气候变化、氮沉降、生物入侵和土地利用变化已经成为当前全球生物多样性的主要威胁(Sala et al ., 2000 ), 因此许多学者开始研究全球变化对生物多样性和物种分布的潜在影响。在这样的背景下, 理解限制物种分布的过程和机制就显得尤为重要(Travis et al ., 2006 )。附生地衣是森林生态系统的重要组成部分, 占据着林冠相当份量的生物量(Sipman & Harris, 1989 ; Gradstein, 1992 ), 其多样性和分布自20世纪70年代起就得到了众多学者的关注(Cornelissen & Steege, 1999 ; Holz & Gradstein, 2005 ; Hauck & Spribille, 2005 )。Aptroot和Sipman (1997)估计在热带雨林中每km2 有300种地衣, Aptroot (1997)报道在巴布亚新几内亚的热带山地雨林一株树上就有173种地衣。地衣的分布受众多因子的综合影响(Gustafasson & Eriksson, 1995 ; Goward & Arsenawlt, 2000 ; Hauck & Spribille, 2005 ), 除自身的繁殖能力和扩散能力以外, 光照、温度、湿度、干扰程度、群落类型以及宿主特征等均对附生地衣物种的组成和分布产生一定的影响(李苏等, 2007 ; Li et al ., 2011 , 2013a, 2013b )。地衣由于其形态结构的特殊性和变水植物的特性, 对环境变化非常敏感, 因此地衣多样性的减少经常被用来指示空气污染(Cislaghi & Nimis, 1997 ; Giordani, 2007 )、气候条件(Jovan & McCune, 2004 ; Geiser & Neitlich, 2007 ; Giordani & Incerti, 2008 )、森林结构和动态(Johansson, 2008 ; Nascimbene et al ., 2010 )。但是通过比较地衣多样性来比较不同区域间的环境条件常常遇到困难, 因为环境条件的不同经常形成高度差异的物种组成, 而如果我们利用地衣的功能特征来指示环境条件则可以很好地解决这一问题(Giordani et al ., 2012 )。
根据国外的相关研究, 地衣按照共生藻、生活型和繁殖策略的不同可以划分为蓝藻地衣(cyano-区性lichens, CYL)、壳状地衣(crustose lichens, CRL)、枝状地衣(fruticose lichens, FRL)、狭叶地衣(narrowly lobed foliose lichens, NFL)、阔叶地衣(broadly lobed foliose lichens, BFL)、裂芽地衣(isidiate lichens, ISL)、粉芽地衣(sorediate lichens, SOL)和有性地衣(sexual lichens, SEL) 8个功能群(McCune, 1993 ; Nimis & Martellos, 2008 )。调查研究显示地衣呈现出与它们功能特征相关的分布格局(Ellis & Coppins, 2006 ; Lakatos et al ., 2006 ; Zedda et al ., 2011 )。比如在热带山地雨林分布着更多的枝状和叶状地衣, 但在低地雨林却是壳状地衣占据优势, 枝状地衣和叶状地衣很稀少(Montfoot & Ek, 1990 ; Sipman & Tan, 1990 ; Lakatos et al ., 2004 )。Hauck等(2007)在蒙古的针叶林发现, 绿藻地衣在群落中处于优势地位, 蓝藻地衣只在有着较高可利用水源的地方有分布。在最近的研究中, 地衣的生活型、繁殖策略、共生藻等功能特征被广泛用来指示地衣群落对环境条件的适应(DÍaz & Cabido, 2001 ; Cornelissen et al ., 2007 ; Glime, 2007 ), 因为地衣的功能特征可以不依赖物种多样性和组成而直接和环境因子建立关联(Ricotta & Bacaro, 2010 ; Webb et al ., 2010 )。Lange等(1994)发现在蓝藻地衣较多的生境中经常有高的积水。Giordani等(2012)基于意大利森林生态系统网络研究了地衣功能特征和环境条件的关系, 结果表明: 壳状和叶状地衣的分布和降水量密切相关; 枝状地衣的分布受海拔的影响较大, 同时和森林的密闭度负相关; 狭叶和阔叶地衣都受到酸性沉积物的负面影响。Giordani等(2014)调查发现阔叶地衣只有在高潜在径流的条件下才能在高辐射的生境中分布, 同时他们还观察到在枝状地衣的生境中有高的潜在径流。
在哀牢山国家级自然保护区分布着我国目前保存最完整、面积最大的山地湿性常绿阔叶林(Young & Herwitz, 1995 ; 朱华和闫丽春, 2009 ), 林内分布着丰富的附生地衣。Li等(2013b)在本地区记录到了217种附生地衣。由于人为的干扰, 在本地区原生林的周围形成了栎类萌生林、滇山杨(Populus bonatii )林、旱冬瓜(Alnus nepalensis )林等次生林(游承侠, 1983 ), 从而形成了不同的生境。在不同森林类型及它们之间的过渡带, 附生植物的多样性和丰富度是不一样的(马文章, 2009 ; 姚元林等, 2012 )。Li等(2013a)对本地区附生地衣的调查研究发现, 不同森林类型内附生地衣功能团的盖度和物种丰富度均明显不同, 在滇山杨林中阔叶地衣、蓝藻地衣及枝状地衣等的物种丰富度最高, 而在旱冬瓜林中, 壳状地衣和狭叶地衣的丰富度最高。李苏等(2015)对本地区4种附生蓝藻地衣分布的研究发现, 4种蓝藻地衣中有3种蓝藻地衣的盖度和频度最高值出现于次生林中, 并且4种蓝藻地衣表现出明显对厚皮香(Ternstroemia gymnanthera )、滇山杨和硬壳柯(Lithocarpus hancei )等树种的偏好性, 以及对小花山茶(Camellia forrestii )等的排斥性, 但目前还无法解释本地区附生地衣功能群的分布格局。Zots和Winder (1994)从碳平衡的角度解释了壳状地衣、枝状地衣和叶状地衣在热带地区海拔梯度上的分布格局。Lakatos等(2006)从形态学和生理学的角度解释了壳状绿藻地衣在法属圭亚那热带低地雨林林下存在的机制。所以, 地衣生理生态学研究是用来解释地衣功能群分布格局的有效途径之一。本研究拟在控制条件下分析比较不同附生地衣功能群的持水力和失水速度及光合作用对水分、光照的响应, 以期从内在机理上解释本地区附生地衣功能群的分布格局, 同时为利用附生地衣功能群指示环境变化提供理论依据。
1 材料和方法
1.1 研究区概况
哀牢山位于云贵高原西南部、横断山区南段, 属于云岭山脉向西南分支的余脉。本研究区位于哀牢山北段的徐家坝地区(23.35°-24.44° N, 100.54°- 101.30° E), 海拔2000-2750 m, 该区降雨充沛, 云雾缭绕, 常年温凉潮湿, 保存着我国亚热带地区目前面积(5000多hm2 )最大、以云南特有植物种为优势种的亚热带原始山地湿性常绿阔叶林(Liu et al ., 2002 ; 徐海清和刘文耀, 2005 ), 附生植物非常丰富, 是国家级自然保护区。根据哀牢山生态站多年气象观测结果显示, 年降水量1947 mm, 年蒸发量为1192 mm, 年平均相对湿度85%, 年平均气温11.3 ℃ (李苏等, 2007 )。该地区受到西南季风的影响, 有着明显的干湿季之分, 其中5-10月为雨季, 其降水量占全年降水量的85%以上(刘洋等, 2009 )。
1.2 研究材料
由于生活型和共生藻类型可能控制群落的结构(Ellis & Coppins, 2006 , 2007, 2010 ; Marini et al ., 2011 ), 结合Li等(2013a, 2013b)对本地区附生地衣分布的调查研究, 我们选择更能指示环境状况的蓝藻地衣、阔叶地衣、狭叶地衣、枝状地衣4种地衣功能群作为研究对象, 4种附生地衣功能群的划分依据见表1 。每种附生地衣功能群选择2种常见种(或代表种)作为研究对象。其中蓝藻地衣为网肺衣(Lobaria retigera )和猫耳衣(Leptogium menziesii ), 阔叶地衣为针芽肺衣(Lobaria isidiophora )和皮革肾岛衣(Nephromopsis pallescens ), 狭叶地衣为白绵腹衣(Anzia leucobatoides )和云南袋衣(Hypogymnia yunnanensis ), 枝状地衣为裂髓树花(Ramalina conduplicans )和多花松萝(Usnea florida )。其中网肺衣和猫耳衣多生长于潮湿的生境中, 针芽肺衣、皮革肾岛衣、白棉腹衣和云南袋衣多生长于阳光充足的生境中, 裂髓树花和多花松萝多分布于干燥、高光的生境中(Li et al ., 2013a )。我们在每个物种分布较多的生境中采集各研究物种材料, 带回哀牢山生态站, 清洗干净后, 进行相关实验。
1.3 研究方法
本研究在2015年7-9月期间在哀牢山生态站开展, 此时期正处于哀牢山雨季, 林内外湿度都能达到90%以上, 林内与林外湿度、温度的差异也不大(刘玉洪, 1993 ), 林内、外湿度差异对本实验的影响很小, 高湿环境也能防止地衣过快失水。地衣光合作用的测定在10:00-14:00进行, 此时间段的气温变化小于2 ℃ (刘玉洪, 1993 ), 所以温度变化对地衣光合作用的影响也很小。
1.3.1 不同附生地衣功能群的水分关系
不同附生地衣功能群的最大含水量(maximum water content, MWC )和失水速率可用来反映其水分关系。在测量MWC 时, 每种附生地衣各选择10个样品进行试验, 将各样品置入去离子水中, 保证其完全浸没, 1 h后取出地衣样品, 用滤纸吸去样品表面的多余水分, 利用精度0.001 g的电子天平(AL204- IC, Mettler Toledo Instruments, 上海)测量各样品鲜质量, 最后将样品放于80 ℃烘箱中烘干48 h, 重新称干质量, 用所测得的样品鲜质量减去干质量即为样品的最大含水量。
测量各附生地衣功能群失水速率时, 每种附生地衣各选择5个样品, 在去离子水中浸没1 h, 之后取出地衣样品, 用滤纸吸去其表面多余水分后称质量, 然后将样品静置于温度约20 ℃、光照约30 µmol·m-2 ·s-1 的恒定环境中, 让其自然失水, 每隔1 h称一次质量, 当样品质量不再变化时停止测量。最后将所有样品放于80 ℃烘箱中烘干48 h后称取干质量。
1.3.2 不同附生地衣功能群光合作用对水分变化的响应
每种附生地衣各选择5个样品, 将地衣样品淹没进去离子水中, 1 h后取出样品, 用滤纸吸去其表面多余水分后, 利用LI-6400便携式光合测定系统(LI-COR, Lincoln, USA)测定各样品的光合速率(光照设为800 µmol·m-2 ·s-1 )。在每次测量达到平衡后(3-5 min)立即记录净光合速率(P n ), 并马上取出称质量, 在空气中干燥30 min后重新放入叶室进行下一次测量, 如此反复直到地衣质量不再变化时停止测量。测量完成后利用V700扫描仪(Seiko Epson, Nagano, Japan)扫描各附生地衣样品, 扫描时利用平滑的玻璃尽量将样品压平整, 然后用根系分析系统(WhiRHIZO, Regent, Canada)分析扫描图片, 得出各地衣样品的投影面积, 其中枝状地衣中的多花松萝没有背腹结构, 所以取它的表面积。最后将样品放于80 ℃烘箱中烘干48 h后称质量。
1.3.3 不同附生地衣功能群光合作用对光强变化的响应
根据不同附生地衣功能群光合作用的水分响应测量结果, 在各附生地衣含水量稍高于其光合作用最适水分含量的条件下进行光合作用光响应曲线的测定, 以此来反映各附生地衣功能群对光照的响应。每种附生地衣各选择5个样品, 为了不损伤地衣样品, 利用硅胶干燥剂干燥48 h至恒质量。然后将地衣样品淹没进去离子水中, 1 h后取出样品, 用滤纸吸去其表面多余水分后称质量, 这样就可以计算出样品的大致含水量。在样品含水量稍高于其光合作用最适水分含量时, 利用LI-6400测量其光响应曲线。光响应曲线的光强梯度设置为0、20、50、100、150、200、400、600、800、1000、1200、1400 µmol·m-2 ·s-1 , 每个光照梯度保持3 min, 以保证气体交换达到稳定状态后换到下一个光照水平, 每次测量前后都对样品称质量, 以确保整个测量过程都在最适水分 条件左右进行。测量完成后测量各地衣样品的表 面积。
1.4 数据处理和分析
不同附生地衣单位干质量持水量的差异利用单因素方差分析和Tukey’s HSD多重配对进行分析, 并利用线性模型(GLM)来分析附生地衣物种、干质量和持水力(WHC)之间的关系(Gauslaa & Coxson, 2011 ; Esseen et al ., 2015 )。各附生地衣的失水速率利用指数递减回归模型(Song et al ., 2015 )进行拟合:
WC = y 0 + a e- bt
式中WC 是样品相对含水量(%), 是绝对含水量占干质量的百分比。t 是水分散失时间(h), y 0 、a、b 是回归系数。样品MWC (t = 0)为y 0 + a 。根据Pintado等(1997)的研究, 通过含水量减少到地衣最大含水量50%和20%所需的时间来反映各附生地衣的失水速率, 为了更好地反映整个失水过程, 我们增加了含水量减少到MWC 80%所需的时间, 并据此绘制水合状态和时间的关系图。
各附生地衣光合作用和相对含水量之间的关系利用三次多项式回归模型进行拟合(Zotzet al ., 2003 ):
P n = y 0 + aWC + bWC 2 + cWC 3
式中P n 是净光合速率(μmol·m-2 ·s-1 ), y 0 、a、b、c 是回归系数。根据回归方程求得光合作用最适水分含量为 P n = 0时的水分含量)。为了更好地说明光合作用和水分含量的关系, 我们参照Lange (1980) 的处理方法, 利用上述方程求出各附生地衣在水分补偿点、最适水分含量及最大含水量10%、20%、30%、40%、50%、60%时的光合速率, 同样利用三次多项式回归模型模拟相对光合速率(光合速率占最适水分含量下光合速率的百分比)和水分含量的关系, 并求出模型曲线的初始斜率, 用来说明附生地衣在干燥状态下对水分含量变化的敏感性。
植物光合作用对光响应模型的研究进行得比较早, 已经研究出许多经典的模型。其中直角双曲线修正模型不但可以拟合植物在饱和光强之后光合速率不下降的光响应曲线, 而且还可以拟合植物在饱和光强之后光合速率随光强的增加而下降这一段的光响应曲线, 且估算的饱和光强和最大净光合速率与实测值最为接近(Ye, 2007 ; 叶子飘, 2010 ; Song et al ., 2015 )。所以我们利用直角双曲线修正模型来拟合各附生地衣光合作用和光照强度的关系:
式中I 是光照强度(μmol·m-2 ·s-1 ), R d 暗呼吸速率(μmol·m-2 ·s-1 ), a、b、c 是回归系数。主要的光合作用特征参数根据叶子飘(2010)进行计算, 包括光照饱和时的光合速率(P nmax ; μmol·m-2 ·s-1 )、光饱和点(LSP; μmol·m-2 ·s-1 )、光补偿点(LCP; μmol·m-2 ·s-1 ), 其中我们参照Lange等(1991) 将光饱和点定义为净光合速率达到最大净光合速率90%时的最小光强。不同附生地衣光响应曲线参数的差异性利用单因素方差分析和Tukey’s HSD多重配对进行分析。
所有数据利用Excel 2013来进行整理, 所有的统计分析和图形绘制都是利用SigmaPlot 12.5进行。
2 结果
2.1 不同功能群附生地衣的水分关系
每种附生地衣的持水量和其干质量表现出线性关联(图1 , 相关系数R 2 为0.65-0.98), 随着地衣干质量的增加, 其含水量表现出线性的增加。不同附生地衣单位干质量的持水量存在差异(表2 ), 比如猫耳衣1 g干质量的持水量能达到10.49 g, 而针芽肺衣只有2.84 g。附生地衣单位干质量的持水量和功能特征之间并没有表现出明显的相关关系, 其中蓝藻地衣和枝状地衣的两种附生地衣间单位干质量持水量存在显著差异, 而阔叶地衣和狭叶地衣的两种附生地衣间单位干质量持水量相差不大, 其中狭叶地衣单位干质量的持水量将近阔叶地衣的2倍。作为蓝藻地衣的猫耳衣其持水量能够达到其干质量的10倍, 另外, 作为同属的两种附生地衣, 属于蓝藻地衣的网肺衣比属于绿藻地衣的针芽肺衣的单位干质量持水量要高出许多, 可见蓝藻地衣有着较高的持水能力。指数递减模型很好地模拟了各附生地衣的失水动力学过程(图2 , R 2 为0.87-0.95), 不同附生地衣的失水动力学曲线是不同的, 可以看到多花松萝、裂髓树花等在开始时失水很快, 后来失水速度逐渐减缓, 而白绵腹衣的失水速度比较均匀, 这点在图3 上显示得更明显。通过比较不同附生地衣含水量减少到最大含水量80%、50%及20%所需的时间(表2 )可以得知, 多花松萝的失水速率最快, 1 h左右其含水量就下降到最大含水量的一半, 而白绵腹衣则需将近5 h其含水量才下降到最大含水量的一半。比较各附生地衣含水量从80%减少到50%和从50%减少到30%所需时间的差异, 发现白绵腹衣仅差0.41 h, 而多花松萝相差1.02 h, 其他附生地衣相差为0.71-0.93。就同一功能群而言, 两种阔叶地衣的失水速率相差不大, 两种蓝藻地衣和狭叶地衣相差较大。总体上枝状地衣的失水速率要比其他附生地衣功能群失水速率快。
图1 各附生地衣干质量和持水量之间的关系(n = 10)。
Fig. 1 Relationship between water-holding capacity (WHC ) and dry mass in different epiphytic lichens (n = 10).
图2 不同附生地衣失水动力学曲线。A, 皮革肾岛衣。B, 针芽肺衣。C, 网肺衣。D, 猫耳衣。E, 云南袋衣。F, 白绵腹衣。G, 多花松萝。H, 裂髓树花。WC 表示含水量。不同形状的图列符号表示不同的样品(n = 3-5)。
Fig. 2 Water release curves for different epiphytic lichens. A, Nephromopsis pallescens . B, Lobaria isidiophora . C, Lobaria retigera . D, Leptogium menziesii . E, Hypogymnia yunnanensis . F, Anzia leucobatoides . G, Usnea florida . H, Ramalina conduplicans . WC in graphs refers to water content. Different shapes of symbols represent different samples (n = 3-5).
2.2 不同功能群附生地衣光合作用对水分变化的响应
各附生地衣的光合速率对水分变化的响应趋势是相同的(图4 ), 均表现为先随含水量的增加而升高, 当含水量达到一定程度后, 光合速率随含水量的增加而降低。但是, 高水分含量对不同附生地衣光合作用的影响程度是不同的, 其中对皮革肾岛衣的影响最小, 而对针芽肺衣、网肺衣、云南袋衣、多花松萝光合速率的抑制能达到最大光合速率的50%, 甚至更多。利用三次多项式回归模型能够较好地模拟各附生地衣光合速率随水分含量的变化(图4 , R 2 为0.73-0.93)。根据模型方程可以计算出各附生地衣光合作用的最适水分含量、水分补偿点。不同附生地衣光合作用的最适水分含量是存在差异的, 其中针芽肺衣、多花松萝、裂髓树花的最适含水量在100%左右, 而猫耳衣和白棉腹衣则超过了300% (表3 )。附生地衣光合速率的最适含水量近似线性(图5 , R 2 = 0.63)地随最大含水量增加。而光合作用的水分补偿点和最适水分含量无明显的关联性(图5 ), 最适含水量较高的猫耳衣和白绵腹衣的水分补偿点反而较低, 不到干质量的20%, 而网肺衣的水分补偿点将近达到干质量的50%, 分布于较干旱生境中枝状地衣的水分补偿点则比较高, 都达到其干质量的30%左右(表3 )。相对净光合速率(净光合速率占最大净光合速率的百分比)和水分含量关系曲线(图5 )的初始斜率能在一定程度上说明附生地衣在干燥情况下光合作用对水分的敏感性(Lange, 1980 )。在本研究中, 光合作用最适水分含量相对较高的白绵腹衣和猫耳衣的初始斜率(0.5-0.64)都比较低, 反而最适水分含量较低的针芽肺衣、多花松萝和裂髓树花的初始斜率(1.92-2.59)较高, 说明在干燥状态下它们的光合作用对水分变化较敏感。
图3 不同附生地衣失水过程中不同时刻的含水量。含水量表示为地衣含水量占最大含水量的百分比。
Fig. 3 Water content in different epiphytic lichens at different times during water loss. Hydration (%) refers to the percentage of maximum water content.
图4 不同附生地衣光合作用水分响应曲线。A, 皮革肾岛衣。B, 针芽肺衣。C, 网肺衣。D, 猫耳衣。E, 云南袋衣(R 2 = 0.83, p < 0.001)。F, 白绵腹衣。G, 多花松萝。H, 裂髓树花。P n 和WC 分别表示净光合速率和含水量。不同形状的图列符号表示不同的样品(n = 4, 5)。
Fig. 4 Photosynthetic water response curves for different epiphytic lichens. A, Nephromopsis pallescens . B, Lobaria isidiophora . C, Lobaria retigera . D, Leptogium menziesii . E, Hypogymnia yunnanensis . F, Anzia leucobatoides . G, Usnea florida . H, Ramalina conduplicans . P n and WC in graphs refer to net photosynthesis rate and water content, respectively. Different shapes of symbols represent different samples (n = 4, 5).
图5 附生地衣光合最适水分含量和最大水分含量的关系(A), 水分补偿点和最适水分补偿点的关系(B), 水分含量和光合作用的关系(C)。图C中净光合速率比表示为占最大净光合速率的百分比。
Fig. 5 The relationship between optimal water content and maximum water content (A), the relationship between water compensation point and optimal water content (B), and the relationship between photosynthesis and water content in different epiphytic lichens (C). Photosynthesis is expressed as the percentage of max net photosynthesis.
2.3 不同功能群附生地衣光合作用对光强变化的响应
通过光合作用光响应曲线(图6 , R 2 为0.95-0.98) 可以看到净光合速率在开始时几乎呈线性地增加, 当光照达到400 μmol·m-2 ·s-1 时, 净光合速率就能达到最大净光合速率的80%以上, 之后光合速率平稳地增加, 并且高光照对各附生地衣的光合作用没有明显的抑制作用。根据光响应曲线的拟合方程计算出各附生地衣光响应曲线参数。通过光响应曲线参数可以看到蓝藻地衣的光补偿点约为20 μmol·m-2 ·s-1 , 而其他功能群附生地衣约为40 μmol·m-2 ·s-1 , 两者之间存在显著性差异。枝状地衣的光饱和点(分别为610.9和613.4 μmol·m-2 ·s-1 )要显著高于其他功能群附生地衣(322.4-466.4 μmol·m-2 ·s-1 )。附生地衣间最大光合速率的差异更多体现在物种之间, 两两之间都存在显著性差异, 而和功能特征没有太大关联。在呼吸速率上, 两种枝状地衣存在显著性差异, 其他附生地衣功能群内两种附生地衣的呼吸速率差异都不显著, 同样呼吸速率的差异和功能特征的关联也不大。
图6 各附生地衣光响应曲线。A, 皮革肾岛衣。B, 针芽肺衣。C, 网肺衣。D, 猫耳衣。E, 云南袋衣(R 2 = 0.83, p < 0.001)。F, 白绵腹衣。G, 多花松萝。H, 裂髓树花。不同形状的图列符号表示不同的样品(n = 4, 5)。
Fig. 6 Photosynthetic light response curves for different epiphytic lichens. A, Nephromopsis pallescens . B, Lobaria isidiophora . C, Lobaria retigera . D, Leptogium menziesii . E, Hypogymnia yunnanensis . F, Anzia leucobatoides . G, Usnea florida . H, Ramalina conduplicans . P n and I in graphs refer to net photosynthesis and light intensity, respectively. Different shapes of symbols represent different samples (n = 4, 5).
