版权声明: 2016 植物生态学报编辑部 本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。
基金资助:
国家自然科学基金(30770365和31070383)。
展开
摘要
探究叶片性状的空间变异性和环境关联性有助于我们理解植物对环境的适应策略和群落建构机制。该研究以浙江天童5 hm2大型动态样地所有胸径≥1 cm的木本植物为对象, 测定了20 253株个体的单叶面积、比叶面积和叶片干物质含量, 土壤总氮、总磷、总碳、pH值、体积含水率、容重和腐殖质, 以及海拔、坡度和凹凸度等; 并用地统计学等方法分析了叶片性状的空间变异性及其与环境因子的相关性。结果表明: 1)单叶面积的空间变异最大, 比叶面积次之, 叶片干物质含量最小。三者在0-5.16 m空间范围内表现出较弱的空间自相关, 其半变异函数的最优模型分别为高斯模型、指数模型和指数模型。2)叶片性状空间变异具有方向性, 单叶面积空间变异在东北-西南方向上最大, 在西北-东南方向上最小; 比叶面积和叶片干物质含量的空间变异均在西北-东南方向最大, 在东北-西南方向最小。3)单叶面积与地形因子显著负相关(r = -0.12, p < 0.0001); 比叶面积与土壤养分显著负相关(r = -0.16, p < 0.0001), 叶片干物质含量与土壤养分显著正相关(r = 0.13, p < 0.0001)。4)东北-西南方向上, 地形因子对单叶面积、比叶面积和叶片干物质含量空间变异的影响大于土壤养分; 西北-东南方向上, 地形因子对单叶面积空间变异的影响相对较大, 而土壤养分对比叶面积和叶片干物质含量空间变异的影响较大。总之, 在研究样地内, 植物叶片性状随空间距离和方向存在很大的变异性, 叶片性状与地形因子、土壤养分的关联性间接表明了环境过滤对群落构建的影响。
关键词:
Abstract
Keywords:
植物性状是指一切对植物的定居、生存和适应有着潜在重要影响, 或与获取、利用和保护资源的能力有关的属性(Reich et al., 2003; Violle et al., 2007), 反映了植物对不同环境的适应及植物内部不同功能之间的生理或进化权衡后的一种属性表现(Grime et al., 1997; Díaz et al., 2004; 孟婷婷等, 2007)。近年来, 由于植物性状能够作为联系植物生理适应性和环境梯度间的桥梁, 在重建群落生态学、预测生态系统响应等方面广泛应用(Wright et al., 2004; Beaumont & Burns, 2009)。
由于性状的空间变异(spatial variation)能解释环境对性状的影响, 可联系生态系统的结构和功能过程, 因而近年来受到生态学家的普遍关注。一般认为, 引起植物性状空间变异的因素主要有环境差异和物种组成, 而物种间的关系比较复杂, 因此, 本文只探讨环境异质性对植物性状空间变异的影响。植物性状与环境的关系已经是性状研究的核心问题(丁佳等, 2011)。例如, Wright等(2004)发现气候因子大约能够解释植物叶片性状变异的18%; 降水梯度和资源利用能够影响中国东北样带草原植物性状的变化(Wang & Ni, 2005a; 2005b)。同时, 研究认为, 环境的空间异质性能够解释性状的空间变异, 随空间距离的增加, 受环境筛选的影响, 适应性或功能上差异较大的物种分布在空间距离较远的位置, 而性状值相近的物种分布在空间距离较近的位置(Weiher & Keddy, 1999; Cornwell et al., 2006)。因此, 植物性状会发生空间变异(Villar & Merino, 2001; Reich et al., 2003; 刘璐等, 2010; 张莉等, 2013)。譬如: 植物性状随土壤肥力和酸碱程度在空间上出现变化(Liu et al., 2012), 叶片厚度、气孔密度随海拔升高而增加, 叶片大小和叶面积随海拔升高而降低(Hölscher et al., 2002; Craine & Lee, 2003)。同时, 在局域尺度上, 由于山体、沟谷的分布和走向, 土壤养分随地形的变化, 使得性状在不同方向的空间变异性表现出差异。如沿沟谷方向分布的物种叶面积大、叶片干物质含量小; 山脊方向, 由于土壤养分、水分的限制, 叶面积较小, 叶片干物质含量较大。而植物性状空间变异是一个连续的过程, 为了精确探究叶片性状的空间变异性, 需要用基于每木个体的空间差异进行连续采样的方法。
为了精确探究植物叶片性状在局域环境中的空间变异及其与环境因子的相关性, 我们在浙江天童国家森林公园20 hm2常绿阔叶林动态样地内建立了5 hm2固定样地, 开展植物性状空间变异以及与环境关系的研究。以样地内所有胸径≥1 cm的植株的叶片性状(单叶面积、比叶面积、叶片干物质含量)为研究对象, 提出以下科学假说: 1)植物叶片性状存在空间变异性; 2)局域尺度上的微地形, 由于不同方向上地形、土壤养分的差异, 使得植物叶片性状存在空间方向变异性; 3)环境因子可以解释部分植物叶片性状的空间变异。研究这些问题对理解环境过滤对植物群落构建以及物种对环境的适应策略具有重要意义。
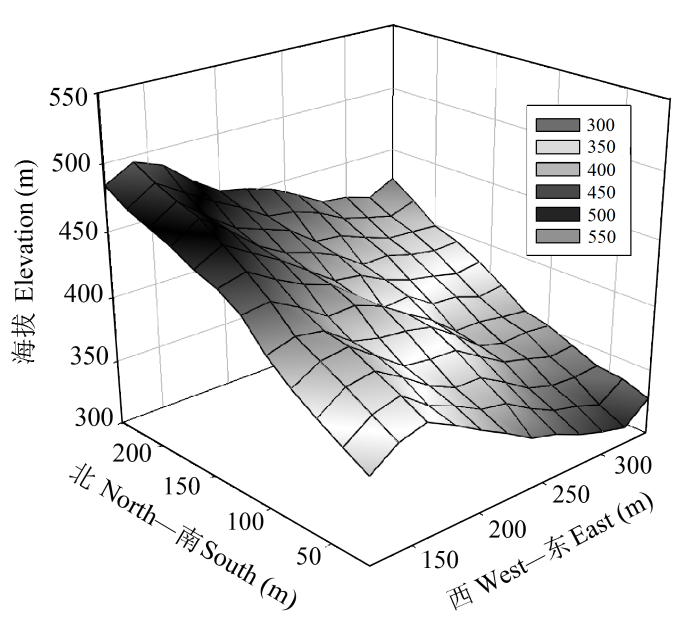
研究区域位于浙江天童国家森林公园(29.80° N, 121.78° E)内, 属于典型的亚热带季风气候, 全年温暖潮湿, 年平均气温16.2 ℃, 年降水量1374.7 mm, 土壤为山地黄红壤, 土层薄厚不一, 约为1 m。地形复杂, 最高海拔为602.89 m (宋永昌和王祥荣, 1995)。地带性植被主要为亚热带常绿阔叶林, 物种组成丰富。选择天童国家森林公园20 hm2大型动态监测样地(29.81°-29.82° N, 121.78°-121.79° E)中的5 hm2区域(相对横坐标为120-340 m, 相对纵坐标为20-240 m, 此外, 四角还有4个样地, 地形图中没有显示)作为研究样地(图1)。样地北高南低, 东西两端各有一条大致呈南北走向的山谷, 中间有两条呈东南-西北走向的山脊, 地形较为复杂。海拔在300-500 m之间, 坡度大多在10°-30°之间。优势种和常见木本植物物种名录见表1。
图1 浙江天童5 hm2研究样地地形图。
Fig. 1 Topography map of the 5 hm2 study plot in Tiantong, Zhejiang Province.
