柽柳(Tamarix chinensis )属于典型的泌盐植物, 叶片和嫩枝可以将吸收于植物体内的盐分排出, 是黄河三角洲盐碱地植被恢复与生态重建的重要树种, 以其独特的生理和形态特点适应着盐碱地生境(Armas et al ., 2010 )。在黄河三角洲泥质海岸盐碱地, 地下水位浅且矿化度高, 水分和盐分是影响该区域柽柳空间分布及其生态位的主要因素(贺强等, 2008 ; Cui et al ., 2009 ), 土壤水分、盐分、地下水水位及其矿化度是表征盐碱地水盐环境的主要指标, 而地下水通过“饱和带-包气带-植被”间的垂向联系由点及面产生极为重要的生态环境效应(Singh et al ., 2010 ; Gou & Miller, 2014 ; 夏江宝等, 2015 ), 特别是地下水矿化度的不同, 是导致土壤盐分和水分变化的主要因素。地下水通过影响土壤水分和盐分变化, 进而影响与植物光合生产力形成有关的物质运输和能量平衡(Álvarez & Sánchez-Blanco, 2014 ), 从而对植物生长及其光合生理生态过程产生重要影响。植物光合特性和树干液流速率可较好地反映植物生长及其水分利用对逆境条件的适应能力(夏江宝等, 2014 ; Xia et al ., 2015 )。植物光合效率能够表征植物在一定环境下的健康状况和光合生产能力, 可用来预测和表征植物对逆境的适应性和可塑性(Zhang et al ., 2014 ; Sanchez et al ., 2015 )。树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 )。目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 )。从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨。研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 )。而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面。上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施。鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考。
1 材料和方法
1.1 试验材料
本研究在山东省黄河三角洲生态环境重点实验室的科研温室(37.38° N, 117.98° E)内进行, 栽植柽柳的土壤取自黄河三角洲的黄河滩地, 土壤质地为粉砂壤土, 其初始pH值为7.54, 平均含盐量为0.1%, 田间持水量为37.86%, 土壤容重为1.32 g·cm-3 。黄河三角洲泥质海岸带土壤为氯化物型盐土, NaCl占全盐含量的70%以上, 是土壤中最主要的盐分来源(吴志芬等, 1994 ; 曹建荣等, 2014 ), 因此, 为模拟设置该区域的地下水矿化度, 地下水以黄河三角洲NaCl为主的海盐自行配置, 主要依据天然水矿化度的数值分类标准, 称取各地下水矿化度模拟设置数值对应换算的海盐量, 溶解于特定容积的水桶中, 然后将配置好的不同矿化度水按照模拟设计水位分别浇灌于设计排列好的水桶中(图1 )。柽柳苗木栽植时采用三年生实生苗, 茎基部直径平均为(14.0 ± 0.5) mm, 苗高统一截杆至50 mm。
图1 不同地下水矿化度栽植柽柳的土柱模拟示意图和实景图。 A, 土柱模拟示意图。 B, 土柱试验照片。1, 柽柳; 2, 土壤; 3, 地下水; 4, 淹水区550 mm; 5, 潜水水位1200 mm; 6, 空隙层30 mm。
Fig. 1 Schematic diagram and photo of soil columns with Tamarix chinensis under different groundwater salinity. A, Schematic of soil column. B, Photo of experiment with soil columns. 1, Tamarix chinensis; 2, soil; 3, groundwater; 4, flooding depth at 550 mm; 5, groundwater level at 1200 mm; 6, interstice layer of 30 mm.
1.2 试验设计
黄河三角洲柽柳生长地带地下水位主要集中在300-2000 mm (马玉蕾等, 2013 ; 夏江宝等, 2015 ), 结合在科研温室内利于实验控制, 因此本研究以泥质海岸带柽柳大面积生长的地下水水位1200 mm为准, 模拟设置淡水、微咸水、咸水和盐水4种地下水矿化度, 其对应矿化度数值分别为0 g∙L-1 、3.00 g∙L-1 、8.00 g∙L-1 和20.00 g∙L-1 。2014年3月5日将柽柳栽植于直径为300 mm、高度为1780 mm的PVC圆管中, 其中PVC管实际淹水区为550 mm, 管底部设有纱网, 防止土壤渗漏。将PVC圆管放置于700 mm高, 桶底内径455 mm, 桶上口内径570 mm的水桶中, 为使地下水温度一致, 将水桶埋置于土壤中。不同地下水矿化度下栽植柽柳的土柱模拟示意图和实景图见图1 。为维持地下水水位及其矿化度的稳定性, 定期监测地下水盐度, 依次来补充所需盐分。柽柳苗木正常生长1月后对其进行不同地下水矿化度处理, 在此期间从地上部共浇水12.00 L, 分4次灌溉, 地下水矿化度处理后, 不再进行水分灌溉等措施。每种地下水矿化度处理下3个重复, 共计12个栽植柽柳的土壤柱体, 每土壤柱体栽植柽柳1株。在地下水矿化度处理80天后, 苗木平均高度为(67 ± 0.6) mm, 茎基部直径平均为(15.0 ± 0.7) mm, 6月27日开始柽柳苗木光合作用和树干液流的测定, 7月6日测定结束。
1.3 测定指标及方法
1.3.1 气体交换参数的测定
应用LI-6400XT便携式光合作用仪(LI-COR, Lincoln, USA)测定不同地下水矿化度处理下柽柳叶片的光响应曲线, 测定在晴朗天气的8:30-11:30进行, 使用缓冲瓶维持CO2 浓度的稳定性((390 ± 5) μmol∙mol-1 ), 设定人工光源的光合有效辐射(PAR )分别为: 1400、1200、1000、800、500、300、200、100、50、20、0 μmol∙m-2 ∙s-1 , 记录柽柳叶片的净光合速率(P n )、蒸腾速率(T r )、气孔导度(G s )、胞间CO2 浓度(C i )和大气CO2 浓度(C a )等参数。叶片瞬时水分利用效率(WUE )为P n 与T r 的比值, 即WUE = P n /T r (Nijs et al ., 1997 ); 气孔限制值(L s )为C i 与C a 的比值, 即L s = 1 - C i /C a (Wong et al ., 1979 )。同时在7:00- 19:00用LI-6400XT光合仪进行柽柳光合参数日动态的测定, 仪器自动记录T r 、G s 、PAR 和水汽压亏缺(VPD )等参数, 每2 h测定1次。以上测定每个处理测定3株苗木, 每株选取中上部生长健壮的成熟叶片2-3个, 在具体测定时, 采用不同处理、不同植株和叶片重复之间的交替测定法, 因此, 每株苗木依据测定的叶片数可测定2-3次, 以尽量避免时间波动对光合参数的影响。由于柽柳叶片形状不规则, 为得到精确的测量数据, 测定时需将标记的叶片平铺于整个测定叶室内, 同时参考邓雄(2003)和闫海龙(2010)等对不规则植物叶片光合参数的校正方法, 将观测的植物叶片剪下, 用扫描仪扫描后, 使用面积分析软件Delta-T Scan (CB50EJ, Cambridge, UK)计算实际的叶表面积, 按计算后的实际光合有效面积重新测算光合参数。
1.3.2 树干液流的测定
使用基于茎热平衡法的包裹式茎流计(Flow 32, Dynamax, Houston, USA)测定供试苗木树干液流动态。安装部位位于苗木茎干300-400 mm处。测定前, 用小刀将离地面300-400 mm处柽柳苗木的小枝条和萌芽去除, 用砂纸将茎干打磨光滑, 然后用游标卡尺测量样株所测部位的茎干直径为(7.0 ± 1.8) mm, 茎流探头以SGA5 (适合安装范围在茎干直径为5.0-7.0 mm)和SGB9 (适合安装范围在茎干直径为8.0-12.0 mm)为主。在打磨好的安装区涂抹G4 混合油, 将加热片安装于被测区, 用铝箔包裹安装探头所在的茎干部分, 并用胶带密封。数据采集通过该茎流计的数据采集器(Delta-T Logger, Cambridge, UK)完成, 每30 min记录一次。树干液流速率计算公式为F = Qf / (Cp ·d T ), 式中: F 为树干液流速率(g·h-1 ), Qf 为茎流带走的热量, Cp 为水的比热, d T 为竖向两热电偶电压和的平均值。具体计算过程可见刘鑫等(2014)。
1.3.3 土壤水盐含量的测定
对模拟土柱进行0-700 mm土壤样品采集, 随机对土柱样品进行3次取样。烘干法测定土壤质量含水量, 质量法测定土壤全盐含量, 水土体积比5:1。土壤溶液绝对浓度(C s , %) =土壤含盐量(占干土质量%)/土壤质量含水量(占干土质量%) × 100。
1.4 数据处理
将柽柳叶片光合光响应曲线使用直角双曲线修正模型拟合, 根据模型公式获取部分光响应特征参数光补偿点(LCP )、光饱和点(LSP )和表观光合量子效率(AQY )等。
直角双曲线修正模型公式(Ye, 2007 ):
式中: I 为光合有效辐射, 且均大于0, P n (I )为净光合速率, I c 为光补偿点, α 、β 、γ 是独立于I 的系数。其中α 为初始量子效率, β、γ 为修正系数。
数据处理依据3株供试苗木的重复测定进行指标均值求解, 数据的统计分析应用SPSS软件进行。对本研究中实验数据用单因素方差分析检验地下水矿化度的效应, 不同处理间的比较采用邓肯式新复极差法进行检验。
2 结果和分析
2.1 不同地下水矿化度下的土壤水盐特征
由表1 可知: 随地下水矿化度升高, 土壤水盐含量变化差异较大, 其中土壤含盐量和土壤溶液绝对浓度显著升高, 与淡水处理相比, 微咸水、咸水和盐水处理下土壤含盐量分别增加3.5倍、8.5倍和9.4倍, 差异显著(p < 0.05); 土壤溶液绝对浓度分别增加3.2倍、7.4倍和9.5倍, 差异显著(p < 0.05), 而地下水矿化度对土壤含水量无显著影响(p > 0.05)。
2.2 不同地下水矿化度对柽柳叶片光合光响应曲线的影响
由图2 可知: 随地下水矿化度增加, 柽柳叶片P n 先升高后降低, 咸水处理下最高(46.32 µmol∙m-2 ∙ s-1 ), 咸水处理下P n 光响应平均值也最大(25.90µmol∙m-2 ∙s-1 ), 淡水、微咸水、盐水处理下P n 光响应平均值分别比咸水处理降低44.1%、15.1%、62.6%。可见地下水矿化度过高会抑制柽柳叶片的光合作用, 适度的盐分胁迫可增强柽柳的光合能力。由表2 可知, 不同地下水矿化度下, 柽柳叶片光响应曲线参数差异显著(p < 0.05)。随地下水矿化度增大, 柽柳叶片AQY 、P nmax 先升高后降低, 各处理间差异显著(p < 0.05), 咸水处理下两者均达最高, 而盐水处理下均最低, 其中淡水、微咸水和盐水处理下, AQY 分别比咸水处理降低59.6%、18.3%和65.4%; P nmax 分别比咸水处理降低44.3%、16.6%和60.7%, 表明咸水处理下, 柽柳叶片利用弱光能力较弱, 但光合能力较强, 而盐水处理严重抑制了柽柳的最大光合能力。随地下水矿化度增加, 柽柳叶片R d 先升高后降低, 咸水处理下达最高, 其中, 淡水、微咸水和盐水处理下R d 分别比咸水处理降低92.0%、59.5%和35.0%。可见, 淡水处理下柽柳叶片呼吸作用较弱, 而咸水和盐水处理下呼吸作用旺盛。随地下水矿化度增大, 柽柳叶片LCP 逐渐升高且差异显著(p < 0.05), 而LSP 先升高后降低, 微咸水处理下LSP 达最高值。其中, 淡水、微咸水和咸水处理下LCP 分别比盐水处理降低68.2%、50.6%和16.0%; 淡水、咸水和盐水处理下LSP 分别比微咸水处理降低18.0%、12.7%和13.8%。
图2 不同地下水矿化度下柽柳叶片净光合速率的光响应曲线(平均值±标准误差, n = 3)。
Fig. 2 Light response curves of net photosynthetic rate in leaves of Tamarix chinensis under different groundwater salinities (mean ± SE, n = 3).
2.3 不同地下水矿化度对不同光强度下柽柳叶片蒸腾速率的影响
由图3 可知: 不同地下水矿化度下, 柽柳叶片T r 的光响应曲线差异显著, 其中在淡水和盐水处理下, T r 对PAR 响应不敏感, 较强的PAR 并没有导致T r 显著增加。但不同地下水矿化度下T r 差别较大, 随地下水矿化度增加, T r 先升高后降低, 咸水处理下达最高(15.80 mmol∙m-2 ∙s-1 ), 淡水、微咸水和盐水处理下T r 最大值分别比咸水处理降低59.3%、11.1%和49.9%。咸水处理下T r 光响应平均值也达最高(12.50 mmol∙m-2 ∙s-1 ), 淡水、微咸水和盐水处理下T r 光响应平均值分别比咸水处理降低55.4%、16.0%和50.2%。可见地下水矿化度过高或过低会抑制柽柳叶片的蒸腾作用, 适度的盐分胁迫会增强其蒸腾能力。
图3 同地下水矿化度下柽柳叶片蒸腾速率的光响应(平均值±标准误差, n = 3)。
Fig. 3 Light response curves of transpiration rate in leaves of Tamarix chinensis under different groundwater salinities (mean ± SE, n = 3).
2.4 不同地下水矿化度对不同光强度下柽柳叶片水分利用效率的影响
由图4 可知, 不同地下水矿化度下, 柽柳叶片WUE 的光响应过程类似。低光强(PAR ≤ 200 、µmol∙m-2 ∙s-1 )时, 随PAR 增强, WUE 上升较快, 达到光饱和点(1000-1400 µmol∙m-2 ∙s-1 )后, 随光强增强WUE 变化较小。不同地下水矿化度下WUE 差异显著(p < 0.05), 淡水处理下WUE 最高; 随地下水矿化度增加, WUE 逐渐降低, 微咸水、咸水和盐水处理下WUE 最大值分别比淡水处理(23.99 µmol∙mmol-1 )降低26.1%、27.8%和43.4%, WUE 光响应平均值分别比淡水处理(2.40 µmol∙mmol-1 )降低25.0%、29.2%和41.7%。
图4 不同地下水矿化度下柽柳叶片水分利用效率的光响应(平均值±标准误差, n = 3)。
Fig. 4 Light response curves of water use efficiency in leaves of Tamarix chinensis under different groundwater salinities (mean ± SE, n = 3).
2.5 不同地下水矿化度对不同光强度下柽柳叶片的气孔导度、胞间CO2 浓度和气孔限制值的影响
由图5 可知, 不同地下水矿化度下柽柳叶片G s 、C i 和L s 的光响应过程差异显著。盐水处理下, 柽柳G s 随PAR 增加变化比较平稳, 说明盐分胁迫对柽柳生长造成了一定的影响, 柽柳叶片气孔基本上失去了调节作用, 导致G s 对PAR 的变化响应不敏感。咸水至淡水处理下, 随地下水矿化度降低, 柽柳叶片P n 下降, G s 下降, C i 上升, L s 下降; 咸水至盐水处理下, 随地下水矿化度增加, 柽柳叶片P n 下降, G s 下降, C i 下降, L s 上升。依据气孔限制理论(Farquhar & Sharkey, 1982 ), 咸水至淡水处理下柽柳叶片P n 下降以非气孔限制为主, 咸水至盐水处理下柽柳叶片P n 下降以气孔限制为主。
图5 不同地下水矿化度下柽柳叶片气孔导度(A)、胞间CO2 浓度(B)和气孔限制值(C)的光响应(平均值±标准误差, n = 3)。
Fig. 5 Light response curves of stomatal conductance (A), intercellular CO2 concentration (B), stomatal limitation (C) in leaves of Tamarix chinensis under different groundwater salinities (mean ± SE, n = 3).
2.6 不同地下水矿化度对柽柳苗木蒸腾速率和液流速率日动态的影响
由图6 可知, 不同地下水矿化度下PAR 和VPD 日变化均呈单峰型, 分别在11:00-13:00和15:00达到峰值。不同地下水矿化度下, 随PAR 和VPD 的先升高后降低, 柽柳叶片T r 和树干液流速率总体也呈先升高后降低趋势, 但咸水和微咸水处理下T r 和液流速率呈双峰曲线, 在13:00-15:00左右出现“午休”, 且受PAR 和VPD 变化影响较大; 而淡水和盐水处理下T r 和液流速率日变化呈单峰曲线且变化相对平缓, 但峰值出现时刻有较大差异。由图6 C可知, 柽柳树干昼夜液流速率差异较大, 白天液流速率明显高于夜间。不同地下水矿化度下, 柽柳在0:00-6:00液流速率较低, 6:00-18:00液流速率上升较快, 随后液流速率保持平稳变化趋势, 即树干液流变化微弱, 夜间(19:30-5:30)柽柳仍可进行微弱的液流活动。随地下水矿化度升高, 日液流时间先延长后缩短, 淡水和盐水处理下, 日液流速率变化较小且启动时间延后至6:30, 结束时间提前至19:00, 日液流时间明显缩短; 微咸水和咸水处理下, 日液流速率变化较大且启动时间提前至6:00, 结束时间延后至19:30, 日液流时间明显延长。随地下水矿化度升高, 柽柳叶片T r 和液流速率均表现为先升高后降低, 咸水处理下两者均达最高, 其次是微咸水和盐水, 而淡水处理最低, 其中淡水、微咸水和盐水处理下日液流速率平均值分别比咸水处理(16.96 g·h-1 )降低61.8%、13.1%和41.9%。
图6 观测期(7月3-5日)不同地下水矿化度下柽柳光合有效辐射、水气压亏缺(A)、蒸腾速率(B)和液流速率(C)的平均日动态(平均值±标准误差, n = 3)。
Fig. 6 The mean daily change of photosynthetically available radiation (PAR ), vapor pressure deficit (VPD ) (A), transpiration rate (B) and stem sap flow rate (C) in Tamarix chinensis under different groundwater salinities during study period on 3 to 5, July (mean ± SE, n = 3).
