随着全球气候变化加剧, 碳循环直接影响着人类的生存和各种生物环境的稳定性, 逐渐成为生态系统研究的重点(杨景成等, 2003 ; Bardgett et al ., 2013 )。陆地生态系统在全球碳循环中起着重要的作用, 因此对陆地生态系统碳循环的理解非常必要(Canadell et al ., 2000 )。我国现有草地面积约4亿hm2 , 是最大的陆地生态系统, 也是生物地球循环的重要组成部分, 但其容易受到气候变化和各种人类活动(如: 放牧、施肥等)的影响(齐玉春等, 2003 ; Xu & Guo, 2015 )。目前, 由于放牧和农垦活动频繁, 草地生态系统成为受人类活动影响较为严重的区域。因此, 对自然和人为因素干扰下草地生态系统碳循环过程的研究具有十分重要的科学意义。
草地凋落物是指由植物地上部分和地下根系形成并归还到土壤的所有有机质的总称, 是陆地生态系统养分循环和碳循环过程中极为重要的环节(高永恒等, 2007 )。陆地生态系统中约90%的净初级生产量以凋落物的形式归还给土壤, 植物的生长发育在很大程度上依赖于植被归还给土壤的养分再循环(Wardle et al ., 2004 )。凋落物的分解是一个非常复杂的过程, 深受气候、凋落物质量和土壤生物的影响, 在很大程度上决定了生态系统的固碳速率及动态(Tateno et al ., 2007 )。因此, 在全球变化背景下, 对草地生态系统凋落物分解的研究是草地生态系统动态机理以及全球变化生态学研究的重要内容(张艳博等, 2012 )。另外, 地下凋落物的碳输入是地上部分凋落物碳输入的3倍以上, 但是过去仅有2%的植物凋落物研究关注于地下凋落物分解, 很少有研究对干旱、半干旱草原的地上和地下凋落物同时进行系统研究(Zhang et al ., 2008 ; Freschet et al ., 2013 )。因此研究不同干扰下草地地下凋落物分解的变化, 对准确估算全球变化背景下不同尺度碳收支至关重要(Solly et al ., 2014 )。
放牧和围栏封育是草地最基本的两种管理方式, 它们直接改变地表的植被特征, 也改变土壤环境(如温度、含水量等)及其养分含量, 使植物碳分配模式和土壤微生物代谢过程等发生改变, 进而影响凋落物分解和生态系统碳循环(Raich & Tufekciogiu, 2000 )。放牧是草地生态系统生物地球化学循环的重要驱动因子, 不同放牧强度下, 牲畜通过采食、排泄和践踏等方式影响植物群落结构和土壤特性, 使凋落物形成和分解发生变化(Stark et al ., 2015 )。但是目前关于放牧强度对地下凋落物分解的影响研究不足。围栏封育是保持草地生态系统功能的有效管理方式。放牧草地经过围栏封育后, 使草地植被得到恢复, 土壤特性、碳输入、植物组成以及凋落物基质质量发生了变化, 这些因素都影响了草地凋落物的形成和分解(王蕾等, 2013 ; Lu et al ., 2015 )。目前, 国内外学者对草地围栏封育的研究仅局限在不同年限的围封对群落结构和凋落物的影响。研究表明, 围封时间过长会引起群落地上生物量降低、凋落物堆积, 抑制植物的再生, 一部分优势种开始衰退, 阻碍草地资源的更新利用(Eldridge et al ., 1992 ; 左万庆等, 2009 ), 因此研究放牧后的短期围封对草地恢复更具指导意义。目前, 草地不同退化强度基础上的短期围封研究尚且不足(Yeo, 2005 ; Hu et al ., 2016 )。因此, 我们研究不同放牧强度下的短期围栏封育对凋落物分解和草地生产力恢复的影响, 以期为防止草地退化、改善生态环境提供参考依据。
凋落物基质质量能够表征凋落物在自然界中的可分解性。衡量凋落物质量的指标主要是C含量、N含量、木质素浓度、C:N、木质素:N等。一般来说, 凋落物中N含量越高、C:N越小、木质素含量越低, 凋落物分解速率越快(Koukoura et al ., 2003 )。在干旱和半干旱草地生态系统中, 降水是影响凋落物分解的重要气候因子, 它可以通过影响植物群落组成和分解者的活性来影响凋落物质量, 进而影响凋落物分解。但降水和凋落物质量可能存在交互作用, 使得我们无法准确评估某一因子对分解的影响(Bontti et al ., 2009 )。本研究在内蒙古温带草原上开展, 2011年和2012年该区域的降水量分别是188 mm和465 mm, 存在明显差异, 使得本文恰好能够探究降水差异对草地凋落物分解的影响。本文在内蒙古温带典型草原中测定了干湿年份内4种样地(轻度放牧样地、轻度放牧后围封样地、重度放牧样地、重度放牧后围封样地)的土壤养分含量、凋落物产量、质量及分解速率来探讨不同放牧强度对凋落物形成和分解的影响, 以及围栏封育对生态系统恢复的作用, 具体研究目标如下: 1)分析不同放牧强度和围栏封育下凋落物分解的差异; 2)探讨调控地上、地下凋落物分解的主要因子; 3)揭示降水变化对凋落物产量和分解的作用。通过讨论分析, 以期为草地生态系统和草地畜牧业的可持续发展提供理论依据。
1 材料和方法
1.1 环境概况和样地设计
研究样地位于内蒙古大学草地生态系统研究站(116.03°-116.50° E, 44.80°-44.82° N), 海拔约1055 m, 属于温带干旱大陆性气候, 年平均气温0-1 ℃, ≥0 ℃的积温1800 ℃, 年降水量300-360 mm, 无霜期90-115天。生长季从5月到9月, 非生长季从10月到次年4月。气象站观测数据显示: 2011-2012年年平均气温1.7 ℃, 2011年、2012年的累积降水量分别为188 mm和465 mm, 降水多集中在6-8月份。土壤以栗钙土为主。该实验样地植被类型属于温带典型草原, 大针茅(Stipa grandis )、羊草(Leymus chinensis )、冷蒿(Artemisia frigida )在该区的植物群落中占优势地位。样地选在以居民点为中心的自由放牧方式家庭牧场, 该牧场为长期放牧区, 整个生长季都进行放牧。这一地区的主要特点是以居民点为中心,呈条带放射状向外构成不同放牧梯度。根据与居民点的距离远近, 将放牧区划分为轻度放牧样地和重度放牧样地。划分方法是以居民点为中心在放牧地上放射状拉3条样带, 样带之间有一定的距离, 使样带所包含的信息尽可能代表整个草地(王明军等, 2007 )。实验样地整体放牧强度为每hm2 、每年、每天674标准羊单位。实验开始前, 对样地进行基本群落调查, 收获地上生物量, 尤其是对样地内的羊粪便进行计数。每个样地羊粪数和群落调查基本数据均作为样地划分的依据, 样地基本信息见表1 。2011年实验开始时, 在每一条退化样带上的轻度和重度放牧样地各围封了3个退化样地, 每个样地围封面积为100 m × 100 m, 分别为轻度放牧后围封样地和重度放牧后围封样地。
1.2 凋落物产量的测定
我们用生长季内地上生物量的最大值代表该年地上凋落物的产量(Zhang et al ., 2013 )。分别于2011年和2012年生长高峰期8月下旬, 在轻度放牧后围封、轻度放牧、重度放牧后围封、重度放牧4个样地内随机布设5个样方(1 m × 1 m), 收集样方内全部地上生物量, 不包括直立或倒伏的凋落物, 去除植物体表面的泥土和杂质, 在105 ℃下杀青15 min, 然后在65 ℃下烘干至恒质量后进行称量。在生长季末, 植物体将全部凋亡, 以凋落物的形式归还到土壤中。地下凋落物产量采用内生长土芯法测定, 于2011年4月中旬在4个样地内随机布设5个样点, 按照0-10、10-20、20-30、30-40 cm 4个垂直层次, 用根钻打钻, 每钻土壤不混合, 分层收集, 带回实验室立即处理。在室内尽量挑出每层土壤的全部根系, 然后将处理好的无根土壤对应回填。回填时, 按照土层顺序对应将每两层装入一个圆柱形尼龙网袋中, 然后按上下两层对应放回原位。于每年生长季初和生长季末, 每个样点各取样一次, 每个样点取上下两层土芯网袋。带回实验室后清洗网袋中全部活根系, 在105 ℃下杀青15 min, 然后在65 ℃下 烘干至恒质量。用生长季初和生长季末根新生量 的差值作为该年地下凋落物产量(Montané et al ., 2010 )。
1.3 凋落物分解速率的测定
2010年8月上旬, 分别在4个样地内随机选择5个样点, 随机布设5个样方(1 m × 1 m)采集地上凋落物, 采集的凋落物在自然条件下风干, 去除植物体表面的泥土和杂质。用于地上凋落物分解实验。去除地表植被后, 按照0-10、10-20、20-30、30-40 cm 4个垂直层次, 用根钻采集地下完整的活根系, 自然风干。采用凋落物网袋法进行实验, 将地上、地下凋落物剪成20 cm的小段, 取10 g地上凋落物或者5 g地下凋落物装入1 mm孔径的尼龙网袋(10 cm × 10 cm), 每个样地36袋地上凋落物网袋和36袋地下凋落物网袋。在每个样地内选取3个样点, 去除地表植被, 在每个样点将12袋地上凋落物网袋均匀平铺在地表与土壤直接接触。在样点附近, 将12袋地下凋落物网袋均匀埋放在土深10 cm左右的土层。2011-2012年5月和9月下旬, 在每样地3个样点内各收集3袋凋落物。将网袋内凋落物全部取出洗净, 在65 ℃下烘干至恒质量, 进行称量。利用每次收获得到的残留凋落物干质量, 根据Olson (1963) 指数分解模型公式, 计算分解速率k 值:
X /X 0 = e- kt
式中, X 表示经过t 年分解时间后凋落物的残留量, X 0 为凋落物初始量。
1.4 凋落物质量的测定
将2011年和2012年每次洗净烘干的凋落物样品进行粉碎研磨, 再通过40目网筛。一部分用于凋落物C、N含量的测定, 另一部分用于木质素和纤维素含量的测定。全C含量的测定采用重铬酸钾法(Nelson & Sommers, 1982 ), 全N含量的测定采用凯氏定氮法(Bremner, 1960 )。木质素和纤维素的测定依据van Soest (1963, 1967)的实验原理, 参照AOAC international (2000)的具体实验操作标准, 经过消煮-酸洗-灼烧等步骤, 测定各样地初始凋落物和残留凋落物中的木质素和纤维素的含量。
1.5 土壤养分的测定
于2011-2012年每月收获凋落物网袋的同时, 在每个样地的实验样点附近布设1 m × 1 m的样方, 去除地表植被后, 利用土钻打5钻采集0-40 cm层次的土壤样品, 带回实验室进行处理。肉眼挑出土壤中可见的植物根系和杂质, 然后通过网筛将细根系和土壤区分开来, 尽量挑出土壤中的全部根系, 在自然状态下风干数日。对土壤样品进行研磨, 土壤全C、全N测量方法与凋落物C、N的测定方法相同。
1.6 数据统计与分析
实验数据首先利用SPSS 17.0进行重复测量方差分析, 为了解同一年内单变量是否存在显著性差异, 进行了单因素方差分析和LSD 多重比较(p = 0.05), 并运用双因素方差分析进一步分析各变量及其相互作用对凋落物产量和分解的影响。利用线性回归对不同放牧强度和围封条件下凋落物产量、质量、土壤养分与分解速率的关系进行分析。所有图件使用Origin 8.5软件制作。
2 结果和分析
2.1 不同放牧强度和围封样地的凋落物产量
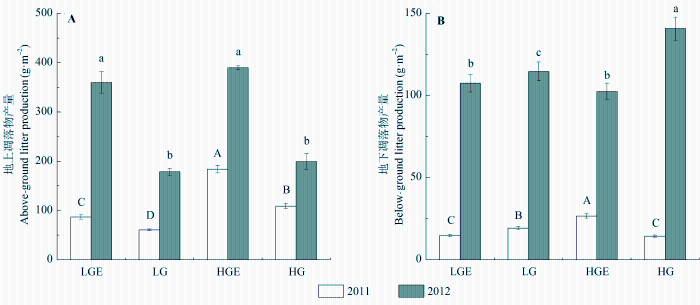
地上和地下凋落物产量在样地间差异显著, 在干旱的2011年和湿润的2012年间也存在显著差异, 且样地和年份的交互作用对凋落物产量有显著影响(表2 , 表3 ; p < 0.01)。在干旱年, 重度放牧样地地上凋落物产量显著高于轻度放牧样地(p < 0.05), 但在湿润年差异不显著。两年中, 围封样地地上凋落物产量均显著高于对应的放牧样地(图1A ; p < 0.05), 尤其在湿润年, 轻度放牧后围封样地产量是轻度放牧样地的2.01倍, 重度放牧后围封样地产量是重度放牧样地的1.96倍。地下凋落物产量表现为干旱年重度放牧后围封样地最高, 轻度放牧高于重度放牧(图1B ; p < 0.01), 湿润年重度放牧样地最高。相同样地湿润年凋落物产量极显著高于干旱年(p < 0.01), 2012年轻度放牧后围封样地地上凋落物产量是2011年的4.14倍, 2012年重度放牧地下凋落物产量是2011年的9.83倍。
图1 2011和2012年4个样地地上凋落物(A)和地下凋落物(B)产量(平均值±标准误差)。不同大写字母表示2011年凋落物产量在不同样地之间差异显著, 不同小写字母表示2012年凋落物产量在不同样地之间差异显著(p < 0.05)。HG, 重度放牧样地; HGE, 重度放牧后围封样地; LG, 轻度放牧样地; LGE, 轻度放牧后围封样地。
Fig. 1 Above-ground (A) and below-ground (B) litter production in the four treatment plots in 2011 and 2012 (mean ± SE ). Capital letters indicate significant difference in litter production between different plots in 2011 at 0.05 levels, and lowercase letters indicate significant difference in litter production between different plots in 2012 at 0.05 levels. HG, heavy-grazing; HGE, heavy-grazing exclusion; LG, light-grazing; LGE, light-grazing exclusion .
2.2 不同放牧强度和围封样地的初始凋落物质量
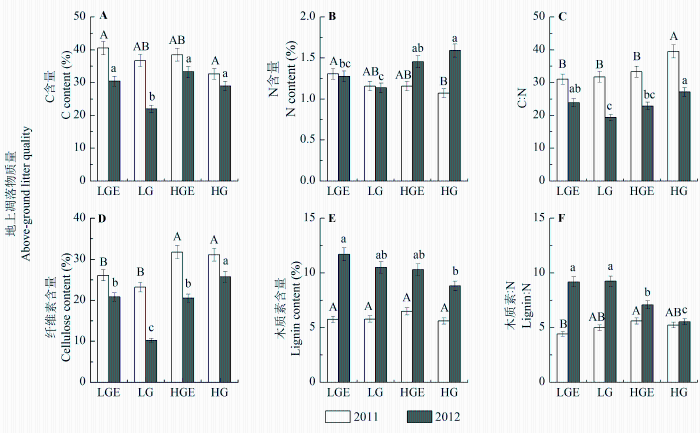
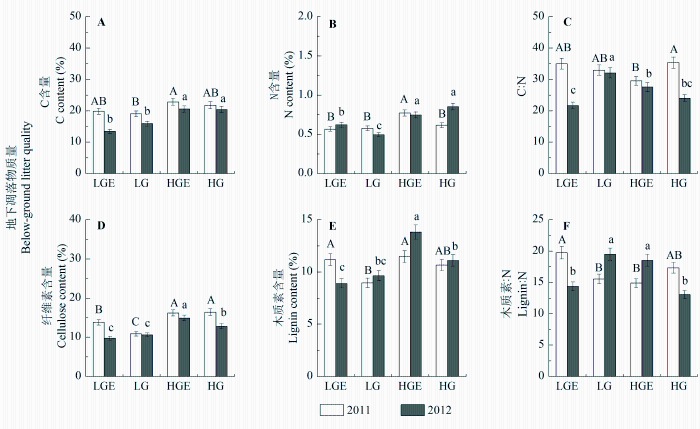
地上和地下凋落物质量指标在样地和年份间普遍存在显著差异(p < 0.01), 且样地和年份的交互作用对地下凋落物质量有显著影响(p < 0.01; 表2 , 表3 )。在干旱年, 轻度放牧样地地上凋落物的C:N显著低于重度放牧(p < 0.05), 轻度放牧后围封样地与轻度放牧地上凋落物质量无显著差异, 重度放牧后围封样地地上凋落物C:N显著低于重度放牧(p < 0.05); 在湿润年, 轻度放牧样地地上凋落物的C含量、N含量和C:N均显著低于重度放牧样地(p < 0.05), 轻度放牧后围封样地地上凋落物C含量和C:N显著高于轻度放牧(p < 0.05), 重度放牧后围封样地较重度放牧样地地上凋落物具有高木质素:N (p < 0.05)、低C:N (p < 0.05)。对地下凋落物而言, 干旱年轻度放牧和重度放牧各指标无显著差异, 轻度放牧后围封样地凋落物质量比轻度放牧差(木质素、木质素:N高), 重度放牧后围封样地质量比重度放牧好(高N含量、低C:N); 湿润年重度放牧样地地下凋落物质量较轻度放牧样地好(低C:N和木质素:N)。地上、地下凋落物分解速率受到凋落物质量指标的影响不同。地上凋落物分解速率与地上凋落物N含量显著正相关(R 2 = 0.489, p < 0.01); 地下凋落物分解与地下凋落物C含量(R 2 = 0.263, p < 0.01)、C:N (R 2 = 0.349, p < 0.01)和纤维素含量(R 2 = 0.460, p < 0.01)显著负相关。
图2 2011和2012年4个样地地上凋落物C含量(A)、N含量(B)、C:N (C)、纤维素含量(D)、木质素含量(E)和木质素:N (F)(平均值±标准误差), 不同大写字母表示2011年地上凋落物质量在不同样地之间差异显著, 不同小写字母表示2012年地上凋落物质量在不同样地之间差异显著(p < 0.05)。HG, 重度放牧样地; HGE, 重度放牧后围封样地; LG, 轻度放牧样地; LGE, 轻度放牧后围封样地。
Fig. 2 Above-ground litter C content (A), N content (B), C:N (C), cellulose content (D), lignin content (E) and lignin:N (F) in four plots in 2011 and 2012 (mean ± SE ). Capital letters indicate significant difference in litter quality between different plots in 2011 at 0.05 levels, and lowercase letters indicate significant difference in litter quality between different plots in 2012 at 0.05 levels. HG, heavy-grazing; HGE, heavy-grazing exclusion; LG, light-grazing; LGE, light-grazing exclusion.
