Effects of nitrogen addition on leaf traits of common species in natural Pinus tabuliformis forests in Taiyue Mountain, Shanxi Province, China
XIAO Di1, WANG Xiao-Jie1, ZHANG Kai1, HE Nian-Peng2, HOU Ji-Hua1, *,
1Key Laboratory for Forest Resources & Ecosystem Processes of Beijing Forestry University, Beijing 100083, China and 2Key Laboratory of Ecosystem Network Observation and Modeling, Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China
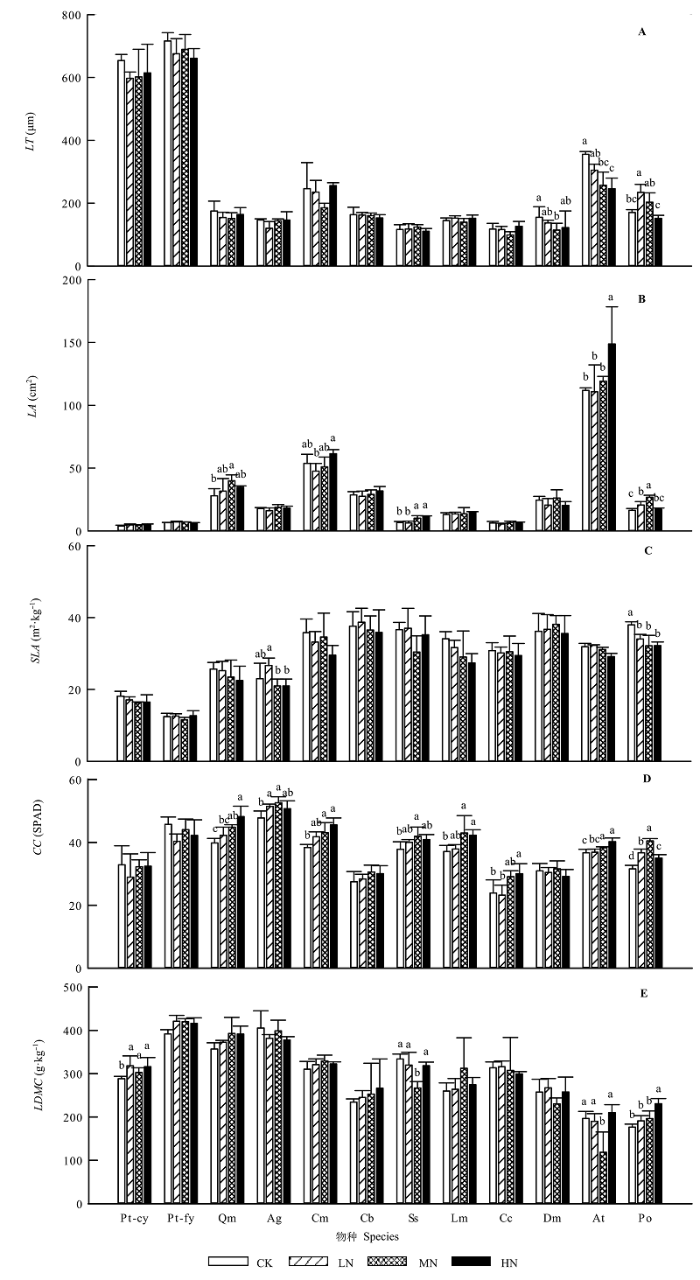
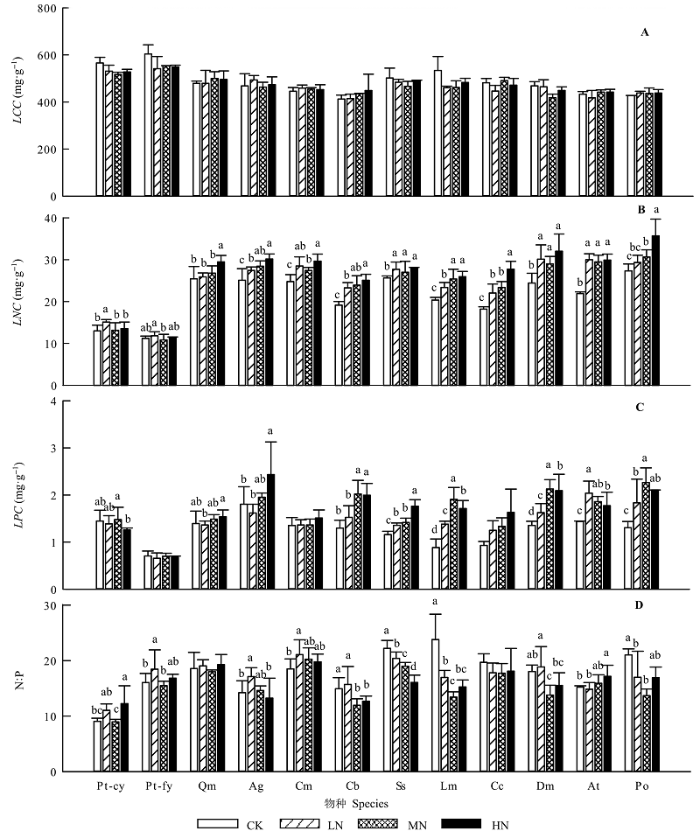
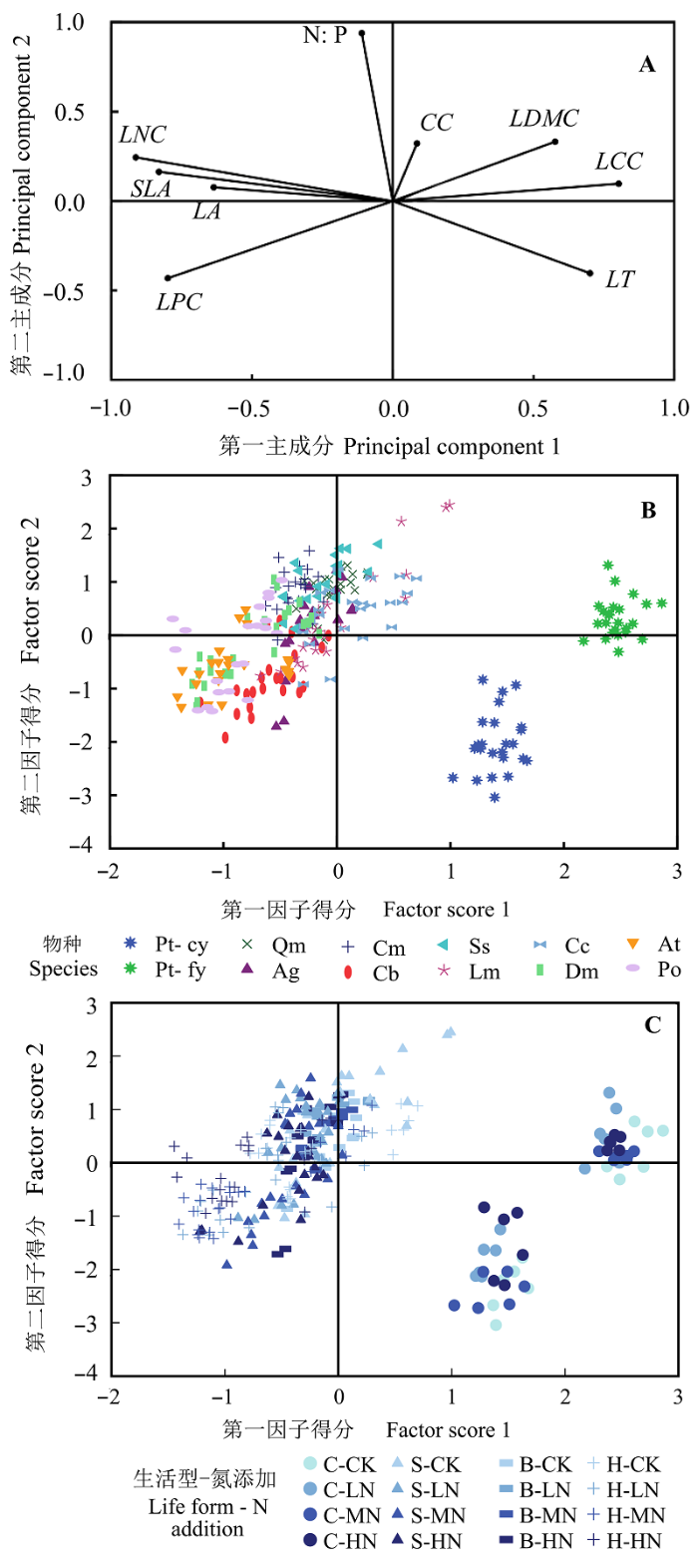
Aims Our objectives were to explore the effects of simulated nitrogen deposition on leaf traits and ecological stoichiometry characteristics of common species in natural Pinus tabuliformis forests. Methods We conducted the experiment of nitrogen (N) addition from 2009 to 2013 in the natural Pinus tabuliformis forests in Taiyue Mountain, Shanxi, China. The levels of N addition were 0 (control), 50 (low-N), 100 (medium-N) and 150 (high-N) kg·hm-2·a-1, respectively. Eleven common plant species in 12 20 m × 20 m plots were selected, including Pinus tabuliformis, Quercus mongolica, Acer ginnala, Corylus mandshurica, Cornus bretschneideri, Spiraea salicifolia, Lonicera maackii, Carex callitrichos, Diarrhena mandshurica, Anemone tomentosa, and Polygonatum odoratum. Nine leaf traits were measured, including leaf thickness (LT), specific leaf area (SLA), leaf dry matter content (LDMC), leaf nitrogen content (LNC), leaf phosphorus content (LPC), and other four. Important findings We found that: 1) LT and SLA of Polygonatum odoratum significantly differed among four levels of N addition. Leaf area (LA) and LDMC of several species, such as Spiraea salicifolia, had significant difference among the N addition concentration. LNC of all species, chlorophyll content (CC) and LPC of most species increased significantly with the addition of N. Leaf N:P of 9 species varied significantly, and leaves with different types and ages showed different responses to N addition. 2) Leaf traits were significantly correlated with each other. For instance, SLA was significantly positively correlated with LNC and LPC. In contrast LT was negatively connected with LNC and LPC. In addition, the degree of correlation changed with the level of N addition. 3) The pattern of species distribution in leaf trait space was consistent with the prediction from the theory of Leaf Economic Spectrum (LES). N addition drove species moving along axis 1 in the trait space, and propelled them towards different directions along axis 2, which indicated that these species tended to take the “fast investment-return” strategy. These results suggested that with the change of environmental conditions, plants changed their survival strategy and adjusted resource allocation to maintain the stability of communities. This is the inherent characteristic of plants, thus the formation of LES did not depend on the environment change.