2010-2013年的6-11月, 对5 hm2样地内所有胸径≥1 cm的木本植物个体, 逐株进行叶片的采集(因样本量太大, 历时4年完成数据收集)。取样方案主要参照Cornelissen等(2003)编写的《植物性状测量手册》。具体如下: 在没有明显遮阴的树冠层外部, 用高枝剪剪切1段生长良好的枝条(株高>5 m, 用20 m高枝剪, 树高<5 m, 用3 m高枝剪), 在其上摘取完整、未有损伤的40片叶子, 置于密封袋中, 记录编号, 带回实验室。随后, 在12 h内从密封袋中随机挑选20片成熟叶片, 称其鲜质量, 再用叶面积仪(LI- COR 3100C, LI-COR, Lincoln, USA)测量叶面积。待完成后, 将叶片按编号装入信封, 置于80 ℃干燥箱中烘干48 h至恒定质量, 称量样品干质量。用总的叶面积除以叶片数得到单叶面积(individual leaf area, ILA), 用总叶面积与叶片干质量的比值得到比叶面积(specific leaf area, SLA), 用叶片干质量与叶片鲜质量的比值得到叶片干物质含量(leaf dry matter content, LDMC)。各物种的叶片性状均值见表1。
表1 5 hm2样地木本植物名录和叶片性状
Table 1 Species list and leaf traits of woody plants in the 5 hm2 plot
| 生活型 Life form | 物种 Species | 多度 Abundance | 单叶面积 Individual leaf area (cm2) | 比叶面积 Specific leaf area (cm2·g-1) | 叶片干物质含量 Leaf dry matter content (mg·g-1) |
|---|---|---|---|---|---|
| 常绿乔木 Evergreen trees | 薄叶山矾 Symplocos anomala | 436 | 10.70 ± 0.16 | 133.07 ± 2.06 | 34.30 ± 0.24 |
| 豹皮樟 Litsea coreana | 49 | 12.83 ± 0.54 | 120.38 ± 3.14 | 46.05 ± 1.06 | |
| 赤皮青冈 Cyclobalanopsis gilva | 23 | 17.27 ± 1.07 | 113.25 ± 3.99 | 48.03 ± 0.67 | |
| 刺叶桂樱 Prunus spinulosa | 2 | 10.81 ± 1.18 | 168.21 ± 12.64 | 35.07 ± 2.70 | |
| 大叶冬青 Ilex latifolia | 48 | 58.18 ± 2.74 | 73.81 ± 3.17 | 39.39 ± 0.80 | |
| 冬青 Ilex chinensis | 4 | 16.56 ± 1.52 | 137.96 ± 19.09 | 38.21 ± 1.76 | |
| 杜英 Elaeocarpus decipiens | 1 | 31.07 | 184.93 | 31.88 | |
| 短梗冬青 Ilex buergeri | 95 | 9.56 ± 0.33 | 128.73 ± 3.91 | 42.11 ± 0.59 | |
| 光叶石楠 Photinia glabra | 174 | 14.95 ± 0.33 | 85.49 ± 2.54 | 46.95 ± 0.59 | |
| 光枝刺叶冬青 Ilex hylonoma | 4 | 26.06 ± 3.60 | 113.88 ± 6.89 | 35.00 ± 1.51 | |
| 红楠 Machilus thunbergii | 401 | 18.86 ± 0.37 | 95.36 ± 1.07 | 42.93 ± 0.29 | |
| 虎皮楠 Daphniphyllum oldhamii | 139 | 26.16 ± 0.62 | 121.71 ± 3.31 | 35.09 ± 0.58 | |
| 华东木犀 Osmanthus cooperi | 103 | 21.39 ± 1.20 | 90.46 ± 4.85 | 44.88 ± 1.03 | |
| 华东楠 Machilus leptophylla | 408 | 78.79 ± 1.41 | 141.74 ± 1.58 | 33.94 ± 0.33 | |
| 黄丹木姜子 Litsea elongata | 3 161 | 22.86 ± 0.13 | 123.33 ± 0.52 | 45.27 ± 0.10 | |
| 黄牛奶树 Symplocos cochinchinensis | 149 | 22.08 ± 0.63 | 141.81 ± 6.36 | 32.94 ± 0.38 | |
| 栲树 Castanopsis fargesii | 200 | 17.56 ± 0.44 | 114.02 ± 2.68 | 42.89 ± 0.43 | |
| 苦槠 Castanopsis sclerophylla | 14 | 33.76 ± 3.00 | 109.86 ± 9.13 | 44.61 ± 1.28 | |
| 米槠 Castanopsis carlesii | 69 | 9.97 ± 0.39 | 143.54 ± 6.41 | 43.73 ± 0.84 | |
| 木荷 Schima superba | 277 | 29.74 ± 0.64 | 99.25 ± 1.78 | 41.79 ± 0.43 | |
| 木犀 Osmanthus fragrans | 11 | 33.27 ± 1.84 | 84.25 ± 2.16 | 46.94 ± 0.93 | |
| 披针叶茴香 Illicium lanceolatum | 258 | 16.31 ± 0.42 | 114.59 ± 3.48 | 32.18 ± 0.45 | |
| 朴树 Celtis sinensis | 1 | 9.26 | 308.8 | 27.52 | |
| 青冈 Cyclobalanopsis glauca | 19 | 27.14 ± 2.11 | 121.83 ± 12.62 | 42.68 ± 1.92 | |
| 三尖杉 Cephalotaxus fortunei | 2 | 65.32 ± 40.05 | 82.90 ± 2.29 | 28.18 ± 0.36 | |
| 石栎 Lithocarpus glaber | 13 | 29.05 ± 3.87 | 95.09 ± 3.70 | 49.60 ± 2.17 | |
| 四川山矾 Symplocos setchuensis | 173 | 18.96 ± 0.44 | 86.32 ± 2.07 | 39.95 ± 0.57 | |
| 铁冬青 Ilex rotunda | 44 | 14.38 ± 0.71 | 161.62 ± 8.80 | 30.02 ± 0.91 | |
| 细叶青冈 Cyclobalanopsis gracilis | 23 | 18.34 ± 1.19 | 119.78 ± 2.666 | 44.28 ± 0.84 | |
