伴随着水文条件的恢复, 芦苇(Phragmites australis )在许多湿地迅速建群扩展, 限制了自然植被的恢复(Zedler & Kercher, 2004 )。芦苇具有植株高大、致密, 凋落物积累多等特点, 导致到达地表的有效光辐射减少(Larkin et al ., 2012 )。不同植物对弱光生境的适应性不同, 最终决定群落物种组成; 那些在竞争光资源方面较弱的本土物种便被外来种竞争替代(Perry & Galatowitsch, 2003 ; Kotowski et al ., 2013 )。因此, 在湿地生态系统研究芦苇对不同光照条件的生态响应, 有助于理解芦苇成功扩张的机制, 为植被管理实践提供科学依据。
植物种子萌发信息对于预测野外植物群落动态和植被管理十分重要(Grime et al ., 1981 )。Nikolaje- vskij (1971) 和Haslam (1973) 研究表明高光照是促进芦苇种子萌发的必要条件, 而另外一个研究则发现芦苇种子萌发结果在黑暗和光照条件下无明显差异(Greenwood & MacFarlane, 2006 )。可见, 芦苇种子萌发与光照之间的关系还需进一步探究。不同研究报道的芦苇种子萌发对水文条件的响应也有差别, 如Haslam (1972) 报道允许芦苇萌发的最深水位不超过5 cm, Yu等(2012) 则证实芦苇在15 cm水深仍有2%的种子能够萌发。并且, 湿地植物的表现往往还受光照、水深交互作用的影响(Thouveno et al ., 2013 ), 而芦苇种子萌发与两者交互作用关系的研究鲜有报道。
莫莫格自然保护区位于松嫩平原西部, 面积14.4 × 104 hm2 , 设立该保护区的主要目的是保护湿地及在此栖息的白鹤(Grus leucogeranus )、丹顶鹤(Grus japonensis )等珍稀水禽(Pan et al ., 2006 )。扁秆藨草(Scirpus planiculmis )是莫莫格湿地的重要植物, 也是白鹤在该地停栖的主要食源植物(Hui et al ., 2009 )。由于气候变化和人类活动的影响, 扁秆藨草湿地退化严重。当地从2010年开始生态补水以恢复白鹤觅食生境。然而, 扁秆藨草在恢复三年的湿地很少出现, 而芦苇等高大湿地植物快速扩展, 成为群落建群种。目前关于该区域芦苇种子萌发特征的报道十分少见, 限制了对芦苇群落动态的认识, 制约了湿地植被的管理。
为此, 我们在实验室模拟野外情景, 研究光照和水深对芦苇种子萌发的影响, 以回答以下问题: (1)光照和水位变化对松嫩平原湿地芦苇种子萌发有何影响?(2)如何通过控制水文及光照条件有效限制芦苇通过有性繁殖更新?
1 材料和方法
1.1 植物材料
实验用芦苇种子来自莫莫格湿地保护区。莫莫格湿地位于吉林省镇赉县境内, 地理坐标为45.75°- 46.17° N, 122.45°-124.07° E, 属于嫩江及其支流冲积而成的低平原, 平均海拔142 m; 研究区年平均气温为4.2 ℃, 年蒸发量1553 mm, 年降水量391.88 mm, 降水主要集中在6-8月(Wang et al ., 2010 )。于2013年9月, 选择不同位置的30株芦苇, 整穗采集, 装入布袋中; 回室内后, 将种子置于黑暗条件下风干, 一个月后进行休眠检测。萌发实验前, 将种子用0.1%的KMnO4 浸泡12 h, 然后用蒸馏水反复清洗以除掉残余药液。
1.2 萌发试验设计
为了明确光照和水深对芦苇种子萌发的影响, 我们设计了3种光照(全光、遮阴、黑暗)和4种水深(0, 5, 15和30 cm)共12个处理组合, 每处理5次重复(共60组培养皿)。遮阴及水位的选择均基于我们在野外的观测结果。采用两层绿色塑料网遮盖模拟野外遮阴条件(50 μmol·m-2 ·s-1 ), 通过双层铝包装实现完全黑暗条件。通过补充蒸馏水维持实验水深。
用玻璃胶分别将50粒种子均匀固定于直径11 cm的培养皿底部, 置于不同处理条件下。萌发实验在培养箱(HPG-280HX, 哈尔滨市东联电子技术开发有限公司)进行, 培养箱设置的光温条件为光照12 h/25 ℃和黑暗12 h/15 ℃。每隔一天记录一次种子萌发情况, 在暗处绿光条件下进行, 幼芽伸长约1 mm视为成功萌发。