3 讨论
3.1 地下水矿化度对柽柳光合参数的影响
水分是盐分运移的载体, 潜水蒸发作用下, 浅埋深地下水通过毛管作用进入包气带土壤层, 致使土壤水盐发生变化, 进而影响植物根系生长和叶片的光合作用(宫兆宁等, 2006 ; Gou & Miller, 2014 )。本研究发现相同潜水水位下, 土壤盐分和溶液绝对浓度差异较大成为影响柽柳苗木光合作用的主要因素。王鹏等(2012)研究发现, 多枝柽柳幼苗P n 随地下水矿化度增大而升高, 咸水处理下P n 最高, 光照生态幅最宽, 微咸水处理下AQY 和R d 最高; 而本研究发现, 随地下水矿化度增大, 柽柳P n 先升高后降低, 咸水处理下P n 、P nmax 、AQY 和R d 均达到最大值, 微咸水处理下光照生态幅最宽。王伟华等(2009)研究发现, 多枝柽柳P n 日平均值随盐水浓度的增加先升高后降低, 盐水处理下P n 日平均值达最高。可见, 淡水处理下柽柳并非有最好的光合性能, 而适度提高地下水至咸水矿化度会促进柽柳苗木的光合特性, 应该与盐生植物柽柳的泌盐生理特性和较强的耐盐性有关, 这可能是柽柳长期适应盐碱地生境的一种竞争策略。地下水矿化度的增加使土壤含盐量增加, 柽柳苗木通过增强叶片的光能转化效率、R d 和光照生态幅来提高光合作用以适应盐胁迫, 并且抗氧化物酶系统可清除体内因盐分过多而产生的活性氧, 保持体内活性氧的动态平衡(苏华等, 2012 ; 朱金方等, 2012 , 2015 ), 这可能是咸水处理下柽柳光合能力显著提高的主要原因。柽柳的泌盐特性通过泌盐腺将体内过多的Na+ 、Cl- 以盐粒结晶的形式排出体外, 同时还将其他有害微量元素进行离子区隔化, 可溶性糖、脯氨酸等可维持细胞渗透压, 能使柽柳在高盐度环境下生长(Scholander et al ., 1996 ; Anderson et al ., 2005 )。但盐水处理下, 土壤含盐量和溶液绝对浓度均达最高, 致使土壤渗透势升高, 根系吸水困难, 导致柽柳AQY 、R d 和P nmax 降低, 光照生态幅变窄。李紫薇等(2014)也发现, 重度盐分胁迫使蒺藜苜蓿(Medicago truncatula )叶片光能转化效率降低, AQY 降低进而导致P nmax 、LSP 降低, LCP 升高, 光照生态幅变窄, R d 降低, 本研究结果与之类似。这主要是因为柽柳对Na+ 、Cl- 的吸收, 排斥了对另一些营养元素的吸收, 幼苗生长受到威胁; 同时重度盐分胁迫促进了叶绿素酶对叶绿素的分解使叶绿素合成系统受到破坏, 活性氧自由基超过超氧化物歧化酶清除量, 细胞受损严重(朱金方等, 2012 , 2015 ), 渗透调节物质的功能显著降低(朱金方等, 2013 )。
气孔限制和非气孔限制是抑制光合作用下降的主要原因(Farquhar & Sharkey, 1982 )。王伟华等(2009)研究发现, 盐水浓度4 g∙L-1 处理下多枝柽柳光合作用受到促进, 其他盐水浓度(12、20 和28 g∙L-1 )处理下, 多枝柽柳植株光合能力下降受气孔限制影响, 并且灌溉盐水的浓度越大气孔限制越强烈。本研究也发现类似规律, 在盐水和淡水处理下, 柽柳P n 下降的原因分别是气孔限制和非气孔限制。但也有研究发现, 随地下水矿化度增大, 多枝柽柳幼苗P n 下降受非气孔限制影响(王鹏等, 2012 )。可见, 地下水水位及其矿化度以及土壤介质内盐胁迫处理方式不同时, 随盐分胁迫的增加, 柽柳叶片光合作用下降的气孔限制机理有一定差异。淡水至咸水处理下, 土壤含盐量和溶液绝对浓度分别为0.11%-1.05%, 0.64%-5.38%, 柽柳的泌盐特性可以缓解盐分胁迫对柽柳生长的影响; 咸水至盐水处理下, 土壤含盐量和溶液绝对浓度分别为1.05%- 1.17%, 5.38%-6.70%, 过高的盐胁迫降低了柽柳的光合作用, 柽柳为保证自身水分利用和避免光合机构受损而关闭气孔。
3.2 地下水矿化度对柽柳蒸腾耗水和水分利用效率的影响
树干液流速率是反映植物液流瞬时耗水规律的主要参数, 不但能够反映植物生理用水对环境因子的响应过程和规律, 也是确定树体储存水对蒸腾耗水贡献程度的主要指标(金鹰等, 2011 ; Wang et al ., 2012 )。光照和VPD 是影响树木蒸腾耗水的主要驱动因子(倪广艳等, 2015 ; 徐世琴等, 2015 ), 相关研究发现, 荷木(Schima superba )蒸腾在干、湿季均与PAR 和VPD 显著正相关(倪广艳等, 2015 ); 梭梭(Haloxylon ammodendron )液流密度日变化过程受PAR 、温度和VPD 的共同影响呈多峰特征(徐世琴等, 2015 )。本研究发现, 地下水矿化度会显著影响柽柳树干液流速率的日动态, 柽柳T r 和液流速率日动态随PAR 和VPD 的变化呈现单峰或双峰特征, 在咸水和微咸水处理下, 柽柳叶片气孔调节和蒸腾耗水受光照、空气温湿度等环境因子的协同作用影响较大, 通过“午休”有效调节了光合生理过程, 利于其对水盐的有效利用; 而淡水和盐水处理下, 环境因子的协同效应对其蒸腾耗水过程影响相对较小, 气孔调节相对较弱。可见随地下水矿化度的不同, 影响柽柳蒸腾耗水日动态的诸多环境因子的协同效应有较大差异。在土壤盐分为1.05%的咸水矿化度下, 柽柳T r 和日液流量最大, 而地下水矿化度过高或过低均会导致柽柳树干液流速率日变幅和日耗水量降低, 适度提高地下水矿化度会增加柽柳树干的液流速率和日作用时间。已有研究表明: 柽柳蒸腾速率随盐胁迫增加而降低(米文精等, 2011 ); 微咸水和咸水处理下, 柽柳T r 高于淡水处理(王鹏等, 2012 )。可见, 地下水矿化度与土壤盐分胁迫可显著影响树木耗水特性, 适度提高地下水矿化度可增强柽柳的蒸腾生理活性, 这可能是因为作为泌盐植物的柽柳, 适度土壤盐分起到类似营养盐的作用, 在水分不受限制的条件下, 盐分增高导致其蒸腾耗水有升高趋势。但随咸水至盐水处理下盐分渗透胁迫增强, 进入植物体内的水分减少, 蒸腾作用和液流速率显著降低。过高的地下水矿化度下, 柽柳苗木表现出通过关闭气孔降低蒸腾耗水来维持自身水分有效利用的生理策略。植物能否较好地协调碳同化与水分耗散之间的关系是辨识植物生长适应逆境胁迫的主要方法, 水分利用效率是判定植物生长适应逆境生存与否的关键因子(曹生奎等, 2009 )。本研究发现: 淡水和盐水处理下, 柽柳WUE 分别达最高和最低, 地下水矿化度的增加可显著降低柽柳WUE, 但地下水矿化度对柽柳WUE 的影响不如P n 和T r 明显, 特别是咸水和微咸水处理下, 柽柳WUE 无显著差异(p > 0.05), 这可能是柽柳通过调节自身生理状况以适应不同地下水矿化度和土壤盐分环境的水分利用策略。而已有研究表明: 柽柳WUE 随盐胁迫的加重有所增加(刘玉娟等, 2015 ); 咸水处理下多枝柽柳WUE 始终大于微咸水和淡水处理(王鹏等, 2012 )。可见, 随盐分处理方式的不同, 柽柳WUE 对盐胁迫引起的渗透胁迫表现出一定差异, 这主要与柽柳P n 和T r 对盐胁迫的响应不一致有关。在微咸水和咸水处理下, 柽柳P n 、T r 和液流活动均保持在较高状态, 生理活动旺盛, 柽柳通过调节光合特性和日液流量的变化来适应盐胁迫, 能保持高的水分利用效率; 盐水处理下, 柽柳P n 最低, 但蒸腾耗水并非最低, 导致WUE 最低; 淡水处理下由于T r 对地下水矿化度响应更加敏感, T r 下降幅度高于P n , 导致WUE 最高, 但此时P n 和T r 均较低, 液流活动微弱, 易导致蒸腾耗水过低而使叶片温度升高, 长时间的这种生理过程对植物正常的生理活动不利。
综上所述, 地下水矿化度可显著影响土壤盐分, 进而影响柽柳叶片的光合特性和树干液流量。柽柳苗木对盐胁迫适应能力较强, 适度提高地下水矿化度可显著增强柽柳的光合能力、耗水量和光能利用率, 但降低了其水分利用效率。淡水和盐水处理下柽柳叶片P n 下降分别受非气孔限制和气孔限制的影响。咸水处理下柽柳苗木可维持最高的光合效率和较高的水分利用效率, 更有利于柽柳苗木的生长。盐分胁迫下柽柳叶片光合特性和树木耗水能力表现出较高的可塑性, 在泥质海岸带对地下咸水矿化度的适应能力最好, 在低于咸水矿化度的地下水浅埋区具有较强的潜在适应能力, 这对于柽柳在黄河三角洲泥质海岸带的生存和竞争生长具有重要意义。但因本研究为在科研温室内实施的单一地下水矿化度处理的模拟控制试验, 特别是对温室内的温度和湿度等微环境因子进行了控制, 与野外实际生境下的天气变化有较大差异, 而柽柳光合和耗水特征在受地下水矿化度影响的同时, 可能与其他因子存在交互效应, 尤其是柽柳蒸腾耗水日动态的变化受外界环境因子的综合影响较大。因此, 在下一步的研究中, 需在野外选取不同地下水水位及其矿化度生境, 深入分析典型季节柽柳光合效率和蒸腾耗水变化及其对环境因子的响应规律。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Álvarez S Sánchez-Blanco MJ 2014 ). Long-term effect of salinity on plant quality, water relations, photosynthetic parameters and iron distribution in
Callistemon citrinus . Plant Biology 16 , 757 -764 .
[本文引用: 1]
[2]
An LS Zhao QS Ye SY Liu GQ Ding XG 2011 ). Water-salt interactions factors and vegetation effects in the groundwater ecosystem in Yellow River Delta
.Advances in Water Science 22 , 689 -695 .(in Chinese with English abstract) [安乐生 , 赵全生 , 叶思源 , 刘贯群 , 丁喜桂 (2011 ). 黄河三角洲地下水关键水盐因子及其植被效应. 水科学进展,
22 , 689 -695 .]
[本文引用: 1]
[3]
Anderson GL Garruthers RI Ge SK Gong P 2005 ). Monitoring of invasive
Tamarix distribution and effects of biological control with airborne hyperspectral remote sensing. International Journal of Remote Sensing 26 , 2487 -2489 .
[本文引用: 1]
[4]
Armas C Padilla FM Pugnaire FI Jackson RB 2010 ). Hydraulic lift and tolerance to salinity of semiarid species: Consequences for species interactions
.Oecologia 162 , 11 -21 .
[本文引用: 1]
[5]
Cao D Shi FC Takayoshi K Lu ZH Sun JK 2014 ). Halophyte plant communities affecting enzyme activity and microbes in saline soils of the Yellow River Delta in China
. Clean —Soil, Air,Water 42 , 1433 -1440 .
[本文引用: 1]
[6]
Cao JR Xü YX Yü HJ Huang C 2014 ). Research on the variation and evolution of groundwater chemistry in the Yellow River Delta
. Marine Sciences 38 , 78 -85 .(in Chinese with English abstract) [曹建荣 , 徐永兴 , 于洪军 , 黄翀 (2014 ). 黄河三角洲浅层地下水化学特征与演化. 海洋科学,
38 , 78 -85 .]
[本文引用: 1]
[7]
Cao SK Feng Q Si JH Chang ZQ Zhuo MC Xi HY Su YH 2009 ). Summary on the plant water use efficiency at leaf lever
.Acta Ecological Sinica 29 , 3882 -3892 .(in Chinese with English abstract) [曹生奎 , 冯起 , 司建华 , 常宗强 , 卓玛错 , 席海洋 , 苏永红 (2009 ). 植物叶片水分利用效率研究综述. 生态学报,
29 , 3882 -3892 .]
[本文引用: 1]
[8]
Chen YP Chen YN Xu CC Li WH 2011 ). Photosynthesis and water use efficiency of
Populus euphratica in response to changing groundwater depth and CO2 concentration. Environmental Earth Sciences 62 , 119 -125 .
[本文引用: 2]
[9]
Cui BS Tang N Zhao XS Bai JH 2009 ). A management- oriented valuation method to determine ecological water requirement for wetlands in Yellow River Delta of China
.Journal for Nature Conservation 17 , 129 -141 .
[本文引用: 1]
[10]
Deng X Li XM Zhang XM Ye WH Foezki A Runge M 2003 ). Studies on gas exchange of
Tamarix ramosissima Lbd. Acta Ecologica Sinica 23 , 180 -187 .(in Chinese with English abstract) [邓雄 , 李小明 , 张希明 , 叶万辉 , Andrea Foezki Michael Runge 2003 ). 多枝柽柳气体交换特性研究. 生态学报,
23 , 180 -187 .]
[11]
Farquhar GD Sharkey TD 1982 ). Stomatal conductance and photosynthesis
.Annual Review of Plant Physiology 33 , 317 -345 .
[本文引用: 2]
[12]
Glenn EP Nelson SG Ambrose B Martinez K Soliz D 2012 ). Comparison of salinity tolerance of three
Atriplex spp. in well-watered and drying soil. Environmental and Experimental Botany 83 , 62 -72 .
[本文引用: 1]
[13]
Gong ZN Gong HL Deng W Zhao WJ 2006 ). An overview of water movement in groundwater-soil-plant-atmosphere continuum with shallow water table
.Journal of Agro-Environment Science 25 , 365 -373 .(in Chinese with English abstract) [宫兆宁 , 宮辉力 , 邓伟 , 赵文吉 (2006 ). 浅埋条件下地下水-土壤-植物-大气连续体中水分运移研究规律, 农业环境科学学报,
25 , 365 -373 .]
[本文引用: 1]
[14]
Gou S Miller G 2014 ). A groundwater-soil-plant-atmosphere continuum approach for modelling water stress, uptake, and hydraulic redistribution in phreatophytic vegetation
.Ecohydrology 7 , 1029 -1041 .
[本文引用: 2]
[15]
He Q Cui BS Zhao XS Fu HL 2008 ). Niches of plant species in wetlands of the Yellow River Delta under gradients of water table depth and soil salinity
.Chinese Journal of Applied Ecology 19 , 969 -975 .(in Chinese with English abstract) [贺强 , 崔宝山 , 赵欣胜 , 付华龄 (2008 ). 水、盐梯度下黄河三角洲湿地植物种的生态位. 应用生态学报,
19 , 969 -975 .]
[本文引用: 1]
[16]
He Q Cui BS Zhao XS Fu HL Xiong X Feng GH 2007 ). Vegetation distribution patterns to the gradients of water depth and soil salinity in wetlands of Yellow River Delta, China
.Wetland Science 5 , 208 -214 .(in Chinese with English abstract) [贺强 , 崔保山 , 赵欣胜 , 付华龄 , 雄熊 , 冯光海 (2007 ). 水盐梯度下黄河三角洲湿地植被空间分异规律的定量分析. 湿地科学,
5 , 208 -214 .]
[本文引用: 1]
[17]
Jin Y Wang CK Sang Y 2011 ). Contribution of stem water storage to daily transpiration of three temperate trees in northeastern China
.Chinese Journal of Plant Ecology 35 , 1310 -1317 .(in Chinese with English abstract) [金鹰 , 王传宽 , 桑英 (2011 ). 三种温带树种树干储存水对蒸腾的贡献. 植物生态学报,
35 , 1310 -1317 .]
[本文引用: 1]
[18]
Li J Yu B Zhao C Nowak RS Zhao Z Sheng Y Li J 2013 ). Physiological and morphological responses of
Tamarix ramosissima and Populus euphratica to altered groundwater availability. Tree Physiology 33 , 57 -68 .
[本文引用: 1]
[19]
Li ZW Ma TY Liang GT Guo J 2014 ). Photosynthesis in leaves of
Medicago truncatula under salt stress. Acta Botanica Boreali-Occidentalia Sinica 34 , 2070 -2077 .(in Chinese with English abstract) [李紫薇 , 马天意 ,梁国婷 , 国静 (2014 ). 蒺藜苜蓿叶片光合作用对盐胁迫的响应. 西北植物学报,
34 , 2070 -2077 .]
[20]
Liu X Zhang JC Zhuang JY Gu ZY Han C Wu YW 2014 ). Impact factors of sap flow of Chinese fir sapling and its contribution to transpiration of Chinese fir forest
.Bulletin of Soil and Water Conservation 34 (6 ), 73 -78 .(in Chinese with English abstract) [刘鑫 , 张金池 , 庄家尧 , 顾哲衍 , 韩诚 , 吴雁雯 (2014 ). 杉木幼树树干液流影响因子及其对杉木林蒸腾量的贡献. 水土保持通报,
34(6) , 73 -78 .]
[21]
Liu YJ He KN Wang LW Wang H Xu T Wang Y 2015 ). Effect of salt stress on diurnal variation of photosynthesis of
Nitraria tangutorum and Tamarix chinensis . Chinese Agricultural Science Bulletin 31 , 6 -12 .(in Chinese with English abstract) [刘玉娟 , 贺康宁 , 王伟路 , 王辉 , 徐特 , 王颖 (2015 ). 盐胁迫对柽柳和白刺光合日变化的影响. 中国农学通报,
31 , 6 -12 .]
[本文引用: 1]
[22]
Ma JX Chen YN Li WH Huang X Zhu CG Ma XD 2010 ). Response of sap flow in
Populus euphratica to changes in groundwater depth in the middle and lower reaches of the Tarim River of northwestern China. Chinese Journal of Plant Ecology 34 , 915 -923 .(in Chinese with English abstract) [马建新 , 陈亚宁 , 李卫红 , 黄湘 , 朱成刚 , 马晓东 (2010 ). 胡杨液流对地下水埋深变化的响应. 植物生态学报,
34 , 915 -923 .]
[本文引用: 1]
[23]
Ma YL Wang D Liu JM Wen XH Gao M Shao HB 2013 ). Relationships between typical vegetations, soil salinity, and groundwater depth in the Yellow River Delta of China
.Chinese Journal of Applied Ecology 24 , 2423 -2430 .(in Chinese with English abstract) [马玉蕾 , 王德 , 刘俊民 , 温小虎 , 高猛 , 邵宏波 (2013 ). 黄河三角洲典型植被与地下水埋深和土壤盐分的关系. 应用生态学报,
24 , 2423 -2430 .]
[本文引用: 1]
[24]
Mi WJ Liu KD Zhao YG Zheng CX Shi C Hao JQ Lü K 2011 ). Initial selection of plant species for ecological restoration in salinized soil in Datong Basin
.Journal of Beijing Forestry University 33 (1 ), 49 -54 .(in Chinese with English abstract) [米文精 , 刘克东 , 赵永刚 , 郑彩霞 , 时朝 , 郝建卿 , 吕坤 (2011 ). 大同盆地盐碱地生态修复利用植物的初步选择. 北京林业大学学报,
33(1) , 49 -54 .]
[本文引用: 1]
[25]
Ni GY Zhao P Zhu LW Niu JF Zhao XH Zeng XP 2015 ). Hydraulic responses of whole tree transpiration of
Schima superba to soil moisture in dry and wet seasons. Acta Ecologica Sinica 35 , 652 -662 .(in Chinese with English abstract) [倪广艳 , 赵平 , 朱丽薇 , 牛俊峰 , 赵秀华 , 曾小平 (2015 ). 荷木整树蒸腾对干湿季土壤水分的水力响应. 生态学报,
35 , 652 -662 .]
[本文引用: 2]
[26]
Nijs I Ferris R Blum H Hendrey G Impens I 1997 ). Stomatal regulation in a changing climate: A field study using free air temperature increase (FATI) and free air CO2 enrichment (FACE)
.Plant, Cell & Environment 20 , 1041 -1050 .
[本文引用: 1]
[27]
Qin J Dong WY He KN Yu Y Tan GD Han L Dong M Zhang YY Zhang D Li AZ Wang ZL 2010 ). NaCl salinity-induced changes in water status, iron contents and photosynthetic properties of
Shepherdia argentea Pursh) Nutt. seedings. Plant Soil Environment , 56 , 325 -332 .
[本文引用: 1]
[28]
Rong LS Liu GH Shu LC Song CY Huang C 2010 ). Study on ecologic water lever cover depth in Yellow River Delta
.Water Resource and Power 28 (6 ), 92 -95, 148 .(in Chinese with English abstract) [荣丽杉 , 刘高焕 , 束龙仓 , 宋创业 , 黄翀 (2010 ). 黄河三角洲地下水生态水位埋深研究. 水电能源科学,
28(6) , 92 -95, 148 .]
[本文引用: 1]
[29]
Sanchez E Scordia D Lino C Arias C Cosentino SL Nogues S 2015 ). Salinity and water stress effects on biomass production in different
Arundo donax L. Clones. Bioenergy Research 8 , 1461 -1479 .
[本文引用: 1]
[30]
Scholander PF Bradstreet ED Hammel HT Hemmingsen EA 1996 ). Sap concentrations in halophytes and some other plants
.Plant Physiology 41 , 529 -532 .
[本文引用: 1]
[31]
Singh V Brar MS Sharma P Malhi SS 2010 ). Arsenic in water, soil, and rice plants in the Indo-Gangetic plains of northwestern India
.Communication in Soil Science and Plant Analysis 41 , 1350 -1360 .
[本文引用: 1]
[32]
Su H Li YG Su BY Sun JX 2012 ). Effects of groundwater decline on photosynthetic characteristic and stress tolerance of
Ulmus pumila in Hunshandake Sandy Land, China. Chinese Journal of Plant Ecology 36 , 178 -186 .(in Chinese with English abstract) [苏华 , 李永庚 , 苏本营 , 孙建新 (2012 ). 地下水位下降对浑善达克沙地榆树光合及抗逆性的影响. 植物生态学报,
36 , 178 -186 .]
[本文引用: 2]
[33]
Wang H Zhao P Holscher D Wang Q Lu P Cai XA Zeng XP 2012 ). Nighttime sap flow of
Acacia mangium and its implications for nighttime transpiration and stem water storage. Journal of Plant Ecology 5 , 294 -304 .
[本文引用: 1]
[34]
Wang P Zhao CY Li J 2012 ). Effects of groundwater depth and mineralization degree on photosynthesis and growth of
Tamarix ramosissima seedlings. Bulletin of Soil and Water Conservation 32 (2 ), 84 -89 .(in Chinese with English abstract) [王鹏 , 赵成义 , 李君 (2012 ). 地下水埋深及矿化度对多枝柽柳幼苗光合特性及生长的影响. 水土保持通报,
32(2) , 84 -89 .]
[本文引用: 4]
[35]
Wang W Wang RQ Yuan YF Du N Guo WH 2011 ). Effects of salt and water stress on plant biomass and photosynthetic characteristics of
Tamarix (Tamarix chinensis L.) seedlings. Africa Journal of Biotechnical 10 , 17981 -17989 .
[本文引用: 1]
[36]
Wang WH Zhang XM Yan HL Ling SM Yang XL 2009 ). Effects of salt stress on photosynthetic and osmoregulation substances of
Tamarix ramosissima Ledeb. Arid Zone Research 26 , 561 -568 .(in Chinese with English abstract) [王伟华 , 张希明 , 闫海龙 , 梁少民 , 杨小林 (2009 ). 盐处理对多枝柽柳光合作用和渗调物质的影响. 干旱区研究,
26 , 561 -568 .]
[37]
Wang ZR Zhao GX Gao MX Jiang SQ Chang CY Jia JC 2015 ). Characteristics of soil water and salt spatial variation in the spring season in typical Yellow River Delta areas of Kenli County, China
.Journal of Agricultural Resource and Environment 32 (2 ), 154 -161 .(in Chinese with English abstract) [王卓然 , 赵庚兴 , 高明秀 , 姜曙千 , 常春艳 , 贾吉超 (2015 ). 黄河三角洲典型地区春季土壤水盐空间分异特征研究——以垦利县为例. 农业资源与环境学报,
32(2) , 154 -161 .]
[本文引用: 1]
[38]
Wong SC Cowan IR Farquhar GD 1979 ). Stomatal conductance correlates with photosynthetic capacity
.Nature 282 , 424 -426 .
[本文引用: 1]
[39]
Wu ZF Zhao SL Zhang XL 1994 ). Studies on interrelation between salt vegetation and soil salinity in the Yellow River Delta
.Acta Phytoecologica Sinica 18 , 184 -193 .(in Chinese with English abstract) [吴志芬 , 赵善伦 , 张学雷 (1994 ). 黄河三角洲盐生植被与土壤盐分的相关性研究. 植物生态学报,
18 , 184 -193 .]
[本文引用: 1]
[40]
Xia JB Zhang SY Guo J Kong QQ Zhang GC 2015 ). Critical effects of gas exchange parameters in
Tamarix chinensis on soil water and its relevant environmental factors on a shell ridge island in China’s Yellow River Delta. Ecological Engineering 76 , 36 -46 .
[本文引用: 1]
[41]
Xia JB Zhang SY Zhu LP Zhao ZG Zhao YY 2014 ). Responds characteristics of stem sap flow and leaf photosynthesis of
Ziziphus jujuba var. spinosus in responds to soil moisture in shell ridge island. Scientia Silvae Sinicae 50 (10 ), 24 -32 .(in Chinese with English abstract) [夏江宝 , 张淑勇 , 朱丽平 , 赵自国 , 赵艳云 (2014 ). 贝壳堤岛酸枣树干液流及光合参数对土壤水分的响应特征. 林业科学,
50(10) , 24 -32 .]
