Effects of elevated temperature on soil respiration in a coastal wetland during the non- growing season in the Yellow River Delta, China
SUN Bao-Yu1, 2, HAN Guang-Xuan1, *, , CHEN Liang3, CHU Xiao-Jing1, 2, XING Qing-Hui1, 2, WU Li-Xin4, ZHU Shu-Yu4
1Key Laboratory of Coastal Environmental Processes and Ecological Remediation, Chinese Academy of Sciences, Yantai Institute of Coastal Zone Research, Chinese Academy of Sciences, Yantai, Shandong 264003, China2University of Chinese Academy of Sciences, Beijing 100049, China3College of Environment and Planning, Liaocheng University, Liaocheng, Shandong 252059, Chinaand 4Administration Bureau of the Yellow River Delta National Nature Reserve, Dongying, Shandong 257091, China
通讯作者: * 通信作者Author for correspondence (E-mail: gxhan@yic.ac.cn)
Aims Winter soil respiration plays a crucial role in terrestrial carbon cycle, which could lose carbon gained in the growing season. With global warming, the average near-surface air temperatures will rise by 0.3 to 4.8 °C. Winter is expected to be warmer obviously than other seasons. Thus, the elevated temperature can significantly affect soil respiration. The coastal wetland has shallow underground water level and is affected by the fresh water and salt water. Elevated temperature can cause the increase of soil salinity, and as a result high salinity can limit soil respiration. Our objectives were to determine the diurnal and seasonal dynamics of soil respiration in a coastal wetland during the non-growing season, and to explore the responses of soil respiration to environmental factors, especially soil temperature and salinity. Methods A manipulative warming experiment was conducted in a costal wetland in the Yellow River Delta using the infrared heaters. A complete random block design with two treatments, including control and warming, and each treatment was replicated each treatment four times. Soil respiration was measured twice a month during the non-growing season by a LI-8100 soil CO2 efflux system. The measurements were taken every 2 h for 24 h at clear days. During each soil respiration measurement, soil environmental parameters were determined simultaneously, including soil temperature, moisture and salinity. Important findings The diurnal variation of soil respiration in the warming plots was closely coupled with that in the control plots, and both exhibited single-peak curves. The daily soil respiration in the warming was higher than that in the control from November 2014 to January 2015. Contrarily, from March to April 2015. During the non-growing seasons, there were no significant differences in the daily mean soil respiration between the two treatments. However, soil temperature and soil salt content in the warming plots were significantly higher than those in the control plots. The non-growing season was divided into the no salt restriction period (November 2014 to middle February 2015) and salt restriction period (middle February 2015 to April 2015). During non-growing season, soil respiration in the warming had no significant difference compared with that in control. During the no salt restriction period, soil respiration in the warming was 22.9% (p < 0.01) greater than the control when soil temperature at 10 cm depth in warming was elevated by 4.0 °C compared with that in control. However, experimental warming decreased temperature sensitivity of soil respiration (Q10). During salt restriction period, soil warming decreased soil respiration by 20.7% compared with the control although with higher temperature (3.3 °C), which may be attributed to the increased soil salt content (Soil electric conductivity increased from 4.4 ds·m-1 to 5.3 ds·m-1). The high water content can limit soil respiration in some extent. In addition, the Q10 value in the warming had no significant difference compared with that in control during this period. Therefore, soil warming can not only increase soil respiration by elevating soil temperature, but also decrease soil respiration by increasing soil salt content due to evaporation, which consequently regulating the soil carbon balance of coastal wetlands.
Keywords:elevated temperature
;
soil respiration
;
non-growing season
;
soil salt content
;
coastal wetland
;
Yellow River Delta
SUNBao-Yu, HANGuang-Xuan, CHENLiang, CHUXiao-Jing, XINGQing-Hui, WULi-Xin, ZHUShu-Yu. Effects of elevated temperature on soil respiration in a coastal wetland during the non- growing season in the Yellow River Delta, China. Chinese Journal of Plant Ecology, 2016, 40(11): 1111-1123 https://doi.org/10.17521/cjpe.2015.0414
Fig. 2 Average diurnal variation of soil respiration rate (A), soil temperature (B), soil water content (C), and soil salt content (D) at 10 cm depth under warming and control treatments (mean ± SE).
Fig. 3 Seasonal variation of soil respiration rate (A), soil temperature (B), soil water content (C), and soil electric conductivity (D) at 10 cm depth under warming and control treatments (White expresses non-salt restriction period, gray expresses salt restriction period).
图4 增温对非盐分限制阶段、盐分限制阶段和整个非生长季土壤呼吸速率(A)、10 cm土壤温度(B)、土壤含水量(C)、土壤电导率(D)的影响(平均值±标准误差)。*, p < 0.05; **, p < 0.01。
Fig. 4 Soil respiration rate (A), soil temperature (B), soil water content (C), and soil electric conductivity (D) at 10 cm depth of non- salt restriction period, salt restriction period and non-growing season under warming and control treatments (mean ± SE).
IPCC (Intergovernmental Panel on Climate Change) (2013). Contribution of working group 1 to the fifth assessment report of the intergovernmental panel on climate change. In: Stocker TF, Qin DH, Plattner GK, Tignor M, Allen SK, Boschung J, Nauels A, Xia Y, Bex V, Midgley PM eds. Climate Change in 2013: The Physical Science Basis. Cambridge University Press, Cambridge, UK.
Response of Soil Respiration and Microbial Community Structure to Soil Warming and Throughfall Exclusion in Warm-temperature Oak (Quercus aliena var. acuteserrata) Forest
. PhD dissertation, Chinese Academy of Forestry Sciences, Beijing. (in Chinese with English abstract)[刘彦春 (2013).
Climatic influences on net ecosystem CO2 exchange during the transition from wintertime carbon source to springtime carbon sink in a high elevation, subalpine forest
Heterotrophic microbial activities and nutritional status of microbial communities in tropical marsh sediments of different salinities: The effects of phosphorus addition and plant species
Capacity of fatty acid profiles and substrate utilization patterns to describe differences in soil microbial communities associated with increased salinity or alkalinity at three locations in South Australia
The short-term effects of salinization on anaerobic nutrient cycling and microbial community structure in sediment from a freshwater wetland
1
2006
... 盐分影响着微生物异养呼吸作用, 从而改变土壤呼吸速率.一方面, 盐分抑制土壤微生物活性, 使得土壤呼吸随盐度的升高而降低(Wichern et al., 2006; Wong et al., 2008; Iwai et al., 2012; 李凤霞等, 2012); 同时, 高的含盐量会降低微生物数量(Garcia & Hernandez, 1996; Pattnaik et al., 2000; Pivnickova et al., 2010; Kiehn et al., 2013)以及微生物群落的多样性(Baldwin et al., 2006); 并且, 高盐度能使微生物生理形态发生明显变化, 从而使其分解有机质的能力受到强烈影响(Thottathil et al., 2008), 微生物受到盐分的影响时, 一般能通过自身的渗透压调节机制来平衡细胞内的渗透压, 而在高的渗透压条件下, 微生物耗氧速率增加, 但耗氧速率的增加不是为了有机物的降解, 而是为了能够抵御高盐环境对微生物所产生的伤害(Pankhurst et al., 2001; Rietz & Haynes, 2003; 崔有为等, 2004; Thottathil et al., 2008).另一方面, 多数土壤水解酶与氧化还原酶类活性均随盐渍化的水平升高而明显下降(Rietz & Haynes, 2003; 张建锋等, 2005), 低盐分对酶的活性有促进作用, 而高盐分条件下其活性反而下降(Yan et al., 2013). ...
Experimental warming and burn severity alter soil CO2 flux and soil functional groups in a recently burned boreal forest
2
2004
... 野外自然条件下的增温实验是研究全球变化重要的信息来源(徐振峰等, 2010), 是研究全球变暖与陆地生态系统相互关系的一个主要方法(牛书丽等, 2007; Hoeppner & Dukes, 2012; Hou et al., 2013), 不仅能够获取气候变暖对生态系统影响的直接证据, 还可以解释陆地生态系统对气候变化响应的内在机制(Hou et al., 2013).目前, 有关全球变暖对陆地生态系统影响的增温装置主要有被动增温和主动增温两类.其中, 红外辐射器能更真实有效地模拟全球变暖, 对土壤无物理干扰, 也不改变小气候状况, 是现有的模拟增温的理想装置, 近年来被广泛应用于森林、草地、苔原等生态系统(牛书丽等, 2007).增温影响微生物或根系的代谢活性从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004), 大多数研究表明增温明显促进了土壤呼吸(Lin et al., 2001; Wan et al., 2007; Zhou et al., 2007; Xia et al., 2009; Wang et al., 2014).然而, 也有研究表明随着温度的升高, 土壤呼吸速率出现降低的趋势(Pajari, 1995; Saleska et al., 1999; Melillo et al., 2002; Yin et al., 2013)或不变(Wan et al., 2007), 这可能是因为土壤有限的活性碳库(Oechel et al., 2000; Giardina & Ryan, 2000; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005)和土壤呼吸温度适应性的存在 (Luo et al., 2001). ...
... 众多学者认为是由于温度升高影响了微生物的代谢活性, 从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004).冬季植物根系呼吸显著减少, 土壤呼吸主要是微生物的呼吸(Schindlbacher et al., 2007; Ruehr & Buchmann, 2010).增温一方面会增加土壤无机氮库(Luo et al., 2001; Rustad et al., 2001; Melillo et al., 2002), 而无机氮库的增加在一定程度上弥补了冬季活性养分的缺失, 从而促进了微生物的生长; 另一方面, 增温能够提高土壤微生物和酶的活性(Bokhorst et al., 2010), 从而提高土壤中CO2的释放速率.有研究发现, 冬季的土壤呼吸平均能释放掉生态系统在生长季所固存碳的50% (Brooks et al., 2004), 在此情况下, 温度的升高将进一步导致土壤向大气排放更多CO2.因此, 气候变化引起的冬季土壤呼吸速率的升高可能会显著降低高海拔、高纬度生态系统的碳储量(Melillo et al., 2002). ...
Impacts of extreme winter warming events on litter decomposition in a sub-Arctic heathland
1
2010
... 众多学者认为是由于温度升高影响了微生物的代谢活性, 从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004).冬季植物根系呼吸显著减少, 土壤呼吸主要是微生物的呼吸(Schindlbacher et al., 2007; Ruehr & Buchmann, 2010).增温一方面会增加土壤无机氮库(Luo et al., 2001; Rustad et al., 2001; Melillo et al., 2002), 而无机氮库的增加在一定程度上弥补了冬季活性养分的缺失, 从而促进了微生物的生长; 另一方面, 增温能够提高土壤微生物和酶的活性(Bokhorst et al., 2010), 从而提高土壤中CO2的释放速率.有研究发现, 冬季的土壤呼吸平均能释放掉生态系统在生长季所固存碳的50% (Brooks et al., 2004), 在此情况下, 温度的升高将进一步导致土壤向大气排放更多CO2.因此, 气候变化引起的冬季土壤呼吸速率的升高可能会显著降低高海拔、高纬度生态系统的碳储量(Melillo et al., 2002). ...
Soil carbon stocks in experimental mesocosms are dependent on the rate of labile carbon, nitrogen and phosphorus inputs to soils
3
2008
... 这是由于土壤活性碳库是有限的, 当这部分碳释放消耗后, 增温不再刺激土壤呼吸(Melillo et al., 2002); 同时, 随着温度的进一步升高或较高温度持续时间的延长, 呼吸底物的有效性降低(Atkin & Tjoeller, 2003; Bradford et al., 2008; Yuste et al., 2010), 从而降低了土壤呼吸温度敏感性, 进而减缓土壤呼吸随温度升高而增加的量(Jarvis & Linder, 2000; Giardina & Ryan, 2000; Oechel et al., 2000; Luo et al., 2001; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005); 而且, 长期增温可能改变了土壤酶的活性和微生物的种群结构, 从而使土壤呼吸速率降低(Grogan & Jonasson, 2005; Bradford et al., 2008; Hartley et al., 2008); 此外, 因增温引起的其他生态因子的变化也能引起土壤呼吸速率的降低(陈全胜等, 2003a), 这样导致的结果是潜在地减弱了增温-土壤呼吸-大气CO2浓度这个环节的正反馈(Luo et al., 2001). ...
... ; Bradford et al., 2008; Hartley et al., 2008); 此外, 因增温引起的其他生态因子的变化也能引起土壤呼吸速率的降低(陈全胜等, 2003a), 这样导致的结果是潜在地减弱了增温-土壤呼吸-大气CO2浓度这个环节的正反馈(Luo et al., 2001). ...
Carbon limitation of soil respiration under winter snow packs: Potential feedbacks between growing season and winter carbon fluxes
2
2004
... 湿地生态系统在全球碳收支平衡中扮演着重要角色(Huntingford et al., 2009).一方面, 湿地具有较高的初级生产力, 因而具有较强的固碳能力(Gorham, 1991).另一方面, 全球变暖背景下, 温度升高促进土壤有机碳分解, 可能会使湿地碳库由碳汇变为碳源(孔雨光等, 2009).然而, 与其他生态系统类型相比, 增温对湿地生态系统土壤呼吸的研究相对较少.同时, 目前土壤呼吸测定大多集中在生长季, 对年土壤呼吸量的估算大多基于非生长季土壤呼吸为0的假设(Grogan & Jonasson, 2006).然而已有研究表明土壤呼吸是全年性的过程, 非生长季土壤呼吸不仅不为0, 而且能占到年土壤呼吸总量的14%-30% (Jones, 1999), 同时冬季会释放生长季固存碳的50%甚至更多(Oechel et al., 2000; Brooks et al., 2004; Monson, 2005), 是区域碳收支非常重要的组成部分(Wickland et al., 2001; Schimel et al., 2006; Han et al., 2012), 能够显著地影响生态系统的碳平衡(Hubbard et al., 2005; Wang et al., 2014).因此, 研究模拟增温对非生长季土壤呼吸很有必要.同时, 湿地生态系统典型样地一般都分布在偏远地区, 且非生长季观测期间易受雪霜以及低温条件的限制, 环境条件恶劣, 因此相关的研究开展得并不多, 是湿地生态系统碳通量研究的薄弱环节. ...
... 众多学者认为是由于温度升高影响了微生物的代谢活性, 从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004).冬季植物根系呼吸显著减少, 土壤呼吸主要是微生物的呼吸(Schindlbacher et al., 2007; Ruehr & Buchmann, 2010).增温一方面会增加土壤无机氮库(Luo et al., 2001; Rustad et al., 2001; Melillo et al., 2002), 而无机氮库的增加在一定程度上弥补了冬季活性养分的缺失, 从而促进了微生物的生长; 另一方面, 增温能够提高土壤微生物和酶的活性(Bokhorst et al., 2010), 从而提高土壤中CO2的释放速率.有研究发现, 冬季的土壤呼吸平均能释放掉生态系统在生长季所固存碳的50% (Brooks et al., 2004), 在此情况下, 温度的升高将进一步导致土壤向大气排放更多CO2.因此, 气候变化引起的冬季土壤呼吸速率的升高可能会显著降低高海拔、高纬度生态系统的碳储量(Melillo et al., 2002). ...
基于空间化技术对中国近50年平均气温时空演变特征的研究
1
2006
... 土壤呼吸作为陆地生态系统碳循环的重要环节(Raich & Schlesinger, 1992), 是陆地生态系统碳释放的主要途径, 可达总释放量的2/3 (Davidson et al., 2006).全球气候变暖会促进土壤呼吸(Fang & Moncrieff, 2001; Fouche et al., 2014), 进而使大气CO2浓度增加, 在陆地生态系统和大气之间产生一个强烈的正反馈作用(Cox et al., 2000; Davidson et al., 2000), 使全球变暖的情况更趋严重(Peterjohn et al., 1994; Carney et al., 2007).模拟研究表明, 2016- 2035年全球平均地表温度将升高0.3-0.7 ℃, 2018- 2100年将升高0.3-4.8 ℃ (IPCC, 2013).在全球尺度上, 冬季和春季增温速度较快, 而夏季和秋季的增温速率明显低于冬季和春季以及年平均增温速率(Nagato & Tanakab, 2012).北半球高纬度和高海拔地区温度升幅更大, 而且冬季被认为是增温幅度最大的季节(IPCC, 2013).近50年中, 中国年、季的全国平均气温均表现出显著增高趋势, 而冬季和春季地表增温最为显著, 其中冬季增幅最大, 为0.03 ℃·a-1 (蔡福等, 2006).因此, 研究增温特别是冬季增温对土壤呼吸的影响对正确评估全球变化背景下的陆地生态系统碳循环具有重要意义. ...
Altered soil microbial community at elevated CO2 leads to loss of soil carbon
1
2007
... 土壤呼吸作为陆地生态系统碳循环的重要环节(Raich & Schlesinger, 1992), 是陆地生态系统碳释放的主要途径, 可达总释放量的2/3 (Davidson et al., 2006).全球气候变暖会促进土壤呼吸(Fang & Moncrieff, 2001; Fouche et al., 2014), 进而使大气CO2浓度增加, 在陆地生态系统和大气之间产生一个强烈的正反馈作用(Cox et al., 2000; Davidson et al., 2000), 使全球变暖的情况更趋严重(Peterjohn et al., 1994; Carney et al., 2007).模拟研究表明, 2016- 2035年全球平均地表温度将升高0.3-0.7 ℃, 2018- 2100年将升高0.3-4.8 ℃ (IPCC, 2013).在全球尺度上, 冬季和春季增温速度较快, 而夏季和秋季的增温速率明显低于冬季和春季以及年平均增温速率(Nagato & Tanakab, 2012).北半球高纬度和高海拔地区温度升幅更大, 而且冬季被认为是增温幅度最大的季节(IPCC, 2013).近50年中, 中国年、季的全国平均气温均表现出显著增高趋势, 而冬季和春季地表增温最为显著, 其中冬季增幅最大, 为0.03 ℃·a-1 (蔡福等, 2006).因此, 研究增温特别是冬季增温对土壤呼吸的影响对正确评估全球变化背景下的陆地生态系统碳循环具有重要意义. ...
Summer fluxes of atmospheric greenhouse gases N2O, CH4 and CO2 from mangrove soil in South China
1
2010
... 增温不仅能直接影响土壤呼吸, 也能通过影响土壤盐分进而影响土壤呼吸.黄河三角洲滨海湿地咸淡水交互作用明显, 且地下水位浅(Fan et al., 2011; Zhong & Du, 2013), 当土壤温度升高时, 土壤水分的蒸发加速, 从而促进了地下咸水向土壤表面的输送(Wichern et al., 2006; Setia et al., 2011), 导致的结果是土壤表层盐分含量高.因此, 盐度也可能是影响滨海湿地CO2产生与排放的重要环境因子(仝川等, 2011).本研究中, 在盐分限制阶段, 增温处理下土壤盐分抑制了滨海湿地土壤呼吸速率(图3).很多研究表明, 随着土壤含盐量的增加, 土壤呼吸速率呈现下降的趋势(Setia et al., 2010; Wong et al., 2010; Hugler & Sievert, 2011; Yan et al., 2013).例如, 海水入侵淡水潮汐沼泽时, 高盐度水平下土壤CO2产生速率随着盐度的升高而减小(Marton et al., 2012); 长江口滩涂土壤呼吸呈现出近岸水体年平均盐度越高, 土壤呼吸速率越低的趋势, 即距海越近土壤呼吸速率越小(聂明华等, 2011); 珠江三角地区近陆红树林湿地CO2排放通量显著高于近海红树林湿地, 这可能是由土壤盐分的差异性引起的(Chen et al., 2010); 当土壤盐分含量比对照高出4倍时, 南澳大利亚莫纳托湿地土壤呼吸速率降低20% (Hasbullah & Petra, 2015).另外, 盐分还可能导致土壤碳源、汇的转化, 例如美国Nueces三角洲湿地在土壤含水量较高、土壤盐度较低的情况下表现为吸收CO2, 而在土壤含水量较低、土壤盐度较高的情况下则表现为排放CO2 (Heinsch et al., 2004). ...
