竹子(竹亚科植物的总称)生长快速、材质优良, 被广泛用于建材与家居; 竹笋味道鲜美、营养丰富, 备受人们青睐(江泽慧, 2007 ; Chongtham et al ., 2011 )。仅2001年全球竹材消费就高达3000万t、竹笋消费200万t (Kleinhenz et al ., 2000 ; Kleinhenz & Midmore, 2001 ), 年贸易额高达45亿美元(Kleinhenz & Midmore, 2001 )。我国竹林面积近600万hm2 , 产值1600亿元, 并呈逐年上升的趋势(董杰和张燕, 2015 )。竹产品需求持续增长, 大大促进了竹林发展, 但不合理经营导致了部分竹林衰退(楼一平等, 1997 )、产量下降(孟赐福等, 2009 )、地力退化(楼一平, 2001 ; Guo et al ., 2014 )、环境污染(吴家森等, 2009 )等生产和生态问题。这些问题既与竹子自身的生长发育、开花结实等生物习性有关, 也与人类采收笋材、垦复施肥等经营活动相关。然而, 前人仅从竹子生物学、生态学和培育学等方面进行了零散的研究, 不利于以上问题的综合解决。因此, 竹子领域研究亟需一个统一化理论做指导。
生态化学计量学是从元素计量的角度来探讨生命运动的内在规律的理论(曾德慧和陈广生, 2005 ; 贺金生和韩兴国, 2010 )。它通过有机体内元素含量及比率关系, 将生物的生长发育、健康状况、行为方式、生态系统动态、生态环境保护等多层次、多学科问题联系起来, 现已成为生态学、环境科学和生物学领域最流行的理论框架(Sterner & Elser, 2002 ; Hillebrand et al ., 2014 )。在竹子经营与研究过程中, 虽然人们早已发现竹子开花(何奇江等, 2005 ; 丁兴萃, 2006 )、发笋大小年(费世民, 2011 )、竹笋品质形成(林海萍等, 2004 )与体内养分含量及比例有关, 竹林土壤污染主要是因为养分搭配不当和总量盈余(孟赐福等, 2009 ), 也提出了平衡施肥与养分管理技术(郭晓敏等, 2013 ), 但尚未形成一个从竹子基础生物学到生产经营的有关养分元素的统一化理论, 极大地影响了竹林生产和生态环境建设。
本文试从生态化学计量学角度, 总结近年竹子生物学、生理学、生态学、营养学、培育学等方面的研究成果, 重点分析竹子生命元素的组成及内稳性特征、元素化学计量与竹子生理生态学过程的关系、竹子化学计量平衡的主要内外影响因子、生态化学计量学在竹林养分诊断与平衡施肥领域的应用等竹子生理、生态与生产问题, 以期进一步丰富生态化学计量学内容、扩大生态化学计量学在生产实践中的应用, 也为竹林这一特殊森林的生产经营及环境保护提供理论指导, 实现我国“少投入、多产出、保环境”的战略构想。
1 生态化学计量学的概念与理论基础
生态化学计量学主要是从元素组成及比率的角度来研究生物与环境关系, 是近年生态学中飞速发展的领域。自1840年德国农业化学家Justus von Liebig发现植物生长与元素含量的关系并提出了“最小因子定律”以来, 经过生态位理论(Grubb, 1977 )、最佳取食理论(Belovsky, 1978 )、资源比理论(Tilman, 1982 )、养分利用理论(Vitousek, 1982 )、Redfield比值(Redfield, 1958 )等理论的发展, Reiners (1986)提出“Chemical Stoichiometry of the Biota”, 将化学计量学理论用于研究生态系统结构与功能, 到2002年, Sterner和Elser出版专著《Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere》, 系统地阐述了生态化学计量学的定义、基本理论及其应用范畴, 标志着生态化学计量学体系的逐渐成熟, 也为生物学统一化理论构建提供了新思路(Sterner & Elser, 2002 ; 曾德慧和陈广生, 2005 ; 贺金生和韩兴国, 2010 )。当前, 生态化学计量学已发展成从生物大分子到生态系统, 从微生物、动物到植物, 从营养学、生态学到环境科学多层次、多学科都相互关联的综合性理论, 并逐渐被应用于预测或解决区域乃至全球的生态环境问题(曾德慧和陈广生, 2005 )。其理论逻辑如下:
(1)生命是物质运动的最高级形式, 一切有机体都是由元素构成;
(2)生物有机体都有其特定的元素组成及其比率关系, 即元素计量内稳定性(stoichiometric homeostasis);
(3)有机体内元素平衡是相对的, 它易受外界环境(食物)影响而发生变化(甚至偏离);
(4)体内元素平衡变化或偏离会影响有机体的新陈代谢、生长发育、健康状况和行为方式(Schatz & McCauley, 2007 ; Rivas-Ubach et al ., 2012 );
(5)有机体具有维持元素内平衡的机制(hom- eostatic mechanism), 它可通过生理或行为等适应性调节, 在变化的环境(食物)中可以维持或恢复自身化学计量的相对平衡;
(6)不同有机体的化学计量内平衡能力有所差异, 能保持平衡者正常生长发育, 反之则异常, 甚至死亡;
(7)群体中有机体的此消彼长会影响到种群动态、群落演替和生态系统的发展及其生产、生态功能的发挥(Sterner & Elser, 2002 ; Güsewell, 2004 ; Yan et al ., 2015 )。
通过生命元素这一线索, 生态化学计量学将不同层次的生命活动统一起来, 也将不同生命学科统一起来, 并形成两个被人们广泛接受的基础理论(或假说)(Sterner & Elser, 2002 ; Anderson et al ., 2004 ), 即化学计量内稳性假说和生长速率假说。
化学计量内稳性假说(stoichiometric homeostasis hypothesis)是指在环境(食物)化学元素组成发生变化的情况下, 生物有机体可通过一系列生理或行为调节以保持其元素组成的相对稳定(Sterner & Elser, 2002 ; Elser et al ., 2010 )。Sterner和Elser (2002)提出有机体-环境化学计量关系公式: y = cx 1/ H y 、x 分别表示有机体、环境元素浓度(或比率), c 是常数, H 是化学计量内稳性指数, 反映有机体化学计量内稳性维持能力的大小。当H = 1时, 该有机体没有任何的化学计量内稳性; H > 1时, 有机体具有维持自身元素平衡的能力, H 越大有机体内稳性越高(Sterner & Elser, 2002 )。
生长速率假说(growth rate hypothesis)是解析有机体维持内平衡的机制性假说。该假说认为有机体生长速率与其体内元素化学计量比率紧密联系。高生长速率往往对应高N:C、P:C以及较低的N:P。因为有机体快速生长需要大量富含P的核糖体(蛋白质的合成场所)和RNA、DNA、ATP等物质(Elser et al ., 2003 ; Reef et al ., 2010 )。生长速率假说是解析细胞生物学、生物新陈代谢、种群动态和生态系统功能的理论框架(Niklas et al ., 2005 ; Ågren, 2008 )。
另外, 生态化学计量学特别重视生物与环境的矛盾统一, 认为生物元素内平衡需求与环境供应间的适配(match)与错配(mismatch)关系是推动生命发展的根本动力(Sterner & Elser, 2002 ), 也是环境变化的重要原因之一。一方面环境元素会影响生物的内平衡和生长速率(Méndez & Karlsson, 2005 ; Rivas-Ubach et al ., 2012 ); 另一方面生物会通过吸收和释放体内元素而改变环境, 进而对生物产生影响(Sterner & Elser, 2002 )。因此, 生物与环境间连续而复杂的元素化学计量适配或错配反馈关系推动着生物与环境的协同进化(Abbas et al ., 2013 )。
2 生态化学计量学在竹子基础研究中的应用
2.1 竹子养分元素需求与化学计量内平衡
竹子体内含有30-40种大量元素和微量元素, 它们都与竹子生长发育、竹材(笋)品质等生物学特性密切相关(蒋式洪等, 2000 ; 费世民, 2011 )。C、N、P、K是竹子的基本组成元素, C是生命的骨架元素与能量元素, 毛竹(Phyllostachys edulis )叶片C含量((512.13 ± 11.13) mg·g-1 )高于世界陆生植物的平均水平((464.32 ± 32.10) mg·g-1 )。N、P、K不仅影响竹子发笋数量, 也会影响竹笋品质(郑郁善等, 1998 ; 蒋式洪等, 2000 ); Ca、Mg分别与竹子根系生长和叶绿素形成有密切关系。另外, Si尽管不是生命必需元素, 但竹子体内Si含量特别高, 近年备受关注(潘月等, 2013 ; 戎洁庆等, 2013 ; Umemura & Takenaka, 2014 )。
竹子具有较强的维持体内元素相对平衡的能力。郭子武等(2011)发现施肥对红哺鸡竹(Phyllostachys iridescens )叶片C、N、P含量及比值影响不大。其叶片C:N、C:P可保持在18.71-35.02、304.41-458.52, N:P变幅更小, 仅为15.28-17.12。郭宝华等(2014)野外调查发现, 尽管土壤N:P差异明显(4.50-13.69), 但毛竹叶片N:P变化不大(12.60- 13.69)。顾大形等(2011)通过盆栽实验发现, 土壤N或P增加2-3倍, 四季竹(Oligostachyum lubricum )仍可保持叶片N:P基本不变, 其内稳性指数H (N:P) = 2.85>1。庄明浩等(2013b)发现当CO2 浓度由360 μmol·mol-1 增至500 μmol·mol-1 时, 毛竹和四季竹叶片养分C、N、P含量及其比例都没有明显变化, 说明竹子在变化的土壤或大气环境中, 都能保持体内元素的相对平衡, 符合生态化学计量学的“内稳性假说”。综合多种竹子的研究结果(周国模和姜培坤, 2004 ; 费世民, 2011 ; 庄明浩等, 2013b ), 发现竹子叶片平均化学计量比值为C:N:P = 380:16:1, 其中N:P高于自然界陆生植物平均值(10:1) (Knecht & Göransson, 2004 ), 也高于国内一般草本植物(13.5:1) (Han et al ., 2005 ), 而与阔叶植物(15.1:1.0)和常绿木本植物的平均值(15.2:1.0)相近(Han et al ., 2005 ), 说明竹子与常绿阔叶木本植物具有相似的“植物-环境”化学计量学响应特征。
2.2 竹子化学计量内平衡的影响因子
生物有机体生态化学计量内平衡易受到许多内因和外因的影响, 不同物种、器官、年龄、发育阶段化学计量特征有所不同, 而且土壤养分、气候、水分、人类活动等环境因子都会直接或间接地影响化学计量内平衡(Aerts & Chapin, 2000 ; Ågren & Weih, 2012 )(图1 )。
图1 竹子化学计量内平衡的影响因子及对植物功能的级联效应模型(仿Méndez & Karlsson (2005) )。
Fig. 1 A conceptual model of the factors influencing the ecological stoichiometry in bamboos and the cascading effects on plant functions (modified after Méndez & Karlsson (2005) ).
首先, 不同竹种的化学计量学特征有所差异。毛竹叶片N、P、K含量分别为21.44 g·kg-1 、1.33 g·kg-1 、13.15 g·kg-1 (费世民, 2011 ), 雷竹(Phyllos- tachys praecox ‘ Prevernalis’)叶片N、P、K含量分别为8.86 g·kg-1 、1.07 g·kg-1 、6.55 g·kg-1 (吴家森等, 2005a )。毛竹叶片N:P大于雷竹, 而P:K小于雷竹。麻竹(Dendrocalamus latiflorus )叶片N、P、K含量分别为56.9 g·kg-1 、4.4 g·kg-1 、8.1 g·kg-1 (邱尔发等, 2004 ), 苦竹(Pleioblastus amarus )叶片N、P、K含量分别为24.26 g·kg-1 、1.51 g·kg-1 、11.56 g·kg-1 (蒋俊明等, 2007 ), 麻竹叶片N、P、K含量都明显高于苦竹。不同竹种对环境养分变化的化学计量学响应的程度也有差异(Piouceau et al ., 2014 )。
其次, 不同器官的元素组成及比率存在较大差异。毛竹叶、枝、秆、蔸、根中N含量变异系数高达85.5%, 叶N含量(21.44 g·kg-1 )是秆(2.86 g·kg-1 )的6.0倍。Si含量变化更大, 根Si含量为70.78 g·kg-1 , 秆Si含量仅1.89 g·kg-1 , 相差近37倍(费世民, 2011 )。毛竹的N:P、P:Ca表现为叶>枝>秆>根, P:K则为叶>根>枝>秆(刘广路等, 2010 )。苦竹的N含量及N:P均呈现出叶>鞭>蔸>秆; K:P和Si:P则表现出鞭>蔸>叶>秆(刘力等, 2004 )。雷竹、麻竹等竹种的K、P、Ca、Mg等多重元素的含量及比例也都存在器官差异规律(邱尔发等, 2004 ; 吴家森等, 2005a )。以上说明竹子元素分布规律遵循植物元素“功能性不均衡分布”
原理(Yu et al ., 2011 ; Minden & Kleyer, 2014 )。
再次, 年龄也是影响竹子化学计量特征的因子。1-6年内, 毛竹秆C含量随秆龄增长呈上升趋势, N、P含量呈下降趋势(吴家森等, 2005b ; 刘广路等, 2010 ), 故C:N、C:P随年龄增长而明显增加。不同竹叶(叶龄)差异也较大, 一年生新叶N:P、N:K为13.35、4.78, 2年老叶为21.45、5.23, 老叶N:P比值明显大于幼叶(刘广路等, 2010 )。杨清培发现毛竹幼龄鞭N:P (7.0)明显低于老龄鞭(15.0) (未发表)。糙花少穗竹(Oligostachyum scabriflorum )也表现出同样的规律, 1-2年竹秆N:P为13.68-13.86, 而3、4年竹秆分别为20.18和27.88 (陈世品等, 2007 )。另外, 不同季节竹子也表现出不同的化学计量特征, 生长季N:P明显偏低(黄伯惠, 1983 )。说明竹子化学计量特征遵循“生长速率假说”。
除遗传因素外, 土壤、气候、人类活动等环境因素也会直接或间接影响竹子化学计量特征。顾大形等(2011)发现, 土壤N、P养分供应水平及土壤N:P会影响四季竹叶片N:P值, 但土壤对竹子养分计量特征的影响较复杂(陈志阳等, 2009 )。除养分含量外, 土壤有机质含量、pH值、土壤微生物等都会影响竹子体内养分含量与比率(郑郁善等, 1998 ), 所以竹林的地下-地上化学计量关系有待深入研究。
庄明浩等(2013b) 发现CO2 升高至700 μmol·mol-1 时, 竹叶主要养分元素含量及其化学计量比会发生明显的适应性变化, 毛竹叶C:N:P由406:15:1变为565:24:1, 四季竹C:N:P由468:17:1变为614:19:1, 表现出明显的P素限制作用。刘玉芳等(2015)发现长期水淹会显著降低河竹(Phyllostachys rivalis )鞭根的N、P、K含量, 从而增加C:N、C:P、C:K比值。周先容等(2012)通过氮沉降模拟实验发现, 氮沉降会显著降低青川箭竹(Fargesia rufa )叶片中C的含量, 并增加N的含量, 但没有改变P的含量, 从而C:N减小, 而N:P增加。
另外, 竹子采伐、竹笋采收等人类活动也会影响竹林化学计量特征(Raghubanshi, 1994 )。然而, 目前对竹子化学计量内平衡的影响因子及作用规律研究仍停留在少数单因素实验, 养分、水分与光照等多种因子耦合交互作用有待深入研究。
2.3 化学计量内平衡的变化对竹子生物学过程的影响
化学计量内平衡变化会影响到竹子开花结实、发笋成竹等生物学过程, 也会影响光合作用、养分吸收、克隆觅食行为、群落稳定性维持等生理生态过程(图1 )。
2.3.1 竹子开花与死亡
竹子很少开花, 但一旦开花就造成竹子大面积的突发性死亡, 因此竹子开花一直是一个有趣而神秘的话题(Janzen, 1976 ; Takahashi et al ., 2007 )。近100年来, 有关竹子开花分别从气候、管理、环境等方面提出了不同的假说(郑郁善和洪伟, 1998 ), 其中营养计量学说是目前比较普遍认可的一个重要机制性假说。
研究表明, 体内元素含量状况会影响竹子花芽分化与成花过程。如P、Ca、Mg等元素含量增加会诱导花芽分化; N、K等元素含量增加会促进营养生长, 而延迟竹子开花(何奇江等, 2005 ; 詹爱军和李兆华, 2007 )。
同时, 元素比例变化也是竹子开花的另一个重要原因(何奇江等, 2005 ; 丁兴萃, 2006 )。丁兴萃(2006)发现, 早竹(Phyllostachys violascens )开花体内C:N明显上升, 符合C:N花芽分化机理(Corbesier et al ., 2002 ); 何奇江等(2005)也发现, 开花雷竹叶、秆和鞭的P:K分别是未开花竹的1.97倍、2.80倍和2.24倍。另外, 开花往往导致养分流失、比率失衡, 促使竹子死亡(何奇江等, 2005 ; 丁兴萃, 2006 ; 詹爱军和李兆华, 2007 )。
2.3.2 发笋成竹与退笋
发笋数量、成竹质量都直接影响到竹林生态功能与生产价值, 然而有些竹子(如毛竹、毛环竹(Phyllostachys meyeri ))存在明显的发笋成竹大小年现象, 即一年大量发笋长竹, 一年行鞭换叶, 产笋较少, 每两年为一周期(南京林产工业学院竹类研究室, 1974 )。大小年现象虽有遗传原因, 但与体内N、P、K等主要矿质营养元素的化学计量特征密切相关, 如吴家森(2005b)发现小年毛竹叶中N、P、K含量均显著高于大年。Li等(1998a)发现: 竹叶被竹蝗取食后, 光合产物减少; 改变竹子体内养分状况, 可使大年变小年, 小年变大年。在实际生产实践中, 通过施肥、小年留笋养竹、大年疏笋等措施, 可将大小年竹林改造成“花年”竹林(黄伯惠, 1983 )。
另外, 并不是所有的竹笋都能长成竹子, 许多竹笋出土前就会死亡, 或虽能出土却死在成竹之前, 这叫退笋。大量研究认为退笋率高低与养分供应水平紧密相关, 增加养分供给可以降低退笋率、提高成竹率(傅懋毅等, 1988 ; Li et al ., 1998b ; 郑郁善等, 1998 ; Li et al ., 2000 )。
2.3.3 养分吸收与分配
生态化学计量特征变化会影响竹子对矿质养分的选择吸收、运输与分配。庄明浩等(2013a)发现毛竹叶片C:N下降、C:P、N:P升高, 可以促进竹叶对Mg、Ca的分配, 增强竹根Na积累, 提高Fe、Ca、Mg向上选择性运输的能力。顾大形等(2011)认为四季竹在土壤N供应充足时, 叶片也会对N进行奢侈性吸收。同时, 化学计量特征还会影响竹子的养分克隆生理整合过程, 一般“富养”分株(或部位)供应“贫养”分株(或部位), 老龄竹将N、P等元素转移到低龄竹及幼嫩部位, 但不同元素的整合强度与体内化学计量状态有关(Li et al ., 1998b ; Saitoh et al ., 2006 )。
2.3.4 光合作用与代谢
化学计量特征变化会直接影响到竹子光合作用。当施N量为250 kg·hm-2 时, 与对照相比, 1年、3年、5年生毛竹叶片的光饱和点均达最大, 分别增加了5.2%、9.1%和7.0%, 同时光补偿点降至最低, 分别降低了49.5%、20.4%和31.8% (高培军等, 2014 ), 土壤养分会影响叶片N、P含量与比例, 从而对光合作用产生直接或间接影响(宋艳冬等, 2010 ; 顾大形等, 2011 )。顾大形等(2011)发现一定范围内增加土壤N含量, 四季竹叶片的N含量增加, N:P上升, 提高了叶绿素含量, 促进其光合作用。而当施肥量不足或过量时会降低毛竹叶片光合色素含量, 结果其光合能力受到影响。苏文会(2012)发现当毛竹叶片N:P从20.70下降至15.29时, 其光合产物淀粉含量从35.47 g·kg-1 降至16.21 g·kg-1 , 降幅达54.3%。说明竹子叶片N、P化学计量特征对其光合作用影响较大。
2.3.5 觅食行为与扩张
土壤养分不足时, 竹子可调整鞭根形态、分布格局而表现出明显的觅食行为(Li et al ., 2000 ; 刘骏等, 2013 ), 以获取养分资源, 维持体内养分平衡。施肥试验表明, 毛竹竹鞭穿越养分异质环境时, 它会避开贫养斑块, 将大量竹笋有选择地放置在富养斑块(Li et al ., 2000 )。随着土壤N含量的增加, 雷竹地上竹子数量增多、生物量增大, 而且地下竹鞭长度和鞭节长度显著降低(Yue et al ., 2005 ), 表现出明显的觅食特征。毛竹扩鞭试验表明, 林缘施肥不仅可以增加新竹数量、降低退笋率, 而且可扩大新竹扩展范围, 施肥组每年平均扩张5 m 以上(董晨玲, 2003 )。这些可间接说明, 竹林向邻近森林扩张可能是为了维持体内养分平衡而表现出来的一种觅食 行为。
2.3.6 竹林(竹阔混交林)健康与稳定
竹子作为典型的克隆植物, 单株间个体大小和发育阶段不同, 其元素化学计量调节能力存在差异。当林地养分(或光照)资源不足或失衡时, 老竹个体C:N、C:P明显增加而逐渐死亡, 新竹数量减少, 竹林衰老退化。如果能及时地进行养分补充或疏伐, 会大大改善林分健康状况(郑郁善和洪伟, 1998 )。
竹子扩张会降低邻近森林植物多样性, 影响生态系统的稳定性(白尚斌等, 2013 )。初步研究认为毛竹扩张间接影响化学计量内平衡是植物多样性下降的重要机制。毛竹扩张改变了邻近森林土壤的养分状况(宋庆妮等, 2013 ), 而不同植物对土壤养分变化的响应差异较大, 其中栲(Castanopsis fargesii )、红楠(Machilus thunbergii )、交让木(Daphniphyllum macropodum )、赤杨叶(Alniphyllum fortunei )、金钱松(Pseudolarix amabilis )等内平衡调节能力较弱的植物最终退出竹木混交林, 而南方红豆杉(Taxus chinensis var. mairei )、钩锥(Castanopsis tibetana )、黄牛奶树(Symplocos laurina )等内平衡调节和可塑性能力较强的植物生长良好(杨清培, 未发表)。然而, 这方面的研究才刚刚起步, 今后应借鉴入侵植物化学计量学的研究成果(González et al ., 2010 )深入开展竹阔混交林组成与稳定性的生态化学计量学机制研究。
3 竹子生态化学计量学的初步实践
生态化学计量学不仅是探讨生物新陈代谢、生长发育、种群变化、群落演替的重要理论, 而且还是林业生产的直接指南(曾德慧和陈广生, 2005 )。前期的竹林养分诊断、平衡施肥、优质高产竹林培育等都不自觉地运用了生态化学计量学的相关原理。
3.1 指导养分诊断
营养不足容易引起植物组织老化, 而过剩又引起植物毒害, 造成资源浪费与环境污染(Güsewell, 2004 )。因此, 养分诊断是保证林农业产品优质高产的重要措施, 并将体内营养元素的含量及比例作为衡量植物营养状况的重要指标(Sinclair et al ., 1997 ; Güsewell et al ., 2003 ; Tessier & Raynal, 2003 )。
洪顺山等(1989) 和张献义等(1995)发现竹子叶片养分状况与新竹产量密切相关, 并将之作为竹林养分诊断的重要指标。此后, 陈卫文等(2004) 采用“临界值”法, 制定了毛竹不同养分诊断的化学计量标准(表1 )。
Li等(1998b) 认为, 除竹叶外, 竹秆、竹鞭、竹根等组织的养分化学计量比也都可用于养分诊断, 并初步提出毛竹叶、秆、鞭、根中N:P:K浓度含量比分别为19:1:6、15:1:26、23:1:11和44:1:54。郭宝华(2014)建议采用N:P作为竹林养分诊断的生态指标, 认为土壤N:P < 14时, 毛竹林主要受到N素的限制。因此, 今后竹林养分诊断应由单一的元素含量向多重元素比例转变, 由单一叶片向多个器官转变, 由单一的植物体测定向植物-土壤联合诊断转变。
3.2 高产优质培育
合理计量施肥不但可以增加竹材(笋)产量, 而且可以提高品质。傅懋毅等(1991)发现每年沟施复合肥3945 kg·hm-2 (N:P:K = 4:3:1), 则每度(两年)可增产春笋20000 kg·hm-2 , 并认为这是毛竹笋用林最佳培育方式; 在澳大利亚每年折合施N 250、375、500 kg·hm-2 的复合肥(N:P:K = 5:1:2.8), 毛竹笋产量分别为8300、10200、14200 kg·hm-2 (Kleinhenz et al ., 2003 )。陈孝丑等(2012)发现施肥可以显著提高毛竹林的胸径。林海萍等(2004)筛选了优质雷竹笋用林最佳配方, 当施肥量为尿素975 kg·hm-2 、复合肥1500 kg·hm-2 、厩肥112500 kg·hm-2 (N:P:K = 10:2:1)时, 最有利于雷竹笋总糖、脂肪及淀粉的积累, 而减少灰分、蛋白质及游离氨基酸的含量, 品质提高效果最好。另外, 孟勇等(2014)发现添加一些非必需元素(如硒Se)对竹笋品质具有明显的改善作用。但也有实验发现仅施化肥会使竹壁变薄, 竹材品质下降(洪顺山等, 1992 )。有机肥对竹材(笋)品质形成化学计量学机制值得关注。
3.3 平衡施肥试验
为了达到竹林优质高产的目标, 近年许多学者在养分诊断的基础上, 开展了竹林平衡施肥试验与推广。洪顺山(1987)认为毛竹林N:P:K最适施肥比率为10:6:4.7; 野中重之在1989年提出了N:P:K:Si配比为10:5:6:8 (郭晓敏等, 2013 ); 胡冬南等(2004)认为经营目标不同, 竹林需求养分的数量和比例有所差异, 认为毛竹笋用林N:P:K最佳配比为10:3:2.2, 材用林为10:6.8:9.8。2006年, 我国编制了“毛竹林丰产技术国家标准GB/T 20391-2006”, 建议对I立地级的材用林施N肥100-120 kg·hm-2 、P肥20 -25 kg·hm-2 、K肥40-45 kg·hm-2 , N:P:K为10:2.0:4.0。对II、III立地级的材用林施肥量翻倍, 笋材两用林施N肥150 kg·hm-2 、P肥40-50 kg·hm-2 、K 肥80-100 kg·hm-2 , N:P:K为10:3.0:6.0。但这些研究都还停留在经验上, 缺乏真正的计量学机制研究, 如洪顺山(1992)认为毛竹笋竹两用林最佳配方为: N肥310 kg·hm-2 、P肥83 kg·hm-2 、K 肥118 kg·hm-2 , 但如按苏文会(2012)竹材(笋)的养分含量与采收输出方法计算, 林分N、P、K利用率仅分别为28.9%、20.3%、62.5 %, 养分供需并不平衡, 因此平衡施肥需要引入化学计量学机制研究。
4 竹子化学计量学研究展望
生态化学计量学理论初步解释了竹子开花、发笋成竹等生物学问题, 也解释了竹林退化、群落动态等生态学问题, 但这些研究还很不完善, 仍停留在定性、理论和表观层面。如何使竹子生态化学计量学走向定量、应用和机制研究, 对每一位从事竹子理论或应用的研究者来说, 无疑是机遇和挑战并存。为满足社会对竹材(笋)优质高产、安全高效和环境保护的需求, 我们认为今后的竹子生态化学计量学应重点加强以下研究: (1)竹子生态化学计量
内平衡维持机制与影响因子; (2)竹材(笋)品质形成的化学计量学机制; (3)竹子-土壤化学计量学互作过程的定量化; (4)养分诊断与生态化学计量平衡施肥; (5)竹林生态系统的生态化学计量优化管理(图2 )。
图2 竹子生态化学计量学研究展望。
Fig. 2 Research outlook on ecological stoichiometry in bamboos.