3 讨论
作为变水植物, 地衣的含水量随着环境条件迅速变化, 而其所有的代谢活动, 特别是净光合速率和呼吸速率也随之发生变化(Green & Lange, 1994 )。所以地衣水分关系的研究格外受到关注(Galun, 1988 )。地衣不能主动地调节水分的吸收、储存和流失(Green et al ., 2011 ), 它们利用形态学特征来促进叶状体从不同来源(雨水、潮湿空气、露水)吸收和贮存水分(Gauslaa & Coxson, 2011 )。形态结构和解剖结构是影响地衣水分贮藏的重要因素(Larson & Kershaw, 1976 ; Larson, 1981 ; Sancho & Kappen, 1989 )。在本研究中, 附生地衣的最大含水量和功能特征相关性不大, 属于同一功能团的两种附生地衣间的最大含水量也可能相差很大, 而枝状地衣的最大含水量也可能比叶状地衣的大, 这就说明解剖结构可能在附生地衣持水量上发挥了更重要的作用。Gauslaa和Coxson (2011)认为共生藻层的厚度是影响地衣物种间持水量的重要因素, 而共生藻的类型是影响其厚度的主要因素。我们所研究的两种蓝藻地衣都有着较高的持水量, 尤其是猫耳衣, 其持水量明显高于其他附生地衣。而网肺衣的持水量也比同属的针芽肺衣高100%左右。蓝藻地衣之所以有较高的持水量是因为在它们的共生藻细胞外有一层较厚的凝胶状保护鞘(Honegger et al ., 1996 ; Gauslaa & Coxson, 2011 ), 能够帮助蓝藻地衣贮存更多的水分。水分贮存能力、形态结构、解剖结构以及颜色是影响地衣吸收和丢失水分的主要因素(Palmqvist, 2000 )。比如, 相比于叶状地衣, 枝状地衣和环境之间的水分交换更快(Hartard et al ., 2009 )。在本研究中也是枝状地衣的失水速度要快于其他功能群附生地衣。枝状地衣的这种形态特征也使得它们能够从湿润的空气中快速吸收水分(Lange & Ziegler, 1986 ), 也就使得它们能够适应林外干燥的环境。相对于枝状地衣, 叶状地衣则是通过增大持水量和水分贮存能力来保持体内的水分含量从而延长生理活动的时间(Lange & Ziegler, 1986 ), 比如我们所研究的白绵腹衣, 其失水速度要明显低于其他附生地衣。对于蓝藻地衣而言, 由于只能利用液态水来进行光合作用(Lange & Ziegler, 1986 ), 所以它们通过凝胶状物质来增加持水量。
水分含量和光照是决定地衣光合速率的主要因素(Lange, 2002 )。作为变水植物, 地衣的生理代谢, 特别是光合作用, 受到水分含量的强烈影响, 过低和过高的水分含量都将限制地衣的光合作用(Lange et al ., 1993 )。Zotz等(1998)在对热带阔叶地衣Sticta tomentosa 气体交换的研究中发现, 在最大含水量时, 其净光合速率比最大光合速率的50%还小。Lange (2002)在对石附生地衣Lecanora muralis 的研究中发现, 在干燥和过饱和状态下, Lecanora muralis 的净光合速率都等于0或接近于0。在本研究中, 我们同样发现过高或过低的含水量都明显地限制着各附生地衣的光合作用。在陈克等(2014)对本地区3种附生地衣的光合生理研究中, 网肺衣在含水量为150%-200%时拥有最适光合水分条件, 这和我们所得到的网肺衣最适光合含水量为172.9%相符。过高含水量对不同附生地衣光合作用的抑制程度是不同的, 其中对云南袋衣的限制作用最明显, 而对皮革肾岛衣的影响则比较小, 这可能是由于不同附生地衣适应环境的策略不同造成的。云南袋衣可能通过增加叶状体厚度及扩大细胞间隙来增加持水量和降低失水速率, 这样带来的结果是细胞间隙间过多的水分阻碍了CO2 扩散, 从而影响了光合作用。皮革肾岛衣则可能通过增加表面积来加快叶状体和环境水分的交换, 这样就能降低过多水分对CO2 扩散的影响, 但是也降低持水量和加快了失水速率。通过分析各附生地衣光合作用的最适含水量, 枝状地衣具有较低的光合作用最适含水量, 这可能与它们分布于干燥生境有密切关系, 并且我们的研究还发现枝状地衣在干燥状态下对水分变化很敏感, 这就意味
着枝状地衣能够在林外相对较干的生境中迅速有效地利用昼夜温差所形成的露水。对于蓝藻地衣, 较高的光合最适含水量和只能利用液态水进行光合作用的特征可能是它们较多分布于湿润地区的主要 原因。
地衣的变水特征使得它们的光响应特征比较复杂。地衣水分含量和净光合速率(P n )的关系能够改变它们的P n 对光照的响应(Lange & Matthes, 1981 )。低的水分含量使得地衣的P n 获得低数值的光补偿点(LCP )及光饱和点(LSP ), 而高水分含量对P n 的抑制作用也能够降低地衣的LSP 值, 但对LCP 的影响不大(Lange et al ., 1996 , 2004 ; Green et al ., 1997 ; 陈克等, 2014 )。Green等(1997)对新西兰热带雨林9种地衣光适应的研究指出, 光响应曲线参数和所测定地衣的生境相关。他们的研究发现, 林内地衣的光饱和点要低于300 μmol·m-2 ·s-1 , 而在开阔生境中地衣的光饱和点要大于549 μmol·m-2 ·s-1 。在本研究中, 8种附生地衣的LSP 都比较高(333.9-616.0 μmol·m-2 ·s-1 , 表4 ), 并且即使光强高于1000 μmol·m-2 ·s-1 也没有明显的光抑制现象, 表现出明显的“阳生植物”特性, 这在一定程度上解释了为什么在哀牢山地区附生地衣在次生林比在原生林中有着更高的丰富度(李苏等, 2007 ; Li et al ., 2013a , 2013b )。就各功能群附生地衣的LSP 而言, 枝状地衣的LSP 要明显高于其他功能群附生地衣, 这也很好地解释了它们在林冠上层和林外空旷地有较高多样性的分布格局(Li et al ., 2015 )。Green等(1997)和陈克等(2014)的研究中都指出相比于绿藻地衣, 蓝藻地衣有着较低的LSP 。在我们的研究中虽然网肺衣也有着较高的LSP, 但是猫耳衣的LSP 还是低于其他功能群附生地衣。LCP 是衡量植物利用弱光能力的重要指标。在我们的研究中, 8种附生植物的LCP 都比较高(20.1-43.1 μmol·m-2 ·
s-1 , 表4 ), 除了蓝藻地衣外, 其他功能群附生地衣的LCP 都要高于Green等(1997)在新西兰热带雨林所研究的林内林缘地衣LCP (4.3-29 μmol·m-2 ·s-1 ), 而低于林外地衣LCP (≥50 μmol·m-2 ·s-1 )。在本研究中, 蓝藻地衣的LCP 显著低于其他功能群的附生地衣, 这和在哀牢山地区观察到的蓝藻地衣能分布在较阴暗生境相符(Li et al ., 2015 )。总之, 蓝藻地衣能够利用的光谱比较宽(较低的LCP 和较高的LSP ), 所以蓝藻地衣既能分布在较强太阳辐射的生境中, 又对弱光有一定的适应能力(Rikkien, 2009 ), 而绿藻地衣大多只能分布在太阳辐射较高的生境中。地衣的光合能力和其氮素含量密切相关(Lange et al ., 2004 )。在Lange等(2004)对巴拿马热带雨林6种地衣碳获取的研究中, 具有高浓度氮含量的牛皮叶属地衣的P n (4.9-5.8 μmol·m-2 ·s-1 )要比肺衣属的(2.0-2.9 μmol·m-2 ·s-1 )高很多。我们所研究的8种附生地衣的P n 都比较小(0.92-2.84 μmol·m-2 ·s-1 , 表4 ), 两种肺衣属(针芽肺衣和网肺衣)地衣的P n (分别为1.77 和2.84 μmol·m-2 ·s-1 )和Lange等(2004)测得的同属地衣的P n 相差不大, 并且属于蓝藻地衣的网肺衣的 P n 要比属于绿藻地衣的针芽肺衣的高, 这可能和网肺衣具有固氮能力有关。光合能力的差异和功能群的相关性不大, 可能更多地和地衣氮浓度含量相关。
不同附生地衣对环境有着自己的适应方式, 还表现出与其功能特征相关。作为变水植物, 附生地衣生理活动的强弱依赖于水分含量的多少。蓝藻地衣能通过增加持水量来延长生理活动时间; 枝状地衣能够快速地与环境进行水分交换进而延长生理活动时间; 狭叶和阔叶地衣则因物种而异, 可能通过增加持水力, 也可能通过加快和环境的水分交换速率延长生理活动时间。蓝藻地衣的光合生理活动对水分条件要求较高, 它们的光合最适水分含量较高, 所以它们广泛分布于潮湿的生境中。同时蓝藻地衣还有着低的光补偿点和高的光饱和点, 使得它们对光照条件有着较高的容忍度, 能够同时分布在高光和弱光生境中。枝状地衣的光合生理活动对光照条件要求比较高, 它们的光补偿点和光饱和点都很高, 而光合最适水分含量较低, 所以它们广泛分布于光照很强而相对干燥的生境中。阔叶和狭叶地衣的光合活动需要一定的水分条件, 并且它们的光补偿点比较高, 所以它们广泛分布于光照充足的生境中(如林冠外层)。不同功能群附生地衣在更小和更大尺度上分布的差异还需要进行更为深入的研究, 比如吸水速率、呼吸和水分含量的关系、生理活动和温度的关系等, 同时还需要研究不同功能群附生地衣在不同生境中生理活动时间的长短, 及对不同生境的适应能力。但是从我们的研究可以得知不同附生地衣功能群的生理特征存在一定的差异, 这样使得我们能够从生理生态学的角度去解释不同功能群附生地衣的分布格局, 如果结合不同功能群附生地衣生理特性对环境变化的响应, 我们就能够预测在全球变化背景下附生地衣分布格局的变化。
致谢 感谢中国科学院西双版纳热带植物园付培立博士在仪器使用方面给予的帮助, 同时感谢哀牢山生态站武传胜博士在野外工作中给予的帮助。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Aptroot A Lichen biodiversity in Papua New Guinea, with the report of 173 species on one tree. In: Türk R, Zore R eds. Progress and Problems in Lichenology in the Nineties . Bibliotheca Lichenologica , Berlin, Germany . 203 -213 .
[2]
Aptroot A Sipman HJM Diversity of lichenized fungi in the tropics. In: Hyde KD ed. Biodiversity of Tropical Microfungi . Hong Kong University Press , Hong Kong, China . 93 -106 .
[3]
Chen K Liu WY Li S Song L 2014 ). Photosynthetic characteristics of three epiphytic lichens under different water conditions.
Plant Diversity and Resources , 36 , 603 -610 . (in Chinese with English abstract)[陈克 , 刘文耀 , 李苏 , 宋亮 (2014 ). 不同水分条件下三种附生地衣的光合作用特性
. 植物分类与资源学报 , 36 , 603 -610 .]
[4]
Cislaghi C Nimis PL 1997 ). Lichens, air pollution and lung cancer.
Nature , 387 , 463 -464 .
[本文引用: 1]
[5]
Cornelissen JHC Lang SI Soudzilovskaia NA During HJ 2007 ). Comparative cryptogam ecology: A review of bryophyte and lichen traits that drive biogeochemistry.
Annals of Botany , 99 , 987 -1001 .
[本文引用: 1]
[6]
Cornelissen JHC Steege HT 1989 ). Distribution and ecology of epiphytic bryophytes and lichens in dry evergreen forest of Guyana.
Journal of Tropical Ecology , 5 , 29 -35 .
[本文引用: 2]
[7]
DÍaz S Cabido M 2001 ). Vive la difference: Plant functional diversity matters to ecosystem processes.
Tends in Ecology & Evolution , 16 , 646 -655 .
[8]
Ellis CJ Coppins BJ 2006 ). Contrasting functional traits maintain lichen epiphyte diversity in response to climate and autogenic succession.
Journal of Biogeography , 33 , 1643 -1656 .
[本文引用: 1]
[9]
Ellis CJ Coppins BJ 2007 ). Changing climate and historic-woodland structure interact to control species diversity of the ‘Lobarion’ epiphyte community in Scotland.
Journal of Vegetation Science , 18 , 725 -34 .
[本文引用: 2]
[10]
Ellis CJ Coppins BJ 2010 ). Integrating multiple landscape- scale drivers in the lichen epiphyte response: Climatic setting, pollution regime and woodland spatial-temporal structure.
Diversityand Distributions , 16 , 43 -52 .
[本文引用: 1]
[11]
Esseen PA Olsson T Coxson D Gauslaa Y 2015 ). Morphology influences water storage in hair lichens from boreal forest canopies.
Fungal Ecology , 18 , 26 -35 .
[12]
Galun M Handbook of Lichenology . CRC Press , Boca Raton, USA .
[本文引用: 2]
[13]
Gauslaa Y Coxson DS 2011 ). Interspecific and intraspecific variations in water storage in epiphytic old forest foliose lichens.
Botany , 89 , 787 -798 .
[本文引用: 1]
[14]
Geiser LH Neitlich PN 2007 ). Air pollution and climate gradients in western Oregon and Washington indicated by epiphytic macrolichens.
Environment Pollution , 145 , 203 -218 .
[本文引用: 3]
[15]
Giordani P 2007 ). Is the diversity of epiphytic lichens a reliable indicator of air pollution? A case study from Italy.
Environmental Pollution , 146 , 317 -323 .
[本文引用: 1]
[16]
Giordani P Brunialti G Bacaro G Nascimbene J 2012 ). Functional traits of epiphytic lichens as potential indicators of environmental conditions in forest ecosystems.
Ecological Indicators , 18 , 413 -420 .
[本文引用: 1]
[17]
Giordani P Incerti G 2008 ). The influence of climate on the distribution of lichens: A case study in a borderline area (Liguria, NW Italy).
Plant Ecology , 195 , 257 -272 .
[本文引用: 1]
[18]
Giordani P Incerti G Rizzl G Rellini I Nimis PL Modenesi P 2014 ). Functional traits of cryptogams in Mediterranean ecosystems are driven by water, light and substrate interactions.
Journal of Vegetation Science , 25 , 778 -792 .
[本文引用: 1]
[19]
Glime JM
URL
[本文引用: 1]
[20]
Goward T Arsenault A 2000 ). Cyanolichen distribution in young unmanaged forests: A dripzone effect?
The Bryologist , 103 , 28 -37 .
[本文引用: 1]
[21]
Gradstein SR The vanishing tropical rain forest as an environment for bryophytes and lichens. In: Bates JW, Farvner AM eds. Bryophytes and Lichens in a Changing Environment . Clarendon Press , Oxford, UK . 234 -258 .
[本文引用: 1]
[22]
Green TGA Büdel B Meyer A Zellner H Lange OL 1997 ). Temperate rainforest lichens in New Zealand: Light response of photosynthesis.
New Zealand Journal of Botany , 35 , 493 -504 .
[本文引用: 1]
[23]
Green TGA Lange OL Photosynthesis of poikilohydric plants: Lichens and bryophytes—A comparison. In: Schulze DE, Caldwell MM eds . Ecophysiology of Photosynthesis . Springer-Verlag, Berlin . 319 -342 .
[本文引用: 1]
[24]
Green TGA Sancho LG Pintado A Ecophysiology of desiccation/rehydration cycles in mosses and lichens. In: Lüttge U, Beck E, Bartels D eds . Plant Desiccation Tolerance . Springer-Verlag, Berlin, Germany . 89 -120 .
[25]
Gustafsson L Eriksson I 1995 ). Factors of importance for the epiphytic vegetation aspen Populus tremula with special emphasis on chemistry and soil chemistry.
Journal of Applied Ecology , 32 , 412 -424 .
[本文引用: 1]
[26]
Hartard B Cuntz M Maguas C Lakatos M 2009 ). Water isotopes in desiccating lichens.
Planta , 231 , 179 -193 .
[本文引用: 1]
[27]
Hauck M Dulamsuren C Mühlenberg M 2007 ). Lichen diversity on steppe slopes in the northern Mongolian mountain taiga and its dependence on microclimate.
Flora , 202 , 530 -546 .
[28]
Hauck M Spribille T 2005 ). The significance of precipitation and substrate chemistry for epiphytic lichen diversity in spruce-fir forests of the Salish Mountains, northwestern Montana.
Flora , 200 , 547 -562 .
[本文引用: 2]
[29]
Holz I Gradstein SR 2005 ). Cryptogamic epiphytes in primary and recovering upper montane oak forests of Costa Rica—Species richness, community composition and ecology.
Plant Ecology , 178 , 89 -109 .
[本文引用: 1]
[30]
Honegger R Peter M Scherrer S 1996 ). Drought-induced structural alterations at the mycobiont-photobiont interface in a range of foliose macrolichens.
Protoplasma , 190 , 221 -232 .
[本文引用: 1]
[31]
Johansson P 2008 ). Consequences of disturbance on epiphytic lichens in boreal and near boreal forests.
Biological Conservation , 141 , 1933 -1944 .
[本文引用: 1]
[32]
Jovan S McCune B 2004 ). Regional variation in epiphytic macrolichen communities in northern and central California forests.
Bryologist , 107 , 328 -339 .
[本文引用: 1]
[33]
Lakatos M Lange-Bertalot H Büdel B 2004 ). Diatoms living inside the thallus of the green algal lichen Coenogonium linkii in neotropical lowland rain forests.
Journal of Phycology , 40 , 70 -73 .
[本文引用: 1]
[34]
Lakatos M Rascher U Büdel B. 2006 ). Functional character- istics of corticolous lichens in the understory of a tropical lowland and rain forest.
New Phytologist , 172 , 679 -695 .
[本文引用: 1]
[35]
Lange OL 1980 ). Moisture content and CO2 exchange in lichens. I. Influence of temperature on moisture dependent net photosynthesis and dark respiration in Ramalina maciformis.
Oecologia , 45 , 82 -87 .
[本文引用: 2]
[36]
Lange OL 2002 ). Photosynthetic productivity of the epilithic lichen Lecanora muralis: Long-term field monitoring of CO2 exchange and its physiological interpretation. I. Dependence of photosynthesis on water content, light, temperature, and CO2 concentration from laboratory measurements.
Flora , 197 , 233 -249 .
[本文引用: 1]
[37]
Lange OL Büdel B Heber U Meyer A Zellner H Green TGA 1993 ). Temperate rainforest lichens in New Zealand: High thallus water content can severely limit photosynthetic CO2 exchange.
Oecologia , 95 , 303 -313 .
[本文引用: 1]
[38]
Lange OL Büdel B Meyer A Zellner H Zotz G 2004 ). Lichen carbon gain under tropical conditions: Water relations and CO2 exchange of three Lobariaceae species of a lower montane rainforest in Panama.
Lichenologist , 36 , 329 -334 .
[本文引用: 2]
[39]
Lange OL Green TGA Reichenberger H Meyer A 1996 ). Photosynthetic depression at high thallus water content in lichens: Concurrent use of gas exchange and fluorescence techniques with a cyanobacterial and a green algal Peltigera species.
Botanica Acta , 109 , 43 -50 .
[本文引用: 1]
[40]
Lange OL Matthes U 1981 ). Moisture-dependent CO2 exchange of lichens.
Photosynthetica , 15 , 555 -574 .
[本文引用: 1]
[41]
Lange OL Meyer A Ullmann I Zellner H 1991 ). Mikroklima, Wassergehalt und Photosynthese von Flechten in der küstennahen Nebelzone der Namib-Wüste: Messungen während der herbstlichen Witterungsperiod.
Flora , 185 , 233 -266 .
[本文引用: 1]
[42]
Lange OL Meyer A Zellner H Heber U 1994 ). Photosynthesis and water relations of lichen soil crusts—Field measurements in the coastal fog zone of the Namib Desert.
Functional Ecology , 8 , 253 -264 .
[43]
Lange OL Ziegler H Different limiting processes of photosynthesis in lichens. In: Marcelle R, Clijsters H, van Poucke M eds. Biological Control of Photosynthesis . Martinus Nijhoff Publishers , Dordrecht . 147 -161 .
[本文引用: 3]
[44]
Larson DW 1981 ). Differential wetting in some lichens and mosses: The role of morphology.
The Bryologist , 84 , 1 -15 .
[本文引用: 1]
[45]
Larson DW Kershaw KA 1976 ). Studies on lichen-dominated systems XVIII. Morphological control of evaporation in lichens.
Canadian Journal of Botany , 54 , 2061 -2073 .
[本文引用: 1]
[46]
Li S Liu WY Li DW 2013 a). Bole epiphytic lichens as potential indicators of environmental change in subtropical forest ecosystems in southwest China.
Ecological Indicators , 29 , 93 -104 .
[本文引用: 3]
[47]
Li S Liu WY Li DW 2013 b). Epiphytic lichens in subtropical forest ecosystems in southwest China: Species diversity and implications for conservation.
Biological Conservation ,159 , 88 -95 .
[本文引用: 1]
[48]
Li S Liu WY Shi XM Liu S Hu T Huang JB Chen X Song L Wu CS 2015 ). Responses of the distribution of four epiphytic cyanolichens to habitat change in subtropical forests.
Chinese Journal of Plant Ecology , 39 , 217 -228 . (in Chinese with English abstract)[李苏 , 刘文耀 , 石贤萌 , 柳帅 , 胡涛 , 黄俊彪 , 陈曦 , 宋亮 , 武传胜 (2015 ). 亚热带森林生态系统4种附生蓝藻地衣的分布对生境变化的响应
. 植物生态学报 , 39 , 217 -228 .]
[本文引用: 2]
[49]
Li S Liu WY Wang LS Ma WZ Song L 2011 ). Biomass, diversity and composition of epiphytic macrolichens in primary and secondary forests in the subtropical Ailao Mountains, southwest China.
Forest Ecology and Management , 261 , 1760 -1770 .
[50]
Li S Liu WY Wang LS Yang GP Li DW 2007 ). Species diversity and distribution of epiphytic lichens in the primary and secondary forests in Ailao Mountain, Yunnan.
Biodiversity Science , 15 , 445 -455 . (in Chinese with English abstract)[李苏 , 刘文耀 , 王立松 , 杨国平 , 李达文 (2007 ). 云南哀牢山原生林及次生林群落附生地衣物种多样性与分布
. 生物多样性 , 15 , 445 -455 .]
[本文引用: 1]
[51]
Liu WY Fox JED Xu ZF 2002 ). Nutrient fluxes in bulk precipitation, throughfall and stemflow in montane moist evergreen broad-leaved forest on Ailao Mountain in Yunnan, SW China.
Journal of Tropical Ecology , 18 , 527 -548 .
[52]
Liu Y Zhang YP Liu YH Gao F Gong HD 2009 ). Climate change from 1980 to 2005 in the Ailao Mountains, Southwest China.
Journal of Mountain Research , 27 , 203 -210 . (in Chinese with English abstract)[刘洋 , 张一平 , 刘玉洪 , 高富 , 巩合德 (2009 ). 哀牢山北段地区气候特征及变化趋势
. 山地学报 , 27 , 203 -210 .]
[本文引用: 3]
[53]
Liu YH 1993 ). Study on climate characteristics of evergreen broad-leaf forest on Ailao Mountain.
Scientia Silvae Sinicae , 29 , 547 -552 . (in Chinese with English abstract)[刘玉洪 (1993 ). 哀牢山常绿阔叶林地的气候特征研究
. 林业科学 , 29 , 547 -552 .]
[本文引用: 1]
[54]
Ma WZ 2009 ). The Composition and Biomass of Epiphytic Materials and Their Relationships with Ecological Factors in Xujiaba Region form Ailao Mountain, Yunnan
. PhD dissertation, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Kunming . (in Chinese)[马文章 (2009 ). 云南哀牢山徐家坝地区附生(植)物的组成、生物量及其与生态因子的关系
. 博士学位论文, 中国科学院西双版纳热带植物园, 昆明.]
[55]
Marini L Nascimbene J Nimis PL 2011 ). Large-scale patterns of epiphytic lichen species richness: Photobiont-dependent response to climate and forest structure.
Science of the Total Environment , 409 , 4381 -4386 .
[本文引用: 1]
[56]
McCune B 1993 ). Gradients in epiphyte biomass in three Pseudotsuga-Tsuga forests of different ages in western Oregon and Washington.
The Bryologist , 96 , 405 -411 .
[57]
Montfoort D Ek R 1990 ). Vertical distribution and ecology of epiphytic bryophytes and lichens in a lowland rain forest in French Guyana. PhD dissertation, University of Utrecht, Utrecht, the
Netherlands.
[本文引用: 3]
[58]
Nascimbene J Brunialti G Ravera S Frati L Caniglia G 2010 ). Testing Lobaria pulmonaria (L.) Hoffm as an indicator of lichen conservation importance of Italian forests.
Ecological Indicators , 10 , 353 -360 .
[59]
Nimis PL Martellos S
URL
[本文引用: 2]
[60]
Palmqvist K 2000 ). Carbon economy in lichens.
New Phytologist , 148 , 11 -36 .
[本文引用: 1]
[61]
Pintado A Valladares F Sancho LG 1997 ). Exploring phenol- typic plasticity in the lichen Ramalina capitata: Morphol- ogy, water relations and chlorophyll content in north- and south-facing populations.
Annals of Botany , 80 , 345 -353 .
[本文引用: 3]
[62]
Ricotta C Bacaro (2010 ). On plot to plot dissimilarity measures based on species functional traits.
Community Ecology , 11 , 113 -119 .
[本文引用: 1]
[63]
Rikkinen J Relations between cyanobacterial symbionts in lichens and plants. In: Pawlowski K ed. Prokaryotic Symbionts in Plants . Springer , Berlin . 265 -270 .
[本文引用: 2]
[64]
Sala OE Chapin III FS Armesto JJ Berlow E Bloomfield J Dirzo R Huber-Sanwald E Huenneke LF Jackson RB Kinzig A Leemans R Lodge DM Mooney HA Oesterheld M Poff NL Sykes MT Walker BH Walker M Wall DH 2000 ). Global biodiversity scenarios for the year 2100.
Science , 287 , 1770 -1774 .
[本文引用: 1]
[65]
Sancho LG Kappen L 1989 ). Photosynthesis and water rela- tions and the role of anatomy in Umbilicariaceae (lichens) from Central Spain.
Oecologia , 81 , 473 -480 .
[66]
Sipman HJM Harris RC 1989). Lichens. In: Lieth H, Werger MJA eds. Tropical Rain Forest Ecosystems (Bio- geographical and Ecological Studies), Ecosystems of the World 14B. Elsevier, Amsterdam , USA . 303 -309 .
[本文引用: 1]
[67]
Sipman HJM Tan BC 1990 ). A field impression of the lichen and bryophyte zonation on Mount Kinabalu.
Flora Malesiana Bulletin , 10 , 241 -244 .
[68]
Song L Zhang YJ Chen X Li S LU HZ Wu CS Tan ZH Liu WY Shi XM 2015 ). Water relations and gas exchange of fan bryophytes and their adaptations to microhabitats in an Asian subtropical montane cloud forest.
Journal of Plant Research , 128 , 573 -584 .
[本文引用: 1]
[69]
Travis JMJ Brooker RW Clark , EJ , Dytham C 2006 ). The distribution of positive and negative species interactions across environmental gradients on a dual-lattice model.
Journal of Theoretical Biology , 241 , 896 -902 .
[本文引用: 1]
[70]
Webb CT Hoeting JA Arns GM Pyne MI Poff NL 2010 ). A structured and dynamic framework to advance traits-based theory and prediction in ecology.
Ecology Letters , 13 , 267 -283 .
[71]
Xu HQ Liu WY 2005 ). Species diversity and distribution of epiphytes in the montane moist evergreen broad-leaved forest in Ailao Mountain, Yunnan.
Biodiversity Science , 13 , 137 -147 .[徐海清 , 刘文耀 (2005 ). 云南哀牢山山地湿性常绿阔叶林附生植物的多样性与分布
. 生物多样性 , 13 , 137 -147 .]
[本文引用: 1]
[72]
Yao YL Liu WY Ma WZ Song L 2012 ). Species composition and diversity of epiphytes of several ecotones in Ailao Mountain National Nature Reserve, Yunnan.
Biodiversity Science , 20 , 654 -664 . (in Chinese with English abstract)[姚元林 , 刘文耀 , 马文章 , 宋亮 (2012 ). 云南哀牢山国家保护区几个过渡带树干附生苔藓的物种组成与多样性
. 生物多样性 , 20 , 654 -664 .]
[本文引用: 2]
[73]
Ye ZP 2007 ). A new model for relationship between light intensity and the rate of photosynthesis in Oryza sativa.
Photosynthetica , 45 , 637 -640 .
[本文引用: 1]
[74]
Ye ZP 2010 ). A review on modeling of responses of photosynthesis to light and CO2.
Chinese Journal of Plant Ecology , 34 , 727 -740 . (in Chinese with English abstract)[叶子飘 (2010 ). 光合作用对光和CO2响应模型的研究进展
. 植物生态学报 , 34 , 722 -740 .]
[本文引用: 1]
[75]
You CX Classification of vegetation in Xujiaba region in Ailao Mts. In: Wu ZY ed. Research of Forest Ecosystems on Ailao Mountains, Yunnan. Yunnan Science and Technology Press , Kunming . 74 -117 .(in Chinese). [游承侠 (1983). 哀牢山徐家坝地区的植被分类. 见: 吴征镒主编. 云南哀牢山森林生态系统研究 . 云南科技出版社 , 昆明 . 74 -117 .]
[本文引用: 2]
[76]
Young SS Herwitz SR 1995 ). Floristic diversity and co- occurrences in a subtropical broad-leaved forest and two contrasting regrowth stands in central-west Yunnan Province, China.
Vegetatio , 119 , 1 -13 .
[77]
Zedda L Kong SM Rambold G Morphological groups as a surrogate for soil lichen biodiversity in Southern Africa. In: Bates ST, Bungartz F, Lücking R, Herrera- Campos MA, Zambrano A eds. Biomonitoring, Ecology, and Systematics of Lichens Festschrift Thomas H. Nash III, Bibliotheca Lichenologica 106 . Borntraeger Verlagsbuchhandlung , Stuttgart . 391 -408 .
[本文引用: 2]
[78]
Zhu H Yan LC List of Seed Plants in the Ailao Mts. of Yunnan Province, China . Yunnan Science and Technology Press , Kunming . (in Chinese)[朱华 , 闫丽春 (2009). 云南哀牢山种子植物 . 云南科技出版社 , 昆明.]
[本文引用: 2]
[79]
Zotz G Büdel B Meyer A Zellner H Lange OL 1998 ). In situ studies of water relations and CO2 exchange of the tropical macrolichen, Sticta tomentosa.
New Phytologist , 139 , 525 -535 .
[80]
Zotz G Schultz S Rottenberger S 2003 ). Are tropical lowlands a marginal habitat for macrolichens? Evidence from a field study with Parmotrema endosulphureum in Panama.
Flora , 198 , 71 -77 .
[本文引用: 2]
[81]
Zotz Z Winter K 1994 ). Photosynthesis and carbon gain of the lichen, Leptogium azureum, in lowland tropical forest.
Flora , 189 , 179 -186 .
不同水分条件下三种附生地衣的光合作用特性
2014
Lichens, air pollution and lung cancer.
