植物要适应盐渍化的生境, 必须具备克服盐离子毒害(离子胁迫)、抵抗低水势(渗透胁迫)和平衡养分吸收的能力(Greeway & Munns, 1980 ; Munns & Termaat, 1986 ), 于是在生理上形成一系列抗盐特征—稀盐、泌盐、积盐和拒盐(赵可夫, 2002 ; Flowers & Colmer, 2008 ; Wang et al ., 2009 ; Zhang & Shi, 2013 ; Shabala et al ., 2014 ), 来减轻盐胁迫对组织产生的伤害, 同样所有的高等植物也都可以通过这些生理方面的调节及导入耐盐基因来增强自身对盐胁迫的耐受性(张金林等, 2015 )。
碳(C)、氮(N)、磷(P)作为植物体重要的组成元素, 对植物体的生长和生理机能的调节起着十分重要的作用(荣戗戗等, 2012 ), 而且它们之间的化学计量比强烈影响着植物在生态系统中的生长(Song et al ., 2014 )。C:N和C:P反映了植物的生长速率和对N、P养分的利用效率, 而N:P既反映了植物的生长受N或P或N、P共同的限制, 还可以作为一个关键指标决定群落的结构和功能(Aerts & Chapin, 2000 )。植物C、N、P的特征不仅体现了植物的特性, 也反映了植物对环境的长期适应和响应(侯学煜, 1982 )。
近年来研究者结合植物生理生态响应和生态化学计量特征进行了大量的研究(Hikosaka, 2004 ; Lambers & Poorter, 2004 ; Hidaka & Kitayama, 2009 ; Yu et al ., 2010 ; 郭淑青等, 2014 )。例如, 马玉(2015)发现植物光合特性与植物叶片养分有一定的关系, 在一定范围内的N浓度会促进植物光合特性与叶片养分间的关系, C含量与叶绿素呈负相关关系。这些研究对于了解盐生植物生态适应性及其对养分的利用策略具有一定的意义。因此, 本文从盐生植物体盐分对其化学计量特征影响的角度来揭示盐生植物在盐渍化生境中对养分和盐分的分配策略, 为理解盐碱地植物的生态过程和盐碱地改良利用提供科学依据。
1 材料和方法
1.1 研究区概况
研究区位于玛纳斯河流域扇缘带(44.37°- 44.48° N, 85.90-85.98° E), 属于典型的温带大陆性干旱半干旱气候区, 具有冬季严寒、夏季酷热、日照充足、干旱少雨等特点, 平均海拔377 m, 年平均气温6.9 ℃ (7月最热, 平均气温26 ℃, 1月最冷, 最低气温-18.5 ℃), 年降水量140-180 mm, 年蒸发量1500-2100 mm (其中4-9月蒸发量最大, 约占全年蒸发量的88%)。该区域土壤盐渍化程度偏重(7月份土壤基本理化性质: 电导率13.36 ms·cm-1 ; 含水率12.13%; 有机质5.35 g·kg-1 ; 全N 0.46 g·kg-1 ; 全P 0.53 g·kg-1 ; 碱解氮0.044 g·kg-1 ; 速效磷0.012 g·kg-1 ; 速效钾0.320 g·kg-1 ), 地表具有2-5 cm的盐结皮, 积盐厚度10-20 cm, 土壤类型为氯化物-硫酸盐潮盐土(王遵亲等, 1993 )。该区植物群落结构通常表现为由不连续的灌木层和相对连续的草本层组成的典型灌-草型二元结构群落(涂锦娜等, 2011 ), 主要包括盐节木(Halocnemum strobilaceum )、小叶碱蓬(Suaeda microphylla )、盐爪爪(Kalidium foliatum )、盐角草(Salicornia europaea )、多枝柽柳(Tamarix ramosissima )、花花柴(Karelinia caspia )等。
1.2 研究方法
1.2.1 样品采集
通过2014年对样地的观测, 于2015年7月下旬在样区选取了处于同一盐渍化生境下的4种藜科单优群落(半灌木盐节木和小叶碱蓬、小灌木盐爪爪、一年生草本盐角草), 每种植物分别选取生长健壮的植株5株(5个1 m × 1 m盐角草样方), 相邻株距3 m, 5株基本呈“Z”字形分布。采用全株挖取法取样, 分离根、茎(上中下)、叶, 用清水冲洗干净后, 用滤纸吸干, 在105 ℃烘箱中杀青15 min, 然后将温度调至65 ℃烘至恒质量, 用植物组织粉碎机粉碎过100目筛, 装入自封袋并标记, 待测。
1.2.2 测定方法
植株全C采用重铬酸钾容量法-外加热法测定; 全P采用(高氯酸-浓硫酸)钼锑抗比色法测定; 全N采用凯氏半微量滴定法测定; 电导率(EC)采用中国上海三信仪表厂MP521型电导仪测定(称2 g植物干样灰化后冷却至恒质量, 用热的无CO2 蒸馏水溶解, 转移至100 mL容量瓶中定容, 测定EC); 盐分离子的测定参考《土壤农化分析》(鲍士旦, 2011 )。
1.2.3 数据处理
采用Excel和SPSS 17统计分析软件处理数据, 使用Origin 8作图, 应用单因素方差分析中的LSD 法对盐生植物各盐分离子和C、N、P、C:N、C:P、N:P进行显著性检验, 并用多重比较法检验其相关性。
2 结果
2.1 4种盐生植物盐分离子含量和EC的变化
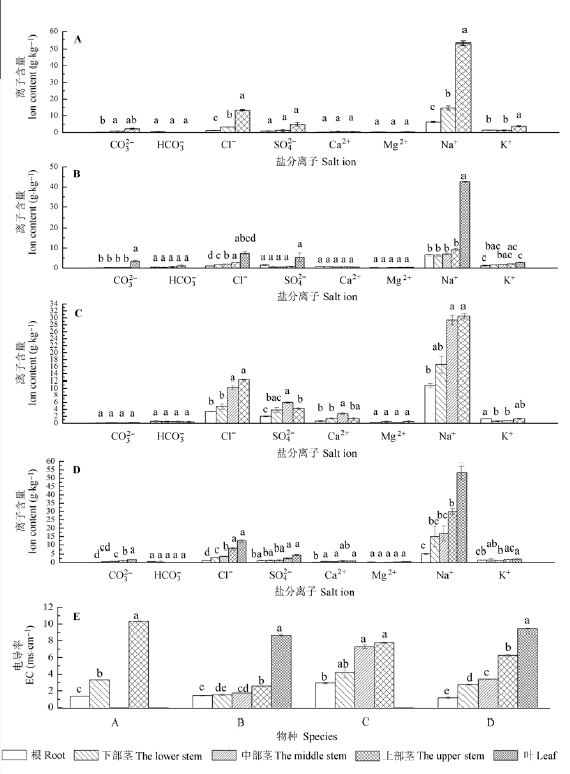
4种盐生植物不同部位和同一部位盐分离子含量、EC呈现不同的变化趋势。4种植物体中Na+ 含量最大, 其次是Cl- , 并且和EC随盐离子从植物体根部向顶端运输的过程呈显著增加的趋势(图1 )。方差分析表明, 器官和物种以及器官和物种交互作用除对Mg2+ 无影响之外, 对CO3 2- 、HCO3 2- 、Cl- 、SO4 2- 、Ca2+ 、Na+ 、K+ 、EC及C、N、P、C:N、C:P、N:P均具有显著影响(表1 )。
图1 4种盐生植物不同部位盐离子浓度和电导率的变化(平均值±标准偏差, n = 51)。A, 盐节木。B, 小叶碱蓬。C, 盐角草。D, 盐爪爪。同行不同字母表示同种植物不同部位差异显著(p < 0.05)。
Fig. 1 Changes in salt ion concentrations and electrical conductivity (EC) in different organs of the four halophytes (mean ± SD, n = 51). A, Halocnemum strobilaceum . B, Suaeda microphylla . C, Salicornia europaea . D, Kalidium foliatum . Different letters in the same row represent significant differences in different organ of the same plant (p < 0.05).
2.2 4种盐生植物C、N、P及其化学计量比
4种盐生植物同一部位和不同部位间C、N、P含量及C:N、C:P、N:P呈现不同的变化趋势(表2 )。在盐离子从植物根部向顶端运输的过程中, C含量除了在盐节木中呈下降趋势, 在其他3种盐生植物中均呈降-升-降趋势; N含量在盐节木和小叶碱蓬中呈降-升趋势, 在盐爪爪和盐角草中呈上升趋势; P含量在4种盐生植物中均呈降-升趋势; C:N除在小叶碱蓬中呈升-降趋势外, 在其他3种盐生植物中均呈下降趋势; C:P在盐节木和小叶碱蓬中呈下降趋势, 在盐爪爪和盐角草中呈升-降趋势; N:P在盐节木和盐角草中呈上升趋势, 在小叶碱蓬中呈降-升趋势, 在盐爪爪中呈升-降趋势。总体来讲, 盐节木、盐爪爪和盐角草地下C、P含量明显高于地上, 地下N含量明显低于地上; 盐爪爪、小叶碱蓬和盐爪爪地上(下) N含量和N:P均高于盐角草地上(下), 而其地上(下) C:N均小于盐角草地上(下); 盐爪爪在3种灌木中地下C、N、N:P最小, C:N最高; 小叶碱蓬在3种灌木顶端N、P含量最高, C:N、C:P、N:P最低。
2.3 盐生植物盐分离子与C、N、P化学计量特征间的相关性
除Na+ 与P之间和EC与P之间相关性不显著外, Na+ 、K+ 、CO3 2- 和EC与C、N、P、C:N、C:P、N:P之间均显著相关; 除Mg2+ 、SO4 2- 与N:P之间和SO4 2- 与P之间相关性不显著外, Mg2+ 、SO4 2- 与C、N、P、C:N、C:P之间均显著相关; Ca2+ 除了与P之间负相关外, 与C、N、C:N、C:P、N:P之间相关性均不显著; Cl- 与C、C:N之间显著负相关, 与N、N:P之间显著正相关; HCO3 - 与N、C:N、N:P显著负相关, 与C:N显著正相关。全C与全N、全P之间和全N与全P之间显著负相关; C:N与C:P之间显著正相关, 与N:P之间显著负相关, 而C:P与N:P之间相关性不显著(表3 )。
3 讨论
3.1 盐生植物吸盐特征
有研究显示, 盐生植物能够在其植丛下形成“盐岛”效应(郗金标等, 2004 ; 尹传华等, 2007 ; 张立华和陈小兵, 2015 ), 而该研究显示盐生植物还能够在其植丛上形成同样的规律。该研究表明4种盐生植物在植物体内全盐量随盐离子从根部向顶端运输过程中呈显著增加趋势, 推测在植物生长旺盛期蒸腾作用的增强大大促进了体内盐分离子(尤其是Na+ 、Cl- )的运输, 造成盐生植物体内出现“盐岛”效应。而Na+ 、Cl- 是重要的无机渗透调节物质, 4种盐生植物通过吸收和积累无机离子, 将Na+ 、Cl- 区域化于液泡中, 达到渗透调节和降低离子的毒害作用。同时4种盐生植物还通过叶、茎的肉质化或叶退化成鳞片状, 促使在盐渍环境下贮存水分, 保证光合作用的进行, 完成了正常的生理功能。然而彭飞等(2015)对盐爪爪体内的总盐分积累的研究与此结果并不一致, 可能是因为同一物种由于采摘时期不一致导致盐分运输规律不同, 或者因生活环境的不同发生了变异。该研究还表明器官和物种及二者的交互作用影响植物对盐离子(除Mg2+ 外)的吸收和积累, 但4种盐生植物对盐离子的吸收具有一致的选择作用(吸收强度从高到低: Na+ >> Cl- > K+ 、SO4 2- > Ca2+ 、Mg2+ 、CO3 2- 、HCO3 - ), 这种对Na+ 、Cl- 较强的集聚特征使这些盐生植物更适用于氯化物盐渍土的改良, 同时植物本身还可以作为优良的饲料供牲畜 食用。
3.2 盐生植物C、N、P生态化学计量特征
特定组织中的养分含量是衡量植物营养状况的一个重要指标, 它能反映植株的生长状况(Donohue & Brown, 1984 ), 并且它在植物各器官间的分配受植物自身的结构特点和生长节律的影响及生境地的养分有效性制约, 是物种系统发育和环境共同作用的结果(Baldwin et al ., 2006 ), 同时植物器官养分元素的含量也可以反映生境土壤的营养水平。研究表明4种盐生植物(灌木盐节木、盐爪爪、小叶碱蓬和草本盐角草)不同组织部位的C、N、P含量存有明显的差异, 表明了同一生活型(牛得草等, 2013 )和不同生活型(张珂等, 2014 )盐生植物存在明显的种间特异性, 体现了物种进化过程中植物种属对养分利用策略的分异特征。草本盐生植物盐角草顶端N和P含量均低于其他3种灌木盐生植物顶端N和P含量, 与Whight等(2004)和张珂等(2014)的研究结果正好相反, 反映该盐生灌木对N、P的吸收利用率高于盐生草本, 可能是因为灌木光合面积大, 光合作用强, 使顶端蒸腾作用加强, 全盐量积累大, 进而吸收更多的N和P来维持正常的生理功能。同时4种盐生植物C、N、P的变异系数分别为19%、54%、35%, 反映出C在这4种植物组织内最稳定, P含量次之, N含量最不稳定。一般而言, C在植物体内主要起骨架作用, 基本不参与植物生产活动, 所以变异最小, 最稳定; N来源广泛, 变异系数次之; P元素来源局限又易损失, 变异系数最大, 最不稳定(Krner, 1989 ; Reich & Oleksyn, 2004 ; 洪江涛等, 2014 ), 而该4种盐生植物却表现出P含量比N含量更稳定, 推测是植物自身属性和环境相互作用的结果。C含量与N、P含量之间均显著负相关, N与P之间显著正相关, 体现了盐生植物与其他高等陆地植物C、N、P化学计量特征(Sterner & Elser, 2002 ; 阎凯等, 2011 ; 李征等, 2012 )一样具备同样的规律, 反映了它们不同组织属性间的经济策略相同(有机体元素组成平衡), 是盐生植物种群能够在盐渍化生境中稳定生长发育的重要保障。