| 细叶香桂 Cinnamomum subavenium | 219 | 16.41 ± 0.44 | 98.95 ± 1.66 | 47.36 ± 0.42 | |
| 小叶青冈 Cyclobalanopsis myrsinifolia | 140 | 16.58 ± 0.43 | 116.90 ± 2.48 | 45.77 ± 0.54 | |
| 杨梅 Myrica rubra | 24 | 19.67 ± 1.28 | 141.67 ± 7.86 | 41.97 ± 1.90 | |
| 杨梅叶蚊母树 Distylium myricoides | 1 394 | 13.79 ± 0.29 | 90.37 ± 0.86 | 43.34 ± 0.17 | |
| 杨桐 Adinandra millettii | 489 | 20.08 ± 0.32 | 90.39 ± 1.36 | 38.98 ± 0.29 | |
| 云山青冈 Cyclobalanopsis sessilifolia | 241 | 19.74 ± 0.50 | 102.39 ± 2.38 | 43.23 ± 0.41 | |
| 长叶石栎 Lithocarpus henryi | 352 | 56.12 ± 1.22 | 91.20 ± 1.11 | 44.19 ± 0.37 | |
| 浙江新木姜子 Neolitsea aurata | 698 | 14.01 ± 0.21 | 122.51 ± 1.35 | 48.84 ± 0.23 | |
| 中华杜英 Elaeocarpus chinensis | 3 | 16.16 ± 0.92 | 157.27 ± 19.36 | 33.63 ± 4.49 | |
| 皱柄冬青 Ilex kengii | 31 | 10.35 ± 0.41 | 121.31 ± 4.13 | 40.39 ± 0.86 | |
| 紫楠 Phoebe sheareri | 12 | 65.04 ± 10.68 | 138.24 ± 12.14 | 40.86 ± 0.97 | |
| 总状山矾 Symplocos botryantha | 34 | 14.12 ± 0.67 | 127.06 ± 6.50 | 34.66 ± 0.83 | |
| 常绿灌木 Evergreen shrubs | 白花苦灯笼 Tarenna mollissima | 3 | 19.02 ± 2.54 | 276.47 ± 13.12 | 26.68 ± 4.44 |
| 百齿卫矛 Euonymus centidens | 3 | 6.13 ± 0.60 | 192.55 ± 17.67 | 32.96 ± 0.26 | |
| 赤楠 Syzygium buxifolium | 120 | 5.89 ± 0.29 | 122.34 ± 5.48 | 39.31 ± 1.04 | |
| 格药柃 Eurya muricata | 127 | 15.92 ± 0.39 | 92.56 ± 2.5 | 38.23 ± 0.61 | |
| 赤楠 Syzygium buxifolium | 120 | 5.89 ± 0.29 | 122.34 ± 5.48 | 39.31 ± 1.04 | |
| 格药柃 Eurya muricata | 127 | 15.92 ± 0.39 | 92.56 ±2.5 | 38.23 ± 0.61 | |
| 光叶山矾 Symplocos lancifolia | 171 | 9.99 ± 0.29 | 193.55 ± 3.47 | 32.78 ± 0.38 | |
| 红皮树 Stytax suberifolius | 2 | 23.47 ± 3.08 | 164.01 ± 7.79 | 31.80 ± 7.92 | |
| 常绿灌木 Evergreen shrubs | 红山茶 Camellia japonica | 1 | 18.16 | 85.46 | 32.77 |
| 厚皮香 Ternstroemia gymnanthera | 24 | 14.01 ± 2.12 | 102.90 ± 8.49 | 35.33 ± 1.26 | |
| 檵木 Loropetalum chinensis | 37 | 5.83 ± 0.47 | 187.39 ± 11.97 | 37.05 ± 1.17 | |
| 老鼠矢 Symplocos stellaris | 27 | 31.57 ± 2.22 | 81.81 ± 2.86 | 34.79 ± 0.63 | |
| 连蕊茶 Camellia cuspidata | 1 782 | 7.18 ± 0.18 | 130.68 ± 1.89 | 37.65 ± 1.22 | |
| 马银花 Rhododendron ovatum | 337 | 8.95 ± 0.30 | 135.34 ± 2.71 | 39.95 ± 0.39 | |
| 米饭 Vaccinium mandarinorum | 45 | 12.28 ± 0.55 | 138.91 ± 6.65 | 4.41 ± 0.66 | |
| 南天竹 Nandina domestica | 1 | 109.06 | 152.32 | 41.87 | |
| 山矾 Symplocos sumuntia | 316 | 10.71 ± 0.19 | 113.47 ± 1.68 | 34.95 ± 0.28 | |
| 石斑木 Rhaphiolepis indica | 20 | 8.04 ± 0.81 | 121.99 ± 13.52 | 43.23 ± 2.36 | |
| 细枝柃 Eurya loquaiana | 5 688 | 8.74 ± 0.41 | 153.57 ± 0.52 | 38.12 ± 0.07 | |
| 腺点樱 Prunus phaeosticta | 203 | 13.02 ± 0.37 | 147.94 ± 2.26 | 39.05 ± 0.35 | |
| 窄基红褐柃 Eurya rubiginosa | 194 | 13.07 ± 0.33 | 108.61 ± 2.34 | 37.45 ± 0.39 | |
| 落叶乔木 Deciduous trees | 灯台树 Cornus controversa | 4 | 42.25 ± 5.89 | 247.28 ± 26.56 | 26.45 ± 2.89 |
| 红枝柴 Meliosma oldhamii | 1 | 89.44 | 160.75 | 29.73 | |
| 华东野核桃 Juglans cathayensis | 7 | 461.32 ± 114.81 | 155.39 ± 12.90 | 32.86 ± 1.99 | |
| 华东油柿 Diospyros oleifera | 7 | 69.34 ± 8.16 | 236.62 ± 27.88 | 31.22 ± 2.20 | |
| 青皮木 Schoepfia jasminodora | 9 | 21.44 ± 3.85 | 170.58 ± 13.46 | 32.27 ± 1.51 | |