4.1 加强竹子生态化学计量学内平衡维持机制与影响因子研究
生态化学计量学认为元素组成相对稳定是一切生物的基本特征。本文发现竹子叶片多重元素比率平均为C:N:P = 380:16:1。然而, 竹种、器官、年龄和发育阶段等遗传因素, 土壤、气候、人类活动等环境因素都会直接或间接地影响竹子的生态化学计量内平衡(Reich & Oleksyn, 2004 ; Sardans et al ., 2012 ), 进而影响其新陈代谢、生长发育等生命过程。同时, 植物也会通过组织老化、凋落、转移与储藏等形式, 调整养分吸收与释放, 实现整体对多重元素(如C:N:P)比率的调节, 但竹子化学计量平衡的影响因子及作用机制尚不清楚。以后应加强多竹种(尤其是经济竹种和珍稀竹种)化学计量内平衡特征研究; 加强开花、发笋、大小年等基础生物学过程与元素化学计量的关系研究; 结合分子、同位素等先进技术, 深入开展竹子化学计量内稳性与其光合作用、养分分配、克隆整合等生理生态学研究; 开展多因子(养分与水分)对元素内平衡的交互影响及竹子的响应机制; 开展竹子化学计量特征与新陈代谢活性物质的关系研究(Rivas-Ubach et al ., 2012 ), 以提高生态化学计量学对竹子生物学现象、过程和机制的解释能力。
4.2 加强竹材(笋)品质形成的化学计量学基础研究
竹材(笋)品质是市场竞争力的核心。大径级、节间长、壁厚、枝下高长的竹材价格明显要高; 外观漂亮、营养丰富的竹笋更受市场欢迎。尽管目前已开展了部分养分元素(N、P、K、Se、Si)对竹材(笋)品质形成的影响实验(孟勇等, 2014 ), 但相关的基础研究仍很滞后, 高产优质的形成规律还未被充分地揭示与认识, 可推广的高产优质培育技术尚未确立, 制约了竹林产量提高和品质的改善。因此建议开展以下研究: 1)竹笋品质形成规律及化学计量学机制研究, 尤其是Se、Zn等非必需元素、有机肥与竹笋品质形成的生理机制; 2)节长、壁厚等优质材用林定向培育的化学计量学原理与技术研究; 3)基于光、温、肥、水等多种环境因子的竹材(笋)高产、优质和资源高效利用的化学计量调控机制研究。
4.3 强化竹子-土壤化学计量学互作过程的定量化研究
植物养分主要来源于土壤, 土壤养分也受到植物的反作用。因此, 加强竹子与土壤养分元素的化学计量适配与错配互作研究, 对竹林生产、环境保护至关重要。首先, 土壤环境决定着养分形态、数量及其有效性, 从而影响到竹子的生长发育(Fan et al ., 2015 ; Zechmeister-Boltenstern et al ., 2015 )。建议开展土壤环境因子与土壤、竹子养分元素化学计量特征动态关系的长期监测与研究。其次, 凋落物分解和根际效应是影响土壤养分的一个重要过程, 尤其是根际过程更应受到关注(Bell et al ., 2014 , 2015 ; Carrillo et al ., 2014 ), 所以应采用稳定同位素技术、分子生态技术, 聚焦于竹子根际过程及养分活化机制(如根系吸收与分泌、根际酸碱度变化、根际微生物活动) , 实现“竹子-根际-土壤”三者元素关系的量化分析。
4.4 加强养分诊断与生态化学计量平衡施肥研究
养分诊断与平衡施肥是提高林业生产、维护系统稳定、选择合适管理策略的重要手段(Park et al ., 2014 ), 但当前竹林养分诊断仍停留在单一的养分测定(陈卫文等, 2004 ; 郭晓敏等, 2013 ), 很少考虑多养分、多组织的耦合效应, 失衡超量施肥强烈影响到竹林土壤理化性质, 并引起竹笋食用安全、水土环境恶化等问题。今后应以生态化学计量平衡理论为基础, 改进养分诊断方法, 根据不同竹种、不同林分、不同时期养分限制性的差异特征, 筛选养分丰缺指标, 建立多元素、多组分的养分诊断技术, 提高监测精度与预测能力; 形成生态化学计量平衡施肥技术, 根据竹子和土壤的营养特点, 筛选肥料种类、确定肥料用量与配比, 实施时空精准施肥, 提高肥料利用效率, 实现多重元素间的平衡, 竹子养分需求与环境养分供给间的平衡, 肥料投入与经济产出间的平衡, 产品生产与环境保护间的平衡。
4.5 强化竹林生态系统生态化学计量优化管理
生命元素通过物质循环、能量流动等形式将生态系统的各组分、多层次紧密联系起来(Carnicer et al ., 2015 )。竹林是一个人工或半人工的开放生态系统, 几乎每年都有竹材(或竹笋)等产品输出和外源养分的输入。产品输出和养分输入的失衡会改变竹子、土壤、微生物等不同组分的化学计量特征(Raghubanshi, 1994 ; Venkatesh et al ., 2005 ; Zhang et al ., 2013 )。因此, 今后应充分考虑生物与环境间的相互渗透性, 根据物质守恒与能量守恒定律, 综合考虑土壤、气候、经营活动等因素, 加强竹林生态系统的生态化学计量模型与理论研究, 构建“竹子-土壤-微生物”多组分的“生态化学计量匹配模型”, 竹子化学计量特征与群落组成结构、生态系统生产力多层次的多重养分优化计量模型, 并通过长期野外监测与微宇宙模拟实验, 找到具有竹子特色的“特征参数”, 最后形成可视化的竹林养分预测和管理系统, 指导竹林健康经营和环境综合效应评估, 实现因地制宜、以产定肥的竹林生态系统生态化学计量优化管理模式。
总之, 竹子通过“化学计量内稳性”维持和“生长速率”调节, 影响着竹子生长发育、开花死亡等生物学过程, 也影响到竹材(笋)品质、竹林产量、群落动态和环境效应等生态学过程。随着人们对竹子优质高产、资源持续利用、环境友好等多目标需求增长, 今后应当借鉴生态化学计量学领域的最新研究成果, 紧紧围绕竹子与环境间重要生命元素的互动耦合机制、竹林生态系统物质循环与竹材(笋)品质及产量形成过程, 加强竹林生态系统“计量平衡-养分管理-竹林生产-环境效应”动态过程的长期监测与研究。
可以预言, 竹子生态化学计量学必将进一步整合竹子生物学、营养学、培育学以及生态学等领域的研究成果, 广泛应用于竹子基础研究与生产实践, 为我国竹产业发展与生态保护做出应有的贡献。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Abbas M Ebeling A Oelmann Y Ptacnik R Roscher C Weigelt A Weisser WW Wilcke W Hillebrand H 2013 ). Biodiversity effects on plant stoichiometry.
PLoS ONE , 8 , e58179 .
[本文引用: 1]
[2]
Aerts R Chapin F 2000 ). The mineral nutrition of wildland plants re-visited: A re-evaluation of processes and patterns.
Advances in Ecological Research , 30 , 1 -67 .
[本文引用: 1]
[3]
Ågren GI 2008 ). Stoichiometry and nutrition of plant growth in natural communities.
Annual Review of Ecology, Evolution, and Systematics , 39 , 153 -170 .
[本文引用: 1]
[4]
Ågren GI Weih M 2012 ). Plant stoichiometry at different scales: Element concentration patterns reflect environment more than genotype.
New Phytologist , 194 , 944 -952 .
[本文引用: 1]
[5]
Anderson TR Boersma M Raubenheimer D 2004 ). Stoichio- metry: Linking elements to biochemicals.
Ecology , 85 , 1193 -1202 .
[本文引用: 1]
[6]
Bai SB Zhou GM Wang YX Liang QQ Chen J Cheng YY Shen R 2013 ). Plant species diversity and its dynamics in forests invaded by Moso bamboo (Phyllostachys edulis).
Biodiversity Science , 21 , 1 -8 . (in Chinese with English abstract)[白尚斌 , 周国模 , 王懿祥 , 梁倩倩 , 陈娟 , 程艳艳 , 沈蕊 (2013 ). 森林群落植物多样性对毛竹入侵的响应及动态变化
. 生物多样性 , 21 , 1 -8 .]
[本文引用: 1]
[7]
Bamboo Research Institute of Nanjing Technological College of Forest Products (1974 ). Bamboo Cultivation . China Agriculture Press, Beijing . (in Chinese)[南京林产工业学院竹类研究室 (1974 ). 竹林培育. 中国农业出版社 , 北京.]
[8]
Bell C Carrillo Y Boot CM Rocca JD Pendall E Wallenstein MD 2014 ). Rhizosphere stoichiometry: Are C:N: P ratios of plants, soils, and enzymes conserved at the plant species-level?
New Phytologist , 201 , 505 -517 .
[本文引用: 1]
[9]
Bell CW Asao S Calderon F Wolk B Wallenstein MD 2015 ). Plant nitrogen uptake drives rhizosphere bacterial community assembly during plant growth.
Soil Biology & Biochemistry , 85 , 170 -182 .
[本文引用: 1]
[10]
Belovsky GE 1978 ). Diet optimization in a generalist herbivore: The moose.
Theoretical Population Biology , 14 , 105 -134 .
[本文引用: 1]
[11]
Carnicer J Sardans J Stefanescu C Ubach A Bartrons M Asensio D Peñuelas J 2015 ). Global biodiversity, stoichiometry and ecosystem function responses to human-induced C-N-P imbalances.
Journal of Plant Physiology , 172 , 82 -91 .
[本文引用: 1]
[12]
Carrillo Y Dijkstra F Pendall E LeCain D Tucker C 2014 ). Plant rhizosphere influence on microbial C metabolism: The role of elevated CO2 , N availability and root stoichiometry.
Biogeochemistry , 117 , 229 -240 .
[本文引用: 1]
[13]
Chen SP Li MB Chen AL 2007 ). Difference of nutrient allocation in different slope positions in Oligostachyum scabriflorum forest.
Journal of Fujian College of Forestry , 27 , 193 -198 . (in Chinese with English abstract)[陈世品 , 黎茂彪 , 陈爱玲 (2007 ). 不同坡位糙花少穗竹林养分分配格局
. 福建林学院学报 , 27 , 193 -198 .]
[本文引用: 1]
[14]
Chen WW Luo ZJ Chen F Lu JW Ding CP 2004 ). The nutrition status and nutrition diagnosis of bamboo leaves in the south of Hubei.
Journal of Northeast Forestry University , 32 (2 ), 41 -44 . (in Chinese with English abstract)[陈卫文 , 罗治建 , 陈防 , 鲁剑巍 , 丁次平 (2004 ). 鄂南毛竹林的养分状况与营养诊断标准
. 东北林业大学学报 , 32 (2 ), 41 -44 .]
[本文引用: 4]
[15]
Chen XC Liu GL Fang SH Guan FY Su WH Huang JH 2012 ). Growth characteristics and biomass distribution of Phyllostachys pubescens stands with changes in fertilization time.
Journal of Zhejiang A & F University , 29 , 52 -57 . (in Chinese with English abstract)[陈孝丑 , 刘广路 , 范少辉 , 官凤英 , 苏文会 , 黄金华 (2012 ). 连续施肥对毛竹林生长特征及生物量空间构型的影响
. 浙江农林大学学报 , 29 , 52 -57 .]
[16]
Chen ZY Yao XM Tian XM 2009 ). Establishment of leaf nutrition, soil fertility and output models in Phyllostachys pubescens.
Nonwood Forest Research , 27 (3 ), 53 -56 . (in Chinese with English abstract)[陈志阳 , 姚先铭 , 田小梅 (2009 ). 毛竹叶片营养与土壤肥力及产量模型的建立
. 经济林研究 , 27 (3 ), 53 -56 .]
[本文引用: 1]
[17]
Chongtham N Bisht MS Haorongbam S 2011 ). Nutritional properties of bamboo shoots: Potential and prospects for utilization as a health food.
Comprehensive Reviews in Food Science and Food Safety , 10 (3 ), 153 -168 .
[本文引用: 1]
[18]
Corbesier L Bernier G Périlleux C 2002 ). C:N ratio increases in the phloem sap during floral transition of the long-day plants Sinapis alba and Arabidopsis thaliana.
Plant and Cell Physiology , 43 , 684 -688 .
[本文引用: 1]
[19]
Ding XC 2006 ). Physiology of Aging and Flowering for Phyllostachys praecox. PhD dissertation, Nanjing Forestry University, Nanjing.
(in Chinese with English abstract)[丁兴萃 (2006 ). 早竹衰老开花生理学基础研究
. 博士学位论文, 南京林业大学, 南京.]
[本文引用: 3]
[20]
Dong CL 2003 ). Study on growth effect on new stands of Phyllostachys heterocycla cv. Pubescens through rhizome expanding and growing.
Journal of Bamboo Research , 22 (4 ), 30 -33 . (in Chinese with English abstract)[董晨玲 (2003 ). 毛竹扩鞭成林新竹生长效果研究
. 竹子研究汇刊 , 22 (4 ), 30 -33 .]
[本文引用: 1]
[21]
Dong J Zhang Y 2015 ). Developing bamboo industry to promote China-Africa South-South cooperation.
Forestry Economics , 6 , 94 -97 . (in Chinese with English abstract)[董杰 , 张燕 (2015 ). 发展竹产业促进中非南南合作
. 林业经济 , 6 , 94 -97 .]
[本文引用: 1]
[22]
Elser JJ Acharya K Kyle M Cotner J Makino W Markow T Watts T Hobbie S Fagan W Schade J Hood J Sterner RW 2003 ). Growth rate-stoichiometry couplings in diverse biota.
Ecology Letters , 6 , 936 -943 .
[本文引用: 1]
[23]
Elser JJ Fagan WF Kerkhoff AJ Swenson NG Enquist BJ 2010 ). Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change.
New Phytologist , 186 , 593 -608 .
[本文引用: 1]
[24]
Fan H Wu J Liu W Yuan Y Hu L Cai Q 2015 ). Linkages of plant and soil C:N:P stoichiometry and their relationships to forest growth in subtropical plantations.
Plant and Soil , 392 , 127 -138 .
[本文引用: 1]
[25]
Fei SM 2011). Bamboo Forest Ecological Research . China Forestry Publishing House, Beijing . (in Chinese)[费世民 (2011). 竹林生态研究. 中国林业出版社 , 北京.]
[26]
Fu MY Xie JZ Fang XY 1991 ). Fertilization studies in bamboo stands with different end uses II. High yield management method for bamboo shoot stands.
Forest Research , 4 , 238 -245 . (in Chinese with English abstract)[傅懋毅 , 谢锦忠 , 方敏瑜 (1991 ). 不同用途毛竹林的施肥研究II. 毛竹笋用林丰产经营技术
. 林业科学研究 , 4 , 238 -245 .]
[27]
Fu MY Xie JZ Fang XY Ren XY Li DY 1988 ). Fertilization studies in bamboo stands with different end uses I. For bamboo timber stand.
Forest Research , 1 , 541 -547 . (in Chinese with English abstract)[傅懋毅 , 谢锦忠 , 方敏瑜 , 任晓京 , 李岱一 (1988 ). 不同用途毛竹林的施肥研究Ⅰ. 毛竹材用林的施肥
. 林业科学研究 , 1 , 541 -547 .]
[本文引用: 1]
[28]
Gao PJ Qiu YH Zhou ZQ He RH Xu J 2014 ). Productivity and photosynthetic ability of Phyllostachys edulis with nitrogen fertilization.
Journal of Zhejiang A & F University , 31 , 697 -703 . (in Chinese with English abstract)[高培军 , 邱永华 , 周紫球 , 何仁华 , 徐佳 (2014 ). 氮素施肥对毛竹生产力与光合能力的影响
. 浙江农林大学学报 , 31 , 697 -703 .]
[本文引用: 1]
[29]
González AL Kominoski JS Danger M Ishida S Iwai N Rubach A 2010 ). Can ecological stoichiometry help explain patterns of biological invasions?
Oikos , 119 , 779 -790 .
[本文引用: 1]
[30]
Grubb PJ 1977 ). The maintenance of species-richness in plant communities: The importance of the regeneration niche.
Biological Reviews , 52 , 107 -145 .
[本文引用: 1]
[31]
Gu DX Chen SL Huang YQ 2011 ). Effects of soil nitrogen and phosphonium on leaf nitrogen and phosphonium stoichiometric characteristics and chlorophyll content of Oligostachyum lubricum.
