 林窗边缘木将出现明显的生长释放(王晓春和赵玉芳, 2011 ).例如, 研究区春季(4-5月)多风且风速较大, 春季最大风速平均值前10个最大年份分别为1971、1976、1987、1989-1990、1994-1996和1998-1999年, 这些年份当年或后几年红松生长变化百分率均出现明显增加的趋势.臭冷杉由于是喜阴树种且多处于林冠下层, 生长释放不太明显.此外, 林窗内形成木数量、腐烂等级以及林窗边缘木生长释放特征, 很好地证明了森林遭大风袭击后摧毁了部分林冠层树木, 造成林冠参差不齐.林窗形成后, 该区对风的抵抗力便会大大削弱, 林分很容易发生持续的风倒, 产生多林窗潮(multiple-gap episodes)(阳含熙和谢海生, 1994 ; 臧润国和徐化成, 1998 ). ...
... ), 林窗的出现解除了周围树木生长的竞争, 林窗边缘木将出现明显的生长释放(王晓春和赵玉芳, 2011 ).例如, 研究区春季(4-5月)多风且风速较大, 春季最大风速平均值前10个最大年份分别为1971、1976、1987、1989-1990、1994-1996和1998-1999年, 这些年份当年或后几年红松生长变化百分率均出现明显增加的趋势.臭冷杉由于是喜阴树种且多处于林冠下层, 生长释放不太明显.此外, 林窗内形成木数量、腐烂等级以及林窗边缘木生长释放特征, 很好地证明了森林遭大风袭击后摧毁了部分林冠层树木, 造成林冠参差不齐.林窗形成后, 该区对风的抵抗力便会大大削弱, 林分很容易发生持续的风倒, 产生多林窗潮(multiple-gap episodes)(阳含熙和谢海生, 1994 ; 臧润国和徐化成, 1998 ). ...
干扰生态学: 一门必须重视的学科
1
2010
... 本研究发现小兴安岭地区林窗木与非林窗木的年生长变化规律(平均生长变化百分率)基本一致, 而以林窗为单位得到的林窗木生长变化百分率与非林窗木之间的差异却很明显, 可能是不同林窗或同一林窗不同树木对同一干扰事件的响应存在差异的结果.自然干扰及其所产生的影响存在巨大的变异性(魏晓华, 2010 ), 可能因海拔、坡度和坡位等因素而出现不同的格局(沈泽昊等, 2001 ).刘妍妍和金光泽(2009)在分析本研究样地内粗木质残体与地形因子的关系时, 也得到了类似的结论.林窗木生长释放特征也具有极大的变异性, 如臭冷杉发生中等以上释放每株2.16次,介于每株0-6次之间; 初次释放平均年龄为32.14 a, 介于15-116 a; 平均间隔期为16.56 a, 介于2-67 a, 从另一方面印证了小兴安岭地区典型阔叶红松林干扰及其所产生的影响存在巨大的变异性.夏冰等(1997)在亚高山云冷杉混交林得到了类似的研究结果.此外, 本研究还发现了一些连续多年的生长释放, 如1889-1904、1932-1938、1947-1973年等, 这可能是连续发生释放或者间隔释放相叠加的结果, 也可能是区域尺度上大范围干扰事件造成的. ...
干扰生态学: 一门必须重视的学科
1
2010
... 本研究发现小兴安岭地区林窗木与非林窗木的年生长变化规律(平均生长变化百分率)基本一致, 而以林窗为单位得到的林窗木生长变化百分率与非林窗木之间的差异却很明显, 可能是不同林窗或同一林窗不同树木对同一干扰事件的响应存在差异的结果.自然干扰及其所产生的影响存在巨大的变异性(魏晓华, 2010 ), 可能因海拔、坡度和坡位等因素而出现不同的格局(沈泽昊等, 2001 ).刘妍妍和金光泽(2009)在分析本研究样地内粗木质残体与地形因子的关系时, 也得到了类似的结论.林窗木生长释放特征也具有极大的变异性, 如臭冷杉发生中等以上释放每株2.16次,介于每株0-6次之间; 初次释放平均年龄为32.14 a, 介于15-116 a; 平均间隔期为16.56 a, 介于2-67 a, 从另一方面印证了小兴安岭地区典型阔叶红松林干扰及其所产生的影响存在巨大的变异性.夏冰等(1997)在亚高山云冷杉混交林得到了类似的研究结果.此外, 本研究还发现了一些连续多年的生长释放, 如1889-1904、1932-1938、1947-1973年等, 这可能是连续发生释放或者间隔释放相叠加的结果, 也可能是区域尺度上大范围干扰事件造成的. ...
树轮分析用于森林干扰强度推测的实例研究
2
2001
... 国外已有一些研究利用树木年轮重建森林干扰历史(Nowacki & Abrams, 1997 ; Ishikawa et al ., 1999 ; Rubino & McCarthyz, 2004 ; Brown & Giesecke, 2014 ), 而国内这方面研究起步较晚, 且相对较少.20世纪90年代初, 阳含熙和谢海生(1994)最先利用树木生长释放和抑制对长白山红松(Pinus koraiensis )混交林干扰历史进行了重建, 并指出该方法是以风作为主要干扰类型, 重建干扰历史最准确、最可靠、提供信息量最大的方法; 侯向阳等(2000)研究长白山红松林干扰节律时, 也得到了类似的结论.乔佃锋(2005)利用该方法(阳含熙和谢海生, 1994 )重建了长白山敦化地区阔叶红松林的干扰历史, 发现该区红松林干扰以小规模的风干扰为主, 1980年出现了明显的干扰峰, 干扰峰之间60年的时间间隔正好与红松的更替世代相吻合.王晓春和赵玉芳(2011)发现前一年径向生长对生长释放事件的判定很重要, 小兴安岭北端黑河地区的红松和红皮云杉(Picea koraiensis )的生长释放可能受林窗产生、微环境变化以及温度和降水等气候因素影响.夏冰等(1995)在天目山建立了生长释放和抑制的判定标准, 并重建了黄山松(Pinus taiwanensis )阔叶林树冠的干扰历史; 同时, 还对不同树种年轮宽度的生长释放判别方法进行了研究, 并做了重建干扰强度的尝试(夏冰等, 1997 , 2001 ).此外, 不同学者还利用树轮生态学方法对火灾(张磊和王晓春, 2013 )、泥石流(铁永波等, 2014 )、山体滑坡(洪婷等, 2012 )和冰川活动(Zhu et al ., 2013 )等干扰进行了重建. ...
... 本文采用王晓春和赵玉芳(2011)的方法计算当年年轮宽度序列的生长变化百分率(Percentage Growth Change, GC).该方法在Nowacki和Abrams (1997)的方法基础上, 将滑动平均值法修正为中值法, 并将滑动窗口由10 a调整为5 a, 以凸显非正常干扰事件的影响(Rubino & McCarthyz, 2004 ; 王晓春和赵玉芳, 2011 ).研究中以林窗为单位计算臭冷杉和红松的生长释放标准曲线, 并利用该标准曲线判断每一边缘木的生长释放情况.将GC介于25%-50%之间且持续5 a以上的释放记为“中等释放”, GC > 50%且持续5 a以上的释放记为“主要释放”, GC < 25%则不考虑为真正释放.林窗木与非林窗木生长变化百分率的差值可以在很大程度上反映林窗的干扰情况.首先, 求出样地内每一个林窗所有边缘木生长变化百分率的平均值; 其次, 将各平均值逐一与非林窗木生长变化百分率的平均值作差, 得出各个林窗干扰情况随时间的变化序列; 最后, 再将得到的44个时间序列每一年的值, 按正、负进行求和后便得出柱状累计图(图1 ).本文以平均每株树木经历生长释放次数的倒数乘以生长释放频率, 推测过去树木的死亡率即干扰强度(夏冰等, 1997 , 2001 ).例如, 臭冷杉经历的平均主要生长释放次数为1.05次, 其平均生长释放频率为每10 a 8.79%, 则其平均干扰强度是每10 a 8.37% (表1 ). ...
树轮分析用于森林干扰强度推测的实例研究
2
2001
... 国外已有一些研究利用树木年轮重建森林干扰历史(Nowacki & Abrams, 1997 ; Ishikawa et al ., 1999 ; Rubino & McCarthyz, 2004 ; Brown & Giesecke, 2014 ), 而国内这方面研究起步较晚, 且相对较少.20世纪90年代初, 阳含熙和谢海生(1994)最先利用树木生长释放和抑制对长白山红松(Pinus koraiensis )混交林干扰历史进行了重建, 并指出该方法是以风作为主要干扰类型, 重建干扰历史最准确、最可靠、提供信息量最大的方法; 侯向阳等(2000)研究长白山红松林干扰节律时, 也得到了类似的结论.乔佃锋(2005)利用该方法(阳含熙和谢海生, 1994 )重建了长白山敦化地区阔叶红松林的干扰历史, 发现该区红松林干扰以小规模的风干扰为主, 1980年出现了明显的干扰峰, 干扰峰之间60年的时间间隔正好与红松的更替世代相吻合.王晓春和赵玉芳(2011)发现前一年径向生长对生长释放事件的判定很重要, 小兴安岭北端黑河地区的红松和红皮云杉(Picea koraiensis )的生长释放可能受林窗产生、微环境变化以及温度和降水等气候因素影响.夏冰等(1995)在天目山建立了生长释放和抑制的判定标准, 并重建了黄山松(Pinus taiwanensis )阔叶林树冠的干扰历史; 同时, 还对不同树种年轮宽度的生长释放判别方法进行了研究, 并做了重建干扰强度的尝试(夏冰等, 1997 , 2001 ).此外, 不同学者还利用树轮生态学方法对火灾(张磊和王晓春, 2013 )、泥石流(铁永波等, 2014 )、山体滑坡(洪婷等, 2012 )和冰川活动(Zhu et al ., 2013 )等干扰进行了重建. ...
... 本文采用王晓春和赵玉芳(2011)的方法计算当年年轮宽度序列的生长变化百分率(Percentage Growth Change, GC).该方法在Nowacki和Abrams (1997)的方法基础上, 将滑动平均值法修正为中值法, 并将滑动窗口由10 a调整为5 a, 以凸显非正常干扰事件的影响(Rubino & McCarthyz, 2004 ; 王晓春和赵玉芳, 2011 ).研究中以林窗为单位计算臭冷杉和红松的生长释放标准曲线, 并利用该标准曲线判断每一边缘木的生长释放情况.将GC介于25%-50%之间且持续5 a以上的释放记为“中等释放”, GC > 50%且持续5 a以上的释放记为“主要释放”, GC < 25%则不考虑为真正释放.林窗木与非林窗木生长变化百分率的差值可以在很大程度上反映林窗的干扰情况.首先, 求出样地内每一个林窗所有边缘木生长变化百分率的平均值; 其次, 将各平均值逐一与非林窗木生长变化百分率的平均值作差, 得出各个林窗干扰情况随时间的变化序列; 最后, 再将得到的44个时间序列每一年的值, 按正、负进行求和后便得出柱状累计图(图1 ).本文以平均每株树木经历生长释放次数的倒数乘以生长释放频率, 推测过去树木的死亡率即干扰强度(夏冰等, 1997 , 2001 ).例如, 臭冷杉经历的平均主要生长释放次数为1.05次, 其平均生长释放频率为每10 a 8.79%, 则其平均干扰强度是每10 a 8.37% (表1 ). ...
亚高山云冷杉混交林树木生长释放与干扰分析
3
1997
... 国外已有一些研究利用树木年轮重建森林干扰历史(Nowacki & Abrams, 1997 ; Ishikawa et al ., 1999 ; Rubino & McCarthyz, 2004 ; Brown & Giesecke, 2014 ), 而国内这方面研究起步较晚, 且相对较少.20世纪90年代初, 阳含熙和谢海生(1994)最先利用树木生长释放和抑制对长白山红松(Pinus koraiensis )混交林干扰历史进行了重建, 并指出该方法是以风作为主要干扰类型, 重建干扰历史最准确、最可靠、提供信息量最大的方法; 侯向阳等(2000)研究长白山红松林干扰节律时, 也得到了类似的结论.乔佃锋(2005)利用该方法(阳含熙和谢海生, 1994 )重建了长白山敦化地区阔叶红松林的干扰历史, 发现该区红松林干扰以小规模的风干扰为主, 1980年出现了明显的干扰峰, 干扰峰之间60年的时间间隔正好与红松的更替世代相吻合.王晓春和赵玉芳(2011)发现前一年径向生长对生长释放事件的判定很重要, 小兴安岭北端黑河地区的红松和红皮云杉(Picea koraiensis )的生长释放可能受林窗产生、微环境变化以及温度和降水等气候因素影响.夏冰等(1995)在天目山建立了生长释放和抑制的判定标准, 并重建了黄山松(Pinus taiwanensis )阔叶林树冠的干扰历史; 同时, 还对不同树种年轮宽度的生长释放判别方法进行了研究, 并做了重建干扰强度的尝试(夏冰等, 1997 , 2001 ).此外, 不同学者还利用树轮生态学方法对火灾(张磊和王晓春, 2013 )、泥石流(铁永波等, 2014 )、山体滑坡(洪婷等, 2012 )和冰川活动(Zhu et al ., 2013 )等干扰进行了重建. ...
... 本文采用王晓春和赵玉芳(2011)的方法计算当年年轮宽度序列的生长变化百分率(Percentage Growth Change, GC).该方法在Nowacki和Abrams (1997)的方法基础上, 将滑动平均值法修正为中值法, 并将滑动窗口由10 a调整为5 a, 以凸显非正常干扰事件的影响(Rubino & McCarthyz, 2004 ; 王晓春和赵玉芳, 2011 ).研究中以林窗为单位计算臭冷杉和红松的生长释放标准曲线, 并利用该标准曲线判断每一边缘木的生长释放情况.将GC介于25%-50%之间且持续5 a以上的释放记为“中等释放”, GC > 50%且持续5 a以上的释放记为“主要释放”, GC < 25%则不考虑为真正释放.林窗木与非林窗木生长变化百分率的差值可以在很大程度上反映林窗的干扰情况.首先, 求出样地内每一个林窗所有边缘木生长变化百分率的平均值; 其次, 将各平均值逐一与非林窗木生长变化百分率的平均值作差, 得出各个林窗干扰情况随时间的变化序列; 最后, 再将得到的44个时间序列每一年的值, 按正、负进行求和后便得出柱状累计图(图1 ).本文以平均每株树木经历生长释放次数的倒数乘以生长释放频率, 推测过去树木的死亡率即干扰强度(夏冰等, 1997 , 2001 ).例如, 臭冷杉经历的平均主要生长释放次数为1.05次, 其平均生长释放频率为每10 a 8.79%, 则其平均干扰强度是每10 a 8.37% (表1 ). ...
