Woody-tissue respiration (Rw) is well known to be a large component of the terrestrial ecosystem carbon balance. In order to quantify the intra- and inter-specific variations in stem respiration, investigations were made on the temporal and vertical variations in Rw in order to reveal the vertical pattern and the regulatory mechanisms of the temperature-sensitivity coefficient (Q10).
Methods
CO2 release rates and stem temperature were measured in two typical deciduous species (Sophora japonica and Salix matsudana) from March through December 2013 in a suburban park in Beijing. All measurements were carried out at three heights (10 cm, 140 cm and 270 cm) corresponding to the base, breast height and first branch of the trees.
Important findings
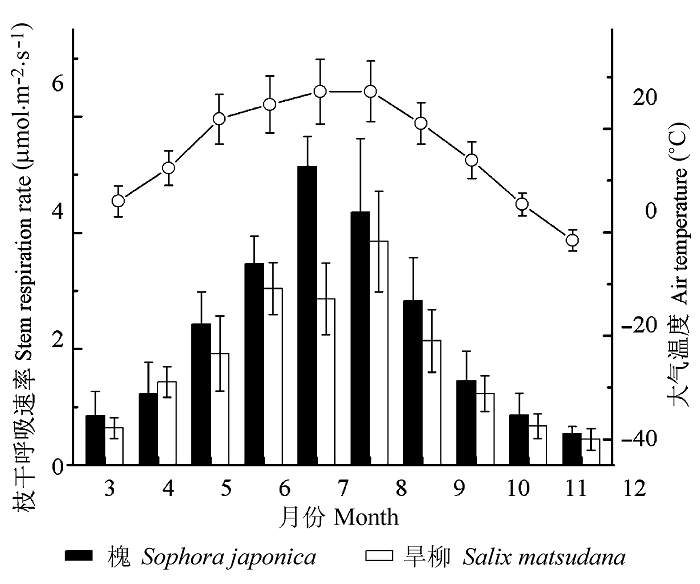
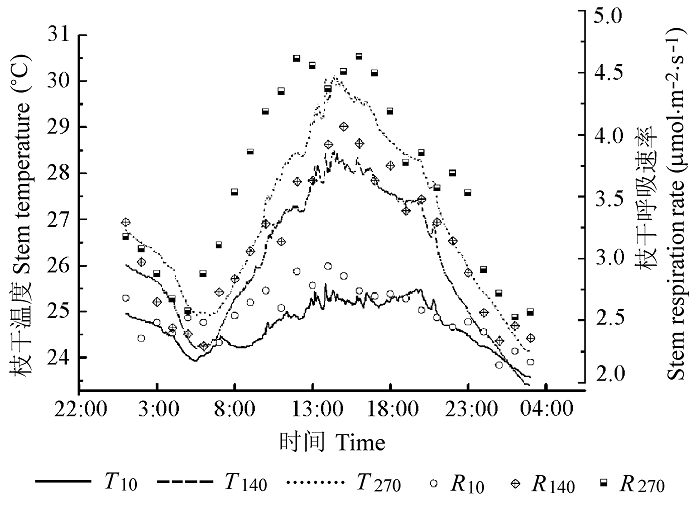
It was found that the stem respiration differed significantly between the two tree species. The Rw in Sophora japonica was 1.12 (July) to 1.79 (May) times of that in Salix matsudana for the same months except in April. Clear diurnal cycles and strong seasonal variations were found in the stem respiration per unit surface area (RS). The seasonal variation patterns of Rw were unimodal for both species; however, the peak month differed between the two species, i.e. July ((5.13 ± 0.24) μmol·m-2·s-1) for Sophora japonica and August ((3.85 ± 0.17) μmol·m-2·s-1) for Salix matsudana. Stem respiration during the growing season (July and August) was high- er than in the dormant season (November and December); whereas the seasonal variations of Q10 showed opposite trend, i.e. higher in the dormant season than during the growing season. RW increased and the diel patterns of Rw varied with height; the pattern of stem respiration was unimodal for trunk and diauxie for branches, respectively. Stem temperature was found to be the dominant factor regulating the diurnal dynamics of stem respiration at a daily scale. In addition, Q10 higher at the top than at the base. Stem temperature and Q10 collectively determined the temporal and vertical patters of stem respiration. During the growing months, daily accumulated respiration per volume of woody tissue (mmol·m-3·d-1) was linearly related to the inverse of stem diameter measured at breast height. The level of respiration was better expressed on area base (μmol·m-2·s-1) for comparisons among individuals and examination of temporal and spatial variations of the same individual. Therefore, the spatial and temporal variability of Rw should be considered in the construction of city forest carbon budget model so as to reduce the estimation error.
HANFeng-Sen, HUDan, WANGXiao-Lin, ZHOUHong-Xuan. Respiration rates of stems at different heights and their sensitivity to temperature in two broad-leaved trees in Beijing[J]. Chinese Journal of Plant Ecology, 2015, 39(2): 197-205 https://doi.org/10.17521/cjpe.2015.0019
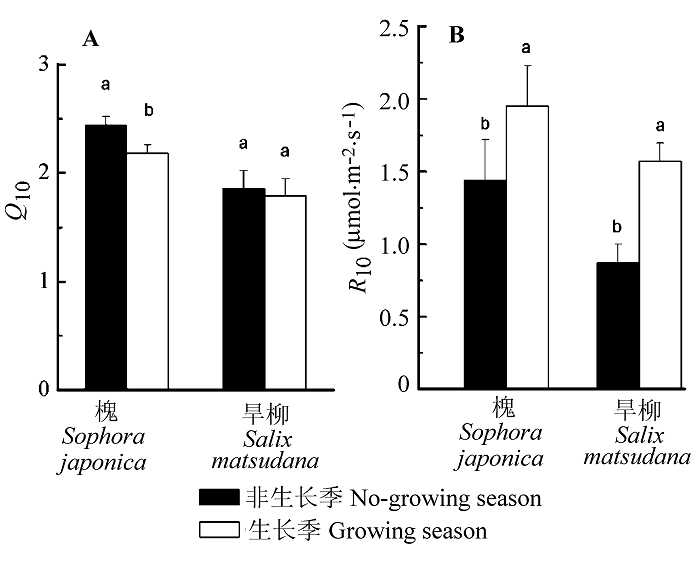
Fig. 2 Seasonal variations of temperature-sensitivity coefficient (Q10, A) and stem respiration at a reference temperature of 10 °C (R10, B) on stem surface area base for the non-growing season (Oct.-Dec.) and growing season (Jun.-Aug.) in Sophora japonica and Salix matsudana (mean ± SD).