植物C:N反映了C积累的能力, 不同组织C:N差异很大(Donohue & Brown, 1984 ), 研究表明4种盐生植物茎顶端C:N远小于其余茎和根, 与王冬梅和杨惠敏(2011)所研究的牧草叶片C:N均低于茎、根的结果一致, 可能是由于茎和根的纤维化程度较高导致C含量高, 也反映了在植物生长旺盛期, 顶端全N含量最高。植物叶片N:P能够判断环境为植物生长所供应的养分状况(吴统贵等, 2010 ), Koerselaman和Meuleman (1996)研究表明, 植物生长受N和P限制的N:P阈值分别为14和16。即N:P < 14 时, 群落水平上的植物生长主要受N限制; 当N:P > 16时, 植物生长主要受P限制; 当N:P在14-16时, 则受N和P的共同限制或不受限。该研究表明小叶碱蓬、盐爪爪、盐节木顶端N:P均大于16, 受P限制; 盐角草顶端N:P接近16, 大于全国范围内陆地草本植物叶片平均N:P (13.5)(Han et al ., 2005 ), 受N、P共同限制(偏P), 进一步表明该盐碱地植物生长与荒漠(牛得草等, 2013 )和草原(杨阔等, 2010 ; 张文彦等, 2010 )一样主要受P的限制。
3.3 盐生植物体内盐分离子对植物化学计量特征的影响
盐生植物盐分离子运输过程中必然伴随着C、N、P元素含量的变化。已有研究认为, 增施N肥能够显著增加盐生植物的总盐分在各器官的积累, 增施P肥能够在一定程度上增加盐生植物各器官中的总盐分含量(王界平和田长彦, 2011 ), 对于生长在盐渍土上的植物, 增施N、P肥不仅可以明显提高植物的耐盐能力, 缓解盐分胁迫对植物的伤害, 还可以改善植株体内的N、P养分状况, 促进植株的生长发育(Rrezboem & Thomas, 1998 ; 冯固等, 2000 ; 段德玉等, 2005 ; 邵晶等, 2005 )。研究表明4种盐生植物盐离子及总盐分(除Ca2+ 与N之间无相关性和CO3 2- 、HCO3 - 与N显著负相关外)均与N显著正相关, 说明了盐离子和N元素具有相互促进的作用, 推测N素营养可以有效地促进盐生植物对盐离子的吸收, 进而有助于盐生植物对生境土壤的盐碱性的改良。而4种盐生植物盐离子及总盐分(除Ca2+ 和HCO3 - 外)均与C元素显著负相关, 表明了光合作用增强促进这些盐离子向上运输并使光合作用产物碳水化合物向下运输, 保证植物维持正常的生理功能。同时K+ 和CO3 2- 与P之间显著正相关, 那么增施P肥有助于盐生植物对其的吸收, K+ 和CO3 2- 增加不仅大大促进了光合作用, 还可以减少水分的蒸腾作用, 同时K+ 还可以维持细胞的渗透性, 抵御盐胁迫对植物造成的细胞损害和死亡。Na+ 、K+ 、CO3 2- 、总盐分与C:N、C:P之间显著负相关, 与N:P显著正相关, 进一步说明了N素营养有助于这些盐离子(尤其是Na+ )的吸收, K素营养有助于盐生植物对N和P的吸收。因此, 本研究表明了盐生植物体内离子分配与对养分的吸收利用具有一定的相互作用, 一定规律的化学计量比使其在盐渍化的环境中能正常生长发育, 成为盐生植物对盐渍化环境的生理生态适应的理化基础。同时也说明在盐生植物生境中若增加N素营养, 会有助于降低氯化物盐碱土对植物的盐碱性伤害, 有利于土壤的改良和盐生植被的重建。
4 结论
1) 在生长旺盛期4种盐生植物体内全盐量随盐离子从植物根部向顶端运输呈显著增加的趋势, 说明盐生植物体内能够形成“盐岛”效应。
2) 4种盐生植物C与N、P之间显著负相关, N与P之间显著正相关, 体现了盐生植物和其他高等陆地植物一样具有相同的养分利用策略。
3) 4种盐生植物中灌木盐生植物顶端N:P > 16, 草本盐生植物顶端N:P在14-16之间, 说明同一研究区不同生活型盐生植物植丛下的土壤养分有效性可能存在差异。
4) 4种盐生植物体内总盐分、Na+ 和K+ 与N、N:P之间显著正相关, 与C、C:N、C:P之间显著负相关, K+ 和CO3 2- 与P之间显著正相关, 说明了盐生植物器官盐分积累与其C、N、P化学计量特征之间存在一定的相互促进关系。
致谢 感谢新疆石河子大学王振楠、孟凡翔、李旭东等在野外实验中给予的大力帮助!
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Aerts R Chapin FS 2000 ). The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns
.Advances in Ecological Research 30 , 1 -67 .
[本文引用: 1]
[2]
Baldwin DS Rees GN Mitchell AM 2006 ). The short-term effects of salinization on anaerobic nutrient cycling and microbial community structure in sediment from freshwater wetland
.Wetlands 26 , 455 -464 .
[本文引用: 1]
[3]
Bao SD 2011). Soil Agricultural Chemistry Analysis . Chinese Agricultural Press, Beijing .(in Chinese ) [鲍士旦 (2011). 土壤农化分析. 中国农业出版社 , 北京.]
[4]
Donohue SJ Brown DE 1984 ). Optimum N concentration in winter wheat grown in coastal region of Virginia
.Communications in Soil Science and Plant Analysis 15 , 651 -661 .
[本文引用: 2]
[5]
Duan DY Liu XJ Li CZ Qiao HL 2005 ). Effects of nitrogen nutrition on the seedlings growth and the change of osmotic regulation substances in
Suaeda salsa under NaCl stress. Acta Prataculturae Sinica 14 (5 ), 63 -68 .(in Chinese with English abstract) [段德玉 , 刘小京 , 李存桢 , 乔海龙 (2005 ). N素营养对NaCl胁迫下盐地碱蓬幼苗生长及渗透调节物质变化的影响. 草业学报,
14(5) , 63 -68 .]
[本文引用: 1]
[6]
Feng G Li XL Zhang FS Li SX 2000 ). Effect of phosphorus and arbuscular mycorrhizal fungus on response of maize plant to saline environment
.Journal of Plant Resources and Environment 9 (2 ), 22 -26 .(in Chinese with English abstract) [冯固 , 李晓林 , 张福锁 , 李生秀 (2000 ). 施磷和接种AM真菌对玉米耐盐性的影响. 植物资源与环境学报,
9(2) , 22 -26 .]
[本文引用: 1]
[7]
Flowers TJ Colmer TD 2008 ). Salinity tolerance in halophytes
.New Phytologist 179 , 945 -963 .
[本文引用: 1]
[8]
Greenway H Munns R 1980 ). Mechanisms of salt tolerance in non-halophytes
.Plant Physiology 131 , 149 -190 .
[9]
Guo SQ Li WJ Zhang RY Wang G 2014 ). Effects of N and P additions on foliar stoichiometry and photosynthetic characteristics of
Potentilla fruiticosa. Guihaia 34 , 629 -634 .(in Chinese with English abstract) [郭淑青 , 李文金 , 张仁懿 , 王刚 (2014 ). 氮磷添加对金露梅叶片化学计量及光合特征的影响. 广西植物,
34 , 629 -634 .]
[本文引用: 1]
[10]
Han WX Fang JY Guo DJ Zhang Y 2005 ). Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China
.New Phytologist 168 , 337 -385 .
[本文引用: 1]
[11]
Hidaka A Kitayama K 2009 ). Divergent patterns of photosynthetic phosphorus-use efficiency versus nitrogen-use efficiency of tree leaves along nutrient-availability gradients
.Journal of Ecology 97 , 984 -991 .
[本文引用: 1]
[12]
Hikosaka K 2004 ). Interspecific difference in the photosynthesis- nitrogen relationship: Patterns, physiological causes, and ecological importance
.Journal of Plant Research 117 , 481 -494 .
[本文引用: 1]
[13]
Hong JT Wu JB Wang XD 2014 ). Purple flower needle couch grass root of carbon and phosphorus ecological stoichiometry characteristics
. Journal of Mountain Science 32 , 467 -474 .(in Chinese with English abstract) [洪江涛 , 吴建波 , 王小丹 (2014 ). 藏北高寒草原紫花针茅根系碳氮磷生态化学计量特征. 山地学报,
32 , 467 -474 .]
[本文引用: 1]
[14]
Hou XY 1982). Vegetation Geography and Advantages of Plant Chemical Composition . Science Press, Beijing .(in Chinese) [侯学煜 (1982). 中国植被地理及优势植物化学成分. 科学出版社 , 北京.]
[15]
Koerselman W Meuleman AFM 1996 ). The vegetation N/P ratios: A new tool to detect the nature of nutrient limitation
.Journal of Applied Ecology 33 , 1441 -1450 .
[16]
Krner C 1989 ). The nutritional status of plants from high altitudes
.Oecologia 81 , 379 -391 .
[本文引用: 1]
[17]
Lambers H Poorter H 2004 ). Inherent variation in growth rate between higher plants: A search for physiological causes and ecological consequences
. Advances in Ecological Research, 283-362.
[本文引用: 1]
[18]
Li Z Han L Liu YH An SQ Leng X 2012 ). C, N and P stoichiometric characteristics in leaves of
Suaeda salsa during different growth phase in coastal wetlands of China. Chinese Journal of Plant Ecology 36 , 1054 -1061 .(in Chinese with English abstract) [李征 , 韩琳 , 刘玉虹 , 安树青 , 冷欣 (2012 ). 滨海盐地碱蓬不同生长阶段叶片C、N、P化学计量特征. 植物生态学报,
36 , 1054 -1061 .]
