人类活动导致大气氮(N)沉降不断增强(Galloway et al ., 2004 ; Dentener et al ., 2006 ; Mahowald et al ., 2008 ), 正深刻影响着生态系统及其养分循环过程(Vitousek & Tilman, 1997 ; Galloway & Sutton, 2008 ; Treseder, 2008 )。野外人工N添加实验是模拟自然N沉降增加对生态系统影响的重要手段。自20世纪80年代起, 研究者在温带和北方森林地区陆续开展了大量长时间的野外N添加实验(Aber & Melillo, 1989 ; Wright & Rasmussen, 1998 ; Magill et al ., 2000 , 2004 ), 而N添加实验在亚热带常绿阔叶林地区开展得相对较少(Mo et al ., 2008 ; Lu et al ., 2010 )。我国的亚热带常绿阔叶林面积广阔、物种组成丰富(Fang et al ., 2014 ), 是全球亚热带常绿阔叶林的主要分布区, 在我国亚热带常绿阔叶林地区开展N添加的生态学效应的研究十分必要。
在全球变化的背景下, 植物的化学计量特征可以表征植物维持体内化学计量内稳性的能力(Sterner & Elser, 2002 ; Demars & Edwards, 2007 ), 并反映植物对环境变化的适应性(Elser et al ., 2010a )。以往的研究表明, 植物叶片N、磷(P)化学计量特征因植被类型、功能群、物种的差异而不同(Sterner & Elser, 2002 ; Reich, 2003 ; Ågren, 2004 ), 不同功能群和物种对N添加的响应也存在差异(Güsewell, 2004 )。自20世纪80年代起, 研究人员在温带和北方森林地区陆续开展了一些经典的野外N添加实验(Aber & Melillo, 1989 ; Wright & Rasmussen, 1998 ; Magill et al ., 2000 ), 植物化学计量学作为研究N循环的新手段, 也被应用于N添加生态学效应的研究中。例如: Magill等(2004)在美国哈佛森林开展了长期的N添加试验, 经过15年的研究发现: N添加显著增加了乔木优势种的叶片N含量, 并且叶片化学计量特征对N添加的响应存在明显的年际变化; Huang等(2012)在模拟的亚热带常绿阔叶林生态系统中开展了N添加实验, 对群落内5种乔木树种叶片N、P化学计量特征进行了研究, 结果显示N添加显著影响了多数物种的叶片N、P含量和N:P; Cui等(2010)对温带草原8种优势植物叶片化学计量特征对N添加的响应进行了研究, 发现不同物种的叶片化学计量特征对N添加的响应也存在差异。
已有的研究几乎只针对几种优势乔木物种或是同一功能群的物种展开, 对于群落内不同功能群、不同层次物种叶片N、P化学计量特征对N添加的响应缺乏研究。本研究旨在系统地评估群落内不同物种、不同功能群植物叶片化学计量特征对N沉降的响应, 并结合年际变化综合分析N添加的时间效应。
本研究选择武夷山亚热带常绿阔叶林作为研究对象, 通过设立人工N添加野外实验, 在已进行5年的人工N添加实验样地内, 对群落内的优势种以及其他常见种的叶片N、P化学计量特征进行3年的连续观测, 试图回答以下两个问题: 1)武夷山植物叶片N、P化学计量特征对N添加的响应及其年际动态如何? 2)不同功能群物种的叶片N、P化学计量特征对N添加的响应有何差异?
1 材料和方法
1.1 样地概况
N添加样地设在福建武夷山森林生态系统定位观测站(27.71° N, 117.75° E), 海拔630 m, 平均坡度31°, 坡向南偏西10°。该地区为亚热带季风气候, 夏季高温多雨, 冬季温和湿润。年平均气温17.5 ℃, 年平均相对湿度78%-84%, 年降水量1486-2150 mm, 年雾日平均达120天(福建武夷山森林生态定位观测研究站气象观测数据)。实验样地土壤类型为黄壤, 平均土层厚度为60 cm, 样地局部区域可达100 cm以上。土壤pH值4-5。
实验样地为中亚热带常绿阔叶米槠(Cas- tanopsis carlesii )林, 这里植被保存相对完好, 层次分明, 物种丰富。森林垂直结构分为3层: (1)乔木层(>5 m)植物主要有米槠、刨花润楠(Machilus pauhoi )、丝栗栲(Castanopsis fargesii )、罗浮锥(Castan- opsis faberi )、少叶黄杞(Engelhardtia fenzlii )、杉木(Cunninghamia lanceolata )等; (2)灌木层(0.3-5.0 m)植物主要有树参(Dendropanax dentiger )、柃木(Eurya japonica )、弯蒴杜鹃(Rhododendron henryi )、榕叶冬青(Ilex ficoidea )、虎皮楠(Daphniphyllum oldhami )等; (3)草本层(<0.3 m)物种稀少, 主要有砂仁(Amomum villosum )、狗脊(Woodwardia japonica )、绿色白发藓(Leucobryum chlorophyllosum )等。
1.2 实验设计
武夷山N添加实验始于2011年7月, 共设置3个N添加梯度和1个对照, N添加梯度分别为低N (LN, 50 kg Nhm-2 a-1 )、中N (MN, 100 kg Nhm-2 a-1 )和高N (HN, 150 kg Nhm-2 a-1 )。每种处理设3个重复, 共12块样地。样地大小为20 m × 20 m, 重复内相邻样地预留≥5 m的间隔, 不同重复间相距10-15 m。在每一样地内距4条边线留2.5 m边缘带, 样地中心带划分9个5 m × 5 m的小样格, 从底线自西向东依次编号1-9, 在样格内进行调查和采样。N添加安排在每年生长季3-11月进行。每月初, 按照不同处理梯度的要求, 将相应质量的NH4 NO3 溶于30 L水中, 用肩背式喷雾器在实验样地来回均匀地喷洒, 对照样地喷洒相同量的水。
1.3 样品采集
新鲜叶片采集于2013年、2014年和2015年8月初。乔木和灌木利用高枝剪或爬树采集向阳、林冠1/3处优势种的叶片, 林下植物直接用剪刀进行采集。采样时, 在样地内对每个物种随机选取3个成熟个体, 分别采集10-20片生长健康、完全展开的成熟叶片, 每个物种的叶片混合后取50 g左右作为一份样品。
经过优势度分析和筛选, 以下9种植物为群落内各层植物的代表性物种, 选择乔木层植物米槠、刨花润楠、杉木, 灌木层植物树参、柃木、弯蒴杜鹃, 草本层植物砂仁、狗脊(蕨类植物)、绿色白发藓(苔藓植物)作为各层植物的代表。
土壤样品采集于2015年8月初。表层土壤样品(0-5 cm、5-10 cm)用直径5 cm的土钻在每一样地的1、3、5、7、9号样格内采集, 同一块样地的土壤样品同层混合作为一份样品。
1.4 样品测定
将采集的新鲜叶片样品洗净、去叶柄后装入信封, 在65 ℃恒温烘干48 h至恒质量。烘干后的叶片样品用球磨仪磨碎(<0.5 mm), 用于测定叶片的全碳(C)、全N和全P含量。全C和全N含量用元素分析仪(Elementar Vario EL III, Hanau, Germany)测定, 全P含量采用钼锑抗比色法(LY/T 1271-1999)测定。叶片N:P为叶片全N和全P含量的质量比。
土壤样品充分风干后, 去除根系和砾石等, 并过2 mm筛。称取15 g土壤样品, 以50 mL 1 molL-1 的KCl溶液浸提, 用酸度计(pH S-3B, 上海精科, 上海)测定浸提液的pH值。另取一份预处理的土壤样品在65 ℃下烘干48 h, 用球磨仪研磨后, 用元素分析仪测定土壤样品全C、全N含量; 用钼锑抗比色法测定土壤样品全P含量。
1.5 数据分析
采用相对效应指数(RE )表征N添加样地植物叶片化学计量特征对N添加的响应(Duval et al ., 2011 )。计算公式为: 相对效应指数(RE ) = (处理样地值-对照样地值)/对照样地值×100%。
实验数据利用SPSS 20.0进行统计分析。方差分析和多重比较分别采用单因素方差分析法和一般线性模型General Linear Model完成。数据分析显著性水平采用p < 0.05, 绘图在SigmaPlot 12.5中完成。
2 结果
2.1 叶片N含量对N添加的响应
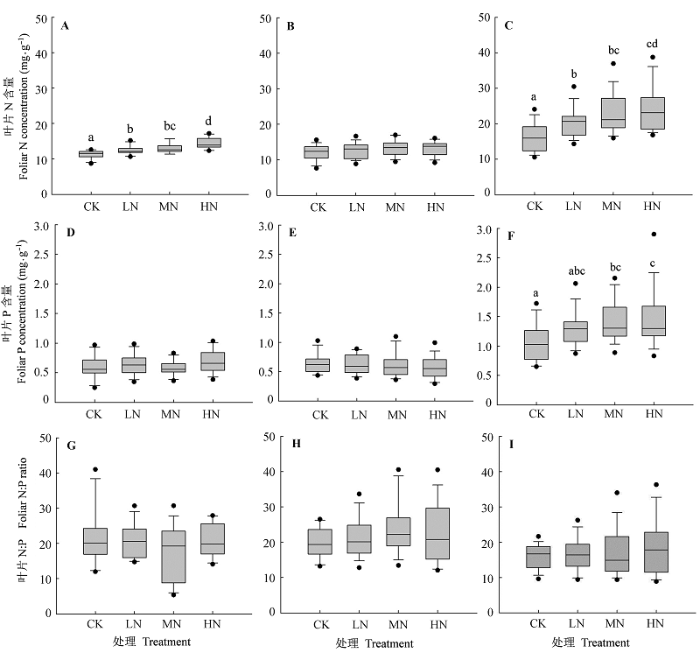
整体来看, N添加显著增加了植物叶片N含量(p < 0.05), 但不同功能群植物叶片N含量对N添加的响应存在差异, 草本层植物叶片N含量对N添加的响应比乔木层和灌木层植物更加敏感(图1 )。
图1 N添加对不同功能群物种叶片N含量、P含量和N:P的影响(平均值±标准误差)。CK、LN、MN和HN指N添加量分别为0、50、100和150 kg Nhm-2 a-1 。不同小写字母表示不同处理间差异显著(p < 0.05)。A, D, G, 乔木层植物。B, E, H, 灌木层植物。C, F, I, 林下草本层植物。
Fig. 1 Effects of N addition on foliar N concentrations, P concentrations and N:P ratios in different functional groups (mean ± SE ). CK, LN, MN and HN indicate that N addition were 0, 50, 100 and 150 kg Nhm-2 a-1 , respectively. Different lowercase letters indicate significant differences between treatments at p < 0.05 levels. A, D, G, Trees. B, E, H, Shrubs. C, F, I, Understory plants.
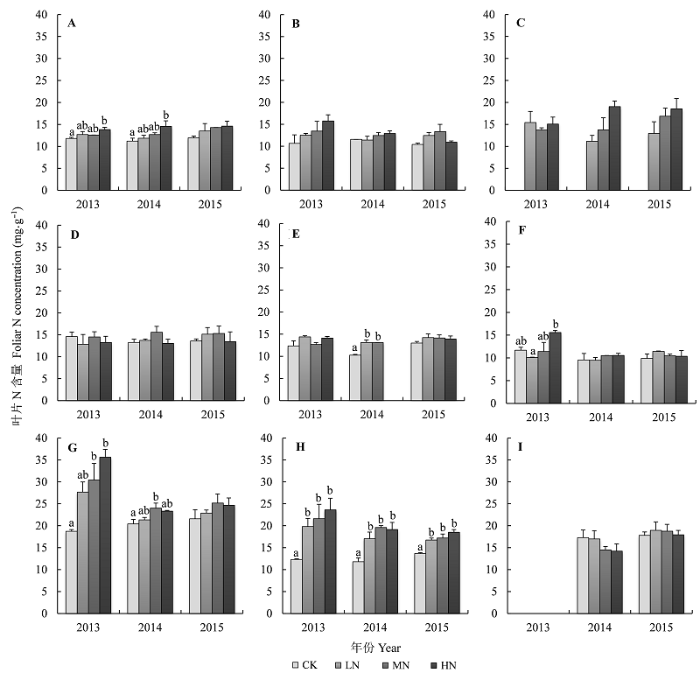
乔木层优势种米槠对N添加响应敏感, 2013年和2014年米槠叶片N含量增加显著(p < 0.05), HN样地米槠叶片N含量均显著高于对照样地, 叶片N含量平均从11.65 mgg-1 增加至14.35 mgg-1 , 增加了23%; 刨花润楠和杉木叶片N含量有增加趋势, 但增加不显著(图2 )。
图2 N添加对植物叶片N含量的影响(平均值±标准误差)。A, 米槠。B, 刨花润楠。C, 杉木。D, 树参。E, 柃木。F, 弯蒴杜鹃。G, 砂仁。H, 狗脊。I, 绿色白发藓。CK、LN、MN和HN指N添加量分别为0、50、100和150 kg Nhm-2 a-1 。不同小写字母表示不同处理间差异显著(p < 0.05)。
Fig. 2 Effects of N addition on foliar N concentrations of nine species (mean ± SE ). A, Castanopsis carlesii . B, Machilus pauhoi . C, Cunninghamia lanceolata . D, Dendropanax dentiger . E, Eurya japonica . F, Rhododendron henryi . G, Amomum villosum . H, Woodwardia japonica . I, Leucobryum chlorophyllosum . CK, LN, MN and HN indicate that N addition were 0, 50, 100 and 150 kg Nhm-2 a-1 , respectively. Different lowercase letters indicate significant differences between treatments at p < 0.05 levels.
N添加整体上对灌木层植物叶片N含量没有显著影响, 但在2014年和2013年分别显著提高了柃木和弯蒴杜鹃叶片的N含量。例如: 2014年N添加导致柃木叶片N含量显著增加(p < 0.05), 由10.28 mgg-1 上升至13.15 mgg-1 ; 在2013年, 与对照相比, 施肥样地弯蒴杜鹃叶片N含量增加了15%- 50% (图2 )。
草本层物种对N添加响应敏感, N添加显著增加了砂仁和狗脊叶片N含量(p < 0.05)。砂仁对N添加的响应在不同年份间具有显著差异, 与对照样地相比, 2013年砂仁叶片N含量增加了45%-90%, 2014年砂仁叶片N含量增加了5%-20%, 而2015年砂仁叶片N含量对N添加没有显著响应。狗脊叶片N含量随着N添加梯度增加而显著上升, 3年平均来看, N添加使得N浓度由1.25 mgg-1 上升到2.04 mgg-1 , 且随着N添加的进行, 不同N添加梯度间叶片N含量分异不断减小。N添加导致绿色白发藓叶片N含量有微弱下降的趋势, 但下降不显著(图2 )。
2.2 叶片P含量对N添加的响应
不同功能群植物叶片P含量对N添加的响应存在差异, 草本层植物叶片P含量显著增加(p < 0.05), 而乔木层植物和灌木层植物叶片P含量没有显著变化(图1 )。
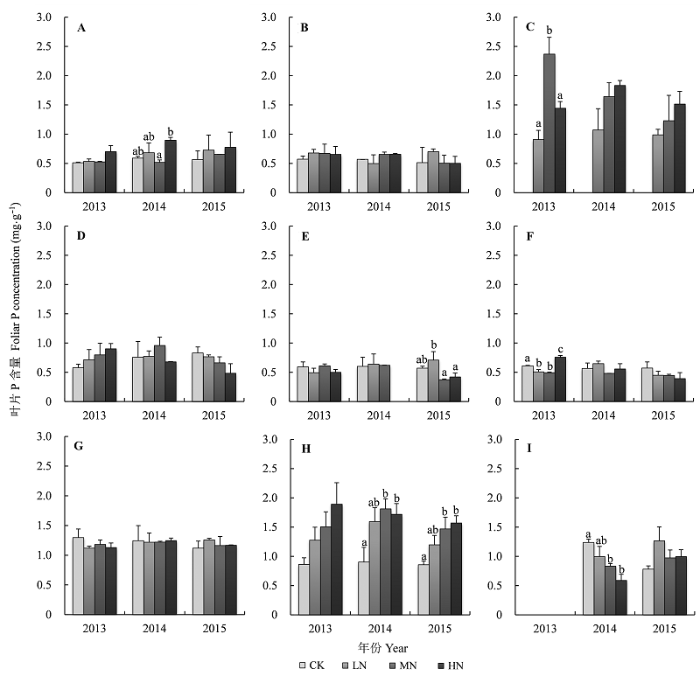
乔木层裸子植物杉木叶片P含量(0.99 mgg-1 )显著高于被子植物米槠(0.55 mgg-1 )和刨花润楠(0.54 mgg-1 )。N添加显著增加了优势种米槠以及杉木的叶片P含量(p < 0.05), 这两个物种叶片P含量随着N添加梯度增加而增加, 并分别在2014年和2013年达到显著水平(p < 0.05); 各年N添加对刨花润楠叶片P含量均无显著影响(图3 )。灌木层常见植物叶片P含量平均为0.61 mgg-1 , 不同物种间叶片P含量无显著差异, 并且P含量对N添加的响应也不敏感(图3 )。
图3 N添加对植物叶片P含量的影响(平均值±标准误差)。A, 米槠。B, 刨花润楠。C, 杉木。D, 树参。E, 柃木。F, 弯蒴杜鹃。G, 砂仁。H, 狗脊。I, 绿色白发藓。CK、LN、MN和HN指N添加量分别为0、50、100和150 kg Nhm-2 a-1 。不同小写字母表示不同处理间差异显著(p < 0.05)。
Fig. 3 Effects of N addition on foliar P concentrations of nine species (mean ± SE ). A, Castanopsis carlesii . B, Machilus pauhoi . C, Cunninghamia lanceolata . D, Dendropanax dentiger . E, Eurya japonica . F, Rhododendron henryi . G, Amomum villosum . H, Woodwardia japonica . I, Leucobryum chlorophyllosum . CK, LN, MN and HN indicate that N addition were 0, 50, 100 and 150 kg Nhm-2 a-1 , respectively. Different lowercase letters indicate significant differences between treatments at p < 0.05 levels.