1.3 统计分析
采用双因素方差分析明确光照、水深及其互作对芦苇种子萌发的影响。方差分析前, 先进行数据正态性和方差齐性检验(Shapiro-Wilk和Homogeneity-of- variance), 若数据不满足正态性, 对数据进行对数转换。方差分析后对数据进行多重比较(Tukey)。数据处理和分析在Excel和SPSS 16.0软件下完成。
2 结果
2.1 光照的影响
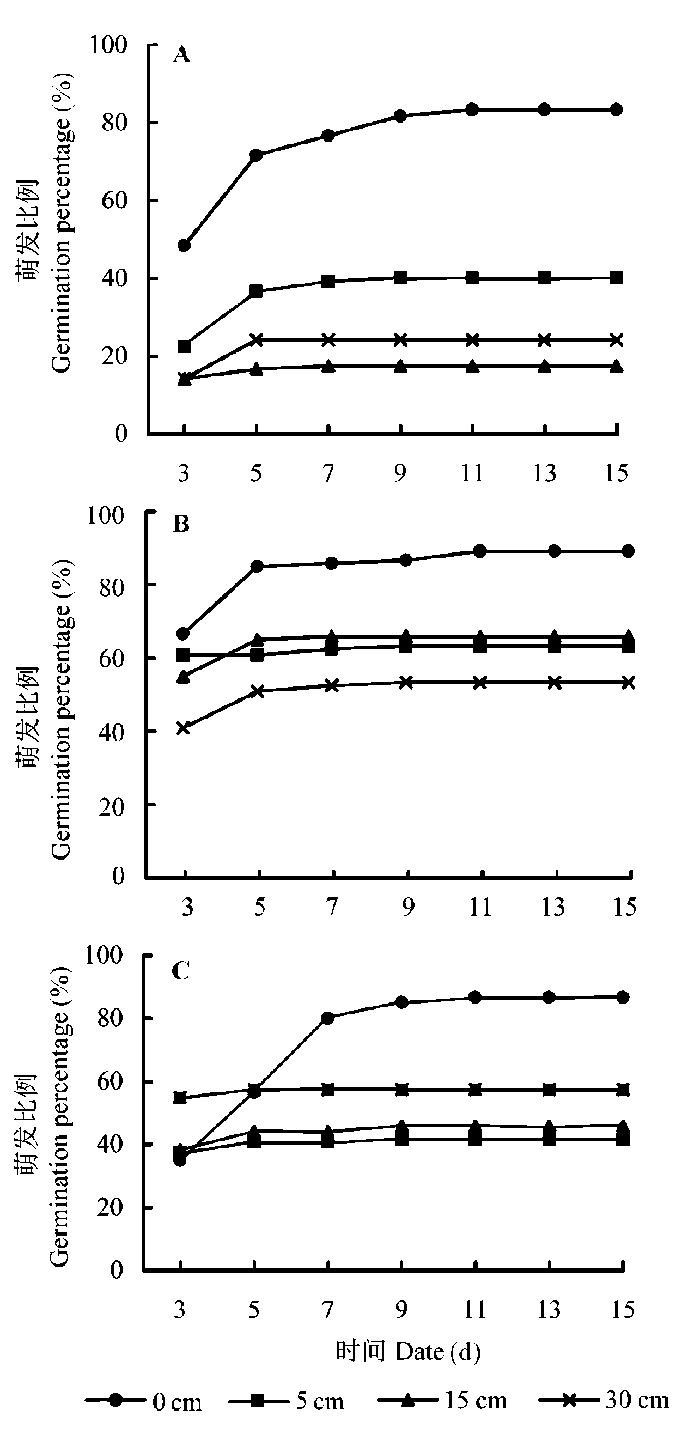
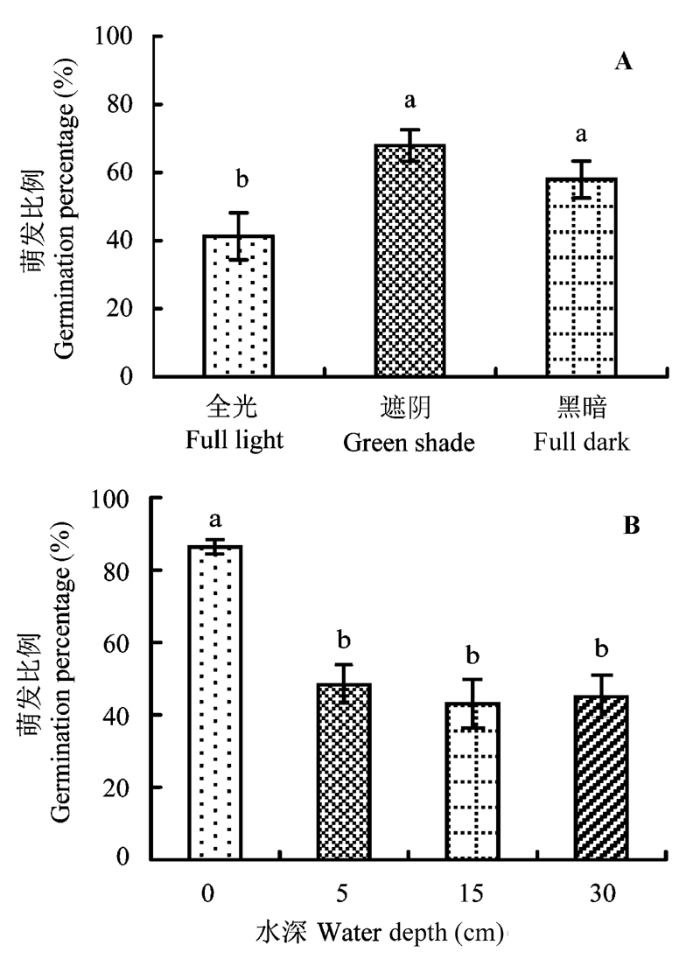
由图1 可以看出, 在不同处理组合下, 芦苇种子在实验开始后的第3天开始萌发, 大多数处理的种子在接下来的2天达到萌发高峰, 随后只有少量种子萌发, 共持续15天。统计分析表明, 单独光照条件明显影响芦苇种子的萌发(F = 19.14, p < 0.001)。最终萌发比例在全光下显著低于在遮阴和黑暗环境, 而萌发种子数量在遮阴和黑暗条件下差异不显著(图2 )。
图1 芦苇种子萌发特征在不同光照和水深下的变化。A, 全光。B, 遮阴。C, 黑暗。
Fig. 1 Accumulative percentage in seed germination of Phragmites australis through time responding to light and water depth conditions. A, Full light. B, Green shade. C, Full dark.
图2 光照和水深条件对芦苇种子萌发的影响(平均值±标准误差)。不同小写字母表示差异显著(p < 0.05)。
Fig. 2 Comparisons of light (A) and water depth (B) on seed germination rates of Phragmites australis (mean ± SE ). Differ- ent lower-case letters indicate significant difference (p < 0.05).
2.2 水深的作用
不考虑互作因素, 水深变化对芦苇种子萌发也有极显著影响(F = 33.50, p < 0.001), 即较深水位明显抑制种子萌芽。由图2 可以看出, 最终萌发比例在0 cm浅水处理最高(86.39% ± 1.99%), 且显著高于其他较深水位下的数值; 而较深水位(5-30 cm)下的萌发种子数量无显著性差异。
2.3 交互作用对芦苇种子萌发的影响
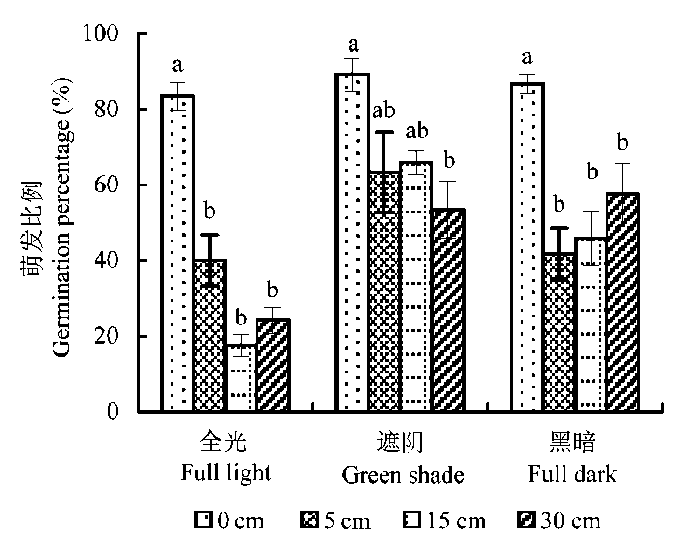
在所有3种光照处理下, 芦苇种子均在0 cm水深萌发最多, 萌发种子数量在5-30 cm较深水位差异不显著。统计分析表明, 芦苇种子萌发明显受光照和水深交互作用的影响(F = 3.27, p < 0.05)。在全光照条件下, 种子萌发比例在0 cm水深为(86.67 ± 2.36)%, 明显高于其他较深水位下的数值(图3 ); 在遮阴和黑暗环境, 0 cm水深与其他水深间萌发比例的变化范围比在全光条件下小, 且在较深水位(5-30 cm)下的萌发比例要高于在全光照条件下同水深下的数值。
图3 不同光照条件下水深变化对芦苇种子萌发的影响(平均值±标准误差)。不同小写字母表示差异显著(p < 0.05)。
Fig. 3 Comparisons of water depth on the final germination rates of Phragmites australis at full light, green shade and full dark conditions (mean ± SE ). Different lower-case letters indicate significant differences (p < 0.05).