水分对土壤呼吸的影响及机理
1
2003
... 这是由于土壤活性碳库是有限的, 当这部分碳释放消耗后, 增温不再刺激土壤呼吸(Melillo et al., 2002); 同时, 随着温度的进一步升高或较高温度持续时间的延长, 呼吸底物的有效性降低(Atkin & Tjoeller, 2003; Bradford et al., 2008; Yuste et al., 2010), 从而降低了土壤呼吸温度敏感性, 进而减缓土壤呼吸随温度升高而增加的量(Jarvis & Linder, 2000; Giardina & Ryan, 2000; Oechel et al., 2000; Luo et al., 2001; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005); 而且, 长期增温可能改变了土壤酶的活性和微生物的种群结构, 从而使土壤呼吸速率降低(Grogan & Jonasson, 2005; Bradford et al., 2008; Hartley et al., 2008); 此外, 因增温引起的其他生态因子的变化也能引起土壤呼吸速率的降低(陈全胜等, 2003a), 这样导致的结果是潜在地减弱了增温-土壤呼吸-大气CO2浓度这个环节的正反馈(Luo et al., 2001). ...
Acceleration of global warming due to carbon-cycle feedbacks in a coupled climate model
1
2000
... 土壤呼吸作为陆地生态系统碳循环的重要环节(Raich & Schlesinger, 1992), 是陆地生态系统碳释放的主要途径, 可达总释放量的2/3 (Davidson et al., 2006).全球气候变暖会促进土壤呼吸(Fang & Moncrieff, 2001; Fouche et al., 2014), 进而使大气CO2浓度增加, 在陆地生态系统和大气之间产生一个强烈的正反馈作用(Cox et al., 2000; Davidson et al., 2000), 使全球变暖的情况更趋严重(Peterjohn et al., 1994; Carney et al., 2007).模拟研究表明, 2016- 2035年全球平均地表温度将升高0.3-0.7 ℃, 2018- 2100年将升高0.3-4.8 ℃ (IPCC, 2013).在全球尺度上, 冬季和春季增温速度较快, 而夏季和秋季的增温速率明显低于冬季和春季以及年平均增温速率(Nagato & Tanakab, 2012).北半球高纬度和高海拔地区温度升幅更大, 而且冬季被认为是增温幅度最大的季节(IPCC, 2013).近50年中, 中国年、季的全国平均气温均表现出显著增高趋势, 而冬季和春季地表增温最为显著, 其中冬季增幅最大, 为0.03 ℃·a-1 (蔡福等, 2006).因此, 研究增温特别是冬季增温对土壤呼吸的影响对正确评估全球变化背景下的陆地生态系统碳循环具有重要意义. ...
NaCl盐度对活性污泥处理系统的影响
1
2004
... 盐分影响着微生物异养呼吸作用, 从而改变土壤呼吸速率.一方面, 盐分抑制土壤微生物活性, 使得土壤呼吸随盐度的升高而降低(Wichern et al., 2006; Wong et al., 2008; Iwai et al., 2012; 李凤霞等, 2012); 同时, 高的含盐量会降低微生物数量(Garcia & Hernandez, 1996; Pattnaik et al., 2000; Pivnickova et al., 2010; Kiehn et al., 2013)以及微生物群落的多样性(Baldwin et al., 2006); 并且, 高盐度能使微生物生理形态发生明显变化, 从而使其分解有机质的能力受到强烈影响(Thottathil et al., 2008), 微生物受到盐分的影响时, 一般能通过自身的渗透压调节机制来平衡细胞内的渗透压, 而在高的渗透压条件下, 微生物耗氧速率增加, 但耗氧速率的增加不是为了有机物的降解, 而是为了能够抵御高盐环境对微生物所产生的伤害(Pankhurst et al., 2001; Rietz & Haynes, 2003; 崔有为等, 2004; Thottathil et al., 2008).另一方面, 多数土壤水解酶与氧化还原酶类活性均随盐渍化的水平升高而明显下降(Rietz & Haynes, 2003; 张建锋等, 2005), 低盐分对酶的活性有促进作用, 而高盐分条件下其活性反而下降(Yan et al., 2013). ...
Soil warming and organic carbon content
1
2000
... 土壤呼吸作为陆地生态系统碳循环的重要环节(Raich & Schlesinger, 1992), 是陆地生态系统碳释放的主要途径, 可达总释放量的2/3 (Davidson et al., 2006).全球气候变暖会促进土壤呼吸(Fang & Moncrieff, 2001; Fouche et al., 2014), 进而使大气CO2浓度增加, 在陆地生态系统和大气之间产生一个强烈的正反馈作用(Cox et al., 2000; Davidson et al., 2000), 使全球变暖的情况更趋严重(Peterjohn et al., 1994; Carney et al., 2007).模拟研究表明, 2016- 2035年全球平均地表温度将升高0.3-0.7 ℃, 2018- 2100年将升高0.3-4.8 ℃ (IPCC, 2013).在全球尺度上, 冬季和春季增温速度较快, 而夏季和秋季的增温速率明显低于冬季和春季以及年平均增温速率(Nagato & Tanakab, 2012).北半球高纬度和高海拔地区温度升幅更大, 而且冬季被认为是增温幅度最大的季节(IPCC, 2013).近50年中, 中国年、季的全国平均气温均表现出显著增高趋势, 而冬季和春季地表增温最为显著, 其中冬季增幅最大, 为0.03 ℃·a-1 (蔡福等, 2006).因此, 研究增温特别是冬季增温对土壤呼吸的影响对正确评估全球变化背景下的陆地生态系统碳循环具有重要意义. ...
A distinct seasonal pattern of the ratio of soil respiration to total ecosystem respiration in a spruce- dominated forest
1
2006
... 土壤呼吸作为陆地生态系统碳循环的重要环节(Raich & Schlesinger, 1992), 是陆地生态系统碳释放的主要途径, 可达总释放量的2/3 (Davidson et al., 2006).全球气候变暖会促进土壤呼吸(Fang & Moncrieff, 2001; Fouche et al., 2014), 进而使大气CO2浓度增加, 在陆地生态系统和大气之间产生一个强烈的正反馈作用(Cox et al., 2000; Davidson et al., 2000), 使全球变暖的情况更趋严重(Peterjohn et al., 1994; Carney et al., 2007).模拟研究表明, 2016- 2035年全球平均地表温度将升高0.3-0.7 ℃, 2018- 2100年将升高0.3-4.8 ℃ (IPCC, 2013).在全球尺度上, 冬季和春季增温速度较快, 而夏季和秋季的增温速率明显低于冬季和春季以及年平均增温速率(Nagato & Tanakab, 2012).北半球高纬度和高海拔地区温度升幅更大, 而且冬季被认为是增温幅度最大的季节(IPCC, 2013).近50年中, 中国年、季的全国平均气温均表现出显著增高趋势, 而冬季和春季地表增温最为显著, 其中冬季增幅最大, 为0.03 ℃·a-1 (蔡福等, 2006).因此, 研究增温特别是冬季增温对土壤呼吸的影响对正确评估全球变化背景下的陆地生态系统碳循环具有重要意义. ...
The response of heterotrophic CO2 flux to soil warming
2
2005
... 野外自然条件下的增温实验是研究全球变化重要的信息来源(徐振峰等, 2010), 是研究全球变暖与陆地生态系统相互关系的一个主要方法(牛书丽等, 2007; Hoeppner & Dukes, 2012; Hou et al., 2013), 不仅能够获取气候变暖对生态系统影响的直接证据, 还可以解释陆地生态系统对气候变化响应的内在机制(Hou et al., 2013).目前, 有关全球变暖对陆地生态系统影响的增温装置主要有被动增温和主动增温两类.其中, 红外辐射器能更真实有效地模拟全球变暖, 对土壤无物理干扰, 也不改变小气候状况, 是现有的模拟增温的理想装置, 近年来被广泛应用于森林、草地、苔原等生态系统(牛书丽等, 2007).增温影响微生物或根系的代谢活性从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004), 大多数研究表明增温明显促进了土壤呼吸(Lin et al., 2001; Wan et al., 2007; Zhou et al., 2007; Xia et al., 2009; Wang et al., 2014).然而, 也有研究表明随着温度的升高, 土壤呼吸速率出现降低的趋势(Pajari, 1995; Saleska et al., 1999; Melillo et al., 2002; Yin et al., 2013)或不变(Wan et al., 2007), 这可能是因为土壤有限的活性碳库(Oechel et al., 2000; Giardina & Ryan, 2000; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005)和土壤呼吸温度适应性的存在 (Luo et al., 2001). ...
... 这是由于土壤活性碳库是有限的, 当这部分碳释放消耗后, 增温不再刺激土壤呼吸(Melillo et al., 2002); 同时, 随着温度的进一步升高或较高温度持续时间的延长, 呼吸底物的有效性降低(Atkin & Tjoeller, 2003; Bradford et al., 2008; Yuste et al., 2010), 从而降低了土壤呼吸温度敏感性, 进而减缓土壤呼吸随温度升高而增加的量(Jarvis & Linder, 2000; Giardina & Ryan, 2000; Oechel et al., 2000; Luo et al., 2001; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005); 而且, 长期增温可能改变了土壤酶的活性和微生物的种群结构, 从而使土壤呼吸速率降低(Grogan & Jonasson, 2005; Bradford et al., 2008; Hartley et al., 2008); 此外, 因增温引起的其他生态因子的变化也能引起土壤呼吸速率的降低(陈全胜等, 2003a), 这样导致的结果是潜在地减弱了增温-土壤呼吸-大气CO2浓度这个环节的正反馈(Luo et al., 2001). ...
Potential plant species distribution in the Yellow River Delta under the influence of groundwater level and soil salinity
2
2011
... 滨海湿地作为湿地生态系统的重要类型, 其地下水位浅且受淡咸水交互作用(Fan et al., 2011; Zhong & Du, 2013), 温度升高增强土壤水分蒸发, 带动浅层地下水可溶性盐向地表输送(Yao & Yang, 2010; Zhang et al., 2011), 从而引起土壤表层盐分的变化, 而盐分的升高会影响土壤呼吸(Wichern et al., 2006; Yang et al., 2009; Setia et al., 2011), 然而关于温度、盐分是如何影响滨海湿地土壤呼吸的, 目前还没有明确结论.因此本研究选取黄河三角洲滨海湿地为研究对象, 采用红外辐射加热器模拟土壤增温, 对非生长季(2014年11月-2015年4月)的土壤呼吸速率、土壤温湿度和土壤盐分等进行监测.分析增温对非生长季土壤呼吸日变化及季节变化的影响, 并探讨土壤呼吸对土壤温度、盐分变化的响应机制, 可为了解未来气候变暖对土壤碳循环的影响规律提供基础数据和理论依据. ...
... 增温不仅能直接影响土壤呼吸, 也能通过影响土壤盐分进而影响土壤呼吸.黄河三角洲滨海湿地咸淡水交互作用明显, 且地下水位浅(Fan et al., 2011; Zhong & Du, 2013), 当土壤温度升高时, 土壤水分的蒸发加速, 从而促进了地下咸水向土壤表面的输送(Wichern et al., 2006; Setia et al., 2011), 导致的结果是土壤表层盐分含量高.因此, 盐度也可能是影响滨海湿地CO2产生与排放的重要环境因子(仝川等, 2011).本研究中, 在盐分限制阶段, 增温处理下土壤盐分抑制了滨海湿地土壤呼吸速率(图3).很多研究表明, 随着土壤含盐量的增加, 土壤呼吸速率呈现下降的趋势(Setia et al., 2010; Wong et al., 2010; Hugler & Sievert, 2011; Yan et al., 2013).例如, 海水入侵淡水潮汐沼泽时, 高盐度水平下土壤CO2产生速率随着盐度的升高而减小(Marton et al., 2012); 长江口滩涂土壤呼吸呈现出近岸水体年平均盐度越高, 土壤呼吸速率越低的趋势, 即距海越近土壤呼吸速率越小(聂明华等, 2011); 珠江三角地区近陆红树林湿地CO2排放通量显著高于近海红树林湿地, 这可能是由土壤盐分的差异性引起的(Chen et al., 2010); 当土壤盐分含量比对照高出4倍时, 南澳大利亚莫纳托湿地土壤呼吸速率降低20% (Hasbullah & Petra, 2015).另外, 盐分还可能导致土壤碳源、汇的转化, 例如美国Nueces三角洲湿地在土壤含水量较高、土壤盐度较低的情况下表现为吸收CO2, 而在土壤含水量较低、土壤盐度较高的情况下则表现为排放CO2 (Heinsch et al., 2004). ...
The dependence of soil CO2 efflux on temperature
1
2001
... 土壤呼吸作为陆地生态系统碳循环的重要环节(Raich & Schlesinger, 1992), 是陆地生态系统碳释放的主要途径, 可达总释放量的2/3 (Davidson et al., 2006).全球气候变暖会促进土壤呼吸(Fang & Moncrieff, 2001; Fouche et al., 2014), 进而使大气CO2浓度增加, 在陆地生态系统和大气之间产生一个强烈的正反馈作用(Cox et al., 2000; Davidson et al., 2000), 使全球变暖的情况更趋严重(Peterjohn et al., 1994; Carney et al., 2007).模拟研究表明, 2016- 2035年全球平均地表温度将升高0.3-0.7 ℃, 2018- 2100年将升高0.3-4.8 ℃ (IPCC, 2013).在全球尺度上, 冬季和春季增温速度较快, 而夏季和秋季的增温速率明显低于冬季和春季以及年平均增温速率(Nagato & Tanakab, 2012).北半球高纬度和高海拔地区温度升幅更大, 而且冬季被认为是增温幅度最大的季节(IPCC, 2013).近50年中, 中国年、季的全国平均气温均表现出显著增高趋势, 而冬季和春季地表增温最为显著, 其中冬季增幅最大, 为0.03 ℃·a-1 (蔡福等, 2006).因此, 研究增温特别是冬季增温对土壤呼吸的影响对正确评估全球变化背景下的陆地生态系统碳循环具有重要意义. ...
Increased CO2 fluxes under warming tests and soil solution chemistry in Histic and Turbic Cryosols, Salluit, Nunavik, Canada
1
2014
... 土壤呼吸作为陆地生态系统碳循环的重要环节(Raich & Schlesinger, 1992), 是陆地生态系统碳释放的主要途径, 可达总释放量的2/3 (Davidson et al., 2006).全球气候变暖会促进土壤呼吸(Fang & Moncrieff, 2001; Fouche et al., 2014), 进而使大气CO2浓度增加, 在陆地生态系统和大气之间产生一个强烈的正反馈作用(Cox et al., 2000; Davidson et al., 2000), 使全球变暖的情况更趋严重(Peterjohn et al., 1994; Carney et al., 2007).模拟研究表明, 2016- 2035年全球平均地表温度将升高0.3-0.7 ℃, 2018- 2100年将升高0.3-4.8 ℃ (IPCC, 2013).在全球尺度上, 冬季和春季增温速度较快, 而夏季和秋季的增温速率明显低于冬季和春季以及年平均增温速率(Nagato & Tanakab, 2012).北半球高纬度和高海拔地区温度升幅更大, 而且冬季被认为是增温幅度最大的季节(IPCC, 2013).近50年中, 中国年、季的全国平均气温均表现出显著增高趋势, 而冬季和春季地表增温最为显著, 其中冬季增幅最大, 为0.03 ℃·a-1 (蔡福等, 2006).因此, 研究增温特别是冬季增温对土壤呼吸的影响对正确评估全球变化背景下的陆地生态系统碳循环具有重要意义. ...
Influence of salinity on the biological and biochemical activity of a calciorthird soil
1
1996
... 盐分影响着微生物异养呼吸作用, 从而改变土壤呼吸速率.一方面, 盐分抑制土壤微生物活性, 使得土壤呼吸随盐度的升高而降低(Wichern et al., 2006; Wong et al., 2008; Iwai et al., 2012; 李凤霞等, 2012); 同时, 高的含盐量会降低微生物数量(Garcia & Hernandez, 1996; Pattnaik et al., 2000; Pivnickova et al., 2010; Kiehn et al., 2013)以及微生物群落的多样性(Baldwin et al., 2006); 并且, 高盐度能使微生物生理形态发生明显变化, 从而使其分解有机质的能力受到强烈影响(Thottathil et al., 2008), 微生物受到盐分的影响时, 一般能通过自身的渗透压调节机制来平衡细胞内的渗透压, 而在高的渗透压条件下, 微生物耗氧速率增加, 但耗氧速率的增加不是为了有机物的降解, 而是为了能够抵御高盐环境对微生物所产生的伤害(Pankhurst et al., 2001; Rietz & Haynes, 2003; 崔有为等, 2004; Thottathil et al., 2008).另一方面, 多数土壤水解酶与氧化还原酶类活性均随盐渍化的水平升高而明显下降(Rietz & Haynes, 2003; 张建锋等, 2005), 低盐分对酶的活性有促进作用, 而高盐分条件下其活性反而下降(Yan et al., 2013). ...
Interpreting the dependence of soil respiration on soil temperature and water content in a boreal aspen stand
Evidence that decomposition rates of organic carbon in mineral soil do not vary with temperature
3
2000
... 野外自然条件下的增温实验是研究全球变化重要的信息来源(徐振峰等, 2010), 是研究全球变暖与陆地生态系统相互关系的一个主要方法(牛书丽等, 2007; Hoeppner & Dukes, 2012; Hou et al., 2013), 不仅能够获取气候变暖对生态系统影响的直接证据, 还可以解释陆地生态系统对气候变化响应的内在机制(Hou et al., 2013).目前, 有关全球变暖对陆地生态系统影响的增温装置主要有被动增温和主动增温两类.其中, 红外辐射器能更真实有效地模拟全球变暖, 对土壤无物理干扰, 也不改变小气候状况, 是现有的模拟增温的理想装置, 近年来被广泛应用于森林、草地、苔原等生态系统(牛书丽等, 2007).增温影响微生物或根系的代谢活性从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004), 大多数研究表明增温明显促进了土壤呼吸(Lin et al., 2001; Wan et al., 2007; Zhou et al., 2007; Xia et al., 2009; Wang et al., 2014).然而, 也有研究表明随着温度的升高, 土壤呼吸速率出现降低的趋势(Pajari, 1995; Saleska et al., 1999; Melillo et al., 2002; Yin et al., 2013)或不变(Wan et al., 2007), 这可能是因为土壤有限的活性碳库(Oechel et al., 2000; Giardina & Ryan, 2000; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005)和土壤呼吸温度适应性的存在 (Luo et al., 2001). ...
... 这是由于土壤活性碳库是有限的, 当这部分碳释放消耗后, 增温不再刺激土壤呼吸(Melillo et al., 2002); 同时, 随着温度的进一步升高或较高温度持续时间的延长, 呼吸底物的有效性降低(Atkin & Tjoeller, 2003; Bradford et al., 2008; Yuste et al., 2010), 从而降低了土壤呼吸温度敏感性, 进而减缓土壤呼吸随温度升高而增加的量(Jarvis & Linder, 2000; Giardina & Ryan, 2000; Oechel et al., 2000; Luo et al., 2001; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005); 而且, 长期增温可能改变了土壤酶的活性和微生物的种群结构, 从而使土壤呼吸速率降低(Grogan & Jonasson, 2005; Bradford et al., 2008; Hartley et al., 2008); 此外, 因增温引起的其他生态因子的变化也能引起土壤呼吸速率的降低(陈全胜等, 2003a), 这样导致的结果是潜在地减弱了增温-土壤呼吸-大气CO2浓度这个环节的正反馈(Luo et al., 2001). ...