草本层蕨类植物和苔藓植物叶片P含量对N添加的响应较为敏感。N添加显著增加了狗脊叶片P含量, 由0.87 mgg-1 增至1.73 mgg-1 , 增加了55%- 95%。绿色白发藓却表现出相反的变化趋势, 2014年N添加显著降低了绿色白发藓叶片P含量, 由1.24 mgg-1 降至0.59 mgg-1 , 下降了20%-50% (图3 )。
2.3 叶片N:P对N添加的响应
研究结果表明, N添加对武夷山乔木层植物叶片N:P无显著影响, 且在不同年份间无显著差异, 乔木层植物叶片N:P (18.67)基本保持稳定(图1 )。
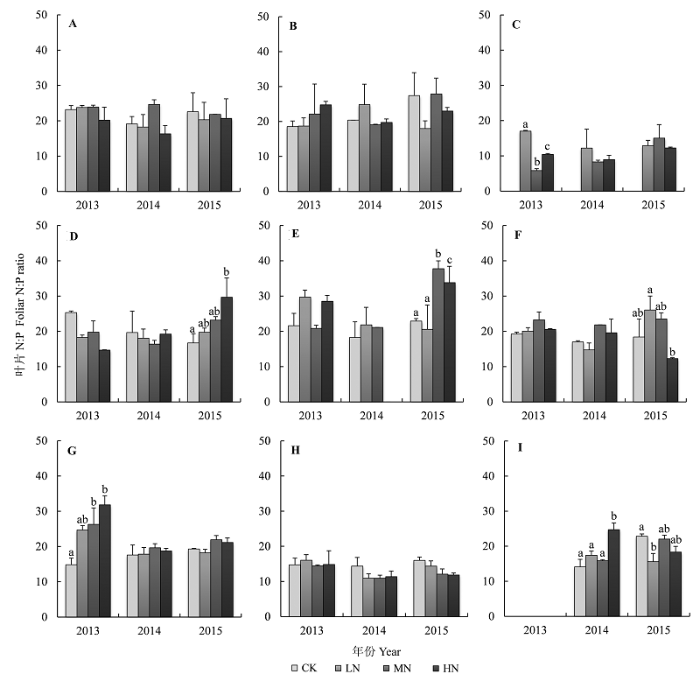
N添加逐步提高了灌木植物叶片N:P。2013年和2014年, N添加对灌木层各植物叶片N:P没有显著影响, 但2015年N添加显著提高了树参和柃木两种灌木层植物叶片N:P (p < 0.05)。例如, 在2015年, 树参和柃木两种植物叶片N:P分别由对照样地(CK)的16.69和22.88增加至HN样地的29.66和37.77 (图4 )。
图4 N添加对植物叶片N:P的影响(平均值±标准误差)。A, 米槠。B, 刨花润楠。C, 杉木。D, 树参。E, 柃木。F, 弯蒴杜鹃。G, 砂仁。H, 狗脊。I, 绿色白发藓。CK、LN、MN和HN指N添加量分别为0、50、100和150 kg Nhm-2 a-1 。不同小写字母表示不同处理间差异显著(p < 0.05)。
Fig. 4 Effects of N addition on foliar N:P ratios of nine species (mean ± SE ). A, Castanopsis carlesii . B, Machilus pauhoi . C, Cunninghamia lanceolata . D, Dendropanax dentiger . E, Eurya japonica . F, Rhododendron henryi . G, Amomum villosum . H, Woodwardia japonica . I, Leucobryum chlorophyllosum . CK, LN, MN and HN indicate that N addition were 0, 50, 100 and 150 kg Nhm-2 a-1 , respectively. Different lowercase letters indicate significant differences between treatments at p < 0.05 levels.
草本植物砂仁和苔藓植物绿色白发藓的叶片N:P对N添加的响应敏感。在2013年, N添加使得砂仁叶片N:P (14.85)增加了60%-110%, 从多年结果来看, 随着N添加量的增加, 砂仁叶片N:P也呈现上升趋势。N添加提高了绿色白发藓叶片N:P, 但个别N添加处理对N:P有降低的效果。狗脊叶片N:P对N添加响应不明显, 基本维持在13.52左右(图4 )。
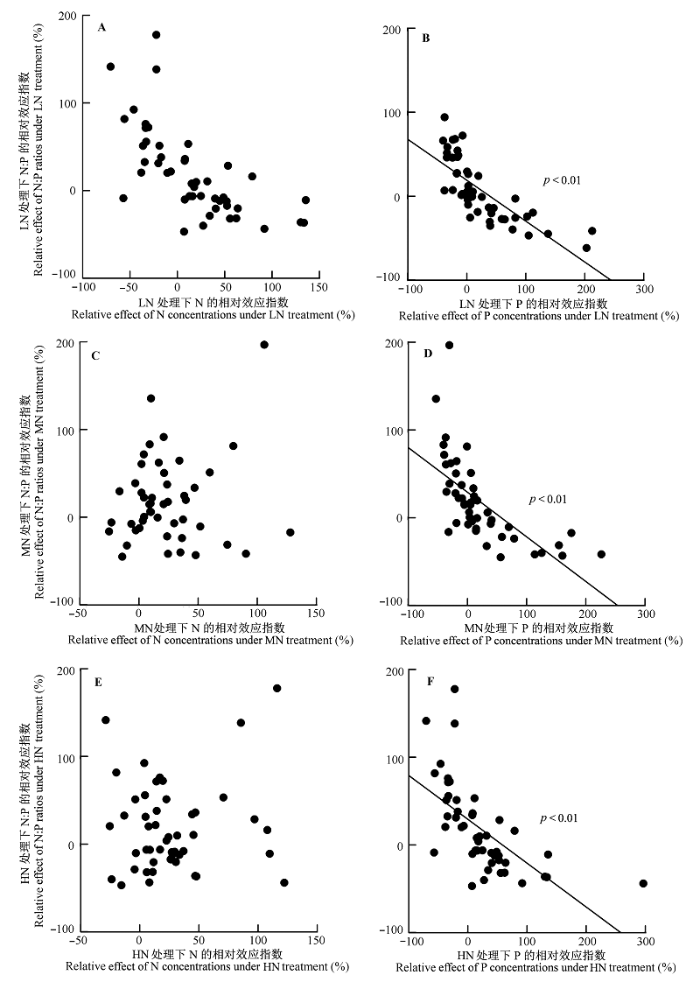
将N添加样地内物种叶片N:P的相对效应指数与其对应的N、P相对效应指数建立相关关系, 结果显示: 武夷山常绿阔叶林植物叶片的N:P相对效应指数与植物叶片的N相对效应指数无显著的相关性, 而与植物叶片的P相对效应指数存在极显著的负相关关系(p < 0.01)(图5 )。这说明N添加样地内植物叶片N:P的变化主要受到叶片P含量变化的影响, 而非叶片N含量变化的影响。
图5 叶片N:P变化与叶片N、P含量变化的关系。HN、LN、MN指N添加量分别为150、50和100 kg N·hm-2 ·a-1 。
Fig. 5 Correlations between the relative effect of foliar N:P ratios and the relative effect of N or P concentrations. HN, LN, MN indicate that N addition were 150, 50 and 100 kg N·hm-2 ·a-1 , respectively.
3 讨论
3.1 N添加对植物叶片N、P元素含量的影响
武夷山米槠群落植物叶片N、P含量平均值低于世界和全国平均水平, 叶片N:P则高于世界和全国平均水平(Reich & Oleksyn, 2004 ; Han et al ., 2005 )。本研究区植物生长受P元素限制更加明显。这与亚热带地区较低的土壤养分含量、较快的养分周转速率以及较强的淋溶等环境特点相适应(方运霆等, 2004 ; Reich & Oleksyn, 2004 ; Lu et al ., 2010 )。
不同功能群物种叶片的N、P含量均存在显著差异, 这与以往的研究结果(Reich & Bowman, 1999 ; Sterner & Elser, 2002 ; Sardans et al ., 2012 )较为一致。本研究显示, 草本层植物叶片的N、P含量显著高于木本植物(包括乔木和灌木植物), 针叶植物叶片的N、P含量显著高于阔叶植物, 这一结果与其他研究中得到的中国草本和木本植物的N、P化学计量模式基本一致(高三平等, 2007 ; He et al ., 2008 ; Han et al ., 2011 ), 并且证明了生命周期短而生长快速的植物要比生命周期长而生长缓慢的植物具有更高的叶片N、P含量(Aerts, 1996 ; Thompson et al ., 1997 ; Güsewell & Koerselman, 2002 )。
N添加整体上促进了植物对N的吸收, 这与以往的研究结果(Magill et al ., 2000 ; Mcnulty et al ., 2005 ; Rowe et al ., 2008 )一致。植物叶片的矿质元素含量取决于土壤养分的可利用性(Townsend & Bustamante, 2007 )。以往的研究也显示, 植物组织内的矿质元素含量与土壤中相应元素含量呈现显著正相关关系(Reich & Oleksyn, 2004 ), 当土壤中某种矿质元素含量增加时, 植物体内相应的矿质元素含量也会随之增加。功能群和物种则是影响叶片N、P化学计量特征及其对N添加响应的重要因素(Reich & Bowman, 1999 ; Sterner & Elser, 2002 ; Sardans et al ., 2012 )。N添加情况下, 植物对N、P等矿质元素的利用策略因需求、竞争力和优势度的不同而存在差异(Tilman, 1990 ; Güsewell & Koerselman, 2002 )。本研究中, 乔木优势种米槠对N添加响应敏感, N添加显著增加了其叶片N含量(图2 )。这可能是由于当养分条件发生变化时, 生态系统趋向于将N、P等矿质元素运输到生产力较高的植物中去, 优先保证其生长需求(Aber & Melillo, 1989 ; Elliott & White, 1994 )。本研究中林下草本层植物N含量对N添加的响应也较为敏感(图2 )。以往的研究显示, 草本、蕨类、苔藓等速生型植物对N添加的敏感性明显高于其他物种, 这可能与速生物种较快的养分转换速率 及其养分策略可以循环利用更多的N有关(Aerts & Peijl, 1993 )。
本研究表明N添加对植物P元素的吸收有促进作用, 这与鼎湖山亚热带N添加实验的结果(Huang et al ., 2012 )相一致, 但与一些温带森林的研究结果 (Nilsson & Wallander, 2003 ; Braun et al ., 2010 )不同。N添加引起植物P含量增加的原因通常包含两方面: 一方面, N添加通过提高土壤中P的可利用性, 提高植物体内的P含量, 这与本研究中表层土壤P含量显著增加的结果(表1 )相一致。另一方面, Fujita等(2010)的研究表明, N添加会通过促进根系磷酸酶活性等方式提高植物对土壤中P的吸收能力。此外, N添加条件下植物可以将自身多余的N用于构造更多的磷酸酶, 从而促进对土壤P元素的吸收, 进而导致叶片P含量上升(Kathleen & Vitousek, 2001 )。
3.2 N添加对植物叶片N:P的影响
植物N:P可以用来指示N、P元素对生长限制的相对强弱(Güsewell & Koerselman, 2002 ; Tessier & Raynal, 2003 ; Güsewell, 2004 )。Koerselman和Meu- leman (1996)提出判断植物N、P营养限制的标准: 当植物叶片N:P小于14时, 植物生长受N元素限制; 当N:P介于14和16之间时, 植物生长受N和P元素共同限制; 当N:P大于16时, 植物生长受P元素限制。本实验中对照样地的植物叶片平均N:P为18.67, 各物种N:P均大于16, 表明N添加样地土壤P元素供应短缺, 植物生长主要受P元素限制。这与Aerts等(2003)关于亚热带地区森林生长主要受土壤P限制的论述一致, 并与Han等(2005)关于我国大部分地区P元素缺乏的结果相符合。本研究发现, 随着N添加量的增加, 植物叶片N:P逐渐升高, N:P由对照样地的18.67增加到19.72, 说明随着N添加量的增加, 植物生长的P限制增强。这与其他研究中关于N沉降使陆地生态系统从N限制向P限制转变的结论(Elser et al ., 2007 ; Sardans et al ., 2012 )相一致。
植物叶片N:P与物种的生态策略有关(Reich, 2003 ; Jeyasingh et al ., 2009 ), 植物叶片N:P的变化表征了植物维持体内化学计量内稳性的能力(Sterner & Elser, 2002 ; Demars & Edwards, 2007 ), 反映了生物对环境变化的适应性(Elser et al ., 2010a )。当外界环境发生改变时, 植物通常倾向于保持相对稳定的N:P来满足自身功能的稳定(Demars & Edwards, 2007 ; Elser et al ., 2010b ; Irakli & Elser, 2011 )。本研究5年的N添加实验表明: 乔木叶片的N:P相对稳定, 变化不明显; 而草本层植物叶片的N:P提高显著, 这预示着短期N添加条件下, 乔木植物叶片N:P较林下植物更加稳定。就灌木层植物而言, 其N:P的N添加效果表现出明显的时间效应(图4 )。随着N添加年限的增加, 灌木植物N:P对N添加的响应逐渐增强, 这符合Aber等(1998)提出的N滞留假设, 即: N添加的影响存在累积效应, 过去添加的N仍会对现在的植物化学计量特征产生影响。
研究显示N添加背景下, 武夷山植物叶片N:P的变化主要受叶片P含量变化影响, 而非受N含量变化影响(图3 )。其他研究结果也显示, 在N元素相对充足而P元素相对匮乏的生态系统, N添加会明显改变该生态系统的P循环(Güsewell, 2004 ; Liu et al ., 2013 )。N、P元素从细胞到生态系统尺度, 都存在紧密的耦合作用(Elser et al ., 2000 ; Sterner & Elser, 2002 ; Vitousek et al ., 2010 ), 其中一种元素的吸收受到另一种元素可利用性的影响(Ågren et al ., 2012 ), 因此N添加会通过影响土壤中P的可利用性、促进根系磷酸酶活性提高植物对P的吸收等方式改变整个生态系统的P循环(Treseder & Vitousek, 2001 ; Marklein & Houlton, 2012 )。本研究结果进一步表明, N添加对武夷山植物群落P循环的影响明显, 植物叶片P含量变化是叶片N:P变化的主要原因。
致谢 感谢北京大学方精云、吉成均、朱彪等老师对本研究的悉心指导, 以及李鹏、王逸然、田地等同学在实验和文章撰写过程中给予的无私帮助。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Aber JD McDowell W Nadelhoffer KJ Magill AH Berntson G Kamakea M McNulty S Currie W Rustad L Fernandez I 1998 ). Nitrogen saturation in temperate forest ecosystems: Hypotheses revisited
.Bioscience , 48 , 921 -934 .
[2]
Aber JD Melillo JM 1989 ). Nitrogen saturation in northern forest ecosystems
.Bioscience , 39 , 378 -386 .
[本文引用: 3]
[3]
Aerts R 1996 ). Nutrient resorption from senescing leaves of perennials: Are there general patterns?
Journal of Ecology , 84 , 597 -608 .
[本文引用: 1]
[4]
Aerts R Caluwe HD Beltman B 2003 ). Is the relation between nutrient supply and biodiversity co-determined by the type of nutrient limitation?
Oikos , 101 , 489 -498 .
[5]
Aerts R Peijl MJVD 1993 ). A simple model to explain the dominance of low productive perennials in nutrient poor habitats
.Oikos , 66 , 144 -147 .
[本文引用: 1]
[6]
Ågren GI 2004 ). The C:N:P stoichiometry of autotrophs— Theory and observations
.Ecology Letters , 7 , 185 -191 .
[本文引用: 1]
[7]
Ågren GI Wetterstedt JÅM Billberger MFK 2012 ). Nutrient limitation on terrestrial plant growth—modeling the interaction between nitrogen and phosphorus
.New Phytologist , 194 , 953 -960 .