3 讨论
3.1 光照对芦苇种子萌发的影响
种子萌发对光照、水分等环境因子的响应有助于预测物种在野外的更新机制, 为湿地植被管理提供可靠信息(Laubhan & Shaffer, 2006 ; Chauhan & Johnson, 2011 )。在本研究中, 芦苇种子在遮阴和黑暗条件下的萌发情况明显好于在全光条件下(图2 ), 这与李有志(2007) 、Nikolajevskij (1971) 、Haslam (1973) 及Ekstam等(1999) 的研究不同; 这是因为本研究综合考虑了光照和不同水深对芦苇种子萌发的影响, 而已有文献仅在湿润条件下研究萌发和光照间的关系。弱光条件下较高的萌发比例意味着较强的适应遮阴环境的能力, 有利于芦苇在湿地恢复初期从土壤中成功萌发、定居, 然后通过根茎营养繁殖迅速建群扩张。并且, 在莫莫格湿地, 与扁秆藨草相比, 芦苇植株高大(1.8-2.4 m), 致密(盖度80%-95%), 凋落物积累较多, 大量累积的凋落物不仅降低了到达地表的有效光辐射, 也通过机械阻碍作用抑制其他植物的生存生长(Bergelson, 1990 ; Larkin et al ., 2012 )。
多数研究表明芦苇种子在遮阴、黑暗和全光条件下均能萌发(Ekstam et al ., 1999 ; Greenwood & MacFarlane, 2006 ), 本研究结果与之类似, 说明尽管芦苇种子生产量大, 但不易在土壤中形成持久种子库, 在这些湿地通过改变光照条件来限制芦苇的更新也是行不通的。不过, 也有研究发现芦苇种子在有光条件下才能萌发(Nikolajevskij, 1971 ; Haslam, 1973 ), 这可能与芦苇适应不同气候、湿地类型等生态条件有关。
3.2 水深对芦苇种子萌发的影响
在湿地生态系统, 较深的水位往往导致植物不能萌发或萌发比例下降(Li et al ., 2009 )。本研究表明, 芦苇在0 cm水深下的萌发明显高于其他较深水位(图2 ), 这种趋势与Yu等(2012) 在黄河三角洲的研究类似。另外, 本研究发现在5-30 cm水深有超过40%的芦苇种子能够萌发, 而Haslam (1972) 报道允许芦苇萌发的最深水位不超过5 cm, 这可能与盐碱湿地芦苇种子特殊的生理适应特征有关。
在利用洪水控制非目标种时, 需要考虑目标种的适宜水位。洪水对扁秆藨草萌发有明显的抑制作用, 在20 cm水深只有3%的种子萌发, 而在1 cm水深萌发比例达81% (吴竟仑和周恒昌, 2006 )。李慧芳和章光新(2013) 的研究表明扁秆藨草在0-5 cm水深条件下更新生长最佳。因此, 在莫莫格湿地采用洪水处理(5-30 cm)难以有效地控制芦苇扩张, 促进扁秆藨草更新。
3.3 光照、水深交互作用
芦苇幼苗的生长除受光照、水深单独作用外, 也受两者交互作用的影响(Thouveno et al ., 2013 )。我们发现, 水位和光照也同时影响芦苇种子的萌发, 因为种子萌发随水位变化的格局在不同光照条件下有差别。在0 cm水深有超过80%的芦苇种子能够萌发、且与光照条件无关, 而弱光条件能够缓解较深水位对芦苇种子萌芽的抑制作用(图3 ), 这可能是本研究芦苇种子萌发比例在遮阴和黑暗条件下(不考虑水深)也较高的重要原因。
莫莫格湿地位于干旱半干旱区, 水资源十分珍贵。因此, 提高湿地恢复的水分利用效率十分重要。另外, 芦苇湿地土壤含有较多的扁秆藨草种子, 且扁秆藨草种子浮力较高, 可以通过水传播远距离扩散, 这为扁秆藨草的恢复提供了种源保障(未发表数据)。因此, 在恢复湿地芦苇建群初期, 及时进行清除处理并保持一定的地表水位可以限制芦苇通过有性繁殖更新, 同时改善地表光辐射等生境条件, 促进扁秆藨草恢复。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Bergelson J 1990 ). Life after death: Site pre-emption by the remains of Poa annua.
Ecology , 71 , 2157 -2165 .
[本文引用: 1]
[2]
Chauhan BS Johnson DE 2011 ). Ecological studies on Echinochloa crus-galli and the implications for weed management in direct-seeded rice.
Crop Protection , 30 , 1385 -1391 .
[本文引用: 1]
[3]
Ekstam B Johannesson R Milberg P 1999 ). The effect of light and number of diurnal temperature fluctuations on germination of Phragmites australis.
Seed Science Research , 9 , 165 -170 .
[本文引用: 2]
[4]
Greenwood ME MacFarlane GR 2006 ). Effects of salinity and temperature on the germination of Phragmites australis, Juncus kraussii, and Juncus acutus: Implications for estuarine restoration initiatives.
Wetlands , 26 , 854 -861 .
[本文引用: 2]
[5]
Grime JP Mason G Curtis AV Rodman J Band SR 1981 ). A comparative study of germination characteristics in a local flora.
Journal of Ecology , 69 , 1017 -1059 .
[本文引用: 1]
[6]
Haslam SM 1972 ). Biological flora of the British Isles. Phragmites communis Trin.
Journal of Ecology , 60 , 585 -610 .
[本文引用: 1]
[7]
Haslam SM 1973 ). Some aspects of the life history and autecology of Phragmites communis Trin. a review.
Polskie Archiwum Hydrobiologii , 20 , 79 -100 .
[本文引用: 3]
[8]
Hui SR Li X Zhang ZX Ning Y 2009 ). Influences of soil salt contents on growth of Scirpus bicincavys.
Journal of Liaoning Forestry Science & Technology , (1 ), 25 -28 . (in Chinese with English abstract)
[本文引用: 1]
[惠淑荣 , 李响 , 张志翔 , 宁宇 (2009 ). 土壤含盐量对扁秆藨草生长的影响
. 辽宁林业科技 , (1 ), 25 -28 .]
[本文引用: 1]
[9]
Kotowski W Dzierża P Czerwiński M Kozub Ł Śnieg S 2013 ). Shrub removal facilitates recovery of wetland species in a rewetted fen.
Journal for Nature Conservation , 21 , 294 -308 .
[本文引用: 1]
[10]
Larkin DJ Freyman MJ Lishawa SC Geddes P Tuchman NC 2012 ). Mechanisms of dominance by the invasive hybrid cattail Typha × glauca.
Biological Invasions , 14 , 65 -77 .
[本文引用: 2]
[11]
Laubhan MK Shaffer TL 2006 ). Seed germination of Cirsium arvense and Lepidium latifolium: Implications for management of montane wetlands.
Wetlands , 26 , 69 -78 .
[本文引用: 1]
[12]
Li HF Zhang GX 2013 ). Influence of water depth and salinity coupling on growth of Scirpus planiculmis seedlings in Momoge national nature reserve.