Chinese Journal of Plant Ecology , 35 , 1219 -1225 . (in Chinese with English abstract)[顾大形 , 陈双林 , 黄玉清 (2011 ). 土壤氮磷对四季竹叶片氮磷化学计量特征和叶绿素含量的影响
. 植物生态学报 , 35 , 1219 -1225 .]
[本文引用: 1]
[32]
Guo BH Liu GL Fang SH Du MY Su WH 2014 ). Distribution patterns and stoichiometry characteristics of C, N, P in Phyllostachys edulis forests of different productivity levels.
Scientia Silvae Sinica , 50 (6 ), 1 -9 . (in Chinese with English abstract)[郭宝华 , 刘广路 , 范少辉 , 杜满义 , 苏文会 (2014 ). 不同生产力水平毛竹林碳氮磷的分布格局和计量特征
. 林业科学 , 50 (6 ), 1 -9 .]
[本文引用: 1]
[33]
Guo XM Niu DK Chen F Zhang WY 2013). Nutrient Management for Bamboo Forest . Science Press, Beijing . (in Chinese)[郭晓敏 , 牛德奎 , 陈防 , 张文元 (2013). 毛竹林平衡施肥与营养管理. 科学出版社 , 北京.]
[34]
Guo Z Chen S Yang Q Li Y Zhuang M 2014 ). Effects of mulching management on soil and foliar C, N and P stoichiometry in bamboo (Phyllostachys violascens) plantations.
Journal of Tropical Forest Science , 26 , 572 -580 .
[35]
Guo ZW Yu MZ Zheng LX Yang QP Li YC Chen XL 2011 ). Stoichiometry of C, N and P in Phyllostachys iridescens leaves under long-term application of different fertilizers.
Chinese Journal of Ecology , 30 , 2667 -2671 . (in Chinese with English abstract)[郭子武 , 虞敏之 , 郑连喜 , 杨清平 , 李迎春 , 陈双林 (2011 ). 长期施用不同肥料对红哺鸡竹林叶片碳、氮、磷化学计量特征的影响
. 生态学杂志 , 30 , 2667 -2671 .]
[36]
Güsewell S 2004 ). N:P ratios in terrestrial plants: Variation and functional significance.
New Phytologist , 164 , 243 -266 .
[本文引用: 2]
[37]
Güsewell S Koerselman W Verhoeven JTA 2003 ). Biomass N:P ratios as indicators of nutrient limitation for plant populations in wetlands.
Ecological Applications , 13 , 372 -384 .
[本文引用: 1]
[38]
Han W Fang J Guo D Zhang Y 2005 ). Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China.
New Phytologist , 168 , 377 -385 .
[本文引用: 2]
[39]
He JS Han XG 2010 ). Ecological stoichiometry: Searching for unifying principles from individuals to ecosystem.
Chinese Journal of Plant Ecology , 34 , 2 -6 . (in Chinese with English abstract)[贺金生 , 韩兴国 (2010 ). 生态化学计量学: 探索从个体到生态系统的统一化理论
. 植物生态学报 , 34 , 2 -6 .]
[本文引用: 2]
[40]
He QJ Wang KH Hua XQ Tong XQ 2005 ). Change of endogenous hormones, amino-acid and nutrition in flowering stage of Phyllostachys praecox f. prevernalis.
Scientia Silvae Sinicae , 41 (2 ), 169 -173 . (in Chinese with English abstract)[何奇江 , 汪奎宏 , 华锡奇 , 童晓青 (2005 ). 雷竹开花期内源激素、氨基酸和营养成分含量变化
. 林业科学 , 41 (2 ), 169 -173 .]
[本文引用: 4]
[41]
Hillebrand H Cowles JM Lewandowska A van de Waal DB Plum C 2014 ). Think ratio! A stoichiometric view on biodiversity-ecosystem functioning research.
Basic and Applied Ecology , 15 , 465 -474 .
[本文引用: 1]
[42]
Hong SS 1987 ). A preliminary study on optimum amounts and proportions of fertilizers for the growth of Phyllostachys pubescens.
Journal of Bamboo Research , 6 (1 ), 35 -41 . (in Chinese with English abstract)[洪顺山 (1987 ). 毛竹配方施肥研究初报
. 竹子研究汇刊 , 6 (1 ), 35 -41 .]
[43]
Hong SS Hu BT Jiang YG 1989 ). A study on nutrient diagnoses of Phyllostachys pubescens.
Forest Research , 2 , 15 -24 . (in Chinese with English abstract)[洪顺山 , 胡炳堂 , 江业根 (1989 ). 毛竹营养诊断的研究
. 林业科学研究 , 2 , 15 -24 .]
[本文引用: 1]
[44]
Hong SS Hu BT Jiang YG 1992 ). The responses of fertilization on the plantations of Phyllostachys pubescens.
Forest Research , 5 , 371 -378 . (in Chinese with English abstract)[洪顺山 , 胡炳堂 , 江业根 (1992 ). 毛竹林施肥效应研究
. 林业科学研究 , 5 , 371 -378 .]
[本文引用: 1]
[45]
Hu DN Chen LX Li FK Guo XM Lei J 2004 ). Effect of optimum amount and proportion of fertilizer on bamboo shoots and timbers.
Acta Agriculturae Universitatis Jiangxiensis , 26 , 196 -199 . (in Chinese with English abstract)[胡冬南 , 陈立新 , 李发凯 , 郭晓敏 , 雷俊 (2004 ). 配方施肥对毛竹笋材的影响
. 江西农业大学学报 , 26 , 196 -199 .]
[46]
Huang BH 1983 ). A dynamic study on the elements of mineral nutrition in Phyllostachys pubescens.
Journal of Bamboo Research , 2 (1 ), 87 -111 . (in Chinese with English abstract)[黄伯惠 (1983 ). 毛竹矿质营养元素动态的研究
. 竹子研究汇刊 , 2 (1 ), 87 -111 .]
[本文引用: 2]
[47]
Janzen DH 1976 ). Why bamboos wait so long to flower.
Annual Review of Ecology and Systematics , 7 , 347 -391 .
[本文引用: 1]
[48]
Jiang JM Fei SM Li JY Tang SM Ye X 2007 ). The distribution of main nutrient elements in the organs of Pleioblastus amarus and the nutrient output of its harvesting.
Journal of Sichuan Forestry Science and Technology , 28 (2 ), 15 -19 . (in Chinese with English abstract)[蒋俊明 , 费世民 , 李吉跃 , 唐森强 , 叶梓 (2007 ). 苦竹各器官主要营养元素分布及采伐的养分输出
. 四川林业科技 , 28 (2 ), 15 -19 .]
[本文引用: 1]
[49]
Jiang SH Zhong CS Zhang GZ Zhang XL Ye DZ Wang WP 2000 ). Study on the effects of phosphorus, potassium and boron fertilizers on shoot qualities of Phyllostachys iridenscens.
Journal of Bamboo Research , 19 (1 ), 48 -51 . (in Chinese with English abstract)[蒋式洪 , 钟传声 , 张国镇 , 张贤林 , 叶德柱 , 王卫平 (2000 ). 磷钾硼肥对红竹笋品质效应的研究
. 竹子研究汇刊 , 19 (1 ), 48 -51 .]
[本文引用: 2]
[50]
Jiang ZH 2007). Bamboo and Rattan in the World . China Forestry Publishing House, Beijing . (in Chinese)[江泽慧 (2007). 世界竹藤. 中国林业出版社 , 北京.]
[51]
Kleinhenz V Gosbee M Elsmore S Lyall TW Blackburn K Harrower K Midmore DJ 2000 ). Storage methods for extending shelf life of fresh, edible bamboo shoots [Bambusa oldhamii (Munro)].
Postharvest Biology and Technology , 19 , 253 -264 .
[本文引用: 1]
[52]
Kleinhenz V Midmore DJ 2001 ). Aspects of bamboo agronomy.
Advances in Agronomy , 74 , 99 -153 .
[本文引用: 2]
[53]
Kleinhenz V Milne J Walsh KB Midmore DJ 2003 ). A case study on the effects of irrigation and fertilization on soil water and soil nutrient status, and on growth and yield of bamboo (Phyllostachys pubescens) shoots.
Journal of Bamboo and Rattan , 2 , 281 -293 .
[本文引用: 1]
[54]
Knecht MF Göransson A 2004 ). Terrestrial plants require nutrients in similar proportions.
Tree Physiology , 24 , 447 -460 .
[本文引用: 1]
[55]
Li R Werger MJA During HJ Zhong ZC 1998 a). Biennial variation in production of new shoots in groves of the giant bamboo Phyllostachys pubescens in Sichuan, China.
Plant Ecology , 135 , 103 -112 .
[56]
Li R Werger MJA During HJ Zhong ZC 1998 b). Carbon and nutrient dynamics in relation to growth rhythm in the giant bamboo Phyllostachys pubescens.
Plant and Soil , 201 , 113 -123 .
[57]
Li R Werger MJA Kroon HD During HJ Zhong ZC 2000 ). Interactions between shoot age structure, nutrient availability and physiological integration in the giant bamboo Phyllostachys pubescens.
Plant Biology , 2 , 437 -446 .
[本文引用: 3]
[58]
Lin HP Zhou SH Shen ZP 2004 ). The influence of intensive management on the bamboo shoot’s quality.
Journal of Jiangsu Forestry Science & Technology , 31 (2 ), 9 -11 . (in Chinese with English abstract)[林海萍 , 周淑红 , 沈振民 (2004 ). 集约经营对毛竹笋品质的影响
. 江苏林业科技 , 31 (2 ), 9 -11 .]
[本文引用: 1]
[59]
Liu GL Fang SH Guan FY Du MY Huang YN 2010 ). Distribution pattern of nutrient elements and its relationship with soil environment in different aged Phyllostachys edulis.
Forest Research , 23 , 252 -258 . (in Chinese with English abstract)[刘广路 , 范少辉 , 官凤英 , 杜满意 , 黄永南 (2010 ). 不同年龄毛竹营养器官主要养分元素分布及与土壤环境的关系
. 林业科学研究 , 23 , 252 -258 .]
[本文引用: 3]
[60]
Liu J Yang QP Song QP Yu DK Yang GY Qi HY Shi JM 2013 ). Strategy of fine root expansion of Phyllostachys pubescens population into evergreen broad-leaved forest.
Chinese Journal of Plant Ecology , 37 , 230 -238 . (in Chinese with English abstract)[刘骏 , 杨清培 , 宋庆妮 , 余定坤 , 杨光耀 , 祁红艳 , 施建敏 (2013 ). 毛竹种群向常绿阔叶林扩张的细根策略
. 植物生态学报 , 37 , 230 -238 .]
[本文引用: 1]
[61]
Liu L Lin XC Jin AW Feng TX Zhou CP Ji ZF 2004 ). Analysis of nutrient elements in various organs of Pleioblastus amarus.
Journal of Zhejiang Forestry College , 21 , 172 -175 . (in Chinese with English abstract)[刘力 , 林新春 , 金爱武 , 冯天喜 , 周昌平 , 季宗富 (2004 ). 苦竹各器官营养元素分析
. 浙江林学院学报 , 21 , 172 -175 .]
[本文引用: 1]
[62]
Liu YF Chen SL Li YC Chen S Guo ZW 2015 ). Effect of long-term flooded conditions on nutrient stoichiometric characteristics of Phyllostachys rivalis rhizome roots.
Acta Botanica Boreali-Occidentalia Sinica , 35 , 350 -355 . (in Chinese with English abstract)[刘玉芳 , 陈双林 , 李迎春 , 陈珊 , 郭子武 , 杨清平 (2015 ). 长期淹水对河竹鞭根养分化学计量特征的影响
. 西北植物学报 , 35 , 350 -355 .]
[63]
Lou YP 2001 ). The Maintenance of Sustainable Site Productivity of Managed Moso Bamboo Forests in China. PhD dissertation, Chinese Academy of Forestry Sciences, Beijing.
(in Chinese with English abstract)[楼一平 (2001 ). 毛竹人工林持续立地生产力的研究
. 博士学位论文, 中国林业科学研究院, 北京.]
[本文引用: 1]
[64]
Lou YP Wu LR Li RC Liu ZJ Liu YR 1997 ). Growth dynamics of pure Phyllostaehys pubeseens stands transformed from mixed stands.
Forest Research , 10 , 35 -41 . (in Chinese with English abstract)[楼一平 , 吴良如 , 李瑞成 , 刘仲君 , 刘耀荣 (1997 ). 竹木混交林改为毛竹纯林经营后的林分生长动态
. 林业科学研究 , 10 , 35 -41 .]
[本文引用: 1]
[65]
Méndez M Karlsson PS 2005 ). Nutrient stoichiometry in Pinguicula vulgaris: Nutrient availability, plant size, and reproductive status.
Ecology , 86 , 982 -991 .
[本文引用: 1]
[66]
Meng CF Shen Q Jiang PK Xu QF 2009 ). Effects of different fertilization on soil nutrient balance and bamboo shoot yield of Phyllostachys praecox stands.
Journal of Bamboo Research , 28 (4 ), 11 -17 . (in Chinese with English abstract)[孟赐福 , 沈菁 , 姜培坤 , 徐秋芳 (2009 ). 不同施肥处理对雷竹林土壤养分平衡和竹笋产量的影响
. 竹子研究汇刊 , 28 (4 ), 11 -17 .]
[本文引用: 2]
[67]
Meng Y Ai WS Yang SL Yang M Liu SS Li MQ Xu J Pu XY 2014 ). Effect of selenium fertilization for bamboo cavity of Phyllostachys pubescens on the quality of bamboo shoots.
Hunan Forestry Science & Technology , 41 (2 ), 20 -23 . (in Chinese with English abstract)[孟勇 , 艾文胜 , 杨书林 , 杨明 , 刘少山 , 李美群 , 涂佳 , 蒲湘云 (2014 ). 毛竹竹腔富硒施肥对竹笋品质的影响
. 湖南林业科技 , 41 (2 ), 20 -23 .]
[本文引用: 1]
[68]
Minden V Kleyer M 2014 ). Internal and external regulation of plant organ stoichiometry.
Plant Biology , 16 , 897 -907 .
[本文引用: 1]
[69]
Niklas KJ Owens T Reich PB Cobb ED 2005 ). Nitrogen/ phosphorus leaf stoichiometry and the scaling of plant growth.
Ecology Letters , 8 , 636 -642 .
[本文引用: 1]
[70]
Pan Y Rong JQ Sheng WX Gui RY 2013 ). Silicon and drought resistance of Phyllostachys violascens.
Journal of Zhejiang A & F University , 30 , 852 -857 . (in Chinese with English abstract)[潘月 , 戎洁庆 , 盛卫星 , 桂仁意 (2013 ). 硅对雷竹抗旱性的影响
. 浙江农林大学学报 , 30 , 852 -857 .]
[本文引用: 1]
[71]
Park BB Park GE Bae K 2014 ). Diagnosis of plant nutrient and growth responses on fertilization with vector analysis and morphological index.
Forest Science and Technology , 11 , 1 -10 .
[本文引用: 1]
[72]
Piouceau J Bois G Panfili F Anastase M Dufossé L Arfi V 2014 ). Effects of high nutrient supply on the growth of seven bamboo species.
International Journal of Phytoremediation , 16 , 1042 -1057 .
[本文引用: 1]
[73]
Qiu EF Chen ZM Hong W Zheng YS Huang BL Yang ZQ 2004 ). Nutrient allocation pattern of Dendrocalamus latiflorus forest ecosystem planted on hill.
Acta Ecologica Sinica , 24 , 2693 -2699 . (in Chinese with English abstract)[邱尔发 , 陈卓梅 , 洪伟 , 郑郁善 , 黄宝龙 , 杨主泉 (2004 ). 山地麻竹林生态系统养分分配格局
. 生态学报 , 24 , 2693 -2699 .]
[本文引用: 2]
[74]
Raghubanshi AS 1994 ). Effect of bamboo harvest on dynamics of nutrient pools, N mineralization, and microbial biomass in soil.
Biology and Fertility of Soils , 18 , 137 -142 .
[本文引用: 2]
[75]
Redfield AC 1958 ). The biological control of chemical factors in the environment.
American Scientist , 46 , 205 -221 .
[本文引用: 1]
[76]
Reef R Ball MC Feller IC Lovelock CE 2010 ). Relationships among RNA:DNA ratio, growth and elemental stoichio- metry in mangrove trees.
Functional Ecology , 24 , 1064 -1072 .
[本文引用: 1]
[77]
Reich PB Oleksyn J 2004 ). Global patterns of plant leaf N and P in relation to temperature and latitude.
Proceedings of the National Academy of Sciences of the United States of America , 101 , 11001 -11006 .
[本文引用: 1]
[78]
Reiners WA 1986 ). Complementary models for ecosystems.
The American Naturalist , 127 , 59 -73 .
[79]
Rivas-Ubach A Sardans J Pérez-Trujillo M Estiarte M Peñuelas J 2012 ). Strong relationship between elemental stoichiometry and metabolome in plants.
Proceedings of the National Academy of Sciences of the United States of America , 109 , 4181 -4186 .
[本文引用: 3]
[80]
Rong JQ Fan Y Gui RY 2013 ). Effect of silicon fertilizer application on plantation of Phyllostachys violascens.
Acta Agriculturae Universitatis Jiangxiensis , 35 , 473 -479 . (in Chinese with English abstract)[戎洁庆 , 潘月 , 桂仁意 (2013 ). 硅肥对雷竹林生长影响研究
. 江西农业大学学报 , 35 , 473 -479 .]
[本文引用: 1]
[81]
Saitoh T Seiwa K Nishiwaki A 2006 ). Effects of resource heterogeneity on nitrogen translocation within clonal fragments of Sasa palmata: An isotopic (15 N) assessment.
Annals of Botany , 98 , 657 -663 .
[本文引用: 1]
[82]
Sardans J Rivas-Ubach A Peñuelas J 2012 ). The C:N:P stoichiometry of organisms and ecosystems in a changing world: A review and perspectives.
Perspectives in Plant Ecology, Evolution and Systematics , 14 , 33 -47 .
[本文引用: 1]
[83]
Schatz G McCauley E 2007 ). Foraging behavior by Daphnia in stoichiometric gradients of food quality.
Oecologia , 153 , 1021 -1030 .
[本文引用: 1]
[84]
Sinclair AG Morrison JD Smith LC Dodds KG 1997 ). Determination of optimum nutrient element ratios in plant tissue.
Journal of Plant Nutrition , 20 , 1069 -1083 .
[本文引用: 1]
[85]
Song QN Yang QP Liu J Yu DK Fang K Xu P He YJ 2013 ). Effects of Phyllostachys edulis expansion on soil nitrogen mineralization and its availability in evergreen broadleaf forest.
Chinese Journal of Applied Ecology , 24 , 338 -344 . (in Chinese with English abstract)[宋庆妮 , 杨清培 , 刘骏 , 余定坤 , 方楷 , 徐佩 , 何宇娟 (2013 ). 毛竹扩张对常绿阔叶林土壤氮素矿化及有效性的影响
. 应用生态学报 , 24 , 338 -344 .]
[本文引用: 1]
[86]
Song YD Jin AW Jin XC Hu YB Du LL Jiang ZY 2010 ). Physiology of leaf photosynthesis with fertilization in Phyllostachys pubescens.
Journal of Zhejiang Forestry College , 27 , 334 -339 . (in Chinese with English abstract)[宋艳冬 , 金爱武 , 金晓春 , 胡元斌 , 杜亮亮 , 江志友 (2010 ). 施肥对毛竹叶片光合生理的影响
. 浙江林学院学报 , 27 , 334 -339 .]
[本文引用: 1]
[87]
Sterner RW Elser JJ 2002). Ecological Stoichiometry : The Biology of Elements from Molecules to the Biosphere. Princeton University Press, Princeton, USA .
[88]
Su WH 2012 ). Fertilization Theory and Practice for Phyllostachys edulis Stand Based on Growth and Nutrient Accumulation Rule. PhD dissertation, Chinese Academy of Forestry Sciences, Beijing.
(in Chinese with English abstract)[苏文会 (2012 ). 基于生长和养分积累规律的毛竹林施肥理论与实践研究
. 博士学位论文, 中国林业科学研究院, 北京.]
[89]
Takahashi M Furusawa H Limtong P Sunanthapongsuk V Marod D Panuthai S 2007 ). Soil nutrient status after bamboo flowering and death in a seasonal tropical forest in western Thailand.
Ecological Research , 22 (1 ), 160 -164 .
[本文引用: 1]
[90]
Tessier JT Raynal DJ 2003 ). Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation.
Journal of Applied Ecology , 40 , 523 -534 .
[本文引用: 1]
[91]
Tilman D 1982). Resource Competition and Community Structure . Princeton University Press Princeton, Princeton, USA .
[92]
Umemura , M , Takenaka C 2014 ). Biological cycle of silicon in moso bamboo (Phyllostachys pubescens) forests in central Japan.
Ecological Research , 29 , 501 -510 .
[93]
Venkatesh MS Bhatt BP Kumar K Majumdar B Singh K 2005 ). Soil properties influenced by some important edible bamboo species in the North Eastern Himalayan region, India.
Journal of Bamboo and Rattan , 4 , 221 -230
[本文引用: 1]
[94]
Vitousek P 1982 ). Nutrient cycling and nutrient use efficiency.
The American Naturalist , 119 , 553 -527 .
[本文引用: 1]
[95]
Wu JS Jiang PK Sheng WY Xie BL 2009 ). Effect of bamboo (Phyllostachys praecox f. prevernalis) intensive cultivation on water quality in water systems.
Scientia Silvae Sinicae , 45 (8 ), 76 -81 . (in Chinese with English abstract)[吴家森 , 姜培坤 , 盛卫星 , 谢秉楼 (2009 ). 雷竹集约栽培对周边河流水质的影响
. 林业科学 , 45 (8 ), 76 -81 .]
[本文引用: 1]
[96]
Wu JS Wu XH Ye F 2005 a). Nutrient distribution and accumulation in Phyllostacys praecox f. prevernalis.
Journal of Bamboo Research , 24 , 29 -31 . (in Chinese with English abstract)[吴家森 , 吴夏华 , 叶飞 (2005 a). 雷竹林营养元素分配与积累
. 竹子研究汇刊 , 24 , 29 -31 .]