[本文引用: 1]
[19]
Ma Y 2015 ). Responses of Desert Halophyte Foliar Stoichio- metry and Photosynthetic Characteristics to N and P Additions
. Master degree dissertation, Xinjiang University, Ürümqi. [马玉 (2015). 两种荒漠盐生植物叶片生态化学计量及光合特性对氮磷添加的相应. 硕士学位论文, 新疆大学, 乌鲁木齐.]
[20]
Munns R Termaat A 1986 ). Whole plant response to salinity
.Plant Physiology 13 , 143 -160 .
[本文引用: 1]
[21]
Niu DC Li Q Jing SG Chang PJ Fu H 2013 ). Seasonal variations of leaf C:N:P stoichiometry of six shrubs in desert of China’s Alxa Plateau
.Chinese Journal of Pant Ecology 37 , 317 -325 .(in Chinese with English abstract) [牛得草 , 李茜 , 江世高 , 常佩静 , 傅华 (2013 ). 阿拉善荒漠区6种主要灌木植物叶片C:N:P化学计量比的季节变化. 植物生态学报,
37 , 317 -325 .]
[本文引用: 2]
[22]
Peng F Huang CH Liao J Xue X Wang T 2015 ). Effect of
Kalidium foliatum community on soil salinization. Journal of Desert Research 35 (1 ), 90 -93 .(in Chinese with English abstract) [彭飞 , 黄翠华 , 廖杰 , 薛娴 , 王涛 (2015 ). 盐爪爪群落对土壤盐分影响. 中国沙漠,
35(1) , 90 -93 .]
[23]
Reich PB Oleksyn J 2004 ). Global patterns of plant leaf N and P in relation to temperature and latitude
.Proceedings of the National Academy of Sciences of the United States of America 101 , 11001 -11006 .
[本文引用: 1]
[24]
Rong QQ Liu JT Xia JB LU ZH Guo CH 2012 ). Leaf N and P stoichiometry of
Tamarix chinensis L in Laizhou Bay wetland. Chinese Journal of Ecology 31 , 3032 -3037 .(in Chinese with English abstract) [荣戗戗 , 刘京涛 , 夏江宝 , 陆兆华 , 郭彩虹 (2012 ). 莱州湾湿地柽柳叶片N、P生态化学计量特征. 生态学杂志,
31 , 3032 -3037 .]
[本文引用: 1]
[25]
Rrezboem FH Thomas GW 1998 ). Phosphorus nutrition affects wheat response to water deficit
.Agron 90 , 166 -171 .
[本文引用: 1]
[26]
Shabala S Bose J Hedrich R 2014 ). Salt bladders: Do they matter
.Trends in Plant Science 19 , 687 -691 .
[本文引用: 1]
[27]
Shao J Zheng QS Liu ZP Ning JF 2005 ). Effects of phosphorus application on ion distribution in aloe seedlings under seawater stress
.Acta Ecologica Sinica 25 , 3167 -3171 .(in Chinese with English abstract) [邵晶 , 郑青松 , 刘兆普 , 宁建凤 (2005 ). 磷对海水胁迫下芦荟幼苗离子分布的影响. 生态学报,
25 , 3167 -3171 .]
[本文引用: 1]
[28]
Song ZL Liu HY Zhao FJ Xu CY 2014 ). Ecological stoichiometry of N:P:Si in China’s grasslands
. Plant and Soil 380 , 165 -174 .
[本文引用: 1]
[29]
Sterner RW Elser JJ 2002). Ecological Stoichiometry : the Biology of Elements from Molecules to the Biosphere. Princeton University Press , Princeton . 87 -104 .
[30]
Tu JN Xiong YC Zhang X Yang Y Zhu LJ Chen JH Li ZH Wang SM 2011 ). “Fertile island” features of soil available nutrients around
Halostachys caspica shrub in the alluvial fan area of Manas River Watershed. Acta Ecologica Sinica 31 , 2461 -2470 .(in Chinese with English abstract) [涂锦娜 , 熊友才 , 张霞 , 杨岩 , 朱丽洁 , 陈接华 , 李志华 , 王绍明 (2011 ). 玛河流域扇缘带盐穗木土壤速效养分的“肥岛”特征. 生态学报,
31 , 2461 -2470 .]
[本文引用: 1]
[31]
Wang CM Zhang JL Liu XS 2009 ). Puccinellia tenuiflora maintains a low Na+ level under salinity by limiting unidirectional Na+ influx resulting in a high selectivity for K+ over Na+
.Plant, Cell & Environment 32 , 486 -496 .
[本文引用: 1]
[32]
Wang DM Yang HM 2011 ). C and N ecological characteristics of chemical metrology in different growth period of four kinds of forages
.Pratacultural Science 28 , 921 -925 .(in Chinese with English abstract) [王冬梅 , 杨惠敏 (2011 ). 4种牧草不同生长期C、N生态化学计量特征. 草业科学,
28 , 921 -925 .]
[33]
Wang JP Tian CY 2011 ). The Characteristics analysis of the growth and salt accumulation of
Salicornia europaea under different levels of nitrogen and phosphorus. Acta Prataculturae Sinica 20 , 232 -243 .(in Chinese with English abstract) [王界平 , 田长彦 (2011 ). 不同氮磷水平下盐角草生长及盐分积累特征分析. 草业学报,
20 , 232 -243 .]
[本文引用: 1]
[34]
Wang ZQ Zu SQ Yu RP 1993). Saline Soil in China . Science Press , Beijing . 400 -511 .(in Chinese) [王遵亲 , 祝寿泉 , 俞仁培 (1993). 中国盐渍土. 科学出版社 , 北京 . 400 -511 .]
[35]
Wright IJ Reich PB Westoby M Ackerly DD Baruch Z Bongers F Cavender-Bares J Chapin T Cornelissen JHC Diemer M Flexas J Garnier E Groom PK Gulias J Hikosaka K Lamont BB Lee T Lee W Lusk C Midgley JJ Navas M Niinemets , Oleksyn J Osada N Poorter H Poot P Prior L Pyankov VI Roumet C Thomas SC Tjoelker MG Veneklaas EJ Villar R 2004 ). The worldwide leaf economics spectrum
.Nature 428 , 821 -827 .
[36]
Wu TG Chen BF Xiao YH Pan YJ Chen Y Xiao YH 2010 ). Leaf stoichiometry of tree forest types in Pearl River Delta, South China
. Chinese Journal of Plant Ecology 34 , 58 -63 .(in Chinese with English abstract) [吴统贵 , 陈步峰 , 肖以华 , 潘勇军 , 陈勇 , 萧江华 (2010 ). 珠江三角洲3种典型森林类型乔木叶片生态化学计量学. 植物生态学报,
34 , 58 -63 .]
[本文引用: 1]
[37]
Xi JB Zhang FS Chen Y Mao DR Yin CH Tian CY 2004 ). A preliminary study on salt contents of soil in root-canopy area of halophytes
.Journal of Applied Ecology 15 , 53 -58 .(in Chinese with English abstract) [郗金标 , 张福锁 , 陈阳 , 毛达如 , 尹传华 , 田长彦 (2004 ). 盐生植物根冠区土壤盐分变化的初步研究. 应用生态学报,
15 , 53 -58 .]
[本文引用: 1]
[38]
Yan K Fu DG He F Duan CQ 2011 ). Leaf nutrient stoichiometry of plants in the phosphorus-enriched soils of the Lake Dianchi watershed, southwestern China
.Chinese Journal of Plant Ecology 35 , 353 -361 .(in Chinese with English abstract) [阎凯 , 付登高 , 何峰 , 段昌群 (2011 ). 滇池流域富磷区不同土壤磷水平下植物叶片的养分化学计量特征. 植物生态学报,
35 , 353 -361 .]
[本文引用: 1]
[39]
Yang K Huang JH Dong D Ma WH He JS 2010 ). Canopy leaf N and P stoichiometry in grassland communities of Qinghai-Tibetan Plateau, China
.Chinese Journal of Plant Ecology 34 , 17 -22 .(in Chinese with English abstract) [杨阔 , 黄建辉 , 董丹 , 马文红 , 贺金生 (2010 ). 青藏高原草地植物群落冠层叶片氮磷化学计量学分析. 植物生态学报,
34 , 17 -22 .]
[本文引用: 1]
[40]
Yin CH Feng G Tian CY Bai DS Zhang FS 2007 ). Influence of tamarisk shrub on the distribution of soil salinity and moisture on the edge of Taklamakan Desert
. China Environmental Science 27 , 670 -675 .(in Chinese with English abstract) [尹传华 , 冯固 , 田长彦 , 白灯莎 , 张福锁 (2007 ). 塔克拉玛干沙漠边缘柽柳对土壤水盐分布的影响. 中国环境科学,
27 , 670 -675 .]
[本文引用: 1]
[41]
Yu Q Chen Q Elser JJ 2010 ). Linking stoichiometric homoeostasis with ecosystem structure, functioning and stability
.Ecology Letters 13 , 1390 -1399 .
[本文引用: 1]
[42]
Zhang JL Li HR Guo SY Wang SM Shi HZ Han QQ Bao AK Ma Q 2015 ). Research advances in higher plant adaptation to salt stress
.Acta Prataculturae Sinica 24 (12 ), 220 -236 .(in Chinese with English abstract) [张金林 , 李慧茹 , 郭姝媛 , 王锁民 , 施华中 , 韩庆庆 , 包爱科 , 马青 (2015 ). 高等植物适应盐逆境研究进展. 草业学报,
24(12) , 220 -236 .]
[本文引用: 1]
[43]
Zhang JL Shi HZ 2013 ). Physiological and molecular mechanisms of plant salt tolerance
.Photosynthesis Research 115 , 1 -22 .
[本文引用: 1]
[44]
Zhang K He MZ LI XR Tan HJ Gao YH LI G Han GJ Wu YY 2014 ). Foliar carton, nitrogen and phosphorus stoichiometry of typical desert plants across the Alashan Desert
. Acta Ecologica Sinica 34 , 6538 -6547 .(in Chinese with English abstract) [张珂 , 何明珠 , 李新荣 , 谭会娟 , 高艳红 , 李刚 , 韩国君 , 吴杨杨 (2014 ). 阿拉善荒漠典型植物叶片碳氮磷化学计量特征. 生态学报,
34 , 6538 -6547 .]
[本文引用: 1]
[45]
Zhang LH Chen XB 2015 ). Characteristics of “salt island” and “fertile island” for
Tamarix chinensis and soil carton, nitrogen and phosphorus ecological stoichiometry in saline-alkali land. Journal of Applied Ecology 26 , 653 -658 .(in Chinese with English abstract) [张立华 , 陈小兵 (2015 ). 盐碱地柽柳“盐岛”和“肥岛”效应及其碳氮磷生态化学计量学特征. 应用生态学报,
26 , 653 -658 .]
[本文引用: 1]
[46]
Zhang WY Fan JW Zhong HP Hu ZM Song LL Wang N 2010 ). The nitrogen phosphorus stoichiometry of different plant functional groups four dominant species of typical steppes in China
.Acta Prataculturae Sinica 18 , 503 -509 .(in Chinese with English abstract) [张文彦 , 樊江文 , 钟华平 , 胡中民 , 宋璐璐 , 王宁 (2010 ). 中国典型草原优势植物功能群氮磷化学计量学特征研究. 草业学报,
18 , 503 -509 .]
[本文引用: 1]
[47]
Zhao KF 2002 ). Plants adapt to salt adversity
.Bulletin of Biology 37 (6 ), 7 -10 .(in Chinese with English abstract) [赵可夫 (2002 ). 植物对盐渍逆境的适应. 生物学通报,
37(6) , 7 -10 .]
[本文引用: 1]
The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns
1
2000
... 碳(C)、氮(N)、磷(P)作为植物体重要的组成元素, 对植物体的生长和生理机能的调节起着十分重要的作用(荣戗戗等, 2012 ), 而且它们之间的化学计量比强烈影响着植物在生态系统中的生长(Song et al ., 2014 ).C:N和C:P反映了植物的生长速率和对N、P养分的利用效率, 而N:P既反映了植物的生长受N或P或N、P共同的限制, 还可以作为一个关键指标决定群落的结构和功能(Aerts & Chapin, 2000 ).植物C、N、P的特征不仅体现了植物的特性, 也反映了植物对环境的长期适应和响应(侯学煜, 1982 ). ...