[本文引用: 1]
[8]
Braun S Thomas VFD Quiring R Flückiger W 2010 ). Does nitrogen deposition increase forest production? The role of phosphorus
.Environmental Pollution , 158 , 2043 -2052 .
[本文引用: 1]
[9]
Cui Q Lü XT Wang QB Han XG 2010 ). Nitrogen fertilization and fire act independently on foliar stoichiometry in a temperate steppe
.Plant and Soil , 334 , 209 -219 .
[10]
Demars BOL Edwards AC 2007 ). Tissue nutrient concentrations in freshwater aquatic macrophytes: High inter-taxon differences and low phenotypic response to nutrient supply
.Freshwater Biology , 52 , 2073 -2086 .
[本文引用: 3]
[11]
Dentener F Drevet J Lamarque JF Bey I Eickhout B Fiore AM Hauglustaine D Horowitz LW Krol M Kulshrestha UC Lawrence M Galy-Lacaux C Rast S Shindell D Stevenson D Noije TV Atherton C Bell N Bergman D Butler T Cofala J Collins B Doherty R Ellingsen K Galloway J Gauss M Montanaro V Müller JF Pitari G Rodriguez J Sanderson M Solmon F Strahan S Schultz M Sudo K Szopa S Wild O 2006 ). Nitrogen and sulfur deposition on regional and global scales: A multimodel evaluation
.Global Biogeochemical Cycles , 20 , 16615 .
[本文引用: 1]
[12]
Duval BD Dijkstra P Natali SM Megonigal JP Ketterer ME Drake BG Lerdau MT Gordon G Anbar AD Hungate BA 2011 ). Plant-soil distribution of potentially toxic elements in response to elevated atmospheric CO2
.Environmental Science & Technology , 45 , 2570 -2574 .
[本文引用: 1]
[13]
Elliott KJ White AS 1994 ). Effects of light, nitrogen, and phosphorus on red pine seedling growth and nutrient use efficiency
.Forest Science , 40 , 47 -58 .
[本文引用: 1]
[14]
Elser JJ Bracken MES Cleland EE Gruner DS Harpole WS Hillebrand H Ngai JT Seabloom EW Shurin JB Smith JE 2007 ). Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems
. Ecology Letters , 10 , 1135 -1142 .
[本文引用: 1]
[15]
Elser JJ Fagan WF Kerkhoff AJ Swenson NG Enquist BJ 2010 a). Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change
.New Phytologist , 186 , 593 -608 .
[本文引用: 2]
[16]
Elser JJ Peace AL Kyle M Wojewodzic M Mccrackin ML Andersen T Hessen DO 2010 b). Atmospheric nitrogen deposition is associated with elevated phosphorus limitation of lake zooplankton
.Ecology Letters , 13 , 1256 -1261 .
[本文引用: 1]
[17]
Elser JJ Sterner RW Gorokhova E Fagan WF Markow TA Cotner JB Harrison JF Hobbie SE Odell GM Weide LJ 2000 ). Biological stoichiometry from genes to ecosystems
.Ecology Letters , 3 , 540 -550 .
[本文引用: 1]
[18]
Fang JY Guo ZD Hu HF Kato T Muraoka H Son Y 2014 ). Forest biomass carbon sinks in East Asia, with special reference to the relative contributions of forest expansion and forest growth
.Global Change Biology , 20 , 2019 -2030 .
[本文引用: 1]
[19]
Fang YT Mo JM Zhou GY Gundersen P Li DJ Jiang YQ 2004 ). The short-term responses of soil available nitrogen of Dinghushan forests to simulated N deposition in subtropical China
.Acta Ecologica Sinica , 24 , 2353 -2359 . (in Chinese with English abstract)[方运霆 , 莫江明 , 周国逸 , Gundersen P 李德军 , 江远清 (2004 ). 南亚热带森林土壤有效氮含量及其对模拟氮沉降增加的初期响应
. 生态学报 , 24 , 2353 -2359 .]
[本文引用: 1]
[20]
Fujita Y Robroek BJM Ruiter PCD Heil GW Wassen MJ 2010 ). Increased N affects P uptake of eight grassland species: The role of root surface phosphatase activity
.Oikos , 119 , 1665 -1673 .
[21]
Galloway JN Dentener FJ Capone DG Boyer EW Howarth RW Seitzinger SP Vöosmarty CJ 2004 ). Nitrogen cycles: Past, present and future
.Biogeochemistry , 70 , 153 -226 .
[本文引用: 1]
[22]
Galloway JN Sutton MA 2008 ). Transformation of the nitrogen cycle: Recent trends, questions, and potential solutions
.Science , 320 , 889 -892 .
[本文引用: 1]
[23]
Gao SP Li JX Xu MC Chen X Dai J 2007 ). Leaf N and P stoichiometry of common species in successional stages of the evergreen broad-leaved forest in Tiantong National Forest Park, Zhejiang Province, China
.Acta Ecologica Sinica , 27 , 947 -952 . (in Chinese with English abstract)[高三平 , 李俊祥 , 徐明策 , 陈熙 , 戴洁 (2007 ). 天童常绿阔叶林不同演替阶段常见种叶片N、P化学计量学特征
. 生态学报 , 27 , 947 -952 .]
[本文引用: 1]
[24]
Güsewell S 2004 ). N:P ratios in terrestrial plants: Variation and functional significance
.New Phytologist , 164 , 243 -266 .
[本文引用: 3]
[25]
Güsewell S Koerselman W 2002 ). Variation in nitrogen and phosphorus concentrations of wetland plants
.Perspectives in Plant Ecology Evolution & Systematics , 5 , 37 -61 .
[本文引用: 3]
[26]
Han WX Fang JY Guo DL Zhang Y 2005 ). Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China
.New Phytologist , 168 , 377 -385 .
[本文引用: 1]
[27]
Han WX Fang JY Reich PB Woodward FI Wang ZH 2011 ). Biogeography and variability of eleven mineral elements in plant leaves across gradients of climate, soil and plant functional type in China
.Ecology Letters , 14 , 788 -796 .
[本文引用: 1]
[28]
He JS Wang L Dan FBF Wang XP Ma WH Fang JY 2008 ). Leaf nitrogen:phosphorus stoichiometry across Chinese grassland biomes
.Oecologia , 155 , 301 -310 .
[本文引用: 1]
[29]
Huang WJ Zhou GY Liu JX Zhang DQ Xu ZH Liu SZ 2012 ). Effects of elevated carbon dioxide and nitrogen addition on foliar stoichiometry of nitrogen and phosphorus of five tree species in subtropical model forest
.Environmental Pollution , 168 , 113 -120 .
[本文引用: 1]
[30]
Irakli L Elser JJ 2011 ). The origins of the red field nitrogento- phosphorus ratio are in a homoeostatic protein-to-rRNA ratio
.Ecology Letters , 14 , 244 -250 .
[本文引用: 1]
[31]
Jeyasingh PD Weider LJ Sterner RW 2009 ). Genetically- based trade-offs in response to stoichiometric food quality influence competition in a keystone aquatic herbivore
.Ecology Letters , 12 , 1229 -1237 .
[本文引用: 1]
[32]
Kathleen KT Vitousek PM 2001 ). Effects of soil nutrient availability on investment in acquisition of N and P in Hawaiian Rain Forests
. Ecology , 82 , 946 -954 .
[本文引用: 1]
[33]
Koerselman W Meuleman AFM 1996 ). The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation
.Journal of Applied Ecology , 33 , 1441 -1450 .
[34]
Liu JX Huang WJ Zhou G Zhang D Liu S Li Y 2013 ). Nitrogen to phosphorus ratios of tree species in response to elevated carbon dioxide and nitrogen addition in subtropical forests
.Global Change Biology , 19 , 208 -216 .
[本文引用: 1]
[35]
Lu XK Mo JM Gilliam FS Zhou GY Fang YT 2010 ). Effects of experimental nitrogen additions on plant diversity in an old-growth tropical forest
.Global Change Biology , 16 , 2688 -2700 .
[本文引用: 2]
[36]
Magill AH Aber JD Berntson GM Mcdowell WH Nadelhoffer KJ Melillo JM Steudler P 2000 ). Long-term nitrogen additions and nitrogen saturation in two temperate forests
.Ecosystems , 3 , 238 -253 .
[本文引用: 3]
[37]
Magill AH Aber JD Currie WS Nadelhoffer KJ Martin ME Mcdowell WH Melillo JM Steudler P 2004 ). Ecosystem response to 15 years of chronic nitrogen additions at the Harvard forest LTER, Massachusetts, USA
.Forest Ecology & Management , 196 , 7 -28 .
[本文引用: 1]
[38]
Mahowald N Jickells TD Baker AR Artaxo P Benitez-Nelson CR Bergametti G Bond TC Chen Y Cohen DD Herut B Kubilay N Losno R Luo C Maenhaut W McGee KA Okin GS Siefert RL Tsukuda S 2008 ). Global distribution of atmospheric phosphorus sources, concentrations and deposition rates, and anthropogenic impacts
.Global Biogeochemical Cycles , 22 , 37 -42 .
[本文引用: 1]
[39]
Marklein AR Houlton BZ 2012 ). Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems
.New Phytologist , 193 , 696 -704 .
[本文引用: 1]
[40]
Mcnulty SG Boggs J Aber JD Rustad L Magill A 2005 ). Red spruce ecosystem level changes following 14 years of chronic N fertilization
.Forest Ecology & Management , 219 , 279 -291 .
[本文引用: 1]
[41]
Mo JM Li DJ Gundersen P 2008 ). Seedling growth response of two tropical tree species to nitrogen deposition in southern China
.European Journal of Forest Research , 127 , 275 -283 .
[本文引用: 1]
[42]
Nilsson LO Wallander H 2003 ). Production of external mycelium by ectomycorrhizal fungi in a Norway spruce forest was reduced in response to nitrogen fertilization
.New Phytologist , 158 , 409 -416 .
[本文引用: 1]
[43]
Reich PB 2003 ). The evolution of plant functional variation: Traits, spectra and strategies
.International Journal of Plant Sciences , 164 , 143 -164 .
[本文引用: 2]
[44]
Reich PB Bowman WD 1999 ). Generality of leaf trait relationships: A test across six biomes
.Ecology , 80 , 1955 -1969 .
[本文引用: 2]
[45]
Reich PB Oleksyn J 2004 ). Global patterns of plant leaf N and P in relation to temperature and latitude
.Proceedings of the National Academy of Sciences of the United States of America , 101 , 11001 -11006 .
[本文引用: 3]
[46]
Rowe EC Smart SM Kennedy VH Emmett BA Evans CD 2008 ). Nitrogen deposition increases the acquisition of phosphorus and potassium by heather calluna vulgaris
.Environmental Pollution , 155 , 201 -207 .
[本文引用: 1]
[47]
Sardans J Rivas-Ubach A Peñuelas J 2012 ). The C:N:P stoichiometry of organisms and ecosystems in a changing world: A review and perspectives
.Perspectives in Plant Ecology Evolution & Systematics , 14 , 33 -47 .
[本文引用: 3]
[48]
Sterner RW Elser JJ 2002 ). Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere . Princeton University Press , Princeton, USA .
[本文引用: 6]
[49]
Tessier JT Raynal DJ 2003 ). Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation
.Journal of Applied Ecology , 40 , 523 -534 .
[本文引用: 1]
[50]
Thompson K Parkinson JA Band SR Spencer RE 1997 ). A comparative study of leaf nutrient concentrations in a regional herbaceous flora
.New Phytologist , 136 , 679 -689 .
[本文引用: 1]
[51]
Tilman D 1990 ). Constraints and tradeoffs: Toward a predictive theory of competition and succession
.Oikos , 58 , 3 -15 .
[本文引用: 1]
[52]
Townsend AR Bustamante MMC 2007 ). Controls over foliar N:P ratios in tropical rain forests
.Ecology , 88 , 107 -118 .
[本文引用: 1]
[53]
Treseder KK 2008 ). Nitrogen additions and microbial biomass: A meta-analysis of ecosystem studies
.Ecology Letters , 11 , 1111 -1120 .
[本文引用: 1]
[54]
Treseder KK Vitousek PM 2001 ). Effects of soil nutrient availability on investment in acquisition of N and P in Hawaiian rain forests
.Ecology , 82 , 946 -954 .
[本文引用: 1]
[55]
Vitousek PM Stephen P Houlton BZ Chadwick OA 2010 ). Terrestrial phosphorus limitation: Mechanisms, implications and nitrogen-phosphorus interactions
.Ecological Applications , 20 , 5 -15 .
[本文引用: 1]
[56]
Vitousek PM Tilman DG 1997 ). Technical report: Human alteration of the global nitrogen cycle: Sources and consequences
.Ecological Applications , 7 , 737 -750 .
[本文引用: 1]
[57]
Wright RF Rasmussen L 1998 ). Introduction to the NITREX and EXMAN projects
.Forest Ecology & Management , 101 , 1 -7 .
[本文引用: 2]
Nitrogen saturation in temperate forest ecosystems: Hypotheses revisited
1998
Nitrogen saturation in northern forest ecosystems
3
1989
... 人类活动导致大气氮(N)沉降不断增强(Galloway et al ., 2004 ; Dentener et al ., 2006 ; Mahowald et al ., 2008 ), 正深刻影响着生态系统及其养分循环过程(Vitousek & Tilman, 1997 ; Galloway & Sutton, 2008 ; Treseder, 2008 ).野外人工N添加实验是模拟自然N沉降增加对生态系统影响的重要手段.自20世纪80年代起, 研究者在温带和北方森林地区陆续开展了大量长时间的野外N添加实验(Aber & Melillo, 1989 ; Wright & Rasmussen, 1998 ; Magill et al ., 2000 , 2004 ), 而N添加实验在亚热带常绿阔叶林地区开展得相对较少(Mo et al ., 2008 ; Lu et al ., 2010 ).我国的亚热带常绿阔叶林面积广阔、物种组成丰富(Fang et al ., 2014 ), 是全球亚热带常绿阔叶林的主要分布区, 在我国亚热带常绿阔叶林地区开展N添加的生态学效应的研究十分必要. ...
... 在全球变化的背景下, 植物的化学计量特征可以表征植物维持体内化学计量内稳性的能力(Sterner & Elser, 2002 ; Demars & Edwards, 2007 ), 并反映植物对环境变化的适应性(Elser et al ., 2010a ).以往的研究表明, 植物叶片N、磷(P)化学计量特征因植被类型、功能群、物种的差异而不同(Sterner & Elser, 2002 ; Reich, 2003 ; Ågren, 2004 ), 不同功能群和物种对N添加的响应也存在差异(Güsewell, 2004 ).自20世纪80年代起, 研究人员在温带和北方森林地区陆续开展了一些经典的野外N添加实验(Aber & Melillo, 1989 ; Wright & Rasmussen, 1998 ; Magill et al ., 2000 ), 植物化学计量学作为研究N循环的新手段, 也被应用于N添加生态学效应的研究中.例如: Magill等(2004)在美国哈佛森林开展了长期的N添加试验, 经过15年的研究发现: N添加显著增加了乔木优势种的叶片N含量, 并且叶片化学计量特征对N添加的响应存在明显的年际变化; Huang等(2012)在模拟的亚热带常绿阔叶林生态系统中开展了N添加实验, 对群落内5种乔木树种叶片N、P化学计量特征进行了研究, 结果显示N添加显著影响了多数物种的叶片N、P含量和N:P; Cui等(2010)对温带草原8种优势植物叶片化学计量特征对N添加的响应进行了研究, 发现不同物种的叶片化学计量特征对N添加的响应也存在差异. ...
... N添加整体上促进了植物对N的吸收, 这与以往的研究结果(Magill et al ., 2000 ; Mcnulty et al ., 2005 ; Rowe et al ., 2008 )一致.植物叶片的矿质元素含量取决于土壤养分的可利用性(Townsend & Bustamante, 2007 ).以往的研究也显示, 植物组织内的矿质元素含量与土壤中相应元素含量呈现显著正相关关系(Reich & Oleksyn, 2004 ), 当土壤中某种矿质元素含量增加时, 植物体内相应的矿质元素含量也会随之增加.功能群和物种则是影响叶片N、P化学计量特征及其对N添加响应的重要因素(Reich & Bowman, 1999 ; Sterner & Elser, 2002 ; Sardans et al ., 2012 ).N添加情况下, 植物对N、P等矿质元素的利用策略因需求、竞争力和优势度的不同而存在差异(Tilman, 1990 ; Güsewell & Koerselman, 2002 ).本研究中, 乔木优势种米槠对N添加响应敏感, N添加显著增加了其叶片N含量(图2 ).这可能是由于当养分条件发生变化时, 生态系统趋向于将N、P等矿质元素运输到生产力较高的植物中去, 优先保证其生长需求(Aber & Melillo, 1989 ; Elliott & White, 1994 ).本研究中林下草本层植物N含量对N添加的响应也较为敏感(图2 ).以往的研究显示, 草本、蕨类、苔藓等速生型植物对N添加的敏感性明显高于其他物种, 这可能与速生物种较快的养分转换速率 及其养分策略可以循环利用更多的N有关(Aerts & Peijl, 1993 ). ...
