1College of Animal Science and Technology, Southwest University, Chongqing 400715, China2College of Horticulture and Landscape Architecture, Southwest University, Chongqing 400715, China
Leaf mass per area (LMA) is a composite structural parameter as well as a basic leaf functional trait in the leaf economics spectrum (LES). It is not only closely related to many physiological responses of plants, but also can measure the investment of dry mass per unit of light-intercepting leaf area. LMA is considered an important indicator of plant ecological strategies and has been studied widely in plant ecology, agronomy, forestry, and plant physiology. This paper elucidates the structural analysis and computational methods of LMA at the organizational scales of whole leaf, tissues and cells, examines the influence of LMA on photosynthesis, and discusses the inherent differences in LMA and the responses of LMA to environmental stresses (temperature, water and light), aiming for clarifying research frameworks and methods in studies of LMA and providing guidance on future research.
Keywords:leaf mass per area (LMA)
;
leaf density
;
leaf thickness
;
photosynthesis
;
mesophyll conductance to CO2 (gm)
;
environmental stress
LIUMing-Xiu, LIANGGuo-Lu. Research progress on leaf mass per area. Chinese Journal of Plant Ecology, 2016, 40(8): 847-860 https://doi.org/10.17521/cjpe.2015.0428
自然界中的每一种植物都有自己独特的叶片形态和结构, 这种特异性并不是偶然的, 而是自然界选择进化的结果。每一种叶片的形态和组织结构都有与之相协调的生理过程紧密联系。因此, 从叶片形态和结构入手揭示植物的生态策略和生理过程是一种行之有效的研究手段。在植物的叶片解剖研究过程中, 大量研究资料显示比叶质量(LMA)是一个非常重要的反应植物功能的结构参数。LMA指叶片干质量与面积之间的比率(g·m-2), 是叶片众多解剖结构特征的综合体, 例如叶肉组织厚度、细胞壁厚度、细胞形状、细胞内含物等(Muir et al., 2014)。在大量的叶片形态解剖结构参数中, LMA是最容易定量测定的参数之一, 它能定量测定植物在单位叶面积的投入成本, 因而被认为是反映植物生态策略的重要指标(Westoby et al., 2002; Wright et al., 2004)。LMA的高低反映了植物对资源的利用效率以及对环境的应对策略。总的来讲, 低LMA的植物光合效率高, 单位叶面积的投入成本低, 显示出较快的资源流动速度及回报效率; 而高LMA的植物光合效率低, 单位叶面积的投入成本高, 资源的流动速度及回报效率低(Niinemets et al., 2007; Hikosaka & Shigeno, 2009)。
Fig. 1 Conceptual diagram of the anatomical characteristics at three integration levels (leaf, tissue, and cell) (marked with different gray) to show variation in leaf mass per area (LMA). Cited from Villar et al. (2013). LD, leaf density; LVA, leaf volume per area.
图2 不同植物的叶肉细胞导度(gm)与其比叶质量(LMA)之间的关系(非胁迫条件下)。向上的实心三角形代表高海拔植物, 向下的实心三角形代表老叶, 实心的正方形代表阴生叶, 其他的叶用圆圈表示。引自Flexas et al. (2008)。
Fig. 2 The relationship between mesophyll conductance (gm) and leaf mass per area (LMA) in different species, in the absence of stress. Data from high-altitude plants are indicated by filled upward triangles, old leaves by filled downward triangles and shade leaves by filled squares. Other leaves are indicated by open circles. Cited from Flexas et al. (2008).
草本植物与木本植物相比, 叶片薄而柔软, LMA低, 有较高的gm和光合速率(Flexas et al., 2008; Scafaro et al., 2011; Tomás et al., 2013; Muir et al., 2014)。Flexas等(2008)研究表明: LMA低的植物, 其gm较高且变化幅度较大, 也就是不同种类或功能群的低LMA植物的LMA与其gm之间没有显著的相关性(Flexas et al., 2008)。但是, 近年来也有不同的研究结果, 通过对番茄野生近缘种(Solanum sect. Lycopersicon, sect. Lycopersicoides) (Muir et al., 2014)、水稻野生近缘种(Oryza spp.) (Scafaro et al., 2011)、向日葵(Helianthus annuus)、辣椒(Capsicum annuum)、菜豆(Phaseolus vulgaris)、菠菜(Spinacea oleracea)、罗勒(Ocimum basilicum) (Tomás et al., 2013)的研究表明草本植物的LMA与gm也呈负相关关系, 但是导致gm下降的原因却各有不同。Muir等(2014)通过对8种番茄的研究表明, LMA与gm的关系主要受细胞内含物和叶片厚度的调控。Scafaro等(2011)研究表明, 在3种水稻中, 叶肉细胞壁增厚是gm下降, 并导致光合速率下降的重要原因。Tomás等(2013)对5种不同科属的草本植物的研究指出: 草本植物的细胞壁厚度对gm没有显著的影响, 细胞质和基质对gm影响约占60%, 是影响叶肉细胞导度的重要原因。由此来看, 导致草本植物gm下降的原因应该与植物的种的特异性有关, 具体原因还需要进一步研究。总之, 根据目前的资料报道, 无论是木本植物还是草本植物, LMA与gm之间的负相关关系是导致高LMA的植物光合效率下降的主要原因。
其次, 高LMA植物叶片中氮浓度低可能也是植物光合能力(Amass)降低的原因。植物叶片中大部分的氮参与光合作用的运转, LMA高的植物叶片中的关键营养元素的浓度都比较低, 例如氮、磷(Wright et al., 2004)。目前研究表明, 植物叶片中氮的浓度低会导致光合作用的氮利用效率以及分配到核酮糖-1,5-二磷酸羧化酶/加氧酶(Rubisco)和类囊体的氮显著下降, 从而引起光合能力的下降(Hikosaka, 2004; Hassiotou et al., 2010)。Niinemets等(2004)研究表明冬青栎的LMA随叶龄的增长而增加, 而氮含量随LMA增加而下降, 氮含量的降低导致Rubisco的最大羧化活性以及光合作用的电子传递能力下降。Xu等(2012)的研究表明柳叶桉(Eucalyptus saligna)的叶氮含量随LMA的增加而降低, 单位质量的最大光合效率随氮含量降低而降低。但并非所有情况下氮含量都随LMA增加而下降, Han (2011)发现LMA随赤松(Pinus densiflora)树高的增加而增加的过程中, Nmass和Narea均表现出增加的趋势, 这可能是植物为了减少因高度引起的光合效率的下降而采取的补偿机制。
在低温环境中, 植物的LMA增加是适应环境的表现。Ball等(2002)的研究表明, 拥有较高的LVA的植物可以通过减缓冰冻速率来减少冻害的发生和减轻冻害胁迫。此外, 细胞层数和数量增多导致单位叶面积的蛋白质含量以及次生化合物(例如: 脯氨酸)增加, 这些物质的增加有助于提高植物的耐寒能力(Usadel et al., 2008)。
生长在温度较高的环境中的植物通常拥有较低的LMA, 但LMA与温度并不呈线性相关(p < 0.01) (Atkin et al., 2006; Poorter et al., 2009)。温度升高引起LMA降低的直接原因是叶片变薄。目前大量实验表明, 生长在高温环境中的植物, 叶面积增大但叶片显著变薄, 这与叶片的表皮层、栅栏组织、海绵组织的细胞层数减少及细胞变小引起的叶组织厚度的降低有关(Hartikainen et al., 2009; Jin et al., 2011)。杨兵等(2010)研究发现长期模拟增温条件下, 岷江冷杉(Abies faxoniana)幼苗LMA显著下降。Xu等(2012)通过模拟温室效应来研究气温升高对柳叶桉形态解剖结构的影响, 结果表明随温度的升高柳叶桉叶片LMA呈下降趋势。Zhang等(2015)的研究表明长期热胁迫会导致云杉属(Picea)植物的LMA下降。
Acclimation of lamina mass per unit area, photosynthetic characteristics and dark respiration to growth light regimes in fourtropical rainforest species.
Acta Ecologica Sinica, 22, 901-910. (in Chinese with English abstract)[冯玉龙, 曹坤芳, 冯志立, 马玲 (2002).
Leaf responses to drought stress in Mediterranean accessions of Solanum lycopersicum: Anatomical adaptations in relation to gas exchange parameters. Plant,
Emissions of volatile organic compounds and leaf structural characteristics of European aspen (Populus tremula) grown under elevated ozone and temperature.
Tree Physiology, 29, 1163-1173.
[37]
HanbaYT, KogamiH, TerashimaI (2002).
The effect of growth irradiance on leaf anatomy and photosynthesis in Acer species differing in light demand. Plant,
Heigh related decreases in mesophyll conductance, leaf photosynthesis and compensating adjustments associated with leaf nitrogen concentrations in Pinus densiflora.
Leaf-age effects on seasonal variability in photosynthetic parameters and its relationships with leaf mass per area and leaf nitrogen concentration within a Pinus densiflora crown.
Comparison of leaf mass per area, photosynthetic capacity and chemical defense traits of four evergreen broad-leaved tree species under contrasting light conditions.
Chinese Journal of Applied and Environmental Biology, 16, 462-467. (in Chinese with English abstract)[柳凤娟, 向双, 阳小成, 孙书存 (2010). [
b). Submergence-induced morphological, anatomical, and biochemical responses in a terrestrial species affect gas diffusion resistance and photosynthetic performance.
Morphological and anatomical determinants of mesophyll conductance in wild relatives of tomato (Solanum sect. Lycopersicon, sect. Lycopersicoides; Solanaceae). Plant,
Cell & Environment, 37, 1415-1426.
[61]
MyersDA, VogelmannTC, BornmanJF (1994).
Epidermal focusing and effects on light utilization in Oxalis acetosella.
Physiologia Plantarum, 91, 651-656.
[62]
NiinemetsÜ (1999).
Components of leaf dry mass per area- thickness and density-alter leaf photosynthetic capacity in reverse directions in woody plants.
New Phytologist, 144, 35-47.
[63]
NiinemetsÜ, KullO, TenhunenJD (1998).
An analysis of light effects on foliar morphology, physiology, and light interception in temperate deciduous woody species of contrasting shade tolerance.
Tree Physiology, 18, 681-696.
[64]
NiinemetsÜ (2001).
Global-scale climatic controls of leaf dry mass per area, density, and thickness in trees and shrubs.
Within canopy variation in the rate of development of photosynthetic capacity is proportional to integrated quantum flux density in temperate deciduous trees. Plant,
Complex adjustments of photosynthetic potentials and internal diffusion conductance to current and previous light availabilities and leaf age in Mediterranean evergreen species Quercus ilex. Plant,
Leaf anatomical properties in relation to differences in mesophyll conductance to CO2 and photosynthesis in two related Mediterranean Abies species. Plant,
Regulation of photosynthesis and stomatal and mesophyll conductance under water stress and recovery in olive trees: Correlation with gene expression of carbonic anhydrase and aquaporins.
Variations in nitrogen use efficiency reflect the biochemical subtype while variations in water use efficiency reflect the evolutionary lineage of C4 grasses at inter-glacial CO2. Plant,
Developmental changes in mesophyll diffusion conductance and photosynthetic capacity under different light and water availabilities in Populus tremula: How structure constrains function. Plant,
TsujiiY, OnodaY, IzunoA, IsagiY, Kitayama, K (2016).
A quantitative analysis of phenotypic variations of Metrosideros polymorpha within and across populations along environmental gradients on Mauna Loa, Hawaii.
Multilevel genomic analysis of the response of transcripts, enzyme activities and metabolites in Arabidopsis rosettes to a progressive decrease of temperature in the nonfreezing range. Plant,
Differences in construction costs and chemical composition between deciduous and evergreen woody species are small as compared to differences between families. Plant,
Responses of leaf structure and photosynthetic properties to intra-canopy light gradients: A common garden test with four broadleaf deciduous angiosperm and seven evergreen conifer tree species.
Leaf structural characteristics are less important than leaf chemical properties in determining the response of leaf mass per area and photosynthesis of Eucalyptus saligna to industrial-age changes in [CO2] and temperature.
The impact of long-term water stress on relative growth rate and morphology of needles and shoots of Metasequoia glyptostro- boides seedlings: Research toward identifying mechanistic models.
Differential leaf expansion can enable hydraulic acclimation to sun and shade. Plant,
1
2012
... 植物的LMA随光照强度的增加而增加, 但LMA由总的光照度/天(DPI)决定, 而不是瞬时的高的光照强度决定(Chabot et al., 1979; Niinemets et al., 2004).LMA随DPI的增加而显著增加, 但这种响应呈非线性相关, 因为在低光条件下LMA随DPI的增加而显著增加, 但是当DPI超过20 mol·m-2·d-1时, 其LMA的增加会变缓慢(Poorter et al., 2009).LMA 随光照度增加而增加的过程中, LD和LVA均扮演重要角色.Coble和Cavaleri (2014)的研究表明, 光照增加导致LMA的增加主要与LVA的增加有关.但在LVA增加的过程中, 并不是所有的叶片组织的体积都增加, 一般情况下, 栅栏组织的体积随光强度的增加而显著增加, 而表皮组织的体积保持不变(Hanson, 1917; Onoda et al., 2008).而高光条件下, LD的增加与叶脉密度的显著增加有关(Carins Murphy et al., 2012); 此外, 在一定程度上还与栅栏组织细胞排列的紧密度的增加及总的非结构性碳水化合物(TNC)浓度的增加有关(Niinemets et al., 1998; Poorter et al., 2009). ...
Leaf structure and anatomy as related to leaf mass per area variation in seedlings of a wide range of woody plant species and types.
6
2000
... 20世纪90年代, 很多学者对不同植物的LMA做了大量研究, 并探讨了LVA和LD与LMA之间的关系.研究表明在同一可控环境条件下, 草本植物(Garnier & Laurent, 1994; van Arendonk & Poorter, 1994)和木本植物(Castro-Diez et al., 2000)的LMA主要受LD的影响, Poorter等(2009)对这些数据作了统计和分析, 表明LMA差异的80%由LD决定, 20%由LVA决定.然而在复杂的自然环境条件下, LD和LVA与LMA之间的关系是多样化的(Choong et al., 1992; Niinemets, 1999; Wilson et al., 1999).由此可见, 仅仅把LMA分解为LVA和LD并不能很好地解释导致植物种类之间LMA差异的原因. ...
... 目前植物解剖学的研究大多采用较薄的组织切片, 因此LMA的细胞水平的研究大多用横切切片中的细胞面积来代表细胞体积(Pyankov et al., 1999; Castro-Díez et al., 2000; Villar et al., 2013), 但采用这种方法很难准确反映细胞的真实的体积.随着技术的发展, 在近期的一些文献中也采用激光扫描共聚焦显微镜形成的3D图像计算叶肉细胞的体积(Xu et al., 2012).LMA的细胞水平的研究表明, 木本植物和草本植物的叶肉细胞增大是导致叶片增厚的主要原因, 同时细胞大小与LD呈负相关关系, 但细胞大小与LMA无明显的相关性(Pyankov et al., 1999; Castro-Díez et al., 2000).Villar等(2013)研究发现, 常绿和落叶的木本植物的叶片LD相似, 常绿木本植物叶肉细胞增大引起的叶片增厚是导致其LMA值普遍高于落叶木本植物的原因.可见, 只有从细胞水平分解叶片, 才能更好地分析引起LMA变化的原因. ...
... 从目前的研究结果来看, 木本植物比草本植物的LMA高, 在同一环境控制下, 两者之间的差异大约为35%; 在野外环境下, 两者之间的差异约83% (Poorter et al., 2009).LMA在叶片水平可分解为叶片密度和厚度.根据目前的研究文献报道, 密度和厚度都有可能是决定木本植物LMA高低的原因(Castro-Díez et al., 2000; Wright & Cannon, 2001; Villar et al., 2013).LMA高的木本植物, 其硬化组织占整个叶片的比例高, 细胞小而紧凑、叶片含水量和氮含量较低(Wright & Cannon, 2001).常绿木本植物普遍比落叶木本植物的LMA高(Castro-Díez et al., 2000; Villar & Merino, 2001; Wright et al., 2005; Li et al., 2013; Villar et al., 2013).Castro-Díez (2000)和Villar等(2013)认为常绿和落叶木本植物的叶片密度之间没有显著的差异, 常绿木本植物较高的LMA主要与叶片厚度有关, 同科属中的常绿木本植物的叶片通常比落叶木本植物的叶片厚, 而叶肉组织增厚是导致叶片变厚的主要原因.然而, 不同科属的常绿和落叶植物之间的LMA也存在交差和重叠, 这主要是由植物的系统发育决定的; 也就是不同科属之间植物的LMA是有差异的, 而在同一科属中, 常绿植物比落叶植物的LMA高(Poorter et al., 2009; Villar et al., 2013). ...
... 高(Castro-Díez et al., 2000; Villar & Merino, 2001; Wright et al., 2005; Li et al., 2013; Villar et al., 2013).Castro-Díez (2000)和Villar等(2013)认为常绿和落叶木本植物的叶片密度之间没有显著的差异, 常绿木本植物较高的LMA主要与叶片厚度有关, 同科属中的常绿木本植物的叶片通常比落叶木本植物的叶片厚, 而叶肉组织增厚是导致叶片变厚的主要原因.然而, 不同科属的常绿和落叶植物之间的LMA也存在交差和重叠, 这主要是由植物的系统发育决定的; 也就是不同科属之间植物的LMA是有差异的, 而在同一科属中, 常绿植物比落叶植物的LMA高(Poorter et al., 2009; Villar et al., 2013). ...
... 树木冠层内的LMA的差异也是森林生态系统研究的重要内容.LMA与叶片在冠层中的位置有关, 一般处于冠层顶部的叶片LMA值比冠层底部的高(Koch et al., 2004; Lichtenthaler et al., 2007; Poorter et al., 2009; Weerasinghe et al., 2014).据Koch等(2004)报道100 m高的红杉(Sequoia sempervirens)的冠层底部LMA为150 g·m-2, 而冠层顶部LMA值达800 g·m-2, 这是比较极端的例子.一般来说, 不管是木本植物还是草本植物, 其冠层LMA大约是底层LMA的两倍(Poorter et al., 2009).这种底层和冠层之间的差异在一定程度上是由物种的特异性决定的(Lichtenthaler et al., 2007).此外, LMA还随叶寿命的不同表现出较大的差异.叶寿命是植物在长期适应环境的过程中为获得最大光合生产所形成的适应策略(Reich et al., 1991; 张林和罗天祥, 2004).LMA反映了植物单位叶面积的构建成本, 通常与叶寿命呈正相关关系.LMA低的植物, 其光合效率高, 生长速度快, 环境胁迫应答策略表现为逃避, 因而用于防御投资的物质较少, 叶寿命较短, 如草本植物、落叶乔灌木; 而LMA高的植物, 光合效率慢, 生长速度慢, 环境胁迫应答的策略表现为抵御和耐受, 需要不断积累更多的物质构建保卫构造, 因此表现出较长的叶寿命, 如常绿木本植物(Reich et al., 1991; Wright et al., 2004; Wyka et al., 2012; 王瑞雪等, 2014).常绿木本植物的LMA通常随叶龄的增长而逐渐增加, LMA的增加主要与叶肉细胞的细胞壁变厚、木质素含量增加、叶片变厚、叶片的机械组织含量增加有关(Castro-Díez et al., 2000; Hanba et al., 2002; Wright et al., 2004).Niinemets等(2005, 2006)报道地中海3种阔叶常绿树种月桂(Laurus nobilis)、油橄榄(Olea europea)、冬青栎(Quercus ilex)的LMA随叶龄的增长而增加.而且叶龄对LMA的影响还与叶片在冠层中的位置有关, Han等(2008)报道赤松冠层底部LMA随叶龄的增加而增加的幅度为27%, 但叶龄对冠层顶部LMA无影响.此外, 在常绿木本植物中, 叶寿命不同的植物, 其LMA也有较大的差异, Wyka等(2012)报道松柏类比阔叶类植物的叶寿命长, 其LMA也显著高于阔叶类植物. ...
Light drives vertical gradients of leaf morphology in a sugar maple (Acer saccharum) forest.
2014
Light acclimation optimizes leaf functional traits despite height-related constraints in a canopy shading experiment.
3
2015
... 关于植物与光照的关系, 目前研究的比较多的是森林生态系统中光照不足对植物产生的影响.在森林生态系统中, 对光的需求的差异是决定植物生长和生存的主要因素(Hanba et al., 2002).大多研究表明植物在适应光环境的过程中主要通过调节叶片形态和结构适应光照的变化, 主要表现在LMA的增大或减小(Niinemets, 2007; Lusk et al., 2008; Coble & Cavaleri, 2015).目前的研究表明, 对于同一种植物, 遮阴导致叶片LMA、厚度、密度下降, 而单位干质量的叶面积增加, 增大了其叶片捕光面积, 利于在弱光环境下生存(冯玉龙等, 2002; Lusk et al., 2008; 柳凤娟等, 2010; 王博轶等, 2012; Coble & Cavaleri, 2015).在植物的冠层内部, 阴生叶比阳生叶的LMA低, 差异可达2-4倍, LMA降低主要与叶肉细胞层厚度减少, 叶片变薄, 叶片密度显著降低, 细胞内含物显著下降有关(Niinemets, 2007; Lusk et al., 2010; Chen et al., 2014).Meng等(2013, 2014)的研究发现, 遮阴不仅会导致叶片的LMA下降, 还会使叶片倾角变小, 以截获更多的光照.值得关注的是, 在很多胁迫条件下(比如干旱、低温), 叶片LMA的增加伴随细胞壁的增厚, 然而在遮阴条件下, 虽然LMA降低了, 但其细胞壁反而增厚, 这可能是植物在低光条件下为增强其叶片硬度和韧度, 避免食草动物的取食而采取的防御机制(Hanba et al., 2002; Coble & Cavaleri, 2015).此外, 很多学者在对热带常绿和落叶木本植物的研究中发现, 在同一低光环境中, 耐阴性强的植物与需光植物相比, 具有较低的光合能力和较高的LMA, LMA的增加在一定程度上与叶肉细胞组织孔隙度的下降、叶片表皮毛密度的增加有关(Hanba et al., 2002; Lusk et al., 2008).LMA的增加使叶片更加强韧, 可增强植物对食草动物和病原菌的防御能力, 提高其在弱光下的生存几率(Kitajima et al., 1994). ...
... ; Coble & Cavaleri, 2015).在植物的冠层内部, 阴生叶比阳生叶的LMA低, 差异可达2-4倍, LMA降低主要与叶肉细胞层厚度减少, 叶片变薄, 叶片密度显著降低, 细胞内含物显著下降有关(Niinemets, 2007; Lusk et al., 2010; Chen et al., 2014).Meng等(2013, 2014)的研究发现, 遮阴不仅会导致叶片的LMA下降, 还会使叶片倾角变小, 以截获更多的光照.值得关注的是, 在很多胁迫条件下(比如干旱、低温), 叶片LMA的增加伴随细胞壁的增厚, 然而在遮阴条件下, 虽然LMA降低了, 但其细胞壁反而增厚, 这可能是植物在低光条件下为增强其叶片硬度和韧度, 避免食草动物的取食而采取的防御机制(Hanba et al., 2002; Coble & Cavaleri, 2015).此外, 很多学者在对热带常绿和落叶木本植物的研究中发现, 在同一低光环境中, 耐阴性强的植物与需光植物相比, 具有较低的光合能力和较高的LMA, LMA的增加在一定程度上与叶肉细胞组织孔隙度的下降、叶片表皮毛密度的增加有关(Hanba et al., 2002; Lusk et al., 2008).LMA的增加使叶片更加强韧, 可增强植物对食草动物和病原菌的防御能力, 提高其在弱光下的生存几率(Kitajima et al., 1994). ...
... ; Coble & Cavaleri, 2015).此外, 很多学者在对热带常绿和落叶木本植物的研究中发现, 在同一低光环境中, 耐阴性强的植物与需光植物相比, 具有较低的光合能力和较高的LMA, LMA的增加在一定程度上与叶肉细胞组织孔隙度的下降、叶片表皮毛密度的增加有关(Hanba et al., 2002; Lusk et al., 2008).LMA的增加使叶片更加强韧, 可增强植物对食草动物和病原菌的防御能力, 提高其在弱光下的生存几率(Kitajima et al., 1994). ...
Leaf anatomical and photosynthetic acclimation to cool temperature and high light in two winter versus two summer annuals.
Influence of instantaneous and integrated light-flux density on leaf anatomy and photosynthesis.
1
1979
... 植物的LMA随光照强度的增加而增加, 但LMA由总的光照度/天(DPI)决定, 而不是瞬时的高的光照强度决定(Chabot et al., 1979; Niinemets et al., 2004).LMA随DPI的增加而显著增加, 但这种响应呈非线性相关, 因为在低光条件下LMA随DPI的增加而显著增加, 但是当DPI超过20 mol·m-2·d-1时, 其LMA的增加会变缓慢(Poorter et al., 2009).LMA 随光照度增加而增加的过程中, LD和LVA均扮演重要角色.Coble和Cavaleri (2014)的研究表明, 光照增加导致LMA的增加主要与LVA的增加有关.但在LVA增加的过程中, 并不是所有的叶片组织的体积都增加, 一般情况下, 栅栏组织的体积随光强度的增加而显著增加, 而表皮组织的体积保持不变(Hanson, 1917; Onoda et al., 2008).而高光条件下, LD的增加与叶脉密度的显著增加有关(Carins Murphy et al., 2012); 此外, 在一定程度上还与栅栏组织细胞排列的紧密度的增加及总的非结构性碳水化合物(TNC)浓度的增加有关(Niinemets et al., 1998; Poorter et al., 2009). ...
Species- independent down-regulation of leaf photosynthesis and respiration in response to shading: Evidence from six temperate tree species.
