Foliar nitrogen and phosphorus stoichiometry of alien invasive plants and co-occurring natives in Xishuangbanna
HU Chao-Chen1, 2, LIU Xue-Yan1, 3, *, , LEI Yan-Bao4, TAN Yun-Hong5, ZHANG Peng1, DONG Yu-Ping3, LIU Cong-Qiang1, 3
1State Key Laboratory of Environmental Geochemistry, Institute of Geochemistry, Chinese Academy of Sciences, Guiyang 550081, China2University of Chinese Academy of Sciences, Beijing 100049, China3Institute of Surface-Earth System Science, Tianjin University, Tianjin 300072, China4Institute of Mountain Hazards and Environment, Chinese Academy of Sciences, Chengdu 610041, Chinaand 5Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Xishuangbanna, Yunnan 666303, China
Aims How alien invasive plants and co-occurring native plants utilize nutrients is one of major issues in invasion ecology. Foliar nitrogen (N) and phosphorus (P) contents and stoichiometry can elucidate the uptake ability and limitation status of nutrients in plants, which provides basic knowledge for understanding the invading ability and co-occurrence or disappearance of plants. Methods Based on typical alien invasive plants (Chromolaena odorata, Ageratina adenophora) and native plants in southwestern China, this study focused on strategies of N and P utilization among invasive plants and native plants under different invasion conditions. The species compositions, aboveground biomass, leaf N and P contents and leaf N:P were investigated for plants in plots with no invasion and with different invasion extents (estimated by the plot-based percentage of invaders’ biomass in total community) at Mt. Kongming in Xishuangbanna region, Yunnan Province, China. Important findings The species number decreased significantly with the invasion extent of both C. odorata and A. adenophora, although the aboveground biomass was greatly enhanced. Leaf N and P contents did not differ between the two studied invaders, but they showed significantly higher N and P levels than both co-occurring and only native species (p < 0.05). Besides, leaf N and P contents of invaders increased with the invasion extent, and leaf N of native plants also showed an increasing trend with the invasion extent. When the influence of invasion was checked for the same species, leaf P contents decreased, whereas leaf N and N:P increased for most native plants under invasion. Based on the absolute foliar N and P contents, N:P values, we inferred that native plants were still limited by N, although N availability might be enhanced by invasion. Both invasive plants had leaf N:P values lower than 10, suggesting a higher P uptake relative to N uptake. All above results highlighted a higher N and P uptake of typical alien invasive plants in southwestern China.
HUChao-Chen, LIUXue-Yan, LEIYan-Bao, TANYun-Hong, ZHANGPeng, DONGYu-Ping, LIUCong-Qiang. Foliar nitrogen and phosphorus stoichiometry of alien invasive plants and co-occurring natives in Xishuangbanna. Chinese Journal of Plant Ecology, 2016, 40(11): 1145-1153 https://doi.org/10.17521/cjpe.2016.0052
Table 1 Native plant species across the study plots with no invasion (n = 3), invaded by Chromolaena odorata (n = 6) and Ageratina adenophora (n = 6), respectively, at Mt. Kongming in Xishuangbanna, Yunnan Province, China
科 Family
无入侵样方本地植物(出现的样方数) Species in plots with no invasion (n)
飞机草入侵样方本地 植物(出现的样方数) Native species in plots invaded by C. odorata (n)
紫茎泽兰入侵样方本地植物 (出现的样方数) Native species in plots invaded by A. adenophora (n)
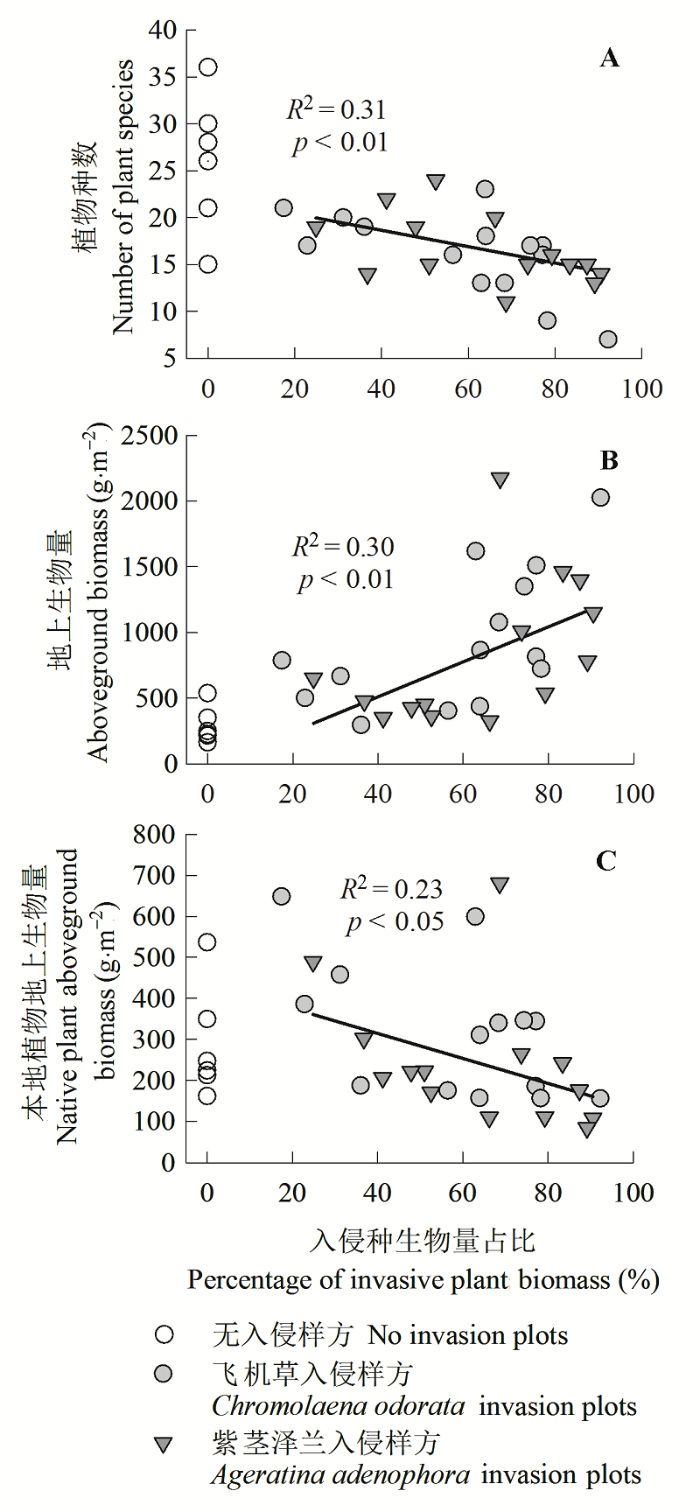
Fig. 1 Variations of plot-based species number (A), community aboveground biomass and native plants aboveground biomass (B, C) with the biomass percentage of alien invasive plants in total community.