Residue properties influence the impact of salinity on soil respiration
1
2015
... 增温不仅能直接影响土壤呼吸, 也能通过影响土壤盐分进而影响土壤呼吸.黄河三角洲滨海湿地咸淡水交互作用明显, 且地下水位浅(Fan et al., 2011; Zhong & Du, 2013), 当土壤温度升高时, 土壤水分的蒸发加速, 从而促进了地下咸水向土壤表面的输送(Wichern et al., 2006; Setia et al., 2011), 导致的结果是土壤表层盐分含量高.因此, 盐度也可能是影响滨海湿地CO2产生与排放的重要环境因子(仝川等, 2011).本研究中, 在盐分限制阶段, 增温处理下土壤盐分抑制了滨海湿地土壤呼吸速率(图3).很多研究表明, 随着土壤含盐量的增加, 土壤呼吸速率呈现下降的趋势(Setia et al., 2010; Wong et al., 2010; Hugler & Sievert, 2011; Yan et al., 2013).例如, 海水入侵淡水潮汐沼泽时, 高盐度水平下土壤CO2产生速率随着盐度的升高而减小(Marton et al., 2012); 长江口滩涂土壤呼吸呈现出近岸水体年平均盐度越高, 土壤呼吸速率越低的趋势, 即距海越近土壤呼吸速率越小(聂明华等, 2011); 珠江三角地区近陆红树林湿地CO2排放通量显著高于近海红树林湿地, 这可能是由土壤盐分的差异性引起的(Chen et al., 2010); 当土壤盐分含量比对照高出4倍时, 南澳大利亚莫纳托湿地土壤呼吸速率降低20% (Hasbullah & Petra, 2015).另外, 盐分还可能导致土壤碳源、汇的转化, 例如美国Nueces三角洲湿地在土壤含水量较高、土壤盐度较低的情况下表现为吸收CO2, 而在土壤含水量较低、土壤盐度较高的情况下则表现为排放CO2 (Heinsch et al., 2004). ...
Carbon dioxide exchange in a high marsh on the Texas Gulf Coast: Effects of freshwater availability
1
2004
... 增温不仅能直接影响土壤呼吸, 也能通过影响土壤盐分进而影响土壤呼吸.黄河三角洲滨海湿地咸淡水交互作用明显, 且地下水位浅(Fan et al., 2011; Zhong & Du, 2013), 当土壤温度升高时, 土壤水分的蒸发加速, 从而促进了地下咸水向土壤表面的输送(Wichern et al., 2006; Setia et al., 2011), 导致的结果是土壤表层盐分含量高.因此, 盐度也可能是影响滨海湿地CO2产生与排放的重要环境因子(仝川等, 2011).本研究中, 在盐分限制阶段, 增温处理下土壤盐分抑制了滨海湿地土壤呼吸速率(图3).很多研究表明, 随着土壤含盐量的增加, 土壤呼吸速率呈现下降的趋势(Setia et al., 2010; Wong et al., 2010; Hugler & Sievert, 2011; Yan et al., 2013).例如, 海水入侵淡水潮汐沼泽时, 高盐度水平下土壤CO2产生速率随着盐度的升高而减小(Marton et al., 2012); 长江口滩涂土壤呼吸呈现出近岸水体年平均盐度越高, 土壤呼吸速率越低的趋势, 即距海越近土壤呼吸速率越小(聂明华等, 2011); 珠江三角地区近陆红树林湿地CO2排放通量显著高于近海红树林湿地, 这可能是由土壤盐分的差异性引起的(Chen et al., 2010); 当土壤盐分含量比对照高出4倍时, 南澳大利亚莫纳托湿地土壤呼吸速率降低20% (Hasbullah & Petra, 2015).另外, 盐分还可能导致土壤碳源、汇的转化, 例如美国Nueces三角洲湿地在土壤含水量较高、土壤盐度较低的情况下表现为吸收CO2, 而在土壤含水量较低、土壤盐度较高的情况下则表现为排放CO2 (Heinsch et al., 2004). ...
Interactive responses of old- field plant growth and composition to warming and precipitation
1
2012
... 野外自然条件下的增温实验是研究全球变化重要的信息来源(徐振峰等, 2010), 是研究全球变暖与陆地生态系统相互关系的一个主要方法(牛书丽等, 2007; Hoeppner & Dukes, 2012; Hou et al., 2013), 不仅能够获取气候变暖对生态系统影响的直接证据, 还可以解释陆地生态系统对气候变化响应的内在机制(Hou et al., 2013).目前, 有关全球变暖对陆地生态系统影响的增温装置主要有被动增温和主动增温两类.其中, 红外辐射器能更真实有效地模拟全球变暖, 对土壤无物理干扰, 也不改变小气候状况, 是现有的模拟增温的理想装置, 近年来被广泛应用于森林、草地、苔原等生态系统(牛书丽等, 2007).增温影响微生物或根系的代谢活性从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004), 大多数研究表明增温明显促进了土壤呼吸(Lin et al., 2001; Wan et al., 2007; Zhou et al., 2007; Xia et al., 2009; Wang et al., 2014).然而, 也有研究表明随着温度的升高, 土壤呼吸速率出现降低的趋势(Pajari, 1995; Saleska et al., 1999; Melillo et al., 2002; Yin et al., 2013)或不变(Wan et al., 2007), 这可能是因为土壤有限的活性碳库(Oechel et al., 2000; Giardina & Ryan, 2000; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005)和土壤呼吸温度适应性的存在 (Luo et al., 2001). ...
Interactive effects of warming and increased precipitation on community structure and composition in an annual forb dominated desert steppe
2
2013
... 野外自然条件下的增温实验是研究全球变化重要的信息来源(徐振峰等, 2010), 是研究全球变暖与陆地生态系统相互关系的一个主要方法(牛书丽等, 2007; Hoeppner & Dukes, 2012; Hou et al., 2013), 不仅能够获取气候变暖对生态系统影响的直接证据, 还可以解释陆地生态系统对气候变化响应的内在机制(Hou et al., 2013).目前, 有关全球变暖对陆地生态系统影响的增温装置主要有被动增温和主动增温两类.其中, 红外辐射器能更真实有效地模拟全球变暖, 对土壤无物理干扰, 也不改变小气候状况, 是现有的模拟增温的理想装置, 近年来被广泛应用于森林、草地、苔原等生态系统(牛书丽等, 2007).增温影响微生物或根系的代谢活性从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004), 大多数研究表明增温明显促进了土壤呼吸(Lin et al., 2001; Wan et al., 2007; Zhou et al., 2007; Xia et al., 2009; Wang et al., 2014).然而, 也有研究表明随着温度的升高, 土壤呼吸速率出现降低的趋势(Pajari, 1995; Saleska et al., 1999; Melillo et al., 2002; Yin et al., 2013)或不变(Wan et al., 2007), 这可能是因为土壤有限的活性碳库(Oechel et al., 2000; Giardina & Ryan, 2000; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005)和土壤呼吸温度适应性的存在 (Luo et al., 2001). ...
... ), 不仅能够获取气候变暖对生态系统影响的直接证据, 还可以解释陆地生态系统对气候变化响应的内在机制(Hou et al., 2013).目前, 有关全球变暖对陆地生态系统影响的增温装置主要有被动增温和主动增温两类.其中, 红外辐射器能更真实有效地模拟全球变暖, 对土壤无物理干扰, 也不改变小气候状况, 是现有的模拟增温的理想装置, 近年来被广泛应用于森林、草地、苔原等生态系统(牛书丽等, 2007).增温影响微生物或根系的代谢活性从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004), 大多数研究表明增温明显促进了土壤呼吸(Lin et al., 2001; Wan et al., 2007; Zhou et al., 2007; Xia et al., 2009; Wang et al., 2014).然而, 也有研究表明随着温度的升高, 土壤呼吸速率出现降低的趋势(Pajari, 1995; Saleska et al., 1999; Melillo et al., 2002; Yin et al., 2013)或不变(Wan et al., 2007), 这可能是因为土壤有限的活性碳库(Oechel et al., 2000; Giardina & Ryan, 2000; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005)和土壤呼吸温度适应性的存在 (Luo et al., 2001). ...
Seasonal patterns in soil surface CO2 flux under snow cover in 50 and 300 year old subalpine forest
1
2005
... 湿地生态系统在全球碳收支平衡中扮演着重要角色(Huntingford et al., 2009).一方面, 湿地具有较高的初级生产力, 因而具有较强的固碳能力(Gorham, 1991).另一方面, 全球变暖背景下, 温度升高促进土壤有机碳分解, 可能会使湿地碳库由碳汇变为碳源(孔雨光等, 2009).然而, 与其他生态系统类型相比, 增温对湿地生态系统土壤呼吸的研究相对较少.同时, 目前土壤呼吸测定大多集中在生长季, 对年土壤呼吸量的估算大多基于非生长季土壤呼吸为0的假设(Grogan & Jonasson, 2006).然而已有研究表明土壤呼吸是全年性的过程, 非生长季土壤呼吸不仅不为0, 而且能占到年土壤呼吸总量的14%-30% (Jones, 1999), 同时冬季会释放生长季固存碳的50%甚至更多(Oechel et al., 2000; Brooks et al., 2004; Monson, 2005), 是区域碳收支非常重要的组成部分(Wickland et al., 2001; Schimel et al., 2006; Han et al., 2012), 能够显著地影响生态系统的碳平衡(Hubbard et al., 2005; Wang et al., 2014).因此, 研究模拟增温对非生长季土壤呼吸很有必要.同时, 湿地生态系统典型样地一般都分布在偏远地区, 且非生长季观测期间易受雪霜以及低温条件的限制, 环境条件恶劣, 因此相关的研究开展得并不多, 是湿地生态系统碳通量研究的薄弱环节. ...
Beyond the Calvin cycle: Autotrophic carbon fixation in the ocean
1
2011
... 增温不仅能直接影响土壤呼吸, 也能通过影响土壤盐分进而影响土壤呼吸.黄河三角洲滨海湿地咸淡水交互作用明显, 且地下水位浅(Fan et al., 2011; Zhong & Du, 2013), 当土壤温度升高时, 土壤水分的蒸发加速, 从而促进了地下咸水向土壤表面的输送(Wichern et al., 2006; Setia et al., 2011), 导致的结果是土壤表层盐分含量高.因此, 盐度也可能是影响滨海湿地CO2产生与排放的重要环境因子(仝川等, 2011).本研究中, 在盐分限制阶段, 增温处理下土壤盐分抑制了滨海湿地土壤呼吸速率(图3).很多研究表明, 随着土壤含盐量的增加, 土壤呼吸速率呈现下降的趋势(Setia et al., 2010; Wong et al., 2010; Hugler & Sievert, 2011; Yan et al., 2013).例如, 海水入侵淡水潮汐沼泽时, 高盐度水平下土壤CO2产生速率随着盐度的升高而减小(Marton et al., 2012); 长江口滩涂土壤呼吸呈现出近岸水体年平均盐度越高, 土壤呼吸速率越低的趋势, 即距海越近土壤呼吸速率越小(聂明华等, 2011); 珠江三角地区近陆红树林湿地CO2排放通量显著高于近海红树林湿地, 这可能是由土壤盐分的差异性引起的(Chen et al., 2010); 当土壤盐分含量比对照高出4倍时, 南澳大利亚莫纳托湿地土壤呼吸速率降低20% (Hasbullah & Petra, 2015).另外, 盐分还可能导致土壤碳源、汇的转化, 例如美国Nueces三角洲湿地在土壤含水量较高、土壤盐度较低的情况下表现为吸收CO2, 而在土壤含水量较低、土壤盐度较高的情况下则表现为排放CO2 (Heinsch et al., 2004). ...
Contributions of carbon cycle uncertainty to future climate projection spread
1
2009
... 湿地生态系统在全球碳收支平衡中扮演着重要角色(Huntingford et al., 2009).一方面, 湿地具有较高的初级生产力, 因而具有较强的固碳能力(Gorham, 1991).另一方面, 全球变暖背景下, 温度升高促进土壤有机碳分解, 可能会使湿地碳库由碳汇变为碳源(孔雨光等, 2009).然而, 与其他生态系统类型相比, 增温对湿地生态系统土壤呼吸的研究相对较少.同时, 目前土壤呼吸测定大多集中在生长季, 对年土壤呼吸量的估算大多基于非生长季土壤呼吸为0的假设(Grogan & Jonasson, 2006).然而已有研究表明土壤呼吸是全年性的过程, 非生长季土壤呼吸不仅不为0, 而且能占到年土壤呼吸总量的14%-30% (Jones, 1999), 同时冬季会释放生长季固存碳的50%甚至更多(Oechel et al., 2000; Brooks et al., 2004; Monson, 2005), 是区域碳收支非常重要的组成部分(Wickland et al., 2001; Schimel et al., 2006; Han et al., 2012), 能够显著地影响生态系统的碳平衡(Hubbard et al., 2005; Wang et al., 2014).因此, 研究模拟增温对非生长季土壤呼吸很有必要.同时, 湿地生态系统典型样地一般都分布在偏远地区, 且非生长季观测期间易受雪霜以及低温条件的限制, 环境条件恶劣, 因此相关的研究开展得并不多, 是湿地生态系统碳通量研究的薄弱环节. ...
3
2013
... 土壤呼吸作为陆地生态系统碳循环的重要环节(Raich & Schlesinger, 1992), 是陆地生态系统碳释放的主要途径, 可达总释放量的2/3 (Davidson et al., 2006).全球气候变暖会促进土壤呼吸(Fang & Moncrieff, 2001; Fouche et al., 2014), 进而使大气CO2浓度增加, 在陆地生态系统和大气之间产生一个强烈的正反馈作用(Cox et al., 2000; Davidson et al., 2000), 使全球变暖的情况更趋严重(Peterjohn et al., 1994; Carney et al., 2007).模拟研究表明, 2016- 2035年全球平均地表温度将升高0.3-0.7 ℃, 2018- 2100年将升高0.3-4.8 ℃ (IPCC, 2013).在全球尺度上, 冬季和春季增温速度较快, 而夏季和秋季的增温速率明显低于冬季和春季以及年平均增温速率(Nagato & Tanakab, 2012).北半球高纬度和高海拔地区温度升幅更大, 而且冬季被认为是增温幅度最大的季节(IPCC, 2013).近50年中, 中国年、季的全国平均气温均表现出显著增高趋势, 而冬季和春季地表增温最为显著, 其中冬季增幅最大, 为0.03 ℃·a-1 (蔡福等, 2006).因此, 研究增温特别是冬季增温对土壤呼吸的影响对正确评估全球变化背景下的陆地生态系统碳循环具有重要意义. ...
... 盐分影响着微生物异养呼吸作用, 从而改变土壤呼吸速率.一方面, 盐分抑制土壤微生物活性, 使得土壤呼吸随盐度的升高而降低(Wichern et al., 2006; Wong et al., 2008; Iwai et al., 2012; 李凤霞等, 2012); 同时, 高的含盐量会降低微生物数量(Garcia & Hernandez, 1996; Pattnaik et al., 2000; Pivnickova et al., 2010; Kiehn et al., 2013)以及微生物群落的多样性(Baldwin et al., 2006); 并且, 高盐度能使微生物生理形态发生明显变化, 从而使其分解有机质的能力受到强烈影响(Thottathil et al., 2008), 微生物受到盐分的影响时, 一般能通过自身的渗透压调节机制来平衡细胞内的渗透压, 而在高的渗透压条件下, 微生物耗氧速率增加, 但耗氧速率的增加不是为了有机物的降解, 而是为了能够抵御高盐环境对微生物所产生的伤害(Pankhurst et al., 2001; Rietz & Haynes, 2003; 崔有为等, 2004; Thottathil et al., 2008).另一方面, 多数土壤水解酶与氧化还原酶类活性均随盐渍化的水平升高而明显下降(Rietz & Haynes, 2003; 张建锋等, 2005), 低盐分对酶的活性有促进作用, 而高盐分条件下其活性反而下降(Yan et al., 2013). ...
Soil property and microbial activity in natural salt affected soils in an alternating wet-dry tropical climate
2
2012
... 本研究中, 在非盐分限制阶段, 增温使得土壤呼吸温度敏感性降低(表1), 这与众多研究的结果相一致.土壤呼吸的温度敏感性随着土壤温度的升高而下降(Janssens & Pilegaard, 2003; Wan et al., 2007; Zhou et al., 2007; Zhong & Du, 2013).北美高草草原没有加温的实验点的Q10值显著高于加温的实验点的Q10值(Luo et al., 2001; Wan et al., 2007; Zhou et al., 2007); 当温度升高幅度在1.8-3.1 ℃时, 芬兰东部北方针叶林土壤呼吸温度敏感性降低了2.7%- 12.2% (Niinisto et al., 2004); 中国长江口崇明岛盐沼湿地非生长季温度升高0.2 ℃, 土壤呼吸温度敏感性降低13.2% (Zhong & Du, 2013). ...
Nitrous oxide and methane emissions from the restored mangrove ecosystem of the Ciénaga Grande de Santa Marta, Colombia
1
2014
... 盐分影响着微生物异养呼吸作用, 从而改变土壤呼吸速率.一方面, 盐分抑制土壤微生物活性, 使得土壤呼吸随盐度的升高而降低(Wichern et al., 2006; Wong et al., 2008; Iwai et al., 2012; 李凤霞等, 2012); 同时, 高的含盐量会降低微生物数量(Garcia & Hernandez, 1996; Pattnaik et al., 2000; Pivnickova et al., 2010; Kiehn et al., 2013)以及微生物群落的多样性(Baldwin et al., 2006); 并且, 高盐度能使微生物生理形态发生明显变化, 从而使其分解有机质的能力受到强烈影响(Thottathil et al., 2008), 微生物受到盐分的影响时, 一般能通过自身的渗透压调节机制来平衡细胞内的渗透压, 而在高的渗透压条件下, 微生物耗氧速率增加, 但耗氧速率的增加不是为了有机物的降解, 而是为了能够抵御高盐环境对微生物所产生的伤害(Pankhurst et al., 2001; Rietz & Haynes, 2003; 崔有为等, 2004; Thottathil et al., 2008).另一方面, 多数土壤水解酶与氧化还原酶类活性均随盐渍化的水平升高而明显下降(Rietz & Haynes, 2003; 张建锋等, 2005), 低盐分对酶的活性有促进作用, 而高盐分条件下其活性反而下降(Yan et al., 2013). ...
宁夏不同类型盐渍化土壤微生物区系及多样性
1
2012
... 野外自然条件下的增温实验是研究全球变化重要的信息来源(徐振峰等, 2010), 是研究全球变暖与陆地生态系统相互关系的一个主要方法(牛书丽等, 2007; Hoeppner & Dukes, 2012; Hou et al., 2013), 不仅能够获取气候变暖对生态系统影响的直接证据, 还可以解释陆地生态系统对气候变化响应的内在机制(Hou et al., 2013).目前, 有关全球变暖对陆地生态系统影响的增温装置主要有被动增温和主动增温两类.其中, 红外辐射器能更真实有效地模拟全球变暖, 对土壤无物理干扰, 也不改变小气候状况, 是现有的模拟增温的理想装置, 近年来被广泛应用于森林、草地、苔原等生态系统(牛书丽等, 2007).增温影响微生物或根系的代谢活性从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004), 大多数研究表明增温明显促进了土壤呼吸(Lin et al., 2001; Wan et al., 2007; Zhou et al., 2007; Xia et al., 2009; Wang et al., 2014).然而, 也有研究表明随着温度的升高, 土壤呼吸速率出现降低的趋势(Pajari, 1995; Saleska et al., 1999; Melillo et al., 2002; Yin et al., 2013)或不变(Wan et al., 2007), 这可能是因为土壤有限的活性碳库(Oechel et al., 2000; Giardina & Ryan, 2000; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005)和土壤呼吸温度适应性的存在 (Luo et al., 2001). ...
Time-dependent responses of soil CO2 efflux components to elevated atmospheric CO2 and temperature in experimental forest mesocosms
The role of organic matter in the sorption capacity of marine sediments
5
2007
... 野外自然条件下的增温实验是研究全球变化重要的信息来源(徐振峰等, 2010), 是研究全球变暖与陆地生态系统相互关系的一个主要方法(牛书丽等, 2007; Hoeppner & Dukes, 2012; Hou et al., 2013), 不仅能够获取气候变暖对生态系统影响的直接证据, 还可以解释陆地生态系统对气候变化响应的内在机制(Hou et al., 2013).目前, 有关全球变暖对陆地生态系统影响的增温装置主要有被动增温和主动增温两类.其中, 红外辐射器能更真实有效地模拟全球变暖, 对土壤无物理干扰, 也不改变小气候状况, 是现有的模拟增温的理想装置, 近年来被广泛应用于森林、草地、苔原等生态系统(牛书丽等, 2007).增温影响微生物或根系的代谢活性从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004), 大多数研究表明增温明显促进了土壤呼吸(Lin et al., 2001; Wan et al., 2007; Zhou et al., 2007; Xia et al., 2009; Wang et al., 2014).然而, 也有研究表明随着温度的升高, 土壤呼吸速率出现降低的趋势(Pajari, 1995; Saleska et al., 1999; Melillo et al., 2002; Yin et al., 2013)或不变(Wan et al., 2007), 这可能是因为土壤有限的活性碳库(Oechel et al., 2000; Giardina & Ryan, 2000; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005)和土壤呼吸温度适应性的存在 (Luo et al., 2001). ...