Wetland Science , 11 , 173 -177 . (in Chinese with English abstract)
[本文引用: 1]
[李惠芳 , 章光新 (2013 ). 水盐交互作用对莫莫格国家级自然保护区扁秆藨草幼苗生长的影响
. 湿地科学 , 11 , 173 -177 .]
[本文引用: 1]
[13]
Li YZ 2007 ). Studies on the Responses of Seed Germination and Seedling Growth of Calamagrostis angustifolia, Phragmites Australis to Environmental Factors.
Master degree dissertation, Hunan Agricultural University, Cha- ngsha . 10 -15 . (in Chinese).
[本文引用: 1]
[李有志 (2007 ). 小叶章和芦苇种子萌发以及幼苗生长对环境因子的响应研究
. 硕士学位论文, 湖南农业大学, 长沙 . 10 -15 .]
[本文引用: 1]
[14]
Li YZ Zhang CM Xie YH Liu F 2009 ). Germination of Deyeuxia angustifolia as affected by soil type, burial depth, water depth and oxygen level.
Mitigation and Adaptation Strategies for Global Change , 14 , 537 -545 .
[本文引用: 1]
[15]
Nikolajevskij VG 1971 ). Research into the biology of the common reed (Phragmites communis Trin.) in the U.S.S.R.
Folia Geobotanica et Phytotaxonomica , 6 , 221 -230 .
[本文引用: 3]
[16]
Pan XL Zhang DY Quan L 2006 ). Interactive factors leading to dying-off Carex tato in Momoge wetland polluted by crude oil, Western Jilin, China.
Chemosphere , 65 , 1772 -1777 .
[本文引用: 1]
[17]
Perry LG Galatowitsch SM 2003 ). A test of two annual cover crops for controlling Phalaris arundinacea invasion in restored sedge meadow wetlands.
Restoration Ecology , 11 , 297 -307 .
[本文引用: 1]
[18]
Thouvenot L Haury J Thiébaut G 2013 ). Seasonal plasticity of Ludwigia grandiflora under light and water depth gradients: An outdoor mesocosm experiment.
Flora , 208 , 430 -437 .
[本文引用: 2]
[19]
Wang XY Feng J Zhao JM 2010 ). Effects of crude oil residuals on soil chemical properties in oil sites, Momoge Wetland, China.
Environmental Monitoring and Assessment , 161 , 271 -280 .
[本文引用: 1]
[20]
Wu JL Zhou HC 2006 ). Seed bank of perennial weeds in paddy fields.
Chinese Journal of Rice Science , 20 , 89 -96 . (in Chinese with English abstract)
[本文引用: 1]
[吴竞仑 , 周恒昌 (2006 ). 稻田土壤多年生杂草种子库研究
. 中国水稻科学 , 20 , 89 -96 .]
[本文引用: 1]
[21]
Yu JB Wang XH Ning K Li YZ Wu HF Fu YQ Zhou D Guan B Lin QX 2012 ). Effects of salinity and water depth on germination of Phragmites australis in coastal wetland of the Yellow River Delta.
CLEAN-Soil, Air, Water , 40 , 1154 -1158 .
[本文引用: 2]
[22]
Zedler JB Kercher S 2004 ). Causes and consequences of invasive plants in wetlands: Opportunities, opportunists, and outcomes.
Critical Reviews in Plant Sciences , 23 , 431 -452 .
[本文引用: 1]
Life after death: Site pre-emption by the remains of Poa annua.
1
1990
... 种子萌发对光照、水分等环境因子的响应有助于预测物种在野外的更新机制, 为湿地植被管理提供可靠信息(Laubhan & Shaffer, 2006 ; Chauhan & Johnson, 2011 ).在本研究中, 芦苇种子在遮阴和黑暗条件下的萌发情况明显好于在全光条件下(图2 ), 这与李有志(2007) 、Nikolajevskij (1971) 、Haslam (1973) 及Ekstam等(1999) 的研究不同; 这是因为本研究综合考虑了光照和不同水深对芦苇种子萌发的影响, 而已有文献仅在湿润条件下研究萌发和光照间的关系.弱光条件下较高的萌发比例意味着较强的适应遮阴环境的能力, 有利于芦苇在湿地恢复初期从土壤中成功萌发、定居, 然后通过根茎营养繁殖迅速建群扩张.并且, 在莫莫格湿地, 与扁秆藨草相比, 芦苇植株高大(1.8-2.4 m), 致密(盖度80%-95%), 凋落物积累较多, 大量累积的凋落物不仅降低了到达地表的有效光辐射, 也通过机械阻碍作用抑制其他植物的生存生长(Bergelson, 1990 ; Larkin et al ., 2012 ). ...
Ecological studies on Echinochloa crus-galli and the implications for weed management in direct-seeded rice.
1
2011
... 种子萌发对光照、水分等环境因子的响应有助于预测物种在野外的更新机制, 为湿地植被管理提供可靠信息(Laubhan & Shaffer, 2006 ; Chauhan & Johnson, 2011 ).在本研究中, 芦苇种子在遮阴和黑暗条件下的萌发情况明显好于在全光条件下(图2 ), 这与李有志(2007) 、Nikolajevskij (1971) 、Haslam (1973) 及Ekstam等(1999) 的研究不同; 这是因为本研究综合考虑了光照和不同水深对芦苇种子萌发的影响, 而已有文献仅在湿润条件下研究萌发和光照间的关系.弱光条件下较高的萌发比例意味着较强的适应遮阴环境的能力, 有利于芦苇在湿地恢复初期从土壤中成功萌发、定居, 然后通过根茎营养繁殖迅速建群扩张.并且, 在莫莫格湿地, 与扁秆藨草相比, 芦苇植株高大(1.8-2.4 m), 致密(盖度80%-95%), 凋落物积累较多, 大量累积的凋落物不仅降低了到达地表的有效光辐射, 也通过机械阻碍作用抑制其他植物的生存生长(Bergelson, 1990 ; Larkin et al ., 2012 ). ...
The effect of light and number of diurnal temperature fluctuations on germination of Phragmites australis.
2
1999
... 种子萌发对光照、水分等环境因子的响应有助于预测物种在野外的更新机制, 为湿地植被管理提供可靠信息(Laubhan & Shaffer, 2006 ; Chauhan & Johnson, 2011 ).在本研究中, 芦苇种子在遮阴和黑暗条件下的萌发情况明显好于在全光条件下(图2 ), 这与李有志(2007) 、Nikolajevskij (1971) 、Haslam (1973) 及Ekstam等(1999) 的研究不同; 这是因为本研究综合考虑了光照和不同水深对芦苇种子萌发的影响, 而已有文献仅在湿润条件下研究萌发和光照间的关系.弱光条件下较高的萌发比例意味着较强的适应遮阴环境的能力, 有利于芦苇在湿地恢复初期从土壤中成功萌发、定居, 然后通过根茎营养繁殖迅速建群扩张.并且, 在莫莫格湿地, 与扁秆藨草相比, 芦苇植株高大(1.8-2.4 m), 致密(盖度80%-95%), 凋落物积累较多, 大量累积的凋落物不仅降低了到达地表的有效光辐射, 也通过机械阻碍作用抑制其他植物的生存生长(Bergelson, 1990 ; Larkin et al ., 2012 ). ...