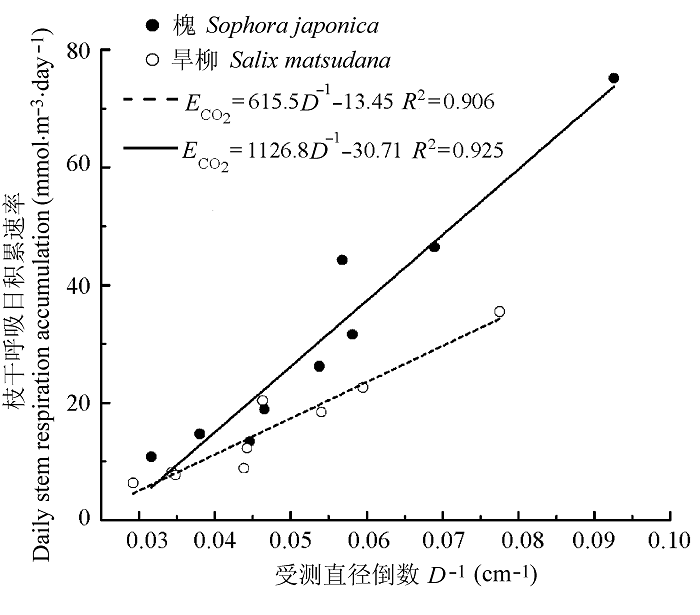
Fig. 5 Analysis of the relationship between the daily stem respiration accumulation per tree expressed on a volume basis and the inverse of stem diameter measured at the breast height (D-1) in Sophora japonica and Salix matsudana.
The McCree-de Wit-Penning de Vries- Thornley respiration paradigms: 30 years later.
1
2000
... 温度一直被认为是造成枝干呼吸时空变化的主要原因(Amthor, 2000).在枝干呼吸研究中, 通常采用温度敏感系数(Q10, 温度每升高10 ℃, 呼吸变化的倍数)表达呼吸对温度的敏感性.研究表明, Q10在不同树种之间差异很大(Zha et al., 2004; Brito et al., 2010; Yang et al., 2011).木质组织Q10的研究报告显示, 90%以上的Q10值处于1.0-3.0之间, 集中分布范围为1.5-2.0, 占总体的42% (王文杰等, 2005).其中, 测量地域、年份和季节等都会影响木质组织Q10值的确定(Atkin & Tjeolker, 2003; Wang et al., 2003; Zha et al., 2004; Brito et al., 2010), 甚至一天之内不同时刻的Q10也有很大不同(Yang et al., 2014).为研究方便, 绝大多数生态系统估测模型将其设置为常数2.0 (Ryan, 1991; Ryan et al., 1994), 给陆地生态系统的碳平衡估测带来了较大误差.因此, 精细量化树木枝干呼吸及其温度敏感性的时间动态和空间变异性是理解森林生态系统碳循环机制的前提条件. ...
Estimation of whole-stem respiration, incorporating vertical and seasonal variations in stem CO2 efflux rate, of Chamaecyparis obtusa trees.
1
2010
... 木质组织呼吸是森林生态系统碳平衡的重要组成部分, 可占植物个体地上自养呼吸(包括叶片呼吸和枝干呼吸)的14%-48% (Vose & Ryan, 2002), 在特定条件下超过了叶片呼吸对系统总碳通量的贡献率(Edwards & Hanson, 1995), 对陆地碳循环起着至关重要的作用(Ryan et al., 1996).由于受测量方法的限制, 枝干呼吸在以往研究中未能引起足够的重视(王秀伟等, 2011).近几十年来, 随着原位测量技术的快速发展以及人们对陆地生态系统碳平衡过程的不断认识, 枝干呼吸逐渐成为森林生态系统碳循环研究中的热点问题之一(Damesin et al., 2002; Zha et al., 2004; Teskey et al., 2008; Araki et al., 2010). ...
Thermal acclimation and the dynamic response of plant respiration to temperature.
1
2003
... 温度一直被认为是造成枝干呼吸时空变化的主要原因(Amthor, 2000).在枝干呼吸研究中, 通常采用温度敏感系数(Q10, 温度每升高10 ℃, 呼吸变化的倍数)表达呼吸对温度的敏感性.研究表明, Q10在不同树种之间差异很大(Zha et al., 2004; Brito et al., 2010; Yang et al., 2011).木质组织Q10的研究报告显示, 90%以上的Q10值处于1.0-3.0之间, 集中分布范围为1.5-2.0, 占总体的42% (王文杰等, 2005).其中, 测量地域、年份和季节等都会影响木质组织Q10值的确定(Atkin & Tjeolker, 2003; Wang et al., 2003; Zha et al., 2004; Brito et al., 2010), 甚至一天之内不同时刻的Q10也有很大不同(Yang et al., 2014).为研究方便, 绝大多数生态系统估测模型将其设置为常数2.0 (Ryan, 1991; Ryan et al., 1994), 给陆地生态系统的碳平衡估测带来了较大误差.因此, 精细量化树木枝干呼吸及其温度敏感性的时间动态和空间变异性是理解森林生态系统碳循环机制的前提条件. ...
Spatial and seasonal variations in stem CO2 efflux of Pinus canariensis at their upper distribution limit.
2
2010
... 温度一直被认为是造成枝干呼吸时空变化的主要原因(Amthor, 2000).在枝干呼吸研究中, 通常采用温度敏感系数(Q10, 温度每升高10 ℃, 呼吸变化的倍数)表达呼吸对温度的敏感性.研究表明, Q10在不同树种之间差异很大(Zha et al., 2004; Brito et al., 2010; Yang et al., 2011).木质组织Q10的研究报告显示, 90%以上的Q10值处于1.0-3.0之间, 集中分布范围为1.5-2.0, 占总体的42% (王文杰等, 2005).其中, 测量地域、年份和季节等都会影响木质组织Q10值的确定(Atkin & Tjeolker, 2003; Wang et al., 2003; Zha et al., 2004; Brito et al., 2010), 甚至一天之内不同时刻的Q10也有很大不同(Yang et al., 2014).为研究方便, 绝大多数生态系统估测模型将其设置为常数2.0 (Ryan, 1991; Ryan et al., 1994), 给陆地生态系统的碳平衡估测带来了较大误差.因此, 精细量化树木枝干呼吸及其温度敏感性的时间动态和空间变异性是理解森林生态系统碳循环机制的前提条件. ...
... ; Brito et al., 2010), 甚至一天之内不同时刻的Q10也有很大不同(Yang et al., 2014).为研究方便, 绝大多数生态系统估测模型将其设置为常数2.0 (Ryan, 1991; Ryan et al., 1994), 给陆地生态系统的碳平衡估测带来了较大误差.因此, 精细量化树木枝干呼吸及其温度敏感性的时间动态和空间变异性是理解森林生态系统碳循环机制的前提条件. ...
Spatial and seasonal variations in stem respiration of beech trees (Fagus sylvatica).