1
1997
... 地衣的变水特征使得它们的光响应特征比较复杂.地衣水分含量和净光合速率(P n )的关系能够改变它们的P n 对光照的响应(Lange & Matthes, 1981 ).低的水分含量使得地衣的P n 获得低数值的光补偿点(LCP )及光饱和点(LSP ), 而高水分含量对P n 的抑制作用也能够降低地衣的LSP 值, 但对LCP 的影响不大(Lange et al ., 1996 , 2004 ; Green et al ., 1997 ; 陈克等, 2014 ).Green等(1997)对新西兰热带雨林9种地衣光适应的研究指出, 光响应曲线参数和所测定地衣的生境相关.他们的研究发现, 林内地衣的光饱和点要低于300 μmol·m-2 ·s-1 , 而在开阔生境中地衣的光饱和点要大于549 μmol·m-2 ·s-1 .在本研究中, 8种附生地衣的LSP 都比较高(333.9-616.0 μmol·m-2 ·s-1 , 表4 ), 并且即使光强高于1000 μmol·m-2 ·s-1 也没有明显的光抑制现象, 表现出明显的“阳生植物”特性, 这在一定程度上解释了为什么在哀牢山地区附生地衣在次生林比在原生林中有着更高的丰富度(李苏等, 2007 ; Li et al ., 2013a , 2013b ).就各功能群附生地衣的LSP 而言, 枝状地衣的LSP 要明显高于其他功能群附生地衣, 这也很好地解释了它们在林冠上层和林外空旷地有较高多样性的分布格局(Li et al ., 2015 ).Green等(1997)和陈克等(2014)的研究中都指出相比于绿藻地衣, 蓝藻地衣有着较低的LSP .在我们的研究中虽然网肺衣也有着较高的LSP, 但是猫耳衣的LSP 还是低于其他功能群附生地衣.LCP 是衡量植物利用弱光能力的重要指标.在我们的研究中, 8种附生植物的LCP 都比较高(20.1-43.1 μmol·m-2 · ...
Comparative cryptogam ecology: A review of bryophyte and lichen traits that drive biogeochemistry.
1
2007
... 气候变化、氮沉降、生物入侵和土地利用变化已经成为当前全球生物多样性的主要威胁(Sala et al ., 2000 ), 因此许多学者开始研究全球变化对生物多样性和物种分布的潜在影响.在这样的背景下, 理解限制物种分布的过程和机制就显得尤为重要(Travis et al ., 2006 ).附生地衣是森林生态系统的重要组成部分, 占据着林冠相当份量的生物量(Sipman & Harris, 1989 ; Gradstein, 1992 ), 其多样性和分布自20世纪70年代起就得到了众多学者的关注(Cornelissen & Steege, 1999 ; Holz & Gradstein, 2005 ; Hauck & Spribille, 2005 ).Aptroot和Sipman (1997)估计在热带雨林中每km2 有300种地衣, Aptroot (1997)报道在巴布亚新几内亚的热带山地雨林一株树上就有173种地衣.地衣的分布受众多因子的综合影响(Gustafasson & Eriksson, 1995 ; Goward & Arsenawlt, 2000 ; Hauck & Spribille, 2005 ), 除自身的繁殖能力和扩散能力以外, 光照、温度、湿度、干扰程度、群落类型以及宿主特征等均对附生地衣物种的组成和分布产生一定的影响(李苏等, 2007 ; Li et al ., 2011 , 2013a, 2013b ).地衣由于其形态结构的特殊性和变水植物的特性, 对环境变化非常敏感, 因此地衣多样性的减少经常被用来指示空气污染(Cislaghi & Nimis, 1997 ; Giordani, 2007 )、气候条件(Jovan & McCune, 2004 ; Geiser & Neitlich, 2007 ; Giordani & Incerti, 2008 )、森林结构和动态(Johansson, 2008 ; Nascimbene et al ., 2010 ).但是通过比较地衣多样性来比较不同区域间的环境条件常常遇到困难, 因为环境条件的不同经常形成高度差异的物种组成, 而如果我们利用地衣的功能特征来指示环境条件则可以很好地解决这一问题(Giordani et al ., 2012 ). ...
Distribution and ecology of epiphytic bryophytes and lichens in dry evergreen forest of Guyana.
2
1989
... 气候变化、氮沉降、生物入侵和土地利用变化已经成为当前全球生物多样性的主要威胁(Sala et al ., 2000 ), 因此许多学者开始研究全球变化对生物多样性和物种分布的潜在影响.在这样的背景下, 理解限制物种分布的过程和机制就显得尤为重要(Travis et al ., 2006 ).附生地衣是森林生态系统的重要组成部分, 占据着林冠相当份量的生物量(Sipman & Harris, 1989 ; Gradstein, 1992 ), 其多样性和分布自20世纪70年代起就得到了众多学者的关注(Cornelissen & Steege, 1999 ; Holz & Gradstein, 2005 ; Hauck & Spribille, 2005 ).Aptroot和Sipman (1997)估计在热带雨林中每km2 有300种地衣, Aptroot (1997)报道在巴布亚新几内亚的热带山地雨林一株树上就有173种地衣.地衣的分布受众多因子的综合影响(Gustafasson & Eriksson, 1995 ; Goward & Arsenawlt, 2000 ; Hauck & Spribille, 2005 ), 除自身的繁殖能力和扩散能力以外, 光照、温度、湿度、干扰程度、群落类型以及宿主特征等均对附生地衣物种的组成和分布产生一定的影响(李苏等, 2007 ; Li et al ., 2011 , 2013a, 2013b ).地衣由于其形态结构的特殊性和变水植物的特性, 对环境变化非常敏感, 因此地衣多样性的减少经常被用来指示空气污染(Cislaghi & Nimis, 1997 ; Giordani, 2007 )、气候条件(Jovan & McCune, 2004 ; Geiser & Neitlich, 2007 ; Giordani & Incerti, 2008 )、森林结构和动态(Johansson, 2008 ; Nascimbene et al ., 2010 ).但是通过比较地衣多样性来比较不同区域间的环境条件常常遇到困难, 因为环境条件的不同经常形成高度差异的物种组成, 而如果我们利用地衣的功能特征来指示环境条件则可以很好地解决这一问题(Giordani et al ., 2012 ). ...
... 根据国外的相关研究, 地衣按照共生藻、生活型和繁殖策略的不同可以划分为蓝藻地衣(cyano-区性lichens, CYL)、壳状地衣(crustose lichens, CRL)、枝状地衣(fruticose lichens, FRL)、狭叶地衣(narrowly lobed foliose lichens, NFL)、阔叶地衣(broadly lobed foliose lichens, BFL)、裂芽地衣(isidiate lichens, ISL)、粉芽地衣(sorediate lichens, SOL)和有性地衣(sexual lichens, SEL) 8个功能群(McCune, 1993 ; Nimis & Martellos, 2008 ).调查研究显示地衣呈现出与它们功能特征相关的分布格局(Ellis & Coppins, 2006 ; Lakatos et al ., 2006 ; Zedda et al ., 2011 ).比如在热带山地雨林分布着更多的枝状和叶状地衣, 但在低地雨林却是壳状地衣占据优势, 枝状地衣和叶状地衣很稀少(Montfoot & Ek, 1990 ; Sipman & Tan, 1990 ; Lakatos et al ., 2004 ).Hauck等(2007)在蒙古的针叶林发现, 绿藻地衣在群落中处于优势地位, 蓝藻地衣只在有着较高可利用水源的地方有分布.在最近的研究中, 地衣的生活型、繁殖策略、共生藻等功能特征被广泛用来指示地衣群落对环境条件的适应(DÍaz & Cabido, 2001 ; Cornelissen et al ., 2007 ; Glime, 2007 ), 因为地衣的功能特征可以不依赖物种多样性和组成而直接和环境因子建立关联(Ricotta & Bacaro, 2010 ; Webb et al ., 2010 ).Lange等(1994)发现在蓝藻地衣较多的生境中经常有高的积水.Giordani等(2012)基于意大利森林生态系统网络研究了地衣功能特征和环境条件的关系, 结果表明: 壳状和叶状地衣的分布和降水量密切相关; 枝状地衣的分布受海拔的影响较大, 同时和森林的密闭度负相关; 狭叶和阔叶地衣都受到酸性沉积物的负面影响.Giordani等(2014)调查发现阔叶地衣只有在高潜在径流的条件下才能在高辐射的生境中分布, 同时他们还观察到在枝状地衣的生境中有高的潜在径流. ...
Vive la difference: Plant functional diversity matters to ecosystem processes.
2001
Contrasting functional traits maintain lichen epiphyte diversity in response to climate and autogenic succession.
1
2006
... 根据国外的相关研究, 地衣按照共生藻、生活型和繁殖策略的不同可以划分为蓝藻地衣(cyano-区性lichens, CYL)、壳状地衣(crustose lichens, CRL)、枝状地衣(fruticose lichens, FRL)、狭叶地衣(narrowly lobed foliose lichens, NFL)、阔叶地衣(broadly lobed foliose lichens, BFL)、裂芽地衣(isidiate lichens, ISL)、粉芽地衣(sorediate lichens, SOL)和有性地衣(sexual lichens, SEL) 8个功能群(McCune, 1993 ; Nimis & Martellos, 2008 ).调查研究显示地衣呈现出与它们功能特征相关的分布格局(Ellis & Coppins, 2006 ; Lakatos et al ., 2006 ; Zedda et al ., 2011 ).比如在热带山地雨林分布着更多的枝状和叶状地衣, 但在低地雨林却是壳状地衣占据优势, 枝状地衣和叶状地衣很稀少(Montfoot & Ek, 1990 ; Sipman & Tan, 1990 ; Lakatos et al ., 2004 ).Hauck等(2007)在蒙古的针叶林发现, 绿藻地衣在群落中处于优势地位, 蓝藻地衣只在有着较高可利用水源的地方有分布.在最近的研究中, 地衣的生活型、繁殖策略、共生藻等功能特征被广泛用来指示地衣群落对环境条件的适应(DÍaz & Cabido, 2001 ; Cornelissen et al ., 2007 ; Glime, 2007 ), 因为地衣的功能特征可以不依赖物种多样性和组成而直接和环境因子建立关联(Ricotta & Bacaro, 2010 ; Webb et al ., 2010 ).Lange等(1994)发现在蓝藻地衣较多的生境中经常有高的积水.Giordani等(2012)基于意大利森林生态系统网络研究了地衣功能特征和环境条件的关系, 结果表明: 壳状和叶状地衣的分布和降水量密切相关; 枝状地衣的分布受海拔的影响较大, 同时和森林的密闭度负相关; 狭叶和阔叶地衣都受到酸性沉积物的负面影响.Giordani等(2014)调查发现阔叶地衣只有在高潜在径流的条件下才能在高辐射的生境中分布, 同时他们还观察到在枝状地衣的生境中有高的潜在径流. ...
Changing climate and historic-woodland structure interact to control species diversity of the ‘Lobarion’ epiphyte community in Scotland.
2
2007
... 根据国外的相关研究, 地衣按照共生藻、生活型和繁殖策略的不同可以划分为蓝藻地衣(cyano-区性lichens, CYL)、壳状地衣(crustose lichens, CRL)、枝状地衣(fruticose lichens, FRL)、狭叶地衣(narrowly lobed foliose lichens, NFL)、阔叶地衣(broadly lobed foliose lichens, BFL)、裂芽地衣(isidiate lichens, ISL)、粉芽地衣(sorediate lichens, SOL)和有性地衣(sexual lichens, SEL) 8个功能群(McCune, 1993 ; Nimis & Martellos, 2008 ).调查研究显示地衣呈现出与它们功能特征相关的分布格局(Ellis & Coppins, 2006 ; Lakatos et al ., 2006 ; Zedda et al ., 2011 ).比如在热带山地雨林分布着更多的枝状和叶状地衣, 但在低地雨林却是壳状地衣占据优势, 枝状地衣和叶状地衣很稀少(Montfoot & Ek, 1990 ; Sipman & Tan, 1990 ; Lakatos et al ., 2004 ).Hauck等(2007)在蒙古的针叶林发现, 绿藻地衣在群落中处于优势地位, 蓝藻地衣只在有着较高可利用水源的地方有分布.在最近的研究中, 地衣的生活型、繁殖策略、共生藻等功能特征被广泛用来指示地衣群落对环境条件的适应(DÍaz & Cabido, 2001 ; Cornelissen et al ., 2007 ; Glime, 2007 ), 因为地衣的功能特征可以不依赖物种多样性和组成而直接和环境因子建立关联(Ricotta & Bacaro, 2010 ; Webb et al ., 2010 ).Lange等(1994)发现在蓝藻地衣较多的生境中经常有高的积水.Giordani等(2012)基于意大利森林生态系统网络研究了地衣功能特征和环境条件的关系, 结果表明: 壳状和叶状地衣的分布和降水量密切相关; 枝状地衣的分布受海拔的影响较大, 同时和森林的密闭度负相关; 狭叶和阔叶地衣都受到酸性沉积物的负面影响.Giordani等(2014)调查发现阔叶地衣只有在高潜在径流的条件下才能在高辐射的生境中分布, 同时他们还观察到在枝状地衣的生境中有高的潜在径流. ...
... 由于生活型和共生藻类型可能控制群落的结构(Ellis & Coppins, 2006 , 2007, 2010 ; Marini et al ., 2011 ), 结合Li等(2013a, 2013b)对本地区附生地衣分布的调查研究, 我们选择更能指示环境状况的蓝藻地衣、阔叶地衣、狭叶地衣、枝状地衣4种地衣功能群作为研究对象, 4种附生地衣功能群的划分依据见表1 .每种附生地衣功能群选择2种常见种(或代表种)作为研究对象.其中蓝藻地衣为网肺衣(Lobaria retigera )和猫耳衣(Leptogium menziesii ), 阔叶地衣为针芽肺衣(Lobaria isidiophora )和皮革肾岛衣(Nephromopsis pallescens ), 狭叶地衣为白绵腹衣(Anzia leucobatoides )和云南袋衣(Hypogymnia yunnanensis ), 枝状地衣为裂髓树花(Ramalina conduplicans )和多花松萝(Usnea florida ).其中网肺衣和猫耳衣多生长于潮湿的生境中, 针芽肺衣、皮革肾岛衣、白棉腹衣和云南袋衣多生长于阳光充足的生境中, 裂髓树花和多花松萝多分布于干燥、高光的生境中(Li et al ., 2013a ).我们在每个物种分布较多的生境中采集各研究物种材料, 带回哀牢山生态站, 清洗干净后, 进行相关实验. ...
Integrating multiple landscape- scale drivers in the lichen epiphyte response: Climatic setting, pollution regime and woodland spatial-temporal structure.
1
2010
... 由于生活型和共生藻类型可能控制群落的结构(Ellis & Coppins, 2006 , 2007, 2010 ; Marini et al ., 2011 ), 结合Li等(2013a, 2013b)对本地区附生地衣分布的调查研究, 我们选择更能指示环境状况的蓝藻地衣、阔叶地衣、狭叶地衣、枝状地衣4种地衣功能群作为研究对象, 4种附生地衣功能群的划分依据见表1 .每种附生地衣功能群选择2种常见种(或代表种)作为研究对象.其中蓝藻地衣为网肺衣(Lobaria retigera )和猫耳衣(Leptogium menziesii ), 阔叶地衣为针芽肺衣(Lobaria isidiophora )和皮革肾岛衣(Nephromopsis pallescens ), 狭叶地衣为白绵腹衣(Anzia leucobatoides )和云南袋衣(Hypogymnia yunnanensis ), 枝状地衣为裂髓树花(Ramalina conduplicans )和多花松萝(Usnea florida ).其中网肺衣和猫耳衣多生长于潮湿的生境中, 针芽肺衣、皮革肾岛衣、白棉腹衣和云南袋衣多生长于阳光充足的生境中, 裂髓树花和多花松萝多分布于干燥、高光的生境中(Li et al ., 2013a ).我们在每个物种分布较多的生境中采集各研究物种材料, 带回哀牢山生态站, 清洗干净后, 进行相关实验. ...
Morphology influences water storage in hair lichens from boreal forest canopies.
2015
2
... 不同附生地衣单位干质量持水量的差异利用单因素方差分析和Tukey’s HSD多重配对进行分析, 并利用线性模型(GLM)来分析附生地衣物种、干质量和持水力(WHC)之间的关系(Gauslaa & Coxson, 2011 ; Esseen et al ., 2015 ).各附生地衣的失水速率利用指数递减回归模型(Song et al ., 2015 )进行拟合: ...
... 作为变水植物, 地衣的含水量随着环境条件迅速变化, 而其所有的代谢活动, 特别是净光合速率和呼吸速率也随之发生变化(Green & Lange, 1994 ).所以地衣水分关系的研究格外受到关注(Galun, 1988 ).地衣不能主动地调节水分的吸收、储存和流失(Green et al ., 2011 ), 它们利用形态学特征来促进叶状体从不同来源(雨水、潮湿空气、露水)吸收和贮存水分(Gauslaa & Coxson, 2011 ).形态结构和解剖结构是影响地衣水分贮藏的重要因素(Larson & Kershaw, 1976 ; Larson, 1981 ; Sancho & Kappen, 1989 ).在本研究中, 附生地衣的最大含水量和功能特征相关性不大, 属于同一功能团的两种附生地衣间的最大含水量也可能相差很大, 而枝状地衣的最大含水量也可能比叶状地衣的大, 这就说明解剖结构可能在附生地衣持水量上发挥了更重要的作用.Gauslaa和Coxson (2011)认为共生藻层的厚度是影响地衣物种间持水量的重要因素, 而共生藻的类型是影响其厚度的主要因素.我们所研究的两种蓝藻地衣都有着较高的持水量, 尤其是猫耳衣, 其持水量明显高于其他附生地衣.而网肺衣的持水量也比同属的针芽肺衣高100%左右.蓝藻地衣之所以有较高的持水量是因为在它们的共生藻细胞外有一层较厚的凝胶状保护鞘(Honegger et al ., 1996 ; Gauslaa & Coxson, 2011 ), 能够帮助蓝藻地衣贮存更多的水分.水分贮存能力、形态结构、解剖结构以及颜色是影响地衣吸收和丢失水分的主要因素(Palmqvist, 2000 ).比如, 相比于叶状地衣, 枝状地衣和环境之间的水分交换更快(Hartard et al ., 2009 ).在本研究中也是枝状地衣的失水速度要快于其他功能群附生地衣.枝状地衣的这种形态特征也使得它们能够从湿润的空气中快速吸收水分(Lange & Ziegler, 1986 ), 也就使得它们能够适应林外干燥的环境.相对于枝状地衣, 叶状地衣则是通过增大持水量和水分贮存能力来保持体内的水分含量从而延长生理活动的时间(Lange & Ziegler, 1986 ), 比如我们所研究的白绵腹衣, 其失水速度要明显低于其他附生地衣.对于蓝藻地衣而言, 由于只能利用液态水来进行光合作用(Lange & Ziegler, 1986 ), 所以它们通过凝胶状物质来增加持水量. ...
Interspecific and intraspecific variations in water storage in epiphytic old forest foliose lichens.
1
2011
... 作为变水植物, 地衣的含水量随着环境条件迅速变化, 而其所有的代谢活动, 特别是净光合速率和呼吸速率也随之发生变化(Green & Lange, 1994 ).所以地衣水分关系的研究格外受到关注(Galun, 1988 ).地衣不能主动地调节水分的吸收、储存和流失(Green et al ., 2011 ), 它们利用形态学特征来促进叶状体从不同来源(雨水、潮湿空气、露水)吸收和贮存水分(Gauslaa & Coxson, 2011 ).形态结构和解剖结构是影响地衣水分贮藏的重要因素(Larson & Kershaw, 1976 ; Larson, 1981 ; Sancho & Kappen, 1989 ).在本研究中, 附生地衣的最大含水量和功能特征相关性不大, 属于同一功能团的两种附生地衣间的最大含水量也可能相差很大, 而枝状地衣的最大含水量也可能比叶状地衣的大, 这就说明解剖结构可能在附生地衣持水量上发挥了更重要的作用.Gauslaa和Coxson (2011)认为共生藻层的厚度是影响地衣物种间持水量的重要因素, 而共生藻的类型是影响其厚度的主要因素.我们所研究的两种蓝藻地衣都有着较高的持水量, 尤其是猫耳衣, 其持水量明显高于其他附生地衣.而网肺衣的持水量也比同属的针芽肺衣高100%左右.蓝藻地衣之所以有较高的持水量是因为在它们的共生藻细胞外有一层较厚的凝胶状保护鞘(Honegger et al ., 1996 ; Gauslaa & Coxson, 2011 ), 能够帮助蓝藻地衣贮存更多的水分.水分贮存能力、形态结构、解剖结构以及颜色是影响地衣吸收和丢失水分的主要因素(Palmqvist, 2000 ).比如, 相比于叶状地衣, 枝状地衣和环境之间的水分交换更快(Hartard et al ., 2009 ).在本研究中也是枝状地衣的失水速度要快于其他功能群附生地衣.枝状地衣的这种形态特征也使得它们能够从湿润的空气中快速吸收水分(Lange & Ziegler, 1986 ), 也就使得它们能够适应林外干燥的环境.相对于枝状地衣, 叶状地衣则是通过增大持水量和水分贮存能力来保持体内的水分含量从而延长生理活动的时间(Lange & Ziegler, 1986 ), 比如我们所研究的白绵腹衣, 其失水速度要明显低于其他附生地衣.对于蓝藻地衣而言, 由于只能利用液态水来进行光合作用(Lange & Ziegler, 1986 ), 所以它们通过凝胶状物质来增加持水量. ...
Air pollution and climate gradients in western Oregon and Washington indicated by epiphytic macrolichens.
3
2007
... 不同附生地衣单位干质量持水量的差异利用单因素方差分析和Tukey’s HSD多重配对进行分析, 并利用线性模型(GLM)来分析附生地衣物种、干质量和持水力(WHC)之间的关系(Gauslaa & Coxson, 2011 ; Esseen et al ., 2015 ).各附生地衣的失水速率利用指数递减回归模型(Song et al ., 2015 )进行拟合: ...
... 作为变水植物, 地衣的含水量随着环境条件迅速变化, 而其所有的代谢活动, 特别是净光合速率和呼吸速率也随之发生变化(Green & Lange, 1994 ).所以地衣水分关系的研究格外受到关注(Galun, 1988 ).地衣不能主动地调节水分的吸收、储存和流失(Green et al ., 2011 ), 它们利用形态学特征来促进叶状体从不同来源(雨水、潮湿空气、露水)吸收和贮存水分(Gauslaa & Coxson, 2011 ).形态结构和解剖结构是影响地衣水分贮藏的重要因素(Larson & Kershaw, 1976 ; Larson, 1981 ; Sancho & Kappen, 1989 ).在本研究中, 附生地衣的最大含水量和功能特征相关性不大, 属于同一功能团的两种附生地衣间的最大含水量也可能相差很大, 而枝状地衣的最大含水量也可能比叶状地衣的大, 这就说明解剖结构可能在附生地衣持水量上发挥了更重要的作用.Gauslaa和Coxson (2011)认为共生藻层的厚度是影响地衣物种间持水量的重要因素, 而共生藻的类型是影响其厚度的主要因素.我们所研究的两种蓝藻地衣都有着较高的持水量, 尤其是猫耳衣, 其持水量明显高于其他附生地衣.而网肺衣的持水量也比同属的针芽肺衣高100%左右.蓝藻地衣之所以有较高的持水量是因为在它们的共生藻细胞外有一层较厚的凝胶状保护鞘(Honegger et al ., 1996 ; Gauslaa & Coxson, 2011 ), 能够帮助蓝藻地衣贮存更多的水分.水分贮存能力、形态结构、解剖结构以及颜色是影响地衣吸收和丢失水分的主要因素(Palmqvist, 2000 ).比如, 相比于叶状地衣, 枝状地衣和环境之间的水分交换更快(Hartard et al ., 2009 ).在本研究中也是枝状地衣的失水速度要快于其他功能群附生地衣.枝状地衣的这种形态特征也使得它们能够从湿润的空气中快速吸收水分(Lange & Ziegler, 1986 ), 也就使得它们能够适应林外干燥的环境.相对于枝状地衣, 叶状地衣则是通过增大持水量和水分贮存能力来保持体内的水分含量从而延长生理活动的时间(Lange & Ziegler, 1986 ), 比如我们所研究的白绵腹衣, 其失水速度要明显低于其他附生地衣.对于蓝藻地衣而言, 由于只能利用液态水来进行光合作用(Lange & Ziegler, 1986 ), 所以它们通过凝胶状物质来增加持水量. ...
... ; Gauslaa & Coxson, 2011 ), 能够帮助蓝藻地衣贮存更多的水分.水分贮存能力、形态结构、解剖结构以及颜色是影响地衣吸收和丢失水分的主要因素(Palmqvist, 2000 ).比如, 相比于叶状地衣, 枝状地衣和环境之间的水分交换更快(Hartard et al ., 2009 ).在本研究中也是枝状地衣的失水速度要快于其他功能群附生地衣.枝状地衣的这种形态特征也使得它们能够从湿润的空气中快速吸收水分(Lange & Ziegler, 1986 ), 也就使得它们能够适应林外干燥的环境.相对于枝状地衣, 叶状地衣则是通过增大持水量和水分贮存能力来保持体内的水分含量从而延长生理活动的时间(Lange & Ziegler, 1986 ), 比如我们所研究的白绵腹衣, 其失水速度要明显低于其他附生地衣.对于蓝藻地衣而言, 由于只能利用液态水来进行光合作用(Lange & Ziegler, 1986 ), 所以它们通过凝胶状物质来增加持水量. ...
Is the diversity of epiphytic lichens a reliable indicator of air pollution? A case study from Italy.
1
2007
... 气候变化、氮沉降、生物入侵和土地利用变化已经成为当前全球生物多样性的主要威胁(Sala et al ., 2000 ), 因此许多学者开始研究全球变化对生物多样性和物种分布的潜在影响.在这样的背景下, 理解限制物种分布的过程和机制就显得尤为重要(Travis et al ., 2006 ).附生地衣是森林生态系统的重要组成部分, 占据着林冠相当份量的生物量(Sipman & Harris, 1989 ; Gradstein, 1992 ), 其多样性和分布自20世纪70年代起就得到了众多学者的关注(Cornelissen & Steege, 1999 ; Holz & Gradstein, 2005 ; Hauck & Spribille, 2005 ).Aptroot和Sipman (1997)估计在热带雨林中每km2 有300种地衣, Aptroot (1997)报道在巴布亚新几内亚的热带山地雨林一株树上就有173种地衣.地衣的分布受众多因子的综合影响(Gustafasson & Eriksson, 1995 ; Goward & Arsenawlt, 2000 ; Hauck & Spribille, 2005 ), 除自身的繁殖能力和扩散能力以外, 光照、温度、湿度、干扰程度、群落类型以及宿主特征等均对附生地衣物种的组成和分布产生一定的影响(李苏等, 2007 ; Li et al ., 2011 , 2013a, 2013b ).地衣由于其形态结构的特殊性和变水植物的特性, 对环境变化非常敏感, 因此地衣多样性的减少经常被用来指示空气污染(Cislaghi & Nimis, 1997 ; Giordani, 2007 )、气候条件(Jovan & McCune, 2004 ; Geiser & Neitlich, 2007 ; Giordani & Incerti, 2008 )、森林结构和动态(Johansson, 2008 ; Nascimbene et al ., 2010 ).但是通过比较地衣多样性来比较不同区域间的环境条件常常遇到困难, 因为环境条件的不同经常形成高度差异的物种组成, 而如果我们利用地衣的功能特征来指示环境条件则可以很好地解决这一问题(Giordani et al ., 2012 ). ...
Functional traits of epiphytic lichens as potential indicators of environmental conditions in forest ecosystems.