Nutrient resorption from senescing leaves of perennials: Are there general patterns?
1
1996
... 不同功能群物种叶片的N、P含量均存在显著差异, 这与以往的研究结果(Reich & Bowman, 1999 ; Sterner & Elser, 2002 ; Sardans et al ., 2012 )较为一致.本研究显示, 草本层植物叶片的N、P含量显著高于木本植物(包括乔木和灌木植物), 针叶植物叶片的N、P含量显著高于阔叶植物, 这一结果与其他研究中得到的中国草本和木本植物的N、P化学计量模式基本一致(高三平等, 2007 ; He et al ., 2008 ; Han et al ., 2011 ), 并且证明了生命周期短而生长快速的植物要比生命周期长而生长缓慢的植物具有更高的叶片N、P含量(Aerts, 1996 ; Thompson et al ., 1997 ; Güsewell & Koerselman, 2002 ). ...
Is the relation between nutrient supply and biodiversity co-determined by the type of nutrient limitation?
2003
A simple model to explain the dominance of low productive perennials in nutrient poor habitats
1
1993
... N添加整体上促进了植物对N的吸收, 这与以往的研究结果(Magill et al ., 2000 ; Mcnulty et al ., 2005 ; Rowe et al ., 2008 )一致.植物叶片的矿质元素含量取决于土壤养分的可利用性(Townsend & Bustamante, 2007 ).以往的研究也显示, 植物组织内的矿质元素含量与土壤中相应元素含量呈现显著正相关关系(Reich & Oleksyn, 2004 ), 当土壤中某种矿质元素含量增加时, 植物体内相应的矿质元素含量也会随之增加.功能群和物种则是影响叶片N、P化学计量特征及其对N添加响应的重要因素(Reich & Bowman, 1999 ; Sterner & Elser, 2002 ; Sardans et al ., 2012 ).N添加情况下, 植物对N、P等矿质元素的利用策略因需求、竞争力和优势度的不同而存在差异(Tilman, 1990 ; Güsewell & Koerselman, 2002 ).本研究中, 乔木优势种米槠对N添加响应敏感, N添加显著增加了其叶片N含量(图2 ).这可能是由于当养分条件发生变化时, 生态系统趋向于将N、P等矿质元素运输到生产力较高的植物中去, 优先保证其生长需求(Aber & Melillo, 1989 ; Elliott & White, 1994 ).本研究中林下草本层植物N含量对N添加的响应也较为敏感(图2 ).以往的研究显示, 草本、蕨类、苔藓等速生型植物对N添加的敏感性明显高于其他物种, 这可能与速生物种较快的养分转换速率 及其养分策略可以循环利用更多的N有关(Aerts & Peijl, 1993 ). ...
The C:N:P stoichiometry of autotrophs— Theory and observations
1
2004
... 在全球变化的背景下, 植物的化学计量特征可以表征植物维持体内化学计量内稳性的能力(Sterner & Elser, 2002 ; Demars & Edwards, 2007 ), 并反映植物对环境变化的适应性(Elser et al ., 2010a ).以往的研究表明, 植物叶片N、磷(P)化学计量特征因植被类型、功能群、物种的差异而不同(Sterner & Elser, 2002 ; Reich, 2003 ; Ågren, 2004 ), 不同功能群和物种对N添加的响应也存在差异(Güsewell, 2004 ).自20世纪80年代起, 研究人员在温带和北方森林地区陆续开展了一些经典的野外N添加实验(Aber & Melillo, 1989 ; Wright & Rasmussen, 1998 ; Magill et al ., 2000 ), 植物化学计量学作为研究N循环的新手段, 也被应用于N添加生态学效应的研究中.例如: Magill等(2004)在美国哈佛森林开展了长期的N添加试验, 经过15年的研究发现: N添加显著增加了乔木优势种的叶片N含量, 并且叶片化学计量特征对N添加的响应存在明显的年际变化; Huang等(2012)在模拟的亚热带常绿阔叶林生态系统中开展了N添加实验, 对群落内5种乔木树种叶片N、P化学计量特征进行了研究, 结果显示N添加显著影响了多数物种的叶片N、P含量和N:P; Cui等(2010)对温带草原8种优势植物叶片化学计量特征对N添加的响应进行了研究, 发现不同物种的叶片化学计量特征对N添加的响应也存在差异. ...
Nutrient limitation on terrestrial plant growth—modeling the interaction between nitrogen and phosphorus
1
2012
... 研究显示N添加背景下, 武夷山植物叶片N:P的变化主要受叶片P含量变化影响, 而非受N含量变化影响(图3 ).其他研究结果也显示, 在N元素相对充足而P元素相对匮乏的生态系统, N添加会明显改变该生态系统的P循环(Güsewell, 2004 ; Liu et al ., 2013 ).N、P元素从细胞到生态系统尺度, 都存在紧密的耦合作用(Elser et al ., 2000 ; Sterner & Elser, 2002 ; Vitousek et al ., 2010 ), 其中一种元素的吸收受到另一种元素可利用性的影响(Ågren et al ., 2012 ), 因此N添加会通过影响土壤中P的可利用性、促进根系磷酸酶活性提高植物对P的吸收等方式改变整个生态系统的P循环(Treseder & Vitousek, 2001 ; Marklein & Houlton, 2012 ).本研究结果进一步表明, N添加对武夷山植物群落P循环的影响明显, 植物叶片P含量变化是叶片N:P变化的主要原因. ...
Does nitrogen deposition increase forest production? The role of phosphorus
1
2010
... 本研究表明N添加对植物P元素的吸收有促进作用, 这与鼎湖山亚热带N添加实验的结果(Huang et al ., 2012 )相一致, 但与一些温带森林的研究结果 (Nilsson & Wallander, 2003 ; Braun et al ., 2010 )不同.N添加引起植物P含量增加的原因通常包含两方面: 一方面, N添加通过提高土壤中P的可利用性, 提高植物体内的P含量, 这与本研究中表层土壤P含量显著增加的结果(表1 )相一致.另一方面, Fujita等(2010)的研究表明, N添加会通过促进根系磷酸酶活性等方式提高植物对土壤中P的吸收能力.此外, N添加条件下植物可以将自身多余的N用于构造更多的磷酸酶, 从而促进对土壤P元素的吸收, 进而导致叶片P含量上升(Kathleen & Vitousek, 2001 ). ...
Nitrogen fertilization and fire act independently on foliar stoichiometry in a temperate steppe
2010
Tissue nutrient concentrations in freshwater aquatic macrophytes: High inter-taxon differences and low phenotypic response to nutrient supply
3
2007
... 在全球变化的背景下, 植物的化学计量特征可以表征植物维持体内化学计量内稳性的能力(Sterner & Elser, 2002 ; Demars & Edwards, 2007 ), 并反映植物对环境变化的适应性(Elser et al ., 2010a ).以往的研究表明, 植物叶片N、磷(P)化学计量特征因植被类型、功能群、物种的差异而不同(Sterner & Elser, 2002 ; Reich, 2003 ; Ågren, 2004 ), 不同功能群和物种对N添加的响应也存在差异(Güsewell, 2004 ).自20世纪80年代起, 研究人员在温带和北方森林地区陆续开展了一些经典的野外N添加实验(Aber & Melillo, 1989 ; Wright & Rasmussen, 1998 ; Magill et al ., 2000 ), 植物化学计量学作为研究N循环的新手段, 也被应用于N添加生态学效应的研究中.例如: Magill等(2004)在美国哈佛森林开展了长期的N添加试验, 经过15年的研究发现: N添加显著增加了乔木优势种的叶片N含量, 并且叶片化学计量特征对N添加的响应存在明显的年际变化; Huang等(2012)在模拟的亚热带常绿阔叶林生态系统中开展了N添加实验, 对群落内5种乔木树种叶片N、P化学计量特征进行了研究, 结果显示N添加显著影响了多数物种的叶片N、P含量和N:P; Cui等(2010)对温带草原8种优势植物叶片化学计量特征对N添加的响应进行了研究, 发现不同物种的叶片化学计量特征对N添加的响应也存在差异. ...
... 植物叶片N:P与物种的生态策略有关(Reich, 2003 ; Jeyasingh et al ., 2009 ), 植物叶片N:P的变化表征了植物维持体内化学计量内稳性的能力(Sterner & Elser, 2002 ; Demars & Edwards, 2007 ), 反映了生物对环境变化的适应性(Elser et al ., 2010a ).当外界环境发生改变时, 植物通常倾向于保持相对稳定的N:P来满足自身功能的稳定(Demars & Edwards, 2007 ; Elser et al ., 2010b ; Irakli & Elser, 2011 ).本研究5年的N添加实验表明: 乔木叶片的N:P相对稳定, 变化不明显; 而草本层植物叶片的N:P提高显著, 这预示着短期N添加条件下, 乔木植物叶片N:P较林下植物更加稳定.就灌木层植物而言, 其N:P的N添加效果表现出明显的时间效应(图4 ).随着N添加年限的增加, 灌木植物N:P对N添加的响应逐渐增强, 这符合Aber等(1998)提出的N滞留假设, 即: N添加的影响存在累积效应, 过去添加的N仍会对现在的植物化学计量特征产生影响. ...
... ).当外界环境发生改变时, 植物通常倾向于保持相对稳定的N:P来满足自身功能的稳定(Demars & Edwards, 2007 ; Elser et al ., 2010b ; Irakli & Elser, 2011 ).本研究5年的N添加实验表明: 乔木叶片的N:P相对稳定, 变化不明显; 而草本层植物叶片的N:P提高显著, 这预示着短期N添加条件下, 乔木植物叶片N:P较林下植物更加稳定.就灌木层植物而言, 其N:P的N添加效果表现出明显的时间效应(图4 ).随着N添加年限的增加, 灌木植物N:P对N添加的响应逐渐增强, 这符合Aber等(1998)提出的N滞留假设, 即: N添加的影响存在累积效应, 过去添加的N仍会对现在的植物化学计量特征产生影响. ...
Nitrogen and sulfur deposition on regional and global scales: A multimodel evaluation
1
2006
... 人类活动导致大气氮(N)沉降不断增强(Galloway et al ., 2004 ; Dentener et al ., 2006 ; Mahowald et al ., 2008 ), 正深刻影响着生态系统及其养分循环过程(Vitousek & Tilman, 1997 ; Galloway & Sutton, 2008 ; Treseder, 2008 ).野外人工N添加实验是模拟自然N沉降增加对生态系统影响的重要手段.自20世纪80年代起, 研究者在温带和北方森林地区陆续开展了大量长时间的野外N添加实验(Aber & Melillo, 1989 ; Wright & Rasmussen, 1998 ; Magill et al ., 2000 , 2004 ), 而N添加实验在亚热带常绿阔叶林地区开展得相对较少(Mo et al ., 2008 ; Lu et al ., 2010 ).我国的亚热带常绿阔叶林面积广阔、物种组成丰富(Fang et al ., 2014 ), 是全球亚热带常绿阔叶林的主要分布区, 在我国亚热带常绿阔叶林地区开展N添加的生态学效应的研究十分必要. ...
Plant-soil distribution of potentially toxic elements in response to elevated atmospheric CO2
1
2011
... 采用相对效应指数(RE )表征N添加样地植物叶片化学计量特征对N添加的响应(Duval et al ., 2011 ).计算公式为: 相对效应指数(RE ) = (处理样地值-对照样地值)/对照样地值×100%. ...
Effects of light, nitrogen, and phosphorus on red pine seedling growth and nutrient use efficiency
1
1994
... N添加整体上促进了植物对N的吸收, 这与以往的研究结果(Magill et al ., 2000 ; Mcnulty et al ., 2005 ; Rowe et al ., 2008 )一致.植物叶片的矿质元素含量取决于土壤养分的可利用性(Townsend & Bustamante, 2007 ).以往的研究也显示, 植物组织内的矿质元素含量与土壤中相应元素含量呈现显著正相关关系(Reich & Oleksyn, 2004 ), 当土壤中某种矿质元素含量增加时, 植物体内相应的矿质元素含量也会随之增加.功能群和物种则是影响叶片N、P化学计量特征及其对N添加响应的重要因素(Reich & Bowman, 1999 ; Sterner & Elser, 2002 ; Sardans et al ., 2012 ).N添加情况下, 植物对N、P等矿质元素的利用策略因需求、竞争力和优势度的不同而存在差异(Tilman, 1990 ; Güsewell & Koerselman, 2002 ).本研究中, 乔木优势种米槠对N添加响应敏感, N添加显著增加了其叶片N含量(图2 ).这可能是由于当养分条件发生变化时, 生态系统趋向于将N、P等矿质元素运输到生产力较高的植物中去, 优先保证其生长需求(Aber & Melillo, 1989 ; Elliott & White, 1994 ).本研究中林下草本层植物N含量对N添加的响应也较为敏感(图2 ).以往的研究显示, 草本、蕨类、苔藓等速生型植物对N添加的敏感性明显高于其他物种, 这可能与速生物种较快的养分转换速率 及其养分策略可以循环利用更多的N有关(Aerts & Peijl, 1993 ). ...
Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems
1
2007
... 植物N:P可以用来指示N、P元素对生长限制的相对强弱(Güsewell & Koerselman, 2002 ; Tessier & Raynal, 2003 ; Güsewell, 2004 ).Koerselman和Meu- leman (1996)提出判断植物N、P营养限制的标准: 当植物叶片N:P小于14时, 植物生长受N元素限制; 当N:P介于14和16之间时, 植物生长受N和P元素共同限制; 当N:P大于16时, 植物生长受P元素限制.本实验中对照样地的植物叶片平均N:P为18.67, 各物种N:P均大于16, 表明N添加样地土壤P元素供应短缺, 植物生长主要受P元素限制.这与Aerts等(2003)关于亚热带地区森林生长主要受土壤P限制的论述一致, 并与Han等(2005)关于我国大部分地区P元素缺乏的结果相符合.本研究发现, 随着N添加量的增加, 植物叶片N:P逐渐升高, N:P由对照样地的18.67增加到19.72, 说明随着N添加量的增加, 植物生长的P限制增强.这与其他研究中关于N沉降使陆地生态系统从N限制向P限制转变的结论(Elser et al ., 2007 ; Sardans et al ., 2012 )相一致. ...
Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change
2
2010
... 在全球变化的背景下, 植物的化学计量特征可以表征植物维持体内化学计量内稳性的能力(Sterner & Elser, 2002 ; Demars & Edwards, 2007 ), 并反映植物对环境变化的适应性(Elser et al ., 2010a ).以往的研究表明, 植物叶片N、磷(P)化学计量特征因植被类型、功能群、物种的差异而不同(Sterner & Elser, 2002 ; Reich, 2003 ; Ågren, 2004 ), 不同功能群和物种对N添加的响应也存在差异(Güsewell, 2004 ).自20世纪80年代起, 研究人员在温带和北方森林地区陆续开展了一些经典的野外N添加实验(Aber & Melillo, 1989 ; Wright & Rasmussen, 1998 ; Magill et al ., 2000 ), 植物化学计量学作为研究N循环的新手段, 也被应用于N添加生态学效应的研究中.例如: Magill等(2004)在美国哈佛森林开展了长期的N添加试验, 经过15年的研究发现: N添加显著增加了乔木优势种的叶片N含量, 并且叶片化学计量特征对N添加的响应存在明显的年际变化; Huang等(2012)在模拟的亚热带常绿阔叶林生态系统中开展了N添加实验, 对群落内5种乔木树种叶片N、P化学计量特征进行了研究, 结果显示N添加显著影响了多数物种的叶片N、P含量和N:P; Cui等(2010)对温带草原8种优势植物叶片化学计量特征对N添加的响应进行了研究, 发现不同物种的叶片化学计量特征对N添加的响应也存在差异. ...