... 生长释放判定方法的选择及判定标准的确定是树轮干扰历史重建的关键.判定方法和标准不同, 得出的生长释放结果亦不同(Rubino & McCarthyz, 2004 ), 每一种方法都有各自的优缺点及使用范围(封晓辉等, 2011 ).在实践中, 通过主观判断去选择生长释放判定方法及判定标准, 往往会因主观原因放大或弱化干扰事件.因此, 在判定方法及判定标准选择时, 在充分考虑研究需要的同时, 还需结合研究地点及环境条件等合理选择(阳含熙和谢海生, 1994 ; 夏冰等, 1997 ; 王晓春和赵玉芳, 2011 ).为了保留更多的干扰信号, 本文选用生长释放标准曲线法(王晓春和赵玉芳, 2011 ), 即生长变化率超过一定的标准便可视为生长释放发生, 该方法是目前应用最多的方法(封晓辉等, 2011 ).本研究中林窗木与非林窗木的生长变化百分率有很强的一致性(仅局部存在部分差异), 说明林窗木生长变化百分率序列中包含了部分非干扰信号.通过生长释放判定得到的生长释放中必定会有部分噪音即非干扰信号, 即并不是所有的释放(包括中等释放和主要释放)都能代表林窗干扰(王晓春和赵玉芳, 2011 ), 因此主要释放或部分主要释放才能代表真正意义上的干扰. 生长变化百分率差值图(图1 )与干扰年表(图2 , 图3 )谷、峰值具有很强的一致性, 且干扰年表在一定程度上对生长变化百分率差值图中不太明显的峰值具有局部放大作用, 经局部放大的部分信号可能是噪音的主要来源.因此, 通过林窗边缘木生长释放判定得出的干扰年表, 能够很好地反映该区阔叶红松林的干扰历史, 且能将一些不太明显的微小干扰包括其中. ...
亚高山云冷杉混交林树木生长释放与干扰分析
3
1997
... 国外已有一些研究利用树木年轮重建森林干扰历史(Nowacki & Abrams, 1997 ; Ishikawa et al ., 1999 ; Rubino & McCarthyz, 2004 ; Brown & Giesecke, 2014 ), 而国内这方面研究起步较晚, 且相对较少.20世纪90年代初, 阳含熙和谢海生(1994)最先利用树木生长释放和抑制对长白山红松(Pinus koraiensis )混交林干扰历史进行了重建, 并指出该方法是以风作为主要干扰类型, 重建干扰历史最准确、最可靠、提供信息量最大的方法; 侯向阳等(2000)研究长白山红松林干扰节律时, 也得到了类似的结论.乔佃锋(2005)利用该方法(阳含熙和谢海生, 1994 )重建了长白山敦化地区阔叶红松林的干扰历史, 发现该区红松林干扰以小规模的风干扰为主, 1980年出现了明显的干扰峰, 干扰峰之间60年的时间间隔正好与红松的更替世代相吻合.王晓春和赵玉芳(2011)发现前一年径向生长对生长释放事件的判定很重要, 小兴安岭北端黑河地区的红松和红皮云杉(Picea koraiensis )的生长释放可能受林窗产生、微环境变化以及温度和降水等气候因素影响.夏冰等(1995)在天目山建立了生长释放和抑制的判定标准, 并重建了黄山松(Pinus taiwanensis )阔叶林树冠的干扰历史; 同时, 还对不同树种年轮宽度的生长释放判别方法进行了研究, 并做了重建干扰强度的尝试(夏冰等, 1997 , 2001 ).此外, 不同学者还利用树轮生态学方法对火灾(张磊和王晓春, 2013 )、泥石流(铁永波等, 2014 )、山体滑坡(洪婷等, 2012 )和冰川活动(Zhu et al ., 2013 )等干扰进行了重建. ...
... 本文采用王晓春和赵玉芳(2011)的方法计算当年年轮宽度序列的生长变化百分率(Percentage Growth Change, GC).该方法在Nowacki和Abrams (1997)的方法基础上, 将滑动平均值法修正为中值法, 并将滑动窗口由10 a调整为5 a, 以凸显非正常干扰事件的影响(Rubino & McCarthyz, 2004 ; 王晓春和赵玉芳, 2011 ).研究中以林窗为单位计算臭冷杉和红松的生长释放标准曲线, 并利用该标准曲线判断每一边缘木的生长释放情况.将GC介于25%-50%之间且持续5 a以上的释放记为“中等释放”, GC > 50%且持续5 a以上的释放记为“主要释放”, GC < 25%则不考虑为真正释放.林窗木与非林窗木生长变化百分率的差值可以在很大程度上反映林窗的干扰情况.首先, 求出样地内每一个林窗所有边缘木生长变化百分率的平均值; 其次, 将各平均值逐一与非林窗木生长变化百分率的平均值作差, 得出各个林窗干扰情况随时间的变化序列; 最后, 再将得到的44个时间序列每一年的值, 按正、负进行求和后便得出柱状累计图(图1 ).本文以平均每株树木经历生长释放次数的倒数乘以生长释放频率, 推测过去树木的死亡率即干扰强度(夏冰等, 1997 , 2001 ).例如, 臭冷杉经历的平均主要生长释放次数为1.05次, 其平均生长释放频率为每10 a 8.79%, 则其平均干扰强度是每10 a 8.37% (表1 ). ...
... 生长释放判定方法的选择及判定标准的确定是树轮干扰历史重建的关键.判定方法和标准不同, 得出的生长释放结果亦不同(Rubino & McCarthyz, 2004 ), 每一种方法都有各自的优缺点及使用范围(封晓辉等, 2011 ).在实践中, 通过主观判断去选择生长释放判定方法及判定标准, 往往会因主观原因放大或弱化干扰事件.因此, 在判定方法及判定标准选择时, 在充分考虑研究需要的同时, 还需结合研究地点及环境条件等合理选择(阳含熙和谢海生, 1994 ; 夏冰等, 1997 ; 王晓春和赵玉芳, 2011 ).为了保留更多的干扰信号, 本文选用生长释放标准曲线法(王晓春和赵玉芳, 2011 ), 即生长变化率超过一定的标准便可视为生长释放发生, 该方法是目前应用最多的方法(封晓辉等, 2011 ).本研究中林窗木与非林窗木的生长变化百分率有很强的一致性(仅局部存在部分差异), 说明林窗木生长变化百分率序列中包含了部分非干扰信号.通过生长释放判定得到的生长释放中必定会有部分噪音即非干扰信号, 即并不是所有的释放(包括中等释放和主要释放)都能代表林窗干扰(王晓春和赵玉芳, 2011 ), 因此主要释放或部分主要释放才能代表真正意义上的干扰. 生长变化百分率差值图(图1 )与干扰年表(图2 , 图3 )谷、峰值具有很强的一致性, 且干扰年表在一定程度上对生长变化百分率差值图中不太明显的峰值具有局部放大作用, 经局部放大的部分信号可能是噪音的主要来源.因此, 通过林窗边缘木生长释放判定得出的干扰年表, 能够很好地反映该区阔叶红松林的干扰历史, 且能将一些不太明显的微小干扰包括其中. ...
西天目山黄山松阔叶林的冠层干扰与动态推测
1
1995
... 林窗干扰是自然界中最常见的一种森林干扰(臧润国和徐化成, 1998 ; 臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).林窗干扰发生后, 周围树木由于竞争去除而产生生长释放(王晓春和赵玉芳, 2011 ), 并且这种释放可以持续多年.通常将大于一定阈值且持续5 a或10 a以上的生长释放视为“真正释放”, 并将生长释放开始的时间视为干扰的发生时间(Black & Abrams, 2004 ; Rubino & McCarthyz, 2004 ; 王晓春和赵玉芳, 2011 ).确定生长释放的发生时间是重建森林干扰历史的最基本过程(Black & Abrams, 2004 ), 林窗形成时间常以林窗内幼苗更新时间(刘少冲等, 2013 )、倒木倾倒时间(用腐烂等级确定, 韩路等, 2011 )以及林窗周边树木生长释放时间(阳含熙和谢海生, 1994 ; 夏冰等, 1995 )等确定.本研究林窗干扰时间的确定主要用树木生长释放来判断, 同时还参考了林窗内的幼苗更新及倒木腐烂等级, 但有些林窗更新的非均一性以及林窗内倒木腐烂情况比较复杂也影响林窗形成时间的确定. ...
西天目山黄山松阔叶林的冠层干扰与动态推测
1
1995
... 林窗干扰是自然界中最常见的一种森林干扰(臧润国和徐化成, 1998 ; 臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).林窗干扰发生后, 周围树木由于竞争去除而产生生长释放(王晓春和赵玉芳, 2011 ), 并且这种释放可以持续多年.通常将大于一定阈值且持续5 a或10 a以上的生长释放视为“真正释放”, 并将生长释放开始的时间视为干扰的发生时间(Black & Abrams, 2004 ; Rubino & McCarthyz, 2004 ; 王晓春和赵玉芳, 2011 ).确定生长释放的发生时间是重建森林干扰历史的最基本过程(Black & Abrams, 2004 ), 林窗形成时间常以林窗内幼苗更新时间(刘少冲等, 2013 )、倒木倾倒时间(用腐烂等级确定, 韩路等, 2011 )以及林窗周边树木生长释放时间(阳含熙和谢海生, 1994 ; 夏冰等, 1995 )等确定.本研究林窗干扰时间的确定主要用树木生长释放来判断, 同时还参考了林窗内的幼苗更新及倒木腐烂等级, 但有些林窗更新的非均一性以及林窗内倒木腐烂情况比较复杂也影响林窗形成时间的确定. ...
安图红松树轮稳定δ13 C、δ18 O序列记录的气候变化信息
2
2002
... 小兴安岭地区阔叶红松林内红松干扰事件主要发生在18世纪30年代、19世纪10年代、19世纪30-40年代、20世纪20-30年代, 臭冷杉主要发生在19世纪90年代、20世纪30年代、20世纪50-60年代、20世纪90年代.该结果与其他学者在长白山地区(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭地区(赵玉芳, 2011 )以及俄罗斯远东地区(Ishikawa et al ., 1999 )等阔叶红松林主要分布区的研究结果相一致, 说明阔叶红松林干扰事件不只是局限在研究区域内的小范围事件, 而且可能为区域尺度上(整个阔叶红松林分布区)的大范围干扰事件.红松和臭冷杉干扰事件可能受区域性气候模态周期影响(图4 ), 进一步说明阔叶红松林干扰事件是区域尺度上的大范围干扰事件.小兴安岭丰林自然保护区臭冷杉径向生长与AMO和太平洋年代际振荡(Pacific Decadal Oscillation, PDO)有直接关系, AMO、PDO的相位转变可能是谷地臭冷杉衰退的原因(姚启超和王晓春, 2013 ).长白山安图(徐海等, 2002 )和小兴安岭五营(尹红等, 2009 )红松树轮重建的降水或气温序列中也发现了与ENSO等区域性气候模态对应的周期, 也说明红松径向生长受区域性气候模态的影响.大尺度气候波动虽然不会直接影响某一区域树木的生长, 但能通过影响局地气候间接影响树木生长(张先亮等, 2010 ).当这种影响超过一定的阈值时, 势必会导致树木出现生长衰退或死亡.因此, 外力作用下更易形成林窗, 造成周围树体生长释放.另外, 区域气候模态(尤其是ENSO)与东亚季风关系密切(徐袁和钱维宏, 2003 ), 由此形成的风暴更是触发林窗形成的关键原因. ...
... 红松和臭冷杉干扰事件的周期性中, 存在11 a的极显著(p < 0.01)太阳活动周期, 而且红松生长变化百分率与太阳黑子异常值呈显著负相关(p < 0.05) (图10 ), 也表明太阳活动很大程度上影响着该区树木的生长释放.太阳黑子爆发年与红松、臭冷杉生长变化百分率的SEA分析(图11 )表明: 小兴安岭地区红松树轮记录的生长变化百分率由太阳黑子爆发前2-4 a的显著(p < 0.05)负相关, 转变为爆发后1-4 a的正相关; 臭冷杉叠加事件分析的结果与红松基本一致, 但未达到显著水平.太阳活动可通过影响局地气候间接影响树木的生长(徐海等, 2002 ; 尹红等, 2009 ).太阳活动剧烈时, 气温异常值随之增大或减小, 风速异常值也随之增大或减小, 太阳活动通过影响局地的气温和风速变化(表2 )间接影响该区树木生长, 具体表现为造成树木衰退或死亡, 抗干扰能力降低, 最终导致树体间竞争解除或减弱, 产生生长释放.太阳活动不仅直接影响着区域尺度下的气温、降水和风速变化, 还在很大程度上影响着区域性气候模态(赵亮等, 2011 ).因此, 太阳活动通过影响气温、降水和风速等局地气候以及其他大尺度气候模态干扰该地区森林动态, 是小兴安岭典型阔叶红松林可能存在的干扰机制. ...
安图红松树轮稳定δ13 C、δ18 O序列记录的气候变化信息
2
2002
... 小兴安岭地区阔叶红松林内红松干扰事件主要发生在18世纪30年代、19世纪10年代、19世纪30-40年代、20世纪20-30年代, 臭冷杉主要发生在19世纪90年代、20世纪30年代、20世纪50-60年代、20世纪90年代.该结果与其他学者在长白山地区(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭地区(赵玉芳, 2011 )以及俄罗斯远东地区(Ishikawa et al ., 1999 )等阔叶红松林主要分布区的研究结果相一致, 说明阔叶红松林干扰事件不只是局限在研究区域内的小范围事件, 而且可能为区域尺度上(整个阔叶红松林分布区)的大范围干扰事件.红松和臭冷杉干扰事件可能受区域性气候模态周期影响(图4 ), 进一步说明阔叶红松林干扰事件是区域尺度上的大范围干扰事件.小兴安岭丰林自然保护区臭冷杉径向生长与AMO和太平洋年代际振荡(Pacific Decadal Oscillation, PDO)有直接关系, AMO、PDO的相位转变可能是谷地臭冷杉衰退的原因(姚启超和王晓春, 2013 ).长白山安图(徐海等, 2002 )和小兴安岭五营(尹红等, 2009 )红松树轮重建的降水或气温序列中也发现了与ENSO等区域性气候模态对应的周期, 也说明红松径向生长受区域性气候模态的影响.大尺度气候波动虽然不会直接影响某一区域树木的生长, 但能通过影响局地气候间接影响树木生长(张先亮等, 2010 ).当这种影响超过一定的阈值时, 势必会导致树木出现生长衰退或死亡.因此, 外力作用下更易形成林窗, 造成周围树体生长释放.另外, 区域气候模态(尤其是ENSO)与东亚季风关系密切(徐袁和钱维宏, 2003 ), 由此形成的风暴更是触发林窗形成的关键原因. ...