... 众多学者认为是由于温度升高影响了微生物的代谢活性, 从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004).冬季植物根系呼吸显著减少, 土壤呼吸主要是微生物的呼吸(Schindlbacher et al., 2007; Ruehr & Buchmann, 2010).增温一方面会增加土壤无机氮库(Luo et al., 2001; Rustad et al., 2001; Melillo et al., 2002), 而无机氮库的增加在一定程度上弥补了冬季活性养分的缺失, 从而促进了微生物的生长; 另一方面, 增温能够提高土壤微生物和酶的活性(Bokhorst et al., 2010), 从而提高土壤中CO2的释放速率.有研究发现, 冬季的土壤呼吸平均能释放掉生态系统在生长季所固存碳的50% (Brooks et al., 2004), 在此情况下, 温度的升高将进一步导致土壤向大气排放更多CO2.因此, 气候变化引起的冬季土壤呼吸速率的升高可能会显著降低高海拔、高纬度生态系统的碳储量(Melillo et al., 2002). ...
... 这是由于土壤活性碳库是有限的, 当这部分碳释放消耗后, 增温不再刺激土壤呼吸(Melillo et al., 2002); 同时, 随着温度的进一步升高或较高温度持续时间的延长, 呼吸底物的有效性降低(Atkin & Tjoeller, 2003; Bradford et al., 2008; Yuste et al., 2010), 从而降低了土壤呼吸温度敏感性, 进而减缓土壤呼吸随温度升高而增加的量(Jarvis & Linder, 2000; Giardina & Ryan, 2000; Oechel et al., 2000; Luo et al., 2001; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005); 而且, 长期增温可能改变了土壤酶的活性和微生物的种群结构, 从而使土壤呼吸速率降低(Grogan & Jonasson, 2005; Bradford et al., 2008; Hartley et al., 2008); 此外, 因增温引起的其他生态因子的变化也能引起土壤呼吸速率的降低(陈全胜等, 2003a), 这样导致的结果是潜在地减弱了增温-土壤呼吸-大气CO2浓度这个环节的正反馈(Luo et al., 2001). ...
... 浓度这个环节的正反馈(Luo et al., 2001). ...
... 本研究中, 在非盐分限制阶段, 增温使得土壤呼吸温度敏感性降低(表1), 这与众多研究的结果相一致.土壤呼吸的温度敏感性随着土壤温度的升高而下降(Janssens & Pilegaard, 2003; Wan et al., 2007; Zhou et al., 2007; Zhong & Du, 2013).北美高草草原没有加温的实验点的Q10值显著高于加温的实验点的Q10值(Luo et al., 2001; Wan et al., 2007; Zhou et al., 2007); 当温度升高幅度在1.8-3.1 ℃时, 芬兰东部北方针叶林土壤呼吸温度敏感性降低了2.7%- 12.2% (Niinisto et al., 2004); 中国长江口崇明岛盐沼湿地非生长季温度升高0.2 ℃, 土壤呼吸温度敏感性降低13.2% (Zhong & Du, 2013). ...
Acclimatization of soil respiration to warming in a tall grass prairie
... 增温不仅能直接影响土壤呼吸, 也能通过影响土壤盐分进而影响土壤呼吸.黄河三角洲滨海湿地咸淡水交互作用明显, 且地下水位浅(Fan et al., 2011; Zhong & Du, 2013), 当土壤温度升高时, 土壤水分的蒸发加速, 从而促进了地下咸水向土壤表面的输送(Wichern et al., 2006; Setia et al., 2011), 导致的结果是土壤表层盐分含量高.因此, 盐度也可能是影响滨海湿地CO2产生与排放的重要环境因子(仝川等, 2011).本研究中, 在盐分限制阶段, 增温处理下土壤盐分抑制了滨海湿地土壤呼吸速率(图3).很多研究表明, 随着土壤含盐量的增加, 土壤呼吸速率呈现下降的趋势(Setia et al., 2010; Wong et al., 2010; Hugler & Sievert, 2011; Yan et al., 2013).例如, 海水入侵淡水潮汐沼泽时, 高盐度水平下土壤CO2产生速率随着盐度的升高而减小(Marton et al., 2012); 长江口滩涂土壤呼吸呈现出近岸水体年平均盐度越高, 土壤呼吸速率越低的趋势, 即距海越近土壤呼吸速率越小(聂明华等, 2011); 珠江三角地区近陆红树林湿地CO2排放通量显著高于近海红树林湿地, 这可能是由土壤盐分的差异性引起的(Chen et al., 2010); 当土壤盐分含量比对照高出4倍时, 南澳大利亚莫纳托湿地土壤呼吸速率降低20% (Hasbullah & Petra, 2015).另外, 盐分还可能导致土壤碳源、汇的转化, 例如美国Nueces三角洲湿地在土壤含水量较高、土壤盐度较低的情况下表现为吸收CO2, 而在土壤含水量较低、土壤盐度较高的情况下则表现为排放CO2 (Heinsch et al., 2004). ...
Effects of salinity on denitrification and greenhouse gas production from laboratory-incubated tidal forest soils
7
2012
... 野外自然条件下的增温实验是研究全球变化重要的信息来源(徐振峰等, 2010), 是研究全球变暖与陆地生态系统相互关系的一个主要方法(牛书丽等, 2007; Hoeppner & Dukes, 2012; Hou et al., 2013), 不仅能够获取气候变暖对生态系统影响的直接证据, 还可以解释陆地生态系统对气候变化响应的内在机制(Hou et al., 2013).目前, 有关全球变暖对陆地生态系统影响的增温装置主要有被动增温和主动增温两类.其中, 红外辐射器能更真实有效地模拟全球变暖, 对土壤无物理干扰, 也不改变小气候状况, 是现有的模拟增温的理想装置, 近年来被广泛应用于森林、草地、苔原等生态系统(牛书丽等, 2007).增温影响微生物或根系的代谢活性从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004), 大多数研究表明增温明显促进了土壤呼吸(Lin et al., 2001; Wan et al., 2007; Zhou et al., 2007; Xia et al., 2009; Wang et al., 2014).然而, 也有研究表明随着温度的升高, 土壤呼吸速率出现降低的趋势(Pajari, 1995; Saleska et al., 1999; Melillo et al., 2002; Yin et al., 2013)或不变(Wan et al., 2007), 这可能是因为土壤有限的活性碳库(Oechel et al., 2000; Giardina & Ryan, 2000; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005)和土壤呼吸温度适应性的存在 (Luo et al., 2001). ...
... ; Melillo et al., 2002; Eliasson et al., 2005)和土壤呼吸温度适应性的存在 (Luo et al., 2001). ...
... 众多学者认为是由于温度升高影响了微生物的代谢活性, 从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004).冬季植物根系呼吸显著减少, 土壤呼吸主要是微生物的呼吸(Schindlbacher et al., 2007; Ruehr & Buchmann, 2010).增温一方面会增加土壤无机氮库(Luo et al., 2001; Rustad et al., 2001; Melillo et al., 2002), 而无机氮库的增加在一定程度上弥补了冬季活性养分的缺失, 从而促进了微生物的生长; 另一方面, 增温能够提高土壤微生物和酶的活性(Bokhorst et al., 2010), 从而提高土壤中CO2的释放速率.有研究发现, 冬季的土壤呼吸平均能释放掉生态系统在生长季所固存碳的50% (Brooks et al., 2004), 在此情况下, 温度的升高将进一步导致土壤向大气排放更多CO2.因此, 气候变化引起的冬季土壤呼吸速率的升高可能会显著降低高海拔、高纬度生态系统的碳储量(Melillo et al., 2002). ...
... .因此, 气候变化引起的冬季土壤呼吸速率的升高可能会显著降低高海拔、高纬度生态系统的碳储量(Melillo et al., 2002). ...
... 然而, 也有研究表明, 土壤呼吸与增温的关系并没有固定的反应模式, 土壤呼吸速率随着土壤温度的升高也可能会降低(Pajari, 1995; Saleska et al., 1999)或者不变(Wan et al., 2007).本研究中, 整个非生长季增温对土壤呼吸速率无显著影响, 并且在盐分限制阶段, 增温使土壤呼吸速率降低(图3).这与以下研究结果相似: 美国Harvard森林在试验前6年, 温度升高使CO2通量平均增加了28%, 而在后4年, 温度升高对土壤呼吸的影响明显降低(Melillo et al., 2002); 长江口崇明东滩围垦湿地生态系统中, 非生长季温度升高0.66 ℃时, 土壤呼吸速率降低了16% (Zhong & Du, 2013). ...
... 这是由于土壤活性碳库是有限的, 当这部分碳释放消耗后, 增温不再刺激土壤呼吸(Melillo et al., 2002); 同时, 随着温度的进一步升高或较高温度持续时间的延长, 呼吸底物的有效性降低(Atkin & Tjoeller, 2003; Bradford et al., 2008; Yuste et al., 2010), 从而降低了土壤呼吸温度敏感性, 进而减缓土壤呼吸随温度升高而增加的量(Jarvis & Linder, 2000; Giardina & Ryan, 2000; Oechel et al., 2000; Luo et al., 2001; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005); 而且, 长期增温可能改变了土壤酶的活性和微生物的种群结构, 从而使土壤呼吸速率降低(Grogan & Jonasson, 2005; Bradford et al., 2008; Hartley et al., 2008); 此外, 因增温引起的其他生态因子的变化也能引起土壤呼吸速率的降低(陈全胜等, 2003a), 这样导致的结果是潜在地减弱了增温-土壤呼吸-大气CO2浓度这个环节的正反馈(Luo et al., 2001). ...
... ; Melillo et al., 2002; Eliasson et al., 2005); 而且, 长期增温可能改变了土壤酶的活性和微生物的种群结构, 从而使土壤呼吸速率降低(Grogan & Jonasson, 2005; Bradford et al., 2008; Hartley et al., 2008); 此外, 因增温引起的其他生态因子的变化也能引起土壤呼吸速率的降低(陈全胜等, 2003a), 这样导致的结果是潜在地减弱了增温-土壤呼吸-大气CO2浓度这个环节的正反馈(Luo et al., 2001). ...
Soil warming and carbon-cycle feedbacks to the climate systems
1
2002
... 湿地生态系统在全球碳收支平衡中扮演着重要角色(Huntingford et al., 2009).一方面, 湿地具有较高的初级生产力, 因而具有较强的固碳能力(Gorham, 1991).另一方面, 全球变暖背景下, 温度升高促进土壤有机碳分解, 可能会使湿地碳库由碳汇变为碳源(孔雨光等, 2009).然而, 与其他生态系统类型相比, 增温对湿地生态系统土壤呼吸的研究相对较少.同时, 目前土壤呼吸测定大多集中在生长季, 对年土壤呼吸量的估算大多基于非生长季土壤呼吸为0的假设(Grogan & Jonasson, 2006).然而已有研究表明土壤呼吸是全年性的过程, 非生长季土壤呼吸不仅不为0, 而且能占到年土壤呼吸总量的14%-30% (Jones, 1999), 同时冬季会释放生长季固存碳的50%甚至更多(Oechel et al., 2000; Brooks et al., 2004; Monson, 2005), 是区域碳收支非常重要的组成部分(Wickland et al., 2001; Schimel et al., 2006; Han et al., 2012), 能够显著地影响生态系统的碳平衡(Hubbard et al., 2005; Wang et al., 2014).因此, 研究模拟增温对非生长季土壤呼吸很有必要.同时, 湿地生态系统典型样地一般都分布在偏远地区, 且非生长季观测期间易受雪霜以及低温条件的限制, 环境条件恶劣, 因此相关的研究开展得并不多, 是湿地生态系统碳通量研究的薄弱环节. ...
Climatic influences on net ecosystem CO2 exchange during the transition from wintertime carbon source to springtime carbon sink in a high elevation, subalpine forest
1
2005
... 土壤呼吸作为陆地生态系统碳循环的重要环节(Raich & Schlesinger, 1992), 是陆地生态系统碳释放的主要途径, 可达总释放量的2/3 (Davidson et al., 2006).全球气候变暖会促进土壤呼吸(Fang & Moncrieff, 2001; Fouche et al., 2014), 进而使大气CO2浓度增加, 在陆地生态系统和大气之间产生一个强烈的正反馈作用(Cox et al., 2000; Davidson et al., 2000), 使全球变暖的情况更趋严重(Peterjohn et al., 1994; Carney et al., 2007).模拟研究表明, 2016- 2035年全球平均地表温度将升高0.3-0.7 ℃, 2018- 2100年将升高0.3-4.8 ℃ (IPCC, 2013).在全球尺度上, 冬季和春季增温速度较快, 而夏季和秋季的增温速率明显低于冬季和春季以及年平均增温速率(Nagato & Tanakab, 2012).北半球高纬度和高海拔地区温度升幅更大, 而且冬季被认为是增温幅度最大的季节(IPCC, 2013).近50年中, 中国年、季的全国平均气温均表现出显著增高趋势, 而冬季和春季地表增温最为显著, 其中冬季增幅最大, 为0.03 ℃·a-1 (蔡福等, 2006).因此, 研究增温特别是冬季增温对土壤呼吸的影响对正确评估全球变化背景下的陆地生态系统碳循环具有重要意义. ...
Global warming trend without the contributions from decadal variability of the Arctic Oscillation
1
2012
... 研究区位于山东省东营市的中国科学院黄河三角洲滨海湿地生态试验站(37.76° N, 118.99° E).该研究区属于温带半湿润大陆性季风气候, 阳光充足, 四季分明, 雨热同期.年平均气温为12.9 ℃, 最高气温41.9 ℃, 最低气温-23.3 ℃, 年降水量为550-640 mm (Han et al., 2014), 70%降水集中于5-9月, 降水量的季节和年际变化较大, 年蒸发量为1962 mm (Han et al., 2013).该地区地势平坦, 植物生长茂盛, 土壤质地以轻壤土和中壤土为主, 土壤类型以潮土和盐碱土为主(Nie et al., 2009).主要的植被为芦苇(Phragmites australis)、盐地碱蓬(Suaeda salsa)、柽柳(Tamarix chinensis)和白茅(Imperata cylindrical var. major). ...
Effects of salt marsh invasion by Spartina alterniflora on sulfate-reducing bacteria in the Yangtze River estuary, China
1
2009
... 增温不仅能直接影响土壤呼吸, 也能通过影响土壤盐分进而影响土壤呼吸.黄河三角洲滨海湿地咸淡水交互作用明显, 且地下水位浅(Fan et al., 2011; Zhong & Du, 2013), 当土壤温度升高时, 土壤水分的蒸发加速, 从而促进了地下咸水向土壤表面的输送(Wichern et al., 2006; Setia et al., 2011), 导致的结果是土壤表层盐分含量高.因此, 盐度也可能是影响滨海湿地CO2产生与排放的重要环境因子(仝川等, 2011).本研究中, 在盐分限制阶段, 增温处理下土壤盐分抑制了滨海湿地土壤呼吸速率(图3).很多研究表明, 随着土壤含盐量的增加, 土壤呼吸速率呈现下降的趋势(Setia et al., 2010; Wong et al., 2010; Hugler & Sievert, 2011; Yan et al., 2013).例如, 海水入侵淡水潮汐沼泽时, 高盐度水平下土壤CO2产生速率随着盐度的升高而减小(Marton et al., 2012); 长江口滩涂土壤呼吸呈现出近岸水体年平均盐度越高, 土壤呼吸速率越低的趋势, 即距海越近土壤呼吸速率越小(聂明华等, 2011); 珠江三角地区近陆红树林湿地CO2排放通量显著高于近海红树林湿地, 这可能是由土壤盐分的差异性引起的(Chen et al., 2010); 当土壤盐分含量比对照高出4倍时, 南澳大利亚莫纳托湿地土壤呼吸速率降低20% (Hasbullah & Petra, 2015).另外, 盐分还可能导致土壤碳源、汇的转化, 例如美国Nueces三角洲湿地在土壤含水量较高、土壤盐度较低的情况下表现为吸收CO2, 而在土壤含水量较低、土壤盐度较高的情况下则表现为排放CO2 (Heinsch et al., 2004). ...
长江口潮滩土壤呼吸季节变化及其影响因素
1
2011
... 本研究中, 在非盐分限制阶段, 增温使得土壤呼吸温度敏感性降低(表1), 这与众多研究的结果相一致.土壤呼吸的温度敏感性随着土壤温度的升高而下降(Janssens & Pilegaard, 2003; Wan et al., 2007; Zhou et al., 2007; Zhong & Du, 2013).北美高草草原没有加温的实验点的Q10值显著高于加温的实验点的Q10值(Luo et al., 2001; Wan et al., 2007; Zhou et al., 2007); 当温度升高幅度在1.8-3.1 ℃时, 芬兰东部北方针叶林土壤呼吸温度敏感性降低了2.7%- 12.2% (Niinisto et al., 2004); 中国长江口崇明岛盐沼湿地非生长季温度升高0.2 ℃, 土壤呼吸温度敏感性降低13.2% (Zhong & Du, 2013). ...
Soil CO2 efflux in a boreal pine forest under atmospheric CO2 enrichment and air warming
2
2004
... 野外自然条件下的增温实验是研究全球变化重要的信息来源(徐振峰等, 2010), 是研究全球变暖与陆地生态系统相互关系的一个主要方法(牛书丽等, 2007; Hoeppner & Dukes, 2012; Hou et al., 2013), 不仅能够获取气候变暖对生态系统影响的直接证据, 还可以解释陆地生态系统对气候变化响应的内在机制(Hou et al., 2013).目前, 有关全球变暖对陆地生态系统影响的增温装置主要有被动增温和主动增温两类.其中, 红外辐射器能更真实有效地模拟全球变暖, 对土壤无物理干扰, 也不改变小气候状况, 是现有的模拟增温的理想装置, 近年来被广泛应用于森林、草地、苔原等生态系统(牛书丽等, 2007).增温影响微生物或根系的代谢活性从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004), 大多数研究表明增温明显促进了土壤呼吸(Lin et al., 2001; Wan et al., 2007; Zhou et al., 2007; Xia et al., 2009; Wang et al., 2014).然而, 也有研究表明随着温度的升高, 土壤呼吸速率出现降低的趋势(Pajari, 1995; Saleska et al., 1999; Melillo et al., 2002; Yin et al., 2013)或不变(Wan et al., 2007), 这可能是因为土壤有限的活性碳库(Oechel et al., 2000; Giardina & Ryan, 2000; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005)和土壤呼吸温度适应性的存在 (Luo et al., 2001). ...
... ).目前, 有关全球变暖对陆地生态系统影响的增温装置主要有被动增温和主动增温两类.其中, 红外辐射器能更真实有效地模拟全球变暖, 对土壤无物理干扰, 也不改变小气候状况, 是现有的模拟增温的理想装置, 近年来被广泛应用于森林、草地、苔原等生态系统(牛书丽等, 2007).增温影响微生物或根系的代谢活性从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004), 大多数研究表明增温明显促进了土壤呼吸(Lin et al., 2001; Wan et al., 2007; Zhou et al., 2007; Xia et al., 2009; Wang et al., 2014).然而, 也有研究表明随着温度的升高, 土壤呼吸速率出现降低的趋势(Pajari, 1995; Saleska et al., 1999; Melillo et al., 2002; Yin et al., 2013)或不变(Wan et al., 2007), 这可能是因为土壤有限的活性碳库(Oechel et al., 2000; Giardina & Ryan, 2000; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005)和土壤呼吸温度适应性的存在 (Luo et al., 2001). ...