[97]
Wu JS Zhou GM Xu QF Yang F 2005 b). Spatial distribution of nutrition element and its relationship with soil nutrients in different years of Phyllostachys pubescens.
Scientia Silvae Sinicae , 41 (3 ), 171 -173 . (in Chinese with English abstract)[吴家森 , 周国模 , 徐秋芳 , 杨芳 (2005 b). 不同年份毛竹营养元素的空间分布及与土壤养分的关系
. 林业科学 , 41 (3 ), 171 -173 .]
[98]
Yan J Li K Peng X Huang Z Liu S Zhang Q 2015 ). The mechanism for exclusion of Pinus massoniana during the succession in subtropical forest ecosystems: Light competition or stoichiometric homoeostasis?
Scientific Report , 5 ,10994
[本文引用: 1]
[99]
Yu Q Elser J He N Wu H Chen Q Zhang G Han X 2011 ). Stoichiometric homeostasis of vascular plants in the Inner Mongolia grassland.
Oecologia , 166 , 1 -10 .
[本文引用: 1]
[100]
Yue CL Wang KH Zhu YM 2005 ). Morphological plasticity of clonal plant Phyllostachys praecox f. prevernalis (Poaceae) in response to nitrogen availability.
Annales Botanici Fennici , 42 , 123 -127 .
[本文引用: 1]
[101]
Zechmeister-Boltenstern S Keiblinger KM Mooshammer M Peñuelas J Richter A Sardans J Wanek W 2015 ). The application of ecological stoichiometry to plant-microbial- soil organic matter transformations.
Ecological Monographs , 85 , 133 -155 .
[本文引用: 1]
[102]
Zeng DH Cheng GS 2005 ). Ecological stoichiometry: A science to explore the complexity of living systems.
Acta Phytoecologica Sinica , 29 , 1007 -1019 .(in Chinese with English abstract) [曾德慧 , 陈广生 (2005 ). 生态化学计量学: 复杂生命系统奥秘的探索. 植物生态学报,
29 ,1007 -1019 .]
[本文引用: 4]
[103]
Zhan AJ Li ZH 2007 ). The nutrition dynamic of N, P, K in umbrella bamboo (Fargesia murielae) before and after flowering.
Journal of Wuhan Botanical Research , 25 , 213 -216 . (in Chinese with English abstract)[詹爱军 , 李兆华 (2007 ). 神农箭竹开花前后植物体内N、P、K元素的动态
. 武汉植物学研究 , 25 , 213 -216 .]
[本文引用: 2]
[104]
Zhang T Li Y Chang S Jiang P Zhou G Liu J Lin L 2013 ). Converting paddy fields to Lei Bamboo (Phyllostachys praecox) stands affected soil nutrient concentrations, labile organic carbon pools, and organic carbon chemical compositions.
Plant and Soil , 367 , 249 -261
[本文引用: 1]
[105]
Zhang XY Chen JL Ye CQ Liang WX Zhang BS Li QP 1995 ). Study on the relationship between nutrition dynamics and yield of Phyllostachys pubescens.
Forest Research , 8 , 477 -482 . (in Chinese with English abstract)[张献义 , 陈金林 , 叶长青 , 梁文焰 , 张碧松 , 李启鹏 (1995 ). 毛竹林养分动态与产量关系的研究
. 林业科学研究 , 8 , 477 -482 .]
[106]
Zheng YS Hong W 1998). Moso Bamboo Management . Xiamen University Press, Xiamen . (in Chinese)[郑郁善 , 洪伟 (1998). 毛竹经营学. 厦门大学出版社 , 厦门.]
[107]
Zheng YS Hong W Chen LG Qiu EF 1998 ). Study on correlation between soil fertility and bamboo growth and leaves nutrient.
Scientia Silvae Sinicae , 34 (S1 ), 65 -68 .(in Chinese with English abstract) [郑郁善 , 洪伟 , 陈礼光 , 邱尔发 (1998 ). 竹林生长及竹叶养分和土壤肥力相关研究. 林业科学,
34(S1) , 65 -68 .]
[本文引用: 5]
[108]
Zhou GM Jiang PK 2004 ). Density, storage and spatial distribution of carbon in Phyllostachy pubescens forest.
Scientia Silvae Sinicae , 40 (6 ), 20 -24 . (in Chinese with English abstract)[周国模 , 姜培坤 (2004 ). 毛竹林的碳密度和碳贮量及其空间分布
. 林业科学 , 40 (6 ), 20 -24 .]
[本文引用: 1]
[109]
Zhou XR Wang JH Zhang H Wang JC Zhang YB 2012 ). Nutrient allocation pattern of Dendrocalamus latiflorus forest ecosystem planted on hill.
Acta Ecologica Sinica , 24 , 2693 -2699 . (in Chinese with English abstract)[周先容 , 汪建华 , 张红 , 王进闯 , 张远彬 (2012 ). CO2 浓度升高和模拟氮沉降对青川箭竹叶营养质量的影响
. 生态学报 , 24 , 2693 -2699 .]
[110]
Zhuang MH Chen SL Li YC Guo ZW Yang QP 2013 a). Effects of increased concentrations of gas CO2 on mineral ion uptake, transportation and distribution in Phyllostachys edulis.
Acta Ecologica Sinica , 33 , 4297 -4305 . (in Chinese with English abstract)[庄明浩 , 陈双林 , 李迎春 , 郭子武 , 杨清平 (2013 a). CO2 浓度升高对毛竹器官矿质离子吸收、运输和分配的影响
. 生态学报 , 33 , 4297 -4305 .]
[111]
Zhuang MH Li YC Guo ZW Yang QP Chen SM 2013 b). Effects of elevated CO2 on the leaf nutrient stoichiometrical characteristics in Phyllostachys edulis and Oligostachyum lubricum.
Plant Nutrition and Fertilizer Science , 19 , 239 -245 . (in Chinese with English abstract)[庄明浩 , 李迎春 , 郭子武 , 杨清平 , 陈双林 (2013 b). CO2 浓度升高对毛竹和四季竹叶片主要养分化学计量特征的影响
. 植物营养与肥料学报 , 19 , 239 -245 .]
Biodiversity effects on plant stoichiometry.
1
2013
... 另外, 生态化学计量学特别重视生物与环境的矛盾统一, 认为生物元素内平衡需求与环境供应间的适配(match)与错配(mismatch)关系是推动生命发展的根本动力(Sterner & Elser, 2002 ), 也是环境变化的重要原因之一.一方面环境元素会影响生物的内平衡和生长速率(Méndez & Karlsson, 2005 ; Rivas-Ubach et al ., 2012 ); 另一方面生物会通过吸收和释放体内元素而改变环境, 进而对生物产生影响(Sterner & Elser, 2002 ).因此, 生物与环境间连续而复杂的元素化学计量适配或错配反馈关系推动着生物与环境的协同进化(Abbas et al ., 2013 ). ...
The mineral nutrition of wildland plants re-visited: A re-evaluation of processes and patterns.
1
2000
... 生物有机体生态化学计量内平衡易受到许多内因和外因的影响, 不同物种、器官、年龄、发育阶段化学计量特征有所不同, 而且土壤养分、气候、水分、人类活动等环境因子都会直接或间接地影响化学计量内平衡(Aerts & Chapin, 2000 ; Ågren & Weih, 2012 )(图1 ). ...
Stoichiometry and nutrition of plant growth in natural communities.
1
2008
... 生长速率假说(growth rate hypothesis)是解析有机体维持内平衡的机制性假说.该假说认为有机体生长速率与其体内元素化学计量比率紧密联系.高生长速率往往对应高N:C、P:C以及较低的N:P.因为有机体快速生长需要大量富含P的核糖体(蛋白质的合成场所)和RNA、DNA、ATP等物质(Elser et al ., 2003 ; Reef et al ., 2010 ).生长速率假说是解析细胞生物学、生物新陈代谢、种群动态和生态系统功能的理论框架(Niklas et al ., 2005 ; Ågren, 2008 ). ...
Plant stoichiometry at different scales: Element concentration patterns reflect environment more than genotype.
1
2012
... 生物有机体生态化学计量内平衡易受到许多内因和外因的影响, 不同物种、器官、年龄、发育阶段化学计量特征有所不同, 而且土壤养分、气候、水分、人类活动等环境因子都会直接或间接地影响化学计量内平衡(Aerts & Chapin, 2000 ; Ågren & Weih, 2012 )(图1 ). ...
Stoichio- metry: Linking elements to biochemicals.
1
2004
... 通过生命元素这一线索, 生态化学计量学将不同层次的生命活动统一起来, 也将不同生命学科统一起来, 并形成两个被人们广泛接受的基础理论(或假说)(Sterner & Elser, 2002 ; Anderson et al ., 2004 ), 即化学计量内稳性假说和生长速率假说. ...
森林群落植物多样性对毛竹入侵的响应及动态变化
1
2013
... 竹子扩张会降低邻近森林植物多样性, 影响生态系统的稳定性(白尚斌等, 2013 ).初步研究认为毛竹扩张间接影响化学计量内平衡是植物多样性下降的重要机制.毛竹扩张改变了邻近森林土壤的养分状况(宋庆妮等, 2013 ), 而不同植物对土壤养分变化的响应差异较大, 其中栲(Castanopsis fargesii )、红楠(Machilus thunbergii )、交让木(Daphniphyllum macropodum )、赤杨叶(Alniphyllum fortunei )、金钱松(Pseudolarix amabilis )等内平衡调节能力较弱的植物最终退出竹木混交林, 而南方红豆杉(Taxus chinensis var. mairei )、钩锥(Castanopsis tibetana )、黄牛奶树(Symplocos laurina )等内平衡调节和可塑性能力较强的植物生长良好(杨清培, 未发表).然而, 这方面的研究才刚刚起步, 今后应借鉴入侵植物化学计量学的研究成果(González et al ., 2010 )深入开展竹阔混交林组成与稳定性的生态化学计量学机制研究. ...
Rhizosphere stoichiometry: Are C:N: P ratios of plants, soils, and enzymes conserved at the plant species-level?
1
2014
... 植物养分主要来源于土壤, 土壤养分也受到植物的反作用.因此, 加强竹子与土壤养分元素的化学计量适配与错配互作研究, 对竹林生产、环境保护至关重要.首先, 土壤环境决定着养分形态、数量及其有效性, 从而影响到竹子的生长发育(Fan et al ., 2015 ; Zechmeister-Boltenstern et al ., 2015 ).建议开展土壤环境因子与土壤、竹子养分元素化学计量特征动态关系的长期监测与研究.其次, 凋落物分解和根际效应是影响土壤养分的一个重要过程, 尤其是根际过程更应受到关注(Bell et al ., 2014 , 2015 ; Carrillo et al ., 2014 ), 所以应采用稳定同位素技术、分子生态技术, 聚焦于竹子根际过程及养分活化机制(如根系吸收与分泌、根际酸碱度变化、根际微生物活动) , 实现“竹子-根际-土壤”三者元素关系的量化分析. ...
Plant nitrogen uptake drives rhizosphere bacterial community assembly during plant growth.
1
2015
... 植物养分主要来源于土壤, 土壤养分也受到植物的反作用.因此, 加强竹子与土壤养分元素的化学计量适配与错配互作研究, 对竹林生产、环境保护至关重要.首先, 土壤环境决定着养分形态、数量及其有效性, 从而影响到竹子的生长发育(Fan et al ., 2015 ; Zechmeister-Boltenstern et al ., 2015 ).建议开展土壤环境因子与土壤、竹子养分元素化学计量特征动态关系的长期监测与研究.其次, 凋落物分解和根际效应是影响土壤养分的一个重要过程, 尤其是根际过程更应受到关注(Bell et al ., 2014 , 2015 ; Carrillo et al ., 2014 ), 所以应采用稳定同位素技术、分子生态技术, 聚焦于竹子根际过程及养分活化机制(如根系吸收与分泌、根际酸碱度变化、根际微生物活动) , 实现“竹子-根际-土壤”三者元素关系的量化分析. ...
Diet optimization in a generalist herbivore: The moose.
1
1978
... 生态化学计量学主要是从元素组成及比率的角度来研究生物与环境关系, 是近年生态学中飞速发展的领域.自1840年德国农业化学家Justus von Liebig发现植物生长与元素含量的关系并提出了“最小因子定律”以来, 经过生态位理论(Grubb, 1977 )、最佳取食理论(Belovsky, 1978 )、资源比理论(Tilman, 1982 )、养分利用理论(Vitousek, 1982 )、Redfield比值(Redfield, 1958 )等理论的发展, Reiners (1986)提出“Chemical Stoichiometry of the Biota”, 将化学计量学理论用于研究生态系统结构与功能, 到2002年, Sterner和Elser出版专著《Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere》, 系统地阐述了生态化学计量学的定义、基本理论及其应用范畴, 标志着生态化学计量学体系的逐渐成熟, 也为生物学统一化理论构建提供了新思路(Sterner & Elser, 2002 ; 曾德慧和陈广生, 2005 ; 贺金生和韩兴国, 2010 ).当前, 生态化学计量学已发展成从生物大分子到生态系统, 从微生物、动物到植物, 从营养学、生态学到环境科学多层次、多学科都相互关联的综合性理论, 并逐渐被应用于预测或解决区域乃至全球的生态环境问题(曾德慧和陈广生, 2005 ).其理论逻辑如下: ...
Global biodiversity, stoichiometry and ecosystem function responses to human-induced C-N-P imbalances.
1
2015
... 生命元素通过物质循环、能量流动等形式将生态系统的各组分、多层次紧密联系起来(Carnicer et al ., 2015 ).竹林是一个人工或半人工的开放生态系统, 几乎每年都有竹材(或竹笋)等产品输出和外源养分的输入.产品输出和养分输入的失衡会改变竹子、土壤、微生物等不同组分的化学计量特征(Raghubanshi, 1994 ; Venkatesh et al ., 2005 ; Zhang et al ., 2013 ).因此, 今后应充分考虑生物与环境间的相互渗透性, 根据物质守恒与能量守恒定律, 综合考虑土壤、气候、经营活动等因素, 加强竹林生态系统的生态化学计量模型与理论研究, 构建“竹子-土壤-微生物”多组分的“生态化学计量匹配模型”, 竹子化学计量特征与群落组成结构、生态系统生产力多层次的多重养分优化计量模型, 并通过长期野外监测与微宇宙模拟实验, 找到具有竹子特色的“特征参数”, 最后形成可视化的竹林养分预测和管理系统, 指导竹林健康经营和环境综合效应评估, 实现因地制宜、以产定肥的竹林生态系统生态化学计量优化管理模式. ...
Plant rhizosphere influence on microbial C metabolism: The role of elevated CO2 , N availability and root stoichiometry.
1
2014
... 植物养分主要来源于土壤, 土壤养分也受到植物的反作用.因此, 加强竹子与土壤养分元素的化学计量适配与错配互作研究, 对竹林生产、环境保护至关重要.首先, 土壤环境决定着养分形态、数量及其有效性, 从而影响到竹子的生长发育(Fan et al ., 2015 ; Zechmeister-Boltenstern et al ., 2015 ).建议开展土壤环境因子与土壤、竹子养分元素化学计量特征动态关系的长期监测与研究.其次, 凋落物分解和根际效应是影响土壤养分的一个重要过程, 尤其是根际过程更应受到关注(Bell et al ., 2014 , 2015 ; Carrillo et al ., 2014 ), 所以应采用稳定同位素技术、分子生态技术, 聚焦于竹子根际过程及养分活化机制(如根系吸收与分泌、根际酸碱度变化、根际微生物活动) , 实现“竹子-根际-土壤”三者元素关系的量化分析. ...
不同坡位糙花少穗竹林养分分配格局
1
2007
... 再次, 年龄也是影响竹子化学计量特征的因子.1-6年内, 毛竹秆C含量随秆龄增长呈上升趋势, N、P含量呈下降趋势(吴家森等, 2005b ; 刘广路等, 2010 ), 故C:N、C:P随年龄增长而明显增加.不同竹叶(叶龄)差异也较大, 一年生新叶N:P、N:K为13.35、4.78, 2年老叶为21.45、5.23, 老叶N:P比值明显大于幼叶(刘广路等, 2010 ).杨清培发现毛竹幼龄鞭N:P (7.0)明显低于老龄鞭(15.0) (未发表).糙花少穗竹(Oligostachyum scabriflorum )也表现出同样的规律, 1-2年竹秆N:P为13.68-13.86, 而3、4年竹秆分别为20.18和27.88 (陈世品等, 2007 ).另外, 不同季节竹子也表现出不同的化学计量特征, 生长季N:P明显偏低(黄伯惠, 1983 ).说明竹子化学计量特征遵循“生长速率假说”. ...
鄂南毛竹林的养分状况与营养诊断标准
4
2004
... 洪顺山等(1989) 和张献义等(1995)发现竹子叶片养分状况与新竹产量密切相关, 并将之作为竹林养分诊断的重要指标.此后, 陈卫文等(2004) 采用“临界值”法, 制定了毛竹不同养分诊断的化学计量标准(表1 ). ...
... 毛竹叶片养分水平分级标准(陈卫文等, 2004 ) ...
... Criteria for classification of nutrient levels in Phyllostachys edulis leaves (Chen et al ., 2004 ) ...
... 养分诊断与平衡施肥是提高林业生产、维护系统稳定、选择合适管理策略的重要手段(Park et al ., 2014 ), 但当前竹林养分诊断仍停留在单一的养分测定(陈卫文等, 2004 ; 郭晓敏等, 2013 ), 很少考虑多养分、多组织的耦合效应, 失衡超量施肥强烈影响到竹林土壤理化性质, 并引起竹笋食用安全、水土环境恶化等问题.今后应以生态化学计量平衡理论为基础, 改进养分诊断方法, 根据不同竹种、不同林分、不同时期养分限制性的差异特征, 筛选养分丰缺指标, 建立多元素、多组分的养分诊断技术, 提高监测精度与预测能力; 形成生态化学计量平衡施肥技术, 根据竹子和土壤的营养特点, 筛选肥料种类、确定肥料用量与配比, 实施时空精准施肥, 提高肥料利用效率, 实现多重元素间的平衡, 竹子养分需求与环境养分供给间的平衡, 肥料投入与经济产出间的平衡, 产品生产与环境保护间的平衡. ...
连续施肥对毛竹林生长特征及生物量空间构型的影响
2012
毛竹叶片营养与土壤肥力及产量模型的建立
1
2009
... 除遗传因素外, 土壤、气候、人类活动等环境因素也会直接或间接影响竹子化学计量特征.顾大形等(2011)发现, 土壤N、P养分供应水平及土壤N:P会影响四季竹叶片N:P值, 但土壤对竹子养分计量特征的影响较复杂(陈志阳等, 2009 ).除养分含量外, 土壤有机质含量、pH值、土壤微生物等都会影响竹子体内养分含量与比率(郑郁善等, 1998 ), 所以竹林的地下-地上化学计量关系有待深入研究. ...
Nutritional properties of bamboo shoots: Potential and prospects for utilization as a health food.
1
2011
... 竹子(竹亚科植物的总称)生长快速、材质优良, 被广泛用于建材与家居; 竹笋味道鲜美、营养丰富, 备受人们青睐(江泽慧, 2007 ; Chongtham et al ., 2011 ).仅2001年全球竹材消费就高达3000万t、竹笋消费200万t (Kleinhenz et al ., 2000 ; Kleinhenz & Midmore, 2001 ), 年贸易额高达45亿美元(Kleinhenz & Midmore, 2001 ).我国竹林面积近600万hm2 , 产值1600亿元, 并呈逐年上升的趋势(董杰和张燕, 2015 ).竹产品需求持续增长, 大大促进了竹林发展, 但不合理经营导致了部分竹林衰退(楼一平等, 1997 )、产量下降(孟赐福等, 2009 )、地力退化(楼一平, 2001 ; Guo et al ., 2014 )、环境污染(吴家森等, 2009 )等生产和生态问题.这些问题既与竹子自身的生长发育、开花结实等生物习性有关, 也与人类采收笋材、垦复施肥等经营活动相关.然而, 前人仅从竹子生物学、生态学和培育学等方面进行了零散的研究, 不利于以上问题的综合解决.因此, 竹子领域研究亟需一个统一化理论做指导. ...
C:N ratio increases in the phloem sap during floral transition of the long-day plants Sinapis alba and Arabidopsis thaliana.
1
2002
... 同时, 元素比例变化也是竹子开花的另一个重要原因(何奇江等, 2005 ; 丁兴萃, 2006 ).丁兴萃(2006)发现, 早竹(Phyllostachys violascens )开花体内C:N明显上升, 符合C:N花芽分化机理(Corbesier et al ., 2002 ); 何奇江等(2005)也发现, 开花雷竹叶、秆和鞭的P:K分别是未开花竹的1.97倍、2.80倍和2.24倍.另外, 开花往往导致养分流失、比率失衡, 促使竹子死亡(何奇江等, 2005 ; 丁兴萃, 2006 ; 詹爱军和李兆华, 2007 ). ...
早竹衰老开花生理学基础研究
3
2006
... 生态化学计量学是从元素计量的角度来探讨生命运动的内在规律的理论(曾德慧和陈广生, 2005 ; 贺金生和韩兴国, 2010 ).它通过有机体内元素含量及比率关系, 将生物的生长发育、健康状况、行为方式、生态系统动态、生态环境保护等多层次、多学科问题联系起来, 现已成为生态学、环境科学和生物学领域最流行的理论框架(Sterner & Elser, 2002 ; Hillebrand et al ., 2014 ).在竹子经营与研究过程中, 虽然人们早已发现竹子开花(何奇江等, 2005 ; 丁兴萃, 2006 )、发笋大小年(费世民, 2011 )、竹笋品质形成(林海萍等, 2004 )与体内养分含量及比例有关, 竹林土壤污染主要是因为养分搭配不当和总量盈余(孟赐福等, 2009 ), 也提出了平衡施肥与养分管理技术(郭晓敏等, 2013 ), 但尚未形成一个从竹子基础生物学到生产经营的有关养分元素的统一化理论, 极大地影响了竹林生产和生态环境建设. ...