2
2002
... 目前, 对树木整体枝干呼吸的估测方法通常采用局部通量测定法(王秀伟等, 2011), 即通过测量树干局部CO2排放速率(通常是胸高位置), 并以此作为树干呼吸的表征参数上推至树木整体、群落, 乃至整个生态系统(Ryan, 1990).该方法基于一个基本假设: 局部测得的树干CO2释放速率与对应呼吸组织的生物化学反应速率相当, 且不随测量部位的高度发生变化(Sprugel, 1990).然而, 越来越多的研究表明, 局部通量法在枝干呼吸的估测过程中存在极大的不确定性(Ceschia et al., 2002; Teskey et al., 2008).基于该理论开展的气体交换研究结果在不同个体间或者同一个体的不同部位间差异很大(Damesin, 2002; Teskey et al., 2008).Amthor (2000)和王文杰等(2005)先后在部分针叶树种不同高度处的枝干CO2排放规律研究中发现, 单位表面积枝干CO2释放速率在树冠内部远远高于树干, 且顶部呼吸高于基部(Sprugel, 1990; Ceschia et al., 2002).但由于缺乏枝干高度梯度上CO2排放规律的长期连续观测, 枝干呼吸的垂直分布格局及其影响因素并未真正被了解. ...
... ; Ceschia et al., 2002).但由于缺乏枝干高度梯度上CO2排放规律的长期连续观测, 枝干呼吸的垂直分布格局及其影响因素并未真正被了解. ...
Stem and branch respiration of beech: From tree measurements to estimations at the stand level.
2
2002
... 木质组织呼吸是森林生态系统碳平衡的重要组成部分, 可占植物个体地上自养呼吸(包括叶片呼吸和枝干呼吸)的14%-48% (Vose & Ryan, 2002), 在特定条件下超过了叶片呼吸对系统总碳通量的贡献率(Edwards & Hanson, 1995), 对陆地碳循环起着至关重要的作用(Ryan et al., 1996).由于受测量方法的限制, 枝干呼吸在以往研究中未能引起足够的重视(王秀伟等, 2011).近几十年来, 随着原位测量技术的快速发展以及人们对陆地生态系统碳平衡过程的不断认识, 枝干呼吸逐渐成为森林生态系统碳循环研究中的热点问题之一(Damesin et al., 2002; Zha et al., 2004; Teskey et al., 2008; Araki et al., 2010). ...
... 目前, 对树木整体枝干呼吸的估测方法通常采用局部通量测定法(王秀伟等, 2011), 即通过测量树干局部CO2排放速率(通常是胸高位置), 并以此作为树干呼吸的表征参数上推至树木整体、群落, 乃至整个生态系统(Ryan, 1990).该方法基于一个基本假设: 局部测得的树干CO2释放速率与对应呼吸组织的生物化学反应速率相当, 且不随测量部位的高度发生变化(Sprugel, 1990).然而, 越来越多的研究表明, 局部通量法在枝干呼吸的估测过程中存在极大的不确定性(Ceschia et al., 2002; Teskey et al., 2008).基于该理论开展的气体交换研究结果在不同个体间或者同一个体的不同部位间差异很大(Damesin, 2002; Teskey et al., 2008).Amthor (2000)和王文杰等(2005)先后在部分针叶树种不同高度处的枝干CO2排放规律研究中发现, 单位表面积枝干CO2释放速率在树冠内部远远高于树干, 且顶部呼吸高于基部(Sprugel, 1990; Ceschia et al., 2002).但由于缺乏枝干高度梯度上CO2排放规律的长期连续观测, 枝干呼吸的垂直分布格局及其影响因素并未真正被了解. ...
Stem respiration in a closed-canopy upland oak forest.
1
1995
... 木质组织呼吸是森林生态系统碳平衡的重要组成部分, 可占植物个体地上自养呼吸(包括叶片呼吸和枝干呼吸)的14%-48% (Vose & Ryan, 2002), 在特定条件下超过了叶片呼吸对系统总碳通量的贡献率(Edwards & Hanson, 1995), 对陆地碳循环起着至关重要的作用(Ryan et al., 1996).由于受测量方法的限制, 枝干呼吸在以往研究中未能引起足够的重视(王秀伟等, 2011).近几十年来, 随着原位测量技术的快速发展以及人们对陆地生态系统碳平衡过程的不断认识, 枝干呼吸逐渐成为森林生态系统碳循环研究中的热点问题之一(Damesin et al., 2002; Zha et al., 2004; Teskey et al., 2008; Araki et al., 2010). ...
Stem/branch maintenance respiration of Japanese red pine stand.
1
2007
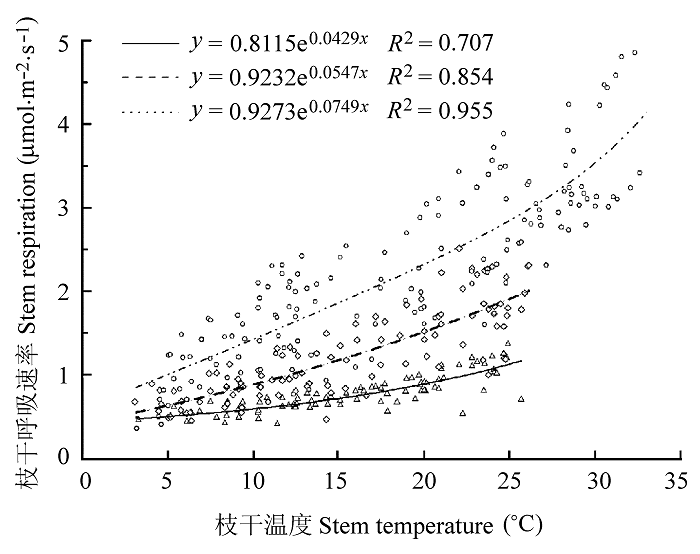
... 木质组织Q10不仅是局部通量法中的关键指标, 也是众多生态系统观测模型的重要参数.本研究着重对两树种枝干呼吸Q10值做了全面的种间比较和种内时空分析.研究结果显示, 槐对温度变化的敏感程度高于旱柳, 具体表现为前者RW年平均Q10值大于后者(2.32 > 1.98).对同一树种而言, 非生长季枝干呼吸对温度变化的敏感性高于生长季, 这是由于非生长季枝干呼吸以维持呼吸为主, 而维持呼吸比生长呼吸对温度变化更为敏感(Saxe et al., 2001).在树干高度梯度上, Q10值随测量高度的增加呈上升趋势, 枝干形成层的生理活性和枝干含水量是影响Q10值的主要原因, 与之前报道的幼林树干呼吸Q10值高于成熟林, 且随林木径级的增加呈降低的趋势的结论(Kim et al., 2007)相吻合.然而, 以往绝大多数研究认为, 树干内部温度及Q10值在树干的所有位置恒定不变(Lavigne, 1996; Ryan et al., 1996), 生态系统模型采用统一的Q10值(2.0)来代表木质组织对温度敏感性的平均水平.但越来越多的研究证明, 年平均Q10值不再是枝干呼吸对温度响应的真实度量, 而是对温度、树干边材比例及活性和枝干含水量等的综合反应(王淼等, 2008), 在枝干呼吸应用中有一定的局限性(Tjoelker et al., 2001).所以, 在今后的研究中, 要准确估测枝干呼吸的整体水平需要更为详细的检测数据, 选择合适的表达单位, 并同步考虑树干温度及其温度敏感系数的时空变异性, 以提高模拟精度. ...