[本文引用: 3]
[42]
Xia JB Zhao XM Zhao ZG Chen YP Liu JH 2015 ). Migration characteristics of soil water and salt and their interaction under different groundwater levers
.Transactions of the Chinese Society of Agricultural Engineering 31 (15 ), 93 -100 .(in Chinese with English abstract) [夏江宝 , 赵西梅 , 赵自国 , 陈印平 , 刘俊华 (2015 ). 不同潜水埋深下土壤水盐运移特征及其交互效应. 农业工程学报,
31(15) , 93 -100 .]
[本文引用: 2]
[43]
Xu SQ Ji XB Jin BW 2015 ). Dynamics and responses of sap flow of typical sand binding plants
Haloxylon ammodendron to environmental variables. Chinese Journal of Plant Ecology 39 , 890 -900 .(in Chinese with English abstract) [徐世琴 , 吉喜斌 , 金博文 (2015 ). 典型固沙植物梭梭生长季蒸腾变化及其对环境因子的响应. 植物生态学报,
39 , 890 -900 .]
[本文引用: 2]
[44]
Yan HL Zhang XM Xu H Liu Y 2010 ). Photosynthetic characteristics responds of three plants to drought stress in Tarim Desert Highway shelterbelt
.Acta Ecologica Sinica 30 , 2519 -2528 .(in Chinese with English abstract) [闫海龙 , 张希明 , 许浩 , 刘英 (2010 ). 塔里木沙漠公路防护林3种植物光合特性对干旱胁迫的响应. 生态学报,
30 , 2519 -2528 .]
[45]
Ye ZP 2007 ). A new model for relationship between irradiance and the rate of photosynthesis in
Oryza sativa . Photosynthetica 45 , 637 -640 .
[本文引用: 1]
[46]
Zhang SY Xia JB Zhang GC Zhao ZG Zhao YY Shao HB Sun JK Shao CY Liu Q 2014 ). Threshold effects of photosynthetic efficiency parameters of wild
jujube in response to soil moisture variation on shell beach ridges, Shandong, China. Plant Biosystems 148 , 140 -149 .
[本文引用: 1]
[47]
Zhao CY Si JH Feng Q Yu TF Li W 2015 ). Research progress and prospect of stem sap flow
.Journal of Northwest Forestry University 30 (5 ), 98 -105 .(in Chinese with English abstract) [赵春彦 , 司建华 , 冯起 , 鱼腾飞 , 李炜 (2015 ). 树干液流研究进展与展望. 西北林学院学报,
30(5) , 98 -105 .]
[本文引用: 1]
[48]
Zhao Y Zhao CY Xu ZL Liu YY Wang Y Wang C Peng HH Zheng XL 2012 ). Physiological responses of
Populus euphratica Oliv. to groundwater table variation in the lower reaches of Heihe River, Northwest China. Journal of Arid Land 4 , 281 -291 .
[本文引用: 1]
[49]
Zhu JF Liu JT Lu ZH Xia JB Liu HN Jin Y 2015 ). Effects of salt stress on physiological characteristics of
Tamarix chinensis Lour. seedlings. Acta Ecologica Sinica 35 , 5141 -5146 .(in Chinese with English abstract) [朱金方 , 刘京涛 , 陆兆华 , 夏江宝 , 柳海宁 , 金悦 (2015 ). 盐胁迫对中国柽柳幼苗生理特性的影响. 生态学报,
35 , 5141 -5146 .]
[本文引用: 2]
[50]
Zhu JF Lu ZH Xia JB Chen X Zhang M Liu JT 2013 ). Changes of osmotic adjusting substances in leaves of
Tamarix chinensis seedlings under salt and drought stress. Acta Botanica Boreali-Occidentalia Sinica 33 , 357 -363 .(in Chinese with English abstract) [朱金方 , 陆兆华 , 夏江宝 , 陈曦 , 张萌 , 刘京涛 (2013 ). 盐旱交叉胁迫对柽柳幼苗渗透调节物质含量的影响. 西北植物学报,
33 , 357 -363 .]
[本文引用: 1]
[51]
Zhu JF Xia JB Lu ZH Liu JT Sun JK 2012 ). Growth, physiological and biochemical characteristics of
Tamarix chinensis seedlings under salt-drought intercross stress. Acta Botanica Boreali-Occidentalia Sinica 32 , 124 -130 .(in Chinese with English abstract) [朱金方 , 夏江宝 , 陆兆华 , 刘京涛 , 孙景宽 (2012 ). 盐旱交叉胁迫对柽柳幼苗生长及生理生化特性的影响. 西北植物学报,
32 , 124 -130 .]
[本文引用: 2]
Long-term effect of salinity on plant quality, water relations, photosynthetic parameters and iron distribution in
1
2014
... 柽柳(Tamarix chinensis )属于典型的泌盐植物, 叶片和嫩枝可以将吸收于植物体内的盐分排出, 是黄河三角洲盐碱地植被恢复与生态重建的重要树种, 以其独特的生理和形态特点适应着盐碱地生境(Armas et al ., 2010 ).在黄河三角洲泥质海岸盐碱地, 地下水位浅且矿化度高, 水分和盐分是影响该区域柽柳空间分布及其生态位的主要因素(贺强等, 2008 ; Cui et al ., 2009 ), 土壤水分、盐分、地下水水位及其矿化度是表征盐碱地水盐环境的主要指标, 而地下水通过“饱和带-包气带-植被”间的垂向联系由点及面产生极为重要的生态环境效应(Singh et al ., 2010 ; Gou & Miller, 2014 ; 夏江宝等, 2015 ), 特别是地下水矿化度的不同, 是导致土壤盐分和水分变化的主要因素.地下水通过影响土壤水分和盐分变化, 进而影响与植物光合生产力形成有关的物质运输和能量平衡(Álvarez & Sánchez-Blanco, 2014 ), 从而对植物生长及其光合生理生态过程产生重要影响.植物光合特性和树干液流速率可较好地反映植物生长及其水分利用对逆境条件的适应能力(夏江宝等, 2014 ; Xia et al ., 2015 ).植物光合效率能够表征植物在一定环境下的健康状况和光合生产能力, 可用来预测和表征植物对逆境的适应性和可塑性(Zhang et al ., 2014 ; Sanchez et al ., 2015 ).树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 ).目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
黄河三角洲地下水关键水盐因子及其植被效应. 水科学进展,
1
2011
... 柽柳(Tamarix chinensis )属于典型的泌盐植物, 叶片和嫩枝可以将吸收于植物体内的盐分排出, 是黄河三角洲盐碱地植被恢复与生态重建的重要树种, 以其独特的生理和形态特点适应着盐碱地生境(Armas et al ., 2010 ).在黄河三角洲泥质海岸盐碱地, 地下水位浅且矿化度高, 水分和盐分是影响该区域柽柳空间分布及其生态位的主要因素(贺强等, 2008 ; Cui et al ., 2009 ), 土壤水分、盐分、地下水水位及其矿化度是表征盐碱地水盐环境的主要指标, 而地下水通过“饱和带-包气带-植被”间的垂向联系由点及面产生极为重要的生态环境效应(Singh et al ., 2010 ; Gou & Miller, 2014 ; 夏江宝等, 2015 ), 特别是地下水矿化度的不同, 是导致土壤盐分和水分变化的主要因素.地下水通过影响土壤水分和盐分变化, 进而影响与植物光合生产力形成有关的物质运输和能量平衡(Álvarez & Sánchez-Blanco, 2014 ), 从而对植物生长及其光合生理生态过程产生重要影响.植物光合特性和树干液流速率可较好地反映植物生长及其水分利用对逆境条件的适应能力(夏江宝等, 2014 ; Xia et al ., 2015 ).植物光合效率能够表征植物在一定环境下的健康状况和光合生产能力, 可用来预测和表征植物对逆境的适应性和可塑性(Zhang et al ., 2014 ; Sanchez et al ., 2015 ).树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 ).目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
Monitoring of invasive
1
2005
... 水分是盐分运移的载体, 潜水蒸发作用下, 浅埋深地下水通过毛管作用进入包气带土壤层, 致使土壤水盐发生变化, 进而影响植物根系生长和叶片的光合作用(宫兆宁等, 2006 ; Gou & Miller, 2014 ).本研究发现相同潜水水位下, 土壤盐分和溶液绝对浓度差异较大成为影响柽柳苗木光合作用的主要因素.王鹏等(2012)研究发现, 多枝柽柳幼苗P n 随地下水矿化度增大而升高, 咸水处理下P n 最高, 光照生态幅最宽, 微咸水处理下AQY 和R d 最高; 而本研究发现, 随地下水矿化度增大, 柽柳P n 先升高后降低, 咸水处理下P n 、P nmax 、AQY 和R d 均达到最大值, 微咸水处理下光照生态幅最宽.王伟华等(2009)研究发现, 多枝柽柳P n 日平均值随盐水浓度的增加先升高后降低, 盐水处理下P n 日平均值达最高.可见, 淡水处理下柽柳并非有最好的光合性能, 而适度提高地下水至咸水矿化度会促进柽柳苗木的光合特性, 应该与盐生植物柽柳的泌盐生理特性和较强的耐盐性有关, 这可能是柽柳长期适应盐碱地生境的一种竞争策略.地下水矿化度的增加使土壤含盐量增加, 柽柳苗木通过增强叶片的光能转化效率、R d 和光照生态幅来提高光合作用以适应盐胁迫, 并且抗氧化物酶系统可清除体内因盐分过多而产生的活性氧, 保持体内活性氧的动态平衡(苏华等, 2012 ; 朱金方等, 2012 , 2015 ), 这可能是咸水处理下柽柳光合能力显著提高的主要原因.柽柳的泌盐特性通过泌盐腺将体内过多的Na+ 、Cl- 以盐粒结晶的形式排出体外, 同时还将其他有害微量元素进行离子区隔化, 可溶性糖、脯氨酸等可维持细胞渗透压, 能使柽柳在高盐度环境下生长(Scholander et al ., 1996 ; Anderson et al ., 2005 ).但盐水处理下, 土壤含盐量和溶液绝对浓度均达最高, 致使土壤渗透势升高, 根系吸水困难, 导致柽柳AQY 、R d 和P nmax 降低, 光照生态幅变窄.李紫薇等(2014)也发现, 重度盐分胁迫使蒺藜苜蓿(Medicago truncatula )叶片光能转化效率降低, AQY 降低进而导致P nmax 、LSP 降低, LCP 升高, 光照生态幅变窄, R d 降低, 本研究结果与之类似.这主要是因为柽柳对Na+ 、Cl- 的吸收, 排斥了对另一些营养元素的吸收, 幼苗生长受到威胁; 同时重度盐分胁迫促进了叶绿素酶对叶绿素的分解使叶绿素合成系统受到破坏, 活性氧自由基超过超氧化物歧化酶清除量, 细胞受损严重(朱金方等, 2012 , 2015 ), 渗透调节物质的功能显著降低(朱金方等, 2013 ). ...
Hydraulic lift and tolerance to salinity of semiarid species: Consequences for species interactions
1
2010
... 柽柳(Tamarix chinensis )属于典型的泌盐植物, 叶片和嫩枝可以将吸收于植物体内的盐分排出, 是黄河三角洲盐碱地植被恢复与生态重建的重要树种, 以其独特的生理和形态特点适应着盐碱地生境(Armas et al ., 2010 ).在黄河三角洲泥质海岸盐碱地, 地下水位浅且矿化度高, 水分和盐分是影响该区域柽柳空间分布及其生态位的主要因素(贺强等, 2008 ; Cui et al ., 2009 ), 土壤水分、盐分、地下水水位及其矿化度是表征盐碱地水盐环境的主要指标, 而地下水通过“饱和带-包气带-植被”间的垂向联系由点及面产生极为重要的生态环境效应(Singh et al ., 2010 ; Gou & Miller, 2014 ; 夏江宝等, 2015 ), 特别是地下水矿化度的不同, 是导致土壤盐分和水分变化的主要因素.地下水通过影响土壤水分和盐分变化, 进而影响与植物光合生产力形成有关的物质运输和能量平衡(Álvarez & Sánchez-Blanco, 2014 ), 从而对植物生长及其光合生理生态过程产生重要影响.植物光合特性和树干液流速率可较好地反映植物生长及其水分利用对逆境条件的适应能力(夏江宝等, 2014 ; Xia et al ., 2015 ).植物光合效率能够表征植物在一定环境下的健康状况和光合生产能力, 可用来预测和表征植物对逆境的适应性和可塑性(Zhang et al ., 2014 ; Sanchez et al ., 2015 ).树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 ).目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
Halophyte plant communities affecting enzyme activity and microbes in saline soils of the Yellow River Delta in China
1
2014
... 柽柳(Tamarix chinensis )属于典型的泌盐植物, 叶片和嫩枝可以将吸收于植物体内的盐分排出, 是黄河三角洲盐碱地植被恢复与生态重建的重要树种, 以其独特的生理和形态特点适应着盐碱地生境(Armas et al ., 2010 ).在黄河三角洲泥质海岸盐碱地, 地下水位浅且矿化度高, 水分和盐分是影响该区域柽柳空间分布及其生态位的主要因素(贺强等, 2008 ; Cui et al ., 2009 ), 土壤水分、盐分、地下水水位及其矿化度是表征盐碱地水盐环境的主要指标, 而地下水通过“饱和带-包气带-植被”间的垂向联系由点及面产生极为重要的生态环境效应(Singh et al ., 2010 ; Gou & Miller, 2014 ; 夏江宝等, 2015 ), 特别是地下水矿化度的不同, 是导致土壤盐分和水分变化的主要因素.地下水通过影响土壤水分和盐分变化, 进而影响与植物光合生产力形成有关的物质运输和能量平衡(Álvarez & Sánchez-Blanco, 2014 ), 从而对植物生长及其光合生理生态过程产生重要影响.植物光合特性和树干液流速率可较好地反映植物生长及其水分利用对逆境条件的适应能力(夏江宝等, 2014 ; Xia et al ., 2015 ).植物光合效率能够表征植物在一定环境下的健康状况和光合生产能力, 可用来预测和表征植物对逆境的适应性和可塑性(Zhang et al ., 2014 ; Sanchez et al ., 2015 ).树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 ).目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
黄河三角洲浅层地下水化学特征与演化. 海洋科学,
1
2014
... 本研究在山东省黄河三角洲生态环境重点实验室的科研温室(37.38° N, 117.98° E)内进行, 栽植柽柳的土壤取自黄河三角洲的黄河滩地, 土壤质地为粉砂壤土, 其初始pH值为7.54, 平均含盐量为0.1%, 田间持水量为37.86%, 土壤容重为1.32 g·cm-3 .黄河三角洲泥质海岸带土壤为氯化物型盐土, NaCl占全盐含量的70%以上, 是土壤中最主要的盐分来源(吴志芬等, 1994 ; 曹建荣等, 2014 ), 因此, 为模拟设置该区域的地下水矿化度, 地下水以黄河三角洲NaCl为主的海盐自行配置, 主要依据天然水矿化度的数值分类标准, 称取各地下水矿化度模拟设置数值对应换算的海盐量, 溶解于特定容积的水桶中, 然后将配置好的不同矿化度水按照模拟设计水位分别浇灌于设计排列好的水桶中(图1 ).柽柳苗木栽植时采用三年生实生苗, 茎基部直径平均为(14.0 ± 0.5) mm, 苗高统一截杆至50 mm. ...
植物叶片水分利用效率研究综述. 生态学报,
1
2009
... 树干液流速率是反映植物液流瞬时耗水规律的主要参数, 不但能够反映植物生理用水对环境因子的响应过程和规律, 也是确定树体储存水对蒸腾耗水贡献程度的主要指标(金鹰等, 2011 ; Wang et al ., 2012 ).光照和VPD 是影响树木蒸腾耗水的主要驱动因子(倪广艳等, 2015 ; 徐世琴等, 2015 ), 相关研究发现, 荷木(Schima superba )蒸腾在干、湿季均与PAR 和VPD 显著正相关(倪广艳等, 2015 ); 梭梭(Haloxylon ammodendron )液流密度日变化过程受PAR 、温度和VPD 的共同影响呈多峰特征(徐世琴等, 2015 ).本研究发现, 地下水矿化度会显著影响柽柳树干液流速率的日动态, 柽柳T r 和液流速率日动态随PAR 和VPD 的变化呈现单峰或双峰特征, 在咸水和微咸水处理下, 柽柳叶片气孔调节和蒸腾耗水受光照、空气温湿度等环境因子的协同作用影响较大, 通过“午休”有效调节了光合生理过程, 利于其对水盐的有效利用; 而淡水和盐水处理下, 环境因子的协同效应对其蒸腾耗水过程影响相对较小, 气孔调节相对较弱.可见随地下水矿化度的不同, 影响柽柳蒸腾耗水日动态的诸多环境因子的协同效应有较大差异.在土壤盐分为1.05%的咸水矿化度下, 柽柳T r 和日液流量最大, 而地下水矿化度过高或过低均会导致柽柳树干液流速率日变幅和日耗水量降低, 适度提高地下水矿化度会增加柽柳树干的液流速率和日作用时间.已有研究表明: 柽柳蒸腾速率随盐胁迫增加而降低(米文精等, 2011 ); 微咸水和咸水处理下, 柽柳T r 高于淡水处理(王鹏等, 2012 ).可见, 地下水矿化度与土壤盐分胁迫可显著影响树木耗水特性, 适度提高地下水矿化度可增强柽柳的蒸腾生理活性, 这可能是因为作为泌盐植物的柽柳, 适度土壤盐分起到类似营养盐的作用, 在水分不受限制的条件下, 盐分增高导致其蒸腾耗水有升高趋势.但随咸水至盐水处理下盐分渗透胁迫增强, 进入植物体内的水分减少, 蒸腾作用和液流速率显著降低.过高的地下水矿化度下, 柽柳苗木表现出通过关闭气孔降低蒸腾耗水来维持自身水分有效利用的生理策略.植物能否较好地协调碳同化与水分耗散之间的关系是辨识植物生长适应逆境胁迫的主要方法, 水分利用效率是判定植物生长适应逆境生存与否的关键因子(曹生奎等, 2009 ).本研究发现: 淡水和盐水处理下, 柽柳WUE 分别达最高和最低, 地下水矿化度的增加可显著降低柽柳WUE, 但地下水矿化度对柽柳WUE 的影响不如P n 和T r 明显, 特别是咸水和微咸水处理下, 柽柳WUE 无显著差异(p > 0.05), 这可能是柽柳通过调节自身生理状况以适应不同地下水矿化度和土壤盐分环境的水分利用策略.而已有研究表明: 柽柳WUE 随盐胁迫的加重有所增加(刘玉娟等, 2015 ); 咸水处理下多枝柽柳WUE 始终大于微咸水和淡水处理(王鹏等, 2012 ).可见, 随盐分处理方式的不同, 柽柳WUE 对盐胁迫引起的渗透胁迫表现出一定差异, 这主要与柽柳P n 和T r 对盐胁迫的响应不一致有关.在微咸水和咸水处理下, 柽柳P n 、T r 和液流活动均保持在较高状态, 生理活动旺盛, 柽柳通过调节光合特性和日液流量的变化来适应盐胁迫, 能保持高的水分利用效率; 盐水处理下, 柽柳P n 最低, 但蒸腾耗水并非最低, 导致WUE 最低; 淡水处理下由于T r 对地下水矿化度响应更加敏感, T r 下降幅度高于P n , 导致WUE 最高, 但此时P n 和T r 均较低, 液流活动微弱, 易导致蒸腾耗水过低而使叶片温度升高, 长时间的这种生理过程对植物正常的生理活动不利. ...
Photosynthesis and water use efficiency of
2
2011
... 柽柳(Tamarix chinensis )属于典型的泌盐植物, 叶片和嫩枝可以将吸收于植物体内的盐分排出, 是黄河三角洲盐碱地植被恢复与生态重建的重要树种, 以其独特的生理和形态特点适应着盐碱地生境(Armas et al ., 2010 ).在黄河三角洲泥质海岸盐碱地, 地下水位浅且矿化度高, 水分和盐分是影响该区域柽柳空间分布及其生态位的主要因素(贺强等, 2008 ; Cui et al ., 2009 ), 土壤水分、盐分、地下水水位及其矿化度是表征盐碱地水盐环境的主要指标, 而地下水通过“饱和带-包气带-植被”间的垂向联系由点及面产生极为重要的生态环境效应(Singh et al ., 2010 ; Gou & Miller, 2014 ; 夏江宝等, 2015 ), 特别是地下水矿化度的不同, 是导致土壤盐分和水分变化的主要因素.地下水通过影响土壤水分和盐分变化, 进而影响与植物光合生产力形成有关的物质运输和能量平衡(Álvarez & Sánchez-Blanco, 2014 ), 从而对植物生长及其光合生理生态过程产生重要影响.植物光合特性和树干液流速率可较好地反映植物生长及其水分利用对逆境条件的适应能力(夏江宝等, 2014 ; Xia et al ., 2015 ).植物光合效率能够表征植物在一定环境下的健康状况和光合生产能力, 可用来预测和表征植物对逆境的适应性和可塑性(Zhang et al ., 2014 ; Sanchez et al ., 2015 ).树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 ).目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
... ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
A management- oriented valuation method to determine ecological water requirement for wetlands in Yellow River Delta of China
1
2009
... 柽柳(Tamarix chinensis )属于典型的泌盐植物, 叶片和嫩枝可以将吸收于植物体内的盐分排出, 是黄河三角洲盐碱地植被恢复与生态重建的重要树种, 以其独特的生理和形态特点适应着盐碱地生境(Armas et al ., 2010 ).在黄河三角洲泥质海岸盐碱地, 地下水位浅且矿化度高, 水分和盐分是影响该区域柽柳空间分布及其生态位的主要因素(贺强等, 2008 ; Cui et al ., 2009 ), 土壤水分、盐分、地下水水位及其矿化度是表征盐碱地水盐环境的主要指标, 而地下水通过“饱和带-包气带-植被”间的垂向联系由点及面产生极为重要的生态环境效应(Singh et al ., 2010 ; Gou & Miller, 2014 ; 夏江宝等, 2015 ), 特别是地下水矿化度的不同, 是导致土壤盐分和水分变化的主要因素.地下水通过影响土壤水分和盐分变化, 进而影响与植物光合生产力形成有关的物质运输和能量平衡(Álvarez & Sánchez-Blanco, 2014 ), 从而对植物生长及其光合生理生态过程产生重要影响.植物光合特性和树干液流速率可较好地反映植物生长及其水分利用对逆境条件的适应能力(夏江宝等, 2014 ; Xia et al ., 2015 ).植物光合效率能够表征植物在一定环境下的健康状况和光合生产能力, 可用来预测和表征植物对逆境的适应性和可塑性(Zhang et al ., 2014 ; Sanchez et al ., 2015 ).树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 ).目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
Stomatal conductance and photosynthesis
2
1982
... 由图5 可知, 不同地下水矿化度下柽柳叶片G s 、C i 和L s 的光响应过程差异显著.盐水处理下, 柽柳G s 随PAR 增加变化比较平稳, 说明盐分胁迫对柽柳生长造成了一定的影响, 柽柳叶片气孔基本上失去了调节作用, 导致G s 对PAR 的变化响应不敏感.咸水至淡水处理下, 随地下水矿化度降低, 柽柳叶片P n 下降, G s 下降, C i 上升, L s 下降; 咸水至盐水处理下, 随地下水矿化度增加, 柽柳叶片P n 下降, G s 下降, C i 下降, L s 上升.依据气孔限制理论(Farquhar & Sharkey, 1982 ), 咸水至淡水处理下柽柳叶片P n 下降以非气孔限制为主, 咸水至盐水处理下柽柳叶片P n 下降以气孔限制为主. ...