... 同时, 元素比例变化也是竹子开花的另一个重要原因(何奇江等, 2005 ; 丁兴萃, 2006 ).丁兴萃(2006)发现, 早竹(Phyllostachys violascens )开花体内C:N明显上升, 符合C:N花芽分化机理(Corbesier et al ., 2002 ); 何奇江等(2005)也发现, 开花雷竹叶、秆和鞭的P:K分别是未开花竹的1.97倍、2.80倍和2.24倍.另外, 开花往往导致养分流失、比率失衡, 促使竹子死亡(何奇江等, 2005 ; 丁兴萃, 2006 ; 詹爱军和李兆华, 2007 ). ...
... ; 丁兴萃, 2006 ; 詹爱军和李兆华, 2007 ). ...
毛竹扩鞭成林新竹生长效果研究
1
2003
... 土壤养分不足时, 竹子可调整鞭根形态、分布格局而表现出明显的觅食行为(Li et al ., 2000 ; 刘骏等, 2013 ), 以获取养分资源, 维持体内养分平衡.施肥试验表明, 毛竹竹鞭穿越养分异质环境时, 它会避开贫养斑块, 将大量竹笋有选择地放置在富养斑块(Li et al ., 2000 ).随着土壤N含量的增加, 雷竹地上竹子数量增多、生物量增大, 而且地下竹鞭长度和鞭节长度显著降低(Yue et al ., 2005 ), 表现出明显的觅食特征.毛竹扩鞭试验表明, 林缘施肥不仅可以增加新竹数量、降低退笋率, 而且可扩大新竹扩展范围, 施肥组每年平均扩张5 m 以上(董晨玲, 2003 ).这些可间接说明, 竹林向邻近森林扩张可能是为了维持体内养分平衡而表现出来的一种觅食 行为. ...
发展竹产业促进中非南南合作
1
2015
... 竹子(竹亚科植物的总称)生长快速、材质优良, 被广泛用于建材与家居; 竹笋味道鲜美、营养丰富, 备受人们青睐(江泽慧, 2007 ; Chongtham et al ., 2011 ).仅2001年全球竹材消费就高达3000万t、竹笋消费200万t (Kleinhenz et al ., 2000 ; Kleinhenz & Midmore, 2001 ), 年贸易额高达45亿美元(Kleinhenz & Midmore, 2001 ).我国竹林面积近600万hm2 , 产值1600亿元, 并呈逐年上升的趋势(董杰和张燕, 2015 ).竹产品需求持续增长, 大大促进了竹林发展, 但不合理经营导致了部分竹林衰退(楼一平等, 1997 )、产量下降(孟赐福等, 2009 )、地力退化(楼一平, 2001 ; Guo et al ., 2014 )、环境污染(吴家森等, 2009 )等生产和生态问题.这些问题既与竹子自身的生长发育、开花结实等生物习性有关, 也与人类采收笋材、垦复施肥等经营活动相关.然而, 前人仅从竹子生物学、生态学和培育学等方面进行了零散的研究, 不利于以上问题的综合解决.因此, 竹子领域研究亟需一个统一化理论做指导. ...
Growth rate-stoichiometry couplings in diverse biota.
1
2003
... 生长速率假说(growth rate hypothesis)是解析有机体维持内平衡的机制性假说.该假说认为有机体生长速率与其体内元素化学计量比率紧密联系.高生长速率往往对应高N:C、P:C以及较低的N:P.因为有机体快速生长需要大量富含P的核糖体(蛋白质的合成场所)和RNA、DNA、ATP等物质(Elser et al ., 2003 ; Reef et al ., 2010 ).生长速率假说是解析细胞生物学、生物新陈代谢、种群动态和生态系统功能的理论框架(Niklas et al ., 2005 ; Ågren, 2008 ). ...
Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change.
1
2010
... 化学计量内稳性假说(stoichiometric homeostasis hypothesis)是指在环境(食物)化学元素组成发生变化的情况下, 生物有机体可通过一系列生理或行为调节以保持其元素组成的相对稳定(Sterner & Elser, 2002 ; Elser et al ., 2010 ).Sterner和Elser (2002)提出有机体-环境化学计量关系公式: y = cx 1/ H y 、x 分别表示有机体、环境元素浓度(或比率), c 是常数, H 是化学计量内稳性指数, 反映有机体化学计量内稳性维持能力的大小.当H = 1时, 该有机体没有任何的化学计量内稳性; H > 1时, 有机体具有维持自身元素平衡的能力, H 越大有机体内稳性越高(Sterner & Elser, 2002 ). ...
Linkages of plant and soil C:N:P stoichiometry and their relationships to forest growth in subtropical plantations.
1
2015
... 植物养分主要来源于土壤, 土壤养分也受到植物的反作用.因此, 加强竹子与土壤养分元素的化学计量适配与错配互作研究, 对竹林生产、环境保护至关重要.首先, 土壤环境决定着养分形态、数量及其有效性, 从而影响到竹子的生长发育(Fan et al ., 2015 ; Zechmeister-Boltenstern et al ., 2015 ).建议开展土壤环境因子与土壤、竹子养分元素化学计量特征动态关系的长期监测与研究.其次, 凋落物分解和根际效应是影响土壤养分的一个重要过程, 尤其是根际过程更应受到关注(Bell et al ., 2014 , 2015 ; Carrillo et al ., 2014 ), 所以应采用稳定同位素技术、分子生态技术, 聚焦于竹子根际过程及养分活化机制(如根系吸收与分泌、根际酸碱度变化、根际微生物活动) , 实现“竹子-根际-土壤”三者元素关系的量化分析. ...
不同用途毛竹林的施肥研究II. 毛竹笋用林丰产经营技术
1991
不同用途毛竹林的施肥研究Ⅰ. 毛竹材用林的施肥
1
1988
... 另外, 并不是所有的竹笋都能长成竹子, 许多竹笋出土前就会死亡, 或虽能出土却死在成竹之前, 这叫退笋.大量研究认为退笋率高低与养分供应水平紧密相关, 增加养分供给可以降低退笋率、提高成竹率(傅懋毅等, 1988 ; Li et al ., 1998b ; 郑郁善等, 1998 ; Li et al ., 2000 ). ...
氮素施肥对毛竹生产力与光合能力的影响
1
2014
... 化学计量特征变化会直接影响到竹子光合作用.当施N量为250 kg·hm-2 时, 与对照相比, 1年、3年、5年生毛竹叶片的光饱和点均达最大, 分别增加了5.2%、9.1%和7.0%, 同时光补偿点降至最低, 分别降低了49.5%、20.4%和31.8% (高培军等, 2014 ), 土壤养分会影响叶片N、P含量与比例, 从而对光合作用产生直接或间接影响(宋艳冬等, 2010 ; 顾大形等, 2011 ).顾大形等(2011)发现一定范围内增加土壤N含量, 四季竹叶片的N含量增加, N:P上升, 提高了叶绿素含量, 促进其光合作用.而当施肥量不足或过量时会降低毛竹叶片光合色素含量, 结果其光合能力受到影响.苏文会(2012)发现当毛竹叶片N:P从20.70下降至15.29时, 其光合产物淀粉含量从35.47 g·kg-1 降至16.21 g·kg-1 , 降幅达54.3%.说明竹子叶片N、P化学计量特征对其光合作用影响较大. ...
Can ecological stoichiometry help explain patterns of biological invasions?
1
2010
... 竹子扩张会降低邻近森林植物多样性, 影响生态系统的稳定性(白尚斌等, 2013 ).初步研究认为毛竹扩张间接影响化学计量内平衡是植物多样性下降的重要机制.毛竹扩张改变了邻近森林土壤的养分状况(宋庆妮等, 2013 ), 而不同植物对土壤养分变化的响应差异较大, 其中栲(Castanopsis fargesii )、红楠(Machilus thunbergii )、交让木(Daphniphyllum macropodum )、赤杨叶(Alniphyllum fortunei )、金钱松(Pseudolarix amabilis )等内平衡调节能力较弱的植物最终退出竹木混交林, 而南方红豆杉(Taxus chinensis var. mairei )、钩锥(Castanopsis tibetana )、黄牛奶树(Symplocos laurina )等内平衡调节和可塑性能力较强的植物生长良好(杨清培, 未发表).然而, 这方面的研究才刚刚起步, 今后应借鉴入侵植物化学计量学的研究成果(González et al ., 2010 )深入开展竹阔混交林组成与稳定性的生态化学计量学机制研究. ...
The maintenance of species-richness in plant communities: The importance of the regeneration niche.
1
1977
... 生态化学计量学主要是从元素组成及比率的角度来研究生物与环境关系, 是近年生态学中飞速发展的领域.自1840年德国农业化学家Justus von Liebig发现植物生长与元素含量的关系并提出了“最小因子定律”以来, 经过生态位理论(Grubb, 1977 )、最佳取食理论(Belovsky, 1978 )、资源比理论(Tilman, 1982 )、养分利用理论(Vitousek, 1982 )、Redfield比值(Redfield, 1958 )等理论的发展, Reiners (1986)提出“Chemical Stoichiometry of the Biota”, 将化学计量学理论用于研究生态系统结构与功能, 到2002年, Sterner和Elser出版专著《Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere》, 系统地阐述了生态化学计量学的定义、基本理论及其应用范畴, 标志着生态化学计量学体系的逐渐成熟, 也为生物学统一化理论构建提供了新思路(Sterner & Elser, 2002 ; 曾德慧和陈广生, 2005 ; 贺金生和韩兴国, 2010 ).当前, 生态化学计量学已发展成从生物大分子到生态系统, 从微生物、动物到植物, 从营养学、生态学到环境科学多层次、多学科都相互关联的综合性理论, 并逐渐被应用于预测或解决区域乃至全球的生态环境问题(曾德慧和陈广生, 2005 ).其理论逻辑如下: ...
土壤氮磷对四季竹叶片氮磷化学计量特征和叶绿素含量的影响
1
2011
... 化学计量特征变化会直接影响到竹子光合作用.当施N量为250 kg·hm-2 时, 与对照相比, 1年、3年、5年生毛竹叶片的光饱和点均达最大, 分别增加了5.2%、9.1%和7.0%, 同时光补偿点降至最低, 分别降低了49.5%、20.4%和31.8% (高培军等, 2014 ), 土壤养分会影响叶片N、P含量与比例, 从而对光合作用产生直接或间接影响(宋艳冬等, 2010 ; 顾大形等, 2011 ).顾大形等(2011)发现一定范围内增加土壤N含量, 四季竹叶片的N含量增加, N:P上升, 提高了叶绿素含量, 促进其光合作用.而当施肥量不足或过量时会降低毛竹叶片光合色素含量, 结果其光合能力受到影响.苏文会(2012)发现当毛竹叶片N:P从20.70下降至15.29时, 其光合产物淀粉含量从35.47 g·kg-1 降至16.21 g·kg-1 , 降幅达54.3%.说明竹子叶片N、P化学计量特征对其光合作用影响较大. ...
不同生产力水平毛竹林碳氮磷的分布格局和计量特征
1
2014
... 竹子(竹亚科植物的总称)生长快速、材质优良, 被广泛用于建材与家居; 竹笋味道鲜美、营养丰富, 备受人们青睐(江泽慧, 2007 ; Chongtham et al ., 2011 ).仅2001年全球竹材消费就高达3000万t、竹笋消费200万t (Kleinhenz et al ., 2000 ; Kleinhenz & Midmore, 2001 ), 年贸易额高达45亿美元(Kleinhenz & Midmore, 2001 ).我国竹林面积近600万hm2 , 产值1600亿元, 并呈逐年上升的趋势(董杰和张燕, 2015 ).竹产品需求持续增长, 大大促进了竹林发展, 但不合理经营导致了部分竹林衰退(楼一平等, 1997 )、产量下降(孟赐福等, 2009 )、地力退化(楼一平, 2001 ; Guo et al ., 2014 )、环境污染(吴家森等, 2009 )等生产和生态问题.这些问题既与竹子自身的生长发育、开花结实等生物习性有关, 也与人类采收笋材、垦复施肥等经营活动相关.然而, 前人仅从竹子生物学、生态学和培育学等方面进行了零散的研究, 不利于以上问题的综合解决.因此, 竹子领域研究亟需一个统一化理论做指导. ...
Effects of mulching management on soil and foliar C, N and P stoichiometry in bamboo (Phyllostachys violascens) plantations.
2014
长期施用不同肥料对红哺鸡竹林叶片碳、氮、磷化学计量特征的影响
2011
N:P ratios in terrestrial plants: Variation and functional significance.
2
2004
... (7)群体中有机体的此消彼长会影响到种群动态、群落演替和生态系统的发展及其生产、生态功能的发挥(Sterner & Elser, 2002 ; Güsewell, 2004 ; Yan et al ., 2015 ). ...
... 营养不足容易引起植物组织老化, 而过剩又引起植物毒害, 造成资源浪费与环境污染(Güsewell, 2004 ).因此, 养分诊断是保证林农业产品优质高产的重要措施, 并将体内营养元素的含量及比例作为衡量植物营养状况的重要指标(Sinclair et al ., 1997 ; Güsewell et al ., 2003 ; Tessier & Raynal, 2003 ). ...
Biomass N:P ratios as indicators of nutrient limitation for plant populations in wetlands.
1
2003
... 营养不足容易引起植物组织老化, 而过剩又引起植物毒害, 造成资源浪费与环境污染(Güsewell, 2004 ).因此, 养分诊断是保证林农业产品优质高产的重要措施, 并将体内营养元素的含量及比例作为衡量植物营养状况的重要指标(Sinclair et al ., 1997 ; Güsewell et al ., 2003 ; Tessier & Raynal, 2003 ). ...
Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China.
2
2005
... 竹子具有较强的维持体内元素相对平衡的能力.郭子武等(2011)发现施肥对红哺鸡竹(Phyllostachys iridescens )叶片C、N、P含量及比值影响不大.其叶片C:N、C:P可保持在18.71-35.02、304.41-458.52, N:P变幅更小, 仅为15.28-17.12.郭宝华等(2014)野外调查发现, 尽管土壤N:P差异明显(4.50-13.69), 但毛竹叶片N:P变化不大(12.60- 13.69).顾大形等(2011)通过盆栽实验发现, 土壤N或P增加2-3倍, 四季竹(Oligostachyum lubricum )仍可保持叶片N:P基本不变, 其内稳性指数H (N:P) = 2.85>1.庄明浩等(2013b)发现当CO2 浓度由360 μmol·mol-1 增至500 μmol·mol-1 时, 毛竹和四季竹叶片养分C、N、P含量及其比例都没有明显变化, 说明竹子在变化的土壤或大气环境中, 都能保持体内元素的相对平衡, 符合生态化学计量学的“内稳性假说”.综合多种竹子的研究结果(周国模和姜培坤, 2004 ; 费世民, 2011 ; 庄明浩等, 2013b ), 发现竹子叶片平均化学计量比值为C:N:P = 380:16:1, 其中N:P高于自然界陆生植物平均值(10:1) (Knecht & Göransson, 2004 ), 也高于国内一般草本植物(13.5:1) (Han et al ., 2005 ), 而与阔叶植物(15.1:1.0)和常绿木本植物的平均值(15.2:1.0)相近(Han et al ., 2005 ), 说明竹子与常绿阔叶木本植物具有相似的“植物-环境”化学计量学响应特征. ...
... ), 而与阔叶植物(15.1:1.0)和常绿木本植物的平均值(15.2:1.0)相近(Han et al ., 2005 ), 说明竹子与常绿阔叶木本植物具有相似的“植物-环境”化学计量学响应特征. ...
生态化学计量学: 探索从个体到生态系统的统一化理论
2
2010
... 生态化学计量学是从元素计量的角度来探讨生命运动的内在规律的理论(曾德慧和陈广生, 2005 ; 贺金生和韩兴国, 2010 ).它通过有机体内元素含量及比率关系, 将生物的生长发育、健康状况、行为方式、生态系统动态、生态环境保护等多层次、多学科问题联系起来, 现已成为生态学、环境科学和生物学领域最流行的理论框架(Sterner & Elser, 2002 ; Hillebrand et al ., 2014 ).在竹子经营与研究过程中, 虽然人们早已发现竹子开花(何奇江等, 2005 ; 丁兴萃, 2006 )、发笋大小年(费世民, 2011 )、竹笋品质形成(林海萍等, 2004 )与体内养分含量及比例有关, 竹林土壤污染主要是因为养分搭配不当和总量盈余(孟赐福等, 2009 ), 也提出了平衡施肥与养分管理技术(郭晓敏等, 2013 ), 但尚未形成一个从竹子基础生物学到生产经营的有关养分元素的统一化理论, 极大地影响了竹林生产和生态环境建设. ...
... 生态化学计量学主要是从元素组成及比率的角度来研究生物与环境关系, 是近年生态学中飞速发展的领域.自1840年德国农业化学家Justus von Liebig发现植物生长与元素含量的关系并提出了“最小因子定律”以来, 经过生态位理论(Grubb, 1977 )、最佳取食理论(Belovsky, 1978 )、资源比理论(Tilman, 1982 )、养分利用理论(Vitousek, 1982 )、Redfield比值(Redfield, 1958 )等理论的发展, Reiners (1986)提出“Chemical Stoichiometry of the Biota”, 将化学计量学理论用于研究生态系统结构与功能, 到2002年, Sterner和Elser出版专著《Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere》, 系统地阐述了生态化学计量学的定义、基本理论及其应用范畴, 标志着生态化学计量学体系的逐渐成熟, 也为生物学统一化理论构建提供了新思路(Sterner & Elser, 2002 ; 曾德慧和陈广生, 2005 ; 贺金生和韩兴国, 2010 ).当前, 生态化学计量学已发展成从生物大分子到生态系统, 从微生物、动物到植物, 从营养学、生态学到环境科学多层次、多学科都相互关联的综合性理论, 并逐渐被应用于预测或解决区域乃至全球的生态环境问题(曾德慧和陈广生, 2005 ).其理论逻辑如下: ...
雷竹开花期内源激素、氨基酸和营养成分含量变化
4
2005
... 生态化学计量学是从元素计量的角度来探讨生命运动的内在规律的理论(曾德慧和陈广生, 2005 ; 贺金生和韩兴国, 2010 ).它通过有机体内元素含量及比率关系, 将生物的生长发育、健康状况、行为方式、生态系统动态、生态环境保护等多层次、多学科问题联系起来, 现已成为生态学、环境科学和生物学领域最流行的理论框架(Sterner & Elser, 2002 ; Hillebrand et al ., 2014 ).在竹子经营与研究过程中, 虽然人们早已发现竹子开花(何奇江等, 2005 ; 丁兴萃, 2006 )、发笋大小年(费世民, 2011 )、竹笋品质形成(林海萍等, 2004 )与体内养分含量及比例有关, 竹林土壤污染主要是因为养分搭配不当和总量盈余(孟赐福等, 2009 ), 也提出了平衡施肥与养分管理技术(郭晓敏等, 2013 ), 但尚未形成一个从竹子基础生物学到生产经营的有关养分元素的统一化理论, 极大地影响了竹林生产和生态环境建设. ...
... 研究表明, 体内元素含量状况会影响竹子花芽分化与成花过程.如P、Ca、Mg等元素含量增加会诱导花芽分化; N、K等元素含量增加会促进营养生长, 而延迟竹子开花(何奇江等, 2005 ; 詹爱军和李兆华, 2007 ). ...
... 同时, 元素比例变化也是竹子开花的另一个重要原因(何奇江等, 2005 ; 丁兴萃, 2006 ).丁兴萃(2006)发现, 早竹(Phyllostachys violascens )开花体内C:N明显上升, 符合C:N花芽分化机理(Corbesier et al ., 2002 ); 何奇江等(2005)也发现, 开花雷竹叶、秆和鞭的P:K分别是未开花竹的1.97倍、2.80倍和2.24倍.另外, 开花往往导致养分流失、比率失衡, 促使竹子死亡(何奇江等, 2005 ; 丁兴萃, 2006 ; 詹爱军和李兆华, 2007 ). ...
... ); 何奇江等(2005)也发现, 开花雷竹叶、秆和鞭的P:K分别是未开花竹的1.97倍、2.80倍和2.24倍.另外, 开花往往导致养分流失、比率失衡, 促使竹子死亡(何奇江等, 2005 ; 丁兴萃, 2006 ; 詹爱军和李兆华, 2007 ). ...
Think ratio! A stoichiometric view on biodiversity-ecosystem functioning research.
1
2014
... 生态化学计量学是从元素计量的角度来探讨生命运动的内在规律的理论(曾德慧和陈广生, 2005 ; 贺金生和韩兴国, 2010 ).它通过有机体内元素含量及比率关系, 将生物的生长发育、健康状况、行为方式、生态系统动态、生态环境保护等多层次、多学科问题联系起来, 现已成为生态学、环境科学和生物学领域最流行的理论框架(Sterner & Elser, 2002 ; Hillebrand et al ., 2014 ).在竹子经营与研究过程中, 虽然人们早已发现竹子开花(何奇江等, 2005 ; 丁兴萃, 2006 )、发笋大小年(费世民, 2011 )、竹笋品质形成(林海萍等, 2004 )与体内养分含量及比例有关, 竹林土壤污染主要是因为养分搭配不当和总量盈余(孟赐福等, 2009 ), 也提出了平衡施肥与养分管理技术(郭晓敏等, 2013 ), 但尚未形成一个从竹子基础生物学到生产经营的有关养分元素的统一化理论, 极大地影响了竹林生产和生态环境建设. ...
毛竹营养诊断的研究
1
1989
... 洪顺山等(1989) 和张献义等(1995)发现竹子叶片养分状况与新竹产量密切相关, 并将之作为竹林养分诊断的重要指标.此后, 陈卫文等(2004) 采用“临界值”法, 制定了毛竹不同养分诊断的化学计量标准(表1 ). ...
毛竹林施肥效应研究
1
1992
... 合理计量施肥不但可以增加竹材(笋)产量, 而且可以提高品质.傅懋毅等(1991)发现每年沟施复合肥3945 kg·hm-2 (N:P:K = 4:3:1), 则每度(两年)可增产春笋20000 kg·hm-2 , 并认为这是毛竹笋用林最佳培育方式; 在澳大利亚每年折合施N 250、375、500 kg·hm-2 的复合肥(N:P:K = 5:1:2.8), 毛竹笋产量分别为8300、10200、14200 kg·hm-2 (Kleinhenz et al ., 2003 ).陈孝丑等(2012)发现施肥可以显著提高毛竹林的胸径.林海萍等(2004)筛选了优质雷竹笋用林最佳配方, 当施肥量为尿素975 kg·hm-2 、复合肥1500 kg·hm-2 、厩肥112500 kg·hm-2 (N:P:K = 10:2:1)时, 最有利于雷竹笋总糖、脂肪及淀粉的积累, 而减少灰分、蛋白质及游离氨基酸的含量, 品质提高效果最好.另外, 孟勇等(2014)发现添加一些非必需元素(如硒Se)对竹笋品质具有明显的改善作用.但也有实验发现仅施化肥会使竹壁变薄, 竹材品质下降(洪顺山等, 1992 ).有机肥对竹材(笋)品质形成化学计量学机制值得关注. ...