The short-term effects of salinization on anaerobic nutrient cycling and microbial community structure in sediment from freshwater wetland
1
2006
... 特定组织中的养分含量是衡量植物营养状况的一个重要指标, 它能反映植株的生长状况(Donohue & Brown, 1984 ), 并且它在植物各器官间的分配受植物自身的结构特点和生长节律的影响及生境地的养分有效性制约, 是物种系统发育和环境共同作用的结果(Baldwin et al ., 2006 ), 同时植物器官养分元素的含量也可以反映生境土壤的营养水平.研究表明4种盐生植物(灌木盐节木、盐爪爪、小叶碱蓬和草本盐角草)不同组织部位的C、N、P含量存有明显的差异, 表明了同一生活型(牛得草等, 2013 )和不同生活型(张珂等, 2014 )盐生植物存在明显的种间特异性, 体现了物种进化过程中植物种属对养分利用策略的分异特征.草本盐生植物盐角草顶端N和P含量均低于其他3种灌木盐生植物顶端N和P含量, 与Whight等(2004)和张珂等(2014)的研究结果正好相反, 反映该盐生灌木对N、P的吸收利用率高于盐生草本, 可能是因为灌木光合面积大, 光合作用强, 使顶端蒸腾作用加强, 全盐量积累大, 进而吸收更多的N和P来维持正常的生理功能.同时4种盐生植物C、N、P的变异系数分别为19%、54%、35%, 反映出C在这4种植物组织内最稳定, P含量次之, N含量最不稳定.一般而言, C在植物体内主要起骨架作用, 基本不参与植物生产活动, 所以变异最小, 最稳定; N来源广泛, 变异系数次之; P元素来源局限又易损失, 变异系数最大, 最不稳定(Krner, 1989 ; Reich & Oleksyn, 2004 ; 洪江涛等, 2014 ), 而该4种盐生植物却表现出P含量比N含量更稳定, 推测是植物自身属性和环境相互作用的结果.C含量与N、P含量之间均显著负相关, N与P之间显著正相关, 体现了盐生植物与其他高等陆地植物C、N、P化学计量特征(Sterner & Elser, 2002 ; 阎凯等, 2011 ; 李征等, 2012 )一样具备同样的规律, 反映了它们不同组织属性间的经济策略相同(有机体元素组成平衡), 是盐生植物种群能够在盐渍化生境中稳定生长发育的重要保障. ...
Optimum N concentration in winter wheat grown in coastal region of Virginia
2
1984
... 特定组织中的养分含量是衡量植物营养状况的一个重要指标, 它能反映植株的生长状况(Donohue & Brown, 1984 ), 并且它在植物各器官间的分配受植物自身的结构特点和生长节律的影响及生境地的养分有效性制约, 是物种系统发育和环境共同作用的结果(Baldwin et al ., 2006 ), 同时植物器官养分元素的含量也可以反映生境土壤的营养水平.研究表明4种盐生植物(灌木盐节木、盐爪爪、小叶碱蓬和草本盐角草)不同组织部位的C、N、P含量存有明显的差异, 表明了同一生活型(牛得草等, 2013 )和不同生活型(张珂等, 2014 )盐生植物存在明显的种间特异性, 体现了物种进化过程中植物种属对养分利用策略的分异特征.草本盐生植物盐角草顶端N和P含量均低于其他3种灌木盐生植物顶端N和P含量, 与Whight等(2004)和张珂等(2014)的研究结果正好相反, 反映该盐生灌木对N、P的吸收利用率高于盐生草本, 可能是因为灌木光合面积大, 光合作用强, 使顶端蒸腾作用加强, 全盐量积累大, 进而吸收更多的N和P来维持正常的生理功能.同时4种盐生植物C、N、P的变异系数分别为19%、54%、35%, 反映出C在这4种植物组织内最稳定, P含量次之, N含量最不稳定.一般而言, C在植物体内主要起骨架作用, 基本不参与植物生产活动, 所以变异最小, 最稳定; N来源广泛, 变异系数次之; P元素来源局限又易损失, 变异系数最大, 最不稳定(Krner, 1989 ; Reich & Oleksyn, 2004 ; 洪江涛等, 2014 ), 而该4种盐生植物却表现出P含量比N含量更稳定, 推测是植物自身属性和环境相互作用的结果.C含量与N、P含量之间均显著负相关, N与P之间显著正相关, 体现了盐生植物与其他高等陆地植物C、N、P化学计量特征(Sterner & Elser, 2002 ; 阎凯等, 2011 ; 李征等, 2012 )一样具备同样的规律, 反映了它们不同组织属性间的经济策略相同(有机体元素组成平衡), 是盐生植物种群能够在盐渍化生境中稳定生长发育的重要保障. ...
... 植物C:N反映了C积累的能力, 不同组织C:N差异很大(Donohue & Brown, 1984 ), 研究表明4种盐生植物茎顶端C:N远小于其余茎和根, 与王冬梅和杨惠敏(2011)所研究的牧草叶片C:N均低于茎、根的结果一致, 可能是由于茎和根的纤维化程度较高导致C含量高, 也反映了在植物生长旺盛期, 顶端全N含量最高.植物叶片N:P能够判断环境为植物生长所供应的养分状况(吴统贵等, 2010 ), Koerselaman和Meuleman (1996)研究表明, 植物生长受N和P限制的N:P阈值分别为14和16.即N:P < 14 时, 群落水平上的植物生长主要受N限制; 当N:P > 16时, 植物生长主要受P限制; 当N:P在14-16时, 则受N和P的共同限制或不受限.该研究表明小叶碱蓬、盐爪爪、盐节木顶端N:P均大于16, 受P限制; 盐角草顶端N:P接近16, 大于全国范围内陆地草本植物叶片平均N:P (13.5)(Han et al ., 2005 ), 受N、P共同限制(偏P), 进一步表明该盐碱地植物生长与荒漠(牛得草等, 2013 )和草原(杨阔等, 2010 ; 张文彦等, 2010 )一样主要受P的限制. ...
N素营养对NaCl胁迫下盐地碱蓬幼苗生长及渗透调节物质变化的影响. 草业学报,
1
2005
... 盐生植物盐分离子运输过程中必然伴随着C、N、P元素含量的变化.已有研究认为, 增施N肥能够显著增加盐生植物的总盐分在各器官的积累, 增施P肥能够在一定程度上增加盐生植物各器官中的总盐分含量(王界平和田长彦, 2011 ), 对于生长在盐渍土上的植物, 增施N、P肥不仅可以明显提高植物的耐盐能力, 缓解盐分胁迫对植物的伤害, 还可以改善植株体内的N、P养分状况, 促进植株的生长发育(Rrezboem & Thomas, 1998 ; 冯固等, 2000 ; 段德玉等, 2005 ; 邵晶等, 2005 ).研究表明4种盐生植物盐离子及总盐分(除Ca2+ 与N之间无相关性和CO3 2- 、HCO3 - 与N显著负相关外)均与N显著正相关, 说明了盐离子和N元素具有相互促进的作用, 推测N素营养可以有效地促进盐生植物对盐离子的吸收, 进而有助于盐生植物对生境土壤的盐碱性的改良.而4种盐生植物盐离子及总盐分(除Ca2+ 和HCO3 - 外)均与C元素显著负相关, 表明了光合作用增强促进这些盐离子向上运输并使光合作用产物碳水化合物向下运输, 保证植物维持正常的生理功能.同时K+ 和CO3 2- 与P之间显著正相关, 那么增施P肥有助于盐生植物对其的吸收, K+ 和CO3 2- 增加不仅大大促进了光合作用, 还可以减少水分的蒸腾作用, 同时K+ 还可以维持细胞的渗透性, 抵御盐胁迫对植物造成的细胞损害和死亡.Na+ 、K+ 、CO3 2- 、总盐分与C:N、C:P之间显著负相关, 与N:P显著正相关, 进一步说明了N素营养有助于这些盐离子(尤其是Na+ )的吸收, K素营养有助于盐生植物对N和P的吸收.因此, 本研究表明了盐生植物体内离子分配与对养分的吸收利用具有一定的相互作用, 一定规律的化学计量比使其在盐渍化的环境中能正常生长发育, 成为盐生植物对盐渍化环境的生理生态适应的理化基础.同时也说明在盐生植物生境中若增加N素营养, 会有助于降低氯化物盐碱土对植物的盐碱性伤害, 有利于土壤的改良和盐生植被的重建. ...
施磷和接种AM真菌对玉米耐盐性的影响. 植物资源与环境学报,
1
2000
... 盐生植物盐分离子运输过程中必然伴随着C、N、P元素含量的变化.已有研究认为, 增施N肥能够显著增加盐生植物的总盐分在各器官的积累, 增施P肥能够在一定程度上增加盐生植物各器官中的总盐分含量(王界平和田长彦, 2011 ), 对于生长在盐渍土上的植物, 增施N、P肥不仅可以明显提高植物的耐盐能力, 缓解盐分胁迫对植物的伤害, 还可以改善植株体内的N、P养分状况, 促进植株的生长发育(Rrezboem & Thomas, 1998 ; 冯固等, 2000 ; 段德玉等, 2005 ; 邵晶等, 2005 ).研究表明4种盐生植物盐离子及总盐分(除Ca2+ 与N之间无相关性和CO3 2- 、HCO3 - 与N显著负相关外)均与N显著正相关, 说明了盐离子和N元素具有相互促进的作用, 推测N素营养可以有效地促进盐生植物对盐离子的吸收, 进而有助于盐生植物对生境土壤的盐碱性的改良.而4种盐生植物盐离子及总盐分(除Ca2+ 和HCO3 - 外)均与C元素显著负相关, 表明了光合作用增强促进这些盐离子向上运输并使光合作用产物碳水化合物向下运输, 保证植物维持正常的生理功能.同时K+ 和CO3 2- 与P之间显著正相关, 那么增施P肥有助于盐生植物对其的吸收, K+ 和CO3 2- 增加不仅大大促进了光合作用, 还可以减少水分的蒸腾作用, 同时K+ 还可以维持细胞的渗透性, 抵御盐胁迫对植物造成的细胞损害和死亡.Na+ 、K+ 、CO3 2- 、总盐分与C:N、C:P之间显著负相关, 与N:P显著正相关, 进一步说明了N素营养有助于这些盐离子(尤其是Na+ )的吸收, K素营养有助于盐生植物对N和P的吸收.因此, 本研究表明了盐生植物体内离子分配与对养分的吸收利用具有一定的相互作用, 一定规律的化学计量比使其在盐渍化的环境中能正常生长发育, 成为盐生植物对盐渍化环境的生理生态适应的理化基础.同时也说明在盐生植物生境中若增加N素营养, 会有助于降低氯化物盐碱土对植物的盐碱性伤害, 有利于土壤的改良和盐生植被的重建. ...
Salinity tolerance in halophytes
1
2008
... 植物要适应盐渍化的生境, 必须具备克服盐离子毒害(离子胁迫)、抵抗低水势(渗透胁迫)和平衡养分吸收的能力(Greeway & Munns, 1980 ; Munns & Termaat, 1986 ), 于是在生理上形成一系列抗盐特征—稀盐、泌盐、积盐和拒盐(赵可夫, 2002 ; Flowers & Colmer, 2008 ; Wang et al ., 2009 ; Zhang & Shi, 2013 ; Shabala et al ., 2014 ), 来减轻盐胁迫对组织产生的伤害, 同样所有的高等植物也都可以通过这些生理方面的调节及导入耐盐基因来增强自身对盐胁迫的耐受性(张金林等, 2015 ). ...