XIAODi, WANGXiao-Jie, ZHANGKai, HENian-Peng, HOUJi-Hua. Effects of nitrogen addition on leaf traits of common species in natural Pinus tabuliformis forests in Taiyue Mountain, Shanxi Province, China. Chinese Journal of Plant Ecology, 2016, 40(7): 686-701 https://doi.org/10.17521/cjpe.2015.1043
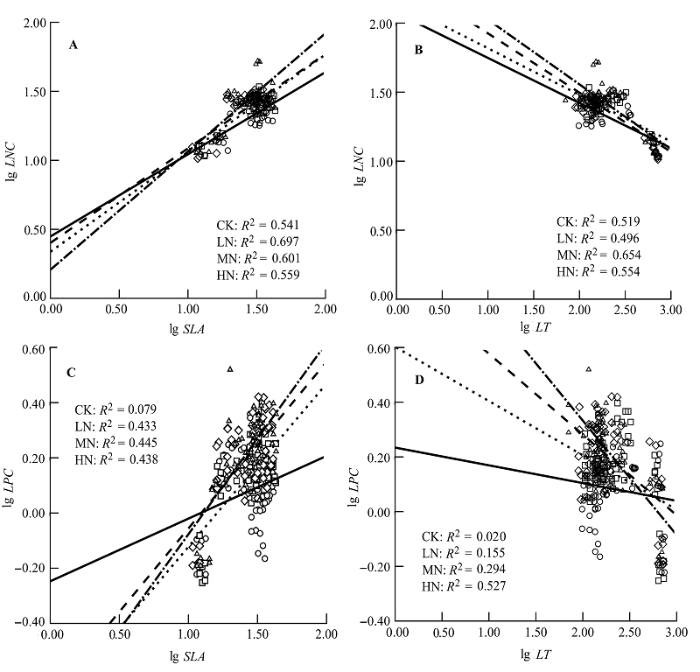
对于与构成叶经济谱相关的关联性状LNC vs. SLA、LPC vs. SLA、LNC vs. LT和LPC vs. LT进行分析发现, 氮添加强度增大时, LNC、LPC vs. SLA、LT的拟合线斜率均发生了明显变化, 且LPC vs. SLA、LT的斜率变化幅度大于LNC vs. SLA、LT的变化幅度(图3)。此外, 随着氮添加浓度的增加, LNC:SLA、LPC:SLA和LNC:LT、LPC:LT的值均显著增大(表4), 说明这4组相关关系受氮添加影响显著。
Fig. 3 Effects of nitrogen addition on leaf nitrogen content (LNC) vs. specific leaf area (SLA) (A), LNC vs. leaf thickness (LT) (B), leaf phosphorus content (LPC) vs. SLA (C) and LPC vs. LT (D). CK, 0 kg·hm-2·a-1 nitrogen; LN, 50 kg·hm-2·a-1 nitrogen; MN, 100 kg·hm-2·a-1 nitrogen; HN, 150 kg·hm-2·a-1 nitrogen.
Table 4
表4
表4 氮添加处理对叶氮含量、叶磷含量与比叶面积、叶厚度比值影响的单因素方差分析(平均值±标准误差, n = 36)
Table 4 Results of one-way ANOVA for the effect of nitrogen addition on the ratio of leaf nitrogen content (LNC) over specific leaf area (SLA) and leaf thickness (LT), and leaf phosphorus content (LPC) over specific leaf area (SLA) and leaf thickness (LT) (mean ± SE, n = 36)
Table 5 Loadings of leaf traits in principal components analyses
载荷 Loading
叶厚度 Leaf thickness
叶面积 Leaf area
比叶面积 Specific leaf area
叶绿素含量 Chlorophyll content
叶干物质含量 Leaf dry matter content
叶碳含量 Leaf carbon content
叶氮含量 Leaf nitrogen content
叶磷含量 Leaf phosphorus content
氮磷比 N-P ratio
氮添加处理 N addition treatment
主成分1 Principal component 1
0.70
-0.64
-0.83
0.09
0.58
0.80
-0.91
-0.80
-0.11
-0.20
主成分2 Principal component 2
-0.40
0.08
0.16
0.32
0.33
0.10
0.24
-0.43
0.94
-0.57
The total explanation of two principal component is 67.66%, the explanation of principal component 1 and component 2 are 52.04% and 15.62%, respectively.两个主成分总的解释力为67.66%, 其中主成分1和主成分2解释力分别为52.04%和15.62%。
Effects of N, P Addition on N:P Stoichiometry of Different Functional Groups in Potentilla fruticosa Community in a Sub-alpine Meadow. Master degree dissertation, Lanzhou University, Lanzhou
Leaf trait relationships of native and invasive plants: Community- and global-scale comparisons
.New Phytologist, 176, 635-643.
[36]
LiDJ, MoJM, FangYT, LiZA (2005).
Effects of simulated nitrogen deposition on biomass production and allocation in Schima superba and Cryptocarya concinna seedlings in subtropical China
.Acta Phytoecologica Sinica, 29, 543-549. (in Chinese with English abstract)[李德军, 莫江明, 方运霆, 李志安 (2005).
Variations in leaf functional traits of six species along a nitrogen addition gradient in Leymus chinensis steppe in Inner Mongolia. Journal of Plant Ecology (Chinese Version)
, 32, 611-621. (in Chinese with English abstract)[万宏伟, 杨阳, 白世勤, 徐云虎, 白永飞 (2008).
羊草草原群落6种植物叶片功能特性对氮素添加的响应
. 植物生态学报, 32, 611-621.]
[67]
WangCS, WangSP (2015).
A review of research on responses of leaf traits to climate change
.Chinese Journal of Plant Ecology, 39, 206-216. (in Chinese with English abstract)[王常顺, 汪诗平 (2015).
Nitrogen saturation in temperate forest ecosystems: Hypothesis revisited
1
1998
... 以欧洲氮饱和实验NITREX项目(Wright & Rasmussen, 1998)和北美哈佛森林(Harvard Forest)等(Aber et al., 1998)研究设计为参照, 在未被破坏的天然油松林(112.02°-112.25° E, 36.52°-36.72° N, 平均海拔1500 m)中建立12个20 m × 20 m的固定试验样地, 设置对照(CK, 0 kg·hm-2·a-1)、低氮(LN, 50 kg·hm-2·a-1)、中氮(MN, 100 kg·hm-2·a-1)和高氮(HN, 150 kg·hm-2·a-1) 4个处理, 3次重复.样地林分特征及0-20 cm土壤理化性质详见表1.从2009年5月起, 用尿素(CO(NH2)2)作为氮源, 将年施用量平均分成12等份, 每月下旬分别将所需的尿素溶解于2 L水中, 用背式喷雾器在对应处理的20 m × 20 m样地内均匀喷洒(对照组喷施等量的水), 其他水源来自自然降水. ...
Nitrogen saturation in northern forest ecosystems
1
1989
... 自工业革命以来, 大气氮沉降呈现急剧增加的趋势(Holland et al., 1999; Zheng et al., 2002; Adams et al., 2004; Suding et al., 2005), 这直接影响到人类和生态系统的健康、温室气体的平衡及生物的多样性, 引起了科学家们的广泛关注与担忧(Vitousek et al., 1997; Richter et al., 2005; Clark & Tilman, 2008; Liu et al., 2013).研究表明, 氮素作为陆地生态系统中最主要的限制元素(贺金生和韩兴国, 2010), 对森林植物有重要影响(李德军等, 2005).在其他养分成为限制因子之前, 氮的增加可以提高植物的净初级生产力(Aber et al., 1989; Tamm, 1990), 但长期、连续、过量的氮输入会降低植物的净光合速率和净初级生产力, 引起森林生产力的下降和森林生态系统功能的退化(Ulrich, 1995; Fenn et al., 1996; Magill et al., 2004; 方运霆等, 2005), 这被认为是森林衰退的主要原因(Nihlgard, 1985; Bobbink et al., 1998; 张维娜和廖周瑜, 2009).因此, 研究大气氮沉降对森林植物的影响具有十分重要的意义. ...