全球变暖与陆地生态系统研究中的野外增温装置
3
2007
... 野外自然条件下的增温实验是研究全球变化重要的信息来源(徐振峰等, 2010), 是研究全球变暖与陆地生态系统相互关系的一个主要方法(牛书丽等, 2007; Hoeppner & Dukes, 2012; Hou et al., 2013), 不仅能够获取气候变暖对生态系统影响的直接证据, 还可以解释陆地生态系统对气候变化响应的内在机制(Hou et al., 2013).目前, 有关全球变暖对陆地生态系统影响的增温装置主要有被动增温和主动增温两类.其中, 红外辐射器能更真实有效地模拟全球变暖, 对土壤无物理干扰, 也不改变小气候状况, 是现有的模拟增温的理想装置, 近年来被广泛应用于森林、草地、苔原等生态系统(牛书丽等, 2007).增温影响微生物或根系的代谢活性从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004), 大多数研究表明增温明显促进了土壤呼吸(Lin et al., 2001; Wan et al., 2007; Zhou et al., 2007; Xia et al., 2009; Wang et al., 2014).然而, 也有研究表明随着温度的升高, 土壤呼吸速率出现降低的趋势(Pajari, 1995; Saleska et al., 1999; Melillo et al., 2002; Yin et al., 2013)或不变(Wan et al., 2007), 这可能是因为土壤有限的活性碳库(Oechel et al., 2000; Giardina & Ryan, 2000; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005)和土壤呼吸温度适应性的存在 (Luo et al., 2001). ...
... 湿地生态系统在全球碳收支平衡中扮演着重要角色(Huntingford et al., 2009).一方面, 湿地具有较高的初级生产力, 因而具有较强的固碳能力(Gorham, 1991).另一方面, 全球变暖背景下, 温度升高促进土壤有机碳分解, 可能会使湿地碳库由碳汇变为碳源(孔雨光等, 2009).然而, 与其他生态系统类型相比, 增温对湿地生态系统土壤呼吸的研究相对较少.同时, 目前土壤呼吸测定大多集中在生长季, 对年土壤呼吸量的估算大多基于非生长季土壤呼吸为0的假设(Grogan & Jonasson, 2006).然而已有研究表明土壤呼吸是全年性的过程, 非生长季土壤呼吸不仅不为0, 而且能占到年土壤呼吸总量的14%-30% (Jones, 1999), 同时冬季会释放生长季固存碳的50%甚至更多(Oechel et al., 2000; Brooks et al., 2004; Monson, 2005), 是区域碳收支非常重要的组成部分(Wickland et al., 2001; Schimel et al., 2006; Han et al., 2012), 能够显著地影响生态系统的碳平衡(Hubbard et al., 2005; Wang et al., 2014).因此, 研究模拟增温对非生长季土壤呼吸很有必要.同时, 湿地生态系统典型样地一般都分布在偏远地区, 且非生长季观测期间易受雪霜以及低温条件的限制, 环境条件恶劣, 因此相关的研究开展得并不多, 是湿地生态系统碳通量研究的薄弱环节. ...
... 这是由于土壤活性碳库是有限的, 当这部分碳释放消耗后, 增温不再刺激土壤呼吸(Melillo et al., 2002); 同时, 随着温度的进一步升高或较高温度持续时间的延长, 呼吸底物的有效性降低(Atkin & Tjoeller, 2003; Bradford et al., 2008; Yuste et al., 2010), 从而降低了土壤呼吸温度敏感性, 进而减缓土壤呼吸随温度升高而增加的量(Jarvis & Linder, 2000; Giardina & Ryan, 2000; Oechel et al., 2000; Luo et al., 2001; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005); 而且, 长期增温可能改变了土壤酶的活性和微生物的种群结构, 从而使土壤呼吸速率降低(Grogan & Jonasson, 2005; Bradford et al., 2008; Hartley et al., 2008); 此外, 因增温引起的其他生态因子的变化也能引起土壤呼吸速率的降低(陈全胜等, 2003a), 这样导致的结果是潜在地减弱了增温-土壤呼吸-大气CO2浓度这个环节的正反馈(Luo et al., 2001). ...
Acclimation of ecosystem CO2 exchange in the Alaskan Artic in response to decadal climate warming
2
2000
... 野外自然条件下的增温实验是研究全球变化重要的信息来源(徐振峰等, 2010), 是研究全球变暖与陆地生态系统相互关系的一个主要方法(牛书丽等, 2007; Hoeppner & Dukes, 2012; Hou et al., 2013), 不仅能够获取气候变暖对生态系统影响的直接证据, 还可以解释陆地生态系统对气候变化响应的内在机制(Hou et al., 2013).目前, 有关全球变暖对陆地生态系统影响的增温装置主要有被动增温和主动增温两类.其中, 红外辐射器能更真实有效地模拟全球变暖, 对土壤无物理干扰, 也不改变小气候状况, 是现有的模拟增温的理想装置, 近年来被广泛应用于森林、草地、苔原等生态系统(牛书丽等, 2007).增温影响微生物或根系的代谢活性从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004), 大多数研究表明增温明显促进了土壤呼吸(Lin et al., 2001; Wan et al., 2007; Zhou et al., 2007; Xia et al., 2009; Wang et al., 2014).然而, 也有研究表明随着温度的升高, 土壤呼吸速率出现降低的趋势(Pajari, 1995; Saleska et al., 1999; Melillo et al., 2002; Yin et al., 2013)或不变(Wan et al., 2007), 这可能是因为土壤有限的活性碳库(Oechel et al., 2000; Giardina & Ryan, 2000; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005)和土壤呼吸温度适应性的存在 (Luo et al., 2001). ...
... 然而, 也有研究表明, 土壤呼吸与增温的关系并没有固定的反应模式, 土壤呼吸速率随着土壤温度的升高也可能会降低(Pajari, 1995; Saleska et al., 1999)或者不变(Wan et al., 2007).本研究中, 整个非生长季增温对土壤呼吸速率无显著影响, 并且在盐分限制阶段, 增温使土壤呼吸速率降低(图3).这与以下研究结果相似: 美国Harvard森林在试验前6年, 温度升高使CO2通量平均增加了28%, 而在后4年, 温度升高对土壤呼吸的影响明显降低(Melillo et al., 2002); 长江口崇明东滩围垦湿地生态系统中, 非生长季温度升高0.66 ℃时, 土壤呼吸速率降低了16% (Zhong & Du, 2013). ...
Soil respiration in a poor upland site of Scots pine stand subjected to elevated temperatures and atmospheric carbon concentration
1
1995
... 盐分影响着微生物异养呼吸作用, 从而改变土壤呼吸速率.一方面, 盐分抑制土壤微生物活性, 使得土壤呼吸随盐度的升高而降低(Wichern et al., 2006; Wong et al., 2008; Iwai et al., 2012; 李凤霞等, 2012); 同时, 高的含盐量会降低微生物数量(Garcia & Hernandez, 1996; Pattnaik et al., 2000; Pivnickova et al., 2010; Kiehn et al., 2013)以及微生物群落的多样性(Baldwin et al., 2006); 并且, 高盐度能使微生物生理形态发生明显变化, 从而使其分解有机质的能力受到强烈影响(Thottathil et al., 2008), 微生物受到盐分的影响时, 一般能通过自身的渗透压调节机制来平衡细胞内的渗透压, 而在高的渗透压条件下, 微生物耗氧速率增加, 但耗氧速率的增加不是为了有机物的降解, 而是为了能够抵御高盐环境对微生物所产生的伤害(Pankhurst et al., 2001; Rietz & Haynes, 2003; 崔有为等, 2004; Thottathil et al., 2008).另一方面, 多数土壤水解酶与氧化还原酶类活性均随盐渍化的水平升高而明显下降(Rietz & Haynes, 2003; 张建锋等, 2005), 低盐分对酶的活性有促进作用, 而高盐分条件下其活性反而下降(Yan et al., 2013). ...
Influence of salinity on methanogenesis and associated microflora in tropical rice soils
1
2000
... 土壤呼吸作为陆地生态系统碳循环的重要环节(Raich & Schlesinger, 1992), 是陆地生态系统碳释放的主要途径, 可达总释放量的2/3 (Davidson et al., 2006).全球气候变暖会促进土壤呼吸(Fang & Moncrieff, 2001; Fouche et al., 2014), 进而使大气CO2浓度增加, 在陆地生态系统和大气之间产生一个强烈的正反馈作用(Cox et al., 2000; Davidson et al., 2000), 使全球变暖的情况更趋严重(Peterjohn et al., 1994; Carney et al., 2007).模拟研究表明, 2016- 2035年全球平均地表温度将升高0.3-0.7 ℃, 2018- 2100年将升高0.3-4.8 ℃ (IPCC, 2013).在全球尺度上, 冬季和春季增温速度较快, 而夏季和秋季的增温速率明显低于冬季和春季以及年平均增温速率(Nagato & Tanakab, 2012).北半球高纬度和高海拔地区温度升幅更大, 而且冬季被认为是增温幅度最大的季节(IPCC, 2013).近50年中, 中国年、季的全国平均气温均表现出显著增高趋势, 而冬季和春季地表增温最为显著, 其中冬季增幅最大, 为0.03 ℃·a-1 (蔡福等, 2006).因此, 研究增温特别是冬季增温对土壤呼吸的影响对正确评估全球变化背景下的陆地生态系统碳循环具有重要意义. ...
Responses of trace gas fluxes and N availability to experimentally elevated soil temperatures
1
1994
... 盐分影响着微生物异养呼吸作用, 从而改变土壤呼吸速率.一方面, 盐分抑制土壤微生物活性, 使得土壤呼吸随盐度的升高而降低(Wichern et al., 2006; Wong et al., 2008; Iwai et al., 2012; 李凤霞等, 2012); 同时, 高的含盐量会降低微生物数量(Garcia & Hernandez, 1996; Pattnaik et al., 2000; Pivnickova et al., 2010; Kiehn et al., 2013)以及微生物群落的多样性(Baldwin et al., 2006); 并且, 高盐度能使微生物生理形态发生明显变化, 从而使其分解有机质的能力受到强烈影响(Thottathil et al., 2008), 微生物受到盐分的影响时, 一般能通过自身的渗透压调节机制来平衡细胞内的渗透压, 而在高的渗透压条件下, 微生物耗氧速率增加, 但耗氧速率的增加不是为了有机物的降解, 而是为了能够抵御高盐环境对微生物所产生的伤害(Pankhurst et al., 2001; Rietz & Haynes, 2003; 崔有为等, 2004; Thottathil et al., 2008).另一方面, 多数土壤水解酶与氧化还原酶类活性均随盐渍化的水平升高而明显下降(Rietz & Haynes, 2003; 张建锋等, 2005), 低盐分对酶的活性有促进作用, 而高盐分条件下其活性反而下降(Yan et al., 2013). ...
Heterotrophic microbial activities and nutritional status of microbial communities in tropical marsh sediments of different salinities: The effects of phosphorus addition and plant species
1
2010
... 盐分影响着微生物异养呼吸作用, 从而改变土壤呼吸速率.一方面, 盐分抑制土壤微生物活性, 使得土壤呼吸随盐度的升高而降低(Wichern et al., 2006; Wong et al., 2008; Iwai et al., 2012; 李凤霞等, 2012); 同时, 高的含盐量会降低微生物数量(Garcia & Hernandez, 1996; Pattnaik et al., 2000; Pivnickova et al., 2010; Kiehn et al., 2013)以及微生物群落的多样性(Baldwin et al., 2006); 并且, 高盐度能使微生物生理形态发生明显变化, 从而使其分解有机质的能力受到强烈影响(Thottathil et al., 2008), 微生物受到盐分的影响时, 一般能通过自身的渗透压调节机制来平衡细胞内的渗透压, 而在高的渗透压条件下, 微生物耗氧速率增加, 但耗氧速率的增加不是为了有机物的降解, 而是为了能够抵御高盐环境对微生物所产生的伤害(Pankhurst et al., 2001; Rietz & Haynes, 2003; 崔有为等, 2004; Thottathil et al., 2008).另一方面, 多数土壤水解酶与氧化还原酶类活性均随盐渍化的水平升高而明显下降(Rietz & Haynes, 2003; 张建锋等, 2005), 低盐分对酶的活性有促进作用, 而高盐分条件下其活性反而下降(Yan et al., 2013). ...
Capacity of fatty acid profiles and substrate utilization patterns to describe differences in soil microbial communities associated with increased salinity or alkalinity at three locations in South Australia
1
2001
... 土壤呼吸作为陆地生态系统碳循环的重要环节(Raich & Schlesinger, 1992), 是陆地生态系统碳释放的主要途径, 可达总释放量的2/3 (Davidson et al., 2006).全球气候变暖会促进土壤呼吸(Fang & Moncrieff, 2001; Fouche et al., 2014), 进而使大气CO2浓度增加, 在陆地生态系统和大气之间产生一个强烈的正反馈作用(Cox et al., 2000; Davidson et al., 2000), 使全球变暖的情况更趋严重(Peterjohn et al., 1994; Carney et al., 2007).模拟研究表明, 2016- 2035年全球平均地表温度将升高0.3-0.7 ℃, 2018- 2100年将升高0.3-4.8 ℃ (IPCC, 2013).在全球尺度上, 冬季和春季增温速度较快, 而夏季和秋季的增温速率明显低于冬季和春季以及年平均增温速率(Nagato & Tanakab, 2012).北半球高纬度和高海拔地区温度升幅更大, 而且冬季被认为是增温幅度最大的季节(IPCC, 2013).近50年中, 中国年、季的全国平均气温均表现出显著增高趋势, 而冬季和春季地表增温最为显著, 其中冬季增幅最大, 为0.03 ℃·a-1 (蔡福等, 2006).因此, 研究增温特别是冬季增温对土壤呼吸的影响对正确评估全球变化背景下的陆地生态系统碳循环具有重要意义. ...
The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate
2
1992
... 盐分影响着微生物异养呼吸作用, 从而改变土壤呼吸速率.一方面, 盐分抑制土壤微生物活性, 使得土壤呼吸随盐度的升高而降低(Wichern et al., 2006; Wong et al., 2008; Iwai et al., 2012; 李凤霞等, 2012); 同时, 高的含盐量会降低微生物数量(Garcia & Hernandez, 1996; Pattnaik et al., 2000; Pivnickova et al., 2010; Kiehn et al., 2013)以及微生物群落的多样性(Baldwin et al., 2006); 并且, 高盐度能使微生物生理形态发生明显变化, 从而使其分解有机质的能力受到强烈影响(Thottathil et al., 2008), 微生物受到盐分的影响时, 一般能通过自身的渗透压调节机制来平衡细胞内的渗透压, 而在高的渗透压条件下, 微生物耗氧速率增加, 但耗氧速率的增加不是为了有机物的降解, 而是为了能够抵御高盐环境对微生物所产生的伤害(Pankhurst et al., 2001; Rietz & Haynes, 2003; 崔有为等, 2004; Thottathil et al., 2008).另一方面, 多数土壤水解酶与氧化还原酶类活性均随盐渍化的水平升高而明显下降(Rietz & Haynes, 2003; 张建锋等, 2005), 低盐分对酶的活性有促进作用, 而高盐分条件下其活性反而下降(Yan et al., 2013). ...
... 众多学者认为是由于温度升高影响了微生物的代谢活性, 从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004).冬季植物根系呼吸显著减少, 土壤呼吸主要是微生物的呼吸(Schindlbacher et al., 2007; Ruehr & Buchmann, 2010).增温一方面会增加土壤无机氮库(Luo et al., 2001; Rustad et al., 2001; Melillo et al., 2002), 而无机氮库的增加在一定程度上弥补了冬季活性养分的缺失, 从而促进了微生物的生长; 另一方面, 增温能够提高土壤微生物和酶的活性(Bokhorst et al., 2010), 从而提高土壤中CO2的释放速率.有研究发现, 冬季的土壤呼吸平均能释放掉生态系统在生长季所固存碳的50% (Brooks et al., 2004), 在此情况下, 温度的升高将进一步导致土壤向大气排放更多CO2.因此, 气候变化引起的冬季土壤呼吸速率的升高可能会显著降低高海拔、高纬度生态系统的碳储量(Melillo et al., 2002). ...
Soil respiration fluxes in a temperate mixed forest: Seasonality and temperature sensitivities differ among microbial and root-rhizosphere respiration
3
2010
... 野外自然条件下的增温实验是研究全球变化重要的信息来源(徐振峰等, 2010), 是研究全球变暖与陆地生态系统相互关系的一个主要方法(牛书丽等, 2007; Hoeppner & Dukes, 2012; Hou et al., 2013), 不仅能够获取气候变暖对生态系统影响的直接证据, 还可以解释陆地生态系统对气候变化响应的内在机制(Hou et al., 2013).目前, 有关全球变暖对陆地生态系统影响的增温装置主要有被动增温和主动增温两类.其中, 红外辐射器能更真实有效地模拟全球变暖, 对土壤无物理干扰, 也不改变小气候状况, 是现有的模拟增温的理想装置, 近年来被广泛应用于森林、草地、苔原等生态系统(牛书丽等, 2007).增温影响微生物或根系的代谢活性从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004), 大多数研究表明增温明显促进了土壤呼吸(Lin et al., 2001; Wan et al., 2007; Zhou et al., 2007; Xia et al., 2009; Wang et al., 2014).然而, 也有研究表明随着温度的升高, 土壤呼吸速率出现降低的趋势(Pajari, 1995; Saleska et al., 1999; Melillo et al., 2002; Yin et al., 2013)或不变(Wan et al., 2007), 这可能是因为土壤有限的活性碳库(Oechel et al., 2000; Giardina & Ryan, 2000; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005)和土壤呼吸温度适应性的存在 (Luo et al., 2001). ...
... 众多学者认为是由于温度升高影响了微生物的代谢活性, 从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004).冬季植物根系呼吸显著减少, 土壤呼吸主要是微生物的呼吸(Schindlbacher et al., 2007; Ruehr & Buchmann, 2010).增温一方面会增加土壤无机氮库(Luo et al., 2001; Rustad et al., 2001; Melillo et al., 2002), 而无机氮库的增加在一定程度上弥补了冬季活性养分的缺失, 从而促进了微生物的生长; 另一方面, 增温能够提高土壤微生物和酶的活性(Bokhorst et al., 2010), 从而提高土壤中CO2的释放速率.有研究发现, 冬季的土壤呼吸平均能释放掉生态系统在生长季所固存碳的50% (Brooks et al., 2004), 在此情况下, 温度的升高将进一步导致土壤向大气排放更多CO2.因此, 气候变化引起的冬季土壤呼吸速率的升高可能会显著降低高海拔、高纬度生态系统的碳储量(Melillo et al., 2002). ...
... 这是由于土壤活性碳库是有限的, 当这部分碳释放消耗后, 增温不再刺激土壤呼吸(Melillo et al., 2002); 同时, 随着温度的进一步升高或较高温度持续时间的延长, 呼吸底物的有效性降低(Atkin & Tjoeller, 2003; Bradford et al., 2008; Yuste et al., 2010), 从而降低了土壤呼吸温度敏感性, 进而减缓土壤呼吸随温度升高而增加的量(Jarvis & Linder, 2000; Giardina & Ryan, 2000; Oechel et al., 2000; Luo et al., 2001; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005); 而且, 长期增温可能改变了土壤酶的活性和微生物的种群结构, 从而使土壤呼吸速率降低(Grogan & Jonasson, 2005; Bradford et al., 2008; Hartley et al., 2008); 此外, 因增温引起的其他生态因子的变化也能引起土壤呼吸速率的降低(陈全胜等, 2003a), 这样导致的结果是潜在地减弱了增温-土壤呼吸-大气CO2浓度这个环节的正反馈(Luo et al., 2001). ...
A meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming
4
2001
... 野外自然条件下的增温实验是研究全球变化重要的信息来源(徐振峰等, 2010), 是研究全球变暖与陆地生态系统相互关系的一个主要方法(牛书丽等, 2007; Hoeppner & Dukes, 2012; Hou et al., 2013), 不仅能够获取气候变暖对生态系统影响的直接证据, 还可以解释陆地生态系统对气候变化响应的内在机制(Hou et al., 2013).目前, 有关全球变暖对陆地生态系统影响的增温装置主要有被动增温和主动增温两类.其中, 红外辐射器能更真实有效地模拟全球变暖, 对土壤无物理干扰, 也不改变小气候状况, 是现有的模拟增温的理想装置, 近年来被广泛应用于森林、草地、苔原等生态系统(牛书丽等, 2007).增温影响微生物或根系的代谢活性从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004), 大多数研究表明增温明显促进了土壤呼吸(Lin et al., 2001; Wan et al., 2007; Zhou et al., 2007; Xia et al., 2009; Wang et al., 2014).然而, 也有研究表明随着温度的升高, 土壤呼吸速率出现降低的趋势(Pajari, 1995; Saleska et al., 1999; Melillo et al., 2002; Yin et al., 2013)或不变(Wan et al., 2007), 这可能是因为土壤有限的活性碳库(Oechel et al., 2000; Giardina & Ryan, 2000; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005)和土壤呼吸温度适应性的存在 (Luo et al., 2001). ...