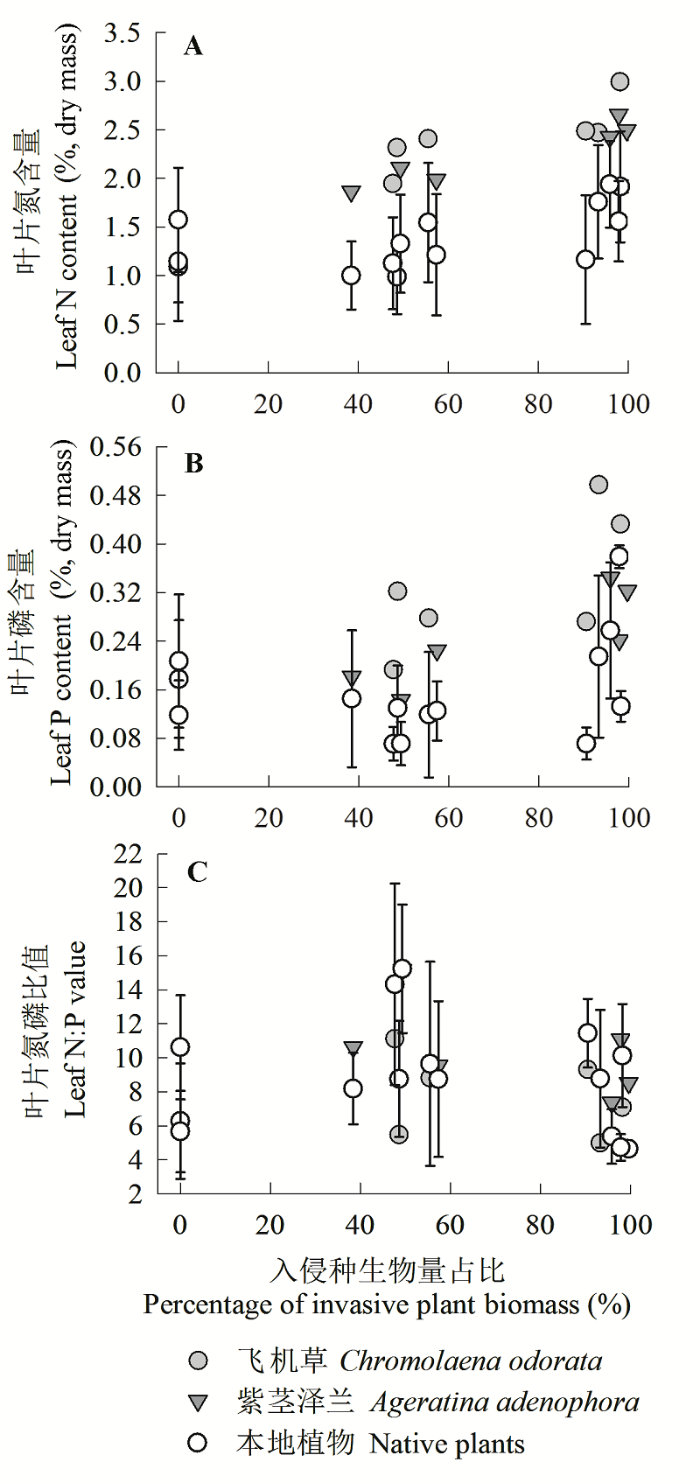
Table 2 Leaf N and P concentration, N:P values (mean ± SD) of native and invasive plants under invasion and no invasion (values of each parameter with different letter a or b differ significantly from each other)
Fig. 2 Variations of plot-based leaf N (A), P (B), N:P (C) in invasive and native plants with the biomass percentage of invasive plants in total community (mean ± SD).