图3 2011和2012年4个样地地下凋落物C含量(A)、N含量(B)、C:N (C)、纤维素含量(D)、木质素含量(E)和木质素:N (F) (平均值±标准误差)。 不同大写字母表示2011年地下凋落物质量在不同样地之间差异显著, 不同小写字母表示2012年地下凋落物质量在不同样地之间差异显著(p < 0.05)。HG, 重度放牧样地; HGE, 重度放牧后围封样地; LG, 轻度放牧样地; LGE, 轻度放牧后围封样地。
Fig. 3 Below-ground litter C content (A), N content (B), C:N (C), cellulose content (D), lignin content (E) and lignin:N (F) in four plots in 2011 and 2012 (mean ± SE ). Different capital letters indicate significant difference in litter quality between different plots in 2011 at 0.05 levels, and different lowercase letters indicate significant difference in litter quality between different plots in 2012 at 0.05 levels. HG, heavy-grazing; HGE, heavy-grazing exclusion; LG, light-grazing; LGE, light-grazing exclusion.
2.3 不同放牧强度和围封样地的凋落物分解速率
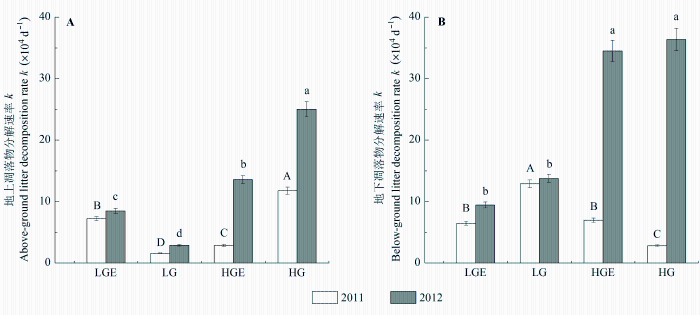
不同样地和年份间地上和地下凋落物分解速率均差异显著, 且样地和年份的交互作用也显著影响凋落物分解(p < 0.01; 表2 , 表3 )。在干旱年和湿润年, 地上凋落物分解速率均表现为重度放牧>轻度放牧(p = 0.05), 轻度放牧后围封>轻度放牧(p < 0.01), 重度放牧>重度放牧后围封(图4A ; p < 0.01)。干旱年地下凋落物分解速率在轻度放牧样地最高, 重度放牧样地最低; 湿润年重度放牧样地分解速率显著高于轻度放牧样地(图4B ; p < 0.01), 但围封与放牧样地间差异不显著。降水对地上凋落物分解速率有极显著促进作用(p < 0.01)。除轻度放牧样地外, 随着降水的增加, 地下凋落物分解也显著加快(p < 0.01)。
2.4 不同放牧强度和围封样地的土壤养分含量
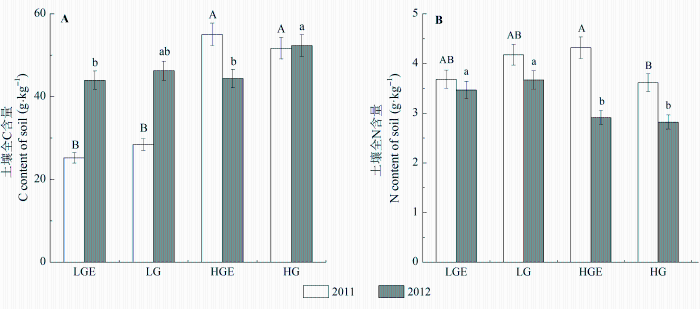
不同样地和年份间土壤全C、全N含量差异显著, 且样地和年份间交互作用也显著影响土壤全C含量(表2 , 表3 ; p < 0.01)。土壤全C含量在干旱年表现为轻度放牧样地显著低于重度放牧样地(p < 0.01), 而轻度放牧后围封和轻度放牧、重度放牧后围封和重度放牧样地之间差异不显著; 在湿润年表现为重度放牧样地显著高于重度放牧后围封样地(p < 0.05), 而轻度放牧与重度放牧、轻度放牧后围封与轻度放牧样地之间差异不显著(图5A )。土壤全N含量在干旱年表现为重度放牧后围封样地显著高于重度放牧样地(p < 0.05), 但在轻度放牧后围封和轻度放牧样地之间、轻度放牧和重度放牧样地之间不存在显著差异; 湿润年轻度放牧样地土壤全N显著高于重度放牧(p < 0.05), 但轻度放牧后围封与轻度放牧样地之间、重度放牧后围封与重度放牧样地之间不存在显著差异(图5B )。另外, 地上凋落物分解速率与土壤全N呈显著负相关(R 2 = 0.450, p < 0.01)。
图4 2011和2012年4个样地地上凋落物(A)和地下凋落物(B)分解速率k (平均值±标准误差)。不同大写字母表示2011年凋落物分解速率在不同样地之间差异显著; 不同小写字母表示2012年凋落物分解速率在不同样地之间差异显著(p < 0.05)。HG, 重度放牧样地; HGE, 重度放牧后围封样地; LG, 轻度放牧样地; LGE, 轻度放牧后围封样地。
Fig. 4 Aboveground (A) and belowground (B) litter decomposition rates (k ) in four plots in 2011 and 2012 (mean ± SE ). Different capital letters indicate significant difference in litter decomposition rate between different plots in 2011 at 0.05 levels, and different lowercase letters indicate significant difference in litter decomposition rate between different plots in 2012 at 0.05 levels. HG, heavy-grazing; HGE, heavy-grazing exclusion; LG, light-grazing; LGE, light-grazing exclusion.
图5 2011和2012年4个样地土壤全C (A)和全N (B)含量(平均值±标准误差)。不同大写字母表示2011年土壤C、N在不同样地之间差异显著, 不同小写字母表示2012年土壤C、N在不同样地之间差异显著(p < 0.05)。HG, 重度放牧样地; HGE, 重度放牧后围封样地; LG, 轻度放牧样地; LGE, 轻度放牧后围封样地。
Fig. 5 Soil total C (A) and total N content (B) in four plots in 2011 and 2012 (mean ± SE ). Different capital letters indicate significant difference in soil nutrient content between different plots in 2011 at 0.05 levels, and different lowercase letters indicate significant difference in soil nutrient content between different plots in 2012 at 0.05 levels. HG, heavy-grazing; HGE, heavy-grazing exclusion; LG, light-grazing; LGE, light-grazing exclusion.
3 讨论
3.1 不同放牧强度对凋落物形成和分解的影响
放牧牲畜通过采食、践踏和排泄作用影响凋落物形成和分解。放牧对凋落物的影响主要表现在两个方面: 一是放牧使植物物种组成、牧草本身的形态学特征(如根冠比、叶片/叶鞘等)发生改变, 从而影响凋落物起始质量(Holland & Detling, 1990 ; Semmartin & Ghersa, 2006 ); 二是放牧使凋落物分解的立地土壤物理和生物环境发生改变, 直接影响凋落物分解(Shariff & Grygiel, 1994 )。本研究中, 重度放牧样地地上凋落物产量显著高于轻度放牧, 因为放牧可以削减草地群落的物种冗余程度, 牲畜唾液可以引入生长刺激物, 使植物产生补偿效应(Gong et al ., 2015 ); 两年中重度放牧比轻度放牧更能促进地上凋落物分解, 重度放牧样地地上凋落物质量好, C、N含量高、木质素:N低, 加上牲畜的踩踏作用加快凋落物的碎裂并与土壤紧密接触和混合。同时牲畜啃食会使植被盖度减少, 地表太阳辐射增多, 表层土壤温度和微生物活性提高, 都可以促进地上凋落物分解(张成霞和南志标, 2010 ; Hewins et al ., 2013 )。另外, 长期重度放牧使叶片有变小的趋势, 这是植物在功能属性上忍受放牧和加快更新的措施, 这些变化使其凋落后的分解更加容易(Cingolani et al ., 2005 ; An & Li, 2014 )。综上所述, 重度放牧加快了地上凋落物的产生和分解, 有利于凋落物中碳氮参与草地生态系统中的碳氮循环, 对维持草地生态系统中碳氮平衡具有重要意义。然而, 重度放牧可能会使土壤物理性质变差, 这将造成重度放牧对草地生态系统物质循环的整体效应降低, 不利于草地的健康发展。因此, 合理放牧才是保证草地健康发展的管理方式。
植物分配一定的光合物质到地下以维持根系的产量, 同时根系吸收土壤中的水分和养分供给地上部分生长。因此植物地上和根系的生长紧密相连, 地上和地下凋落物的产生和分解也是相互影响的(Bai et al ., 2015 )。植物根系是土壤有机质的主要来源, 放牧主要通过影响土壤微环境(温度和湿度)和根系特征来影响地下凋落物产生和分解, 进而影响草地生态系统土壤养分循环(Smith et al ., 2014 )。本研究中, 干旱年重度放牧样地地下凋落物产量显著低于轻度放牧样地, 湿润年则相反。这可能是由于重度放牧样地家畜的践踏作用使土壤孔隙度和水稳性团聚体减少, 引起土壤透水性、透气性和水导率下降, 加上在干旱年水分亏缺严重, 严重阻碍根系的生长(侯扶江等, 2004 )。而湿润年降水充足, 土壤理化性质变好, 适宜根系生长, 重度放牧样地地下凋落物产量迅速增加, 显著高于轻度放牧样地。另外在本研究中, 干旱年重度放牧地下凋落物分解速率显著低于轻度放牧, 湿润年则相反。在干旱年, 重度放牧使植被盖度显著下降, 导致地表裸露、土温上升、土壤水分散失加剧、土壤物理性质变差; 另一方面, 土壤可利用性水分减少, 加上内蒙古草原的高海拔使重度放牧样地裸露的地表UV-B辐射增多, 大幅降低土壤分解者的活性, 这些都会严重阻碍重度放牧样地地下凋落物的分解(Dormaar & Willms, 1990 ; Giese et al ., 2009 )。但在湿润年, 重度放牧地下凋落物质量变好, 低C:N和木质素:N使重度放牧样地地下分解速率显著高于轻度放牧, 这也说明重度放牧样地地下凋落物分解对降水条件的响应十分敏感。因此我们的实验结果显示放牧对半干旱草地生态系统凋落物产生和分解的影响主要取决于环境的降水条件。
3.2 围栏封育对凋落物形成和分解的影响
围栏封育是草地有效管理方式之一, 它对植被资源恢复、土壤理化性质恢复和生物多样性保护具有重要意义。围封后植被生长状况、土壤无脊椎动物和微生物活性以及凋落物分解的关键过程都会发生改变(Lindsay & Cunningham, 2009 )。本研究中, 两年内放牧样地地上凋落物产量在围封后均有显著提高。由于围栏封育后排除人为和牲畜干扰, 植被得到恢复, 地上生物量、凋落物积累和土壤有机质输入显著提高(Tessier et al ., 2003 )。围封后除了植被得到恢复外, 土壤条件也得到明显改善。放牧样地经过围封后, 土壤容重降低, 养分含量增加, 土壤酶活性增强, 这些都可以促进围封样地地上凋落物形成(文海燕等, 2005 )。在本研究两年围封期间, 轻度放牧和轻度放牧后围封样地凋落物质量和土壤养分之间的差异并不显著, 但轻度放牧后围封样地地上凋落物分解速率加快, 可能是土壤物理和生物特性发生变化的结果(Mekuria et al ., 2007 )。与之相反, 重度放牧围封后地上凋落物分解速率减慢, 凋落物质量变差, 说明重度放牧围封后植被组成发生了变化, 植物叶片结构性碳(木质素)合成增加, 导致地上凋落物木质素含量和木质素:N升高, 凋落物中难分解物质比例增加(Bontti et al ., 2009 )。围封后排除了牲畜的排泄作用, 土壤有机质含量减少, 土壤酶活性和微生物活性降低, 这都将导致重度放牧样地围封后地上凋落物分解减慢(Aldezabal et al ., 2015 )。由此可以推测, 长期轻度放牧样地生态系统的恢复作用更强, 短期围封后地上凋落物分解速率和养分循环加快。
在本研究中, 短期围封后地下凋落物产量反而显著低于对应的放牧样地。放牧活动能促使根系向土壤上层集中, 且它对根系的生长有促进作用, 因此围封后根系生长变缓(Greenwood & Hutchinson, 2003 ; Pucheta et al ., 2004 )。但只有在干旱年, 重度放牧后围封样地地下凋落物产量显著高于重度放牧。这可能是由于重度放牧样地牲畜的踩踏作用使土壤遭到破坏, 加上干旱年降水稀少, 限制了地下根系生长。在干旱年, 放牧和围封样地地下凋落物分解速率存在显著差异主要是凋落物质量差异所致。轻度放牧样地围封后, 地下凋落物质量变差, 高木质素含量和木质素:N使得轻度放牧后围封样地地下凋落物分解速率显著低于轻度放牧样地; 而重度放牧样地相反, 围封后凋落物质量和土壤养分状况变好(图3 , 图5 ), 使得重度放牧样地围封后分解速率显著高于重度放牧样地。湿润年各样地的地下凋落物质量和土壤养分之间虽然存在差异, 但各样地地下凋落物分解速率并无显著差异。相比于凋落物质量和土壤养分的作用, 降水对地下凋落物分解过程的调控作用更强(Bontti et al ., 2009 ; Giese et al ., 2009 )。综上所述, 本研究表明短期围栏封育能使植被生长环境、土壤理化性质得到明显改善, 轻度放牧围封后还能提高凋落物分解速率和养分循环。不过有研究表明, 长期围封会使草地非禾本科牧草比例大幅增加, 不利于禾本科牧草产量的积累, 这将会阻碍畜牧业发展, 造成严重的经济损失(Shi et al ., 2010 )。因此, 在草地放牧强度和围封年限的确定上, 我们应注意寻找草地经济效益和生态系统保护之间的平衡点。
3.3 降水对凋落物分解的作用
除放牧强度和围栏封育外, 降水变化对凋落物分解也有显著影响。降水通过影响土壤温湿度, 改变植物群落组成和功能, 从而影响凋落物质量和土壤微生物活性, 调控凋落物形成和分解过程(Austin & Vitousek, 2001 )。本研究中, 湿润年地上、地下凋落物产量和分解速率均高于干旱年。干旱年植物生长受水分限制, 导致气孔关闭、光合能力下降, 植物生产力降低(Sanaullah et al ., 2014 ), 产生的凋落物也较湿润年少。在轻度放牧和轻度放牧后围封样地, 湿润年土壤全C含量是干旱年的近2倍。湿润年降水增多可提高土壤含水量和酶活性(Mikan et al ., 2002 )、加快凋落物化学组分的淋溶, 从地表输入土壤的C增多, 根系的分解也随之加快, 土壤C输入显著提高。并且土壤可利用水增多可加快土壤中的养分向微生物种群扩散, 使微生物活性增强(Wang et al ., 2015 )。这都可以显著促进凋落物分解过程, 加快草地生态系统的碳循环和养分循环。另外, 在草地生态系统中, 由于植物根生物量比其他生态系统大, 较快的根系分解速率和周转速率有助于维持土壤养分的供应和植物的生长(Liu et al ., 2009 )。因此降水还可以通过促进地下凋落物分解而增加土壤养分, 进而促进植物地上部分生长和凋落物分解过程。
4 结论
放牧通过影响草地植被和土壤特性, 导致土壤养分、凋落物生产、质量及分解过程发生变化。与轻度放牧相比, 重度放牧更能促进地上凋落物形成和分解, 有利于半干旱草地生态系统碳循环和养分循环。短期围栏封育能使植被生长环境、土壤理化性质得到改善。轻度放牧草地围封后地上凋落物分解速率加快, 重度放牧草地围封后分解变慢。因此轻度放牧草地的生态恢复更适合围栏封育, 而重度放牧草地的生态恢复仅凭围封是不够的, 可能还需辅以必要的人工恢复措施。降水是调控凋落物形成和分解的关键因子, 降水增多可以显著促进地上、地下凋落物的形成和分解。不同放牧强度地下凋落物的生产和分解受降水年际波动影响很大, 重度放牧草地地下凋落物对降水变化的敏感度较轻度放牧草地高。总之, 凋落物分解是一个复杂的生态过程, 影响其分解过程的因子有环境因子、凋落物基质质量、土壤物理和生物特性及其交互作用等。由于围封时间较短和降水的影响, 本实验不能很好地评估凋落物质量和土壤养分对分解的影响, 需进一步深入地研究。
致谢 感谢内蒙古大学对本研究的大力支持以及宝音陶格涛教授在野外工作中提供的帮助。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Aldezabal A Moragues L Odriozola I Mijangos I 2015 ). Impact of grazing abandonment on plant and soil microbial communities in an Atlantic mountain grassland.