... 气孔限制和非气孔限制是抑制光合作用下降的主要原因(Farquhar & Sharkey, 1982 ).王伟华等(2009)研究发现, 盐水浓度4 g∙L-1 处理下多枝柽柳光合作用受到促进, 其他盐水浓度(12、20 和28 g∙L-1 )处理下, 多枝柽柳植株光合能力下降受气孔限制影响, 并且灌溉盐水的浓度越大气孔限制越强烈.本研究也发现类似规律, 在盐水和淡水处理下, 柽柳P n 下降的原因分别是气孔限制和非气孔限制.但也有研究发现, 随地下水矿化度增大, 多枝柽柳幼苗P n 下降受非气孔限制影响(王鹏等, 2012 ).可见, 地下水水位及其矿化度以及土壤介质内盐胁迫处理方式不同时, 随盐分胁迫的增加, 柽柳叶片光合作用下降的气孔限制机理有一定差异.淡水至咸水处理下, 土壤含盐量和溶液绝对浓度分别为0.11%-1.05%, 0.64%-5.38%, 柽柳的泌盐特性可以缓解盐分胁迫对柽柳生长的影响; 咸水至盐水处理下, 土壤含盐量和溶液绝对浓度分别为1.05%- 1.17%, 5.38%-6.70%, 过高的盐胁迫降低了柽柳的光合作用, 柽柳为保证自身水分利用和避免光合机构受损而关闭气孔. ...
Comparison of salinity tolerance of three
1
2012
... 柽柳(Tamarix chinensis )属于典型的泌盐植物, 叶片和嫩枝可以将吸收于植物体内的盐分排出, 是黄河三角洲盐碱地植被恢复与生态重建的重要树种, 以其独特的生理和形态特点适应着盐碱地生境(Armas et al ., 2010 ).在黄河三角洲泥质海岸盐碱地, 地下水位浅且矿化度高, 水分和盐分是影响该区域柽柳空间分布及其生态位的主要因素(贺强等, 2008 ; Cui et al ., 2009 ), 土壤水分、盐分、地下水水位及其矿化度是表征盐碱地水盐环境的主要指标, 而地下水通过“饱和带-包气带-植被”间的垂向联系由点及面产生极为重要的生态环境效应(Singh et al ., 2010 ; Gou & Miller, 2014 ; 夏江宝等, 2015 ), 特别是地下水矿化度的不同, 是导致土壤盐分和水分变化的主要因素.地下水通过影响土壤水分和盐分变化, 进而影响与植物光合生产力形成有关的物质运输和能量平衡(Álvarez & Sánchez-Blanco, 2014 ), 从而对植物生长及其光合生理生态过程产生重要影响.植物光合特性和树干液流速率可较好地反映植物生长及其水分利用对逆境条件的适应能力(夏江宝等, 2014 ; Xia et al ., 2015 ).植物光合效率能够表征植物在一定环境下的健康状况和光合生产能力, 可用来预测和表征植物对逆境的适应性和可塑性(Zhang et al ., 2014 ; Sanchez et al ., 2015 ).树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 ).目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
浅埋条件下地下水-土壤-植物-大气连续体中水分运移研究规律, 农业环境科学学报,
1
2006
... 水分是盐分运移的载体, 潜水蒸发作用下, 浅埋深地下水通过毛管作用进入包气带土壤层, 致使土壤水盐发生变化, 进而影响植物根系生长和叶片的光合作用(宫兆宁等, 2006 ; Gou & Miller, 2014 ).本研究发现相同潜水水位下, 土壤盐分和溶液绝对浓度差异较大成为影响柽柳苗木光合作用的主要因素.王鹏等(2012)研究发现, 多枝柽柳幼苗P n 随地下水矿化度增大而升高, 咸水处理下P n 最高, 光照生态幅最宽, 微咸水处理下AQY 和R d 最高; 而本研究发现, 随地下水矿化度增大, 柽柳P n 先升高后降低, 咸水处理下P n 、P nmax 、AQY 和R d 均达到最大值, 微咸水处理下光照生态幅最宽.王伟华等(2009)研究发现, 多枝柽柳P n 日平均值随盐水浓度的增加先升高后降低, 盐水处理下P n 日平均值达最高.可见, 淡水处理下柽柳并非有最好的光合性能, 而适度提高地下水至咸水矿化度会促进柽柳苗木的光合特性, 应该与盐生植物柽柳的泌盐生理特性和较强的耐盐性有关, 这可能是柽柳长期适应盐碱地生境的一种竞争策略.地下水矿化度的增加使土壤含盐量增加, 柽柳苗木通过增强叶片的光能转化效率、R d 和光照生态幅来提高光合作用以适应盐胁迫, 并且抗氧化物酶系统可清除体内因盐分过多而产生的活性氧, 保持体内活性氧的动态平衡(苏华等, 2012 ; 朱金方等, 2012 , 2015 ), 这可能是咸水处理下柽柳光合能力显著提高的主要原因.柽柳的泌盐特性通过泌盐腺将体内过多的Na+ 、Cl- 以盐粒结晶的形式排出体外, 同时还将其他有害微量元素进行离子区隔化, 可溶性糖、脯氨酸等可维持细胞渗透压, 能使柽柳在高盐度环境下生长(Scholander et al ., 1996 ; Anderson et al ., 2005 ).但盐水处理下, 土壤含盐量和溶液绝对浓度均达最高, 致使土壤渗透势升高, 根系吸水困难, 导致柽柳AQY 、R d 和P nmax 降低, 光照生态幅变窄.李紫薇等(2014)也发现, 重度盐分胁迫使蒺藜苜蓿(Medicago truncatula )叶片光能转化效率降低, AQY 降低进而导致P nmax 、LSP 降低, LCP 升高, 光照生态幅变窄, R d 降低, 本研究结果与之类似.这主要是因为柽柳对Na+ 、Cl- 的吸收, 排斥了对另一些营养元素的吸收, 幼苗生长受到威胁; 同时重度盐分胁迫促进了叶绿素酶对叶绿素的分解使叶绿素合成系统受到破坏, 活性氧自由基超过超氧化物歧化酶清除量, 细胞受损严重(朱金方等, 2012 , 2015 ), 渗透调节物质的功能显著降低(朱金方等, 2013 ). ...
A groundwater-soil-plant-atmosphere continuum approach for modelling water stress, uptake, and hydraulic redistribution in phreatophytic vegetation
2
2014
... 柽柳(Tamarix chinensis )属于典型的泌盐植物, 叶片和嫩枝可以将吸收于植物体内的盐分排出, 是黄河三角洲盐碱地植被恢复与生态重建的重要树种, 以其独特的生理和形态特点适应着盐碱地生境(Armas et al ., 2010 ).在黄河三角洲泥质海岸盐碱地, 地下水位浅且矿化度高, 水分和盐分是影响该区域柽柳空间分布及其生态位的主要因素(贺强等, 2008 ; Cui et al ., 2009 ), 土壤水分、盐分、地下水水位及其矿化度是表征盐碱地水盐环境的主要指标, 而地下水通过“饱和带-包气带-植被”间的垂向联系由点及面产生极为重要的生态环境效应(Singh et al ., 2010 ; Gou & Miller, 2014 ; 夏江宝等, 2015 ), 特别是地下水矿化度的不同, 是导致土壤盐分和水分变化的主要因素.地下水通过影响土壤水分和盐分变化, 进而影响与植物光合生产力形成有关的物质运输和能量平衡(Álvarez & Sánchez-Blanco, 2014 ), 从而对植物生长及其光合生理生态过程产生重要影响.植物光合特性和树干液流速率可较好地反映植物生长及其水分利用对逆境条件的适应能力(夏江宝等, 2014 ; Xia et al ., 2015 ).植物光合效率能够表征植物在一定环境下的健康状况和光合生产能力, 可用来预测和表征植物对逆境的适应性和可塑性(Zhang et al ., 2014 ; Sanchez et al ., 2015 ).树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 ).目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
... 水分是盐分运移的载体, 潜水蒸发作用下, 浅埋深地下水通过毛管作用进入包气带土壤层, 致使土壤水盐发生变化, 进而影响植物根系生长和叶片的光合作用(宫兆宁等, 2006 ; Gou & Miller, 2014 ).本研究发现相同潜水水位下, 土壤盐分和溶液绝对浓度差异较大成为影响柽柳苗木光合作用的主要因素.王鹏等(2012)研究发现, 多枝柽柳幼苗P n 随地下水矿化度增大而升高, 咸水处理下P n 最高, 光照生态幅最宽, 微咸水处理下AQY 和R d 最高; 而本研究发现, 随地下水矿化度增大, 柽柳P n 先升高后降低, 咸水处理下P n 、P nmax 、AQY 和R d 均达到最大值, 微咸水处理下光照生态幅最宽.王伟华等(2009)研究发现, 多枝柽柳P n 日平均值随盐水浓度的增加先升高后降低, 盐水处理下P n 日平均值达最高.可见, 淡水处理下柽柳并非有最好的光合性能, 而适度提高地下水至咸水矿化度会促进柽柳苗木的光合特性, 应该与盐生植物柽柳的泌盐生理特性和较强的耐盐性有关, 这可能是柽柳长期适应盐碱地生境的一种竞争策略.地下水矿化度的增加使土壤含盐量增加, 柽柳苗木通过增强叶片的光能转化效率、R d 和光照生态幅来提高光合作用以适应盐胁迫, 并且抗氧化物酶系统可清除体内因盐分过多而产生的活性氧, 保持体内活性氧的动态平衡(苏华等, 2012 ; 朱金方等, 2012 , 2015 ), 这可能是咸水处理下柽柳光合能力显著提高的主要原因.柽柳的泌盐特性通过泌盐腺将体内过多的Na+ 、Cl- 以盐粒结晶的形式排出体外, 同时还将其他有害微量元素进行离子区隔化, 可溶性糖、脯氨酸等可维持细胞渗透压, 能使柽柳在高盐度环境下生长(Scholander et al ., 1996 ; Anderson et al ., 2005 ).但盐水处理下, 土壤含盐量和溶液绝对浓度均达最高, 致使土壤渗透势升高, 根系吸水困难, 导致柽柳AQY 、R d 和P nmax 降低, 光照生态幅变窄.李紫薇等(2014)也发现, 重度盐分胁迫使蒺藜苜蓿(Medicago truncatula )叶片光能转化效率降低, AQY 降低进而导致P nmax 、LSP 降低, LCP 升高, 光照生态幅变窄, R d 降低, 本研究结果与之类似.这主要是因为柽柳对Na+ 、Cl- 的吸收, 排斥了对另一些营养元素的吸收, 幼苗生长受到威胁; 同时重度盐分胁迫促进了叶绿素酶对叶绿素的分解使叶绿素合成系统受到破坏, 活性氧自由基超过超氧化物歧化酶清除量, 细胞受损严重(朱金方等, 2012 , 2015 ), 渗透调节物质的功能显著降低(朱金方等, 2013 ). ...
水、盐梯度下黄河三角洲湿地植物种的生态位. 应用生态学报,
1
2008
... 柽柳(Tamarix chinensis )属于典型的泌盐植物, 叶片和嫩枝可以将吸收于植物体内的盐分排出, 是黄河三角洲盐碱地植被恢复与生态重建的重要树种, 以其独特的生理和形态特点适应着盐碱地生境(Armas et al ., 2010 ).在黄河三角洲泥质海岸盐碱地, 地下水位浅且矿化度高, 水分和盐分是影响该区域柽柳空间分布及其生态位的主要因素(贺强等, 2008 ; Cui et al ., 2009 ), 土壤水分、盐分、地下水水位及其矿化度是表征盐碱地水盐环境的主要指标, 而地下水通过“饱和带-包气带-植被”间的垂向联系由点及面产生极为重要的生态环境效应(Singh et al ., 2010 ; Gou & Miller, 2014 ; 夏江宝等, 2015 ), 特别是地下水矿化度的不同, 是导致土壤盐分和水分变化的主要因素.地下水通过影响土壤水分和盐分变化, 进而影响与植物光合生产力形成有关的物质运输和能量平衡(Álvarez & Sánchez-Blanco, 2014 ), 从而对植物生长及其光合生理生态过程产生重要影响.植物光合特性和树干液流速率可较好地反映植物生长及其水分利用对逆境条件的适应能力(夏江宝等, 2014 ; Xia et al ., 2015 ).植物光合效率能够表征植物在一定环境下的健康状况和光合生产能力, 可用来预测和表征植物对逆境的适应性和可塑性(Zhang et al ., 2014 ; Sanchez et al ., 2015 ).树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 ).目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
水盐梯度下黄河三角洲湿地植被空间分异规律的定量分析. 湿地科学,
1
2007
... 柽柳(Tamarix chinensis )属于典型的泌盐植物, 叶片和嫩枝可以将吸收于植物体内的盐分排出, 是黄河三角洲盐碱地植被恢复与生态重建的重要树种, 以其独特的生理和形态特点适应着盐碱地生境(Armas et al ., 2010 ).在黄河三角洲泥质海岸盐碱地, 地下水位浅且矿化度高, 水分和盐分是影响该区域柽柳空间分布及其生态位的主要因素(贺强等, 2008 ; Cui et al ., 2009 ), 土壤水分、盐分、地下水水位及其矿化度是表征盐碱地水盐环境的主要指标, 而地下水通过“饱和带-包气带-植被”间的垂向联系由点及面产生极为重要的生态环境效应(Singh et al ., 2010 ; Gou & Miller, 2014 ; 夏江宝等, 2015 ), 特别是地下水矿化度的不同, 是导致土壤盐分和水分变化的主要因素.地下水通过影响土壤水分和盐分变化, 进而影响与植物光合生产力形成有关的物质运输和能量平衡(Álvarez & Sánchez-Blanco, 2014 ), 从而对植物生长及其光合生理生态过程产生重要影响.植物光合特性和树干液流速率可较好地反映植物生长及其水分利用对逆境条件的适应能力(夏江宝等, 2014 ; Xia et al ., 2015 ).植物光合效率能够表征植物在一定环境下的健康状况和光合生产能力, 可用来预测和表征植物对逆境的适应性和可塑性(Zhang et al ., 2014 ; Sanchez et al ., 2015 ).树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 ).目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
三种温带树种树干储存水对蒸腾的贡献. 植物生态学报,
1
2011
... 树干液流速率是反映植物液流瞬时耗水规律的主要参数, 不但能够反映植物生理用水对环境因子的响应过程和规律, 也是确定树体储存水对蒸腾耗水贡献程度的主要指标(金鹰等, 2011 ; Wang et al ., 2012 ).光照和VPD 是影响树木蒸腾耗水的主要驱动因子(倪广艳等, 2015 ; 徐世琴等, 2015 ), 相关研究发现, 荷木(Schima superba )蒸腾在干、湿季均与PAR 和VPD 显著正相关(倪广艳等, 2015 ); 梭梭(Haloxylon ammodendron )液流密度日变化过程受PAR 、温度和VPD 的共同影响呈多峰特征(徐世琴等, 2015 ).本研究发现, 地下水矿化度会显著影响柽柳树干液流速率的日动态, 柽柳T r 和液流速率日动态随PAR 和VPD 的变化呈现单峰或双峰特征, 在咸水和微咸水处理下, 柽柳叶片气孔调节和蒸腾耗水受光照、空气温湿度等环境因子的协同作用影响较大, 通过“午休”有效调节了光合生理过程, 利于其对水盐的有效利用; 而淡水和盐水处理下, 环境因子的协同效应对其蒸腾耗水过程影响相对较小, 气孔调节相对较弱.可见随地下水矿化度的不同, 影响柽柳蒸腾耗水日动态的诸多环境因子的协同效应有较大差异.在土壤盐分为1.05%的咸水矿化度下, 柽柳T r 和日液流量最大, 而地下水矿化度过高或过低均会导致柽柳树干液流速率日变幅和日耗水量降低, 适度提高地下水矿化度会增加柽柳树干的液流速率和日作用时间.已有研究表明: 柽柳蒸腾速率随盐胁迫增加而降低(米文精等, 2011 ); 微咸水和咸水处理下, 柽柳T r 高于淡水处理(王鹏等, 2012 ).可见, 地下水矿化度与土壤盐分胁迫可显著影响树木耗水特性, 适度提高地下水矿化度可增强柽柳的蒸腾生理活性, 这可能是因为作为泌盐植物的柽柳, 适度土壤盐分起到类似营养盐的作用, 在水分不受限制的条件下, 盐分增高导致其蒸腾耗水有升高趋势.但随咸水至盐水处理下盐分渗透胁迫增强, 进入植物体内的水分减少, 蒸腾作用和液流速率显著降低.过高的地下水矿化度下, 柽柳苗木表现出通过关闭气孔降低蒸腾耗水来维持自身水分有效利用的生理策略.植物能否较好地协调碳同化与水分耗散之间的关系是辨识植物生长适应逆境胁迫的主要方法, 水分利用效率是判定植物生长适应逆境生存与否的关键因子(曹生奎等, 2009 ).本研究发现: 淡水和盐水处理下, 柽柳WUE 分别达最高和最低, 地下水矿化度的增加可显著降低柽柳WUE, 但地下水矿化度对柽柳WUE 的影响不如P n 和T r 明显, 特别是咸水和微咸水处理下, 柽柳WUE 无显著差异(p > 0.05), 这可能是柽柳通过调节自身生理状况以适应不同地下水矿化度和土壤盐分环境的水分利用策略.而已有研究表明: 柽柳WUE 随盐胁迫的加重有所增加(刘玉娟等, 2015 ); 咸水处理下多枝柽柳WUE 始终大于微咸水和淡水处理(王鹏等, 2012 ).可见, 随盐分处理方式的不同, 柽柳WUE 对盐胁迫引起的渗透胁迫表现出一定差异, 这主要与柽柳P n 和T r 对盐胁迫的响应不一致有关.在微咸水和咸水处理下, 柽柳P n 、T r 和液流活动均保持在较高状态, 生理活动旺盛, 柽柳通过调节光合特性和日液流量的变化来适应盐胁迫, 能保持高的水分利用效率; 盐水处理下, 柽柳P n 最低, 但蒸腾耗水并非最低, 导致WUE 最低; 淡水处理下由于T r 对地下水矿化度响应更加敏感, T r 下降幅度高于P n , 导致WUE 最高, 但此时P n 和T r 均较低, 液流活动微弱, 易导致蒸腾耗水过低而使叶片温度升高, 长时间的这种生理过程对植物正常的生理活动不利. ...