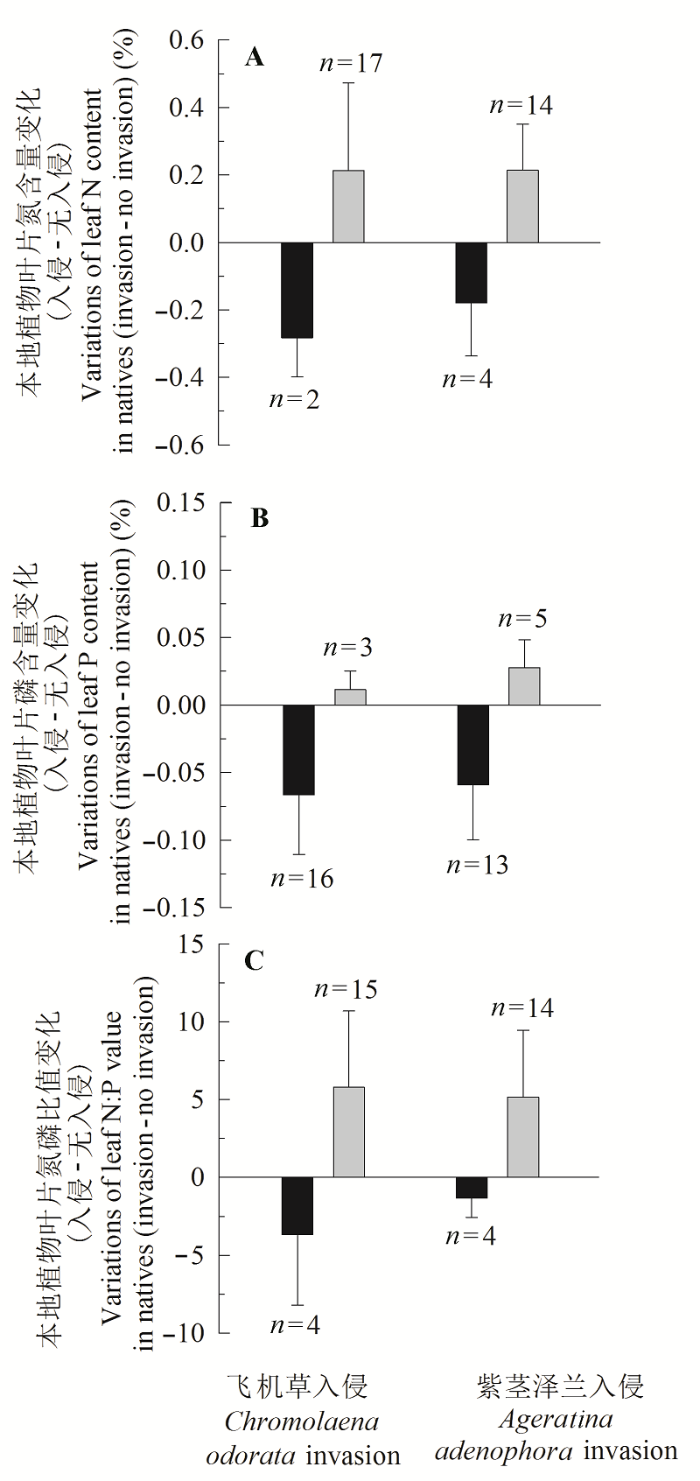
Fig. 3 Differences of leaf N and P concentration, N:P values in the same native plants between non-invasion and invasion plots, respectively (Error bars represent SD). The gray and black columns denote increases and decreases in N, P contents or N:P in co-occuring plants, respectively, n denotes species number with increases or decreases in N, P, N:P, respectively.
Photosynthesis, nitrogen allocation and specific leaf area in invasive Eupatorium adenophorum and native Eupatorium japonicum grown at different irradiances
Comparative studies on seasonal dynamics of soil enzymatic activities and soil nutrient availability in mono- and mixed-culture plant communities of Ageratina adenophora and Seiaria sphaclata
. Journal of Plant Ecology (Chinese Version), 32, 900-907. (in Chinese with English abstract)[蒋智林, 刘万学, 万方浩, 李正跃 (2008).
Innate and evolutionarily increased advantages of invasive Eupatorium adenophorum over native E. japonicum under ambient and doubled atmospheric CO2 concentrations
An invasive aster (Ageratina adenophora) invades and dominates forest understories in China: Altered soil microbial communities facilitate the invader and inhibit natives
Ecological stoichiometry of C, N, and P of invasive Phragmites australis and native Cyperus malaccensis species in the Minjiang River tidal estuarine wetlands of China
Long-term ecosystem impacts of an introduced grass in the northern Great Plains
1
1999
... 氮(N)、磷(P)是影响植物生长的重要营养元素, 也是生态系统生产力的主要限制因子(Elser et al., 2007; Vitousek et al., 2010; Lü et al., 2014; Li et al., 2015).植物叶片N、P含量能够反映植物、土壤的营养状况、种间竞争性吸收、储存能力, N、P化学计量关系能够反映植物受N、P限制的状况(Elser et al., 2007).目前, 叶片N、P含量和化学计量特征已被用于解释入侵生态系统植物营养元素的分配策略(王满莲和冯玉龙, 2005), 探讨外来植物与入侵地土壤N、P有效性的联系(陆建忠等, 2005; 于兴军等, 2005).另外, 也有研究涉及入侵植物叶片N、P含量及N:P值的生境异质性(屠臣阳等, 2013)和季节性变化规律(Wang et al., 2015)、入侵种化学计量特征和生物量变化对生态系统C、N、P循环的调控作用(Christian & Willson, 1999)、外来植物入侵与全球变化因子(如模拟增温、降水减少等)(Liao et al., 2008)和土壤N、P可利用性变化(包括自然和模拟的N沉降, P添加等)的相互关系(Li et al., 2006; Niu et al., 2007).这些研究对利用N、P化学计量学方法理解植物种群间的生态关系提供了重要的见解.然而, 对自然条件下不同入侵程度外来入侵植物及其共存种的N、P利用特征研究还不够, 需要更详细的原位研究, 才能准确地理解决定入侵程度和本地植物留存的生物学机制(Feng et al., 2009). ...
Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems
3
2007
... 氮(N)、磷(P)是影响植物生长的重要营养元素, 也是生态系统生产力的主要限制因子(Elser et al., 2007; Vitousek et al., 2010; Lü et al., 2014; Li et al., 2015).植物叶片N、P含量能够反映植物、土壤的营养状况、种间竞争性吸收、储存能力, N、P化学计量关系能够反映植物受N、P限制的状况(Elser et al., 2007).目前, 叶片N、P含量和化学计量特征已被用于解释入侵生态系统植物营养元素的分配策略(王满莲和冯玉龙, 2005), 探讨外来植物与入侵地土壤N、P有效性的联系(陆建忠等, 2005; 于兴军等, 2005).另外, 也有研究涉及入侵植物叶片N、P含量及N:P值的生境异质性(屠臣阳等, 2013)和季节性变化规律(Wang et al., 2015)、入侵种化学计量特征和生物量变化对生态系统C、N、P循环的调控作用(Christian & Willson, 1999)、外来植物入侵与全球变化因子(如模拟增温、降水减少等)(Liao et al., 2008)和土壤N、P可利用性变化(包括自然和模拟的N沉降, P添加等)的相互关系(Li et al., 2006; Niu et al., 2007).这些研究对利用N、P化学计量学方法理解植物种群间的生态关系提供了重要的见解.然而, 对自然条件下不同入侵程度外来入侵植物及其共存种的N、P利用特征研究还不够, 需要更详细的原位研究, 才能准确地理解决定入侵程度和本地植物留存的生物学机制(Feng et al., 2009). ...