1
2014
... 关于植物与光照的关系, 目前研究的比较多的是森林生态系统中光照不足对植物产生的影响.在森林生态系统中, 对光的需求的差异是决定植物生长和生存的主要因素(Hanba et al., 2002).大多研究表明植物在适应光环境的过程中主要通过调节叶片形态和结构适应光照的变化, 主要表现在LMA的增大或减小(Niinemets, 2007; Lusk et al., 2008; Coble & Cavaleri, 2015).目前的研究表明, 对于同一种植物, 遮阴导致叶片LMA、厚度、密度下降, 而单位干质量的叶面积增加, 增大了其叶片捕光面积, 利于在弱光环境下生存(冯玉龙等, 2002; Lusk et al., 2008; 柳凤娟等, 2010; 王博轶等, 2012; Coble & Cavaleri, 2015).在植物的冠层内部, 阴生叶比阳生叶的LMA低, 差异可达2-4倍, LMA降低主要与叶肉细胞层厚度减少, 叶片变薄, 叶片密度显著降低, 细胞内含物显著下降有关(Niinemets, 2007; Lusk et al., 2010; Chen et al., 2014).Meng等(2013, 2014)的研究发现, 遮阴不仅会导致叶片的LMA下降, 还会使叶片倾角变小, 以截获更多的光照.值得关注的是, 在很多胁迫条件下(比如干旱、低温), 叶片LMA的增加伴随细胞壁的增厚, 然而在遮阴条件下, 虽然LMA降低了, 但其细胞壁反而增厚, 这可能是植物在低光条件下为增强其叶片硬度和韧度, 避免食草动物的取食而采取的防御机制(Hanba et al., 2002; Coble & Cavaleri, 2015).此外, 很多学者在对热带常绿和落叶木本植物的研究中发现, 在同一低光环境中, 耐阴性强的植物与需光植物相比, 具有较低的光合能力和较高的LMA, LMA的增加在一定程度上与叶肉细胞组织孔隙度的下降、叶片表皮毛密度的增加有关(Hanba et al., 2002; Lusk et al., 2008).LMA的增加使叶片更加强韧, 可增强植物对食草动物和病原菌的防御能力, 提高其在弱光下的生存几率(Kitajima et al., 1994). ...
Leaf fracture toughness and sclerophylly: Their correlation and ecological implications.
1
1992
... 20世纪90年代, 很多学者对不同植物的LMA做了大量研究, 并探讨了LVA和LD与LMA之间的关系.研究表明在同一可控环境条件下, 草本植物(Garnier & Laurent, 1994; van Arendonk & Poorter, 1994)和木本植物(Castro-Diez et al., 2000)的LMA主要受LD的影响, Poorter等(2009)对这些数据作了统计和分析, 表明LMA差异的80%由LD决定, 20%由LVA决定.然而在复杂的自然环境条件下, LD和LVA与LMA之间的关系是多样化的(Choong et al., 1992; Niinemets, 1999; Wilson et al., 1999).由此可见, 仅仅把LMA分解为LVA和LD并不能很好地解释导致植物种类之间LMA差异的原因. ...
The plant traits that drive ecosystems: Evidence from three continents.
1
2004
... LMA反映了植物在单位叶面积上的投入成本, 是叶片经济型谱中最基础的叶功能性状.植物LMA也是最易定量测定的复合结构参数, 与植物学的很多研究领域紧密相关.LMA在农艺学中可作为反映作物生产力的指标; 在生态学中是反映植物生态策略的重要指标; 在生理学上与植物的光合作用及抗逆生理生化反应密切相关.LMA在个体水平与植物的细胞结构及生理反应相关; 而在群落水平可以通过改变群落的初级生产力、凋落物的分解速率、碳氮的循环对生态系统功能产生重要的影响(Díaz et al., 2004; Poorter et al., 2009).因此, 无论从植物的个体还是群落水平研究LMA均具有重要意义.到目前为止, 关于植物LMA已在解剖结构、生理生态学方面展开了深入的研究, 但还有一些问题值得进一步关注和继续完善研究. ...
Incorporating plant functional diversity effects in ecosystem service assessments.
1
2007
... (2)植物LMA反映了碳资源的获取能力和资源的利用方式, 与其生态位息息相关.当环境发生改变时, 植物通过调节LMA的大小来改变调整资源的获取方式和生存对策, 最终可能导致其生态位的改变, 由此对整个生态群落产生较大的影响(Garnier et al., 2004; Díaz et al., 2007).因此, 面对全球高温、干旱、水涝等极端气候的频繁出现, 研究逆境条件下植物的生理生化反应及生存策略对于预测气候变化对植物生态的系统的影响具有重要意义.目前, 关于逆境条件下植物LMA的研究相对较少, 逆境引起的LMA的改变最终导致生理反应的改变, 尤其对光合作用的影响最大.研究表明, gm是限制光合作用的一个重要因素, 而LMA的改变总伴随gm的改变.LMA高、叶片厚的植物, 其细胞壁导度是限制gm的主要因素; 而LMA低、叶片薄的植物, 其限制gm的主要因素各不相同, 其机制仍然不是很清楚.有学者提出, 水通道蛋白(aquaporins)和碳酸酐酶(carbonic anhydrases)可能对低LMA植物的gm具有重要调控作用(Tomás et al., 2013).关于两者对gm的调节机制已有一些初步的报道(Perez-Martin et al., 2014; Boudichevskaia et al., 2015), 但是还需要进一步完善研究以明确其调控机理.此外, 研究环境改变对植物LMA、gm、光合效率的影响及三者之间的相互的调控机制对于理解植物的进化适应性以及气候变化对植物的影响具有重要价值, 这不仅是目前研究的重点, 也是将来研究的重要内容. ...
干旱和湿润生境中主要优势树种叶片功能性状的比较
2012
Resistances along the CO2 diffusion pathway inside leaves.
2009
Intraspecific trait variation and covariation in a widespread tree species (Nothofagus pumilio) in southern Chile.
3
2010
... 目前研究表明, 具有不同形态和生境的植物种间或种内的gm差异很大(Flexas et al., 2008; Warren, 2008; Niinemets et al., 2009a, 2011), 而且特定种类或基因型的植物, 其最大gm与其叶片的解剖学特征有很大的关系(Niinemets et al., 2009a; Tosens et al., 2012).叶片内的CO2的扩散路径主要由两种相组成, 一种是细胞间的气相, 另一种是细胞的液相(主要由水和脂类组成) (Niinemets & Reichstein, 2003; Evans et al., 2009).细胞间气相的变化对CO2的扩散能力影响不大, 因此对整个光合能力没有显著的影响(Parkhurst & Mott, 1990; Evans et al., 2009).细胞相主要由细胞壁、质膜、胞液、叶绿体被膜、基质组成, 其中细胞壁和叶绿体被膜对gm的限制最大(Terashima et al., 2011).据目前的研究估算, 细胞壁厚度(Tcw)对gm的影响占25%-50%, 两者呈负相关关系(Evans et al., 2009; Terashima et al., 2011; Tosens et al., 2012).LMA高的植物, 总是伴随细胞壁的增厚, gm下降, CO2的同化速率降低, 最终导致光合能力下降(Niinemets, 1999; Terashima et al., 2006; Niinemets et al., 2007; Hassiotou et al., 2009).关于这一观点, 目前已在很多硬叶木本植物中得到广泛的证实, 如澳大利亚49种山龙眼科(Proteaceae)的植物(Hassiotou et al., 2010), Abies alba和Abies pinsapo (Peguero-Pina et al., 2012), 冬青栎(Quercus ilex)、柑橘(Citrus reticulate)、橡皮树(Ficus elastica)、海桐(Pittosporum tobira)、华盛顿棕榈(Washingtonia filifera) (Tomás et al., 2013).而且在冠层内, LMA随树木高度增加而增加的过程中所伴随的最大光合效率的下降的主要因素也是CO2的扩散阻力增加, gm下降(Han, 2011). ...
... ; Evans et al., 2009).细胞相主要由细胞壁、质膜、胞液、叶绿体被膜、基质组成, 其中细胞壁和叶绿体被膜对gm的限制最大(Terashima et al., 2011).据目前的研究估算, 细胞壁厚度(Tcw)对gm的影响占25%-50%, 两者呈负相关关系(Evans et al., 2009; Terashima et al., 2011; Tosens et al., 2012).LMA高的植物, 总是伴随细胞壁的增厚, gm下降, CO2的同化速率降低, 最终导致光合能力下降(Niinemets, 1999; Terashima et al., 2006; Niinemets et al., 2007; Hassiotou et al., 2009).关于这一观点, 目前已在很多硬叶木本植物中得到广泛的证实, 如澳大利亚49种山龙眼科(Proteaceae)的植物(Hassiotou et al., 2010), Abies alba和Abies pinsapo (Peguero-Pina et al., 2012), 冬青栎(Quercus ilex)、柑橘(Citrus reticulate)、橡皮树(Ficus elastica)、海桐(Pittosporum tobira)、华盛顿棕榈(Washingtonia filifera) (Tomás et al., 2013).而且在冠层内, LMA随树木高度增加而增加的过程中所伴随的最大光合效率的下降的主要因素也是CO2的扩散阻力增加, gm下降(Han, 2011). ...
... 的影响占25%-50%, 两者呈负相关关系(Evans et al., 2009; Terashima et al., 2011; Tosens et al., 2012).LMA高的植物, 总是伴随细胞壁的增厚, gm下降, CO2的同化速率降低, 最终导致光合能力下降(Niinemets, 1999; Terashima et al., 2006; Niinemets et al., 2007; Hassiotou et al., 2009).关于这一观点, 目前已在很多硬叶木本植物中得到广泛的证实, 如澳大利亚49种山龙眼科(Proteaceae)的植物(Hassiotou et al., 2010), Abies alba和Abies pinsapo (Peguero-Pina et al., 2012), 冬青栎(Quercus ilex)、柑橘(Citrus reticulate)、橡皮树(Ficus elastica)、海桐(Pittosporum tobira)、华盛顿棕榈(Washingtonia filifera) (Tomás et al., 2013).而且在冠层内, LMA随树木高度增加而增加的过程中所伴随的最大光合效率的下降的主要因素也是CO2的扩散阻力增加, gm下降(Han, 2011). ...
四种热带雨林树种幼苗比叶重, 光合特性和暗呼吸对生长光环境的适应
1
2002
... 要进一步研究引起LMA变化的原因, 需要在组织水平进一步分解LMA.20世纪50年代以来, 很多学者在植物解剖结构方面开展了大量的研究, 表明不同的组织在整个叶片中的比例不一样, 同一组织在叶片中的比例受植物种类和环境的影响(Witkowski & Lamont, 1991; Garnier & Laurent, 1994; Poorter et al., 2009; Villar et al., 2013).在环境梯度上, 植物种间的LMA与其叶片组织特征(叶厚度、叶密度)的变化趋势、变化程度不一致(Fajardo & Piper, 2010; Richardson et al., 2013), 因此要想揭示种间LMA差异的原因, 必须分解LMA.在一些研究中, 常常将组织厚度当作组织体积, 这对于阔叶型的扁平状叶片的组织(叶肉组织或上表皮)是适当的, 但很多植物的叶片本身不规则(如松科、柏科), 而且叶片本身一些组织的形状也不规则, 例如维管组织, 因此将组织体积理解为组织厚度并不合适(Poorter et al., 2009).Roderick等(1999)提出LMA的计算公式中的体积应该是各部分组织的体积加上气室体积.Poorter等(2009)从组织水平提出LMA的计算公式: ...
Seasonal patterns and control of gas exchange in local populations of the Mediterranean evergreen shrub Pistacia lentiscus L.
2001
Mesophyll conductance to CO2: Current knowledge and future prospects. Plant,
1
2008
... 关于植物与光照的关系, 目前研究的比较多的是森林生态系统中光照不足对植物产生的影响.在森林生态系统中, 对光的需求的差异是决定植物生长和生存的主要因素(Hanba et al., 2002).大多研究表明植物在适应光环境的过程中主要通过调节叶片形态和结构适应光照的变化, 主要表现在LMA的增大或减小(Niinemets, 2007; Lusk et al., 2008; Coble & Cavaleri, 2015).目前的研究表明, 对于同一种植物, 遮阴导致叶片LMA、厚度、密度下降, 而单位干质量的叶面积增加, 增大了其叶片捕光面积, 利于在弱光环境下生存(冯玉龙等, 2002; Lusk et al., 2008; 柳凤娟等, 2010; 王博轶等, 2012; Coble & Cavaleri, 2015).在植物的冠层内部, 阴生叶比阳生叶的LMA低, 差异可达2-4倍, LMA降低主要与叶肉细胞层厚度减少, 叶片变薄, 叶片密度显著降低, 细胞内含物显著下降有关(Niinemets, 2007; Lusk et al., 2010; Chen et al., 2014).Meng等(2013, 2014)的研究发现, 遮阴不仅会导致叶片的LMA下降, 还会使叶片倾角变小, 以截获更多的光照.值得关注的是, 在很多胁迫条件下(比如干旱、低温), 叶片LMA的增加伴随细胞壁的增厚, 然而在遮阴条件下, 虽然LMA降低了, 但其细胞壁反而增厚, 这可能是植物在低光条件下为增强其叶片硬度和韧度, 避免食草动物的取食而采取的防御机制(Hanba et al., 2002; Coble & Cavaleri, 2015).此外, 很多学者在对热带常绿和落叶木本植物的研究中发现, 在同一低光环境中, 耐阴性强的植物与需光植物相比, 具有较低的光合能力和较高的LMA, LMA的增加在一定程度上与叶肉细胞组织孔隙度的下降、叶片表皮毛密度的增加有关(Hanba et al., 2002; Lusk et al., 2008).LMA的增加使叶片更加强韧, 可增强植物对食草动物和病原菌的防御能力, 提高其在弱光下的生存几率(Kitajima et al., 1994). ...
An evolutionary perspective on leaf economics: Phylogenetics of leaf mass per area in vascular plants.
1
2014
... 短时间的水分胁迫对植物的叶片解剖结构没有影响, 植物只有处于长期缺水状态才会使叶片的解剖结构发生实质性的改变(Poorter et al., 2009).目前, 大多数学者的研究表明耐旱性强的植物一般表现出较高的LMA, 这与叶片厚度和密度的增加有关(Niinemets et al., 2009b).龚春梅和王根轩(2010)通过研究干旱半干旱地区植物群落特征发现植物的LMA随干旱胁迫的加剧而增大.董廷发等(2012)的研究表明在植物群落水平上, 干旱生境中植物的LMA显著高于湿润生境.在长期水分胁迫条件下, 叶片的各组织和细胞结构的改变是导致LMA增加的直接原因.植物经过长期的水分胁迫, 叶片的厚度显著增加, 这与叶片上表皮组织、栅栏组织、海绵组织的增厚有关, 尤其是栅栏组织(Boughalleb & Hajlaoui, 2011; Galmés et al., 2013).此外, 叶片细胞的大小和结构也会发生适应性改变.水分胁迫下发育的叶片其扩展速率较低, 细胞因此更小, 更紧凑, 气室变小(Shield, 1950).Zhang等(2011)的研究发现水杉(Metasequoia glyptostroboides)幼苗叶片在长期水分胁迫下, 其叶肉细胞及上表皮细胞显著变小.细胞结构的改变主要表现为细胞壁增厚, 木质素含量增加(Sobrado, 1992; Henry et al., 2000).以上这些叶片组织和细胞的改变最终表现为叶片的LD及LMA逐渐增加(Nogués & Baker, 2000; Boughalleb & Hajlaoui, 2011).即使水分胁迫解除, 植物仍然会维持叶片解剖结构的适应性的改变; 而且经过干旱胁迫后的植物拥有更强韧的叶片及更高的LMA (Kalapos et al., 1996; Niinemets et al., 2009b; Boughalleb & Hajlaoui, 2011).这种适应性的改变最终对植物的生理过程尤其是光合作用产生较大的影响.很多研究表明植物遭受干旱胁迫后的光合速率会显著下降, 即使胁迫解除后也很难再恢复到胁迫前的值, 这与叶片解剖结构适应性的改变有关(Flexas et al., 2001; Gallé et al., 2009; Galmés et al., 2011).近年来的研究强调gm在限制植物光合速率从水分胁迫恢复中扮演重要角色; 而且大多研究认为叶片解剖学的适应性的改变是引起gm下降的重要原因, 如水分胁迫引起的LMA、LD增加、尤其是细胞壁增厚(Gallé et al., 2009; Boughalleb & Hajlaoui, 2011; Galmés et al., 2013). ...
The role of mesophyll conductance during water stress and recovery in tobacco (Nicotiana sylvestris): Acclimation or limitation?
3
2009
... 据目前的文献报道总结, 多数学者认为高LMA往往伴随较低的叶肉细胞导度(gm), 这是导致高LMA的植物光合作用下降的重要原因(Flexas et al., 2008; Hassiotou et al., 2009; Niinemets et al., 2009a).gm指CO2从气孔下腔到叶绿体羧化位点的扩散能力, 是限制植物光合能力的重要因素 (Hassiotou et al., 2009, 2010; Niinemets et al., 2009a).Flexas等(2008)根据发表文献整理了不同植物的LMA所对应的gm (图2), 结果表明高LMA往往导致较低的gm. ...
... 目前研究表明, 具有不同形态和生境的植物种间或种内的gm差异很大(Flexas et al., 2008; Warren, 2008; Niinemets et al., 2009a, 2011), 而且特定种类或基因型的植物, 其最大gm与其叶片的解剖学特征有很大的关系(Niinemets et al., 2009a; Tosens et al., 2012).叶片内的CO2的扩散路径主要由两种相组成, 一种是细胞间的气相, 另一种是细胞的液相(主要由水和脂类组成) (Niinemets & Reichstein, 2003; Evans et al., 2009).细胞间气相的变化对CO2的扩散能力影响不大, 因此对整个光合能力没有显著的影响(Parkhurst & Mott, 1990; Evans et al., 2009).细胞相主要由细胞壁、质膜、胞液、叶绿体被膜、基质组成, 其中细胞壁和叶绿体被膜对gm的限制最大(Terashima et al., 2011).据目前的研究估算, 细胞壁厚度(Tcw)对gm的影响占25%-50%, 两者呈负相关关系(Evans et al., 2009; Terashima et al., 2011; Tosens et al., 2012).LMA高的植物, 总是伴随细胞壁的增厚, gm下降, CO2的同化速率降低, 最终导致光合能力下降(Niinemets, 1999; Terashima et al., 2006; Niinemets et al., 2007; Hassiotou et al., 2009).关于这一观点, 目前已在很多硬叶木本植物中得到广泛的证实, 如澳大利亚49种山龙眼科(Proteaceae)的植物(Hassiotou et al., 2010), Abies alba和Abies pinsapo (Peguero-Pina et al., 2012), 冬青栎(Quercus ilex)、柑橘(Citrus reticulate)、橡皮树(Ficus elastica)、海桐(Pittosporum tobira)、华盛顿棕榈(Washingtonia filifera) (Tomás et al., 2013).而且在冠层内, LMA随树木高度增加而增加的过程中所伴随的最大光合效率的下降的主要因素也是CO2的扩散阻力增加, gm下降(Han, 2011). ...
... 草本植物与木本植物相比, 叶片薄而柔软, LMA低, 有较高的gm和光合速率(Flexas et al., 2008; Scafaro et al., 2011; Tomás et al., 2013; Muir et al., 2014).Flexas等(2008)研究表明: LMA低的植物, 其gm较高且变化幅度较大, 也就是不同种类或功能群的低LMA植物的LMA与其gm之间没有显著的相关性(Flexas et al., 2008).但是, 近年来也有不同的研究结果, 通过对番茄野生近缘种(Solanum sect. Lycopersicon, sect. Lycopersicoides) (Muir et al., 2014)、水稻野生近缘种(Oryza spp.) (Scafaro et al., 2011)、向日葵(Helianthus annuus)、辣椒(Capsicum annuum)、菜豆(Phaseolus vulgaris)、菠菜(Spinacea oleracea)、罗勒(Ocimum basilicum) (Tomás et al., 2013)的研究表明草本植物的LMA与gm也呈负相关关系, 但是导致gm下降的原因却各有不同.Muir等(2014)通过对8种番茄的研究表明, LMA与gm的关系主要受细胞内含物和叶片厚度的调控.Scafaro等(2011)研究表明, 在3种水稻中, 叶肉细胞壁增厚是gm下降, 并导致光合速率下降的重要原因.Tomás等(2013)对5种不同科属的草本植物的研究指出: 草本植物的细胞壁厚度对gm没有显著的影响, 细胞质和基质对gm影响约占60%, 是影响叶肉细胞导度的重要原因.由此来看, 导致草本植物gm下降的原因应该与植物的种的特异性有关, 具体原因还需要进一步研究.总之, 根据目前的资料报道, 无论是木本植物还是草本植物, LMA与gm之间的负相关关系是导致高LMA的植物光合效率下降的主要原因. ...
Physiological and morphological adaptations in relation to water use efficiency in Mediterranean accessions of Solanum lycopersicum. Plant,
4
2011
... LMA不仅决定植物潜在的生长速率, 反映植物生态策略, 而且与许多生理过程息息相关(Antúnez et al., 2001; Villar et al., 2006).全球叶片经济型谱的研究明确了LMA与叶片单位干物质的最大光合能力(Amass)密切相关, Amass随LMA的增加有明显的下降趋势.LMA的大小主要由植物叶片的厚度和密度决定(Witkowski & Lamont, 1991), 两者的改变都会影响光的利用效率以及叶肉细胞导度(gm)的大小, 最终导致光合效率的改变(Muir et al., 2014).当受到各种选择压力时, 植物能通过调节LMA来改变光合效率, 使植物的投入和收益重新达到最佳的平衡点.研究表明, LMA是自然选择压力下的稳定遗传的结果, 反映了植物与环境共进化的过程, 具有遗传进化上的意义(Asner et al., 2011; Aspinwall et al., 2013; Flores et al., 2014; Tsujii et al., 2016).因此, 研究LMA具有生理、生态及进化的重要意义.从目前已发表的文章来看, 国内还缺乏关于LMA较深入的综合性报道.本文系统梳理了LMA在叶片整体、组织、细胞三个水平的结构解析和计算方法; 重点分析了LMA对光合作用的影响; 讨论了LMA的内在的遗传差异以及外部的环境胁迫因子(温度、水分、光照)对LMA的影响, 以期为今后更深入的研究提供借鉴和参考. ...
... 不同种类的植物, 其LMA也不相同, LMA究竟由环境决定还是植物的系统发育决定的呢?Asner等(2011)研究2873株植物在不同地点、不同气候条件的LMA差异原因, 发现LMA主要由植物分类地位决定, 环境因素处于次要地位.当面对气候变化时, 植物种间LMA的适应性的改变也是有差异的, Aspinwall等(2013)认为这种差异主要由植物的遗传因素决定.Flores等(2014)通过建立5401种维管植物的LMA数据资料库对LMA的进化历史进行了大规模的数据分析, 实验结果表明, LMA中系统发育的信号虽然低, 但却非常保守, 也就是具有亲缘关系的物种, 其LMA也会更相似.此外LMA的保守性与植物的生长型显著相关, 也就是选择强度与表型的进化速率是一致的, 木本植物比草本植物在稳定化选择方面表现出更高的保守性, 因此木本植物的进化速率比草本植物低(Flores et al., 2014).Tsujii等(2016)通过对夏威夷群岛的铁心木(Metrosideros polymorpha) LMA的研究表明环境梯度导致的表型遗传变异是物种进化的主要驱动力.目前的研究均赋予了LMA遗传进化上的重要意义.由此可见, 植物的遗传基础是决定LMA的内在因素, 而环境是外在的推动力.据目前文献资料的报道分析, 5%-95%的陆生植物的LMA的分布范围为30 g·m-2至330 g·m-2, 不同物种的LMA相差约100倍(Poorter et al., 2009).不同功能群植物LMA总的趋势为水生植物<蕨类植物<草本植物<落叶灌木或乔木<常绿灌木、乔木或多浆植物(郑淑霞和上官周平, 2007; Poorter et al., 2009; Flores et al., 2014). ...
... ; Flores et al., 2014). ...
... 草本植物的LMA普遍比木本植物的低, 叶片生长速度快, 资源获取和流动的速度快, 进化程度比木本植物高(Flores et al., 2014).一般来说, 多年生草本植物比一年生草本植物的LMA高.但在双子叶草本植物中, 一年生与多年生植物之间的LMA差异不大; 而单子叶植物中, 禾本科多年生植物比一年生植物的LMA平均高60% (Poorter et al., 2009).而且同一片叶不同部位的LMA也不同, Niinemets等(2007)研究了大量的草本植物叶片, 表明叶片中脉部分的LMA比叶片其余部分的LMA高, 之间的差异约为6倍.然而, 这种差异可能是由物种的特异性决定的, 小麦叶片中脉的LMA与叶片其余部分的LMA差异不大(Rawson et al., 1987).而且大多数禾本科植物基部叶片的LMA比末端叶片的LMA高(Poorter et al., 2009).Pinto等(2015)的研究表明C4禾本科植物的LMA主要与其亚型有关.但目前关于草本植物的LMA的相关的研究还不是很多, 尤其是关于禾本科植物的研究. ...
Leaf responses to drought stress in Mediterranean accessions of Solanum lycopersicum: Anatomical adaptations in relation to gas exchange parameters. Plant,
2
2013
... 短时间的水分胁迫对植物的叶片解剖结构没有影响, 植物只有处于长期缺水状态才会使叶片的解剖结构发生实质性的改变(Poorter et al., 2009).目前, 大多数学者的研究表明耐旱性强的植物一般表现出较高的LMA, 这与叶片厚度和密度的增加有关(Niinemets et al., 2009b).龚春梅和王根轩(2010)通过研究干旱半干旱地区植物群落特征发现植物的LMA随干旱胁迫的加剧而增大.董廷发等(2012)的研究表明在植物群落水平上, 干旱生境中植物的LMA显著高于湿润生境.在长期水分胁迫条件下, 叶片的各组织和细胞结构的改变是导致LMA增加的直接原因.植物经过长期的水分胁迫, 叶片的厚度显著增加, 这与叶片上表皮组织、栅栏组织、海绵组织的增厚有关, 尤其是栅栏组织(Boughalleb & Hajlaoui, 2011; Galmés et al., 2013).此外, 叶片细胞的大小和结构也会发生适应性改变.水分胁迫下发育的叶片其扩展速率较低, 细胞因此更小, 更紧凑, 气室变小(Shield, 1950).Zhang等(2011)的研究发现水杉(Metasequoia glyptostroboides)幼苗叶片在长期水分胁迫下, 其叶肉细胞及上表皮细胞显著变小.细胞结构的改变主要表现为细胞壁增厚, 木质素含量增加(Sobrado, 1992; Henry et al., 2000).以上这些叶片组织和细胞的改变最终表现为叶片的LD及LMA逐渐增加(Nogués & Baker, 2000; Boughalleb & Hajlaoui, 2011).即使水分胁迫解除, 植物仍然会维持叶片解剖结构的适应性的改变; 而且经过干旱胁迫后的植物拥有更强韧的叶片及更高的LMA (Kalapos et al., 1996; Niinemets et al., 2009b; Boughalleb & Hajlaoui, 2011).这种适应性的改变最终对植物的生理过程尤其是光合作用产生较大的影响.很多研究表明植物遭受干旱胁迫后的光合速率会显著下降, 即使胁迫解除后也很难再恢复到胁迫前的值, 这与叶片解剖结构适应性的改变有关(Flexas et al., 2001; Gallé et al., 2009; Galmés et al., 2011).近年来的研究强调gm在限制植物光合速率从水分胁迫恢复中扮演重要角色; 而且大多研究认为叶片解剖学的适应性的改变是引起gm下降的重要原因, 如水分胁迫引起的LMA、LD增加、尤其是细胞壁增厚(Gallé et al., 2009; Boughalleb & Hajlaoui, 2011; Galmés et al., 2013). ...