毛竹矿质营养元素动态的研究
2
1983
... 再次, 年龄也是影响竹子化学计量特征的因子.1-6年内, 毛竹秆C含量随秆龄增长呈上升趋势, N、P含量呈下降趋势(吴家森等, 2005b ; 刘广路等, 2010 ), 故C:N、C:P随年龄增长而明显增加.不同竹叶(叶龄)差异也较大, 一年生新叶N:P、N:K为13.35、4.78, 2年老叶为21.45、5.23, 老叶N:P比值明显大于幼叶(刘广路等, 2010 ).杨清培发现毛竹幼龄鞭N:P (7.0)明显低于老龄鞭(15.0) (未发表).糙花少穗竹(Oligostachyum scabriflorum )也表现出同样的规律, 1-2年竹秆N:P为13.68-13.86, 而3、4年竹秆分别为20.18和27.88 (陈世品等, 2007 ).另外, 不同季节竹子也表现出不同的化学计量特征, 生长季N:P明显偏低(黄伯惠, 1983 ).说明竹子化学计量特征遵循“生长速率假说”. ...
... 发笋数量、成竹质量都直接影响到竹林生态功能与生产价值, 然而有些竹子(如毛竹、毛环竹(Phyllostachys meyeri ))存在明显的发笋成竹大小年现象, 即一年大量发笋长竹, 一年行鞭换叶, 产笋较少, 每两年为一周期(南京林产工业学院竹类研究室, 1974 ).大小年现象虽有遗传原因, 但与体内N、P、K等主要矿质营养元素的化学计量特征密切相关, 如吴家森(2005b)发现小年毛竹叶中N、P、K含量均显著高于大年.Li等(1998a)发现: 竹叶被竹蝗取食后, 光合产物减少; 改变竹子体内养分状况, 可使大年变小年, 小年变大年.在实际生产实践中, 通过施肥、小年留笋养竹、大年疏笋等措施, 可将大小年竹林改造成“花年”竹林(黄伯惠, 1983 ). ...
Why bamboos wait so long to flower.
1
1976
... 竹子很少开花, 但一旦开花就造成竹子大面积的突发性死亡, 因此竹子开花一直是一个有趣而神秘的话题(Janzen, 1976 ; Takahashi et al ., 2007 ).近100年来, 有关竹子开花分别从气候、管理、环境等方面提出了不同的假说(郑郁善和洪伟, 1998 ), 其中营养计量学说是目前比较普遍认可的一个重要机制性假说. ...
苦竹各器官主要营养元素分布及采伐的养分输出
1
2007
... 首先, 不同竹种的化学计量学特征有所差异.毛竹叶片N、P、K含量分别为21.44 g·kg-1 、1.33 g·kg-1 、13.15 g·kg-1 (费世民, 2011 ), 雷竹(Phyllos- tachys praecox ‘ Prevernalis’)叶片N、P、K含量分别为8.86 g·kg-1 、1.07 g·kg-1 、6.55 g·kg-1 (吴家森等, 2005a ).毛竹叶片N:P大于雷竹, 而P:K小于雷竹.麻竹(Dendrocalamus latiflorus )叶片N、P、K含量分别为56.9 g·kg-1 、4.4 g·kg-1 、8.1 g·kg-1 (邱尔发等, 2004 ), 苦竹(Pleioblastus amarus )叶片N、P、K含量分别为24.26 g·kg-1 、1.51 g·kg-1 、11.56 g·kg-1 (蒋俊明等, 2007 ), 麻竹叶片N、P、K含量都明显高于苦竹.不同竹种对环境养分变化的化学计量学响应的程度也有差异(Piouceau et al ., 2014 ). ...
磷钾硼肥对红竹笋品质效应的研究
2
2000
... 竹子体内含有30-40种大量元素和微量元素, 它们都与竹子生长发育、竹材(笋)品质等生物学特性密切相关(蒋式洪等, 2000 ; 费世民, 2011 ).C、N、P、K是竹子的基本组成元素, C是生命的骨架元素与能量元素, 毛竹(Phyllostachys edulis )叶片C含量((512.13 ± 11.13) mg·g-1 )高于世界陆生植物的平均水平((464.32 ± 32.10) mg·g-1 ).N、P、K不仅影响竹子发笋数量, 也会影响竹笋品质(郑郁善等, 1998 ; 蒋式洪等, 2000 ); Ca、Mg分别与竹子根系生长和叶绿素形成有密切关系.另外, Si尽管不是生命必需元素, 但竹子体内Si含量特别高, 近年备受关注(潘月等, 2013 ; 戎洁庆等, 2013 ; Umemura & Takenaka, 2014 ). ...
... ; 蒋式洪等, 2000 ); Ca、Mg分别与竹子根系生长和叶绿素形成有密切关系.另外, Si尽管不是生命必需元素, 但竹子体内Si含量特别高, 近年备受关注(潘月等, 2013 ; 戎洁庆等, 2013 ; Umemura & Takenaka, 2014 ). ...
Storage methods for extending shelf life of fresh, edible bamboo shoots [Bambusa oldhamii (Munro)].
1
2000
... 竹子(竹亚科植物的总称)生长快速、材质优良, 被广泛用于建材与家居; 竹笋味道鲜美、营养丰富, 备受人们青睐(江泽慧, 2007 ; Chongtham et al ., 2011 ).仅2001年全球竹材消费就高达3000万t、竹笋消费200万t (Kleinhenz et al ., 2000 ; Kleinhenz & Midmore, 2001 ), 年贸易额高达45亿美元(Kleinhenz & Midmore, 2001 ).我国竹林面积近600万hm2 , 产值1600亿元, 并呈逐年上升的趋势(董杰和张燕, 2015 ).竹产品需求持续增长, 大大促进了竹林发展, 但不合理经营导致了部分竹林衰退(楼一平等, 1997 )、产量下降(孟赐福等, 2009 )、地力退化(楼一平, 2001 ; Guo et al ., 2014 )、环境污染(吴家森等, 2009 )等生产和生态问题.这些问题既与竹子自身的生长发育、开花结实等生物习性有关, 也与人类采收笋材、垦复施肥等经营活动相关.然而, 前人仅从竹子生物学、生态学和培育学等方面进行了零散的研究, 不利于以上问题的综合解决.因此, 竹子领域研究亟需一个统一化理论做指导. ...
Aspects of bamboo agronomy.
2
2001
... 竹子(竹亚科植物的总称)生长快速、材质优良, 被广泛用于建材与家居; 竹笋味道鲜美、营养丰富, 备受人们青睐(江泽慧, 2007 ; Chongtham et al ., 2011 ).仅2001年全球竹材消费就高达3000万t、竹笋消费200万t (Kleinhenz et al ., 2000 ; Kleinhenz & Midmore, 2001 ), 年贸易额高达45亿美元(Kleinhenz & Midmore, 2001 ).我国竹林面积近600万hm2 , 产值1600亿元, 并呈逐年上升的趋势(董杰和张燕, 2015 ).竹产品需求持续增长, 大大促进了竹林发展, 但不合理经营导致了部分竹林衰退(楼一平等, 1997 )、产量下降(孟赐福等, 2009 )、地力退化(楼一平, 2001 ; Guo et al ., 2014 )、环境污染(吴家森等, 2009 )等生产和生态问题.这些问题既与竹子自身的生长发育、开花结实等生物习性有关, 也与人类采收笋材、垦复施肥等经营活动相关.然而, 前人仅从竹子生物学、生态学和培育学等方面进行了零散的研究, 不利于以上问题的综合解决.因此, 竹子领域研究亟需一个统一化理论做指导. ...
... ), 年贸易额高达45亿美元(Kleinhenz & Midmore, 2001 ).我国竹林面积近600万hm2 , 产值1600亿元, 并呈逐年上升的趋势(董杰和张燕, 2015 ).竹产品需求持续增长, 大大促进了竹林发展, 但不合理经营导致了部分竹林衰退(楼一平等, 1997 )、产量下降(孟赐福等, 2009 )、地力退化(楼一平, 2001 ; Guo et al ., 2014 )、环境污染(吴家森等, 2009 )等生产和生态问题.这些问题既与竹子自身的生长发育、开花结实等生物习性有关, 也与人类采收笋材、垦复施肥等经营活动相关.然而, 前人仅从竹子生物学、生态学和培育学等方面进行了零散的研究, 不利于以上问题的综合解决.因此, 竹子领域研究亟需一个统一化理论做指导. ...
A case study on the effects of irrigation and fertilization on soil water and soil nutrient status, and on growth and yield of bamboo (Phyllostachys pubescens) shoots.
1
2003
... 合理计量施肥不但可以增加竹材(笋)产量, 而且可以提高品质.傅懋毅等(1991)发现每年沟施复合肥3945 kg·hm-2 (N:P:K = 4:3:1), 则每度(两年)可增产春笋20000 kg·hm-2 , 并认为这是毛竹笋用林最佳培育方式; 在澳大利亚每年折合施N 250、375、500 kg·hm-2 的复合肥(N:P:K = 5:1:2.8), 毛竹笋产量分别为8300、10200、14200 kg·hm-2 (Kleinhenz et al ., 2003 ).陈孝丑等(2012)发现施肥可以显著提高毛竹林的胸径.林海萍等(2004)筛选了优质雷竹笋用林最佳配方, 当施肥量为尿素975 kg·hm-2 、复合肥1500 kg·hm-2 、厩肥112500 kg·hm-2 (N:P:K = 10:2:1)时, 最有利于雷竹笋总糖、脂肪及淀粉的积累, 而减少灰分、蛋白质及游离氨基酸的含量, 品质提高效果最好.另外, 孟勇等(2014)发现添加一些非必需元素(如硒Se)对竹笋品质具有明显的改善作用.但也有实验发现仅施化肥会使竹壁变薄, 竹材品质下降(洪顺山等, 1992 ).有机肥对竹材(笋)品质形成化学计量学机制值得关注. ...
Terrestrial plants require nutrients in similar proportions.
1
2004
... 竹子具有较强的维持体内元素相对平衡的能力.郭子武等(2011)发现施肥对红哺鸡竹(Phyllostachys iridescens )叶片C、N、P含量及比值影响不大.其叶片C:N、C:P可保持在18.71-35.02、304.41-458.52, N:P变幅更小, 仅为15.28-17.12.郭宝华等(2014)野外调查发现, 尽管土壤N:P差异明显(4.50-13.69), 但毛竹叶片N:P变化不大(12.60- 13.69).顾大形等(2011)通过盆栽实验发现, 土壤N或P增加2-3倍, 四季竹(Oligostachyum lubricum )仍可保持叶片N:P基本不变, 其内稳性指数H (N:P) = 2.85>1.庄明浩等(2013b)发现当CO2 浓度由360 μmol·mol-1 增至500 μmol·mol-1 时, 毛竹和四季竹叶片养分C、N、P含量及其比例都没有明显变化, 说明竹子在变化的土壤或大气环境中, 都能保持体内元素的相对平衡, 符合生态化学计量学的“内稳性假说”.综合多种竹子的研究结果(周国模和姜培坤, 2004 ; 费世民, 2011 ; 庄明浩等, 2013b ), 发现竹子叶片平均化学计量比值为C:N:P = 380:16:1, 其中N:P高于自然界陆生植物平均值(10:1) (Knecht & Göransson, 2004 ), 也高于国内一般草本植物(13.5:1) (Han et al ., 2005 ), 而与阔叶植物(15.1:1.0)和常绿木本植物的平均值(15.2:1.0)相近(Han et al ., 2005 ), 说明竹子与常绿阔叶木本植物具有相似的“植物-环境”化学计量学响应特征. ...
a). Biennial variation in production of new shoots in groves of the giant bamboo Phyllostachys pubescens in Sichuan, China.
1998
b). Carbon and nutrient dynamics in relation to growth rhythm in the giant bamboo Phyllostachys pubescens.
1998
Interactions between shoot age structure, nutrient availability and physiological integration in the giant bamboo Phyllostachys pubescens.
3
2000
... 另外, 并不是所有的竹笋都能长成竹子, 许多竹笋出土前就会死亡, 或虽能出土却死在成竹之前, 这叫退笋.大量研究认为退笋率高低与养分供应水平紧密相关, 增加养分供给可以降低退笋率、提高成竹率(傅懋毅等, 1988 ; Li et al ., 1998b ; 郑郁善等, 1998 ; Li et al ., 2000 ). ...
... 土壤养分不足时, 竹子可调整鞭根形态、分布格局而表现出明显的觅食行为(Li et al ., 2000 ; 刘骏等, 2013 ), 以获取养分资源, 维持体内养分平衡.施肥试验表明, 毛竹竹鞭穿越养分异质环境时, 它会避开贫养斑块, 将大量竹笋有选择地放置在富养斑块(Li et al ., 2000 ).随着土壤N含量的增加, 雷竹地上竹子数量增多、生物量增大, 而且地下竹鞭长度和鞭节长度显著降低(Yue et al ., 2005 ), 表现出明显的觅食特征.毛竹扩鞭试验表明, 林缘施肥不仅可以增加新竹数量、降低退笋率, 而且可扩大新竹扩展范围, 施肥组每年平均扩张5 m 以上(董晨玲, 2003 ).这些可间接说明, 竹林向邻近森林扩张可能是为了维持体内养分平衡而表现出来的一种觅食 行为. ...
... ), 以获取养分资源, 维持体内养分平衡.施肥试验表明, 毛竹竹鞭穿越养分异质环境时, 它会避开贫养斑块, 将大量竹笋有选择地放置在富养斑块(Li et al ., 2000 ).随着土壤N含量的增加, 雷竹地上竹子数量增多、生物量增大, 而且地下竹鞭长度和鞭节长度显著降低(Yue et al ., 2005 ), 表现出明显的觅食特征.毛竹扩鞭试验表明, 林缘施肥不仅可以增加新竹数量、降低退笋率, 而且可扩大新竹扩展范围, 施肥组每年平均扩张5 m 以上(董晨玲, 2003 ).这些可间接说明, 竹林向邻近森林扩张可能是为了维持体内养分平衡而表现出来的一种觅食 行为. ...
集约经营对毛竹笋品质的影响
1
2004
... 生态化学计量学是从元素计量的角度来探讨生命运动的内在规律的理论(曾德慧和陈广生, 2005 ; 贺金生和韩兴国, 2010 ).它通过有机体内元素含量及比率关系, 将生物的生长发育、健康状况、行为方式、生态系统动态、生态环境保护等多层次、多学科问题联系起来, 现已成为生态学、环境科学和生物学领域最流行的理论框架(Sterner & Elser, 2002 ; Hillebrand et al ., 2014 ).在竹子经营与研究过程中, 虽然人们早已发现竹子开花(何奇江等, 2005 ; 丁兴萃, 2006 )、发笋大小年(费世民, 2011 )、竹笋品质形成(林海萍等, 2004 )与体内养分含量及比例有关, 竹林土壤污染主要是因为养分搭配不当和总量盈余(孟赐福等, 2009 ), 也提出了平衡施肥与养分管理技术(郭晓敏等, 2013 ), 但尚未形成一个从竹子基础生物学到生产经营的有关养分元素的统一化理论, 极大地影响了竹林生产和生态环境建设. ...
不同年龄毛竹营养器官主要养分元素分布及与土壤环境的关系
3
2010
... 其次, 不同器官的元素组成及比率存在较大差异.毛竹叶、枝、秆、蔸、根中N含量变异系数高达85.5%, 叶N含量(21.44 g·kg-1 )是秆(2.86 g·kg-1 )的6.0倍.Si含量变化更大, 根Si含量为70.78 g·kg-1 , 秆Si含量仅1.89 g·kg-1 , 相差近37倍(费世民, 2011 ).毛竹的N:P、P:Ca表现为叶>枝>秆>根, P:K则为叶>根>枝>秆(刘广路等, 2010 ).苦竹的N含量及N:P均呈现出叶>鞭>蔸>秆; K:P和Si:P则表现出鞭>蔸>叶>秆(刘力等, 2004 ).雷竹、麻竹等竹种的K、P、Ca、Mg等多重元素的含量及比例也都存在器官差异规律(邱尔发等, 2004 ; 吴家森等, 2005a ).以上说明竹子元素分布规律遵循植物元素“功能性不均衡分布” ...
... 再次, 年龄也是影响竹子化学计量特征的因子.1-6年内, 毛竹秆C含量随秆龄增长呈上升趋势, N、P含量呈下降趋势(吴家森等, 2005b ; 刘广路等, 2010 ), 故C:N、C:P随年龄增长而明显增加.不同竹叶(叶龄)差异也较大, 一年生新叶N:P、N:K为13.35、4.78, 2年老叶为21.45、5.23, 老叶N:P比值明显大于幼叶(刘广路等, 2010 ).杨清培发现毛竹幼龄鞭N:P (7.0)明显低于老龄鞭(15.0) (未发表).糙花少穗竹(Oligostachyum scabriflorum )也表现出同样的规律, 1-2年竹秆N:P为13.68-13.86, 而3、4年竹秆分别为20.18和27.88 (陈世品等, 2007 ).另外, 不同季节竹子也表现出不同的化学计量特征, 生长季N:P明显偏低(黄伯惠, 1983 ).说明竹子化学计量特征遵循“生长速率假说”. ...
... ), 故C:N、C:P随年龄增长而明显增加.不同竹叶(叶龄)差异也较大, 一年生新叶N:P、N:K为13.35、4.78, 2年老叶为21.45、5.23, 老叶N:P比值明显大于幼叶(刘广路等, 2010 ).杨清培发现毛竹幼龄鞭N:P (7.0)明显低于老龄鞭(15.0) (未发表).糙花少穗竹(Oligostachyum scabriflorum )也表现出同样的规律, 1-2年竹秆N:P为13.68-13.86, 而3、4年竹秆分别为20.18和27.88 (陈世品等, 2007 ).另外, 不同季节竹子也表现出不同的化学计量特征, 生长季N:P明显偏低(黄伯惠, 1983 ).说明竹子化学计量特征遵循“生长速率假说”. ...
毛竹种群向常绿阔叶林扩张的细根策略
1
2013
... 土壤养分不足时, 竹子可调整鞭根形态、分布格局而表现出明显的觅食行为(Li et al ., 2000 ; 刘骏等, 2013 ), 以获取养分资源, 维持体内养分平衡.施肥试验表明, 毛竹竹鞭穿越养分异质环境时, 它会避开贫养斑块, 将大量竹笋有选择地放置在富养斑块(Li et al ., 2000 ).随着土壤N含量的增加, 雷竹地上竹子数量增多、生物量增大, 而且地下竹鞭长度和鞭节长度显著降低(Yue et al ., 2005 ), 表现出明显的觅食特征.毛竹扩鞭试验表明, 林缘施肥不仅可以增加新竹数量、降低退笋率, 而且可扩大新竹扩展范围, 施肥组每年平均扩张5 m 以上(董晨玲, 2003 ).这些可间接说明, 竹林向邻近森林扩张可能是为了维持体内养分平衡而表现出来的一种觅食 行为. ...
苦竹各器官营养元素分析
1
2004
... 其次, 不同器官的元素组成及比率存在较大差异.毛竹叶、枝、秆、蔸、根中N含量变异系数高达85.5%, 叶N含量(21.44 g·kg-1 )是秆(2.86 g·kg-1 )的6.0倍.Si含量变化更大, 根Si含量为70.78 g·kg-1 , 秆Si含量仅1.89 g·kg-1 , 相差近37倍(费世民, 2011 ).毛竹的N:P、P:Ca表现为叶>枝>秆>根, P:K则为叶>根>枝>秆(刘广路等, 2010 ).苦竹的N含量及N:P均呈现出叶>鞭>蔸>秆; K:P和Si:P则表现出鞭>蔸>叶>秆(刘力等, 2004 ).雷竹、麻竹等竹种的K、P、Ca、Mg等多重元素的含量及比例也都存在器官差异规律(邱尔发等, 2004 ; 吴家森等, 2005a ).以上说明竹子元素分布规律遵循植物元素“功能性不均衡分布” ...
长期淹水对河竹鞭根养分化学计量特征的影响
2015
毛竹人工林持续立地生产力的研究
1
2001
... 竹子(竹亚科植物的总称)生长快速、材质优良, 被广泛用于建材与家居; 竹笋味道鲜美、营养丰富, 备受人们青睐(江泽慧, 2007 ; Chongtham et al ., 2011 ).仅2001年全球竹材消费就高达3000万t、竹笋消费200万t (Kleinhenz et al ., 2000 ; Kleinhenz & Midmore, 2001 ), 年贸易额高达45亿美元(Kleinhenz & Midmore, 2001 ).我国竹林面积近600万hm2 , 产值1600亿元, 并呈逐年上升的趋势(董杰和张燕, 2015 ).竹产品需求持续增长, 大大促进了竹林发展, 但不合理经营导致了部分竹林衰退(楼一平等, 1997 )、产量下降(孟赐福等, 2009 )、地力退化(楼一平, 2001 ; Guo et al ., 2014 )、环境污染(吴家森等, 2009 )等生产和生态问题.这些问题既与竹子自身的生长发育、开花结实等生物习性有关, 也与人类采收笋材、垦复施肥等经营活动相关.然而, 前人仅从竹子生物学、生态学和培育学等方面进行了零散的研究, 不利于以上问题的综合解决.因此, 竹子领域研究亟需一个统一化理论做指导. ...