... ).植物叶片N、P含量能够反映植物、土壤的营养状况、种间竞争性吸收、储存能力, N、P化学计量关系能够反映植物受N、P限制的状况(Elser et al., 2007).目前, 叶片N、P含量和化学计量特征已被用于解释入侵生态系统植物营养元素的分配策略(王满莲和冯玉龙, 2005), 探讨外来植物与入侵地土壤N、P有效性的联系(陆建忠等, 2005; 于兴军等, 2005).另外, 也有研究涉及入侵植物叶片N、P含量及N:P值的生境异质性(屠臣阳等, 2013)和季节性变化规律(Wang et al., 2015)、入侵种化学计量特征和生物量变化对生态系统C、N、P循环的调控作用(Christian & Willson, 1999)、外来植物入侵与全球变化因子(如模拟增温、降水减少等)(Liao et al., 2008)和土壤N、P可利用性变化(包括自然和模拟的N沉降, P添加等)的相互关系(Li et al., 2006; Niu et al., 2007).这些研究对利用N、P化学计量学方法理解植物种群间的生态关系提供了重要的见解.然而, 对自然条件下不同入侵程度外来入侵植物及其共存种的N、P利用特征研究还不够, 需要更详细的原位研究, 才能准确地理解决定入侵程度和本地植物留存的生物学机制(Feng et al., 2009). ...
Photosynthesis, nitrogen allocation and specific leaf area in invasive Eupatorium adenophorum and native Eupatorium japonicum grown at different irradiances
... 氮(N)、磷(P)是影响植物生长的重要营养元素, 也是生态系统生产力的主要限制因子(Elser et al., 2007; Vitousek et al., 2010; Lü et al., 2014; Li et al., 2015).植物叶片N、P含量能够反映植物、土壤的营养状况、种间竞争性吸收、储存能力, N、P化学计量关系能够反映植物受N、P限制的状况(Elser et al., 2007).目前, 叶片N、P含量和化学计量特征已被用于解释入侵生态系统植物营养元素的分配策略(王满莲和冯玉龙, 2005), 探讨外来植物与入侵地土壤N、P有效性的联系(陆建忠等, 2005; 于兴军等, 2005).另外, 也有研究涉及入侵植物叶片N、P含量及N:P值的生境异质性(屠臣阳等, 2013)和季节性变化规律(Wang et al., 2015)、入侵种化学计量特征和生物量变化对生态系统C、N、P循环的调控作用(Christian & Willson, 1999)、外来植物入侵与全球变化因子(如模拟增温、降水减少等)(Liao et al., 2008)和土壤N、P可利用性变化(包括自然和模拟的N沉降, P添加等)的相互关系(Li et al., 2006; Niu et al., 2007).这些研究对利用N、P化学计量学方法理解植物种群间的生态关系提供了重要的见解.然而, 对自然条件下不同入侵程度外来入侵植物及其共存种的N、P利用特征研究还不够, 需要更详细的原位研究, 才能准确地理解决定入侵程度和本地植物留存的生物学机制(Feng et al., 2009). ...
... 飞机草(Chromolaena odorata)和紫茎泽兰(又叫破坏草, Ageratina adenophora)原产于中美洲, 是菊科多年生草本植物或亚灌木, 自20世纪40年代从我国云南西双版纳入侵以来, 已经成为我国危害最严重的外来入侵植物(万方浩等, 2002).它们具有明显的先锋植物特性, 土壤种子库持久, 极易形成单优群落.已发现紫茎泽兰具有对资源养分的强吸收能力, 而且还能活化土壤养分(牛红榜等, 2007; 蒋智林等, 2008).而将N更多地分配到叶片光合机构, 并和防御系统间进行权衡是紫茎泽兰的一个重要入侵策略(Feng et al., 2009; Lei et al., 2011).此外, 化感作用也被认为是紫茎泽兰在与土著植物种群竞争中获得优势的策略之一(Niu et al., 2007; Zheng et al., 2014).然而, 除了对紫茎泽兰的同质园实验, 对自然条件下紫茎泽兰、飞机草相对于本地植物的N、P吸收能力, 以及在不同入侵程度下植物的N、P利用方式需要更系统和详细的调查.据此, 本研究以受飞机草、紫茎泽兰不同程度入侵的生态系统为研究对象, 通过对物种组成和生物量变化、主要植物叶片N、P含量和化学计量特征的详细分析, 探讨外来植物与本地种N、P吸收特征及其随入侵程度的变化规律.这对于准确地把握我国西南地区典型外来菊科植物的入侵机制、理解土著物种群落的生物多样性和生产力变化具有科学价值. ...
N:P ratios in terrestrial plants: Variation and functional significance
Innate and evolutionarily increased advantages of invasive Eupatorium adenophorum over native E. japonicum under ambient and doubled atmospheric CO2 concentrations
1
2011
... 飞机草(Chromolaena odorata)和紫茎泽兰(又叫破坏草, Ageratina adenophora)原产于中美洲, 是菊科多年生草本植物或亚灌木, 自20世纪40年代从我国云南西双版纳入侵以来, 已经成为我国危害最严重的外来入侵植物(万方浩等, 2002).它们具有明显的先锋植物特性, 土壤种子库持久, 极易形成单优群落.已发现紫茎泽兰具有对资源养分的强吸收能力, 而且还能活化土壤养分(牛红榜等, 2007; 蒋智林等, 2008).而将N更多地分配到叶片光合机构, 并和防御系统间进行权衡是紫茎泽兰的一个重要入侵策略(Feng et al., 2009; Lei et al., 2011).此外, 化感作用也被认为是紫茎泽兰在与土著植物种群竞争中获得优势的策略之一(Niu et al., 2007; Zheng et al., 2014).然而, 除了对紫茎泽兰的同质园实验, 对自然条件下紫茎泽兰、飞机草相对于本地植物的N、P吸收能力, 以及在不同入侵程度下植物的N、P利用方式需要更系统和详细的调查.据此, 本研究以受飞机草、紫茎泽兰不同程度入侵的生态系统为研究对象, 通过对物种组成和生物量变化、主要植物叶片N、P含量和化学计量特征的详细分析, 探讨外来植物与本地种N、P吸收特征及其随入侵程度的变化规律.这对于准确地把握我国西南地区典型外来菊科植物的入侵机制、理解土著物种群落的生物多样性和生产力变化具有科学价值. ...