Soil functional responses to excess nitrogen inputs at global scales
1
2004
... 自工业革命以来, 大气氮沉降呈现急剧增加的趋势(Holland et al., 1999; Zheng et al., 2002; Adams et al., 2004; Suding et al., 2005), 这直接影响到人类和生态系统的健康、温室气体的平衡及生物的多样性, 引起了科学家们的广泛关注与担忧(Vitousek et al., 1997; Richter et al., 2005; Clark & Tilman, 2008; Liu et al., 2013).研究表明, 氮素作为陆地生态系统中最主要的限制元素(贺金生和韩兴国, 2010), 对森林植物有重要影响(李德军等, 2005).在其他养分成为限制因子之前, 氮的增加可以提高植物的净初级生产力(Aber et al., 1989; Tamm, 1990), 但长期、连续、过量的氮输入会降低植物的净光合速率和净初级生产力, 引起森林生产力的下降和森林生态系统功能的退化(Ulrich, 1995; Fenn et al., 1996; Magill et al., 2004; 方运霆等, 2005), 这被认为是森林衰退的主要原因(Nihlgard, 1985; Bobbink et al., 1998; 张维娜和廖周瑜, 2009).因此, 研究大气氮沉降对森林植物的影响具有十分重要的意义. ...
The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns
2
2000
... 一般认为, 氮添加会导致SLA的增大(Rose et al., 2013), 因为氮添加促进了植物的生长而提高了植物的光合效率(Ordoñez et al., 2009).万宏伟等(2008)和黄菊莹等(2009)研究发现, 羊草(Leymus chinensis)等几种草本植物的SLA因氮添加而显著增大; Bassin等(2009)的氮添加实验也表明, 虽然大多数植物叶片的比叶重(LMA, SLA的倒数)无显著变化, 但两种植物叶片的LMA显著降低(即SLA显著增大).然而, 并非所有研究结果都是如此, 赵新风等(2014)发现, 钠猪毛菜(Salsola nitraria)受氮添加影响, SLA呈下降趋势, LDMC呈增大趋势, 与上述结果相反.研究表明, 较小的SLA和较高的LDMC使得植物具有更好的资源获取能力、碳氮固持效率及水分利用效率, 在逆境中更具优势(Wilson et al., 1999; Aerts & Chapin, 2000; Zhang et al., 2007).因此, 这两项指标常被用来指示植物对环境变化的响应策略, SLA的减小和LDMC的增大往往意味着环境条件的恶化, 因为植物开始趋向于保守的生长策略(Rose et al., 2013).本研究结果表明, 大部分植物的SLA和LDMC并没有表现出显著差异, 只有茶条枫和玉竹的SLA呈显著减小趋势, 油松当年生叶和玉竹叶片的LDMC显著增大, 与赵新风等(2014)的结果相一致.这说明不同植物对氮添加的响应不同.并且, 氮添加对油松、茶条枫和玉竹这3种植物的生长是不利的. ...
Effects of combined ozone and nitrogen deposition on the in situ properties of eleven key plant species of a subalpine pasture
2009
The evolutionary basis of leaf senescence: Method to the madness?
1
1998
... 黄海侠等(2013)发现, 同种植物叶片的物质分配策略在异龄叶间存在显著差异: 当年生叶的SLA显著大于往年生叶、LDMC比往年生叶小, 且当年生叶性状的变异系数往往高于往年生叶.本研究也从油松当年生叶和往年生叶的性状比较中, 发现了类似的结果.这是因为叶片在生长初期往往需要较多的资源来优化同化器官、最大程度地获取光资源(Sultan, 2000); 但在老化过程中, 内部养分元素(如氮和磷)发生迁移且光合速率降低(Bleecker, 1998; Ono et al., 2001), 结构充分形成并固化, 对环境变化的响应就变得相对迟缓(Oguchi et al., 2003); 老叶为忍受多年环境变化, 在机械和防御构造上的额外投资较多(Chabot & David, 1982).此外, 新叶对环境变化敏感, 轻微的环境扰动就能诱导其性状发生改变, 表现出较强的可塑性(Wyka et al., 2000; Frak et al., 2001).因此, 叶片的物质投资和分配格局是随叶龄变化的, 这是植物适应环境的一个重要途径(Silvertown et al., 1997). ...
Assessing the causes and scales of the leaf economics spectrum using venation networks in Populus tremuloides
2013
The effects of air-borne nitrogen pollutants on species diversity in natural and semi-natural European vegetation
1
1998
... 自工业革命以来, 大气氮沉降呈现急剧增加的趋势(Holland et al., 1999; Zheng et al., 2002; Adams et al., 2004; Suding et al., 2005), 这直接影响到人类和生态系统的健康、温室气体的平衡及生物的多样性, 引起了科学家们的广泛关注与担忧(Vitousek et al., 1997; Richter et al., 2005; Clark & Tilman, 2008; Liu et al., 2013).研究表明, 氮素作为陆地生态系统中最主要的限制元素(贺金生和韩兴国, 2010), 对森林植物有重要影响(李德军等, 2005).在其他养分成为限制因子之前, 氮的增加可以提高植物的净初级生产力(Aber et al., 1989; Tamm, 1990), 但长期、连续、过量的氮输入会降低植物的净光合速率和净初级生产力, 引起森林生产力的下降和森林生态系统功能的退化(Ulrich, 1995; Fenn et al., 1996; Magill et al., 2004; 方运霆等, 2005), 这被认为是森林衰退的主要原因(Nihlgard, 1985; Bobbink et al., 1998; 张维娜和廖周瑜, 2009).因此, 研究大气氮沉降对森林植物的影响具有十分重要的意义. ...
The ecology of leaf life spans
1
1982
... 黄海侠等(2013)发现, 同种植物叶片的物质分配策略在异龄叶间存在显著差异: 当年生叶的SLA显著大于往年生叶、LDMC比往年生叶小, 且当年生叶性状的变异系数往往高于往年生叶.本研究也从油松当年生叶和往年生叶的性状比较中, 发现了类似的结果.这是因为叶片在生长初期往往需要较多的资源来优化同化器官、最大程度地获取光资源(Sultan, 2000); 但在老化过程中, 内部养分元素(如氮和磷)发生迁移且光合速率降低(Bleecker, 1998; Ono et al., 2001), 结构充分形成并固化, 对环境变化的响应就变得相对迟缓(Oguchi et al., 2003); 老叶为忍受多年环境变化, 在机械和防御构造上的额外投资较多(Chabot & David, 1982).此外, 新叶对环境变化敏感, 轻微的环境扰动就能诱导其性状发生改变, 表现出较强的可塑性(Wyka et al., 2000; Frak et al., 2001).因此, 叶片的物质投资和分配格局是随叶龄变化的, 这是植物适应环境的一个重要途径(Silvertown et al., 1997). ...
Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands
1
2008
... 自工业革命以来, 大气氮沉降呈现急剧增加的趋势(Holland et al., 1999; Zheng et al., 2002; Adams et al., 2004; Suding et al., 2005), 这直接影响到人类和生态系统的健康、温室气体的平衡及生物的多样性, 引起了科学家们的广泛关注与担忧(Vitousek et al., 1997; Richter et al., 2005; Clark & Tilman, 2008; Liu et al., 2013).研究表明, 氮素作为陆地生态系统中最主要的限制元素(贺金生和韩兴国, 2010), 对森林植物有重要影响(李德军等, 2005).在其他养分成为限制因子之前, 氮的增加可以提高植物的净初级生产力(Aber et al., 1989; Tamm, 1990), 但长期、连续、过量的氮输入会降低植物的净光合速率和净初级生产力, 引起森林生产力的下降和森林生态系统功能的退化(Ulrich, 1995; Fenn et al., 1996; Magill et al., 2004; 方运霆等, 2005), 这被认为是森林衰退的主要原因(Nihlgard, 1985; Bobbink et al., 1998; 张维娜和廖周瑜, 2009).因此, 研究大气氮沉降对森林植物的影响具有十分重要的意义. ...
Functional traits of woody plants: Correspondence of species rankings between field adults and laboratory-grown seedlings?
1
2003
... 植物通常会对自身资源进行配置、补偿和平衡, 以最大限度地减小环境变化对植物的不利影响, 植物功能性状(functional traits)就是这种调节机制的具体体现(Comelissen et al., 2003), 因此成为探索植物与环境之间关系的重要纽带, 也为预测在全球变化背景下的生态系统响应提供了方法和依据(Wright et al., 2005).其中, 叶片性状对气候变化十分敏感, 能较为准确地反映植物对气候变化的响应与适应机制(Royer et al., 2010; Scoffoni et al., 2011).研究表明, 氮沉降可导致植物比叶面积(SLA)、叶片氮含量(LNC)、叶片磷含量(LPC)及氮磷比(N:P)等发生显著改变(Pontes et al., 2007; 黄菊莹等, 2009; Ordoñez et al., 2009; Rose et al., 2013; 王常顺和汪诗平, 2015). ...