| 山合欢 Albizia kalkora | 2 | 60.92 ± 22.08 | 112.61 ± 14.73 | 36.86 ± 0.98 | |
| 迎春樱 Prunus discoidea | 7 | 22.01 ± 3.74 | 191.59 ± 38.88 | 44.36 ± 5.95 | |
| 浙江柿 Diospyros glaucifolia | 12 | 70.55 ± 8.81 | 195.19 ± 19.52 | 32.07 ± 1.92 | |
| 糙叶树 Aphananthe aspera | 11 | 27.17 ± 5.54 | 213.89 ± 27.17 | 37.17 ± 1.50 | |
| 檫木 Sassafras tzumu | 82 | 71.02 ± 3.27 | 177.23 ± 5.76 | 33.86 ± 0.69 | |
| 臭辣树 Euodia fargesii | 1 | 81.39 | 152.99 | 38.97 | |
| 枫香 Liquidambar formosana | 33 | 43.02 ± 2.78 | 226.51 ± 13.65 | 33.13 ± 1.02 | |
| 杭州榆 Ulmus changii | 4 | 12.72 ± 2.03 | 197.71 ± 26.37 | 32.62 ± 0.63 | |
| 厚壳树 Ehretia acuminata | 3 | 54.95 ± 5.99 | 228.32 ± 21.04 | 27.61 ± 2.67 | |
| 化香 Platycarya strobilacea | 5 | 129.33 ± 12.11 | 206.12 ± 20.67 | 34.89 ± 0.77 | |
| 黄檀 Dalbergia hupeana | 11 | 61.82 ± 6.75 | 229.09 ± 27.29 | 32.14 ± 1.62 | |
| 苦枥木 Fraxinus insularis | 41 | 63.75 ± 5.21 | 186.60 ± 8.99 | 34.16 ± 0.98 | |
| 雷公鹅耳枥 Carpinus viminea | 96 | 20.43 ± 1.97 | 253.86 ± 9.06 | 37.70 ± 0.76 | |
| 毛八角枫 Alangium kurzii | 4 | 53.65 ± 6.56 | 288.76 ± 27.21 | 26.22 ± 1.70 | |
| 毛脉槭 Acer pubinerve | 48 | 41.42 ± 2.81 | 206.96 ± 8.82 | 37.91 ± 1.31 | |
| 南酸枣 Choerospondias axillaris | 263 | 128.98 ± 3.61 | 160.29 ± 4.74 | 34.88 ± 0.31 | |
| 拟赤杨 Alniphyllum fortunei | 117 | 52.20 ± 2.03 | 263.67 ± 9.78 | 28.01 ± 0.69 | |
| 青钱柳 Cyclocarya paliurus | 10 | 87.84 ± 11.97 | 219.88 ± 18.23 | 33.35 ± 1.39 | |
| 锐角槭 Acer acutum | 7 | 127.05 ± 27.05 | 229.32 ± 37.71 | 33.54 ± 2.08 | |
| 山黄皮 Randia cochinchinensis | 2 | 16.67 ± 1.46 | 137.36 ± 21.11 | 44.72 ± 0.93 | |
| 山鸡椒 Litsea cubeba | 15 | 20.01 ± 1.30 | 298.63 ± 22.63 | 28.01 ± 1.37 | |
| 山桐子 Idesia polycarpa | 7 | 71.93 ± 11.57 | 282.54 ± 35.72 | 37.51 ± 3.89 | |
| 无患子 Sapindus saponaria | 18 | 298.93 ± 29.12 | 233.64 ± 12.56 | 32.85 ± 1.12 | |
| 西川朴 Celtis vandervoetiana | 16 | 45.58 ± 4.33 | 221.86 ± 20.72 | 37.55 ± 1.76 | |
| 小果冬青 Ilex micrococca | 2 | 20.53 ± 9.25 | 133.61 ± 28.22 | 37.09 ± 3.07 | |
| 野漆树 Toxicodendron succedaneum | 21 | 147.20 ± 11.24 | 152.28 ± 6.47 | 36.37 ± 1.47 | |
| 油桐 Vernicia fordii | 75 | 238.53 ± 17.24 | 182.18 ± 7.19 | 32.94 ± 0.94 | |
| 紫弹树 Celtis biondii | 6 | 12.86 ± 1.21 | 201.41 ± 35.63 | 33.26 ± 1.60 | |
| 落叶灌木 Deciduous shrubs | 豆腐柴 Premna microphylla | 4 | 12.61 ± 2.20 | 304.82 ± 26.20 | 24.80 ± 1.87 |
| 山胡椒 Lindera glauca | 4 | 10.98 ± 0.64 | 240.99 ± 46.23 | 39.09 ± 2.40 | |
| 乌饭 Vaccinium bracteatum | 13 | 10.58 ± 0.93 | 150.69 ± 4.60 | 30.53 ± 0.89 | |
| 白背叶 Machilus apeltus | 24 | 113.97 ± 12.50 | 247.95 ± 15.19 | 30.83 ± 1.15 | |
| 落叶灌木 Deciduous shrubs | 刺毛越橘 Vaccinium trichocladum | 14 | 8.97 ± 0.51 | 129.16 ± 8.89 | 32.73 ± 1.37 |
| 大青 Clerodendrum cyrtophyllum | 60 | 66.10 ± 4.59 | 240.95 ± 13.38 | 28.39 ± 1.01 | |
| 大叶白纸扇 Mussaenda shikokiana | 5 | 105.86 ± 11.95 | 361.34 ± 82.37 | 24.38 ± 3.80 | |
| 杜鹃花 Rhododendron simsii | 4 | 14.07 ± 3.51 | 383.14 ± 90.13 | 26.87 ± 3.22 | |
| 红脉钓樟 Lindera rubronervia | 35 | 15.76 ± 0.87 | 307.86 ± 14.30 | 30.66 ± 0.85 | |