... 红松和臭冷杉干扰事件的周期性中, 存在11 a的极显著(p < 0.01)太阳活动周期, 而且红松生长变化百分率与太阳黑子异常值呈显著负相关(p < 0.05) (图10 ), 也表明太阳活动很大程度上影响着该区树木的生长释放.太阳黑子爆发年与红松、臭冷杉生长变化百分率的SEA分析(图11 )表明: 小兴安岭地区红松树轮记录的生长变化百分率由太阳黑子爆发前2-4 a的显著(p < 0.05)负相关, 转变为爆发后1-4 a的正相关; 臭冷杉叠加事件分析的结果与红松基本一致, 但未达到显著水平.太阳活动可通过影响局地气候间接影响树木的生长(徐海等, 2002 ; 尹红等, 2009 ).太阳活动剧烈时, 气温异常值随之增大或减小, 风速异常值也随之增大或减小, 太阳活动通过影响局地的气温和风速变化(表2 )间接影响该区树木生长, 具体表现为造成树木衰退或死亡, 抗干扰能力降低, 最终导致树体间竞争解除或减弱, 产生生长释放.太阳活动不仅直接影响着区域尺度下的气温、降水和风速变化, 还在很大程度上影响着区域性气候模态(赵亮等, 2011 ).因此, 太阳活动通过影响气温、降水和风速等局地气候以及其他大尺度气候模态干扰该地区森林动态, 是小兴安岭典型阔叶红松林可能存在的干扰机制. ...
小兴安岭凉水典型阔叶红松林动态监测样地: 物种组成与群落结构
1
2012
... 本研究在已有的9 hm2 典型阔叶红松林森林动态监测样地内进行.该样地位于保护区的中段, 从凉水沟谷地到海拔600 m的中山山脊.样地以红松为优势种, 伴生有臭冷杉、水曲柳(Fraxinus mandshurica )、紫椴(Tilia amurensis )、枫桦(Betula costata )、大青杨(Populus ussuriensis )、裂叶榆(Ulmus laciniata )、色木槭(Acer mono )和花楷槭(Acer ukurunduense )等20余种针阔叶树种; 主要下木有刺五加(Acanthopanax senticosus )、毛榛子(Corylus mand- shurica )、东北山梅花(Philadelphus schrenkii )等.草本植物主要有薹草(Carex spp.)、东北蹄盖蕨(Athyrium brevifrons )和东北羊角芹(Aegopodium alpestre )等, 藤本植物有五味子(Schisandra chinensis )、狗枣猕猴桃(Actinidia kolomikta )等(徐丽娜和金光泽, 2012 ). ...
小兴安岭凉水典型阔叶红松林动态监测样地: 物种组成与群落结构
1
2012
... 本研究在已有的9 hm2 典型阔叶红松林森林动态监测样地内进行.该样地位于保护区的中段, 从凉水沟谷地到海拔600 m的中山山脊.样地以红松为优势种, 伴生有臭冷杉、水曲柳(Fraxinus mandshurica )、紫椴(Tilia amurensis )、枫桦(Betula costata )、大青杨(Populus ussuriensis )、裂叶榆(Ulmus laciniata )、色木槭(Acer mono )和花楷槭(Acer ukurunduense )等20余种针阔叶树种; 主要下木有刺五加(Acanthopanax senticosus )、毛榛子(Corylus mand- shurica )、东北山梅花(Philadelphus schrenkii )等.草本植物主要有薹草(Carex spp.)、东北蹄盖蕨(Athyrium brevifrons )和东北羊角芹(Aegopodium alpestre )等, 藤本植物有五味子(Schisandra chinensis )、狗枣猕猴桃(Actinidia kolomikta )等(徐丽娜和金光泽, 2012 ). ...
东亚季风边缘活动带研究综述
1
2003
... 小兴安岭地区阔叶红松林内红松干扰事件主要发生在18世纪30年代、19世纪10年代、19世纪30-40年代、20世纪20-30年代, 臭冷杉主要发生在19世纪90年代、20世纪30年代、20世纪50-60年代、20世纪90年代.该结果与其他学者在长白山地区(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭地区(赵玉芳, 2011 )以及俄罗斯远东地区(Ishikawa et al ., 1999 )等阔叶红松林主要分布区的研究结果相一致, 说明阔叶红松林干扰事件不只是局限在研究区域内的小范围事件, 而且可能为区域尺度上(整个阔叶红松林分布区)的大范围干扰事件.红松和臭冷杉干扰事件可能受区域性气候模态周期影响(图4 ), 进一步说明阔叶红松林干扰事件是区域尺度上的大范围干扰事件.小兴安岭丰林自然保护区臭冷杉径向生长与AMO和太平洋年代际振荡(Pacific Decadal Oscillation, PDO)有直接关系, AMO、PDO的相位转变可能是谷地臭冷杉衰退的原因(姚启超和王晓春, 2013 ).长白山安图(徐海等, 2002 )和小兴安岭五营(尹红等, 2009 )红松树轮重建的降水或气温序列中也发现了与ENSO等区域性气候模态对应的周期, 也说明红松径向生长受区域性气候模态的影响.大尺度气候波动虽然不会直接影响某一区域树木的生长, 但能通过影响局地气候间接影响树木生长(张先亮等, 2010 ).当这种影响超过一定的阈值时, 势必会导致树木出现生长衰退或死亡.因此, 外力作用下更易形成林窗, 造成周围树体生长释放.另外, 区域气候模态(尤其是ENSO)与东亚季风关系密切(徐袁和钱维宏, 2003 ), 由此形成的风暴更是触发林窗形成的关键原因. ...
东亚季风边缘活动带研究综述
1
2003
... 小兴安岭地区阔叶红松林内红松干扰事件主要发生在18世纪30年代、19世纪10年代、19世纪30-40年代、20世纪20-30年代, 臭冷杉主要发生在19世纪90年代、20世纪30年代、20世纪50-60年代、20世纪90年代.该结果与其他学者在长白山地区(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭地区(赵玉芳, 2011 )以及俄罗斯远东地区(Ishikawa et al ., 1999 )等阔叶红松林主要分布区的研究结果相一致, 说明阔叶红松林干扰事件不只是局限在研究区域内的小范围事件, 而且可能为区域尺度上(整个阔叶红松林分布区)的大范围干扰事件.红松和臭冷杉干扰事件可能受区域性气候模态周期影响(图4 ), 进一步说明阔叶红松林干扰事件是区域尺度上的大范围干扰事件.小兴安岭丰林自然保护区臭冷杉径向生长与AMO和太平洋年代际振荡(Pacific Decadal Oscillation, PDO)有直接关系, AMO、PDO的相位转变可能是谷地臭冷杉衰退的原因(姚启超和王晓春, 2013 ).长白山安图(徐海等, 2002 )和小兴安岭五营(尹红等, 2009 )红松树轮重建的降水或气温序列中也发现了与ENSO等区域性气候模态对应的周期, 也说明红松径向生长受区域性气候模态的影响.大尺度气候波动虽然不会直接影响某一区域树木的生长, 但能通过影响局地气候间接影响树木生长(张先亮等, 2010 ).当这种影响超过一定的阈值时, 势必会导致树木出现生长衰退或死亡.因此, 外力作用下更易形成林窗, 造成周围树体生长释放.另外, 区域气候模态(尤其是ENSO)与东亚季风关系密切(徐袁和钱维宏, 2003 ), 由此形成的风暴更是触发林窗形成的关键原因. ...
长白山红松混交林干扰历史的重构研究
7
1994
... 国外已有一些研究利用树木年轮重建森林干扰历史(Nowacki & Abrams, 1997 ; Ishikawa et al ., 1999 ; Rubino & McCarthyz, 2004 ; Brown & Giesecke, 2014 ), 而国内这方面研究起步较晚, 且相对较少.20世纪90年代初, 阳含熙和谢海生(1994)最先利用树木生长释放和抑制对长白山红松(Pinus koraiensis )混交林干扰历史进行了重建, 并指出该方法是以风作为主要干扰类型, 重建干扰历史最准确、最可靠、提供信息量最大的方法; 侯向阳等(2000)研究长白山红松林干扰节律时, 也得到了类似的结论.乔佃锋(2005)利用该方法(阳含熙和谢海生, 1994 )重建了长白山敦化地区阔叶红松林的干扰历史, 发现该区红松林干扰以小规模的风干扰为主, 1980年出现了明显的干扰峰, 干扰峰之间60年的时间间隔正好与红松的更替世代相吻合.王晓春和赵玉芳(2011)发现前一年径向生长对生长释放事件的判定很重要, 小兴安岭北端黑河地区的红松和红皮云杉(Picea koraiensis )的生长释放可能受林窗产生、微环境变化以及温度和降水等气候因素影响.夏冰等(1995)在天目山建立了生长释放和抑制的判定标准, 并重建了黄山松(Pinus taiwanensis )阔叶林树冠的干扰历史; 同时, 还对不同树种年轮宽度的生长释放判别方法进行了研究, 并做了重建干扰强度的尝试(夏冰等, 1997 , 2001 ).此外, 不同学者还利用树轮生态学方法对火灾(张磊和王晓春, 2013 )、泥石流(铁永波等, 2014 )、山体滑坡(洪婷等, 2012 )和冰川活动(Zhu et al ., 2013 )等干扰进行了重建. ...
... 林窗干扰是自然界中最常见的一种森林干扰(臧润国和徐化成, 1998 ; 臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).林窗干扰发生后, 周围树木由于竞争去除而产生生长释放(王晓春和赵玉芳, 2011 ), 并且这种释放可以持续多年.通常将大于一定阈值且持续5 a或10 a以上的生长释放视为“真正释放”, 并将生长释放开始的时间视为干扰的发生时间(Black & Abrams, 2004 ; Rubino & McCarthyz, 2004 ; 王晓春和赵玉芳, 2011 ).确定生长释放的发生时间是重建森林干扰历史的最基本过程(Black & Abrams, 2004 ), 林窗形成时间常以林窗内幼苗更新时间(刘少冲等, 2013 )、倒木倾倒时间(用腐烂等级确定, 韩路等, 2011 )以及林窗周边树木生长释放时间(阳含熙和谢海生, 1994 ; 夏冰等, 1995 )等确定.本研究林窗干扰时间的确定主要用树木生长释放来判断, 同时还参考了林窗内的幼苗更新及倒木腐烂等级, 但有些林窗更新的非均一性以及林窗内倒木腐烂情况比较复杂也影响林窗形成时间的确定. ...
... 生长释放判定方法的选择及判定标准的确定是树轮干扰历史重建的关键.判定方法和标准不同, 得出的生长释放结果亦不同(Rubino & McCarthyz, 2004 ), 每一种方法都有各自的优缺点及使用范围(封晓辉等, 2011 ).在实践中, 通过主观判断去选择生长释放判定方法及判定标准, 往往会因主观原因放大或弱化干扰事件.因此, 在判定方法及判定标准选择时, 在充分考虑研究需要的同时, 还需结合研究地点及环境条件等合理选择(阳含熙和谢海生, 1994 ; 夏冰等, 1997 ; 王晓春和赵玉芳, 2011 ).为了保留更多的干扰信号, 本文选用生长释放标准曲线法(王晓春和赵玉芳, 2011 ), 即生长变化率超过一定的标准便可视为生长释放发生, 该方法是目前应用最多的方法(封晓辉等, 2011 ).本研究中林窗木与非林窗木的生长变化百分率有很强的一致性(仅局部存在部分差异), 说明林窗木生长变化百分率序列中包含了部分非干扰信号.通过生长释放判定得到的生长释放中必定会有部分噪音即非干扰信号, 即并不是所有的释放(包括中等释放和主要释放)都能代表林窗干扰(王晓春和赵玉芳, 2011 ), 因此主要释放或部分主要释放才能代表真正意义上的干扰. 生长变化百分率差值图(图1 )与干扰年表(图2 , 图3 )谷、峰值具有很强的一致性, 且干扰年表在一定程度上对生长变化百分率差值图中不太明显的峰值具有局部放大作用, 经局部放大的部分信号可能是噪音的主要来源.因此, 通过林窗边缘木生长释放判定得出的干扰年表, 能够很好地反映该区阔叶红松林的干扰历史, 且能将一些不太明显的微小干扰包括其中. ...
... 小兴安岭地区阔叶红松林内红松干扰事件主要发生在18世纪30年代、19世纪10年代、19世纪30-40年代、20世纪20-30年代, 臭冷杉主要发生在19世纪90年代、20世纪30年代、20世纪50-60年代、20世纪90年代.该结果与其他学者在长白山地区(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭地区(赵玉芳, 2011 )以及俄罗斯远东地区(Ishikawa et al ., 1999 )等阔叶红松林主要分布区的研究结果相一致, 说明阔叶红松林干扰事件不只是局限在研究区域内的小范围事件, 而且可能为区域尺度上(整个阔叶红松林分布区)的大范围干扰事件.红松和臭冷杉干扰事件可能受区域性气候模态周期影响(图4 ), 进一步说明阔叶红松林干扰事件是区域尺度上的大范围干扰事件.小兴安岭丰林自然保护区臭冷杉径向生长与AMO和太平洋年代际振荡(Pacific Decadal Oscillation, PDO)有直接关系, AMO、PDO的相位转变可能是谷地臭冷杉衰退的原因(姚启超和王晓春, 2013 ).长白山安图(徐海等, 2002 )和小兴安岭五营(尹红等, 2009 )红松树轮重建的降水或气温序列中也发现了与ENSO等区域性气候模态对应的周期, 也说明红松径向生长受区域性气候模态的影响.大尺度气候波动虽然不会直接影响某一区域树木的生长, 但能通过影响局地气候间接影响树木生长(张先亮等, 2010 ).当这种影响超过一定的阈值时, 势必会导致树木出现生长衰退或死亡.因此, 外力作用下更易形成林窗, 造成周围树体生长释放.另外, 区域气候模态(尤其是ENSO)与东亚季风关系密切(徐袁和钱维宏, 2003 ), 由此形成的风暴更是触发林窗形成的关键原因. ...