Herbivory: The Dynamics of Animal-Plant Interactions
... 自工业革命以来, 大气氮沉降呈现急剧增加的趋势(Holland et al., 1999; Zheng et al., 2002; Adams et al., 2004; Suding et al., 2005), 这直接影响到人类和生态系统的健康、温室气体的平衡及生物的多样性, 引起了科学家们的广泛关注与担忧(Vitousek et al., 1997; Richter et al., 2005; Clark & Tilman, 2008; Liu et al., 2013).研究表明, 氮素作为陆地生态系统中最主要的限制元素(贺金生和韩兴国, 2010), 对森林植物有重要影响(李德军等, 2005).在其他养分成为限制因子之前, 氮的增加可以提高植物的净初级生产力(Aber et al., 1989; Tamm, 1990), 但长期、连续、过量的氮输入会降低植物的净光合速率和净初级生产力, 引起森林生产力的下降和森林生态系统功能的退化(Ulrich, 1995; Fenn et al., 1996; Magill et al., 2004; 方运霆等, 2005), 这被认为是森林衰退的主要原因(Nihlgard, 1985; Bobbink et al., 1998; 张维娜和廖周瑜, 2009).因此, 研究大气氮沉降对森林植物的影响具有十分重要的意义. ...
Evidence for nitrogen saturation in the San Bernardino Mountains in Southern California
1
1996
... 自工业革命以来, 大气氮沉降呈现急剧增加的趋势(Holland et al., 1999; Zheng et al., 2002; Adams et al., 2004; Suding et al., 2005), 这直接影响到人类和生态系统的健康、温室气体的平衡及生物的多样性, 引起了科学家们的广泛关注与担忧(Vitousek et al., 1997; Richter et al., 2005; Clark & Tilman, 2008; Liu et al., 2013).研究表明, 氮素作为陆地生态系统中最主要的限制元素(贺金生和韩兴国, 2010), 对森林植物有重要影响(李德军等, 2005).在其他养分成为限制因子之前, 氮的增加可以提高植物的净初级生产力(Aber et al., 1989; Tamm, 1990), 但长期、连续、过量的氮输入会降低植物的净光合速率和净初级生产力, 引起森林生产力的下降和森林生态系统功能的退化(Ulrich, 1995; Fenn et al., 1996; Magill et al., 2004; 方运霆等, 2005), 这被认为是森林衰退的主要原因(Nihlgard, 1985; Bobbink et al., 1998; 张维娜和廖周瑜, 2009).因此, 研究大气氮沉降对森林植物的影响具有十分重要的意义. ...
Shifts in trait combinations along rainfall and phosphorus gradients
2000
Changes in total leaf nitrogen and partitioning of leaf nitrogen drive photosynthetic acclimation to light in fully developed walnut leaves
1
2001
... 黄海侠等(2013)发现, 同种植物叶片的物质分配策略在异龄叶间存在显著差异: 当年生叶的SLA显著大于往年生叶、LDMC比往年生叶小, 且当年生叶性状的变异系数往往高于往年生叶.本研究也从油松当年生叶和往年生叶的性状比较中, 发现了类似的结果.这是因为叶片在生长初期往往需要较多的资源来优化同化器官、最大程度地获取光资源(Sultan, 2000); 但在老化过程中, 内部养分元素(如氮和磷)发生迁移且光合速率降低(Bleecker, 1998; Ono et al., 2001), 结构充分形成并固化, 对环境变化的响应就变得相对迟缓(Oguchi et al., 2003); 老叶为忍受多年环境变化, 在机械和防御构造上的额外投资较多(Chabot & David, 1982).此外, 新叶对环境变化敏感, 轻微的环境扰动就能诱导其性状发生改变, 表现出较强的可塑性(Wyka et al., 2000; Frak et al., 2001).因此, 叶片的物质投资和分配格局是随叶龄变化的, 这是植物适应环境的一个重要途径(Silvertown et al., 1997). ...
Evidence of the ‘plant economics spectrum’ in a subarctic flora
1
2010
... Wright等(2004)以叶片的形态、生理、化学性状对叶片的资源权衡策略进行量化, 发现植物叶片性状沿着一条连续变化的功能性状组合谱(即叶经济谱, LES)有序排列, 其一端代表着比叶重小、含氮量高、光合速率大、呼吸速率高、叶寿命短的“快速投资-收益型”策略; 而另一端代表着寿命长、比叶重大、含氮量、光合速率和呼吸速率都偏低的“缓慢投资-收益型”策略.叶经济谱为解释碳、氮元素在植物体内的固定和利用、营养物质的流动, 以及植被的分布如何随环境条件变化而改变等问题提供了理论依据(Royer, 2009; 陈莹婷和许振柱, 2014).之后, 关于植物的资源权衡关系及全球尺度上植物叶片性状随环境的变化规律的研究大量涌现(Reich et al., 2003; Freschet et al., 2010; Laughlin, 2011; Osnas et al., 2013). ...
Plant Strategies, Vegetation Processes and Ecosystem Properties
... 自工业革命以来, 大气氮沉降呈现急剧增加的趋势(Holland et al., 1999; Zheng et al., 2002; Adams et al., 2004; Suding et al., 2005), 这直接影响到人类和生态系统的健康、温室气体的平衡及生物的多样性, 引起了科学家们的广泛关注与担忧(Vitousek et al., 1997; Richter et al., 2005; Clark & Tilman, 2008; Liu et al., 2013).研究表明, 氮素作为陆地生态系统中最主要的限制元素(贺金生和韩兴国, 2010), 对森林植物有重要影响(李德军等, 2005).在其他养分成为限制因子之前, 氮的增加可以提高植物的净初级生产力(Aber et al., 1989; Tamm, 1990), 但长期、连续、过量的氮输入会降低植物的净光合速率和净初级生产力, 引起森林生产力的下降和森林生态系统功能的退化(Ulrich, 1995; Fenn et al., 1996; Magill et al., 2004; 方运霆等, 2005), 这被认为是森林衰退的主要原因(Nihlgard, 1985; Bobbink et al., 1998; 张维娜和廖周瑜, 2009).因此, 研究大气氮沉降对森林植物的影响具有十分重要的意义. ...
Leaf nitrogen:phosphorus stoichiometry across Chinese grassland biomes
Contemporary and pre-industrial global reactive nitrogen budgets
1
1999
... 自工业革命以来, 大气氮沉降呈现急剧增加的趋势(Holland et al., 1999; Zheng et al., 2002; Adams et al., 2004; Suding et al., 2005), 这直接影响到人类和生态系统的健康、温室气体的平衡及生物的多样性, 引起了科学家们的广泛关注与担忧(Vitousek et al., 1997; Richter et al., 2005; Clark & Tilman, 2008; Liu et al., 2013).研究表明, 氮素作为陆地生态系统中最主要的限制元素(贺金生和韩兴国, 2010), 对森林植物有重要影响(李德军等, 2005).在其他养分成为限制因子之前, 氮的增加可以提高植物的净初级生产力(Aber et al., 1989; Tamm, 1990), 但长期、连续、过量的氮输入会降低植物的净光合速率和净初级生产力, 引起森林生产力的下降和森林生态系统功能的退化(Ulrich, 1995; Fenn et al., 1996; Magill et al., 2004; 方运霆等, 2005), 这被认为是森林衰退的主要原因(Nihlgard, 1985; Bobbink et al., 1998; 张维娜和廖周瑜, 2009).因此, 研究大气氮沉降对森林植物的影响具有十分重要的意义. ...
浙江天童常绿植物当年生与往年生叶片性状的变异与关联
2013
羊草绿叶氮、磷浓度和比叶面积沿氮、磷和水分梯度的变化
1
2009
... 植物通常会对自身资源进行配置、补偿和平衡, 以最大限度地减小环境变化对植物的不利影响, 植物功能性状(functional traits)就是这种调节机制的具体体现(Comelissen et al., 2003), 因此成为探索植物与环境之间关系的重要纽带, 也为预测在全球变化背景下的生态系统响应提供了方法和依据(Wright et al., 2005).其中, 叶片性状对气候变化十分敏感, 能较为准确地反映植物对气候变化的响应与适应机制(Royer et al., 2010; Scoffoni et al., 2011).研究表明, 氮沉降可导致植物比叶面积(SLA)、叶片氮含量(LNC)、叶片磷含量(LPC)及氮磷比(N:P)等发生显著改变(Pontes et al., 2007; 黄菊莹等, 2009; Ordoñez et al., 2009; Rose et al., 2013; 王常顺和汪诗平, 2015). ...
A cost-benefit analysis of leaf habit and leaf longevity of trees and their geographical pattern
Nitrification is linked to dominant leaf traits rather than functional diversity
1
2011
... Wright等(2004)以叶片的形态、生理、化学性状对叶片的资源权衡策略进行量化, 发现植物叶片性状沿着一条连续变化的功能性状组合谱(即叶经济谱, LES)有序排列, 其一端代表着比叶重小、含氮量高、光合速率大、呼吸速率高、叶寿命短的“快速投资-收益型”策略; 而另一端代表着寿命长、比叶重大、含氮量、光合速率和呼吸速率都偏低的“缓慢投资-收益型”策略.叶经济谱为解释碳、氮元素在植物体内的固定和利用、营养物质的流动, 以及植被的分布如何随环境条件变化而改变等问题提供了理论依据(Royer, 2009; 陈莹婷和许振柱, 2014).之后, 关于植物的资源权衡关系及全球尺度上植物叶片性状随环境的变化规律的研究大量涌现(Reich et al., 2003; Freschet et al., 2010; Laughlin, 2011; Osnas et al., 2013). ...