... 增加、尤其是细胞壁增厚(Gallé et al., 2009; Boughalleb & Hajlaoui, 2011; Galmés et al., 2013). ...
Leaf anatomy, specific mass and water content in congeneric annual and perennial grass species.
1
1994
... 短时间的水分胁迫对植物的叶片解剖结构没有影响, 植物只有处于长期缺水状态才会使叶片的解剖结构发生实质性的改变(Poorter et al., 2009).目前, 大多数学者的研究表明耐旱性强的植物一般表现出较高的LMA, 这与叶片厚度和密度的增加有关(Niinemets et al., 2009b).龚春梅和王根轩(2010)通过研究干旱半干旱地区植物群落特征发现植物的LMA随干旱胁迫的加剧而增大.董廷发等(2012)的研究表明在植物群落水平上, 干旱生境中植物的LMA显著高于湿润生境.在长期水分胁迫条件下, 叶片的各组织和细胞结构的改变是导致LMA增加的直接原因.植物经过长期的水分胁迫, 叶片的厚度显著增加, 这与叶片上表皮组织、栅栏组织、海绵组织的增厚有关, 尤其是栅栏组织(Boughalleb & Hajlaoui, 2011; Galmés et al., 2013).此外, 叶片细胞的大小和结构也会发生适应性改变.水分胁迫下发育的叶片其扩展速率较低, 细胞因此更小, 更紧凑, 气室变小(Shield, 1950).Zhang等(2011)的研究发现水杉(Metasequoia glyptostroboides)幼苗叶片在长期水分胁迫下, 其叶肉细胞及上表皮细胞显著变小.细胞结构的改变主要表现为细胞壁增厚, 木质素含量增加(Sobrado, 1992; Henry et al., 2000).以上这些叶片组织和细胞的改变最终表现为叶片的LD及LMA逐渐增加(Nogués & Baker, 2000; Boughalleb & Hajlaoui, 2011).即使水分胁迫解除, 植物仍然会维持叶片解剖结构的适应性的改变; 而且经过干旱胁迫后的植物拥有更强韧的叶片及更高的LMA (Kalapos et al., 1996; Niinemets et al., 2009b; Boughalleb & Hajlaoui, 2011).这种适应性的改变最终对植物的生理过程尤其是光合作用产生较大的影响.很多研究表明植物遭受干旱胁迫后的光合速率会显著下降, 即使胁迫解除后也很难再恢复到胁迫前的值, 这与叶片解剖结构适应性的改变有关(Flexas et al., 2001; Gallé et al., 2009; Galmés et al., 2011).近年来的研究强调gm在限制植物光合速率从水分胁迫恢复中扮演重要角色; 而且大多研究认为叶片解剖学的适应性的改变是引起gm下降的重要原因, 如水分胁迫引起的LMA、LD增加、尤其是细胞壁增厚(Gallé et al., 2009; Boughalleb & Hajlaoui, 2011; Galmés et al., 2013). ...
Plant functional markers capture ecosystem properties during secondary succession.
2
2004
... 短时间的水分胁迫对植物的叶片解剖结构没有影响, 植物只有处于长期缺水状态才会使叶片的解剖结构发生实质性的改变(Poorter et al., 2009).目前, 大多数学者的研究表明耐旱性强的植物一般表现出较高的LMA, 这与叶片厚度和密度的增加有关(Niinemets et al., 2009b).龚春梅和王根轩(2010)通过研究干旱半干旱地区植物群落特征发现植物的LMA随干旱胁迫的加剧而增大.董廷发等(2012)的研究表明在植物群落水平上, 干旱生境中植物的LMA显著高于湿润生境.在长期水分胁迫条件下, 叶片的各组织和细胞结构的改变是导致LMA增加的直接原因.植物经过长期的水分胁迫, 叶片的厚度显著增加, 这与叶片上表皮组织、栅栏组织、海绵组织的增厚有关, 尤其是栅栏组织(Boughalleb & Hajlaoui, 2011; Galmés et al., 2013).此外, 叶片细胞的大小和结构也会发生适应性改变.水分胁迫下发育的叶片其扩展速率较低, 细胞因此更小, 更紧凑, 气室变小(Shield, 1950).Zhang等(2011)的研究发现水杉(Metasequoia glyptostroboides)幼苗叶片在长期水分胁迫下, 其叶肉细胞及上表皮细胞显著变小.细胞结构的改变主要表现为细胞壁增厚, 木质素含量增加(Sobrado, 1992; Henry et al., 2000).以上这些叶片组织和细胞的改变最终表现为叶片的LD及LMA逐渐增加(Nogués & Baker, 2000; Boughalleb & Hajlaoui, 2011).即使水分胁迫解除, 植物仍然会维持叶片解剖结构的适应性的改变; 而且经过干旱胁迫后的植物拥有更强韧的叶片及更高的LMA (Kalapos et al., 1996; Niinemets et al., 2009b; Boughalleb & Hajlaoui, 2011).这种适应性的改变最终对植物的生理过程尤其是光合作用产生较大的影响.很多研究表明植物遭受干旱胁迫后的光合速率会显著下降, 即使胁迫解除后也很难再恢复到胁迫前的值, 这与叶片解剖结构适应性的改变有关(Flexas et al., 2001; Gallé et al., 2009; Galmés et al., 2011).近年来的研究强调gm在限制植物光合速率从水分胁迫恢复中扮演重要角色; 而且大多研究认为叶片解剖学的适应性的改变是引起gm下降的重要原因, 如水分胁迫引起的LMA、LD增加、尤其是细胞壁增厚(Gallé et al., 2009; Boughalleb & Hajlaoui, 2011; Galmés et al., 2013). ...
... ; Galmés et al., 2013). ...
干旱半干旱区降水量梯度下植物群落的叶、茎属性特征
2
2010
... 20世纪90年代, 很多学者对不同植物的LMA做了大量研究, 并探讨了LVA和LD与LMA之间的关系.研究表明在同一可控环境条件下, 草本植物(Garnier & Laurent, 1994; van Arendonk & Poorter, 1994)和木本植物(Castro-Diez et al., 2000)的LMA主要受LD的影响, Poorter等(2009)对这些数据作了统计和分析, 表明LMA差异的80%由LD决定, 20%由LVA决定.然而在复杂的自然环境条件下, LD和LVA与LMA之间的关系是多样化的(Choong et al., 1992; Niinemets, 1999; Wilson et al., 1999).由此可见, 仅仅把LMA分解为LVA和LD并不能很好地解释导致植物种类之间LMA差异的原因. ...
... 要进一步研究引起LMA变化的原因, 需要在组织水平进一步分解LMA.20世纪50年代以来, 很多学者在植物解剖结构方面开展了大量的研究, 表明不同的组织在整个叶片中的比例不一样, 同一组织在叶片中的比例受植物种类和环境的影响(Witkowski & Lamont, 1991; Garnier & Laurent, 1994; Poorter et al., 2009; Villar et al., 2013).在环境梯度上, 植物种间的LMA与其叶片组织特征(叶厚度、叶密度)的变化趋势、变化程度不一致(Fajardo & Piper, 2010; Richardson et al., 2013), 因此要想揭示种间LMA差异的原因, 必须分解LMA.在一些研究中, 常常将组织厚度当作组织体积, 这对于阔叶型的扁平状叶片的组织(叶肉组织或上表皮)是适当的, 但很多植物的叶片本身不规则(如松科、柏科), 而且叶片本身一些组织的形状也不规则, 例如维管组织, 因此将组织体积理解为组织厚度并不合适(Poorter et al., 2009).Roderick等(1999)提出LMA的计算公式中的体积应该是各部分组织的体积加上气室体积.Poorter等(2009)从组织水平提出LMA的计算公式: ...
Temporal hetero- geneity of cold acclimation phenotypes in Arabidopsis leaves. Plant,
1
2010
... (2)植物LMA反映了碳资源的获取能力和资源的利用方式, 与其生态位息息相关.当环境发生改变时, 植物通过调节LMA的大小来改变调整资源的获取方式和生存对策, 最终可能导致其生态位的改变, 由此对整个生态群落产生较大的影响(Garnier et al., 2004; Díaz et al., 2007).因此, 面对全球高温、干旱、水涝等极端气候的频繁出现, 研究逆境条件下植物的生理生化反应及生存策略对于预测气候变化对植物生态的系统的影响具有重要意义.目前, 关于逆境条件下植物LMA的研究相对较少, 逆境引起的LMA的改变最终导致生理反应的改变, 尤其对光合作用的影响最大.研究表明, gm是限制光合作用的一个重要因素, 而LMA的改变总伴随gm的改变.LMA高、叶片厚的植物, 其细胞壁导度是限制gm的主要因素; 而LMA低、叶片薄的植物, 其限制gm的主要因素各不相同, 其机制仍然不是很清楚.有学者提出, 水通道蛋白(aquaporins)和碳酸酐酶(carbonic anhydrases)可能对低LMA植物的gm具有重要调控作用(Tomás et al., 2013).关于两者对gm的调节机制已有一些初步的报道(Perez-Martin et al., 2014; Boudichevskaia et al., 2015), 但是还需要进一步完善研究以明确其调控机理.此外, 研究环境改变对植物LMA、gm、光合效率的影响及三者之间的相互的调控机制对于理解植物的进化适应性以及气候变化对植物的影响具有重要价值, 这不仅是目前研究的重点, 也是将来研究的重要内容. ...
Influence of leaf dry mass per area, CO2 and irradiance on mesophyll conductance in sclerophylls.
2009
Photosynthesis at an extreme end of the leaf trait spectrum: How does it relate to high leaf dry mass per area and associated structural parameters?
2010
Emissions of volatile organic compounds and leaf structural characteristics of European aspen (Populus tremula) grown under elevated ozone and temperature.
2009
The effect of growth irradiance on leaf anatomy and photosynthesis in Acer species differing in light demand. Plant,
3
2002
... 据目前的文献报道总结, 多数学者认为高LMA往往伴随较低的叶肉细胞导度(gm), 这是导致高LMA的植物光合作用下降的重要原因(Flexas et al., 2008; Hassiotou et al., 2009; Niinemets et al., 2009a).gm指CO2从气孔下腔到叶绿体羧化位点的扩散能力, 是限制植物光合能力的重要因素 (Hassiotou et al., 2009, 2010; Niinemets et al., 2009a).Flexas等(2008)根据发表文献整理了不同植物的LMA所对应的gm (图2), 结果表明高LMA往往导致较低的gm. ...
... 从气孔下腔到叶绿体羧化位点的扩散能力, 是限制植物光合能力的重要因素 (Hassiotou et al., 2009, 2010; Niinemets et al., 2009a).Flexas等(2008)根据发表文献整理了不同植物的LMA所对应的gm (图2), 结果表明高LMA往往导致较低的gm. ...
... 目前研究表明, 具有不同形态和生境的植物种间或种内的gm差异很大(Flexas et al., 2008; Warren, 2008; Niinemets et al., 2009a, 2011), 而且特定种类或基因型的植物, 其最大gm与其叶片的解剖学特征有很大的关系(Niinemets et al., 2009a; Tosens et al., 2012).叶片内的CO2的扩散路径主要由两种相组成, 一种是细胞间的气相, 另一种是细胞的液相(主要由水和脂类组成) (Niinemets & Reichstein, 2003; Evans et al., 2009).细胞间气相的变化对CO2的扩散能力影响不大, 因此对整个光合能力没有显著的影响(Parkhurst & Mott, 1990; Evans et al., 2009).细胞相主要由细胞壁、质膜、胞液、叶绿体被膜、基质组成, 其中细胞壁和叶绿体被膜对gm的限制最大(Terashima et al., 2011).据目前的研究估算, 细胞壁厚度(Tcw)对gm的影响占25%-50%, 两者呈负相关关系(Evans et al., 2009; Terashima et al., 2011; Tosens et al., 2012).LMA高的植物, 总是伴随细胞壁的增厚, gm下降, CO2的同化速率降低, 最终导致光合能力下降(Niinemets, 1999; Terashima et al., 2006; Niinemets et al., 2007; Hassiotou et al., 2009).关于这一观点, 目前已在很多硬叶木本植物中得到广泛的证实, 如澳大利亚49种山龙眼科(Proteaceae)的植物(Hassiotou et al., 2010), Abies alba和Abies pinsapo (Peguero-Pina et al., 2012), 冬青栎(Quercus ilex)、柑橘(Citrus reticulate)、橡皮树(Ficus elastica)、海桐(Pittosporum tobira)、华盛顿棕榈(Washingtonia filifera) (Tomás et al., 2013).而且在冠层内, LMA随树木高度增加而增加的过程中所伴随的最大光合效率的下降的主要因素也是CO2的扩散阻力增加, gm下降(Han, 2011). ...
Heigh related decreases in mesophyll conductance, leaf photosynthesis and compensating adjustments associated with leaf nitrogen concentrations in Pinus densiflora.
4
2011
... LMA是一个复合的结构参数, 与叶片的很多解剖结构有关, 因而对光合作用有潜在的影响(Niinemets et al., 2009a; Scafaro et al., 2011; Terashima et al., 2011).近年来, LMA对光合作用的影响引起了生理生态学家的广泛关注.全球叶片经济型谱的建立已明确LMA与叶片单位干物质的最大光合能力、叶龄、单位干物质的氮、磷含量等密切相关(Wright et al., 2004).LMA低的植物, 其叶片生命周期短、单位干物质营养元素含量高, 光合能力强、单位叶面积的经济投入较低, 导致植物资源的利用效率高而生长速度快; LMA高的植物, 其单位叶面积的投入较高, 尤其增加了叶片的维管组织和细胞壁的投入(Niinemets et al., 2007; Hikosaka & Shigeno, 2009), 虽然增强了植物耐贫瘠、耐干旱的能力, 但导致较低的光合能力和缓慢的生长速率(Niinemets, 2001; Wright et al., 2004).近年的研究进一步表明, LMA与植物的单位叶面积的光合能力(Aarea)没有明显的相关性; 但随LMA的增加, 单位叶干质量的最大光合能力(Amass)有明显的下降趋势(Niinemets & Sack, 2006; Hassiotou et al., 2010). ...
... 据目前的文献报道总结, 多数学者认为高LMA往往伴随较低的叶肉细胞导度(gm), 这是导致高LMA的植物光合作用下降的重要原因(Flexas et al., 2008; Hassiotou et al., 2009; Niinemets et al., 2009a).gm指CO2从气孔下腔到叶绿体羧化位点的扩散能力, 是限制植物光合能力的重要因素 (Hassiotou et al., 2009, 2010; Niinemets et al., 2009a).Flexas等(2008)根据发表文献整理了不同植物的LMA所对应的gm (图2), 结果表明高LMA往往导致较低的gm. ...
... 目前研究表明, 具有不同形态和生境的植物种间或种内的gm差异很大(Flexas et al., 2008; Warren, 2008; Niinemets et al., 2009a, 2011), 而且特定种类或基因型的植物, 其最大gm与其叶片的解剖学特征有很大的关系(Niinemets et al., 2009a; Tosens et al., 2012).叶片内的CO2的扩散路径主要由两种相组成, 一种是细胞间的气相, 另一种是细胞的液相(主要由水和脂类组成) (Niinemets & Reichstein, 2003; Evans et al., 2009).细胞间气相的变化对CO2的扩散能力影响不大, 因此对整个光合能力没有显著的影响(Parkhurst & Mott, 1990; Evans et al., 2009).细胞相主要由细胞壁、质膜、胞液、叶绿体被膜、基质组成, 其中细胞壁和叶绿体被膜对gm的限制最大(Terashima et al., 2011).据目前的研究估算, 细胞壁厚度(Tcw)对gm的影响占25%-50%, 两者呈负相关关系(Evans et al., 2009; Terashima et al., 2011; Tosens et al., 2012).LMA高的植物, 总是伴随细胞壁的增厚, gm下降, CO2的同化速率降低, 最终导致光合能力下降(Niinemets, 1999; Terashima et al., 2006; Niinemets et al., 2007; Hassiotou et al., 2009).关于这一观点, 目前已在很多硬叶木本植物中得到广泛的证实, 如澳大利亚49种山龙眼科(Proteaceae)的植物(Hassiotou et al., 2010), Abies alba和Abies pinsapo (Peguero-Pina et al., 2012), 冬青栎(Quercus ilex)、柑橘(Citrus reticulate)、橡皮树(Ficus elastica)、海桐(Pittosporum tobira)、华盛顿棕榈(Washingtonia filifera) (Tomás et al., 2013).而且在冠层内, LMA随树木高度增加而增加的过程中所伴随的最大光合效率的下降的主要因素也是CO2的扩散阻力增加, gm下降(Han, 2011). ...
... 其次, 高LMA植物叶片中氮浓度低可能也是植物光合能力(Amass)降低的原因.植物叶片中大部分的氮参与光合作用的运转, LMA高的植物叶片中的关键营养元素的浓度都比较低, 例如氮、磷(Wright et al., 2004).目前研究表明, 植物叶片中氮的浓度低会导致光合作用的氮利用效率以及分配到核酮糖-1,5-二磷酸羧化酶/加氧酶(Rubisco)和类囊体的氮显著下降, 从而引起光合能力的下降(Hikosaka, 2004; Hassiotou et al., 2010).Niinemets等(2004)研究表明冬青栎的LMA随叶龄的增长而增加, 而氮含量随LMA增加而下降, 氮含量的降低导致Rubisco的最大羧化活性以及光合作用的电子传递能力下降.Xu等(2012)的研究表明柳叶桉(Eucalyptus saligna)的叶氮含量随LMA的增加而降低, 单位质量的最大光合效率随氮含量降低而降低.但并非所有情况下氮含量都随LMA增加而下降, Han (2011)发现LMA随赤松(Pinus densiflora)树高的增加而增加的过程中, Nmass和Narea均表现出增加的趋势, 这可能是植物为了减少因高度引起的光合效率的下降而采取的补偿机制. ...
Leaf-age effects on seasonal variability in photosynthetic parameters and its relationships with leaf mass per area and leaf nitrogen concentration within a Pinus densiflora crown.
1
2008
... 生长在温度较高的环境中的植物通常拥有较低的LMA, 但LMA与温度并不呈线性相关(p < 0.01) (Atkin et al., 2006; Poorter et al., 2009).温度升高引起LMA降低的直接原因是叶片变薄.目前大量实验表明, 生长在高温环境中的植物, 叶面积增大但叶片显著变薄, 这与叶片的表皮层、栅栏组织、海绵组织的细胞层数减少及细胞变小引起的叶组织厚度的降低有关(Hartikainen et al., 2009; Jin et al., 2011).杨兵等(2010)研究发现长期模拟增温条件下, 岷江冷杉(Abies faxoniana)幼苗LMA显著下降.Xu等(2012)通过模拟温室效应来研究气温升高对柳叶桉形态解剖结构的影响, 结果表明随温度的升高柳叶桉叶片LMA呈下降趋势.Zhang等(2015)的研究表明长期热胁迫会导致云杉属(Picea)植物的LMA下降. ...
Leaf structure as related to environment.
4
1917
... 树木冠层内的LMA的差异也是森林生态系统研究的重要内容.LMA与叶片在冠层中的位置有关, 一般处于冠层顶部的叶片LMA值比冠层底部的高(Koch et al., 2004; Lichtenthaler et al., 2007; Poorter et al., 2009; Weerasinghe et al., 2014).据Koch等(2004)报道100 m高的红杉(Sequoia sempervirens)的冠层底部LMA为150 g·m-2, 而冠层顶部LMA值达800 g·m-2, 这是比较极端的例子.一般来说, 不管是木本植物还是草本植物, 其冠层LMA大约是底层LMA的两倍(Poorter et al., 2009).这种底层和冠层之间的差异在一定程度上是由物种的特异性决定的(Lichtenthaler et al., 2007).此外, LMA还随叶寿命的不同表现出较大的差异.叶寿命是植物在长期适应环境的过程中为获得最大光合生产所形成的适应策略(Reich et al., 1991; 张林和罗天祥, 2004).LMA反映了植物单位叶面积的构建成本, 通常与叶寿命呈正相关关系.LMA低的植物, 其光合效率高, 生长速度快, 环境胁迫应答策略表现为逃避, 因而用于防御投资的物质较少, 叶寿命较短, 如草本植物、落叶乔灌木; 而LMA高的植物, 光合效率慢, 生长速度慢, 环境胁迫应答的策略表现为抵御和耐受, 需要不断积累更多的物质构建保卫构造, 因此表现出较长的叶寿命, 如常绿木本植物(Reich et al., 1991; Wright et al., 2004; Wyka et al., 2012; 王瑞雪等, 2014).常绿木本植物的LMA通常随叶龄的增长而逐渐增加, LMA的增加主要与叶肉细胞的细胞壁变厚、木质素含量增加、叶片变厚、叶片的机械组织含量增加有关(Castro-Díez et al., 2000; Hanba et al., 2002; Wright et al., 2004).Niinemets等(2005, 2006)报道地中海3种阔叶常绿树种月桂(Laurus nobilis)、油橄榄(Olea europea)、冬青栎(Quercus ilex)的LMA随叶龄的增长而增加.而且叶龄对LMA的影响还与叶片在冠层中的位置有关, Han等(2008)报道赤松冠层底部LMA随叶龄的增加而增加的幅度为27%, 但叶龄对冠层顶部LMA无影响.此外, 在常绿木本植物中, 叶寿命不同的植物, 其LMA也有较大的差异, Wyka等(2012)报道松柏类比阔叶类植物的叶寿命长, 其LMA也显著高于阔叶类植物. ...
... 关于植物与光照的关系, 目前研究的比较多的是森林生态系统中光照不足对植物产生的影响.在森林生态系统中, 对光的需求的差异是决定植物生长和生存的主要因素(Hanba et al., 2002).大多研究表明植物在适应光环境的过程中主要通过调节叶片形态和结构适应光照的变化, 主要表现在LMA的增大或减小(Niinemets, 2007; Lusk et al., 2008; Coble & Cavaleri, 2015).目前的研究表明, 对于同一种植物, 遮阴导致叶片LMA、厚度、密度下降, 而单位干质量的叶面积增加, 增大了其叶片捕光面积, 利于在弱光环境下生存(冯玉龙等, 2002; Lusk et al., 2008; 柳凤娟等, 2010; 王博轶等, 2012; Coble & Cavaleri, 2015).在植物的冠层内部, 阴生叶比阳生叶的LMA低, 差异可达2-4倍, LMA降低主要与叶肉细胞层厚度减少, 叶片变薄, 叶片密度显著降低, 细胞内含物显著下降有关(Niinemets, 2007; Lusk et al., 2010; Chen et al., 2014).Meng等(2013, 2014)的研究发现, 遮阴不仅会导致叶片的LMA下降, 还会使叶片倾角变小, 以截获更多的光照.值得关注的是, 在很多胁迫条件下(比如干旱、低温), 叶片LMA的增加伴随细胞壁的增厚, 然而在遮阴条件下, 虽然LMA降低了, 但其细胞壁反而增厚, 这可能是植物在低光条件下为增强其叶片硬度和韧度, 避免食草动物的取食而采取的防御机制(Hanba et al., 2002; Coble & Cavaleri, 2015).此外, 很多学者在对热带常绿和落叶木本植物的研究中发现, 在同一低光环境中, 耐阴性强的植物与需光植物相比, 具有较低的光合能力和较高的LMA, LMA的增加在一定程度上与叶肉细胞组织孔隙度的下降、叶片表皮毛密度的增加有关(Hanba et al., 2002; Lusk et al., 2008).LMA的增加使叶片更加强韧, 可增强植物对食草动物和病原菌的防御能力, 提高其在弱光下的生存几率(Kitajima et al., 1994). ...
... 降低了, 但其细胞壁反而增厚, 这可能是植物在低光条件下为增强其叶片硬度和韧度, 避免食草动物的取食而采取的防御机制(Hanba et al., 2002; Coble & Cavaleri, 2015).此外, 很多学者在对热带常绿和落叶木本植物的研究中发现, 在同一低光环境中, 耐阴性强的植物与需光植物相比, 具有较低的光合能力和较高的LMA, LMA的增加在一定程度上与叶肉细胞组织孔隙度的下降、叶片表皮毛密度的增加有关(Hanba et al., 2002; Lusk et al., 2008).LMA的增加使叶片更加强韧, 可增强植物对食草动物和病原菌的防御能力, 提高其在弱光下的生存几率(Kitajima et al., 1994). ...
... 的增加在一定程度上与叶肉细胞组织孔隙度的下降、叶片表皮毛密度的增加有关(Hanba et al., 2002; Lusk et al., 2008).LMA的增加使叶片更加强韧, 可增强植物对食草动物和病原菌的防御能力, 提高其在弱光下的生存几率(Kitajima et al., 1994). ...
Seasonal changes and the effect of temperature and leaf moisture content on intrinsic shear strength of leaves of pasture grasses.