... 植物叶片N:P与物种的生态策略有关(Reich, 2003 ; Jeyasingh et al ., 2009 ), 植物叶片N:P的变化表征了植物维持体内化学计量内稳性的能力(Sterner & Elser, 2002 ; Demars & Edwards, 2007 ), 反映了生物对环境变化的适应性(Elser et al ., 2010a ).当外界环境发生改变时, 植物通常倾向于保持相对稳定的N:P来满足自身功能的稳定(Demars & Edwards, 2007 ; Elser et al ., 2010b ; Irakli & Elser, 2011 ).本研究5年的N添加实验表明: 乔木叶片的N:P相对稳定, 变化不明显; 而草本层植物叶片的N:P提高显著, 这预示着短期N添加条件下, 乔木植物叶片N:P较林下植物更加稳定.就灌木层植物而言, 其N:P的N添加效果表现出明显的时间效应(图4 ).随着N添加年限的增加, 灌木植物N:P对N添加的响应逐渐增强, 这符合Aber等(1998)提出的N滞留假设, 即: N添加的影响存在累积效应, 过去添加的N仍会对现在的植物化学计量特征产生影响. ...
b). Atmospheric nitrogen deposition is associated with elevated phosphorus limitation of lake zooplankton
1
2010
... 植物叶片N:P与物种的生态策略有关(Reich, 2003 ; Jeyasingh et al ., 2009 ), 植物叶片N:P的变化表征了植物维持体内化学计量内稳性的能力(Sterner & Elser, 2002 ; Demars & Edwards, 2007 ), 反映了生物对环境变化的适应性(Elser et al ., 2010a ).当外界环境发生改变时, 植物通常倾向于保持相对稳定的N:P来满足自身功能的稳定(Demars & Edwards, 2007 ; Elser et al ., 2010b ; Irakli & Elser, 2011 ).本研究5年的N添加实验表明: 乔木叶片的N:P相对稳定, 变化不明显; 而草本层植物叶片的N:P提高显著, 这预示着短期N添加条件下, 乔木植物叶片N:P较林下植物更加稳定.就灌木层植物而言, 其N:P的N添加效果表现出明显的时间效应(图4 ).随着N添加年限的增加, 灌木植物N:P对N添加的响应逐渐增强, 这符合Aber等(1998)提出的N滞留假设, 即: N添加的影响存在累积效应, 过去添加的N仍会对现在的植物化学计量特征产生影响. ...
Biological stoichiometry from genes to ecosystems
1
2000
... 研究显示N添加背景下, 武夷山植物叶片N:P的变化主要受叶片P含量变化影响, 而非受N含量变化影响(图3 ).其他研究结果也显示, 在N元素相对充足而P元素相对匮乏的生态系统, N添加会明显改变该生态系统的P循环(Güsewell, 2004 ; Liu et al ., 2013 ).N、P元素从细胞到生态系统尺度, 都存在紧密的耦合作用(Elser et al ., 2000 ; Sterner & Elser, 2002 ; Vitousek et al ., 2010 ), 其中一种元素的吸收受到另一种元素可利用性的影响(Ågren et al ., 2012 ), 因此N添加会通过影响土壤中P的可利用性、促进根系磷酸酶活性提高植物对P的吸收等方式改变整个生态系统的P循环(Treseder & Vitousek, 2001 ; Marklein & Houlton, 2012 ).本研究结果进一步表明, N添加对武夷山植物群落P循环的影响明显, 植物叶片P含量变化是叶片N:P变化的主要原因. ...
Forest biomass carbon sinks in East Asia, with special reference to the relative contributions of forest expansion and forest growth
1
2014
... 人类活动导致大气氮(N)沉降不断增强(Galloway et al ., 2004 ; Dentener et al ., 2006 ; Mahowald et al ., 2008 ), 正深刻影响着生态系统及其养分循环过程(Vitousek & Tilman, 1997 ; Galloway & Sutton, 2008 ; Treseder, 2008 ).野外人工N添加实验是模拟自然N沉降增加对生态系统影响的重要手段.自20世纪80年代起, 研究者在温带和北方森林地区陆续开展了大量长时间的野外N添加实验(Aber & Melillo, 1989 ; Wright & Rasmussen, 1998 ; Magill et al ., 2000 , 2004 ), 而N添加实验在亚热带常绿阔叶林地区开展得相对较少(Mo et al ., 2008 ; Lu et al ., 2010 ).我国的亚热带常绿阔叶林面积广阔、物种组成丰富(Fang et al ., 2014 ), 是全球亚热带常绿阔叶林的主要分布区, 在我国亚热带常绿阔叶林地区开展N添加的生态学效应的研究十分必要. ...
南亚热带森林土壤有效氮含量及其对模拟氮沉降增加的初期响应
1
2004
... 武夷山米槠群落植物叶片N、P含量平均值低于世界和全国平均水平, 叶片N:P则高于世界和全国平均水平(Reich & Oleksyn, 2004 ; Han et al ., 2005 ).本研究区植物生长受P元素限制更加明显.这与亚热带地区较低的土壤养分含量、较快的养分周转速率以及较强的淋溶等环境特点相适应(方运霆等, 2004 ; Reich & Oleksyn, 2004 ; Lu et al ., 2010 ). ...
Increased N affects P uptake of eight grassland species: The role of root surface phosphatase activity
2010
Nitrogen cycles: Past, present and future
1
2004
... 人类活动导致大气氮(N)沉降不断增强(Galloway et al ., 2004 ; Dentener et al ., 2006 ; Mahowald et al ., 2008 ), 正深刻影响着生态系统及其养分循环过程(Vitousek & Tilman, 1997 ; Galloway & Sutton, 2008 ; Treseder, 2008 ).野外人工N添加实验是模拟自然N沉降增加对生态系统影响的重要手段.自20世纪80年代起, 研究者在温带和北方森林地区陆续开展了大量长时间的野外N添加实验(Aber & Melillo, 1989 ; Wright & Rasmussen, 1998 ; Magill et al ., 2000 , 2004 ), 而N添加实验在亚热带常绿阔叶林地区开展得相对较少(Mo et al ., 2008 ; Lu et al ., 2010 ).我国的亚热带常绿阔叶林面积广阔、物种组成丰富(Fang et al ., 2014 ), 是全球亚热带常绿阔叶林的主要分布区, 在我国亚热带常绿阔叶林地区开展N添加的生态学效应的研究十分必要. ...
Transformation of the nitrogen cycle: Recent trends, questions, and potential solutions
1
2008
... 人类活动导致大气氮(N)沉降不断增强(Galloway et al ., 2004 ; Dentener et al ., 2006 ; Mahowald et al ., 2008 ), 正深刻影响着生态系统及其养分循环过程(Vitousek & Tilman, 1997 ; Galloway & Sutton, 2008 ; Treseder, 2008 ).野外人工N添加实验是模拟自然N沉降增加对生态系统影响的重要手段.自20世纪80年代起, 研究者在温带和北方森林地区陆续开展了大量长时间的野外N添加实验(Aber & Melillo, 1989 ; Wright & Rasmussen, 1998 ; Magill et al ., 2000 , 2004 ), 而N添加实验在亚热带常绿阔叶林地区开展得相对较少(Mo et al ., 2008 ; Lu et al ., 2010 ).我国的亚热带常绿阔叶林面积广阔、物种组成丰富(Fang et al ., 2014 ), 是全球亚热带常绿阔叶林的主要分布区, 在我国亚热带常绿阔叶林地区开展N添加的生态学效应的研究十分必要. ...
天童常绿阔叶林不同演替阶段常见种叶片N、P化学计量学特征
1
2007
... 不同功能群物种叶片的N、P含量均存在显著差异, 这与以往的研究结果(Reich & Bowman, 1999 ; Sterner & Elser, 2002 ; Sardans et al ., 2012 )较为一致.本研究显示, 草本层植物叶片的N、P含量显著高于木本植物(包括乔木和灌木植物), 针叶植物叶片的N、P含量显著高于阔叶植物, 这一结果与其他研究中得到的中国草本和木本植物的N、P化学计量模式基本一致(高三平等, 2007 ; He et al ., 2008 ; Han et al ., 2011 ), 并且证明了生命周期短而生长快速的植物要比生命周期长而生长缓慢的植物具有更高的叶片N、P含量(Aerts, 1996 ; Thompson et al ., 1997 ; Güsewell & Koerselman, 2002 ). ...
N:P ratios in terrestrial plants: Variation and functional significance
3
2004
... 在全球变化的背景下, 植物的化学计量特征可以表征植物维持体内化学计量内稳性的能力(Sterner & Elser, 2002 ; Demars & Edwards, 2007 ), 并反映植物对环境变化的适应性(Elser et al ., 2010a ).以往的研究表明, 植物叶片N、磷(P)化学计量特征因植被类型、功能群、物种的差异而不同(Sterner & Elser, 2002 ; Reich, 2003 ; Ågren, 2004 ), 不同功能群和物种对N添加的响应也存在差异(Güsewell, 2004 ).自20世纪80年代起, 研究人员在温带和北方森林地区陆续开展了一些经典的野外N添加实验(Aber & Melillo, 1989 ; Wright & Rasmussen, 1998 ; Magill et al ., 2000 ), 植物化学计量学作为研究N循环的新手段, 也被应用于N添加生态学效应的研究中.例如: Magill等(2004)在美国哈佛森林开展了长期的N添加试验, 经过15年的研究发现: N添加显著增加了乔木优势种的叶片N含量, 并且叶片化学计量特征对N添加的响应存在明显的年际变化; Huang等(2012)在模拟的亚热带常绿阔叶林生态系统中开展了N添加实验, 对群落内5种乔木树种叶片N、P化学计量特征进行了研究, 结果显示N添加显著影响了多数物种的叶片N、P含量和N:P; Cui等(2010)对温带草原8种优势植物叶片化学计量特征对N添加的响应进行了研究, 发现不同物种的叶片化学计量特征对N添加的响应也存在差异. ...
... 植物N:P可以用来指示N、P元素对生长限制的相对强弱(Güsewell & Koerselman, 2002 ; Tessier & Raynal, 2003 ; Güsewell, 2004 ).Koerselman和Meu- leman (1996)提出判断植物N、P营养限制的标准: 当植物叶片N:P小于14时, 植物生长受N元素限制; 当N:P介于14和16之间时, 植物生长受N和P元素共同限制; 当N:P大于16时, 植物生长受P元素限制.本实验中对照样地的植物叶片平均N:P为18.67, 各物种N:P均大于16, 表明N添加样地土壤P元素供应短缺, 植物生长主要受P元素限制.这与Aerts等(2003)关于亚热带地区森林生长主要受土壤P限制的论述一致, 并与Han等(2005)关于我国大部分地区P元素缺乏的结果相符合.本研究发现, 随着N添加量的增加, 植物叶片N:P逐渐升高, N:P由对照样地的18.67增加到19.72, 说明随着N添加量的增加, 植物生长的P限制增强.这与其他研究中关于N沉降使陆地生态系统从N限制向P限制转变的结论(Elser et al ., 2007 ; Sardans et al ., 2012 )相一致. ...
... 研究显示N添加背景下, 武夷山植物叶片N:P的变化主要受叶片P含量变化影响, 而非受N含量变化影响(图3 ).其他研究结果也显示, 在N元素相对充足而P元素相对匮乏的生态系统, N添加会明显改变该生态系统的P循环(Güsewell, 2004 ; Liu et al ., 2013 ).N、P元素从细胞到生态系统尺度, 都存在紧密的耦合作用(Elser et al ., 2000 ; Sterner & Elser, 2002 ; Vitousek et al ., 2010 ), 其中一种元素的吸收受到另一种元素可利用性的影响(Ågren et al ., 2012 ), 因此N添加会通过影响土壤中P的可利用性、促进根系磷酸酶活性提高植物对P的吸收等方式改变整个生态系统的P循环(Treseder & Vitousek, 2001 ; Marklein & Houlton, 2012 ).本研究结果进一步表明, N添加对武夷山植物群落P循环的影响明显, 植物叶片P含量变化是叶片N:P变化的主要原因. ...
Variation in nitrogen and phosphorus concentrations of wetland plants
3
2002
... 不同功能群物种叶片的N、P含量均存在显著差异, 这与以往的研究结果(Reich & Bowman, 1999 ; Sterner & Elser, 2002 ; Sardans et al ., 2012 )较为一致.本研究显示, 草本层植物叶片的N、P含量显著高于木本植物(包括乔木和灌木植物), 针叶植物叶片的N、P含量显著高于阔叶植物, 这一结果与其他研究中得到的中国草本和木本植物的N、P化学计量模式基本一致(高三平等, 2007 ; He et al ., 2008 ; Han et al ., 2011 ), 并且证明了生命周期短而生长快速的植物要比生命周期长而生长缓慢的植物具有更高的叶片N、P含量(Aerts, 1996 ; Thompson et al ., 1997 ; Güsewell & Koerselman, 2002 ). ...
... N添加整体上促进了植物对N的吸收, 这与以往的研究结果(Magill et al ., 2000 ; Mcnulty et al ., 2005 ; Rowe et al ., 2008 )一致.植物叶片的矿质元素含量取决于土壤养分的可利用性(Townsend & Bustamante, 2007 ).以往的研究也显示, 植物组织内的矿质元素含量与土壤中相应元素含量呈现显著正相关关系(Reich & Oleksyn, 2004 ), 当土壤中某种矿质元素含量增加时, 植物体内相应的矿质元素含量也会随之增加.功能群和物种则是影响叶片N、P化学计量特征及其对N添加响应的重要因素(Reich & Bowman, 1999 ; Sterner & Elser, 2002 ; Sardans et al ., 2012 ).N添加情况下, 植物对N、P等矿质元素的利用策略因需求、竞争力和优势度的不同而存在差异(Tilman, 1990 ; Güsewell & Koerselman, 2002 ).本研究中, 乔木优势种米槠对N添加响应敏感, N添加显著增加了其叶片N含量(图2 ).这可能是由于当养分条件发生变化时, 生态系统趋向于将N、P等矿质元素运输到生产力较高的植物中去, 优先保证其生长需求(Aber & Melillo, 1989 ; Elliott & White, 1994 ).本研究中林下草本层植物N含量对N添加的响应也较为敏感(图2 ).以往的研究显示, 草本、蕨类、苔藓等速生型植物对N添加的敏感性明显高于其他物种, 这可能与速生物种较快的养分转换速率 及其养分策略可以循环利用更多的N有关(Aerts & Peijl, 1993 ). ...
... 植物N:P可以用来指示N、P元素对生长限制的相对强弱(Güsewell & Koerselman, 2002 ; Tessier & Raynal, 2003 ; Güsewell, 2004 ).Koerselman和Meu- leman (1996)提出判断植物N、P营养限制的标准: 当植物叶片N:P小于14时, 植物生长受N元素限制; 当N:P介于14和16之间时, 植物生长受N和P元素共同限制; 当N:P大于16时, 植物生长受P元素限制.本实验中对照样地的植物叶片平均N:P为18.67, 各物种N:P均大于16, 表明N添加样地土壤P元素供应短缺, 植物生长主要受P元素限制.这与Aerts等(2003)关于亚热带地区森林生长主要受土壤P限制的论述一致, 并与Han等(2005)关于我国大部分地区P元素缺乏的结果相符合.本研究发现, 随着N添加量的增加, 植物叶片N:P逐渐升高, N:P由对照样地的18.67增加到19.72, 说明随着N添加量的增加, 植物生长的P限制增强.这与其他研究中关于N沉降使陆地生态系统从N限制向P限制转变的结论(Elser et al ., 2007 ; Sardans et al ., 2012 )相一致. ...
Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China
1
2005
... 武夷山米槠群落植物叶片N、P含量平均值低于世界和全国平均水平, 叶片N:P则高于世界和全国平均水平(Reich & Oleksyn, 2004 ; Han et al ., 2005 ).本研究区植物生长受P元素限制更加明显.这与亚热带地区较低的土壤养分含量、较快的养分周转速率以及较强的淋溶等环境特点相适应(方运霆等, 2004 ; Reich & Oleksyn, 2004 ; Lu et al ., 2010 ). ...
Biogeography and variability of eleven mineral elements in plant leaves across gradients of climate, soil and plant functional type in China
1
2011
... 不同功能群物种叶片的N、P含量均存在显著差异, 这与以往的研究结果(Reich & Bowman, 1999 ; Sterner & Elser, 2002 ; Sardans et al ., 2012 )较为一致.本研究显示, 草本层植物叶片的N、P含量显著高于木本植物(包括乔木和灌木植物), 针叶植物叶片的N、P含量显著高于阔叶植物, 这一结果与其他研究中得到的中国草本和木本植物的N、P化学计量模式基本一致(高三平等, 2007 ; He et al ., 2008 ; Han et al ., 2011 ), 并且证明了生命周期短而生长快速的植物要比生命周期长而生长缓慢的植物具有更高的叶片N、P含量(Aerts, 1996 ; Thompson et al ., 1997 ; Güsewell & Koerselman, 2002 ). ...
Leaf nitrogen:phosphorus stoichiometry across Chinese grassland biomes
1
2008
... 不同功能群物种叶片的N、P含量均存在显著差异, 这与以往的研究结果(Reich & Bowman, 1999 ; Sterner & Elser, 2002 ; Sardans et al ., 2012 )较为一致.本研究显示, 草本层植物叶片的N、P含量显著高于木本植物(包括乔木和灌木植物), 针叶植物叶片的N、P含量显著高于阔叶植物, 这一结果与其他研究中得到的中国草本和木本植物的N、P化学计量模式基本一致(高三平等, 2007 ; He et al ., 2008 ; Han et al ., 2011 ), 并且证明了生命周期短而生长快速的植物要比生命周期长而生长缓慢的植物具有更高的叶片N、P含量(Aerts, 1996 ; Thompson et al ., 1997 ; Güsewell & Koerselman, 2002 ). ...