... 本研究发现风干扰是小兴安岭原始阔叶红松林树木生长释放的一个主要因素, 风干扰是该区阔叶红松林干扰的主要类型, 这与其他学者在长白山(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭(王晓春和赵玉芳, 2011 ; 赵玉芳, 2011 )及俄罗斯远东(Ishikawa et al ., 1999 )等地阔叶红松林干扰研究的结果一致.风干扰造成树体倾倒(尤其是林冠层乔木, 如红松)形成林窗(图9 ), 林窗的出现解除了周围树木生长的竞争, 林窗边缘木将出现明显的生长释放(王晓春和赵玉芳, 2011 ).例如, 研究区春季(4-5月)多风且风速较大, 春季最大风速平均值前10个最大年份分别为1971、1976、1987、1989-1990、1994-1996和1998-1999年, 这些年份当年或后几年红松生长变化百分率均出现明显增加的趋势.臭冷杉由于是喜阴树种且多处于林冠下层, 生长释放不太明显.此外, 林窗内形成木数量、腐烂等级以及林窗边缘木生长释放特征, 很好地证明了森林遭大风袭击后摧毁了部分林冠层树木, 造成林冠参差不齐.林窗形成后, 该区对风的抵抗力便会大大削弱, 林分很容易发生持续的风倒, 产生多林窗潮(multiple-gap episodes)(阳含熙和谢海生, 1994 ; 臧润国和徐化成, 1998 ). ...
... ).例如, 研究区春季(4-5月)多风且风速较大, 春季最大风速平均值前10个最大年份分别为1971、1976、1987、1989-1990、1994-1996和1998-1999年, 这些年份当年或后几年红松生长变化百分率均出现明显增加的趋势.臭冷杉由于是喜阴树种且多处于林冠下层, 生长释放不太明显.此外, 林窗内形成木数量、腐烂等级以及林窗边缘木生长释放特征, 很好地证明了森林遭大风袭击后摧毁了部分林冠层树木, 造成林冠参差不齐.林窗形成后, 该区对风的抵抗力便会大大削弱, 林分很容易发生持续的风倒, 产生多林窗潮(multiple-gap episodes)(阳含熙和谢海生, 1994 ; 臧润国和徐化成, 1998 ). ...
... 风干扰是阔叶红松林干扰的主要因素之一(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 ), 然而并不意味着风速越大, 形成林窗的数量就越多, 生长释放就越明显.风只是林窗干扰发生的一个外因, 林窗干扰是否发生以及干扰的强弱还取决于树体自身的生长情况及其生存的环境条件.此外, 异常气温也在很大程度上影响着该区树木的生长释放, 这一结果与王晓春和赵玉芳(2011)的研究结果一致.异常气温影响研究区的干旱化现象(贺伟等, 2013 ), 导致该区树木生长衰退或死亡, 对外界干扰的抵抗能力降低或丧失, 在外力作用下更易形成倒木, 进而造成林窗边缘树木发生释放. ...
长白山红松混交林干扰历史的重构研究
7
1994
... 国外已有一些研究利用树木年轮重建森林干扰历史(Nowacki & Abrams, 1997 ; Ishikawa et al ., 1999 ; Rubino & McCarthyz, 2004 ; Brown & Giesecke, 2014 ), 而国内这方面研究起步较晚, 且相对较少.20世纪90年代初, 阳含熙和谢海生(1994)最先利用树木生长释放和抑制对长白山红松(Pinus koraiensis )混交林干扰历史进行了重建, 并指出该方法是以风作为主要干扰类型, 重建干扰历史最准确、最可靠、提供信息量最大的方法; 侯向阳等(2000)研究长白山红松林干扰节律时, 也得到了类似的结论.乔佃锋(2005)利用该方法(阳含熙和谢海生, 1994 )重建了长白山敦化地区阔叶红松林的干扰历史, 发现该区红松林干扰以小规模的风干扰为主, 1980年出现了明显的干扰峰, 干扰峰之间60年的时间间隔正好与红松的更替世代相吻合.王晓春和赵玉芳(2011)发现前一年径向生长对生长释放事件的判定很重要, 小兴安岭北端黑河地区的红松和红皮云杉(Picea koraiensis )的生长释放可能受林窗产生、微环境变化以及温度和降水等气候因素影响.夏冰等(1995)在天目山建立了生长释放和抑制的判定标准, 并重建了黄山松(Pinus taiwanensis )阔叶林树冠的干扰历史; 同时, 还对不同树种年轮宽度的生长释放判别方法进行了研究, 并做了重建干扰强度的尝试(夏冰等, 1997 , 2001 ).此外, 不同学者还利用树轮生态学方法对火灾(张磊和王晓春, 2013 )、泥石流(铁永波等, 2014 )、山体滑坡(洪婷等, 2012 )和冰川活动(Zhu et al ., 2013 )等干扰进行了重建. ...
... 林窗干扰是自然界中最常见的一种森林干扰(臧润国和徐化成, 1998 ; 臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).林窗干扰发生后, 周围树木由于竞争去除而产生生长释放(王晓春和赵玉芳, 2011 ), 并且这种释放可以持续多年.通常将大于一定阈值且持续5 a或10 a以上的生长释放视为“真正释放”, 并将生长释放开始的时间视为干扰的发生时间(Black & Abrams, 2004 ; Rubino & McCarthyz, 2004 ; 王晓春和赵玉芳, 2011 ).确定生长释放的发生时间是重建森林干扰历史的最基本过程(Black & Abrams, 2004 ), 林窗形成时间常以林窗内幼苗更新时间(刘少冲等, 2013 )、倒木倾倒时间(用腐烂等级确定, 韩路等, 2011 )以及林窗周边树木生长释放时间(阳含熙和谢海生, 1994 ; 夏冰等, 1995 )等确定.本研究林窗干扰时间的确定主要用树木生长释放来判断, 同时还参考了林窗内的幼苗更新及倒木腐烂等级, 但有些林窗更新的非均一性以及林窗内倒木腐烂情况比较复杂也影响林窗形成时间的确定. ...
... 生长释放判定方法的选择及判定标准的确定是树轮干扰历史重建的关键.判定方法和标准不同, 得出的生长释放结果亦不同(Rubino & McCarthyz, 2004 ), 每一种方法都有各自的优缺点及使用范围(封晓辉等, 2011 ).在实践中, 通过主观判断去选择生长释放判定方法及判定标准, 往往会因主观原因放大或弱化干扰事件.因此, 在判定方法及判定标准选择时, 在充分考虑研究需要的同时, 还需结合研究地点及环境条件等合理选择(阳含熙和谢海生, 1994 ; 夏冰等, 1997 ; 王晓春和赵玉芳, 2011 ).为了保留更多的干扰信号, 本文选用生长释放标准曲线法(王晓春和赵玉芳, 2011 ), 即生长变化率超过一定的标准便可视为生长释放发生, 该方法是目前应用最多的方法(封晓辉等, 2011 ).本研究中林窗木与非林窗木的生长变化百分率有很强的一致性(仅局部存在部分差异), 说明林窗木生长变化百分率序列中包含了部分非干扰信号.通过生长释放判定得到的生长释放中必定会有部分噪音即非干扰信号, 即并不是所有的释放(包括中等释放和主要释放)都能代表林窗干扰(王晓春和赵玉芳, 2011 ), 因此主要释放或部分主要释放才能代表真正意义上的干扰. 生长变化百分率差值图(图1 )与干扰年表(图2 , 图3 )谷、峰值具有很强的一致性, 且干扰年表在一定程度上对生长变化百分率差值图中不太明显的峰值具有局部放大作用, 经局部放大的部分信号可能是噪音的主要来源.因此, 通过林窗边缘木生长释放判定得出的干扰年表, 能够很好地反映该区阔叶红松林的干扰历史, 且能将一些不太明显的微小干扰包括其中. ...
... 小兴安岭地区阔叶红松林内红松干扰事件主要发生在18世纪30年代、19世纪10年代、19世纪30-40年代、20世纪20-30年代, 臭冷杉主要发生在19世纪90年代、20世纪30年代、20世纪50-60年代、20世纪90年代.该结果与其他学者在长白山地区(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭地区(赵玉芳, 2011 )以及俄罗斯远东地区(Ishikawa et al ., 1999 )等阔叶红松林主要分布区的研究结果相一致, 说明阔叶红松林干扰事件不只是局限在研究区域内的小范围事件, 而且可能为区域尺度上(整个阔叶红松林分布区)的大范围干扰事件.红松和臭冷杉干扰事件可能受区域性气候模态周期影响(图4 ), 进一步说明阔叶红松林干扰事件是区域尺度上的大范围干扰事件.小兴安岭丰林自然保护区臭冷杉径向生长与AMO和太平洋年代际振荡(Pacific Decadal Oscillation, PDO)有直接关系, AMO、PDO的相位转变可能是谷地臭冷杉衰退的原因(姚启超和王晓春, 2013 ).长白山安图(徐海等, 2002 )和小兴安岭五营(尹红等, 2009 )红松树轮重建的降水或气温序列中也发现了与ENSO等区域性气候模态对应的周期, 也说明红松径向生长受区域性气候模态的影响.大尺度气候波动虽然不会直接影响某一区域树木的生长, 但能通过影响局地气候间接影响树木生长(张先亮等, 2010 ).当这种影响超过一定的阈值时, 势必会导致树木出现生长衰退或死亡.因此, 外力作用下更易形成林窗, 造成周围树体生长释放.另外, 区域气候模态(尤其是ENSO)与东亚季风关系密切(徐袁和钱维宏, 2003 ), 由此形成的风暴更是触发林窗形成的关键原因. ...
... 本研究发现风干扰是小兴安岭原始阔叶红松林树木生长释放的一个主要因素, 风干扰是该区阔叶红松林干扰的主要类型, 这与其他学者在长白山(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭(王晓春和赵玉芳, 2011 ; 赵玉芳, 2011 )及俄罗斯远东(Ishikawa et al ., 1999 )等地阔叶红松林干扰研究的结果一致.风干扰造成树体倾倒(尤其是林冠层乔木, 如红松)形成林窗(图9 ), 林窗的出现解除了周围树木生长的竞争, 林窗边缘木将出现明显的生长释放(王晓春和赵玉芳, 2011 ).例如, 研究区春季(4-5月)多风且风速较大, 春季最大风速平均值前10个最大年份分别为1971、1976、1987、1989-1990、1994-1996和1998-1999年, 这些年份当年或后几年红松生长变化百分率均出现明显增加的趋势.臭冷杉由于是喜阴树种且多处于林冠下层, 生长释放不太明显.此外, 林窗内形成木数量、腐烂等级以及林窗边缘木生长释放特征, 很好地证明了森林遭大风袭击后摧毁了部分林冠层树木, 造成林冠参差不齐.林窗形成后, 该区对风的抵抗力便会大大削弱, 林分很容易发生持续的风倒, 产生多林窗潮(multiple-gap episodes)(阳含熙和谢海生, 1994 ; 臧润国和徐化成, 1998 ). ...
... ).例如, 研究区春季(4-5月)多风且风速较大, 春季最大风速平均值前10个最大年份分别为1971、1976、1987、1989-1990、1994-1996和1998-1999年, 这些年份当年或后几年红松生长变化百分率均出现明显增加的趋势.臭冷杉由于是喜阴树种且多处于林冠下层, 生长释放不太明显.此外, 林窗内形成木数量、腐烂等级以及林窗边缘木生长释放特征, 很好地证明了森林遭大风袭击后摧毁了部分林冠层树木, 造成林冠参差不齐.林窗形成后, 该区对风的抵抗力便会大大削弱, 林分很容易发生持续的风倒, 产生多林窗潮(multiple-gap episodes)(阳含熙和谢海生, 1994 ; 臧润国和徐化成, 1998 ). ...
... 风干扰是阔叶红松林干扰的主要因素之一(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 ), 然而并不意味着风速越大, 形成林窗的数量就越多, 生长释放就越明显.风只是林窗干扰发生的一个外因, 林窗干扰是否发生以及干扰的强弱还取决于树体自身的生长情况及其生存的环境条件.此外, 异常气温也在很大程度上影响着该区树木的生长释放, 这一结果与王晓春和赵玉芳(2011)的研究结果一致.异常气温影响研究区的干旱化现象(贺伟等, 2013 ), 导致该区树木生长衰退或死亡, 对外界干扰的抵抗能力降低或丧失, 在外力作用下更易形成倒木, 进而造成林窗边缘树木发生释放. ...
小兴安岭不同海拔臭冷杉年轮-气候关系及大尺度气候影响
2
2013
... 红松干扰年表(生长释放树木数量占总采样树木的比例随时间的变化序列)存在9.1-18.2 a、7.3-7.9 a、3.8 a、3.5 a和2.0 a的极显著(p < 0.01)周期, 臭冷杉干扰年表(生长释放树木数量占总采样树木的比例随时间的变化序列)存在65-85 a、7.5-48.8 a和3.5-3.6 a的极显著周期(图4 ).通常气象因子波动和植物生长具有2-3 a的周期, 红松与臭冷杉干扰年表也记录了该周期; 2-7 a的干扰周期也可能与厄尔尼诺-南方涛动(El Niño-Southern Oscillation, ENSO)事件有关; 9.1-18.2 a的干扰周期可能受太阳活动的影响.而臭冷杉干扰年表中65-85 a的干扰周期可能受北大西洋年代际振荡(Atlantic Multidecadal Oscillation, AMO)的影响, 已有研究表明AMO会影响东北亚树木生长(Wang et al ., 2011 ; 姚启超和王晓春, 2013 ). ...
... 小兴安岭地区阔叶红松林内红松干扰事件主要发生在18世纪30年代、19世纪10年代、19世纪30-40年代、20世纪20-30年代, 臭冷杉主要发生在19世纪90年代、20世纪30年代、20世纪50-60年代、20世纪90年代.该结果与其他学者在长白山地区(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭地区(赵玉芳, 2011 )以及俄罗斯远东地区(Ishikawa et al ., 1999 )等阔叶红松林主要分布区的研究结果相一致, 说明阔叶红松林干扰事件不只是局限在研究区域内的小范围事件, 而且可能为区域尺度上(整个阔叶红松林分布区)的大范围干扰事件.红松和臭冷杉干扰事件可能受区域性气候模态周期影响(图4 ), 进一步说明阔叶红松林干扰事件是区域尺度上的大范围干扰事件.小兴安岭丰林自然保护区臭冷杉径向生长与AMO和太平洋年代际振荡(Pacific Decadal Oscillation, PDO)有直接关系, AMO、PDO的相位转变可能是谷地臭冷杉衰退的原因(姚启超和王晓春, 2013 ).长白山安图(徐海等, 2002 )和小兴安岭五营(尹红等, 2009 )红松树轮重建的降水或气温序列中也发现了与ENSO等区域性气候模态对应的周期, 也说明红松径向生长受区域性气候模态的影响.大尺度气候波动虽然不会直接影响某一区域树木的生长, 但能通过影响局地气候间接影响树木生长(张先亮等, 2010 ).当这种影响超过一定的阈值时, 势必会导致树木出现生长衰退或死亡.因此, 外力作用下更易形成林窗, 造成周围树体生长释放.另外, 区域气候模态(尤其是ENSO)与东亚季风关系密切(徐袁和钱维宏, 2003 ), 由此形成的风暴更是触发林窗形成的关键原因. ...