竹木混交林改为毛竹纯林经营后的林分生长动态
1
1997
... 竹子(竹亚科植物的总称)生长快速、材质优良, 被广泛用于建材与家居; 竹笋味道鲜美、营养丰富, 备受人们青睐(江泽慧, 2007 ; Chongtham et al ., 2011 ).仅2001年全球竹材消费就高达3000万t、竹笋消费200万t (Kleinhenz et al ., 2000 ; Kleinhenz & Midmore, 2001 ), 年贸易额高达45亿美元(Kleinhenz & Midmore, 2001 ).我国竹林面积近600万hm2 , 产值1600亿元, 并呈逐年上升的趋势(董杰和张燕, 2015 ).竹产品需求持续增长, 大大促进了竹林发展, 但不合理经营导致了部分竹林衰退(楼一平等, 1997 )、产量下降(孟赐福等, 2009 )、地力退化(楼一平, 2001 ; Guo et al ., 2014 )、环境污染(吴家森等, 2009 )等生产和生态问题.这些问题既与竹子自身的生长发育、开花结实等生物习性有关, 也与人类采收笋材、垦复施肥等经营活动相关.然而, 前人仅从竹子生物学、生态学和培育学等方面进行了零散的研究, 不利于以上问题的综合解决.因此, 竹子领域研究亟需一个统一化理论做指导. ...
Nutrient stoichiometry in Pinguicula vulgaris: Nutrient availability, plant size, and reproductive status.
1
2005
... 另外, 生态化学计量学特别重视生物与环境的矛盾统一, 认为生物元素内平衡需求与环境供应间的适配(match)与错配(mismatch)关系是推动生命发展的根本动力(Sterner & Elser, 2002 ), 也是环境变化的重要原因之一.一方面环境元素会影响生物的内平衡和生长速率(Méndez & Karlsson, 2005 ; Rivas-Ubach et al ., 2012 ); 另一方面生物会通过吸收和释放体内元素而改变环境, 进而对生物产生影响(Sterner & Elser, 2002 ).因此, 生物与环境间连续而复杂的元素化学计量适配或错配反馈关系推动着生物与环境的协同进化(Abbas et al ., 2013 ). ...
不同施肥处理对雷竹林土壤养分平衡和竹笋产量的影响
2
2009
... 竹子(竹亚科植物的总称)生长快速、材质优良, 被广泛用于建材与家居; 竹笋味道鲜美、营养丰富, 备受人们青睐(江泽慧, 2007 ; Chongtham et al ., 2011 ).仅2001年全球竹材消费就高达3000万t、竹笋消费200万t (Kleinhenz et al ., 2000 ; Kleinhenz & Midmore, 2001 ), 年贸易额高达45亿美元(Kleinhenz & Midmore, 2001 ).我国竹林面积近600万hm2 , 产值1600亿元, 并呈逐年上升的趋势(董杰和张燕, 2015 ).竹产品需求持续增长, 大大促进了竹林发展, 但不合理经营导致了部分竹林衰退(楼一平等, 1997 )、产量下降(孟赐福等, 2009 )、地力退化(楼一平, 2001 ; Guo et al ., 2014 )、环境污染(吴家森等, 2009 )等生产和生态问题.这些问题既与竹子自身的生长发育、开花结实等生物习性有关, 也与人类采收笋材、垦复施肥等经营活动相关.然而, 前人仅从竹子生物学、生态学和培育学等方面进行了零散的研究, 不利于以上问题的综合解决.因此, 竹子领域研究亟需一个统一化理论做指导. ...
... 生态化学计量学是从元素计量的角度来探讨生命运动的内在规律的理论(曾德慧和陈广生, 2005 ; 贺金生和韩兴国, 2010 ).它通过有机体内元素含量及比率关系, 将生物的生长发育、健康状况、行为方式、生态系统动态、生态环境保护等多层次、多学科问题联系起来, 现已成为生态学、环境科学和生物学领域最流行的理论框架(Sterner & Elser, 2002 ; Hillebrand et al ., 2014 ).在竹子经营与研究过程中, 虽然人们早已发现竹子开花(何奇江等, 2005 ; 丁兴萃, 2006 )、发笋大小年(费世民, 2011 )、竹笋品质形成(林海萍等, 2004 )与体内养分含量及比例有关, 竹林土壤污染主要是因为养分搭配不当和总量盈余(孟赐福等, 2009 ), 也提出了平衡施肥与养分管理技术(郭晓敏等, 2013 ), 但尚未形成一个从竹子基础生物学到生产经营的有关养分元素的统一化理论, 极大地影响了竹林生产和生态环境建设. ...
毛竹竹腔富硒施肥对竹笋品质的影响
1
2014
... 竹材(笋)品质是市场竞争力的核心.大径级、节间长、壁厚、枝下高长的竹材价格明显要高; 外观漂亮、营养丰富的竹笋更受市场欢迎.尽管目前已开展了部分养分元素(N、P、K、Se、Si)对竹材(笋)品质形成的影响实验(孟勇等, 2014 ), 但相关的基础研究仍很滞后, 高产优质的形成规律还未被充分地揭示与认识, 可推广的高产优质培育技术尚未确立, 制约了竹林产量提高和品质的改善.因此建议开展以下研究: 1)竹笋品质形成规律及化学计量学机制研究, 尤其是Se、Zn等非必需元素、有机肥与竹笋品质形成的生理机制; 2)节长、壁厚等优质材用林定向培育的化学计量学原理与技术研究; 3)基于光、温、肥、水等多种环境因子的竹材(笋)高产、优质和资源高效利用的化学计量调控机制研究. ...
Internal and external regulation of plant organ stoichiometry.
1
2014
... 原理(Yu et al ., 2011 ; Minden & Kleyer, 2014 ). ...
Nitrogen/ phosphorus leaf stoichiometry and the scaling of plant growth.
1
2005
... 生长速率假说(growth rate hypothesis)是解析有机体维持内平衡的机制性假说.该假说认为有机体生长速率与其体内元素化学计量比率紧密联系.高生长速率往往对应高N:C、P:C以及较低的N:P.因为有机体快速生长需要大量富含P的核糖体(蛋白质的合成场所)和RNA、DNA、ATP等物质(Elser et al ., 2003 ; Reef et al ., 2010 ).生长速率假说是解析细胞生物学、生物新陈代谢、种群动态和生态系统功能的理论框架(Niklas et al ., 2005 ; Ågren, 2008 ). ...
硅对雷竹抗旱性的影响
1
2013
... 竹子体内含有30-40种大量元素和微量元素, 它们都与竹子生长发育、竹材(笋)品质等生物学特性密切相关(蒋式洪等, 2000 ; 费世民, 2011 ).C、N、P、K是竹子的基本组成元素, C是生命的骨架元素与能量元素, 毛竹(Phyllostachys edulis )叶片C含量((512.13 ± 11.13) mg·g-1 )高于世界陆生植物的平均水平((464.32 ± 32.10) mg·g-1 ).N、P、K不仅影响竹子发笋数量, 也会影响竹笋品质(郑郁善等, 1998 ; 蒋式洪等, 2000 ); Ca、Mg分别与竹子根系生长和叶绿素形成有密切关系.另外, Si尽管不是生命必需元素, 但竹子体内Si含量特别高, 近年备受关注(潘月等, 2013 ; 戎洁庆等, 2013 ; Umemura & Takenaka, 2014 ). ...
Diagnosis of plant nutrient and growth responses on fertilization with vector analysis and morphological index.
1
2014
... 养分诊断与平衡施肥是提高林业生产、维护系统稳定、选择合适管理策略的重要手段(Park et al ., 2014 ), 但当前竹林养分诊断仍停留在单一的养分测定(陈卫文等, 2004 ; 郭晓敏等, 2013 ), 很少考虑多养分、多组织的耦合效应, 失衡超量施肥强烈影响到竹林土壤理化性质, 并引起竹笋食用安全、水土环境恶化等问题.今后应以生态化学计量平衡理论为基础, 改进养分诊断方法, 根据不同竹种、不同林分、不同时期养分限制性的差异特征, 筛选养分丰缺指标, 建立多元素、多组分的养分诊断技术, 提高监测精度与预测能力; 形成生态化学计量平衡施肥技术, 根据竹子和土壤的营养特点, 筛选肥料种类、确定肥料用量与配比, 实施时空精准施肥, 提高肥料利用效率, 实现多重元素间的平衡, 竹子养分需求与环境养分供给间的平衡, 肥料投入与经济产出间的平衡, 产品生产与环境保护间的平衡. ...
Effects of high nutrient supply on the growth of seven bamboo species.
1
2014
... 首先, 不同竹种的化学计量学特征有所差异.毛竹叶片N、P、K含量分别为21.44 g·kg-1 、1.33 g·kg-1 、13.15 g·kg-1 (费世民, 2011 ), 雷竹(Phyllos- tachys praecox ‘ Prevernalis’)叶片N、P、K含量分别为8.86 g·kg-1 、1.07 g·kg-1 、6.55 g·kg-1 (吴家森等, 2005a ).毛竹叶片N:P大于雷竹, 而P:K小于雷竹.麻竹(Dendrocalamus latiflorus )叶片N、P、K含量分别为56.9 g·kg-1 、4.4 g·kg-1 、8.1 g·kg-1 (邱尔发等, 2004 ), 苦竹(Pleioblastus amarus )叶片N、P、K含量分别为24.26 g·kg-1 、1.51 g·kg-1 、11.56 g·kg-1 (蒋俊明等, 2007 ), 麻竹叶片N、P、K含量都明显高于苦竹.不同竹种对环境养分变化的化学计量学响应的程度也有差异(Piouceau et al ., 2014 ). ...
山地麻竹林生态系统养分分配格局
2
2004
... 首先, 不同竹种的化学计量学特征有所差异.毛竹叶片N、P、K含量分别为21.44 g·kg-1 、1.33 g·kg-1 、13.15 g·kg-1 (费世民, 2011 ), 雷竹(Phyllos- tachys praecox ‘ Prevernalis’)叶片N、P、K含量分别为8.86 g·kg-1 、1.07 g·kg-1 、6.55 g·kg-1 (吴家森等, 2005a ).毛竹叶片N:P大于雷竹, 而P:K小于雷竹.麻竹(Dendrocalamus latiflorus )叶片N、P、K含量分别为56.9 g·kg-1 、4.4 g·kg-1 、8.1 g·kg-1 (邱尔发等, 2004 ), 苦竹(Pleioblastus amarus )叶片N、P、K含量分别为24.26 g·kg-1 、1.51 g·kg-1 、11.56 g·kg-1 (蒋俊明等, 2007 ), 麻竹叶片N、P、K含量都明显高于苦竹.不同竹种对环境养分变化的化学计量学响应的程度也有差异(Piouceau et al ., 2014 ). ...
... 其次, 不同器官的元素组成及比率存在较大差异.毛竹叶、枝、秆、蔸、根中N含量变异系数高达85.5%, 叶N含量(21.44 g·kg-1 )是秆(2.86 g·kg-1 )的6.0倍.Si含量变化更大, 根Si含量为70.78 g·kg-1 , 秆Si含量仅1.89 g·kg-1 , 相差近37倍(费世民, 2011 ).毛竹的N:P、P:Ca表现为叶>枝>秆>根, P:K则为叶>根>枝>秆(刘广路等, 2010 ).苦竹的N含量及N:P均呈现出叶>鞭>蔸>秆; K:P和Si:P则表现出鞭>蔸>叶>秆(刘力等, 2004 ).雷竹、麻竹等竹种的K、P、Ca、Mg等多重元素的含量及比例也都存在器官差异规律(邱尔发等, 2004 ; 吴家森等, 2005a ).以上说明竹子元素分布规律遵循植物元素“功能性不均衡分布” ...
Effect of bamboo harvest on dynamics of nutrient pools, N mineralization, and microbial biomass in soil.
2
1994
... 另外, 竹子采伐、竹笋采收等人类活动也会影响竹林化学计量特征(Raghubanshi, 1994 ).然而, 目前对竹子化学计量内平衡的影响因子及作用规律研究仍停留在少数单因素实验, 养分、水分与光照等多种因子耦合交互作用有待深入研究. ...
... 生命元素通过物质循环、能量流动等形式将生态系统的各组分、多层次紧密联系起来(Carnicer et al ., 2015 ).竹林是一个人工或半人工的开放生态系统, 几乎每年都有竹材(或竹笋)等产品输出和外源养分的输入.产品输出和养分输入的失衡会改变竹子、土壤、微生物等不同组分的化学计量特征(Raghubanshi, 1994 ; Venkatesh et al ., 2005 ; Zhang et al ., 2013 ).因此, 今后应充分考虑生物与环境间的相互渗透性, 根据物质守恒与能量守恒定律, 综合考虑土壤、气候、经营活动等因素, 加强竹林生态系统的生态化学计量模型与理论研究, 构建“竹子-土壤-微生物”多组分的“生态化学计量匹配模型”, 竹子化学计量特征与群落组成结构、生态系统生产力多层次的多重养分优化计量模型, 并通过长期野外监测与微宇宙模拟实验, 找到具有竹子特色的“特征参数”, 最后形成可视化的竹林养分预测和管理系统, 指导竹林健康经营和环境综合效应评估, 实现因地制宜、以产定肥的竹林生态系统生态化学计量优化管理模式. ...
The biological control of chemical factors in the environment.
1
1958
... 生态化学计量学主要是从元素组成及比率的角度来研究生物与环境关系, 是近年生态学中飞速发展的领域.自1840年德国农业化学家Justus von Liebig发现植物生长与元素含量的关系并提出了“最小因子定律”以来, 经过生态位理论(Grubb, 1977 )、最佳取食理论(Belovsky, 1978 )、资源比理论(Tilman, 1982 )、养分利用理论(Vitousek, 1982 )、Redfield比值(Redfield, 1958 )等理论的发展, Reiners (1986)提出“Chemical Stoichiometry of the Biota”, 将化学计量学理论用于研究生态系统结构与功能, 到2002年, Sterner和Elser出版专著《Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere》, 系统地阐述了生态化学计量学的定义、基本理论及其应用范畴, 标志着生态化学计量学体系的逐渐成熟, 也为生物学统一化理论构建提供了新思路(Sterner & Elser, 2002 ; 曾德慧和陈广生, 2005 ; 贺金生和韩兴国, 2010 ).当前, 生态化学计量学已发展成从生物大分子到生态系统, 从微生物、动物到植物, 从营养学、生态学到环境科学多层次、多学科都相互关联的综合性理论, 并逐渐被应用于预测或解决区域乃至全球的生态环境问题(曾德慧和陈广生, 2005 ).其理论逻辑如下: ...
Relationships among RNA:DNA ratio, growth and elemental stoichio- metry in mangrove trees.
1
2010
... 生长速率假说(growth rate hypothesis)是解析有机体维持内平衡的机制性假说.该假说认为有机体生长速率与其体内元素化学计量比率紧密联系.高生长速率往往对应高N:C、P:C以及较低的N:P.因为有机体快速生长需要大量富含P的核糖体(蛋白质的合成场所)和RNA、DNA、ATP等物质(Elser et al ., 2003 ; Reef et al ., 2010 ).生长速率假说是解析细胞生物学、生物新陈代谢、种群动态和生态系统功能的理论框架(Niklas et al ., 2005 ; Ågren, 2008 ). ...
Global patterns of plant leaf N and P in relation to temperature and latitude.
1
2004
... 生态化学计量学认为元素组成相对稳定是一切生物的基本特征.本文发现竹子叶片多重元素比率平均为C:N:P = 380:16:1.然而, 竹种、器官、年龄和发育阶段等遗传因素, 土壤、气候、人类活动等环境因素都会直接或间接地影响竹子的生态化学计量内平衡(Reich & Oleksyn, 2004 ; Sardans et al ., 2012 ), 进而影响其新陈代谢、生长发育等生命过程.同时, 植物也会通过组织老化、凋落、转移与储藏等形式, 调整养分吸收与释放, 实现整体对多重元素(如C:N:P)比率的调节, 但竹子化学计量平衡的影响因子及作用机制尚不清楚.以后应加强多竹种(尤其是经济竹种和珍稀竹种)化学计量内平衡特征研究; 加强开花、发笋、大小年等基础生物学过程与元素化学计量的关系研究; 结合分子、同位素等先进技术, 深入开展竹子化学计量内稳性与其光合作用、养分分配、克隆整合等生理生态学研究; 开展多因子(养分与水分)对元素内平衡的交互影响及竹子的响应机制; 开展竹子化学计量特征与新陈代谢活性物质的关系研究(Rivas-Ubach et al ., 2012 ), 以提高生态化学计量学对竹子生物学现象、过程和机制的解释能力. ...
Complementary models for ecosystems.
1986
Strong relationship between elemental stoichiometry and metabolome in plants.
3
2012
... (4)体内元素平衡变化或偏离会影响有机体的新陈代谢、生长发育、健康状况和行为方式(Schatz & McCauley, 2007 ; Rivas-Ubach et al ., 2012 ); ...
... 另外, 生态化学计量学特别重视生物与环境的矛盾统一, 认为生物元素内平衡需求与环境供应间的适配(match)与错配(mismatch)关系是推动生命发展的根本动力(Sterner & Elser, 2002 ), 也是环境变化的重要原因之一.一方面环境元素会影响生物的内平衡和生长速率(Méndez & Karlsson, 2005 ; Rivas-Ubach et al ., 2012 ); 另一方面生物会通过吸收和释放体内元素而改变环境, 进而对生物产生影响(Sterner & Elser, 2002 ).因此, 生物与环境间连续而复杂的元素化学计量适配或错配反馈关系推动着生物与环境的协同进化(Abbas et al ., 2013 ). ...
... 生态化学计量学认为元素组成相对稳定是一切生物的基本特征.本文发现竹子叶片多重元素比率平均为C:N:P = 380:16:1.然而, 竹种、器官、年龄和发育阶段等遗传因素, 土壤、气候、人类活动等环境因素都会直接或间接地影响竹子的生态化学计量内平衡(Reich & Oleksyn, 2004 ; Sardans et al ., 2012 ), 进而影响其新陈代谢、生长发育等生命过程.同时, 植物也会通过组织老化、凋落、转移与储藏等形式, 调整养分吸收与释放, 实现整体对多重元素(如C:N:P)比率的调节, 但竹子化学计量平衡的影响因子及作用机制尚不清楚.以后应加强多竹种(尤其是经济竹种和珍稀竹种)化学计量内平衡特征研究; 加强开花、发笋、大小年等基础生物学过程与元素化学计量的关系研究; 结合分子、同位素等先进技术, 深入开展竹子化学计量内稳性与其光合作用、养分分配、克隆整合等生理生态学研究; 开展多因子(养分与水分)对元素内平衡的交互影响及竹子的响应机制; 开展竹子化学计量特征与新陈代谢活性物质的关系研究(Rivas-Ubach et al ., 2012 ), 以提高生态化学计量学对竹子生物学现象、过程和机制的解释能力. ...
硅肥对雷竹林生长影响研究
1
2013
... 竹子体内含有30-40种大量元素和微量元素, 它们都与竹子生长发育、竹材(笋)品质等生物学特性密切相关(蒋式洪等, 2000 ; 费世民, 2011 ).C、N、P、K是竹子的基本组成元素, C是生命的骨架元素与能量元素, 毛竹(Phyllostachys edulis )叶片C含量((512.13 ± 11.13) mg·g-1 )高于世界陆生植物的平均水平((464.32 ± 32.10) mg·g-1 ).N、P、K不仅影响竹子发笋数量, 也会影响竹笋品质(郑郁善等, 1998 ; 蒋式洪等, 2000 ); Ca、Mg分别与竹子根系生长和叶绿素形成有密切关系.另外, Si尽管不是生命必需元素, 但竹子体内Si含量特别高, 近年备受关注(潘月等, 2013 ; 戎洁庆等, 2013 ; Umemura & Takenaka, 2014 ). ...
Effects of resource heterogeneity on nitrogen translocation within clonal fragments of Sasa palmata: An isotopic (15 N) assessment.
1
2006
... 生态化学计量特征变化会影响竹子对矿质养分的选择吸收、运输与分配.庄明浩等(2013a)发现毛竹叶片C:N下降、C:P、N:P升高, 可以促进竹叶对Mg、Ca的分配, 增强竹根Na积累, 提高Fe、Ca、Mg向上选择性运输的能力.顾大形等(2011)认为四季竹在土壤N供应充足时, 叶片也会对N进行奢侈性吸收.同时, 化学计量特征还会影响竹子的养分克隆生理整合过程, 一般“富养”分株(或部位)供应“贫养”分株(或部位), 老龄竹将N、P等元素转移到低龄竹及幼嫩部位, 但不同元素的整合强度与体内化学计量状态有关(Li et al ., 1998b ; Saitoh et al ., 2006 ). ...
The C:N:P stoichiometry of organisms and ecosystems in a changing world: A review and perspectives.
1
2012
... 生态化学计量学认为元素组成相对稳定是一切生物的基本特征.本文发现竹子叶片多重元素比率平均为C:N:P = 380:16:1.然而, 竹种、器官、年龄和发育阶段等遗传因素, 土壤、气候、人类活动等环境因素都会直接或间接地影响竹子的生态化学计量内平衡(Reich & Oleksyn, 2004 ; Sardans et al ., 2012 ), 进而影响其新陈代谢、生长发育等生命过程.同时, 植物也会通过组织老化、凋落、转移与储藏等形式, 调整养分吸收与释放, 实现整体对多重元素(如C:N:P)比率的调节, 但竹子化学计量平衡的影响因子及作用机制尚不清楚.以后应加强多竹种(尤其是经济竹种和珍稀竹种)化学计量内平衡特征研究; 加强开花、发笋、大小年等基础生物学过程与元素化学计量的关系研究; 结合分子、同位素等先进技术, 深入开展竹子化学计量内稳性与其光合作用、养分分配、克隆整合等生理生态学研究; 开展多因子(养分与水分)对元素内平衡的交互影响及竹子的响应机制; 开展竹子化学计量特征与新陈代谢活性物质的关系研究(Rivas-Ubach et al ., 2012 ), 以提高生态化学计量学对竹子生物学现象、过程和机制的解释能力. ...
Foraging behavior by Daphnia in stoichiometric gradients of food quality.
1
2007
... (4)体内元素平衡变化或偏离会影响有机体的新陈代谢、生长发育、健康状况和行为方式(Schatz & McCauley, 2007 ; Rivas-Ubach et al ., 2012 ); ...
Determination of optimum nutrient element ratios in plant tissue.
1
1997
... 营养不足容易引起植物组织老化, 而过剩又引起植物毒害, 造成资源浪费与环境污染(Güsewell, 2004 ).因此, 养分诊断是保证林农业产品优质高产的重要措施, 并将体内营养元素的含量及比例作为衡量植物营养状况的重要指标(Sinclair et al ., 1997 ; Güsewell et al ., 2003 ; Tessier & Raynal, 2003 ). ...