Leaf trait relationships of native and invasive plants: Community- and global-scale comparisons
2007
模拟氮沉降对南亚热带两种乔木幼苗生物量及其分配的影响
2
2005
... 自工业革命以来, 大气氮沉降呈现急剧增加的趋势(Holland et al., 1999; Zheng et al., 2002; Adams et al., 2004; Suding et al., 2005), 这直接影响到人类和生态系统的健康、温室气体的平衡及生物的多样性, 引起了科学家们的广泛关注与担忧(Vitousek et al., 1997; Richter et al., 2005; Clark & Tilman, 2008; Liu et al., 2013).研究表明, 氮素作为陆地生态系统中最主要的限制元素(贺金生和韩兴国, 2010), 对森林植物有重要影响(李德军等, 2005).在其他养分成为限制因子之前, 氮的增加可以提高植物的净初级生产力(Aber et al., 1989; Tamm, 1990), 但长期、连续、过量的氮输入会降低植物的净光合速率和净初级生产力, 引起森林生产力的下降和森林生态系统功能的退化(Ulrich, 1995; Fenn et al., 1996; Magill et al., 2004; 方运霆等, 2005), 这被认为是森林衰退的主要原因(Nihlgard, 1985; Bobbink et al., 1998; 张维娜和廖周瑜, 2009).因此, 研究大气氮沉降对森林植物的影响具有十分重要的意义. ...
... 自工业革命以来, 大气氮沉降呈现急剧增加的趋势(Holland et al., 1999; Zheng et al., 2002; Adams et al., 2004; Suding et al., 2005), 这直接影响到人类和生态系统的健康、温室气体的平衡及生物的多样性, 引起了科学家们的广泛关注与担忧(Vitousek et al., 1997; Richter et al., 2005; Clark & Tilman, 2008; Liu et al., 2013).研究表明, 氮素作为陆地生态系统中最主要的限制元素(贺金生和韩兴国, 2010), 对森林植物有重要影响(李德军等, 2005).在其他养分成为限制因子之前, 氮的增加可以提高植物的净初级生产力(Aber et al., 1989; Tamm, 1990), 但长期、连续、过量的氮输入会降低植物的净光合速率和净初级生产力, 引起森林生产力的下降和森林生态系统功能的退化(Ulrich, 1995; Fenn et al., 1996; Magill et al., 2004; 方运霆等, 2005), 这被认为是森林衰退的主要原因(Nihlgard, 1985; Bobbink et al., 1998; 张维娜和廖周瑜, 2009).因此, 研究大气氮沉降对森林植物的影响具有十分重要的意义. ...
Ecosystem response to 15 years of chronic nitrogen additions at the Harvard Forest LTER, Massachusetts, USA
1
2004
... 自工业革命以来, 大气氮沉降呈现急剧增加的趋势(Holland et al., 1999; Zheng et al., 2002; Adams et al., 2004; Suding et al., 2005), 这直接影响到人类和生态系统的健康、温室气体的平衡及生物的多样性, 引起了科学家们的广泛关注与担忧(Vitousek et al., 1997; Richter et al., 2005; Clark & Tilman, 2008; Liu et al., 2013).研究表明, 氮素作为陆地生态系统中最主要的限制元素(贺金生和韩兴国, 2010), 对森林植物有重要影响(李德军等, 2005).在其他养分成为限制因子之前, 氮的增加可以提高植物的净初级生产力(Aber et al., 1989; Tamm, 1990), 但长期、连续、过量的氮输入会降低植物的净光合速率和净初级生产力, 引起森林生产力的下降和森林生态系统功能的退化(Ulrich, 1995; Fenn et al., 1996; Magill et al., 2004; 方运霆等, 2005), 这被认为是森林衰退的主要原因(Nihlgard, 1985; Bobbink et al., 1998; 张维娜和廖周瑜, 2009).因此, 研究大气氮沉降对森林植物的影响具有十分重要的意义. ...
Disentangling coordination among functional traits using an individual-centred model: Impact on plant performance at intra- and inter-specific levels
2013
Rebuilding community ecology from functional traits
The ammonium hypothesis—An additional explanation to the forest dieback in Europe
1
1985
... 自工业革命以来, 大气氮沉降呈现急剧增加的趋势(Holland et al., 1999; Zheng et al., 2002; Adams et al., 2004; Suding et al., 2005), 这直接影响到人类和生态系统的健康、温室气体的平衡及生物的多样性, 引起了科学家们的广泛关注与担忧(Vitousek et al., 1997; Richter et al., 2005; Clark & Tilman, 2008; Liu et al., 2013).研究表明, 氮素作为陆地生态系统中最主要的限制元素(贺金生和韩兴国, 2010), 对森林植物有重要影响(李德军等, 2005).在其他养分成为限制因子之前, 氮的增加可以提高植物的净初级生产力(Aber et al., 1989; Tamm, 1990), 但长期、连续、过量的氮输入会降低植物的净光合速率和净初级生产力, 引起森林生产力的下降和森林生态系统功能的退化(Ulrich, 1995; Fenn et al., 1996; Magill et al., 2004; 方运霆等, 2005), 这被认为是森林衰退的主要原因(Nihlgard, 1985; Bobbink et al., 1998; 张维娜和廖周瑜, 2009).因此, 研究大气氮沉降对森林植物的影响具有十分重要的意义. ...
Does the photosynthetic light-acclimation need change in leaf anatomy?
1
2003
... 黄海侠等(2013)发现, 同种植物叶片的物质分配策略在异龄叶间存在显著差异: 当年生叶的SLA显著大于往年生叶、LDMC比往年生叶小, 且当年生叶性状的变异系数往往高于往年生叶.本研究也从油松当年生叶和往年生叶的性状比较中, 发现了类似的结果.这是因为叶片在生长初期往往需要较多的资源来优化同化器官、最大程度地获取光资源(Sultan, 2000); 但在老化过程中, 内部养分元素(如氮和磷)发生迁移且光合速率降低(Bleecker, 1998; Ono et al., 2001), 结构充分形成并固化, 对环境变化的响应就变得相对迟缓(Oguchi et al., 2003); 老叶为忍受多年环境变化, 在机械和防御构造上的额外投资较多(Chabot & David, 1982).此外, 新叶对环境变化敏感, 轻微的环境扰动就能诱导其性状发生改变, 表现出较强的可塑性(Wyka et al., 2000; Frak et al., 2001).因此, 叶片的物质投资和分配格局是随叶龄变化的, 这是植物适应环境的一个重要途径(Silvertown et al., 1997). ...
Possible mechanisms of adaptive leaf senescence
1
2001
... 黄海侠等(2013)发现, 同种植物叶片的物质分配策略在异龄叶间存在显著差异: 当年生叶的SLA显著大于往年生叶、LDMC比往年生叶小, 且当年生叶性状的变异系数往往高于往年生叶.本研究也从油松当年生叶和往年生叶的性状比较中, 发现了类似的结果.这是因为叶片在生长初期往往需要较多的资源来优化同化器官、最大程度地获取光资源(Sultan, 2000); 但在老化过程中, 内部养分元素(如氮和磷)发生迁移且光合速率降低(Bleecker, 1998; Ono et al., 2001), 结构充分形成并固化, 对环境变化的响应就变得相对迟缓(Oguchi et al., 2003); 老叶为忍受多年环境变化, 在机械和防御构造上的额外投资较多(Chabot & David, 1982).此外, 新叶对环境变化敏感, 轻微的环境扰动就能诱导其性状发生改变, 表现出较强的可塑性(Wyka et al., 2000; Frak et al., 2001).因此, 叶片的物质投资和分配格局是随叶龄变化的, 这是植物适应环境的一个重要途径(Silvertown et al., 1997). ...
A global study of relationships between leaf traits, climate and soil measures of nutrient fertility
3
2009
... 植物通常会对自身资源进行配置、补偿和平衡, 以最大限度地减小环境变化对植物的不利影响, 植物功能性状(functional traits)就是这种调节机制的具体体现(Comelissen et al., 2003), 因此成为探索植物与环境之间关系的重要纽带, 也为预测在全球变化背景下的生态系统响应提供了方法和依据(Wright et al., 2005).其中, 叶片性状对气候变化十分敏感, 能较为准确地反映植物对气候变化的响应与适应机制(Royer et al., 2010; Scoffoni et al., 2011).研究表明, 氮沉降可导致植物比叶面积(SLA)、叶片氮含量(LNC)、叶片磷含量(LPC)及氮磷比(N:P)等发生显著改变(Pontes et al., 2007; 黄菊莹等, 2009; Ordoñez et al., 2009; Rose et al., 2013; 王常顺和汪诗平, 2015). ...