Changes in soil microbial community associated with invasion of the exotic weed,Mikania micrantha H.B.K
1
2006
... 氮(N)、磷(P)是影响植物生长的重要营养元素, 也是生态系统生产力的主要限制因子(Elser et al., 2007; Vitousek et al., 2010; Lü et al., 2014; Li et al., 2015).植物叶片N、P含量能够反映植物、土壤的营养状况、种间竞争性吸收、储存能力, N、P化学计量关系能够反映植物受N、P限制的状况(Elser et al., 2007).目前, 叶片N、P含量和化学计量特征已被用于解释入侵生态系统植物营养元素的分配策略(王满莲和冯玉龙, 2005), 探讨外来植物与入侵地土壤N、P有效性的联系(陆建忠等, 2005; 于兴军等, 2005).另外, 也有研究涉及入侵植物叶片N、P含量及N:P值的生境异质性(屠臣阳等, 2013)和季节性变化规律(Wang et al., 2015)、入侵种化学计量特征和生物量变化对生态系统C、N、P循环的调控作用(Christian & Willson, 1999)、外来植物入侵与全球变化因子(如模拟增温、降水减少等)(Liao et al., 2008)和土壤N、P可利用性变化(包括自然和模拟的N沉降, P添加等)的相互关系(Li et al., 2006; Niu et al., 2007).这些研究对利用N、P化学计量学方法理解植物种群间的生态关系提供了重要的见解.然而, 对自然条件下不同入侵程度外来入侵植物及其共存种的N、P利用特征研究还不够, 需要更详细的原位研究, 才能准确地理解决定入侵程度和本地植物留存的生物学机制(Feng et al., 2009). ...
Aggravated phosphorus limitation on biomass production under increasing N addition: A meta-analysis
1
2015
... 氮(N)、磷(P)是影响植物生长的重要营养元素, 也是生态系统生产力的主要限制因子(Elser et al., 2007; Vitousek et al., 2010; Lü et al., 2014; Li et al., 2015).植物叶片N、P含量能够反映植物、土壤的营养状况、种间竞争性吸收、储存能力, N、P化学计量关系能够反映植物受N、P限制的状况(Elser et al., 2007).目前, 叶片N、P含量和化学计量特征已被用于解释入侵生态系统植物营养元素的分配策略(王满莲和冯玉龙, 2005), 探讨外来植物与入侵地土壤N、P有效性的联系(陆建忠等, 2005; 于兴军等, 2005).另外, 也有研究涉及入侵植物叶片N、P含量及N:P值的生境异质性(屠臣阳等, 2013)和季节性变化规律(Wang et al., 2015)、入侵种化学计量特征和生物量变化对生态系统C、N、P循环的调控作用(Christian & Willson, 1999)、外来植物入侵与全球变化因子(如模拟增温、降水减少等)(Liao et al., 2008)和土壤N、P可利用性变化(包括自然和模拟的N沉降, P添加等)的相互关系(Li et al., 2006; Niu et al., 2007).这些研究对利用N、P化学计量学方法理解植物种群间的生态关系提供了重要的见解.然而, 对自然条件下不同入侵程度外来入侵植物及其共存种的N、P利用特征研究还不够, 需要更详细的原位研究, 才能准确地理解决定入侵程度和本地植物留存的生物学机制(Feng et al., 2009). ...
Altered ecosystem carbon and nitrogen cycles by plant invasion: A meta-analysis
2
2008
... 氮(N)、磷(P)是影响植物生长的重要营养元素, 也是生态系统生产力的主要限制因子(Elser et al., 2007; Vitousek et al., 2010; Lü et al., 2014; Li et al., 2015).植物叶片N、P含量能够反映植物、土壤的营养状况、种间竞争性吸收、储存能力, N、P化学计量关系能够反映植物受N、P限制的状况(Elser et al., 2007).目前, 叶片N、P含量和化学计量特征已被用于解释入侵生态系统植物营养元素的分配策略(王满莲和冯玉龙, 2005), 探讨外来植物与入侵地土壤N、P有效性的联系(陆建忠等, 2005; 于兴军等, 2005).另外, 也有研究涉及入侵植物叶片N、P含量及N:P值的生境异质性(屠臣阳等, 2013)和季节性变化规律(Wang et al., 2015)、入侵种化学计量特征和生物量变化对生态系统C、N、P循环的调控作用(Christian & Willson, 1999)、外来植物入侵与全球变化因子(如模拟增温、降水减少等)(Liao et al., 2008)和土壤N、P可利用性变化(包括自然和模拟的N沉降, P添加等)的相互关系(Li et al., 2006; Niu et al., 2007).这些研究对利用N、P化学计量学方法理解植物种群间的生态关系提供了重要的见解.然而, 对自然条件下不同入侵程度外来入侵植物及其共存种的N、P利用特征研究还不够, 需要更详细的原位研究, 才能准确地理解决定入侵程度和本地植物留存的生物学机制(Feng et al., 2009). ...
... 植物N、P吸收增加通常反映了土壤N、P可利用性增加, 这主要归因于外来植物入侵促进了土壤N、P微生物活性.于兴军等(2005)发现紫茎泽兰入侵改变了土壤微生物群落结构、提高了土壤无机N (NH4+-N和NO3--N)、P的可利用性.牛红榜等(2007)也认为紫茎泽兰入侵能够改变一些土壤菌根的数量, 提高养分的可利用性, 这是紫茎泽兰入侵地土壤无机N浓度高于无入侵地的主要原因.Liao等(2008)通过整合分析, 总结了植物入侵会降低植物根冠比、增加地上/地下净初级生产力、凋落物N库、加速凋落物分解, 进而提高土壤N净矿化和硝化速率的机理.我国学者也在互花米草与土著植物(芦苇)的N竞争研究中揭示了入侵植物这种较强的“表型可塑性” (Zhao et al., 2010).这些反映了入侵植物N、P吸收和代谢能力与入侵程度或与其对土壤N、P可利用性之间存在正反馈关系(Niu et al., 2007; Liao et al., 2008).我们的研究通过紫茎泽兰、飞机草以及更详细的共存植物种对比(图2, 图3), 证实了入侵植物的N、P吸收优势, 这种优势还会随入侵程度增加而变大.尽管本研究结果也支持上述反馈机制, 但仍需对研究区土壤N、P营养状态和变化进行详细的调查来验证. ...