Comparing stem respiration and growth of jack pine provenances from northern and southern locations.
1
1996
... 木质组织Q10不仅是局部通量法中的关键指标, 也是众多生态系统观测模型的重要参数.本研究着重对两树种枝干呼吸Q10值做了全面的种间比较和种内时空分析.研究结果显示, 槐对温度变化的敏感程度高于旱柳, 具体表现为前者RW年平均Q10值大于后者(2.32 > 1.98).对同一树种而言, 非生长季枝干呼吸对温度变化的敏感性高于生长季, 这是由于非生长季枝干呼吸以维持呼吸为主, 而维持呼吸比生长呼吸对温度变化更为敏感(Saxe et al., 2001).在树干高度梯度上, Q10值随测量高度的增加呈上升趋势, 枝干形成层的生理活性和枝干含水量是影响Q10值的主要原因, 与之前报道的幼林树干呼吸Q10值高于成熟林, 且随林木径级的增加呈降低的趋势的结论(Kim et al., 2007)相吻合.然而, 以往绝大多数研究认为, 树干内部温度及Q10值在树干的所有位置恒定不变(Lavigne, 1996; Ryan et al., 1996), 生态系统模型采用统一的Q10值(2.0)来代表木质组织对温度敏感性的平均水平.但越来越多的研究证明, 年平均Q10值不再是枝干呼吸对温度响应的真实度量, 而是对温度、树干边材比例及活性和枝干含水量等的综合反应(王淼等, 2008), 在枝干呼吸应用中有一定的局限性(Tjoelker et al., 2001).所以, 在今后的研究中, 要准确估测枝干呼吸的整体水平需要更为详细的检测数据, 选择合适的表达单位, 并同步考虑树干温度及其温度敏感系数的时空变异性, 以提高模拟精度. ...
Stem CO2 fluxes in two Sahelian shrub species (Guiera senegalensis and Combretum micranthum).
Growth and maintenance respiration in stems of Pinus contorta and Picea engelmannii.
1
1990
... 目前, 对树木整体枝干呼吸的估测方法通常采用局部通量测定法(王秀伟等, 2011), 即通过测量树干局部CO2排放速率(通常是胸高位置), 并以此作为树干呼吸的表征参数上推至树木整体、群落, 乃至整个生态系统(Ryan, 1990).该方法基于一个基本假设: 局部测得的树干CO2释放速率与对应呼吸组织的生物化学反应速率相当, 且不随测量部位的高度发生变化(Sprugel, 1990).然而, 越来越多的研究表明, 局部通量法在枝干呼吸的估测过程中存在极大的不确定性(Ceschia et al., 2002; Teskey et al., 2008).基于该理论开展的气体交换研究结果在不同个体间或者同一个体的不同部位间差异很大(Damesin, 2002; Teskey et al., 2008).Amthor (2000)和王文杰等(2005)先后在部分针叶树种不同高度处的枝干CO2排放规律研究中发现, 单位表面积枝干CO2释放速率在树冠内部远远高于树干, 且顶部呼吸高于基部(Sprugel, 1990; Ceschia et al., 2002).但由于缺乏枝干高度梯度上CO2排放规律的长期连续观测, 枝干呼吸的垂直分布格局及其影响因素并未真正被了解. ...
Effects of climate change on plant respiration.
2
1991
... 温度一直被认为是造成枝干呼吸时空变化的主要原因(Amthor, 2000).在枝干呼吸研究中, 通常采用温度敏感系数(Q10, 温度每升高10 ℃, 呼吸变化的倍数)表达呼吸对温度的敏感性.研究表明, Q10在不同树种之间差异很大(Zha et al., 2004; Brito et al., 2010; Yang et al., 2011).木质组织Q10的研究报告显示, 90%以上的Q10值处于1.0-3.0之间, 集中分布范围为1.5-2.0, 占总体的42% (王文杰等, 2005).其中, 测量地域、年份和季节等都会影响木质组织Q10值的确定(Atkin & Tjeolker, 2003; Wang et al., 2003; Zha et al., 2004; Brito et al., 2010), 甚至一天之内不同时刻的Q10也有很大不同(Yang et al., 2014).为研究方便, 绝大多数生态系统估测模型将其设置为常数2.0 (Ryan, 1991; Ryan et al., 1994), 给陆地生态系统的碳平衡估测带来了较大误差.因此, 精细量化树木枝干呼吸及其温度敏感性的时间动态和空间变异性是理解森林生态系统碳循环机制的前提条件. ...
Woody-tissue respiration for Simarouba amara and Minquartia guianensis, two tropical wet forest trees with different growth habits.
1
1994
... 温度一直被认为是造成枝干呼吸时空变化的主要原因(Amthor, 2000).在枝干呼吸研究中, 通常采用温度敏感系数(Q10, 温度每升高10 ℃, 呼吸变化的倍数)表达呼吸对温度的敏感性.研究表明, Q10在不同树种之间差异很大(Zha et al., 2004; Brito et al., 2010; Yang et al., 2011).木质组织Q10的研究报告显示, 90%以上的Q10值处于1.0-3.0之间, 集中分布范围为1.5-2.0, 占总体的42% (王文杰等, 2005).其中, 测量地域、年份和季节等都会影响木质组织Q10值的确定(Atkin & Tjeolker, 2003; Wang et al., 2003; Zha et al., 2004; Brito et al., 2010), 甚至一天之内不同时刻的Q10也有很大不同(Yang et al., 2014).为研究方便, 绝大多数生态系统估测模型将其设置为常数2.0 (Ryan, 1991; Ryan et al., 1994), 给陆地生态系统的碳平衡估测带来了较大误差.因此, 精细量化树木枝干呼吸及其温度敏感性的时间动态和空间变异性是理解森林生态系统碳循环机制的前提条件. ...
Foliage, fine-root, woody-tissue and stand respiration in Pinus radiata in relation to nitrogen status.
2
1996
... 木质组织呼吸是森林生态系统碳平衡的重要组成部分, 可占植物个体地上自养呼吸(包括叶片呼吸和枝干呼吸)的14%-48% (Vose & Ryan, 2002), 在特定条件下超过了叶片呼吸对系统总碳通量的贡献率(Edwards & Hanson, 1995), 对陆地碳循环起着至关重要的作用(Ryan et al., 1996).由于受测量方法的限制, 枝干呼吸在以往研究中未能引起足够的重视(王秀伟等, 2011).近几十年来, 随着原位测量技术的快速发展以及人们对陆地生态系统碳平衡过程的不断认识, 枝干呼吸逐渐成为森林生态系统碳循环研究中的热点问题之一(Damesin et al., 2002; Zha et al., 2004; Teskey et al., 2008; Araki et al., 2010). ...