... ; Saleska et al., 1999; Melillo et al., 2002; Yin et al., 2013)或不变(Wan et al., 2007), 这可能是因为土壤有限的活性碳库(Oechel et al., 2000; Giardina & Ryan, 2000; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005)和土壤呼吸温度适应性的存在 (Luo et al., 2001). ...
... 众多学者认为是由于温度升高影响了微生物的代谢活性, 从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004).冬季植物根系呼吸显著减少, 土壤呼吸主要是微生物的呼吸(Schindlbacher et al., 2007; Ruehr & Buchmann, 2010).增温一方面会增加土壤无机氮库(Luo et al., 2001; Rustad et al., 2001; Melillo et al., 2002), 而无机氮库的增加在一定程度上弥补了冬季活性养分的缺失, 从而促进了微生物的生长; 另一方面, 增温能够提高土壤微生物和酶的活性(Bokhorst et al., 2010), 从而提高土壤中CO2的释放速率.有研究发现, 冬季的土壤呼吸平均能释放掉生态系统在生长季所固存碳的50% (Brooks et al., 2004), 在此情况下, 温度的升高将进一步导致土壤向大气排放更多CO2.因此, 气候变化引起的冬季土壤呼吸速率的升高可能会显著降低高海拔、高纬度生态系统的碳储量(Melillo et al., 2002). ...
... 然而, 也有研究表明, 土壤呼吸与增温的关系并没有固定的反应模式, 土壤呼吸速率随着土壤温度的升高也可能会降低(Pajari, 1995; Saleska et al., 1999)或者不变(Wan et al., 2007).本研究中, 整个非生长季增温对土壤呼吸速率无显著影响, 并且在盐分限制阶段, 增温使土壤呼吸速率降低(图3).这与以下研究结果相似: 美国Harvard森林在试验前6年, 温度升高使CO2通量平均增加了28%, 而在后4年, 温度升高对土壤呼吸的影响明显降低(Melillo et al., 2002); 长江口崇明东滩围垦湿地生态系统中, 非生长季温度升高0.66 ℃时, 土壤呼吸速率降低了16% (Zhong & Du, 2013). ...
The effect of experimental ecosystem warming on CO2 fluxes in a montane meadow
1
1999
... 湿地生态系统在全球碳收支平衡中扮演着重要角色(Huntingford et al., 2009).一方面, 湿地具有较高的初级生产力, 因而具有较强的固碳能力(Gorham, 1991).另一方面, 全球变暖背景下, 温度升高促进土壤有机碳分解, 可能会使湿地碳库由碳汇变为碳源(孔雨光等, 2009).然而, 与其他生态系统类型相比, 增温对湿地生态系统土壤呼吸的研究相对较少.同时, 目前土壤呼吸测定大多集中在生长季, 对年土壤呼吸量的估算大多基于非生长季土壤呼吸为0的假设(Grogan & Jonasson, 2006).然而已有研究表明土壤呼吸是全年性的过程, 非生长季土壤呼吸不仅不为0, 而且能占到年土壤呼吸总量的14%-30% (Jones, 1999), 同时冬季会释放生长季固存碳的50%甚至更多(Oechel et al., 2000; Brooks et al., 2004; Monson, 2005), 是区域碳收支非常重要的组成部分(Wickland et al., 2001; Schimel et al., 2006; Han et al., 2012), 能够显著地影响生态系统的碳平衡(Hubbard et al., 2005; Wang et al., 2014).因此, 研究模拟增温对非生长季土壤呼吸很有必要.同时, 湿地生态系统典型样地一般都分布在偏远地区, 且非生长季观测期间易受雪霜以及低温条件的限制, 环境条件恶劣, 因此相关的研究开展得并不多, 是湿地生态系统碳通量研究的薄弱环节. ...
Cold-season production of CO2 in arctic soils: Can laboratory and field estimates be reconciled through a simple modeling approach?
1
2006
... 众多学者认为是由于温度升高影响了微生物的代谢活性, 从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004).冬季植物根系呼吸显著减少, 土壤呼吸主要是微生物的呼吸(Schindlbacher et al., 2007; Ruehr & Buchmann, 2010).增温一方面会增加土壤无机氮库(Luo et al., 2001; Rustad et al., 2001; Melillo et al., 2002), 而无机氮库的增加在一定程度上弥补了冬季活性养分的缺失, 从而促进了微生物的生长; 另一方面, 增温能够提高土壤微生物和酶的活性(Bokhorst et al., 2010), 从而提高土壤中CO2的释放速率.有研究发现, 冬季的土壤呼吸平均能释放掉生态系统在生长季所固存碳的50% (Brooks et al., 2004), 在此情况下, 温度的升高将进一步导致土壤向大气排放更多CO2.因此, 气候变化引起的冬季土壤呼吸速率的升高可能会显著降低高海拔、高纬度生态系统的碳储量(Melillo et al., 2002). ...
Winter soil respiration from an Austrian mountain forest
2
2007
... 滨海湿地作为湿地生态系统的重要类型, 其地下水位浅且受淡咸水交互作用(Fan et al., 2011; Zhong & Du, 2013), 温度升高增强土壤水分蒸发, 带动浅层地下水可溶性盐向地表输送(Yao & Yang, 2010; Zhang et al., 2011), 从而引起土壤表层盐分的变化, 而盐分的升高会影响土壤呼吸(Wichern et al., 2006; Yang et al., 2009; Setia et al., 2011), 然而关于温度、盐分是如何影响滨海湿地土壤呼吸的, 目前还没有明确结论.因此本研究选取黄河三角洲滨海湿地为研究对象, 采用红外辐射加热器模拟土壤增温, 对非生长季(2014年11月-2015年4月)的土壤呼吸速率、土壤温湿度和土壤盐分等进行监测.分析增温对非生长季土壤呼吸日变化及季节变化的影响, 并探讨土壤呼吸对土壤温度、盐分变化的响应机制, 可为了解未来气候变暖对土壤碳循环的影响规律提供基础数据和理论依据. ...
... 增温不仅能直接影响土壤呼吸, 也能通过影响土壤盐分进而影响土壤呼吸.黄河三角洲滨海湿地咸淡水交互作用明显, 且地下水位浅(Fan et al., 2011; Zhong & Du, 2013), 当土壤温度升高时, 土壤水分的蒸发加速, 从而促进了地下咸水向土壤表面的输送(Wichern et al., 2006; Setia et al., 2011), 导致的结果是土壤表层盐分含量高.因此, 盐度也可能是影响滨海湿地CO2产生与排放的重要环境因子(仝川等, 2011).本研究中, 在盐分限制阶段, 增温处理下土壤盐分抑制了滨海湿地土壤呼吸速率(图3).很多研究表明, 随着土壤含盐量的增加, 土壤呼吸速率呈现下降的趋势(Setia et al., 2010; Wong et al., 2010; Hugler & Sievert, 2011; Yan et al., 2013).例如, 海水入侵淡水潮汐沼泽时, 高盐度水平下土壤CO2产生速率随着盐度的升高而减小(Marton et al., 2012); 长江口滩涂土壤呼吸呈现出近岸水体年平均盐度越高, 土壤呼吸速率越低的趋势, 即距海越近土壤呼吸速率越小(聂明华等, 2011); 珠江三角地区近陆红树林湿地CO2排放通量显著高于近海红树林湿地, 这可能是由土壤盐分的差异性引起的(Chen et al., 2010); 当土壤盐分含量比对照高出4倍时, 南澳大利亚莫纳托湿地土壤呼吸速率降低20% (Hasbullah & Petra, 2015).另外, 盐分还可能导致土壤碳源、汇的转化, 例如美国Nueces三角洲湿地在土壤含水量较高、土壤盐度较低的情况下表现为吸收CO2, 而在土壤含水量较低、土壤盐度较高的情况下则表现为排放CO2 (Heinsch et al., 2004). ...
Relationships between carbon dioxide emission and soil properties in salt-affected landscapes
1
2011
... 增温不仅能直接影响土壤呼吸, 也能通过影响土壤盐分进而影响土壤呼吸.黄河三角洲滨海湿地咸淡水交互作用明显, 且地下水位浅(Fan et al., 2011; Zhong & Du, 2013), 当土壤温度升高时, 土壤水分的蒸发加速, 从而促进了地下咸水向土壤表面的输送(Wichern et al., 2006; Setia et al., 2011), 导致的结果是土壤表层盐分含量高.因此, 盐度也可能是影响滨海湿地CO2产生与排放的重要环境因子(仝川等, 2011).本研究中, 在盐分限制阶段, 增温处理下土壤盐分抑制了滨海湿地土壤呼吸速率(图3).很多研究表明, 随着土壤含盐量的增加, 土壤呼吸速率呈现下降的趋势(Setia et al., 2010; Wong et al., 2010; Hugler & Sievert, 2011; Yan et al., 2013).例如, 海水入侵淡水潮汐沼泽时, 高盐度水平下土壤CO2产生速率随着盐度的升高而减小(Marton et al., 2012); 长江口滩涂土壤呼吸呈现出近岸水体年平均盐度越高, 土壤呼吸速率越低的趋势, 即距海越近土壤呼吸速率越小(聂明华等, 2011); 珠江三角地区近陆红树林湿地CO2排放通量显著高于近海红树林湿地, 这可能是由土壤盐分的差异性引起的(Chen et al., 2010); 当土壤盐分含量比对照高出4倍时, 南澳大利亚莫纳托湿地土壤呼吸速率降低20% (Hasbullah & Petra, 2015).另外, 盐分还可能导致土壤碳源、汇的转化, 例如美国Nueces三角洲湿地在土壤含水量较高、土壤盐度较低的情况下表现为吸收CO2, 而在土壤含水量较低、土壤盐度较高的情况下则表现为排放CO2 (Heinsch et al., 2004). ...
Is CO2 evolution in saline soils affected by an osmotic effect and calcium carbonate?
2
2010
... 盐分影响着微生物异养呼吸作用, 从而改变土壤呼吸速率.一方面, 盐分抑制土壤微生物活性, 使得土壤呼吸随盐度的升高而降低(Wichern et al., 2006; Wong et al., 2008; Iwai et al., 2012; 李凤霞等, 2012); 同时, 高的含盐量会降低微生物数量(Garcia & Hernandez, 1996; Pattnaik et al., 2000; Pivnickova et al., 2010; Kiehn et al., 2013)以及微生物群落的多样性(Baldwin et al., 2006); 并且, 高盐度能使微生物生理形态发生明显变化, 从而使其分解有机质的能力受到强烈影响(Thottathil et al., 2008), 微生物受到盐分的影响时, 一般能通过自身的渗透压调节机制来平衡细胞内的渗透压, 而在高的渗透压条件下, 微生物耗氧速率增加, 但耗氧速率的增加不是为了有机物的降解, 而是为了能够抵御高盐环境对微生物所产生的伤害(Pankhurst et al., 2001; Rietz & Haynes, 2003; 崔有为等, 2004; Thottathil et al., 2008).另一方面, 多数土壤水解酶与氧化还原酶类活性均随盐渍化的水平升高而明显下降(Rietz & Haynes, 2003; 张建锋等, 2005), 低盐分对酶的活性有促进作用, 而高盐分条件下其活性反而下降(Yan et al., 2013). ...
... ; Thottathil et al., 2008).另一方面, 多数土壤水解酶与氧化还原酶类活性均随盐渍化的水平升高而明显下降(Rietz & Haynes, 2003; 张建锋等, 2005), 低盐分对酶的活性有促进作用, 而高盐分条件下其活性反而下降(Yan et al., 2013). ...
Tidal switch on metabolic activity: Salinity induced responses on bacterioplankton metabolic capabilities in a tropical estuary. Estuarine,
1
2008
... 增温不仅能直接影响土壤呼吸, 也能通过影响土壤盐分进而影响土壤呼吸.黄河三角洲滨海湿地咸淡水交互作用明显, 且地下水位浅(Fan et al., 2011; Zhong & Du, 2013), 当土壤温度升高时, 土壤水分的蒸发加速, 从而促进了地下咸水向土壤表面的输送(Wichern et al., 2006; Setia et al., 2011), 导致的结果是土壤表层盐分含量高.因此, 盐度也可能是影响滨海湿地CO2产生与排放的重要环境因子(仝川等, 2011).本研究中, 在盐分限制阶段, 增温处理下土壤盐分抑制了滨海湿地土壤呼吸速率(图3).很多研究表明, 随着土壤含盐量的增加, 土壤呼吸速率呈现下降的趋势(Setia et al., 2010; Wong et al., 2010; Hugler & Sievert, 2011; Yan et al., 2013).例如, 海水入侵淡水潮汐沼泽时, 高盐度水平下土壤CO2产生速率随着盐度的升高而减小(Marton et al., 2012); 长江口滩涂土壤呼吸呈现出近岸水体年平均盐度越高, 土壤呼吸速率越低的趋势, 即距海越近土壤呼吸速率越小(聂明华等, 2011); 珠江三角地区近陆红树林湿地CO2排放通量显著高于近海红树林湿地, 这可能是由土壤盐分的差异性引起的(Chen et al., 2010); 当土壤盐分含量比对照高出4倍时, 南澳大利亚莫纳托湿地土壤呼吸速率降低20% (Hasbullah & Petra, 2015).另外, 盐分还可能导致土壤碳源、汇的转化, 例如美国Nueces三角洲湿地在土壤含水量较高、土壤盐度较低的情况下表现为吸收CO2, 而在土壤含水量较低、土壤盐度较高的情况下则表现为排放CO2 (Heinsch et al., 2004). ...
闽江河口潮汐沼泽湿地CO2排放通量特征
5
2011
... 野外自然条件下的增温实验是研究全球变化重要的信息来源(徐振峰等, 2010), 是研究全球变暖与陆地生态系统相互关系的一个主要方法(牛书丽等, 2007; Hoeppner & Dukes, 2012; Hou et al., 2013), 不仅能够获取气候变暖对生态系统影响的直接证据, 还可以解释陆地生态系统对气候变化响应的内在机制(Hou et al., 2013).目前, 有关全球变暖对陆地生态系统影响的增温装置主要有被动增温和主动增温两类.其中, 红外辐射器能更真实有效地模拟全球变暖, 对土壤无物理干扰, 也不改变小气候状况, 是现有的模拟增温的理想装置, 近年来被广泛应用于森林、草地、苔原等生态系统(牛书丽等, 2007).增温影响微生物或根系的代谢活性从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004), 大多数研究表明增温明显促进了土壤呼吸(Lin et al., 2001; Wan et al., 2007; Zhou et al., 2007; Xia et al., 2009; Wang et al., 2014).然而, 也有研究表明随着温度的升高, 土壤呼吸速率出现降低的趋势(Pajari, 1995; Saleska et al., 1999; Melillo et al., 2002; Yin et al., 2013)或不变(Wan et al., 2007), 这可能是因为土壤有限的活性碳库(Oechel et al., 2000; Giardina & Ryan, 2000; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005)和土壤呼吸温度适应性的存在 (Luo et al., 2001). ...
... )或不变(Wan et al., 2007), 这可能是因为土壤有限的活性碳库(Oechel et al., 2000; Giardina & Ryan, 2000; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005)和土壤呼吸温度适应性的存在 (Luo et al., 2001). ...
... 然而, 也有研究表明, 土壤呼吸与增温的关系并没有固定的反应模式, 土壤呼吸速率随着土壤温度的升高也可能会降低(Pajari, 1995; Saleska et al., 1999)或者不变(Wan et al., 2007).本研究中, 整个非生长季增温对土壤呼吸速率无显著影响, 并且在盐分限制阶段, 增温使土壤呼吸速率降低(图3).这与以下研究结果相似: 美国Harvard森林在试验前6年, 温度升高使CO2通量平均增加了28%, 而在后4年, 温度升高对土壤呼吸的影响明显降低(Melillo et al., 2002); 长江口崇明东滩围垦湿地生态系统中, 非生长季温度升高0.66 ℃时, 土壤呼吸速率降低了16% (Zhong & Du, 2013). ...
... 本研究中, 在非盐分限制阶段, 增温使得土壤呼吸温度敏感性降低(表1), 这与众多研究的结果相一致.土壤呼吸的温度敏感性随着土壤温度的升高而下降(Janssens & Pilegaard, 2003; Wan et al., 2007; Zhou et al., 2007; Zhong & Du, 2013).北美高草草原没有加温的实验点的Q10值显著高于加温的实验点的Q10值(Luo et al., 2001; Wan et al., 2007; Zhou et al., 2007); 当温度升高幅度在1.8-3.1 ℃时, 芬兰东部北方针叶林土壤呼吸温度敏感性降低了2.7%- 12.2% (Niinisto et al., 2004); 中国长江口崇明岛盐沼湿地非生长季温度升高0.2 ℃, 土壤呼吸温度敏感性降低13.2% (Zhong & Du, 2013). ...
Responses of soil respiration to elevated CO2, air warming, and changing soil water availability in a model old-field grassland
3
2007
... 野外自然条件下的增温实验是研究全球变化重要的信息来源(徐振峰等, 2010), 是研究全球变暖与陆地生态系统相互关系的一个主要方法(牛书丽等, 2007; Hoeppner & Dukes, 2012; Hou et al., 2013), 不仅能够获取气候变暖对生态系统影响的直接证据, 还可以解释陆地生态系统对气候变化响应的内在机制(Hou et al., 2013).目前, 有关全球变暖对陆地生态系统影响的增温装置主要有被动增温和主动增温两类.其中, 红外辐射器能更真实有效地模拟全球变暖, 对土壤无物理干扰, 也不改变小气候状况, 是现有的模拟增温的理想装置, 近年来被广泛应用于森林、草地、苔原等生态系统(牛书丽等, 2007).增温影响微生物或根系的代谢活性从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004), 大多数研究表明增温明显促进了土壤呼吸(Lin et al., 2001; Wan et al., 2007; Zhou et al., 2007; Xia et al., 2009; Wang et al., 2014).然而, 也有研究表明随着温度的升高, 土壤呼吸速率出现降低的趋势(Pajari, 1995; Saleska et al., 1999; Melillo et al., 2002; Yin et al., 2013)或不变(Wan et al., 2007), 这可能是因为土壤有限的活性碳库(Oechel et al., 2000; Giardina & Ryan, 2000; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005)和土壤呼吸温度适应性的存在 (Luo et al., 2001). ...
... 湿地生态系统在全球碳收支平衡中扮演着重要角色(Huntingford et al., 2009).一方面, 湿地具有较高的初级生产力, 因而具有较强的固碳能力(Gorham, 1991).另一方面, 全球变暖背景下, 温度升高促进土壤有机碳分解, 可能会使湿地碳库由碳汇变为碳源(孔雨光等, 2009).然而, 与其他生态系统类型相比, 增温对湿地生态系统土壤呼吸的研究相对较少.同时, 目前土壤呼吸测定大多集中在生长季, 对年土壤呼吸量的估算大多基于非生长季土壤呼吸为0的假设(Grogan & Jonasson, 2006).然而已有研究表明土壤呼吸是全年性的过程, 非生长季土壤呼吸不仅不为0, 而且能占到年土壤呼吸总量的14%-30% (Jones, 1999), 同时冬季会释放生长季固存碳的50%甚至更多(Oechel et al., 2000; Brooks et al., 2004; Monson, 2005), 是区域碳收支非常重要的组成部分(Wickland et al., 2001; Schimel et al., 2006; Han et al., 2012), 能够显著地影响生态系统的碳平衡(Hubbard et al., 2005; Wang et al., 2014).因此, 研究模拟增温对非生长季土壤呼吸很有必要.同时, 湿地生态系统典型样地一般都分布在偏远地区, 且非生长季观测期间易受雪霜以及低温条件的限制, 环境条件恶劣, 因此相关的研究开展得并不多, 是湿地生态系统碳通量研究的薄弱环节. ...