Physiological and morphological responses of
1
2013
... 柽柳(Tamarix chinensis )属于典型的泌盐植物, 叶片和嫩枝可以将吸收于植物体内的盐分排出, 是黄河三角洲盐碱地植被恢复与生态重建的重要树种, 以其独特的生理和形态特点适应着盐碱地生境(Armas et al ., 2010 ).在黄河三角洲泥质海岸盐碱地, 地下水位浅且矿化度高, 水分和盐分是影响该区域柽柳空间分布及其生态位的主要因素(贺强等, 2008 ; Cui et al ., 2009 ), 土壤水分、盐分、地下水水位及其矿化度是表征盐碱地水盐环境的主要指标, 而地下水通过“饱和带-包气带-植被”间的垂向联系由点及面产生极为重要的生态环境效应(Singh et al ., 2010 ; Gou & Miller, 2014 ; 夏江宝等, 2015 ), 特别是地下水矿化度的不同, 是导致土壤盐分和水分变化的主要因素.地下水通过影响土壤水分和盐分变化, 进而影响与植物光合生产力形成有关的物质运输和能量平衡(Álvarez & Sánchez-Blanco, 2014 ), 从而对植物生长及其光合生理生态过程产生重要影响.植物光合特性和树干液流速率可较好地反映植物生长及其水分利用对逆境条件的适应能力(夏江宝等, 2014 ; Xia et al ., 2015 ).植物光合效率能够表征植物在一定环境下的健康状况和光合生产能力, 可用来预测和表征植物对逆境的适应性和可塑性(Zhang et al ., 2014 ; Sanchez et al ., 2015 ).树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 ).目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
蒺藜苜蓿叶片光合作用对盐胁迫的响应. 西北植物学报,
2014
杉木幼树树干液流影响因子及其对杉木林蒸腾量的贡献. 水土保持通报,
2014
盐胁迫对柽柳和白刺光合日变化的影响. 中国农学通报,
1
2015
... 树干液流速率是反映植物液流瞬时耗水规律的主要参数, 不但能够反映植物生理用水对环境因子的响应过程和规律, 也是确定树体储存水对蒸腾耗水贡献程度的主要指标(金鹰等, 2011 ; Wang et al ., 2012 ).光照和VPD 是影响树木蒸腾耗水的主要驱动因子(倪广艳等, 2015 ; 徐世琴等, 2015 ), 相关研究发现, 荷木(Schima superba )蒸腾在干、湿季均与PAR 和VPD 显著正相关(倪广艳等, 2015 ); 梭梭(Haloxylon ammodendron )液流密度日变化过程受PAR 、温度和VPD 的共同影响呈多峰特征(徐世琴等, 2015 ).本研究发现, 地下水矿化度会显著影响柽柳树干液流速率的日动态, 柽柳T r 和液流速率日动态随PAR 和VPD 的变化呈现单峰或双峰特征, 在咸水和微咸水处理下, 柽柳叶片气孔调节和蒸腾耗水受光照、空气温湿度等环境因子的协同作用影响较大, 通过“午休”有效调节了光合生理过程, 利于其对水盐的有效利用; 而淡水和盐水处理下, 环境因子的协同效应对其蒸腾耗水过程影响相对较小, 气孔调节相对较弱.可见随地下水矿化度的不同, 影响柽柳蒸腾耗水日动态的诸多环境因子的协同效应有较大差异.在土壤盐分为1.05%的咸水矿化度下, 柽柳T r 和日液流量最大, 而地下水矿化度过高或过低均会导致柽柳树干液流速率日变幅和日耗水量降低, 适度提高地下水矿化度会增加柽柳树干的液流速率和日作用时间.已有研究表明: 柽柳蒸腾速率随盐胁迫增加而降低(米文精等, 2011 ); 微咸水和咸水处理下, 柽柳T r 高于淡水处理(王鹏等, 2012 ).可见, 地下水矿化度与土壤盐分胁迫可显著影响树木耗水特性, 适度提高地下水矿化度可增强柽柳的蒸腾生理活性, 这可能是因为作为泌盐植物的柽柳, 适度土壤盐分起到类似营养盐的作用, 在水分不受限制的条件下, 盐分增高导致其蒸腾耗水有升高趋势.但随咸水至盐水处理下盐分渗透胁迫增强, 进入植物体内的水分减少, 蒸腾作用和液流速率显著降低.过高的地下水矿化度下, 柽柳苗木表现出通过关闭气孔降低蒸腾耗水来维持自身水分有效利用的生理策略.植物能否较好地协调碳同化与水分耗散之间的关系是辨识植物生长适应逆境胁迫的主要方法, 水分利用效率是判定植物生长适应逆境生存与否的关键因子(曹生奎等, 2009 ).本研究发现: 淡水和盐水处理下, 柽柳WUE 分别达最高和最低, 地下水矿化度的增加可显著降低柽柳WUE, 但地下水矿化度对柽柳WUE 的影响不如P n 和T r 明显, 特别是咸水和微咸水处理下, 柽柳WUE 无显著差异(p > 0.05), 这可能是柽柳通过调节自身生理状况以适应不同地下水矿化度和土壤盐分环境的水分利用策略.而已有研究表明: 柽柳WUE 随盐胁迫的加重有所增加(刘玉娟等, 2015 ); 咸水处理下多枝柽柳WUE 始终大于微咸水和淡水处理(王鹏等, 2012 ).可见, 随盐分处理方式的不同, 柽柳WUE 对盐胁迫引起的渗透胁迫表现出一定差异, 这主要与柽柳P n 和T r 对盐胁迫的响应不一致有关.在微咸水和咸水处理下, 柽柳P n 、T r 和液流活动均保持在较高状态, 生理活动旺盛, 柽柳通过调节光合特性和日液流量的变化来适应盐胁迫, 能保持高的水分利用效率; 盐水处理下, 柽柳P n 最低, 但蒸腾耗水并非最低, 导致WUE 最低; 淡水处理下由于T r 对地下水矿化度响应更加敏感, T r 下降幅度高于P n , 导致WUE 最高, 但此时P n 和T r 均较低, 液流活动微弱, 易导致蒸腾耗水过低而使叶片温度升高, 长时间的这种生理过程对植物正常的生理活动不利. ...
胡杨液流对地下水埋深变化的响应. 植物生态学报,
1
2010
... 柽柳(Tamarix chinensis )属于典型的泌盐植物, 叶片和嫩枝可以将吸收于植物体内的盐分排出, 是黄河三角洲盐碱地植被恢复与生态重建的重要树种, 以其独特的生理和形态特点适应着盐碱地生境(Armas et al ., 2010 ).在黄河三角洲泥质海岸盐碱地, 地下水位浅且矿化度高, 水分和盐分是影响该区域柽柳空间分布及其生态位的主要因素(贺强等, 2008 ; Cui et al ., 2009 ), 土壤水分、盐分、地下水水位及其矿化度是表征盐碱地水盐环境的主要指标, 而地下水通过“饱和带-包气带-植被”间的垂向联系由点及面产生极为重要的生态环境效应(Singh et al ., 2010 ; Gou & Miller, 2014 ; 夏江宝等, 2015 ), 特别是地下水矿化度的不同, 是导致土壤盐分和水分变化的主要因素.地下水通过影响土壤水分和盐分变化, 进而影响与植物光合生产力形成有关的物质运输和能量平衡(Álvarez & Sánchez-Blanco, 2014 ), 从而对植物生长及其光合生理生态过程产生重要影响.植物光合特性和树干液流速率可较好地反映植物生长及其水分利用对逆境条件的适应能力(夏江宝等, 2014 ; Xia et al ., 2015 ).植物光合效率能够表征植物在一定环境下的健康状况和光合生产能力, 可用来预测和表征植物对逆境的适应性和可塑性(Zhang et al ., 2014 ; Sanchez et al ., 2015 ).树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 ).目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
黄河三角洲典型植被与地下水埋深和土壤盐分的关系. 应用生态学报,
1
2013
... 黄河三角洲柽柳生长地带地下水位主要集中在300-2000 mm (马玉蕾等, 2013 ; 夏江宝等, 2015 ), 结合在科研温室内利于实验控制, 因此本研究以泥质海岸带柽柳大面积生长的地下水水位1200 mm为准, 模拟设置淡水、微咸水、咸水和盐水4种地下水矿化度, 其对应矿化度数值分别为0 g∙L-1 、3.00 g∙L-1 、8.00 g∙L-1 和20.00 g∙L-1 .2014年3月5日将柽柳栽植于直径为300 mm、高度为1780 mm的PVC圆管中, 其中PVC管实际淹水区为550 mm, 管底部设有纱网, 防止土壤渗漏.将PVC圆管放置于700 mm高, 桶底内径455 mm, 桶上口内径570 mm的水桶中, 为使地下水温度一致, 将水桶埋置于土壤中.不同地下水矿化度下栽植柽柳的土柱模拟示意图和实景图见图1 .为维持地下水水位及其矿化度的稳定性, 定期监测地下水盐度, 依次来补充所需盐分.柽柳苗木正常生长1月后对其进行不同地下水矿化度处理, 在此期间从地上部共浇水12.00 L, 分4次灌溉, 地下水矿化度处理后, 不再进行水分灌溉等措施.每种地下水矿化度处理下3个重复, 共计12个栽植柽柳的土壤柱体, 每土壤柱体栽植柽柳1株.在地下水矿化度处理80天后, 苗木平均高度为(67 ± 0.6) mm, 茎基部直径平均为(15.0 ± 0.7) mm, 6月27日开始柽柳苗木光合作用和树干液流的测定, 7月6日测定结束. ...
大同盆地盐碱地生态修复利用植物的初步选择. 北京林业大学学报,
1
2011
... 树干液流速率是反映植物液流瞬时耗水规律的主要参数, 不但能够反映植物生理用水对环境因子的响应过程和规律, 也是确定树体储存水对蒸腾耗水贡献程度的主要指标(金鹰等, 2011 ; Wang et al ., 2012 ).光照和VPD 是影响树木蒸腾耗水的主要驱动因子(倪广艳等, 2015 ; 徐世琴等, 2015 ), 相关研究发现, 荷木(Schima superba )蒸腾在干、湿季均与PAR 和VPD 显著正相关(倪广艳等, 2015 ); 梭梭(Haloxylon ammodendron )液流密度日变化过程受PAR 、温度和VPD 的共同影响呈多峰特征(徐世琴等, 2015 ).本研究发现, 地下水矿化度会显著影响柽柳树干液流速率的日动态, 柽柳T r 和液流速率日动态随PAR 和VPD 的变化呈现单峰或双峰特征, 在咸水和微咸水处理下, 柽柳叶片气孔调节和蒸腾耗水受光照、空气温湿度等环境因子的协同作用影响较大, 通过“午休”有效调节了光合生理过程, 利于其对水盐的有效利用; 而淡水和盐水处理下, 环境因子的协同效应对其蒸腾耗水过程影响相对较小, 气孔调节相对较弱.可见随地下水矿化度的不同, 影响柽柳蒸腾耗水日动态的诸多环境因子的协同效应有较大差异.在土壤盐分为1.05%的咸水矿化度下, 柽柳T r 和日液流量最大, 而地下水矿化度过高或过低均会导致柽柳树干液流速率日变幅和日耗水量降低, 适度提高地下水矿化度会增加柽柳树干的液流速率和日作用时间.已有研究表明: 柽柳蒸腾速率随盐胁迫增加而降低(米文精等, 2011 ); 微咸水和咸水处理下, 柽柳T r 高于淡水处理(王鹏等, 2012 ).可见, 地下水矿化度与土壤盐分胁迫可显著影响树木耗水特性, 适度提高地下水矿化度可增强柽柳的蒸腾生理活性, 这可能是因为作为泌盐植物的柽柳, 适度土壤盐分起到类似营养盐的作用, 在水分不受限制的条件下, 盐分增高导致其蒸腾耗水有升高趋势.但随咸水至盐水处理下盐分渗透胁迫增强, 进入植物体内的水分减少, 蒸腾作用和液流速率显著降低.过高的地下水矿化度下, 柽柳苗木表现出通过关闭气孔降低蒸腾耗水来维持自身水分有效利用的生理策略.植物能否较好地协调碳同化与水分耗散之间的关系是辨识植物生长适应逆境胁迫的主要方法, 水分利用效率是判定植物生长适应逆境生存与否的关键因子(曹生奎等, 2009 ).本研究发现: 淡水和盐水处理下, 柽柳WUE 分别达最高和最低, 地下水矿化度的增加可显著降低柽柳WUE, 但地下水矿化度对柽柳WUE 的影响不如P n 和T r 明显, 特别是咸水和微咸水处理下, 柽柳WUE 无显著差异(p > 0.05), 这可能是柽柳通过调节自身生理状况以适应不同地下水矿化度和土壤盐分环境的水分利用策略.而已有研究表明: 柽柳WUE 随盐胁迫的加重有所增加(刘玉娟等, 2015 ); 咸水处理下多枝柽柳WUE 始终大于微咸水和淡水处理(王鹏等, 2012 ).可见, 随盐分处理方式的不同, 柽柳WUE 对盐胁迫引起的渗透胁迫表现出一定差异, 这主要与柽柳P n 和T r 对盐胁迫的响应不一致有关.在微咸水和咸水处理下, 柽柳P n 、T r 和液流活动均保持在较高状态, 生理活动旺盛, 柽柳通过调节光合特性和日液流量的变化来适应盐胁迫, 能保持高的水分利用效率; 盐水处理下, 柽柳P n 最低, 但蒸腾耗水并非最低, 导致WUE 最低; 淡水处理下由于T r 对地下水矿化度响应更加敏感, T r 下降幅度高于P n , 导致WUE 最高, 但此时P n 和T r 均较低, 液流活动微弱, 易导致蒸腾耗水过低而使叶片温度升高, 长时间的这种生理过程对植物正常的生理活动不利. ...
荷木整树蒸腾对干湿季土壤水分的水力响应. 生态学报,
2
2015
... 树干液流速率是反映植物液流瞬时耗水规律的主要参数, 不但能够反映植物生理用水对环境因子的响应过程和规律, 也是确定树体储存水对蒸腾耗水贡献程度的主要指标(金鹰等, 2011 ; Wang et al ., 2012 ).光照和VPD 是影响树木蒸腾耗水的主要驱动因子(倪广艳等, 2015 ; 徐世琴等, 2015 ), 相关研究发现, 荷木(Schima superba )蒸腾在干、湿季均与PAR 和VPD 显著正相关(倪广艳等, 2015 ); 梭梭(Haloxylon ammodendron )液流密度日变化过程受PAR 、温度和VPD 的共同影响呈多峰特征(徐世琴等, 2015 ).本研究发现, 地下水矿化度会显著影响柽柳树干液流速率的日动态, 柽柳T r 和液流速率日动态随PAR 和VPD 的变化呈现单峰或双峰特征, 在咸水和微咸水处理下, 柽柳叶片气孔调节和蒸腾耗水受光照、空气温湿度等环境因子的协同作用影响较大, 通过“午休”有效调节了光合生理过程, 利于其对水盐的有效利用; 而淡水和盐水处理下, 环境因子的协同效应对其蒸腾耗水过程影响相对较小, 气孔调节相对较弱.可见随地下水矿化度的不同, 影响柽柳蒸腾耗水日动态的诸多环境因子的协同效应有较大差异.在土壤盐分为1.05%的咸水矿化度下, 柽柳T r 和日液流量最大, 而地下水矿化度过高或过低均会导致柽柳树干液流速率日变幅和日耗水量降低, 适度提高地下水矿化度会增加柽柳树干的液流速率和日作用时间.已有研究表明: 柽柳蒸腾速率随盐胁迫增加而降低(米文精等, 2011 ); 微咸水和咸水处理下, 柽柳T r 高于淡水处理(王鹏等, 2012 ).可见, 地下水矿化度与土壤盐分胁迫可显著影响树木耗水特性, 适度提高地下水矿化度可增强柽柳的蒸腾生理活性, 这可能是因为作为泌盐植物的柽柳, 适度土壤盐分起到类似营养盐的作用, 在水分不受限制的条件下, 盐分增高导致其蒸腾耗水有升高趋势.但随咸水至盐水处理下盐分渗透胁迫增强, 进入植物体内的水分减少, 蒸腾作用和液流速率显著降低.过高的地下水矿化度下, 柽柳苗木表现出通过关闭气孔降低蒸腾耗水来维持自身水分有效利用的生理策略.植物能否较好地协调碳同化与水分耗散之间的关系是辨识植物生长适应逆境胁迫的主要方法, 水分利用效率是判定植物生长适应逆境生存与否的关键因子(曹生奎等, 2009 ).本研究发现: 淡水和盐水处理下, 柽柳WUE 分别达最高和最低, 地下水矿化度的增加可显著降低柽柳WUE, 但地下水矿化度对柽柳WUE 的影响不如P n 和T r 明显, 特别是咸水和微咸水处理下, 柽柳WUE 无显著差异(p > 0.05), 这可能是柽柳通过调节自身生理状况以适应不同地下水矿化度和土壤盐分环境的水分利用策略.而已有研究表明: 柽柳WUE 随盐胁迫的加重有所增加(刘玉娟等, 2015 ); 咸水处理下多枝柽柳WUE 始终大于微咸水和淡水处理(王鹏等, 2012 ).可见, 随盐分处理方式的不同, 柽柳WUE 对盐胁迫引起的渗透胁迫表现出一定差异, 这主要与柽柳P n 和T r 对盐胁迫的响应不一致有关.在微咸水和咸水处理下, 柽柳P n 、T r 和液流活动均保持在较高状态, 生理活动旺盛, 柽柳通过调节光合特性和日液流量的变化来适应盐胁迫, 能保持高的水分利用效率; 盐水处理下, 柽柳P n 最低, 但蒸腾耗水并非最低, 导致WUE 最低; 淡水处理下由于T r 对地下水矿化度响应更加敏感, T r 下降幅度高于P n , 导致WUE 最高, 但此时P n 和T r 均较低, 液流活动微弱, 易导致蒸腾耗水过低而使叶片温度升高, 长时间的这种生理过程对植物正常的生理活动不利. ...
... 显著正相关(倪广艳等, 2015 ); 梭梭(Haloxylon ammodendron )液流密度日变化过程受PAR 、温度和VPD 的共同影响呈多峰特征(徐世琴等, 2015 ).本研究发现, 地下水矿化度会显著影响柽柳树干液流速率的日动态, 柽柳T r 和液流速率日动态随PAR 和VPD 的变化呈现单峰或双峰特征, 在咸水和微咸水处理下, 柽柳叶片气孔调节和蒸腾耗水受光照、空气温湿度等环境因子的协同作用影响较大, 通过“午休”有效调节了光合生理过程, 利于其对水盐的有效利用; 而淡水和盐水处理下, 环境因子的协同效应对其蒸腾耗水过程影响相对较小, 气孔调节相对较弱.可见随地下水矿化度的不同, 影响柽柳蒸腾耗水日动态的诸多环境因子的协同效应有较大差异.在土壤盐分为1.05%的咸水矿化度下, 柽柳T r 和日液流量最大, 而地下水矿化度过高或过低均会导致柽柳树干液流速率日变幅和日耗水量降低, 适度提高地下水矿化度会增加柽柳树干的液流速率和日作用时间.已有研究表明: 柽柳蒸腾速率随盐胁迫增加而降低(米文精等, 2011 ); 微咸水和咸水处理下, 柽柳T r 高于淡水处理(王鹏等, 2012 ).可见, 地下水矿化度与土壤盐分胁迫可显著影响树木耗水特性, 适度提高地下水矿化度可增强柽柳的蒸腾生理活性, 这可能是因为作为泌盐植物的柽柳, 适度土壤盐分起到类似营养盐的作用, 在水分不受限制的条件下, 盐分增高导致其蒸腾耗水有升高趋势.但随咸水至盐水处理下盐分渗透胁迫增强, 进入植物体内的水分减少, 蒸腾作用和液流速率显著降低.过高的地下水矿化度下, 柽柳苗木表现出通过关闭气孔降低蒸腾耗水来维持自身水分有效利用的生理策略.植物能否较好地协调碳同化与水分耗散之间的关系是辨识植物生长适应逆境胁迫的主要方法, 水分利用效率是判定植物生长适应逆境生存与否的关键因子(曹生奎等, 2009 ).本研究发现: 淡水和盐水处理下, 柽柳WUE 分别达最高和最低, 地下水矿化度的增加可显著降低柽柳WUE, 但地下水矿化度对柽柳WUE 的影响不如P n 和T r 明显, 特别是咸水和微咸水处理下, 柽柳WUE 无显著差异(p > 0.05), 这可能是柽柳通过调节自身生理状况以适应不同地下水矿化度和土壤盐分环境的水分利用策略.而已有研究表明: 柽柳WUE 随盐胁迫的加重有所增加(刘玉娟等, 2015 ); 咸水处理下多枝柽柳WUE 始终大于微咸水和淡水处理(王鹏等, 2012 ).可见, 随盐分处理方式的不同, 柽柳WUE 对盐胁迫引起的渗透胁迫表现出一定差异, 这主要与柽柳P n 和T r 对盐胁迫的响应不一致有关.在微咸水和咸水处理下, 柽柳P n 、T r 和液流活动均保持在较高状态, 生理活动旺盛, 柽柳通过调节光合特性和日液流量的变化来适应盐胁迫, 能保持高的水分利用效率; 盐水处理下, 柽柳P n 最低, 但蒸腾耗水并非最低, 导致WUE 最低; 淡水处理下由于T r 对地下水矿化度响应更加敏感, T r 下降幅度高于P n , 导致WUE 最高, 但此时P n 和T r 均较低, 液流活动微弱, 易导致蒸腾耗水过低而使叶片温度升高, 长时间的这种生理过程对植物正常的生理活动不利. ...
Stomatal regulation in a changing climate: A field study using free air temperature increase (FATI) and free air CO2 enrichment (FACE)
1
1997
... 应用LI-6400XT便携式光合作用仪(LI-COR, Lincoln, USA)测定不同地下水矿化度处理下柽柳叶片的光响应曲线, 测定在晴朗天气的8:30-11:30进行, 使用缓冲瓶维持CO2 浓度的稳定性((390 ± 5) μmol∙mol-1 ), 设定人工光源的光合有效辐射(PAR )分别为: 1400、1200、1000、800、500、300、200、100、50、20、0 μmol∙m-2 ∙s-1 , 记录柽柳叶片的净光合速率(P n )、蒸腾速率(T r )、气孔导度(G s )、胞间CO2 浓度(C i )和大气CO2 浓度(C a )等参数.叶片瞬时水分利用效率(WUE )为P n 与T r 的比值, 即WUE = P n /T r (Nijs et al ., 1997 ); 气孔限制值(L s )为C i 与C a 的比值, 即L s = 1 - C i /C a (Wong et al ., 1979 ).同时在7:00- 19:00用LI-6400XT光合仪进行柽柳光合参数日动态的测定, 仪器自动记录T r 、G s 、PAR 和水汽压亏缺(VPD )等参数, 每2 h测定1次.以上测定每个处理测定3株苗木, 每株选取中上部生长健壮的成熟叶片2-3个, 在具体测定时, 采用不同处理、不同植株和叶片重复之间的交替测定法, 因此, 每株苗木依据测定的叶片数可测定2-3次, 以尽量避免时间波动对光合参数的影响.由于柽柳叶片形状不规则, 为得到精确的测量数据, 测定时需将标记的叶片平铺于整个测定叶室内, 同时参考邓雄(2003)和闫海龙(2010)等对不规则植物叶片光合参数的校正方法, 将观测的植物叶片剪下, 用扫描仪扫描后, 使用面积分析软件Delta-T Scan (CB50EJ, Cambridge, UK)计算实际的叶表面积, 按计算后的实际光合有效面积重新测算光合参数. ...