小兴安岭不同海拔臭冷杉年轮-气候关系及大尺度气候影响
2
2013
... 红松干扰年表(生长释放树木数量占总采样树木的比例随时间的变化序列)存在9.1-18.2 a、7.3-7.9 a、3.8 a、3.5 a和2.0 a的极显著(p < 0.01)周期, 臭冷杉干扰年表(生长释放树木数量占总采样树木的比例随时间的变化序列)存在65-85 a、7.5-48.8 a和3.5-3.6 a的极显著周期(图4 ).通常气象因子波动和植物生长具有2-3 a的周期, 红松与臭冷杉干扰年表也记录了该周期; 2-7 a的干扰周期也可能与厄尔尼诺-南方涛动(El Niño-Southern Oscillation, ENSO)事件有关; 9.1-18.2 a的干扰周期可能受太阳活动的影响.而臭冷杉干扰年表中65-85 a的干扰周期可能受北大西洋年代际振荡(Atlantic Multidecadal Oscillation, AMO)的影响, 已有研究表明AMO会影响东北亚树木生长(Wang et al ., 2011 ; 姚启超和王晓春, 2013 ). ...
... 小兴安岭地区阔叶红松林内红松干扰事件主要发生在18世纪30年代、19世纪10年代、19世纪30-40年代、20世纪20-30年代, 臭冷杉主要发生在19世纪90年代、20世纪30年代、20世纪50-60年代、20世纪90年代.该结果与其他学者在长白山地区(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭地区(赵玉芳, 2011 )以及俄罗斯远东地区(Ishikawa et al ., 1999 )等阔叶红松林主要分布区的研究结果相一致, 说明阔叶红松林干扰事件不只是局限在研究区域内的小范围事件, 而且可能为区域尺度上(整个阔叶红松林分布区)的大范围干扰事件.红松和臭冷杉干扰事件可能受区域性气候模态周期影响(图4 ), 进一步说明阔叶红松林干扰事件是区域尺度上的大范围干扰事件.小兴安岭丰林自然保护区臭冷杉径向生长与AMO和太平洋年代际振荡(Pacific Decadal Oscillation, PDO)有直接关系, AMO、PDO的相位转变可能是谷地臭冷杉衰退的原因(姚启超和王晓春, 2013 ).长白山安图(徐海等, 2002 )和小兴安岭五营(尹红等, 2009 )红松树轮重建的降水或气温序列中也发现了与ENSO等区域性气候模态对应的周期, 也说明红松径向生长受区域性气候模态的影响.大尺度气候波动虽然不会直接影响某一区域树木的生长, 但能通过影响局地气候间接影响树木生长(张先亮等, 2010 ).当这种影响超过一定的阈值时, 势必会导致树木出现生长衰退或死亡.因此, 外力作用下更易形成林窗, 造成周围树体生长释放.另外, 区域气候模态(尤其是ENSO)与东亚季风关系密切(徐袁和钱维宏, 2003 ), 由此形成的风暴更是触发林窗形成的关键原因. ...
利用树轮重建小兴安岭五营1796年以来的温度变化
2
2009
... 小兴安岭地区阔叶红松林内红松干扰事件主要发生在18世纪30年代、19世纪10年代、19世纪30-40年代、20世纪20-30年代, 臭冷杉主要发生在19世纪90年代、20世纪30年代、20世纪50-60年代、20世纪90年代.该结果与其他学者在长白山地区(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭地区(赵玉芳, 2011 )以及俄罗斯远东地区(Ishikawa et al ., 1999 )等阔叶红松林主要分布区的研究结果相一致, 说明阔叶红松林干扰事件不只是局限在研究区域内的小范围事件, 而且可能为区域尺度上(整个阔叶红松林分布区)的大范围干扰事件.红松和臭冷杉干扰事件可能受区域性气候模态周期影响(图4 ), 进一步说明阔叶红松林干扰事件是区域尺度上的大范围干扰事件.小兴安岭丰林自然保护区臭冷杉径向生长与AMO和太平洋年代际振荡(Pacific Decadal Oscillation, PDO)有直接关系, AMO、PDO的相位转变可能是谷地臭冷杉衰退的原因(姚启超和王晓春, 2013 ).长白山安图(徐海等, 2002 )和小兴安岭五营(尹红等, 2009 )红松树轮重建的降水或气温序列中也发现了与ENSO等区域性气候模态对应的周期, 也说明红松径向生长受区域性气候模态的影响.大尺度气候波动虽然不会直接影响某一区域树木的生长, 但能通过影响局地气候间接影响树木生长(张先亮等, 2010 ).当这种影响超过一定的阈值时, 势必会导致树木出现生长衰退或死亡.因此, 外力作用下更易形成林窗, 造成周围树体生长释放.另外, 区域气候模态(尤其是ENSO)与东亚季风关系密切(徐袁和钱维宏, 2003 ), 由此形成的风暴更是触发林窗形成的关键原因. ...
... 红松和臭冷杉干扰事件的周期性中, 存在11 a的极显著(p < 0.01)太阳活动周期, 而且红松生长变化百分率与太阳黑子异常值呈显著负相关(p < 0.05) (图10 ), 也表明太阳活动很大程度上影响着该区树木的生长释放.太阳黑子爆发年与红松、臭冷杉生长变化百分率的SEA分析(图11 )表明: 小兴安岭地区红松树轮记录的生长变化百分率由太阳黑子爆发前2-4 a的显著(p < 0.05)负相关, 转变为爆发后1-4 a的正相关; 臭冷杉叠加事件分析的结果与红松基本一致, 但未达到显著水平.太阳活动可通过影响局地气候间接影响树木的生长(徐海等, 2002 ; 尹红等, 2009 ).太阳活动剧烈时, 气温异常值随之增大或减小, 风速异常值也随之增大或减小, 太阳活动通过影响局地的气温和风速变化(表2 )间接影响该区树木生长, 具体表现为造成树木衰退或死亡, 抗干扰能力降低, 最终导致树体间竞争解除或减弱, 产生生长释放.太阳活动不仅直接影响着区域尺度下的气温、降水和风速变化, 还在很大程度上影响着区域性气候模态(赵亮等, 2011 ).因此, 太阳活动通过影响气温、降水和风速等局地气候以及其他大尺度气候模态干扰该地区森林动态, 是小兴安岭典型阔叶红松林可能存在的干扰机制. ...
利用树轮重建小兴安岭五营1796年以来的温度变化
2
2009
... 小兴安岭地区阔叶红松林内红松干扰事件主要发生在18世纪30年代、19世纪10年代、19世纪30-40年代、20世纪20-30年代, 臭冷杉主要发生在19世纪90年代、20世纪30年代、20世纪50-60年代、20世纪90年代.该结果与其他学者在长白山地区(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭地区(赵玉芳, 2011 )以及俄罗斯远东地区(Ishikawa et al ., 1999 )等阔叶红松林主要分布区的研究结果相一致, 说明阔叶红松林干扰事件不只是局限在研究区域内的小范围事件, 而且可能为区域尺度上(整个阔叶红松林分布区)的大范围干扰事件.红松和臭冷杉干扰事件可能受区域性气候模态周期影响(图4 ), 进一步说明阔叶红松林干扰事件是区域尺度上的大范围干扰事件.小兴安岭丰林自然保护区臭冷杉径向生长与AMO和太平洋年代际振荡(Pacific Decadal Oscillation, PDO)有直接关系, AMO、PDO的相位转变可能是谷地臭冷杉衰退的原因(姚启超和王晓春, 2013 ).长白山安图(徐海等, 2002 )和小兴安岭五营(尹红等, 2009 )红松树轮重建的降水或气温序列中也发现了与ENSO等区域性气候模态对应的周期, 也说明红松径向生长受区域性气候模态的影响.大尺度气候波动虽然不会直接影响某一区域树木的生长, 但能通过影响局地气候间接影响树木生长(张先亮等, 2010 ).当这种影响超过一定的阈值时, 势必会导致树木出现生长衰退或死亡.因此, 外力作用下更易形成林窗, 造成周围树体生长释放.另外, 区域气候模态(尤其是ENSO)与东亚季风关系密切(徐袁和钱维宏, 2003 ), 由此形成的风暴更是触发林窗形成的关键原因. ...
... 红松和臭冷杉干扰事件的周期性中, 存在11 a的极显著(p < 0.01)太阳活动周期, 而且红松生长变化百分率与太阳黑子异常值呈显著负相关(p < 0.05) (图10 ), 也表明太阳活动很大程度上影响着该区树木的生长释放.太阳黑子爆发年与红松、臭冷杉生长变化百分率的SEA分析(图11 )表明: 小兴安岭地区红松树轮记录的生长变化百分率由太阳黑子爆发前2-4 a的显著(p < 0.05)负相关, 转变为爆发后1-4 a的正相关; 臭冷杉叠加事件分析的结果与红松基本一致, 但未达到显著水平.太阳活动可通过影响局地气候间接影响树木的生长(徐海等, 2002 ; 尹红等, 2009 ).太阳活动剧烈时, 气温异常值随之增大或减小, 风速异常值也随之增大或减小, 太阳活动通过影响局地的气温和风速变化(表2 )间接影响该区树木生长, 具体表现为造成树木衰退或死亡, 抗干扰能力降低, 最终导致树体间竞争解除或减弱, 产生生长释放.太阳活动不仅直接影响着区域尺度下的气温、降水和风速变化, 还在很大程度上影响着区域性气候模态(赵亮等, 2011 ).因此, 太阳活动通过影响气温、降水和风速等局地气候以及其他大尺度气候模态干扰该地区森林动态, 是小兴安岭典型阔叶红松林可能存在的干扰机制. ...
林隙(GAP)干扰研究进展
2
1998
... 林窗干扰是自然界中最常见的一种森林干扰(臧润国和徐化成, 1998 ; 臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).林窗干扰发生后, 周围树木由于竞争去除而产生生长释放(王晓春和赵玉芳, 2011 ), 并且这种释放可以持续多年.通常将大于一定阈值且持续5 a或10 a以上的生长释放视为“真正释放”, 并将生长释放开始的时间视为干扰的发生时间(Black & Abrams, 2004 ; Rubino & McCarthyz, 2004 ; 王晓春和赵玉芳, 2011 ).确定生长释放的发生时间是重建森林干扰历史的最基本过程(Black & Abrams, 2004 ), 林窗形成时间常以林窗内幼苗更新时间(刘少冲等, 2013 )、倒木倾倒时间(用腐烂等级确定, 韩路等, 2011 )以及林窗周边树木生长释放时间(阳含熙和谢海生, 1994 ; 夏冰等, 1995 )等确定.本研究林窗干扰时间的确定主要用树木生长释放来判断, 同时还参考了林窗内的幼苗更新及倒木腐烂等级, 但有些林窗更新的非均一性以及林窗内倒木腐烂情况比较复杂也影响林窗形成时间的确定. ...
... 本研究发现风干扰是小兴安岭原始阔叶红松林树木生长释放的一个主要因素, 风干扰是该区阔叶红松林干扰的主要类型, 这与其他学者在长白山(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭(王晓春和赵玉芳, 2011 ; 赵玉芳, 2011 )及俄罗斯远东(Ishikawa et al ., 1999 )等地阔叶红松林干扰研究的结果一致.风干扰造成树体倾倒(尤其是林冠层乔木, 如红松)形成林窗(图9 ), 林窗的出现解除了周围树木生长的竞争, 林窗边缘木将出现明显的生长释放(王晓春和赵玉芳, 2011 ).例如, 研究区春季(4-5月)多风且风速较大, 春季最大风速平均值前10个最大年份分别为1971、1976、1987、1989-1990、1994-1996和1998-1999年, 这些年份当年或后几年红松生长变化百分率均出现明显增加的趋势.臭冷杉由于是喜阴树种且多处于林冠下层, 生长释放不太明显.此外, 林窗内形成木数量、腐烂等级以及林窗边缘木生长释放特征, 很好地证明了森林遭大风袭击后摧毁了部分林冠层树木, 造成林冠参差不齐.林窗形成后, 该区对风的抵抗力便会大大削弱, 林分很容易发生持续的风倒, 产生多林窗潮(multiple-gap episodes)(阳含熙和谢海生, 1994 ; 臧润国和徐化成, 1998 ). ...
林隙(GAP)干扰研究进展
2
1998
... 林窗干扰是自然界中最常见的一种森林干扰(臧润国和徐化成, 1998 ; 臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).林窗干扰发生后, 周围树木由于竞争去除而产生生长释放(王晓春和赵玉芳, 2011 ), 并且这种释放可以持续多年.通常将大于一定阈值且持续5 a或10 a以上的生长释放视为“真正释放”, 并将生长释放开始的时间视为干扰的发生时间(Black & Abrams, 2004 ; Rubino & McCarthyz, 2004 ; 王晓春和赵玉芳, 2011 ).确定生长释放的发生时间是重建森林干扰历史的最基本过程(Black & Abrams, 2004 ), 林窗形成时间常以林窗内幼苗更新时间(刘少冲等, 2013 )、倒木倾倒时间(用腐烂等级确定, 韩路等, 2011 )以及林窗周边树木生长释放时间(阳含熙和谢海生, 1994 ; 夏冰等, 1995 )等确定.本研究林窗干扰时间的确定主要用树木生长释放来判断, 同时还参考了林窗内的幼苗更新及倒木腐烂等级, 但有些林窗更新的非均一性以及林窗内倒木腐烂情况比较复杂也影响林窗形成时间的确定. ...