Soil respiration under climate warming: Differential response of heterotrophic and autotrophic respiration
3
2014
... 滨海湿地作为湿地生态系统的重要类型, 其地下水位浅且受淡咸水交互作用(Fan et al., 2011; Zhong & Du, 2013), 温度升高增强土壤水分蒸发, 带动浅层地下水可溶性盐向地表输送(Yao & Yang, 2010; Zhang et al., 2011), 从而引起土壤表层盐分的变化, 而盐分的升高会影响土壤呼吸(Wichern et al., 2006; Yang et al., 2009; Setia et al., 2011), 然而关于温度、盐分是如何影响滨海湿地土壤呼吸的, 目前还没有明确结论.因此本研究选取黄河三角洲滨海湿地为研究对象, 采用红外辐射加热器模拟土壤增温, 对非生长季(2014年11月-2015年4月)的土壤呼吸速率、土壤温湿度和土壤盐分等进行监测.分析增温对非生长季土壤呼吸日变化及季节变化的影响, 并探讨土壤呼吸对土壤温度、盐分变化的响应机制, 可为了解未来气候变暖对土壤碳循环的影响规律提供基础数据和理论依据. ...
... 增温不仅能直接影响土壤呼吸, 也能通过影响土壤盐分进而影响土壤呼吸.黄河三角洲滨海湿地咸淡水交互作用明显, 且地下水位浅(Fan et al., 2011; Zhong & Du, 2013), 当土壤温度升高时, 土壤水分的蒸发加速, 从而促进了地下咸水向土壤表面的输送(Wichern et al., 2006; Setia et al., 2011), 导致的结果是土壤表层盐分含量高.因此, 盐度也可能是影响滨海湿地CO2产生与排放的重要环境因子(仝川等, 2011).本研究中, 在盐分限制阶段, 增温处理下土壤盐分抑制了滨海湿地土壤呼吸速率(图3).很多研究表明, 随着土壤含盐量的增加, 土壤呼吸速率呈现下降的趋势(Setia et al., 2010; Wong et al., 2010; Hugler & Sievert, 2011; Yan et al., 2013).例如, 海水入侵淡水潮汐沼泽时, 高盐度水平下土壤CO2产生速率随着盐度的升高而减小(Marton et al., 2012); 长江口滩涂土壤呼吸呈现出近岸水体年平均盐度越高, 土壤呼吸速率越低的趋势, 即距海越近土壤呼吸速率越小(聂明华等, 2011); 珠江三角地区近陆红树林湿地CO2排放通量显著高于近海红树林湿地, 这可能是由土壤盐分的差异性引起的(Chen et al., 2010); 当土壤盐分含量比对照高出4倍时, 南澳大利亚莫纳托湿地土壤呼吸速率降低20% (Hasbullah & Petra, 2015).另外, 盐分还可能导致土壤碳源、汇的转化, 例如美国Nueces三角洲湿地在土壤含水量较高、土壤盐度较低的情况下表现为吸收CO2, 而在土壤含水量较低、土壤盐度较高的情况下则表现为排放CO2 (Heinsch et al., 2004). ...
... 盐分影响着微生物异养呼吸作用, 从而改变土壤呼吸速率.一方面, 盐分抑制土壤微生物活性, 使得土壤呼吸随盐度的升高而降低(Wichern et al., 2006; Wong et al., 2008; Iwai et al., 2012; 李凤霞等, 2012); 同时, 高的含盐量会降低微生物数量(Garcia & Hernandez, 1996; Pattnaik et al., 2000; Pivnickova et al., 2010; Kiehn et al., 2013)以及微生物群落的多样性(Baldwin et al., 2006); 并且, 高盐度能使微生物生理形态发生明显变化, 从而使其分解有机质的能力受到强烈影响(Thottathil et al., 2008), 微生物受到盐分的影响时, 一般能通过自身的渗透压调节机制来平衡细胞内的渗透压, 而在高的渗透压条件下, 微生物耗氧速率增加, 但耗氧速率的增加不是为了有机物的降解, 而是为了能够抵御高盐环境对微生物所产生的伤害(Pankhurst et al., 2001; Rietz & Haynes, 2003; 崔有为等, 2004; Thottathil et al., 2008).另一方面, 多数土壤水解酶与氧化还原酶类活性均随盐渍化的水平升高而明显下降(Rietz & Haynes, 2003; 张建锋等, 2005), 低盐分对酶的活性有促进作用, 而高盐分条件下其活性反而下降(Yan et al., 2013). ...
Impact of salinity on soil microbial communities and the decomposition of maize in acidic soils
1
2006
... 湿地生态系统在全球碳收支平衡中扮演着重要角色(Huntingford et al., 2009).一方面, 湿地具有较高的初级生产力, 因而具有较强的固碳能力(Gorham, 1991).另一方面, 全球变暖背景下, 温度升高促进土壤有机碳分解, 可能会使湿地碳库由碳汇变为碳源(孔雨光等, 2009).然而, 与其他生态系统类型相比, 增温对湿地生态系统土壤呼吸的研究相对较少.同时, 目前土壤呼吸测定大多集中在生长季, 对年土壤呼吸量的估算大多基于非生长季土壤呼吸为0的假设(Grogan & Jonasson, 2006).然而已有研究表明土壤呼吸是全年性的过程, 非生长季土壤呼吸不仅不为0, 而且能占到年土壤呼吸总量的14%-30% (Jones, 1999), 同时冬季会释放生长季固存碳的50%甚至更多(Oechel et al., 2000; Brooks et al., 2004; Monson, 2005), 是区域碳收支非常重要的组成部分(Wickland et al., 2001; Schimel et al., 2006; Han et al., 2012), 能够显著地影响生态系统的碳平衡(Hubbard et al., 2005; Wang et al., 2014).因此, 研究模拟增温对非生长季土壤呼吸很有必要.同时, 湿地生态系统典型样地一般都分布在偏远地区, 且非生长季观测期间易受雪霜以及低温条件的限制, 环境条件恶劣, 因此相关的研究开展得并不多, 是湿地生态系统碳通量研究的薄弱环节. ...
Carbon gas exchange at a southern Rocky Mountain wetland, 1996- 1998
1
2001
... 增温不仅能直接影响土壤呼吸, 也能通过影响土壤盐分进而影响土壤呼吸.黄河三角洲滨海湿地咸淡水交互作用明显, 且地下水位浅(Fan et al., 2011; Zhong & Du, 2013), 当土壤温度升高时, 土壤水分的蒸发加速, 从而促进了地下咸水向土壤表面的输送(Wichern et al., 2006; Setia et al., 2011), 导致的结果是土壤表层盐分含量高.因此, 盐度也可能是影响滨海湿地CO2产生与排放的重要环境因子(仝川等, 2011).本研究中, 在盐分限制阶段, 增温处理下土壤盐分抑制了滨海湿地土壤呼吸速率(图3).很多研究表明, 随着土壤含盐量的增加, 土壤呼吸速率呈现下降的趋势(Setia et al., 2010; Wong et al., 2010; Hugler & Sievert, 2011; Yan et al., 2013).例如, 海水入侵淡水潮汐沼泽时, 高盐度水平下土壤CO2产生速率随着盐度的升高而减小(Marton et al., 2012); 长江口滩涂土壤呼吸呈现出近岸水体年平均盐度越高, 土壤呼吸速率越低的趋势, 即距海越近土壤呼吸速率越小(聂明华等, 2011); 珠江三角地区近陆红树林湿地CO2排放通量显著高于近海红树林湿地, 这可能是由土壤盐分的差异性引起的(Chen et al., 2010); 当土壤盐分含量比对照高出4倍时, 南澳大利亚莫纳托湿地土壤呼吸速率降低20% (Hasbullah & Petra, 2015).另外, 盐分还可能导致土壤碳源、汇的转化, 例如美国Nueces三角洲湿地在土壤含水量较高、土壤盐度较低的情况下表现为吸收CO2, 而在土壤含水量较低、土壤盐度较高的情况下则表现为排放CO2 (Heinsch et al., 2004). ...
Soil carbon dynamics in saline and sodic soils: A review
1
2010
... 盐分影响着微生物异养呼吸作用, 从而改变土壤呼吸速率.一方面, 盐分抑制土壤微生物活性, 使得土壤呼吸随盐度的升高而降低(Wichern et al., 2006; Wong et al., 2008; Iwai et al., 2012; 李凤霞等, 2012); 同时, 高的含盐量会降低微生物数量(Garcia & Hernandez, 1996; Pattnaik et al., 2000; Pivnickova et al., 2010; Kiehn et al., 2013)以及微生物群落的多样性(Baldwin et al., 2006); 并且, 高盐度能使微生物生理形态发生明显变化, 从而使其分解有机质的能力受到强烈影响(Thottathil et al., 2008), 微生物受到盐分的影响时, 一般能通过自身的渗透压调节机制来平衡细胞内的渗透压, 而在高的渗透压条件下, 微生物耗氧速率增加, 但耗氧速率的增加不是为了有机物的降解, 而是为了能够抵御高盐环境对微生物所产生的伤害(Pankhurst et al., 2001; Rietz & Haynes, 2003; 崔有为等, 2004; Thottathil et al., 2008).另一方面, 多数土壤水解酶与氧化还原酶类活性均随盐渍化的水平升高而明显下降(Rietz & Haynes, 2003; 张建锋等, 2005), 低盐分对酶的活性有促进作用, 而高盐分条件下其活性反而下降(Yan et al., 2013). ...
Salinity and sodi- city effects on respiration and microbial biomass of soil
1
2008
... 野外自然条件下的增温实验是研究全球变化重要的信息来源(徐振峰等, 2010), 是研究全球变暖与陆地生态系统相互关系的一个主要方法(牛书丽等, 2007; Hoeppner & Dukes, 2012; Hou et al., 2013), 不仅能够获取气候变暖对生态系统影响的直接证据, 还可以解释陆地生态系统对气候变化响应的内在机制(Hou et al., 2013).目前, 有关全球变暖对陆地生态系统影响的增温装置主要有被动增温和主动增温两类.其中, 红外辐射器能更真实有效地模拟全球变暖, 对土壤无物理干扰, 也不改变小气候状况, 是现有的模拟增温的理想装置, 近年来被广泛应用于森林、草地、苔原等生态系统(牛书丽等, 2007).增温影响微生物或根系的代谢活性从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004), 大多数研究表明增温明显促进了土壤呼吸(Lin et al., 2001; Wan et al., 2007; Zhou et al., 2007; Xia et al., 2009; Wang et al., 2014).然而, 也有研究表明随着温度的升高, 土壤呼吸速率出现降低的趋势(Pajari, 1995; Saleska et al., 1999; Melillo et al., 2002; Yin et al., 2013)或不变(Wan et al., 2007), 这可能是因为土壤有限的活性碳库(Oechel et al., 2000; Giardina & Ryan, 2000; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005)和土壤呼吸温度适应性的存在 (Luo et al., 2001). ...
Effects of diurnal warming on soil respiration are not equal to the summed effects of day and night warming in a temperate steppe
... 野外自然条件下的增温实验是研究全球变化重要的信息来源(徐振峰等, 2010), 是研究全球变暖与陆地生态系统相互关系的一个主要方法(牛书丽等, 2007; Hoeppner & Dukes, 2012; Hou et al., 2013), 不仅能够获取气候变暖对生态系统影响的直接证据, 还可以解释陆地生态系统对气候变化响应的内在机制(Hou et al., 2013).目前, 有关全球变暖对陆地生态系统影响的增温装置主要有被动增温和主动增温两类.其中, 红外辐射器能更真实有效地模拟全球变暖, 对土壤无物理干扰, 也不改变小气候状况, 是现有的模拟增温的理想装置, 近年来被广泛应用于森林、草地、苔原等生态系统(牛书丽等, 2007).增温影响微生物或根系的代谢活性从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004), 大多数研究表明增温明显促进了土壤呼吸(Lin et al., 2001; Wan et al., 2007; Zhou et al., 2007; Xia et al., 2009; Wang et al., 2014).然而, 也有研究表明随着温度的升高, 土壤呼吸速率出现降低的趋势(Pajari, 1995; Saleska et al., 1999; Melillo et al., 2002; Yin et al., 2013)或不变(Wan et al., 2007), 这可能是因为土壤有限的活性碳库(Oechel et al., 2000; Giardina & Ryan, 2000; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005)和土壤呼吸温度适应性的存在 (Luo et al., 2001). ...
... 增温不仅能直接影响土壤呼吸, 也能通过影响土壤盐分进而影响土壤呼吸.黄河三角洲滨海湿地咸淡水交互作用明显, 且地下水位浅(Fan et al., 2011; Zhong & Du, 2013), 当土壤温度升高时, 土壤水分的蒸发加速, 从而促进了地下咸水向土壤表面的输送(Wichern et al., 2006; Setia et al., 2011), 导致的结果是土壤表层盐分含量高.因此, 盐度也可能是影响滨海湿地CO2产生与排放的重要环境因子(仝川等, 2011).本研究中, 在盐分限制阶段, 增温处理下土壤盐分抑制了滨海湿地土壤呼吸速率(图3).很多研究表明, 随着土壤含盐量的增加, 土壤呼吸速率呈现下降的趋势(Setia et al., 2010; Wong et al., 2010; Hugler & Sievert, 2011; Yan et al., 2013).例如, 海水入侵淡水潮汐沼泽时, 高盐度水平下土壤CO2产生速率随着盐度的升高而减小(Marton et al., 2012); 长江口滩涂土壤呼吸呈现出近岸水体年平均盐度越高, 土壤呼吸速率越低的趋势, 即距海越近土壤呼吸速率越小(聂明华等, 2011); 珠江三角地区近陆红树林湿地CO2排放通量显著高于近海红树林湿地, 这可能是由土壤盐分的差异性引起的(Chen et al., 2010); 当土壤盐分含量比对照高出4倍时, 南澳大利亚莫纳托湿地土壤呼吸速率降低20% (Hasbullah & Petra, 2015).另外, 盐分还可能导致土壤碳源、汇的转化, 例如美国Nueces三角洲湿地在土壤含水量较高、土壤盐度较低的情况下表现为吸收CO2, 而在土壤含水量较低、土壤盐度较高的情况下则表现为排放CO2 (Heinsch et al., 2004). ...
... 盐分影响着微生物异养呼吸作用, 从而改变土壤呼吸速率.一方面, 盐分抑制土壤微生物活性, 使得土壤呼吸随盐度的升高而降低(Wichern et al., 2006; Wong et al., 2008; Iwai et al., 2012; 李凤霞等, 2012); 同时, 高的含盐量会降低微生物数量(Garcia & Hernandez, 1996; Pattnaik et al., 2000; Pivnickova et al., 2010; Kiehn et al., 2013)以及微生物群落的多样性(Baldwin et al., 2006); 并且, 高盐度能使微生物生理形态发生明显变化, 从而使其分解有机质的能力受到强烈影响(Thottathil et al., 2008), 微生物受到盐分的影响时, 一般能通过自身的渗透压调节机制来平衡细胞内的渗透压, 而在高的渗透压条件下, 微生物耗氧速率增加, 但耗氧速率的增加不是为了有机物的降解, 而是为了能够抵御高盐环境对微生物所产生的伤害(Pankhurst et al., 2001; Rietz & Haynes, 2003; 崔有为等, 2004; Thottathil et al., 2008).另一方面, 多数土壤水解酶与氧化还原酶类活性均随盐渍化的水平升高而明显下降(Rietz & Haynes, 2003; 张建锋等, 2005), 低盐分对酶的活性有促进作用, 而高盐分条件下其活性反而下降(Yan et al., 2013). ...
Five year soil respiration reflected soil quality evolution in different forest and grassland vegetation types in the Eastern Loess Plateau of China
... 滨海湿地作为湿地生态系统的重要类型, 其地下水位浅且受淡咸水交互作用(Fan et al., 2011; Zhong & Du, 2013), 温度升高增强土壤水分蒸发, 带动浅层地下水可溶性盐向地表输送(Yao & Yang, 2010; Zhang et al., 2011), 从而引起土壤表层盐分的变化, 而盐分的升高会影响土壤呼吸(Wichern et al., 2006; Yang et al., 2009; Setia et al., 2011), 然而关于温度、盐分是如何影响滨海湿地土壤呼吸的, 目前还没有明确结论.因此本研究选取黄河三角洲滨海湿地为研究对象, 采用红外辐射加热器模拟土壤增温, 对非生长季(2014年11月-2015年4月)的土壤呼吸速率、土壤温湿度和土壤盐分等进行监测.分析增温对非生长季土壤呼吸日变化及季节变化的影响, 并探讨土壤呼吸对土壤温度、盐分变化的响应机制, 可为了解未来气候变暖对土壤碳循环的影响规律提供基础数据和理论依据. ...
Above- and belowground biomass allocation in Tibetan grasslands
1
2009
... 滨海湿地作为湿地生态系统的重要类型, 其地下水位浅且受淡咸水交互作用(Fan et al., 2011; Zhong & Du, 2013), 温度升高增强土壤水分蒸发, 带动浅层地下水可溶性盐向地表输送(Yao & Yang, 2010; Zhang et al., 2011), 从而引起土壤表层盐分的变化, 而盐分的升高会影响土壤呼吸(Wichern et al., 2006; Yang et al., 2009; Setia et al., 2011), 然而关于温度、盐分是如何影响滨海湿地土壤呼吸的, 目前还没有明确结论.因此本研究选取黄河三角洲滨海湿地为研究对象, 采用红外辐射加热器模拟土壤增温, 对非生长季(2014年11月-2015年4月)的土壤呼吸速率、土壤温湿度和土壤盐分等进行监测.分析增温对非生长季土壤呼吸日变化及季节变化的影响, 并探讨土壤呼吸对土壤温度、盐分变化的响应机制, 可为了解未来气候变暖对土壤碳循环的影响规律提供基础数据和理论依据. ...
Quantitative evaluation of soil salinity and its spatial distribution using electromagnetic induction method
1
2010
... 野外自然条件下的增温实验是研究全球变化重要的信息来源(徐振峰等, 2010), 是研究全球变暖与陆地生态系统相互关系的一个主要方法(牛书丽等, 2007; Hoeppner & Dukes, 2012; Hou et al., 2013), 不仅能够获取气候变暖对生态系统影响的直接证据, 还可以解释陆地生态系统对气候变化响应的内在机制(Hou et al., 2013).目前, 有关全球变暖对陆地生态系统影响的增温装置主要有被动增温和主动增温两类.其中, 红外辐射器能更真实有效地模拟全球变暖, 对土壤无物理干扰, 也不改变小气候状况, 是现有的模拟增温的理想装置, 近年来被广泛应用于森林、草地、苔原等生态系统(牛书丽等, 2007).增温影响微生物或根系的代谢活性从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004), 大多数研究表明增温明显促进了土壤呼吸(Lin et al., 2001; Wan et al., 2007; Zhou et al., 2007; Xia et al., 2009; Wang et al., 2014).然而, 也有研究表明随着温度的升高, 土壤呼吸速率出现降低的趋势(Pajari, 1995; Saleska et al., 1999; Melillo et al., 2002; Yin et al., 2013)或不变(Wan et al., 2007), 这可能是因为土壤有限的活性碳库(Oechel et al., 2000; Giardina & Ryan, 2000; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005)和土壤呼吸温度适应性的存在 (Luo et al., 2001). ...
Enhanced root exudation stimulates soil nitrogen transformations in a subalpine coniferous forest under experimental warming
2
2013
... 这是由于土壤活性碳库是有限的, 当这部分碳释放消耗后, 增温不再刺激土壤呼吸(Melillo et al., 2002); 同时, 随着温度的进一步升高或较高温度持续时间的延长, 呼吸底物的有效性降低(Atkin & Tjoeller, 2003; Bradford et al., 2008; Yuste et al., 2010), 从而降低了土壤呼吸温度敏感性, 进而减缓土壤呼吸随温度升高而增加的量(Jarvis & Linder, 2000; Giardina & Ryan, 2000; Oechel et al., 2000; Luo et al., 2001; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005); 而且, 长期增温可能改变了土壤酶的活性和微生物的种群结构, 从而使土壤呼吸速率降低(Grogan & Jonasson, 2005; Bradford et al., 2008; Hartley et al., 2008); 此外, 因增温引起的其他生态因子的变化也能引起土壤呼吸速率的降低(陈全胜等, 2003a), 这样导致的结果是潜在地减弱了增温-土壤呼吸-大气CO2浓度这个环节的正反馈(Luo et al., 2001). ...