1
2012
... 气候变化、氮沉降、生物入侵和土地利用变化已经成为当前全球生物多样性的主要威胁(Sala et al ., 2000 ), 因此许多学者开始研究全球变化对生物多样性和物种分布的潜在影响.在这样的背景下, 理解限制物种分布的过程和机制就显得尤为重要(Travis et al ., 2006 ).附生地衣是森林生态系统的重要组成部分, 占据着林冠相当份量的生物量(Sipman & Harris, 1989 ; Gradstein, 1992 ), 其多样性和分布自20世纪70年代起就得到了众多学者的关注(Cornelissen & Steege, 1999 ; Holz & Gradstein, 2005 ; Hauck & Spribille, 2005 ).Aptroot和Sipman (1997)估计在热带雨林中每km2 有300种地衣, Aptroot (1997)报道在巴布亚新几内亚的热带山地雨林一株树上就有173种地衣.地衣的分布受众多因子的综合影响(Gustafasson & Eriksson, 1995 ; Goward & Arsenawlt, 2000 ; Hauck & Spribille, 2005 ), 除自身的繁殖能力和扩散能力以外, 光照、温度、湿度、干扰程度、群落类型以及宿主特征等均对附生地衣物种的组成和分布产生一定的影响(李苏等, 2007 ; Li et al ., 2011 , 2013a, 2013b ).地衣由于其形态结构的特殊性和变水植物的特性, 对环境变化非常敏感, 因此地衣多样性的减少经常被用来指示空气污染(Cislaghi & Nimis, 1997 ; Giordani, 2007 )、气候条件(Jovan & McCune, 2004 ; Geiser & Neitlich, 2007 ; Giordani & Incerti, 2008 )、森林结构和动态(Johansson, 2008 ; Nascimbene et al ., 2010 ).但是通过比较地衣多样性来比较不同区域间的环境条件常常遇到困难, 因为环境条件的不同经常形成高度差异的物种组成, 而如果我们利用地衣的功能特征来指示环境条件则可以很好地解决这一问题(Giordani et al ., 2012 ). ...
The influence of climate on the distribution of lichens: A case study in a borderline area (Liguria, NW Italy).
1
2008
... 气候变化、氮沉降、生物入侵和土地利用变化已经成为当前全球生物多样性的主要威胁(Sala et al ., 2000 ), 因此许多学者开始研究全球变化对生物多样性和物种分布的潜在影响.在这样的背景下, 理解限制物种分布的过程和机制就显得尤为重要(Travis et al ., 2006 ).附生地衣是森林生态系统的重要组成部分, 占据着林冠相当份量的生物量(Sipman & Harris, 1989 ; Gradstein, 1992 ), 其多样性和分布自20世纪70年代起就得到了众多学者的关注(Cornelissen & Steege, 1999 ; Holz & Gradstein, 2005 ; Hauck & Spribille, 2005 ).Aptroot和Sipman (1997)估计在热带雨林中每km2 有300种地衣, Aptroot (1997)报道在巴布亚新几内亚的热带山地雨林一株树上就有173种地衣.地衣的分布受众多因子的综合影响(Gustafasson & Eriksson, 1995 ; Goward & Arsenawlt, 2000 ; Hauck & Spribille, 2005 ), 除自身的繁殖能力和扩散能力以外, 光照、温度、湿度、干扰程度、群落类型以及宿主特征等均对附生地衣物种的组成和分布产生一定的影响(李苏等, 2007 ; Li et al ., 2011 , 2013a, 2013b ).地衣由于其形态结构的特殊性和变水植物的特性, 对环境变化非常敏感, 因此地衣多样性的减少经常被用来指示空气污染(Cislaghi & Nimis, 1997 ; Giordani, 2007 )、气候条件(Jovan & McCune, 2004 ; Geiser & Neitlich, 2007 ; Giordani & Incerti, 2008 )、森林结构和动态(Johansson, 2008 ; Nascimbene et al ., 2010 ).但是通过比较地衣多样性来比较不同区域间的环境条件常常遇到困难, 因为环境条件的不同经常形成高度差异的物种组成, 而如果我们利用地衣的功能特征来指示环境条件则可以很好地解决这一问题(Giordani et al ., 2012 ). ...
Functional traits of cryptogams in Mediterranean ecosystems are driven by water, light and substrate interactions.
1
2014
... 气候变化、氮沉降、生物入侵和土地利用变化已经成为当前全球生物多样性的主要威胁(Sala et al ., 2000 ), 因此许多学者开始研究全球变化对生物多样性和物种分布的潜在影响.在这样的背景下, 理解限制物种分布的过程和机制就显得尤为重要(Travis et al ., 2006 ).附生地衣是森林生态系统的重要组成部分, 占据着林冠相当份量的生物量(Sipman & Harris, 1989 ; Gradstein, 1992 ), 其多样性和分布自20世纪70年代起就得到了众多学者的关注(Cornelissen & Steege, 1999 ; Holz & Gradstein, 2005 ; Hauck & Spribille, 2005 ).Aptroot和Sipman (1997)估计在热带雨林中每km2 有300种地衣, Aptroot (1997)报道在巴布亚新几内亚的热带山地雨林一株树上就有173种地衣.地衣的分布受众多因子的综合影响(Gustafasson & Eriksson, 1995 ; Goward & Arsenawlt, 2000 ; Hauck & Spribille, 2005 ), 除自身的繁殖能力和扩散能力以外, 光照、温度、湿度、干扰程度、群落类型以及宿主特征等均对附生地衣物种的组成和分布产生一定的影响(李苏等, 2007 ; Li et al ., 2011 , 2013a, 2013b ).地衣由于其形态结构的特殊性和变水植物的特性, 对环境变化非常敏感, 因此地衣多样性的减少经常被用来指示空气污染(Cislaghi & Nimis, 1997 ; Giordani, 2007 )、气候条件(Jovan & McCune, 2004 ; Geiser & Neitlich, 2007 ; Giordani & Incerti, 2008 )、森林结构和动态(Johansson, 2008 ; Nascimbene et al ., 2010 ).但是通过比较地衣多样性来比较不同区域间的环境条件常常遇到困难, 因为环境条件的不同经常形成高度差异的物种组成, 而如果我们利用地衣的功能特征来指示环境条件则可以很好地解决这一问题(Giordani et al ., 2012 ). ...
1
... 根据国外的相关研究, 地衣按照共生藻、生活型和繁殖策略的不同可以划分为蓝藻地衣(cyano-区性lichens, CYL)、壳状地衣(crustose lichens, CRL)、枝状地衣(fruticose lichens, FRL)、狭叶地衣(narrowly lobed foliose lichens, NFL)、阔叶地衣(broadly lobed foliose lichens, BFL)、裂芽地衣(isidiate lichens, ISL)、粉芽地衣(sorediate lichens, SOL)和有性地衣(sexual lichens, SEL) 8个功能群(McCune, 1993 ; Nimis & Martellos, 2008 ).调查研究显示地衣呈现出与它们功能特征相关的分布格局(Ellis & Coppins, 2006 ; Lakatos et al ., 2006 ; Zedda et al ., 2011 ).比如在热带山地雨林分布着更多的枝状和叶状地衣, 但在低地雨林却是壳状地衣占据优势, 枝状地衣和叶状地衣很稀少(Montfoot & Ek, 1990 ; Sipman & Tan, 1990 ; Lakatos et al ., 2004 ).Hauck等(2007)在蒙古的针叶林发现, 绿藻地衣在群落中处于优势地位, 蓝藻地衣只在有着较高可利用水源的地方有分布.在最近的研究中, 地衣的生活型、繁殖策略、共生藻等功能特征被广泛用来指示地衣群落对环境条件的适应(DÍaz & Cabido, 2001 ; Cornelissen et al ., 2007 ; Glime, 2007 ), 因为地衣的功能特征可以不依赖物种多样性和组成而直接和环境因子建立关联(Ricotta & Bacaro, 2010 ; Webb et al ., 2010 ).Lange等(1994)发现在蓝藻地衣较多的生境中经常有高的积水.Giordani等(2012)基于意大利森林生态系统网络研究了地衣功能特征和环境条件的关系, 结果表明: 壳状和叶状地衣的分布和降水量密切相关; 枝状地衣的分布受海拔的影响较大, 同时和森林的密闭度负相关; 狭叶和阔叶地衣都受到酸性沉积物的负面影响.Giordani等(2014)调查发现阔叶地衣只有在高潜在径流的条件下才能在高辐射的生境中分布, 同时他们还观察到在枝状地衣的生境中有高的潜在径流. ...
Cyanolichen distribution in young unmanaged forests: A dripzone effect?
1
2000
... 气候变化、氮沉降、生物入侵和土地利用变化已经成为当前全球生物多样性的主要威胁(Sala et al ., 2000 ), 因此许多学者开始研究全球变化对生物多样性和物种分布的潜在影响.在这样的背景下, 理解限制物种分布的过程和机制就显得尤为重要(Travis et al ., 2006 ).附生地衣是森林生态系统的重要组成部分, 占据着林冠相当份量的生物量(Sipman & Harris, 1989 ; Gradstein, 1992 ), 其多样性和分布自20世纪70年代起就得到了众多学者的关注(Cornelissen & Steege, 1999 ; Holz & Gradstein, 2005 ; Hauck & Spribille, 2005 ).Aptroot和Sipman (1997)估计在热带雨林中每km2 有300种地衣, Aptroot (1997)报道在巴布亚新几内亚的热带山地雨林一株树上就有173种地衣.地衣的分布受众多因子的综合影响(Gustafasson & Eriksson, 1995 ; Goward & Arsenawlt, 2000 ; Hauck & Spribille, 2005 ), 除自身的繁殖能力和扩散能力以外, 光照、温度、湿度、干扰程度、群落类型以及宿主特征等均对附生地衣物种的组成和分布产生一定的影响(李苏等, 2007 ; Li et al ., 2011 , 2013a, 2013b ).地衣由于其形态结构的特殊性和变水植物的特性, 对环境变化非常敏感, 因此地衣多样性的减少经常被用来指示空气污染(Cislaghi & Nimis, 1997 ; Giordani, 2007 )、气候条件(Jovan & McCune, 2004 ; Geiser & Neitlich, 2007 ; Giordani & Incerti, 2008 )、森林结构和动态(Johansson, 2008 ; Nascimbene et al ., 2010 ).但是通过比较地衣多样性来比较不同区域间的环境条件常常遇到困难, 因为环境条件的不同经常形成高度差异的物种组成, 而如果我们利用地衣的功能特征来指示环境条件则可以很好地解决这一问题(Giordani et al ., 2012 ). ...
1
... 气候变化、氮沉降、生物入侵和土地利用变化已经成为当前全球生物多样性的主要威胁(Sala et al ., 2000 ), 因此许多学者开始研究全球变化对生物多样性和物种分布的潜在影响.在这样的背景下, 理解限制物种分布的过程和机制就显得尤为重要(Travis et al ., 2006 ).附生地衣是森林生态系统的重要组成部分, 占据着林冠相当份量的生物量(Sipman & Harris, 1989 ; Gradstein, 1992 ), 其多样性和分布自20世纪70年代起就得到了众多学者的关注(Cornelissen & Steege, 1999 ; Holz & Gradstein, 2005 ; Hauck & Spribille, 2005 ).Aptroot和Sipman (1997)估计在热带雨林中每km2 有300种地衣, Aptroot (1997)报道在巴布亚新几内亚的热带山地雨林一株树上就有173种地衣.地衣的分布受众多因子的综合影响(Gustafasson & Eriksson, 1995 ; Goward & Arsenawlt, 2000 ; Hauck & Spribille, 2005 ), 除自身的繁殖能力和扩散能力以外, 光照、温度、湿度、干扰程度、群落类型以及宿主特征等均对附生地衣物种的组成和分布产生一定的影响(李苏等, 2007 ; Li et al ., 2011 , 2013a, 2013b ).地衣由于其形态结构的特殊性和变水植物的特性, 对环境变化非常敏感, 因此地衣多样性的减少经常被用来指示空气污染(Cislaghi & Nimis, 1997 ; Giordani, 2007 )、气候条件(Jovan & McCune, 2004 ; Geiser & Neitlich, 2007 ; Giordani & Incerti, 2008 )、森林结构和动态(Johansson, 2008 ; Nascimbene et al ., 2010 ).但是通过比较地衣多样性来比较不同区域间的环境条件常常遇到困难, 因为环境条件的不同经常形成高度差异的物种组成, 而如果我们利用地衣的功能特征来指示环境条件则可以很好地解决这一问题(Giordani et al ., 2012 ). ...
Temperate rainforest lichens in New Zealand: Light response of photosynthesis.
1
1997
... 地衣的变水特征使得它们的光响应特征比较复杂.地衣水分含量和净光合速率(P n )的关系能够改变它们的P n 对光照的响应(Lange & Matthes, 1981 ).低的水分含量使得地衣的P n 获得低数值的光补偿点(LCP )及光饱和点(LSP ), 而高水分含量对P n 的抑制作用也能够降低地衣的LSP 值, 但对LCP 的影响不大(Lange et al ., 1996 , 2004 ; Green et al ., 1997 ; 陈克等, 2014 ).Green等(1997)对新西兰热带雨林9种地衣光适应的研究指出, 光响应曲线参数和所测定地衣的生境相关.他们的研究发现, 林内地衣的光饱和点要低于300 μmol·m-2 ·s-1 , 而在开阔生境中地衣的光饱和点要大于549 μmol·m-2 ·s-1 .在本研究中, 8种附生地衣的LSP 都比较高(333.9-616.0 μmol·m-2 ·s-1 , 表4 ), 并且即使光强高于1000 μmol·m-2 ·s-1 也没有明显的光抑制现象, 表现出明显的“阳生植物”特性, 这在一定程度上解释了为什么在哀牢山地区附生地衣在次生林比在原生林中有着更高的丰富度(李苏等, 2007 ; Li et al ., 2013a , 2013b ).就各功能群附生地衣的LSP 而言, 枝状地衣的LSP 要明显高于其他功能群附生地衣, 这也很好地解释了它们在林冠上层和林外空旷地有较高多样性的分布格局(Li et al ., 2015 ).Green等(1997)和陈克等(2014)的研究中都指出相比于绿藻地衣, 蓝藻地衣有着较低的LSP .在我们的研究中虽然网肺衣也有着较高的LSP, 但是猫耳衣的LSP 还是低于其他功能群附生地衣.LCP 是衡量植物利用弱光能力的重要指标.在我们的研究中, 8种附生植物的LCP 都比较高(20.1-43.1 μmol·m-2 · ...
1
... 作为变水植物, 地衣的含水量随着环境条件迅速变化, 而其所有的代谢活动, 特别是净光合速率和呼吸速率也随之发生变化(Green & Lange, 1994 ).所以地衣水分关系的研究格外受到关注(Galun, 1988 ).地衣不能主动地调节水分的吸收、储存和流失(Green et al ., 2011 ), 它们利用形态学特征来促进叶状体从不同来源(雨水、潮湿空气、露水)吸收和贮存水分(Gauslaa & Coxson, 2011 ).形态结构和解剖结构是影响地衣水分贮藏的重要因素(Larson & Kershaw, 1976 ; Larson, 1981 ; Sancho & Kappen, 1989 ).在本研究中, 附生地衣的最大含水量和功能特征相关性不大, 属于同一功能团的两种附生地衣间的最大含水量也可能相差很大, 而枝状地衣的最大含水量也可能比叶状地衣的大, 这就说明解剖结构可能在附生地衣持水量上发挥了更重要的作用.Gauslaa和Coxson (2011)认为共生藻层的厚度是影响地衣物种间持水量的重要因素, 而共生藻的类型是影响其厚度的主要因素.我们所研究的两种蓝藻地衣都有着较高的持水量, 尤其是猫耳衣, 其持水量明显高于其他附生地衣.而网肺衣的持水量也比同属的针芽肺衣高100%左右.蓝藻地衣之所以有较高的持水量是因为在它们的共生藻细胞外有一层较厚的凝胶状保护鞘(Honegger et al ., 1996 ; Gauslaa & Coxson, 2011 ), 能够帮助蓝藻地衣贮存更多的水分.水分贮存能力、形态结构、解剖结构以及颜色是影响地衣吸收和丢失水分的主要因素(Palmqvist, 2000 ).比如, 相比于叶状地衣, 枝状地衣和环境之间的水分交换更快(Hartard et al ., 2009 ).在本研究中也是枝状地衣的失水速度要快于其他功能群附生地衣.枝状地衣的这种形态特征也使得它们能够从湿润的空气中快速吸收水分(Lange & Ziegler, 1986 ), 也就使得它们能够适应林外干燥的环境.相对于枝状地衣, 叶状地衣则是通过增大持水量和水分贮存能力来保持体内的水分含量从而延长生理活动的时间(Lange & Ziegler, 1986 ), 比如我们所研究的白绵腹衣, 其失水速度要明显低于其他附生地衣.对于蓝藻地衣而言, 由于只能利用液态水来进行光合作用(Lange & Ziegler, 1986 ), 所以它们通过凝胶状物质来增加持水量. ...
Factors of importance for the epiphytic vegetation aspen Populus tremula with special emphasis on chemistry and soil chemistry.
1
1995
... 气候变化、氮沉降、生物入侵和土地利用变化已经成为当前全球生物多样性的主要威胁(Sala et al ., 2000 ), 因此许多学者开始研究全球变化对生物多样性和物种分布的潜在影响.在这样的背景下, 理解限制物种分布的过程和机制就显得尤为重要(Travis et al ., 2006 ).附生地衣是森林生态系统的重要组成部分, 占据着林冠相当份量的生物量(Sipman & Harris, 1989 ; Gradstein, 1992 ), 其多样性和分布自20世纪70年代起就得到了众多学者的关注(Cornelissen & Steege, 1999 ; Holz & Gradstein, 2005 ; Hauck & Spribille, 2005 ).Aptroot和Sipman (1997)估计在热带雨林中每km2 有300种地衣, Aptroot (1997)报道在巴布亚新几内亚的热带山地雨林一株树上就有173种地衣.地衣的分布受众多因子的综合影响(Gustafasson & Eriksson, 1995 ; Goward & Arsenawlt, 2000 ; Hauck & Spribille, 2005 ), 除自身的繁殖能力和扩散能力以外, 光照、温度、湿度、干扰程度、群落类型以及宿主特征等均对附生地衣物种的组成和分布产生一定的影响(李苏等, 2007 ; Li et al ., 2011 , 2013a, 2013b ).地衣由于其形态结构的特殊性和变水植物的特性, 对环境变化非常敏感, 因此地衣多样性的减少经常被用来指示空气污染(Cislaghi & Nimis, 1997 ; Giordani, 2007 )、气候条件(Jovan & McCune, 2004 ; Geiser & Neitlich, 2007 ; Giordani & Incerti, 2008 )、森林结构和动态(Johansson, 2008 ; Nascimbene et al ., 2010 ).但是通过比较地衣多样性来比较不同区域间的环境条件常常遇到困难, 因为环境条件的不同经常形成高度差异的物种组成, 而如果我们利用地衣的功能特征来指示环境条件则可以很好地解决这一问题(Giordani et al ., 2012 ). ...
Water isotopes in desiccating lichens.
1
2009
... 作为变水植物, 地衣的含水量随着环境条件迅速变化, 而其所有的代谢活动, 特别是净光合速率和呼吸速率也随之发生变化(Green & Lange, 1994 ).所以地衣水分关系的研究格外受到关注(Galun, 1988 ).地衣不能主动地调节水分的吸收、储存和流失(Green et al ., 2011 ), 它们利用形态学特征来促进叶状体从不同来源(雨水、潮湿空气、露水)吸收和贮存水分(Gauslaa & Coxson, 2011 ).形态结构和解剖结构是影响地衣水分贮藏的重要因素(Larson & Kershaw, 1976 ; Larson, 1981 ; Sancho & Kappen, 1989 ).在本研究中, 附生地衣的最大含水量和功能特征相关性不大, 属于同一功能团的两种附生地衣间的最大含水量也可能相差很大, 而枝状地衣的最大含水量也可能比叶状地衣的大, 这就说明解剖结构可能在附生地衣持水量上发挥了更重要的作用.Gauslaa和Coxson (2011)认为共生藻层的厚度是影响地衣物种间持水量的重要因素, 而共生藻的类型是影响其厚度的主要因素.我们所研究的两种蓝藻地衣都有着较高的持水量, 尤其是猫耳衣, 其持水量明显高于其他附生地衣.而网肺衣的持水量也比同属的针芽肺衣高100%左右.蓝藻地衣之所以有较高的持水量是因为在它们的共生藻细胞外有一层较厚的凝胶状保护鞘(Honegger et al ., 1996 ; Gauslaa & Coxson, 2011 ), 能够帮助蓝藻地衣贮存更多的水分.水分贮存能力、形态结构、解剖结构以及颜色是影响地衣吸收和丢失水分的主要因素(Palmqvist, 2000 ).比如, 相比于叶状地衣, 枝状地衣和环境之间的水分交换更快(Hartard et al ., 2009 ).在本研究中也是枝状地衣的失水速度要快于其他功能群附生地衣.枝状地衣的这种形态特征也使得它们能够从湿润的空气中快速吸收水分(Lange & Ziegler, 1986 ), 也就使得它们能够适应林外干燥的环境.相对于枝状地衣, 叶状地衣则是通过增大持水量和水分贮存能力来保持体内的水分含量从而延长生理活动的时间(Lange & Ziegler, 1986 ), 比如我们所研究的白绵腹衣, 其失水速度要明显低于其他附生地衣.对于蓝藻地衣而言, 由于只能利用液态水来进行光合作用(Lange & Ziegler, 1986 ), 所以它们通过凝胶状物质来增加持水量. ...
Lichen diversity on steppe slopes in the northern Mongolian mountain taiga and its dependence on microclimate.
2007
The significance of precipitation and substrate chemistry for epiphytic lichen diversity in spruce-fir forests of the Salish Mountains, northwestern Montana.
2
2005
... 气候变化、氮沉降、生物入侵和土地利用变化已经成为当前全球生物多样性的主要威胁(Sala et al ., 2000 ), 因此许多学者开始研究全球变化对生物多样性和物种分布的潜在影响.在这样的背景下, 理解限制物种分布的过程和机制就显得尤为重要(Travis et al ., 2006 ).附生地衣是森林生态系统的重要组成部分, 占据着林冠相当份量的生物量(Sipman & Harris, 1989 ; Gradstein, 1992 ), 其多样性和分布自20世纪70年代起就得到了众多学者的关注(Cornelissen & Steege, 1999 ; Holz & Gradstein, 2005 ; Hauck & Spribille, 2005 ).Aptroot和Sipman (1997)估计在热带雨林中每km2 有300种地衣, Aptroot (1997)报道在巴布亚新几内亚的热带山地雨林一株树上就有173种地衣.地衣的分布受众多因子的综合影响(Gustafasson & Eriksson, 1995 ; Goward & Arsenawlt, 2000 ; Hauck & Spribille, 2005 ), 除自身的繁殖能力和扩散能力以外, 光照、温度、湿度、干扰程度、群落类型以及宿主特征等均对附生地衣物种的组成和分布产生一定的影响(李苏等, 2007 ; Li et al ., 2011 , 2013a, 2013b ).地衣由于其形态结构的特殊性和变水植物的特性, 对环境变化非常敏感, 因此地衣多样性的减少经常被用来指示空气污染(Cislaghi & Nimis, 1997 ; Giordani, 2007 )、气候条件(Jovan & McCune, 2004 ; Geiser & Neitlich, 2007 ; Giordani & Incerti, 2008 )、森林结构和动态(Johansson, 2008 ; Nascimbene et al ., 2010 ).但是通过比较地衣多样性来比较不同区域间的环境条件常常遇到困难, 因为环境条件的不同经常形成高度差异的物种组成, 而如果我们利用地衣的功能特征来指示环境条件则可以很好地解决这一问题(Giordani et al ., 2012 ). ...
... ; Hauck & Spribille, 2005 ), 除自身的繁殖能力和扩散能力以外, 光照、温度、湿度、干扰程度、群落类型以及宿主特征等均对附生地衣物种的组成和分布产生一定的影响(李苏等, 2007 ; Li et al ., 2011 , 2013a, 2013b ).地衣由于其形态结构的特殊性和变水植物的特性, 对环境变化非常敏感, 因此地衣多样性的减少经常被用来指示空气污染(Cislaghi & Nimis, 1997 ; Giordani, 2007 )、气候条件(Jovan & McCune, 2004 ; Geiser & Neitlich, 2007 ; Giordani & Incerti, 2008 )、森林结构和动态(Johansson, 2008 ; Nascimbene et al ., 2010 ).但是通过比较地衣多样性来比较不同区域间的环境条件常常遇到困难, 因为环境条件的不同经常形成高度差异的物种组成, 而如果我们利用地衣的功能特征来指示环境条件则可以很好地解决这一问题(Giordani et al ., 2012 ). ...
Cryptogamic epiphytes in primary and recovering upper montane oak forests of Costa Rica—Species richness, community composition and ecology.
1
2005
... 气候变化、氮沉降、生物入侵和土地利用变化已经成为当前全球生物多样性的主要威胁(Sala et al ., 2000 ), 因此许多学者开始研究全球变化对生物多样性和物种分布的潜在影响.在这样的背景下, 理解限制物种分布的过程和机制就显得尤为重要(Travis et al ., 2006 ).附生地衣是森林生态系统的重要组成部分, 占据着林冠相当份量的生物量(Sipman & Harris, 1989 ; Gradstein, 1992 ), 其多样性和分布自20世纪70年代起就得到了众多学者的关注(Cornelissen & Steege, 1999 ; Holz & Gradstein, 2005 ; Hauck & Spribille, 2005 ).Aptroot和Sipman (1997)估计在热带雨林中每km2 有300种地衣, Aptroot (1997)报道在巴布亚新几内亚的热带山地雨林一株树上就有173种地衣.地衣的分布受众多因子的综合影响(Gustafasson & Eriksson, 1995 ; Goward & Arsenawlt, 2000 ; Hauck & Spribille, 2005 ), 除自身的繁殖能力和扩散能力以外, 光照、温度、湿度、干扰程度、群落类型以及宿主特征等均对附生地衣物种的组成和分布产生一定的影响(李苏等, 2007 ; Li et al ., 2011 , 2013a, 2013b ).地衣由于其形态结构的特殊性和变水植物的特性, 对环境变化非常敏感, 因此地衣多样性的减少经常被用来指示空气污染(Cislaghi & Nimis, 1997 ; Giordani, 2007 )、气候条件(Jovan & McCune, 2004 ; Geiser & Neitlich, 2007 ; Giordani & Incerti, 2008 )、森林结构和动态(Johansson, 2008 ; Nascimbene et al ., 2010 ).但是通过比较地衣多样性来比较不同区域间的环境条件常常遇到困难, 因为环境条件的不同经常形成高度差异的物种组成, 而如果我们利用地衣的功能特征来指示环境条件则可以很好地解决这一问题(Giordani et al ., 2012 ). ...
Drought-induced structural alterations at the mycobiont-photobiont interface in a range of foliose macrolichens.
1
1996
... 作为变水植物, 地衣的含水量随着环境条件迅速变化, 而其所有的代谢活动, 特别是净光合速率和呼吸速率也随之发生变化(Green & Lange, 1994 ).所以地衣水分关系的研究格外受到关注(Galun, 1988 ).地衣不能主动地调节水分的吸收、储存和流失(Green et al ., 2011 ), 它们利用形态学特征来促进叶状体从不同来源(雨水、潮湿空气、露水)吸收和贮存水分(Gauslaa & Coxson, 2011 ).形态结构和解剖结构是影响地衣水分贮藏的重要因素(Larson & Kershaw, 1976 ; Larson, 1981 ; Sancho & Kappen, 1989 ).在本研究中, 附生地衣的最大含水量和功能特征相关性不大, 属于同一功能团的两种附生地衣间的最大含水量也可能相差很大, 而枝状地衣的最大含水量也可能比叶状地衣的大, 这就说明解剖结构可能在附生地衣持水量上发挥了更重要的作用.Gauslaa和Coxson (2011)认为共生藻层的厚度是影响地衣物种间持水量的重要因素, 而共生藻的类型是影响其厚度的主要因素.我们所研究的两种蓝藻地衣都有着较高的持水量, 尤其是猫耳衣, 其持水量明显高于其他附生地衣.而网肺衣的持水量也比同属的针芽肺衣高100%左右.蓝藻地衣之所以有较高的持水量是因为在它们的共生藻细胞外有一层较厚的凝胶状保护鞘(Honegger et al ., 1996 ; Gauslaa & Coxson, 2011 ), 能够帮助蓝藻地衣贮存更多的水分.水分贮存能力、形态结构、解剖结构以及颜色是影响地衣吸收和丢失水分的主要因素(Palmqvist, 2000 ).比如, 相比于叶状地衣, 枝状地衣和环境之间的水分交换更快(Hartard et al ., 2009 ).在本研究中也是枝状地衣的失水速度要快于其他功能群附生地衣.枝状地衣的这种形态特征也使得它们能够从湿润的空气中快速吸收水分(Lange & Ziegler, 1986 ), 也就使得它们能够适应林外干燥的环境.相对于枝状地衣, 叶状地衣则是通过增大持水量和水分贮存能力来保持体内的水分含量从而延长生理活动的时间(Lange & Ziegler, 1986 ), 比如我们所研究的白绵腹衣, 其失水速度要明显低于其他附生地衣.对于蓝藻地衣而言, 由于只能利用液态水来进行光合作用(Lange & Ziegler, 1986 ), 所以它们通过凝胶状物质来增加持水量. ...