... 本研究发现风干扰是小兴安岭原始阔叶红松林树木生长释放的一个主要因素, 风干扰是该区阔叶红松林干扰的主要类型, 这与其他学者在长白山(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭(王晓春和赵玉芳, 2011 ; 赵玉芳, 2011 )及俄罗斯远东(Ishikawa et al ., 1999 )等地阔叶红松林干扰研究的结果一致.风干扰造成树体倾倒(尤其是林冠层乔木, 如红松)形成林窗(图9 ), 林窗的出现解除了周围树木生长的竞争, 林窗边缘木将出现明显的生长释放(王晓春和赵玉芳, 2011 ).例如, 研究区春季(4-5月)多风且风速较大, 春季最大风速平均值前10个最大年份分别为1971、1976、1987、1989-1990、1994-1996和1998-1999年, 这些年份当年或后几年红松生长变化百分率均出现明显增加的趋势.臭冷杉由于是喜阴树种且多处于林冠下层, 生长释放不太明显.此外, 林窗内形成木数量、腐烂等级以及林窗边缘木生长释放特征, 很好地证明了森林遭大风袭击后摧毁了部分林冠层树木, 造成林冠参差不齐.林窗形成后, 该区对风的抵抗力便会大大削弱, 林分很容易发生持续的风倒, 产生多林窗潮(multiple-gap episodes)(阳含熙和谢海生, 1994 ; 臧润国和徐化成, 1998 ). ...
海南岛热带山地雨林林隙及其自然干扰特征
3
1999
... 随着频繁的人类活动和异常自然干扰的发生, 森林生态系统结构与功能正受到严重的威胁(Vitousek et al ., 1997 ).干扰生态学已成为生态学研究最活跃的领域之一(朱教君和刘世荣, 2007 ; Bond-Lamberty et al ., 2014 ), 其重点是从森林群落的干扰历史和林窗(隙)动态两个方面对中小规模干扰的特征及影响开展研究, 且有关林窗动态的研究相对较多(臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).重建干扰历史是植物群落演替和更新研究的基础, 越来越受到重视.森林群落的干扰动态可以通过固定样地的长期观测获得(Cooke & Lorenzetti, 2006 ), 但对于缺乏长期观测的森林群落, 人们无法得知其干扰历史.树木年轮是树木生长的历史记录(Smith, 2008 ), 其宽度、密度、同位素含量以及解剖结构等记载了树木逐年的生长信息及内、外部环境变化的影响(Cook & Kairiuktis, 1990 ).树轮具有生长时间长、定年准确、材料广泛、容易获取等优点, 是研究树木生长环境变化的重要手段, 也是重建森林群落动态的重要材料. ...
... 2013年6月, 在对9 hm2 固定样地大面积踏查的基础上, 选择比较典型的300 m × 200 m地块作为实验样地.采用样方调查法(臧润国等, 1999 ), 以林内东北点作为调查起点, 沿罗盘仪所指方向, 由北向南穿行, 调查每一个林窗的基本特征(本文定义树冠空隙面积10 m2 以上的林冠空隙为林窗).对样地内每一株林窗边缘木(gap tree)(距林窗边缘3 m以内立木)进行生长锥取样.取样位置在树高约1.2 m处, 取样方向均朝向林窗中心点处, 所取样芯尽量达到髓心.在符合要求的44个林窗中共获得606个边缘木样芯, 其中红松461个(平均年龄约160年), 臭冷杉145个(平均年龄约80年).因红松与臭冷杉均存在腐心现象, 部分样芯未取到髓心, 因此实际树龄比样芯年龄更长.此外, 还在固定样地外随机钻取非林窗木(non-gap tree)样芯(每个树种≥40个样芯)作为对照.将钻取的树芯装入孔径相当的塑料管并进行编号, 同时记录每一个样本的特征及生境条件. ...
... 林窗干扰是自然界中最常见的一种森林干扰(臧润国和徐化成, 1998 ; 臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).林窗干扰发生后, 周围树木由于竞争去除而产生生长释放(王晓春和赵玉芳, 2011 ), 并且这种释放可以持续多年.通常将大于一定阈值且持续5 a或10 a以上的生长释放视为“真正释放”, 并将生长释放开始的时间视为干扰的发生时间(Black & Abrams, 2004 ; Rubino & McCarthyz, 2004 ; 王晓春和赵玉芳, 2011 ).确定生长释放的发生时间是重建森林干扰历史的最基本过程(Black & Abrams, 2004 ), 林窗形成时间常以林窗内幼苗更新时间(刘少冲等, 2013 )、倒木倾倒时间(用腐烂等级确定, 韩路等, 2011 )以及林窗周边树木生长释放时间(阳含熙和谢海生, 1994 ; 夏冰等, 1995 )等确定.本研究林窗干扰时间的确定主要用树木生长释放来判断, 同时还参考了林窗内的幼苗更新及倒木腐烂等级, 但有些林窗更新的非均一性以及林窗内倒木腐烂情况比较复杂也影响林窗形成时间的确定. ...
海南岛热带山地雨林林隙及其自然干扰特征
3
1999
... 随着频繁的人类活动和异常自然干扰的发生, 森林生态系统结构与功能正受到严重的威胁(Vitousek et al ., 1997 ).干扰生态学已成为生态学研究最活跃的领域之一(朱教君和刘世荣, 2007 ; Bond-Lamberty et al ., 2014 ), 其重点是从森林群落的干扰历史和林窗(隙)动态两个方面对中小规模干扰的特征及影响开展研究, 且有关林窗动态的研究相对较多(臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).重建干扰历史是植物群落演替和更新研究的基础, 越来越受到重视.森林群落的干扰动态可以通过固定样地的长期观测获得(Cooke & Lorenzetti, 2006 ), 但对于缺乏长期观测的森林群落, 人们无法得知其干扰历史.树木年轮是树木生长的历史记录(Smith, 2008 ), 其宽度、密度、同位素含量以及解剖结构等记载了树木逐年的生长信息及内、外部环境变化的影响(Cook & Kairiuktis, 1990 ).树轮具有生长时间长、定年准确、材料广泛、容易获取等优点, 是研究树木生长环境变化的重要手段, 也是重建森林群落动态的重要材料. ...
... 2013年6月, 在对9 hm2 固定样地大面积踏查的基础上, 选择比较典型的300 m × 200 m地块作为实验样地.采用样方调查法(臧润国等, 1999 ), 以林内东北点作为调查起点, 沿罗盘仪所指方向, 由北向南穿行, 调查每一个林窗的基本特征(本文定义树冠空隙面积10 m2 以上的林冠空隙为林窗).对样地内每一株林窗边缘木(gap tree)(距林窗边缘3 m以内立木)进行生长锥取样.取样位置在树高约1.2 m处, 取样方向均朝向林窗中心点处, 所取样芯尽量达到髓心.在符合要求的44个林窗中共获得606个边缘木样芯, 其中红松461个(平均年龄约160年), 臭冷杉145个(平均年龄约80年).因红松与臭冷杉均存在腐心现象, 部分样芯未取到髓心, 因此实际树龄比样芯年龄更长.此外, 还在固定样地外随机钻取非林窗木(non-gap tree)样芯(每个树种≥40个样芯)作为对照.将钻取的树芯装入孔径相当的塑料管并进行编号, 同时记录每一个样本的特征及生境条件. ...
... 林窗干扰是自然界中最常见的一种森林干扰(臧润国和徐化成, 1998 ; 臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).林窗干扰发生后, 周围树木由于竞争去除而产生生长释放(王晓春和赵玉芳, 2011 ), 并且这种释放可以持续多年.通常将大于一定阈值且持续5 a或10 a以上的生长释放视为“真正释放”, 并将生长释放开始的时间视为干扰的发生时间(Black & Abrams, 2004 ; Rubino & McCarthyz, 2004 ; 王晓春和赵玉芳, 2011 ).确定生长释放的发生时间是重建森林干扰历史的最基本过程(Black & Abrams, 2004 ), 林窗形成时间常以林窗内幼苗更新时间(刘少冲等, 2013 )、倒木倾倒时间(用腐烂等级确定, 韩路等, 2011 )以及林窗周边树木生长释放时间(阳含熙和谢海生, 1994 ; 夏冰等, 1995 )等确定.本研究林窗干扰时间的确定主要用树木生长释放来判断, 同时还参考了林窗内的幼苗更新及倒木腐烂等级, 但有些林窗更新的非均一性以及林窗内倒木腐烂情况比较复杂也影响林窗形成时间的确定. ...
基于树轮火疤重建大兴安岭盘古兴安落叶松林近200年火灾历史
1
2013
... 国外已有一些研究利用树木年轮重建森林干扰历史(Nowacki & Abrams, 1997 ; Ishikawa et al ., 1999 ; Rubino & McCarthyz, 2004 ; Brown & Giesecke, 2014 ), 而国内这方面研究起步较晚, 且相对较少.20世纪90年代初, 阳含熙和谢海生(1994)最先利用树木生长释放和抑制对长白山红松(Pinus koraiensis )混交林干扰历史进行了重建, 并指出该方法是以风作为主要干扰类型, 重建干扰历史最准确、最可靠、提供信息量最大的方法; 侯向阳等(2000)研究长白山红松林干扰节律时, 也得到了类似的结论.乔佃锋(2005)利用该方法(阳含熙和谢海生, 1994 )重建了长白山敦化地区阔叶红松林的干扰历史, 发现该区红松林干扰以小规模的风干扰为主, 1980年出现了明显的干扰峰, 干扰峰之间60年的时间间隔正好与红松的更替世代相吻合.王晓春和赵玉芳(2011)发现前一年径向生长对生长释放事件的判定很重要, 小兴安岭北端黑河地区的红松和红皮云杉(Picea koraiensis )的生长释放可能受林窗产生、微环境变化以及温度和降水等气候因素影响.夏冰等(1995)在天目山建立了生长释放和抑制的判定标准, 并重建了黄山松(Pinus taiwanensis )阔叶林树冠的干扰历史; 同时, 还对不同树种年轮宽度的生长释放判别方法进行了研究, 并做了重建干扰强度的尝试(夏冰等, 1997 , 2001 ).此外, 不同学者还利用树轮生态学方法对火灾(张磊和王晓春, 2013 )、泥石流(铁永波等, 2014 )、山体滑坡(洪婷等, 2012 )和冰川活动(Zhu et al ., 2013 )等干扰进行了重建. ...
基于树轮火疤重建大兴安岭盘古兴安落叶松林近200年火灾历史
1
2013
... 国外已有一些研究利用树木年轮重建森林干扰历史(Nowacki & Abrams, 1997 ; Ishikawa et al ., 1999 ; Rubino & McCarthyz, 2004 ; Brown & Giesecke, 2014 ), 而国内这方面研究起步较晚, 且相对较少.20世纪90年代初, 阳含熙和谢海生(1994)最先利用树木生长释放和抑制对长白山红松(Pinus koraiensis )混交林干扰历史进行了重建, 并指出该方法是以风作为主要干扰类型, 重建干扰历史最准确、最可靠、提供信息量最大的方法; 侯向阳等(2000)研究长白山红松林干扰节律时, 也得到了类似的结论.乔佃锋(2005)利用该方法(阳含熙和谢海生, 1994 )重建了长白山敦化地区阔叶红松林的干扰历史, 发现该区红松林干扰以小规模的风干扰为主, 1980年出现了明显的干扰峰, 干扰峰之间60年的时间间隔正好与红松的更替世代相吻合.王晓春和赵玉芳(2011)发现前一年径向生长对生长释放事件的判定很重要, 小兴安岭北端黑河地区的红松和红皮云杉(Picea koraiensis )的生长释放可能受林窗产生、微环境变化以及温度和降水等气候因素影响.夏冰等(1995)在天目山建立了生长释放和抑制的判定标准, 并重建了黄山松(Pinus taiwanensis )阔叶林树冠的干扰历史; 同时, 还对不同树种年轮宽度的生长释放判别方法进行了研究, 并做了重建干扰强度的尝试(夏冰等, 1997 , 2001 ).此外, 不同学者还利用树轮生态学方法对火灾(张磊和王晓春, 2013 )、泥石流(铁永波等, 2014 )、山体滑坡(洪婷等, 2012 )和冰川活动(Zhu et al ., 2013 )等干扰进行了重建. ...
大兴安岭库都尔地区兴安落叶松年轮宽度年表及其与气候变化的关系
1
2010
... 小兴安岭地区阔叶红松林内红松干扰事件主要发生在18世纪30年代、19世纪10年代、19世纪30-40年代、20世纪20-30年代, 臭冷杉主要发生在19世纪90年代、20世纪30年代、20世纪50-60年代、20世纪90年代.该结果与其他学者在长白山地区(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭地区(赵玉芳, 2011 )以及俄罗斯远东地区(Ishikawa et al ., 1999 )等阔叶红松林主要分布区的研究结果相一致, 说明阔叶红松林干扰事件不只是局限在研究区域内的小范围事件, 而且可能为区域尺度上(整个阔叶红松林分布区)的大范围干扰事件.红松和臭冷杉干扰事件可能受区域性气候模态周期影响(图4 ), 进一步说明阔叶红松林干扰事件是区域尺度上的大范围干扰事件.小兴安岭丰林自然保护区臭冷杉径向生长与AMO和太平洋年代际振荡(Pacific Decadal Oscillation, PDO)有直接关系, AMO、PDO的相位转变可能是谷地臭冷杉衰退的原因(姚启超和王晓春, 2013 ).长白山安图(徐海等, 2002 )和小兴安岭五营(尹红等, 2009 )红松树轮重建的降水或气温序列中也发现了与ENSO等区域性气候模态对应的周期, 也说明红松径向生长受区域性气候模态的影响.大尺度气候波动虽然不会直接影响某一区域树木的生长, 但能通过影响局地气候间接影响树木生长(张先亮等, 2010 ).当这种影响超过一定的阈值时, 势必会导致树木出现生长衰退或死亡.因此, 外力作用下更易形成林窗, 造成周围树体生长释放.另外, 区域气候模态(尤其是ENSO)与东亚季风关系密切(徐袁和钱维宏, 2003 ), 由此形成的风暴更是触发林窗形成的关键原因. ...