1
2000
... 目前研究表明, 具有不同形态和生境的植物种间或种内的gm差异很大(Flexas et al., 2008; Warren, 2008; Niinemets et al., 2009a, 2011), 而且特定种类或基因型的植物, 其最大gm与其叶片的解剖学特征有很大的关系(Niinemets et al., 2009a; Tosens et al., 2012).叶片内的CO2的扩散路径主要由两种相组成, 一种是细胞间的气相, 另一种是细胞的液相(主要由水和脂类组成) (Niinemets & Reichstein, 2003; Evans et al., 2009).细胞间气相的变化对CO2的扩散能力影响不大, 因此对整个光合能力没有显著的影响(Parkhurst & Mott, 1990; Evans et al., 2009).细胞相主要由细胞壁、质膜、胞液、叶绿体被膜、基质组成, 其中细胞壁和叶绿体被膜对gm的限制最大(Terashima et al., 2011).据目前的研究估算, 细胞壁厚度(Tcw)对gm的影响占25%-50%, 两者呈负相关关系(Evans et al., 2009; Terashima et al., 2011; Tosens et al., 2012).LMA高的植物, 总是伴随细胞壁的增厚, gm下降, CO2的同化速率降低, 最终导致光合能力下降(Niinemets, 1999; Terashima et al., 2006; Niinemets et al., 2007; Hassiotou et al., 2009).关于这一观点, 目前已在很多硬叶木本植物中得到广泛的证实, 如澳大利亚49种山龙眼科(Proteaceae)的植物(Hassiotou et al., 2010), Abies alba和Abies pinsapo (Peguero-Pina et al., 2012), 冬青栎(Quercus ilex)、柑橘(Citrus reticulate)、橡皮树(Ficus elastica)、海桐(Pittosporum tobira)、华盛顿棕榈(Washingtonia filifera) (Tomás et al., 2013).而且在冠层内, LMA随树木高度增加而增加的过程中所伴随的最大光合效率的下降的主要因素也是CO2的扩散阻力增加, gm下降(Han, 2011). ...
Interspecific difference in the photosynthesis- nitrogen relationship: Patterns, physiological causes, and ecological importance.
2004
The role of Rubisco and cell walls in the interspecific variation in photosynthetic capacity.
1
2009
... 植物的LMA随光照强度的增加而增加, 但LMA由总的光照度/天(DPI)决定, 而不是瞬时的高的光照强度决定(Chabot et al., 1979; Niinemets et al., 2004).LMA随DPI的增加而显著增加, 但这种响应呈非线性相关, 因为在低光条件下LMA随DPI的增加而显著增加, 但是当DPI超过20 mol·m-2·d-1时, 其LMA的增加会变缓慢(Poorter et al., 2009).LMA 随光照度增加而增加的过程中, LD和LVA均扮演重要角色.Coble和Cavaleri (2014)的研究表明, 光照增加导致LMA的增加主要与LVA的增加有关.但在LVA增加的过程中, 并不是所有的叶片组织的体积都增加, 一般情况下, 栅栏组织的体积随光强度的增加而显著增加, 而表皮组织的体积保持不变(Hanson, 1917; Onoda et al., 2008).而高光条件下, LD的增加与叶脉密度的显著增加有关(Carins Murphy et al., 2012); 此外, 在一定程度上还与栅栏组织细胞排列的紧密度的增加及总的非结构性碳水化合物(TNC)浓度的增加有关(Niinemets et al., 1998; Poorter et al., 2009). ...
三峡水库消落带植物叶片光合与营养性状特征
1
2012
... 短时间的水分胁迫对植物的叶片解剖结构没有影响, 植物只有处于长期缺水状态才会使叶片的解剖结构发生实质性的改变(Poorter et al., 2009).目前, 大多数学者的研究表明耐旱性强的植物一般表现出较高的LMA, 这与叶片厚度和密度的增加有关(Niinemets et al., 2009b).龚春梅和王根轩(2010)通过研究干旱半干旱地区植物群落特征发现植物的LMA随干旱胁迫的加剧而增大.董廷发等(2012)的研究表明在植物群落水平上, 干旱生境中植物的LMA显著高于湿润生境.在长期水分胁迫条件下, 叶片的各组织和细胞结构的改变是导致LMA增加的直接原因.植物经过长期的水分胁迫, 叶片的厚度显著增加, 这与叶片上表皮组织、栅栏组织、海绵组织的增厚有关, 尤其是栅栏组织(Boughalleb & Hajlaoui, 2011; Galmés et al., 2013).此外, 叶片细胞的大小和结构也会发生适应性改变.水分胁迫下发育的叶片其扩展速率较低, 细胞因此更小, 更紧凑, 气室变小(Shield, 1950).Zhang等(2011)的研究发现水杉(Metasequoia glyptostroboides)幼苗叶片在长期水分胁迫下, 其叶肉细胞及上表皮细胞显著变小.细胞结构的改变主要表现为细胞壁增厚, 木质素含量增加(Sobrado, 1992; Henry et al., 2000).以上这些叶片组织和细胞的改变最终表现为叶片的LD及LMA逐渐增加(Nogués & Baker, 2000; Boughalleb & Hajlaoui, 2011).即使水分胁迫解除, 植物仍然会维持叶片解剖结构的适应性的改变; 而且经过干旱胁迫后的植物拥有更强韧的叶片及更高的LMA (Kalapos et al., 1996; Niinemets et al., 2009b; Boughalleb & Hajlaoui, 2011).这种适应性的改变最终对植物的生理过程尤其是光合作用产生较大的影响.很多研究表明植物遭受干旱胁迫后的光合速率会显著下降, 即使胁迫解除后也很难再恢复到胁迫前的值, 这与叶片解剖结构适应性的改变有关(Flexas et al., 2001; Gallé et al., 2009; Galmés et al., 2011).近年来的研究强调gm在限制植物光合速率从水分胁迫恢复中扮演重要角色; 而且大多研究认为叶片解剖学的适应性的改变是引起gm下降的重要原因, 如水分胁迫引起的LMA、LD增加、尤其是细胞壁增厚(Gallé et al., 2009; Boughalleb & Hajlaoui, 2011; Galmés et al., 2013). ...
The effect of experimental warming on leaf functional traits, leaf structure and leaf biochemistry in Arabidopsis thaliana.
1
2011
... 其次, 高LMA植物叶片中氮浓度低可能也是植物光合能力(Amass)降低的原因.植物叶片中大部分的氮参与光合作用的运转, LMA高的植物叶片中的关键营养元素的浓度都比较低, 例如氮、磷(Wright et al., 2004).目前研究表明, 植物叶片中氮的浓度低会导致光合作用的氮利用效率以及分配到核酮糖-1,5-二磷酸羧化酶/加氧酶(Rubisco)和类囊体的氮显著下降, 从而引起光合能力的下降(Hikosaka, 2004; Hassiotou et al., 2010).Niinemets等(2004)研究表明冬青栎的LMA随叶龄的增长而增加, 而氮含量随LMA增加而下降, 氮含量的降低导致Rubisco的最大羧化活性以及光合作用的电子传递能力下降.Xu等(2012)的研究表明柳叶桉(Eucalyptus saligna)的叶氮含量随LMA的增加而降低, 单位质量的最大光合效率随氮含量降低而降低.但并非所有情况下氮含量都随LMA增加而下降, Han (2011)发现LMA随赤松(Pinus densiflora)树高的增加而增加的过程中, Nmass和Narea均表现出增加的趋势, 这可能是植物为了减少因高度引起的光合效率的下降而采取的补偿机制. ...
Effect of soil drying on growth, biomass allocation and leaf gas exchange of two annual grass species.
2
1996
... 自然界中的每一种植物都有自己独特的叶片形态和结构, 这种特异性并不是偶然的, 而是自然界选择进化的结果.每一种叶片的形态和组织结构都有与之相协调的生理过程紧密联系.因此, 从叶片形态和结构入手揭示植物的生态策略和生理过程是一种行之有效的研究手段.在植物的叶片解剖研究过程中, 大量研究资料显示比叶质量(LMA)是一个非常重要的反应植物功能的结构参数.LMA指叶片干质量与面积之间的比率(g·m-2), 是叶片众多解剖结构特征的综合体, 例如叶肉组织厚度、细胞壁厚度、细胞形状、细胞内含物等(Muir et al., 2014).在大量的叶片形态解剖结构参数中, LMA是最容易定量测定的参数之一, 它能定量测定植物在单位叶面积的投入成本, 因而被认为是反映植物生态策略的重要指标(Westoby et al., 2002; Wright et al., 2004).LMA的高低反映了植物对资源的利用效率以及对环境的应对策略.总的来讲, 低LMA的植物光合效率高, 单位叶面积的投入成本低, 显示出较快的资源流动速度及回报效率; 而高LMA的植物光合效率低, 单位叶面积的投入成本高, 资源的流动速度及回报效率低(Niinemets et al., 2007; Hikosaka & Shigeno, 2009). ...
... LMA是一个复合的结构参数, 与叶片的很多解剖结构有关, 因而对光合作用有潜在的影响(Niinemets et al., 2009a; Scafaro et al., 2011; Terashima et al., 2011).近年来, LMA对光合作用的影响引起了生理生态学家的广泛关注.全球叶片经济型谱的建立已明确LMA与叶片单位干物质的最大光合能力、叶龄、单位干物质的氮、磷含量等密切相关(Wright et al., 2004).LMA低的植物, 其叶片生命周期短、单位干物质营养元素含量高, 光合能力强、单位叶面积的经济投入较低, 导致植物资源的利用效率高而生长速度快; LMA高的植物, 其单位叶面积的投入较高, 尤其增加了叶片的维管组织和细胞壁的投入(Niinemets et al., 2007; Hikosaka & Shigeno, 2009), 虽然增强了植物耐贫瘠、耐干旱的能力, 但导致较低的光合能力和缓慢的生长速率(Niinemets, 2001; Wright et al., 2004).近年的研究进一步表明, LMA与植物的单位叶面积的光合能力(Aarea)没有明显的相关性; 但随LMA的增加, 单位叶干质量的最大光合能力(Amass)有明显的下降趋势(Niinemets & Sack, 2006; Hassiotou et al., 2010). ...
Relative importance of photosynthetic traits and allocation patterns as correlates of seedling shade tolerance of 13 tropical trees.
1994
The limits to tree height.
2004
Unbiased estimation of chloroplast number in mesophyll cells: Advantage of a genuine three- dimensional approach.
1
2014
... 生长在温度较高的环境中的植物通常拥有较低的LMA, 但LMA与温度并不呈线性相关(p < 0.01) (Atkin et al., 2006; Poorter et al., 2009).温度升高引起LMA降低的直接原因是叶片变薄.目前大量实验表明, 生长在高温环境中的植物, 叶面积增大但叶片显著变薄, 这与叶片的表皮层、栅栏组织、海绵组织的细胞层数减少及细胞变小引起的叶组织厚度的降低有关(Hartikainen et al., 2009; Jin et al., 2011).杨兵等(2010)研究发现长期模拟增温条件下, 岷江冷杉(Abies faxoniana)幼苗LMA显著下降.Xu等(2012)通过模拟温室效应来研究气温升高对柳叶桉形态解剖结构的影响, 结果表明随温度的升高柳叶桉叶片LMA呈下降趋势.Zhang等(2015)的研究表明长期热胁迫会导致云杉属(Picea)植物的LMA下降. ...
Different responses in leaf pigments and leaf mass per area to altitudebetween evergreen and deciduous woody species.
1
2013
... 短时间的水分胁迫对植物的叶片解剖结构没有影响, 植物只有处于长期缺水状态才会使叶片的解剖结构发生实质性的改变(Poorter et al., 2009).目前, 大多数学者的研究表明耐旱性强的植物一般表现出较高的LMA, 这与叶片厚度和密度的增加有关(Niinemets et al., 2009b).龚春梅和王根轩(2010)通过研究干旱半干旱地区植物群落特征发现植物的LMA随干旱胁迫的加剧而增大.董廷发等(2012)的研究表明在植物群落水平上, 干旱生境中植物的LMA显著高于湿润生境.在长期水分胁迫条件下, 叶片的各组织和细胞结构的改变是导致LMA增加的直接原因.植物经过长期的水分胁迫, 叶片的厚度显著增加, 这与叶片上表皮组织、栅栏组织、海绵组织的增厚有关, 尤其是栅栏组织(Boughalleb & Hajlaoui, 2011; Galmés et al., 2013).此外, 叶片细胞的大小和结构也会发生适应性改变.水分胁迫下发育的叶片其扩展速率较低, 细胞因此更小, 更紧凑, 气室变小(Shield, 1950).Zhang等(2011)的研究发现水杉(Metasequoia glyptostroboides)幼苗叶片在长期水分胁迫下, 其叶肉细胞及上表皮细胞显著变小.细胞结构的改变主要表现为细胞壁增厚, 木质素含量增加(Sobrado, 1992; Henry et al., 2000).以上这些叶片组织和细胞的改变最终表现为叶片的LD及LMA逐渐增加(Nogués & Baker, 2000; Boughalleb & Hajlaoui, 2011).即使水分胁迫解除, 植物仍然会维持叶片解剖结构的适应性的改变; 而且经过干旱胁迫后的植物拥有更强韧的叶片及更高的LMA (Kalapos et al., 1996; Niinemets et al., 2009b; Boughalleb & Hajlaoui, 2011).这种适应性的改变最终对植物的生理过程尤其是光合作用产生较大的影响.很多研究表明植物遭受干旱胁迫后的光合速率会显著下降, 即使胁迫解除后也很难再恢复到胁迫前的值, 这与叶片解剖结构适应性的改变有关(Flexas et al., 2001; Gallé et al., 2009; Galmés et al., 2011).近年来的研究强调gm在限制植物光合速率从水分胁迫恢复中扮演重要角色; 而且大多研究认为叶片解剖学的适应性的改变是引起gm下降的重要原因, 如水分胁迫引起的LMA、LD增加、尤其是细胞壁增厚(Gallé et al., 2009; Boughalleb & Hajlaoui, 2011; Galmés et al., 2013). ...
Chlorophyll fluorescence imaging of photosynthetic activity in sun and shade leaves of trees.
1
2007
... 关于植物与光照的关系, 目前研究的比较多的是森林生态系统中光照不足对植物产生的影响.在森林生态系统中, 对光的需求的差异是决定植物生长和生存的主要因素(Hanba et al., 2002).大多研究表明植物在适应光环境的过程中主要通过调节叶片形态和结构适应光照的变化, 主要表现在LMA的增大或减小(Niinemets, 2007; Lusk et al., 2008; Coble & Cavaleri, 2015).目前的研究表明, 对于同一种植物, 遮阴导致叶片LMA、厚度、密度下降, 而单位干质量的叶面积增加, 增大了其叶片捕光面积, 利于在弱光环境下生存(冯玉龙等, 2002; Lusk et al., 2008; 柳凤娟等, 2010; 王博轶等, 2012; Coble & Cavaleri, 2015).在植物的冠层内部, 阴生叶比阳生叶的LMA低, 差异可达2-4倍, LMA降低主要与叶肉细胞层厚度减少, 叶片变薄, 叶片密度显著降低, 细胞内含物显著下降有关(Niinemets, 2007; Lusk et al., 2010; Chen et al., 2014).Meng等(2013, 2014)的研究发现, 遮阴不仅会导致叶片的LMA下降, 还会使叶片倾角变小, 以截获更多的光照.值得关注的是, 在很多胁迫条件下(比如干旱、低温), 叶片LMA的增加伴随细胞壁的增厚, 然而在遮阴条件下, 虽然LMA降低了, 但其细胞壁反而增厚, 这可能是植物在低光条件下为增强其叶片硬度和韧度, 避免食草动物的取食而采取的防御机制(Hanba et al., 2002; Coble & Cavaleri, 2015).此外, 很多学者在对热带常绿和落叶木本植物的研究中发现, 在同一低光环境中, 耐阴性强的植物与需光植物相比, 具有较低的光合能力和较高的LMA, LMA的增加在一定程度上与叶肉细胞组织孔隙度的下降、叶片表皮毛密度的增加有关(Hanba et al., 2002; Lusk et al., 2008).LMA的增加使叶片更加强韧, 可增强植物对食草动物和病原菌的防御能力, 提高其在弱光下的生存几率(Kitajima et al., 1994). ...
两种光照生境下4种常绿阔叶树的单位叶面积干重、光合能力与化学防御物质含量比较
1
2010
... 树木冠层内的LMA的差异也是森林生态系统研究的重要内容.LMA与叶片在冠层中的位置有关, 一般处于冠层顶部的叶片LMA值比冠层底部的高(Koch et al., 2004; Lichtenthaler et al., 2007; Poorter et al., 2009; Weerasinghe et al., 2014).据Koch等(2004)报道100 m高的红杉(Sequoia sempervirens)的冠层底部LMA为150 g·m-2, 而冠层顶部LMA值达800 g·m-2, 这是比较极端的例子.一般来说, 不管是木本植物还是草本植物, 其冠层LMA大约是底层LMA的两倍(Poorter et al., 2009).这种底层和冠层之间的差异在一定程度上是由物种的特异性决定的(Lichtenthaler et al., 2007).此外, LMA还随叶寿命的不同表现出较大的差异.叶寿命是植物在长期适应环境的过程中为获得最大光合生产所形成的适应策略(Reich et al., 1991; 张林和罗天祥, 2004).LMA反映了植物单位叶面积的构建成本, 通常与叶寿命呈正相关关系.LMA低的植物, 其光合效率高, 生长速度快, 环境胁迫应答策略表现为逃避, 因而用于防御投资的物质较少, 叶寿命较短, 如草本植物、落叶乔灌木; 而LMA高的植物, 光合效率慢, 生长速度慢, 环境胁迫应答的策略表现为抵御和耐受, 需要不断积累更多的物质构建保卫构造, 因此表现出较长的叶寿命, 如常绿木本植物(Reich et al., 1991; Wright et al., 2004; Wyka et al., 2012; 王瑞雪等, 2014).常绿木本植物的LMA通常随叶龄的增长而逐渐增加, LMA的增加主要与叶肉细胞的细胞壁变厚、木质素含量增加、叶片变厚、叶片的机械组织含量增加有关(Castro-Díez et al., 2000; Hanba et al., 2002; Wright et al., 2004).Niinemets等(2005, 2006)报道地中海3种阔叶常绿树种月桂(Laurus nobilis)、油橄榄(Olea europea)、冬青栎(Quercus ilex)的LMA随叶龄的增长而增加.而且叶龄对LMA的影响还与叶片在冠层中的位置有关, Han等(2008)报道赤松冠层底部LMA随叶龄的增加而增加的幅度为27%, 但叶龄对冠层顶部LMA无影响.此外, 在常绿木本植物中, 叶寿命不同的植物, 其LMA也有较大的差异, Wyka等(2012)报道松柏类比阔叶类植物的叶寿命长, 其LMA也显著高于阔叶类植物. ...
Why are evergreen leaves so contrary about shade?
1
2008
... (1)植物LMA并不是完全由遗传或者环境来决定的, 而是由内在的遗传基础和外在的环境共同作用形成的.因此, LMA的大小在很大程度上由植物的内在遗传基础决定.不同功能群、种间、科属的植物LMA存在较大的差异, 而通过在叶片水平、组织解剖水平、细胞水平的比较研究能进一步解释造成不同科属或功能群的植物LMA差异的原因.目前已经针对不同科属的植物做了很多组织解剖学方面的比较研究, 但在细胞水平的比较研究还很有限, 主要由于实验技术方面的原因, 难以对不同组织的细胞大小和数量的改变做定量比较研究.传统的组织切片只能反映细胞某个横截面的面积, 难以真实反映细胞的体积、形状、数量, 而采用3D成像技术与计算机空间模型(Xu et al., 2012; Kubínová et al., 2014)的结合能展现叶片组织、细胞的立体结构, 这可能成为未来定量研究细胞和细胞器的重要手段. ...
Reconciling species-level vs plastic responses of evergreen leaf structure to light gradients: Shade leaves punch above their weight.
1
2010
... 从目前的研究结果来看, 木本植物比草本植物的LMA高, 在同一环境控制下, 两者之间的差异大约为35%; 在野外环境下, 两者之间的差异约83% (Poorter et al., 2009).LMA在叶片水平可分解为叶片密度和厚度.根据目前的研究文献报道, 密度和厚度都有可能是决定木本植物LMA高低的原因(Castro-Díez et al., 2000; Wright & Cannon, 2001; Villar et al., 2013).LMA高的木本植物, 其硬化组织占整个叶片的比例高, 细胞小而紧凑、叶片含水量和氮含量较低(Wright & Cannon, 2001).常绿木本植物普遍比落叶木本植物的LMA高(Castro-Díez et al., 2000; Villar & Merino, 2001; Wright et al., 2005; Li et al., 2013; Villar et al., 2013).Castro-Díez (2000)和Villar等(2013)认为常绿和落叶木本植物的叶片密度之间没有显著的差异, 常绿木本植物较高的LMA主要与叶片厚度有关, 同科属中的常绿木本植物的叶片通常比落叶木本植物的叶片厚, 而叶肉组织增厚是导致叶片变厚的主要原因.然而, 不同科属的常绿和落叶植物之间的LMA也存在交差和重叠, 这主要是由植物的系统发育决定的; 也就是不同科属之间植物的LMA是有差异的, 而在同一科属中, 常绿植物比落叶植物的LMA高(Poorter et al., 2009; Villar et al., 2013). ...
Within- twig leaf distribution patterns differ among plantlife- forms in a subtropical Chinese forest.
2
2013
... 树木冠层内的LMA的差异也是森林生态系统研究的重要内容.LMA与叶片在冠层中的位置有关, 一般处于冠层顶部的叶片LMA值比冠层底部的高(Koch et al., 2004; Lichtenthaler et al., 2007; Poorter et al., 2009; Weerasinghe et al., 2014).据Koch等(2004)报道100 m高的红杉(Sequoia sempervirens)的冠层底部LMA为150 g·m-2, 而冠层顶部LMA值达800 g·m-2, 这是比较极端的例子.一般来说, 不管是木本植物还是草本植物, 其冠层LMA大约是底层LMA的两倍(Poorter et al., 2009).这种底层和冠层之间的差异在一定程度上是由物种的特异性决定的(Lichtenthaler et al., 2007).此外, LMA还随叶寿命的不同表现出较大的差异.叶寿命是植物在长期适应环境的过程中为获得最大光合生产所形成的适应策略(Reich et al., 1991; 张林和罗天祥, 2004).LMA反映了植物单位叶面积的构建成本, 通常与叶寿命呈正相关关系.LMA低的植物, 其光合效率高, 生长速度快, 环境胁迫应答策略表现为逃避, 因而用于防御投资的物质较少, 叶寿命较短, 如草本植物、落叶乔灌木; 而LMA高的植物, 光合效率慢, 生长速度慢, 环境胁迫应答的策略表现为抵御和耐受, 需要不断积累更多的物质构建保卫构造, 因此表现出较长的叶寿命, 如常绿木本植物(Reich et al., 1991; Wright et al., 2004; Wyka et al., 2012; 王瑞雪等, 2014).常绿木本植物的LMA通常随叶龄的增长而逐渐增加, LMA的增加主要与叶肉细胞的细胞壁变厚、木质素含量增加、叶片变厚、叶片的机械组织含量增加有关(Castro-Díez et al., 2000; Hanba et al., 2002; Wright et al., 2004).Niinemets等(2005, 2006)报道地中海3种阔叶常绿树种月桂(Laurus nobilis)、油橄榄(Olea europea)、冬青栎(Quercus ilex)的LMA随叶龄的增长而增加.而且叶龄对LMA的影响还与叶片在冠层中的位置有关, Han等(2008)报道赤松冠层底部LMA随叶龄的增加而增加的幅度为27%, 但叶龄对冠层顶部LMA无影响.此外, 在常绿木本植物中, 叶寿命不同的植物, 其LMA也有较大的差异, Wyka等(2012)报道松柏类比阔叶类植物的叶寿命长, 其LMA也显著高于阔叶类植物. ...
... ).这种底层和冠层之间的差异在一定程度上是由物种的特异性决定的(Lichtenthaler et al., 2007).此外, LMA还随叶寿命的不同表现出较大的差异.叶寿命是植物在长期适应环境的过程中为获得最大光合生产所形成的适应策略(Reich et al., 1991; 张林和罗天祥, 2004).LMA反映了植物单位叶面积的构建成本, 通常与叶寿命呈正相关关系.LMA低的植物, 其光合效率高, 生长速度快, 环境胁迫应答策略表现为逃避, 因而用于防御投资的物质较少, 叶寿命较短, 如草本植物、落叶乔灌木; 而LMA高的植物, 光合效率慢, 生长速度慢, 环境胁迫应答的策略表现为抵御和耐受, 需要不断积累更多的物质构建保卫构造, 因此表现出较长的叶寿命, 如常绿木本植物(Reich et al., 1991; Wright et al., 2004; Wyka et al., 2012; 王瑞雪等, 2014).常绿木本植物的LMA通常随叶龄的增长而逐渐增加, LMA的增加主要与叶肉细胞的细胞壁变厚、木质素含量增加、叶片变厚、叶片的机械组织含量增加有关(Castro-Díez et al., 2000; Hanba et al., 2002; Wright et al., 2004).Niinemets等(2005, 2006)报道地中海3种阔叶常绿树种月桂(Laurus nobilis)、油橄榄(Olea europea)、冬青栎(Quercus ilex)的LMA随叶龄的增长而增加.而且叶龄对LMA的影响还与叶片在冠层中的位置有关, Han等(2008)报道赤松冠层底部LMA随叶龄的增加而增加的幅度为27%, 但叶龄对冠层顶部LMA无影响.此外, 在常绿木本植物中, 叶寿命不同的植物, 其LMA也有较大的差异, Wyka等(2012)报道松柏类比阔叶类植物的叶寿命长, 其LMA也显著高于阔叶类植物. ...
Trade-offs between light interception and leaf water shedding: Acomparison of shade-and sun-adapted species in a subtropical rainforest.
2014
a). A functional comparison of acclimation to shade and submergence in two terrestrial plant species.
1
2005
... 关于植物与光照的关系, 目前研究的比较多的是森林生态系统中光照不足对植物产生的影响.在森林生态系统中, 对光的需求的差异是决定植物生长和生存的主要因素(Hanba et al., 2002).大多研究表明植物在适应光环境的过程中主要通过调节叶片形态和结构适应光照的变化, 主要表现在LMA的增大或减小(Niinemets, 2007; Lusk et al., 2008; Coble & Cavaleri, 2015).目前的研究表明, 对于同一种植物, 遮阴导致叶片LMA、厚度、密度下降, 而单位干质量的叶面积增加, 增大了其叶片捕光面积, 利于在弱光环境下生存(冯玉龙等, 2002; Lusk et al., 2008; 柳凤娟等, 2010; 王博轶等, 2012; Coble & Cavaleri, 2015).在植物的冠层内部, 阴生叶比阳生叶的LMA低, 差异可达2-4倍, LMA降低主要与叶肉细胞层厚度减少, 叶片变薄, 叶片密度显著降低, 细胞内含物显著下降有关(Niinemets, 2007; Lusk et al., 2010; Chen et al., 2014).Meng等(2013, 2014)的研究发现, 遮阴不仅会导致叶片的LMA下降, 还会使叶片倾角变小, 以截获更多的光照.值得关注的是, 在很多胁迫条件下(比如干旱、低温), 叶片LMA的增加伴随细胞壁的增厚, 然而在遮阴条件下, 虽然LMA降低了, 但其细胞壁反而增厚, 这可能是植物在低光条件下为增强其叶片硬度和韧度, 避免食草动物的取食而采取的防御机制(Hanba et al., 2002; Coble & Cavaleri, 2015).此外, 很多学者在对热带常绿和落叶木本植物的研究中发现, 在同一低光环境中, 耐阴性强的植物与需光植物相比, 具有较低的光合能力和较高的LMA, LMA的增加在一定程度上与叶肉细胞组织孔隙度的下降、叶片表皮毛密度的增加有关(Hanba et al., 2002; Lusk et al., 2008).LMA的增加使叶片更加强韧, 可增强植物对食草动物和病原菌的防御能力, 提高其在弱光下的生存几率(Kitajima et al., 1994). ...
b). Submergence-induced morphological, anatomical, and biochemical responses in a terrestrial species affect gas diffusion resistance and photosynthetic performance.