Mechanisms of salt tolerance in non-halophytes
1980
氮磷添加对金露梅叶片化学计量及光合特征的影响. 广西植物,
1
2014
... 近年来研究者结合植物生理生态响应和生态化学计量特征进行了大量的研究(Hikosaka, 2004 ; Lambers & Poorter, 2004 ; Hidaka & Kitayama, 2009 ; Yu et al ., 2010 ; 郭淑青等, 2014 ).例如, 马玉(2015)发现植物光合特性与植物叶片养分有一定的关系, 在一定范围内的N浓度会促进植物光合特性与叶片养分间的关系, C含量与叶绿素呈负相关关系.这些研究对于了解盐生植物生态适应性及其对养分的利用策略具有一定的意义.因此, 本文从盐生植物体盐分对其化学计量特征影响的角度来揭示盐生植物在盐渍化生境中对养分和盐分的分配策略, 为理解盐碱地植物的生态过程和盐碱地改良利用提供科学依据. ...
Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China
1
2005
... 植物C:N反映了C积累的能力, 不同组织C:N差异很大(Donohue & Brown, 1984 ), 研究表明4种盐生植物茎顶端C:N远小于其余茎和根, 与王冬梅和杨惠敏(2011)所研究的牧草叶片C:N均低于茎、根的结果一致, 可能是由于茎和根的纤维化程度较高导致C含量高, 也反映了在植物生长旺盛期, 顶端全N含量最高.植物叶片N:P能够判断环境为植物生长所供应的养分状况(吴统贵等, 2010 ), Koerselaman和Meuleman (1996)研究表明, 植物生长受N和P限制的N:P阈值分别为14和16.即N:P < 14 时, 群落水平上的植物生长主要受N限制; 当N:P > 16时, 植物生长主要受P限制; 当N:P在14-16时, 则受N和P的共同限制或不受限.该研究表明小叶碱蓬、盐爪爪、盐节木顶端N:P均大于16, 受P限制; 盐角草顶端N:P接近16, 大于全国范围内陆地草本植物叶片平均N:P (13.5)(Han et al ., 2005 ), 受N、P共同限制(偏P), 进一步表明该盐碱地植物生长与荒漠(牛得草等, 2013 )和草原(杨阔等, 2010 ; 张文彦等, 2010 )一样主要受P的限制. ...
Divergent patterns of photosynthetic phosphorus-use efficiency versus nitrogen-use efficiency of tree leaves along nutrient-availability gradients
1
2009
... 近年来研究者结合植物生理生态响应和生态化学计量特征进行了大量的研究(Hikosaka, 2004 ; Lambers & Poorter, 2004 ; Hidaka & Kitayama, 2009 ; Yu et al ., 2010 ; 郭淑青等, 2014 ).例如, 马玉(2015)发现植物光合特性与植物叶片养分有一定的关系, 在一定范围内的N浓度会促进植物光合特性与叶片养分间的关系, C含量与叶绿素呈负相关关系.这些研究对于了解盐生植物生态适应性及其对养分的利用策略具有一定的意义.因此, 本文从盐生植物体盐分对其化学计量特征影响的角度来揭示盐生植物在盐渍化生境中对养分和盐分的分配策略, 为理解盐碱地植物的生态过程和盐碱地改良利用提供科学依据. ...
Interspecific difference in the photosynthesis- nitrogen relationship: Patterns, physiological causes, and ecological importance
1
2004
... 近年来研究者结合植物生理生态响应和生态化学计量特征进行了大量的研究(Hikosaka, 2004 ; Lambers & Poorter, 2004 ; Hidaka & Kitayama, 2009 ; Yu et al ., 2010 ; 郭淑青等, 2014 ).例如, 马玉(2015)发现植物光合特性与植物叶片养分有一定的关系, 在一定范围内的N浓度会促进植物光合特性与叶片养分间的关系, C含量与叶绿素呈负相关关系.这些研究对于了解盐生植物生态适应性及其对养分的利用策略具有一定的意义.因此, 本文从盐生植物体盐分对其化学计量特征影响的角度来揭示盐生植物在盐渍化生境中对养分和盐分的分配策略, 为理解盐碱地植物的生态过程和盐碱地改良利用提供科学依据. ...
藏北高寒草原紫花针茅根系碳氮磷生态化学计量特征. 山地学报,
1
2014
... 特定组织中的养分含量是衡量植物营养状况的一个重要指标, 它能反映植株的生长状况(Donohue & Brown, 1984 ), 并且它在植物各器官间的分配受植物自身的结构特点和生长节律的影响及生境地的养分有效性制约, 是物种系统发育和环境共同作用的结果(Baldwin et al ., 2006 ), 同时植物器官养分元素的含量也可以反映生境土壤的营养水平.研究表明4种盐生植物(灌木盐节木、盐爪爪、小叶碱蓬和草本盐角草)不同组织部位的C、N、P含量存有明显的差异, 表明了同一生活型(牛得草等, 2013 )和不同生活型(张珂等, 2014 )盐生植物存在明显的种间特异性, 体现了物种进化过程中植物种属对养分利用策略的分异特征.草本盐生植物盐角草顶端N和P含量均低于其他3种灌木盐生植物顶端N和P含量, 与Whight等(2004)和张珂等(2014)的研究结果正好相反, 反映该盐生灌木对N、P的吸收利用率高于盐生草本, 可能是因为灌木光合面积大, 光合作用强, 使顶端蒸腾作用加强, 全盐量积累大, 进而吸收更多的N和P来维持正常的生理功能.同时4种盐生植物C、N、P的变异系数分别为19%、54%、35%, 反映出C在这4种植物组织内最稳定, P含量次之, N含量最不稳定.一般而言, C在植物体内主要起骨架作用, 基本不参与植物生产活动, 所以变异最小, 最稳定; N来源广泛, 变异系数次之; P元素来源局限又易损失, 变异系数最大, 最不稳定(Krner, 1989 ; Reich & Oleksyn, 2004 ; 洪江涛等, 2014 ), 而该4种盐生植物却表现出P含量比N含量更稳定, 推测是植物自身属性和环境相互作用的结果.C含量与N、P含量之间均显著负相关, N与P之间显著正相关, 体现了盐生植物与其他高等陆地植物C、N、P化学计量特征(Sterner & Elser, 2002 ; 阎凯等, 2011 ; 李征等, 2012 )一样具备同样的规律, 反映了它们不同组织属性间的经济策略相同(有机体元素组成平衡), 是盐生植物种群能够在盐渍化生境中稳定生长发育的重要保障. ...
The vegetation N/P ratios: A new tool to detect the nature of nutrient limitation
1996
The nutritional status of plants from high altitudes
1
1989
... 特定组织中的养分含量是衡量植物营养状况的一个重要指标, 它能反映植株的生长状况(Donohue & Brown, 1984 ), 并且它在植物各器官间的分配受植物自身的结构特点和生长节律的影响及生境地的养分有效性制约, 是物种系统发育和环境共同作用的结果(Baldwin et al ., 2006 ), 同时植物器官养分元素的含量也可以反映生境土壤的营养水平.研究表明4种盐生植物(灌木盐节木、盐爪爪、小叶碱蓬和草本盐角草)不同组织部位的C、N、P含量存有明显的差异, 表明了同一生活型(牛得草等, 2013 )和不同生活型(张珂等, 2014 )盐生植物存在明显的种间特异性, 体现了物种进化过程中植物种属对养分利用策略的分异特征.草本盐生植物盐角草顶端N和P含量均低于其他3种灌木盐生植物顶端N和P含量, 与Whight等(2004)和张珂等(2014)的研究结果正好相反, 反映该盐生灌木对N、P的吸收利用率高于盐生草本, 可能是因为灌木光合面积大, 光合作用强, 使顶端蒸腾作用加强, 全盐量积累大, 进而吸收更多的N和P来维持正常的生理功能.同时4种盐生植物C、N、P的变异系数分别为19%、54%、35%, 反映出C在这4种植物组织内最稳定, P含量次之, N含量最不稳定.一般而言, C在植物体内主要起骨架作用, 基本不参与植物生产活动, 所以变异最小, 最稳定; N来源广泛, 变异系数次之; P元素来源局限又易损失, 变异系数最大, 最不稳定(Krner, 1989 ; Reich & Oleksyn, 2004 ; 洪江涛等, 2014 ), 而该4种盐生植物却表现出P含量比N含量更稳定, 推测是植物自身属性和环境相互作用的结果.C含量与N、P含量之间均显著负相关, N与P之间显著正相关, 体现了盐生植物与其他高等陆地植物C、N、P化学计量特征(Sterner & Elser, 2002 ; 阎凯等, 2011 ; 李征等, 2012 )一样具备同样的规律, 反映了它们不同组织属性间的经济策略相同(有机体元素组成平衡), 是盐生植物种群能够在盐渍化生境中稳定生长发育的重要保障. ...
Inherent variation in growth rate between higher plants: A search for physiological causes and ecological consequences
1
2004
... 近年来研究者结合植物生理生态响应和生态化学计量特征进行了大量的研究(Hikosaka, 2004 ; Lambers & Poorter, 2004 ; Hidaka & Kitayama, 2009 ; Yu et al ., 2010 ; 郭淑青等, 2014 ).例如, 马玉(2015)发现植物光合特性与植物叶片养分有一定的关系, 在一定范围内的N浓度会促进植物光合特性与叶片养分间的关系, C含量与叶绿素呈负相关关系.这些研究对于了解盐生植物生态适应性及其对养分的利用策略具有一定的意义.因此, 本文从盐生植物体盐分对其化学计量特征影响的角度来揭示盐生植物在盐渍化生境中对养分和盐分的分配策略, 为理解盐碱地植物的生态过程和盐碱地改良利用提供科学依据. ...
滨海盐地碱蓬不同生长阶段叶片C、N、P化学计量特征. 植物生态学报,
1
2012
... 特定组织中的养分含量是衡量植物营养状况的一个重要指标, 它能反映植株的生长状况(Donohue & Brown, 1984 ), 并且它在植物各器官间的分配受植物自身的结构特点和生长节律的影响及生境地的养分有效性制约, 是物种系统发育和环境共同作用的结果(Baldwin et al ., 2006 ), 同时植物器官养分元素的含量也可以反映生境土壤的营养水平.研究表明4种盐生植物(灌木盐节木、盐爪爪、小叶碱蓬和草本盐角草)不同组织部位的C、N、P含量存有明显的差异, 表明了同一生活型(牛得草等, 2013 )和不同生活型(张珂等, 2014 )盐生植物存在明显的种间特异性, 体现了物种进化过程中植物种属对养分利用策略的分异特征.草本盐生植物盐角草顶端N和P含量均低于其他3种灌木盐生植物顶端N和P含量, 与Whight等(2004)和张珂等(2014)的研究结果正好相反, 反映该盐生灌木对N、P的吸收利用率高于盐生草本, 可能是因为灌木光合面积大, 光合作用强, 使顶端蒸腾作用加强, 全盐量积累大, 进而吸收更多的N和P来维持正常的生理功能.同时4种盐生植物C、N、P的变异系数分别为19%、54%、35%, 反映出C在这4种植物组织内最稳定, P含量次之, N含量最不稳定.一般而言, C在植物体内主要起骨架作用, 基本不参与植物生产活动, 所以变异最小, 最稳定; N来源广泛, 变异系数次之; P元素来源局限又易损失, 变异系数最大, 最不稳定(Krner, 1989 ; Reich & Oleksyn, 2004 ; 洪江涛等, 2014 ), 而该4种盐生植物却表现出P含量比N含量更稳定, 推测是植物自身属性和环境相互作用的结果.C含量与N、P含量之间均显著负相关, N与P之间显著正相关, 体现了盐生植物与其他高等陆地植物C、N、P化学计量特征(Sterner & Elser, 2002 ; 阎凯等, 2011 ; 李征等, 2012 )一样具备同样的规律, 反映了它们不同组织属性间的经济策略相同(有机体元素组成平衡), 是盐生植物种群能够在盐渍化生境中稳定生长发育的重要保障. ...