NaCl salinity-induced changes in water status, iron contents and photosynthetic properties of
1
2010
... 柽柳(Tamarix chinensis )属于典型的泌盐植物, 叶片和嫩枝可以将吸收于植物体内的盐分排出, 是黄河三角洲盐碱地植被恢复与生态重建的重要树种, 以其独特的生理和形态特点适应着盐碱地生境(Armas et al ., 2010 ).在黄河三角洲泥质海岸盐碱地, 地下水位浅且矿化度高, 水分和盐分是影响该区域柽柳空间分布及其生态位的主要因素(贺强等, 2008 ; Cui et al ., 2009 ), 土壤水分、盐分、地下水水位及其矿化度是表征盐碱地水盐环境的主要指标, 而地下水通过“饱和带-包气带-植被”间的垂向联系由点及面产生极为重要的生态环境效应(Singh et al ., 2010 ; Gou & Miller, 2014 ; 夏江宝等, 2015 ), 特别是地下水矿化度的不同, 是导致土壤盐分和水分变化的主要因素.地下水通过影响土壤水分和盐分变化, 进而影响与植物光合生产力形成有关的物质运输和能量平衡(Álvarez & Sánchez-Blanco, 2014 ), 从而对植物生长及其光合生理生态过程产生重要影响.植物光合特性和树干液流速率可较好地反映植物生长及其水分利用对逆境条件的适应能力(夏江宝等, 2014 ; Xia et al ., 2015 ).植物光合效率能够表征植物在一定环境下的健康状况和光合生产能力, 可用来预测和表征植物对逆境的适应性和可塑性(Zhang et al ., 2014 ; Sanchez et al ., 2015 ).树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 ).目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
黄河三角洲地下水生态水位埋深研究. 水电能源科学,
1
2010
... 柽柳(Tamarix chinensis )属于典型的泌盐植物, 叶片和嫩枝可以将吸收于植物体内的盐分排出, 是黄河三角洲盐碱地植被恢复与生态重建的重要树种, 以其独特的生理和形态特点适应着盐碱地生境(Armas et al ., 2010 ).在黄河三角洲泥质海岸盐碱地, 地下水位浅且矿化度高, 水分和盐分是影响该区域柽柳空间分布及其生态位的主要因素(贺强等, 2008 ; Cui et al ., 2009 ), 土壤水分、盐分、地下水水位及其矿化度是表征盐碱地水盐环境的主要指标, 而地下水通过“饱和带-包气带-植被”间的垂向联系由点及面产生极为重要的生态环境效应(Singh et al ., 2010 ; Gou & Miller, 2014 ; 夏江宝等, 2015 ), 特别是地下水矿化度的不同, 是导致土壤盐分和水分变化的主要因素.地下水通过影响土壤水分和盐分变化, 进而影响与植物光合生产力形成有关的物质运输和能量平衡(Álvarez & Sánchez-Blanco, 2014 ), 从而对植物生长及其光合生理生态过程产生重要影响.植物光合特性和树干液流速率可较好地反映植物生长及其水分利用对逆境条件的适应能力(夏江宝等, 2014 ; Xia et al ., 2015 ).植物光合效率能够表征植物在一定环境下的健康状况和光合生产能力, 可用来预测和表征植物对逆境的适应性和可塑性(Zhang et al ., 2014 ; Sanchez et al ., 2015 ).树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 ).目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
Salinity and water stress effects on biomass production in different
1
2015
... 柽柳(Tamarix chinensis )属于典型的泌盐植物, 叶片和嫩枝可以将吸收于植物体内的盐分排出, 是黄河三角洲盐碱地植被恢复与生态重建的重要树种, 以其独特的生理和形态特点适应着盐碱地生境(Armas et al ., 2010 ).在黄河三角洲泥质海岸盐碱地, 地下水位浅且矿化度高, 水分和盐分是影响该区域柽柳空间分布及其生态位的主要因素(贺强等, 2008 ; Cui et al ., 2009 ), 土壤水分、盐分、地下水水位及其矿化度是表征盐碱地水盐环境的主要指标, 而地下水通过“饱和带-包气带-植被”间的垂向联系由点及面产生极为重要的生态环境效应(Singh et al ., 2010 ; Gou & Miller, 2014 ; 夏江宝等, 2015 ), 特别是地下水矿化度的不同, 是导致土壤盐分和水分变化的主要因素.地下水通过影响土壤水分和盐分变化, 进而影响与植物光合生产力形成有关的物质运输和能量平衡(Álvarez & Sánchez-Blanco, 2014 ), 从而对植物生长及其光合生理生态过程产生重要影响.植物光合特性和树干液流速率可较好地反映植物生长及其水分利用对逆境条件的适应能力(夏江宝等, 2014 ; Xia et al ., 2015 ).植物光合效率能够表征植物在一定环境下的健康状况和光合生产能力, 可用来预测和表征植物对逆境的适应性和可塑性(Zhang et al ., 2014 ; Sanchez et al ., 2015 ).树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 ).目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
Sap concentrations in halophytes and some other plants
1
1996
... 水分是盐分运移的载体, 潜水蒸发作用下, 浅埋深地下水通过毛管作用进入包气带土壤层, 致使土壤水盐发生变化, 进而影响植物根系生长和叶片的光合作用(宫兆宁等, 2006 ; Gou & Miller, 2014 ).本研究发现相同潜水水位下, 土壤盐分和溶液绝对浓度差异较大成为影响柽柳苗木光合作用的主要因素.王鹏等(2012)研究发现, 多枝柽柳幼苗P n 随地下水矿化度增大而升高, 咸水处理下P n 最高, 光照生态幅最宽, 微咸水处理下AQY 和R d 最高; 而本研究发现, 随地下水矿化度增大, 柽柳P n 先升高后降低, 咸水处理下P n 、P nmax 、AQY 和R d 均达到最大值, 微咸水处理下光照生态幅最宽.王伟华等(2009)研究发现, 多枝柽柳P n 日平均值随盐水浓度的增加先升高后降低, 盐水处理下P n 日平均值达最高.可见, 淡水处理下柽柳并非有最好的光合性能, 而适度提高地下水至咸水矿化度会促进柽柳苗木的光合特性, 应该与盐生植物柽柳的泌盐生理特性和较强的耐盐性有关, 这可能是柽柳长期适应盐碱地生境的一种竞争策略.地下水矿化度的增加使土壤含盐量增加, 柽柳苗木通过增强叶片的光能转化效率、R d 和光照生态幅来提高光合作用以适应盐胁迫, 并且抗氧化物酶系统可清除体内因盐分过多而产生的活性氧, 保持体内活性氧的动态平衡(苏华等, 2012 ; 朱金方等, 2012 , 2015 ), 这可能是咸水处理下柽柳光合能力显著提高的主要原因.柽柳的泌盐特性通过泌盐腺将体内过多的Na+ 、Cl- 以盐粒结晶的形式排出体外, 同时还将其他有害微量元素进行离子区隔化, 可溶性糖、脯氨酸等可维持细胞渗透压, 能使柽柳在高盐度环境下生长(Scholander et al ., 1996 ; Anderson et al ., 2005 ).但盐水处理下, 土壤含盐量和溶液绝对浓度均达最高, 致使土壤渗透势升高, 根系吸水困难, 导致柽柳AQY 、R d 和P nmax 降低, 光照生态幅变窄.李紫薇等(2014)也发现, 重度盐分胁迫使蒺藜苜蓿(Medicago truncatula )叶片光能转化效率降低, AQY 降低进而导致P nmax 、LSP 降低, LCP 升高, 光照生态幅变窄, R d 降低, 本研究结果与之类似.这主要是因为柽柳对Na+ 、Cl- 的吸收, 排斥了对另一些营养元素的吸收, 幼苗生长受到威胁; 同时重度盐分胁迫促进了叶绿素酶对叶绿素的分解使叶绿素合成系统受到破坏, 活性氧自由基超过超氧化物歧化酶清除量, 细胞受损严重(朱金方等, 2012 , 2015 ), 渗透调节物质的功能显著降低(朱金方等, 2013 ). ...
Arsenic in water, soil, and rice plants in the Indo-Gangetic plains of northwestern India
1
2010
... 柽柳(Tamarix chinensis )属于典型的泌盐植物, 叶片和嫩枝可以将吸收于植物体内的盐分排出, 是黄河三角洲盐碱地植被恢复与生态重建的重要树种, 以其独特的生理和形态特点适应着盐碱地生境(Armas et al ., 2010 ).在黄河三角洲泥质海岸盐碱地, 地下水位浅且矿化度高, 水分和盐分是影响该区域柽柳空间分布及其生态位的主要因素(贺强等, 2008 ; Cui et al ., 2009 ), 土壤水分、盐分、地下水水位及其矿化度是表征盐碱地水盐环境的主要指标, 而地下水通过“饱和带-包气带-植被”间的垂向联系由点及面产生极为重要的生态环境效应(Singh et al ., 2010 ; Gou & Miller, 2014 ; 夏江宝等, 2015 ), 特别是地下水矿化度的不同, 是导致土壤盐分和水分变化的主要因素.地下水通过影响土壤水分和盐分变化, 进而影响与植物光合生产力形成有关的物质运输和能量平衡(Álvarez & Sánchez-Blanco, 2014 ), 从而对植物生长及其光合生理生态过程产生重要影响.植物光合特性和树干液流速率可较好地反映植物生长及其水分利用对逆境条件的适应能力(夏江宝等, 2014 ; Xia et al ., 2015 ).植物光合效率能够表征植物在一定环境下的健康状况和光合生产能力, 可用来预测和表征植物对逆境的适应性和可塑性(Zhang et al ., 2014 ; Sanchez et al ., 2015 ).树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 ).目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
地下水位下降对浑善达克沙地榆树光合及抗逆性的影响. 植物生态学报,
2
2012
... 柽柳(Tamarix chinensis )属于典型的泌盐植物, 叶片和嫩枝可以将吸收于植物体内的盐分排出, 是黄河三角洲盐碱地植被恢复与生态重建的重要树种, 以其独特的生理和形态特点适应着盐碱地生境(Armas et al ., 2010 ).在黄河三角洲泥质海岸盐碱地, 地下水位浅且矿化度高, 水分和盐分是影响该区域柽柳空间分布及其生态位的主要因素(贺强等, 2008 ; Cui et al ., 2009 ), 土壤水分、盐分、地下水水位及其矿化度是表征盐碱地水盐环境的主要指标, 而地下水通过“饱和带-包气带-植被”间的垂向联系由点及面产生极为重要的生态环境效应(Singh et al ., 2010 ; Gou & Miller, 2014 ; 夏江宝等, 2015 ), 特别是地下水矿化度的不同, 是导致土壤盐分和水分变化的主要因素.地下水通过影响土壤水分和盐分变化, 进而影响与植物光合生产力形成有关的物质运输和能量平衡(Álvarez & Sánchez-Blanco, 2014 ), 从而对植物生长及其光合生理生态过程产生重要影响.植物光合特性和树干液流速率可较好地反映植物生长及其水分利用对逆境条件的适应能力(夏江宝等, 2014 ; Xia et al ., 2015 ).植物光合效率能够表征植物在一定环境下的健康状况和光合生产能力, 可用来预测和表征植物对逆境的适应性和可塑性(Zhang et al ., 2014 ; Sanchez et al ., 2015 ).树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 ).目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
... 水分是盐分运移的载体, 潜水蒸发作用下, 浅埋深地下水通过毛管作用进入包气带土壤层, 致使土壤水盐发生变化, 进而影响植物根系生长和叶片的光合作用(宫兆宁等, 2006 ; Gou & Miller, 2014 ).本研究发现相同潜水水位下, 土壤盐分和溶液绝对浓度差异较大成为影响柽柳苗木光合作用的主要因素.王鹏等(2012)研究发现, 多枝柽柳幼苗P n 随地下水矿化度增大而升高, 咸水处理下P n 最高, 光照生态幅最宽, 微咸水处理下AQY 和R d 最高; 而本研究发现, 随地下水矿化度增大, 柽柳P n 先升高后降低, 咸水处理下P n 、P nmax 、AQY 和R d 均达到最大值, 微咸水处理下光照生态幅最宽.王伟华等(2009)研究发现, 多枝柽柳P n 日平均值随盐水浓度的增加先升高后降低, 盐水处理下P n 日平均值达最高.可见, 淡水处理下柽柳并非有最好的光合性能, 而适度提高地下水至咸水矿化度会促进柽柳苗木的光合特性, 应该与盐生植物柽柳的泌盐生理特性和较强的耐盐性有关, 这可能是柽柳长期适应盐碱地生境的一种竞争策略.地下水矿化度的增加使土壤含盐量增加, 柽柳苗木通过增强叶片的光能转化效率、R d 和光照生态幅来提高光合作用以适应盐胁迫, 并且抗氧化物酶系统可清除体内因盐分过多而产生的活性氧, 保持体内活性氧的动态平衡(苏华等, 2012 ; 朱金方等, 2012 , 2015 ), 这可能是咸水处理下柽柳光合能力显著提高的主要原因.柽柳的泌盐特性通过泌盐腺将体内过多的Na+ 、Cl- 以盐粒结晶的形式排出体外, 同时还将其他有害微量元素进行离子区隔化, 可溶性糖、脯氨酸等可维持细胞渗透压, 能使柽柳在高盐度环境下生长(Scholander et al ., 1996 ; Anderson et al ., 2005 ).但盐水处理下, 土壤含盐量和溶液绝对浓度均达最高, 致使土壤渗透势升高, 根系吸水困难, 导致柽柳AQY 、R d 和P nmax 降低, 光照生态幅变窄.李紫薇等(2014)也发现, 重度盐分胁迫使蒺藜苜蓿(Medicago truncatula )叶片光能转化效率降低, AQY 降低进而导致P nmax 、LSP 降低, LCP 升高, 光照生态幅变窄, R d 降低, 本研究结果与之类似.这主要是因为柽柳对Na+ 、Cl- 的吸收, 排斥了对另一些营养元素的吸收, 幼苗生长受到威胁; 同时重度盐分胁迫促进了叶绿素酶对叶绿素的分解使叶绿素合成系统受到破坏, 活性氧自由基超过超氧化物歧化酶清除量, 细胞受损严重(朱金方等, 2012 , 2015 ), 渗透调节物质的功能显著降低(朱金方等, 2013 ). ...
Nighttime sap flow of
1
2012
... 树干液流速率是反映植物液流瞬时耗水规律的主要参数, 不但能够反映植物生理用水对环境因子的响应过程和规律, 也是确定树体储存水对蒸腾耗水贡献程度的主要指标(金鹰等, 2011 ; Wang et al ., 2012 ).光照和VPD 是影响树木蒸腾耗水的主要驱动因子(倪广艳等, 2015 ; 徐世琴等, 2015 ), 相关研究发现, 荷木(Schima superba )蒸腾在干、湿季均与PAR 和VPD 显著正相关(倪广艳等, 2015 ); 梭梭(Haloxylon ammodendron )液流密度日变化过程受PAR 、温度和VPD 的共同影响呈多峰特征(徐世琴等, 2015 ).本研究发现, 地下水矿化度会显著影响柽柳树干液流速率的日动态, 柽柳T r 和液流速率日动态随PAR 和VPD 的变化呈现单峰或双峰特征, 在咸水和微咸水处理下, 柽柳叶片气孔调节和蒸腾耗水受光照、空气温湿度等环境因子的协同作用影响较大, 通过“午休”有效调节了光合生理过程, 利于其对水盐的有效利用; 而淡水和盐水处理下, 环境因子的协同效应对其蒸腾耗水过程影响相对较小, 气孔调节相对较弱.可见随地下水矿化度的不同, 影响柽柳蒸腾耗水日动态的诸多环境因子的协同效应有较大差异.在土壤盐分为1.05%的咸水矿化度下, 柽柳T r 和日液流量最大, 而地下水矿化度过高或过低均会导致柽柳树干液流速率日变幅和日耗水量降低, 适度提高地下水矿化度会增加柽柳树干的液流速率和日作用时间.已有研究表明: 柽柳蒸腾速率随盐胁迫增加而降低(米文精等, 2011 ); 微咸水和咸水处理下, 柽柳T r 高于淡水处理(王鹏等, 2012 ).可见, 地下水矿化度与土壤盐分胁迫可显著影响树木耗水特性, 适度提高地下水矿化度可增强柽柳的蒸腾生理活性, 这可能是因为作为泌盐植物的柽柳, 适度土壤盐分起到类似营养盐的作用, 在水分不受限制的条件下, 盐分增高导致其蒸腾耗水有升高趋势.但随咸水至盐水处理下盐分渗透胁迫增强, 进入植物体内的水分减少, 蒸腾作用和液流速率显著降低.过高的地下水矿化度下, 柽柳苗木表现出通过关闭气孔降低蒸腾耗水来维持自身水分有效利用的生理策略.植物能否较好地协调碳同化与水分耗散之间的关系是辨识植物生长适应逆境胁迫的主要方法, 水分利用效率是判定植物生长适应逆境生存与否的关键因子(曹生奎等, 2009 ).本研究发现: 淡水和盐水处理下, 柽柳WUE 分别达最高和最低, 地下水矿化度的增加可显著降低柽柳WUE, 但地下水矿化度对柽柳WUE 的影响不如P n 和T r 明显, 特别是咸水和微咸水处理下, 柽柳WUE 无显著差异(p > 0.05), 这可能是柽柳通过调节自身生理状况以适应不同地下水矿化度和土壤盐分环境的水分利用策略.而已有研究表明: 柽柳WUE 随盐胁迫的加重有所增加(刘玉娟等, 2015 ); 咸水处理下多枝柽柳WUE 始终大于微咸水和淡水处理(王鹏等, 2012 ).可见, 随盐分处理方式的不同, 柽柳WUE 对盐胁迫引起的渗透胁迫表现出一定差异, 这主要与柽柳P n 和T r 对盐胁迫的响应不一致有关.在微咸水和咸水处理下, 柽柳P n 、T r 和液流活动均保持在较高状态, 生理活动旺盛, 柽柳通过调节光合特性和日液流量的变化来适应盐胁迫, 能保持高的水分利用效率; 盐水处理下, 柽柳P n 最低, 但蒸腾耗水并非最低, 导致WUE 最低; 淡水处理下由于T r 对地下水矿化度响应更加敏感, T r 下降幅度高于P n , 导致WUE 最高, 但此时P n 和T r 均较低, 液流活动微弱, 易导致蒸腾耗水过低而使叶片温度升高, 长时间的这种生理过程对植物正常的生理活动不利. ...
地下水埋深及矿化度对多枝柽柳幼苗光合特性及生长的影响. 水土保持通报,
4
2012
... 柽柳(Tamarix chinensis )属于典型的泌盐植物, 叶片和嫩枝可以将吸收于植物体内的盐分排出, 是黄河三角洲盐碱地植被恢复与生态重建的重要树种, 以其独特的生理和形态特点适应着盐碱地生境(Armas et al ., 2010 ).在黄河三角洲泥质海岸盐碱地, 地下水位浅且矿化度高, 水分和盐分是影响该区域柽柳空间分布及其生态位的主要因素(贺强等, 2008 ; Cui et al ., 2009 ), 土壤水分、盐分、地下水水位及其矿化度是表征盐碱地水盐环境的主要指标, 而地下水通过“饱和带-包气带-植被”间的垂向联系由点及面产生极为重要的生态环境效应(Singh et al ., 2010 ; Gou & Miller, 2014 ; 夏江宝等, 2015 ), 特别是地下水矿化度的不同, 是导致土壤盐分和水分变化的主要因素.地下水通过影响土壤水分和盐分变化, 进而影响与植物光合生产力形成有关的物质运输和能量平衡(Álvarez & Sánchez-Blanco, 2014 ), 从而对植物生长及其光合生理生态过程产生重要影响.植物光合特性和树干液流速率可较好地反映植物生长及其水分利用对逆境条件的适应能力(夏江宝等, 2014 ; Xia et al ., 2015 ).植物光合效率能够表征植物在一定环境下的健康状况和光合生产能力, 可用来预测和表征植物对逆境的适应性和可塑性(Zhang et al ., 2014 ; Sanchez et al ., 2015 ).树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 ).目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
... 气孔限制和非气孔限制是抑制光合作用下降的主要原因(Farquhar & Sharkey, 1982 ).王伟华等(2009)研究发现, 盐水浓度4 g∙L-1 处理下多枝柽柳光合作用受到促进, 其他盐水浓度(12、20 和28 g∙L-1 )处理下, 多枝柽柳植株光合能力下降受气孔限制影响, 并且灌溉盐水的浓度越大气孔限制越强烈.本研究也发现类似规律, 在盐水和淡水处理下, 柽柳P n 下降的原因分别是气孔限制和非气孔限制.但也有研究发现, 随地下水矿化度增大, 多枝柽柳幼苗P n 下降受非气孔限制影响(王鹏等, 2012 ).可见, 地下水水位及其矿化度以及土壤介质内盐胁迫处理方式不同时, 随盐分胁迫的增加, 柽柳叶片光合作用下降的气孔限制机理有一定差异.淡水至咸水处理下, 土壤含盐量和溶液绝对浓度分别为0.11%-1.05%, 0.64%-5.38%, 柽柳的泌盐特性可以缓解盐分胁迫对柽柳生长的影响; 咸水至盐水处理下, 土壤含盐量和溶液绝对浓度分别为1.05%- 1.17%, 5.38%-6.70%, 过高的盐胁迫降低了柽柳的光合作用, 柽柳为保证自身水分利用和避免光合机构受损而关闭气孔. ...
... 树干液流速率是反映植物液流瞬时耗水规律的主要参数, 不但能够反映植物生理用水对环境因子的响应过程和规律, 也是确定树体储存水对蒸腾耗水贡献程度的主要指标(金鹰等, 2011 ; Wang et al ., 2012 ).光照和VPD 是影响树木蒸腾耗水的主要驱动因子(倪广艳等, 2015 ; 徐世琴等, 2015 ), 相关研究发现, 荷木(Schima superba )蒸腾在干、湿季均与PAR 和VPD 显著正相关(倪广艳等, 2015 ); 梭梭(Haloxylon ammodendron )液流密度日变化过程受PAR 、温度和VPD 的共同影响呈多峰特征(徐世琴等, 2015 ).本研究发现, 地下水矿化度会显著影响柽柳树干液流速率的日动态, 柽柳T r 和液流速率日动态随PAR 和VPD 的变化呈现单峰或双峰特征, 在咸水和微咸水处理下, 柽柳叶片气孔调节和蒸腾耗水受光照、空气温湿度等环境因子的协同作用影响较大, 通过“午休”有效调节了光合生理过程, 利于其对水盐的有效利用; 而淡水和盐水处理下, 环境因子的协同效应对其蒸腾耗水过程影响相对较小, 气孔调节相对较弱.可见随地下水矿化度的不同, 影响柽柳蒸腾耗水日动态的诸多环境因子的协同效应有较大差异.在土壤盐分为1.05%的咸水矿化度下, 柽柳T r 和日液流量最大, 而地下水矿化度过高或过低均会导致柽柳树干液流速率日变幅和日耗水量降低, 适度提高地下水矿化度会增加柽柳树干的液流速率和日作用时间.已有研究表明: 柽柳蒸腾速率随盐胁迫增加而降低(米文精等, 2011 ); 微咸水和咸水处理下, 柽柳T r 高于淡水处理(王鹏等, 2012 ).可见, 地下水矿化度与土壤盐分胁迫可显著影响树木耗水特性, 适度提高地下水矿化度可增强柽柳的蒸腾生理活性, 这可能是因为作为泌盐植物的柽柳, 适度土壤盐分起到类似营养盐的作用, 在水分不受限制的条件下, 盐分增高导致其蒸腾耗水有升高趋势.但随咸水至盐水处理下盐分渗透胁迫增强, 进入植物体内的水分减少, 蒸腾作用和液流速率显著降低.过高的地下水矿化度下, 柽柳苗木表现出通过关闭气孔降低蒸腾耗水来维持自身水分有效利用的生理策略.植物能否较好地协调碳同化与水分耗散之间的关系是辨识植物生长适应逆境胁迫的主要方法, 水分利用效率是判定植物生长适应逆境生存与否的关键因子(曹生奎等, 2009 ).本研究发现: 淡水和盐水处理下, 柽柳WUE 分别达最高和最低, 地下水矿化度的增加可显著降低柽柳WUE, 但地下水矿化度对柽柳WUE 的影响不如P n 和T r 明显, 特别是咸水和微咸水处理下, 柽柳WUE 无显著差异(p > 0.05), 这可能是柽柳通过调节自身生理状况以适应不同地下水矿化度和土壤盐分环境的水分利用策略.而已有研究表明: 柽柳WUE 随盐胁迫的加重有所增加(刘玉娟等, 2015 ); 咸水处理下多枝柽柳WUE 始终大于微咸水和淡水处理(王鹏等, 2012 ).可见, 随盐分处理方式的不同, 柽柳WUE 对盐胁迫引起的渗透胁迫表现出一定差异, 这主要与柽柳P n 和T r 对盐胁迫的响应不一致有关.在微咸水和咸水处理下, 柽柳P n 、T r 和液流活动均保持在较高状态, 生理活动旺盛, 柽柳通过调节光合特性和日液流量的变化来适应盐胁迫, 能保持高的水分利用效率; 盐水处理下, 柽柳P n 最低, 但蒸腾耗水并非最低, 导致WUE 最低; 淡水处理下由于T r 对地下水矿化度响应更加敏感, T r 下降幅度高于P n , 导致WUE 最高, 但此时P n 和T r 均较低, 液流活动微弱, 易导致蒸腾耗水过低而使叶片温度升高, 长时间的这种生理过程对植物正常的生理活动不利. ...