3
2005
... 关于植物与光照的关系, 目前研究的比较多的是森林生态系统中光照不足对植物产生的影响.在森林生态系统中, 对光的需求的差异是决定植物生长和生存的主要因素(Hanba et al., 2002).大多研究表明植物在适应光环境的过程中主要通过调节叶片形态和结构适应光照的变化, 主要表现在LMA的增大或减小(Niinemets, 2007; Lusk et al., 2008; Coble & Cavaleri, 2015).目前的研究表明, 对于同一种植物, 遮阴导致叶片LMA、厚度、密度下降, 而单位干质量的叶面积增加, 增大了其叶片捕光面积, 利于在弱光环境下生存(冯玉龙等, 2002; Lusk et al., 2008; 柳凤娟等, 2010; 王博轶等, 2012; Coble & Cavaleri, 2015).在植物的冠层内部, 阴生叶比阳生叶的LMA低, 差异可达2-4倍, LMA降低主要与叶肉细胞层厚度减少, 叶片变薄, 叶片密度显著降低, 细胞内含物显著下降有关(Niinemets, 2007; Lusk et al., 2010; Chen et al., 2014).Meng等(2013, 2014)的研究发现, 遮阴不仅会导致叶片的LMA下降, 还会使叶片倾角变小, 以截获更多的光照.值得关注的是, 在很多胁迫条件下(比如干旱、低温), 叶片LMA的增加伴随细胞壁的增厚, 然而在遮阴条件下, 虽然LMA降低了, 但其细胞壁反而增厚, 这可能是植物在低光条件下为增强其叶片硬度和韧度, 避免食草动物的取食而采取的防御机制(Hanba et al., 2002; Coble & Cavaleri, 2015).此外, 很多学者在对热带常绿和落叶木本植物的研究中发现, 在同一低光环境中, 耐阴性强的植物与需光植物相比, 具有较低的光合能力和较高的LMA, LMA的增加在一定程度上与叶肉细胞组织孔隙度的下降、叶片表皮毛密度的增加有关(Hanba et al., 2002; Lusk et al., 2008).LMA的增加使叶片更加强韧, 可增强植物对食草动物和病原菌的防御能力, 提高其在弱光下的生存几率(Kitajima et al., 1994). ...
... ; Lusk et al., 2008; 柳凤娟等, 2010; 王博轶等, 2012; Coble & Cavaleri, 2015).在植物的冠层内部, 阴生叶比阳生叶的LMA低, 差异可达2-4倍, LMA降低主要与叶肉细胞层厚度减少, 叶片变薄, 叶片密度显著降低, 细胞内含物显著下降有关(Niinemets, 2007; Lusk et al., 2010; Chen et al., 2014).Meng等(2013, 2014)的研究发现, 遮阴不仅会导致叶片的LMA下降, 还会使叶片倾角变小, 以截获更多的光照.值得关注的是, 在很多胁迫条件下(比如干旱、低温), 叶片LMA的增加伴随细胞壁的增厚, 然而在遮阴条件下, 虽然LMA降低了, 但其细胞壁反而增厚, 这可能是植物在低光条件下为增强其叶片硬度和韧度, 避免食草动物的取食而采取的防御机制(Hanba et al., 2002; Coble & Cavaleri, 2015).此外, 很多学者在对热带常绿和落叶木本植物的研究中发现, 在同一低光环境中, 耐阴性强的植物与需光植物相比, 具有较低的光合能力和较高的LMA, LMA的增加在一定程度上与叶肉细胞组织孔隙度的下降、叶片表皮毛密度的增加有关(Hanba et al., 2002; Lusk et al., 2008).LMA的增加使叶片更加强韧, 可增强植物对食草动物和病原菌的防御能力, 提高其在弱光下的生存几率(Kitajima et al., 1994). ...
... ; Lusk et al., 2008).LMA的增加使叶片更加强韧, 可增强植物对食草动物和病原菌的防御能力, 提高其在弱光下的生存几率(Kitajima et al., 1994). ...
Submergence-induced leaf acclimation in terrestrial species varying in flooding tolerance.
1
2007
... 关于植物与光照的关系, 目前研究的比较多的是森林生态系统中光照不足对植物产生的影响.在森林生态系统中, 对光的需求的差异是决定植物生长和生存的主要因素(Hanba et al., 2002).大多研究表明植物在适应光环境的过程中主要通过调节叶片形态和结构适应光照的变化, 主要表现在LMA的增大或减小(Niinemets, 2007; Lusk et al., 2008; Coble & Cavaleri, 2015).目前的研究表明, 对于同一种植物, 遮阴导致叶片LMA、厚度、密度下降, 而单位干质量的叶面积增加, 增大了其叶片捕光面积, 利于在弱光环境下生存(冯玉龙等, 2002; Lusk et al., 2008; 柳凤娟等, 2010; 王博轶等, 2012; Coble & Cavaleri, 2015).在植物的冠层内部, 阴生叶比阳生叶的LMA低, 差异可达2-4倍, LMA降低主要与叶肉细胞层厚度减少, 叶片变薄, 叶片密度显著降低, 细胞内含物显著下降有关(Niinemets, 2007; Lusk et al., 2010; Chen et al., 2014).Meng等(2013, 2014)的研究发现, 遮阴不仅会导致叶片的LMA下降, 还会使叶片倾角变小, 以截获更多的光照.值得关注的是, 在很多胁迫条件下(比如干旱、低温), 叶片LMA的增加伴随细胞壁的增厚, 然而在遮阴条件下, 虽然LMA降低了, 但其细胞壁反而增厚, 这可能是植物在低光条件下为增强其叶片硬度和韧度, 避免食草动物的取食而采取的防御机制(Hanba et al., 2002; Coble & Cavaleri, 2015).此外, 很多学者在对热带常绿和落叶木本植物的研究中发现, 在同一低光环境中, 耐阴性强的植物与需光植物相比, 具有较低的光合能力和较高的LMA, LMA的增加在一定程度上与叶肉细胞组织孔隙度的下降、叶片表皮毛密度的增加有关(Hanba et al., 2002; Lusk et al., 2008).LMA的增加使叶片更加强韧, 可增强植物对食草动物和病原菌的防御能力, 提高其在弱光下的生存几率(Kitajima et al., 1994). ...
Morphological and anatomical determinants of mesophyll conductance in wild relatives of tomato (Solanum sect. Lycopersicon, sect. Lycopersicoides; Solanaceae). Plant,
2014
Epidermal focusing and effects on light utilization in Oxalis acetosella.
1994
Components of leaf dry mass per area- thickness and density-alter leaf photosynthetic capacity in reverse directions in woody plants.
1999
An analysis of light effects on foliar morphology, physiology, and light interception in temperate deciduous woody species of contrasting shade tolerance.
1998
Global-scale climatic controls of leaf dry mass per area, density, and thickness in trees and shrubs.
Within canopy variation in the rate of development of photosynthetic capacity is proportional to integrated quantum flux density in temperate deciduous trees. Plant,
1
2004
... 此外, LMA高的植物叶片通常具有一些特征, 如: 蜡质层增厚、表皮细胞增厚、表皮的毛状体、气孔隐窝以及一些特殊的细胞内含物, 如石细胞, 这些特征都有可能改变光在叶片内的传导途径(Myers et al., 1994; Baldini et al., 1997), 从而影响光合作用. ...
Leaf internal diffusion conductance limits photosynthesis more strongly in older leaves of Mediterranean evergreen broad-leaved species. Plant,
2
2005
... 20世纪90年代, 很多学者对不同植物的LMA做了大量研究, 并探讨了LVA和LD与LMA之间的关系.研究表明在同一可控环境条件下, 草本植物(Garnier & Laurent, 1994; van Arendonk & Poorter, 1994)和木本植物(Castro-Diez et al., 2000)的LMA主要受LD的影响, Poorter等(2009)对这些数据作了统计和分析, 表明LMA差异的80%由LD决定, 20%由LVA决定.然而在复杂的自然环境条件下, LD和LVA与LMA之间的关系是多样化的(Choong et al., 1992; Niinemets, 1999; Wilson et al., 1999).由此可见, 仅仅把LMA分解为LVA和LD并不能很好地解释导致植物种类之间LMA差异的原因. ...
... 目前研究表明, 具有不同形态和生境的植物种间或种内的gm差异很大(Flexas et al., 2008; Warren, 2008; Niinemets et al., 2009a, 2011), 而且特定种类或基因型的植物, 其最大gm与其叶片的解剖学特征有很大的关系(Niinemets et al., 2009a; Tosens et al., 2012).叶片内的CO2的扩散路径主要由两种相组成, 一种是细胞间的气相, 另一种是细胞的液相(主要由水和脂类组成) (Niinemets & Reichstein, 2003; Evans et al., 2009).细胞间气相的变化对CO2的扩散能力影响不大, 因此对整个光合能力没有显著的影响(Parkhurst & Mott, 1990; Evans et al., 2009).细胞相主要由细胞壁、质膜、胞液、叶绿体被膜、基质组成, 其中细胞壁和叶绿体被膜对gm的限制最大(Terashima et al., 2011).据目前的研究估算, 细胞壁厚度(Tcw)对gm的影响占25%-50%, 两者呈负相关关系(Evans et al., 2009; Terashima et al., 2011; Tosens et al., 2012).LMA高的植物, 总是伴随细胞壁的增厚, gm下降, CO2的同化速率降低, 最终导致光合能力下降(Niinemets, 1999; Terashima et al., 2006; Niinemets et al., 2007; Hassiotou et al., 2009).关于这一观点, 目前已在很多硬叶木本植物中得到广泛的证实, 如澳大利亚49种山龙眼科(Proteaceae)的植物(Hassiotou et al., 2010), Abies alba和Abies pinsapo (Peguero-Pina et al., 2012), 冬青栎(Quercus ilex)、柑橘(Citrus reticulate)、橡皮树(Ficus elastica)、海桐(Pittosporum tobira)、华盛顿棕榈(Washingtonia filifera) (Tomás et al., 2013).而且在冠层内, LMA随树木高度增加而增加的过程中所伴随的最大光合效率的下降的主要因素也是CO2的扩散阻力增加, gm下降(Han, 2011). ...
Complex adjustments of photosynthetic potentials and internal diffusion conductance to current and previous light availabilities and leaf age in Mediterranean evergreen species Quercus ilex. Plant,
1
2006
... 植物的LMA随光照强度的增加而增加, 但LMA由总的光照度/天(DPI)决定, 而不是瞬时的高的光照强度决定(Chabot et al., 1979; Niinemets et al., 2004).LMA随DPI的增加而显著增加, 但这种响应呈非线性相关, 因为在低光条件下LMA随DPI的增加而显著增加, 但是当DPI超过20 mol·m-2·d-1时, 其LMA的增加会变缓慢(Poorter et al., 2009).LMA 随光照度增加而增加的过程中, LD和LVA均扮演重要角色.Coble和Cavaleri (2014)的研究表明, 光照增加导致LMA的增加主要与LVA的增加有关.但在LVA增加的过程中, 并不是所有的叶片组织的体积都增加, 一般情况下, 栅栏组织的体积随光强度的增加而显著增加, 而表皮组织的体积保持不变(Hanson, 1917; Onoda et al., 2008).而高光条件下, LD的增加与叶脉密度的显著增加有关(Carins Murphy et al., 2012); 此外, 在一定程度上还与栅栏组织细胞排列的紧密度的增加及总的非结构性碳水化合物(TNC)浓度的增加有关(Niinemets et al., 1998; Poorter et al., 2009). ...
Structural determinants of leaf light-harvesting capacity and photosynthetic potentials.
1
2006
... LMA是一个复合的结构参数, 与叶片的很多解剖结构有关, 因而对光合作用有潜在的影响(Niinemets et al., 2009a; Scafaro et al., 2011; Terashima et al., 2011).近年来, LMA对光合作用的影响引起了生理生态学家的广泛关注.全球叶片经济型谱的建立已明确LMA与叶片单位干物质的最大光合能力、叶龄、单位干物质的氮、磷含量等密切相关(Wright et al., 2004).LMA低的植物, 其叶片生命周期短、单位干物质营养元素含量高, 光合能力强、单位叶面积的经济投入较低, 导致植物资源的利用效率高而生长速度快; LMA高的植物, 其单位叶面积的投入较高, 尤其增加了叶片的维管组织和细胞壁的投入(Niinemets et al., 2007; Hikosaka & Shigeno, 2009), 虽然增强了植物耐贫瘠、耐干旱的能力, 但导致较低的光合能力和缓慢的生长速率(Niinemets, 2001; Wright et al., 2004).近年的研究进一步表明, LMA与植物的单位叶面积的光合能力(Aarea)没有明显的相关性; 但随LMA的增加, 单位叶干质量的最大光合能力(Amass)有明显的下降趋势(Niinemets & Sack, 2006; Hassiotou et al., 2010). ...
Do we underestimate the importance of leaf size in plant economics? Disproportionate scaling of support costs within the spectrum of leaf physiognomy.
1
2007
... 目前研究表明, 具有不同形态和生境的植物种间或种内的gm差异很大(Flexas et al., 2008; Warren, 2008; Niinemets et al., 2009a, 2011), 而且特定种类或基因型的植物, 其最大gm与其叶片的解剖学特征有很大的关系(Niinemets et al., 2009a; Tosens et al., 2012).叶片内的CO2的扩散路径主要由两种相组成, 一种是细胞间的气相, 另一种是细胞的液相(主要由水和脂类组成) (Niinemets & Reichstein, 2003; Evans et al., 2009).细胞间气相的变化对CO2的扩散能力影响不大, 因此对整个光合能力没有显著的影响(Parkhurst & Mott, 1990; Evans et al., 2009).细胞相主要由细胞壁、质膜、胞液、叶绿体被膜、基质组成, 其中细胞壁和叶绿体被膜对gm的限制最大(Terashima et al., 2011).据目前的研究估算, 细胞壁厚度(Tcw)对gm的影响占25%-50%, 两者呈负相关关系(Evans et al., 2009; Terashima et al., 2011; Tosens et al., 2012).LMA高的植物, 总是伴随细胞壁的增厚, gm下降, CO2的同化速率降低, 最终导致光合能力下降(Niinemets, 1999; Terashima et al., 2006; Niinemets et al., 2007; Hassiotou et al., 2009).关于这一观点, 目前已在很多硬叶木本植物中得到广泛的证实, 如澳大利亚49种山龙眼科(Proteaceae)的植物(Hassiotou et al., 2010), Abies alba和Abies pinsapo (Peguero-Pina et al., 2012), 冬青栎(Quercus ilex)、柑橘(Citrus reticulate)、橡皮树(Ficus elastica)、海桐(Pittosporum tobira)、华盛顿棕榈(Washingtonia filifera) (Tomás et al., 2013).而且在冠层内, LMA随树木高度增加而增加的过程中所伴随的最大光合效率的下降的主要因素也是CO2的扩散阻力增加, gm下降(Han, 2011). ...
Photosynthesis and resource distribution through plant canopies. Plant,
1
2007
... 植物的LMA随光照强度的增加而增加, 但LMA由总的光照度/天(DPI)决定, 而不是瞬时的高的光照强度决定(Chabot et al., 1979; Niinemets et al., 2004).LMA随DPI的增加而显著增加, 但这种响应呈非线性相关, 因为在低光条件下LMA随DPI的增加而显著增加, 但是当DPI超过20 mol·m-2·d-1时, 其LMA的增加会变缓慢(Poorter et al., 2009).LMA 随光照度增加而增加的过程中, LD和LVA均扮演重要角色.Coble和Cavaleri (2014)的研究表明, 光照增加导致LMA的增加主要与LVA的增加有关.但在LVA增加的过程中, 并不是所有的叶片组织的体积都增加, 一般情况下, 栅栏组织的体积随光强度的增加而显著增加, 而表皮组织的体积保持不变(Hanson, 1917; Onoda et al., 2008).而高光条件下, LD的增加与叶脉密度的显著增加有关(Carins Murphy et al., 2012); 此外, 在一定程度上还与栅栏组织细胞排列的紧密度的增加及总的非结构性碳水化合物(TNC)浓度的增加有关(Niinemets et al., 1998; Poorter et al., 2009). ...
a). Leaf mesophyll diffusion conductance in 35 Australian sclerophylls covering a broad range of foliage structural and physiological variation.
5
2009
... LMA是一个复合的结构参数, 与叶片的很多解剖结构有关, 因而对光合作用有潜在的影响(Niinemets et al., 2009a; Scafaro et al., 2011; Terashima et al., 2011).近年来, LMA对光合作用的影响引起了生理生态学家的广泛关注.全球叶片经济型谱的建立已明确LMA与叶片单位干物质的最大光合能力、叶龄、单位干物质的氮、磷含量等密切相关(Wright et al., 2004).LMA低的植物, 其叶片生命周期短、单位干物质营养元素含量高, 光合能力强、单位叶面积的经济投入较低, 导致植物资源的利用效率高而生长速度快; LMA高的植物, 其单位叶面积的投入较高, 尤其增加了叶片的维管组织和细胞壁的投入(Niinemets et al., 2007; Hikosaka & Shigeno, 2009), 虽然增强了植物耐贫瘠、耐干旱的能力, 但导致较低的光合能力和缓慢的生长速率(Niinemets, 2001; Wright et al., 2004).近年的研究进一步表明, LMA与植物的单位叶面积的光合能力(Aarea)没有明显的相关性; 但随LMA的增加, 单位叶干质量的最大光合能力(Amass)有明显的下降趋势(Niinemets & Sack, 2006; Hassiotou et al., 2010). ...
... 据目前的文献报道总结, 多数学者认为高LMA往往伴随较低的叶肉细胞导度(gm), 这是导致高LMA的植物光合作用下降的重要原因(Flexas et al., 2008; Hassiotou et al., 2009; Niinemets et al., 2009a).gm指CO2从气孔下腔到叶绿体羧化位点的扩散能力, 是限制植物光合能力的重要因素 (Hassiotou et al., 2009, 2010; Niinemets et al., 2009a).Flexas等(2008)根据发表文献整理了不同植物的LMA所对应的gm (图2), 结果表明高LMA往往导致较低的gm. ...
... ; Niinemets et al., 2009a).Flexas等(2008)根据发表文献整理了不同植物的LMA所对应的gm (图2), 结果表明高LMA往往导致较低的gm. ...
... 目前研究表明, 具有不同形态和生境的植物种间或种内的gm差异很大(Flexas et al., 2008; Warren, 2008; Niinemets et al., 2009a, 2011), 而且特定种类或基因型的植物, 其最大gm与其叶片的解剖学特征有很大的关系(Niinemets et al., 2009a; Tosens et al., 2012).叶片内的CO2的扩散路径主要由两种相组成, 一种是细胞间的气相, 另一种是细胞的液相(主要由水和脂类组成) (Niinemets & Reichstein, 2003; Evans et al., 2009).细胞间气相的变化对CO2的扩散能力影响不大, 因此对整个光合能力没有显著的影响(Parkhurst & Mott, 1990; Evans et al., 2009).细胞相主要由细胞壁、质膜、胞液、叶绿体被膜、基质组成, 其中细胞壁和叶绿体被膜对gm的限制最大(Terashima et al., 2011).据目前的研究估算, 细胞壁厚度(Tcw)对gm的影响占25%-50%, 两者呈负相关关系(Evans et al., 2009; Terashima et al., 2011; Tosens et al., 2012).LMA高的植物, 总是伴随细胞壁的增厚, gm下降, CO2的同化速率降低, 最终导致光合能力下降(Niinemets, 1999; Terashima et al., 2006; Niinemets et al., 2007; Hassiotou et al., 2009).关于这一观点, 目前已在很多硬叶木本植物中得到广泛的证实, 如澳大利亚49种山龙眼科(Proteaceae)的植物(Hassiotou et al., 2010), Abies alba和Abies pinsapo (Peguero-Pina et al., 2012), 冬青栎(Quercus ilex)、柑橘(Citrus reticulate)、橡皮树(Ficus elastica)、海桐(Pittosporum tobira)、华盛顿棕榈(Washingtonia filifera) (Tomás et al., 2013).而且在冠层内, LMA随树木高度增加而增加的过程中所伴随的最大光合效率的下降的主要因素也是CO2的扩散阻力增加, gm下降(Han, 2011). ...
... 与其叶片的解剖学特征有很大的关系(Niinemets et al., 2009a; Tosens et al., 2012).叶片内的CO2的扩散路径主要由两种相组成, 一种是细胞间的气相, 另一种是细胞的液相(主要由水和脂类组成) (Niinemets & Reichstein, 2003; Evans et al., 2009).细胞间气相的变化对CO2的扩散能力影响不大, 因此对整个光合能力没有显著的影响(Parkhurst & Mott, 1990; Evans et al., 2009).细胞相主要由细胞壁、质膜、胞液、叶绿体被膜、基质组成, 其中细胞壁和叶绿体被膜对gm的限制最大(Terashima et al., 2011).据目前的研究估算, 细胞壁厚度(Tcw)对gm的影响占25%-50%, 两者呈负相关关系(Evans et al., 2009; Terashima et al., 2011; Tosens et al., 2012).LMA高的植物, 总是伴随细胞壁的增厚, gm下降, CO2的同化速率降低, 最终导致光合能力下降(Niinemets, 1999; Terashima et al., 2006; Niinemets et al., 2007; Hassiotou et al., 2009).关于这一观点, 目前已在很多硬叶木本植物中得到广泛的证实, 如澳大利亚49种山龙眼科(Proteaceae)的植物(Hassiotou et al., 2010), Abies alba和Abies pinsapo (Peguero-Pina et al., 2012), 冬青栎(Quercus ilex)、柑橘(Citrus reticulate)、橡皮树(Ficus elastica)、海桐(Pittosporum tobira)、华盛顿棕榈(Washingtonia filifera) (Tomás et al., 2013).而且在冠层内, LMA随树木高度增加而增加的过程中所伴随的最大光合效率的下降的主要因素也是CO2的扩散阻力增加, gm下降(Han, 2011). ...
b). Role of mesophyll diffusion conductance in constraining potential photosynthetic productivity in the field.
2
2009
... 短时间的水分胁迫对植物的叶片解剖结构没有影响, 植物只有处于长期缺水状态才会使叶片的解剖结构发生实质性的改变(Poorter et al., 2009).目前, 大多数学者的研究表明耐旱性强的植物一般表现出较高的LMA, 这与叶片厚度和密度的增加有关(Niinemets et al., 2009b).龚春梅和王根轩(2010)通过研究干旱半干旱地区植物群落特征发现植物的LMA随干旱胁迫的加剧而增大.董廷发等(2012)的研究表明在植物群落水平上, 干旱生境中植物的LMA显著高于湿润生境.在长期水分胁迫条件下, 叶片的各组织和细胞结构的改变是导致LMA增加的直接原因.植物经过长期的水分胁迫, 叶片的厚度显著增加, 这与叶片上表皮组织、栅栏组织、海绵组织的增厚有关, 尤其是栅栏组织(Boughalleb & Hajlaoui, 2011; Galmés et al., 2013).此外, 叶片细胞的大小和结构也会发生适应性改变.水分胁迫下发育的叶片其扩展速率较低, 细胞因此更小, 更紧凑, 气室变小(Shield, 1950).Zhang等(2011)的研究发现水杉(Metasequoia glyptostroboides)幼苗叶片在长期水分胁迫下, 其叶肉细胞及上表皮细胞显著变小.细胞结构的改变主要表现为细胞壁增厚, 木质素含量增加(Sobrado, 1992; Henry et al., 2000).以上这些叶片组织和细胞的改变最终表现为叶片的LD及LMA逐渐增加(Nogués & Baker, 2000; Boughalleb & Hajlaoui, 2011).即使水分胁迫解除, 植物仍然会维持叶片解剖结构的适应性的改变; 而且经过干旱胁迫后的植物拥有更强韧的叶片及更高的LMA (Kalapos et al., 1996; Niinemets et al., 2009b; Boughalleb & Hajlaoui, 2011).这种适应性的改变最终对植物的生理过程尤其是光合作用产生较大的影响.很多研究表明植物遭受干旱胁迫后的光合速率会显著下降, 即使胁迫解除后也很难再恢复到胁迫前的值, 这与叶片解剖结构适应性的改变有关(Flexas et al., 2001; Gallé et al., 2009; Galmés et al., 2011).近年来的研究强调gm在限制植物光合速率从水分胁迫恢复中扮演重要角色; 而且大多研究认为叶片解剖学的适应性的改变是引起gm下降的重要原因, 如水分胁迫引起的LMA、LD增加、尤其是细胞壁增厚(Gallé et al., 2009; Boughalleb & Hajlaoui, 2011; Galmés et al., 2013). ...
... ; Niinemets et al., 2009b; Boughalleb & Hajlaoui, 2011).这种适应性的改变最终对植物的生理过程尤其是光合作用产生较大的影响.很多研究表明植物遭受干旱胁迫后的光合速率会显著下降, 即使胁迫解除后也很难再恢复到胁迫前的值, 这与叶片解剖结构适应性的改变有关(Flexas et al., 2001; Gallé et al., 2009; Galmés et al., 2011).近年来的研究强调gm在限制植物光合速率从水分胁迫恢复中扮演重要角色; 而且大多研究认为叶片解剖学的适应性的改变是引起gm下降的重要原因, 如水分胁迫引起的LMA、LD增加、尤其是细胞壁增厚(Gallé et al., 2009; Boughalleb & Hajlaoui, 2011; Galmés et al., 2013). ...