... 木质组织Q10不仅是局部通量法中的关键指标, 也是众多生态系统观测模型的重要参数.本研究着重对两树种枝干呼吸Q10值做了全面的种间比较和种内时空分析.研究结果显示, 槐对温度变化的敏感程度高于旱柳, 具体表现为前者RW年平均Q10值大于后者(2.32 > 1.98).对同一树种而言, 非生长季枝干呼吸对温度变化的敏感性高于生长季, 这是由于非生长季枝干呼吸以维持呼吸为主, 而维持呼吸比生长呼吸对温度变化更为敏感(Saxe et al., 2001).在树干高度梯度上, Q10值随测量高度的增加呈上升趋势, 枝干形成层的生理活性和枝干含水量是影响Q10值的主要原因, 与之前报道的幼林树干呼吸Q10值高于成熟林, 且随林木径级的增加呈降低的趋势的结论(Kim et al., 2007)相吻合.然而, 以往绝大多数研究认为, 树干内部温度及Q10值在树干的所有位置恒定不变(Lavigne, 1996; Ryan et al., 1996), 生态系统模型采用统一的Q10值(2.0)来代表木质组织对温度敏感性的平均水平.但越来越多的研究证明, 年平均Q10值不再是枝干呼吸对温度响应的真实度量, 而是对温度、树干边材比例及活性和枝干含水量等的综合反应(王淼等, 2008), 在枝干呼吸应用中有一定的局限性(Tjoelker et al., 2001).所以, 在今后的研究中, 要准确估测枝干呼吸的整体水平需要更为详细的检测数据, 选择合适的表达单位, 并同步考虑树干温度及其温度敏感系数的时空变异性, 以提高模拟精度. ...
Tree and forest functioning in response to global warming.
1
2001
... 木质组织Q10不仅是局部通量法中的关键指标, 也是众多生态系统观测模型的重要参数.本研究着重对两树种枝干呼吸Q10值做了全面的种间比较和种内时空分析.研究结果显示, 槐对温度变化的敏感程度高于旱柳, 具体表现为前者RW年平均Q10值大于后者(2.32 > 1.98).对同一树种而言, 非生长季枝干呼吸对温度变化的敏感性高于生长季, 这是由于非生长季枝干呼吸以维持呼吸为主, 而维持呼吸比生长呼吸对温度变化更为敏感(Saxe et al., 2001).在树干高度梯度上, Q10值随测量高度的增加呈上升趋势, 枝干形成层的生理活性和枝干含水量是影响Q10值的主要原因, 与之前报道的幼林树干呼吸Q10值高于成熟林, 且随林木径级的增加呈降低的趋势的结论(Kim et al., 2007)相吻合.然而, 以往绝大多数研究认为, 树干内部温度及Q10值在树干的所有位置恒定不变(Lavigne, 1996; Ryan et al., 1996), 生态系统模型采用统一的Q10值(2.0)来代表木质组织对温度敏感性的平均水平.但越来越多的研究证明, 年平均Q10值不再是枝干呼吸对温度响应的真实度量, 而是对温度、树干边材比例及活性和枝干含水量等的综合反应(王淼等, 2008), 在枝干呼吸应用中有一定的局限性(Tjoelker et al., 2001).所以, 在今后的研究中, 要准确估测枝干呼吸的整体水平需要更为详细的检测数据, 选择合适的表达单位, 并同步考虑树干温度及其温度敏感系数的时空变异性, 以提高模拟精度. ...
Components of woody-tissue respiration in young Abies amabilis (Dougl.) Forbes trees.
2
1990
... 目前, 对树木整体枝干呼吸的估测方法通常采用局部通量测定法(王秀伟等, 2011), 即通过测量树干局部CO2排放速率(通常是胸高位置), 并以此作为树干呼吸的表征参数上推至树木整体、群落, 乃至整个生态系统(Ryan, 1990).该方法基于一个基本假设: 局部测得的树干CO2释放速率与对应呼吸组织的生物化学反应速率相当, 且不随测量部位的高度发生变化(Sprugel, 1990).然而, 越来越多的研究表明, 局部通量法在枝干呼吸的估测过程中存在极大的不确定性(Ceschia et al., 2002; Teskey et al., 2008).基于该理论开展的气体交换研究结果在不同个体间或者同一个体的不同部位间差异很大(Damesin, 2002; Teskey et al., 2008).Amthor (2000)和王文杰等(2005)先后在部分针叶树种不同高度处的枝干CO2排放规律研究中发现, 单位表面积枝干CO2释放速率在树冠内部远远高于树干, 且顶部呼吸高于基部(Sprugel, 1990; Ceschia et al., 2002).但由于缺乏枝干高度梯度上CO2排放规律的长期连续观测, 枝干呼吸的垂直分布格局及其影响因素并未真正被了解. ...
... 释放速率在树冠内部远远高于树干, 且顶部呼吸高于基部(Sprugel, 1990; Ceschia et al., 2002).但由于缺乏枝干高度梯度上CO2排放规律的长期连续观测, 枝干呼吸的垂直分布格局及其影响因素并未真正被了解. ...
Resistance to radial CO2 diffusion contributes to between-tree variation in CO2 efflux of Populus deltoides stems.
Origin, fate and significance of CO2 in tree stems.
2
2008
... 木质组织呼吸是森林生态系统碳平衡的重要组成部分, 可占植物个体地上自养呼吸(包括叶片呼吸和枝干呼吸)的14%-48% (Vose & Ryan, 2002), 在特定条件下超过了叶片呼吸对系统总碳通量的贡献率(Edwards & Hanson, 1995), 对陆地碳循环起着至关重要的作用(Ryan et al., 1996).由于受测量方法的限制, 枝干呼吸在以往研究中未能引起足够的重视(王秀伟等, 2011).近几十年来, 随着原位测量技术的快速发展以及人们对陆地生态系统碳平衡过程的不断认识, 枝干呼吸逐渐成为森林生态系统碳循环研究中的热点问题之一(Damesin et al., 2002; Zha et al., 2004; Teskey et al., 2008; Araki et al., 2010). ...