| 赛山梅 Styrax confusus | 90 | 17.86 ± 0.72 | 235.73 ± 7.89 | 29.22 ± 0.48 | |
| 山油麻 Trema cannabina | 3 | 11.45 ± 3.38 | 459.79 ± 68.34 | 27.68 ± 1.30 | |
| 天仙果 Ficus erecta | 2 | 64.54 ± 29.14 | 407.11 ± 11.82 | 23.31 ± 2.03 | |
| 小叶石楠 Photinia parvifolia | 2 | 11.45 ± 1.26 | 312.44 ± 70.23 | 37.82 ± 5.21 | |
| 宜昌荚蒾 Viburnum erosum | 5 | 15.25 ± 1.68 | 350.13 ± 30.99 | 23.78 ± 1.60 | |
| 枳椇 Hovenia acerba | 2 | 103.89 ± 44.38 | 170.60 ± 63.22 | 13.03 ± 9.21 |
本文环境因子包括土壤养分和地形因子, 数据来源于天童大型动态样地的观测数据库。即: 在大样地建立时, 将样地分割成20 m × 20 m的小样方, 在每个样方内测定了土壤的总磷(TP)、总氮(TN)、总碳(TC)、pH值、土壤体积含水率(SVWC)、容重(BD)和腐殖质(HD)、海拔(E)、坡度(S)、凹凸度(C)。具体土壤采样参照CTFS土壤采样方案, 详细见张娜等(2012)。
地统计学能有效地揭示变量在空间上的分布、变异和相关特征, 还能将空间格局与生态过程联系起来(马风云等, 2006; 张峰等, 2012), 近年来在研究环境与生态特征的空间格局中应用广泛(赵斌和蔡庆华, 2000; 王晓春等, 2002; 刘璐等, 2010; 张娜等, 2012)。本文采用地统计学分析的半变异函数探索植物叶片性状的空间变异性, 用最优模型的块金值、基台值、结构比、变程和分维数表征叶片性状的空间变异程度, 具体为: 基台值是当采样点间的距离h增大时, 半变异函数r(h)从初始的块金值达到一个相对稳定的常数。基台值越大表明总的空间异质性程度越高; 变程是当半变异函数的取值由初始的块金值达到基台值时采样点间的距离, 反映了区域化变量影响的范围大小; 块金值表示随机部分的空间变异性, 较大的块金方差表明较小尺度上的某种过程不可忽视; 结构比反映块金方差占总空间变异的大小。一般认为, 结构比<25%时, 变量具有强烈的空间自相关; 结构比在25%-75%时, 变量间具有中等程度的空间自相关; 结构比>75%时, 空间自相关很弱; 分形维数D是空间相关性程度的另一个重要参数, 其值趋近于2, 空间分布越均质, 空间变异性越小; 其值越远离2, 空间变异性越大。分形维数由公式2r(h) = h(4-2D)得到。而区域化变量的半变异函数r(h)不仅与空间距离h有关, 而且与方向有关。如果在某一范围内半变异函数上升, 说明半变异函数具有方向性。因此, 半变异函数还可以用来分析空间变量的方向变异性。
本研究选择5 hm2样地内所有个体的单叶面积、比叶面积和叶片干物质含量, 用ArcGIS的Geostatis- tical analyst模块得到叶片性状的半变异函数值随样本滞后距变化的散点图, 并用球型、指数、高斯模型拟合, 依据标准均方根预测误差(RMSS)越接近于1模型越好的原则, 得出半变异函数的最优模型。再依据最优模型的块金值、基台值、结构比、变程和分维数等参数判断单叶面积、比叶面积和叶片干物质含量的空间变异程度。
为了探索植物叶片性状的方向变异性。在ArcGIS的Geostatistical analyst模块中做出东-西、东北-西南、南-北和西北-东南4个方向的半变异函数模型, 以及反映各个方向上空间变异程度的块金值、基台值、结构比、变程和分维数。基于这些参数, 得出叶片性状空间变异最大和最小的方向。
为探究环境过滤对叶片性状空间变异的影响程度, 利用Kriging插值法将5 hm2样地中的土壤养分和地形因子插值, 借助ArcGIS分析工具中的提取工具得到每株个体所对应的土壤养分和地形数据。最后, 基于5 hm2样地内每木个体的单叶面积、比叶面积、叶片干物质含量, 以及所对应的土壤养分和地形因子, 利用主成分分析和Pearson相关分析法分析土壤养分、地形因子对叶片性状空间变异性的影响。
此外, 为了探究不同方向上环境因子对叶片性状变异性的影响, 选取地形和土壤养分差异最大的东北-西南方向(沿山脊和沟谷)和西北-东南方向(垂直于山脊和山谷)。分别提取东北-西南和西北-东南方向上的叶片性状、土壤养分和地形因子, 再用主成分分析和Pearson相关分析分析土壤养分、地形因子对叶片性状方向变异性的影响。
叶片性状空间变异性分析在ArcGIS 10.0的Geostatistical Analyst模块中实现, 主成分分析和Pearson相关分析均在R 3.0.2中完成, 绘图在Origin 8.5中完成。
叶片性状的半变异函数模型显示, 单叶面积、比叶面积和叶片干物质含量均存在较大的空间变异性(表2)。高斯模型对单叶面积拟合最优(决定系数(R2) = 0.95, 标准均方根预测误差=1.41), 指数模型对比叶面积(R2 = 0.88, 标准均方根预测误差= 0.90)、叶片干物质含量(R2 = 0.96, 标准均方根预测误差=1.29)拟合最优。单叶面积的块金值(0.0032)最大, 比叶面积次之(0.0006), 叶片干物质含量最小(0.0003)。单叶面积、比叶面积和叶片干物质含量的基台值分别为0.090、0.012和0.007, 均表明单叶面积的空间变异最大, 比叶面积次之, 叶片干物质含量最小。
表2 植物叶片性状的半变异函数模型
Table 2 Semi-variogram model of plant leaf traits
| 叶片性状 Leaf trait | 模型 Model | 块金值 Nugget | 基台值 Still | 结构比 Structure ratio | 变程 Range (m) | 决定系数 Determination coefficient | 标准均方根预测误差 Root mean square standardized |
|---|---|---|---|---|---|---|---|
| 单叶面积 Individual leaf area | 球型 Spherical model | 0.000 1 | 0.090 | 0.999 | 6.07 | 0.95 | 1.43 |
| 指数 Exponential model | 0.000 1 | 0.090 | 0.999 | 4.35 | 0.95 | 2.09 | |
| 高斯 Gaussian model | 0.003 2 | 0.090 | 0.964 | 4.80 | 0.95 | 1.41 | |
| 比叶面积 Specific leaf area | 球型 Spherical model | 0.000 01 | 0.012 | 0.999 | 6.38 | 0.87 | 0.86 |
| 指数 Exponential model | 0.000 6 | 0.020 | 0.969 | 5.16 | 0.88 | 0.90 | |