... 一般认为, 氮添加会导致SLA的增大(Rose et al., 2013), 因为氮添加促进了植物的生长而提高了植物的光合效率(Ordoñez et al., 2009).万宏伟等(2008)和黄菊莹等(2009)研究发现, 羊草(Leymus chinensis)等几种草本植物的SLA因氮添加而显著增大; Bassin等(2009)的氮添加实验也表明, 虽然大多数植物叶片的比叶重(LMA, SLA的倒数)无显著变化, 但两种植物叶片的LMA显著降低(即SLA显著增大).然而, 并非所有研究结果都是如此, 赵新风等(2014)发现, 钠猪毛菜(Salsola nitraria)受氮添加影响, SLA呈下降趋势, LDMC呈增大趋势, 与上述结果相反.研究表明, 较小的SLA和较高的LDMC使得植物具有更好的资源获取能力、碳氮固持效率及水分利用效率, 在逆境中更具优势(Wilson et al., 1999; Aerts & Chapin, 2000; Zhang et al., 2007).因此, 这两项指标常被用来指示植物对环境变化的响应策略, SLA的减小和LDMC的增大往往意味着环境条件的恶化, 因为植物开始趋向于保守的生长策略(Rose et al., 2013).本研究结果表明, 大部分植物的SLA和LDMC并没有表现出显著差异, 只有茶条枫和玉竹的SLA呈显著减小趋势, 油松当年生叶和玉竹叶片的LDMC显著增大, 与赵新风等(2014)的结果相一致.这说明不同植物对氮添加的响应不同.并且, 氮添加对油松、茶条枫和玉竹这3种植物的生长是不利的. ...
Global leaf trait relationships: Mass, area, and the leaf economics spectrum
1
2013
... Wright等(2004)以叶片的形态、生理、化学性状对叶片的资源权衡策略进行量化, 发现植物叶片性状沿着一条连续变化的功能性状组合谱(即叶经济谱, LES)有序排列, 其一端代表着比叶重小、含氮量高、光合速率大、呼吸速率高、叶寿命短的“快速投资-收益型”策略; 而另一端代表着寿命长、比叶重大、含氮量、光合速率和呼吸速率都偏低的“缓慢投资-收益型”策略.叶经济谱为解释碳、氮元素在植物体内的固定和利用、营养物质的流动, 以及植被的分布如何随环境条件变化而改变等问题提供了理论依据(Royer, 2009; 陈莹婷和许振柱, 2014).之后, 关于植物的资源权衡关系及全球尺度上植物叶片性状随环境的变化规律的研究大量涌现(Reich et al., 2003; Freschet et al., 2010; Laughlin, 2011; Osnas et al., 2013). ...
Within-species correlations in leaf traits of three boreal plant species along a latitudinal gradient
2010
Leaf traits affect the above-ground productivity and quality of pasture grasses
1
2007
... 植物通常会对自身资源进行配置、补偿和平衡, 以最大限度地减小环境变化对植物的不利影响, 植物功能性状(functional traits)就是这种调节机制的具体体现(Comelissen et al., 2003), 因此成为探索植物与环境之间关系的重要纽带, 也为预测在全球变化背景下的生态系统响应提供了方法和依据(Wright et al., 2005).其中, 叶片性状对气候变化十分敏感, 能较为准确地反映植物对气候变化的响应与适应机制(Royer et al., 2010; Scoffoni et al., 2011).研究表明, 氮沉降可导致植物比叶面积(SLA)、叶片氮含量(LNC)、叶片磷含量(LPC)及氮磷比(N:P)等发生显著改变(Pontes et al., 2007; 黄菊莹等, 2009; Ordoñez et al., 2009; Rose et al., 2013; 王常顺和汪诗平, 2015). ...
Global patterns of plant leaf N and P in relation to temperature and latitude
The evolution of plant functional variation: Traits, spectra, and strategies
1
2003
... Wright等(2004)以叶片的形态、生理、化学性状对叶片的资源权衡策略进行量化, 发现植物叶片性状沿着一条连续变化的功能性状组合谱(即叶经济谱, LES)有序排列, 其一端代表着比叶重小、含氮量高、光合速率大、呼吸速率高、叶寿命短的“快速投资-收益型”策略; 而另一端代表着寿命长、比叶重大、含氮量、光合速率和呼吸速率都偏低的“缓慢投资-收益型”策略.叶经济谱为解释碳、氮元素在植物体内的固定和利用、营养物质的流动, 以及植被的分布如何随环境条件变化而改变等问题提供了理论依据(Royer, 2009; 陈莹婷和许振柱, 2014).之后, 关于植物的资源权衡关系及全球尺度上植物叶片性状随环境的变化规律的研究大量涌现(Reich et al., 2003; Freschet et al., 2010; Laughlin, 2011; Osnas et al., 2013). ...
Increase in tropospheric nitrogen dioxide over China observed from space
1
2005
... 自工业革命以来, 大气氮沉降呈现急剧增加的趋势(Holland et al., 1999; Zheng et al., 2002; Adams et al., 2004; Suding et al., 2005), 这直接影响到人类和生态系统的健康、温室气体的平衡及生物的多样性, 引起了科学家们的广泛关注与担忧(Vitousek et al., 1997; Richter et al., 2005; Clark & Tilman, 2008; Liu et al., 2013).研究表明, 氮素作为陆地生态系统中最主要的限制元素(贺金生和韩兴国, 2010), 对森林植物有重要影响(李德军等, 2005).在其他养分成为限制因子之前, 氮的增加可以提高植物的净初级生产力(Aber et al., 1989; Tamm, 1990), 但长期、连续、过量的氮输入会降低植物的净光合速率和净初级生产力, 引起森林生产力的下降和森林生态系统功能的退化(Ulrich, 1995; Fenn et al., 1996; Magill et al., 2004; 方运霆等, 2005), 这被认为是森林衰退的主要原因(Nihlgard, 1985; Bobbink et al., 1998; 张维娜和廖周瑜, 2009).因此, 研究大气氮沉降对森林植物的影响具有十分重要的意义. ...
Management alters interspecific leaf trait relationships and trait-based species rankings in permanent meadows
4
2013
... 植物通常会对自身资源进行配置、补偿和平衡, 以最大限度地减小环境变化对植物的不利影响, 植物功能性状(functional traits)就是这种调节机制的具体体现(Comelissen et al., 2003), 因此成为探索植物与环境之间关系的重要纽带, 也为预测在全球变化背景下的生态系统响应提供了方法和依据(Wright et al., 2005).其中, 叶片性状对气候变化十分敏感, 能较为准确地反映植物对气候变化的响应与适应机制(Royer et al., 2010; Scoffoni et al., 2011).研究表明, 氮沉降可导致植物比叶面积(SLA)、叶片氮含量(LNC)、叶片磷含量(LPC)及氮磷比(N:P)等发生显著改变(Pontes et al., 2007; 黄菊莹等, 2009; Ordoñez et al., 2009; Rose et al., 2013; 王常顺和汪诗平, 2015). ...
... 一般认为, 氮添加会导致SLA的增大(Rose et al., 2013), 因为氮添加促进了植物的生长而提高了植物的光合效率(Ordoñez et al., 2009).万宏伟等(2008)和黄菊莹等(2009)研究发现, 羊草(Leymus chinensis)等几种草本植物的SLA因氮添加而显著增大; Bassin等(2009)的氮添加实验也表明, 虽然大多数植物叶片的比叶重(LMA, SLA的倒数)无显著变化, 但两种植物叶片的LMA显著降低(即SLA显著增大).然而, 并非所有研究结果都是如此, 赵新风等(2014)发现, 钠猪毛菜(Salsola nitraria)受氮添加影响, SLA呈下降趋势, LDMC呈增大趋势, 与上述结果相反.研究表明, 较小的SLA和较高的LDMC使得植物具有更好的资源获取能力、碳氮固持效率及水分利用效率, 在逆境中更具优势(Wilson et al., 1999; Aerts & Chapin, 2000; Zhang et al., 2007).因此, 这两项指标常被用来指示植物对环境变化的响应策略, SLA的减小和LDMC的增大往往意味着环境条件的恶化, 因为植物开始趋向于保守的生长策略(Rose et al., 2013).本研究结果表明, 大部分植物的SLA和LDMC并没有表现出显著差异, 只有茶条枫和玉竹的SLA呈显著减小趋势, 油松当年生叶和玉竹叶片的LDMC显著增大, 与赵新风等(2014)的结果相一致.这说明不同植物对氮添加的响应不同.并且, 氮添加对油松、茶条枫和玉竹这3种植物的生长是不利的. ...
Nutrient turnover rates in ancient terrestrial ecosystems
1
2009
... Wright等(2004)以叶片的形态、生理、化学性状对叶片的资源权衡策略进行量化, 发现植物叶片性状沿着一条连续变化的功能性状组合谱(即叶经济谱, LES)有序排列, 其一端代表着比叶重小、含氮量高、光合速率大、呼吸速率高、叶寿命短的“快速投资-收益型”策略; 而另一端代表着寿命长、比叶重大、含氮量、光合速率和呼吸速率都偏低的“缓慢投资-收益型”策略.叶经济谱为解释碳、氮元素在植物体内的固定和利用、营养物质的流动, 以及植被的分布如何随环境条件变化而改变等问题提供了理论依据(Royer, 2009; 陈莹婷和许振柱, 2014).之后, 关于植物的资源权衡关系及全球尺度上植物叶片性状随环境的变化规律的研究大量涌现(Reich et al., 2003; Freschet et al., 2010; Laughlin, 2011; Osnas et al., 2013). ...