Plant soil interactions and acclimation to temperature of microbial-mediated soil respiration may affect predictions of soil CO2 efflux
1
2010
... 盐分影响着微生物异养呼吸作用, 从而改变土壤呼吸速率.一方面, 盐分抑制土壤微生物活性, 使得土壤呼吸随盐度的升高而降低(Wichern et al., 2006; Wong et al., 2008; Iwai et al., 2012; 李凤霞等, 2012); 同时, 高的含盐量会降低微生物数量(Garcia & Hernandez, 1996; Pattnaik et al., 2000; Pivnickova et al., 2010; Kiehn et al., 2013)以及微生物群落的多样性(Baldwin et al., 2006); 并且, 高盐度能使微生物生理形态发生明显变化, 从而使其分解有机质的能力受到强烈影响(Thottathil et al., 2008), 微生物受到盐分的影响时, 一般能通过自身的渗透压调节机制来平衡细胞内的渗透压, 而在高的渗透压条件下, 微生物耗氧速率增加, 但耗氧速率的增加不是为了有机物的降解, 而是为了能够抵御高盐环境对微生物所产生的伤害(Pankhurst et al., 2001; Rietz & Haynes, 2003; 崔有为等, 2004; Thottathil et al., 2008).另一方面, 多数土壤水解酶与氧化还原酶类活性均随盐渍化的水平升高而明显下降(Rietz & Haynes, 2003; 张建锋等, 2005), 低盐分对酶的活性有促进作用, 而高盐分条件下其活性反而下降(Yan et al., 2013). ...
盐分胁迫对杨树苗期生长和土壤酶活性的影响
1
2005
... 滨海湿地作为湿地生态系统的重要类型, 其地下水位浅且受淡咸水交互作用(Fan et al., 2011; Zhong & Du, 2013), 温度升高增强土壤水分蒸发, 带动浅层地下水可溶性盐向地表输送(Yao & Yang, 2010; Zhang et al., 2011), 从而引起土壤表层盐分的变化, 而盐分的升高会影响土壤呼吸(Wichern et al., 2006; Yang et al., 2009; Setia et al., 2011), 然而关于温度、盐分是如何影响滨海湿地土壤呼吸的, 目前还没有明确结论.因此本研究选取黄河三角洲滨海湿地为研究对象, 采用红外辐射加热器模拟土壤增温, 对非生长季(2014年11月-2015年4月)的土壤呼吸速率、土壤温湿度和土壤盐分等进行监测.分析增温对非生长季土壤呼吸日变化及季节变化的影响, 并探讨土壤呼吸对土壤温度、盐分变化的响应机制, 可为了解未来气候变暖对土壤碳循环的影响规律提供基础数据和理论依据. ...
Assessing impact of land uses on land salinization in the Yellow River Delta, China using an integrated and spatial statistical model
5
2011
... 滨海湿地作为湿地生态系统的重要类型, 其地下水位浅且受淡咸水交互作用(Fan et al., 2011; Zhong & Du, 2013), 温度升高增强土壤水分蒸发, 带动浅层地下水可溶性盐向地表输送(Yao & Yang, 2010; Zhang et al., 2011), 从而引起土壤表层盐分的变化, 而盐分的升高会影响土壤呼吸(Wichern et al., 2006; Yang et al., 2009; Setia et al., 2011), 然而关于温度、盐分是如何影响滨海湿地土壤呼吸的, 目前还没有明确结论.因此本研究选取黄河三角洲滨海湿地为研究对象, 采用红外辐射加热器模拟土壤增温, 对非生长季(2014年11月-2015年4月)的土壤呼吸速率、土壤温湿度和土壤盐分等进行监测.分析增温对非生长季土壤呼吸日变化及季节变化的影响, 并探讨土壤呼吸对土壤温度、盐分变化的响应机制, 可为了解未来气候变暖对土壤碳循环的影响规律提供基础数据和理论依据. ...
... 然而, 也有研究表明, 土壤呼吸与增温的关系并没有固定的反应模式, 土壤呼吸速率随着土壤温度的升高也可能会降低(Pajari, 1995; Saleska et al., 1999)或者不变(Wan et al., 2007).本研究中, 整个非生长季增温对土壤呼吸速率无显著影响, 并且在盐分限制阶段, 增温使土壤呼吸速率降低(图3).这与以下研究结果相似: 美国Harvard森林在试验前6年, 温度升高使CO2通量平均增加了28%, 而在后4年, 温度升高对土壤呼吸的影响明显降低(Melillo et al., 2002); 长江口崇明东滩围垦湿地生态系统中, 非生长季温度升高0.66 ℃时, 土壤呼吸速率降低了16% (Zhong & Du, 2013). ...
... 本研究中, 在非盐分限制阶段, 增温使得土壤呼吸温度敏感性降低(表1), 这与众多研究的结果相一致.土壤呼吸的温度敏感性随着土壤温度的升高而下降(Janssens & Pilegaard, 2003; Wan et al., 2007; Zhou et al., 2007; Zhong & Du, 2013).北美高草草原没有加温的实验点的Q10值显著高于加温的实验点的Q10值(Luo et al., 2001; Wan et al., 2007; Zhou et al., 2007); 当温度升高幅度在1.8-3.1 ℃时, 芬兰东部北方针叶林土壤呼吸温度敏感性降低了2.7%- 12.2% (Niinisto et al., 2004); 中国长江口崇明岛盐沼湿地非生长季温度升高0.2 ℃, 土壤呼吸温度敏感性降低13.2% (Zhong & Du, 2013). ...
... ); 中国长江口崇明岛盐沼湿地非生长季温度升高0.2 ℃, 土壤呼吸温度敏感性降低13.2% (Zhong & Du, 2013). ...
... 增温不仅能直接影响土壤呼吸, 也能通过影响土壤盐分进而影响土壤呼吸.黄河三角洲滨海湿地咸淡水交互作用明显, 且地下水位浅(Fan et al., 2011; Zhong & Du, 2013), 当土壤温度升高时, 土壤水分的蒸发加速, 从而促进了地下咸水向土壤表面的输送(Wichern et al., 2006; Setia et al., 2011), 导致的结果是土壤表层盐分含量高.因此, 盐度也可能是影响滨海湿地CO2产生与排放的重要环境因子(仝川等, 2011).本研究中, 在盐分限制阶段, 增温处理下土壤盐分抑制了滨海湿地土壤呼吸速率(图3).很多研究表明, 随着土壤含盐量的增加, 土壤呼吸速率呈现下降的趋势(Setia et al., 2010; Wong et al., 2010; Hugler & Sievert, 2011; Yan et al., 2013).例如, 海水入侵淡水潮汐沼泽时, 高盐度水平下土壤CO2产生速率随着盐度的升高而减小(Marton et al., 2012); 长江口滩涂土壤呼吸呈现出近岸水体年平均盐度越高, 土壤呼吸速率越低的趋势, 即距海越近土壤呼吸速率越小(聂明华等, 2011); 珠江三角地区近陆红树林湿地CO2排放通量显著高于近海红树林湿地, 这可能是由土壤盐分的差异性引起的(Chen et al., 2010); 当土壤盐分含量比对照高出4倍时, 南澳大利亚莫纳托湿地土壤呼吸速率降低20% (Hasbullah & Petra, 2015).另外, 盐分还可能导致土壤碳源、汇的转化, 例如美国Nueces三角洲湿地在土壤含水量较高、土壤盐度较低的情况下表现为吸收CO2, 而在土壤含水量较低、土壤盐度较高的情况下则表现为排放CO2 (Heinsch et al., 2004). ...
Effects of in situ experimental air warming on the soil respiration in a coastal salt marsh reclaimed for agriculture
4
2013
... 野外自然条件下的增温实验是研究全球变化重要的信息来源(徐振峰等, 2010), 是研究全球变暖与陆地生态系统相互关系的一个主要方法(牛书丽等, 2007; Hoeppner & Dukes, 2012; Hou et al., 2013), 不仅能够获取气候变暖对生态系统影响的直接证据, 还可以解释陆地生态系统对气候变化响应的内在机制(Hou et al., 2013).目前, 有关全球变暖对陆地生态系统影响的增温装置主要有被动增温和主动增温两类.其中, 红外辐射器能更真实有效地模拟全球变暖, 对土壤无物理干扰, 也不改变小气候状况, 是现有的模拟增温的理想装置, 近年来被广泛应用于森林、草地、苔原等生态系统(牛书丽等, 2007).增温影响微生物或根系的代谢活性从而提高了土壤呼吸速率(Saleska et al., 1999; Berger et al., 2004), 大多数研究表明增温明显促进了土壤呼吸(Lin et al., 2001; Wan et al., 2007; Zhou et al., 2007; Xia et al., 2009; Wang et al., 2014).然而, 也有研究表明随着温度的升高, 土壤呼吸速率出现降低的趋势(Pajari, 1995; Saleska et al., 1999; Melillo et al., 2002; Yin et al., 2013)或不变(Wan et al., 2007), 这可能是因为土壤有限的活性碳库(Oechel et al., 2000; Giardina & Ryan, 2000; Rustad et al., 2001; Melillo et al., 2002; Eliasson et al., 2005)和土壤呼吸温度适应性的存在 (Luo et al., 2001). ...
... 本研究中, 在非盐分限制阶段, 增温使得土壤呼吸温度敏感性降低(表1), 这与众多研究的结果相一致.土壤呼吸的温度敏感性随着土壤温度的升高而下降(Janssens & Pilegaard, 2003; Wan et al., 2007; Zhou et al., 2007; Zhong & Du, 2013).北美高草草原没有加温的实验点的Q10值显著高于加温的实验点的Q10值(Luo et al., 2001; Wan et al., 2007; Zhou et al., 2007); 当温度升高幅度在1.8-3.1 ℃时, 芬兰东部北方针叶林土壤呼吸温度敏感性降低了2.7%- 12.2% (Niinisto et al., 2004); 中国长江口崇明岛盐沼湿地非生长季温度升高0.2 ℃, 土壤呼吸温度敏感性降低13.2% (Zhong & Du, 2013). ...
... ., 2007; Zhou et al., 2007); 当温度升高幅度在1.8-3.1 ℃时, 芬兰东部北方针叶林土壤呼吸温度敏感性降低了2.7%- 12.2% (Niinisto et al., 2004); 中国长江口崇明岛盐沼湿地非生长季温度升高0.2 ℃, 土壤呼吸温度敏感性降低13.2% (Zhong & Du, 2013). ...
Source components and interannual variability of soil CO2 efflux under experimental warming and clipping in a grassland ecosystem
 , 陈亮
, 陈亮

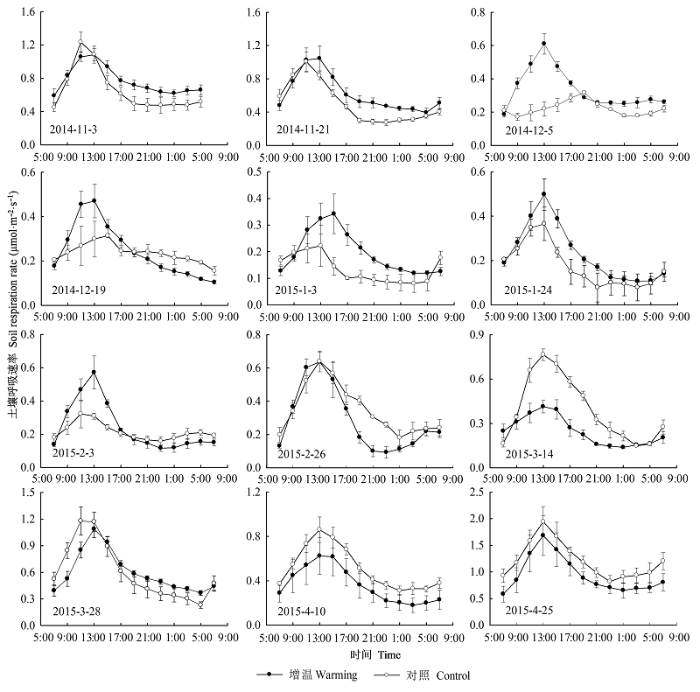
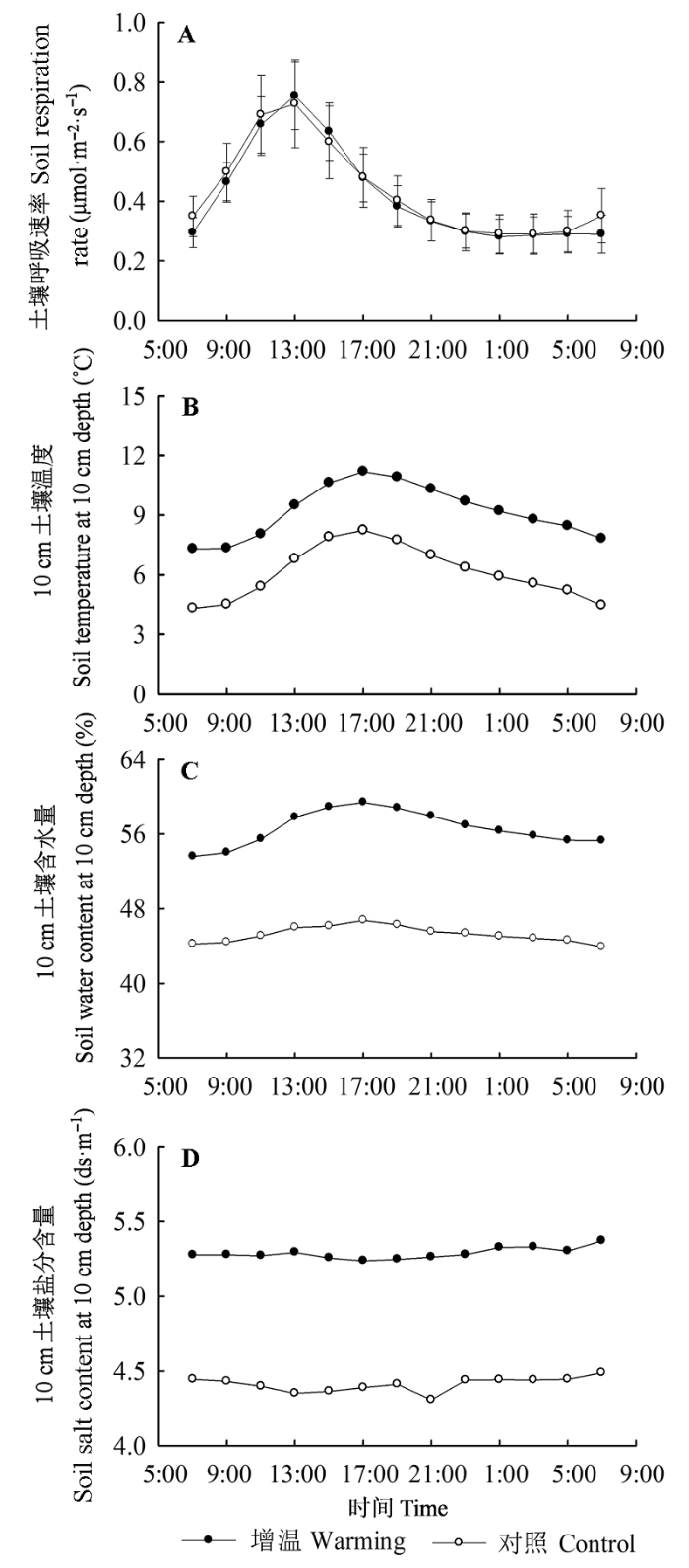
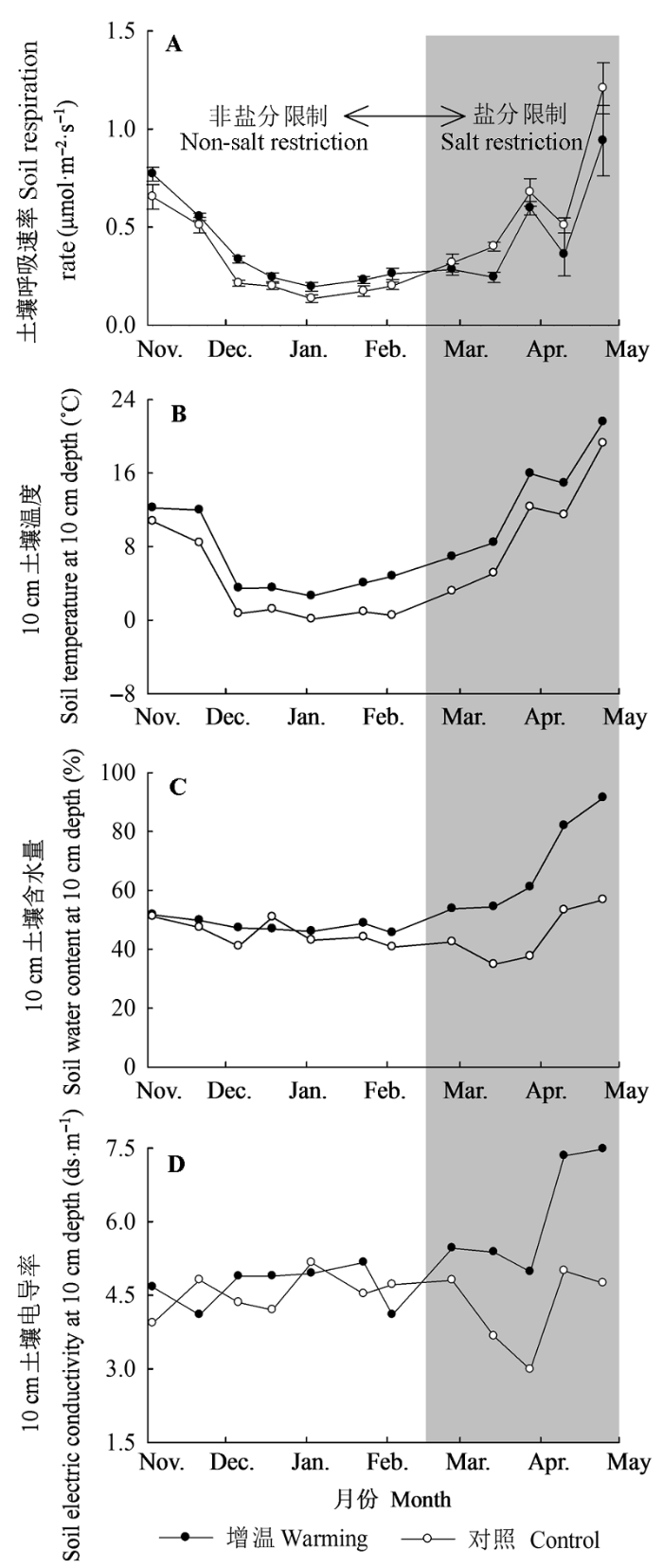
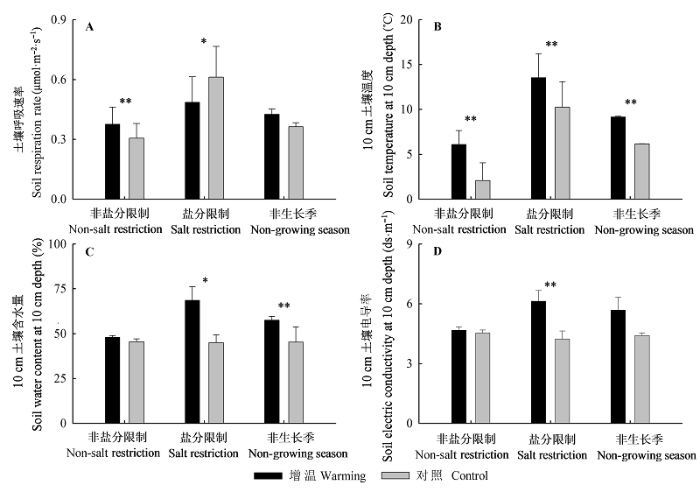
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}