Applied Soil Ecology , 96 , 251 -260 .
[本文引用: 1]
[2]
An H Li GQ 2014 ). Differential effects of grazing on plant functional traits in the desert grassland.
Polish Journal of Ecology , 62 , 239 -251 .
[本文引用: 1]
[3]
AOAC International 2000). AOAC official method 973.18 fiber (acid detergent) and lignin (H2SO4) in animal feed. In: Horwitz W ed. Official Methods of Analysis of AOAC International, 17th edn . Association of Official Analytical Chemists , Gaithersburg, USA .
[4]
Austin AT Vitousek PM 2001 ). Precipitation, decomposition, and litter decomposability of Metrosideros polymorpha on Hawaii.
Journal of Ecology , 88 , 129 -138 .
[本文引用: 1]
[5]
Bai WM Fang Y Zhou M Xie T Li LH Zhang WH 2015 ). Heavily intensified grazing reduces root production in an Inner Mongolia temperate steppe.
Agriculture Ecosystems & Environment , 200 , 143 -150 .
[本文引用: 1]
[6]
Bardgett RD Manning P Elly M Franciska V 2013 ). Hierarchical responses of plant-soil interactions to climate change: Consequences for the global carbon cycle.
Journal of Ecology , 101 , 334 -343 .
[本文引用: 1]
[7]
Bontti EE Decant JP Munson SM Gathany MA Przeszlowska A Haddix ML Owens S Burke IC Parton WJ Harmon ME 2009 ). Litter decomposition in grasslands of Central North America (US Great Plains).
Global Change Biology , 15 , 1356 -1363 .
[本文引用: 3]
[8]
Bremner JM 1960 ). Determination of nitrogen in soil by the Kjeldahl method.
Journal of Agricultural Science , 55 , 11 -33 .
[本文引用: 1]
[9]
Canadell JG Mooney HA Baldocchi DD Berry JA Ehleringer JR Field CB Gower ST Hollinger DY Hunt JE 2000 ). Carbon metabolism of the terrestrial biosphere: A multitechnique approach for improved understanding.
Ecosystems , 3 , 115 -130 .
[本文引用: 1]
[10]
Cingolani AM Posse G Collantes MB 2005 ). Plant functional traits, herbivore selectivity and response to sheep grazing in Patagonian steppe grasslands.
Journal of Applied Ecology , 42 (42 ), 50 -59 .
[本文引用: 1]
[11]
Dormaar JF Willms WD 1990 ). Effect of grazing and cultiva- tion on some chemical properties of soils in the mixed prairie.
Journal of Range Management , 43 , 456 -460 .
[本文引用: 1]
[12]
Eldridge DJ Westoby M Holbrook KM 1992 ). Soil surface characteristics, microtopography and proximity to mature shrubs: Effects on survival of several cohorts of Atriplex vesicaria seedlings.
Journal of Ecology , 78 , 357 -364 .
[本文引用: 1]
[13]
Freschet GT Cornwell WK Wardle DA Elumeeva TG Liu W Jackson BG Onipchenko VG Soudzilovskaia NA Tao J Cornelissen JH 2013 ). Linking litter decomposition of above- and below-ground organs to plant-soil feedbacks worldwide.
Journal of Ecology , 101 , 943 -952 .
[本文引用: 1]
[14]
Giese M Gao YZ Zhao Y Pan QM Lin S Peth S Brueck H 2009 ). Effects of grazing and rainfall variability on root and shoot decomposition in a semi-arid grassland.
Applied Soil Ecology , 41 , 8 -18 .
[本文引用: 2]
[15]
Gao YH Chen H Luo P Wu N Wang GX 2007 ). Effects of grazing intensity on decompositions of two dominant plant species litters in alpine meadow on the Northwester Sichuan.
Ecological Science , 26 (3 ), 193 -198 . (in Chinese with English abstract)[高永恒 , 陈槐 , 罗鹏 , 吴宁 , 王根绪 (2007 ). 放牧强度对川西北高山草甸两个优势物种凋落物分解的影响
. 生态科学 , 26 (3 ), 193 -198 .]
[16]
Gong XY Fanselow N Dittert K Taube F Lin S 2015 ). Re- sponse of primary production and biomass allocation to nitrogen and water supplementation along a grazing intensity gradient in semiarid grassland.
European Journal of Agronomy , 63 , 27 -35 .
[本文引用: 1]
[17]
Greenwood KL Hutchinson KJ 2003 ). Root characteristics of temperate pasture in New South Wales after grazing at three stocking rates for 30 years.
Grass & Forage Science , 53 , 120 -128 .
[本文引用: 1]
[18]
Hewins DB Archer SR Okin GS Mcculley RL Throop HL 2013 ). Soil-Litter mixing accelerates decomposition in a Chihuahuan Desert Grassland.
Ecosystems , 16 , 183 -195 .
[本文引用: 1]
[19]
Holland CA Detling JK 1990 ). Plant response to herbivory and belowground nitrogen cycling.
Ecology , 71 , 1040 -1049 .
[本文引用: 1]
[20]
Hou FJ Chang SH Yu YW Lin HL 2004 ). A review on trampling by grazed livestock.
Acta Ecologica Sinica , 24 , 784 -789 . (in Chinese with English abstract)[侯扶江 , 常生华 , 于应文 , 林慧龙 (2004 ). 放牧家畜的践踏作用研究评述
. 生态学报 , 24 , 784 -789 .]
[本文引用: 1]
[21]
Hu ZM Li S Guo Q Niu SL He NP Li LH Yu GR 2016 ). A synthesis of the effect of grazing exclusion on carbon dynamics in grasslands in China.
Global Change Biology , 22 , 1385 -1393 .
[22]
Koukoura Z Mamolos AP Kalburtji KL 2003 ). Decomposition of dominant plant species litter in a semi-arid grassland.
Applied Soil Ecology , 23 , 13 -23 .
[本文引用: 1]
[23]
Lindsay EA Cunningham SA 2009 ). Livestock grazing exclusion and microhabitat variation affect invertebrates and litter decomposition rates in woodland remnants.
Forest Ecology & Management , 258 , 178 -187 .
[本文引用: 1]
[24]
Liu L King JS Booker FL Giardina CP Allen HL Hu SJ 2009 ). Enhanced litter input rather than changes in litter chemistry drive soil carbon and nitrogen cycles under elevated CO2: A microcosm study.
Global Change Biology , 15 , 441 -453 .
[本文引用: 1]
[25]
Lu X Yan Y Sun J Zhang XK Chen YC Wang XD Cheng GW 2015 ). Carbon, nitrogen, and phosphorus storage in alpine grassland ecosystems of Tibet: Effects of grazing exclusion.
Ecology & Evolution , 5 , 4492 -4504 .
[本文引用: 1]
[26]
Mekuria W Veldkamp E Haile M Nyssen J Muys B Gebrehiwot K 2007 ). Effectiveness of exclosures to restore degraded soils as a result of overgrazing in Tigray, Ethiopia.
Journal of Arid Environments , 69 , 270 -284 .
[本文引用: 1]
[27]
Mikan CJ Schimel JP Doyle AP 2002 ). Temperature controls of microbial respiration in arctic tundra soils above and below freezing.
Soil Biology & Biochemistry , 34 , 1785 -1795 .
[本文引用: 1]
[28]
Montané F Romanyà J Rovira P Casals P 2010 ). Aboveground litter quality changes may drive soil organic carbon increase after shrub encroachment into mountain grasslands.
Plant & Soil , 337 , 151 -165 .
[本文引用: 1]
[29]
Nelson DW Sommers LE Total carbon, organic carbon and organic matter. In: Page AL, Miller RH, Keeney DR eds. Methods of Soil Analysis . American Society of Agronomy , Madison, USA . 539 -579 .
[本文引用: 2]
[30]
Olson JS 1963 ). Energy storage and the balance of producers and decomposers in ecological systems.
Ecology , 44 , 322 -331 .
[本文引用: 2]
[31]
Pucheta E Bonamici I Cabido M Díaz S 2004 ). Below- ground biomass and productivity of a grazed site and a neighbouring ungrazed exclosure in a grassland in central Argentina.
Austral Ecology , 29 , 201 -208 .
[32]
Qi YC Dong YS Geng YB Yang XH Geng HL 2003 ). The progress in the carbon cycle researches in grassland ecosystem in China.
Progress in Geography , 22 , 342 -352 . (in Chinese with English abstract)[齐玉春 , 董云社 , 耿元波 ,杨小红 , 耿会立 (2003 ). 我国草地生态系统碳循环研究进展
. 地理科学进展 , 22 , 342 -352 .]
[33]
Raich JW Tufekciogul A 2000 ). Vegetation and soil respiration: Correlations and controls.
Biogeochemistry , 48 , 71 -90 .
[本文引用: 2]
[34]
Sanaullah M Chabbi A Girardin C Durand JL Poirier M Rumpel C 2014 ). Effects of drought and elevated temperature on biochemical composition of forage plants and their impact on carbon storage in grassland soil.
Plant & Soil , 374 , 767 -778 .
[35]
Semmartin M Ghersa CM 2006 ). Intraspecific changes in plant morphology, associated with grazing, and effects on litter quality, carbon and nutrient dynamics during decomposition.
Austral Ecology , 31 (1 ), 99 -105 .
[本文引用: 1]
[36]
Shariff AR Grygiel CE 1994 ). Grazing intensity effects on litter decomposition and soil nitrogen mineralization.
Journal of Range Management , 47 , 444 -449 .
[37]
Shi FS Chen H Wu Y Wu N 2010 ). Effects of livestock exclusion on vegetation and soil properties under two topographic habitats in an alpine meadow on the eastern Qinghai-Tibetan Plateau.
Polish Journal of Ecology , 58 , 125 -133 .
[本文引用: 1]
[38]
Smith SW Woodin SJ Pakeman RJ David J René VDW 2014 ). Root traits predict decomposition across a landscape-scale grazing experiment.
New Phytologist , 203 , 851 -862 .
[本文引用: 1]
[39]
Solly EF Schöning I Boch S Kandeler E Marhan S Michalzik B Müller J Zscheischler J Trumbore SE Schrumpf M 2014 ). Factors controlling decomposition rates of fine root litter in temperate forests and grasslands.
Plant & Soil , 382 , 203 -218 .
[本文引用: 1]
[40]
Stark S Männistö MK Ganzert L Tiirola M Häggblom MM 2015 ). Grazing intensity in subarctic tundra affects the temperature adaptation of soil microbial communities.
Soil Biology & Biochemistry , 84 , 147 -157 .
[本文引用: 1]
[41]
Tateno R Tokuchi N Yamanaka N Du S Otsuki K Shimamura T Xue Z Wang SQ Hou QC 2007 ). Comparison of litterfall production and leaf litter decomposition between an exotic black locust plantation and an indigenous oak forest near Yan’an on the Loess Plateau, China.
Forest Ecology & Management , 241 (s1-3 ), 84 -90 .
[本文引用: 1]
[42]
Tessier M Vivier JP Ouin A Gloaguen JC Lefeuvre JC 2003 ). Vegetation dynamics and plant species interactions under grazed and ungrazed conditions in a western European salt marsh.
Acta Oecologica , 24 , 103 -111 .
[本文引用: 1]
[43]
van Soest PJ 1963 ). Use of detergents in analysis of fibrous feeds: A rapid method for the determination of fiber and lignin.
Journal of the Association of Official Analytical Chemists , 46 , 829 -835 .
[本文引用: 1]
[44]
van Soest PJ 1967 ). Development of a comprehensive system of feed analyses and its application to forages.
Journal of Animal Science , 26 , 119 -128 .
[本文引用: 1]
[45]
Wang L Zhang Y Xu DM Zhang N 2013 ). Study on litter decomposition rates and N, P, K content of litter in different years of enclosure in desert steppe.
Pratacultural Science , 30 , 1508 -1512 . (in Chinese with English abstract)[王蕾 , 张宇 , 许冬梅 , 张娜 (2013 ). 围封对草地凋落物分解速率和N、P、K含量的影响
. 草业科学 , 30 , 1508 -1512 .]
[本文引用: 1]
[46]
Wang MJ Han GD Zhao ML Chen HJ Wang Z Hao XL Bo T 2007 ). The effects of different grazing intensity on soil organic carbon content in meadow steppe.
Pratacultural Science , 10 , 6 -10 . (in Chinese with English abstract)[王明君 , 韩国栋 , 赵萌莉 , 陈海军 , 王珍 , 郝晓莉 , 薄涛 (2007 ). 草甸草原不同放牧强度对土壤有机碳含量的影响
. 草业科学 , 10 , 6 -10 .]
[本文引用: 1]
[47]
Wang YH Gong JR Liu M Luo QP Xu S Pan Y Zhai ZW 2015 ). Effects of land use and precipitation on above- and below-ground litter decomposition in a semi-arid temperate steppe in Inner Mongolia, China.
Applied Soil Ecology , 96 , 183 -191 .
[48]
Wardle DA Bardgett RD Klironomos JN Heikki S Wim H Wall DH 2004 ). Ecological linkages between aboveground and belowground biota.
Science , 304 , 1629 -1633 .
[49]
Wen HY Zhao HL Fu H 2005 ). Effects of years for reclamation and enclosure years on soil properties of degraded sandy grassland.
Scientia Agricultura Sinica , 14 (1 ), 31 -37 . (in Chinese with English abstract)[文海燕 , 赵哈林 , 傅华 (2005 ). 开垦和封育年限对退化沙质草地土壤性状的影响
. 草业学报 , 14 (1 ), 31 -37 .]
[本文引用: 1]
[50]
Xu DD Guo XL 2015 ). Evaluating the impacts of nearly 30 years of conservation on grassland ecosystem using Landsat TM images.
Grassland Science , 61 , 227 -242 .
[51]
Yang JC Han XG Huang JH Pan QM 2003 ). Effects of land use change on carbon storage in terrestrial ecosystem.
Chinese Journal of Applied Ecology , 14 , 1385 -1390 . (in Chinese with English abstract)[杨景成 , 韩兴国 , 黄建辉 , 潘庆民 (2003 ). 土地利用变化对陆地生态系统碳贮量的影响
. 应用生态学报 , 14 , 1385 -1390 .]
[52]
Yeo JJ 2005 ). Effects of grazing exclusion on rangeland vegetation and soils, East Central Idaho.
Western North American Naturalist , 65 , 91 -102 .
[本文引用: 1]
[53]
Zhang CX Nan ZB 2010 ). Research progress OR effects of grazing on physical and chemical characteristics of grassland soil.
Scientia Agricultura Sinica , 19 (4 ), 204 -211 . (in Chinese with English abstract)[张成霞 , 南志标 (2010 ). 放牧对草地土壤理化特性影响的研究进展
. 草业学报 , 19 (4 ), 204 -211 .]
[本文引用: 1]
[54]
Zhang D Hui D Luo Y Zhou G 2008 ). Rates of litter decom- position in terrestrial ecosystems: Global patterns and controlling factors.
Journal of Plant Ecology , 1 (2 ), 85 -93 .
[55]
Zhang K Cheng X Dang H Ye C Zhang YL Zhang QF 2013 ). Linking litter production, quality and decomposition to vegetation succession following agricultural abandonment.