大兴安岭库都尔地区兴安落叶松年轮宽度年表及其与气候变化的关系
1
2010
... 小兴安岭地区阔叶红松林内红松干扰事件主要发生在18世纪30年代、19世纪10年代、19世纪30-40年代、20世纪20-30年代, 臭冷杉主要发生在19世纪90年代、20世纪30年代、20世纪50-60年代、20世纪90年代.该结果与其他学者在长白山地区(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭地区(赵玉芳, 2011 )以及俄罗斯远东地区(Ishikawa et al ., 1999 )等阔叶红松林主要分布区的研究结果相一致, 说明阔叶红松林干扰事件不只是局限在研究区域内的小范围事件, 而且可能为区域尺度上(整个阔叶红松林分布区)的大范围干扰事件.红松和臭冷杉干扰事件可能受区域性气候模态周期影响(图4 ), 进一步说明阔叶红松林干扰事件是区域尺度上的大范围干扰事件.小兴安岭丰林自然保护区臭冷杉径向生长与AMO和太平洋年代际振荡(Pacific Decadal Oscillation, PDO)有直接关系, AMO、PDO的相位转变可能是谷地臭冷杉衰退的原因(姚启超和王晓春, 2013 ).长白山安图(徐海等, 2002 )和小兴安岭五营(尹红等, 2009 )红松树轮重建的降水或气温序列中也发现了与ENSO等区域性气候模态对应的周期, 也说明红松径向生长受区域性气候模态的影响.大尺度气候波动虽然不会直接影响某一区域树木的生长, 但能通过影响局地气候间接影响树木生长(张先亮等, 2010 ).当这种影响超过一定的阈值时, 势必会导致树木出现生长衰退或死亡.因此, 外力作用下更易形成林窗, 造成周围树体生长释放.另外, 区域气候模态(尤其是ENSO)与东亚季风关系密切(徐袁和钱维宏, 2003 ), 由此形成的风暴更是触发林窗形成的关键原因. ...
Disturbance and regeneration dynamics of a mixed Korean pine dominated forest on Changbai Mountain, North- Eastern China.
3
2014
... 小兴安岭地区阔叶红松林内红松干扰事件主要发生在18世纪30年代、19世纪10年代、19世纪30-40年代、20世纪20-30年代, 臭冷杉主要发生在19世纪90年代、20世纪30年代、20世纪50-60年代、20世纪90年代.该结果与其他学者在长白山地区(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭地区(赵玉芳, 2011 )以及俄罗斯远东地区(Ishikawa et al ., 1999 )等阔叶红松林主要分布区的研究结果相一致, 说明阔叶红松林干扰事件不只是局限在研究区域内的小范围事件, 而且可能为区域尺度上(整个阔叶红松林分布区)的大范围干扰事件.红松和臭冷杉干扰事件可能受区域性气候模态周期影响(图4 ), 进一步说明阔叶红松林干扰事件是区域尺度上的大范围干扰事件.小兴安岭丰林自然保护区臭冷杉径向生长与AMO和太平洋年代际振荡(Pacific Decadal Oscillation, PDO)有直接关系, AMO、PDO的相位转变可能是谷地臭冷杉衰退的原因(姚启超和王晓春, 2013 ).长白山安图(徐海等, 2002 )和小兴安岭五营(尹红等, 2009 )红松树轮重建的降水或气温序列中也发现了与ENSO等区域性气候模态对应的周期, 也说明红松径向生长受区域性气候模态的影响.大尺度气候波动虽然不会直接影响某一区域树木的生长, 但能通过影响局地气候间接影响树木生长(张先亮等, 2010 ).当这种影响超过一定的阈值时, 势必会导致树木出现生长衰退或死亡.因此, 外力作用下更易形成林窗, 造成周围树体生长释放.另外, 区域气候模态(尤其是ENSO)与东亚季风关系密切(徐袁和钱维宏, 2003 ), 由此形成的风暴更是触发林窗形成的关键原因. ...
... 本研究发现风干扰是小兴安岭原始阔叶红松林树木生长释放的一个主要因素, 风干扰是该区阔叶红松林干扰的主要类型, 这与其他学者在长白山(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭(王晓春和赵玉芳, 2011 ; 赵玉芳, 2011 )及俄罗斯远东(Ishikawa et al ., 1999 )等地阔叶红松林干扰研究的结果一致.风干扰造成树体倾倒(尤其是林冠层乔木, 如红松)形成林窗(图9 ), 林窗的出现解除了周围树木生长的竞争, 林窗边缘木将出现明显的生长释放(王晓春和赵玉芳, 2011 ).例如, 研究区春季(4-5月)多风且风速较大, 春季最大风速平均值前10个最大年份分别为1971、1976、1987、1989-1990、1994-1996和1998-1999年, 这些年份当年或后几年红松生长变化百分率均出现明显增加的趋势.臭冷杉由于是喜阴树种且多处于林冠下层, 生长释放不太明显.此外, 林窗内形成木数量、腐烂等级以及林窗边缘木生长释放特征, 很好地证明了森林遭大风袭击后摧毁了部分林冠层树木, 造成林冠参差不齐.林窗形成后, 该区对风的抵抗力便会大大削弱, 林分很容易发生持续的风倒, 产生多林窗潮(multiple-gap episodes)(阳含熙和谢海生, 1994 ; 臧润国和徐化成, 1998 ). ...
... 风干扰是阔叶红松林干扰的主要因素之一(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 ), 然而并不意味着风速越大, 形成林窗的数量就越多, 生长释放就越明显.风只是林窗干扰发生的一个外因, 林窗干扰是否发生以及干扰的强弱还取决于树体自身的生长情况及其生存的环境条件.此外, 异常气温也在很大程度上影响着该区树木的生长释放, 这一结果与王晓春和赵玉芳(2011)的研究结果一致.异常气温影响研究区的干旱化现象(贺伟等, 2013 ), 导致该区树木生长衰退或死亡, 对外界干扰的抵抗能力降低或丧失, 在外力作用下更易形成倒木, 进而造成林窗边缘树木发生释放. ...
太阳活动对近百年气候变化的影响研究进展
1
2011
... 红松和臭冷杉干扰事件的周期性中, 存在11 a的极显著(p < 0.01)太阳活动周期, 而且红松生长变化百分率与太阳黑子异常值呈显著负相关(p < 0.05) (图10 ), 也表明太阳活动很大程度上影响着该区树木的生长释放.太阳黑子爆发年与红松、臭冷杉生长变化百分率的SEA分析(图11 )表明: 小兴安岭地区红松树轮记录的生长变化百分率由太阳黑子爆发前2-4 a的显著(p < 0.05)负相关, 转变为爆发后1-4 a的正相关; 臭冷杉叠加事件分析的结果与红松基本一致, 但未达到显著水平.太阳活动可通过影响局地气候间接影响树木的生长(徐海等, 2002 ; 尹红等, 2009 ).太阳活动剧烈时, 气温异常值随之增大或减小, 风速异常值也随之增大或减小, 太阳活动通过影响局地的气温和风速变化(表2 )间接影响该区树木生长, 具体表现为造成树木衰退或死亡, 抗干扰能力降低, 最终导致树体间竞争解除或减弱, 产生生长释放.太阳活动不仅直接影响着区域尺度下的气温、降水和风速变化, 还在很大程度上影响着区域性气候模态(赵亮等, 2011 ).因此, 太阳活动通过影响气温、降水和风速等局地气候以及其他大尺度气候模态干扰该地区森林动态, 是小兴安岭典型阔叶红松林可能存在的干扰机制. ...
太阳活动对近百年气候变化的影响研究进展
1
2011
... 红松和臭冷杉干扰事件的周期性中, 存在11 a的极显著(p < 0.01)太阳活动周期, 而且红松生长变化百分率与太阳黑子异常值呈显著负相关(p < 0.05) (图10 ), 也表明太阳活动很大程度上影响着该区树木的生长释放.太阳黑子爆发年与红松、臭冷杉生长变化百分率的SEA分析(图11 )表明: 小兴安岭地区红松树轮记录的生长变化百分率由太阳黑子爆发前2-4 a的显著(p < 0.05)负相关, 转变为爆发后1-4 a的正相关; 臭冷杉叠加事件分析的结果与红松基本一致, 但未达到显著水平.太阳活动可通过影响局地气候间接影响树木的生长(徐海等, 2002 ; 尹红等, 2009 ).太阳活动剧烈时, 气温异常值随之增大或减小, 风速异常值也随之增大或减小, 太阳活动通过影响局地的气温和风速变化(表2 )间接影响该区树木生长, 具体表现为造成树木衰退或死亡, 抗干扰能力降低, 最终导致树体间竞争解除或减弱, 产生生长释放.太阳活动不仅直接影响着区域尺度下的气温、降水和风速变化, 还在很大程度上影响着区域性气候模态(赵亮等, 2011 ).因此, 太阳活动通过影响气温、降水和风速等局地气候以及其他大尺度气候模态干扰该地区森林动态, 是小兴安岭典型阔叶红松林可能存在的干扰机制. ...
黑龙江省红松生长释放判定及解释
2
2011
... 小兴安岭地区阔叶红松林内红松干扰事件主要发生在18世纪30年代、19世纪10年代、19世纪30-40年代、20世纪20-30年代, 臭冷杉主要发生在19世纪90年代、20世纪30年代、20世纪50-60年代、20世纪90年代.该结果与其他学者在长白山地区(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭地区(赵玉芳, 2011 )以及俄罗斯远东地区(Ishikawa et al ., 1999 )等阔叶红松林主要分布区的研究结果相一致, 说明阔叶红松林干扰事件不只是局限在研究区域内的小范围事件, 而且可能为区域尺度上(整个阔叶红松林分布区)的大范围干扰事件.红松和臭冷杉干扰事件可能受区域性气候模态周期影响(图4 ), 进一步说明阔叶红松林干扰事件是区域尺度上的大范围干扰事件.小兴安岭丰林自然保护区臭冷杉径向生长与AMO和太平洋年代际振荡(Pacific Decadal Oscillation, PDO)有直接关系, AMO、PDO的相位转变可能是谷地臭冷杉衰退的原因(姚启超和王晓春, 2013 ).长白山安图(徐海等, 2002 )和小兴安岭五营(尹红等, 2009 )红松树轮重建的降水或气温序列中也发现了与ENSO等区域性气候模态对应的周期, 也说明红松径向生长受区域性气候模态的影响.大尺度气候波动虽然不会直接影响某一区域树木的生长, 但能通过影响局地气候间接影响树木生长(张先亮等, 2010 ).当这种影响超过一定的阈值时, 势必会导致树木出现生长衰退或死亡.因此, 外力作用下更易形成林窗, 造成周围树体生长释放.另外, 区域气候模态(尤其是ENSO)与东亚季风关系密切(徐袁和钱维宏, 2003 ), 由此形成的风暴更是触发林窗形成的关键原因. ...
... 本研究发现风干扰是小兴安岭原始阔叶红松林树木生长释放的一个主要因素, 风干扰是该区阔叶红松林干扰的主要类型, 这与其他学者在长白山(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭(王晓春和赵玉芳, 2011 ; 赵玉芳, 2011 )及俄罗斯远东(Ishikawa et al ., 1999 )等地阔叶红松林干扰研究的结果一致.风干扰造成树体倾倒(尤其是林冠层乔木, 如红松)形成林窗(图9 ), 林窗的出现解除了周围树木生长的竞争, 林窗边缘木将出现明显的生长释放(王晓春和赵玉芳, 2011 ).例如, 研究区春季(4-5月)多风且风速较大, 春季最大风速平均值前10个最大年份分别为1971、1976、1987、1989-1990、1994-1996和1998-1999年, 这些年份当年或后几年红松生长变化百分率均出现明显增加的趋势.臭冷杉由于是喜阴树种且多处于林冠下层, 生长释放不太明显.此外, 林窗内形成木数量、腐烂等级以及林窗边缘木生长释放特征, 很好地证明了森林遭大风袭击后摧毁了部分林冠层树木, 造成林冠参差不齐.林窗形成后, 该区对风的抵抗力便会大大削弱, 林分很容易发生持续的风倒, 产生多林窗潮(multiple-gap episodes)(阳含熙和谢海生, 1994 ; 臧润国和徐化成, 1998 ). ...
黑龙江省红松生长释放判定及解释
2
2011
... 小兴安岭地区阔叶红松林内红松干扰事件主要发生在18世纪30年代、19世纪10年代、19世纪30-40年代、20世纪20-30年代, 臭冷杉主要发生在19世纪90年代、20世纪30年代、20世纪50-60年代、20世纪90年代.该结果与其他学者在长白山地区(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭地区(赵玉芳, 2011 )以及俄罗斯远东地区(Ishikawa et al ., 1999 )等阔叶红松林主要分布区的研究结果相一致, 说明阔叶红松林干扰事件不只是局限在研究区域内的小范围事件, 而且可能为区域尺度上(整个阔叶红松林分布区)的大范围干扰事件.红松和臭冷杉干扰事件可能受区域性气候模态周期影响(图4 ), 进一步说明阔叶红松林干扰事件是区域尺度上的大范围干扰事件.小兴安岭丰林自然保护区臭冷杉径向生长与AMO和太平洋年代际振荡(Pacific Decadal Oscillation, PDO)有直接关系, AMO、PDO的相位转变可能是谷地臭冷杉衰退的原因(姚启超和王晓春, 2013 ).长白山安图(徐海等, 2002 )和小兴安岭五营(尹红等, 2009 )红松树轮重建的降水或气温序列中也发现了与ENSO等区域性气候模态对应的周期, 也说明红松径向生长受区域性气候模态的影响.大尺度气候波动虽然不会直接影响某一区域树木的生长, 但能通过影响局地气候间接影响树木生长(张先亮等, 2010 ).当这种影响超过一定的阈值时, 势必会导致树木出现生长衰退或死亡.因此, 外力作用下更易形成林窗, 造成周围树体生长释放.另外, 区域气候模态(尤其是ENSO)与东亚季风关系密切(徐袁和钱维宏, 2003 ), 由此形成的风暴更是触发林窗形成的关键原因. ...
... 本研究发现风干扰是小兴安岭原始阔叶红松林树木生长释放的一个主要因素, 风干扰是该区阔叶红松林干扰的主要类型, 这与其他学者在长白山(阳含熙和谢海生, 1994 ; 侯向阳等, 2000 ; 乔佃锋, 2005 ; Zhang et al ., 2014 )、小兴安岭(王晓春和赵玉芳, 2011 ; 赵玉芳, 2011 )及俄罗斯远东(Ishikawa et al ., 1999 )等地阔叶红松林干扰研究的结果一致.风干扰造成树体倾倒(尤其是林冠层乔木, 如红松)形成林窗(图9 ), 林窗的出现解除了周围树木生长的竞争, 林窗边缘木将出现明显的生长释放(王晓春和赵玉芳, 2011 ).例如, 研究区春季(4-5月)多风且风速较大, 春季最大风速平均值前10个最大年份分别为1971、1976、1987、1989-1990、1994-1996和1998-1999年, 这些年份当年或后几年红松生长变化百分率均出现明显增加的趋势.臭冷杉由于是喜阴树种且多处于林冠下层, 生长释放不太明显.此外, 林窗内形成木数量、腐烂等级以及林窗边缘木生长释放特征, 很好地证明了森林遭大风袭击后摧毁了部分林冠层树木, 造成林冠参差不齐.林窗形成后, 该区对风的抵抗力便会大大削弱, 林分很容易发生持续的风倒, 产生多林窗潮(multiple-gap episodes)(阳含熙和谢海生, 1994 ; 臧润国和徐化成, 1998 ). ...