... 始终大于微咸水和淡水处理(王鹏等, 2012 ).可见, 随盐分处理方式的不同, 柽柳WUE 对盐胁迫引起的渗透胁迫表现出一定差异, 这主要与柽柳P n 和T r 对盐胁迫的响应不一致有关.在微咸水和咸水处理下, 柽柳P n 、T r 和液流活动均保持在较高状态, 生理活动旺盛, 柽柳通过调节光合特性和日液流量的变化来适应盐胁迫, 能保持高的水分利用效率; 盐水处理下, 柽柳P n 最低, 但蒸腾耗水并非最低, 导致WUE 最低; 淡水处理下由于T r 对地下水矿化度响应更加敏感, T r 下降幅度高于P n , 导致WUE 最高, 但此时P n 和T r 均较低, 液流活动微弱, 易导致蒸腾耗水过低而使叶片温度升高, 长时间的这种生理过程对植物正常的生理活动不利. ...
Effects of salt and water stress on plant biomass and photosynthetic characteristics of
1
2011
... 柽柳(Tamarix chinensis )属于典型的泌盐植物, 叶片和嫩枝可以将吸收于植物体内的盐分排出, 是黄河三角洲盐碱地植被恢复与生态重建的重要树种, 以其独特的生理和形态特点适应着盐碱地生境(Armas et al ., 2010 ).在黄河三角洲泥质海岸盐碱地, 地下水位浅且矿化度高, 水分和盐分是影响该区域柽柳空间分布及其生态位的主要因素(贺强等, 2008 ; Cui et al ., 2009 ), 土壤水分、盐分、地下水水位及其矿化度是表征盐碱地水盐环境的主要指标, 而地下水通过“饱和带-包气带-植被”间的垂向联系由点及面产生极为重要的生态环境效应(Singh et al ., 2010 ; Gou & Miller, 2014 ; 夏江宝等, 2015 ), 特别是地下水矿化度的不同, 是导致土壤盐分和水分变化的主要因素.地下水通过影响土壤水分和盐分变化, 进而影响与植物光合生产力形成有关的物质运输和能量平衡(Álvarez & Sánchez-Blanco, 2014 ), 从而对植物生长及其光合生理生态过程产生重要影响.植物光合特性和树干液流速率可较好地反映植物生长及其水分利用对逆境条件的适应能力(夏江宝等, 2014 ; Xia et al ., 2015 ).植物光合效率能够表征植物在一定环境下的健康状况和光合生产能力, 可用来预测和表征植物对逆境的适应性和可塑性(Zhang et al ., 2014 ; Sanchez et al ., 2015 ).树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 ).目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
盐处理对多枝柽柳光合作用和渗调物质的影响. 干旱区研究,
2009
黄河三角洲典型地区春季土壤水盐空间分异特征研究——以垦利县为例. 农业资源与环境学报,
1
2015
... 柽柳(Tamarix chinensis )属于典型的泌盐植物, 叶片和嫩枝可以将吸收于植物体内的盐分排出, 是黄河三角洲盐碱地植被恢复与生态重建的重要树种, 以其独特的生理和形态特点适应着盐碱地生境(Armas et al ., 2010 ).在黄河三角洲泥质海岸盐碱地, 地下水位浅且矿化度高, 水分和盐分是影响该区域柽柳空间分布及其生态位的主要因素(贺强等, 2008 ; Cui et al ., 2009 ), 土壤水分、盐分、地下水水位及其矿化度是表征盐碱地水盐环境的主要指标, 而地下水通过“饱和带-包气带-植被”间的垂向联系由点及面产生极为重要的生态环境效应(Singh et al ., 2010 ; Gou & Miller, 2014 ; 夏江宝等, 2015 ), 特别是地下水矿化度的不同, 是导致土壤盐分和水分变化的主要因素.地下水通过影响土壤水分和盐分变化, 进而影响与植物光合生产力形成有关的物质运输和能量平衡(Álvarez & Sánchez-Blanco, 2014 ), 从而对植物生长及其光合生理生态过程产生重要影响.植物光合特性和树干液流速率可较好地反映植物生长及其水分利用对逆境条件的适应能力(夏江宝等, 2014 ; Xia et al ., 2015 ).植物光合效率能够表征植物在一定环境下的健康状况和光合生产能力, 可用来预测和表征植物对逆境的适应性和可塑性(Zhang et al ., 2014 ; Sanchez et al ., 2015 ).树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 ).目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
Stomatal conductance correlates with photosynthetic capacity
1
1979
... 应用LI-6400XT便携式光合作用仪(LI-COR, Lincoln, USA)测定不同地下水矿化度处理下柽柳叶片的光响应曲线, 测定在晴朗天气的8:30-11:30进行, 使用缓冲瓶维持CO2 浓度的稳定性((390 ± 5) μmol∙mol-1 ), 设定人工光源的光合有效辐射(PAR )分别为: 1400、1200、1000、800、500、300、200、100、50、20、0 μmol∙m-2 ∙s-1 , 记录柽柳叶片的净光合速率(P n )、蒸腾速率(T r )、气孔导度(G s )、胞间CO2 浓度(C i )和大气CO2 浓度(C a )等参数.叶片瞬时水分利用效率(WUE )为P n 与T r 的比值, 即WUE = P n /T r (Nijs et al ., 1997 ); 气孔限制值(L s )为C i 与C a 的比值, 即L s = 1 - C i /C a (Wong et al ., 1979 ).同时在7:00- 19:00用LI-6400XT光合仪进行柽柳光合参数日动态的测定, 仪器自动记录T r 、G s 、PAR 和水汽压亏缺(VPD )等参数, 每2 h测定1次.以上测定每个处理测定3株苗木, 每株选取中上部生长健壮的成熟叶片2-3个, 在具体测定时, 采用不同处理、不同植株和叶片重复之间的交替测定法, 因此, 每株苗木依据测定的叶片数可测定2-3次, 以尽量避免时间波动对光合参数的影响.由于柽柳叶片形状不规则, 为得到精确的测量数据, 测定时需将标记的叶片平铺于整个测定叶室内, 同时参考邓雄(2003)和闫海龙(2010)等对不规则植物叶片光合参数的校正方法, 将观测的植物叶片剪下, 用扫描仪扫描后, 使用面积分析软件Delta-T Scan (CB50EJ, Cambridge, UK)计算实际的叶表面积, 按计算后的实际光合有效面积重新测算光合参数. ...
黄河三角洲盐生植被与土壤盐分的相关性研究. 植物生态学报,
1
1994
... 本研究在山东省黄河三角洲生态环境重点实验室的科研温室(37.38° N, 117.98° E)内进行, 栽植柽柳的土壤取自黄河三角洲的黄河滩地, 土壤质地为粉砂壤土, 其初始pH值为7.54, 平均含盐量为0.1%, 田间持水量为37.86%, 土壤容重为1.32 g·cm-3 .黄河三角洲泥质海岸带土壤为氯化物型盐土, NaCl占全盐含量的70%以上, 是土壤中最主要的盐分来源(吴志芬等, 1994 ; 曹建荣等, 2014 ), 因此, 为模拟设置该区域的地下水矿化度, 地下水以黄河三角洲NaCl为主的海盐自行配置, 主要依据天然水矿化度的数值分类标准, 称取各地下水矿化度模拟设置数值对应换算的海盐量, 溶解于特定容积的水桶中, 然后将配置好的不同矿化度水按照模拟设计水位分别浇灌于设计排列好的水桶中(图1 ).柽柳苗木栽植时采用三年生实生苗, 茎基部直径平均为(14.0 ± 0.5) mm, 苗高统一截杆至50 mm. ...
Critical effects of gas exchange parameters in
1
2015
... 柽柳(Tamarix chinensis )属于典型的泌盐植物, 叶片和嫩枝可以将吸收于植物体内的盐分排出, 是黄河三角洲盐碱地植被恢复与生态重建的重要树种, 以其独特的生理和形态特点适应着盐碱地生境(Armas et al ., 2010 ).在黄河三角洲泥质海岸盐碱地, 地下水位浅且矿化度高, 水分和盐分是影响该区域柽柳空间分布及其生态位的主要因素(贺强等, 2008 ; Cui et al ., 2009 ), 土壤水分、盐分、地下水水位及其矿化度是表征盐碱地水盐环境的主要指标, 而地下水通过“饱和带-包气带-植被”间的垂向联系由点及面产生极为重要的生态环境效应(Singh et al ., 2010 ; Gou & Miller, 2014 ; 夏江宝等, 2015 ), 特别是地下水矿化度的不同, 是导致土壤盐分和水分变化的主要因素.地下水通过影响土壤水分和盐分变化, 进而影响与植物光合生产力形成有关的物质运输和能量平衡(Álvarez & Sánchez-Blanco, 2014 ), 从而对植物生长及其光合生理生态过程产生重要影响.植物光合特性和树干液流速率可较好地反映植物生长及其水分利用对逆境条件的适应能力(夏江宝等, 2014 ; Xia et al ., 2015 ).植物光合效率能够表征植物在一定环境下的健康状况和光合生产能力, 可用来预测和表征植物对逆境的适应性和可塑性(Zhang et al ., 2014 ; Sanchez et al ., 2015 ).树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 ).目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
贝壳堤岛酸枣树干液流及光合参数对土壤水分的响应特征. 林业科学,
3
2014
... 柽柳(Tamarix chinensis )属于典型的泌盐植物, 叶片和嫩枝可以将吸收于植物体内的盐分排出, 是黄河三角洲盐碱地植被恢复与生态重建的重要树种, 以其独特的生理和形态特点适应着盐碱地生境(Armas et al ., 2010 ).在黄河三角洲泥质海岸盐碱地, 地下水位浅且矿化度高, 水分和盐分是影响该区域柽柳空间分布及其生态位的主要因素(贺强等, 2008 ; Cui et al ., 2009 ), 土壤水分、盐分、地下水水位及其矿化度是表征盐碱地水盐环境的主要指标, 而地下水通过“饱和带-包气带-植被”间的垂向联系由点及面产生极为重要的生态环境效应(Singh et al ., 2010 ; Gou & Miller, 2014 ; 夏江宝等, 2015 ), 特别是地下水矿化度的不同, 是导致土壤盐分和水分变化的主要因素.地下水通过影响土壤水分和盐分变化, 进而影响与植物光合生产力形成有关的物质运输和能量平衡(Álvarez & Sánchez-Blanco, 2014 ), 从而对植物生长及其光合生理生态过程产生重要影响.植物光合特性和树干液流速率可较好地反映植物生长及其水分利用对逆境条件的适应能力(夏江宝等, 2014 ; Xia et al ., 2015 ).植物光合效率能够表征植物在一定环境下的健康状况和光合生产能力, 可用来预测和表征植物对逆境的适应性和可塑性(Zhang et al ., 2014 ; Sanchez et al ., 2015 ).树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 ).目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
... ).树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 ).目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
... ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
不同潜水埋深下土壤水盐运移特征及其交互效应. 农业工程学报,
2
2015
... 柽柳(Tamarix chinensis )属于典型的泌盐植物, 叶片和嫩枝可以将吸收于植物体内的盐分排出, 是黄河三角洲盐碱地植被恢复与生态重建的重要树种, 以其独特的生理和形态特点适应着盐碱地生境(Armas et al ., 2010 ).在黄河三角洲泥质海岸盐碱地, 地下水位浅且矿化度高, 水分和盐分是影响该区域柽柳空间分布及其生态位的主要因素(贺强等, 2008 ; Cui et al ., 2009 ), 土壤水分、盐分、地下水水位及其矿化度是表征盐碱地水盐环境的主要指标, 而地下水通过“饱和带-包气带-植被”间的垂向联系由点及面产生极为重要的生态环境效应(Singh et al ., 2010 ; Gou & Miller, 2014 ; 夏江宝等, 2015 ), 特别是地下水矿化度的不同, 是导致土壤盐分和水分变化的主要因素.地下水通过影响土壤水分和盐分变化, 进而影响与植物光合生产力形成有关的物质运输和能量平衡(Álvarez & Sánchez-Blanco, 2014 ), 从而对植物生长及其光合生理生态过程产生重要影响.植物光合特性和树干液流速率可较好地反映植物生长及其水分利用对逆境条件的适应能力(夏江宝等, 2014 ; Xia et al ., 2015 ).植物光合效率能够表征植物在一定环境下的健康状况和光合生产能力, 可用来预测和表征植物对逆境的适应性和可塑性(Zhang et al ., 2014 ; Sanchez et al ., 2015 ).树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 ).目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
... 黄河三角洲柽柳生长地带地下水位主要集中在300-2000 mm (马玉蕾等, 2013 ; 夏江宝等, 2015 ), 结合在科研温室内利于实验控制, 因此本研究以泥质海岸带柽柳大面积生长的地下水水位1200 mm为准, 模拟设置淡水、微咸水、咸水和盐水4种地下水矿化度, 其对应矿化度数值分别为0 g∙L-1 、3.00 g∙L-1 、8.00 g∙L-1 和20.00 g∙L-1 .2014年3月5日将柽柳栽植于直径为300 mm、高度为1780 mm的PVC圆管中, 其中PVC管实际淹水区为550 mm, 管底部设有纱网, 防止土壤渗漏.将PVC圆管放置于700 mm高, 桶底内径455 mm, 桶上口内径570 mm的水桶中, 为使地下水温度一致, 将水桶埋置于土壤中.不同地下水矿化度下栽植柽柳的土柱模拟示意图和实景图见图1 .为维持地下水水位及其矿化度的稳定性, 定期监测地下水盐度, 依次来补充所需盐分.柽柳苗木正常生长1月后对其进行不同地下水矿化度处理, 在此期间从地上部共浇水12.00 L, 分4次灌溉, 地下水矿化度处理后, 不再进行水分灌溉等措施.每种地下水矿化度处理下3个重复, 共计12个栽植柽柳的土壤柱体, 每土壤柱体栽植柽柳1株.在地下水矿化度处理80天后, 苗木平均高度为(67 ± 0.6) mm, 茎基部直径平均为(15.0 ± 0.7) mm, 6月27日开始柽柳苗木光合作用和树干液流的测定, 7月6日测定结束. ...
典型固沙植物梭梭生长季蒸腾变化及其对环境因子的响应. 植物生态学报,
2
2015
... 树干液流速率是反映植物液流瞬时耗水规律的主要参数, 不但能够反映植物生理用水对环境因子的响应过程和规律, 也是确定树体储存水对蒸腾耗水贡献程度的主要指标(金鹰等, 2011 ; Wang et al ., 2012 ).光照和VPD 是影响树木蒸腾耗水的主要驱动因子(倪广艳等, 2015 ; 徐世琴等, 2015 ), 相关研究发现, 荷木(Schima superba )蒸腾在干、湿季均与PAR 和VPD 显著正相关(倪广艳等, 2015 ); 梭梭(Haloxylon ammodendron )液流密度日变化过程受PAR 、温度和VPD 的共同影响呈多峰特征(徐世琴等, 2015 ).本研究发现, 地下水矿化度会显著影响柽柳树干液流速率的日动态, 柽柳T r 和液流速率日动态随PAR 和VPD 的变化呈现单峰或双峰特征, 在咸水和微咸水处理下, 柽柳叶片气孔调节和蒸腾耗水受光照、空气温湿度等环境因子的协同作用影响较大, 通过“午休”有效调节了光合生理过程, 利于其对水盐的有效利用; 而淡水和盐水处理下, 环境因子的协同效应对其蒸腾耗水过程影响相对较小, 气孔调节相对较弱.可见随地下水矿化度的不同, 影响柽柳蒸腾耗水日动态的诸多环境因子的协同效应有较大差异.在土壤盐分为1.05%的咸水矿化度下, 柽柳T r 和日液流量最大, 而地下水矿化度过高或过低均会导致柽柳树干液流速率日变幅和日耗水量降低, 适度提高地下水矿化度会增加柽柳树干的液流速率和日作用时间.已有研究表明: 柽柳蒸腾速率随盐胁迫增加而降低(米文精等, 2011 ); 微咸水和咸水处理下, 柽柳T r 高于淡水处理(王鹏等, 2012 ).可见, 地下水矿化度与土壤盐分胁迫可显著影响树木耗水特性, 适度提高地下水矿化度可增强柽柳的蒸腾生理活性, 这可能是因为作为泌盐植物的柽柳, 适度土壤盐分起到类似营养盐的作用, 在水分不受限制的条件下, 盐分增高导致其蒸腾耗水有升高趋势.但随咸水至盐水处理下盐分渗透胁迫增强, 进入植物体内的水分减少, 蒸腾作用和液流速率显著降低.过高的地下水矿化度下, 柽柳苗木表现出通过关闭气孔降低蒸腾耗水来维持自身水分有效利用的生理策略.植物能否较好地协调碳同化与水分耗散之间的关系是辨识植物生长适应逆境胁迫的主要方法, 水分利用效率是判定植物生长适应逆境生存与否的关键因子(曹生奎等, 2009 ).本研究发现: 淡水和盐水处理下, 柽柳WUE 分别达最高和最低, 地下水矿化度的增加可显著降低柽柳WUE, 但地下水矿化度对柽柳WUE 的影响不如P n 和T r 明显, 特别是咸水和微咸水处理下, 柽柳WUE 无显著差异(p > 0.05), 这可能是柽柳通过调节自身生理状况以适应不同地下水矿化度和土壤盐分环境的水分利用策略.而已有研究表明: 柽柳WUE 随盐胁迫的加重有所增加(刘玉娟等, 2015 ); 咸水处理下多枝柽柳WUE 始终大于微咸水和淡水处理(王鹏等, 2012 ).可见, 随盐分处理方式的不同, 柽柳WUE 对盐胁迫引起的渗透胁迫表现出一定差异, 这主要与柽柳P n 和T r 对盐胁迫的响应不一致有关.在微咸水和咸水处理下, 柽柳P n 、T r 和液流活动均保持在较高状态, 生理活动旺盛, 柽柳通过调节光合特性和日液流量的变化来适应盐胁迫, 能保持高的水分利用效率; 盐水处理下, 柽柳P n 最低, 但蒸腾耗水并非最低, 导致WUE 最低; 淡水处理下由于T r 对地下水矿化度响应更加敏感, T r 下降幅度高于P n , 导致WUE 最高, 但此时P n 和T r 均较低, 液流活动微弱, 易导致蒸腾耗水过低而使叶片温度升高, 长时间的这种生理过程对植物正常的生理活动不利. ...
... 的共同影响呈多峰特征(徐世琴等, 2015 ).本研究发现, 地下水矿化度会显著影响柽柳树干液流速率的日动态, 柽柳T r 和液流速率日动态随PAR 和VPD 的变化呈现单峰或双峰特征, 在咸水和微咸水处理下, 柽柳叶片气孔调节和蒸腾耗水受光照、空气温湿度等环境因子的协同作用影响较大, 通过“午休”有效调节了光合生理过程, 利于其对水盐的有效利用; 而淡水和盐水处理下, 环境因子的协同效应对其蒸腾耗水过程影响相对较小, 气孔调节相对较弱.可见随地下水矿化度的不同, 影响柽柳蒸腾耗水日动态的诸多环境因子的协同效应有较大差异.在土壤盐分为1.05%的咸水矿化度下, 柽柳T r 和日液流量最大, 而地下水矿化度过高或过低均会导致柽柳树干液流速率日变幅和日耗水量降低, 适度提高地下水矿化度会增加柽柳树干的液流速率和日作用时间.已有研究表明: 柽柳蒸腾速率随盐胁迫增加而降低(米文精等, 2011 ); 微咸水和咸水处理下, 柽柳T r 高于淡水处理(王鹏等, 2012 ).可见, 地下水矿化度与土壤盐分胁迫可显著影响树木耗水特性, 适度提高地下水矿化度可增强柽柳的蒸腾生理活性, 这可能是因为作为泌盐植物的柽柳, 适度土壤盐分起到类似营养盐的作用, 在水分不受限制的条件下, 盐分增高导致其蒸腾耗水有升高趋势.但随咸水至盐水处理下盐分渗透胁迫增强, 进入植物体内的水分减少, 蒸腾作用和液流速率显著降低.过高的地下水矿化度下, 柽柳苗木表现出通过关闭气孔降低蒸腾耗水来维持自身水分有效利用的生理策略.植物能否较好地协调碳同化与水分耗散之间的关系是辨识植物生长适应逆境胁迫的主要方法, 水分利用效率是判定植物生长适应逆境生存与否的关键因子(曹生奎等, 2009 ).本研究发现: 淡水和盐水处理下, 柽柳WUE 分别达最高和最低, 地下水矿化度的增加可显著降低柽柳WUE, 但地下水矿化度对柽柳WUE 的影响不如P n 和T r 明显, 特别是咸水和微咸水处理下, 柽柳WUE 无显著差异(p > 0.05), 这可能是柽柳通过调节自身生理状况以适应不同地下水矿化度和土壤盐分环境的水分利用策略.而已有研究表明: 柽柳WUE 随盐胁迫的加重有所增加(刘玉娟等, 2015 ); 咸水处理下多枝柽柳WUE 始终大于微咸水和淡水处理(王鹏等, 2012 ).可见, 随盐分处理方式的不同, 柽柳WUE 对盐胁迫引起的渗透胁迫表现出一定差异, 这主要与柽柳P n 和T r 对盐胁迫的响应不一致有关.在微咸水和咸水处理下, 柽柳P n 、T r 和液流活动均保持在较高状态, 生理活动旺盛, 柽柳通过调节光合特性和日液流量的变化来适应盐胁迫, 能保持高的水分利用效率; 盐水处理下, 柽柳P n 最低, 但蒸腾耗水并非最低, 导致WUE 最低; 淡水处理下由于T r 对地下水矿化度响应更加敏感, T r 下降幅度高于P n , 导致WUE 最高, 但此时P n 和T r 均较低, 液流活动微弱, 易导致蒸腾耗水过低而使叶片温度升高, 长时间的这种生理过程对植物正常的生理活动不利. ...