... 多数研究表明芦苇种子在遮阴、黑暗和全光条件下均能萌发(Ekstam et al ., 1999 ; Greenwood & MacFarlane, 2006 ), 本研究结果与之类似, 说明尽管芦苇种子生产量大, 但不易在土壤中形成持久种子库, 在这些湿地通过改变光照条件来限制芦苇的更新也是行不通的.不过, 也有研究发现芦苇种子在有光条件下才能萌发(Nikolajevskij, 1971 ; Haslam, 1973 ), 这可能与芦苇适应不同气候、湿地类型等生态条件有关. ...
Effects of salinity and temperature on the germination of Phragmites australis, Juncus kraussii, and Juncus acutus: Implications for estuarine restoration initiatives.
2
2006
... 植物种子萌发信息对于预测野外植物群落动态和植被管理十分重要(Grime et al ., 1981 ).Nikolaje- vskij (1971) 和Haslam (1973) 研究表明高光照是促进芦苇种子萌发的必要条件, 而另外一个研究则发现芦苇种子萌发结果在黑暗和光照条件下无明显差异(Greenwood & MacFarlane, 2006 ).可见, 芦苇种子萌发与光照之间的关系还需进一步探究.不同研究报道的芦苇种子萌发对水文条件的响应也有差别, 如Haslam (1972) 报道允许芦苇萌发的最深水位不超过5 cm, Yu等(2012) 则证实芦苇在15 cm水深仍有2%的种子能够萌发.并且, 湿地植物的表现往往还受光照、水深交互作用的影响(Thouveno et al ., 2013 ), 而芦苇种子萌发与两者交互作用关系的研究鲜有报道. ...
... 多数研究表明芦苇种子在遮阴、黑暗和全光条件下均能萌发(Ekstam et al ., 1999 ; Greenwood & MacFarlane, 2006 ), 本研究结果与之类似, 说明尽管芦苇种子生产量大, 但不易在土壤中形成持久种子库, 在这些湿地通过改变光照条件来限制芦苇的更新也是行不通的.不过, 也有研究发现芦苇种子在有光条件下才能萌发(Nikolajevskij, 1971 ; Haslam, 1973 ), 这可能与芦苇适应不同气候、湿地类型等生态条件有关. ...
A comparative study of germination characteristics in a local flora.
1
1981
... 植物种子萌发信息对于预测野外植物群落动态和植被管理十分重要(Grime et al ., 1981 ).Nikolaje- vskij (1971) 和Haslam (1973) 研究表明高光照是促进芦苇种子萌发的必要条件, 而另外一个研究则发现芦苇种子萌发结果在黑暗和光照条件下无明显差异(Greenwood & MacFarlane, 2006 ).可见, 芦苇种子萌发与光照之间的关系还需进一步探究.不同研究报道的芦苇种子萌发对水文条件的响应也有差别, 如Haslam (1972) 报道允许芦苇萌发的最深水位不超过5 cm, Yu等(2012) 则证实芦苇在15 cm水深仍有2%的种子能够萌发.并且, 湿地植物的表现往往还受光照、水深交互作用的影响(Thouveno et al ., 2013 ), 而芦苇种子萌发与两者交互作用关系的研究鲜有报道. ...
Biological flora of the British Isles. Phragmites communis Trin.
1
1972
... 在湿地生态系统, 较深的水位往往导致植物不能萌发或萌发比例下降(Li et al ., 2009 ).本研究表明, 芦苇在0 cm水深下的萌发明显高于其他较深水位(图2 ), 这种趋势与Yu等(2012) 在黄河三角洲的研究类似.另外, 本研究发现在5-30 cm水深有超过40%的芦苇种子能够萌发, 而Haslam (1972) 报道允许芦苇萌发的最深水位不超过5 cm, 这可能与盐碱湿地芦苇种子特殊的生理适应特征有关. ...
Some aspects of the life history and autecology of Phragmites communis Trin. a review.
3
1973
... 植物种子萌发信息对于预测野外植物群落动态和植被管理十分重要(Grime et al ., 1981 ).Nikolaje- vskij (1971) 和Haslam (1973) 研究表明高光照是促进芦苇种子萌发的必要条件, 而另外一个研究则发现芦苇种子萌发结果在黑暗和光照条件下无明显差异(Greenwood & MacFarlane, 2006 ).可见, 芦苇种子萌发与光照之间的关系还需进一步探究.不同研究报道的芦苇种子萌发对水文条件的响应也有差别, 如Haslam (1972) 报道允许芦苇萌发的最深水位不超过5 cm, Yu等(2012) 则证实芦苇在15 cm水深仍有2%的种子能够萌发.并且, 湿地植物的表现往往还受光照、水深交互作用的影响(Thouveno et al ., 2013 ), 而芦苇种子萌发与两者交互作用关系的研究鲜有报道. ...
... 种子萌发对光照、水分等环境因子的响应有助于预测物种在野外的更新机制, 为湿地植被管理提供可靠信息(Laubhan & Shaffer, 2006 ; Chauhan & Johnson, 2011 ).在本研究中, 芦苇种子在遮阴和黑暗条件下的萌发情况明显好于在全光条件下(图2 ), 这与李有志(2007) 、Nikolajevskij (1971) 、Haslam (1973) 及Ekstam等(1999) 的研究不同; 这是因为本研究综合考虑了光照和不同水深对芦苇种子萌发的影响, 而已有文献仅在湿润条件下研究萌发和光照间的关系.弱光条件下较高的萌发比例意味着较强的适应遮阴环境的能力, 有利于芦苇在湿地恢复初期从土壤中成功萌发、定居, 然后通过根茎营养繁殖迅速建群扩张.并且, 在莫莫格湿地, 与扁秆藨草相比, 芦苇植株高大(1.8-2.4 m), 致密(盖度80%-95%), 凋落物积累较多, 大量累积的凋落物不仅降低了到达地表的有效光辐射, 也通过机械阻碍作用抑制其他植物的生存生长(Bergelson, 1990 ; Larkin et al ., 2012 ). ...
... 多数研究表明芦苇种子在遮阴、黑暗和全光条件下均能萌发(Ekstam et al ., 1999 ; Greenwood & MacFarlane, 2006 ), 本研究结果与之类似, 说明尽管芦苇种子生产量大, 但不易在土壤中形成持久种子库, 在这些湿地通过改变光照条件来限制芦苇的更新也是行不通的.不过, 也有研究发现芦苇种子在有光条件下才能萌发(Nikolajevskij, 1971 ; Haslam, 1973 ), 这可能与芦苇适应不同气候、湿地类型等生态条件有关. ...