... 目前, 对树木整体枝干呼吸的估测方法通常采用局部通量测定法(王秀伟等, 2011), 即通过测量树干局部CO2排放速率(通常是胸高位置), 并以此作为树干呼吸的表征参数上推至树木整体、群落, 乃至整个生态系统(Ryan, 1990).该方法基于一个基本假设: 局部测得的树干CO2释放速率与对应呼吸组织的生物化学反应速率相当, 且不随测量部位的高度发生变化(Sprugel, 1990).然而, 越来越多的研究表明, 局部通量法在枝干呼吸的估测过程中存在极大的不确定性(Ceschia et al., 2002; Teskey et al., 2008).基于该理论开展的气体交换研究结果在不同个体间或者同一个体的不同部位间差异很大(Damesin, 2002; Teskey et al., 2008).Amthor (2000)和王文杰等(2005)先后在部分针叶树种不同高度处的枝干CO2排放规律研究中发现, 单位表面积枝干CO2释放速率在树冠内部远远高于树干, 且顶部呼吸高于基部(Sprugel, 1990; Ceschia et al., 2002).但由于缺乏枝干高度梯度上CO2排放规律的长期连续观测, 枝干呼吸的垂直分布格局及其影响因素并未真正被了解. ...
Modelling respiration of vegetation: Evidence for a general temperature- dependent Q10.
1
2001
... 木质组织Q10不仅是局部通量法中的关键指标, 也是众多生态系统观测模型的重要参数.本研究着重对两树种枝干呼吸Q10值做了全面的种间比较和种内时空分析.研究结果显示, 槐对温度变化的敏感程度高于旱柳, 具体表现为前者RW年平均Q10值大于后者(2.32 > 1.98).对同一树种而言, 非生长季枝干呼吸对温度变化的敏感性高于生长季, 这是由于非生长季枝干呼吸以维持呼吸为主, 而维持呼吸比生长呼吸对温度变化更为敏感(Saxe et al., 2001).在树干高度梯度上, Q10值随测量高度的增加呈上升趋势, 枝干形成层的生理活性和枝干含水量是影响Q10值的主要原因, 与之前报道的幼林树干呼吸Q10值高于成熟林, 且随林木径级的增加呈降低的趋势的结论(Kim et al., 2007)相吻合.然而, 以往绝大多数研究认为, 树干内部温度及Q10值在树干的所有位置恒定不变(Lavigne, 1996; Ryan et al., 1996), 生态系统模型采用统一的Q10值(2.0)来代表木质组织对温度敏感性的平均水平.但越来越多的研究证明, 年平均Q10值不再是枝干呼吸对温度响应的真实度量, 而是对温度、树干边材比例及活性和枝干含水量等的综合反应(王淼等, 2008), 在枝干呼吸应用中有一定的局限性(Tjoelker et al., 2001).所以, 在今后的研究中, 要准确估测枝干呼吸的整体水平需要更为详细的检测数据, 选择合适的表达单位, 并同步考虑树干温度及其温度敏感系数的时空变异性, 以提高模拟精度. ...
Seasonal respiration of foliage, fine roots, and woody tissues in relation to growth, tissue N, and photosynthesis.
1
2002
... 木质组织呼吸是森林生态系统碳平衡的重要组成部分, 可占植物个体地上自养呼吸(包括叶片呼吸和枝干呼吸)的14%-48% (Vose & Ryan, 2002), 在特定条件下超过了叶片呼吸对系统总碳通量的贡献率(Edwards & Hanson, 1995), 对陆地碳循环起着至关重要的作用(Ryan et al., 1996).由于受测量方法的限制, 枝干呼吸在以往研究中未能引起足够的重视(王秀伟等, 2011).近几十年来, 随着原位测量技术的快速发展以及人们对陆地生态系统碳平衡过程的不断认识, 枝干呼吸逐渐成为森林生态系统碳循环研究中的热点问题之一(Damesin et al., 2002; Zha et al., 2004; Teskey et al., 2008; Araki et al., 2010). ...
长白山红松针阔叶混交林主要树种树干呼吸速率
1
2008
... 木质组织Q10不仅是局部通量法中的关键指标, 也是众多生态系统观测模型的重要参数.本研究着重对两树种枝干呼吸Q10值做了全面的种间比较和种内时空分析.研究结果显示, 槐对温度变化的敏感程度高于旱柳, 具体表现为前者RW年平均Q10值大于后者(2.32 > 1.98).对同一树种而言, 非生长季枝干呼吸对温度变化的敏感性高于生长季, 这是由于非生长季枝干呼吸以维持呼吸为主, 而维持呼吸比生长呼吸对温度变化更为敏感(Saxe et al., 2001).在树干高度梯度上, Q10值随测量高度的增加呈上升趋势, 枝干形成层的生理活性和枝干含水量是影响Q10值的主要原因, 与之前报道的幼林树干呼吸Q10值高于成熟林, 且随林木径级的增加呈降低的趋势的结论(Kim et al., 2007)相吻合.然而, 以往绝大多数研究认为, 树干内部温度及Q10值在树干的所有位置恒定不变(Lavigne, 1996; Ryan et al., 1996), 生态系统模型采用统一的Q10值(2.0)来代表木质组织对温度敏感性的平均水平.但越来越多的研究证明, 年平均Q10值不再是枝干呼吸对温度响应的真实度量, 而是对温度、树干边材比例及活性和枝干含水量等的综合反应(王淼等, 2008), 在枝干呼吸应用中有一定的局限性(Tjoelker et al., 2001).所以, 在今后的研究中, 要准确估测枝干呼吸的整体水平需要更为详细的检测数据, 选择合适的表达单位, 并同步考虑树干温度及其温度敏感系数的时空变异性, 以提高模拟精度. ...
长白山红松针阔叶混交林主要树种树干呼吸速率
1
2008
... 木质组织Q10不仅是局部通量法中的关键指标, 也是众多生态系统观测模型的重要参数.本研究着重对两树种枝干呼吸Q10值做了全面的种间比较和种内时空分析.研究结果显示, 槐对温度变化的敏感程度高于旱柳, 具体表现为前者RW年平均Q10值大于后者(2.32 > 1.98).对同一树种而言, 非生长季枝干呼吸对温度变化的敏感性高于生长季, 这是由于非生长季枝干呼吸以维持呼吸为主, 而维持呼吸比生长呼吸对温度变化更为敏感(Saxe et al., 2001).在树干高度梯度上, Q10值随测量高度的增加呈上升趋势, 枝干形成层的生理活性和枝干含水量是影响Q10值的主要原因, 与之前报道的幼林树干呼吸Q10值高于成熟林, 且随林木径级的增加呈降低的趋势的结论(Kim et al., 2007)相吻合.然而, 以往绝大多数研究认为, 树干内部温度及Q10值在树干的所有位置恒定不变(Lavigne, 1996; Ryan et al., 1996), 生态系统模型采用统一的Q10值(2.0)来代表木质组织对温度敏感性的平均水平.但越来越多的研究证明, 年平均Q10值不再是枝干呼吸对温度响应的真实度量, 而是对温度、树干边材比例及活性和枝干含水量等的综合反应(王淼等, 2008), 在枝干呼吸应用中有一定的局限性(Tjoelker et al., 2001).所以, 在今后的研究中, 要准确估测枝干呼吸的整体水平需要更为详细的检测数据, 选择合适的表达单位, 并同步考虑树干温度及其温度敏感系数的时空变异性, 以提高模拟精度. ...