Consequences of disturbance on epiphytic lichens in boreal and near boreal forests.
1
2008
... 气候变化、氮沉降、生物入侵和土地利用变化已经成为当前全球生物多样性的主要威胁(Sala et al ., 2000 ), 因此许多学者开始研究全球变化对生物多样性和物种分布的潜在影响.在这样的背景下, 理解限制物种分布的过程和机制就显得尤为重要(Travis et al ., 2006 ).附生地衣是森林生态系统的重要组成部分, 占据着林冠相当份量的生物量(Sipman & Harris, 1989 ; Gradstein, 1992 ), 其多样性和分布自20世纪70年代起就得到了众多学者的关注(Cornelissen & Steege, 1999 ; Holz & Gradstein, 2005 ; Hauck & Spribille, 2005 ).Aptroot和Sipman (1997)估计在热带雨林中每km2 有300种地衣, Aptroot (1997)报道在巴布亚新几内亚的热带山地雨林一株树上就有173种地衣.地衣的分布受众多因子的综合影响(Gustafasson & Eriksson, 1995 ; Goward & Arsenawlt, 2000 ; Hauck & Spribille, 2005 ), 除自身的繁殖能力和扩散能力以外, 光照、温度、湿度、干扰程度、群落类型以及宿主特征等均对附生地衣物种的组成和分布产生一定的影响(李苏等, 2007 ; Li et al ., 2011 , 2013a, 2013b ).地衣由于其形态结构的特殊性和变水植物的特性, 对环境变化非常敏感, 因此地衣多样性的减少经常被用来指示空气污染(Cislaghi & Nimis, 1997 ; Giordani, 2007 )、气候条件(Jovan & McCune, 2004 ; Geiser & Neitlich, 2007 ; Giordani & Incerti, 2008 )、森林结构和动态(Johansson, 2008 ; Nascimbene et al ., 2010 ).但是通过比较地衣多样性来比较不同区域间的环境条件常常遇到困难, 因为环境条件的不同经常形成高度差异的物种组成, 而如果我们利用地衣的功能特征来指示环境条件则可以很好地解决这一问题(Giordani et al ., 2012 ). ...
Regional variation in epiphytic macrolichen communities in northern and central California forests.
1
2004
... 气候变化、氮沉降、生物入侵和土地利用变化已经成为当前全球生物多样性的主要威胁(Sala et al ., 2000 ), 因此许多学者开始研究全球变化对生物多样性和物种分布的潜在影响.在这样的背景下, 理解限制物种分布的过程和机制就显得尤为重要(Travis et al ., 2006 ).附生地衣是森林生态系统的重要组成部分, 占据着林冠相当份量的生物量(Sipman & Harris, 1989 ; Gradstein, 1992 ), 其多样性和分布自20世纪70年代起就得到了众多学者的关注(Cornelissen & Steege, 1999 ; Holz & Gradstein, 2005 ; Hauck & Spribille, 2005 ).Aptroot和Sipman (1997)估计在热带雨林中每km2 有300种地衣, Aptroot (1997)报道在巴布亚新几内亚的热带山地雨林一株树上就有173种地衣.地衣的分布受众多因子的综合影响(Gustafasson & Eriksson, 1995 ; Goward & Arsenawlt, 2000 ; Hauck & Spribille, 2005 ), 除自身的繁殖能力和扩散能力以外, 光照、温度、湿度、干扰程度、群落类型以及宿主特征等均对附生地衣物种的组成和分布产生一定的影响(李苏等, 2007 ; Li et al ., 2011 , 2013a, 2013b ).地衣由于其形态结构的特殊性和变水植物的特性, 对环境变化非常敏感, 因此地衣多样性的减少经常被用来指示空气污染(Cislaghi & Nimis, 1997 ; Giordani, 2007 )、气候条件(Jovan & McCune, 2004 ; Geiser & Neitlich, 2007 ; Giordani & Incerti, 2008 )、森林结构和动态(Johansson, 2008 ; Nascimbene et al ., 2010 ).但是通过比较地衣多样性来比较不同区域间的环境条件常常遇到困难, 因为环境条件的不同经常形成高度差异的物种组成, 而如果我们利用地衣的功能特征来指示环境条件则可以很好地解决这一问题(Giordani et al ., 2012 ). ...
Diatoms living inside the thallus of the green algal lichen Coenogonium linkii in neotropical lowland rain forests.
1
2004
... 根据国外的相关研究, 地衣按照共生藻、生活型和繁殖策略的不同可以划分为蓝藻地衣(cyano-区性lichens, CYL)、壳状地衣(crustose lichens, CRL)、枝状地衣(fruticose lichens, FRL)、狭叶地衣(narrowly lobed foliose lichens, NFL)、阔叶地衣(broadly lobed foliose lichens, BFL)、裂芽地衣(isidiate lichens, ISL)、粉芽地衣(sorediate lichens, SOL)和有性地衣(sexual lichens, SEL) 8个功能群(McCune, 1993 ; Nimis & Martellos, 2008 ).调查研究显示地衣呈现出与它们功能特征相关的分布格局(Ellis & Coppins, 2006 ; Lakatos et al ., 2006 ; Zedda et al ., 2011 ).比如在热带山地雨林分布着更多的枝状和叶状地衣, 但在低地雨林却是壳状地衣占据优势, 枝状地衣和叶状地衣很稀少(Montfoot & Ek, 1990 ; Sipman & Tan, 1990 ; Lakatos et al ., 2004 ).Hauck等(2007)在蒙古的针叶林发现, 绿藻地衣在群落中处于优势地位, 蓝藻地衣只在有着较高可利用水源的地方有分布.在最近的研究中, 地衣的生活型、繁殖策略、共生藻等功能特征被广泛用来指示地衣群落对环境条件的适应(DÍaz & Cabido, 2001 ; Cornelissen et al ., 2007 ; Glime, 2007 ), 因为地衣的功能特征可以不依赖物种多样性和组成而直接和环境因子建立关联(Ricotta & Bacaro, 2010 ; Webb et al ., 2010 ).Lange等(1994)发现在蓝藻地衣较多的生境中经常有高的积水.Giordani等(2012)基于意大利森林生态系统网络研究了地衣功能特征和环境条件的关系, 结果表明: 壳状和叶状地衣的分布和降水量密切相关; 枝状地衣的分布受海拔的影响较大, 同时和森林的密闭度负相关; 狭叶和阔叶地衣都受到酸性沉积物的负面影响.Giordani等(2014)调查发现阔叶地衣只有在高潜在径流的条件下才能在高辐射的生境中分布, 同时他们还观察到在枝状地衣的生境中有高的潜在径流. ...
Functional character- istics of corticolous lichens in the understory of a tropical lowland and rain forest.
1
2006
... 根据国外的相关研究, 地衣按照共生藻、生活型和繁殖策略的不同可以划分为蓝藻地衣(cyano-区性lichens, CYL)、壳状地衣(crustose lichens, CRL)、枝状地衣(fruticose lichens, FRL)、狭叶地衣(narrowly lobed foliose lichens, NFL)、阔叶地衣(broadly lobed foliose lichens, BFL)、裂芽地衣(isidiate lichens, ISL)、粉芽地衣(sorediate lichens, SOL)和有性地衣(sexual lichens, SEL) 8个功能群(McCune, 1993 ; Nimis & Martellos, 2008 ).调查研究显示地衣呈现出与它们功能特征相关的分布格局(Ellis & Coppins, 2006 ; Lakatos et al ., 2006 ; Zedda et al ., 2011 ).比如在热带山地雨林分布着更多的枝状和叶状地衣, 但在低地雨林却是壳状地衣占据优势, 枝状地衣和叶状地衣很稀少(Montfoot & Ek, 1990 ; Sipman & Tan, 1990 ; Lakatos et al ., 2004 ).Hauck等(2007)在蒙古的针叶林发现, 绿藻地衣在群落中处于优势地位, 蓝藻地衣只在有着较高可利用水源的地方有分布.在最近的研究中, 地衣的生活型、繁殖策略、共生藻等功能特征被广泛用来指示地衣群落对环境条件的适应(DÍaz & Cabido, 2001 ; Cornelissen et al ., 2007 ; Glime, 2007 ), 因为地衣的功能特征可以不依赖物种多样性和组成而直接和环境因子建立关联(Ricotta & Bacaro, 2010 ; Webb et al ., 2010 ).Lange等(1994)发现在蓝藻地衣较多的生境中经常有高的积水.Giordani等(2012)基于意大利森林生态系统网络研究了地衣功能特征和环境条件的关系, 结果表明: 壳状和叶状地衣的分布和降水量密切相关; 枝状地衣的分布受海拔的影响较大, 同时和森林的密闭度负相关; 狭叶和阔叶地衣都受到酸性沉积物的负面影响.Giordani等(2014)调查发现阔叶地衣只有在高潜在径流的条件下才能在高辐射的生境中分布, 同时他们还观察到在枝状地衣的生境中有高的潜在径流. ...
Moisture content and CO2 exchange in lichens. I. In?uence of temperature on moisture dependent net photosynthesis and dark respiration in Ramalina maciformis.
2
1980
... 式中P n 是净光合速率(μmol·m-2 ·s-1 ), y 0 、a、b、c 是回归系数.根据回归方程求得光合作用最适水分含量为 P n = 0时的水分含量).为了更好地说明光合作用和水分含量的关系, 我们参照Lange (1980) 的处理方法, 利用上述方程求出各附生地衣在水分补偿点、最适水分含量及最大含水量10%、20%、30%、40%、50%、60%时的光合速率, 同样利用三次多项式回归模型模拟相对光合速率(光合速率占最适水分含量下光合速率的百分比)和水分含量的关系, 并求出模型曲线的初始斜率, 用来说明附生地衣在干燥状态下对水分含量变化的敏感性. ...
... 各附生地衣的光合速率对水分变化的响应趋势是相同的(图4 ), 均表现为先随含水量的增加而升高, 当含水量达到一定程度后, 光合速率随含水量的增加而降低.但是, 高水分含量对不同附生地衣光合作用的影响程度是不同的, 其中对皮革肾岛衣的影响最小, 而对针芽肺衣、网肺衣、云南袋衣、多花松萝光合速率的抑制能达到最大光合速率的50%, 甚至更多.利用三次多项式回归模型能够较好地模拟各附生地衣光合速率随水分含量的变化(图4 , R 2 为0.73-0.93).根据模型方程可以计算出各附生地衣光合作用的最适水分含量、水分补偿点.不同附生地衣光合作用的最适水分含量是存在差异的, 其中针芽肺衣、多花松萝、裂髓树花的最适含水量在100%左右, 而猫耳衣和白棉腹衣则超过了300% (表3 ).附生地衣光合速率的最适含水量近似线性(图5 , R 2 = 0.63)地随最大含水量增加.而光合作用的水分补偿点和最适水分含量无明显的关联性(图5 ), 最适含水量较高的猫耳衣和白绵腹衣的水分补偿点反而较低, 不到干质量的20%, 而网肺衣的水分补偿点将近达到干质量的50%, 分布于较干旱生境中枝状地衣的水分补偿点则比较高, 都达到其干质量的30%左右(表3 ).相对净光合速率(净光合速率占最大净光合速率的百分比)和水分含量关系曲线(图5 )的初始斜率能在一定程度上说明附生地衣在干燥情况下光合作用对水分的敏感性(Lange, 1980 ).在本研究中, 光合作用最适水分含量相对较高的白绵腹衣和猫耳衣的初始斜率(0.5-0.64)都比较低, 反而最适水分含量较低的针芽肺衣、多花松萝和裂髓树花的初始斜率(1.92-2.59)较高, 说明在干燥状态下它们的光合作用对水分变化较敏感. ...
Photosynthetic productivity of the epilithic lichen Lecanora muralis: Long-term field monitoring of CO2 exchange and its physiological interpretation. I. Dependence of photosynthesis on water content, light, temperature, and CO2 concentration from laboratory measurements.
1
2002
... 水分含量和光照是决定地衣光合速率的主要因素(Lange, 2002 ).作为变水植物, 地衣的生理代谢, 特别是光合作用, 受到水分含量的强烈影响, 过低和过高的水分含量都将限制地衣的光合作用(Lange et al ., 1993 ).Zotz等(1998)在对热带阔叶地衣Sticta tomentosa 气体交换的研究中发现, 在最大含水量时, 其净光合速率比最大光合速率的50%还小.Lange (2002)在对石附生地衣Lecanora muralis 的研究中发现, 在干燥和过饱和状态下, Lecanora muralis 的净光合速率都等于0或接近于0.在本研究中, 我们同样发现过高或过低的含水量都明显地限制着各附生地衣的光合作用.在陈克等(2014)对本地区3种附生地衣的光合生理研究中, 网肺衣在含水量为150%-200%时拥有最适光合水分条件, 这和我们所得到的网肺衣最适光合含水量为172.9%相符.过高含水量对不同附生地衣光合作用的抑制程度是不同的, 其中对云南袋衣的限制作用最明显, 而对皮革肾岛衣的影响则比较小, 这可能是由于不同附生地衣适应环境的策略不同造成的.云南袋衣可能通过增加叶状体厚度及扩大细胞间隙来增加持水量和降低失水速率, 这样带来的结果是细胞间隙间过多的水分阻碍了CO2 扩散, 从而影响了光合作用.皮革肾岛衣则可能通过增加表面积来加快叶状体和环境水分的交换, 这样就能降低过多水分对CO2 扩散的影响, 但是也降低持水量和加快了失水速率.通过分析各附生地衣光合作用的最适含水量, 枝状地衣具有较低的光合作用最适含水量, 这可能与它们分布于干燥生境有密切关系, 并且我们的研究还发现枝状地衣在干燥状态下对水分变化很敏感, 这就意味 ...
Temperate rainforest lichens in New Zealand: High thallus water content can severely limit photosynthetic CO2 exchange.
1
1993
... 水分含量和光照是决定地衣光合速率的主要因素(Lange, 2002 ).作为变水植物, 地衣的生理代谢, 特别是光合作用, 受到水分含量的强烈影响, 过低和过高的水分含量都将限制地衣的光合作用(Lange et al ., 1993 ).Zotz等(1998)在对热带阔叶地衣Sticta tomentosa 气体交换的研究中发现, 在最大含水量时, 其净光合速率比最大光合速率的50%还小.Lange (2002)在对石附生地衣Lecanora muralis 的研究中发现, 在干燥和过饱和状态下, Lecanora muralis 的净光合速率都等于0或接近于0.在本研究中, 我们同样发现过高或过低的含水量都明显地限制着各附生地衣的光合作用.在陈克等(2014)对本地区3种附生地衣的光合生理研究中, 网肺衣在含水量为150%-200%时拥有最适光合水分条件, 这和我们所得到的网肺衣最适光合含水量为172.9%相符.过高含水量对不同附生地衣光合作用的抑制程度是不同的, 其中对云南袋衣的限制作用最明显, 而对皮革肾岛衣的影响则比较小, 这可能是由于不同附生地衣适应环境的策略不同造成的.云南袋衣可能通过增加叶状体厚度及扩大细胞间隙来增加持水量和降低失水速率, 这样带来的结果是细胞间隙间过多的水分阻碍了CO2 扩散, 从而影响了光合作用.皮革肾岛衣则可能通过增加表面积来加快叶状体和环境水分的交换, 这样就能降低过多水分对CO2 扩散的影响, 但是也降低持水量和加快了失水速率.通过分析各附生地衣光合作用的最适含水量, 枝状地衣具有较低的光合作用最适含水量, 这可能与它们分布于干燥生境有密切关系, 并且我们的研究还发现枝状地衣在干燥状态下对水分变化很敏感, 这就意味 ...
Lichen carbon gain under tropical conditions: Water relations and CO2 exchange of three Lobariaceae species of a lower montane rainforest in Panama.
2
2004
... 地衣的变水特征使得它们的光响应特征比较复杂.地衣水分含量和净光合速率(P n )的关系能够改变它们的P n 对光照的响应(Lange & Matthes, 1981 ).低的水分含量使得地衣的P n 获得低数值的光补偿点(LCP )及光饱和点(LSP ), 而高水分含量对P n 的抑制作用也能够降低地衣的LSP 值, 但对LCP 的影响不大(Lange et al ., 1996 , 2004 ; Green et al ., 1997 ; 陈克等, 2014 ).Green等(1997)对新西兰热带雨林9种地衣光适应的研究指出, 光响应曲线参数和所测定地衣的生境相关.他们的研究发现, 林内地衣的光饱和点要低于300 μmol·m-2 ·s-1 , 而在开阔生境中地衣的光饱和点要大于549 μmol·m-2 ·s-1 .在本研究中, 8种附生地衣的LSP 都比较高(333.9-616.0 μmol·m-2 ·s-1 , 表4 ), 并且即使光强高于1000 μmol·m-2 ·s-1 也没有明显的光抑制现象, 表现出明显的“阳生植物”特性, 这在一定程度上解释了为什么在哀牢山地区附生地衣在次生林比在原生林中有着更高的丰富度(李苏等, 2007 ; Li et al ., 2013a , 2013b ).就各功能群附生地衣的LSP 而言, 枝状地衣的LSP 要明显高于其他功能群附生地衣, 这也很好地解释了它们在林冠上层和林外空旷地有较高多样性的分布格局(Li et al ., 2015 ).Green等(1997)和陈克等(2014)的研究中都指出相比于绿藻地衣, 蓝藻地衣有着较低的LSP .在我们的研究中虽然网肺衣也有着较高的LSP, 但是猫耳衣的LSP 还是低于其他功能群附生地衣.LCP 是衡量植物利用弱光能力的重要指标.在我们的研究中, 8种附生植物的LCP 都比较高(20.1-43.1 μmol·m-2 · ...
... s-1 , 表4 ), 除了蓝藻地衣外, 其他功能群附生地衣的LCP 都要高于Green等(1997)在新西兰热带雨林所研究的林内林缘地衣LCP (4.3-29 μmol·m-2 ·s-1 ), 而低于林外地衣LCP (≥50 μmol·m-2 ·s-1 ).在本研究中, 蓝藻地衣的LCP 显著低于其他功能群的附生地衣, 这和在哀牢山地区观察到的蓝藻地衣能分布在较阴暗生境相符(Li et al ., 2015 ).总之, 蓝藻地衣能够利用的光谱比较宽(较低的LCP 和较高的LSP ), 所以蓝藻地衣既能分布在较强太阳辐射的生境中, 又对弱光有一定的适应能力(Rikkien, 2009 ), 而绿藻地衣大多只能分布在太阳辐射较高的生境中.地衣的光合能力和其氮素含量密切相关(Lange et al ., 2004 ).在Lange等(2004)对巴拿马热带雨林6种地衣碳获取的研究中, 具有高浓度氮含量的牛皮叶属地衣的P n (4.9-5.8 μmol·m-2 ·s-1 )要比肺衣属的(2.0-2.9 μmol·m-2 ·s-1 )高很多.我们所研究的8种附生地衣的P n 都比较小(0.92-2.84 μmol·m-2 ·s-1 , 表4 ), 两种肺衣属(针芽肺衣和网肺衣)地衣的P n (分别为1.77 和2.84 μmol·m-2 ·s-1 )和Lange等(2004)测得的同属地衣的P n 相差不大, 并且属于蓝藻地衣的网肺衣的 P n 要比属于绿藻地衣的针芽肺衣的高, 这可能和网肺衣具有固氮能力有关.光合能力的差异和功能群的相关性不大, 可能更多地和地衣氮浓度含量相关. ...
Photosynthetic depression at high thallus water content in lichens: Concurrent use of gas exchange and fluorescence techniques with a cyanobacterial and a green algal Peltigera species.
1
1996
... 地衣的变水特征使得它们的光响应特征比较复杂.地衣水分含量和净光合速率(P n )的关系能够改变它们的P n 对光照的响应(Lange & Matthes, 1981 ).低的水分含量使得地衣的P n 获得低数值的光补偿点(LCP )及光饱和点(LSP ), 而高水分含量对P n 的抑制作用也能够降低地衣的LSP 值, 但对LCP 的影响不大(Lange et al ., 1996 , 2004 ; Green et al ., 1997 ; 陈克等, 2014 ).Green等(1997)对新西兰热带雨林9种地衣光适应的研究指出, 光响应曲线参数和所测定地衣的生境相关.他们的研究发现, 林内地衣的光饱和点要低于300 μmol·m-2 ·s-1 , 而在开阔生境中地衣的光饱和点要大于549 μmol·m-2 ·s-1 .在本研究中, 8种附生地衣的LSP 都比较高(333.9-616.0 μmol·m-2 ·s-1 , 表4 ), 并且即使光强高于1000 μmol·m-2 ·s-1 也没有明显的光抑制现象, 表现出明显的“阳生植物”特性, 这在一定程度上解释了为什么在哀牢山地区附生地衣在次生林比在原生林中有着更高的丰富度(李苏等, 2007 ; Li et al ., 2013a , 2013b ).就各功能群附生地衣的LSP 而言, 枝状地衣的LSP 要明显高于其他功能群附生地衣, 这也很好地解释了它们在林冠上层和林外空旷地有较高多样性的分布格局(Li et al ., 2015 ).Green等(1997)和陈克等(2014)的研究中都指出相比于绿藻地衣, 蓝藻地衣有着较低的LSP .在我们的研究中虽然网肺衣也有着较高的LSP, 但是猫耳衣的LSP 还是低于其他功能群附生地衣.LCP 是衡量植物利用弱光能力的重要指标.在我们的研究中, 8种附生植物的LCP 都比较高(20.1-43.1 μmol·m-2 · ...
Moisture-dependent CO2 exchange of lichens.
1
1981
... 地衣的变水特征使得它们的光响应特征比较复杂.地衣水分含量和净光合速率(P n )的关系能够改变它们的P n 对光照的响应(Lange & Matthes, 1981 ).低的水分含量使得地衣的P n 获得低数值的光补偿点(LCP )及光饱和点(LSP ), 而高水分含量对P n 的抑制作用也能够降低地衣的LSP 值, 但对LCP 的影响不大(Lange et al ., 1996 , 2004 ; Green et al ., 1997 ; 陈克等, 2014 ).Green等(1997)对新西兰热带雨林9种地衣光适应的研究指出, 光响应曲线参数和所测定地衣的生境相关.他们的研究发现, 林内地衣的光饱和点要低于300 μmol·m-2 ·s-1 , 而在开阔生境中地衣的光饱和点要大于549 μmol·m-2 ·s-1 .在本研究中, 8种附生地衣的LSP 都比较高(333.9-616.0 μmol·m-2 ·s-1 , 表4 ), 并且即使光强高于1000 μmol·m-2 ·s-1 也没有明显的光抑制现象, 表现出明显的“阳生植物”特性, 这在一定程度上解释了为什么在哀牢山地区附生地衣在次生林比在原生林中有着更高的丰富度(李苏等, 2007 ; Li et al ., 2013a , 2013b ).就各功能群附生地衣的LSP 而言, 枝状地衣的LSP 要明显高于其他功能群附生地衣, 这也很好地解释了它们在林冠上层和林外空旷地有较高多样性的分布格局(Li et al ., 2015 ).Green等(1997)和陈克等(2014)的研究中都指出相比于绿藻地衣, 蓝藻地衣有着较低的LSP .在我们的研究中虽然网肺衣也有着较高的LSP, 但是猫耳衣的LSP 还是低于其他功能群附生地衣.LCP 是衡量植物利用弱光能力的重要指标.在我们的研究中, 8种附生植物的LCP 都比较高(20.1-43.1 μmol·m-2 · ...
Mikroklima, Wassergehalt und Photosynthese von Flechten in der küstennahen Nebelzone der Namib-Wüste: Messungen während der herbstlichen Witterungsperiod.
1
1991
... 式中I 是光照强度(μmol·m-2 ·s-1 ), R d 暗呼吸速率(μmol·m-2 ·s-1 ), a、b、c 是回归系数.主要的光合作用特征参数根据叶子飘(2010)进行计算, 包括光照饱和时的光合速率(P nmax ; μmol·m-2 ·s-1 )、光饱和点(LSP; μmol·m-2 ·s-1 )、光补偿点(LCP; μmol·m-2 ·s-1 ), 其中我们参照Lange等(1991) 将光饱和点定义为净光合速率达到最大净光合速率90%时的最小光强.不同附生地衣光响应曲线参数的差异性利用单因素方差分析和Tukey’s HSD多重配对进行分析. ...
Photosynthesis and water relations of lichen soil crusts—Field measurements in the coastal fog zone of the Namib Desert.
1994
3
... 作为变水植物, 地衣的含水量随着环境条件迅速变化, 而其所有的代谢活动, 特别是净光合速率和呼吸速率也随之发生变化(Green & Lange, 1994 ).所以地衣水分关系的研究格外受到关注(Galun, 1988 ).地衣不能主动地调节水分的吸收、储存和流失(Green et al ., 2011 ), 它们利用形态学特征来促进叶状体从不同来源(雨水、潮湿空气、露水)吸收和贮存水分(Gauslaa & Coxson, 2011 ).形态结构和解剖结构是影响地衣水分贮藏的重要因素(Larson & Kershaw, 1976 ; Larson, 1981 ; Sancho & Kappen, 1989 ).在本研究中, 附生地衣的最大含水量和功能特征相关性不大, 属于同一功能团的两种附生地衣间的最大含水量也可能相差很大, 而枝状地衣的最大含水量也可能比叶状地衣的大, 这就说明解剖结构可能在附生地衣持水量上发挥了更重要的作用.Gauslaa和Coxson (2011)认为共生藻层的厚度是影响地衣物种间持水量的重要因素, 而共生藻的类型是影响其厚度的主要因素.我们所研究的两种蓝藻地衣都有着较高的持水量, 尤其是猫耳衣, 其持水量明显高于其他附生地衣.而网肺衣的持水量也比同属的针芽肺衣高100%左右.蓝藻地衣之所以有较高的持水量是因为在它们的共生藻细胞外有一层较厚的凝胶状保护鞘(Honegger et al ., 1996 ; Gauslaa & Coxson, 2011 ), 能够帮助蓝藻地衣贮存更多的水分.水分贮存能力、形态结构、解剖结构以及颜色是影响地衣吸收和丢失水分的主要因素(Palmqvist, 2000 ).比如, 相比于叶状地衣, 枝状地衣和环境之间的水分交换更快(Hartard et al ., 2009 ).在本研究中也是枝状地衣的失水速度要快于其他功能群附生地衣.枝状地衣的这种形态特征也使得它们能够从湿润的空气中快速吸收水分(Lange & Ziegler, 1986 ), 也就使得它们能够适应林外干燥的环境.相对于枝状地衣, 叶状地衣则是通过增大持水量和水分贮存能力来保持体内的水分含量从而延长生理活动的时间(Lange & Ziegler, 1986 ), 比如我们所研究的白绵腹衣, 其失水速度要明显低于其他附生地衣.对于蓝藻地衣而言, 由于只能利用液态水来进行光合作用(Lange & Ziegler, 1986 ), 所以它们通过凝胶状物质来增加持水量. ...