Effects of elevated carbon dioxide and nitrogen addition on foliar stoichiometry of nitrogen and phosphorus of five tree species in subtropical model forest
1
2012
... 本研究表明N添加对植物P元素的吸收有促进作用, 这与鼎湖山亚热带N添加实验的结果(Huang et al ., 2012 )相一致, 但与一些温带森林的研究结果 (Nilsson & Wallander, 2003 ; Braun et al ., 2010 )不同.N添加引起植物P含量增加的原因通常包含两方面: 一方面, N添加通过提高土壤中P的可利用性, 提高植物体内的P含量, 这与本研究中表层土壤P含量显著增加的结果(表1 )相一致.另一方面, Fujita等(2010)的研究表明, N添加会通过促进根系磷酸酶活性等方式提高植物对土壤中P的吸收能力.此外, N添加条件下植物可以将自身多余的N用于构造更多的磷酸酶, 从而促进对土壤P元素的吸收, 进而导致叶片P含量上升(Kathleen & Vitousek, 2001 ). ...
The origins of the red field nitrogento- phosphorus ratio are in a homoeostatic protein-to-rRNA ratio
1
2011
... 植物叶片N:P与物种的生态策略有关(Reich, 2003 ; Jeyasingh et al ., 2009 ), 植物叶片N:P的变化表征了植物维持体内化学计量内稳性的能力(Sterner & Elser, 2002 ; Demars & Edwards, 2007 ), 反映了生物对环境变化的适应性(Elser et al ., 2010a ).当外界环境发生改变时, 植物通常倾向于保持相对稳定的N:P来满足自身功能的稳定(Demars & Edwards, 2007 ; Elser et al ., 2010b ; Irakli & Elser, 2011 ).本研究5年的N添加实验表明: 乔木叶片的N:P相对稳定, 变化不明显; 而草本层植物叶片的N:P提高显著, 这预示着短期N添加条件下, 乔木植物叶片N:P较林下植物更加稳定.就灌木层植物而言, 其N:P的N添加效果表现出明显的时间效应(图4 ).随着N添加年限的增加, 灌木植物N:P对N添加的响应逐渐增强, 这符合Aber等(1998)提出的N滞留假设, 即: N添加的影响存在累积效应, 过去添加的N仍会对现在的植物化学计量特征产生影响. ...
Genetically- based trade-offs in response to stoichiometric food quality influence competition in a keystone aquatic herbivore
1
2009
... 植物叶片N:P与物种的生态策略有关(Reich, 2003 ; Jeyasingh et al ., 2009 ), 植物叶片N:P的变化表征了植物维持体内化学计量内稳性的能力(Sterner & Elser, 2002 ; Demars & Edwards, 2007 ), 反映了生物对环境变化的适应性(Elser et al ., 2010a ).当外界环境发生改变时, 植物通常倾向于保持相对稳定的N:P来满足自身功能的稳定(Demars & Edwards, 2007 ; Elser et al ., 2010b ; Irakli & Elser, 2011 ).本研究5年的N添加实验表明: 乔木叶片的N:P相对稳定, 变化不明显; 而草本层植物叶片的N:P提高显著, 这预示着短期N添加条件下, 乔木植物叶片N:P较林下植物更加稳定.就灌木层植物而言, 其N:P的N添加效果表现出明显的时间效应(图4 ).随着N添加年限的增加, 灌木植物N:P对N添加的响应逐渐增强, 这符合Aber等(1998)提出的N滞留假设, 即: N添加的影响存在累积效应, 过去添加的N仍会对现在的植物化学计量特征产生影响. ...
Effects of soil nutrient availability on investment in acquisition of N and P in Hawaiian Rain Forests
1
2001
... 本研究表明N添加对植物P元素的吸收有促进作用, 这与鼎湖山亚热带N添加实验的结果(Huang et al ., 2012 )相一致, 但与一些温带森林的研究结果 (Nilsson & Wallander, 2003 ; Braun et al ., 2010 )不同.N添加引起植物P含量增加的原因通常包含两方面: 一方面, N添加通过提高土壤中P的可利用性, 提高植物体内的P含量, 这与本研究中表层土壤P含量显著增加的结果(表1 )相一致.另一方面, Fujita等(2010)的研究表明, N添加会通过促进根系磷酸酶活性等方式提高植物对土壤中P的吸收能力.此外, N添加条件下植物可以将自身多余的N用于构造更多的磷酸酶, 从而促进对土壤P元素的吸收, 进而导致叶片P含量上升(Kathleen & Vitousek, 2001 ). ...
The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation
1996
Nitrogen to phosphorus ratios of tree species in response to elevated carbon dioxide and nitrogen addition in subtropical forests
1
2013
... 研究显示N添加背景下, 武夷山植物叶片N:P的变化主要受叶片P含量变化影响, 而非受N含量变化影响(图3 ).其他研究结果也显示, 在N元素相对充足而P元素相对匮乏的生态系统, N添加会明显改变该生态系统的P循环(Güsewell, 2004 ; Liu et al ., 2013 ).N、P元素从细胞到生态系统尺度, 都存在紧密的耦合作用(Elser et al ., 2000 ; Sterner & Elser, 2002 ; Vitousek et al ., 2010 ), 其中一种元素的吸收受到另一种元素可利用性的影响(Ågren et al ., 2012 ), 因此N添加会通过影响土壤中P的可利用性、促进根系磷酸酶活性提高植物对P的吸收等方式改变整个生态系统的P循环(Treseder & Vitousek, 2001 ; Marklein & Houlton, 2012 ).本研究结果进一步表明, N添加对武夷山植物群落P循环的影响明显, 植物叶片P含量变化是叶片N:P变化的主要原因. ...
Effects of experimental nitrogen additions on plant diversity in an old-growth tropical forest
2
2010
... 人类活动导致大气氮(N)沉降不断增强(Galloway et al ., 2004 ; Dentener et al ., 2006 ; Mahowald et al ., 2008 ), 正深刻影响着生态系统及其养分循环过程(Vitousek & Tilman, 1997 ; Galloway & Sutton, 2008 ; Treseder, 2008 ).野外人工N添加实验是模拟自然N沉降增加对生态系统影响的重要手段.自20世纪80年代起, 研究者在温带和北方森林地区陆续开展了大量长时间的野外N添加实验(Aber & Melillo, 1989 ; Wright & Rasmussen, 1998 ; Magill et al ., 2000 , 2004 ), 而N添加实验在亚热带常绿阔叶林地区开展得相对较少(Mo et al ., 2008 ; Lu et al ., 2010 ).我国的亚热带常绿阔叶林面积广阔、物种组成丰富(Fang et al ., 2014 ), 是全球亚热带常绿阔叶林的主要分布区, 在我国亚热带常绿阔叶林地区开展N添加的生态学效应的研究十分必要. ...
... 武夷山米槠群落植物叶片N、P含量平均值低于世界和全国平均水平, 叶片N:P则高于世界和全国平均水平(Reich & Oleksyn, 2004 ; Han et al ., 2005 ).本研究区植物生长受P元素限制更加明显.这与亚热带地区较低的土壤养分含量、较快的养分周转速率以及较强的淋溶等环境特点相适应(方运霆等, 2004 ; Reich & Oleksyn, 2004 ; Lu et al ., 2010 ). ...
Long-term nitrogen additions and nitrogen saturation in two temperate forests
3
2000
... 人类活动导致大气氮(N)沉降不断增强(Galloway et al ., 2004 ; Dentener et al ., 2006 ; Mahowald et al ., 2008 ), 正深刻影响着生态系统及其养分循环过程(Vitousek & Tilman, 1997 ; Galloway & Sutton, 2008 ; Treseder, 2008 ).野外人工N添加实验是模拟自然N沉降增加对生态系统影响的重要手段.自20世纪80年代起, 研究者在温带和北方森林地区陆续开展了大量长时间的野外N添加实验(Aber & Melillo, 1989 ; Wright & Rasmussen, 1998 ; Magill et al ., 2000 , 2004 ), 而N添加实验在亚热带常绿阔叶林地区开展得相对较少(Mo et al ., 2008 ; Lu et al ., 2010 ).我国的亚热带常绿阔叶林面积广阔、物种组成丰富(Fang et al ., 2014 ), 是全球亚热带常绿阔叶林的主要分布区, 在我国亚热带常绿阔叶林地区开展N添加的生态学效应的研究十分必要. ...
... 在全球变化的背景下, 植物的化学计量特征可以表征植物维持体内化学计量内稳性的能力(Sterner & Elser, 2002 ; Demars & Edwards, 2007 ), 并反映植物对环境变化的适应性(Elser et al ., 2010a ).以往的研究表明, 植物叶片N、磷(P)化学计量特征因植被类型、功能群、物种的差异而不同(Sterner & Elser, 2002 ; Reich, 2003 ; Ågren, 2004 ), 不同功能群和物种对N添加的响应也存在差异(Güsewell, 2004 ).自20世纪80年代起, 研究人员在温带和北方森林地区陆续开展了一些经典的野外N添加实验(Aber & Melillo, 1989 ; Wright & Rasmussen, 1998 ; Magill et al ., 2000 ), 植物化学计量学作为研究N循环的新手段, 也被应用于N添加生态学效应的研究中.例如: Magill等(2004)在美国哈佛森林开展了长期的N添加试验, 经过15年的研究发现: N添加显著增加了乔木优势种的叶片N含量, 并且叶片化学计量特征对N添加的响应存在明显的年际变化; Huang等(2012)在模拟的亚热带常绿阔叶林生态系统中开展了N添加实验, 对群落内5种乔木树种叶片N、P化学计量特征进行了研究, 结果显示N添加显著影响了多数物种的叶片N、P含量和N:P; Cui等(2010)对温带草原8种优势植物叶片化学计量特征对N添加的响应进行了研究, 发现不同物种的叶片化学计量特征对N添加的响应也存在差异. ...
... N添加整体上促进了植物对N的吸收, 这与以往的研究结果(Magill et al ., 2000 ; Mcnulty et al ., 2005 ; Rowe et al ., 2008 )一致.植物叶片的矿质元素含量取决于土壤养分的可利用性(Townsend & Bustamante, 2007 ).以往的研究也显示, 植物组织内的矿质元素含量与土壤中相应元素含量呈现显著正相关关系(Reich & Oleksyn, 2004 ), 当土壤中某种矿质元素含量增加时, 植物体内相应的矿质元素含量也会随之增加.功能群和物种则是影响叶片N、P化学计量特征及其对N添加响应的重要因素(Reich & Bowman, 1999 ; Sterner & Elser, 2002 ; Sardans et al ., 2012 ).N添加情况下, 植物对N、P等矿质元素的利用策略因需求、竞争力和优势度的不同而存在差异(Tilman, 1990 ; Güsewell & Koerselman, 2002 ).本研究中, 乔木优势种米槠对N添加响应敏感, N添加显著增加了其叶片N含量(图2 ).这可能是由于当养分条件发生变化时, 生态系统趋向于将N、P等矿质元素运输到生产力较高的植物中去, 优先保证其生长需求(Aber & Melillo, 1989 ; Elliott & White, 1994 ).本研究中林下草本层植物N含量对N添加的响应也较为敏感(图2 ).以往的研究显示, 草本、蕨类、苔藓等速生型植物对N添加的敏感性明显高于其他物种, 这可能与速生物种较快的养分转换速率 及其养分策略可以循环利用更多的N有关(Aerts & Peijl, 1993 ). ...
Ecosystem response to 15 years of chronic nitrogen additions at the Harvard forest LTER, Massachusetts, USA
1
2004
... 人类活动导致大气氮(N)沉降不断增强(Galloway et al ., 2004 ; Dentener et al ., 2006 ; Mahowald et al ., 2008 ), 正深刻影响着生态系统及其养分循环过程(Vitousek & Tilman, 1997 ; Galloway & Sutton, 2008 ; Treseder, 2008 ).野外人工N添加实验是模拟自然N沉降增加对生态系统影响的重要手段.自20世纪80年代起, 研究者在温带和北方森林地区陆续开展了大量长时间的野外N添加实验(Aber & Melillo, 1989 ; Wright & Rasmussen, 1998 ; Magill et al ., 2000 , 2004 ), 而N添加实验在亚热带常绿阔叶林地区开展得相对较少(Mo et al ., 2008 ; Lu et al ., 2010 ).我国的亚热带常绿阔叶林面积广阔、物种组成丰富(Fang et al ., 2014 ), 是全球亚热带常绿阔叶林的主要分布区, 在我国亚热带常绿阔叶林地区开展N添加的生态学效应的研究十分必要. ...
Global distribution of atmospheric phosphorus sources, concentrations and deposition rates, and anthropogenic impacts
1
2008
... 人类活动导致大气氮(N)沉降不断增强(Galloway et al ., 2004 ; Dentener et al ., 2006 ; Mahowald et al ., 2008 ), 正深刻影响着生态系统及其养分循环过程(Vitousek & Tilman, 1997 ; Galloway & Sutton, 2008 ; Treseder, 2008 ).野外人工N添加实验是模拟自然N沉降增加对生态系统影响的重要手段.自20世纪80年代起, 研究者在温带和北方森林地区陆续开展了大量长时间的野外N添加实验(Aber & Melillo, 1989 ; Wright & Rasmussen, 1998 ; Magill et al ., 2000 , 2004 ), 而N添加实验在亚热带常绿阔叶林地区开展得相对较少(Mo et al ., 2008 ; Lu et al ., 2010 ).我国的亚热带常绿阔叶林面积广阔、物种组成丰富(Fang et al ., 2014 ), 是全球亚热带常绿阔叶林的主要分布区, 在我国亚热带常绿阔叶林地区开展N添加的生态学效应的研究十分必要. ...
Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems
1
2012
... 研究显示N添加背景下, 武夷山植物叶片N:P的变化主要受叶片P含量变化影响, 而非受N含量变化影响(图3 ).其他研究结果也显示, 在N元素相对充足而P元素相对匮乏的生态系统, N添加会明显改变该生态系统的P循环(Güsewell, 2004 ; Liu et al ., 2013 ).N、P元素从细胞到生态系统尺度, 都存在紧密的耦合作用(Elser et al ., 2000 ; Sterner & Elser, 2002 ; Vitousek et al ., 2010 ), 其中一种元素的吸收受到另一种元素可利用性的影响(Ågren et al ., 2012 ), 因此N添加会通过影响土壤中P的可利用性、促进根系磷酸酶活性提高植物对P的吸收等方式改变整个生态系统的P循环(Treseder & Vitousek, 2001 ; Marklein & Houlton, 2012 ).本研究结果进一步表明, N添加对武夷山植物群落P循环的影响明显, 植物叶片P含量变化是叶片N:P变化的主要原因. ...
Red spruce ecosystem level changes following 14 years of chronic N fertilization
1
2005
... N添加整体上促进了植物对N的吸收, 这与以往的研究结果(Magill et al ., 2000 ; Mcnulty et al ., 2005 ; Rowe et al ., 2008 )一致.植物叶片的矿质元素含量取决于土壤养分的可利用性(Townsend & Bustamante, 2007 ).以往的研究也显示, 植物组织内的矿质元素含量与土壤中相应元素含量呈现显著正相关关系(Reich & Oleksyn, 2004 ), 当土壤中某种矿质元素含量增加时, 植物体内相应的矿质元素含量也会随之增加.功能群和物种则是影响叶片N、P化学计量特征及其对N添加响应的重要因素(Reich & Bowman, 1999 ; Sterner & Elser, 2002 ; Sardans et al ., 2012 ).N添加情况下, 植物对N、P等矿质元素的利用策略因需求、竞争力和优势度的不同而存在差异(Tilman, 1990 ; Güsewell & Koerselman, 2002 ).本研究中, 乔木优势种米槠对N添加响应敏感, N添加显著增加了其叶片N含量(图2 ).这可能是由于当养分条件发生变化时, 生态系统趋向于将N、P等矿质元素运输到生产力较高的植物中去, 优先保证其生长需求(Aber & Melillo, 1989 ; Elliott & White, 1994 ).本研究中林下草本层植物N含量对N添加的响应也较为敏感(图2 ).以往的研究显示, 草本、蕨类、苔藓等速生型植物对N添加的敏感性明显高于其他物种, 这可能与速生物种较快的养分转换速率 及其养分策略可以循环利用更多的N有关(Aerts & Peijl, 1993 ). ...
Seedling growth response of two tropical tree species to nitrogen deposition in southern China
1
2008
... 人类活动导致大气氮(N)沉降不断增强(Galloway et al ., 2004 ; Dentener et al ., 2006 ; Mahowald et al ., 2008 ), 正深刻影响着生态系统及其养分循环过程(Vitousek & Tilman, 1997 ; Galloway & Sutton, 2008 ; Treseder, 2008 ).野外人工N添加实验是模拟自然N沉降增加对生态系统影响的重要手段.自20世纪80年代起, 研究者在温带和北方森林地区陆续开展了大量长时间的野外N添加实验(Aber & Melillo, 1989 ; Wright & Rasmussen, 1998 ; Magill et al ., 2000 , 2004 ), 而N添加实验在亚热带常绿阔叶林地区开展得相对较少(Mo et al ., 2008 ; Lu et al ., 2010 ).我国的亚热带常绿阔叶林面积广阔、物种组成丰富(Fang et al ., 2014 ), 是全球亚热带常绿阔叶林的主要分布区, 在我国亚热带常绿阔叶林地区开展N添加的生态学效应的研究十分必要. ...