Evergreens favored by higher responsiveness to increased CO2.
1
2011
... LMA是一个复合的结构参数, 与叶片的很多解剖结构有关, 因而对光合作用有潜在的影响(Niinemets et al., 2009a; Scafaro et al., 2011; Terashima et al., 2011).近年来, LMA对光合作用的影响引起了生理生态学家的广泛关注.全球叶片经济型谱的建立已明确LMA与叶片单位干物质的最大光合能力、叶龄、单位干物质的氮、磷含量等密切相关(Wright et al., 2004).LMA低的植物, 其叶片生命周期短、单位干物质营养元素含量高, 光合能力强、单位叶面积的经济投入较低, 导致植物资源的利用效率高而生长速度快; LMA高的植物, 其单位叶面积的投入较高, 尤其增加了叶片的维管组织和细胞壁的投入(Niinemets et al., 2007; Hikosaka & Shigeno, 2009), 虽然增强了植物耐贫瘠、耐干旱的能力, 但导致较低的光合能力和缓慢的生长速率(Niinemets, 2001; Wright et al., 2004).近年的研究进一步表明, LMA与植物的单位叶面积的光合能力(Aarea)没有明显的相关性; 但随LMA的增加, 单位叶干质量的最大光合能力(Amass)有明显的下降趋势(Niinemets & Sack, 2006; Hassiotou et al., 2010). ...
Effects of drought on photo- synthesis in Mediterranean plants grown under enhanced UV-B radiation.
5
2000
... 自然界中的每一种植物都有自己独特的叶片形态和结构, 这种特异性并不是偶然的, 而是自然界选择进化的结果.每一种叶片的形态和组织结构都有与之相协调的生理过程紧密联系.因此, 从叶片形态和结构入手揭示植物的生态策略和生理过程是一种行之有效的研究手段.在植物的叶片解剖研究过程中, 大量研究资料显示比叶质量(LMA)是一个非常重要的反应植物功能的结构参数.LMA指叶片干质量与面积之间的比率(g·m-2), 是叶片众多解剖结构特征的综合体, 例如叶肉组织厚度、细胞壁厚度、细胞形状、细胞内含物等(Muir et al., 2014).在大量的叶片形态解剖结构参数中, LMA是最容易定量测定的参数之一, 它能定量测定植物在单位叶面积的投入成本, 因而被认为是反映植物生态策略的重要指标(Westoby et al., 2002; Wright et al., 2004).LMA的高低反映了植物对资源的利用效率以及对环境的应对策略.总的来讲, 低LMA的植物光合效率高, 单位叶面积的投入成本低, 显示出较快的资源流动速度及回报效率; 而高LMA的植物光合效率低, 单位叶面积的投入成本高, 资源的流动速度及回报效率低(Niinemets et al., 2007; Hikosaka & Shigeno, 2009). ...
... LMA是一个复合的结构参数, 与叶片的很多解剖结构有关, 因而对光合作用有潜在的影响(Niinemets et al., 2009a; Scafaro et al., 2011; Terashima et al., 2011).近年来, LMA对光合作用的影响引起了生理生态学家的广泛关注.全球叶片经济型谱的建立已明确LMA与叶片单位干物质的最大光合能力、叶龄、单位干物质的氮、磷含量等密切相关(Wright et al., 2004).LMA低的植物, 其叶片生命周期短、单位干物质营养元素含量高, 光合能力强、单位叶面积的经济投入较低, 导致植物资源的利用效率高而生长速度快; LMA高的植物, 其单位叶面积的投入较高, 尤其增加了叶片的维管组织和细胞壁的投入(Niinemets et al., 2007; Hikosaka & Shigeno, 2009), 虽然增强了植物耐贫瘠、耐干旱的能力, 但导致较低的光合能力和缓慢的生长速率(Niinemets, 2001; Wright et al., 2004).近年的研究进一步表明, LMA与植物的单位叶面积的光合能力(Aarea)没有明显的相关性; 但随LMA的增加, 单位叶干质量的最大光合能力(Amass)有明显的下降趋势(Niinemets & Sack, 2006; Hassiotou et al., 2010). ...
... 目前研究表明, 具有不同形态和生境的植物种间或种内的gm差异很大(Flexas et al., 2008; Warren, 2008; Niinemets et al., 2009a, 2011), 而且特定种类或基因型的植物, 其最大gm与其叶片的解剖学特征有很大的关系(Niinemets et al., 2009a; Tosens et al., 2012).叶片内的CO2的扩散路径主要由两种相组成, 一种是细胞间的气相, 另一种是细胞的液相(主要由水和脂类组成) (Niinemets & Reichstein, 2003; Evans et al., 2009).细胞间气相的变化对CO2的扩散能力影响不大, 因此对整个光合能力没有显著的影响(Parkhurst & Mott, 1990; Evans et al., 2009).细胞相主要由细胞壁、质膜、胞液、叶绿体被膜、基质组成, 其中细胞壁和叶绿体被膜对gm的限制最大(Terashima et al., 2011).据目前的研究估算, 细胞壁厚度(Tcw)对gm的影响占25%-50%, 两者呈负相关关系(Evans et al., 2009; Terashima et al., 2011; Tosens et al., 2012).LMA高的植物, 总是伴随细胞壁的增厚, gm下降, CO2的同化速率降低, 最终导致光合能力下降(Niinemets, 1999; Terashima et al., 2006; Niinemets et al., 2007; Hassiotou et al., 2009).关于这一观点, 目前已在很多硬叶木本植物中得到广泛的证实, 如澳大利亚49种山龙眼科(Proteaceae)的植物(Hassiotou et al., 2010), Abies alba和Abies pinsapo (Peguero-Pina et al., 2012), 冬青栎(Quercus ilex)、柑橘(Citrus reticulate)、橡皮树(Ficus elastica)、海桐(Pittosporum tobira)、华盛顿棕榈(Washingtonia filifera) (Tomás et al., 2013).而且在冠层内, LMA随树木高度增加而增加的过程中所伴随的最大光合效率的下降的主要因素也是CO2的扩散阻力增加, gm下降(Han, 2011). ...
... 关于植物与光照的关系, 目前研究的比较多的是森林生态系统中光照不足对植物产生的影响.在森林生态系统中, 对光的需求的差异是决定植物生长和生存的主要因素(Hanba et al., 2002).大多研究表明植物在适应光环境的过程中主要通过调节叶片形态和结构适应光照的变化, 主要表现在LMA的增大或减小(Niinemets, 2007; Lusk et al., 2008; Coble & Cavaleri, 2015).目前的研究表明, 对于同一种植物, 遮阴导致叶片LMA、厚度、密度下降, 而单位干质量的叶面积增加, 增大了其叶片捕光面积, 利于在弱光环境下生存(冯玉龙等, 2002; Lusk et al., 2008; 柳凤娟等, 2010; 王博轶等, 2012; Coble & Cavaleri, 2015).在植物的冠层内部, 阴生叶比阳生叶的LMA低, 差异可达2-4倍, LMA降低主要与叶肉细胞层厚度减少, 叶片变薄, 叶片密度显著降低, 细胞内含物显著下降有关(Niinemets, 2007; Lusk et al., 2010; Chen et al., 2014).Meng等(2013, 2014)的研究发现, 遮阴不仅会导致叶片的LMA下降, 还会使叶片倾角变小, 以截获更多的光照.值得关注的是, 在很多胁迫条件下(比如干旱、低温), 叶片LMA的增加伴随细胞壁的增厚, 然而在遮阴条件下, 虽然LMA降低了, 但其细胞壁反而增厚, 这可能是植物在低光条件下为增强其叶片硬度和韧度, 避免食草动物的取食而采取的防御机制(Hanba et al., 2002; Coble & Cavaleri, 2015).此外, 很多学者在对热带常绿和落叶木本植物的研究中发现, 在同一低光环境中, 耐阴性强的植物与需光植物相比, 具有较低的光合能力和较高的LMA, LMA的增加在一定程度上与叶肉细胞组织孔隙度的下降、叶片表皮毛密度的增加有关(Hanba et al., 2002; Lusk et al., 2008).LMA的增加使叶片更加强韧, 可增强植物对食草动物和病原菌的防御能力, 提高其在弱光下的生存几率(Kitajima et al., 1994). ...
... 降低主要与叶肉细胞层厚度减少, 叶片变薄, 叶片密度显著降低, 细胞内含物显著下降有关(Niinemets, 2007; Lusk et al., 2010; Chen et al., 2014).Meng等(2013, 2014)的研究发现, 遮阴不仅会导致叶片的LMA下降, 还会使叶片倾角变小, 以截获更多的光照.值得关注的是, 在很多胁迫条件下(比如干旱、低温), 叶片LMA的增加伴随细胞壁的增厚, 然而在遮阴条件下, 虽然LMA降低了, 但其细胞壁反而增厚, 这可能是植物在低光条件下为增强其叶片硬度和韧度, 避免食草动物的取食而采取的防御机制(Hanba et al., 2002; Coble & Cavaleri, 2015).此外, 很多学者在对热带常绿和落叶木本植物的研究中发现, 在同一低光环境中, 耐阴性强的植物与需光植物相比, 具有较低的光合能力和较高的LMA, LMA的增加在一定程度上与叶肉细胞组织孔隙度的下降、叶片表皮毛密度的增加有关(Hanba et al., 2002; Lusk et al., 2008).LMA的增加使叶片更加强韧, 可增强植物对食草动物和病原菌的防御能力, 提高其在弱光下的生存几率(Kitajima et al., 1994). ...
Effects of light and nutrient availability on leaf mechanical properties of Plantago major: A conceptual approach.
2008
Intercellular diffusion limits to CO2 uptake in leaves: Studies in air and helox.
1990
Leaf anatomical properties in relation to differences in mesophyll conductance to CO2 and photosynthesis in two related Mediterranean Abies species. Plant,
2012
Regulation of photosynthesis and stomatal and mesophyll conductance under water stress and recovery in olive trees: Correlation with gene expression of carbonic anhydrase and aquaporins.
1
2014
... 目前研究表明, 具有不同形态和生境的植物种间或种内的gm差异很大(Flexas et al., 2008; Warren, 2008; Niinemets et al., 2009a, 2011), 而且特定种类或基因型的植物, 其最大gm与其叶片的解剖学特征有很大的关系(Niinemets et al., 2009a; Tosens et al., 2012).叶片内的CO2的扩散路径主要由两种相组成, 一种是细胞间的气相, 另一种是细胞的液相(主要由水和脂类组成) (Niinemets & Reichstein, 2003; Evans et al., 2009).细胞间气相的变化对CO2的扩散能力影响不大, 因此对整个光合能力没有显著的影响(Parkhurst & Mott, 1990; Evans et al., 2009).细胞相主要由细胞壁、质膜、胞液、叶绿体被膜、基质组成, 其中细胞壁和叶绿体被膜对gm的限制最大(Terashima et al., 2011).据目前的研究估算, 细胞壁厚度(Tcw)对gm的影响占25%-50%, 两者呈负相关关系(Evans et al., 2009; Terashima et al., 2011; Tosens et al., 2012).LMA高的植物, 总是伴随细胞壁的增厚, gm下降, CO2的同化速率降低, 最终导致光合能力下降(Niinemets, 1999; Terashima et al., 2006; Niinemets et al., 2007; Hassiotou et al., 2009).关于这一观点, 目前已在很多硬叶木本植物中得到广泛的证实, 如澳大利亚49种山龙眼科(Proteaceae)的植物(Hassiotou et al., 2010), Abies alba和Abies pinsapo (Peguero-Pina et al., 2012), 冬青栎(Quercus ilex)、柑橘(Citrus reticulate)、橡皮树(Ficus elastica)、海桐(Pittosporum tobira)、华盛顿棕榈(Washingtonia filifera) (Tomás et al., 2013).而且在冠层内, LMA随树木高度增加而增加的过程中所伴随的最大光合效率的下降的主要因素也是CO2的扩散阻力增加, gm下降(Han, 2011). ...
Variations in nitrogen use efficiency reflect the biochemical subtype while variations in water use efficiency reflect the evolutionary lineage of C4 grasses at inter-glacial CO2. Plant,
1
2015
... 短时间的水分胁迫对植物的叶片解剖结构没有影响, 植物只有处于长期缺水状态才会使叶片的解剖结构发生实质性的改变(Poorter et al., 2009).目前, 大多数学者的研究表明耐旱性强的植物一般表现出较高的LMA, 这与叶片厚度和密度的增加有关(Niinemets et al., 2009b).龚春梅和王根轩(2010)通过研究干旱半干旱地区植物群落特征发现植物的LMA随干旱胁迫的加剧而增大.董廷发等(2012)的研究表明在植物群落水平上, 干旱生境中植物的LMA显著高于湿润生境.在长期水分胁迫条件下, 叶片的各组织和细胞结构的改变是导致LMA增加的直接原因.植物经过长期的水分胁迫, 叶片的厚度显著增加, 这与叶片上表皮组织、栅栏组织、海绵组织的增厚有关, 尤其是栅栏组织(Boughalleb & Hajlaoui, 2011; Galmés et al., 2013).此外, 叶片细胞的大小和结构也会发生适应性改变.水分胁迫下发育的叶片其扩展速率较低, 细胞因此更小, 更紧凑, 气室变小(Shield, 1950).Zhang等(2011)的研究发现水杉(Metasequoia glyptostroboides)幼苗叶片在长期水分胁迫下, 其叶肉细胞及上表皮细胞显著变小.细胞结构的改变主要表现为细胞壁增厚, 木质素含量增加(Sobrado, 1992; Henry et al., 2000).以上这些叶片组织和细胞的改变最终表现为叶片的LD及LMA逐渐增加(Nogués & Baker, 2000; Boughalleb & Hajlaoui, 2011).即使水分胁迫解除, 植物仍然会维持叶片解剖结构的适应性的改变; 而且经过干旱胁迫后的植物拥有更强韧的叶片及更高的LMA (Kalapos et al., 1996; Niinemets et al., 2009b; Boughalleb & Hajlaoui, 2011).这种适应性的改变最终对植物的生理过程尤其是光合作用产生较大的影响.很多研究表明植物遭受干旱胁迫后的光合速率会显著下降, 即使胁迫解除后也很难再恢复到胁迫前的值, 这与叶片解剖结构适应性的改变有关(Flexas et al., 2001; Gallé et al., 2009; Galmés et al., 2011).近年来的研究强调gm在限制植物光合速率从水分胁迫恢复中扮演重要角色; 而且大多研究认为叶片解剖学的适应性的改变是引起gm下降的重要原因, 如水分胁迫引起的LMA、LD增加、尤其是细胞壁增厚(Gallé et al., 2009; Boughalleb & Hajlaoui, 2011; Galmés et al., 2013). ...
Leaf traits are good predictors of plant performance across 53 rain forest species.
1
2006
... 植物的LMA随光照强度的增加而增加, 但LMA由总的光照度/天(DPI)决定, 而不是瞬时的高的光照强度决定(Chabot et al., 1979; Niinemets et al., 2004).LMA随DPI的增加而显著增加, 但这种响应呈非线性相关, 因为在低光条件下LMA随DPI的增加而显著增加, 但是当DPI超过20 mol·m-2·d-1时, 其LMA的增加会变缓慢(Poorter et al., 2009).LMA 随光照度增加而增加的过程中, LD和LVA均扮演重要角色.Coble和Cavaleri (2014)的研究表明, 光照增加导致LMA的增加主要与LVA的增加有关.但在LVA增加的过程中, 并不是所有的叶片组织的体积都增加, 一般情况下, 栅栏组织的体积随光强度的增加而显著增加, 而表皮组织的体积保持不变(Hanson, 1917; Onoda et al., 2008).而高光条件下, LD的增加与叶脉密度的显著增加有关(Carins Murphy et al., 2012); 此外, 在一定程度上还与栅栏组织细胞排列的紧密度的增加及总的非结构性碳水化合物(TNC)浓度的增加有关(Niinemets et al., 1998; Poorter et al., 2009). ...
Causes and consequences of variation in leaf mass per area (LMA): A meta-analysis.
1
2009
... 目前研究表明, 具有不同形态和生境的植物种间或种内的gm差异很大(Flexas et al., 2008; Warren, 2008; Niinemets et al., 2009a, 2011), 而且特定种类或基因型的植物, 其最大gm与其叶片的解剖学特征有很大的关系(Niinemets et al., 2009a; Tosens et al., 2012).叶片内的CO2的扩散路径主要由两种相组成, 一种是细胞间的气相, 另一种是细胞的液相(主要由水和脂类组成) (Niinemets & Reichstein, 2003; Evans et al., 2009).细胞间气相的变化对CO2的扩散能力影响不大, 因此对整个光合能力没有显著的影响(Parkhurst & Mott, 1990; Evans et al., 2009).细胞相主要由细胞壁、质膜、胞液、叶绿体被膜、基质组成, 其中细胞壁和叶绿体被膜对gm的限制最大(Terashima et al., 2011).据目前的研究估算, 细胞壁厚度(Tcw)对gm的影响占25%-50%, 两者呈负相关关系(Evans et al., 2009; Terashima et al., 2011; Tosens et al., 2012).LMA高的植物, 总是伴随细胞壁的增厚, gm下降, CO2的同化速率降低, 最终导致光合能力下降(Niinemets, 1999; Terashima et al., 2006; Niinemets et al., 2007; Hassiotou et al., 2009).关于这一观点, 目前已在很多硬叶木本植物中得到广泛的证实, 如澳大利亚49种山龙眼科(Proteaceae)的植物(Hassiotou et al., 2010), Abies alba和Abies pinsapo (Peguero-Pina et al., 2012), 冬青栎(Quercus ilex)、柑橘(Citrus reticulate)、橡皮树(Ficus elastica)、海桐(Pittosporum tobira)、华盛顿棕榈(Washingtonia filifera) (Tomás et al., 2013).而且在冠层内, LMA随树木高度增加而增加的过程中所伴随的最大光合效率的下降的主要因素也是CO2的扩散阻力增加, gm下降(Han, 2011). ...
Leaf structure and specific leaf mass: The alpine desert plants of the Eastern Pamirs, Tadjikistan.
1
1999
... 目前研究表明, 具有不同形态和生境的植物种间或种内的gm差异很大(Flexas et al., 2008; Warren, 2008; Niinemets et al., 2009a, 2011), 而且特定种类或基因型的植物, 其最大gm与其叶片的解剖学特征有很大的关系(Niinemets et al., 2009a; Tosens et al., 2012).叶片内的CO2的扩散路径主要由两种相组成, 一种是细胞间的气相, 另一种是细胞的液相(主要由水和脂类组成) (Niinemets & Reichstein, 2003; Evans et al., 2009).细胞间气相的变化对CO2的扩散能力影响不大, 因此对整个光合能力没有显著的影响(Parkhurst & Mott, 1990; Evans et al., 2009).细胞相主要由细胞壁、质膜、胞液、叶绿体被膜、基质组成, 其中细胞壁和叶绿体被膜对gm的限制最大(Terashima et al., 2011).据目前的研究估算, 细胞壁厚度(Tcw)对gm的影响占25%-50%, 两者呈负相关关系(Evans et al., 2009; Terashima et al., 2011; Tosens et al., 2012).LMA高的植物, 总是伴随细胞壁的增厚, gm下降, CO2的同化速率降低, 最终导致光合能力下降(Niinemets, 1999; Terashima et al., 2006; Niinemets et al., 2007; Hassiotou et al., 2009).关于这一观点, 目前已在很多硬叶木本植物中得到广泛的证实, 如澳大利亚49种山龙眼科(Proteaceae)的植物(Hassiotou et al., 2010), Abies alba和Abies pinsapo (Peguero-Pina et al., 2012), 冬青栎(Quercus ilex)、柑橘(Citrus reticulate)、橡皮树(Ficus elastica)、海桐(Pittosporum tobira)、华盛顿棕榈(Washingtonia filifera) (Tomás et al., 2013).而且在冠层内, LMA随树木高度增加而增加的过程中所伴随的最大光合效率的下降的主要因素也是CO2的扩散阻力增加, gm下降(Han, 2011). ...
Sources of variation in specific leaf area in wheat grown at high temperature.
1
1987
... (2)植物LMA反映了碳资源的获取能力和资源的利用方式, 与其生态位息息相关.当环境发生改变时, 植物通过调节LMA的大小来改变调整资源的获取方式和生存对策, 最终可能导致其生态位的改变, 由此对整个生态群落产生较大的影响(Garnier et al., 2004; Díaz et al., 2007).因此, 面对全球高温、干旱、水涝等极端气候的频繁出现, 研究逆境条件下植物的生理生化反应及生存策略对于预测气候变化对植物生态的系统的影响具有重要意义.目前, 关于逆境条件下植物LMA的研究相对较少, 逆境引起的LMA的改变最终导致生理反应的改变, 尤其对光合作用的影响最大.研究表明, gm是限制光合作用的一个重要因素, 而LMA的改变总伴随gm的改变.LMA高、叶片厚的植物, 其细胞壁导度是限制gm的主要因素; 而LMA低、叶片薄的植物, 其限制gm的主要因素各不相同, 其机制仍然不是很清楚.有学者提出, 水通道蛋白(aquaporins)和碳酸酐酶(carbonic anhydrases)可能对低LMA植物的gm具有重要调控作用(Tomás et al., 2013).关于两者对gm的调节机制已有一些初步的报道(Perez-Martin et al., 2014; Boudichevskaia et al., 2015), 但是还需要进一步完善研究以明确其调控机理.此外, 研究环境改变对植物LMA、gm、光合效率的影响及三者之间的相互的调控机制对于理解植物的进化适应性以及气候变化对植物的影响具有重要价值, 这不仅是目前研究的重点, 也是将来研究的重要内容. ...
Leaf lifespan as a determinant of leaf structure and function among 23 amazonian tree species.
1991
Intraspecific relationships among wood density, leaf structural traits and environment in four co-occurring species of Nothofagus in New Zealand.
... 不同种类的植物, 其LMA也不相同, LMA究竟由环境决定还是植物的系统发育决定的呢?Asner等(2011)研究2873株植物在不同地点、不同气候条件的LMA差异原因, 发现LMA主要由植物分类地位决定, 环境因素处于次要地位.当面对气候变化时, 植物种间LMA的适应性的改变也是有差异的, Aspinwall等(2013)认为这种差异主要由植物的遗传因素决定.Flores等(2014)通过建立5401种维管植物的LMA数据资料库对LMA的进化历史进行了大规模的数据分析, 实验结果表明, LMA中系统发育的信号虽然低, 但却非常保守, 也就是具有亲缘关系的物种, 其LMA也会更相似.此外LMA的保守性与植物的生长型显著相关, 也就是选择强度与表型的进化速率是一致的, 木本植物比草本植物在稳定化选择方面表现出更高的保守性, 因此木本植物的进化速率比草本植物低(Flores et al., 2014).Tsujii等(2016)通过对夏威夷群岛的铁心木(Metrosideros polymorpha) LMA的研究表明环境梯度导致的表型遗传变异是物种进化的主要驱动力.目前的研究均赋予了LMA遗传进化上的重要意义.由此可见, 植物的遗传基础是决定LMA的内在因素, 而环境是外在的推动力.据目前文献资料的报道分析, 5%-95%的陆生植物的LMA的分布范围为30 g·m-2至330 g·m-2, 不同物种的LMA相差约100倍(Poorter et al., 2009).不同功能群植物LMA总的趋势为水生植物<蕨类植物<草本植物<落叶灌木或乔木<常绿灌木、乔木或多浆植物(郑淑霞和上官周平, 2007; Poorter et al., 2009; Flores et al., 2014). ...
... ; Poorter et al., 2009; Flores et al., 2014). ...
... 从目前的研究结果来看, 木本植物比草本植物的LMA高, 在同一环境控制下, 两者之间的差异大约为35%; 在野外环境下, 两者之间的差异约83% (Poorter et al., 2009).LMA在叶片水平可分解为叶片密度和厚度.根据目前的研究文献报道, 密度和厚度都有可能是决定木本植物LMA高低的原因(Castro-Díez et al., 2000; Wright & Cannon, 2001; Villar et al., 2013).LMA高的木本植物, 其硬化组织占整个叶片的比例高, 细胞小而紧凑、叶片含水量和氮含量较低(Wright & Cannon, 2001).常绿木本植物普遍比落叶木本植物的LMA高(Castro-Díez et al., 2000; Villar & Merino, 2001; Wright et al., 2005; Li et al., 2013; Villar et al., 2013).Castro-Díez (2000)和Villar等(2013)认为常绿和落叶木本植物的叶片密度之间没有显著的差异, 常绿木本植物较高的LMA主要与叶片厚度有关, 同科属中的常绿木本植物的叶片通常比落叶木本植物的叶片厚, 而叶肉组织增厚是导致叶片变厚的主要原因.然而, 不同科属的常绿和落叶植物之间的LMA也存在交差和重叠, 这主要是由植物的系统发育决定的; 也就是不同科属之间植物的LMA是有差异的, 而在同一科属中, 常绿植物比落叶植物的LMA高(Poorter et al., 2009; Villar et al., 2013). ...
... 高(Poorter et al., 2009; Villar et al., 2013). ...
... 树木冠层内的LMA的差异也是森林生态系统研究的重要内容.LMA与叶片在冠层中的位置有关, 一般处于冠层顶部的叶片LMA值比冠层底部的高(Koch et al., 2004; Lichtenthaler et al., 2007; Poorter et al., 2009; Weerasinghe et al., 2014).据Koch等(2004)报道100 m高的红杉(Sequoia sempervirens)的冠层底部LMA为150 g·m-2, 而冠层顶部LMA值达800 g·m-2, 这是比较极端的例子.一般来说, 不管是木本植物还是草本植物, 其冠层LMA大约是底层LMA的两倍(Poorter et al., 2009).这种底层和冠层之间的差异在一定程度上是由物种的特异性决定的(Lichtenthaler et al., 2007).此外, LMA还随叶寿命的不同表现出较大的差异.叶寿命是植物在长期适应环境的过程中为获得最大光合生产所形成的适应策略(Reich et al., 1991; 张林和罗天祥, 2004).LMA反映了植物单位叶面积的构建成本, 通常与叶寿命呈正相关关系.LMA低的植物, 其光合效率高, 生长速度快, 环境胁迫应答策略表现为逃避, 因而用于防御投资的物质较少, 叶寿命较短, 如草本植物、落叶乔灌木; 而LMA高的植物, 光合效率慢, 生长速度慢, 环境胁迫应答的策略表现为抵御和耐受, 需要不断积累更多的物质构建保卫构造, 因此表现出较长的叶寿命, 如常绿木本植物(Reich et al., 1991; Wright et al., 2004; Wyka et al., 2012; 王瑞雪等, 2014).常绿木本植物的LMA通常随叶龄的增长而逐渐增加, LMA的增加主要与叶肉细胞的细胞壁变厚、木质素含量增加、叶片变厚、叶片的机械组织含量增加有关(Castro-Díez et al., 2000; Hanba et al., 2002; Wright et al., 2004).Niinemets等(2005, 2006)报道地中海3种阔叶常绿树种月桂(Laurus nobilis)、油橄榄(Olea europea)、冬青栎(Quercus ilex)的LMA随叶龄的增长而增加.而且叶龄对LMA的影响还与叶片在冠层中的位置有关, Han等(2008)报道赤松冠层底部LMA随叶龄的增加而增加的幅度为27%, 但叶龄对冠层顶部LMA无影响.此外, 在常绿木本植物中, 叶寿命不同的植物, 其LMA也有较大的差异, Wyka等(2012)报道松柏类比阔叶类植物的叶寿命长, 其LMA也显著高于阔叶类植物. ...