林木非同化器官与土壤呼吸的温度系数Q10值的特征分析
1
2005
... 温度一直被认为是造成枝干呼吸时空变化的主要原因(Amthor, 2000).在枝干呼吸研究中, 通常采用温度敏感系数(Q10, 温度每升高10 ℃, 呼吸变化的倍数)表达呼吸对温度的敏感性.研究表明, Q10在不同树种之间差异很大(Zha et al., 2004; Brito et al., 2010; Yang et al., 2011).木质组织Q10的研究报告显示, 90%以上的Q10值处于1.0-3.0之间, 集中分布范围为1.5-2.0, 占总体的42% (王文杰等, 2005).其中, 测量地域、年份和季节等都会影响木质组织Q10值的确定(Atkin & Tjeolker, 2003; Wang et al., 2003; Zha et al., 2004; Brito et al., 2010), 甚至一天之内不同时刻的Q10也有很大不同(Yang et al., 2014).为研究方便, 绝大多数生态系统估测模型将其设置为常数2.0 (Ryan, 1991; Ryan et al., 1994), 给陆地生态系统的碳平衡估测带来了较大误差.因此, 精细量化树木枝干呼吸及其温度敏感性的时间动态和空间变异性是理解森林生态系统碳循环机制的前提条件. ...
林木非同化器官与土壤呼吸的温度系数Q10值的特征分析
1
2005
... 温度一直被认为是造成枝干呼吸时空变化的主要原因(Amthor, 2000).在枝干呼吸研究中, 通常采用温度敏感系数(Q10, 温度每升高10 ℃, 呼吸变化的倍数)表达呼吸对温度的敏感性.研究表明, Q10在不同树种之间差异很大(Zha et al., 2004; Brito et al., 2010; Yang et al., 2011).木质组织Q10的研究报告显示, 90%以上的Q10值处于1.0-3.0之间, 集中分布范围为1.5-2.0, 占总体的42% (王文杰等, 2005).其中, 测量地域、年份和季节等都会影响木质组织Q10值的确定(Atkin & Tjeolker, 2003; Wang et al., 2003; Zha et al., 2004; Brito et al., 2010), 甚至一天之内不同时刻的Q10也有很大不同(Yang et al., 2014).为研究方便, 绝大多数生态系统估测模型将其设置为常数2.0 (Ryan, 1991; Ryan et al., 1994), 给陆地生态系统的碳平衡估测带来了较大误差.因此, 精细量化树木枝干呼吸及其温度敏感性的时间动态和空间变异性是理解森林生态系统碳循环机制的前提条件. ...
Stem respiration of a larch (Larix gmelini) plantation in northeast China.
1
2003
... 温度一直被认为是造成枝干呼吸时空变化的主要原因(Amthor, 2000).在枝干呼吸研究中, 通常采用温度敏感系数(Q10, 温度每升高10 ℃, 呼吸变化的倍数)表达呼吸对温度的敏感性.研究表明, Q10在不同树种之间差异很大(Zha et al., 2004; Brito et al., 2010; Yang et al., 2011).木质组织Q10的研究报告显示, 90%以上的Q10值处于1.0-3.0之间, 集中分布范围为1.5-2.0, 占总体的42% (王文杰等, 2005).其中, 测量地域、年份和季节等都会影响木质组织Q10值的确定(Atkin & Tjeolker, 2003; Wang et al., 2003; Zha et al., 2004; Brito et al., 2010), 甚至一天之内不同时刻的Q10也有很大不同(Yang et al., 2014).为研究方便, 绝大多数生态系统估测模型将其设置为常数2.0 (Ryan, 1991; Ryan et al., 1994), 给陆地生态系统的碳平衡估测带来了较大误差.因此, 精细量化树木枝干呼吸及其温度敏感性的时间动态和空间变异性是理解森林生态系统碳循环机制的前提条件. ...
春、秋季节树干温度和液流速度对东北3树种树干表面CO2释放通量的影响
2
2011
... 木质组织呼吸是森林生态系统碳平衡的重要组成部分, 可占植物个体地上自养呼吸(包括叶片呼吸和枝干呼吸)的14%-48% (Vose & Ryan, 2002), 在特定条件下超过了叶片呼吸对系统总碳通量的贡献率(Edwards & Hanson, 1995), 对陆地碳循环起着至关重要的作用(Ryan et al., 1996).由于受测量方法的限制, 枝干呼吸在以往研究中未能引起足够的重视(王秀伟等, 2011).近几十年来, 随着原位测量技术的快速发展以及人们对陆地生态系统碳平衡过程的不断认识, 枝干呼吸逐渐成为森林生态系统碳循环研究中的热点问题之一(Damesin et al., 2002; Zha et al., 2004; Teskey et al., 2008; Araki et al., 2010). ...
... 目前, 对树木整体枝干呼吸的估测方法通常采用局部通量测定法(王秀伟等, 2011), 即通过测量树干局部CO2排放速率(通常是胸高位置), 并以此作为树干呼吸的表征参数上推至树木整体、群落, 乃至整个生态系统(Ryan, 1990).该方法基于一个基本假设: 局部测得的树干CO2释放速率与对应呼吸组织的生物化学反应速率相当, 且不随测量部位的高度发生变化(Sprugel, 1990).然而, 越来越多的研究表明, 局部通量法在枝干呼吸的估测过程中存在极大的不确定性(Ceschia et al., 2002; Teskey et al., 2008).基于该理论开展的气体交换研究结果在不同个体间或者同一个体的不同部位间差异很大(Damesin, 2002; Teskey et al., 2008).Amthor (2000)和王文杰等(2005)先后在部分针叶树种不同高度处的枝干CO2排放规律研究中发现, 单位表面积枝干CO2释放速率在树冠内部远远高于树干, 且顶部呼吸高于基部(Sprugel, 1990; Ceschia et al., 2002).但由于缺乏枝干高度梯度上CO2排放规律的长期连续观测, 枝干呼吸的垂直分布格局及其影响因素并未真正被了解. ...
春、秋季节树干温度和液流速度对东北3树种树干表面CO2释放通量的影响
2
2011
... 木质组织呼吸是森林生态系统碳平衡的重要组成部分, 可占植物个体地上自养呼吸(包括叶片呼吸和枝干呼吸)的14%-48% (Vose & Ryan, 2002), 在特定条件下超过了叶片呼吸对系统总碳通量的贡献率(Edwards & Hanson, 1995), 对陆地碳循环起着至关重要的作用(Ryan et al., 1996).由于受测量方法的限制, 枝干呼吸在以往研究中未能引起足够的重视(王秀伟等, 2011).近几十年来, 随着原位测量技术的快速发展以及人们对陆地生态系统碳平衡过程的不断认识, 枝干呼吸逐渐成为森林生态系统碳循环研究中的热点问题之一(Damesin et al., 2002; Zha et al., 2004; Teskey et al., 2008; Araki et al., 2010). ...