... 氮(N)、磷(P)是影响植物生长的重要营养元素, 也是生态系统生产力的主要限制因子(Elser et al., 2007; Vitousek et al., 2010; Lü et al., 2014; Li et al., 2015).植物叶片N、P含量能够反映植物、土壤的营养状况、种间竞争性吸收、储存能力, N、P化学计量关系能够反映植物受N、P限制的状况(Elser et al., 2007).目前, 叶片N、P含量和化学计量特征已被用于解释入侵生态系统植物营养元素的分配策略(王满莲和冯玉龙, 2005), 探讨外来植物与入侵地土壤N、P有效性的联系(陆建忠等, 2005; 于兴军等, 2005).另外, 也有研究涉及入侵植物叶片N、P含量及N:P值的生境异质性(屠臣阳等, 2013)和季节性变化规律(Wang et al., 2015)、入侵种化学计量特征和生物量变化对生态系统C、N、P循环的调控作用(Christian & Willson, 1999)、外来植物入侵与全球变化因子(如模拟增温、降水减少等)(Liao et al., 2008)和土壤N、P可利用性变化(包括自然和模拟的N沉降, P添加等)的相互关系(Li et al., 2006; Niu et al., 2007).这些研究对利用N、P化学计量学方法理解植物种群间的生态关系提供了重要的见解.然而, 对自然条件下不同入侵程度外来入侵植物及其共存种的N、P利用特征研究还不够, 需要更详细的原位研究, 才能准确地理解决定入侵程度和本地植物留存的生物学机制(Feng et al., 2009). ...
Contrasting responses in leaf nutrient-use strategies of two dominant grass species along a 30-yr temperate steppe grazing exclusion chronosequence
1
2014
... 氮(N)、磷(P)是影响植物生长的重要营养元素, 也是生态系统生产力的主要限制因子(Elser et al., 2007; Vitousek et al., 2010; Lü et al., 2014; Li et al., 2015).植物叶片N、P含量能够反映植物、土壤的营养状况、种间竞争性吸收、储存能力, N、P化学计量关系能够反映植物受N、P限制的状况(Elser et al., 2007).目前, 叶片N、P含量和化学计量特征已被用于解释入侵生态系统植物营养元素的分配策略(王满莲和冯玉龙, 2005), 探讨外来植物与入侵地土壤N、P有效性的联系(陆建忠等, 2005; 于兴军等, 2005).另外, 也有研究涉及入侵植物叶片N、P含量及N:P值的生境异质性(屠臣阳等, 2013)和季节性变化规律(Wang et al., 2015)、入侵种化学计量特征和生物量变化对生态系统C、N、P循环的调控作用(Christian & Willson, 1999)、外来植物入侵与全球变化因子(如模拟增温、降水减少等)(Liao et al., 2008)和土壤N、P可利用性变化(包括自然和模拟的N沉降, P添加等)的相互关系(Li et al., 2006; Niu et al., 2007).这些研究对利用N、P化学计量学方法理解植物种群间的生态关系提供了重要的见解.然而, 对自然条件下不同入侵程度外来入侵植物及其共存种的N、P利用特征研究还不够, 需要更详细的原位研究, 才能准确地理解决定入侵程度和本地植物留存的生物学机制(Feng et al., 2009). ...
An invasive aster (Ageratina adenophora) invades and dominates forest understories in China: Altered soil microbial communities facilitate the invader and inhibit natives
4
2007
... 氮(N)、磷(P)是影响植物生长的重要营养元素, 也是生态系统生产力的主要限制因子(Elser et al., 2007; Vitousek et al., 2010; Lü et al., 2014; Li et al., 2015).植物叶片N、P含量能够反映植物、土壤的营养状况、种间竞争性吸收、储存能力, N、P化学计量关系能够反映植物受N、P限制的状况(Elser et al., 2007).目前, 叶片N、P含量和化学计量特征已被用于解释入侵生态系统植物营养元素的分配策略(王满莲和冯玉龙, 2005), 探讨外来植物与入侵地土壤N、P有效性的联系(陆建忠等, 2005; 于兴军等, 2005).另外, 也有研究涉及入侵植物叶片N、P含量及N:P值的生境异质性(屠臣阳等, 2013)和季节性变化规律(Wang et al., 2015)、入侵种化学计量特征和生物量变化对生态系统C、N、P循环的调控作用(Christian & Willson, 1999)、外来植物入侵与全球变化因子(如模拟增温、降水减少等)(Liao et al., 2008)和土壤N、P可利用性变化(包括自然和模拟的N沉降, P添加等)的相互关系(Li et al., 2006; Niu et al., 2007).这些研究对利用N、P化学计量学方法理解植物种群间的生态关系提供了重要的见解.然而, 对自然条件下不同入侵程度外来入侵植物及其共存种的N、P利用特征研究还不够, 需要更详细的原位研究, 才能准确地理解决定入侵程度和本地植物留存的生物学机制(Feng et al., 2009). ...