毛竹扩张对常绿阔叶林土壤氮素矿化及有效性的影响
1
2013
... 竹子扩张会降低邻近森林植物多样性, 影响生态系统的稳定性(白尚斌等, 2013 ).初步研究认为毛竹扩张间接影响化学计量内平衡是植物多样性下降的重要机制.毛竹扩张改变了邻近森林土壤的养分状况(宋庆妮等, 2013 ), 而不同植物对土壤养分变化的响应差异较大, 其中栲(Castanopsis fargesii )、红楠(Machilus thunbergii )、交让木(Daphniphyllum macropodum )、赤杨叶(Alniphyllum fortunei )、金钱松(Pseudolarix amabilis )等内平衡调节能力较弱的植物最终退出竹木混交林, 而南方红豆杉(Taxus chinensis var. mairei )、钩锥(Castanopsis tibetana )、黄牛奶树(Symplocos laurina )等内平衡调节和可塑性能力较强的植物生长良好(杨清培, 未发表).然而, 这方面的研究才刚刚起步, 今后应借鉴入侵植物化学计量学的研究成果(González et al ., 2010 )深入开展竹阔混交林组成与稳定性的生态化学计量学机制研究. ...
施肥对毛竹叶片光合生理的影响
1
2010
... 化学计量特征变化会直接影响到竹子光合作用.当施N量为250 kg·hm-2 时, 与对照相比, 1年、3年、5年生毛竹叶片的光饱和点均达最大, 分别增加了5.2%、9.1%和7.0%, 同时光补偿点降至最低, 分别降低了49.5%、20.4%和31.8% (高培军等, 2014 ), 土壤养分会影响叶片N、P含量与比例, 从而对光合作用产生直接或间接影响(宋艳冬等, 2010 ; 顾大形等, 2011 ).顾大形等(2011)发现一定范围内增加土壤N含量, 四季竹叶片的N含量增加, N:P上升, 提高了叶绿素含量, 促进其光合作用.而当施肥量不足或过量时会降低毛竹叶片光合色素含量, 结果其光合能力受到影响.苏文会(2012)发现当毛竹叶片N:P从20.70下降至15.29时, 其光合产物淀粉含量从35.47 g·kg-1 降至16.21 g·kg-1 , 降幅达54.3%.说明竹子叶片N、P化学计量特征对其光合作用影响较大. ...
基于生长和养分积累规律的毛竹林施肥理论与实践研究
2012
Soil nutrient status after bamboo flowering and death in a seasonal tropical forest in western Thailand.
1
2007
... 竹子很少开花, 但一旦开花就造成竹子大面积的突发性死亡, 因此竹子开花一直是一个有趣而神秘的话题(Janzen, 1976 ; Takahashi et al ., 2007 ).近100年来, 有关竹子开花分别从气候、管理、环境等方面提出了不同的假说(郑郁善和洪伟, 1998 ), 其中营养计量学说是目前比较普遍认可的一个重要机制性假说. ...
Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation.
1
2003
... 营养不足容易引起植物组织老化, 而过剩又引起植物毒害, 造成资源浪费与环境污染(Güsewell, 2004 ).因此, 养分诊断是保证林农业产品优质高产的重要措施, 并将体内营养元素的含量及比例作为衡量植物营养状况的重要指标(Sinclair et al ., 1997 ; Güsewell et al ., 2003 ; Tessier & Raynal, 2003 ). ...
Biological cycle of silicon in moso bamboo (Phyllostachys pubescens) forests in central Japan.
2014
Soil properties influenced by some important edible bamboo species in the North Eastern Himalayan region, India.
1
2005
... 生命元素通过物质循环、能量流动等形式将生态系统的各组分、多层次紧密联系起来(Carnicer et al ., 2015 ).竹林是一个人工或半人工的开放生态系统, 几乎每年都有竹材(或竹笋)等产品输出和外源养分的输入.产品输出和养分输入的失衡会改变竹子、土壤、微生物等不同组分的化学计量特征(Raghubanshi, 1994 ; Venkatesh et al ., 2005 ; Zhang et al ., 2013 ).因此, 今后应充分考虑生物与环境间的相互渗透性, 根据物质守恒与能量守恒定律, 综合考虑土壤、气候、经营活动等因素, 加强竹林生态系统的生态化学计量模型与理论研究, 构建“竹子-土壤-微生物”多组分的“生态化学计量匹配模型”, 竹子化学计量特征与群落组成结构、生态系统生产力多层次的多重养分优化计量模型, 并通过长期野外监测与微宇宙模拟实验, 找到具有竹子特色的“特征参数”, 最后形成可视化的竹林养分预测和管理系统, 指导竹林健康经营和环境综合效应评估, 实现因地制宜、以产定肥的竹林生态系统生态化学计量优化管理模式. ...
Nutrient cycling and nutrient use efficiency.
1
1982
... 生态化学计量学主要是从元素组成及比率的角度来研究生物与环境关系, 是近年生态学中飞速发展的领域.自1840年德国农业化学家Justus von Liebig发现植物生长与元素含量的关系并提出了“最小因子定律”以来, 经过生态位理论(Grubb, 1977 )、最佳取食理论(Belovsky, 1978 )、资源比理论(Tilman, 1982 )、养分利用理论(Vitousek, 1982 )、Redfield比值(Redfield, 1958 )等理论的发展, Reiners (1986)提出“Chemical Stoichiometry of the Biota”, 将化学计量学理论用于研究生态系统结构与功能, 到2002年, Sterner和Elser出版专著《Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere》, 系统地阐述了生态化学计量学的定义、基本理论及其应用范畴, 标志着生态化学计量学体系的逐渐成熟, 也为生物学统一化理论构建提供了新思路(Sterner & Elser, 2002 ; 曾德慧和陈广生, 2005 ; 贺金生和韩兴国, 2010 ).当前, 生态化学计量学已发展成从生物大分子到生态系统, 从微生物、动物到植物, 从营养学、生态学到环境科学多层次、多学科都相互关联的综合性理论, 并逐渐被应用于预测或解决区域乃至全球的生态环境问题(曾德慧和陈广生, 2005 ).其理论逻辑如下: ...
雷竹集约栽培对周边河流水质的影响
1
2009
... 竹子(竹亚科植物的总称)生长快速、材质优良, 被广泛用于建材与家居; 竹笋味道鲜美、营养丰富, 备受人们青睐(江泽慧, 2007 ; Chongtham et al ., 2011 ).仅2001年全球竹材消费就高达3000万t、竹笋消费200万t (Kleinhenz et al ., 2000 ; Kleinhenz & Midmore, 2001 ), 年贸易额高达45亿美元(Kleinhenz & Midmore, 2001 ).我国竹林面积近600万hm2 , 产值1600亿元, 并呈逐年上升的趋势(董杰和张燕, 2015 ).竹产品需求持续增长, 大大促进了竹林发展, 但不合理经营导致了部分竹林衰退(楼一平等, 1997 )、产量下降(孟赐福等, 2009 )、地力退化(楼一平, 2001 ; Guo et al ., 2014 )、环境污染(吴家森等, 2009 )等生产和生态问题.这些问题既与竹子自身的生长发育、开花结实等生物习性有关, 也与人类采收笋材、垦复施肥等经营活动相关.然而, 前人仅从竹子生物学、生态学和培育学等方面进行了零散的研究, 不利于以上问题的综合解决.因此, 竹子领域研究亟需一个统一化理论做指导. ...
b). 不同年份毛竹营养元素的空间分布及与土壤养分的关系
2005
The mechanism for exclusion of Pinus massoniana during the succession in subtropical forest ecosystems: Light competition or stoichiometric homoeostasis?
1
2015
... (7)群体中有机体的此消彼长会影响到种群动态、群落演替和生态系统的发展及其生产、生态功能的发挥(Sterner & Elser, 2002 ; Güsewell, 2004 ; Yan et al ., 2015 ). ...
Stoichiometric homeostasis of vascular plants in the Inner Mongolia grassland.
1
2011
... 原理(Yu et al ., 2011 ; Minden & Kleyer, 2014 ). ...
Morphological plasticity of clonal plant Phyllostachys praecox f. prevernalis (Poaceae) in response to nitrogen availability.
1
2005
... 土壤养分不足时, 竹子可调整鞭根形态、分布格局而表现出明显的觅食行为(Li et al ., 2000 ; 刘骏等, 2013 ), 以获取养分资源, 维持体内养分平衡.施肥试验表明, 毛竹竹鞭穿越养分异质环境时, 它会避开贫养斑块, 将大量竹笋有选择地放置在富养斑块(Li et al ., 2000 ).随着土壤N含量的增加, 雷竹地上竹子数量增多、生物量增大, 而且地下竹鞭长度和鞭节长度显著降低(Yue et al ., 2005 ), 表现出明显的觅食特征.毛竹扩鞭试验表明, 林缘施肥不仅可以增加新竹数量、降低退笋率, 而且可扩大新竹扩展范围, 施肥组每年平均扩张5 m 以上(董晨玲, 2003 ).这些可间接说明, 竹林向邻近森林扩张可能是为了维持体内养分平衡而表现出来的一种觅食 行为. ...
The application of ecological stoichiometry to plant-microbial- soil organic matter transformations.
1
2015
... 植物养分主要来源于土壤, 土壤养分也受到植物的反作用.因此, 加强竹子与土壤养分元素的化学计量适配与错配互作研究, 对竹林生产、环境保护至关重要.首先, 土壤环境决定着养分形态、数量及其有效性, 从而影响到竹子的生长发育(Fan et al ., 2015 ; Zechmeister-Boltenstern et al ., 2015 ).建议开展土壤环境因子与土壤、竹子养分元素化学计量特征动态关系的长期监测与研究.其次, 凋落物分解和根际效应是影响土壤养分的一个重要过程, 尤其是根际过程更应受到关注(Bell et al ., 2014 , 2015 ; Carrillo et al ., 2014 ), 所以应采用稳定同位素技术、分子生态技术, 聚焦于竹子根际过程及养分活化机制(如根系吸收与分泌、根际酸碱度变化、根际微生物活动) , 实现“竹子-根际-土壤”三者元素关系的量化分析. ...
生态化学计量学: 复杂生命系统奥秘的探索. 植物生态学报,
4
2005
... 生态化学计量学是从元素计量的角度来探讨生命运动的内在规律的理论(曾德慧和陈广生, 2005 ; 贺金生和韩兴国, 2010 ).它通过有机体内元素含量及比率关系, 将生物的生长发育、健康状况、行为方式、生态系统动态、生态环境保护等多层次、多学科问题联系起来, 现已成为生态学、环境科学和生物学领域最流行的理论框架(Sterner & Elser, 2002 ; Hillebrand et al ., 2014 ).在竹子经营与研究过程中, 虽然人们早已发现竹子开花(何奇江等, 2005 ; 丁兴萃, 2006 )、发笋大小年(费世民, 2011 )、竹笋品质形成(林海萍等, 2004 )与体内养分含量及比例有关, 竹林土壤污染主要是因为养分搭配不当和总量盈余(孟赐福等, 2009 ), 也提出了平衡施肥与养分管理技术(郭晓敏等, 2013 ), 但尚未形成一个从竹子基础生物学到生产经营的有关养分元素的统一化理论, 极大地影响了竹林生产和生态环境建设. ...
... 生态化学计量学主要是从元素组成及比率的角度来研究生物与环境关系, 是近年生态学中飞速发展的领域.自1840年德国农业化学家Justus von Liebig发现植物生长与元素含量的关系并提出了“最小因子定律”以来, 经过生态位理论(Grubb, 1977 )、最佳取食理论(Belovsky, 1978 )、资源比理论(Tilman, 1982 )、养分利用理论(Vitousek, 1982 )、Redfield比值(Redfield, 1958 )等理论的发展, Reiners (1986)提出“Chemical Stoichiometry of the Biota”, 将化学计量学理论用于研究生态系统结构与功能, 到2002年, Sterner和Elser出版专著《Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere》, 系统地阐述了生态化学计量学的定义、基本理论及其应用范畴, 标志着生态化学计量学体系的逐渐成熟, 也为生物学统一化理论构建提供了新思路(Sterner & Elser, 2002 ; 曾德慧和陈广生, 2005 ; 贺金生和韩兴国, 2010 ).当前, 生态化学计量学已发展成从生物大分子到生态系统, 从微生物、动物到植物, 从营养学、生态学到环境科学多层次、多学科都相互关联的综合性理论, 并逐渐被应用于预测或解决区域乃至全球的生态环境问题(曾德慧和陈广生, 2005 ).其理论逻辑如下: ...
... ).当前, 生态化学计量学已发展成从生物大分子到生态系统, 从微生物、动物到植物, 从营养学、生态学到环境科学多层次、多学科都相互关联的综合性理论, 并逐渐被应用于预测或解决区域乃至全球的生态环境问题(曾德慧和陈广生, 2005 ).其理论逻辑如下: ...
... 生态化学计量学不仅是探讨生物新陈代谢、生长发育、种群变化、群落演替的重要理论, 而且还是林业生产的直接指南(曾德慧和陈广生, 2005 ).前期的竹林养分诊断、平衡施肥、优质高产竹林培育等都不自觉地运用了生态化学计量学的相关原理. ...
神农箭竹开花前后植物体内N、P、K元素的动态
2
2007
... 研究表明, 体内元素含量状况会影响竹子花芽分化与成花过程.如P、Ca、Mg等元素含量增加会诱导花芽分化; N、K等元素含量增加会促进营养生长, 而延迟竹子开花(何奇江等, 2005 ; 詹爱军和李兆华, 2007 ). ...
... 同时, 元素比例变化也是竹子开花的另一个重要原因(何奇江等, 2005 ; 丁兴萃, 2006 ).丁兴萃(2006)发现, 早竹(Phyllostachys violascens )开花体内C:N明显上升, 符合C:N花芽分化机理(Corbesier et al ., 2002 ); 何奇江等(2005)也发现, 开花雷竹叶、秆和鞭的P:K分别是未开花竹的1.97倍、2.80倍和2.24倍.另外, 开花往往导致养分流失、比率失衡, 促使竹子死亡(何奇江等, 2005 ; 丁兴萃, 2006 ; 詹爱军和李兆华, 2007 ). ...
Converting paddy fields to Lei Bamboo (Phyllostachys praecox) stands affected soil nutrient concentrations, labile organic carbon pools, and organic carbon chemical compositions.
1
2013
... 生命元素通过物质循环、能量流动等形式将生态系统的各组分、多层次紧密联系起来(Carnicer et al ., 2015 ).竹林是一个人工或半人工的开放生态系统, 几乎每年都有竹材(或竹笋)等产品输出和外源养分的输入.产品输出和养分输入的失衡会改变竹子、土壤、微生物等不同组分的化学计量特征(Raghubanshi, 1994 ; Venkatesh et al ., 2005 ; Zhang et al ., 2013 ).因此, 今后应充分考虑生物与环境间的相互渗透性, 根据物质守恒与能量守恒定律, 综合考虑土壤、气候、经营活动等因素, 加强竹林生态系统的生态化学计量模型与理论研究, 构建“竹子-土壤-微生物”多组分的“生态化学计量匹配模型”, 竹子化学计量特征与群落组成结构、生态系统生产力多层次的多重养分优化计量模型, 并通过长期野外监测与微宇宙模拟实验, 找到具有竹子特色的“特征参数”, 最后形成可视化的竹林养分预测和管理系统, 指导竹林健康经营和环境综合效应评估, 实现因地制宜、以产定肥的竹林生态系统生态化学计量优化管理模式. ...
竹林生长及竹叶养分和土壤肥力相关研究. 林业科学,
5
1998
... 竹子体内含有30-40种大量元素和微量元素, 它们都与竹子生长发育、竹材(笋)品质等生物学特性密切相关(蒋式洪等, 2000 ; 费世民, 2011 ).C、N、P、K是竹子的基本组成元素, C是生命的骨架元素与能量元素, 毛竹(Phyllostachys edulis )叶片C含量((512.13 ± 11.13) mg·g-1 )高于世界陆生植物的平均水平((464.32 ± 32.10) mg·g-1 ).N、P、K不仅影响竹子发笋数量, 也会影响竹笋品质(郑郁善等, 1998 ; 蒋式洪等, 2000 ); Ca、Mg分别与竹子根系生长和叶绿素形成有密切关系.另外, Si尽管不是生命必需元素, 但竹子体内Si含量特别高, 近年备受关注(潘月等, 2013 ; 戎洁庆等, 2013 ; Umemura & Takenaka, 2014 ). ...
... 除遗传因素外, 土壤、气候、人类活动等环境因素也会直接或间接影响竹子化学计量特征.顾大形等(2011)发现, 土壤N、P养分供应水平及土壤N:P会影响四季竹叶片N:P值, 但土壤对竹子养分计量特征的影响较复杂(陈志阳等, 2009 ).除养分含量外, 土壤有机质含量、pH值、土壤微生物等都会影响竹子体内养分含量与比率(郑郁善等, 1998 ), 所以竹林的地下-地上化学计量关系有待深入研究. ...
... 竹子很少开花, 但一旦开花就造成竹子大面积的突发性死亡, 因此竹子开花一直是一个有趣而神秘的话题(Janzen, 1976 ; Takahashi et al ., 2007 ).近100年来, 有关竹子开花分别从气候、管理、环境等方面提出了不同的假说(郑郁善和洪伟, 1998 ), 其中营养计量学说是目前比较普遍认可的一个重要机制性假说. ...
... 另外, 并不是所有的竹笋都能长成竹子, 许多竹笋出土前就会死亡, 或虽能出土却死在成竹之前, 这叫退笋.大量研究认为退笋率高低与养分供应水平紧密相关, 增加养分供给可以降低退笋率、提高成竹率(傅懋毅等, 1988 ; Li et al ., 1998b ; 郑郁善等, 1998 ; Li et al ., 2000 ). ...
... 竹子作为典型的克隆植物, 单株间个体大小和发育阶段不同, 其元素化学计量调节能力存在差异.当林地养分(或光照)资源不足或失衡时, 老竹个体C:N、C:P明显增加而逐渐死亡, 新竹数量减少, 竹林衰老退化.如果能及时地进行养分补充或疏伐, 会大大改善林分健康状况(郑郁善和洪伟, 1998 ). ...
毛竹林的碳密度和碳贮量及其空间分布
1
2004
... 竹子具有较强的维持体内元素相对平衡的能力.郭子武等(2011)发现施肥对红哺鸡竹(Phyllostachys iridescens )叶片C、N、P含量及比值影响不大.其叶片C:N、C:P可保持在18.71-35.02、304.41-458.52, N:P变幅更小, 仅为15.28-17.12.郭宝华等(2014)野外调查发现, 尽管土壤N:P差异明显(4.50-13.69), 但毛竹叶片N:P变化不大(12.60- 13.69).顾大形等(2011)通过盆栽实验发现, 土壤N或P增加2-3倍, 四季竹(Oligostachyum lubricum )仍可保持叶片N:P基本不变, 其内稳性指数H (N:P) = 2.85>1.庄明浩等(2013b)发现当CO2 浓度由360 μmol·mol-1 增至500 μmol·mol-1 时, 毛竹和四季竹叶片养分C、N、P含量及其比例都没有明显变化, 说明竹子在变化的土壤或大气环境中, 都能保持体内元素的相对平衡, 符合生态化学计量学的“内稳性假说”.综合多种竹子的研究结果(周国模和姜培坤, 2004 ; 费世民, 2011 ; 庄明浩等, 2013b ), 发现竹子叶片平均化学计量比值为C:N:P = 380:16:1, 其中N:P高于自然界陆生植物平均值(10:1) (Knecht & Göransson, 2004 ), 也高于国内一般草本植物(13.5:1) (Han et al ., 2005 ), 而与阔叶植物(15.1:1.0)和常绿木本植物的平均值(15.2:1.0)相近(Han et al ., 2005 ), 说明竹子与常绿阔叶木本植物具有相似的“植物-环境”化学计量学响应特征. ...
CO2 浓度升高和模拟氮沉降对青川箭竹叶营养质量的影响
2012
a). CO2 浓度升高对毛竹器官矿质离子吸收、运输和分配的影响
2013
b). CO2 浓度升高对毛竹和四季竹叶片主要养分化学计量特征的影响
2013
 , 宋庆妮, 郭春兰, 方向民, 陈昕, 黄兰, 陈伏生
, 宋庆妮, 郭春兰, 方向民, 陈昕, 黄兰, 陈伏生


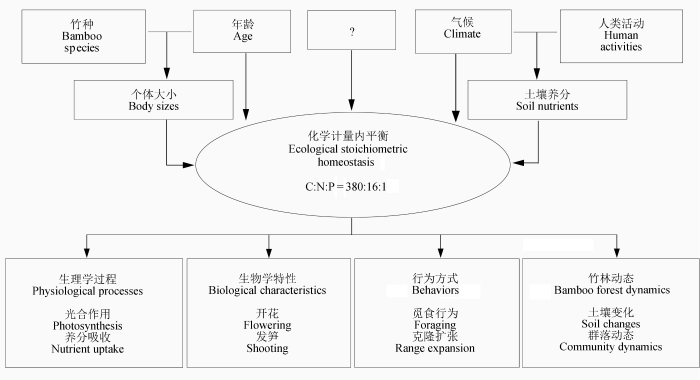
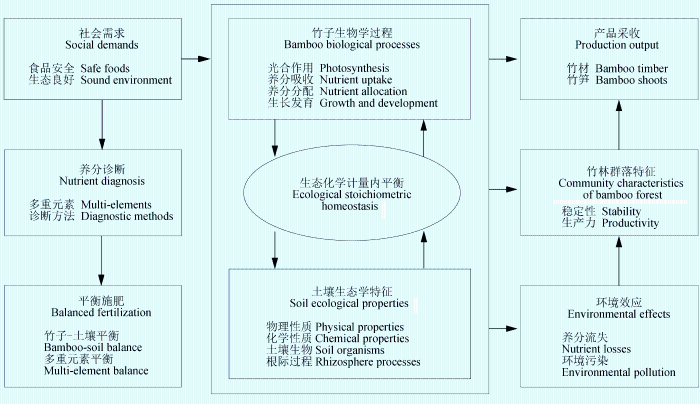
{kind=link}
{kind=link}
{kind=link}
{kind=link}