... 目前, 对树木整体枝干呼吸的估测方法通常采用局部通量测定法(王秀伟等, 2011), 即通过测量树干局部CO2排放速率(通常是胸高位置), 并以此作为树干呼吸的表征参数上推至树木整体、群落, 乃至整个生态系统(Ryan, 1990).该方法基于一个基本假设: 局部测得的树干CO2释放速率与对应呼吸组织的生物化学反应速率相当, 且不随测量部位的高度发生变化(Sprugel, 1990).然而, 越来越多的研究表明, 局部通量法在枝干呼吸的估测过程中存在极大的不确定性(Ceschia et al., 2002; Teskey et al., 2008).基于该理论开展的气体交换研究结果在不同个体间或者同一个体的不同部位间差异很大(Damesin, 2002; Teskey et al., 2008).Amthor (2000)和王文杰等(2005)先后在部分针叶树种不同高度处的枝干CO2排放规律研究中发现, 单位表面积枝干CO2释放速率在树冠内部远远高于树干, 且顶部呼吸高于基部(Sprugel, 1990; Ceschia et al., 2002).但由于缺乏枝干高度梯度上CO2排放规律的长期连续观测, 枝干呼吸的垂直分布格局及其影响因素并未真正被了解. ...
Temporal and spatial variations of stem CO2 efflux of three species in subtropical China.
1
2011
... 温度一直被认为是造成枝干呼吸时空变化的主要原因(Amthor, 2000).在枝干呼吸研究中, 通常采用温度敏感系数(Q10, 温度每升高10 ℃, 呼吸变化的倍数)表达呼吸对温度的敏感性.研究表明, Q10在不同树种之间差异很大(Zha et al., 2004; Brito et al., 2010; Yang et al., 2011).木质组织Q10的研究报告显示, 90%以上的Q10值处于1.0-3.0之间, 集中分布范围为1.5-2.0, 占总体的42% (王文杰等, 2005).其中, 测量地域、年份和季节等都会影响木质组织Q10值的确定(Atkin & Tjeolker, 2003; Wang et al., 2003; Zha et al., 2004; Brito et al., 2010), 甚至一天之内不同时刻的Q10也有很大不同(Yang et al., 2014).为研究方便, 绝大多数生态系统估测模型将其设置为常数2.0 (Ryan, 1991; Ryan et al., 1994), 给陆地生态系统的碳平衡估测带来了较大误差.因此, 精细量化树木枝干呼吸及其温度敏感性的时间动态和空间变异性是理解森林生态系统碳循环机制的前提条件. ...
Diurnal and seasonal change in stem respiration of Larix principis-rupprechtii trees, northern China.
1
2014
... 温度一直被认为是造成枝干呼吸时空变化的主要原因(Amthor, 2000).在枝干呼吸研究中, 通常采用温度敏感系数(Q10, 温度每升高10 ℃, 呼吸变化的倍数)表达呼吸对温度的敏感性.研究表明, Q10在不同树种之间差异很大(Zha et al., 2004; Brito et al., 2010; Yang et al., 2011).木质组织Q10的研究报告显示, 90%以上的Q10值处于1.0-3.0之间, 集中分布范围为1.5-2.0, 占总体的42% (王文杰等, 2005).其中, 测量地域、年份和季节等都会影响木质组织Q10值的确定(Atkin & Tjeolker, 2003; Wang et al., 2003; Zha et al., 2004; Brito et al., 2010), 甚至一天之内不同时刻的Q10也有很大不同(Yang et al., 2014).为研究方便, 绝大多数生态系统估测模型将其设置为常数2.0 (Ryan, 1991; Ryan et al., 1994), 给陆地生态系统的碳平衡估测带来了较大误差.因此, 精细量化树木枝干呼吸及其温度敏感性的时间动态和空间变异性是理解森林生态系统碳循环机制的前提条件. ...
Seasonal and annual stem respiration of Scots pine trees under boreal conditions.
3
2004
... 木质组织呼吸是森林生态系统碳平衡的重要组成部分, 可占植物个体地上自养呼吸(包括叶片呼吸和枝干呼吸)的14%-48% (Vose & Ryan, 2002), 在特定条件下超过了叶片呼吸对系统总碳通量的贡献率(Edwards & Hanson, 1995), 对陆地碳循环起着至关重要的作用(Ryan et al., 1996).由于受测量方法的限制, 枝干呼吸在以往研究中未能引起足够的重视(王秀伟等, 2011).近几十年来, 随着原位测量技术的快速发展以及人们对陆地生态系统碳平衡过程的不断认识, 枝干呼吸逐渐成为森林生态系统碳循环研究中的热点问题之一(Damesin et al., 2002; Zha et al., 2004; Teskey et al., 2008; Araki et al., 2010). ...
... 温度一直被认为是造成枝干呼吸时空变化的主要原因(Amthor, 2000).在枝干呼吸研究中, 通常采用温度敏感系数(Q10, 温度每升高10 ℃, 呼吸变化的倍数)表达呼吸对温度的敏感性.研究表明, Q10在不同树种之间差异很大(Zha et al., 2004; Brito et al., 2010; Yang et al., 2011).木质组织Q10的研究报告显示, 90%以上的Q10值处于1.0-3.0之间, 集中分布范围为1.5-2.0, 占总体的42% (王文杰等, 2005).其中, 测量地域、年份和季节等都会影响木质组织Q10值的确定(Atkin & Tjeolker, 2003; Wang et al., 2003; Zha et al., 2004; Brito et al., 2010), 甚至一天之内不同时刻的Q10也有很大不同(Yang et al., 2014).为研究方便, 绝大多数生态系统估测模型将其设置为常数2.0 (Ryan, 1991; Ryan et al., 1994), 给陆地生态系统的碳平衡估测带来了较大误差.因此, 精细量化树木枝干呼吸及其温度敏感性的时间动态和空间变异性是理解森林生态系统碳循环机制的前提条件. ...
... ; Zha et al., 2004; Brito et al., 2010), 甚至一天之内不同时刻的Q10也有很大不同(Yang et al., 2014).为研究方便, 绝大多数生态系统估测模型将其设置为常数2.0 (Ryan, 1991; Ryan et al., 1994), 给陆地生态系统的碳平衡估测带来了较大误差.因此, 精细量化树木枝干呼吸及其温度敏感性的时间动态和空间变异性是理解森林生态系统碳循环机制的前提条件. ...
 , 王晓琳, 周宏轩
, 王晓琳, 周宏轩
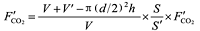


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}