... 飞机草(Chromolaena odorata)和紫茎泽兰(又叫破坏草, Ageratina adenophora)原产于中美洲, 是菊科多年生草本植物或亚灌木, 自20世纪40年代从我国云南西双版纳入侵以来, 已经成为我国危害最严重的外来入侵植物(万方浩等, 2002).它们具有明显的先锋植物特性, 土壤种子库持久, 极易形成单优群落.已发现紫茎泽兰具有对资源养分的强吸收能力, 而且还能活化土壤养分(牛红榜等, 2007; 蒋智林等, 2008).而将N更多地分配到叶片光合机构, 并和防御系统间进行权衡是紫茎泽兰的一个重要入侵策略(Feng et al., 2009; Lei et al., 2011).此外, 化感作用也被认为是紫茎泽兰在与土著植物种群竞争中获得优势的策略之一(Niu et al., 2007; Zheng et al., 2014).然而, 除了对紫茎泽兰的同质园实验, 对自然条件下紫茎泽兰、飞机草相对于本地植物的N、P吸收能力, 以及在不同入侵程度下植物的N、P利用方式需要更系统和详细的调查.据此, 本研究以受飞机草、紫茎泽兰不同程度入侵的生态系统为研究对象, 通过对物种组成和生物量变化、主要植物叶片N、P含量和化学计量特征的详细分析, 探讨外来植物与本地种N、P吸收特征及其随入侵程度的变化规律.这对于准确地把握我国西南地区典型外来菊科植物的入侵机制、理解土著物种群落的生物多样性和生产力变化具有科学价值. ...
... 植物N、P吸收增加通常反映了土壤N、P可利用性增加, 这主要归因于外来植物入侵促进了土壤N、P微生物活性.于兴军等(2005)发现紫茎泽兰入侵改变了土壤微生物群落结构、提高了土壤无机N (NH4+-N和NO3--N)、P的可利用性.牛红榜等(2007)也认为紫茎泽兰入侵能够改变一些土壤菌根的数量, 提高养分的可利用性, 这是紫茎泽兰入侵地土壤无机N浓度高于无入侵地的主要原因.Liao等(2008)通过整合分析, 总结了植物入侵会降低植物根冠比、增加地上/地下净初级生产力、凋落物N库、加速凋落物分解, 进而提高土壤N净矿化和硝化速率的机理.我国学者也在互花米草与土著植物(芦苇)的N竞争研究中揭示了入侵植物这种较强的“表型可塑性” (Zhao et al., 2010).这些反映了入侵植物N、P吸收和代谢能力与入侵程度或与其对土壤N、P可利用性之间存在正反馈关系(Niu et al., 2007; Liao et al., 2008).我们的研究通过紫茎泽兰、飞机草以及更详细的共存植物种对比(图2, 图3), 证实了入侵植物的N、P吸收优势, 这种优势还会随入侵程度增加而变大.尽管本研究结果也支持上述反馈机制, 但仍需对研究区土壤N、P营养状态和变化进行详细的调查来验证. ...
... 飞机草(Chromolaena odorata)和紫茎泽兰(又叫破坏草, Ageratina adenophora)原产于中美洲, 是菊科多年生草本植物或亚灌木, 自20世纪40年代从我国云南西双版纳入侵以来, 已经成为我国危害最严重的外来入侵植物(万方浩等, 2002).它们具有明显的先锋植物特性, 土壤种子库持久, 极易形成单优群落.已发现紫茎泽兰具有对资源养分的强吸收能力, 而且还能活化土壤养分(牛红榜等, 2007; 蒋智林等, 2008).而将N更多地分配到叶片光合机构, 并和防御系统间进行权衡是紫茎泽兰的一个重要入侵策略(Feng et al., 2009; Lei et al., 2011).此外, 化感作用也被认为是紫茎泽兰在与土著植物种群竞争中获得优势的策略之一(Niu et al., 2007; Zheng et al., 2014).然而, 除了对紫茎泽兰的同质园实验, 对自然条件下紫茎泽兰、飞机草相对于本地植物的N、P吸收能力, 以及在不同入侵程度下植物的N、P利用方式需要更系统和详细的调查.据此, 本研究以受飞机草、紫茎泽兰不同程度入侵的生态系统为研究对象, 通过对物种组成和生物量变化、主要植物叶片N、P含量和化学计量特征的详细分析, 探讨外来植物与本地种N、P吸收特征及其随入侵程度的变化规律.这对于准确地把握我国西南地区典型外来菊科植物的入侵机制、理解土著物种群落的生物多样性和生产力变化具有科学价值. ...
From the cover: Global patterns of plant leaf N and P in relation to temperature and latitude
2004
中国东部南北样带654种植物叶片氮和磷的化学计量学特征研究
2007
不同生境黄顶菊碳氮磷化学计量特征
1
2013
... 氮(N)、磷(P)是影响植物生长的重要营养元素, 也是生态系统生产力的主要限制因子(Elser et al., 2007; Vitousek et al., 2010; Lü et al., 2014; Li et al., 2015).植物叶片N、P含量能够反映植物、土壤的营养状况、种间竞争性吸收、储存能力, N、P化学计量关系能够反映植物受N、P限制的状况(Elser et al., 2007).目前, 叶片N、P含量和化学计量特征已被用于解释入侵生态系统植物营养元素的分配策略(王满莲和冯玉龙, 2005), 探讨外来植物与入侵地土壤N、P有效性的联系(陆建忠等, 2005; 于兴军等, 2005).另外, 也有研究涉及入侵植物叶片N、P含量及N:P值的生境异质性(屠臣阳等, 2013)和季节性变化规律(Wang et al., 2015)、入侵种化学计量特征和生物量变化对生态系统C、N、P循环的调控作用(Christian & Willson, 1999)、外来植物入侵与全球变化因子(如模拟增温、降水减少等)(Liao et al., 2008)和土壤N、P可利用性变化(包括自然和模拟的N沉降, P添加等)的相互关系(Li et al., 2006; Niu et al., 2007).这些研究对利用N、P化学计量学方法理解植物种群间的生态关系提供了重要的见解.然而, 对自然条件下不同入侵程度外来入侵植物及其共存种的N、P利用特征研究还不够, 需要更详细的原位研究, 才能准确地理解决定入侵程度和本地植物留存的生物学机制(Feng et al., 2009). ...
Does phosphorus limitation promote species-rich plant communities?
Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen-phosphorus interactions
1
2010
... 氮(N)、磷(P)是影响植物生长的重要营养元素, 也是生态系统生产力的主要限制因子(Elser et al., 2007; Vitousek et al., 2010; Lü et al., 2014; Li et al., 2015).植物叶片N、P含量能够反映植物、土壤的营养状况、种间竞争性吸收、储存能力, N、P化学计量关系能够反映植物受N、P限制的状况(Elser et al., 2007).目前, 叶片N、P含量和化学计量特征已被用于解释入侵生态系统植物营养元素的分配策略(王满莲和冯玉龙, 2005), 探讨外来植物与入侵地土壤N、P有效性的联系(陆建忠等, 2005; 于兴军等, 2005).另外, 也有研究涉及入侵植物叶片N、P含量及N:P值的生境异质性(屠臣阳等, 2013)和季节性变化规律(Wang et al., 2015)、入侵种化学计量特征和生物量变化对生态系统C、N、P循环的调控作用(Christian & Willson, 1999)、外来植物入侵与全球变化因子(如模拟增温、降水减少等)(Liao et al., 2008)和土壤N、P可利用性变化(包括自然和模拟的N沉降, P添加等)的相互关系(Li et al., 2006; Niu et al., 2007).这些研究对利用N、P化学计量学方法理解植物种群间的生态关系提供了重要的见解.然而, 对自然条件下不同入侵程度外来入侵植物及其共存种的N、P利用特征研究还不够, 需要更详细的原位研究, 才能准确地理解决定入侵程度和本地植物留存的生物学机制(Feng et al., 2009). ...
... 氮(N)、磷(P)是影响植物生长的重要营养元素, 也是生态系统生产力的主要限制因子(Elser et al., 2007; Vitousek et al., 2010; Lü et al., 2014; Li et al., 2015).植物叶片N、P含量能够反映植物、土壤的营养状况、种间竞争性吸收、储存能力, N、P化学计量关系能够反映植物受N、P限制的状况(Elser et al., 2007).目前, 叶片N、P含量和化学计量特征已被用于解释入侵生态系统植物营养元素的分配策略(王满莲和冯玉龙, 2005), 探讨外来植物与入侵地土壤N、P有效性的联系(陆建忠等, 2005; 于兴军等, 2005).另外, 也有研究涉及入侵植物叶片N、P含量及N:P值的生境异质性(屠臣阳等, 2013)和季节性变化规律(Wang et al., 2015)、入侵种化学计量特征和生物量变化对生态系统C、N、P循环的调控作用(Christian & Willson, 1999)、外来植物入侵与全球变化因子(如模拟增温、降水减少等)(Liao et al., 2008)和土壤N、P可利用性变化(包括自然和模拟的N沉降, P添加等)的相互关系(Li et al., 2006; Niu et al., 2007).这些研究对利用N、P化学计量学方法理解植物种群间的生态关系提供了重要的见解.然而, 对自然条件下不同入侵程度外来入侵植物及其共存种的N、P利用特征研究还不够, 需要更详细的原位研究, 才能准确地理解决定入侵程度和本地植物留存的生物学机制(Feng et al., 2009). ...
Effects of growing conditions on the growth of and interactions between salt marsh plants: Implications for invasibility of habitats
Ecological stoichiometry of C, N, and P of invasive Phragmites australis and native Cyperus malaccensis species in the Minjiang River tidal estuarine wetlands of China
1
2015
... 氮(N)、磷(P)是影响植物生长的重要营养元素, 也是生态系统生产力的主要限制因子(Elser et al., 2007; Vitousek et al., 2010; Lü et al., 2014; Li et al., 2015).植物叶片N、P含量能够反映植物、土壤的营养状况、种间竞争性吸收、储存能力, N、P化学计量关系能够反映植物受N、P限制的状况(Elser et al., 2007).目前, 叶片N、P含量和化学计量特征已被用于解释入侵生态系统植物营养元素的分配策略(王满莲和冯玉龙, 2005), 探讨外来植物与入侵地土壤N、P有效性的联系(陆建忠等, 2005; 于兴军等, 2005).另外, 也有研究涉及入侵植物叶片N、P含量及N:P值的生境异质性(屠臣阳等, 2013)和季节性变化规律(Wang et al., 2015)、入侵种化学计量特征和生物量变化对生态系统C、N、P循环的调控作用(Christian & Willson, 1999)、外来植物入侵与全球变化因子(如模拟增温、降水减少等)(Liao et al., 2008)和土壤N、P可利用性变化(包括自然和模拟的N沉降, P添加等)的相互关系(Li et al., 2006; Niu et al., 2007).这些研究对利用N、P化学计量学方法理解植物种群间的生态关系提供了重要的见解.然而, 对自然条件下不同入侵程度外来入侵植物及其共存种的N、P利用特征研究还不够, 需要更详细的原位研究, 才能准确地理解决定入侵程度和本地植物留存的生物学机制(Feng et al., 2009). ...
Nutrient concentrations in mire vegetation as a measure of nutrient limitation in mire ecosystems
... 氮(N)、磷(P)是影响植物生长的重要营养元素, 也是生态系统生产力的主要限制因子(Elser et al., 2007; Vitousek et al., 2010; Lü et al., 2014; Li et al., 2015).植物叶片N、P含量能够反映植物、土壤的营养状况、种间竞争性吸收、储存能力, N、P化学计量关系能够反映植物受N、P限制的状况(Elser et al., 2007).目前, 叶片N、P含量和化学计量特征已被用于解释入侵生态系统植物营养元素的分配策略(王满莲和冯玉龙, 2005), 探讨外来植物与入侵地土壤N、P有效性的联系(陆建忠等, 2005; 于兴军等, 2005).另外, 也有研究涉及入侵植物叶片N、P含量及N:P值的生境异质性(屠臣阳等, 2013)和季节性变化规律(Wang et al., 2015)、入侵种化学计量特征和生物量变化对生态系统C、N、P循环的调控作用(Christian & Willson, 1999)、外来植物入侵与全球变化因子(如模拟增温、降水减少等)(Liao et al., 2008)和土壤N、P可利用性变化(包括自然和模拟的N沉降, P添加等)的相互关系(Li et al., 2006; Niu et al., 2007).这些研究对利用N、P化学计量学方法理解植物种群间的生态关系提供了重要的见解.然而, 对自然条件下不同入侵程度外来入侵植物及其共存种的N、P利用特征研究还不够, 需要更详细的原位研究, 才能准确地理解决定入侵程度和本地植物留存的生物学机制(Feng et al., 2009). ...
 , 类延宝
, 类延宝

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}