Soil Biology & Biochemistry , 57 , 803 -813 .
[本文引用: 1]
[56]
Zhang YB Luo P Sun G Mou CX Wang ZY Wu N Luo GR 2012 ). Effects of grazing on litter decomposition in two alpine meadow on the eastern Qinghai-Tibet Plateau.
Acta Ecologica Sinica , 32 , 4605 -4617 . (in Chinese with English abstract)[张艳博 , 罗鹏 , 孙庚 , 牟成香 , 王志远 , 吴宁 , 罗光荣 (2012 ). 放牧对青藏高原东部两种典型高寒草地类型凋落物分解的影响
. 生态学报 , 32 , 4605 -4617 .]
[本文引用: 1]
[57]
Zuo WQ Wang YH Wang FY Shi GX 2009 ). Effects of enclosure on the community characteristics of Leymus chinensis in degenerated steppe.
Scientia Agricultura Sinica , 18 (3 ), 12 -19 . (in Chinese with English abstract)[左万庆 , 王玉辉 , 王风玉 , 师广旭 (2009 ). 围栏封育措施对退化羊草草原植物群落特征影响研究
. 草业学报 , 18 (3 ), 12 -19 .]
Impact of grazing abandonment on plant and soil microbial communities in an Atlantic mountain grassland.
1
2015
... 围栏封育是草地有效管理方式之一, 它对植被资源恢复、土壤理化性质恢复和生物多样性保护具有重要意义.围封后植被生长状况、土壤无脊椎动物和微生物活性以及凋落物分解的关键过程都会发生改变(Lindsay & Cunningham, 2009 ).本研究中, 两年内放牧样地地上凋落物产量在围封后均有显著提高.由于围栏封育后排除人为和牲畜干扰, 植被得到恢复, 地上生物量、凋落物积累和土壤有机质输入显著提高(Tessier et al ., 2003 ).围封后除了植被得到恢复外, 土壤条件也得到明显改善.放牧样地经过围封后, 土壤容重降低, 养分含量增加, 土壤酶活性增强, 这些都可以促进围封样地地上凋落物形成(文海燕等, 2005 ).在本研究两年围封期间, 轻度放牧和轻度放牧后围封样地凋落物质量和土壤养分之间的差异并不显著, 但轻度放牧后围封样地地上凋落物分解速率加快, 可能是土壤物理和生物特性发生变化的结果(Mekuria et al ., 2007 ).与之相反, 重度放牧围封后地上凋落物分解速率减慢, 凋落物质量变差, 说明重度放牧围封后植被组成发生了变化, 植物叶片结构性碳(木质素)合成增加, 导致地上凋落物木质素含量和木质素:N升高, 凋落物中难分解物质比例增加(Bontti et al ., 2009 ).围封后排除了牲畜的排泄作用, 土壤有机质含量减少, 土壤酶活性和微生物活性降低, 这都将导致重度放牧样地围封后地上凋落物分解减慢(Aldezabal et al ., 2015 ).由此可以推测, 长期轻度放牧样地生态系统的恢复作用更强, 短期围封后地上凋落物分解速率和养分循环加快. ...
Differential effects of grazing on plant functional traits in the desert grassland.
1
2014
... 放牧牲畜通过采食、践踏和排泄作用影响凋落物形成和分解.放牧对凋落物的影响主要表现在两个方面: 一是放牧使植物物种组成、牧草本身的形态学特征(如根冠比、叶片/叶鞘等)发生改变, 从而影响凋落物起始质量(Holland & Detling, 1990 ; Semmartin & Ghersa, 2006 ); 二是放牧使凋落物分解的立地土壤物理和生物环境发生改变, 直接影响凋落物分解(Shariff & Grygiel, 1994 ).本研究中, 重度放牧样地地上凋落物产量显著高于轻度放牧, 因为放牧可以削减草地群落的物种冗余程度, 牲畜唾液可以引入生长刺激物, 使植物产生补偿效应(Gong et al ., 2015 ); 两年中重度放牧比轻度放牧更能促进地上凋落物分解, 重度放牧样地地上凋落物质量好, C、N含量高、木质素:N低, 加上牲畜的踩踏作用加快凋落物的碎裂并与土壤紧密接触和混合.同时牲畜啃食会使植被盖度减少, 地表太阳辐射增多, 表层土壤温度和微生物活性提高, 都可以促进地上凋落物分解(张成霞和南志标, 2010 ; Hewins et al ., 2013 ).另外, 长期重度放牧使叶片有变小的趋势, 这是植物在功能属性上忍受放牧和加快更新的措施, 这些变化使其凋落后的分解更加容易(Cingolani et al ., 2005 ; An & Li, 2014 ).综上所述, 重度放牧加快了地上凋落物的产生和分解, 有利于凋落物中碳氮参与草地生态系统中的碳氮循环, 对维持草地生态系统中碳氮平衡具有重要意义.然而, 重度放牧可能会使土壤物理性质变差, 这将造成重度放牧对草地生态系统物质循环的整体效应降低, 不利于草地的健康发展.因此, 合理放牧才是保证草地健康发展的管理方式. ...
Precipitation, decomposition, and litter decomposability of Metrosideros polymorpha on Hawaii.
1
2001
... 除放牧强度和围栏封育外, 降水变化对凋落物分解也有显著影响.降水通过影响土壤温湿度, 改变植物群落组成和功能, 从而影响凋落物质量和土壤微生物活性, 调控凋落物形成和分解过程(Austin & Vitousek, 2001 ).本研究中, 湿润年地上、地下凋落物产量和分解速率均高于干旱年.干旱年植物生长受水分限制, 导致气孔关闭、光合能力下降, 植物生产力降低(Sanaullah et al ., 2014 ), 产生的凋落物也较湿润年少.在轻度放牧和轻度放牧后围封样地, 湿润年土壤全C含量是干旱年的近2倍.湿润年降水增多可提高土壤含水量和酶活性(Mikan et al ., 2002 )、加快凋落物化学组分的淋溶, 从地表输入土壤的C增多, 根系的分解也随之加快, 土壤C输入显著提高.并且土壤可利用水增多可加快土壤中的养分向微生物种群扩散, 使微生物活性增强(Wang et al ., 2015 ).这都可以显著促进凋落物分解过程, 加快草地生态系统的碳循环和养分循环.另外, 在草地生态系统中, 由于植物根生物量比其他生态系统大, 较快的根系分解速率和周转速率有助于维持土壤养分的供应和植物的生长(Liu et al ., 2009 ).因此降水还可以通过促进地下凋落物分解而增加土壤养分, 进而促进植物地上部分生长和凋落物分解过程. ...
Heavily intensified grazing reduces root production in an Inner Mongolia temperate steppe.
1
2015
... 植物分配一定的光合物质到地下以维持根系的产量, 同时根系吸收土壤中的水分和养分供给地上部分生长.因此植物地上和根系的生长紧密相连, 地上和地下凋落物的产生和分解也是相互影响的(Bai et al ., 2015 ).植物根系是土壤有机质的主要来源, 放牧主要通过影响土壤微环境(温度和湿度)和根系特征来影响地下凋落物产生和分解, 进而影响草地生态系统土壤养分循环(Smith et al ., 2014 ).本研究中, 干旱年重度放牧样地地下凋落物产量显著低于轻度放牧样地, 湿润年则相反.这可能是由于重度放牧样地家畜的践踏作用使土壤孔隙度和水稳性团聚体减少, 引起土壤透水性、透气性和水导率下降, 加上在干旱年水分亏缺严重, 严重阻碍根系的生长(侯扶江等, 2004 ).而湿润年降水充足, 土壤理化性质变好, 适宜根系生长, 重度放牧样地地下凋落物产量迅速增加, 显著高于轻度放牧样地.另外在本研究中, 干旱年重度放牧地下凋落物分解速率显著低于轻度放牧, 湿润年则相反.在干旱年, 重度放牧使植被盖度显著下降, 导致地表裸露、土温上升、土壤水分散失加剧、土壤物理性质变差; 另一方面, 土壤可利用性水分减少, 加上内蒙古草原的高海拔使重度放牧样地裸露的地表UV-B辐射增多, 大幅降低土壤分解者的活性, 这些都会严重阻碍重度放牧样地地下凋落物的分解(Dormaar & Willms, 1990 ; Giese et al ., 2009 ).但在湿润年, 重度放牧地下凋落物质量变好, 低C:N和木质素:N使重度放牧样地地下分解速率显著高于轻度放牧, 这也说明重度放牧样地地下凋落物分解对降水条件的响应十分敏感.因此我们的实验结果显示放牧对半干旱草地生态系统凋落物产生和分解的影响主要取决于环境的降水条件. ...
Hierarchical responses of plant-soil interactions to climate change: Consequences for the global carbon cycle.
1
2013
... 随着全球气候变化加剧, 碳循环直接影响着人类的生存和各种生物环境的稳定性, 逐渐成为生态系统研究的重点(杨景成等, 2003 ; Bardgett et al ., 2013 ).陆地生态系统在全球碳循环中起着重要的作用, 因此对陆地生态系统碳循环的理解非常必要(Canadell et al ., 2000 ).我国现有草地面积约4亿hm2 , 是最大的陆地生态系统, 也是生物地球循环的重要组成部分, 但其容易受到气候变化和各种人类活动(如: 放牧、施肥等)的影响(齐玉春等, 2003 ; Xu & Guo, 2015 ).目前, 由于放牧和农垦活动频繁, 草地生态系统成为受人类活动影响较为严重的区域.因此, 对自然和人为因素干扰下草地生态系统碳循环过程的研究具有十分重要的科学意义. ...
Litter decomposition in grasslands of Central North America (US Great Plains).
3
2009
... 凋落物基质质量能够表征凋落物在自然界中的可分解性.衡量凋落物质量的指标主要是C含量、N含量、木质素浓度、C:N、木质素:N等.一般来说, 凋落物中N含量越高、C:N越小、木质素含量越低, 凋落物分解速率越快(Koukoura et al ., 2003 ).在干旱和半干旱草地生态系统中, 降水是影响凋落物分解的重要气候因子, 它可以通过影响植物群落组成和分解者的活性来影响凋落物质量, 进而影响凋落物分解.但降水和凋落物质量可能存在交互作用, 使得我们无法准确评估某一因子对分解的影响(Bontti et al ., 2009 ).本研究在内蒙古温带草原上开展, 2011年和2012年该区域的降水量分别是188 mm和465 mm, 存在明显差异, 使得本文恰好能够探究降水差异对草地凋落物分解的影响.本文在内蒙古温带典型草原中测定了干湿年份内4种样地(轻度放牧样地、轻度放牧后围封样地、重度放牧样地、重度放牧后围封样地)的土壤养分含量、凋落物产量、质量及分解速率来探讨不同放牧强度对凋落物形成和分解的影响, 以及围栏封育对生态系统恢复的作用, 具体研究目标如下: 1)分析不同放牧强度和围栏封育下凋落物分解的差异; 2)探讨调控地上、地下凋落物分解的主要因子; 3)揭示降水变化对凋落物产量和分解的作用.通过讨论分析, 以期为草地生态系统和草地畜牧业的可持续发展提供理论依据. ...
... 围栏封育是草地有效管理方式之一, 它对植被资源恢复、土壤理化性质恢复和生物多样性保护具有重要意义.围封后植被生长状况、土壤无脊椎动物和微生物活性以及凋落物分解的关键过程都会发生改变(Lindsay & Cunningham, 2009 ).本研究中, 两年内放牧样地地上凋落物产量在围封后均有显著提高.由于围栏封育后排除人为和牲畜干扰, 植被得到恢复, 地上生物量、凋落物积累和土壤有机质输入显著提高(Tessier et al ., 2003 ).围封后除了植被得到恢复外, 土壤条件也得到明显改善.放牧样地经过围封后, 土壤容重降低, 养分含量增加, 土壤酶活性增强, 这些都可以促进围封样地地上凋落物形成(文海燕等, 2005 ).在本研究两年围封期间, 轻度放牧和轻度放牧后围封样地凋落物质量和土壤养分之间的差异并不显著, 但轻度放牧后围封样地地上凋落物分解速率加快, 可能是土壤物理和生物特性发生变化的结果(Mekuria et al ., 2007 ).与之相反, 重度放牧围封后地上凋落物分解速率减慢, 凋落物质量变差, 说明重度放牧围封后植被组成发生了变化, 植物叶片结构性碳(木质素)合成增加, 导致地上凋落物木质素含量和木质素:N升高, 凋落物中难分解物质比例增加(Bontti et al ., 2009 ).围封后排除了牲畜的排泄作用, 土壤有机质含量减少, 土壤酶活性和微生物活性降低, 这都将导致重度放牧样地围封后地上凋落物分解减慢(Aldezabal et al ., 2015 ).由此可以推测, 长期轻度放牧样地生态系统的恢复作用更强, 短期围封后地上凋落物分解速率和养分循环加快. ...
... 在本研究中, 短期围封后地下凋落物产量反而显著低于对应的放牧样地.放牧活动能促使根系向土壤上层集中, 且它对根系的生长有促进作用, 因此围封后根系生长变缓(Greenwood & Hutchinson, 2003 ; Pucheta et al ., 2004 ).但只有在干旱年, 重度放牧后围封样地地下凋落物产量显著高于重度放牧.这可能是由于重度放牧样地牲畜的踩踏作用使土壤遭到破坏, 加上干旱年降水稀少, 限制了地下根系生长.在干旱年, 放牧和围封样地地下凋落物分解速率存在显著差异主要是凋落物质量差异所致.轻度放牧样地围封后, 地下凋落物质量变差, 高木质素含量和木质素:N使得轻度放牧后围封样地地下凋落物分解速率显著低于轻度放牧样地; 而重度放牧样地相反, 围封后凋落物质量和土壤养分状况变好(图3 , 图5 ), 使得重度放牧样地围封后分解速率显著高于重度放牧样地.湿润年各样地的地下凋落物质量和土壤养分之间虽然存在差异, 但各样地地下凋落物分解速率并无显著差异.相比于凋落物质量和土壤养分的作用, 降水对地下凋落物分解过程的调控作用更强(Bontti et al ., 2009 ; Giese et al ., 2009 ).综上所述, 本研究表明短期围栏封育能使植被生长环境、土壤理化性质得到明显改善, 轻度放牧围封后还能提高凋落物分解速率和养分循环.不过有研究表明, 长期围封会使草地非禾本科牧草比例大幅增加, 不利于禾本科牧草产量的积累, 这将会阻碍畜牧业发展, 造成严重的经济损失(Shi et al ., 2010 ).因此, 在草地放牧强度和围封年限的确定上, 我们应注意寻找草地经济效益和生态系统保护之间的平衡点. ...
Determination of nitrogen in soil by the Kjeldahl method.
1
1960
... 将2011年和2012年每次洗净烘干的凋落物样品进行粉碎研磨, 再通过40目网筛.一部分用于凋落物C、N含量的测定, 另一部分用于木质素和纤维素含量的测定.全C含量的测定采用重铬酸钾法(Nelson & Sommers, 1982 ), 全N含量的测定采用凯氏定氮法(Bremner, 1960 ).木质素和纤维素的测定依据van Soest (1963, 1967)的实验原理, 参照AOAC international (2000)的具体实验操作标准, 经过消煮-酸洗-灼烧等步骤, 测定各样地初始凋落物和残留凋落物中的木质素和纤维素的含量. ...
Carbon metabolism of the terrestrial biosphere: A multitechnique approach for improved understanding.
1
2000
... 随着全球气候变化加剧, 碳循环直接影响着人类的生存和各种生物环境的稳定性, 逐渐成为生态系统研究的重点(杨景成等, 2003 ; Bardgett et al ., 2013 ).陆地生态系统在全球碳循环中起着重要的作用, 因此对陆地生态系统碳循环的理解非常必要(Canadell et al ., 2000 ).我国现有草地面积约4亿hm2 , 是最大的陆地生态系统, 也是生物地球循环的重要组成部分, 但其容易受到气候变化和各种人类活动(如: 放牧、施肥等)的影响(齐玉春等, 2003 ; Xu & Guo, 2015 ).目前, 由于放牧和农垦活动频繁, 草地生态系统成为受人类活动影响较为严重的区域.因此, 对自然和人为因素干扰下草地生态系统碳循环过程的研究具有十分重要的科学意义. ...
Plant functional traits, herbivore selectivity and response to sheep grazing in Patagonian steppe grasslands.