... 的两倍(Poorter et al., 2009).这种底层和冠层之间的差异在一定程度上是由物种的特异性决定的(Lichtenthaler et al., 2007).此外, LMA还随叶寿命的不同表现出较大的差异.叶寿命是植物在长期适应环境的过程中为获得最大光合生产所形成的适应策略(Reich et al., 1991; 张林和罗天祥, 2004).LMA反映了植物单位叶面积的构建成本, 通常与叶寿命呈正相关关系.LMA低的植物, 其光合效率高, 生长速度快, 环境胁迫应答策略表现为逃避, 因而用于防御投资的物质较少, 叶寿命较短, 如草本植物、落叶乔灌木; 而LMA高的植物, 光合效率慢, 生长速度慢, 环境胁迫应答的策略表现为抵御和耐受, 需要不断积累更多的物质构建保卫构造, 因此表现出较长的叶寿命, 如常绿木本植物(Reich et al., 1991; Wright et al., 2004; Wyka et al., 2012; 王瑞雪等, 2014).常绿木本植物的LMA通常随叶龄的增长而逐渐增加, LMA的增加主要与叶肉细胞的细胞壁变厚、木质素含量增加、叶片变厚、叶片的机械组织含量增加有关(Castro-Díez et al., 2000; Hanba et al., 2002; Wright et al., 2004).Niinemets等(2005, 2006)报道地中海3种阔叶常绿树种月桂(Laurus nobilis)、油橄榄(Olea europea)、冬青栎(Quercus ilex)的LMA随叶龄的增长而增加.而且叶龄对LMA的影响还与叶片在冠层中的位置有关, Han等(2008)报道赤松冠层底部LMA随叶龄的增加而增加的幅度为27%, 但叶龄对冠层顶部LMA无影响.此外, 在常绿木本植物中, 叶寿命不同的植物, 其LMA也有较大的差异, Wyka等(2012)报道松柏类比阔叶类植物的叶寿命长, 其LMA也显著高于阔叶类植物. ...
... 草本植物的LMA普遍比木本植物的低, 叶片生长速度快, 资源获取和流动的速度快, 进化程度比木本植物高(Flores et al., 2014).一般来说, 多年生草本植物比一年生草本植物的LMA高.但在双子叶草本植物中, 一年生与多年生植物之间的LMA差异不大; 而单子叶植物中, 禾本科多年生植物比一年生植物的LMA平均高60% (Poorter et al., 2009).而且同一片叶不同部位的LMA也不同, Niinemets等(2007)研究了大量的草本植物叶片, 表明叶片中脉部分的LMA比叶片其余部分的LMA高, 之间的差异约为6倍.然而, 这种差异可能是由物种的特异性决定的, 小麦叶片中脉的LMA与叶片其余部分的LMA差异不大(Rawson et al., 1987).而且大多数禾本科植物基部叶片的LMA比末端叶片的LMA高(Poorter et al., 2009).Pinto等(2015)的研究表明C4禾本科植物的LMA主要与其亚型有关.但目前关于草本植物的LMA的相关的研究还不是很多, 尤其是关于禾本科植物的研究. ...
... 高(Poorter et al., 2009).Pinto等(2015)的研究表明C4禾本科植物的LMA主要与其亚型有关.但目前关于草本植物的LMA的相关的研究还不是很多, 尤其是关于禾本科植物的研究. ...
... 植物的LMA随光照强度的增加而增加, 但LMA由总的光照度/天(DPI)决定, 而不是瞬时的高的光照强度决定(Chabot et al., 1979; Niinemets et al., 2004).LMA随DPI的增加而显著增加, 但这种响应呈非线性相关, 因为在低光条件下LMA随DPI的增加而显著增加, 但是当DPI超过20 mol·m-2·d-1时, 其LMA的增加会变缓慢(Poorter et al., 2009).LMA 随光照度增加而增加的过程中, LD和LVA均扮演重要角色.Coble和Cavaleri (2014)的研究表明, 光照增加导致LMA的增加主要与LVA的增加有关.但在LVA增加的过程中, 并不是所有的叶片组织的体积都增加, 一般情况下, 栅栏组织的体积随光强度的增加而显著增加, 而表皮组织的体积保持不变(Hanson, 1917; Onoda et al., 2008).而高光条件下, LD的增加与叶脉密度的显著增加有关(Carins Murphy et al., 2012); 此外, 在一定程度上还与栅栏组织细胞排列的紧密度的增加及总的非结构性碳水化合物(TNC)浓度的增加有关(Niinemets et al., 1998; Poorter et al., 2009). ...
... ; Poorter et al., 2009). ...
... LMA反映了植物在单位叶面积上的投入成本, 是叶片经济型谱中最基础的叶功能性状.植物LMA也是最易定量测定的复合结构参数, 与植物学的很多研究领域紧密相关.LMA在农艺学中可作为反映作物生产力的指标; 在生态学中是反映植物生态策略的重要指标; 在生理学上与植物的光合作用及抗逆生理生化反应密切相关.LMA在个体水平与植物的细胞结构及生理反应相关; 而在群落水平可以通过改变群落的初级生产力、凋落物的分解速率、碳氮的循环对生态系统功能产生重要的影响(Díaz et al., 2004; Poorter et al., 2009).因此, 无论从植物的个体还是群落水平研究LMA均具有重要意义.到目前为止, 关于植物LMA已在解剖结构、生理生态学方面展开了深入的研究, 但还有一些问题值得进一步关注和继续完善研究. ...
Temperature response of mesophyll conductance in cultivated and wild Oryza species with contrasting mesophyll cell wall thickness. Plant,
2
2011
... 目前植物解剖学的研究大多采用较薄的组织切片, 因此LMA的细胞水平的研究大多用横切切片中的细胞面积来代表细胞体积(Pyankov et al., 1999; Castro-Díez et al., 2000; Villar et al., 2013), 但采用这种方法很难准确反映细胞的真实的体积.随着技术的发展, 在近期的一些文献中也采用激光扫描共聚焦显微镜形成的3D图像计算叶肉细胞的体积(Xu et al., 2012).LMA的细胞水平的研究表明, 木本植物和草本植物的叶肉细胞增大是导致叶片增厚的主要原因, 同时细胞大小与LD呈负相关关系, 但细胞大小与LMA无明显的相关性(Pyankov et al., 1999; Castro-Díez et al., 2000).Villar等(2013)研究发现, 常绿和落叶的木本植物的叶片LD相似, 常绿木本植物叶肉细胞增大引起的叶片增厚是导致其LMA值普遍高于落叶木本植物的原因.可见, 只有从细胞水平分解叶片, 才能更好地分析引起LMA变化的原因. ...
... 无明显的相关性(Pyankov et al., 1999; Castro-Díez et al., 2000).Villar等(2013)研究发现, 常绿和落叶的木本植物的叶片LD相似, 常绿木本植物叶肉细胞增大引起的叶片增厚是导致其LMA值普遍高于落叶木本植物的原因.可见, 只有从细胞水平分解叶片, 才能更好地分析引起LMA变化的原因. ...
The relationship between nitrogen and photosynthesis in relation to leaf age in a tropical xerophytic tree.
1
1992
... 草本植物的LMA普遍比木本植物的低, 叶片生长速度快, 资源获取和流动的速度快, 进化程度比木本植物高(Flores et al., 2014).一般来说, 多年生草本植物比一年生草本植物的LMA高.但在双子叶草本植物中, 一年生与多年生植物之间的LMA差异不大; 而单子叶植物中, 禾本科多年生植物比一年生植物的LMA平均高60% (Poorter et al., 2009).而且同一片叶不同部位的LMA也不同, Niinemets等(2007)研究了大量的草本植物叶片, 表明叶片中脉部分的LMA比叶片其余部分的LMA高, 之间的差异约为6倍.然而, 这种差异可能是由物种的特异性决定的, 小麦叶片中脉的LMA与叶片其余部分的LMA差异不大(Rawson et al., 1987).而且大多数禾本科植物基部叶片的LMA比末端叶片的LMA高(Poorter et al., 2009).Pinto等(2015)的研究表明C4禾本科植物的LMA主要与其亚型有关.但目前关于草本植物的LMA的相关的研究还不是很多, 尤其是关于禾本科植物的研究. ...
Leaf xeromorphy as related to physiological and structural influences.
2
1950
... 树木冠层内的LMA的差异也是森林生态系统研究的重要内容.LMA与叶片在冠层中的位置有关, 一般处于冠层顶部的叶片LMA值比冠层底部的高(Koch et al., 2004; Lichtenthaler et al., 2007; Poorter et al., 2009; Weerasinghe et al., 2014).据Koch等(2004)报道100 m高的红杉(Sequoia sempervirens)的冠层底部LMA为150 g·m-2, 而冠层顶部LMA值达800 g·m-2, 这是比较极端的例子.一般来说, 不管是木本植物还是草本植物, 其冠层LMA大约是底层LMA的两倍(Poorter et al., 2009).这种底层和冠层之间的差异在一定程度上是由物种的特异性决定的(Lichtenthaler et al., 2007).此外, LMA还随叶寿命的不同表现出较大的差异.叶寿命是植物在长期适应环境的过程中为获得最大光合生产所形成的适应策略(Reich et al., 1991; 张林和罗天祥, 2004).LMA反映了植物单位叶面积的构建成本, 通常与叶寿命呈正相关关系.LMA低的植物, 其光合效率高, 生长速度快, 环境胁迫应答策略表现为逃避, 因而用于防御投资的物质较少, 叶寿命较短, 如草本植物、落叶乔灌木; 而LMA高的植物, 光合效率慢, 生长速度慢, 环境胁迫应答的策略表现为抵御和耐受, 需要不断积累更多的物质构建保卫构造, 因此表现出较长的叶寿命, 如常绿木本植物(Reich et al., 1991; Wright et al., 2004; Wyka et al., 2012; 王瑞雪等, 2014).常绿木本植物的LMA通常随叶龄的增长而逐渐增加, LMA的增加主要与叶肉细胞的细胞壁变厚、木质素含量增加、叶片变厚、叶片的机械组织含量增加有关(Castro-Díez et al., 2000; Hanba et al., 2002; Wright et al., 2004).Niinemets等(2005, 2006)报道地中海3种阔叶常绿树种月桂(Laurus nobilis)、油橄榄(Olea europea)、冬青栎(Quercus ilex)的LMA随叶龄的增长而增加.而且叶龄对LMA的影响还与叶片在冠层中的位置有关, Han等(2008)报道赤松冠层底部LMA随叶龄的增加而增加的幅度为27%, 但叶龄对冠层顶部LMA无影响.此外, 在常绿木本植物中, 叶寿命不同的植物, 其LMA也有较大的差异, Wyka等(2012)报道松柏类比阔叶类植物的叶寿命长, 其LMA也显著高于阔叶类植物. ...
... 高的植物, 光合效率慢, 生长速度慢, 环境胁迫应答的策略表现为抵御和耐受, 需要不断积累更多的物质构建保卫构造, 因此表现出较长的叶寿命, 如常绿木本植物(Reich et al., 1991; Wright et al., 2004; Wyka et al., 2012; 王瑞雪等, 2014).常绿木本植物的LMA通常随叶龄的增长而逐渐增加, LMA的增加主要与叶肉细胞的细胞壁变厚、木质素含量增加、叶片变厚、叶片的机械组织含量增加有关(Castro-Díez et al., 2000; Hanba et al., 2002; Wright et al., 2004).Niinemets等(2005, 2006)报道地中海3种阔叶常绿树种月桂(Laurus nobilis)、油橄榄(Olea europea)、冬青栎(Quercus ilex)的LMA随叶龄的增长而增加.而且叶龄对LMA的影响还与叶片在冠层中的位置有关, Han等(2008)报道赤松冠层底部LMA随叶龄的增加而增加的幅度为27%, 但叶龄对冠层顶部LMA无影响.此外, 在常绿木本植物中, 叶寿命不同的植物, 其LMA也有较大的差异, Wyka等(2012)报道松柏类比阔叶类植物的叶寿命长, 其LMA也显著高于阔叶类植物. ...
Quantitative analysis of cell division in leaves: Methods, developmental patterns and effects of environmental conditions.
1
2000
... 要进一步研究引起LMA变化的原因, 需要在组织水平进一步分解LMA.20世纪50年代以来, 很多学者在植物解剖结构方面开展了大量的研究, 表明不同的组织在整个叶片中的比例不一样, 同一组织在叶片中的比例受植物种类和环境的影响(Witkowski & Lamont, 1991; Garnier & Laurent, 1994; Poorter et al., 2009; Villar et al., 2013).在环境梯度上, 植物种间的LMA与其叶片组织特征(叶厚度、叶密度)的变化趋势、变化程度不一致(Fajardo & Piper, 2010; Richardson et al., 2013), 因此要想揭示种间LMA差异的原因, 必须分解LMA.在一些研究中, 常常将组织厚度当作组织体积, 这对于阔叶型的扁平状叶片的组织(叶肉组织或上表皮)是适当的, 但很多植物的叶片本身不规则(如松科、柏科), 而且叶片本身一些组织的形状也不规则, 例如维管组织, 因此将组织体积理解为组织厚度并不合适(Poorter et al., 2009).Roderick等(1999)提出LMA的计算公式中的体积应该是各部分组织的体积加上气室体积.Poorter等(2009)从组织水平提出LMA的计算公式: ...
Irradiance and phenotype: comparative eco-development of sun and shade leaves in relation to photosynthetic CO2 diffusion.
2006
Leaf functional anatomy in relation to photosynthesis.
3
2011
... LMA是一个复合的结构参数, 与叶片的很多解剖结构有关, 因而对光合作用有潜在的影响(Niinemets et al., 2009a; Scafaro et al., 2011; Terashima et al., 2011).近年来, LMA对光合作用的影响引起了生理生态学家的广泛关注.全球叶片经济型谱的建立已明确LMA与叶片单位干物质的最大光合能力、叶龄、单位干物质的氮、磷含量等密切相关(Wright et al., 2004).LMA低的植物, 其叶片生命周期短、单位干物质营养元素含量高, 光合能力强、单位叶面积的经济投入较低, 导致植物资源的利用效率高而生长速度快; LMA高的植物, 其单位叶面积的投入较高, 尤其增加了叶片的维管组织和细胞壁的投入(Niinemets et al., 2007; Hikosaka & Shigeno, 2009), 虽然增强了植物耐贫瘠、耐干旱的能力, 但导致较低的光合能力和缓慢的生长速率(Niinemets, 2001; Wright et al., 2004).近年的研究进一步表明, LMA与植物的单位叶面积的光合能力(Aarea)没有明显的相关性; 但随LMA的增加, 单位叶干质量的最大光合能力(Amass)有明显的下降趋势(Niinemets & Sack, 2006; Hassiotou et al., 2010). ...
... 草本植物与木本植物相比, 叶片薄而柔软, LMA低, 有较高的gm和光合速率(Flexas et al., 2008; Scafaro et al., 2011; Tomás et al., 2013; Muir et al., 2014).Flexas等(2008)研究表明: LMA低的植物, 其gm较高且变化幅度较大, 也就是不同种类或功能群的低LMA植物的LMA与其gm之间没有显著的相关性(Flexas et al., 2008).但是, 近年来也有不同的研究结果, 通过对番茄野生近缘种(Solanum sect. Lycopersicon, sect. Lycopersicoides) (Muir et al., 2014)、水稻野生近缘种(Oryza spp.) (Scafaro et al., 2011)、向日葵(Helianthus annuus)、辣椒(Capsicum annuum)、菜豆(Phaseolus vulgaris)、菠菜(Spinacea oleracea)、罗勒(Ocimum basilicum) (Tomás et al., 2013)的研究表明草本植物的LMA与gm也呈负相关关系, 但是导致gm下降的原因却各有不同.Muir等(2014)通过对8种番茄的研究表明, LMA与gm的关系主要受细胞内含物和叶片厚度的调控.Scafaro等(2011)研究表明, 在3种水稻中, 叶肉细胞壁增厚是gm下降, 并导致光合速率下降的重要原因.Tomás等(2013)对5种不同科属的草本植物的研究指出: 草本植物的细胞壁厚度对gm没有显著的影响, 细胞质和基质对gm影响约占60%, 是影响叶肉细胞导度的重要原因.由此来看, 导致草本植物gm下降的原因应该与植物的种的特异性有关, 具体原因还需要进一步研究.总之, 根据目前的资料报道, 无论是木本植物还是草本植物, LMA与gm之间的负相关关系是导致高LMA的植物光合效率下降的主要原因. ...
Importance of leaf anatomy in determining mesophyll diffusion conductance to CO2 across species: Quantitative limitations and scaling up by models.
1
2013
... 短时间的水分胁迫对植物的叶片解剖结构没有影响, 植物只有处于长期缺水状态才会使叶片的解剖结构发生实质性的改变(Poorter et al., 2009).目前, 大多数学者的研究表明耐旱性强的植物一般表现出较高的LMA, 这与叶片厚度和密度的增加有关(Niinemets et al., 2009b).龚春梅和王根轩(2010)通过研究干旱半干旱地区植物群落特征发现植物的LMA随干旱胁迫的加剧而增大.董廷发等(2012)的研究表明在植物群落水平上, 干旱生境中植物的LMA显著高于湿润生境.在长期水分胁迫条件下, 叶片的各组织和细胞结构的改变是导致LMA增加的直接原因.植物经过长期的水分胁迫, 叶片的厚度显著增加, 这与叶片上表皮组织、栅栏组织、海绵组织的增厚有关, 尤其是栅栏组织(Boughalleb & Hajlaoui, 2011; Galmés et al., 2013).此外, 叶片细胞的大小和结构也会发生适应性改变.水分胁迫下发育的叶片其扩展速率较低, 细胞因此更小, 更紧凑, 气室变小(Shield, 1950).Zhang等(2011)的研究发现水杉(Metasequoia glyptostroboides)幼苗叶片在长期水分胁迫下, 其叶肉细胞及上表皮细胞显著变小.细胞结构的改变主要表现为细胞壁增厚, 木质素含量增加(Sobrado, 1992; Henry et al., 2000).以上这些叶片组织和细胞的改变最终表现为叶片的LD及LMA逐渐增加(Nogués & Baker, 2000; Boughalleb & Hajlaoui, 2011).即使水分胁迫解除, 植物仍然会维持叶片解剖结构的适应性的改变; 而且经过干旱胁迫后的植物拥有更强韧的叶片及更高的LMA (Kalapos et al., 1996; Niinemets et al., 2009b; Boughalleb & Hajlaoui, 2011).这种适应性的改变最终对植物的生理过程尤其是光合作用产生较大的影响.很多研究表明植物遭受干旱胁迫后的光合速率会显著下降, 即使胁迫解除后也很难再恢复到胁迫前的值, 这与叶片解剖结构适应性的改变有关(Flexas et al., 2001; Gallé et al., 2009; Galmés et al., 2011).近年来的研究强调gm在限制植物光合速率从水分胁迫恢复中扮演重要角色; 而且大多研究认为叶片解剖学的适应性的改变是引起gm下降的重要原因, 如水分胁迫引起的LMA、LD增加、尤其是细胞壁增厚(Gallé et al., 2009; Boughalleb & Hajlaoui, 2011; Galmés et al., 2013). ...
Developmental changes in mesophyll diffusion conductance and photosynthetic capacity under different light and water availabilities in Populus tremula: How structure constrains function. Plant,
1
2012
... 短时间的水分胁迫对植物的叶片解剖结构没有影响, 植物只有处于长期缺水状态才会使叶片的解剖结构发生实质性的改变(Poorter et al., 2009).目前, 大多数学者的研究表明耐旱性强的植物一般表现出较高的LMA, 这与叶片厚度和密度的增加有关(Niinemets et al., 2009b).龚春梅和王根轩(2010)通过研究干旱半干旱地区植物群落特征发现植物的LMA随干旱胁迫的加剧而增大.董廷发等(2012)的研究表明在植物群落水平上, 干旱生境中植物的LMA显著高于湿润生境.在长期水分胁迫条件下, 叶片的各组织和细胞结构的改变是导致LMA增加的直接原因.植物经过长期的水分胁迫, 叶片的厚度显著增加, 这与叶片上表皮组织、栅栏组织、海绵组织的增厚有关, 尤其是栅栏组织(Boughalleb & Hajlaoui, 2011; Galmés et al., 2013).此外, 叶片细胞的大小和结构也会发生适应性改变.水分胁迫下发育的叶片其扩展速率较低, 细胞因此更小, 更紧凑, 气室变小(Shield, 1950).Zhang等(2011)的研究发现水杉(Metasequoia glyptostroboides)幼苗叶片在长期水分胁迫下, 其叶肉细胞及上表皮细胞显著变小.细胞结构的改变主要表现为细胞壁增厚, 木质素含量增加(Sobrado, 1992; Henry et al., 2000).以上这些叶片组织和细胞的改变最终表现为叶片的LD及LMA逐渐增加(Nogués & Baker, 2000; Boughalleb & Hajlaoui, 2011).即使水分胁迫解除, 植物仍然会维持叶片解剖结构的适应性的改变; 而且经过干旱胁迫后的植物拥有更强韧的叶片及更高的LMA (Kalapos et al., 1996; Niinemets et al., 2009b; Boughalleb & Hajlaoui, 2011).这种适应性的改变最终对植物的生理过程尤其是光合作用产生较大的影响.很多研究表明植物遭受干旱胁迫后的光合速率会显著下降, 即使胁迫解除后也很难再恢复到胁迫前的值, 这与叶片解剖结构适应性的改变有关(Flexas et al., 2001; Gallé et al., 2009; Galmés et al., 2011).近年来的研究强调gm在限制植物光合速率从水分胁迫恢复中扮演重要角色; 而且大多研究认为叶片解剖学的适应性的改变是引起gm下降的重要原因, 如水分胁迫引起的LMA、LD增加、尤其是细胞壁增厚(Gallé et al., 2009; Boughalleb & Hajlaoui, 2011; Galmés et al., 2013). ...
A quantitative analysis of phenotypic variations of Metrosideros polymorpha within and across populations along environmental gradients on Mauna Loa, Hawaii.
Multilevel genomic analysis of the response of transcripts, enzyme activities and metabolites in Arabidopsis rosettes to a progressive decrease of temperature in the nonfreezing range. Plant,
1
2008
... 目前研究表明, 具有不同形态和生境的植物种间或种内的gm差异很大(Flexas et al., 2008; Warren, 2008; Niinemets et al., 2009a, 2011), 而且特定种类或基因型的植物, 其最大gm与其叶片的解剖学特征有很大的关系(Niinemets et al., 2009a; Tosens et al., 2012).叶片内的CO2的扩散路径主要由两种相组成, 一种是细胞间的气相, 另一种是细胞的液相(主要由水和脂类组成) (Niinemets & Reichstein, 2003; Evans et al., 2009).细胞间气相的变化对CO2的扩散能力影响不大, 因此对整个光合能力没有显著的影响(Parkhurst & Mott, 1990; Evans et al., 2009).细胞相主要由细胞壁、质膜、胞液、叶绿体被膜、基质组成, 其中细胞壁和叶绿体被膜对gm的限制最大(Terashima et al., 2011).据目前的研究估算, 细胞壁厚度(Tcw)对gm的影响占25%-50%, 两者呈负相关关系(Evans et al., 2009; Terashima et al., 2011; Tosens et al., 2012).LMA高的植物, 总是伴随细胞壁的增厚, gm下降, CO2的同化速率降低, 最终导致光合能力下降(Niinemets, 1999; Terashima et al., 2006; Niinemets et al., 2007; Hassiotou et al., 2009).关于这一观点, 目前已在很多硬叶木本植物中得到广泛的证实, 如澳大利亚49种山龙眼科(Proteaceae)的植物(Hassiotou et al., 2010), Abies alba和Abies pinsapo (Peguero-Pina et al., 2012), 冬青栎(Quercus ilex)、柑橘(Citrus reticulate)、橡皮树(Ficus elastica)、海桐(Pittosporum tobira)、华盛顿棕榈(Washingtonia filifera) (Tomás et al., 2013).而且在冠层内, LMA随树木高度增加而增加的过程中所伴随的最大光合效率的下降的主要因素也是CO2的扩散阻力增加, gm下降(Han, 2011). ...
The chemical composition and anatomical structure of leaves of grass species differing in relative growth rate. Plant,
3
1994
... LMA是一个复合的结构参数, 与叶片的很多解剖结构有关, 因而对光合作用有潜在的影响(Niinemets et al., 2009a; Scafaro et al., 2011; Terashima et al., 2011).近年来, LMA对光合作用的影响引起了生理生态学家的广泛关注.全球叶片经济型谱的建立已明确LMA与叶片单位干物质的最大光合能力、叶龄、单位干物质的氮、磷含量等密切相关(Wright et al., 2004).LMA低的植物, 其叶片生命周期短、单位干物质营养元素含量高, 光合能力强、单位叶面积的经济投入较低, 导致植物资源的利用效率高而生长速度快; LMA高的植物, 其单位叶面积的投入较高, 尤其增加了叶片的维管组织和细胞壁的投入(Niinemets et al., 2007; Hikosaka & Shigeno, 2009), 虽然增强了植物耐贫瘠、耐干旱的能力, 但导致较低的光合能力和缓慢的生长速率(Niinemets, 2001; Wright et al., 2004).近年的研究进一步表明, LMA与植物的单位叶面积的光合能力(Aarea)没有明显的相关性; 但随LMA的增加, 单位叶干质量的最大光合能力(Amass)有明显的下降趋势(Niinemets & Sack, 2006; Hassiotou et al., 2010). ...