Production of external mycelium by ectomycorrhizal fungi in a Norway spruce forest was reduced in response to nitrogen fertilization
1
2003
... 本研究表明N添加对植物P元素的吸收有促进作用, 这与鼎湖山亚热带N添加实验的结果(Huang et al ., 2012 )相一致, 但与一些温带森林的研究结果 (Nilsson & Wallander, 2003 ; Braun et al ., 2010 )不同.N添加引起植物P含量增加的原因通常包含两方面: 一方面, N添加通过提高土壤中P的可利用性, 提高植物体内的P含量, 这与本研究中表层土壤P含量显著增加的结果(表1 )相一致.另一方面, Fujita等(2010)的研究表明, N添加会通过促进根系磷酸酶活性等方式提高植物对土壤中P的吸收能力.此外, N添加条件下植物可以将自身多余的N用于构造更多的磷酸酶, 从而促进对土壤P元素的吸收, 进而导致叶片P含量上升(Kathleen & Vitousek, 2001 ). ...
The evolution of plant functional variation: Traits, spectra and strategies
2
2003
... 在全球变化的背景下, 植物的化学计量特征可以表征植物维持体内化学计量内稳性的能力(Sterner & Elser, 2002 ; Demars & Edwards, 2007 ), 并反映植物对环境变化的适应性(Elser et al ., 2010a ).以往的研究表明, 植物叶片N、磷(P)化学计量特征因植被类型、功能群、物种的差异而不同(Sterner & Elser, 2002 ; Reich, 2003 ; Ågren, 2004 ), 不同功能群和物种对N添加的响应也存在差异(Güsewell, 2004 ).自20世纪80年代起, 研究人员在温带和北方森林地区陆续开展了一些经典的野外N添加实验(Aber & Melillo, 1989 ; Wright & Rasmussen, 1998 ; Magill et al ., 2000 ), 植物化学计量学作为研究N循环的新手段, 也被应用于N添加生态学效应的研究中.例如: Magill等(2004)在美国哈佛森林开展了长期的N添加试验, 经过15年的研究发现: N添加显著增加了乔木优势种的叶片N含量, 并且叶片化学计量特征对N添加的响应存在明显的年际变化; Huang等(2012)在模拟的亚热带常绿阔叶林生态系统中开展了N添加实验, 对群落内5种乔木树种叶片N、P化学计量特征进行了研究, 结果显示N添加显著影响了多数物种的叶片N、P含量和N:P; Cui等(2010)对温带草原8种优势植物叶片化学计量特征对N添加的响应进行了研究, 发现不同物种的叶片化学计量特征对N添加的响应也存在差异. ...
... 植物叶片N:P与物种的生态策略有关(Reich, 2003 ; Jeyasingh et al ., 2009 ), 植物叶片N:P的变化表征了植物维持体内化学计量内稳性的能力(Sterner & Elser, 2002 ; Demars & Edwards, 2007 ), 反映了生物对环境变化的适应性(Elser et al ., 2010a ).当外界环境发生改变时, 植物通常倾向于保持相对稳定的N:P来满足自身功能的稳定(Demars & Edwards, 2007 ; Elser et al ., 2010b ; Irakli & Elser, 2011 ).本研究5年的N添加实验表明: 乔木叶片的N:P相对稳定, 变化不明显; 而草本层植物叶片的N:P提高显著, 这预示着短期N添加条件下, 乔木植物叶片N:P较林下植物更加稳定.就灌木层植物而言, 其N:P的N添加效果表现出明显的时间效应(图4 ).随着N添加年限的增加, 灌木植物N:P对N添加的响应逐渐增强, 这符合Aber等(1998)提出的N滞留假设, 即: N添加的影响存在累积效应, 过去添加的N仍会对现在的植物化学计量特征产生影响. ...
Generality of leaf trait relationships: A test across six biomes
2
1999
... 不同功能群物种叶片的N、P含量均存在显著差异, 这与以往的研究结果(Reich & Bowman, 1999 ; Sterner & Elser, 2002 ; Sardans et al ., 2012 )较为一致.本研究显示, 草本层植物叶片的N、P含量显著高于木本植物(包括乔木和灌木植物), 针叶植物叶片的N、P含量显著高于阔叶植物, 这一结果与其他研究中得到的中国草本和木本植物的N、P化学计量模式基本一致(高三平等, 2007 ; He et al ., 2008 ; Han et al ., 2011 ), 并且证明了生命周期短而生长快速的植物要比生命周期长而生长缓慢的植物具有更高的叶片N、P含量(Aerts, 1996 ; Thompson et al ., 1997 ; Güsewell & Koerselman, 2002 ). ...
... N添加整体上促进了植物对N的吸收, 这与以往的研究结果(Magill et al ., 2000 ; Mcnulty et al ., 2005 ; Rowe et al ., 2008 )一致.植物叶片的矿质元素含量取决于土壤养分的可利用性(Townsend & Bustamante, 2007 ).以往的研究也显示, 植物组织内的矿质元素含量与土壤中相应元素含量呈现显著正相关关系(Reich & Oleksyn, 2004 ), 当土壤中某种矿质元素含量增加时, 植物体内相应的矿质元素含量也会随之增加.功能群和物种则是影响叶片N、P化学计量特征及其对N添加响应的重要因素(Reich & Bowman, 1999 ; Sterner & Elser, 2002 ; Sardans et al ., 2012 ).N添加情况下, 植物对N、P等矿质元素的利用策略因需求、竞争力和优势度的不同而存在差异(Tilman, 1990 ; Güsewell & Koerselman, 2002 ).本研究中, 乔木优势种米槠对N添加响应敏感, N添加显著增加了其叶片N含量(图2 ).这可能是由于当养分条件发生变化时, 生态系统趋向于将N、P等矿质元素运输到生产力较高的植物中去, 优先保证其生长需求(Aber & Melillo, 1989 ; Elliott & White, 1994 ).本研究中林下草本层植物N含量对N添加的响应也较为敏感(图2 ).以往的研究显示, 草本、蕨类、苔藓等速生型植物对N添加的敏感性明显高于其他物种, 这可能与速生物种较快的养分转换速率 及其养分策略可以循环利用更多的N有关(Aerts & Peijl, 1993 ). ...
Global patterns of plant leaf N and P in relation to temperature and latitude
3
2004
... 武夷山米槠群落植物叶片N、P含量平均值低于世界和全国平均水平, 叶片N:P则高于世界和全国平均水平(Reich & Oleksyn, 2004 ; Han et al ., 2005 ).本研究区植物生长受P元素限制更加明显.这与亚热带地区较低的土壤养分含量、较快的养分周转速率以及较强的淋溶等环境特点相适应(方运霆等, 2004 ; Reich & Oleksyn, 2004 ; Lu et al ., 2010 ). ...
... ; Reich & Oleksyn, 2004 ; Lu et al ., 2010 ). ...
... N添加整体上促进了植物对N的吸收, 这与以往的研究结果(Magill et al ., 2000 ; Mcnulty et al ., 2005 ; Rowe et al ., 2008 )一致.植物叶片的矿质元素含量取决于土壤养分的可利用性(Townsend & Bustamante, 2007 ).以往的研究也显示, 植物组织内的矿质元素含量与土壤中相应元素含量呈现显著正相关关系(Reich & Oleksyn, 2004 ), 当土壤中某种矿质元素含量增加时, 植物体内相应的矿质元素含量也会随之增加.功能群和物种则是影响叶片N、P化学计量特征及其对N添加响应的重要因素(Reich & Bowman, 1999 ; Sterner & Elser, 2002 ; Sardans et al ., 2012 ).N添加情况下, 植物对N、P等矿质元素的利用策略因需求、竞争力和优势度的不同而存在差异(Tilman, 1990 ; Güsewell & Koerselman, 2002 ).本研究中, 乔木优势种米槠对N添加响应敏感, N添加显著增加了其叶片N含量(图2 ).这可能是由于当养分条件发生变化时, 生态系统趋向于将N、P等矿质元素运输到生产力较高的植物中去, 优先保证其生长需求(Aber & Melillo, 1989 ; Elliott & White, 1994 ).本研究中林下草本层植物N含量对N添加的响应也较为敏感(图2 ).以往的研究显示, 草本、蕨类、苔藓等速生型植物对N添加的敏感性明显高于其他物种, 这可能与速生物种较快的养分转换速率 及其养分策略可以循环利用更多的N有关(Aerts & Peijl, 1993 ). ...
Nitrogen deposition increases the acquisition of phosphorus and potassium by heather calluna vulgaris
1
2008
... N添加整体上促进了植物对N的吸收, 这与以往的研究结果(Magill et al ., 2000 ; Mcnulty et al ., 2005 ; Rowe et al ., 2008 )一致.植物叶片的矿质元素含量取决于土壤养分的可利用性(Townsend & Bustamante, 2007 ).以往的研究也显示, 植物组织内的矿质元素含量与土壤中相应元素含量呈现显著正相关关系(Reich & Oleksyn, 2004 ), 当土壤中某种矿质元素含量增加时, 植物体内相应的矿质元素含量也会随之增加.功能群和物种则是影响叶片N、P化学计量特征及其对N添加响应的重要因素(Reich & Bowman, 1999 ; Sterner & Elser, 2002 ; Sardans et al ., 2012 ).N添加情况下, 植物对N、P等矿质元素的利用策略因需求、竞争力和优势度的不同而存在差异(Tilman, 1990 ; Güsewell & Koerselman, 2002 ).本研究中, 乔木优势种米槠对N添加响应敏感, N添加显著增加了其叶片N含量(图2 ).这可能是由于当养分条件发生变化时, 生态系统趋向于将N、P等矿质元素运输到生产力较高的植物中去, 优先保证其生长需求(Aber & Melillo, 1989 ; Elliott & White, 1994 ).本研究中林下草本层植物N含量对N添加的响应也较为敏感(图2 ).以往的研究显示, 草本、蕨类、苔藓等速生型植物对N添加的敏感性明显高于其他物种, 这可能与速生物种较快的养分转换速率 及其养分策略可以循环利用更多的N有关(Aerts & Peijl, 1993 ). ...
The C:N:P stoichiometry of organisms and ecosystems in a changing world: A review and perspectives
3
2012
... 不同功能群物种叶片的N、P含量均存在显著差异, 这与以往的研究结果(Reich & Bowman, 1999 ; Sterner & Elser, 2002 ; Sardans et al ., 2012 )较为一致.本研究显示, 草本层植物叶片的N、P含量显著高于木本植物(包括乔木和灌木植物), 针叶植物叶片的N、P含量显著高于阔叶植物, 这一结果与其他研究中得到的中国草本和木本植物的N、P化学计量模式基本一致(高三平等, 2007 ; He et al ., 2008 ; Han et al ., 2011 ), 并且证明了生命周期短而生长快速的植物要比生命周期长而生长缓慢的植物具有更高的叶片N、P含量(Aerts, 1996 ; Thompson et al ., 1997 ; Güsewell & Koerselman, 2002 ). ...
... N添加整体上促进了植物对N的吸收, 这与以往的研究结果(Magill et al ., 2000 ; Mcnulty et al ., 2005 ; Rowe et al ., 2008 )一致.植物叶片的矿质元素含量取决于土壤养分的可利用性(Townsend & Bustamante, 2007 ).以往的研究也显示, 植物组织内的矿质元素含量与土壤中相应元素含量呈现显著正相关关系(Reich & Oleksyn, 2004 ), 当土壤中某种矿质元素含量增加时, 植物体内相应的矿质元素含量也会随之增加.功能群和物种则是影响叶片N、P化学计量特征及其对N添加响应的重要因素(Reich & Bowman, 1999 ; Sterner & Elser, 2002 ; Sardans et al ., 2012 ).N添加情况下, 植物对N、P等矿质元素的利用策略因需求、竞争力和优势度的不同而存在差异(Tilman, 1990 ; Güsewell & Koerselman, 2002 ).本研究中, 乔木优势种米槠对N添加响应敏感, N添加显著增加了其叶片N含量(图2 ).这可能是由于当养分条件发生变化时, 生态系统趋向于将N、P等矿质元素运输到生产力较高的植物中去, 优先保证其生长需求(Aber & Melillo, 1989 ; Elliott & White, 1994 ).本研究中林下草本层植物N含量对N添加的响应也较为敏感(图2 ).以往的研究显示, 草本、蕨类、苔藓等速生型植物对N添加的敏感性明显高于其他物种, 这可能与速生物种较快的养分转换速率 及其养分策略可以循环利用更多的N有关(Aerts & Peijl, 1993 ). ...
... 植物N:P可以用来指示N、P元素对生长限制的相对强弱(Güsewell & Koerselman, 2002 ; Tessier & Raynal, 2003 ; Güsewell, 2004 ).Koerselman和Meu- leman (1996)提出判断植物N、P营养限制的标准: 当植物叶片N:P小于14时, 植物生长受N元素限制; 当N:P介于14和16之间时, 植物生长受N和P元素共同限制; 当N:P大于16时, 植物生长受P元素限制.本实验中对照样地的植物叶片平均N:P为18.67, 各物种N:P均大于16, 表明N添加样地土壤P元素供应短缺, 植物生长主要受P元素限制.这与Aerts等(2003)关于亚热带地区森林生长主要受土壤P限制的论述一致, 并与Han等(2005)关于我国大部分地区P元素缺乏的结果相符合.本研究发现, 随着N添加量的增加, 植物叶片N:P逐渐升高, N:P由对照样地的18.67增加到19.72, 说明随着N添加量的增加, 植物生长的P限制增强.这与其他研究中关于N沉降使陆地生态系统从N限制向P限制转变的结论(Elser et al ., 2007 ; Sardans et al ., 2012 )相一致. ...
6
2002
... 在全球变化的背景下, 植物的化学计量特征可以表征植物维持体内化学计量内稳性的能力(Sterner & Elser, 2002 ; Demars & Edwards, 2007 ), 并反映植物对环境变化的适应性(Elser et al ., 2010a ).以往的研究表明, 植物叶片N、磷(P)化学计量特征因植被类型、功能群、物种的差异而不同(Sterner & Elser, 2002 ; Reich, 2003 ; Ågren, 2004 ), 不同功能群和物种对N添加的响应也存在差异(Güsewell, 2004 ).自20世纪80年代起, 研究人员在温带和北方森林地区陆续开展了一些经典的野外N添加实验(Aber & Melillo, 1989 ; Wright & Rasmussen, 1998 ; Magill et al ., 2000 ), 植物化学计量学作为研究N循环的新手段, 也被应用于N添加生态学效应的研究中.例如: Magill等(2004)在美国哈佛森林开展了长期的N添加试验, 经过15年的研究发现: N添加显著增加了乔木优势种的叶片N含量, 并且叶片化学计量特征对N添加的响应存在明显的年际变化; Huang等(2012)在模拟的亚热带常绿阔叶林生态系统中开展了N添加实验, 对群落内5种乔木树种叶片N、P化学计量特征进行了研究, 结果显示N添加显著影响了多数物种的叶片N、P含量和N:P; Cui等(2010)对温带草原8种优势植物叶片化学计量特征对N添加的响应进行了研究, 发现不同物种的叶片化学计量特征对N添加的响应也存在差异. ...
... ).以往的研究表明, 植物叶片N、磷(P)化学计量特征因植被类型、功能群、物种的差异而不同(Sterner & Elser, 2002 ; Reich, 2003 ; Ågren, 2004 ), 不同功能群和物种对N添加的响应也存在差异(Güsewell, 2004 ).自20世纪80年代起, 研究人员在温带和北方森林地区陆续开展了一些经典的野外N添加实验(Aber & Melillo, 1989 ; Wright & Rasmussen, 1998 ; Magill et al ., 2000 ), 植物化学计量学作为研究N循环的新手段, 也被应用于N添加生态学效应的研究中.例如: Magill等(2004)在美国哈佛森林开展了长期的N添加试验, 经过15年的研究发现: N添加显著增加了乔木优势种的叶片N含量, 并且叶片化学计量特征对N添加的响应存在明显的年际变化; Huang等(2012)在模拟的亚热带常绿阔叶林生态系统中开展了N添加实验, 对群落内5种乔木树种叶片N、P化学计量特征进行了研究, 结果显示N添加显著影响了多数物种的叶片N、P含量和N:P; Cui等(2010)对温带草原8种优势植物叶片化学计量特征对N添加的响应进行了研究, 发现不同物种的叶片化学计量特征对N添加的响应也存在差异. ...