... ), 也就使得它们能够适应林外干燥的环境.相对于枝状地衣, 叶状地衣则是通过增大持水量和水分贮存能力来保持体内的水分含量从而延长生理活动的时间(Lange & Ziegler, 1986 ), 比如我们所研究的白绵腹衣, 其失水速度要明显低于其他附生地衣.对于蓝藻地衣而言, 由于只能利用液态水来进行光合作用(Lange & Ziegler, 1986 ), 所以它们通过凝胶状物质来增加持水量. ...
... ), 比如我们所研究的白绵腹衣, 其失水速度要明显低于其他附生地衣.对于蓝藻地衣而言, 由于只能利用液态水来进行光合作用(Lange & Ziegler, 1986 ), 所以它们通过凝胶状物质来增加持水量. ...
Differential wetting in some lichens and mosses: The role of morphology.
1
1981
... 作为变水植物, 地衣的含水量随着环境条件迅速变化, 而其所有的代谢活动, 特别是净光合速率和呼吸速率也随之发生变化(Green & Lange, 1994 ).所以地衣水分关系的研究格外受到关注(Galun, 1988 ).地衣不能主动地调节水分的吸收、储存和流失(Green et al ., 2011 ), 它们利用形态学特征来促进叶状体从不同来源(雨水、潮湿空气、露水)吸收和贮存水分(Gauslaa & Coxson, 2011 ).形态结构和解剖结构是影响地衣水分贮藏的重要因素(Larson & Kershaw, 1976 ; Larson, 1981 ; Sancho & Kappen, 1989 ).在本研究中, 附生地衣的最大含水量和功能特征相关性不大, 属于同一功能团的两种附生地衣间的最大含水量也可能相差很大, 而枝状地衣的最大含水量也可能比叶状地衣的大, 这就说明解剖结构可能在附生地衣持水量上发挥了更重要的作用.Gauslaa和Coxson (2011)认为共生藻层的厚度是影响地衣物种间持水量的重要因素, 而共生藻的类型是影响其厚度的主要因素.我们所研究的两种蓝藻地衣都有着较高的持水量, 尤其是猫耳衣, 其持水量明显高于其他附生地衣.而网肺衣的持水量也比同属的针芽肺衣高100%左右.蓝藻地衣之所以有较高的持水量是因为在它们的共生藻细胞外有一层较厚的凝胶状保护鞘(Honegger et al ., 1996 ; Gauslaa & Coxson, 2011 ), 能够帮助蓝藻地衣贮存更多的水分.水分贮存能力、形态结构、解剖结构以及颜色是影响地衣吸收和丢失水分的主要因素(Palmqvist, 2000 ).比如, 相比于叶状地衣, 枝状地衣和环境之间的水分交换更快(Hartard et al ., 2009 ).在本研究中也是枝状地衣的失水速度要快于其他功能群附生地衣.枝状地衣的这种形态特征也使得它们能够从湿润的空气中快速吸收水分(Lange & Ziegler, 1986 ), 也就使得它们能够适应林外干燥的环境.相对于枝状地衣, 叶状地衣则是通过增大持水量和水分贮存能力来保持体内的水分含量从而延长生理活动的时间(Lange & Ziegler, 1986 ), 比如我们所研究的白绵腹衣, 其失水速度要明显低于其他附生地衣.对于蓝藻地衣而言, 由于只能利用液态水来进行光合作用(Lange & Ziegler, 1986 ), 所以它们通过凝胶状物质来增加持水量. ...
Studies on lichen-dominated systems XVIII. Morphological control of evaporation in lichens.
1
1976
... 作为变水植物, 地衣的含水量随着环境条件迅速变化, 而其所有的代谢活动, 特别是净光合速率和呼吸速率也随之发生变化(Green & Lange, 1994 ).所以地衣水分关系的研究格外受到关注(Galun, 1988 ).地衣不能主动地调节水分的吸收、储存和流失(Green et al ., 2011 ), 它们利用形态学特征来促进叶状体从不同来源(雨水、潮湿空气、露水)吸收和贮存水分(Gauslaa & Coxson, 2011 ).形态结构和解剖结构是影响地衣水分贮藏的重要因素(Larson & Kershaw, 1976 ; Larson, 1981 ; Sancho & Kappen, 1989 ).在本研究中, 附生地衣的最大含水量和功能特征相关性不大, 属于同一功能团的两种附生地衣间的最大含水量也可能相差很大, 而枝状地衣的最大含水量也可能比叶状地衣的大, 这就说明解剖结构可能在附生地衣持水量上发挥了更重要的作用.Gauslaa和Coxson (2011)认为共生藻层的厚度是影响地衣物种间持水量的重要因素, 而共生藻的类型是影响其厚度的主要因素.我们所研究的两种蓝藻地衣都有着较高的持水量, 尤其是猫耳衣, 其持水量明显高于其他附生地衣.而网肺衣的持水量也比同属的针芽肺衣高100%左右.蓝藻地衣之所以有较高的持水量是因为在它们的共生藻细胞外有一层较厚的凝胶状保护鞘(Honegger et al ., 1996 ; Gauslaa & Coxson, 2011 ), 能够帮助蓝藻地衣贮存更多的水分.水分贮存能力、形态结构、解剖结构以及颜色是影响地衣吸收和丢失水分的主要因素(Palmqvist, 2000 ).比如, 相比于叶状地衣, 枝状地衣和环境之间的水分交换更快(Hartard et al ., 2009 ).在本研究中也是枝状地衣的失水速度要快于其他功能群附生地衣.枝状地衣的这种形态特征也使得它们能够从湿润的空气中快速吸收水分(Lange & Ziegler, 1986 ), 也就使得它们能够适应林外干燥的环境.相对于枝状地衣, 叶状地衣则是通过增大持水量和水分贮存能力来保持体内的水分含量从而延长生理活动的时间(Lange & Ziegler, 1986 ), 比如我们所研究的白绵腹衣, 其失水速度要明显低于其他附生地衣.对于蓝藻地衣而言, 由于只能利用液态水来进行光合作用(Lange & Ziegler, 1986 ), 所以它们通过凝胶状物质来增加持水量. ...
a). Bole epiphytic lichens as potential indicators of environmental change in subtropical forest ecosystems in southwest China.
3
2013
... 气候变化、氮沉降、生物入侵和土地利用变化已经成为当前全球生物多样性的主要威胁(Sala et al ., 2000 ), 因此许多学者开始研究全球变化对生物多样性和物种分布的潜在影响.在这样的背景下, 理解限制物种分布的过程和机制就显得尤为重要(Travis et al ., 2006 ).附生地衣是森林生态系统的重要组成部分, 占据着林冠相当份量的生物量(Sipman & Harris, 1989 ; Gradstein, 1992 ), 其多样性和分布自20世纪70年代起就得到了众多学者的关注(Cornelissen & Steege, 1999 ; Holz & Gradstein, 2005 ; Hauck & Spribille, 2005 ).Aptroot和Sipman (1997)估计在热带雨林中每km2 有300种地衣, Aptroot (1997)报道在巴布亚新几内亚的热带山地雨林一株树上就有173种地衣.地衣的分布受众多因子的综合影响(Gustafasson & Eriksson, 1995 ; Goward & Arsenawlt, 2000 ; Hauck & Spribille, 2005 ), 除自身的繁殖能力和扩散能力以外, 光照、温度、湿度、干扰程度、群落类型以及宿主特征等均对附生地衣物种的组成和分布产生一定的影响(李苏等, 2007 ; Li et al ., 2011 , 2013a, 2013b ).地衣由于其形态结构的特殊性和变水植物的特性, 对环境变化非常敏感, 因此地衣多样性的减少经常被用来指示空气污染(Cislaghi & Nimis, 1997 ; Giordani, 2007 )、气候条件(Jovan & McCune, 2004 ; Geiser & Neitlich, 2007 ; Giordani & Incerti, 2008 )、森林结构和动态(Johansson, 2008 ; Nascimbene et al ., 2010 ).但是通过比较地衣多样性来比较不同区域间的环境条件常常遇到困难, 因为环境条件的不同经常形成高度差异的物种组成, 而如果我们利用地衣的功能特征来指示环境条件则可以很好地解决这一问题(Giordani et al ., 2012 ). ...
... 由于生活型和共生藻类型可能控制群落的结构(Ellis & Coppins, 2006 , 2007, 2010 ; Marini et al ., 2011 ), 结合Li等(2013a, 2013b)对本地区附生地衣分布的调查研究, 我们选择更能指示环境状况的蓝藻地衣、阔叶地衣、狭叶地衣、枝状地衣4种地衣功能群作为研究对象, 4种附生地衣功能群的划分依据见表1 .每种附生地衣功能群选择2种常见种(或代表种)作为研究对象.其中蓝藻地衣为网肺衣(Lobaria retigera )和猫耳衣(Leptogium menziesii ), 阔叶地衣为针芽肺衣(Lobaria isidiophora )和皮革肾岛衣(Nephromopsis pallescens ), 狭叶地衣为白绵腹衣(Anzia leucobatoides )和云南袋衣(Hypogymnia yunnanensis ), 枝状地衣为裂髓树花(Ramalina conduplicans )和多花松萝(Usnea florida ).其中网肺衣和猫耳衣多生长于潮湿的生境中, 针芽肺衣、皮革肾岛衣、白棉腹衣和云南袋衣多生长于阳光充足的生境中, 裂髓树花和多花松萝多分布于干燥、高光的生境中(Li et al ., 2013a ).我们在每个物种分布较多的生境中采集各研究物种材料, 带回哀牢山生态站, 清洗干净后, 进行相关实验. ...
... 地衣的变水特征使得它们的光响应特征比较复杂.地衣水分含量和净光合速率(P n )的关系能够改变它们的P n 对光照的响应(Lange & Matthes, 1981 ).低的水分含量使得地衣的P n 获得低数值的光补偿点(LCP )及光饱和点(LSP ), 而高水分含量对P n 的抑制作用也能够降低地衣的LSP 值, 但对LCP 的影响不大(Lange et al ., 1996 , 2004 ; Green et al ., 1997 ; 陈克等, 2014 ).Green等(1997)对新西兰热带雨林9种地衣光适应的研究指出, 光响应曲线参数和所测定地衣的生境相关.他们的研究发现, 林内地衣的光饱和点要低于300 μmol·m-2 ·s-1 , 而在开阔生境中地衣的光饱和点要大于549 μmol·m-2 ·s-1 .在本研究中, 8种附生地衣的LSP 都比较高(333.9-616.0 μmol·m-2 ·s-1 , 表4 ), 并且即使光强高于1000 μmol·m-2 ·s-1 也没有明显的光抑制现象, 表现出明显的“阳生植物”特性, 这在一定程度上解释了为什么在哀牢山地区附生地衣在次生林比在原生林中有着更高的丰富度(李苏等, 2007 ; Li et al ., 2013a , 2013b ).就各功能群附生地衣的LSP 而言, 枝状地衣的LSP 要明显高于其他功能群附生地衣, 这也很好地解释了它们在林冠上层和林外空旷地有较高多样性的分布格局(Li et al ., 2015 ).Green等(1997)和陈克等(2014)的研究中都指出相比于绿藻地衣, 蓝藻地衣有着较低的LSP .在我们的研究中虽然网肺衣也有着较高的LSP, 但是猫耳衣的LSP 还是低于其他功能群附生地衣.LCP 是衡量植物利用弱光能力的重要指标.在我们的研究中, 8种附生植物的LCP 都比较高(20.1-43.1 μmol·m-2 · ...
b). Epiphytic lichens in subtropical forest ecosystems in southwest China: Species diversity and implications for conservation.
1
2013
... 地衣的变水特征使得它们的光响应特征比较复杂.地衣水分含量和净光合速率(P n )的关系能够改变它们的P n 对光照的响应(Lange & Matthes, 1981 ).低的水分含量使得地衣的P n 获得低数值的光补偿点(LCP )及光饱和点(LSP ), 而高水分含量对P n 的抑制作用也能够降低地衣的LSP 值, 但对LCP 的影响不大(Lange et al ., 1996 , 2004 ; Green et al ., 1997 ; 陈克等, 2014 ).Green等(1997)对新西兰热带雨林9种地衣光适应的研究指出, 光响应曲线参数和所测定地衣的生境相关.他们的研究发现, 林内地衣的光饱和点要低于300 μmol·m-2 ·s-1 , 而在开阔生境中地衣的光饱和点要大于549 μmol·m-2 ·s-1 .在本研究中, 8种附生地衣的LSP 都比较高(333.9-616.0 μmol·m-2 ·s-1 , 表4 ), 并且即使光强高于1000 μmol·m-2 ·s-1 也没有明显的光抑制现象, 表现出明显的“阳生植物”特性, 这在一定程度上解释了为什么在哀牢山地区附生地衣在次生林比在原生林中有着更高的丰富度(李苏等, 2007 ; Li et al ., 2013a , 2013b ).就各功能群附生地衣的LSP 而言, 枝状地衣的LSP 要明显高于其他功能群附生地衣, 这也很好地解释了它们在林冠上层和林外空旷地有较高多样性的分布格局(Li et al ., 2015 ).Green等(1997)和陈克等(2014)的研究中都指出相比于绿藻地衣, 蓝藻地衣有着较低的LSP .在我们的研究中虽然网肺衣也有着较高的LSP, 但是猫耳衣的LSP 还是低于其他功能群附生地衣.LCP 是衡量植物利用弱光能力的重要指标.在我们的研究中, 8种附生植物的LCP 都比较高(20.1-43.1 μmol·m-2 · ...
亚热带森林生态系统4种附生蓝藻地衣的分布对生境变化的响应
2
2015
... 地衣的变水特征使得它们的光响应特征比较复杂.地衣水分含量和净光合速率(P n )的关系能够改变它们的P n 对光照的响应(Lange & Matthes, 1981 ).低的水分含量使得地衣的P n 获得低数值的光补偿点(LCP )及光饱和点(LSP ), 而高水分含量对P n 的抑制作用也能够降低地衣的LSP 值, 但对LCP 的影响不大(Lange et al ., 1996 , 2004 ; Green et al ., 1997 ; 陈克等, 2014 ).Green等(1997)对新西兰热带雨林9种地衣光适应的研究指出, 光响应曲线参数和所测定地衣的生境相关.他们的研究发现, 林内地衣的光饱和点要低于300 μmol·m-2 ·s-1 , 而在开阔生境中地衣的光饱和点要大于549 μmol·m-2 ·s-1 .在本研究中, 8种附生地衣的LSP 都比较高(333.9-616.0 μmol·m-2 ·s-1 , 表4 ), 并且即使光强高于1000 μmol·m-2 ·s-1 也没有明显的光抑制现象, 表现出明显的“阳生植物”特性, 这在一定程度上解释了为什么在哀牢山地区附生地衣在次生林比在原生林中有着更高的丰富度(李苏等, 2007 ; Li et al ., 2013a , 2013b ).就各功能群附生地衣的LSP 而言, 枝状地衣的LSP 要明显高于其他功能群附生地衣, 这也很好地解释了它们在林冠上层和林外空旷地有较高多样性的分布格局(Li et al ., 2015 ).Green等(1997)和陈克等(2014)的研究中都指出相比于绿藻地衣, 蓝藻地衣有着较低的LSP .在我们的研究中虽然网肺衣也有着较高的LSP, 但是猫耳衣的LSP 还是低于其他功能群附生地衣.LCP 是衡量植物利用弱光能力的重要指标.在我们的研究中, 8种附生植物的LCP 都比较高(20.1-43.1 μmol·m-2 · ...
... s-1 , 表4 ), 除了蓝藻地衣外, 其他功能群附生地衣的LCP 都要高于Green等(1997)在新西兰热带雨林所研究的林内林缘地衣LCP (4.3-29 μmol·m-2 ·s-1 ), 而低于林外地衣LCP (≥50 μmol·m-2 ·s-1 ).在本研究中, 蓝藻地衣的LCP 显著低于其他功能群的附生地衣, 这和在哀牢山地区观察到的蓝藻地衣能分布在较阴暗生境相符(Li et al ., 2015 ).总之, 蓝藻地衣能够利用的光谱比较宽(较低的LCP 和较高的LSP ), 所以蓝藻地衣既能分布在较强太阳辐射的生境中, 又对弱光有一定的适应能力(Rikkien, 2009 ), 而绿藻地衣大多只能分布在太阳辐射较高的生境中.地衣的光合能力和其氮素含量密切相关(Lange et al ., 2004 ).在Lange等(2004)对巴拿马热带雨林6种地衣碳获取的研究中, 具有高浓度氮含量的牛皮叶属地衣的P n (4.9-5.8 μmol·m-2 ·s-1 )要比肺衣属的(2.0-2.9 μmol·m-2 ·s-1 )高很多.我们所研究的8种附生地衣的P n 都比较小(0.92-2.84 μmol·m-2 ·s-1 , 表4 ), 两种肺衣属(针芽肺衣和网肺衣)地衣的P n (分别为1.77 和2.84 μmol·m-2 ·s-1 )和Lange等(2004)测得的同属地衣的P n 相差不大, 并且属于蓝藻地衣的网肺衣的 P n 要比属于绿藻地衣的针芽肺衣的高, 这可能和网肺衣具有固氮能力有关.光合能力的差异和功能群的相关性不大, 可能更多地和地衣氮浓度含量相关. ...
Biomass, diversity and composition of epiphytic macrolichens in primary and secondary forests in the subtropical Ailao Mountains, southwest China.
2011
云南哀牢山原生林及次生林群落附生地衣物种多样性与分布
1
2007
... 气候变化、氮沉降、生物入侵和土地利用变化已经成为当前全球生物多样性的主要威胁(Sala et al ., 2000 ), 因此许多学者开始研究全球变化对生物多样性和物种分布的潜在影响.在这样的背景下, 理解限制物种分布的过程和机制就显得尤为重要(Travis et al ., 2006 ).附生地衣是森林生态系统的重要组成部分, 占据着林冠相当份量的生物量(Sipman & Harris, 1989 ; Gradstein, 1992 ), 其多样性和分布自20世纪70年代起就得到了众多学者的关注(Cornelissen & Steege, 1999 ; Holz & Gradstein, 2005 ; Hauck & Spribille, 2005 ).Aptroot和Sipman (1997)估计在热带雨林中每km2 有300种地衣, Aptroot (1997)报道在巴布亚新几内亚的热带山地雨林一株树上就有173种地衣.地衣的分布受众多因子的综合影响(Gustafasson & Eriksson, 1995 ; Goward & Arsenawlt, 2000 ; Hauck & Spribille, 2005 ), 除自身的繁殖能力和扩散能力以外, 光照、温度、湿度、干扰程度、群落类型以及宿主特征等均对附生地衣物种的组成和分布产生一定的影响(李苏等, 2007 ; Li et al ., 2011 , 2013a, 2013b ).地衣由于其形态结构的特殊性和变水植物的特性, 对环境变化非常敏感, 因此地衣多样性的减少经常被用来指示空气污染(Cislaghi & Nimis, 1997 ; Giordani, 2007 )、气候条件(Jovan & McCune, 2004 ; Geiser & Neitlich, 2007 ; Giordani & Incerti, 2008 )、森林结构和动态(Johansson, 2008 ; Nascimbene et al ., 2010 ).但是通过比较地衣多样性来比较不同区域间的环境条件常常遇到困难, 因为环境条件的不同经常形成高度差异的物种组成, 而如果我们利用地衣的功能特征来指示环境条件则可以很好地解决这一问题(Giordani et al ., 2012 ). ...
Nutrient fluxes in bulk precipitation, throughfall and stemflow in montane moist evergreen broad-leaved forest on Ailao Mountain in Yunnan, SW China.
2002
哀牢山北段地区气候特征及变化趋势
3
2009
... 气候变化、氮沉降、生物入侵和土地利用变化已经成为当前全球生物多样性的主要威胁(Sala et al ., 2000 ), 因此许多学者开始研究全球变化对生物多样性和物种分布的潜在影响.在这样的背景下, 理解限制物种分布的过程和机制就显得尤为重要(Travis et al ., 2006 ).附生地衣是森林生态系统的重要组成部分, 占据着林冠相当份量的生物量(Sipman & Harris, 1989 ; Gradstein, 1992 ), 其多样性和分布自20世纪70年代起就得到了众多学者的关注(Cornelissen & Steege, 1999 ; Holz & Gradstein, 2005 ; Hauck & Spribille, 2005 ).Aptroot和Sipman (1997)估计在热带雨林中每km2 有300种地衣, Aptroot (1997)报道在巴布亚新几内亚的热带山地雨林一株树上就有173种地衣.地衣的分布受众多因子的综合影响(Gustafasson & Eriksson, 1995 ; Goward & Arsenawlt, 2000 ; Hauck & Spribille, 2005 ), 除自身的繁殖能力和扩散能力以外, 光照、温度、湿度、干扰程度、群落类型以及宿主特征等均对附生地衣物种的组成和分布产生一定的影响(李苏等, 2007 ; Li et al ., 2011 , 2013a, 2013b ).地衣由于其形态结构的特殊性和变水植物的特性, 对环境变化非常敏感, 因此地衣多样性的减少经常被用来指示空气污染(Cislaghi & Nimis, 1997 ; Giordani, 2007 )、气候条件(Jovan & McCune, 2004 ; Geiser & Neitlich, 2007 ; Giordani & Incerti, 2008 )、森林结构和动态(Johansson, 2008 ; Nascimbene et al ., 2010 ).但是通过比较地衣多样性来比较不同区域间的环境条件常常遇到困难, 因为环境条件的不同经常形成高度差异的物种组成, 而如果我们利用地衣的功能特征来指示环境条件则可以很好地解决这一问题(Giordani et al ., 2012 ). ...
... 哀牢山位于云贵高原西南部、横断山区南段, 属于云岭山脉向西南分支的余脉.本研究区位于哀牢山北段的徐家坝地区(23.35°-24.44° N, 100.54°- 101.30° E), 海拔2000-2750 m, 该区降雨充沛, 云雾缭绕, 常年温凉潮湿, 保存着我国亚热带地区目前面积(5000多hm2 )最大、以云南特有植物种为优势种的亚热带原始山地湿性常绿阔叶林(Liu et al ., 2002 ; 徐海清和刘文耀, 2005 ), 附生植物非常丰富, 是国家级自然保护区.根据哀牢山生态站多年气象观测结果显示, 年降水量1947 mm, 年蒸发量为1192 mm, 年平均相对湿度85%, 年平均气温11.3 ℃ (李苏等, 2007 ).该地区受到西南季风的影响, 有着明显的干湿季之分, 其中5-10月为雨季, 其降水量占全年降水量的85%以上(刘洋等, 2009 ). ...
... 地衣的变水特征使得它们的光响应特征比较复杂.地衣水分含量和净光合速率(P n )的关系能够改变它们的P n 对光照的响应(Lange & Matthes, 1981 ).低的水分含量使得地衣的P n 获得低数值的光补偿点(LCP )及光饱和点(LSP ), 而高水分含量对P n 的抑制作用也能够降低地衣的LSP 值, 但对LCP 的影响不大(Lange et al ., 1996 , 2004 ; Green et al ., 1997 ; 陈克等, 2014 ).Green等(1997)对新西兰热带雨林9种地衣光适应的研究指出, 光响应曲线参数和所测定地衣的生境相关.他们的研究发现, 林内地衣的光饱和点要低于300 μmol·m-2 ·s-1 , 而在开阔生境中地衣的光饱和点要大于549 μmol·m-2 ·s-1 .在本研究中, 8种附生地衣的LSP 都比较高(333.9-616.0 μmol·m-2 ·s-1 , 表4 ), 并且即使光强高于1000 μmol·m-2 ·s-1 也没有明显的光抑制现象, 表现出明显的“阳生植物”特性, 这在一定程度上解释了为什么在哀牢山地区附生地衣在次生林比在原生林中有着更高的丰富度(李苏等, 2007 ; Li et al ., 2013a , 2013b ).就各功能群附生地衣的LSP 而言, 枝状地衣的LSP 要明显高于其他功能群附生地衣, 这也很好地解释了它们在林冠上层和林外空旷地有较高多样性的分布格局(Li et al ., 2015 ).Green等(1997)和陈克等(2014)的研究中都指出相比于绿藻地衣, 蓝藻地衣有着较低的LSP .在我们的研究中虽然网肺衣也有着较高的LSP, 但是猫耳衣的LSP 还是低于其他功能群附生地衣.LCP 是衡量植物利用弱光能力的重要指标.在我们的研究中, 8种附生植物的LCP 都比较高(20.1-43.1 μmol·m-2 · ...
哀牢山常绿阔叶林地的气候特征研究
1
1993
... 哀牢山位于云贵高原西南部、横断山区南段, 属于云岭山脉向西南分支的余脉.本研究区位于哀牢山北段的徐家坝地区(23.35°-24.44° N, 100.54°- 101.30° E), 海拔2000-2750 m, 该区降雨充沛, 云雾缭绕, 常年温凉潮湿, 保存着我国亚热带地区目前面积(5000多hm2 )最大、以云南特有植物种为优势种的亚热带原始山地湿性常绿阔叶林(Liu et al ., 2002 ; 徐海清和刘文耀, 2005 ), 附生植物非常丰富, 是国家级自然保护区.根据哀牢山生态站多年气象观测结果显示, 年降水量1947 mm, 年蒸发量为1192 mm, 年平均相对湿度85%, 年平均气温11.3 ℃ (李苏等, 2007 ).该地区受到西南季风的影响, 有着明显的干湿季之分, 其中5-10月为雨季, 其降水量占全年降水量的85%以上(刘洋等, 2009 ). ...
云南哀牢山徐家坝地区附生(植)物的组成、生物量及其与生态因子的关系
2009
Large-scale patterns of epiphytic lichen species richness: Photobiont-dependent response to climate and forest structure.
1
2011
... 哀牢山位于云贵高原西南部、横断山区南段, 属于云岭山脉向西南分支的余脉.本研究区位于哀牢山北段的徐家坝地区(23.35°-24.44° N, 100.54°- 101.30° E), 海拔2000-2750 m, 该区降雨充沛, 云雾缭绕, 常年温凉潮湿, 保存着我国亚热带地区目前面积(5000多hm2 )最大、以云南特有植物种为优势种的亚热带原始山地湿性常绿阔叶林(Liu et al ., 2002 ; 徐海清和刘文耀, 2005 ), 附生植物非常丰富, 是国家级自然保护区.根据哀牢山生态站多年气象观测结果显示, 年降水量1947 mm, 年蒸发量为1192 mm, 年平均相对湿度85%, 年平均气温11.3 ℃ (李苏等, 2007 ).该地区受到西南季风的影响, 有着明显的干湿季之分, 其中5-10月为雨季, 其降水量占全年降水量的85%以上(刘洋等, 2009 ). ...
Gradients in epiphyte biomass in three Pseudotsuga-Tsuga forests of different ages in western Oregon and Washington.