1
2005
... 放牧牲畜通过采食、践踏和排泄作用影响凋落物形成和分解.放牧对凋落物的影响主要表现在两个方面: 一是放牧使植物物种组成、牧草本身的形态学特征(如根冠比、叶片/叶鞘等)发生改变, 从而影响凋落物起始质量(Holland & Detling, 1990 ; Semmartin & Ghersa, 2006 ); 二是放牧使凋落物分解的立地土壤物理和生物环境发生改变, 直接影响凋落物分解(Shariff & Grygiel, 1994 ).本研究中, 重度放牧样地地上凋落物产量显著高于轻度放牧, 因为放牧可以削减草地群落的物种冗余程度, 牲畜唾液可以引入生长刺激物, 使植物产生补偿效应(Gong et al ., 2015 ); 两年中重度放牧比轻度放牧更能促进地上凋落物分解, 重度放牧样地地上凋落物质量好, C、N含量高、木质素:N低, 加上牲畜的踩踏作用加快凋落物的碎裂并与土壤紧密接触和混合.同时牲畜啃食会使植被盖度减少, 地表太阳辐射增多, 表层土壤温度和微生物活性提高, 都可以促进地上凋落物分解(张成霞和南志标, 2010 ; Hewins et al ., 2013 ).另外, 长期重度放牧使叶片有变小的趋势, 这是植物在功能属性上忍受放牧和加快更新的措施, 这些变化使其凋落后的分解更加容易(Cingolani et al ., 2005 ; An & Li, 2014 ).综上所述, 重度放牧加快了地上凋落物的产生和分解, 有利于凋落物中碳氮参与草地生态系统中的碳氮循环, 对维持草地生态系统中碳氮平衡具有重要意义.然而, 重度放牧可能会使土壤物理性质变差, 这将造成重度放牧对草地生态系统物质循环的整体效应降低, 不利于草地的健康发展.因此, 合理放牧才是保证草地健康发展的管理方式. ...
Effect of grazing and cultiva- tion on some chemical properties of soils in the mixed prairie.
1
1990
... 植物分配一定的光合物质到地下以维持根系的产量, 同时根系吸收土壤中的水分和养分供给地上部分生长.因此植物地上和根系的生长紧密相连, 地上和地下凋落物的产生和分解也是相互影响的(Bai et al ., 2015 ).植物根系是土壤有机质的主要来源, 放牧主要通过影响土壤微环境(温度和湿度)和根系特征来影响地下凋落物产生和分解, 进而影响草地生态系统土壤养分循环(Smith et al ., 2014 ).本研究中, 干旱年重度放牧样地地下凋落物产量显著低于轻度放牧样地, 湿润年则相反.这可能是由于重度放牧样地家畜的践踏作用使土壤孔隙度和水稳性团聚体减少, 引起土壤透水性、透气性和水导率下降, 加上在干旱年水分亏缺严重, 严重阻碍根系的生长(侯扶江等, 2004 ).而湿润年降水充足, 土壤理化性质变好, 适宜根系生长, 重度放牧样地地下凋落物产量迅速增加, 显著高于轻度放牧样地.另外在本研究中, 干旱年重度放牧地下凋落物分解速率显著低于轻度放牧, 湿润年则相反.在干旱年, 重度放牧使植被盖度显著下降, 导致地表裸露、土温上升、土壤水分散失加剧、土壤物理性质变差; 另一方面, 土壤可利用性水分减少, 加上内蒙古草原的高海拔使重度放牧样地裸露的地表UV-B辐射增多, 大幅降低土壤分解者的活性, 这些都会严重阻碍重度放牧样地地下凋落物的分解(Dormaar & Willms, 1990 ; Giese et al ., 2009 ).但在湿润年, 重度放牧地下凋落物质量变好, 低C:N和木质素:N使重度放牧样地地下分解速率显著高于轻度放牧, 这也说明重度放牧样地地下凋落物分解对降水条件的响应十分敏感.因此我们的实验结果显示放牧对半干旱草地生态系统凋落物产生和分解的影响主要取决于环境的降水条件. ...
Soil surface characteristics, microtopography and proximity to mature shrubs: Effects on survival of several cohorts of Atriplex vesicaria seedlings.
1
1992
... 放牧和围栏封育是草地最基本的两种管理方式, 它们直接改变地表的植被特征, 也改变土壤环境(如温度、含水量等)及其养分含量, 使植物碳分配模式和土壤微生物代谢过程等发生改变, 进而影响凋落物分解和生态系统碳循环(Raich & Tufekciogiu, 2000 ).放牧是草地生态系统生物地球化学循环的重要驱动因子, 不同放牧强度下, 牲畜通过采食、排泄和践踏等方式影响植物群落结构和土壤特性, 使凋落物形成和分解发生变化(Stark et al ., 2015 ).但是目前关于放牧强度对地下凋落物分解的影响研究不足.围栏封育是保持草地生态系统功能的有效管理方式.放牧草地经过围栏封育后, 使草地植被得到恢复, 土壤特性、碳输入、植物组成以及凋落物基质质量发生了变化, 这些因素都影响了草地凋落物的形成和分解(王蕾等, 2013 ; Lu et al ., 2015 ).目前, 国内外学者对草地围栏封育的研究仅局限在不同年限的围封对群落结构和凋落物的影响.研究表明, 围封时间过长会引起群落地上生物量降低、凋落物堆积, 抑制植物的再生, 一部分优势种开始衰退, 阻碍草地资源的更新利用(Eldridge et al ., 1992 ; 左万庆等, 2009 ), 因此研究放牧后的短期围封对草地恢复更具指导意义.目前, 草地不同退化强度基础上的短期围封研究尚且不足(Yeo, 2005 ; Hu et al ., 2016 ).因此, 我们研究不同放牧强度下的短期围栏封育对凋落物分解和草地生产力恢复的影响, 以期为防止草地退化、改善生态环境提供参考依据. ...
Linking litter decomposition of above- and below-ground organs to plant-soil feedbacks worldwide.
1
2013
... 草地凋落物是指由植物地上部分和地下根系形成并归还到土壤的所有有机质的总称, 是陆地生态系统养分循环和碳循环过程中极为重要的环节(高永恒等, 2007 ).陆地生态系统中约90%的净初级生产量以凋落物的形式归还给土壤, 植物的生长发育在很大程度上依赖于植被归还给土壤的养分再循环(Wardle et al ., 2004 ).凋落物的分解是一个非常复杂的过程, 深受气候、凋落物质量和土壤生物的影响, 在很大程度上决定了生态系统的固碳速率及动态(Tateno et al ., 2007 ).因此, 在全球变化背景下, 对草地生态系统凋落物分解的研究是草地生态系统动态机理以及全球变化生态学研究的重要内容(张艳博等, 2012 ).另外, 地下凋落物的碳输入是地上部分凋落物碳输入的3倍以上, 但是过去仅有2%的植物凋落物研究关注于地下凋落物分解, 很少有研究对干旱、半干旱草原的地上和地下凋落物同时进行系统研究(Zhang et al ., 2008 ; Freschet et al ., 2013 ).因此研究不同干扰下草地地下凋落物分解的变化, 对准确估算全球变化背景下不同尺度碳收支至关重要(Solly et al ., 2014 ). ...
Effects of grazing and rainfall variability on root and shoot decomposition in a semi-arid grassland.
2
2009
... 植物分配一定的光合物质到地下以维持根系的产量, 同时根系吸收土壤中的水分和养分供给地上部分生长.因此植物地上和根系的生长紧密相连, 地上和地下凋落物的产生和分解也是相互影响的(Bai et al ., 2015 ).植物根系是土壤有机质的主要来源, 放牧主要通过影响土壤微环境(温度和湿度)和根系特征来影响地下凋落物产生和分解, 进而影响草地生态系统土壤养分循环(Smith et al ., 2014 ).本研究中, 干旱年重度放牧样地地下凋落物产量显著低于轻度放牧样地, 湿润年则相反.这可能是由于重度放牧样地家畜的践踏作用使土壤孔隙度和水稳性团聚体减少, 引起土壤透水性、透气性和水导率下降, 加上在干旱年水分亏缺严重, 严重阻碍根系的生长(侯扶江等, 2004 ).而湿润年降水充足, 土壤理化性质变好, 适宜根系生长, 重度放牧样地地下凋落物产量迅速增加, 显著高于轻度放牧样地.另外在本研究中, 干旱年重度放牧地下凋落物分解速率显著低于轻度放牧, 湿润年则相反.在干旱年, 重度放牧使植被盖度显著下降, 导致地表裸露、土温上升、土壤水分散失加剧、土壤物理性质变差; 另一方面, 土壤可利用性水分减少, 加上内蒙古草原的高海拔使重度放牧样地裸露的地表UV-B辐射增多, 大幅降低土壤分解者的活性, 这些都会严重阻碍重度放牧样地地下凋落物的分解(Dormaar & Willms, 1990 ; Giese et al ., 2009 ).但在湿润年, 重度放牧地下凋落物质量变好, 低C:N和木质素:N使重度放牧样地地下分解速率显著高于轻度放牧, 这也说明重度放牧样地地下凋落物分解对降水条件的响应十分敏感.因此我们的实验结果显示放牧对半干旱草地生态系统凋落物产生和分解的影响主要取决于环境的降水条件. ...
... 在本研究中, 短期围封后地下凋落物产量反而显著低于对应的放牧样地.放牧活动能促使根系向土壤上层集中, 且它对根系的生长有促进作用, 因此围封后根系生长变缓(Greenwood & Hutchinson, 2003 ; Pucheta et al ., 2004 ).但只有在干旱年, 重度放牧后围封样地地下凋落物产量显著高于重度放牧.这可能是由于重度放牧样地牲畜的踩踏作用使土壤遭到破坏, 加上干旱年降水稀少, 限制了地下根系生长.在干旱年, 放牧和围封样地地下凋落物分解速率存在显著差异主要是凋落物质量差异所致.轻度放牧样地围封后, 地下凋落物质量变差, 高木质素含量和木质素:N使得轻度放牧后围封样地地下凋落物分解速率显著低于轻度放牧样地; 而重度放牧样地相反, 围封后凋落物质量和土壤养分状况变好(图3 , 图5 ), 使得重度放牧样地围封后分解速率显著高于重度放牧样地.湿润年各样地的地下凋落物质量和土壤养分之间虽然存在差异, 但各样地地下凋落物分解速率并无显著差异.相比于凋落物质量和土壤养分的作用, 降水对地下凋落物分解过程的调控作用更强(Bontti et al ., 2009 ; Giese et al ., 2009 ).综上所述, 本研究表明短期围栏封育能使植被生长环境、土壤理化性质得到明显改善, 轻度放牧围封后还能提高凋落物分解速率和养分循环.不过有研究表明, 长期围封会使草地非禾本科牧草比例大幅增加, 不利于禾本科牧草产量的积累, 这将会阻碍畜牧业发展, 造成严重的经济损失(Shi et al ., 2010 ).因此, 在草地放牧强度和围封年限的确定上, 我们应注意寻找草地经济效益和生态系统保护之间的平衡点. ...
放牧强度对川西北高山草甸两个优势物种凋落物分解的影响
2007
Re- sponse of primary production and biomass allocation to nitrogen and water supplementation along a grazing intensity gradient in semiarid grassland.
1
2015
... 草地凋落物是指由植物地上部分和地下根系形成并归还到土壤的所有有机质的总称, 是陆地生态系统养分循环和碳循环过程中极为重要的环节(高永恒等, 2007 ).陆地生态系统中约90%的净初级生产量以凋落物的形式归还给土壤, 植物的生长发育在很大程度上依赖于植被归还给土壤的养分再循环(Wardle et al ., 2004 ).凋落物的分解是一个非常复杂的过程, 深受气候、凋落物质量和土壤生物的影响, 在很大程度上决定了生态系统的固碳速率及动态(Tateno et al ., 2007 ).因此, 在全球变化背景下, 对草地生态系统凋落物分解的研究是草地生态系统动态机理以及全球变化生态学研究的重要内容(张艳博等, 2012 ).另外, 地下凋落物的碳输入是地上部分凋落物碳输入的3倍以上, 但是过去仅有2%的植物凋落物研究关注于地下凋落物分解, 很少有研究对干旱、半干旱草原的地上和地下凋落物同时进行系统研究(Zhang et al ., 2008 ; Freschet et al ., 2013 ).因此研究不同干扰下草地地下凋落物分解的变化, 对准确估算全球变化背景下不同尺度碳收支至关重要(Solly et al ., 2014 ). ...
Root characteristics of temperate pasture in New South Wales after grazing at three stocking rates for 30 years.
1
2003
... 放牧牲畜通过采食、践踏和排泄作用影响凋落物形成和分解.放牧对凋落物的影响主要表现在两个方面: 一是放牧使植物物种组成、牧草本身的形态学特征(如根冠比、叶片/叶鞘等)发生改变, 从而影响凋落物起始质量(Holland & Detling, 1990 ; Semmartin & Ghersa, 2006 ); 二是放牧使凋落物分解的立地土壤物理和生物环境发生改变, 直接影响凋落物分解(Shariff & Grygiel, 1994 ).本研究中, 重度放牧样地地上凋落物产量显著高于轻度放牧, 因为放牧可以削减草地群落的物种冗余程度, 牲畜唾液可以引入生长刺激物, 使植物产生补偿效应(Gong et al ., 2015 ); 两年中重度放牧比轻度放牧更能促进地上凋落物分解, 重度放牧样地地上凋落物质量好, C、N含量高、木质素:N低, 加上牲畜的踩踏作用加快凋落物的碎裂并与土壤紧密接触和混合.同时牲畜啃食会使植被盖度减少, 地表太阳辐射增多, 表层土壤温度和微生物活性提高, 都可以促进地上凋落物分解(张成霞和南志标, 2010 ; Hewins et al ., 2013 ).另外, 长期重度放牧使叶片有变小的趋势, 这是植物在功能属性上忍受放牧和加快更新的措施, 这些变化使其凋落后的分解更加容易(Cingolani et al ., 2005 ; An & Li, 2014 ).综上所述, 重度放牧加快了地上凋落物的产生和分解, 有利于凋落物中碳氮参与草地生态系统中的碳氮循环, 对维持草地生态系统中碳氮平衡具有重要意义.然而, 重度放牧可能会使土壤物理性质变差, 这将造成重度放牧对草地生态系统物质循环的整体效应降低, 不利于草地的健康发展.因此, 合理放牧才是保证草地健康发展的管理方式. ...
Soil-Litter mixing accelerates decomposition in a Chihuahuan Desert Grassland.
1
2013
... 在本研究中, 短期围封后地下凋落物产量反而显著低于对应的放牧样地.放牧活动能促使根系向土壤上层集中, 且它对根系的生长有促进作用, 因此围封后根系生长变缓(Greenwood & Hutchinson, 2003 ; Pucheta et al ., 2004 ).但只有在干旱年, 重度放牧后围封样地地下凋落物产量显著高于重度放牧.这可能是由于重度放牧样地牲畜的踩踏作用使土壤遭到破坏, 加上干旱年降水稀少, 限制了地下根系生长.在干旱年, 放牧和围封样地地下凋落物分解速率存在显著差异主要是凋落物质量差异所致.轻度放牧样地围封后, 地下凋落物质量变差, 高木质素含量和木质素:N使得轻度放牧后围封样地地下凋落物分解速率显著低于轻度放牧样地; 而重度放牧样地相反, 围封后凋落物质量和土壤养分状况变好(图3 , 图5 ), 使得重度放牧样地围封后分解速率显著高于重度放牧样地.湿润年各样地的地下凋落物质量和土壤养分之间虽然存在差异, 但各样地地下凋落物分解速率并无显著差异.相比于凋落物质量和土壤养分的作用, 降水对地下凋落物分解过程的调控作用更强(Bontti et al ., 2009 ; Giese et al ., 2009 ).综上所述, 本研究表明短期围栏封育能使植被生长环境、土壤理化性质得到明显改善, 轻度放牧围封后还能提高凋落物分解速率和养分循环.不过有研究表明, 长期围封会使草地非禾本科牧草比例大幅增加, 不利于禾本科牧草产量的积累, 这将会阻碍畜牧业发展, 造成严重的经济损失(Shi et al ., 2010 ).因此, 在草地放牧强度和围封年限的确定上, 我们应注意寻找草地经济效益和生态系统保护之间的平衡点. ...
Plant response to herbivory and belowground nitrogen cycling.
1
1990
... 放牧牲畜通过采食、践踏和排泄作用影响凋落物形成和分解.放牧对凋落物的影响主要表现在两个方面: 一是放牧使植物物种组成、牧草本身的形态学特征(如根冠比、叶片/叶鞘等)发生改变, 从而影响凋落物起始质量(Holland & Detling, 1990 ; Semmartin & Ghersa, 2006 ); 二是放牧使凋落物分解的立地土壤物理和生物环境发生改变, 直接影响凋落物分解(Shariff & Grygiel, 1994 ).本研究中, 重度放牧样地地上凋落物产量显著高于轻度放牧, 因为放牧可以削减草地群落的物种冗余程度, 牲畜唾液可以引入生长刺激物, 使植物产生补偿效应(Gong et al ., 2015 ); 两年中重度放牧比轻度放牧更能促进地上凋落物分解, 重度放牧样地地上凋落物质量好, C、N含量高、木质素:N低, 加上牲畜的踩踏作用加快凋落物的碎裂并与土壤紧密接触和混合.同时牲畜啃食会使植被盖度减少, 地表太阳辐射增多, 表层土壤温度和微生物活性提高, 都可以促进地上凋落物分解(张成霞和南志标, 2010 ; Hewins et al ., 2013 ).另外, 长期重度放牧使叶片有变小的趋势, 这是植物在功能属性上忍受放牧和加快更新的措施, 这些变化使其凋落后的分解更加容易(Cingolani et al ., 2005 ; An & Li, 2014 ).综上所述, 重度放牧加快了地上凋落物的产生和分解, 有利于凋落物中碳氮参与草地生态系统中的碳氮循环, 对维持草地生态系统中碳氮平衡具有重要意义.然而, 重度放牧可能会使土壤物理性质变差, 这将造成重度放牧对草地生态系统物质循环的整体效应降低, 不利于草地的健康发展.因此, 合理放牧才是保证草地健康发展的管理方式. ...