Responses of Desert Halophyte Foliar Stoichio- metry and Photosynthetic Characteristics to N and P Additions
2015
Whole plant response to salinity
1
1986
... 植物要适应盐渍化的生境, 必须具备克服盐离子毒害(离子胁迫)、抵抗低水势(渗透胁迫)和平衡养分吸收的能力(Greeway & Munns, 1980 ; Munns & Termaat, 1986 ), 于是在生理上形成一系列抗盐特征—稀盐、泌盐、积盐和拒盐(赵可夫, 2002 ; Flowers & Colmer, 2008 ; Wang et al ., 2009 ; Zhang & Shi, 2013 ; Shabala et al ., 2014 ), 来减轻盐胁迫对组织产生的伤害, 同样所有的高等植物也都可以通过这些生理方面的调节及导入耐盐基因来增强自身对盐胁迫的耐受性(张金林等, 2015 ). ...
阿拉善荒漠区6种主要灌木植物叶片C:N:P化学计量比的季节变化. 植物生态学报,
2
2013
... 特定组织中的养分含量是衡量植物营养状况的一个重要指标, 它能反映植株的生长状况(Donohue & Brown, 1984 ), 并且它在植物各器官间的分配受植物自身的结构特点和生长节律的影响及生境地的养分有效性制约, 是物种系统发育和环境共同作用的结果(Baldwin et al ., 2006 ), 同时植物器官养分元素的含量也可以反映生境土壤的营养水平.研究表明4种盐生植物(灌木盐节木、盐爪爪、小叶碱蓬和草本盐角草)不同组织部位的C、N、P含量存有明显的差异, 表明了同一生活型(牛得草等, 2013 )和不同生活型(张珂等, 2014 )盐生植物存在明显的种间特异性, 体现了物种进化过程中植物种属对养分利用策略的分异特征.草本盐生植物盐角草顶端N和P含量均低于其他3种灌木盐生植物顶端N和P含量, 与Whight等(2004)和张珂等(2014)的研究结果正好相反, 反映该盐生灌木对N、P的吸收利用率高于盐生草本, 可能是因为灌木光合面积大, 光合作用强, 使顶端蒸腾作用加强, 全盐量积累大, 进而吸收更多的N和P来维持正常的生理功能.同时4种盐生植物C、N、P的变异系数分别为19%、54%、35%, 反映出C在这4种植物组织内最稳定, P含量次之, N含量最不稳定.一般而言, C在植物体内主要起骨架作用, 基本不参与植物生产活动, 所以变异最小, 最稳定; N来源广泛, 变异系数次之; P元素来源局限又易损失, 变异系数最大, 最不稳定(Krner, 1989 ; Reich & Oleksyn, 2004 ; 洪江涛等, 2014 ), 而该4种盐生植物却表现出P含量比N含量更稳定, 推测是植物自身属性和环境相互作用的结果.C含量与N、P含量之间均显著负相关, N与P之间显著正相关, 体现了盐生植物与其他高等陆地植物C、N、P化学计量特征(Sterner & Elser, 2002 ; 阎凯等, 2011 ; 李征等, 2012 )一样具备同样的规律, 反映了它们不同组织属性间的经济策略相同(有机体元素组成平衡), 是盐生植物种群能够在盐渍化生境中稳定生长发育的重要保障. ...
... 植物C:N反映了C积累的能力, 不同组织C:N差异很大(Donohue & Brown, 1984 ), 研究表明4种盐生植物茎顶端C:N远小于其余茎和根, 与王冬梅和杨惠敏(2011)所研究的牧草叶片C:N均低于茎、根的结果一致, 可能是由于茎和根的纤维化程度较高导致C含量高, 也反映了在植物生长旺盛期, 顶端全N含量最高.植物叶片N:P能够判断环境为植物生长所供应的养分状况(吴统贵等, 2010 ), Koerselaman和Meuleman (1996)研究表明, 植物生长受N和P限制的N:P阈值分别为14和16.即N:P < 14 时, 群落水平上的植物生长主要受N限制; 当N:P > 16时, 植物生长主要受P限制; 当N:P在14-16时, 则受N和P的共同限制或不受限.该研究表明小叶碱蓬、盐爪爪、盐节木顶端N:P均大于16, 受P限制; 盐角草顶端N:P接近16, 大于全国范围内陆地草本植物叶片平均N:P (13.5)(Han et al ., 2005 ), 受N、P共同限制(偏P), 进一步表明该盐碱地植物生长与荒漠(牛得草等, 2013 )和草原(杨阔等, 2010 ; 张文彦等, 2010 )一样主要受P的限制. ...
Global patterns of plant leaf N and P in relation to temperature and latitude
1
2004
... 特定组织中的养分含量是衡量植物营养状况的一个重要指标, 它能反映植株的生长状况(Donohue & Brown, 1984 ), 并且它在植物各器官间的分配受植物自身的结构特点和生长节律的影响及生境地的养分有效性制约, 是物种系统发育和环境共同作用的结果(Baldwin et al ., 2006 ), 同时植物器官养分元素的含量也可以反映生境土壤的营养水平.研究表明4种盐生植物(灌木盐节木、盐爪爪、小叶碱蓬和草本盐角草)不同组织部位的C、N、P含量存有明显的差异, 表明了同一生活型(牛得草等, 2013 )和不同生活型(张珂等, 2014 )盐生植物存在明显的种间特异性, 体现了物种进化过程中植物种属对养分利用策略的分异特征.草本盐生植物盐角草顶端N和P含量均低于其他3种灌木盐生植物顶端N和P含量, 与Whight等(2004)和张珂等(2014)的研究结果正好相反, 反映该盐生灌木对N、P的吸收利用率高于盐生草本, 可能是因为灌木光合面积大, 光合作用强, 使顶端蒸腾作用加强, 全盐量积累大, 进而吸收更多的N和P来维持正常的生理功能.同时4种盐生植物C、N、P的变异系数分别为19%、54%、35%, 反映出C在这4种植物组织内最稳定, P含量次之, N含量最不稳定.一般而言, C在植物体内主要起骨架作用, 基本不参与植物生产活动, 所以变异最小, 最稳定; N来源广泛, 变异系数次之; P元素来源局限又易损失, 变异系数最大, 最不稳定(Krner, 1989 ; Reich & Oleksyn, 2004 ; 洪江涛等, 2014 ), 而该4种盐生植物却表现出P含量比N含量更稳定, 推测是植物自身属性和环境相互作用的结果.C含量与N、P含量之间均显著负相关, N与P之间显著正相关, 体现了盐生植物与其他高等陆地植物C、N、P化学计量特征(Sterner & Elser, 2002 ; 阎凯等, 2011 ; 李征等, 2012 )一样具备同样的规律, 反映了它们不同组织属性间的经济策略相同(有机体元素组成平衡), 是盐生植物种群能够在盐渍化生境中稳定生长发育的重要保障. ...
莱州湾湿地柽柳叶片N、P生态化学计量特征. 生态学杂志,
1
2012
... 碳(C)、氮(N)、磷(P)作为植物体重要的组成元素, 对植物体的生长和生理机能的调节起着十分重要的作用(荣戗戗等, 2012 ), 而且它们之间的化学计量比强烈影响着植物在生态系统中的生长(Song et al ., 2014 ).C:N和C:P反映了植物的生长速率和对N、P养分的利用效率, 而N:P既反映了植物的生长受N或P或N、P共同的限制, 还可以作为一个关键指标决定群落的结构和功能(Aerts & Chapin, 2000 ).植物C、N、P的特征不仅体现了植物的特性, 也反映了植物对环境的长期适应和响应(侯学煜, 1982 ). ...
Phosphorus nutrition affects wheat response to water deficit
1
1998
... 盐生植物盐分离子运输过程中必然伴随着C、N、P元素含量的变化.已有研究认为, 增施N肥能够显著增加盐生植物的总盐分在各器官的积累, 增施P肥能够在一定程度上增加盐生植物各器官中的总盐分含量(王界平和田长彦, 2011 ), 对于生长在盐渍土上的植物, 增施N、P肥不仅可以明显提高植物的耐盐能力, 缓解盐分胁迫对植物的伤害, 还可以改善植株体内的N、P养分状况, 促进植株的生长发育(Rrezboem & Thomas, 1998 ; 冯固等, 2000 ; 段德玉等, 2005 ; 邵晶等, 2005 ).研究表明4种盐生植物盐离子及总盐分(除Ca2+ 与N之间无相关性和CO3 2- 、HCO3 - 与N显著负相关外)均与N显著正相关, 说明了盐离子和N元素具有相互促进的作用, 推测N素营养可以有效地促进盐生植物对盐离子的吸收, 进而有助于盐生植物对生境土壤的盐碱性的改良.而4种盐生植物盐离子及总盐分(除Ca2+ 和HCO3 - 外)均与C元素显著负相关, 表明了光合作用增强促进这些盐离子向上运输并使光合作用产物碳水化合物向下运输, 保证植物维持正常的生理功能.同时K+ 和CO3 2- 与P之间显著正相关, 那么增施P肥有助于盐生植物对其的吸收, K+ 和CO3 2- 增加不仅大大促进了光合作用, 还可以减少水分的蒸腾作用, 同时K+ 还可以维持细胞的渗透性, 抵御盐胁迫对植物造成的细胞损害和死亡.Na+ 、K+ 、CO3 2- 、总盐分与C:N、C:P之间显著负相关, 与N:P显著正相关, 进一步说明了N素营养有助于这些盐离子(尤其是Na+ )的吸收, K素营养有助于盐生植物对N和P的吸收.因此, 本研究表明了盐生植物体内离子分配与对养分的吸收利用具有一定的相互作用, 一定规律的化学计量比使其在盐渍化的环境中能正常生长发育, 成为盐生植物对盐渍化环境的生理生态适应的理化基础.同时也说明在盐生植物生境中若增加N素营养, 会有助于降低氯化物盐碱土对植物的盐碱性伤害, 有利于土壤的改良和盐生植被的重建. ...
Salt bladders: Do they matter
1
2014
... 植物要适应盐渍化的生境, 必须具备克服盐离子毒害(离子胁迫)、抵抗低水势(渗透胁迫)和平衡养分吸收的能力(Greeway & Munns, 1980 ; Munns & Termaat, 1986 ), 于是在生理上形成一系列抗盐特征—稀盐、泌盐、积盐和拒盐(赵可夫, 2002 ; Flowers & Colmer, 2008 ; Wang et al ., 2009 ; Zhang & Shi, 2013 ; Shabala et al ., 2014 ), 来减轻盐胁迫对组织产生的伤害, 同样所有的高等植物也都可以通过这些生理方面的调节及导入耐盐基因来增强自身对盐胁迫的耐受性(张金林等, 2015 ). ...
磷对海水胁迫下芦荟幼苗离子分布的影响. 生态学报,
1
2005
... 盐生植物盐分离子运输过程中必然伴随着C、N、P元素含量的变化.已有研究认为, 增施N肥能够显著增加盐生植物的总盐分在各器官的积累, 增施P肥能够在一定程度上增加盐生植物各器官中的总盐分含量(王界平和田长彦, 2011 ), 对于生长在盐渍土上的植物, 增施N、P肥不仅可以明显提高植物的耐盐能力, 缓解盐分胁迫对植物的伤害, 还可以改善植株体内的N、P养分状况, 促进植株的生长发育(Rrezboem & Thomas, 1998 ; 冯固等, 2000 ; 段德玉等, 2005 ; 邵晶等, 2005 ).研究表明4种盐生植物盐离子及总盐分(除Ca2+ 与N之间无相关性和CO3 2- 、HCO3 - 与N显著负相关外)均与N显著正相关, 说明了盐离子和N元素具有相互促进的作用, 推测N素营养可以有效地促进盐生植物对盐离子的吸收, 进而有助于盐生植物对生境土壤的盐碱性的改良.而4种盐生植物盐离子及总盐分(除Ca2+ 和HCO3 - 外)均与C元素显著负相关, 表明了光合作用增强促进这些盐离子向上运输并使光合作用产物碳水化合物向下运输, 保证植物维持正常的生理功能.同时K+ 和CO3 2- 与P之间显著正相关, 那么增施P肥有助于盐生植物对其的吸收, K+ 和CO3 2- 增加不仅大大促进了光合作用, 还可以减少水分的蒸腾作用, 同时K+ 还可以维持细胞的渗透性, 抵御盐胁迫对植物造成的细胞损害和死亡.Na+ 、K+ 、CO3 2- 、总盐分与C:N、C:P之间显著负相关, 与N:P显著正相关, 进一步说明了N素营养有助于这些盐离子(尤其是Na+ )的吸收, K素营养有助于盐生植物对N和P的吸收.因此, 本研究表明了盐生植物体内离子分配与对养分的吸收利用具有一定的相互作用, 一定规律的化学计量比使其在盐渍化的环境中能正常生长发育, 成为盐生植物对盐渍化环境的生理生态适应的理化基础.同时也说明在盐生植物生境中若增加N素营养, 会有助于降低氯化物盐碱土对植物的盐碱性伤害, 有利于土壤的改良和盐生植被的重建. ...