土壤含盐量对扁秆藨草生长的影响
1
2009
... 莫莫格自然保护区位于松嫩平原西部, 面积14.4 × 104 hm2 , 设立该保护区的主要目的是保护湿地及在此栖息的白鹤(Grus leucogeranus )、丹顶鹤(Grus japonensis )等珍稀水禽(Pan et al ., 2006 ).扁秆藨草(Scirpus planiculmis )是莫莫格湿地的重要植物, 也是白鹤在该地停栖的主要食源植物(Hui et al ., 2009 ).由于气候变化和人类活动的影响, 扁秆藨草湿地退化严重.当地从2010年开始生态补水以恢复白鹤觅食生境.然而, 扁秆藨草在恢复三年的湿地很少出现, 而芦苇等高大湿地植物快速扩展, 成为群落建群种.目前关于该区域芦苇种子萌发特征的报道十分少见, 限制了对芦苇群落动态的认识, 制约了湿地植被的管理. ...
土壤含盐量对扁秆藨草生长的影响
1
2009
... 莫莫格自然保护区位于松嫩平原西部, 面积14.4 × 104 hm2 , 设立该保护区的主要目的是保护湿地及在此栖息的白鹤(Grus leucogeranus )、丹顶鹤(Grus japonensis )等珍稀水禽(Pan et al ., 2006 ).扁秆藨草(Scirpus planiculmis )是莫莫格湿地的重要植物, 也是白鹤在该地停栖的主要食源植物(Hui et al ., 2009 ).由于气候变化和人类活动的影响, 扁秆藨草湿地退化严重.当地从2010年开始生态补水以恢复白鹤觅食生境.然而, 扁秆藨草在恢复三年的湿地很少出现, 而芦苇等高大湿地植物快速扩展, 成为群落建群种.目前关于该区域芦苇种子萌发特征的报道十分少见, 限制了对芦苇群落动态的认识, 制约了湿地植被的管理. ...
Shrub removal facilitates recovery of wetland species in a rewetted fen.
1
2013
... 伴随着水文条件的恢复, 芦苇(Phragmites australis )在许多湿地迅速建群扩展, 限制了自然植被的恢复(Zedler & Kercher, 2004 ).芦苇具有植株高大、致密, 凋落物积累多等特点, 导致到达地表的有效光辐射减少(Larkin et al ., 2012 ).不同植物对弱光生境的适应性不同, 最终决定群落物种组成; 那些在竞争光资源方面较弱的本土物种便被外来种竞争替代(Perry & Galatowitsch, 2003 ; Kotowski et al ., 2013 ).因此, 在湿地生态系统研究芦苇对不同光照条件的生态响应, 有助于理解芦苇成功扩张的机制, 为植被管理实践提供科学依据. ...
Mechanisms of dominance by the invasive hybrid cattail Typha × glauca.
2
2012
... 伴随着水文条件的恢复, 芦苇(Phragmites australis )在许多湿地迅速建群扩展, 限制了自然植被的恢复(Zedler & Kercher, 2004 ).芦苇具有植株高大、致密, 凋落物积累多等特点, 导致到达地表的有效光辐射减少(Larkin et al ., 2012 ).不同植物对弱光生境的适应性不同, 最终决定群落物种组成; 那些在竞争光资源方面较弱的本土物种便被外来种竞争替代(Perry & Galatowitsch, 2003 ; Kotowski et al ., 2013 ).因此, 在湿地生态系统研究芦苇对不同光照条件的生态响应, 有助于理解芦苇成功扩张的机制, 为植被管理实践提供科学依据. ...
... 种子萌发对光照、水分等环境因子的响应有助于预测物种在野外的更新机制, 为湿地植被管理提供可靠信息(Laubhan & Shaffer, 2006 ; Chauhan & Johnson, 2011 ).在本研究中, 芦苇种子在遮阴和黑暗条件下的萌发情况明显好于在全光条件下(图2 ), 这与李有志(2007) 、Nikolajevskij (1971) 、Haslam (1973) 及Ekstam等(1999) 的研究不同; 这是因为本研究综合考虑了光照和不同水深对芦苇种子萌发的影响, 而已有文献仅在湿润条件下研究萌发和光照间的关系.弱光条件下较高的萌发比例意味着较强的适应遮阴环境的能力, 有利于芦苇在湿地恢复初期从土壤中成功萌发、定居, 然后通过根茎营养繁殖迅速建群扩张.并且, 在莫莫格湿地, 与扁秆藨草相比, 芦苇植株高大(1.8-2.4 m), 致密(盖度80%-95%), 凋落物积累较多, 大量累积的凋落物不仅降低了到达地表的有效光辐射, 也通过机械阻碍作用抑制其他植物的生存生长(Bergelson, 1990 ; Larkin et al ., 2012 ). ...
Seed germination of Cirsium arvense and Lepidium latifolium: Implications for management of montane wetlands.
1
2006
... 种子萌发对光照、水分等环境因子的响应有助于预测物种在野外的更新机制, 为湿地植被管理提供可靠信息(Laubhan & Shaffer, 2006 ; Chauhan & Johnson, 2011 ).在本研究中, 芦苇种子在遮阴和黑暗条件下的萌发情况明显好于在全光条件下(图2 ), 这与李有志(2007) 、Nikolajevskij (1971) 、Haslam (1973) 及Ekstam等(1999) 的研究不同; 这是因为本研究综合考虑了光照和不同水深对芦苇种子萌发的影响, 而已有文献仅在湿润条件下研究萌发和光照间的关系.弱光条件下较高的萌发比例意味着较强的适应遮阴环境的能力, 有利于芦苇在湿地恢复初期从土壤中成功萌发、定居, 然后通过根茎营养繁殖迅速建群扩张.并且, 在莫莫格湿地, 与扁秆藨草相比, 芦苇植株高大(1.8-2.4 m), 致密(盖度80%-95%), 凋落物积累较多, 大量累积的凋落物不仅降低了到达地表的有效光辐射, 也通过机械阻碍作用抑制其他植物的生存生长(Bergelson, 1990 ; Larkin et al ., 2012 ). ...
水盐交互作用对莫莫格国家级自然保护区扁秆藨草幼苗生长的影响
1
2013
... 在利用洪水控制非目标种时, 需要考虑目标种的适宜水位.洪水对扁秆藨草萌发有明显的抑制作用, 在20 cm水深只有3%的种子萌发, 而在1 cm水深萌发比例达81% (吴竟仑和周恒昌, 2006 ).李慧芳和章光新(2013) 的研究表明扁秆藨草在0-5 cm水深条件下更新生长最佳.因此, 在莫莫格湿地采用洪水处理(5-30 cm)难以有效地控制芦苇扩张, 促进扁秆藨草更新. ...
水盐交互作用对莫莫格国家级自然保护区扁秆藨草幼苗生长的影响
1
2013
... 在利用洪水控制非目标种时, 需要考虑目标种的适宜水位.洪水对扁秆藨草萌发有明显的抑制作用, 在20 cm水深只有3%的种子萌发, 而在1 cm水深萌发比例达81% (吴竟仑和周恒昌, 2006 ).李慧芳和章光新(2013) 的研究表明扁秆藨草在0-5 cm水深条件下更新生长最佳.因此, 在莫莫格湿地采用洪水处理(5-30 cm)难以有效地控制芦苇扩张, 促进扁秆藨草更新. ...