1993
Vertical distribution and ecology of epiphytic bryophytes and lichens in a lowland rain forest in French Guyana. PhD dissertation, University of Utrecht, Utrecht, the
3
1990
... 根据国外的相关研究, 地衣按照共生藻、生活型和繁殖策略的不同可以划分为蓝藻地衣(cyano-区性lichens, CYL)、壳状地衣(crustose lichens, CRL)、枝状地衣(fruticose lichens, FRL)、狭叶地衣(narrowly lobed foliose lichens, NFL)、阔叶地衣(broadly lobed foliose lichens, BFL)、裂芽地衣(isidiate lichens, ISL)、粉芽地衣(sorediate lichens, SOL)和有性地衣(sexual lichens, SEL) 8个功能群(McCune, 1993 ; Nimis & Martellos, 2008 ).调查研究显示地衣呈现出与它们功能特征相关的分布格局(Ellis & Coppins, 2006 ; Lakatos et al ., 2006 ; Zedda et al ., 2011 ).比如在热带山地雨林分布着更多的枝状和叶状地衣, 但在低地雨林却是壳状地衣占据优势, 枝状地衣和叶状地衣很稀少(Montfoot & Ek, 1990 ; Sipman & Tan, 1990 ; Lakatos et al ., 2004 ).Hauck等(2007)在蒙古的针叶林发现, 绿藻地衣在群落中处于优势地位, 蓝藻地衣只在有着较高可利用水源的地方有分布.在最近的研究中, 地衣的生活型、繁殖策略、共生藻等功能特征被广泛用来指示地衣群落对环境条件的适应(DÍaz & Cabido, 2001 ; Cornelissen et al ., 2007 ; Glime, 2007 ), 因为地衣的功能特征可以不依赖物种多样性和组成而直接和环境因子建立关联(Ricotta & Bacaro, 2010 ; Webb et al ., 2010 ).Lange等(1994)发现在蓝藻地衣较多的生境中经常有高的积水.Giordani等(2012)基于意大利森林生态系统网络研究了地衣功能特征和环境条件的关系, 结果表明: 壳状和叶状地衣的分布和降水量密切相关; 枝状地衣的分布受海拔的影响较大, 同时和森林的密闭度负相关; 狭叶和阔叶地衣都受到酸性沉积物的负面影响.Giordani等(2014)调查发现阔叶地衣只有在高潜在径流的条件下才能在高辐射的生境中分布, 同时他们还观察到在枝状地衣的生境中有高的潜在径流. ...
... 本研究在2015年7-9月期间在哀牢山生态站开展, 此时期正处于哀牢山雨季, 林内外湿度都能达到90%以上, 林内与林外湿度、温度的差异也不大(刘玉洪, 1993 ), 林内、外湿度差异对本实验的影响很小, 高湿环境也能防止地衣过快失水.地衣光合作用的测定在10:00-14:00进行, 此时间段的气温变化小于2 ℃ (刘玉洪, 1993 ), 所以温度变化对地衣光合作用的影响也很小. ...
... ), 林内、外湿度差异对本实验的影响很小, 高湿环境也能防止地衣过快失水.地衣光合作用的测定在10:00-14:00进行, 此时间段的气温变化小于2 ℃ (刘玉洪, 1993 ), 所以温度变化对地衣光合作用的影响也很小. ...
Testing Lobaria pulmonaria (L.) Hoffm as an indicator of lichen conservation importance of Italian forests.
2010
2
... 根据国外的相关研究, 地衣按照共生藻、生活型和繁殖策略的不同可以划分为蓝藻地衣(cyano-区性lichens, CYL)、壳状地衣(crustose lichens, CRL)、枝状地衣(fruticose lichens, FRL)、狭叶地衣(narrowly lobed foliose lichens, NFL)、阔叶地衣(broadly lobed foliose lichens, BFL)、裂芽地衣(isidiate lichens, ISL)、粉芽地衣(sorediate lichens, SOL)和有性地衣(sexual lichens, SEL) 8个功能群(McCune, 1993 ; Nimis & Martellos, 2008 ).调查研究显示地衣呈现出与它们功能特征相关的分布格局(Ellis & Coppins, 2006 ; Lakatos et al ., 2006 ; Zedda et al ., 2011 ).比如在热带山地雨林分布着更多的枝状和叶状地衣, 但在低地雨林却是壳状地衣占据优势, 枝状地衣和叶状地衣很稀少(Montfoot & Ek, 1990 ; Sipman & Tan, 1990 ; Lakatos et al ., 2004 ).Hauck等(2007)在蒙古的针叶林发现, 绿藻地衣在群落中处于优势地位, 蓝藻地衣只在有着较高可利用水源的地方有分布.在最近的研究中, 地衣的生活型、繁殖策略、共生藻等功能特征被广泛用来指示地衣群落对环境条件的适应(DÍaz & Cabido, 2001 ; Cornelissen et al ., 2007 ; Glime, 2007 ), 因为地衣的功能特征可以不依赖物种多样性和组成而直接和环境因子建立关联(Ricotta & Bacaro, 2010 ; Webb et al ., 2010 ).Lange等(1994)发现在蓝藻地衣较多的生境中经常有高的积水.Giordani等(2012)基于意大利森林生态系统网络研究了地衣功能特征和环境条件的关系, 结果表明: 壳状和叶状地衣的分布和降水量密切相关; 枝状地衣的分布受海拔的影响较大, 同时和森林的密闭度负相关; 狭叶和阔叶地衣都受到酸性沉积物的负面影响.Giordani等(2014)调查发现阔叶地衣只有在高潜在径流的条件下才能在高辐射的生境中分布, 同时他们还观察到在枝状地衣的生境中有高的潜在径流. ...
... 在哀牢山国家级自然保护区分布着我国目前保存最完整、面积最大的山地湿性常绿阔叶林(Young & Herwitz, 1995 ; 朱华和闫丽春, 2009 ), 林内分布着丰富的附生地衣.Li等(2013b)在本地区记录到了217种附生地衣.由于人为的干扰, 在本地区原生林的周围形成了栎类萌生林、滇山杨(Populus bonatii )林、旱冬瓜(Alnus nepalensis )林等次生林(游承侠, 1983 ), 从而形成了不同的生境.在不同森林类型及它们之间的过渡带, 附生植物的多样性和丰富度是不一样的(马文章, 2009 ; 姚元林等, 2012 ).Li等(2013a)对本地区附生地衣的调查研究发现, 不同森林类型内附生地衣功能团的盖度和物种丰富度均明显不同, 在滇山杨林中阔叶地衣、蓝藻地衣及枝状地衣等的物种丰富度最高, 而在旱冬瓜林中, 壳状地衣和狭叶地衣的丰富度最高.李苏等(2015)对本地区4种附生蓝藻地衣分布的研究发现, 4种蓝藻地衣中有3种蓝藻地衣的盖度和频度最高值出现于次生林中, 并且4种蓝藻地衣表现出明显对厚皮香(Ternstroemia gymnanthera )、滇山杨和硬壳柯(Lithocarpus hancei )等树种的偏好性, 以及对小花山茶(Camellia forrestii )等的排斥性, 但目前还无法解释本地区附生地衣功能群的分布格局.Zots和Winder (1994)从碳平衡的角度解释了壳状地衣、枝状地衣和叶状地衣在热带地区海拔梯度上的分布格局.Lakatos等(2006)从形态学和生理学的角度解释了壳状绿藻地衣在法属圭亚那热带低地雨林林下存在的机制.所以, 地衣生理生态学研究是用来解释地衣功能群分布格局的有效途径之一.本研究拟在控制条件下分析比较不同附生地衣功能群的持水力和失水速度及光合作用对水分、光照的响应, 以期从内在机理上解释本地区附生地衣功能群的分布格局, 同时为利用附生地衣功能群指示环境变化提供理论依据. ...
Carbon economy in lichens.
1
2000
... 由于生活型和共生藻类型可能控制群落的结构(Ellis & Coppins, 2006 , 2007, 2010 ; Marini et al ., 2011 ), 结合Li等(2013a, 2013b)对本地区附生地衣分布的调查研究, 我们选择更能指示环境状况的蓝藻地衣、阔叶地衣、狭叶地衣、枝状地衣4种地衣功能群作为研究对象, 4种附生地衣功能群的划分依据见表1 .每种附生地衣功能群选择2种常见种(或代表种)作为研究对象.其中蓝藻地衣为网肺衣(Lobaria retigera )和猫耳衣(Leptogium menziesii ), 阔叶地衣为针芽肺衣(Lobaria isidiophora )和皮革肾岛衣(Nephromopsis pallescens ), 狭叶地衣为白绵腹衣(Anzia leucobatoides )和云南袋衣(Hypogymnia yunnanensis ), 枝状地衣为裂髓树花(Ramalina conduplicans )和多花松萝(Usnea florida ).其中网肺衣和猫耳衣多生长于潮湿的生境中, 针芽肺衣、皮革肾岛衣、白棉腹衣和云南袋衣多生长于阳光充足的生境中, 裂髓树花和多花松萝多分布于干燥、高光的生境中(Li et al ., 2013a ).我们在每个物种分布较多的生境中采集各研究物种材料, 带回哀牢山生态站, 清洗干净后, 进行相关实验. ...
Exploring phenol- typic plasticity in the lichen Ramalina capitata: Morphol- ogy, water relations and chlorophyll content in north- and south-facing populations.
3
1997
... 根据国外的相关研究, 地衣按照共生藻、生活型和繁殖策略的不同可以划分为蓝藻地衣(cyano-区性lichens, CYL)、壳状地衣(crustose lichens, CRL)、枝状地衣(fruticose lichens, FRL)、狭叶地衣(narrowly lobed foliose lichens, NFL)、阔叶地衣(broadly lobed foliose lichens, BFL)、裂芽地衣(isidiate lichens, ISL)、粉芽地衣(sorediate lichens, SOL)和有性地衣(sexual lichens, SEL) 8个功能群(McCune, 1993 ; Nimis & Martellos, 2008 ).调查研究显示地衣呈现出与它们功能特征相关的分布格局(Ellis & Coppins, 2006 ; Lakatos et al ., 2006 ; Zedda et al ., 2011 ).比如在热带山地雨林分布着更多的枝状和叶状地衣, 但在低地雨林却是壳状地衣占据优势, 枝状地衣和叶状地衣很稀少(Montfoot & Ek, 1990 ; Sipman & Tan, 1990 ; Lakatos et al ., 2004 ).Hauck等(2007)在蒙古的针叶林发现, 绿藻地衣在群落中处于优势地位, 蓝藻地衣只在有着较高可利用水源的地方有分布.在最近的研究中, 地衣的生活型、繁殖策略、共生藻等功能特征被广泛用来指示地衣群落对环境条件的适应(DÍaz & Cabido, 2001 ; Cornelissen et al ., 2007 ; Glime, 2007 ), 因为地衣的功能特征可以不依赖物种多样性和组成而直接和环境因子建立关联(Ricotta & Bacaro, 2010 ; Webb et al ., 2010 ).Lange等(1994)发现在蓝藻地衣较多的生境中经常有高的积水.Giordani等(2012)基于意大利森林生态系统网络研究了地衣功能特征和环境条件的关系, 结果表明: 壳状和叶状地衣的分布和降水量密切相关; 枝状地衣的分布受海拔的影响较大, 同时和森林的密闭度负相关; 狭叶和阔叶地衣都受到酸性沉积物的负面影响.Giordani等(2014)调查发现阔叶地衣只有在高潜在径流的条件下才能在高辐射的生境中分布, 同时他们还观察到在枝状地衣的生境中有高的潜在径流. ...
... Modified from McCune (1993) , Nimis & Martellos (2008). ...
... 引自McCune (1993) 、Nimis和Martellos (2008). ...
On plot to plot dissimilarity measures based on species functional traits.
1
2010
... 根据国外的相关研究, 地衣按照共生藻、生活型和繁殖策略的不同可以划分为蓝藻地衣(cyano-区性lichens, CYL)、壳状地衣(crustose lichens, CRL)、枝状地衣(fruticose lichens, FRL)、狭叶地衣(narrowly lobed foliose lichens, NFL)、阔叶地衣(broadly lobed foliose lichens, BFL)、裂芽地衣(isidiate lichens, ISL)、粉芽地衣(sorediate lichens, SOL)和有性地衣(sexual lichens, SEL) 8个功能群(McCune, 1993 ; Nimis & Martellos, 2008 ).调查研究显示地衣呈现出与它们功能特征相关的分布格局(Ellis & Coppins, 2006 ; Lakatos et al ., 2006 ; Zedda et al ., 2011 ).比如在热带山地雨林分布着更多的枝状和叶状地衣, 但在低地雨林却是壳状地衣占据优势, 枝状地衣和叶状地衣很稀少(Montfoot & Ek, 1990 ; Sipman & Tan, 1990 ; Lakatos et al ., 2004 ).Hauck等(2007)在蒙古的针叶林发现, 绿藻地衣在群落中处于优势地位, 蓝藻地衣只在有着较高可利用水源的地方有分布.在最近的研究中, 地衣的生活型、繁殖策略、共生藻等功能特征被广泛用来指示地衣群落对环境条件的适应(DÍaz & Cabido, 2001 ; Cornelissen et al ., 2007 ; Glime, 2007 ), 因为地衣的功能特征可以不依赖物种多样性和组成而直接和环境因子建立关联(Ricotta & Bacaro, 2010 ; Webb et al ., 2010 ).Lange等(1994)发现在蓝藻地衣较多的生境中经常有高的积水.Giordani等(2012)基于意大利森林生态系统网络研究了地衣功能特征和环境条件的关系, 结果表明: 壳状和叶状地衣的分布和降水量密切相关; 枝状地衣的分布受海拔的影响较大, 同时和森林的密闭度负相关; 狭叶和阔叶地衣都受到酸性沉积物的负面影响.Giordani等(2014)调查发现阔叶地衣只有在高潜在径流的条件下才能在高辐射的生境中分布, 同时他们还观察到在枝状地衣的生境中有高的潜在径流. ...
2
... 气候变化、氮沉降、生物入侵和土地利用变化已经成为当前全球生物多样性的主要威胁(Sala et al ., 2000 ), 因此许多学者开始研究全球变化对生物多样性和物种分布的潜在影响.在这样的背景下, 理解限制物种分布的过程和机制就显得尤为重要(Travis et al ., 2006 ).附生地衣是森林生态系统的重要组成部分, 占据着林冠相当份量的生物量(Sipman & Harris, 1989 ; Gradstein, 1992 ), 其多样性和分布自20世纪70年代起就得到了众多学者的关注(Cornelissen & Steege, 1999 ; Holz & Gradstein, 2005 ; Hauck & Spribille, 2005 ).Aptroot和Sipman (1997)估计在热带雨林中每km2 有300种地衣, Aptroot (1997)报道在巴布亚新几内亚的热带山地雨林一株树上就有173种地衣.地衣的分布受众多因子的综合影响(Gustafasson & Eriksson, 1995 ; Goward & Arsenawlt, 2000 ; Hauck & Spribille, 2005 ), 除自身的繁殖能力和扩散能力以外, 光照、温度、湿度、干扰程度、群落类型以及宿主特征等均对附生地衣物种的组成和分布产生一定的影响(李苏等, 2007 ; Li et al ., 2011 , 2013a, 2013b ).地衣由于其形态结构的特殊性和变水植物的特性, 对环境变化非常敏感, 因此地衣多样性的减少经常被用来指示空气污染(Cislaghi & Nimis, 1997 ; Giordani, 2007 )、气候条件(Jovan & McCune, 2004 ; Geiser & Neitlich, 2007 ; Giordani & Incerti, 2008 )、森林结构和动态(Johansson, 2008 ; Nascimbene et al ., 2010 ).但是通过比较地衣多样性来比较不同区域间的环境条件常常遇到困难, 因为环境条件的不同经常形成高度差异的物种组成, 而如果我们利用地衣的功能特征来指示环境条件则可以很好地解决这一问题(Giordani et al ., 2012 ). ...
... s-1 , 表4 ), 除了蓝藻地衣外, 其他功能群附生地衣的LCP 都要高于Green等(1997)在新西兰热带雨林所研究的林内林缘地衣LCP (4.3-29 μmol·m-2 ·s-1 ), 而低于林外地衣LCP (≥50 μmol·m-2 ·s-1 ).在本研究中, 蓝藻地衣的LCP 显著低于其他功能群的附生地衣, 这和在哀牢山地区观察到的蓝藻地衣能分布在较阴暗生境相符(Li et al ., 2015 ).总之, 蓝藻地衣能够利用的光谱比较宽(较低的LCP 和较高的LSP ), 所以蓝藻地衣既能分布在较强太阳辐射的生境中, 又对弱光有一定的适应能力(Rikkien, 2009 ), 而绿藻地衣大多只能分布在太阳辐射较高的生境中.地衣的光合能力和其氮素含量密切相关(Lange et al ., 2004 ).在Lange等(2004)对巴拿马热带雨林6种地衣碳获取的研究中, 具有高浓度氮含量的牛皮叶属地衣的P n (4.9-5.8 μmol·m-2 ·s-1 )要比肺衣属的(2.0-2.9 μmol·m-2 ·s-1 )高很多.我们所研究的8种附生地衣的P n 都比较小(0.92-2.84 μmol·m-2 ·s-1 , 表4 ), 两种肺衣属(针芽肺衣和网肺衣)地衣的P n (分别为1.77 和2.84 μmol·m-2 ·s-1 )和Lange等(2004)测得的同属地衣的P n 相差不大, 并且属于蓝藻地衣的网肺衣的 P n 要比属于绿藻地衣的针芽肺衣的高, 这可能和网肺衣具有固氮能力有关.光合能力的差异和功能群的相关性不大, 可能更多地和地衣氮浓度含量相关. ...
Global biodiversity scenarios for the year 2100.
1
2000
... 作为变水植物, 地衣的含水量随着环境条件迅速变化, 而其所有的代谢活动, 特别是净光合速率和呼吸速率也随之发生变化(Green & Lange, 1994 ).所以地衣水分关系的研究格外受到关注(Galun, 1988 ).地衣不能主动地调节水分的吸收、储存和流失(Green et al ., 2011 ), 它们利用形态学特征来促进叶状体从不同来源(雨水、潮湿空气、露水)吸收和贮存水分(Gauslaa & Coxson, 2011 ).形态结构和解剖结构是影响地衣水分贮藏的重要因素(Larson & Kershaw, 1976 ; Larson, 1981 ; Sancho & Kappen, 1989 ).在本研究中, 附生地衣的最大含水量和功能特征相关性不大, 属于同一功能团的两种附生地衣间的最大含水量也可能相差很大, 而枝状地衣的最大含水量也可能比叶状地衣的大, 这就说明解剖结构可能在附生地衣持水量上发挥了更重要的作用.Gauslaa和Coxson (2011)认为共生藻层的厚度是影响地衣物种间持水量的重要因素, 而共生藻的类型是影响其厚度的主要因素.我们所研究的两种蓝藻地衣都有着较高的持水量, 尤其是猫耳衣, 其持水量明显高于其他附生地衣.而网肺衣的持水量也比同属的针芽肺衣高100%左右.蓝藻地衣之所以有较高的持水量是因为在它们的共生藻细胞外有一层较厚的凝胶状保护鞘(Honegger et al ., 1996 ; Gauslaa & Coxson, 2011 ), 能够帮助蓝藻地衣贮存更多的水分.水分贮存能力、形态结构、解剖结构以及颜色是影响地衣吸收和丢失水分的主要因素(Palmqvist, 2000 ).比如, 相比于叶状地衣, 枝状地衣和环境之间的水分交换更快(Hartard et al ., 2009 ).在本研究中也是枝状地衣的失水速度要快于其他功能群附生地衣.枝状地衣的这种形态特征也使得它们能够从湿润的空气中快速吸收水分(Lange & Ziegler, 1986 ), 也就使得它们能够适应林外干燥的环境.相对于枝状地衣, 叶状地衣则是通过增大持水量和水分贮存能力来保持体内的水分含量从而延长生理活动的时间(Lange & Ziegler, 1986 ), 比如我们所研究的白绵腹衣, 其失水速度要明显低于其他附生地衣.对于蓝藻地衣而言, 由于只能利用液态水来进行光合作用(Lange & Ziegler, 1986 ), 所以它们通过凝胶状物质来增加持水量. ...
Photosynthesis and water rela- tions and the role of anatomy in Umbilicariaceae (lichens) from Central Spain.
1989
1
... 气候变化、氮沉降、生物入侵和土地利用变化已经成为当前全球生物多样性的主要威胁(Sala et al ., 2000 ), 因此许多学者开始研究全球变化对生物多样性和物种分布的潜在影响.在这样的背景下, 理解限制物种分布的过程和机制就显得尤为重要(Travis et al ., 2006 ).附生地衣是森林生态系统的重要组成部分, 占据着林冠相当份量的生物量(Sipman & Harris, 1989 ; Gradstein, 1992 ), 其多样性和分布自20世纪70年代起就得到了众多学者的关注(Cornelissen & Steege, 1999 ; Holz & Gradstein, 2005 ; Hauck & Spribille, 2005 ).Aptroot和Sipman (1997)估计在热带雨林中每km2 有300种地衣, Aptroot (1997)报道在巴布亚新几内亚的热带山地雨林一株树上就有173种地衣.地衣的分布受众多因子的综合影响(Gustafasson & Eriksson, 1995 ; Goward & Arsenawlt, 2000 ; Hauck & Spribille, 2005 ), 除自身的繁殖能力和扩散能力以外, 光照、温度、湿度、干扰程度、群落类型以及宿主特征等均对附生地衣物种的组成和分布产生一定的影响(李苏等, 2007 ; Li et al ., 2011 , 2013a, 2013b ).地衣由于其形态结构的特殊性和变水植物的特性, 对环境变化非常敏感, 因此地衣多样性的减少经常被用来指示空气污染(Cislaghi & Nimis, 1997 ; Giordani, 2007 )、气候条件(Jovan & McCune, 2004 ; Geiser & Neitlich, 2007 ; Giordani & Incerti, 2008 )、森林结构和动态(Johansson, 2008 ; Nascimbene et al ., 2010 ).但是通过比较地衣多样性来比较不同区域间的环境条件常常遇到困难, 因为环境条件的不同经常形成高度差异的物种组成, 而如果我们利用地衣的功能特征来指示环境条件则可以很好地解决这一问题(Giordani et al ., 2012 ). ...
A field impression of the lichen and bryophyte zonation on Mount Kinabalu.
1990
Water relations and gas exchange of fan bryophytes and their adaptations to microhabitats in an Asian subtropical montane cloud forest.
1
2015
... 气候变化、氮沉降、生物入侵和土地利用变化已经成为当前全球生物多样性的主要威胁(Sala et al ., 2000 ), 因此许多学者开始研究全球变化对生物多样性和物种分布的潜在影响.在这样的背景下, 理解限制物种分布的过程和机制就显得尤为重要(Travis et al ., 2006 ).附生地衣是森林生态系统的重要组成部分, 占据着林冠相当份量的生物量(Sipman & Harris, 1989 ; Gradstein, 1992 ), 其多样性和分布自20世纪70年代起就得到了众多学者的关注(Cornelissen & Steege, 1999 ; Holz & Gradstein, 2005 ; Hauck & Spribille, 2005 ).Aptroot和Sipman (1997)估计在热带雨林中每km2 有300种地衣, Aptroot (1997)报道在巴布亚新几内亚的热带山地雨林一株树上就有173种地衣.地衣的分布受众多因子的综合影响(Gustafasson & Eriksson, 1995 ; Goward & Arsenawlt, 2000 ; Hauck & Spribille, 2005 ), 除自身的繁殖能力和扩散能力以外, 光照、温度、湿度、干扰程度、群落类型以及宿主特征等均对附生地衣物种的组成和分布产生一定的影响(李苏等, 2007 ; Li et al ., 2011 , 2013a, 2013b ).地衣由于其形态结构的特殊性和变水植物的特性, 对环境变化非常敏感, 因此地衣多样性的减少经常被用来指示空气污染(Cislaghi & Nimis, 1997 ; Giordani, 2007 )、气候条件(Jovan & McCune, 2004 ; Geiser & Neitlich, 2007 ; Giordani & Incerti, 2008 )、森林结构和动态(Johansson, 2008 ; Nascimbene et al ., 2010 ).但是通过比较地衣多样性来比较不同区域间的环境条件常常遇到困难, 因为环境条件的不同经常形成高度差异的物种组成, 而如果我们利用地衣的功能特征来指示环境条件则可以很好地解决这一问题(Giordani et al ., 2012 ). ...
The distribution of positive and negative species interactions across environmental gradients on a dual-lattice model.
1
2006
... 作为变水植物, 地衣的含水量随着环境条件迅速变化, 而其所有的代谢活动, 特别是净光合速率和呼吸速率也随之发生变化(Green & Lange, 1994 ).所以地衣水分关系的研究格外受到关注(Galun, 1988 ).地衣不能主动地调节水分的吸收、储存和流失(Green et al ., 2011 ), 它们利用形态学特征来促进叶状体从不同来源(雨水、潮湿空气、露水)吸收和贮存水分(Gauslaa & Coxson, 2011 ).形态结构和解剖结构是影响地衣水分贮藏的重要因素(Larson & Kershaw, 1976 ; Larson, 1981 ; Sancho & Kappen, 1989 ).在本研究中, 附生地衣的最大含水量和功能特征相关性不大, 属于同一功能团的两种附生地衣间的最大含水量也可能相差很大, 而枝状地衣的最大含水量也可能比叶状地衣的大, 这就说明解剖结构可能在附生地衣持水量上发挥了更重要的作用.Gauslaa和Coxson (2011)认为共生藻层的厚度是影响地衣物种间持水量的重要因素, 而共生藻的类型是影响其厚度的主要因素.我们所研究的两种蓝藻地衣都有着较高的持水量, 尤其是猫耳衣, 其持水量明显高于其他附生地衣.而网肺衣的持水量也比同属的针芽肺衣高100%左右.蓝藻地衣之所以有较高的持水量是因为在它们的共生藻细胞外有一层较厚的凝胶状保护鞘(Honegger et al ., 1996 ; Gauslaa & Coxson, 2011 ), 能够帮助蓝藻地衣贮存更多的水分.水分贮存能力、形态结构、解剖结构以及颜色是影响地衣吸收和丢失水分的主要因素(Palmqvist, 2000 ).比如, 相比于叶状地衣, 枝状地衣和环境之间的水分交换更快(Hartard et al ., 2009 ).在本研究中也是枝状地衣的失水速度要快于其他功能群附生地衣.枝状地衣的这种形态特征也使得它们能够从湿润的空气中快速吸收水分(Lange & Ziegler, 1986 ), 也就使得它们能够适应林外干燥的环境.相对于枝状地衣, 叶状地衣则是通过增大持水量和水分贮存能力来保持体内的水分含量从而延长生理活动的时间(Lange & Ziegler, 1986 ), 比如我们所研究的白绵腹衣, 其失水速度要明显低于其他附生地衣.对于蓝藻地衣而言, 由于只能利用液态水来进行光合作用(Lange & Ziegler, 1986 ), 所以它们通过凝胶状物质来增加持水量. ...
A structured and dynamic framework to advance traits-based theory and prediction in ecology.
2010
云南哀牢山山地湿性常绿阔叶林附生植物的多样性与分布
1
2005
... 根据国外的相关研究, 地衣按照共生藻、生活型和繁殖策略的不同可以划分为蓝藻地衣(cyano-区性lichens, CYL)、壳状地衣(crustose lichens, CRL)、枝状地衣(fruticose lichens, FRL)、狭叶地衣(narrowly lobed foliose lichens, NFL)、阔叶地衣(broadly lobed foliose lichens, BFL)、裂芽地衣(isidiate lichens, ISL)、粉芽地衣(sorediate lichens, SOL)和有性地衣(sexual lichens, SEL) 8个功能群(McCune, 1993 ; Nimis & Martellos, 2008 ).调查研究显示地衣呈现出与它们功能特征相关的分布格局(Ellis & Coppins, 2006 ; Lakatos et al ., 2006 ; Zedda et al ., 2011 ).比如在热带山地雨林分布着更多的枝状和叶状地衣, 但在低地雨林却是壳状地衣占据优势, 枝状地衣和叶状地衣很稀少(Montfoot & Ek, 1990 ; Sipman & Tan, 1990 ; Lakatos et al ., 2004 ).Hauck等(2007)在蒙古的针叶林发现, 绿藻地衣在群落中处于优势地位, 蓝藻地衣只在有着较高可利用水源的地方有分布.在最近的研究中, 地衣的生活型、繁殖策略、共生藻等功能特征被广泛用来指示地衣群落对环境条件的适应(DÍaz & Cabido, 2001 ; Cornelissen et al ., 2007 ; Glime, 2007 ), 因为地衣的功能特征可以不依赖物种多样性和组成而直接和环境因子建立关联(Ricotta & Bacaro, 2010 ; Webb et al ., 2010 ).Lange等(1994)发现在蓝藻地衣较多的生境中经常有高的积水.Giordani等(2012)基于意大利森林生态系统网络研究了地衣功能特征和环境条件的关系, 结果表明: 壳状和叶状地衣的分布和降水量密切相关; 枝状地衣的分布受海拔的影响较大, 同时和森林的密闭度负相关; 狭叶和阔叶地衣都受到酸性沉积物的负面影响.Giordani等(2014)调查发现阔叶地衣只有在高潜在径流的条件下才能在高辐射的生境中分布, 同时他们还观察到在枝状地衣的生境中有高的潜在径流. ...