放牧家畜的践踏作用研究评述
1
2004
... 放牧牲畜通过采食、践踏和排泄作用影响凋落物形成和分解.放牧对凋落物的影响主要表现在两个方面: 一是放牧使植物物种组成、牧草本身的形态学特征(如根冠比、叶片/叶鞘等)发生改变, 从而影响凋落物起始质量(Holland & Detling, 1990 ; Semmartin & Ghersa, 2006 ); 二是放牧使凋落物分解的立地土壤物理和生物环境发生改变, 直接影响凋落物分解(Shariff & Grygiel, 1994 ).本研究中, 重度放牧样地地上凋落物产量显著高于轻度放牧, 因为放牧可以削减草地群落的物种冗余程度, 牲畜唾液可以引入生长刺激物, 使植物产生补偿效应(Gong et al ., 2015 ); 两年中重度放牧比轻度放牧更能促进地上凋落物分解, 重度放牧样地地上凋落物质量好, C、N含量高、木质素:N低, 加上牲畜的踩踏作用加快凋落物的碎裂并与土壤紧密接触和混合.同时牲畜啃食会使植被盖度减少, 地表太阳辐射增多, 表层土壤温度和微生物活性提高, 都可以促进地上凋落物分解(张成霞和南志标, 2010 ; Hewins et al ., 2013 ).另外, 长期重度放牧使叶片有变小的趋势, 这是植物在功能属性上忍受放牧和加快更新的措施, 这些变化使其凋落后的分解更加容易(Cingolani et al ., 2005 ; An & Li, 2014 ).综上所述, 重度放牧加快了地上凋落物的产生和分解, 有利于凋落物中碳氮参与草地生态系统中的碳氮循环, 对维持草地生态系统中碳氮平衡具有重要意义.然而, 重度放牧可能会使土壤物理性质变差, 这将造成重度放牧对草地生态系统物质循环的整体效应降低, 不利于草地的健康发展.因此, 合理放牧才是保证草地健康发展的管理方式. ...
A synthesis of the effect of grazing exclusion on carbon dynamics in grasslands in China.
2016
Decomposition of dominant plant species litter in a semi-arid grassland.
1
2003
... 植物分配一定的光合物质到地下以维持根系的产量, 同时根系吸收土壤中的水分和养分供给地上部分生长.因此植物地上和根系的生长紧密相连, 地上和地下凋落物的产生和分解也是相互影响的(Bai et al ., 2015 ).植物根系是土壤有机质的主要来源, 放牧主要通过影响土壤微环境(温度和湿度)和根系特征来影响地下凋落物产生和分解, 进而影响草地生态系统土壤养分循环(Smith et al ., 2014 ).本研究中, 干旱年重度放牧样地地下凋落物产量显著低于轻度放牧样地, 湿润年则相反.这可能是由于重度放牧样地家畜的践踏作用使土壤孔隙度和水稳性团聚体减少, 引起土壤透水性、透气性和水导率下降, 加上在干旱年水分亏缺严重, 严重阻碍根系的生长(侯扶江等, 2004 ).而湿润年降水充足, 土壤理化性质变好, 适宜根系生长, 重度放牧样地地下凋落物产量迅速增加, 显著高于轻度放牧样地.另外在本研究中, 干旱年重度放牧地下凋落物分解速率显著低于轻度放牧, 湿润年则相反.在干旱年, 重度放牧使植被盖度显著下降, 导致地表裸露、土温上升、土壤水分散失加剧、土壤物理性质变差; 另一方面, 土壤可利用性水分减少, 加上内蒙古草原的高海拔使重度放牧样地裸露的地表UV-B辐射增多, 大幅降低土壤分解者的活性, 这些都会严重阻碍重度放牧样地地下凋落物的分解(Dormaar & Willms, 1990 ; Giese et al ., 2009 ).但在湿润年, 重度放牧地下凋落物质量变好, 低C:N和木质素:N使重度放牧样地地下分解速率显著高于轻度放牧, 这也说明重度放牧样地地下凋落物分解对降水条件的响应十分敏感.因此我们的实验结果显示放牧对半干旱草地生态系统凋落物产生和分解的影响主要取决于环境的降水条件. ...
Livestock grazing exclusion and microhabitat variation affect invertebrates and litter decomposition rates in woodland remnants.
1
2009
... 放牧和围栏封育是草地最基本的两种管理方式, 它们直接改变地表的植被特征, 也改变土壤环境(如温度、含水量等)及其养分含量, 使植物碳分配模式和土壤微生物代谢过程等发生改变, 进而影响凋落物分解和生态系统碳循环(Raich & Tufekciogiu, 2000 ).放牧是草地生态系统生物地球化学循环的重要驱动因子, 不同放牧强度下, 牲畜通过采食、排泄和践踏等方式影响植物群落结构和土壤特性, 使凋落物形成和分解发生变化(Stark et al ., 2015 ).但是目前关于放牧强度对地下凋落物分解的影响研究不足.围栏封育是保持草地生态系统功能的有效管理方式.放牧草地经过围栏封育后, 使草地植被得到恢复, 土壤特性、碳输入、植物组成以及凋落物基质质量发生了变化, 这些因素都影响了草地凋落物的形成和分解(王蕾等, 2013 ; Lu et al ., 2015 ).目前, 国内外学者对草地围栏封育的研究仅局限在不同年限的围封对群落结构和凋落物的影响.研究表明, 围封时间过长会引起群落地上生物量降低、凋落物堆积, 抑制植物的再生, 一部分优势种开始衰退, 阻碍草地资源的更新利用(Eldridge et al ., 1992 ; 左万庆等, 2009 ), 因此研究放牧后的短期围封对草地恢复更具指导意义.目前, 草地不同退化强度基础上的短期围封研究尚且不足(Yeo, 2005 ; Hu et al ., 2016 ).因此, 我们研究不同放牧强度下的短期围栏封育对凋落物分解和草地生产力恢复的影响, 以期为防止草地退化、改善生态环境提供参考依据. ...
Enhanced litter input rather than changes in litter chemistry drive soil carbon and nitrogen cycles under elevated CO2: A microcosm study.
1
2009
... 凋落物基质质量能够表征凋落物在自然界中的可分解性.衡量凋落物质量的指标主要是C含量、N含量、木质素浓度、C:N、木质素:N等.一般来说, 凋落物中N含量越高、C:N越小、木质素含量越低, 凋落物分解速率越快(Koukoura et al ., 2003 ).在干旱和半干旱草地生态系统中, 降水是影响凋落物分解的重要气候因子, 它可以通过影响植物群落组成和分解者的活性来影响凋落物质量, 进而影响凋落物分解.但降水和凋落物质量可能存在交互作用, 使得我们无法准确评估某一因子对分解的影响(Bontti et al ., 2009 ).本研究在内蒙古温带草原上开展, 2011年和2012年该区域的降水量分别是188 mm和465 mm, 存在明显差异, 使得本文恰好能够探究降水差异对草地凋落物分解的影响.本文在内蒙古温带典型草原中测定了干湿年份内4种样地(轻度放牧样地、轻度放牧后围封样地、重度放牧样地、重度放牧后围封样地)的土壤养分含量、凋落物产量、质量及分解速率来探讨不同放牧强度对凋落物形成和分解的影响, 以及围栏封育对生态系统恢复的作用, 具体研究目标如下: 1)分析不同放牧强度和围栏封育下凋落物分解的差异; 2)探讨调控地上、地下凋落物分解的主要因子; 3)揭示降水变化对凋落物产量和分解的作用.通过讨论分析, 以期为草地生态系统和草地畜牧业的可持续发展提供理论依据. ...
Carbon, nitrogen, and phosphorus storage in alpine grassland ecosystems of Tibet: Effects of grazing exclusion.
1
2015
... 围栏封育是草地有效管理方式之一, 它对植被资源恢复、土壤理化性质恢复和生物多样性保护具有重要意义.围封后植被生长状况、土壤无脊椎动物和微生物活性以及凋落物分解的关键过程都会发生改变(Lindsay & Cunningham, 2009 ).本研究中, 两年内放牧样地地上凋落物产量在围封后均有显著提高.由于围栏封育后排除人为和牲畜干扰, 植被得到恢复, 地上生物量、凋落物积累和土壤有机质输入显著提高(Tessier et al ., 2003 ).围封后除了植被得到恢复外, 土壤条件也得到明显改善.放牧样地经过围封后, 土壤容重降低, 养分含量增加, 土壤酶活性增强, 这些都可以促进围封样地地上凋落物形成(文海燕等, 2005 ).在本研究两年围封期间, 轻度放牧和轻度放牧后围封样地凋落物质量和土壤养分之间的差异并不显著, 但轻度放牧后围封样地地上凋落物分解速率加快, 可能是土壤物理和生物特性发生变化的结果(Mekuria et al ., 2007 ).与之相反, 重度放牧围封后地上凋落物分解速率减慢, 凋落物质量变差, 说明重度放牧围封后植被组成发生了变化, 植物叶片结构性碳(木质素)合成增加, 导致地上凋落物木质素含量和木质素:N升高, 凋落物中难分解物质比例增加(Bontti et al ., 2009 ).围封后排除了牲畜的排泄作用, 土壤有机质含量减少, 土壤酶活性和微生物活性降低, 这都将导致重度放牧样地围封后地上凋落物分解减慢(Aldezabal et al ., 2015 ).由此可以推测, 长期轻度放牧样地生态系统的恢复作用更强, 短期围封后地上凋落物分解速率和养分循环加快. ...
Effectiveness of exclosures to restore degraded soils as a result of overgrazing in Tigray, Ethiopia.
1
2007
... 除放牧强度和围栏封育外, 降水变化对凋落物分解也有显著影响.降水通过影响土壤温湿度, 改变植物群落组成和功能, 从而影响凋落物质量和土壤微生物活性, 调控凋落物形成和分解过程(Austin & Vitousek, 2001 ).本研究中, 湿润年地上、地下凋落物产量和分解速率均高于干旱年.干旱年植物生长受水分限制, 导致气孔关闭、光合能力下降, 植物生产力降低(Sanaullah et al ., 2014 ), 产生的凋落物也较湿润年少.在轻度放牧和轻度放牧后围封样地, 湿润年土壤全C含量是干旱年的近2倍.湿润年降水增多可提高土壤含水量和酶活性(Mikan et al ., 2002 )、加快凋落物化学组分的淋溶, 从地表输入土壤的C增多, 根系的分解也随之加快, 土壤C输入显著提高.并且土壤可利用水增多可加快土壤中的养分向微生物种群扩散, 使微生物活性增强(Wang et al ., 2015 ).这都可以显著促进凋落物分解过程, 加快草地生态系统的碳循环和养分循环.另外, 在草地生态系统中, 由于植物根生物量比其他生态系统大, 较快的根系分解速率和周转速率有助于维持土壤养分的供应和植物的生长(Liu et al ., 2009 ).因此降水还可以通过促进地下凋落物分解而增加土壤养分, 进而促进植物地上部分生长和凋落物分解过程. ...
Temperature controls of microbial respiration in arctic tundra soils above and below freezing.
1
2002
... 放牧和围栏封育是草地最基本的两种管理方式, 它们直接改变地表的植被特征, 也改变土壤环境(如温度、含水量等)及其养分含量, 使植物碳分配模式和土壤微生物代谢过程等发生改变, 进而影响凋落物分解和生态系统碳循环(Raich & Tufekciogiu, 2000 ).放牧是草地生态系统生物地球化学循环的重要驱动因子, 不同放牧强度下, 牲畜通过采食、排泄和践踏等方式影响植物群落结构和土壤特性, 使凋落物形成和分解发生变化(Stark et al ., 2015 ).但是目前关于放牧强度对地下凋落物分解的影响研究不足.围栏封育是保持草地生态系统功能的有效管理方式.放牧草地经过围栏封育后, 使草地植被得到恢复, 土壤特性、碳输入、植物组成以及凋落物基质质量发生了变化, 这些因素都影响了草地凋落物的形成和分解(王蕾等, 2013 ; Lu et al ., 2015 ).目前, 国内外学者对草地围栏封育的研究仅局限在不同年限的围封对群落结构和凋落物的影响.研究表明, 围封时间过长会引起群落地上生物量降低、凋落物堆积, 抑制植物的再生, 一部分优势种开始衰退, 阻碍草地资源的更新利用(Eldridge et al ., 1992 ; 左万庆等, 2009 ), 因此研究放牧后的短期围封对草地恢复更具指导意义.目前, 草地不同退化强度基础上的短期围封研究尚且不足(Yeo, 2005 ; Hu et al ., 2016 ).因此, 我们研究不同放牧强度下的短期围栏封育对凋落物分解和草地生产力恢复的影响, 以期为防止草地退化、改善生态环境提供参考依据. ...
Aboveground litter quality changes may drive soil organic carbon increase after shrub encroachment into mountain grasslands.
1
2010
... 围栏封育是草地有效管理方式之一, 它对植被资源恢复、土壤理化性质恢复和生物多样性保护具有重要意义.围封后植被生长状况、土壤无脊椎动物和微生物活性以及凋落物分解的关键过程都会发生改变(Lindsay & Cunningham, 2009 ).本研究中, 两年内放牧样地地上凋落物产量在围封后均有显著提高.由于围栏封育后排除人为和牲畜干扰, 植被得到恢复, 地上生物量、凋落物积累和土壤有机质输入显著提高(Tessier et al ., 2003 ).围封后除了植被得到恢复外, 土壤条件也得到明显改善.放牧样地经过围封后, 土壤容重降低, 养分含量增加, 土壤酶活性增强, 这些都可以促进围封样地地上凋落物形成(文海燕等, 2005 ).在本研究两年围封期间, 轻度放牧和轻度放牧后围封样地凋落物质量和土壤养分之间的差异并不显著, 但轻度放牧后围封样地地上凋落物分解速率加快, 可能是土壤物理和生物特性发生变化的结果(Mekuria et al ., 2007 ).与之相反, 重度放牧围封后地上凋落物分解速率减慢, 凋落物质量变差, 说明重度放牧围封后植被组成发生了变化, 植物叶片结构性碳(木质素)合成增加, 导致地上凋落物木质素含量和木质素:N升高, 凋落物中难分解物质比例增加(Bontti et al ., 2009 ).围封后排除了牲畜的排泄作用, 土壤有机质含量减少, 土壤酶活性和微生物活性降低, 这都将导致重度放牧样地围封后地上凋落物分解减慢(Aldezabal et al ., 2015 ).由此可以推测, 长期轻度放牧样地生态系统的恢复作用更强, 短期围封后地上凋落物分解速率和养分循环加快. ...
2
... 将2011年和2012年每次洗净烘干的凋落物样品进行粉碎研磨, 再通过40目网筛.一部分用于凋落物C、N含量的测定, 另一部分用于木质素和纤维素含量的测定.全C含量的测定采用重铬酸钾法(Nelson & Sommers, 1982 ), 全N含量的测定采用凯氏定氮法(Bremner, 1960 ).木质素和纤维素的测定依据van Soest (1963, 1967)的实验原理, 参照AOAC international (2000)的具体实验操作标准, 经过消煮-酸洗-灼烧等步骤, 测定各样地初始凋落物和残留凋落物中的木质素和纤维素的含量. ...
... 除放牧强度和围栏封育外, 降水变化对凋落物分解也有显著影响.降水通过影响土壤温湿度, 改变植物群落组成和功能, 从而影响凋落物质量和土壤微生物活性, 调控凋落物形成和分解过程(Austin & Vitousek, 2001 ).本研究中, 湿润年地上、地下凋落物产量和分解速率均高于干旱年.干旱年植物生长受水分限制, 导致气孔关闭、光合能力下降, 植物生产力降低(Sanaullah et al ., 2014 ), 产生的凋落物也较湿润年少.在轻度放牧和轻度放牧后围封样地, 湿润年土壤全C含量是干旱年的近2倍.湿润年降水增多可提高土壤含水量和酶活性(Mikan et al ., 2002 )、加快凋落物化学组分的淋溶, 从地表输入土壤的C增多, 根系的分解也随之加快, 土壤C输入显著提高.并且土壤可利用水增多可加快土壤中的养分向微生物种群扩散, 使微生物活性增强(Wang et al ., 2015 ).这都可以显著促进凋落物分解过程, 加快草地生态系统的碳循环和养分循环.另外, 在草地生态系统中, 由于植物根生物量比其他生态系统大, 较快的根系分解速率和周转速率有助于维持土壤养分的供应和植物的生长(Liu et al ., 2009 ).因此降水还可以通过促进地下凋落物分解而增加土壤养分, 进而促进植物地上部分生长和凋落物分解过程. ...