小叶章和芦苇种子萌发以及幼苗生长对环境因子的响应研究
1
2007
... 种子萌发对光照、水分等环境因子的响应有助于预测物种在野外的更新机制, 为湿地植被管理提供可靠信息(Laubhan & Shaffer, 2006 ; Chauhan & Johnson, 2011 ).在本研究中, 芦苇种子在遮阴和黑暗条件下的萌发情况明显好于在全光条件下(图2 ), 这与李有志(2007) 、Nikolajevskij (1971) 、Haslam (1973) 及Ekstam等(1999) 的研究不同; 这是因为本研究综合考虑了光照和不同水深对芦苇种子萌发的影响, 而已有文献仅在湿润条件下研究萌发和光照间的关系.弱光条件下较高的萌发比例意味着较强的适应遮阴环境的能力, 有利于芦苇在湿地恢复初期从土壤中成功萌发、定居, 然后通过根茎营养繁殖迅速建群扩张.并且, 在莫莫格湿地, 与扁秆藨草相比, 芦苇植株高大(1.8-2.4 m), 致密(盖度80%-95%), 凋落物积累较多, 大量累积的凋落物不仅降低了到达地表的有效光辐射, 也通过机械阻碍作用抑制其他植物的生存生长(Bergelson, 1990 ; Larkin et al ., 2012 ). ...
小叶章和芦苇种子萌发以及幼苗生长对环境因子的响应研究
1
2007
... 种子萌发对光照、水分等环境因子的响应有助于预测物种在野外的更新机制, 为湿地植被管理提供可靠信息(Laubhan & Shaffer, 2006 ; Chauhan & Johnson, 2011 ).在本研究中, 芦苇种子在遮阴和黑暗条件下的萌发情况明显好于在全光条件下(图2 ), 这与李有志(2007) 、Nikolajevskij (1971) 、Haslam (1973) 及Ekstam等(1999) 的研究不同; 这是因为本研究综合考虑了光照和不同水深对芦苇种子萌发的影响, 而已有文献仅在湿润条件下研究萌发和光照间的关系.弱光条件下较高的萌发比例意味着较强的适应遮阴环境的能力, 有利于芦苇在湿地恢复初期从土壤中成功萌发、定居, 然后通过根茎营养繁殖迅速建群扩张.并且, 在莫莫格湿地, 与扁秆藨草相比, 芦苇植株高大(1.8-2.4 m), 致密(盖度80%-95%), 凋落物积累较多, 大量累积的凋落物不仅降低了到达地表的有效光辐射, 也通过机械阻碍作用抑制其他植物的生存生长(Bergelson, 1990 ; Larkin et al ., 2012 ). ...
Germination of Deyeuxia angustifolia as affected by soil type, burial depth, water depth and oxygen level.
1
2009
... 在湿地生态系统, 较深的水位往往导致植物不能萌发或萌发比例下降(Li et al ., 2009 ).本研究表明, 芦苇在0 cm水深下的萌发明显高于其他较深水位(图2 ), 这种趋势与Yu等(2012) 在黄河三角洲的研究类似.另外, 本研究发现在5-30 cm水深有超过40%的芦苇种子能够萌发, 而Haslam (1972) 报道允许芦苇萌发的最深水位不超过5 cm, 这可能与盐碱湿地芦苇种子特殊的生理适应特征有关. ...
Research into the biology of the common reed (Phragmites communis Trin.) in the U.S.S.R.
3
1971
... 植物种子萌发信息对于预测野外植物群落动态和植被管理十分重要(Grime et al ., 1981 ).Nikolaje- vskij (1971) 和Haslam (1973) 研究表明高光照是促进芦苇种子萌发的必要条件, 而另外一个研究则发现芦苇种子萌发结果在黑暗和光照条件下无明显差异(Greenwood & MacFarlane, 2006 ).可见, 芦苇种子萌发与光照之间的关系还需进一步探究.不同研究报道的芦苇种子萌发对水文条件的响应也有差别, 如Haslam (1972) 报道允许芦苇萌发的最深水位不超过5 cm, Yu等(2012) 则证实芦苇在15 cm水深仍有2%的种子能够萌发.并且, 湿地植物的表现往往还受光照、水深交互作用的影响(Thouveno et al ., 2013 ), 而芦苇种子萌发与两者交互作用关系的研究鲜有报道. ...
... 种子萌发对光照、水分等环境因子的响应有助于预测物种在野外的更新机制, 为湿地植被管理提供可靠信息(Laubhan & Shaffer, 2006 ; Chauhan & Johnson, 2011 ).在本研究中, 芦苇种子在遮阴和黑暗条件下的萌发情况明显好于在全光条件下(图2 ), 这与李有志(2007) 、Nikolajevskij (1971) 、Haslam (1973) 及Ekstam等(1999) 的研究不同; 这是因为本研究综合考虑了光照和不同水深对芦苇种子萌发的影响, 而已有文献仅在湿润条件下研究萌发和光照间的关系.弱光条件下较高的萌发比例意味着较强的适应遮阴环境的能力, 有利于芦苇在湿地恢复初期从土壤中成功萌发、定居, 然后通过根茎营养繁殖迅速建群扩张.并且, 在莫莫格湿地, 与扁秆藨草相比, 芦苇植株高大(1.8-2.4 m), 致密(盖度80%-95%), 凋落物积累较多, 大量累积的凋落物不仅降低了到达地表的有效光辐射, 也通过机械阻碍作用抑制其他植物的生存生长(Bergelson, 1990 ; Larkin et al ., 2012 ). ...
... 多数研究表明芦苇种子在遮阴、黑暗和全光条件下均能萌发(Ekstam et al ., 1999 ; Greenwood & MacFarlane, 2006 ), 本研究结果与之类似, 说明尽管芦苇种子生产量大, 但不易在土壤中形成持久种子库, 在这些湿地通过改变光照条件来限制芦苇的更新也是行不通的.不过, 也有研究发现芦苇种子在有光条件下才能萌发(Nikolajevskij, 1971 ; Haslam, 1973 ), 这可能与芦苇适应不同气候、湿地类型等生态条件有关. ...
Interactive factors leading to dying-off Carex tato in Momoge wetland polluted by crude oil, Western Jilin, China.
1
2006
... 莫莫格自然保护区位于松嫩平原西部, 面积14.4 × 104 hm2 , 设立该保护区的主要目的是保护湿地及在此栖息的白鹤(Grus leucogeranus )、丹顶鹤(Grus japonensis )等珍稀水禽(Pan et al ., 2006 ).扁秆藨草(Scirpus planiculmis )是莫莫格湿地的重要植物, 也是白鹤在该地停栖的主要食源植物(Hui et al ., 2009 ).由于气候变化和人类活动的影响, 扁秆藨草湿地退化严重.当地从2010年开始生态补水以恢复白鹤觅食生境.然而, 扁秆藨草在恢复三年的湿地很少出现, 而芦苇等高大湿地植物快速扩展, 成为群落建群种.目前关于该区域芦苇种子萌发特征的报道十分少见, 限制了对芦苇群落动态的认识, 制约了湿地植被的管理. ...