塔里木沙漠公路防护林3种植物光合特性对干旱胁迫的响应. 生态学报,
2010
A new model for relationship between irradiance and the rate of photosynthesis in
1
2007
... 直角双曲线修正模型公式(Ye, 2007 ): ...
Threshold effects of photosynthetic efficiency parameters of wild
1
2014
... 柽柳(Tamarix chinensis )属于典型的泌盐植物, 叶片和嫩枝可以将吸收于植物体内的盐分排出, 是黄河三角洲盐碱地植被恢复与生态重建的重要树种, 以其独特的生理和形态特点适应着盐碱地生境(Armas et al ., 2010 ).在黄河三角洲泥质海岸盐碱地, 地下水位浅且矿化度高, 水分和盐分是影响该区域柽柳空间分布及其生态位的主要因素(贺强等, 2008 ; Cui et al ., 2009 ), 土壤水分、盐分、地下水水位及其矿化度是表征盐碱地水盐环境的主要指标, 而地下水通过“饱和带-包气带-植被”间的垂向联系由点及面产生极为重要的生态环境效应(Singh et al ., 2010 ; Gou & Miller, 2014 ; 夏江宝等, 2015 ), 特别是地下水矿化度的不同, 是导致土壤盐分和水分变化的主要因素.地下水通过影响土壤水分和盐分变化, 进而影响与植物光合生产力形成有关的物质运输和能量平衡(Álvarez & Sánchez-Blanco, 2014 ), 从而对植物生长及其光合生理生态过程产生重要影响.植物光合特性和树干液流速率可较好地反映植物生长及其水分利用对逆境条件的适应能力(夏江宝等, 2014 ; Xia et al ., 2015 ).植物光合效率能够表征植物在一定环境下的健康状况和光合生产能力, 可用来预测和表征植物对逆境的适应性和可塑性(Zhang et al ., 2014 ; Sanchez et al ., 2015 ).树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 ).目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
树干液流研究进展与展望. 西北林学院学报,
1
2015
... 柽柳(Tamarix chinensis )属于典型的泌盐植物, 叶片和嫩枝可以将吸收于植物体内的盐分排出, 是黄河三角洲盐碱地植被恢复与生态重建的重要树种, 以其独特的生理和形态特点适应着盐碱地生境(Armas et al ., 2010 ).在黄河三角洲泥质海岸盐碱地, 地下水位浅且矿化度高, 水分和盐分是影响该区域柽柳空间分布及其生态位的主要因素(贺强等, 2008 ; Cui et al ., 2009 ), 土壤水分、盐分、地下水水位及其矿化度是表征盐碱地水盐环境的主要指标, 而地下水通过“饱和带-包气带-植被”间的垂向联系由点及面产生极为重要的生态环境效应(Singh et al ., 2010 ; Gou & Miller, 2014 ; 夏江宝等, 2015 ), 特别是地下水矿化度的不同, 是导致土壤盐分和水分变化的主要因素.地下水通过影响土壤水分和盐分变化, 进而影响与植物光合生产力形成有关的物质运输和能量平衡(Álvarez & Sánchez-Blanco, 2014 ), 从而对植物生长及其光合生理生态过程产生重要影响.植物光合特性和树干液流速率可较好地反映植物生长及其水分利用对逆境条件的适应能力(夏江宝等, 2014 ; Xia et al ., 2015 ).植物光合效率能够表征植物在一定环境下的健康状况和光合生产能力, 可用来预测和表征植物对逆境的适应性和可塑性(Zhang et al ., 2014 ; Sanchez et al ., 2015 ).树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 ).目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
Physiological responses of
1
2012
... 柽柳(Tamarix chinensis )属于典型的泌盐植物, 叶片和嫩枝可以将吸收于植物体内的盐分排出, 是黄河三角洲盐碱地植被恢复与生态重建的重要树种, 以其独特的生理和形态特点适应着盐碱地生境(Armas et al ., 2010 ).在黄河三角洲泥质海岸盐碱地, 地下水位浅且矿化度高, 水分和盐分是影响该区域柽柳空间分布及其生态位的主要因素(贺强等, 2008 ; Cui et al ., 2009 ), 土壤水分、盐分、地下水水位及其矿化度是表征盐碱地水盐环境的主要指标, 而地下水通过“饱和带-包气带-植被”间的垂向联系由点及面产生极为重要的生态环境效应(Singh et al ., 2010 ; Gou & Miller, 2014 ; 夏江宝等, 2015 ), 特别是地下水矿化度的不同, 是导致土壤盐分和水分变化的主要因素.地下水通过影响土壤水分和盐分变化, 进而影响与植物光合生产力形成有关的物质运输和能量平衡(Álvarez & Sánchez-Blanco, 2014 ), 从而对植物生长及其光合生理生态过程产生重要影响.植物光合特性和树干液流速率可较好地反映植物生长及其水分利用对逆境条件的适应能力(夏江宝等, 2014 ; Xia et al ., 2015 ).植物光合效率能够表征植物在一定环境下的健康状况和光合生产能力, 可用来预测和表征植物对逆境的适应性和可塑性(Zhang et al ., 2014 ; Sanchez et al ., 2015 ).树干液流可准确地反映植物的蒸腾耗水特性, 是土壤-植物-大气连续系统中的关键水分参数(夏江宝等, 2014 ; 赵春彦等, 2015 ).目前, 从水盐角度开展植物光合生理过程研究多集中在土壤这一介质层(Qin et al ., 2010 ; Glenn et al ., 2012 ), 主要以土壤水分或盐分变化对树木光合生理过程及树木耗水机理的影响等方面展开了相关研究(Wang et al ., 2011 ; 夏江宝等, 2014 ).从地下水利用的角度, 主要开展了地下水水位对典型旱生或盐生生境下不同树木的光合生理学和形态学的影响等方面的探讨.研究发现, 在浅层地下水条件下, 胡杨(Populus euphratica )和多枝柽柳(Tamarix ramosissima )可保持较好的生理活性, 而在干旱条件下, 2种树木生长缓慢, 叶面积指数显著降低(Li et al ., 2013 ); 地下水埋深小于2.7 m和大于5.2 m时, 土壤盐分和水分分别成为胡杨叶片气孔密度降低的主要因子(Zhao et al ., 2012 ); 地下水位下降会显著降低胡杨、榆树(Ulmus pumila )的光合性能和水分利用效率(Chen et al ., 2011 ; 苏华等, 2012 ); 当地下水位低于4.5 m时, 胡杨能够正常生长, 且土壤盐分不是影响胡杨液流变化的主要因子(马建新等, 2010 ; Chen et al ., 2011 ); 在地下水埋深20 cm、地下咸水矿化度3.00-10.00 g∙L-1 条件下, 多枝柽柳的净光合速率和水分利用效率最高(王鹏等, 2012 ).而对黄河三角洲柽柳与地下水关系的研究主要集中在水盐生境对柽柳空间分布格局(贺强等, 2007 ; 王卓然等, 2015 )、适宜的生态水位(荣丽杉等, 2010 )以及植被生态效应(安乐生等, 2011 ; Cao et al ., 2014 )等方面.上述研究较少涉及地下水矿化度对柽柳光合生理生态过程及耗水特征等方面的探讨, 致使适宜柽柳生长的地下水矿化度条件及其水盐适应性尚不清晰, 这在一定程度上影响了柽柳幼苗的适地造林和水盐管理措施.鉴于此, 为明确适宜柽柳苗木生长的水盐生境, 本研究模拟设计4种地下水矿化度, 测定分析不同地下水矿化度下柽柳叶片光合光响应参数和蒸腾耗水日动态, 以期阐明柽柳光合效率和树木耗水对地下水矿化度的响应规律, 研究结果可为黄河三角洲地下水浅埋区柽柳的栽培管理和适宜水盐生境选择提供理论依据和技术参考. ...
盐胁迫对中国柽柳幼苗生理特性的影响. 生态学报,
2
2015
... 水分是盐分运移的载体, 潜水蒸发作用下, 浅埋深地下水通过毛管作用进入包气带土壤层, 致使土壤水盐发生变化, 进而影响植物根系生长和叶片的光合作用(宫兆宁等, 2006 ; Gou & Miller, 2014 ).本研究发现相同潜水水位下, 土壤盐分和溶液绝对浓度差异较大成为影响柽柳苗木光合作用的主要因素.王鹏等(2012)研究发现, 多枝柽柳幼苗P n 随地下水矿化度增大而升高, 咸水处理下P n 最高, 光照生态幅最宽, 微咸水处理下AQY 和R d 最高; 而本研究发现, 随地下水矿化度增大, 柽柳P n 先升高后降低, 咸水处理下P n 、P nmax 、AQY 和R d 均达到最大值, 微咸水处理下光照生态幅最宽.王伟华等(2009)研究发现, 多枝柽柳P n 日平均值随盐水浓度的增加先升高后降低, 盐水处理下P n 日平均值达最高.可见, 淡水处理下柽柳并非有最好的光合性能, 而适度提高地下水至咸水矿化度会促进柽柳苗木的光合特性, 应该与盐生植物柽柳的泌盐生理特性和较强的耐盐性有关, 这可能是柽柳长期适应盐碱地生境的一种竞争策略.地下水矿化度的增加使土壤含盐量增加, 柽柳苗木通过增强叶片的光能转化效率、R d 和光照生态幅来提高光合作用以适应盐胁迫, 并且抗氧化物酶系统可清除体内因盐分过多而产生的活性氧, 保持体内活性氧的动态平衡(苏华等, 2012 ; 朱金方等, 2012 , 2015 ), 这可能是咸水处理下柽柳光合能力显著提高的主要原因.柽柳的泌盐特性通过泌盐腺将体内过多的Na+ 、Cl- 以盐粒结晶的形式排出体外, 同时还将其他有害微量元素进行离子区隔化, 可溶性糖、脯氨酸等可维持细胞渗透压, 能使柽柳在高盐度环境下生长(Scholander et al ., 1996 ; Anderson et al ., 2005 ).但盐水处理下, 土壤含盐量和溶液绝对浓度均达最高, 致使土壤渗透势升高, 根系吸水困难, 导致柽柳AQY 、R d 和P nmax 降低, 光照生态幅变窄.李紫薇等(2014)也发现, 重度盐分胁迫使蒺藜苜蓿(Medicago truncatula )叶片光能转化效率降低, AQY 降低进而导致P nmax 、LSP 降低, LCP 升高, 光照生态幅变窄, R d 降低, 本研究结果与之类似.这主要是因为柽柳对Na+ 、Cl- 的吸收, 排斥了对另一些营养元素的吸收, 幼苗生长受到威胁; 同时重度盐分胁迫促进了叶绿素酶对叶绿素的分解使叶绿素合成系统受到破坏, 活性氧自由基超过超氧化物歧化酶清除量, 细胞受损严重(朱金方等, 2012 , 2015 ), 渗透调节物质的功能显著降低(朱金方等, 2013 ). ...
... , 2015 ), 渗透调节物质的功能显著降低(朱金方等, 2013 ). ...
盐旱交叉胁迫对柽柳幼苗渗透调节物质含量的影响. 西北植物学报,
1
2013
... 水分是盐分运移的载体, 潜水蒸发作用下, 浅埋深地下水通过毛管作用进入包气带土壤层, 致使土壤水盐发生变化, 进而影响植物根系生长和叶片的光合作用(宫兆宁等, 2006 ; Gou & Miller, 2014 ).本研究发现相同潜水水位下, 土壤盐分和溶液绝对浓度差异较大成为影响柽柳苗木光合作用的主要因素.王鹏等(2012)研究发现, 多枝柽柳幼苗P n 随地下水矿化度增大而升高, 咸水处理下P n 最高, 光照生态幅最宽, 微咸水处理下AQY 和R d 最高; 而本研究发现, 随地下水矿化度增大, 柽柳P n 先升高后降低, 咸水处理下P n 、P nmax 、AQY 和R d 均达到最大值, 微咸水处理下光照生态幅最宽.王伟华等(2009)研究发现, 多枝柽柳P n 日平均值随盐水浓度的增加先升高后降低, 盐水处理下P n 日平均值达最高.可见, 淡水处理下柽柳并非有最好的光合性能, 而适度提高地下水至咸水矿化度会促进柽柳苗木的光合特性, 应该与盐生植物柽柳的泌盐生理特性和较强的耐盐性有关, 这可能是柽柳长期适应盐碱地生境的一种竞争策略.地下水矿化度的增加使土壤含盐量增加, 柽柳苗木通过增强叶片的光能转化效率、R d 和光照生态幅来提高光合作用以适应盐胁迫, 并且抗氧化物酶系统可清除体内因盐分过多而产生的活性氧, 保持体内活性氧的动态平衡(苏华等, 2012 ; 朱金方等, 2012 , 2015 ), 这可能是咸水处理下柽柳光合能力显著提高的主要原因.柽柳的泌盐特性通过泌盐腺将体内过多的Na+ 、Cl- 以盐粒结晶的形式排出体外, 同时还将其他有害微量元素进行离子区隔化, 可溶性糖、脯氨酸等可维持细胞渗透压, 能使柽柳在高盐度环境下生长(Scholander et al ., 1996 ; Anderson et al ., 2005 ).但盐水处理下, 土壤含盐量和溶液绝对浓度均达最高, 致使土壤渗透势升高, 根系吸水困难, 导致柽柳AQY 、R d 和P nmax 降低, 光照生态幅变窄.李紫薇等(2014)也发现, 重度盐分胁迫使蒺藜苜蓿(Medicago truncatula )叶片光能转化效率降低, AQY 降低进而导致P nmax 、LSP 降低, LCP 升高, 光照生态幅变窄, R d 降低, 本研究结果与之类似.这主要是因为柽柳对Na+ 、Cl- 的吸收, 排斥了对另一些营养元素的吸收, 幼苗生长受到威胁; 同时重度盐分胁迫促进了叶绿素酶对叶绿素的分解使叶绿素合成系统受到破坏, 活性氧自由基超过超氧化物歧化酶清除量, 细胞受损严重(朱金方等, 2012 , 2015 ), 渗透调节物质的功能显著降低(朱金方等, 2013 ). ...
盐旱交叉胁迫对柽柳幼苗生长及生理生化特性的影响. 西北植物学报,
2
2012
... 水分是盐分运移的载体, 潜水蒸发作用下, 浅埋深地下水通过毛管作用进入包气带土壤层, 致使土壤水盐发生变化, 进而影响植物根系生长和叶片的光合作用(宫兆宁等, 2006 ; Gou & Miller, 2014 ).本研究发现相同潜水水位下, 土壤盐分和溶液绝对浓度差异较大成为影响柽柳苗木光合作用的主要因素.王鹏等(2012)研究发现, 多枝柽柳幼苗P n 随地下水矿化度增大而升高, 咸水处理下P n 最高, 光照生态幅最宽, 微咸水处理下AQY 和R d 最高; 而本研究发现, 随地下水矿化度增大, 柽柳P n 先升高后降低, 咸水处理下P n 、P nmax 、AQY 和R d 均达到最大值, 微咸水处理下光照生态幅最宽.王伟华等(2009)研究发现, 多枝柽柳P n 日平均值随盐水浓度的增加先升高后降低, 盐水处理下P n 日平均值达最高.可见, 淡水处理下柽柳并非有最好的光合性能, 而适度提高地下水至咸水矿化度会促进柽柳苗木的光合特性, 应该与盐生植物柽柳的泌盐生理特性和较强的耐盐性有关, 这可能是柽柳长期适应盐碱地生境的一种竞争策略.地下水矿化度的增加使土壤含盐量增加, 柽柳苗木通过增强叶片的光能转化效率、R d 和光照生态幅来提高光合作用以适应盐胁迫, 并且抗氧化物酶系统可清除体内因盐分过多而产生的活性氧, 保持体内活性氧的动态平衡(苏华等, 2012 ; 朱金方等, 2012 , 2015 ), 这可能是咸水处理下柽柳光合能力显著提高的主要原因.柽柳的泌盐特性通过泌盐腺将体内过多的Na+ 、Cl- 以盐粒结晶的形式排出体外, 同时还将其他有害微量元素进行离子区隔化, 可溶性糖、脯氨酸等可维持细胞渗透压, 能使柽柳在高盐度环境下生长(Scholander et al ., 1996 ; Anderson et al ., 2005 ).但盐水处理下, 土壤含盐量和溶液绝对浓度均达最高, 致使土壤渗透势升高, 根系吸水困难, 导致柽柳AQY 、R d 和P nmax 降低, 光照生态幅变窄.李紫薇等(2014)也发现, 重度盐分胁迫使蒺藜苜蓿(Medicago truncatula )叶片光能转化效率降低, AQY 降低进而导致P nmax 、LSP 降低, LCP 升高, 光照生态幅变窄, R d 降低, 本研究结果与之类似.这主要是因为柽柳对Na+ 、Cl- 的吸收, 排斥了对另一些营养元素的吸收, 幼苗生长受到威胁; 同时重度盐分胁迫促进了叶绿素酶对叶绿素的分解使叶绿素合成系统受到破坏, 活性氧自由基超过超氧化物歧化酶清除量, 细胞受损严重(朱金方等, 2012 , 2015 ), 渗透调节物质的功能显著降低(朱金方等, 2013 ). ...
... 的吸收, 排斥了对另一些营养元素的吸收, 幼苗生长受到威胁; 同时重度盐分胁迫促进了叶绿素酶对叶绿素的分解使叶绿素合成系统受到破坏, 活性氧自由基超过超氧化物歧化酶清除量, 细胞受损严重(朱金方等, 2012 , 2015 ), 渗透调节物质的功能显著降低(朱金方等, 2013 ). ...
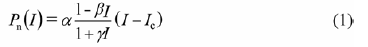


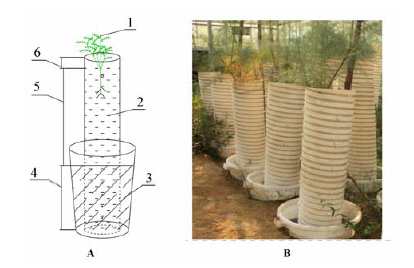
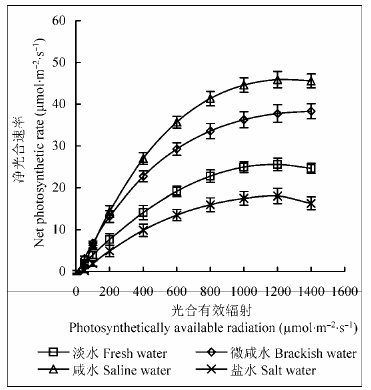
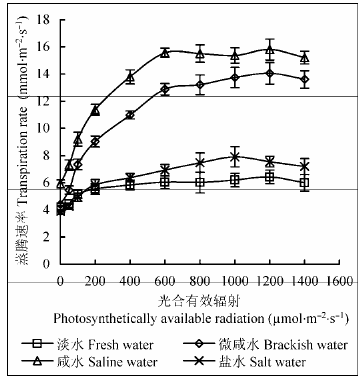
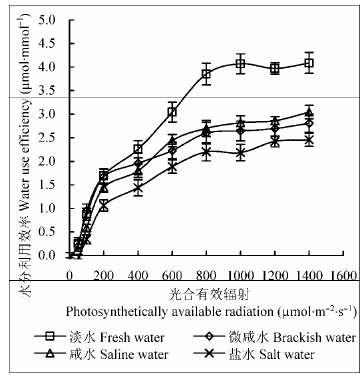
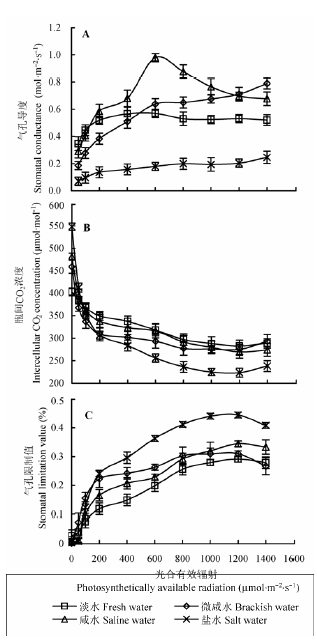
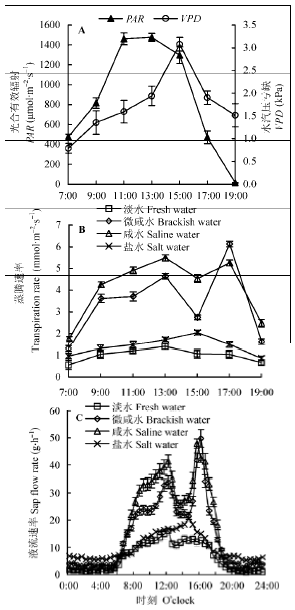
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}