Leaf economic traits from fossils support a weedy habit for early angiosperms
1
2010
... 植物通常会对自身资源进行配置、补偿和平衡, 以最大限度地减小环境变化对植物的不利影响, 植物功能性状(functional traits)就是这种调节机制的具体体现(Comelissen et al., 2003), 因此成为探索植物与环境之间关系的重要纽带, 也为预测在全球变化背景下的生态系统响应提供了方法和依据(Wright et al., 2005).其中, 叶片性状对气候变化十分敏感, 能较为准确地反映植物对气候变化的响应与适应机制(Royer et al., 2010; Scoffoni et al., 2011).研究表明, 氮沉降可导致植物比叶面积(SLA)、叶片氮含量(LNC)、叶片磷含量(LPC)及氮磷比(N:P)等发生显著改变(Pontes et al., 2007; 黄菊莹等, 2009; Ordoñez et al., 2009; Rose et al., 2013; 王常顺和汪诗平, 2015). ...
Decline of leaf hydraulic conductance with dehydration: Relationship to leaf size and venation architecture
1
2011
... 植物通常会对自身资源进行配置、补偿和平衡, 以最大限度地减小环境变化对植物的不利影响, 植物功能性状(functional traits)就是这种调节机制的具体体现(Comelissen et al., 2003), 因此成为探索植物与环境之间关系的重要纽带, 也为预测在全球变化背景下的生态系统响应提供了方法和依据(Wright et al., 2005).其中, 叶片性状对气候变化十分敏感, 能较为准确地反映植物对气候变化的响应与适应机制(Royer et al., 2010; Scoffoni et al., 2011).研究表明, 氮沉降可导致植物比叶面积(SLA)、叶片氮含量(LNC)、叶片磷含量(LPC)及氮磷比(N:P)等发生显著改变(Pontes et al., 2007; 黄菊莹等, 2009; Ordoñez et al., 2009; Rose et al., 2013; 王常顺和汪诗平, 2015). ...
Fundamental trade-offs generating the worldwide leaf economics spectrum
... 黄海侠等(2013)发现, 同种植物叶片的物质分配策略在异龄叶间存在显著差异: 当年生叶的SLA显著大于往年生叶、LDMC比往年生叶小, 且当年生叶性状的变异系数往往高于往年生叶.本研究也从油松当年生叶和往年生叶的性状比较中, 发现了类似的结果.这是因为叶片在生长初期往往需要较多的资源来优化同化器官、最大程度地获取光资源(Sultan, 2000); 但在老化过程中, 内部养分元素(如氮和磷)发生迁移且光合速率降低(Bleecker, 1998; Ono et al., 2001), 结构充分形成并固化, 对环境变化的响应就变得相对迟缓(Oguchi et al., 2003); 老叶为忍受多年环境变化, 在机械和防御构造上的额外投资较多(Chabot & David, 1982).此外, 新叶对环境变化敏感, 轻微的环境扰动就能诱导其性状发生改变, 表现出较强的可塑性(Wyka et al., 2000; Frak et al., 2001).因此, 叶片的物质投资和分配格局是随叶龄变化的, 这是植物适应环境的一个重要途径(Silvertown et al., 1997). ...
Functional- and abundance-based mechanisms explain diversity loss due to N fertilization
1
2005
... 自工业革命以来, 大气氮沉降呈现急剧增加的趋势(Holland et al., 1999; Zheng et al., 2002; Adams et al., 2004; Suding et al., 2005), 这直接影响到人类和生态系统的健康、温室气体的平衡及生物的多样性, 引起了科学家们的广泛关注与担忧(Vitousek et al., 1997; Richter et al., 2005; Clark & Tilman, 2008; Liu et al., 2013).研究表明, 氮素作为陆地生态系统中最主要的限制元素(贺金生和韩兴国, 2010), 对森林植物有重要影响(李德军等, 2005).在其他养分成为限制因子之前, 氮的增加可以提高植物的净初级生产力(Aber et al., 1989; Tamm, 1990), 但长期、连续、过量的氮输入会降低植物的净光合速率和净初级生产力, 引起森林生产力的下降和森林生态系统功能的退化(Ulrich, 1995; Fenn et al., 1996; Magill et al., 2004; 方运霆等, 2005), 这被认为是森林衰退的主要原因(Nihlgard, 1985; Bobbink et al., 1998; 张维娜和廖周瑜, 2009).因此, 研究大气氮沉降对森林植物的影响具有十分重要的意义. ...
Phenotypic plasticity for plant development, function and life history
1
2000
... 黄海侠等(2013)发现, 同种植物叶片的物质分配策略在异龄叶间存在显著差异: 当年生叶的SLA显著大于往年生叶、LDMC比往年生叶小, 且当年生叶性状的变异系数往往高于往年生叶.本研究也从油松当年生叶和往年生叶的性状比较中, 发现了类似的结果.这是因为叶片在生长初期往往需要较多的资源来优化同化器官、最大程度地获取光资源(Sultan, 2000); 但在老化过程中, 内部养分元素(如氮和磷)发生迁移且光合速率降低(Bleecker, 1998; Ono et al., 2001), 结构充分形成并固化, 对环境变化的响应就变得相对迟缓(Oguchi et al., 2003); 老叶为忍受多年环境变化, 在机械和防御构造上的额外投资较多(Chabot & David, 1982).此外, 新叶对环境变化敏感, 轻微的环境扰动就能诱导其性状发生改变, 表现出较强的可塑性(Wyka et al., 2000; Frak et al., 2001).因此, 叶片的物质投资和分配格局是随叶龄变化的, 这是植物适应环境的一个重要途径(Silvertown et al., 1997). ...
Nitrogen in Terrestrial Ecosystems: Questions of Productivity, Vegetational Change, and Ecological Stability
1
1990
... 自工业革命以来, 大气氮沉降呈现急剧增加的趋势(Holland et al., 1999; Zheng et al., 2002; Adams et al., 2004; Suding et al., 2005), 这直接影响到人类和生态系统的健康、温室气体的平衡及生物的多样性, 引起了科学家们的广泛关注与担忧(Vitousek et al., 1997; Richter et al., 2005; Clark & Tilman, 2008; Liu et al., 2013).研究表明, 氮素作为陆地生态系统中最主要的限制元素(贺金生和韩兴国, 2010), 对森林植物有重要影响(李德军等, 2005).在其他养分成为限制因子之前, 氮的增加可以提高植物的净初级生产力(Aber et al., 1989; Tamm, 1990), 但长期、连续、过量的氮输入会降低植物的净光合速率和净初级生产力, 引起森林生产力的下降和森林生态系统功能的退化(Ulrich, 1995; Fenn et al., 1996; Magill et al., 2004; 方运霆等, 2005), 这被认为是森林衰退的主要原因(Nihlgard, 1985; Bobbink et al., 1998; 张维娜和廖周瑜, 2009).因此, 研究大气氮沉降对森林植物的影响具有十分重要的意义. ...
基于氮排放数据的中国大陆大气氮素湿沉降量估算
2010
The history and possible causes of forest decline in central Europe, with particular attention to the German situation
1
1995
... 自工业革命以来, 大气氮沉降呈现急剧增加的趋势(Holland et al., 1999; Zheng et al., 2002; Adams et al., 2004; Suding et al., 2005), 这直接影响到人类和生态系统的健康、温室气体的平衡及生物的多样性, 引起了科学家们的广泛关注与担忧(Vitousek et al., 1997; Richter et al., 2005; Clark & Tilman, 2008; Liu et al., 2013).研究表明, 氮素作为陆地生态系统中最主要的限制元素(贺金生和韩兴国, 2010), 对森林植物有重要影响(李德军等, 2005).在其他养分成为限制因子之前, 氮的增加可以提高植物的净初级生产力(Aber et al., 1989; Tamm, 1990), 但长期、连续、过量的氮输入会降低植物的净光合速率和净初级生产力, 引起森林生产力的下降和森林生态系统功能的退化(Ulrich, 1995; Fenn et al., 1996; Magill et al., 2004; 方运霆等, 2005), 这被认为是森林衰退的主要原因(Nihlgard, 1985; Bobbink et al., 1998; 张维娜和廖周瑜, 2009).因此, 研究大气氮沉降对森林植物的影响具有十分重要的意义. ...