云南哀牢山国家保护区几个过渡带树干附生苔藓的物种组成与多样性
2
2012
... 不同附生地衣单位干质量持水量的差异利用单因素方差分析和Tukey’s HSD多重配对进行分析, 并利用线性模型(GLM)来分析附生地衣物种、干质量和持水力(WHC)之间的关系(Gauslaa & Coxson, 2011 ; Esseen et al ., 2015 ).各附生地衣的失水速率利用指数递减回归模型(Song et al ., 2015 )进行拟合: ...
... 植物光合作用对光响应模型的研究进行得比较早, 已经研究出许多经典的模型.其中直角双曲线修正模型不但可以拟合植物在饱和光强之后光合速率不下降的光响应曲线, 而且还可以拟合植物在饱和光强之后光合速率随光强的增加而下降这一段的光响应曲线, 且估算的饱和光强和最大净光合速率与实测值最为接近(Ye, 2007 ; 叶子飘, 2010 ; Song et al ., 2015 ).所以我们利用直角双曲线修正模型来拟合各附生地衣光合作用和光照强度的关系: ...
A new model for relationship between light intensity and the rate of photosynthesis in Oryza sativa.
1
2007
... 气候变化、氮沉降、生物入侵和土地利用变化已经成为当前全球生物多样性的主要威胁(Sala et al ., 2000 ), 因此许多学者开始研究全球变化对生物多样性和物种分布的潜在影响.在这样的背景下, 理解限制物种分布的过程和机制就显得尤为重要(Travis et al ., 2006 ).附生地衣是森林生态系统的重要组成部分, 占据着林冠相当份量的生物量(Sipman & Harris, 1989 ; Gradstein, 1992 ), 其多样性和分布自20世纪70年代起就得到了众多学者的关注(Cornelissen & Steege, 1999 ; Holz & Gradstein, 2005 ; Hauck & Spribille, 2005 ).Aptroot和Sipman (1997)估计在热带雨林中每km2 有300种地衣, Aptroot (1997)报道在巴布亚新几内亚的热带山地雨林一株树上就有173种地衣.地衣的分布受众多因子的综合影响(Gustafasson & Eriksson, 1995 ; Goward & Arsenawlt, 2000 ; Hauck & Spribille, 2005 ), 除自身的繁殖能力和扩散能力以外, 光照、温度、湿度、干扰程度、群落类型以及宿主特征等均对附生地衣物种的组成和分布产生一定的影响(李苏等, 2007 ; Li et al ., 2011 , 2013a, 2013b ).地衣由于其形态结构的特殊性和变水植物的特性, 对环境变化非常敏感, 因此地衣多样性的减少经常被用来指示空气污染(Cislaghi & Nimis, 1997 ; Giordani, 2007 )、气候条件(Jovan & McCune, 2004 ; Geiser & Neitlich, 2007 ; Giordani & Incerti, 2008 )、森林结构和动态(Johansson, 2008 ; Nascimbene et al ., 2010 ).但是通过比较地衣多样性来比较不同区域间的环境条件常常遇到困难, 因为环境条件的不同经常形成高度差异的物种组成, 而如果我们利用地衣的功能特征来指示环境条件则可以很好地解决这一问题(Giordani et al ., 2012 ). ...
光合作用对光和CO2响应模型的研究进展
1
2010
... 根据国外的相关研究, 地衣按照共生藻、生活型和繁殖策略的不同可以划分为蓝藻地衣(cyano-区性lichens, CYL)、壳状地衣(crustose lichens, CRL)、枝状地衣(fruticose lichens, FRL)、狭叶地衣(narrowly lobed foliose lichens, NFL)、阔叶地衣(broadly lobed foliose lichens, BFL)、裂芽地衣(isidiate lichens, ISL)、粉芽地衣(sorediate lichens, SOL)和有性地衣(sexual lichens, SEL) 8个功能群(McCune, 1993 ; Nimis & Martellos, 2008 ).调查研究显示地衣呈现出与它们功能特征相关的分布格局(Ellis & Coppins, 2006 ; Lakatos et al ., 2006 ; Zedda et al ., 2011 ).比如在热带山地雨林分布着更多的枝状和叶状地衣, 但在低地雨林却是壳状地衣占据优势, 枝状地衣和叶状地衣很稀少(Montfoot & Ek, 1990 ; Sipman & Tan, 1990 ; Lakatos et al ., 2004 ).Hauck等(2007)在蒙古的针叶林发现, 绿藻地衣在群落中处于优势地位, 蓝藻地衣只在有着较高可利用水源的地方有分布.在最近的研究中, 地衣的生活型、繁殖策略、共生藻等功能特征被广泛用来指示地衣群落对环境条件的适应(DÍaz & Cabido, 2001 ; Cornelissen et al ., 2007 ; Glime, 2007 ), 因为地衣的功能特征可以不依赖物种多样性和组成而直接和环境因子建立关联(Ricotta & Bacaro, 2010 ; Webb et al ., 2010 ).Lange等(1994)发现在蓝藻地衣较多的生境中经常有高的积水.Giordani等(2012)基于意大利森林生态系统网络研究了地衣功能特征和环境条件的关系, 结果表明: 壳状和叶状地衣的分布和降水量密切相关; 枝状地衣的分布受海拔的影响较大, 同时和森林的密闭度负相关; 狭叶和阔叶地衣都受到酸性沉积物的负面影响.Giordani等(2014)调查发现阔叶地衣只有在高潜在径流的条件下才能在高辐射的生境中分布, 同时他们还观察到在枝状地衣的生境中有高的潜在径流. ...
2
... 在哀牢山国家级自然保护区分布着我国目前保存最完整、面积最大的山地湿性常绿阔叶林(Young & Herwitz, 1995 ; 朱华和闫丽春, 2009 ), 林内分布着丰富的附生地衣.Li等(2013b)在本地区记录到了217种附生地衣.由于人为的干扰, 在本地区原生林的周围形成了栎类萌生林、滇山杨(Populus bonatii )林、旱冬瓜(Alnus nepalensis )林等次生林(游承侠, 1983 ), 从而形成了不同的生境.在不同森林类型及它们之间的过渡带, 附生植物的多样性和丰富度是不一样的(马文章, 2009 ; 姚元林等, 2012 ).Li等(2013a)对本地区附生地衣的调查研究发现, 不同森林类型内附生地衣功能团的盖度和物种丰富度均明显不同, 在滇山杨林中阔叶地衣、蓝藻地衣及枝状地衣等的物种丰富度最高, 而在旱冬瓜林中, 壳状地衣和狭叶地衣的丰富度最高.李苏等(2015)对本地区4种附生蓝藻地衣分布的研究发现, 4种蓝藻地衣中有3种蓝藻地衣的盖度和频度最高值出现于次生林中, 并且4种蓝藻地衣表现出明显对厚皮香(Ternstroemia gymnanthera )、滇山杨和硬壳柯(Lithocarpus hancei )等树种的偏好性, 以及对小花山茶(Camellia forrestii )等的排斥性, 但目前还无法解释本地区附生地衣功能群的分布格局.Zots和Winder (1994)从碳平衡的角度解释了壳状地衣、枝状地衣和叶状地衣在热带地区海拔梯度上的分布格局.Lakatos等(2006)从形态学和生理学的角度解释了壳状绿藻地衣在法属圭亚那热带低地雨林林下存在的机制.所以, 地衣生理生态学研究是用来解释地衣功能群分布格局的有效途径之一.本研究拟在控制条件下分析比较不同附生地衣功能群的持水力和失水速度及光合作用对水分、光照的响应, 以期从内在机理上解释本地区附生地衣功能群的分布格局, 同时为利用附生地衣功能群指示环境变化提供理论依据. ...
... 哀牢山位于云贵高原西南部、横断山区南段, 属于云岭山脉向西南分支的余脉.本研究区位于哀牢山北段的徐家坝地区(23.35°-24.44° N, 100.54°- 101.30° E), 海拔2000-2750 m, 该区降雨充沛, 云雾缭绕, 常年温凉潮湿, 保存着我国亚热带地区目前面积(5000多hm2 )最大、以云南特有植物种为优势种的亚热带原始山地湿性常绿阔叶林(Liu et al ., 2002 ; 徐海清和刘文耀, 2005 ), 附生植物非常丰富, 是国家级自然保护区.根据哀牢山生态站多年气象观测结果显示, 年降水量1947 mm, 年蒸发量为1192 mm, 年平均相对湿度85%, 年平均气温11.3 ℃ (李苏等, 2007 ).该地区受到西南季风的影响, 有着明显的干湿季之分, 其中5-10月为雨季, 其降水量占全年降水量的85%以上(刘洋等, 2009 ). ...
Floristic diversity and co- occurrences in a subtropical broad-leaved forest and two contrasting regrowth stands in central-west Yunnan Province, China.
1995
2
... 根据国外的相关研究, 地衣按照共生藻、生活型和繁殖策略的不同可以划分为蓝藻地衣(cyano-区性lichens, CYL)、壳状地衣(crustose lichens, CRL)、枝状地衣(fruticose lichens, FRL)、狭叶地衣(narrowly lobed foliose lichens, NFL)、阔叶地衣(broadly lobed foliose lichens, BFL)、裂芽地衣(isidiate lichens, ISL)、粉芽地衣(sorediate lichens, SOL)和有性地衣(sexual lichens, SEL) 8个功能群(McCune, 1993 ; Nimis & Martellos, 2008 ).调查研究显示地衣呈现出与它们功能特征相关的分布格局(Ellis & Coppins, 2006 ; Lakatos et al ., 2006 ; Zedda et al ., 2011 ).比如在热带山地雨林分布着更多的枝状和叶状地衣, 但在低地雨林却是壳状地衣占据优势, 枝状地衣和叶状地衣很稀少(Montfoot & Ek, 1990 ; Sipman & Tan, 1990 ; Lakatos et al ., 2004 ).Hauck等(2007)在蒙古的针叶林发现, 绿藻地衣在群落中处于优势地位, 蓝藻地衣只在有着较高可利用水源的地方有分布.在最近的研究中, 地衣的生活型、繁殖策略、共生藻等功能特征被广泛用来指示地衣群落对环境条件的适应(DÍaz & Cabido, 2001 ; Cornelissen et al ., 2007 ; Glime, 2007 ), 因为地衣的功能特征可以不依赖物种多样性和组成而直接和环境因子建立关联(Ricotta & Bacaro, 2010 ; Webb et al ., 2010 ).Lange等(1994)发现在蓝藻地衣较多的生境中经常有高的积水.Giordani等(2012)基于意大利森林生态系统网络研究了地衣功能特征和环境条件的关系, 结果表明: 壳状和叶状地衣的分布和降水量密切相关; 枝状地衣的分布受海拔的影响较大, 同时和森林的密闭度负相关; 狭叶和阔叶地衣都受到酸性沉积物的负面影响.Giordani等(2014)调查发现阔叶地衣只有在高潜在径流的条件下才能在高辐射的生境中分布, 同时他们还观察到在枝状地衣的生境中有高的潜在径流. ...
... 在哀牢山国家级自然保护区分布着我国目前保存最完整、面积最大的山地湿性常绿阔叶林(Young & Herwitz, 1995 ; 朱华和闫丽春, 2009 ), 林内分布着丰富的附生地衣.Li等(2013b)在本地区记录到了217种附生地衣.由于人为的干扰, 在本地区原生林的周围形成了栎类萌生林、滇山杨(Populus bonatii )林、旱冬瓜(Alnus nepalensis )林等次生林(游承侠, 1983 ), 从而形成了不同的生境.在不同森林类型及它们之间的过渡带, 附生植物的多样性和丰富度是不一样的(马文章, 2009 ; 姚元林等, 2012 ).Li等(2013a)对本地区附生地衣的调查研究发现, 不同森林类型内附生地衣功能团的盖度和物种丰富度均明显不同, 在滇山杨林中阔叶地衣、蓝藻地衣及枝状地衣等的物种丰富度最高, 而在旱冬瓜林中, 壳状地衣和狭叶地衣的丰富度最高.李苏等(2015)对本地区4种附生蓝藻地衣分布的研究发现, 4种蓝藻地衣中有3种蓝藻地衣的盖度和频度最高值出现于次生林中, 并且4种蓝藻地衣表现出明显对厚皮香(Ternstroemia gymnanthera )、滇山杨和硬壳柯(Lithocarpus hancei )等树种的偏好性, 以及对小花山茶(Camellia forrestii )等的排斥性, 但目前还无法解释本地区附生地衣功能群的分布格局.Zots和Winder (1994)从碳平衡的角度解释了壳状地衣、枝状地衣和叶状地衣在热带地区海拔梯度上的分布格局.Lakatos等(2006)从形态学和生理学的角度解释了壳状绿藻地衣在法属圭亚那热带低地雨林林下存在的机制.所以, 地衣生理生态学研究是用来解释地衣功能群分布格局的有效途径之一.本研究拟在控制条件下分析比较不同附生地衣功能群的持水力和失水速度及光合作用对水分、光照的响应, 以期从内在机理上解释本地区附生地衣功能群的分布格局, 同时为利用附生地衣功能群指示环境变化提供理论依据. ...
2
... 在哀牢山国家级自然保护区分布着我国目前保存最完整、面积最大的山地湿性常绿阔叶林(Young & Herwitz, 1995 ; 朱华和闫丽春, 2009 ), 林内分布着丰富的附生地衣.Li等(2013b)在本地区记录到了217种附生地衣.由于人为的干扰, 在本地区原生林的周围形成了栎类萌生林、滇山杨(Populus bonatii )林、旱冬瓜(Alnus nepalensis )林等次生林(游承侠, 1983 ), 从而形成了不同的生境.在不同森林类型及它们之间的过渡带, 附生植物的多样性和丰富度是不一样的(马文章, 2009 ; 姚元林等, 2012 ).Li等(2013a)对本地区附生地衣的调查研究发现, 不同森林类型内附生地衣功能团的盖度和物种丰富度均明显不同, 在滇山杨林中阔叶地衣、蓝藻地衣及枝状地衣等的物种丰富度最高, 而在旱冬瓜林中, 壳状地衣和狭叶地衣的丰富度最高.李苏等(2015)对本地区4种附生蓝藻地衣分布的研究发现, 4种蓝藻地衣中有3种蓝藻地衣的盖度和频度最高值出现于次生林中, 并且4种蓝藻地衣表现出明显对厚皮香(Ternstroemia gymnanthera )、滇山杨和硬壳柯(Lithocarpus hancei )等树种的偏好性, 以及对小花山茶(Camellia forrestii )等的排斥性, 但目前还无法解释本地区附生地衣功能群的分布格局.Zots和Winder (1994)从碳平衡的角度解释了壳状地衣、枝状地衣和叶状地衣在热带地区海拔梯度上的分布格局.Lakatos等(2006)从形态学和生理学的角度解释了壳状绿藻地衣在法属圭亚那热带低地雨林林下存在的机制.所以, 地衣生理生态学研究是用来解释地衣功能群分布格局的有效途径之一.本研究拟在控制条件下分析比较不同附生地衣功能群的持水力和失水速度及光合作用对水分、光照的响应, 以期从内在机理上解释本地区附生地衣功能群的分布格局, 同时为利用附生地衣功能群指示环境变化提供理论依据. ...
... 植物光合作用对光响应模型的研究进行得比较早, 已经研究出许多经典的模型.其中直角双曲线修正模型不但可以拟合植物在饱和光强之后光合速率不下降的光响应曲线, 而且还可以拟合植物在饱和光强之后光合速率随光强的增加而下降这一段的光响应曲线, 且估算的饱和光强和最大净光合速率与实测值最为接近(Ye, 2007 ; 叶子飘, 2010 ; Song et al ., 2015 ).所以我们利用直角双曲线修正模型来拟合各附生地衣光合作用和光照强度的关系: ...
In situ studies of water relations and CO2 exchange of the tropical macrolichen, Sticta tomentosa.
1998
Are tropical lowlands a marginal habitat for macrolichens? Evidence from a field study with Parmotrema endosulphureum in Panama.
2
2003
... 各附生地衣光合作用和相对含水量之间的关系利用三次多项式回归模型进行拟合(Zotzet al ., 2003 ): ...
... 植物光合作用对光响应模型的研究进行得比较早, 已经研究出许多经典的模型.其中直角双曲线修正模型不但可以拟合植物在饱和光强之后光合速率不下降的光响应曲线, 而且还可以拟合植物在饱和光强之后光合速率随光强的增加而下降这一段的光响应曲线, 且估算的饱和光强和最大净光合速率与实测值最为接近(Ye, 2007 ; 叶子飘, 2010 ; Song et al ., 2015 ).所以我们利用直角双曲线修正模型来拟合各附生地衣光合作用和光照强度的关系: ...
Photosynthesis and carbon gain of the lichen, Leptogium azureum, in lowland tropical forest.
1994
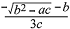
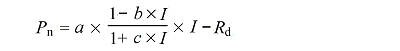


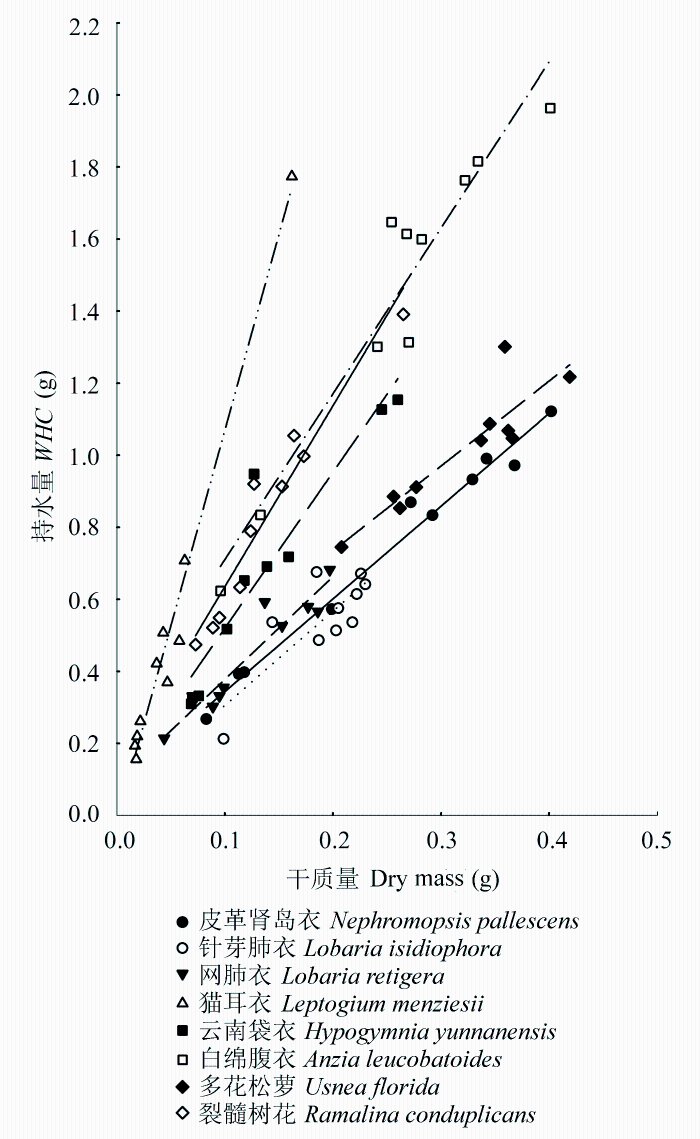
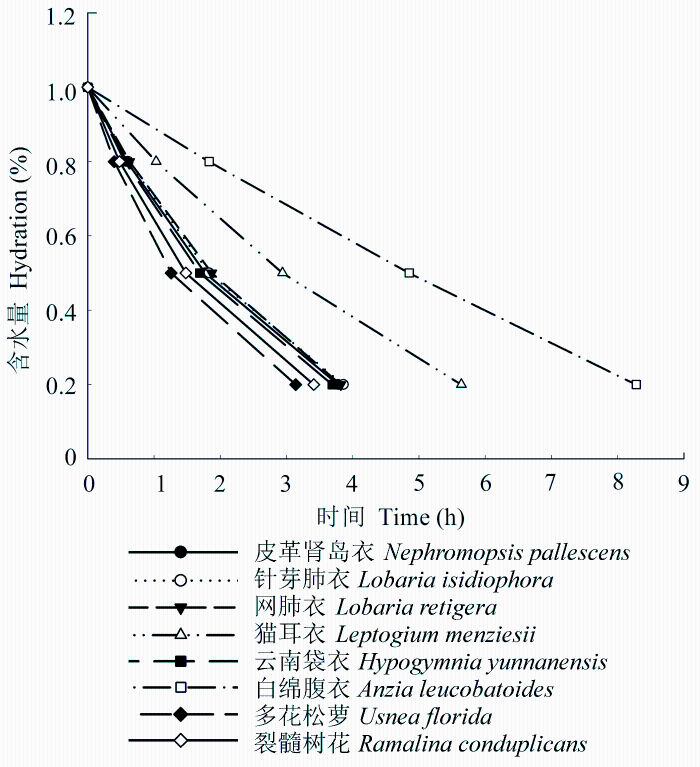
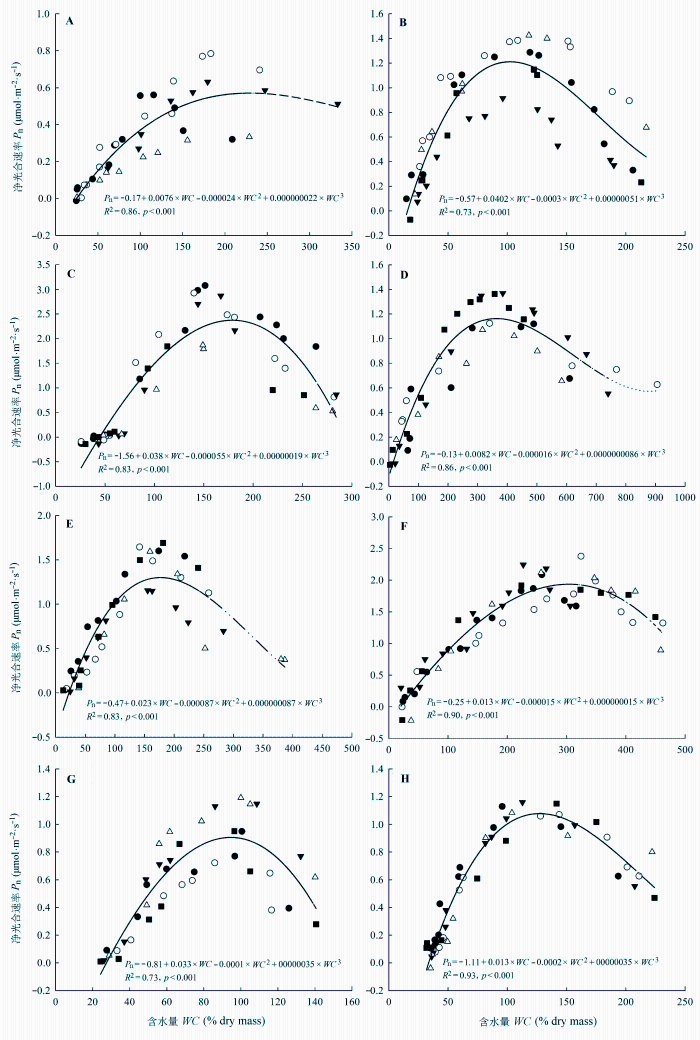
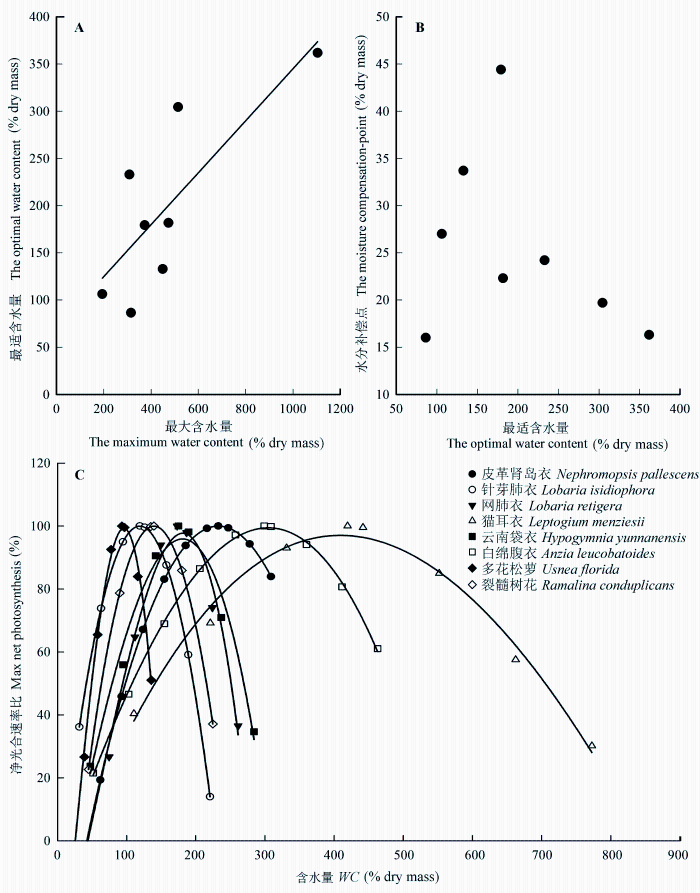
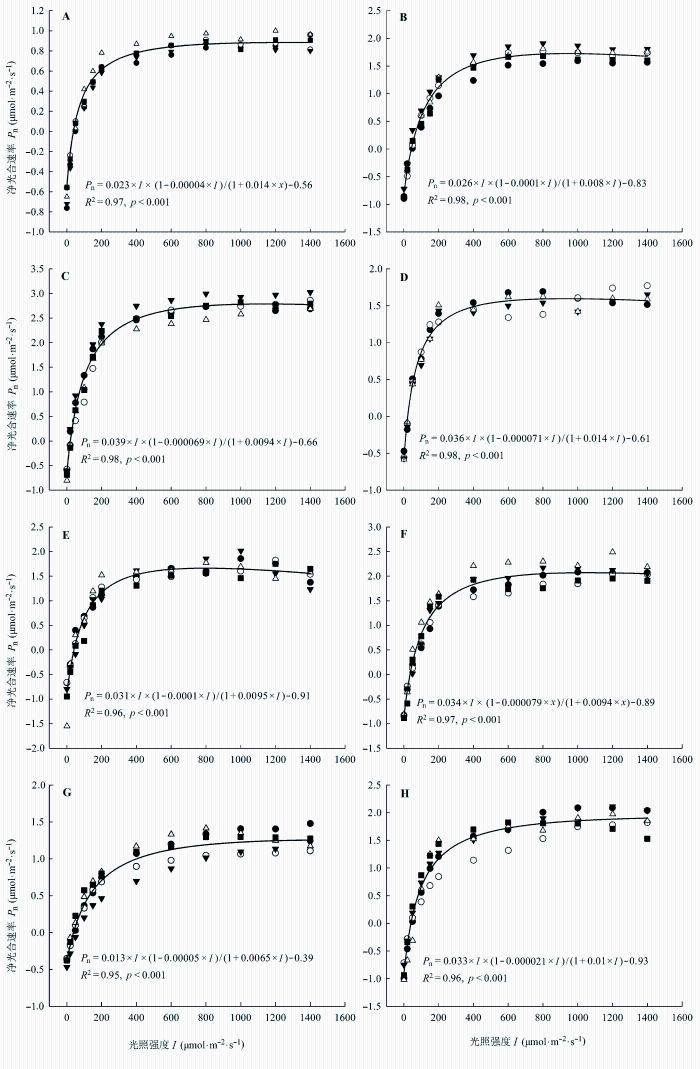
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}