| 高斯 Gaussian model | 0.001 6 | 0.020 | 0.918 | 5.23 | 0.87 | 0.76 | |
| 叶片干物质含量 Leaf dry matter content | 球型 Spherical model | 0.000 01 | 0.007 | 0.999 | 6.27 | 0.96 | 1.29 |
| 指数 Exponential model | 0.000 3 | 0.007 | 0.955 | 4.92 | 0.96 | 1.29 | |
| 高斯 Gaussian model | 0.000 7 | 0.007 | 0.899 | 5.11 | 0.96 | 1.32 |
此外, 单叶面积、比叶面积、叶片干物质含量的结构比均大于75%, 表明它们的空间自相关性很弱。变程参数显示, 单叶面积、比叶面积和叶片干物质含量分别在空间范围为4.80 m、5.16 m、4.92 m以下存在很弱的空间自相关。
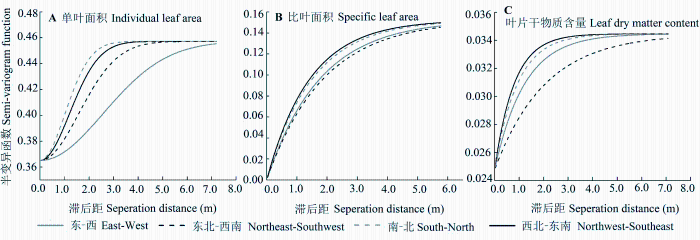
单叶面积、比叶面积、叶片干物质含量在东-西、东北-西南、南-北、西北-东南方向上半变异函数及参数见图2和表3。各方向上, 单叶面积的半变异函数值均高于比叶面积和叶片干物质含量, 这进一步说明了单叶面积的空间变异性大于比叶面积和叶片干物质含量。此外, 4个方向上单叶面积、比叶面积、叶片干物质含量的半变异函数均在变程为6 m和8 m的范围内上升(图2), 这说明单叶面积、比叶面积和叶片干物质含量的空间变异均具有方向性。
表3 植物叶片性状的各向异性参数
Table 3 Anisotropy parameters of plant leaf traits
| 叶片性状 Leaf traits | 东-西 East-West | 东北-西南 Northeast-Southwest | 南-北 South-North | 西北-东南 Northwest-Southeast | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| D | R2 | D | R2 | D | R2 | D | R2 | ||||
| 单叶面积 Individual leaf area | 1.96 | 0.95 | 1.93 | 0.97 | 1.98 | 0.97 | 1.99 | 0.51 | |||
| 比叶面积 Specific leaf area | 1.99 | 0.66 | 1.99 | 0.54 | 1.99 | 0.75 | 1.98 | 0.67 | |||
| 叶片干物质含量 Leaf dry matter content | 1.99 | 0.70 | 1.99 | 0.26 | 1.99 | 0.49 | 1.98 | 0.70 | |||
单叶面积在东北-西南方向上, 空间变异性最大, 单叶面积在此方向上半变异函数最大且最先达到基台值, 变化幅度大且在其周围变化迅速, 分形维数(D = 1.93)最远离2, 决定系数(R2 = 0.97)最大; 相反, 比叶面积和叶片干物质含量的半变异函数在西北-东南方向最大, 且最先达到基台值, 变化幅度大, 分形维数(D = 1.98; D = 1.98 )最远离2, 决定系数(R2 = 0.70; R2 = 0.67)为最大, 因此, 比叶面积和叶片干物质含量在西北-东南方向上空间变异性最大。
环境因子的主成分分析结果如表4所示, 第一和第二主成分累计所占环境因子总变异的70.0%, 因此, 选取前两个主成分度量整个环境信息。第一主成分(PC1)主要由土壤养分组成, 大多数土壤养分有正的载荷, 如总碳(0.35)、体积含水率(0.43)、腐殖质(0.29), 以及负载荷如pH值(-0.36)、容重(-0.40)。第二主成分(PC2)主要是地形因子, 有高的海拔(0.32)、坡度(0.42)、凹凸度(0.32)。
叶片性状与PC1 (土壤养分)、PC2 (地形因子)的相关性分析结果见表5。单叶面积与PC2显著负相关(r = -0.12, p < 0.0001); 比叶面积与PC1显著负相关(r = -0.16, p < 0.0001), 叶片干物质含量与PC1显著正相关(r = 0.13, p < 0.0001)。
在东北-西南和西北-东南方向, 叶片性状与PC1和PC2的相关性见表6。在东北-西南方向上, 叶片性状主要与PC2相关, 但相关性很小。单叶面积与PC2显著负相关(r = -0.07, p = 0.003); 比叶面积与PC2显著负相关(r = -0.08, p = 0.001); 叶片干物质含量与PC2显著正相关(r = 0.11, p = 0.001)。西北-东南方向上, 单叶面积与PC2显著负相关(r = -0.13, p < 0.0001); 比叶面积与PC1显著负相关(r = -0.25, p < 0.0001); 叶片干物质含量与PC1显著负相关(r = -0.22, p < 0.0001)。
整个空间上, 单叶面积的空间变异受地形因子的影响较大, 比叶面积和叶片干物质含量的空间变异受土壤养分的影响较大。在东北-西南方向和西北-东南方向上, 地形差异是单叶面积空间变异的主要因素。而比叶面积和叶片干物质含量的空间变异, 在东北-西南方向上地形因子是主要的影响因素, 在西北-东南方向上, 土壤养分是主要的影响因素。
在环境筛选作用下, 生活在同一生境的物种在性状上保持一定的相似性; 同时, 由于物种间的极限相似性作用, 使得同一生境的物种在性状和功能上保持一定的差异性, 以便达到资源的最优配置和利用。因而在局域尺度上, 物种性状变异较大。张莉等(2013)研究了延河流域植物叶片性状空间变异, 指出比叶面积、叶片氮等均存在很大的空间变异。本文研究的亚热带常绿阔叶林物种丰富, 叶片性状变异大, 单叶面积的变异范围为0.57-920.20 cm2, 变异系数达到了153.35%; 比叶面积在8.59-976.55 cm2·g-1之间, 变异系数为39.67%; 干物质含量值在3.80-89.97 mg·g-1之间, 变异系数为18.50%。整个局域空间尺度上, 单叶面积有很大的空间变异, 其原因主要是研究区域物种丰富, 有叶面积最小的檵木(Loropetalum chinensis)、赤楠(Syzygium buxifolium)等, 也有叶面积最大的华东野核桃(Juglans cathayensis)、无患子(Sapindus mukorossi)等(表1)。地统计学分析表明, 植物叶片性状存在很弱的空间自相关和空间聚集性, 单叶面积、比叶面积、叶片干物质含量分别在4.8 m、5.2 m、4.9 m空间范围以下存在很弱的空间自相关和呈现聚集分布。即: 物种在空间距离为5 m以下受到物种间的相互作用(如竞争排斥)较大, 因而性状差异也较大。整体上, 叶片性状都存在空间变异性, 单叶面积的空间变异最大, 比叶面积次之, 叶片干物质含量最小。单叶面积的空间变异最大主要反映了单叶面积是由物种遗传决定的, 物种的多样性决定了其空间变异的大小。样地内共有108个物种。同时, 样地内环境的高度异质性影响了物种的分布格局, 进而使得单叶面积的空间变异性较大。而比叶面积和叶片干物质含量是质量和面积的综合值, 其受环境的筛选作用较弱, 因而, 在同区域中比叶面积和叶片干物质含量空间变异较小。