Human alteration of the global nitrogen cycle: Sources and consequences
1
1997
... 自工业革命以来, 大气氮沉降呈现急剧增加的趋势(Holland et al., 1999; Zheng et al., 2002; Adams et al., 2004; Suding et al., 2005), 这直接影响到人类和生态系统的健康、温室气体的平衡及生物的多样性, 引起了科学家们的广泛关注与担忧(Vitousek et al., 1997; Richter et al., 2005; Clark & Tilman, 2008; Liu et al., 2013).研究表明, 氮素作为陆地生态系统中最主要的限制元素(贺金生和韩兴国, 2010), 对森林植物有重要影响(李德军等, 2005).在其他养分成为限制因子之前, 氮的增加可以提高植物的净初级生产力(Aber et al., 1989; Tamm, 1990), 但长期、连续、过量的氮输入会降低植物的净光合速率和净初级生产力, 引起森林生产力的下降和森林生态系统功能的退化(Ulrich, 1995; Fenn et al., 1996; Magill et al., 2004; 方运霆等, 2005), 这被认为是森林衰退的主要原因(Nihlgard, 1985; Bobbink et al., 1998; 张维娜和廖周瑜, 2009).因此, 研究大气氮沉降对森林植物的影响具有十分重要的意义. ...
羊草草原群落6种植物叶片功能特性对氮素添加的响应
2008
植物叶片性状对气候变化的响应研究进展
1
2015
... 植物通常会对自身资源进行配置、补偿和平衡, 以最大限度地减小环境变化对植物的不利影响, 植物功能性状(functional traits)就是这种调节机制的具体体现(Comelissen et al., 2003), 因此成为探索植物与环境之间关系的重要纽带, 也为预测在全球变化背景下的生态系统响应提供了方法和依据(Wright et al., 2005).其中, 叶片性状对气候变化十分敏感, 能较为准确地反映植物对气候变化的响应与适应机制(Royer et al., 2010; Scoffoni et al., 2011).研究表明, 氮沉降可导致植物比叶面积(SLA)、叶片氮含量(LNC)、叶片磷含量(LPC)及氮磷比(N:P)等发生显著改变(Pontes et al., 2007; 黄菊莹等, 2009; Ordoñez et al., 2009; Rose et al., 2013; 王常顺和汪诗平, 2015). ...
Plant ecological strategies: Some leading dimensions of variation between species
Modulation of leaf economic traits and trait relationships by climate
3
2005
... 植物通常会对自身资源进行配置、补偿和平衡, 以最大限度地减小环境变化对植物的不利影响, 植物功能性状(functional traits)就是这种调节机制的具体体现(Comelissen et al., 2003), 因此成为探索植物与环境之间关系的重要纽带, 也为预测在全球变化背景下的生态系统响应提供了方法和依据(Wright et al., 2005).其中, 叶片性状对气候变化十分敏感, 能较为准确地反映植物对气候变化的响应与适应机制(Royer et al., 2010; Scoffoni et al., 2011).研究表明, 氮沉降可导致植物比叶面积(SLA)、叶片氮含量(LNC)、叶片磷含量(LPC)及氮磷比(N:P)等发生显著改变(Pontes et al., 2007; 黄菊莹等, 2009; Ordoñez et al., 2009; Rose et al., 2013; 王常顺和汪诗平, 2015). ...
... 以欧洲氮饱和实验NITREX项目(Wright & Rasmussen, 1998)和北美哈佛森林(Harvard Forest)等(Aber et al., 1998)研究设计为参照, 在未被破坏的天然油松林(112.02°-112.25° E, 36.52°-36.72° N, 平均海拔1500 m)中建立12个20 m × 20 m的固定试验样地, 设置对照(CK, 0 kg·hm-2·a-1)、低氮(LN, 50 kg·hm-2·a-1)、中氮(MN, 100 kg·hm-2·a-1)和高氮(HN, 150 kg·hm-2·a-1) 4个处理, 3次重复.样地林分特征及0-20 cm土壤理化性质详见表1.从2009年5月起, 用尿素(CO(NH2)2)作为氮源, 将年施用量平均分成12等份, 每月下旬分别将所需的尿素溶解于2 L水中, 用背式喷雾器在对应处理的20 m × 20 m样地内均匀喷洒(对照组喷施等量的水), 其他水源来自自然降水. ...
Leaf age as a factor in anatomical and physiological acclimative responses of Taxus baccata L. needles to contrasting irradiance environments
1
2000
... 黄海侠等(2013)发现, 同种植物叶片的物质分配策略在异龄叶间存在显著差异: 当年生叶的SLA显著大于往年生叶、LDMC比往年生叶小, 且当年生叶性状的变异系数往往高于往年生叶.本研究也从油松当年生叶和往年生叶的性状比较中, 发现了类似的结果.这是因为叶片在生长初期往往需要较多的资源来优化同化器官、最大程度地获取光资源(Sultan, 2000); 但在老化过程中, 内部养分元素(如氮和磷)发生迁移且光合速率降低(Bleecker, 1998; Ono et al., 2001), 结构充分形成并固化, 对环境变化的响应就变得相对迟缓(Oguchi et al., 2003); 老叶为忍受多年环境变化, 在机械和防御构造上的额外投资较多(Chabot & David, 1982).此外, 新叶对环境变化敏感, 轻微的环境扰动就能诱导其性状发生改变, 表现出较强的可塑性(Wyka et al., 2000; Frak et al., 2001).因此, 叶片的物质投资和分配格局是随叶龄变化的, 这是植物适应环境的一个重要途径(Silvertown et al., 1997). ...
Seasonal variation in photosynthesis in six woody species with different leaf phenology in a valley savanna in southwestern China
1
2007
... 一般认为, 氮添加会导致SLA的增大(Rose et al., 2013), 因为氮添加促进了植物的生长而提高了植物的光合效率(Ordoñez et al., 2009).万宏伟等(2008)和黄菊莹等(2009)研究发现, 羊草(Leymus chinensis)等几种草本植物的SLA因氮添加而显著增大; Bassin等(2009)的氮添加实验也表明, 虽然大多数植物叶片的比叶重(LMA, SLA的倒数)无显著变化, 但两种植物叶片的LMA显著降低(即SLA显著增大).然而, 并非所有研究结果都是如此, 赵新风等(2014)发现, 钠猪毛菜(Salsola nitraria)受氮添加影响, SLA呈下降趋势, LDMC呈增大趋势, 与上述结果相反.研究表明, 较小的SLA和较高的LDMC使得植物具有更好的资源获取能力、碳氮固持效率及水分利用效率, 在逆境中更具优势(Wilson et al., 1999; Aerts & Chapin, 2000; Zhang et al., 2007).因此, 这两项指标常被用来指示植物对环境变化的响应策略, SLA的减小和LDMC的增大往往意味着环境条件的恶化, 因为植物开始趋向于保守的生长策略(Rose et al., 2013).本研究结果表明, 大部分植物的SLA和LDMC并没有表现出显著差异, 只有茶条枫和玉竹的SLA呈显著减小趋势, 油松当年生叶和玉竹叶片的LDMC显著增大, 与赵新风等(2014)的结果相一致.这说明不同植物对氮添加的响应不同.并且, 氮添加对油松、茶条枫和玉竹这3种植物的生长是不利的. ...
氮沉降增加对森林植物影响的研究进展
1
2009
... 自工业革命以来, 大气氮沉降呈现急剧增加的趋势(Holland et al., 1999; Zheng et al., 2002; Adams et al., 2004; Suding et al., 2005), 这直接影响到人类和生态系统的健康、温室气体的平衡及生物的多样性, 引起了科学家们的广泛关注与担忧(Vitousek et al., 1997; Richter et al., 2005; Clark & Tilman, 2008; Liu et al., 2013).研究表明, 氮素作为陆地生态系统中最主要的限制元素(贺金生和韩兴国, 2010), 对森林植物有重要影响(李德军等, 2005).在其他养分成为限制因子之前, 氮的增加可以提高植物的净初级生产力(Aber et al., 1989; Tamm, 1990), 但长期、连续、过量的氮输入会降低植物的净光合速率和净初级生产力, 引起森林生产力的下降和森林生态系统功能的退化(Ulrich, 1995; Fenn et al., 1996; Magill et al., 2004; 方运霆等, 2005), 这被认为是森林衰退的主要原因(Nihlgard, 1985; Bobbink et al., 1998; 张维娜和廖周瑜, 2009).因此, 研究大气氮沉降对森林植物的影响具有十分重要的意义. ...
养分与水分添加对荒漠草地植物钠猪毛菜功能性状的影响
2014
The Asian nitrogen cycle case study
1
2002
... 自工业革命以来, 大气氮沉降呈现急剧增加的趋势(Holland et al., 1999; Zheng et al., 2002; Adams et al., 2004; Suding et al., 2005), 这直接影响到人类和生态系统的健康、温室气体的平衡及生物的多样性, 引起了科学家们的广泛关注与担忧(Vitousek et al., 1997; Richter et al., 2005; Clark & Tilman, 2008; Liu et al., 2013).研究表明, 氮素作为陆地生态系统中最主要的限制元素(贺金生和韩兴国, 2010), 对森林植物有重要影响(李德军等, 2005).在其他养分成为限制因子之前, 氮的增加可以提高植物的净初级生产力(Aber et al., 1989; Tamm, 1990), 但长期、连续、过量的氮输入会降低植物的净光合速率和净初级生产力, 引起森林生产力的下降和森林生态系统功能的退化(Ulrich, 1995; Fenn et al., 1996; Magill et al., 2004; 方运霆等, 2005), 这被认为是森林衰退的主要原因(Nihlgard, 1985; Bobbink et al., 1998; 张维娜和廖周瑜, 2009).因此, 研究大气氮沉降对森林植物的影响具有十分重要的意义. ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}