... 不同功能群物种叶片的N、P含量均存在显著差异, 这与以往的研究结果(Reich & Bowman, 1999 ; Sterner & Elser, 2002 ; Sardans et al ., 2012 )较为一致.本研究显示, 草本层植物叶片的N、P含量显著高于木本植物(包括乔木和灌木植物), 针叶植物叶片的N、P含量显著高于阔叶植物, 这一结果与其他研究中得到的中国草本和木本植物的N、P化学计量模式基本一致(高三平等, 2007 ; He et al ., 2008 ; Han et al ., 2011 ), 并且证明了生命周期短而生长快速的植物要比生命周期长而生长缓慢的植物具有更高的叶片N、P含量(Aerts, 1996 ; Thompson et al ., 1997 ; Güsewell & Koerselman, 2002 ). ...
... N添加整体上促进了植物对N的吸收, 这与以往的研究结果(Magill et al ., 2000 ; Mcnulty et al ., 2005 ; Rowe et al ., 2008 )一致.植物叶片的矿质元素含量取决于土壤养分的可利用性(Townsend & Bustamante, 2007 ).以往的研究也显示, 植物组织内的矿质元素含量与土壤中相应元素含量呈现显著正相关关系(Reich & Oleksyn, 2004 ), 当土壤中某种矿质元素含量增加时, 植物体内相应的矿质元素含量也会随之增加.功能群和物种则是影响叶片N、P化学计量特征及其对N添加响应的重要因素(Reich & Bowman, 1999 ; Sterner & Elser, 2002 ; Sardans et al ., 2012 ).N添加情况下, 植物对N、P等矿质元素的利用策略因需求、竞争力和优势度的不同而存在差异(Tilman, 1990 ; Güsewell & Koerselman, 2002 ).本研究中, 乔木优势种米槠对N添加响应敏感, N添加显著增加了其叶片N含量(图2 ).这可能是由于当养分条件发生变化时, 生态系统趋向于将N、P等矿质元素运输到生产力较高的植物中去, 优先保证其生长需求(Aber & Melillo, 1989 ; Elliott & White, 1994 ).本研究中林下草本层植物N含量对N添加的响应也较为敏感(图2 ).以往的研究显示, 草本、蕨类、苔藓等速生型植物对N添加的敏感性明显高于其他物种, 这可能与速生物种较快的养分转换速率 及其养分策略可以循环利用更多的N有关(Aerts & Peijl, 1993 ). ...
... 植物叶片N:P与物种的生态策略有关(Reich, 2003 ; Jeyasingh et al ., 2009 ), 植物叶片N:P的变化表征了植物维持体内化学计量内稳性的能力(Sterner & Elser, 2002 ; Demars & Edwards, 2007 ), 反映了生物对环境变化的适应性(Elser et al ., 2010a ).当外界环境发生改变时, 植物通常倾向于保持相对稳定的N:P来满足自身功能的稳定(Demars & Edwards, 2007 ; Elser et al ., 2010b ; Irakli & Elser, 2011 ).本研究5年的N添加实验表明: 乔木叶片的N:P相对稳定, 变化不明显; 而草本层植物叶片的N:P提高显著, 这预示着短期N添加条件下, 乔木植物叶片N:P较林下植物更加稳定.就灌木层植物而言, 其N:P的N添加效果表现出明显的时间效应(图4 ).随着N添加年限的增加, 灌木植物N:P对N添加的响应逐渐增强, 这符合Aber等(1998)提出的N滞留假设, 即: N添加的影响存在累积效应, 过去添加的N仍会对现在的植物化学计量特征产生影响. ...
... 研究显示N添加背景下, 武夷山植物叶片N:P的变化主要受叶片P含量变化影响, 而非受N含量变化影响(图3 ).其他研究结果也显示, 在N元素相对充足而P元素相对匮乏的生态系统, N添加会明显改变该生态系统的P循环(Güsewell, 2004 ; Liu et al ., 2013 ).N、P元素从细胞到生态系统尺度, 都存在紧密的耦合作用(Elser et al ., 2000 ; Sterner & Elser, 2002 ; Vitousek et al ., 2010 ), 其中一种元素的吸收受到另一种元素可利用性的影响(Ågren et al ., 2012 ), 因此N添加会通过影响土壤中P的可利用性、促进根系磷酸酶活性提高植物对P的吸收等方式改变整个生态系统的P循环(Treseder & Vitousek, 2001 ; Marklein & Houlton, 2012 ).本研究结果进一步表明, N添加对武夷山植物群落P循环的影响明显, 植物叶片P含量变化是叶片N:P变化的主要原因. ...
Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation
1
2003
... 植物N:P可以用来指示N、P元素对生长限制的相对强弱(Güsewell & Koerselman, 2002 ; Tessier & Raynal, 2003 ; Güsewell, 2004 ).Koerselman和Meu- leman (1996)提出判断植物N、P营养限制的标准: 当植物叶片N:P小于14时, 植物生长受N元素限制; 当N:P介于14和16之间时, 植物生长受N和P元素共同限制; 当N:P大于16时, 植物生长受P元素限制.本实验中对照样地的植物叶片平均N:P为18.67, 各物种N:P均大于16, 表明N添加样地土壤P元素供应短缺, 植物生长主要受P元素限制.这与Aerts等(2003)关于亚热带地区森林生长主要受土壤P限制的论述一致, 并与Han等(2005)关于我国大部分地区P元素缺乏的结果相符合.本研究发现, 随着N添加量的增加, 植物叶片N:P逐渐升高, N:P由对照样地的18.67增加到19.72, 说明随着N添加量的增加, 植物生长的P限制增强.这与其他研究中关于N沉降使陆地生态系统从N限制向P限制转变的结论(Elser et al ., 2007 ; Sardans et al ., 2012 )相一致. ...
A comparative study of leaf nutrient concentrations in a regional herbaceous flora
1
1997
... 不同功能群物种叶片的N、P含量均存在显著差异, 这与以往的研究结果(Reich & Bowman, 1999 ; Sterner & Elser, 2002 ; Sardans et al ., 2012 )较为一致.本研究显示, 草本层植物叶片的N、P含量显著高于木本植物(包括乔木和灌木植物), 针叶植物叶片的N、P含量显著高于阔叶植物, 这一结果与其他研究中得到的中国草本和木本植物的N、P化学计量模式基本一致(高三平等, 2007 ; He et al ., 2008 ; Han et al ., 2011 ), 并且证明了生命周期短而生长快速的植物要比生命周期长而生长缓慢的植物具有更高的叶片N、P含量(Aerts, 1996 ; Thompson et al ., 1997 ; Güsewell & Koerselman, 2002 ). ...
Constraints and tradeoffs: Toward a predictive theory of competition and succession
1
1990
... N添加整体上促进了植物对N的吸收, 这与以往的研究结果(Magill et al ., 2000 ; Mcnulty et al ., 2005 ; Rowe et al ., 2008 )一致.植物叶片的矿质元素含量取决于土壤养分的可利用性(Townsend & Bustamante, 2007 ).以往的研究也显示, 植物组织内的矿质元素含量与土壤中相应元素含量呈现显著正相关关系(Reich & Oleksyn, 2004 ), 当土壤中某种矿质元素含量增加时, 植物体内相应的矿质元素含量也会随之增加.功能群和物种则是影响叶片N、P化学计量特征及其对N添加响应的重要因素(Reich & Bowman, 1999 ; Sterner & Elser, 2002 ; Sardans et al ., 2012 ).N添加情况下, 植物对N、P等矿质元素的利用策略因需求、竞争力和优势度的不同而存在差异(Tilman, 1990 ; Güsewell & Koerselman, 2002 ).本研究中, 乔木优势种米槠对N添加响应敏感, N添加显著增加了其叶片N含量(图2 ).这可能是由于当养分条件发生变化时, 生态系统趋向于将N、P等矿质元素运输到生产力较高的植物中去, 优先保证其生长需求(Aber & Melillo, 1989 ; Elliott & White, 1994 ).本研究中林下草本层植物N含量对N添加的响应也较为敏感(图2 ).以往的研究显示, 草本、蕨类、苔藓等速生型植物对N添加的敏感性明显高于其他物种, 这可能与速生物种较快的养分转换速率 及其养分策略可以循环利用更多的N有关(Aerts & Peijl, 1993 ). ...
Controls over foliar N:P ratios in tropical rain forests
1
2007
... N添加整体上促进了植物对N的吸收, 这与以往的研究结果(Magill et al ., 2000 ; Mcnulty et al ., 2005 ; Rowe et al ., 2008 )一致.植物叶片的矿质元素含量取决于土壤养分的可利用性(Townsend & Bustamante, 2007 ).以往的研究也显示, 植物组织内的矿质元素含量与土壤中相应元素含量呈现显著正相关关系(Reich & Oleksyn, 2004 ), 当土壤中某种矿质元素含量增加时, 植物体内相应的矿质元素含量也会随之增加.功能群和物种则是影响叶片N、P化学计量特征及其对N添加响应的重要因素(Reich & Bowman, 1999 ; Sterner & Elser, 2002 ; Sardans et al ., 2012 ).N添加情况下, 植物对N、P等矿质元素的利用策略因需求、竞争力和优势度的不同而存在差异(Tilman, 1990 ; Güsewell & Koerselman, 2002 ).本研究中, 乔木优势种米槠对N添加响应敏感, N添加显著增加了其叶片N含量(图2 ).这可能是由于当养分条件发生变化时, 生态系统趋向于将N、P等矿质元素运输到生产力较高的植物中去, 优先保证其生长需求(Aber & Melillo, 1989 ; Elliott & White, 1994 ).本研究中林下草本层植物N含量对N添加的响应也较为敏感(图2 ).以往的研究显示, 草本、蕨类、苔藓等速生型植物对N添加的敏感性明显高于其他物种, 这可能与速生物种较快的养分转换速率 及其养分策略可以循环利用更多的N有关(Aerts & Peijl, 1993 ). ...
Nitrogen additions and microbial biomass: A meta-analysis of ecosystem studies
1
2008
... 人类活动导致大气氮(N)沉降不断增强(Galloway et al ., 2004 ; Dentener et al ., 2006 ; Mahowald et al ., 2008 ), 正深刻影响着生态系统及其养分循环过程(Vitousek & Tilman, 1997 ; Galloway & Sutton, 2008 ; Treseder, 2008 ).野外人工N添加实验是模拟自然N沉降增加对生态系统影响的重要手段.自20世纪80年代起, 研究者在温带和北方森林地区陆续开展了大量长时间的野外N添加实验(Aber & Melillo, 1989 ; Wright & Rasmussen, 1998 ; Magill et al ., 2000 , 2004 ), 而N添加实验在亚热带常绿阔叶林地区开展得相对较少(Mo et al ., 2008 ; Lu et al ., 2010 ).我国的亚热带常绿阔叶林面积广阔、物种组成丰富(Fang et al ., 2014 ), 是全球亚热带常绿阔叶林的主要分布区, 在我国亚热带常绿阔叶林地区开展N添加的生态学效应的研究十分必要. ...
Effects of soil nutrient availability on investment in acquisition of N and P in Hawaiian rain forests
1
2001
... 研究显示N添加背景下, 武夷山植物叶片N:P的变化主要受叶片P含量变化影响, 而非受N含量变化影响(图3 ).其他研究结果也显示, 在N元素相对充足而P元素相对匮乏的生态系统, N添加会明显改变该生态系统的P循环(Güsewell, 2004 ; Liu et al ., 2013 ).N、P元素从细胞到生态系统尺度, 都存在紧密的耦合作用(Elser et al ., 2000 ; Sterner & Elser, 2002 ; Vitousek et al ., 2010 ), 其中一种元素的吸收受到另一种元素可利用性的影响(Ågren et al ., 2012 ), 因此N添加会通过影响土壤中P的可利用性、促进根系磷酸酶活性提高植物对P的吸收等方式改变整个生态系统的P循环(Treseder & Vitousek, 2001 ; Marklein & Houlton, 2012 ).本研究结果进一步表明, N添加对武夷山植物群落P循环的影响明显, 植物叶片P含量变化是叶片N:P变化的主要原因. ...
Terrestrial phosphorus limitation: Mechanisms, implications and nitrogen-phosphorus interactions
1
2010
... 研究显示N添加背景下, 武夷山植物叶片N:P的变化主要受叶片P含量变化影响, 而非受N含量变化影响(图3 ).其他研究结果也显示, 在N元素相对充足而P元素相对匮乏的生态系统, N添加会明显改变该生态系统的P循环(Güsewell, 2004 ; Liu et al ., 2013 ).N、P元素从细胞到生态系统尺度, 都存在紧密的耦合作用(Elser et al ., 2000 ; Sterner & Elser, 2002 ; Vitousek et al ., 2010 ), 其中一种元素的吸收受到另一种元素可利用性的影响(Ågren et al ., 2012 ), 因此N添加会通过影响土壤中P的可利用性、促进根系磷酸酶活性提高植物对P的吸收等方式改变整个生态系统的P循环(Treseder & Vitousek, 2001 ; Marklein & Houlton, 2012 ).本研究结果进一步表明, N添加对武夷山植物群落P循环的影响明显, 植物叶片P含量变化是叶片N:P变化的主要原因. ...
Technical report: Human alteration of the global nitrogen cycle: Sources and consequences
1
1997
... 人类活动导致大气氮(N)沉降不断增强(Galloway et al ., 2004 ; Dentener et al ., 2006 ; Mahowald et al ., 2008 ), 正深刻影响着生态系统及其养分循环过程(Vitousek & Tilman, 1997 ; Galloway & Sutton, 2008 ; Treseder, 2008 ).野外人工N添加实验是模拟自然N沉降增加对生态系统影响的重要手段.自20世纪80年代起, 研究者在温带和北方森林地区陆续开展了大量长时间的野外N添加实验(Aber & Melillo, 1989 ; Wright & Rasmussen, 1998 ; Magill et al ., 2000 , 2004 ), 而N添加实验在亚热带常绿阔叶林地区开展得相对较少(Mo et al ., 2008 ; Lu et al ., 2010 ).我国的亚热带常绿阔叶林面积广阔、物种组成丰富(Fang et al ., 2014 ), 是全球亚热带常绿阔叶林的主要分布区, 在我国亚热带常绿阔叶林地区开展N添加的生态学效应的研究十分必要. ...
Introduction to the NITREX and EXMAN projects
2
1998
... 人类活动导致大气氮(N)沉降不断增强(Galloway et al ., 2004 ; Dentener et al ., 2006 ; Mahowald et al ., 2008 ), 正深刻影响着生态系统及其养分循环过程(Vitousek & Tilman, 1997 ; Galloway & Sutton, 2008 ; Treseder, 2008 ).野外人工N添加实验是模拟自然N沉降增加对生态系统影响的重要手段.自20世纪80年代起, 研究者在温带和北方森林地区陆续开展了大量长时间的野外N添加实验(Aber & Melillo, 1989 ; Wright & Rasmussen, 1998 ; Magill et al ., 2000 , 2004 ), 而N添加实验在亚热带常绿阔叶林地区开展得相对较少(Mo et al ., 2008 ; Lu et al ., 2010 ).我国的亚热带常绿阔叶林面积广阔、物种组成丰富(Fang et al ., 2014 ), 是全球亚热带常绿阔叶林的主要分布区, 在我国亚热带常绿阔叶林地区开展N添加的生态学效应的研究十分必要. ...
... 在全球变化的背景下, 植物的化学计量特征可以表征植物维持体内化学计量内稳性的能力(Sterner & Elser, 2002 ; Demars & Edwards, 2007 ), 并反映植物对环境变化的适应性(Elser et al ., 2010a ).以往的研究表明, 植物叶片N、磷(P)化学计量特征因植被类型、功能群、物种的差异而不同(Sterner & Elser, 2002 ; Reich, 2003 ; Ågren, 2004 ), 不同功能群和物种对N添加的响应也存在差异(Güsewell, 2004 ).自20世纪80年代起, 研究人员在温带和北方森林地区陆续开展了一些经典的野外N添加实验(Aber & Melillo, 1989 ; Wright & Rasmussen, 1998 ; Magill et al ., 2000 ), 植物化学计量学作为研究N循环的新手段, 也被应用于N添加生态学效应的研究中.例如: Magill等(2004)在美国哈佛森林开展了长期的N添加试验, 经过15年的研究发现: N添加显著增加了乔木优势种的叶片N含量, 并且叶片化学计量特征对N添加的响应存在明显的年际变化; Huang等(2012)在模拟的亚热带常绿阔叶林生态系统中开展了N添加实验, 对群落内5种乔木树种叶片N、P化学计量特征进行了研究, 结果显示N添加显著影响了多数物种的叶片N、P含量和N:P; Cui等(2010)对温带草原8种优势植物叶片化学计量特征对N添加的响应进行了研究, 发现不同物种的叶片化学计量特征对N添加的响应也存在差异. ...
 , 张歆阳, 曾发旭, 邢娟
, 张歆阳, 曾发旭, 邢娟


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}