各向异性函数得出植物叶片性状存在方向变异性, 表明不同方向上环境差异对植物叶片性状的空间变异影响较大。空间尺度较大的生境中, 环境的异质性塑造了叶片性状的空间变异, 如水分、养分充足的沟谷叶片面积、比叶面积大, 叶片干物质含量小; 贫瘠的山脊上, 土壤养分、水分不足, 以及受光照的影响, 往往分布着叶片面积和比叶面积较小、叶片干物质含量较大的阳性植物。本研究认为地形和土壤养分的差异是叶片性状空间变异的主要环境因子。研究区域主要山脊和山谷走向为西北-东南方向, 此方向及其垂直方向上地形和土壤养分差异最大。此外, 研究区阳坡主要分布在西北-东南方向, 而坡向影响阳光、水分、温度、土壤养分的分布, 进而影响叶片性状, 使得在西北-东南及其垂直方向东北-西南方向叶片性状的空间变异差异最大。
单叶面积间接地反映了叶片的大小, 其对叶片能量和水分平衡有重要意义。总体看, 单叶面积的方向异质性差异最大(图2), 因西北-东南方向及其垂直方向西北-东南地形的差异导致物种的分布差异较大。比叶面积是一定干物质投资所展开的补光表面数量(Wright et al., 2004), 其与植物的光捕获效率有关(Milla & Reich, 2007)。叶片干物质含量反映植物生态行为差异及获取资源能力(Cornelissen et al., 2003), 是植物适应环境所体现出的关键叶性状(张林等, 2008)。比叶面积和叶片干物质含量有相同的最大、最小空间变异方向且与单叶面积相反, 因为比叶面积和叶片干物质含量综合反映了植物对光照、土壤养分的利用效率以及获取碳的策略。而它们本身有显著的负相关关系(r = -0.49, p < 0.01)。西北-东南方向是山脊和山谷方向, 地形复杂, 也是海拔梯度最大的方向, 此方向上土壤养分以及光照梯度差异大, 因而此方向上比叶面积和叶片干物质含量的空间变异最大, 而在其垂直方向——东北-西南方向, 土壤养分分布和获取光照比较均一, 比叶面积和叶片干物质含量的空间变异较小。植物叶片方向变异性的确定, 可以为不同方向的沟谷、山脊物种适应性策略提供理论依据。
植物生长发育不仅取决于自身的生理特性, 而且受到环境梯度的影响, 群落的本质特征之一是群落中的植物和环境因子之间存在一定的相互关系(宋永昌, 2001), 同时, 物种的功能策略能够表征该区域的环境条件, 通过分析环境因子梯度分布可以预测植物的功能策略(宝乐和刘艳红, 2009; Aiba et al., 2012)。
植物性状与环境梯度的关系在大尺度上已经通过大量植物性状集被量化(Wright et al., 2004), 研究表明在大尺度上植物性状与环境有显著的相关性, 植物通过环境的筛选作用塑造其性状, 对环境做出一定的适应策略。叶片性状与资源吸收有关, 如比叶面积、叶片干物质含量与土壤养分含量和地形有关(Ordoñez et al., 2009)。Liu等(2012)得出土壤养分与植物叶片性状在局域尺度上显著相关。本研究中, 从叶片性状与土壤养分和地形因子的相关性(表7)看, 单叶面积与土壤总磷、总氮、总碳、pH值、容重有显著的正相关关系(p < 0.05), 与土壤体积含水率、海拔、坡度、凹凸度有显著的负相关关系(p < 0.05); 比叶面积与土壤总磷、pH值、容重有显著的正相关关系(p < 0.05), 与土壤总氮、总磷、总碳、体积含水率、腐殖质, 海拔、坡度、凹凸度有显著的负相关关系(p < 0.05); 叶片干物质含量与土壤总氮、总碳、体积含水率、腐殖质, 海拔、坡度、凹凸度有显著的正相关关系(p < 0.05), 与土壤pH值、容重有显著的负相关关系(p < 0.05)。总体上, 单个环境因子与叶片性状的相关性显著, 但相关性很小。因此本文对所有环境因子进行主成分分析, 综合分析环境过滤对植物性状的作用。Liu等(2012)预测了性状值高、资源吸收快的植物, 生长在养分梯度丰富的环境中, 经济型谱较低的物种资源吸收率较低、生长环境养分较贫瘠。本研究中PC1主要是土壤养分变量载荷, PC2主要由地形信息组成。由植物叶片性状与PC1、PC2的相关性分析得出, 单叶面积与地形因子有较大的相关性, 比叶面积和叶片干物质含量与土壤养分有较大的相关性。
在东北-西南方向和西北-东南方向上同样得出单叶面积与地形因子相关性较大, 比叶面积和叶片干物质含量与土壤养分相关性较大。东北-西南方向上, 地形空间差异较大, 叶片性状主要与地形因子相关但相关性较小; 西北-东南方向上, 土壤养分的空间差异较大, 比叶面积和叶片干物质含量与土壤养分有较大的显著相关性。在东北-西南方向上地形因子对叶片性状的空间变异影响相对大于土壤养分。在西北-东南方向上土壤养分对叶片性状的空间变异起主要作用。总体上, 单叶面积受地形因子影响较大, 比叶面积和叶片干物质含量受土壤养分的影响较大, 反映在相关性上, 即单叶面积与地形因子的相关系数的绝对值大于其与土壤养分的相关系数的绝对值; 比叶面积、叶片干物质含量和土壤养分的相关系数的绝对值大于其与地形因子的相关系数的绝对值。相关研究也得出同样的结果(Liu et al., 2012)。
整体上, 在局域尺度上环境异质性对植物叶片性状的空间变异的解释度较小, 虽然土壤养分和地形变量在一定程度上可以解释部分叶片性状的空间变异, 但在局域尺度上还有很大一部分空间变异未被解释。相关研究表明: 在局域尺度上, 生物间的相互作用(极限相似性)对物种性状的塑造起主要作用(Herben & Goldberg, 2014), 其主要通过竞争使局域空间的性状趋异达到资源的合理分配与利用。因此, 未来有必要在局域环境中, 探究物种间的相互作用(竞争排斥)对叶片性状空间变异的影响。
致谢 感谢浙江省宁波市重大科技攻关项目(2012- C10027)资助。感谢华东师范大学郑泽梅老师提供土壤养分和地形数据; 感谢华东师范大学郭明、仲强、康蒙、许月、谢一鸣、徐艺露、张志浩、黄海侠、孙宝伟、马文济、黄永贞、王怀耀、刘恋、肖梦馨、欧阳奥等同学在野外收集数据和室内分析中给予的帮助。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
| [21] |
Comparison of leaf construction costs in woody species with differing leaf life-spans in contrasting ecosystems. |
| [22] |
Let the concept of trait be functional! |
| [23] |
a). Plant traits and environmental conditions along the northeast China transect. Ekológia (
|
| [24] |
b). Responses of plant functional types to an environmental gradient on the northeast China transect.