Little Ice Age glacier fluctuations reconstructed for the southeastern Tibetan Plateau using tree rings.
1
2013
... 国外已有一些研究利用树木年轮重建森林干扰历史(Nowacki & Abrams, 1997 ; Ishikawa et al ., 1999 ; Rubino & McCarthyz, 2004 ; Brown & Giesecke, 2014 ), 而国内这方面研究起步较晚, 且相对较少.20世纪90年代初, 阳含熙和谢海生(1994)最先利用树木生长释放和抑制对长白山红松(Pinus koraiensis )混交林干扰历史进行了重建, 并指出该方法是以风作为主要干扰类型, 重建干扰历史最准确、最可靠、提供信息量最大的方法; 侯向阳等(2000)研究长白山红松林干扰节律时, 也得到了类似的结论.乔佃锋(2005)利用该方法(阳含熙和谢海生, 1994 )重建了长白山敦化地区阔叶红松林的干扰历史, 发现该区红松林干扰以小规模的风干扰为主, 1980年出现了明显的干扰峰, 干扰峰之间60年的时间间隔正好与红松的更替世代相吻合.王晓春和赵玉芳(2011)发现前一年径向生长对生长释放事件的判定很重要, 小兴安岭北端黑河地区的红松和红皮云杉(Picea koraiensis )的生长释放可能受林窗产生、微环境变化以及温度和降水等气候因素影响.夏冰等(1995)在天目山建立了生长释放和抑制的判定标准, 并重建了黄山松(Pinus taiwanensis )阔叶林树冠的干扰历史; 同时, 还对不同树种年轮宽度的生长释放判别方法进行了研究, 并做了重建干扰强度的尝试(夏冰等, 1997 , 2001 ).此外, 不同学者还利用树轮生态学方法对火灾(张磊和王晓春, 2013 )、泥石流(铁永波等, 2014 )、山体滑坡(洪婷等, 2012 )和冰川活动(Zhu et al ., 2013 )等干扰进行了重建. ...
1
2007
... 随着频繁的人类活动和异常自然干扰的发生, 森林生态系统结构与功能正受到严重的威胁(Vitousek et al ., 1997 ).干扰生态学已成为生态学研究最活跃的领域之一(朱教君和刘世荣, 2007 ; Bond-Lamberty et al ., 2014 ), 其重点是从森林群落的干扰历史和林窗(隙)动态两个方面对中小规模干扰的特征及影响开展研究, 且有关林窗动态的研究相对较多(臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).重建干扰历史是植物群落演替和更新研究的基础, 越来越受到重视.森林群落的干扰动态可以通过固定样地的长期观测获得(Cooke & Lorenzetti, 2006 ), 但对于缺乏长期观测的森林群落, 人们无法得知其干扰历史.树木年轮是树木生长的历史记录(Smith, 2008 ), 其宽度、密度、同位素含量以及解剖结构等记载了树木逐年的生长信息及内、外部环境变化的影响(Cook & Kairiuktis, 1990 ).树轮具有生长时间长、定年准确、材料广泛、容易获取等优点, 是研究树木生长环境变化的重要手段, 也是重建森林群落动态的重要材料. ...
1
2007
... 随着频繁的人类活动和异常自然干扰的发生, 森林生态系统结构与功能正受到严重的威胁(Vitousek et al ., 1997 ).干扰生态学已成为生态学研究最活跃的领域之一(朱教君和刘世荣, 2007 ; Bond-Lamberty et al ., 2014 ), 其重点是从森林群落的干扰历史和林窗(隙)动态两个方面对中小规模干扰的特征及影响开展研究, 且有关林窗动态的研究相对较多(臧润国等, 1999 ; 韩路等, 2011 ; 刘少冲等, 2013 ).重建干扰历史是植物群落演替和更新研究的基础, 越来越受到重视.森林群落的干扰动态可以通过固定样地的长期观测获得(Cooke & Lorenzetti, 2006 ), 但对于缺乏长期观测的森林群落, 人们无法得知其干扰历史.树木年轮是树木生长的历史记录(Smith, 2008 ), 其宽度、密度、同位素含量以及解剖结构等记载了树木逐年的生长信息及内、外部环境变化的影响(Cook & Kairiuktis, 1990 ).树轮具有生长时间长、定年准确、材料广泛、容易获取等优点, 是研究树木生长环境变化的重要手段, 也是重建森林群落动态的重要材料. ...



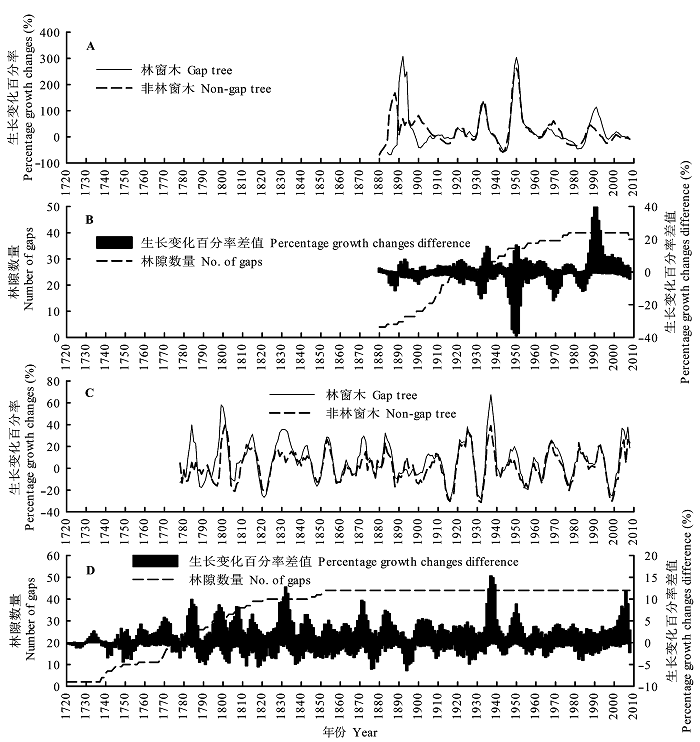
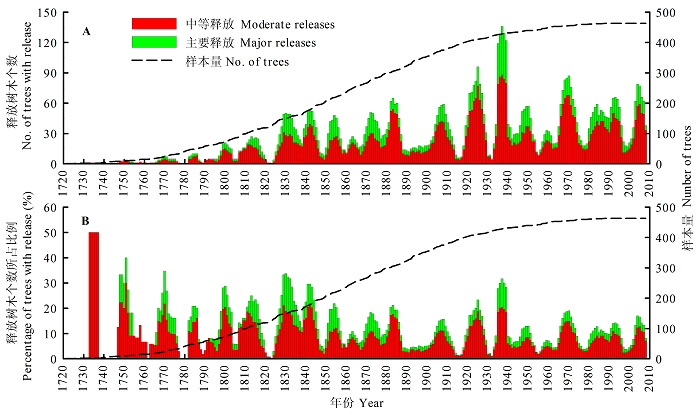
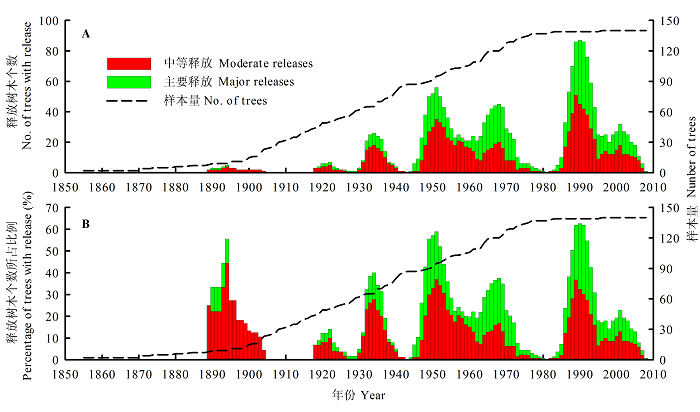
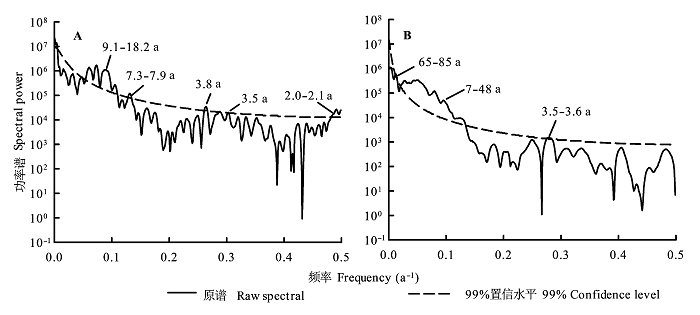
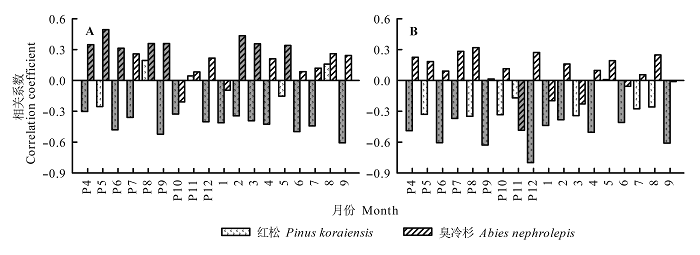
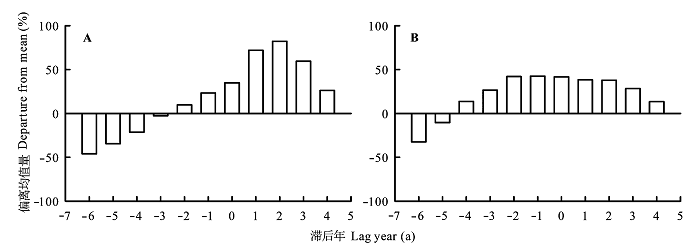
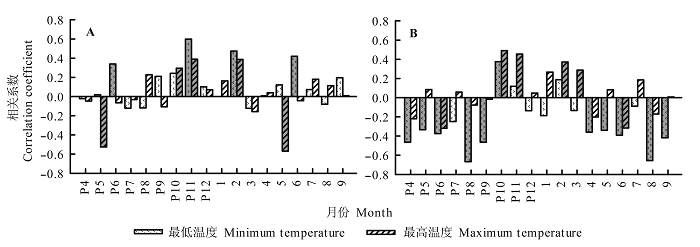
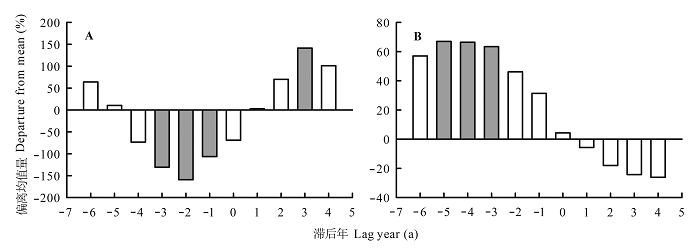

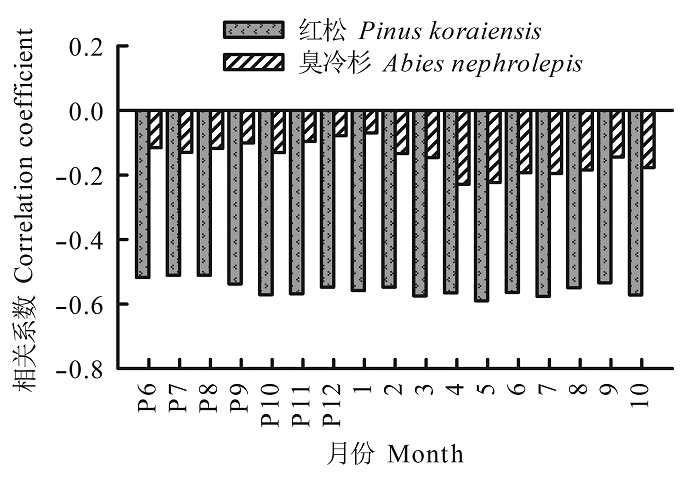
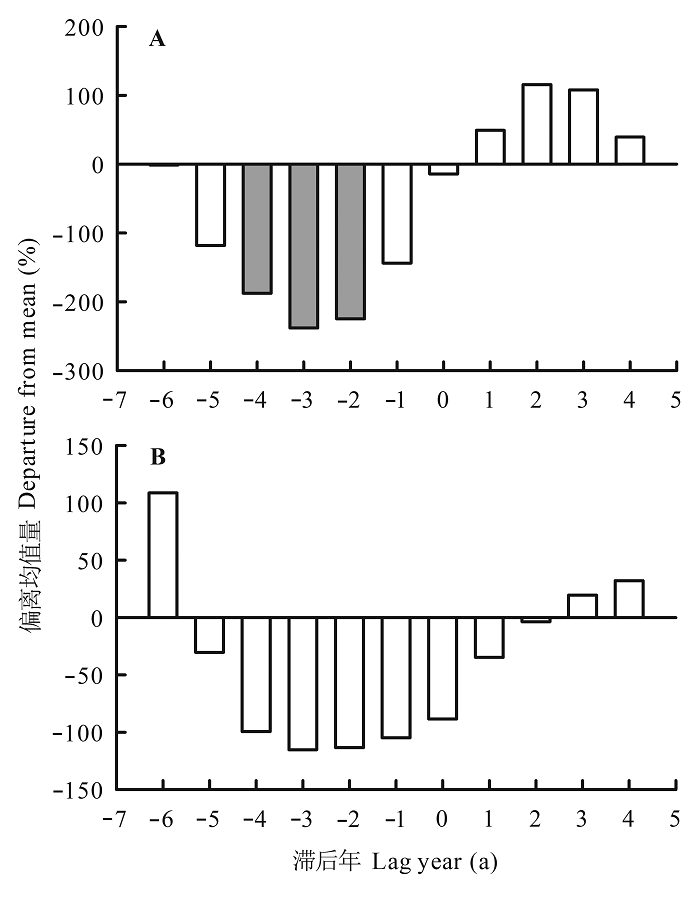
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}