Ecological stoichiometry of N:P:Si in China’s grasslands
1
2014
... 碳(C)、氮(N)、磷(P)作为植物体重要的组成元素, 对植物体的生长和生理机能的调节起着十分重要的作用(荣戗戗等, 2012 ), 而且它们之间的化学计量比强烈影响着植物在生态系统中的生长(Song et al ., 2014 ).C:N和C:P反映了植物的生长速率和对N、P养分的利用效率, 而N:P既反映了植物的生长受N或P或N、P共同的限制, 还可以作为一个关键指标决定群落的结构和功能(Aerts & Chapin, 2000 ).植物C、N、P的特征不仅体现了植物的特性, 也反映了植物对环境的长期适应和响应(侯学煜, 1982 ). ...
玛河流域扇缘带盐穗木土壤速效养分的“肥岛”特征. 生态学报,
1
2011
... 研究区位于玛纳斯河流域扇缘带(44.37°- 44.48° N, 85.90-85.98° E), 属于典型的温带大陆性干旱半干旱气候区, 具有冬季严寒、夏季酷热、日照充足、干旱少雨等特点, 平均海拔377 m, 年平均气温6.9 ℃ (7月最热, 平均气温26 ℃, 1月最冷, 最低气温-18.5 ℃), 年降水量140-180 mm, 年蒸发量1500-2100 mm (其中4-9月蒸发量最大, 约占全年蒸发量的88%).该区域土壤盐渍化程度偏重(7月份土壤基本理化性质: 电导率13.36 ms·cm-1 ; 含水率12.13%; 有机质5.35 g·kg-1 ; 全N 0.46 g·kg-1 ; 全P 0.53 g·kg-1 ; 碱解氮0.044 g·kg-1 ; 速效磷0.012 g·kg-1 ; 速效钾0.320 g·kg-1 ), 地表具有2-5 cm的盐结皮, 积盐厚度10-20 cm, 土壤类型为氯化物-硫酸盐潮盐土(王遵亲等, 1993 ).该区植物群落结构通常表现为由不连续的灌木层和相对连续的草本层组成的典型灌-草型二元结构群落(涂锦娜等, 2011 ), 主要包括盐节木(Halocnemum strobilaceum )、小叶碱蓬(Suaeda microphylla )、盐爪爪(Kalidium foliatum )、盐角草(Salicornia europaea )、多枝柽柳(Tamarix ramosissima )、花花柴(Karelinia caspia )等. ...
Puccinellia tenuiflora maintains a low Na+ level under salinity by limiting unidirectional Na+ influx resulting in a high selectivity for K+ over Na+
1
2009
... 植物要适应盐渍化的生境, 必须具备克服盐离子毒害(离子胁迫)、抵抗低水势(渗透胁迫)和平衡养分吸收的能力(Greeway & Munns, 1980 ; Munns & Termaat, 1986 ), 于是在生理上形成一系列抗盐特征—稀盐、泌盐、积盐和拒盐(赵可夫, 2002 ; Flowers & Colmer, 2008 ; Wang et al ., 2009 ; Zhang & Shi, 2013 ; Shabala et al ., 2014 ), 来减轻盐胁迫对组织产生的伤害, 同样所有的高等植物也都可以通过这些生理方面的调节及导入耐盐基因来增强自身对盐胁迫的耐受性(张金林等, 2015 ). ...
4种牧草不同生长期C、N生态化学计量特征. 草业科学,
2011
不同氮磷水平下盐角草生长及盐分积累特征分析. 草业学报,
1
2011
... 盐生植物盐分离子运输过程中必然伴随着C、N、P元素含量的变化.已有研究认为, 增施N肥能够显著增加盐生植物的总盐分在各器官的积累, 增施P肥能够在一定程度上增加盐生植物各器官中的总盐分含量(王界平和田长彦, 2011 ), 对于生长在盐渍土上的植物, 增施N、P肥不仅可以明显提高植物的耐盐能力, 缓解盐分胁迫对植物的伤害, 还可以改善植株体内的N、P养分状况, 促进植株的生长发育(Rrezboem & Thomas, 1998 ; 冯固等, 2000 ; 段德玉等, 2005 ; 邵晶等, 2005 ).研究表明4种盐生植物盐离子及总盐分(除Ca2+ 与N之间无相关性和CO3 2- 、HCO3 - 与N显著负相关外)均与N显著正相关, 说明了盐离子和N元素具有相互促进的作用, 推测N素营养可以有效地促进盐生植物对盐离子的吸收, 进而有助于盐生植物对生境土壤的盐碱性的改良.而4种盐生植物盐离子及总盐分(除Ca2+ 和HCO3 - 外)均与C元素显著负相关, 表明了光合作用增强促进这些盐离子向上运输并使光合作用产物碳水化合物向下运输, 保证植物维持正常的生理功能.同时K+ 和CO3 2- 与P之间显著正相关, 那么增施P肥有助于盐生植物对其的吸收, K+ 和CO3 2- 增加不仅大大促进了光合作用, 还可以减少水分的蒸腾作用, 同时K+ 还可以维持细胞的渗透性, 抵御盐胁迫对植物造成的细胞损害和死亡.Na+ 、K+ 、CO3 2- 、总盐分与C:N、C:P之间显著负相关, 与N:P显著正相关, 进一步说明了N素营养有助于这些盐离子(尤其是Na+ )的吸收, K素营养有助于盐生植物对N和P的吸收.因此, 本研究表明了盐生植物体内离子分配与对养分的吸收利用具有一定的相互作用, 一定规律的化学计量比使其在盐渍化的环境中能正常生长发育, 成为盐生植物对盐渍化环境的生理生态适应的理化基础.同时也说明在盐生植物生境中若增加N素营养, 会有助于降低氯化物盐碱土对植物的盐碱性伤害, 有利于土壤的改良和盐生植被的重建. ...
The worldwide leaf economics spectrum
2004
珠江三角洲3种典型森林类型乔木叶片生态化学计量学. 植物生态学报,
1
2010
... 植物C:N反映了C积累的能力, 不同组织C:N差异很大(Donohue & Brown, 1984 ), 研究表明4种盐生植物茎顶端C:N远小于其余茎和根, 与王冬梅和杨惠敏(2011)所研究的牧草叶片C:N均低于茎、根的结果一致, 可能是由于茎和根的纤维化程度较高导致C含量高, 也反映了在植物生长旺盛期, 顶端全N含量最高.植物叶片N:P能够判断环境为植物生长所供应的养分状况(吴统贵等, 2010 ), Koerselaman和Meuleman (1996)研究表明, 植物生长受N和P限制的N:P阈值分别为14和16.即N:P < 14 时, 群落水平上的植物生长主要受N限制; 当N:P > 16时, 植物生长主要受P限制; 当N:P在14-16时, 则受N和P的共同限制或不受限.该研究表明小叶碱蓬、盐爪爪、盐节木顶端N:P均大于16, 受P限制; 盐角草顶端N:P接近16, 大于全国范围内陆地草本植物叶片平均N:P (13.5)(Han et al ., 2005 ), 受N、P共同限制(偏P), 进一步表明该盐碱地植物生长与荒漠(牛得草等, 2013 )和草原(杨阔等, 2010 ; 张文彦等, 2010 )一样主要受P的限制. ...
盐生植物根冠区土壤盐分变化的初步研究. 应用生态学报,
1
2004
... 有研究显示, 盐生植物能够在其植丛下形成“盐岛”效应(郗金标等, 2004 ; 尹传华等, 2007 ; 张立华和陈小兵, 2015 ), 而该研究显示盐生植物还能够在其植丛上形成同样的规律.该研究表明4种盐生植物在植物体内全盐量随盐离子从根部向顶端运输过程中呈显著增加趋势, 推测在植物生长旺盛期蒸腾作用的增强大大促进了体内盐分离子(尤其是Na+ 、Cl- )的运输, 造成盐生植物体内出现“盐岛”效应.而Na+ 、Cl- 是重要的无机渗透调节物质, 4种盐生植物通过吸收和积累无机离子, 将Na+ 、Cl- 区域化于液泡中, 达到渗透调节和降低离子的毒害作用.同时4种盐生植物还通过叶、茎的肉质化或叶退化成鳞片状, 促使在盐渍环境下贮存水分, 保证光合作用的进行, 完成了正常的生理功能.然而彭飞等(2015)对盐爪爪体内的总盐分积累的研究与此结果并不一致, 可能是因为同一物种由于采摘时期不一致导致盐分运输规律不同, 或者因生活环境的不同发生了变异.该研究还表明器官和物种及二者的交互作用影响植物对盐离子(除Mg2+ 外)的吸收和积累, 但4种盐生植物对盐离子的吸收具有一致的选择作用(吸收强度从高到低: Na+ >> Cl- > K+ 、SO4 2- > Ca2+ 、Mg2+ 、CO3 2- 、HCO3 - ), 这种对Na+ 、Cl- 较强的集聚特征使这些盐生植物更适用于氯化物盐渍土的改良, 同时植物本身还可以作为优良的饲料供牲畜 食用. ...
滇池流域富磷区不同土壤磷水平下植物叶片的养分化学计量特征. 植物生态学报,
1
2011
... 特定组织中的养分含量是衡量植物营养状况的一个重要指标, 它能反映植株的生长状况(Donohue & Brown, 1984 ), 并且它在植物各器官间的分配受植物自身的结构特点和生长节律的影响及生境地的养分有效性制约, 是物种系统发育和环境共同作用的结果(Baldwin et al ., 2006 ), 同时植物器官养分元素的含量也可以反映生境土壤的营养水平.研究表明4种盐生植物(灌木盐节木、盐爪爪、小叶碱蓬和草本盐角草)不同组织部位的C、N、P含量存有明显的差异, 表明了同一生活型(牛得草等, 2013 )和不同生活型(张珂等, 2014 )盐生植物存在明显的种间特异性, 体现了物种进化过程中植物种属对养分利用策略的分异特征.草本盐生植物盐角草顶端N和P含量均低于其他3种灌木盐生植物顶端N和P含量, 与Whight等(2004)和张珂等(2014)的研究结果正好相反, 反映该盐生灌木对N、P的吸收利用率高于盐生草本, 可能是因为灌木光合面积大, 光合作用强, 使顶端蒸腾作用加强, 全盐量积累大, 进而吸收更多的N和P来维持正常的生理功能.同时4种盐生植物C、N、P的变异系数分别为19%、54%、35%, 反映出C在这4种植物组织内最稳定, P含量次之, N含量最不稳定.一般而言, C在植物体内主要起骨架作用, 基本不参与植物生产活动, 所以变异最小, 最稳定; N来源广泛, 变异系数次之; P元素来源局限又易损失, 变异系数最大, 最不稳定(Krner, 1989 ; Reich & Oleksyn, 2004 ; 洪江涛等, 2014 ), 而该4种盐生植物却表现出P含量比N含量更稳定, 推测是植物自身属性和环境相互作用的结果.C含量与N、P含量之间均显著负相关, N与P之间显著正相关, 体现了盐生植物与其他高等陆地植物C、N、P化学计量特征(Sterner & Elser, 2002 ; 阎凯等, 2011 ; 李征等, 2012 )一样具备同样的规律, 反映了它们不同组织属性间的经济策略相同(有机体元素组成平衡), 是盐生植物种群能够在盐渍化生境中稳定生长发育的重要保障. ...