A test of two annual cover crops for controlling Phalaris arundinacea invasion in restored sedge meadow wetlands.
1
2003
... 伴随着水文条件的恢复, 芦苇(Phragmites australis )在许多湿地迅速建群扩展, 限制了自然植被的恢复(Zedler & Kercher, 2004 ).芦苇具有植株高大、致密, 凋落物积累多等特点, 导致到达地表的有效光辐射减少(Larkin et al ., 2012 ).不同植物对弱光生境的适应性不同, 最终决定群落物种组成; 那些在竞争光资源方面较弱的本土物种便被外来种竞争替代(Perry & Galatowitsch, 2003 ; Kotowski et al ., 2013 ).因此, 在湿地生态系统研究芦苇对不同光照条件的生态响应, 有助于理解芦苇成功扩张的机制, 为植被管理实践提供科学依据. ...
Seasonal plasticity of Ludwigia grandiflora under light and water depth gradients: An outdoor mesocosm experiment.
2
2013
... 植物种子萌发信息对于预测野外植物群落动态和植被管理十分重要(Grime et al ., 1981 ).Nikolaje- vskij (1971) 和Haslam (1973) 研究表明高光照是促进芦苇种子萌发的必要条件, 而另外一个研究则发现芦苇种子萌发结果在黑暗和光照条件下无明显差异(Greenwood & MacFarlane, 2006 ).可见, 芦苇种子萌发与光照之间的关系还需进一步探究.不同研究报道的芦苇种子萌发对水文条件的响应也有差别, 如Haslam (1972) 报道允许芦苇萌发的最深水位不超过5 cm, Yu等(2012) 则证实芦苇在15 cm水深仍有2%的种子能够萌发.并且, 湿地植物的表现往往还受光照、水深交互作用的影响(Thouveno et al ., 2013 ), 而芦苇种子萌发与两者交互作用关系的研究鲜有报道. ...
... 芦苇幼苗的生长除受光照、水深单独作用外, 也受两者交互作用的影响(Thouveno et al ., 2013 ).我们发现, 水位和光照也同时影响芦苇种子的萌发, 因为种子萌发随水位变化的格局在不同光照条件下有差别.在0 cm水深有超过80%的芦苇种子能够萌发、且与光照条件无关, 而弱光条件能够缓解较深水位对芦苇种子萌芽的抑制作用(图3 ), 这可能是本研究芦苇种子萌发比例在遮阴和黑暗条件下(不考虑水深)也较高的重要原因. ...
Effects of crude oil residuals on soil chemical properties in oil sites, Momoge Wetland, China.
1
2010
... 实验用芦苇种子来自莫莫格湿地保护区.莫莫格湿地位于吉林省镇赉县境内, 地理坐标为45.75°- 46.17° N, 122.45°-124.07° E, 属于嫩江及其支流冲积而成的低平原, 平均海拔142 m; 研究区年平均气温为4.2 ℃, 年蒸发量1553 mm, 年降水量391.88 mm, 降水主要集中在6-8月(Wang et al ., 2010 ).于2013年9月, 选择不同位置的30株芦苇, 整穗采集, 装入布袋中; 回室内后, 将种子置于黑暗条件下风干, 一个月后进行休眠检测.萌发实验前, 将种子用0.1%的KMnO4 浸泡12 h, 然后用蒸馏水反复清洗以除掉残余药液. ...
稻田土壤多年生杂草种子库研究
1
2006
... 在利用洪水控制非目标种时, 需要考虑目标种的适宜水位.洪水对扁秆藨草萌发有明显的抑制作用, 在20 cm水深只有3%的种子萌发, 而在1 cm水深萌发比例达81% (吴竟仑和周恒昌, 2006 ).李慧芳和章光新(2013) 的研究表明扁秆藨草在0-5 cm水深条件下更新生长最佳.因此, 在莫莫格湿地采用洪水处理(5-30 cm)难以有效地控制芦苇扩张, 促进扁秆藨草更新. ...
稻田土壤多年生杂草种子库研究
1
2006
... 在利用洪水控制非目标种时, 需要考虑目标种的适宜水位.洪水对扁秆藨草萌发有明显的抑制作用, 在20 cm水深只有3%的种子萌发, 而在1 cm水深萌发比例达81% (吴竟仑和周恒昌, 2006 ).李慧芳和章光新(2013) 的研究表明扁秆藨草在0-5 cm水深条件下更新生长最佳.因此, 在莫莫格湿地采用洪水处理(5-30 cm)难以有效地控制芦苇扩张, 促进扁秆藨草更新. ...
Effects of salinity and water depth on germination of Phragmites australis in coastal wetland of the Yellow River Delta.
2
2012
... 植物种子萌发信息对于预测野外植物群落动态和植被管理十分重要(Grime et al ., 1981 ).Nikolaje- vskij (1971) 和Haslam (1973) 研究表明高光照是促进芦苇种子萌发的必要条件, 而另外一个研究则发现芦苇种子萌发结果在黑暗和光照条件下无明显差异(Greenwood & MacFarlane, 2006 ).可见, 芦苇种子萌发与光照之间的关系还需进一步探究.不同研究报道的芦苇种子萌发对水文条件的响应也有差别, 如Haslam (1972) 报道允许芦苇萌发的最深水位不超过5 cm, Yu等(2012) 则证实芦苇在15 cm水深仍有2%的种子能够萌发.并且, 湿地植物的表现往往还受光照、水深交互作用的影响(Thouveno et al ., 2013 ), 而芦苇种子萌发与两者交互作用关系的研究鲜有报道. ...
... 在湿地生态系统, 较深的水位往往导致植物不能萌发或萌发比例下降(Li et al ., 2009 ).本研究表明, 芦苇在0 cm水深下的萌发明显高于其他较深水位(图2 ), 这种趋势与Yu等(2012) 在黄河三角洲的研究类似.另外, 本研究发现在5-30 cm水深有超过40%的芦苇种子能够萌发, 而Haslam (1972) 报道允许芦苇萌发的最深水位不超过5 cm, 这可能与盐碱湿地芦苇种子特殊的生理适应特征有关. ...
Causes and consequences of invasive plants in wetlands: Opportunities, opportunists, and outcomes.
1
2004
... 伴随着水文条件的恢复, 芦苇(Phragmites australis )在许多湿地迅速建群扩展, 限制了自然植被的恢复(Zedler & Kercher, 2004 ).芦苇具有植株高大、致密, 凋落物积累多等特点, 导致到达地表的有效光辐射减少(Larkin et al ., 2012 ).不同植物对弱光生境的适应性不同, 最终决定群落物种组成; 那些在竞争光资源方面较弱的本土物种便被外来种竞争替代(Perry & Galatowitsch, 2003 ; Kotowski et al ., 2013 ).因此, 在湿地生态系统研究芦苇对不同光照条件的生态响应, 有助于理解芦苇成功扩张的机制, 为植被管理实践提供科学依据. ...
 , 姜明
, 姜明

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}