|
| [25] |
Geostatistical analysis of the pattern of Betula ermanii population in Changbai Mountain. 长白山岳桦种群格局的地统计学分析. 应用生态学报, |
| [26] |
Relative abundance and evenness patterns along diversity and biomass gradients. |
| [27] |
|
| [28] |
The worldwide leaf economics spectrum.
|
| [29] |
Spatial distribution of forest carbon in Zhejiang Province with geostatistics based on CFI sample plots. 基于地统计学和CFI样地的浙江省森林碳空间分布研究. 生态学报, |
| [30] |
Vertical variations in specific leaf area and leaf dry matter content with canopy height in Pinus yunnanensis. 云南松比叶面积和叶干物质含量随冠层高度的垂直变化规律. 北京林业大学学报, |
| [31] |
Source of variation of plant functional traits in the Yanhe river watershed: The influence of environment and phylogenetic background. 延河流域植物功能性状变异来源分析. 生态学报, |
| [32] |
Spatial heterogeneity of soil properties and its relationships with terrain factors in broadleaved forest in Tiantong of Zhejiang Province, East China. 浙江天童常绿阔叶林土壤的空间异质性及其与地形的关系. 应用生态学报, |
| [33] |
An application of geostatistical analysis in fresh water ecosystem. |
| [1] |
Interspecific differences in determinants of plant species distribution and the relationships with functional traits. |
| [2] |
Comparison of leaf functional traits in different forest communities in Mt. Dongling of Beijing. 东灵山地区不同森林群落叶功能性状比较 . |
| [3] |
Vertical gradients in leaf trait diversity in a New Zealand forest. |
| [4] |
A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. |
| [5] |
A trait-based test for habitat filtering: Convex hull volume. |
| [6] |
Covariation in leaf and root traits for native and non-native grasses along an altitudinal gradient in New Zealand. |
| [7] |
The plant traits that drive ecosystems: Evidence from three continents. |
| [8] |
Effects of topographic variations and soil characteristics on plant functional traits in a subtropical evergreen broad-leaved forest. 地形和土壤特性对亚热带常绿阔叶林内植物功能性状的影响 . |
| [9] |
|
| [10] |
Community assembly by limiting similarity vs. competitive hierarchies: Testing the consequences of dispersion of individual traits. |
| [11] |
Growth and leaf traits off our broad-leaved tree species along a hillside gradient. |
| [12] |
Spatial heterogeneity of soil nutrients in karst area’s Mulun National Nature Reserve. 喀斯特木论自然保护区土壤养分的空间变异特征. 应用生态学报, |
| [13] |
Covariation in plant functional traits and soil fertility within two species- rich forests. |
| [14] |
Spatial heterogeneity of soil moisture in Shapotou sand-fixing artificial vegetation area. 沙坡头人工固沙植被土壤水分空间异质性. 应用生态学报, |
| [15] |
Plant functional traits, environments and ecosystem functioning. Journal of Plant Ecology ( 植物功能性状与环境和生态系统功能. 植物生态学报, |
| [16] |
The scaling of leaf area and mass: The cost of light interception increases with leaf size. |
| [17] |
A global study of relationships between leaf traits, climate and soil measures of nutrient fertility. |
| [18] |
The evolution of plant functional variation: Traits, spectra, and strategies. |
| [19] |
|
| [20] |
|
| [33] |
地统计学分析方法在水生态系统研究中的应用. 水生生物学报, |

/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}