青藏高原草地植物群落冠层叶片氮磷化学计量学分析. 植物生态学报,
1
2010
... 植物C:N反映了C积累的能力, 不同组织C:N差异很大(Donohue & Brown, 1984 ), 研究表明4种盐生植物茎顶端C:N远小于其余茎和根, 与王冬梅和杨惠敏(2011)所研究的牧草叶片C:N均低于茎、根的结果一致, 可能是由于茎和根的纤维化程度较高导致C含量高, 也反映了在植物生长旺盛期, 顶端全N含量最高.植物叶片N:P能够判断环境为植物生长所供应的养分状况(吴统贵等, 2010 ), Koerselaman和Meuleman (1996)研究表明, 植物生长受N和P限制的N:P阈值分别为14和16.即N:P < 14 时, 群落水平上的植物生长主要受N限制; 当N:P > 16时, 植物生长主要受P限制; 当N:P在14-16时, 则受N和P的共同限制或不受限.该研究表明小叶碱蓬、盐爪爪、盐节木顶端N:P均大于16, 受P限制; 盐角草顶端N:P接近16, 大于全国范围内陆地草本植物叶片平均N:P (13.5)(Han et al ., 2005 ), 受N、P共同限制(偏P), 进一步表明该盐碱地植物生长与荒漠(牛得草等, 2013 )和草原(杨阔等, 2010 ; 张文彦等, 2010 )一样主要受P的限制. ...
塔克拉玛干沙漠边缘柽柳对土壤水盐分布的影响. 中国环境科学,
1
2007
... 有研究显示, 盐生植物能够在其植丛下形成“盐岛”效应(郗金标等, 2004 ; 尹传华等, 2007 ; 张立华和陈小兵, 2015 ), 而该研究显示盐生植物还能够在其植丛上形成同样的规律.该研究表明4种盐生植物在植物体内全盐量随盐离子从根部向顶端运输过程中呈显著增加趋势, 推测在植物生长旺盛期蒸腾作用的增强大大促进了体内盐分离子(尤其是Na+ 、Cl- )的运输, 造成盐生植物体内出现“盐岛”效应.而Na+ 、Cl- 是重要的无机渗透调节物质, 4种盐生植物通过吸收和积累无机离子, 将Na+ 、Cl- 区域化于液泡中, 达到渗透调节和降低离子的毒害作用.同时4种盐生植物还通过叶、茎的肉质化或叶退化成鳞片状, 促使在盐渍环境下贮存水分, 保证光合作用的进行, 完成了正常的生理功能.然而彭飞等(2015)对盐爪爪体内的总盐分积累的研究与此结果并不一致, 可能是因为同一物种由于采摘时期不一致导致盐分运输规律不同, 或者因生活环境的不同发生了变异.该研究还表明器官和物种及二者的交互作用影响植物对盐离子(除Mg2+ 外)的吸收和积累, 但4种盐生植物对盐离子的吸收具有一致的选择作用(吸收强度从高到低: Na+ >> Cl- > K+ 、SO4 2- > Ca2+ 、Mg2+ 、CO3 2- 、HCO3 - ), 这种对Na+ 、Cl- 较强的集聚特征使这些盐生植物更适用于氯化物盐渍土的改良, 同时植物本身还可以作为优良的饲料供牲畜 食用. ...
Linking stoichiometric homoeostasis with ecosystem structure, functioning and stability
1
2010
... 近年来研究者结合植物生理生态响应和生态化学计量特征进行了大量的研究(Hikosaka, 2004 ; Lambers & Poorter, 2004 ; Hidaka & Kitayama, 2009 ; Yu et al ., 2010 ; 郭淑青等, 2014 ).例如, 马玉(2015)发现植物光合特性与植物叶片养分有一定的关系, 在一定范围内的N浓度会促进植物光合特性与叶片养分间的关系, C含量与叶绿素呈负相关关系.这些研究对于了解盐生植物生态适应性及其对养分的利用策略具有一定的意义.因此, 本文从盐生植物体盐分对其化学计量特征影响的角度来揭示盐生植物在盐渍化生境中对养分和盐分的分配策略, 为理解盐碱地植物的生态过程和盐碱地改良利用提供科学依据. ...
高等植物适应盐逆境研究进展. 草业学报,
1
2015
... 植物要适应盐渍化的生境, 必须具备克服盐离子毒害(离子胁迫)、抵抗低水势(渗透胁迫)和平衡养分吸收的能力(Greeway & Munns, 1980 ; Munns & Termaat, 1986 ), 于是在生理上形成一系列抗盐特征—稀盐、泌盐、积盐和拒盐(赵可夫, 2002 ; Flowers & Colmer, 2008 ; Wang et al ., 2009 ; Zhang & Shi, 2013 ; Shabala et al ., 2014 ), 来减轻盐胁迫对组织产生的伤害, 同样所有的高等植物也都可以通过这些生理方面的调节及导入耐盐基因来增强自身对盐胁迫的耐受性(张金林等, 2015 ). ...
Physiological and molecular mechanisms of plant salt tolerance
1
2013
... 植物要适应盐渍化的生境, 必须具备克服盐离子毒害(离子胁迫)、抵抗低水势(渗透胁迫)和平衡养分吸收的能力(Greeway & Munns, 1980 ; Munns & Termaat, 1986 ), 于是在生理上形成一系列抗盐特征—稀盐、泌盐、积盐和拒盐(赵可夫, 2002 ; Flowers & Colmer, 2008 ; Wang et al ., 2009 ; Zhang & Shi, 2013 ; Shabala et al ., 2014 ), 来减轻盐胁迫对组织产生的伤害, 同样所有的高等植物也都可以通过这些生理方面的调节及导入耐盐基因来增强自身对盐胁迫的耐受性(张金林等, 2015 ). ...
阿拉善荒漠典型植物叶片碳氮磷化学计量特征. 生态学报,
1
2014
... 特定组织中的养分含量是衡量植物营养状况的一个重要指标, 它能反映植株的生长状况(Donohue & Brown, 1984 ), 并且它在植物各器官间的分配受植物自身的结构特点和生长节律的影响及生境地的养分有效性制约, 是物种系统发育和环境共同作用的结果(Baldwin et al ., 2006 ), 同时植物器官养分元素的含量也可以反映生境土壤的营养水平.研究表明4种盐生植物(灌木盐节木、盐爪爪、小叶碱蓬和草本盐角草)不同组织部位的C、N、P含量存有明显的差异, 表明了同一生活型(牛得草等, 2013 )和不同生活型(张珂等, 2014 )盐生植物存在明显的种间特异性, 体现了物种进化过程中植物种属对养分利用策略的分异特征.草本盐生植物盐角草顶端N和P含量均低于其他3种灌木盐生植物顶端N和P含量, 与Whight等(2004)和张珂等(2014)的研究结果正好相反, 反映该盐生灌木对N、P的吸收利用率高于盐生草本, 可能是因为灌木光合面积大, 光合作用强, 使顶端蒸腾作用加强, 全盐量积累大, 进而吸收更多的N和P来维持正常的生理功能.同时4种盐生植物C、N、P的变异系数分别为19%、54%、35%, 反映出C在这4种植物组织内最稳定, P含量次之, N含量最不稳定.一般而言, C在植物体内主要起骨架作用, 基本不参与植物生产活动, 所以变异最小, 最稳定; N来源广泛, 变异系数次之; P元素来源局限又易损失, 变异系数最大, 最不稳定(Krner, 1989 ; Reich & Oleksyn, 2004 ; 洪江涛等, 2014 ), 而该4种盐生植物却表现出P含量比N含量更稳定, 推测是植物自身属性和环境相互作用的结果.C含量与N、P含量之间均显著负相关, N与P之间显著正相关, 体现了盐生植物与其他高等陆地植物C、N、P化学计量特征(Sterner & Elser, 2002 ; 阎凯等, 2011 ; 李征等, 2012 )一样具备同样的规律, 反映了它们不同组织属性间的经济策略相同(有机体元素组成平衡), 是盐生植物种群能够在盐渍化生境中稳定生长发育的重要保障. ...
盐碱地柽柳“盐岛”和“肥岛”效应及其碳氮磷生态化学计量学特征. 应用生态学报,
1
2015
... 有研究显示, 盐生植物能够在其植丛下形成“盐岛”效应(郗金标等, 2004 ; 尹传华等, 2007 ; 张立华和陈小兵, 2015 ), 而该研究显示盐生植物还能够在其植丛上形成同样的规律.该研究表明4种盐生植物在植物体内全盐量随盐离子从根部向顶端运输过程中呈显著增加趋势, 推测在植物生长旺盛期蒸腾作用的增强大大促进了体内盐分离子(尤其是Na+ 、Cl- )的运输, 造成盐生植物体内出现“盐岛”效应.而Na+ 、Cl- 是重要的无机渗透调节物质, 4种盐生植物通过吸收和积累无机离子, 将Na+ 、Cl- 区域化于液泡中, 达到渗透调节和降低离子的毒害作用.同时4种盐生植物还通过叶、茎的肉质化或叶退化成鳞片状, 促使在盐渍环境下贮存水分, 保证光合作用的进行, 完成了正常的生理功能.然而彭飞等(2015)对盐爪爪体内的总盐分积累的研究与此结果并不一致, 可能是因为同一物种由于采摘时期不一致导致盐分运输规律不同, 或者因生活环境的不同发生了变异.该研究还表明器官和物种及二者的交互作用影响植物对盐离子(除Mg2+ 外)的吸收和积累, 但4种盐生植物对盐离子的吸收具有一致的选择作用(吸收强度从高到低: Na+ >> Cl- > K+ 、SO4 2- > Ca2+ 、Mg2+ 、CO3 2- 、HCO3 - ), 这种对Na+ 、Cl- 较强的集聚特征使这些盐生植物更适用于氯化物盐渍土的改良, 同时植物本身还可以作为优良的饲料供牲畜 食用. ...
中国典型草原优势植物功能群氮磷化学计量学特征研究. 草业学报,
1
2010
... 植物C:N反映了C积累的能力, 不同组织C:N差异很大(Donohue & Brown, 1984 ), 研究表明4种盐生植物茎顶端C:N远小于其余茎和根, 与王冬梅和杨惠敏(2011)所研究的牧草叶片C:N均低于茎、根的结果一致, 可能是由于茎和根的纤维化程度较高导致C含量高, 也反映了在植物生长旺盛期, 顶端全N含量最高.植物叶片N:P能够判断环境为植物生长所供应的养分状况(吴统贵等, 2010 ), Koerselaman和Meuleman (1996)研究表明, 植物生长受N和P限制的N:P阈值分别为14和16.即N:P < 14 时, 群落水平上的植物生长主要受N限制; 当N:P > 16时, 植物生长主要受P限制; 当N:P在14-16时, 则受N和P的共同限制或不受限.该研究表明小叶碱蓬、盐爪爪、盐节木顶端N:P均大于16, 受P限制; 盐角草顶端N:P接近16, 大于全国范围内陆地草本植物叶片平均N:P (13.5)(Han et al ., 2005 ), 受N、P共同限制(偏P), 进一步表明该盐碱地植物生长与荒漠(牛得草等, 2013 )和草原(杨阔等, 2010 ; 张文彦等, 2010 )一样主要受P的限制. ...
植物对盐渍逆境的适应. 生物学通报,
1
2002
... 植物要适应盐渍化的生境, 必须具备克服盐离子毒害(离子胁迫)、抵抗低水势(渗透胁迫)和平衡养分吸收的能力(Greeway & Munns, 1980 ; Munns & Termaat, 1986 ), 于是在生理上形成一系列抗盐特征—稀盐、泌盐、积盐和拒盐(赵可夫, 2002 ; Flowers & Colmer, 2008 ; Wang et al ., 2009 ; Zhang & Shi, 2013 ; Shabala et al ., 2014 ), 来减轻盐胁迫对组织产生的伤害, 同样所有的高等植物也都可以通过这些生理方面的调节及导入耐盐基因来增强自身对盐胁迫的耐受性(张金林等, 2015 ). ...


{kind=link}
{kind=link}