Energy storage and the balance of producers and decomposers in ecological systems.
2
1963
... 我们用生长季内地上生物量的最大值代表该年地上凋落物的产量(Zhang et al ., 2013 ).分别于2011年和2012年生长高峰期8月下旬, 在轻度放牧后围封、轻度放牧、重度放牧后围封、重度放牧4个样地内随机布设5个样方(1 m × 1 m), 收集样方内全部地上生物量, 不包括直立或倒伏的凋落物, 去除植物体表面的泥土和杂质, 在105 ℃下杀青15 min, 然后在65 ℃下烘干至恒质量后进行称量.在生长季末, 植物体将全部凋亡, 以凋落物的形式归还到土壤中.地下凋落物产量采用内生长土芯法测定, 于2011年4月中旬在4个样地内随机布设5个样点, 按照0-10、10-20、20-30、30-40 cm 4个垂直层次, 用根钻打钻, 每钻土壤不混合, 分层收集, 带回实验室立即处理.在室内尽量挑出每层土壤的全部根系, 然后将处理好的无根土壤对应回填.回填时, 按照土层顺序对应将每两层装入一个圆柱形尼龙网袋中, 然后按上下两层对应放回原位.于每年生长季初和生长季末, 每个样点各取样一次, 每个样点取上下两层土芯网袋.带回实验室后清洗网袋中全部活根系, 在105 ℃下杀青15 min, 然后在65 ℃下 烘干至恒质量.用生长季初和生长季末根新生量 的差值作为该年地下凋落物产量(Montané et al ., 2010 ). ...
... 2010年8月上旬, 分别在4个样地内随机选择5个样点, 随机布设5个样方(1 m × 1 m)采集地上凋落物, 采集的凋落物在自然条件下风干, 去除植物体表面的泥土和杂质.用于地上凋落物分解实验.去除地表植被后, 按照0-10、10-20、20-30、30-40 cm 4个垂直层次, 用根钻采集地下完整的活根系, 自然风干.采用凋落物网袋法进行实验, 将地上、地下凋落物剪成20 cm的小段, 取10 g地上凋落物或者5 g地下凋落物装入1 mm孔径的尼龙网袋(10 cm × 10 cm), 每个样地36袋地上凋落物网袋和36袋地下凋落物网袋.在每个样地内选取3个样点, 去除地表植被, 在每个样点将12袋地上凋落物网袋均匀平铺在地表与土壤直接接触.在样点附近, 将12袋地下凋落物网袋均匀埋放在土深10 cm左右的土层.2011-2012年5月和9月下旬, 在每样地3个样点内各收集3袋凋落物.将网袋内凋落物全部取出洗净, 在65 ℃下烘干至恒质量, 进行称量.利用每次收获得到的残留凋落物干质量, 根据Olson (1963) 指数分解模型公式, 计算分解速率k 值: ...
Below- ground biomass and productivity of a grazed site and a neighbouring ungrazed exclosure in a grassland in central Argentina.
2004
Vegetation and soil respiration: Correlations and controls.
2
2000
... 放牧和围栏封育是草地最基本的两种管理方式, 它们直接改变地表的植被特征, 也改变土壤环境(如温度、含水量等)及其养分含量, 使植物碳分配模式和土壤微生物代谢过程等发生改变, 进而影响凋落物分解和生态系统碳循环(Raich & Tufekciogiu, 2000 ).放牧是草地生态系统生物地球化学循环的重要驱动因子, 不同放牧强度下, 牲畜通过采食、排泄和践踏等方式影响植物群落结构和土壤特性, 使凋落物形成和分解发生变化(Stark et al ., 2015 ).但是目前关于放牧强度对地下凋落物分解的影响研究不足.围栏封育是保持草地生态系统功能的有效管理方式.放牧草地经过围栏封育后, 使草地植被得到恢复, 土壤特性、碳输入、植物组成以及凋落物基质质量发生了变化, 这些因素都影响了草地凋落物的形成和分解(王蕾等, 2013 ; Lu et al ., 2015 ).目前, 国内外学者对草地围栏封育的研究仅局限在不同年限的围封对群落结构和凋落物的影响.研究表明, 围封时间过长会引起群落地上生物量降低、凋落物堆积, 抑制植物的再生, 一部分优势种开始衰退, 阻碍草地资源的更新利用(Eldridge et al ., 1992 ; 左万庆等, 2009 ), 因此研究放牧后的短期围封对草地恢复更具指导意义.目前, 草地不同退化强度基础上的短期围封研究尚且不足(Yeo, 2005 ; Hu et al ., 2016 ).因此, 我们研究不同放牧强度下的短期围栏封育对凋落物分解和草地生产力恢复的影响, 以期为防止草地退化、改善生态环境提供参考依据. ...
... 在本研究中, 短期围封后地下凋落物产量反而显著低于对应的放牧样地.放牧活动能促使根系向土壤上层集中, 且它对根系的生长有促进作用, 因此围封后根系生长变缓(Greenwood & Hutchinson, 2003 ; Pucheta et al ., 2004 ).但只有在干旱年, 重度放牧后围封样地地下凋落物产量显著高于重度放牧.这可能是由于重度放牧样地牲畜的踩踏作用使土壤遭到破坏, 加上干旱年降水稀少, 限制了地下根系生长.在干旱年, 放牧和围封样地地下凋落物分解速率存在显著差异主要是凋落物质量差异所致.轻度放牧样地围封后, 地下凋落物质量变差, 高木质素含量和木质素:N使得轻度放牧后围封样地地下凋落物分解速率显著低于轻度放牧样地; 而重度放牧样地相反, 围封后凋落物质量和土壤养分状况变好(图3 , 图5 ), 使得重度放牧样地围封后分解速率显著高于重度放牧样地.湿润年各样地的地下凋落物质量和土壤养分之间虽然存在差异, 但各样地地下凋落物分解速率并无显著差异.相比于凋落物质量和土壤养分的作用, 降水对地下凋落物分解过程的调控作用更强(Bontti et al ., 2009 ; Giese et al ., 2009 ).综上所述, 本研究表明短期围栏封育能使植被生长环境、土壤理化性质得到明显改善, 轻度放牧围封后还能提高凋落物分解速率和养分循环.不过有研究表明, 长期围封会使草地非禾本科牧草比例大幅增加, 不利于禾本科牧草产量的积累, 这将会阻碍畜牧业发展, 造成严重的经济损失(Shi et al ., 2010 ).因此, 在草地放牧强度和围封年限的确定上, 我们应注意寻找草地经济效益和生态系统保护之间的平衡点. ...
Effects of drought and elevated temperature on biochemical composition of forage plants and their impact on carbon storage in grassland soil.
2014
Intraspecific changes in plant morphology, associated with grazing, and effects on litter quality, carbon and nutrient dynamics during decomposition.
1
2006
... 随着全球气候变化加剧, 碳循环直接影响着人类的生存和各种生物环境的稳定性, 逐渐成为生态系统研究的重点(杨景成等, 2003 ; Bardgett et al ., 2013 ).陆地生态系统在全球碳循环中起着重要的作用, 因此对陆地生态系统碳循环的理解非常必要(Canadell et al ., 2000 ).我国现有草地面积约4亿hm2 , 是最大的陆地生态系统, 也是生物地球循环的重要组成部分, 但其容易受到气候变化和各种人类活动(如: 放牧、施肥等)的影响(齐玉春等, 2003 ; Xu & Guo, 2015 ).目前, 由于放牧和农垦活动频繁, 草地生态系统成为受人类活动影响较为严重的区域.因此, 对自然和人为因素干扰下草地生态系统碳循环过程的研究具有十分重要的科学意义. ...
Grazing intensity effects on litter decomposition and soil nitrogen mineralization.
1994
Effects of livestock exclusion on vegetation and soil properties under two topographic habitats in an alpine meadow on the eastern Qinghai-Tibetan Plateau.
1
2010
... 除放牧强度和围栏封育外, 降水变化对凋落物分解也有显著影响.降水通过影响土壤温湿度, 改变植物群落组成和功能, 从而影响凋落物质量和土壤微生物活性, 调控凋落物形成和分解过程(Austin & Vitousek, 2001 ).本研究中, 湿润年地上、地下凋落物产量和分解速率均高于干旱年.干旱年植物生长受水分限制, 导致气孔关闭、光合能力下降, 植物生产力降低(Sanaullah et al ., 2014 ), 产生的凋落物也较湿润年少.在轻度放牧和轻度放牧后围封样地, 湿润年土壤全C含量是干旱年的近2倍.湿润年降水增多可提高土壤含水量和酶活性(Mikan et al ., 2002 )、加快凋落物化学组分的淋溶, 从地表输入土壤的C增多, 根系的分解也随之加快, 土壤C输入显著提高.并且土壤可利用水增多可加快土壤中的养分向微生物种群扩散, 使微生物活性增强(Wang et al ., 2015 ).这都可以显著促进凋落物分解过程, 加快草地生态系统的碳循环和养分循环.另外, 在草地生态系统中, 由于植物根生物量比其他生态系统大, 较快的根系分解速率和周转速率有助于维持土壤养分的供应和植物的生长(Liu et al ., 2009 ).因此降水还可以通过促进地下凋落物分解而增加土壤养分, 进而促进植物地上部分生长和凋落物分解过程. ...
Root traits predict decomposition across a landscape-scale grazing experiment.
1
2014
... 放牧牲畜通过采食、践踏和排泄作用影响凋落物形成和分解.放牧对凋落物的影响主要表现在两个方面: 一是放牧使植物物种组成、牧草本身的形态学特征(如根冠比、叶片/叶鞘等)发生改变, 从而影响凋落物起始质量(Holland & Detling, 1990 ; Semmartin & Ghersa, 2006 ); 二是放牧使凋落物分解的立地土壤物理和生物环境发生改变, 直接影响凋落物分解(Shariff & Grygiel, 1994 ).本研究中, 重度放牧样地地上凋落物产量显著高于轻度放牧, 因为放牧可以削减草地群落的物种冗余程度, 牲畜唾液可以引入生长刺激物, 使植物产生补偿效应(Gong et al ., 2015 ); 两年中重度放牧比轻度放牧更能促进地上凋落物分解, 重度放牧样地地上凋落物质量好, C、N含量高、木质素:N低, 加上牲畜的踩踏作用加快凋落物的碎裂并与土壤紧密接触和混合.同时牲畜啃食会使植被盖度减少, 地表太阳辐射增多, 表层土壤温度和微生物活性提高, 都可以促进地上凋落物分解(张成霞和南志标, 2010 ; Hewins et al ., 2013 ).另外, 长期重度放牧使叶片有变小的趋势, 这是植物在功能属性上忍受放牧和加快更新的措施, 这些变化使其凋落后的分解更加容易(Cingolani et al ., 2005 ; An & Li, 2014 ).综上所述, 重度放牧加快了地上凋落物的产生和分解, 有利于凋落物中碳氮参与草地生态系统中的碳氮循环, 对维持草地生态系统中碳氮平衡具有重要意义.然而, 重度放牧可能会使土壤物理性质变差, 这将造成重度放牧对草地生态系统物质循环的整体效应降低, 不利于草地的健康发展.因此, 合理放牧才是保证草地健康发展的管理方式. ...
Factors controlling decomposition rates of fine root litter in temperate forests and grasslands.
1
2014
... 放牧牲畜通过采食、践踏和排泄作用影响凋落物形成和分解.放牧对凋落物的影响主要表现在两个方面: 一是放牧使植物物种组成、牧草本身的形态学特征(如根冠比、叶片/叶鞘等)发生改变, 从而影响凋落物起始质量(Holland & Detling, 1990 ; Semmartin & Ghersa, 2006 ); 二是放牧使凋落物分解的立地土壤物理和生物环境发生改变, 直接影响凋落物分解(Shariff & Grygiel, 1994 ).本研究中, 重度放牧样地地上凋落物产量显著高于轻度放牧, 因为放牧可以削减草地群落的物种冗余程度, 牲畜唾液可以引入生长刺激物, 使植物产生补偿效应(Gong et al ., 2015 ); 两年中重度放牧比轻度放牧更能促进地上凋落物分解, 重度放牧样地地上凋落物质量好, C、N含量高、木质素:N低, 加上牲畜的踩踏作用加快凋落物的碎裂并与土壤紧密接触和混合.同时牲畜啃食会使植被盖度减少, 地表太阳辐射增多, 表层土壤温度和微生物活性提高, 都可以促进地上凋落物分解(张成霞和南志标, 2010 ; Hewins et al ., 2013 ).另外, 长期重度放牧使叶片有变小的趋势, 这是植物在功能属性上忍受放牧和加快更新的措施, 这些变化使其凋落后的分解更加容易(Cingolani et al ., 2005 ; An & Li, 2014 ).综上所述, 重度放牧加快了地上凋落物的产生和分解, 有利于凋落物中碳氮参与草地生态系统中的碳氮循环, 对维持草地生态系统中碳氮平衡具有重要意义.然而, 重度放牧可能会使土壤物理性质变差, 这将造成重度放牧对草地生态系统物质循环的整体效应降低, 不利于草地的健康发展.因此, 合理放牧才是保证草地健康发展的管理方式. ...
Grazing intensity in subarctic tundra affects the temperature adaptation of soil microbial communities.
1
2015
... 在本研究中, 短期围封后地下凋落物产量反而显著低于对应的放牧样地.放牧活动能促使根系向土壤上层集中, 且它对根系的生长有促进作用, 因此围封后根系生长变缓(Greenwood & Hutchinson, 2003 ; Pucheta et al ., 2004 ).但只有在干旱年, 重度放牧后围封样地地下凋落物产量显著高于重度放牧.这可能是由于重度放牧样地牲畜的踩踏作用使土壤遭到破坏, 加上干旱年降水稀少, 限制了地下根系生长.在干旱年, 放牧和围封样地地下凋落物分解速率存在显著差异主要是凋落物质量差异所致.轻度放牧样地围封后, 地下凋落物质量变差, 高木质素含量和木质素:N使得轻度放牧后围封样地地下凋落物分解速率显著低于轻度放牧样地; 而重度放牧样地相反, 围封后凋落物质量和土壤养分状况变好(图3 , 图5 ), 使得重度放牧样地围封后分解速率显著高于重度放牧样地.湿润年各样地的地下凋落物质量和土壤养分之间虽然存在差异, 但各样地地下凋落物分解速率并无显著差异.相比于凋落物质量和土壤养分的作用, 降水对地下凋落物分解过程的调控作用更强(Bontti et al ., 2009 ; Giese et al ., 2009 ).综上所述, 本研究表明短期围栏封育能使植被生长环境、土壤理化性质得到明显改善, 轻度放牧围封后还能提高凋落物分解速率和养分循环.不过有研究表明, 长期围封会使草地非禾本科牧草比例大幅增加, 不利于禾本科牧草产量的积累, 这将会阻碍畜牧业发展, 造成严重的经济损失(Shi et al ., 2010 ).因此, 在草地放牧强度和围封年限的确定上, 我们应注意寻找草地经济效益和生态系统保护之间的平衡点. ...
Comparison of litterfall production and leaf litter decomposition between an exotic black locust plantation and an indigenous oak forest near Yan’an on the Loess Plateau, China.
1
2007
... 植物分配一定的光合物质到地下以维持根系的产量, 同时根系吸收土壤中的水分和养分供给地上部分生长.因此植物地上和根系的生长紧密相连, 地上和地下凋落物的产生和分解也是相互影响的(Bai et al ., 2015 ).植物根系是土壤有机质的主要来源, 放牧主要通过影响土壤微环境(温度和湿度)和根系特征来影响地下凋落物产生和分解, 进而影响草地生态系统土壤养分循环(Smith et al ., 2014 ).本研究中, 干旱年重度放牧样地地下凋落物产量显著低于轻度放牧样地, 湿润年则相反.这可能是由于重度放牧样地家畜的践踏作用使土壤孔隙度和水稳性团聚体减少, 引起土壤透水性、透气性和水导率下降, 加上在干旱年水分亏缺严重, 严重阻碍根系的生长(侯扶江等, 2004 ).而湿润年降水充足, 土壤理化性质变好, 适宜根系生长, 重度放牧样地地下凋落物产量迅速增加, 显著高于轻度放牧样地.另外在本研究中, 干旱年重度放牧地下凋落物分解速率显著低于轻度放牧, 湿润年则相反.在干旱年, 重度放牧使植被盖度显著下降, 导致地表裸露、土温上升、土壤水分散失加剧、土壤物理性质变差; 另一方面, 土壤可利用性水分减少, 加上内蒙古草原的高海拔使重度放牧样地裸露的地表UV-B辐射增多, 大幅降低土壤分解者的活性, 这些都会严重阻碍重度放牧样地地下凋落物的分解(Dormaar & Willms, 1990 ; Giese et al ., 2009 ).但在湿润年, 重度放牧地下凋落物质量变好, 低C:N和木质素:N使重度放牧样地地下分解速率显著高于轻度放牧, 这也说明重度放牧样地地下凋落物分解对降水条件的响应十分敏感.因此我们的实验结果显示放牧对半干旱草地生态系统凋落物产生和分解的影响主要取决于环境的降水条件. ...
Vegetation dynamics and plant species interactions under grazed and ungrazed conditions in a western European salt marsh.
1
2003
... 草地凋落物是指由植物地上部分和地下根系形成并归还到土壤的所有有机质的总称, 是陆地生态系统养分循环和碳循环过程中极为重要的环节(高永恒等, 2007 ).陆地生态系统中约90%的净初级生产量以凋落物的形式归还给土壤, 植物的生长发育在很大程度上依赖于植被归还给土壤的养分再循环(Wardle et al ., 2004 ).凋落物的分解是一个非常复杂的过程, 深受气候、凋落物质量和土壤生物的影响, 在很大程度上决定了生态系统的固碳速率及动态(Tateno et al ., 2007 ).因此, 在全球变化背景下, 对草地生态系统凋落物分解的研究是草地生态系统动态机理以及全球变化生态学研究的重要内容(张艳博等, 2012 ).另外, 地下凋落物的碳输入是地上部分凋落物碳输入的3倍以上, 但是过去仅有2%的植物凋落物研究关注于地下凋落物分解, 很少有研究对干旱、半干旱草原的地上和地下凋落物同时进行系统研究(Zhang et al ., 2008 ; Freschet et al ., 2013 ).因此研究不同干扰下草地地下凋落物分解的变化, 对准确估算全球变化背景下不同尺度碳收支至关重要(Solly et al ., 2014 ). ...
Use of detergents in analysis of fibrous feeds: A rapid method for the determination of fiber and lignin.
1
1963
... 放牧和围栏封育是草地最基本的两种管理方式, 它们直接改变地表的植被特征, 也改变土壤环境(如温度、含水量等)及其养分含量, 使植物碳分配模式和土壤微生物代谢过程等发生改变, 进而影响凋落物分解和生态系统碳循环(Raich & Tufekciogiu, 2000 ).放牧是草地生态系统生物地球化学循环的重要驱动因子, 不同放牧强度下, 牲畜通过采食、排泄和践踏等方式影响植物群落结构和土壤特性, 使凋落物形成和分解发生变化(Stark et al ., 2015 ).但是目前关于放牧强度对地下凋落物分解的影响研究不足.围栏封育是保持草地生态系统功能的有效管理方式.放牧草地经过围栏封育后, 使草地植被得到恢复, 土壤特性、碳输入、植物组成以及凋落物基质质量发生了变化, 这些因素都影响了草地凋落物的形成和分解(王蕾等, 2013 ; Lu et al ., 2015 ).目前, 国内外学者对草地围栏封育的研究仅局限在不同年限的围封对群落结构和凋落物的影响.研究表明, 围封时间过长会引起群落地上生物量降低、凋落物堆积, 抑制植物的再生, 一部分优势种开始衰退, 阻碍草地资源的更新利用(Eldridge et al ., 1992 ; 左万庆等, 2009 ), 因此研究放牧后的短期围封对草地恢复更具指导意义.目前, 草地不同退化强度基础上的短期围封研究尚且不足(Yeo, 2005 ; Hu et al ., 2016 ).因此, 我们研究不同放牧强度下的短期围栏封育对凋落物分解和草地生产力恢复的影响, 以期为防止草地退化、改善生态环境提供参考依据. ...
Development of a comprehensive system of feed analyses and its application to forages.
1
1967
... 草地凋落物是指由植物地上部分和地下根系形成并归还到土壤的所有有机质的总称, 是陆地生态系统养分循环和碳循环过程中极为重要的环节(高永恒等, 2007 ).陆地生态系统中约90%的净初级生产量以凋落物的形式归还给土壤, 植物的生长发育在很大程度上依赖于植被归还给土壤的养分再循环(Wardle et al ., 2004 ).凋落物的分解是一个非常复杂的过程, 深受气候、凋落物质量和土壤生物的影响, 在很大程度上决定了生态系统的固碳速率及动态(Tateno et al ., 2007 ).因此, 在全球变化背景下, 对草地生态系统凋落物分解的研究是草地生态系统动态机理以及全球变化生态学研究的重要内容(张艳博等, 2012 ).另外, 地下凋落物的碳输入是地上部分凋落物碳输入的3倍以上, 但是过去仅有2%的植物凋落物研究关注于地下凋落物分解, 很少有研究对干旱、半干旱草原的地上和地下凋落物同时进行系统研究(Zhang et al ., 2008 ; Freschet et al ., 2013 ).因此研究不同干扰下草地地下凋落物分解的变化, 对准确估算全球变化背景下不同尺度碳收支至关重要(Solly et al ., 2014 ). ...
围封对草地凋落物分解速率和N、P、K含量的影响
1
2013
... 围栏封育是草地有效管理方式之一, 它对植被资源恢复、土壤理化性质恢复和生物多样性保护具有重要意义.围封后植被生长状况、土壤无脊椎动物和微生物活性以及凋落物分解的关键过程都会发生改变(Lindsay & Cunningham, 2009 ).本研究中, 两年内放牧样地地上凋落物产量在围封后均有显著提高.由于围栏封育后排除人为和牲畜干扰, 植被得到恢复, 地上生物量、凋落物积累和土壤有机质输入显著提高(Tessier et al ., 2003 ).围封后除了植被得到恢复外, 土壤条件也得到明显改善.放牧样地经过围封后, 土壤容重降低, 养分含量增加, 土壤酶活性增强, 这些都可以促进围封样地地上凋落物形成(文海燕等, 2005 ).在本研究两年围封期间, 轻度放牧和轻度放牧后围封样地凋落物质量和土壤养分之间的差异并不显著, 但轻度放牧后围封样地地上凋落物分解速率加快, 可能是土壤物理和生物特性发生变化的结果(Mekuria et al ., 2007 ).与之相反, 重度放牧围封后地上凋落物分解速率减慢, 凋落物质量变差, 说明重度放牧围封后植被组成发生了变化, 植物叶片结构性碳(木质素)合成增加, 导致地上凋落物木质素含量和木质素:N升高, 凋落物中难分解物质比例增加(Bontti et al ., 2009 ).围封后排除了牲畜的排泄作用, 土壤有机质含量减少, 土壤酶活性和微生物活性降低, 这都将导致重度放牧样地围封后地上凋落物分解减慢(Aldezabal et al ., 2015 ).由此可以推测, 长期轻度放牧样地生态系统的恢复作用更强, 短期围封后地上凋落物分解速率和养分循环加快. ...
草甸草原不同放牧强度对土壤有机碳含量的影响
1
2007
... 研究样地位于内蒙古大学草地生态系统研究站(116.03°-116.50° E, 44.80°-44.82° N), 海拔约1055 m, 属于温带干旱大陆性气候, 年平均气温0-1 ℃, ≥0 ℃的积温1800 ℃, 年降水量300-360 mm, 无霜期90-115天.生长季从5月到9月, 非生长季从10月到次年4月.气象站观测数据显示: 2011-2012年年平均气温1.7 ℃, 2011年、2012年的累积降水量分别为188 mm和465 mm, 降水多集中在6-8月份.土壤以栗钙土为主.该实验样地植被类型属于温带典型草原, 大针茅(Stipa grandis )、羊草(Leymus chinensis )、冷蒿(Artemisia frigida )在该区的植物群落中占优势地位.样地选在以居民点为中心的自由放牧方式家庭牧场, 该牧场为长期放牧区, 整个生长季都进行放牧.这一地区的主要特点是以居民点为中心,呈条带放射状向外构成不同放牧梯度.根据与居民点的距离远近, 将放牧区划分为轻度放牧样地和重度放牧样地.划分方法是以居民点为中心在放牧地上放射状拉3条样带, 样带之间有一定的距离, 使样带所包含的信息尽可能代表整个草地(王明军等, 2007 ).实验样地整体放牧强度为每hm2 、每年、每天674标准羊单位.实验开始前, 对样地进行基本群落调查, 收获地上生物量, 尤其是对样地内的羊粪便进行计数.每个样地羊粪数和群落调查基本数据均作为样地划分的依据, 样地基本信息见表1 .2011年实验开始时, 在每一条退化样带上的轻度和重度放牧样地各围封了3个退化样地, 每个样地围封面积为100 m × 100 m, 分别为轻度放牧后围封样地和重度放牧后围封样地. ...
Effects of land use and precipitation on above- and below-ground litter decomposition in a semi-arid temperate steppe in Inner Mongolia, China.
2015
Ecological linkages between aboveground and belowground biota.
2004
开垦和封育年限对退化沙质草地土壤性状的影响
1
2005
... 放牧和围栏封育是草地最基本的两种管理方式, 它们直接改变地表的植被特征, 也改变土壤环境(如温度、含水量等)及其养分含量, 使植物碳分配模式和土壤微生物代谢过程等发生改变, 进而影响凋落物分解和生态系统碳循环(Raich & Tufekciogiu, 2000 ).放牧是草地生态系统生物地球化学循环的重要驱动因子, 不同放牧强度下, 牲畜通过采食、排泄和践踏等方式影响植物群落结构和土壤特性, 使凋落物形成和分解发生变化(Stark et al ., 2015 ).但是目前关于放牧强度对地下凋落物分解的影响研究不足.围栏封育是保持草地生态系统功能的有效管理方式.放牧草地经过围栏封育后, 使草地植被得到恢复, 土壤特性、碳输入、植物组成以及凋落物基质质量发生了变化, 这些因素都影响了草地凋落物的形成和分解(王蕾等, 2013 ; Lu et al ., 2015 ).目前, 国内外学者对草地围栏封育的研究仅局限在不同年限的围封对群落结构和凋落物的影响.研究表明, 围封时间过长会引起群落地上生物量降低、凋落物堆积, 抑制植物的再生, 一部分优势种开始衰退, 阻碍草地资源的更新利用(Eldridge et al ., 1992 ; 左万庆等, 2009 ), 因此研究放牧后的短期围封对草地恢复更具指导意义.目前, 草地不同退化强度基础上的短期围封研究尚且不足(Yeo, 2005 ; Hu et al ., 2016 ).因此, 我们研究不同放牧强度下的短期围栏封育对凋落物分解和草地生产力恢复的影响, 以期为防止草地退化、改善生态环境提供参考依据. ...
Evaluating the impacts of nearly 30 years of conservation on grassland ecosystem using Landsat TM images.
2015
Effects of grazing exclusion on rangeland vegetation and soils, East Central Idaho.
1
2005
... 除放牧强度和围栏封育外, 降水变化对凋落物分解也有显著影响.降水通过影响土壤温湿度, 改变植物群落组成和功能, 从而影响凋落物质量和土壤微生物活性, 调控凋落物形成和分解过程(Austin & Vitousek, 2001 ).本研究中, 湿润年地上、地下凋落物产量和分解速率均高于干旱年.干旱年植物生长受水分限制, 导致气孔关闭、光合能力下降, 植物生产力降低(Sanaullah et al ., 2014 ), 产生的凋落物也较湿润年少.在轻度放牧和轻度放牧后围封样地, 湿润年土壤全C含量是干旱年的近2倍.湿润年降水增多可提高土壤含水量和酶活性(Mikan et al ., 2002 )、加快凋落物化学组分的淋溶, 从地表输入土壤的C增多, 根系的分解也随之加快, 土壤C输入显著提高.并且土壤可利用水增多可加快土壤中的养分向微生物种群扩散, 使微生物活性增强(Wang et al ., 2015 ).这都可以显著促进凋落物分解过程, 加快草地生态系统的碳循环和养分循环.另外, 在草地生态系统中, 由于植物根生物量比其他生态系统大, 较快的根系分解速率和周转速率有助于维持土壤养分的供应和植物的生长(Liu et al ., 2009 ).因此降水还可以通过促进地下凋落物分解而增加土壤养分, 进而促进植物地上部分生长和凋落物分解过程. ...
放牧对草地土壤理化特性影响的研究进展
1
2010
... 草地凋落物是指由植物地上部分和地下根系形成并归还到土壤的所有有机质的总称, 是陆地生态系统养分循环和碳循环过程中极为重要的环节(高永恒等, 2007 ).陆地生态系统中约90%的净初级生产量以凋落物的形式归还给土壤, 植物的生长发育在很大程度上依赖于植被归还给土壤的养分再循环(Wardle et al ., 2004 ).凋落物的分解是一个非常复杂的过程, 深受气候、凋落物质量和土壤生物的影响, 在很大程度上决定了生态系统的固碳速率及动态(Tateno et al ., 2007 ).因此, 在全球变化背景下, 对草地生态系统凋落物分解的研究是草地生态系统动态机理以及全球变化生态学研究的重要内容(张艳博等, 2012 ).另外, 地下凋落物的碳输入是地上部分凋落物碳输入的3倍以上, 但是过去仅有2%的植物凋落物研究关注于地下凋落物分解, 很少有研究对干旱、半干旱草原的地上和地下凋落物同时进行系统研究(Zhang et al ., 2008 ; Freschet et al ., 2013 ).因此研究不同干扰下草地地下凋落物分解的变化, 对准确估算全球变化背景下不同尺度碳收支至关重要(Solly et al ., 2014 ). ...
Rates of litter decom- position in terrestrial ecosystems: Global patterns and controlling factors.
2008
Linking litter production, quality and decomposition to vegetation succession following agricultural abandonment.
1
2013
... 围栏封育是草地有效管理方式之一, 它对植被资源恢复、土壤理化性质恢复和生物多样性保护具有重要意义.围封后植被生长状况、土壤无脊椎动物和微生物活性以及凋落物分解的关键过程都会发生改变(Lindsay & Cunningham, 2009 ).本研究中, 两年内放牧样地地上凋落物产量在围封后均有显著提高.由于围栏封育后排除人为和牲畜干扰, 植被得到恢复, 地上生物量、凋落物积累和土壤有机质输入显著提高(Tessier et al ., 2003 ).围封后除了植被得到恢复外, 土壤条件也得到明显改善.放牧样地经过围封后, 土壤容重降低, 养分含量增加, 土壤酶活性增强, 这些都可以促进围封样地地上凋落物形成(文海燕等, 2005 ).在本研究两年围封期间, 轻度放牧和轻度放牧后围封样地凋落物质量和土壤养分之间的差异并不显著, 但轻度放牧后围封样地地上凋落物分解速率加快, 可能是土壤物理和生物特性发生变化的结果(Mekuria et al ., 2007 ).与之相反, 重度放牧围封后地上凋落物分解速率减慢, 凋落物质量变差, 说明重度放牧围封后植被组成发生了变化, 植物叶片结构性碳(木质素)合成增加, 导致地上凋落物木质素含量和木质素:N升高, 凋落物中难分解物质比例增加(Bontti et al ., 2009 ).围封后排除了牲畜的排泄作用, 土壤有机质含量减少, 土壤酶活性和微生物活性降低, 这都将导致重度放牧样地围封后地上凋落物分解减慢(Aldezabal et al ., 2015 ).由此可以推测, 长期轻度放牧样地生态系统的恢复作用更强, 短期围封后地上凋落物分解速率和养分循环加快. ...
放牧对青藏高原东部两种典型高寒草地类型凋落物分解的影响
1
2012
... 随着全球气候变化加剧, 碳循环直接影响着人类的生存和各种生物环境的稳定性, 逐渐成为生态系统研究的重点(杨景成等, 2003 ; Bardgett et al ., 2013 ).陆地生态系统在全球碳循环中起着重要的作用, 因此对陆地生态系统碳循环的理解非常必要(Canadell et al ., 2000 ).我国现有草地面积约4亿hm2 , 是最大的陆地生态系统, 也是生物地球循环的重要组成部分, 但其容易受到气候变化和各种人类活动(如: 放牧、施肥等)的影响(齐玉春等, 2003 ; Xu & Guo, 2015 ).目前, 由于放牧和农垦活动频繁, 草地生态系统成为受人类活动影响较为严重的区域.因此, 对自然和人为因素干扰下草地生态系统碳循环过程的研究具有十分重要的科学意义. ...
围栏封育措施对退化羊草草原植物群落特征影响研究
2009
 , 王忆慧, 刘敏, 罗亲普, 徐沙, 潘琰, 翟占伟
, 王忆慧, 刘敏, 罗亲普, 徐沙, 潘琰, 翟占伟


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}