... 目前研究表明, 具有不同形态和生境的植物种间或种内的gm差异很大(Flexas et al., 2008; Warren, 2008; Niinemets et al., 2009a, 2011), 而且特定种类或基因型的植物, 其最大gm与其叶片的解剖学特征有很大的关系(Niinemets et al., 2009a; Tosens et al., 2012).叶片内的CO2的扩散路径主要由两种相组成, 一种是细胞间的气相, 另一种是细胞的液相(主要由水和脂类组成) (Niinemets & Reichstein, 2003; Evans et al., 2009).细胞间气相的变化对CO2的扩散能力影响不大, 因此对整个光合能力没有显著的影响(Parkhurst & Mott, 1990; Evans et al., 2009).细胞相主要由细胞壁、质膜、胞液、叶绿体被膜、基质组成, 其中细胞壁和叶绿体被膜对gm的限制最大(Terashima et al., 2011).据目前的研究估算, 细胞壁厚度(Tcw)对gm的影响占25%-50%, 两者呈负相关关系(Evans et al., 2009; Terashima et al., 2011; Tosens et al., 2012).LMA高的植物, 总是伴随细胞壁的增厚, gm下降, CO2的同化速率降低, 最终导致光合能力下降(Niinemets, 1999; Terashima et al., 2006; Niinemets et al., 2007; Hassiotou et al., 2009).关于这一观点, 目前已在很多硬叶木本植物中得到广泛的证实, 如澳大利亚49种山龙眼科(Proteaceae)的植物(Hassiotou et al., 2010), Abies alba和Abies pinsapo (Peguero-Pina et al., 2012), 冬青栎(Quercus ilex)、柑橘(Citrus reticulate)、橡皮树(Ficus elastica)、海桐(Pittosporum tobira)、华盛顿棕榈(Washingtonia filifera) (Tomás et al., 2013).而且在冠层内, LMA随树木高度增加而增加的过程中所伴随的最大光合效率的下降的主要因素也是CO2的扩散阻力增加, gm下降(Han, 2011). ...
... ; Terashima et al., 2011; Tosens et al., 2012).LMA高的植物, 总是伴随细胞壁的增厚, gm下降, CO2的同化速率降低, 最终导致光合能力下降(Niinemets, 1999; Terashima et al., 2006; Niinemets et al., 2007; Hassiotou et al., 2009).关于这一观点, 目前已在很多硬叶木本植物中得到广泛的证实, 如澳大利亚49种山龙眼科(Proteaceae)的植物(Hassiotou et al., 2010), Abies alba和Abies pinsapo (Peguero-Pina et al., 2012), 冬青栎(Quercus ilex)、柑橘(Citrus reticulate)、橡皮树(Ficus elastica)、海桐(Pittosporum tobira)、华盛顿棕榈(Washingtonia filifera) (Tomás et al., 2013).而且在冠层内, LMA随树木高度增加而增加的过程中所伴随的最大光合效率的下降的主要因素也是CO2的扩散阻力增加, gm下降(Han, 2011). ...
Comparison of leaf construction cost in woody species with differing leaf life-spans in contrasting ecosystems.
4
2001
... 目前研究表明, 具有不同形态和生境的植物种间或种内的gm差异很大(Flexas et al., 2008; Warren, 2008; Niinemets et al., 2009a, 2011), 而且特定种类或基因型的植物, 其最大gm与其叶片的解剖学特征有很大的关系(Niinemets et al., 2009a; Tosens et al., 2012).叶片内的CO2的扩散路径主要由两种相组成, 一种是细胞间的气相, 另一种是细胞的液相(主要由水和脂类组成) (Niinemets & Reichstein, 2003; Evans et al., 2009).细胞间气相的变化对CO2的扩散能力影响不大, 因此对整个光合能力没有显著的影响(Parkhurst & Mott, 1990; Evans et al., 2009).细胞相主要由细胞壁、质膜、胞液、叶绿体被膜、基质组成, 其中细胞壁和叶绿体被膜对gm的限制最大(Terashima et al., 2011).据目前的研究估算, 细胞壁厚度(Tcw)对gm的影响占25%-50%, 两者呈负相关关系(Evans et al., 2009; Terashima et al., 2011; Tosens et al., 2012).LMA高的植物, 总是伴随细胞壁的增厚, gm下降, CO2的同化速率降低, 最终导致光合能力下降(Niinemets, 1999; Terashima et al., 2006; Niinemets et al., 2007; Hassiotou et al., 2009).关于这一观点, 目前已在很多硬叶木本植物中得到广泛的证实, 如澳大利亚49种山龙眼科(Proteaceae)的植物(Hassiotou et al., 2010), Abies alba和Abies pinsapo (Peguero-Pina et al., 2012), 冬青栎(Quercus ilex)、柑橘(Citrus reticulate)、橡皮树(Ficus elastica)、海桐(Pittosporum tobira)、华盛顿棕榈(Washingtonia filifera) (Tomás et al., 2013).而且在冠层内, LMA随树木高度增加而增加的过程中所伴随的最大光合效率的下降的主要因素也是CO2的扩散阻力增加, gm下降(Han, 2011). ...
... 草本植物与木本植物相比, 叶片薄而柔软, LMA低, 有较高的gm和光合速率(Flexas et al., 2008; Scafaro et al., 2011; Tomás et al., 2013; Muir et al., 2014).Flexas等(2008)研究表明: LMA低的植物, 其gm较高且变化幅度较大, 也就是不同种类或功能群的低LMA植物的LMA与其gm之间没有显著的相关性(Flexas et al., 2008).但是, 近年来也有不同的研究结果, 通过对番茄野生近缘种(Solanum sect. Lycopersicon, sect. Lycopersicoides) (Muir et al., 2014)、水稻野生近缘种(Oryza spp.) (Scafaro et al., 2011)、向日葵(Helianthus annuus)、辣椒(Capsicum annuum)、菜豆(Phaseolus vulgaris)、菠菜(Spinacea oleracea)、罗勒(Ocimum basilicum) (Tomás et al., 2013)的研究表明草本植物的LMA与gm也呈负相关关系, 但是导致gm下降的原因却各有不同.Muir等(2014)通过对8种番茄的研究表明, LMA与gm的关系主要受细胞内含物和叶片厚度的调控.Scafaro等(2011)研究表明, 在3种水稻中, 叶肉细胞壁增厚是gm下降, 并导致光合速率下降的重要原因.Tomás等(2013)对5种不同科属的草本植物的研究指出: 草本植物的细胞壁厚度对gm没有显著的影响, 细胞质和基质对gm影响约占60%, 是影响叶肉细胞导度的重要原因.由此来看, 导致草本植物gm下降的原因应该与植物的种的特异性有关, 具体原因还需要进一步研究.总之, 根据目前的资料报道, 无论是木本植物还是草本植物, LMA与gm之间的负相关关系是导致高LMA的植物光合效率下降的主要原因. ...
... (2)植物LMA反映了碳资源的获取能力和资源的利用方式, 与其生态位息息相关.当环境发生改变时, 植物通过调节LMA的大小来改变调整资源的获取方式和生存对策, 最终可能导致其生态位的改变, 由此对整个生态群落产生较大的影响(Garnier et al., 2004; Díaz et al., 2007).因此, 面对全球高温、干旱、水涝等极端气候的频繁出现, 研究逆境条件下植物的生理生化反应及生存策略对于预测气候变化对植物生态的系统的影响具有重要意义.目前, 关于逆境条件下植物LMA的研究相对较少, 逆境引起的LMA的改变最终导致生理反应的改变, 尤其对光合作用的影响最大.研究表明, gm是限制光合作用的一个重要因素, 而LMA的改变总伴随gm的改变.LMA高、叶片厚的植物, 其细胞壁导度是限制gm的主要因素; 而LMA低、叶片薄的植物, 其限制gm的主要因素各不相同, 其机制仍然不是很清楚.有学者提出, 水通道蛋白(aquaporins)和碳酸酐酶(carbonic anhydrases)可能对低LMA植物的gm具有重要调控作用(Tomás et al., 2013).关于两者对gm的调节机制已有一些初步的报道(Perez-Martin et al., 2014; Boudichevskaia et al., 2015), 但是还需要进一步完善研究以明确其调控机理.此外, 研究环境改变对植物LMA、gm、光合效率的影响及三者之间的相互的调控机制对于理解植物的进化适应性以及气候变化对植物的影响具有重要价值, 这不仅是目前研究的重点, 也是将来研究的重要内容. ...
Differences in construction costs and chemical composition between deciduous and evergreen woody species are small as compared to differences between families. Plant,
2
2006
... 目前研究表明, 具有不同形态和生境的植物种间或种内的gm差异很大(Flexas et al., 2008; Warren, 2008; Niinemets et al., 2009a, 2011), 而且特定种类或基因型的植物, 其最大gm与其叶片的解剖学特征有很大的关系(Niinemets et al., 2009a; Tosens et al., 2012).叶片内的CO2的扩散路径主要由两种相组成, 一种是细胞间的气相, 另一种是细胞的液相(主要由水和脂类组成) (Niinemets & Reichstein, 2003; Evans et al., 2009).细胞间气相的变化对CO2的扩散能力影响不大, 因此对整个光合能力没有显著的影响(Parkhurst & Mott, 1990; Evans et al., 2009).细胞相主要由细胞壁、质膜、胞液、叶绿体被膜、基质组成, 其中细胞壁和叶绿体被膜对gm的限制最大(Terashima et al., 2011).据目前的研究估算, 细胞壁厚度(Tcw)对gm的影响占25%-50%, 两者呈负相关关系(Evans et al., 2009; Terashima et al., 2011; Tosens et al., 2012).LMA高的植物, 总是伴随细胞壁的增厚, gm下降, CO2的同化速率降低, 最终导致光合能力下降(Niinemets, 1999; Terashima et al., 2006; Niinemets et al., 2007; Hassiotou et al., 2009).关于这一观点, 目前已在很多硬叶木本植物中得到广泛的证实, 如澳大利亚49种山龙眼科(Proteaceae)的植物(Hassiotou et al., 2010), Abies alba和Abies pinsapo (Peguero-Pina et al., 2012), 冬青栎(Quercus ilex)、柑橘(Citrus reticulate)、橡皮树(Ficus elastica)、海桐(Pittosporum tobira)、华盛顿棕榈(Washingtonia filifera) (Tomás et al., 2013).而且在冠层内, LMA随树木高度增加而增加的过程中所伴随的最大光合效率的下降的主要因素也是CO2的扩散阻力增加, gm下降(Han, 2011). ...
... ; Tosens et al., 2012).LMA高的植物, 总是伴随细胞壁的增厚, gm下降, CO2的同化速率降低, 最终导致光合能力下降(Niinemets, 1999; Terashima et al., 2006; Niinemets et al., 2007; Hassiotou et al., 2009).关于这一观点, 目前已在很多硬叶木本植物中得到广泛的证实, 如澳大利亚49种山龙眼科(Proteaceae)的植物(Hassiotou et al., 2010), Abies alba和Abies pinsapo (Peguero-Pina et al., 2012), 冬青栎(Quercus ilex)、柑橘(Citrus reticulate)、橡皮树(Ficus elastica)、海桐(Pittosporum tobira)、华盛顿棕榈(Washingtonia filifera) (Tomás et al., 2013).而且在冠层内, LMA随树木高度增加而增加的过程中所伴随的最大光合效率的下降的主要因素也是CO2的扩散阻力增加, gm下降(Han, 2011). ...
Exploring variation in leaf mass per area (LMA) from leaf to cell: An anatomical analysis of 26 woody species.
1
2013
... LMA不仅决定植物潜在的生长速率, 反映植物生态策略, 而且与许多生理过程息息相关(Antúnez et al., 2001; Villar et al., 2006).全球叶片经济型谱的研究明确了LMA与叶片单位干物质的最大光合能力(Amass)密切相关, Amass随LMA的增加有明显的下降趋势.LMA的大小主要由植物叶片的厚度和密度决定(Witkowski & Lamont, 1991), 两者的改变都会影响光的利用效率以及叶肉细胞导度(gm)的大小, 最终导致光合效率的改变(Muir et al., 2014).当受到各种选择压力时, 植物能通过调节LMA来改变光合效率, 使植物的投入和收益重新达到最佳的平衡点.研究表明, LMA是自然选择压力下的稳定遗传的结果, 反映了植物与环境共进化的过程, 具有遗传进化上的意义(Asner et al., 2011; Aspinwall et al., 2013; Flores et al., 2014; Tsujii et al., 2016).因此, 研究LMA具有生理、生态及进化的重要意义.从目前已发表的文章来看, 国内还缺乏关于LMA较深入的综合性报道.本文系统梳理了LMA在叶片整体、组织、细胞三个水平的结构解析和计算方法; 重点分析了LMA对光合作用的影响; 讨论了LMA的内在的遗传差异以及外部的环境胁迫因子(温度、水分、光照)对LMA的影响, 以期为今后更深入的研究提供借鉴和参考. ...
... 20世纪90年代, 很多学者对不同植物的LMA做了大量研究, 并探讨了LVA和LD与LMA之间的关系.研究表明在同一可控环境条件下, 草本植物(Garnier & Laurent, 1994; van Arendonk & Poorter, 1994)和木本植物(Castro-Diez et al., 2000)的LMA主要受LD的影响, Poorter等(2009)对这些数据作了统计和分析, 表明LMA差异的80%由LD决定, 20%由LVA决定.然而在复杂的自然环境条件下, LD和LVA与LMA之间的关系是多样化的(Choong et al., 1992; Niinemets, 1999; Wilson et al., 1999).由此可见, 仅仅把LMA分解为LVA和LD并不能很好地解释导致植物种类之间LMA差异的原因. ...
Stand aside stomata, another actor deserves centre stage: The forgotten role of the internal conductance to CO2 transfer.
1
2008
... 从目前的研究结果来看, 木本植物比草本植物的LMA高, 在同一环境控制下, 两者之间的差异大约为35%; 在野外环境下, 两者之间的差异约83% (Poorter et al., 2009).LMA在叶片水平可分解为叶片密度和厚度.根据目前的研究文献报道, 密度和厚度都有可能是决定木本植物LMA高低的原因(Castro-Díez et al., 2000; Wright & Cannon, 2001; Villar et al., 2013).LMA高的木本植物, 其硬化组织占整个叶片的比例高, 细胞小而紧凑、叶片含水量和氮含量较低(Wright & Cannon, 2001).常绿木本植物普遍比落叶木本植物的LMA高(Castro-Díez et al., 2000; Villar & Merino, 2001; Wright et al., 2005; Li et al., 2013; Villar et al., 2013).Castro-Díez (2000)和Villar等(2013)认为常绿和落叶木本植物的叶片密度之间没有显著的差异, 常绿木本植物较高的LMA主要与叶片厚度有关, 同科属中的常绿木本植物的叶片通常比落叶木本植物的叶片厚, 而叶肉组织增厚是导致叶片变厚的主要原因.然而, 不同科属的常绿和落叶植物之间的LMA也存在交差和重叠, 这主要是由植物的系统发育决定的; 也就是不同科属之间植物的LMA是有差异的, 而在同一科属中, 常绿植物比落叶植物的LMA高(Poorter et al., 2009; Villar et al., 2013). ...
Canopy position affects the relationships between leaf respiration and associated traits in a tropical rainforestin Far North Queensland.
1
2014
... LMA不仅决定植物潜在的生长速率, 反映植物生态策略, 而且与许多生理过程息息相关(Antúnez et al., 2001; Villar et al., 2006).全球叶片经济型谱的研究明确了LMA与叶片单位干物质的最大光合能力(Amass)密切相关, Amass随LMA的增加有明显的下降趋势.LMA的大小主要由植物叶片的厚度和密度决定(Witkowski & Lamont, 1991), 两者的改变都会影响光的利用效率以及叶肉细胞导度(gm)的大小, 最终导致光合效率的改变(Muir et al., 2014).当受到各种选择压力时, 植物能通过调节LMA来改变光合效率, 使植物的投入和收益重新达到最佳的平衡点.研究表明, LMA是自然选择压力下的稳定遗传的结果, 反映了植物与环境共进化的过程, 具有遗传进化上的意义(Asner et al., 2011; Aspinwall et al., 2013; Flores et al., 2014; Tsujii et al., 2016).因此, 研究LMA具有生理、生态及进化的重要意义.从目前已发表的文章来看, 国内还缺乏关于LMA较深入的综合性报道.本文系统梳理了LMA在叶片整体、组织、细胞三个水平的结构解析和计算方法; 重点分析了LMA对光合作用的影响; 讨论了LMA的内在的遗传差异以及外部的环境胁迫因子(温度、水分、光照)对LMA的影响, 以期为今后更深入的研究提供借鉴和参考. ...
Plant ecological strategies: Some leading dimensions of variation between species.
6
2002
... 要进一步研究引起LMA变化的原因, 需要在组织水平进一步分解LMA.20世纪50年代以来, 很多学者在植物解剖结构方面开展了大量的研究, 表明不同的组织在整个叶片中的比例不一样, 同一组织在叶片中的比例受植物种类和环境的影响(Witkowski & Lamont, 1991; Garnier & Laurent, 1994; Poorter et al., 2009; Villar et al., 2013).在环境梯度上, 植物种间的LMA与其叶片组织特征(叶厚度、叶密度)的变化趋势、变化程度不一致(Fajardo & Piper, 2010; Richardson et al., 2013), 因此要想揭示种间LMA差异的原因, 必须分解LMA.在一些研究中, 常常将组织厚度当作组织体积, 这对于阔叶型的扁平状叶片的组织(叶肉组织或上表皮)是适当的, 但很多植物的叶片本身不规则(如松科、柏科), 而且叶片本身一些组织的形状也不规则, 例如维管组织, 因此将组织体积理解为组织厚度并不合适(Poorter et al., 2009).Roderick等(1999)提出LMA的计算公式中的体积应该是各部分组织的体积加上气室体积.Poorter等(2009)从组织水平提出LMA的计算公式: ...
... 近年来, 随着研究方法和手段的发展, 开始在细胞水平对LMA进行分解.LMA的细胞水平分解是将不同组织(t)的单位面积的体积(VAt)分解为该组织单位叶面积(A)细胞数量(Cell#t)乘以该组织细胞的平均大小(Cell sizet) (Villar et al., 2013): ...
... 目前植物解剖学的研究大多采用较薄的组织切片, 因此LMA的细胞水平的研究大多用横切切片中的细胞面积来代表细胞体积(Pyankov et al., 1999; Castro-Díez et al., 2000; Villar et al., 2013), 但采用这种方法很难准确反映细胞的真实的体积.随着技术的发展, 在近期的一些文献中也采用激光扫描共聚焦显微镜形成的3D图像计算叶肉细胞的体积(Xu et al., 2012).LMA的细胞水平的研究表明, 木本植物和草本植物的叶肉细胞增大是导致叶片增厚的主要原因, 同时细胞大小与LD呈负相关关系, 但细胞大小与LMA无明显的相关性(Pyankov et al., 1999; Castro-Díez et al., 2000).Villar等(2013)研究发现, 常绿和落叶的木本植物的叶片LD相似, 常绿木本植物叶肉细胞增大引起的叶片增厚是导致其LMA值普遍高于落叶木本植物的原因.可见, 只有从细胞水平分解叶片, 才能更好地分析引起LMA变化的原因. ...
... 从目前的研究结果来看, 木本植物比草本植物的LMA高, 在同一环境控制下, 两者之间的差异大约为35%; 在野外环境下, 两者之间的差异约83% (Poorter et al., 2009).LMA在叶片水平可分解为叶片密度和厚度.根据目前的研究文献报道, 密度和厚度都有可能是决定木本植物LMA高低的原因(Castro-Díez et al., 2000; Wright & Cannon, 2001; Villar et al., 2013).LMA高的木本植物, 其硬化组织占整个叶片的比例高, 细胞小而紧凑、叶片含水量和氮含量较低(Wright & Cannon, 2001).常绿木本植物普遍比落叶木本植物的LMA高(Castro-Díez et al., 2000; Villar & Merino, 2001; Wright et al., 2005; Li et al., 2013; Villar et al., 2013).Castro-Díez (2000)和Villar等(2013)认为常绿和落叶木本植物的叶片密度之间没有显著的差异, 常绿木本植物较高的LMA主要与叶片厚度有关, 同科属中的常绿木本植物的叶片通常比落叶木本植物的叶片厚, 而叶肉组织增厚是导致叶片变厚的主要原因.然而, 不同科属的常绿和落叶植物之间的LMA也存在交差和重叠, 这主要是由植物的系统发育决定的; 也就是不同科属之间植物的LMA是有差异的, 而在同一科属中, 常绿植物比落叶植物的LMA高(Poorter et al., 2009; Villar et al., 2013). ...
... ; Villar et al., 2013).Castro-Díez (2000)和Villar等(2013)认为常绿和落叶木本植物的叶片密度之间没有显著的差异, 常绿木本植物较高的LMA主要与叶片厚度有关, 同科属中的常绿木本植物的叶片通常比落叶木本植物的叶片厚, 而叶肉组织增厚是导致叶片变厚的主要原因.然而, 不同科属的常绿和落叶植物之间的LMA也存在交差和重叠, 这主要是由植物的系统发育决定的; 也就是不同科属之间植物的LMA是有差异的, 而在同一科属中, 常绿植物比落叶植物的LMA高(Poorter et al., 2009; Villar et al., 2013). ...
... ; Villar et al., 2013). ...
Relationships between leaf lifespan and structural defences in a low-nutrient, sclerophyll flora.
2001
The world- wide leaf economics spectrum.
1
2004
... 关于植物与光照的关系, 目前研究的比较多的是森林生态系统中光照不足对植物产生的影响.在森林生态系统中, 对光的需求的差异是决定植物生长和生存的主要因素(Hanba et al., 2002).大多研究表明植物在适应光环境的过程中主要通过调节叶片形态和结构适应光照的变化, 主要表现在LMA的增大或减小(Niinemets, 2007; Lusk et al., 2008; Coble & Cavaleri, 2015).目前的研究表明, 对于同一种植物, 遮阴导致叶片LMA、厚度、密度下降, 而单位干质量的叶面积增加, 增大了其叶片捕光面积, 利于在弱光环境下生存(冯玉龙等, 2002; Lusk et al., 2008; 柳凤娟等, 2010; 王博轶等, 2012; Coble & Cavaleri, 2015).在植物的冠层内部, 阴生叶比阳生叶的LMA低, 差异可达2-4倍, LMA降低主要与叶肉细胞层厚度减少, 叶片变薄, 叶片密度显著降低, 细胞内含物显著下降有关(Niinemets, 2007; Lusk et al., 2010; Chen et al., 2014).Meng等(2013, 2014)的研究发现, 遮阴不仅会导致叶片的LMA下降, 还会使叶片倾角变小, 以截获更多的光照.值得关注的是, 在很多胁迫条件下(比如干旱、低温), 叶片LMA的增加伴随细胞壁的增厚, 然而在遮阴条件下, 虽然LMA降低了, 但其细胞壁反而增厚, 这可能是植物在低光条件下为增强其叶片硬度和韧度, 避免食草动物的取食而采取的防御机制(Hanba et al., 2002; Coble & Cavaleri, 2015).此外, 很多学者在对热带常绿和落叶木本植物的研究中发现, 在同一低光环境中, 耐阴性强的植物与需光植物相比, 具有较低的光合能力和较高的LMA, LMA的增加在一定程度上与叶肉细胞组织孔隙度的下降、叶片表皮毛密度的增加有关(Hanba et al., 2002; Lusk et al., 2008).LMA的增加使叶片更加强韧, 可增强植物对食草动物和病原菌的防御能力, 提高其在弱光下的生存几率(Kitajima et al., 1994). ...
Assesing the generality of global leaf trait relationships.
2005
Specific leaf area and leaf dry matter content as alternative predictors of plant strategies.
1
1999
... 树木冠层内的LMA的差异也是森林生态系统研究的重要内容.LMA与叶片在冠层中的位置有关, 一般处于冠层顶部的叶片LMA值比冠层底部的高(Koch et al., 2004; Lichtenthaler et al., 2007; Poorter et al., 2009; Weerasinghe et al., 2014).据Koch等(2004)报道100 m高的红杉(Sequoia sempervirens)的冠层底部LMA为150 g·m-2, 而冠层顶部LMA值达800 g·m-2, 这是比较极端的例子.一般来说, 不管是木本植物还是草本植物, 其冠层LMA大约是底层LMA的两倍(Poorter et al., 2009).这种底层和冠层之间的差异在一定程度上是由物种的特异性决定的(Lichtenthaler et al., 2007).此外, LMA还随叶寿命的不同表现出较大的差异.叶寿命是植物在长期适应环境的过程中为获得最大光合生产所形成的适应策略(Reich et al., 1991; 张林和罗天祥, 2004).LMA反映了植物单位叶面积的构建成本, 通常与叶寿命呈正相关关系.LMA低的植物, 其光合效率高, 生长速度快, 环境胁迫应答策略表现为逃避, 因而用于防御投资的物质较少, 叶寿命较短, 如草本植物、落叶乔灌木; 而LMA高的植物, 光合效率慢, 生长速度慢, 环境胁迫应答的策略表现为抵御和耐受, 需要不断积累更多的物质构建保卫构造, 因此表现出较长的叶寿命, 如常绿木本植物(Reich et al., 1991; Wright et al., 2004; Wyka et al., 2012; 王瑞雪等, 2014).常绿木本植物的LMA通常随叶龄的增长而逐渐增加, LMA的增加主要与叶肉细胞的细胞壁变厚、木质素含量增加、叶片变厚、叶片的机械组织含量增加有关(Castro-Díez et al., 2000; Hanba et al., 2002; Wright et al., 2004).Niinemets等(2005, 2006)报道地中海3种阔叶常绿树种月桂(Laurus nobilis)、油橄榄(Olea europea)、冬青栎(Quercus ilex)的LMA随叶龄的增长而增加.而且叶龄对LMA的影响还与叶片在冠层中的位置有关, Han等(2008)报道赤松冠层底部LMA随叶龄的增加而增加的幅度为27%, 但叶龄对冠层顶部LMA无影响.此外, 在常绿木本植物中, 叶寿命不同的植物, 其LMA也有较大的差异, Wyka等(2012)报道松柏类比阔叶类植物的叶寿命长, 其LMA也显著高于阔叶类植物. ...
Leaf specific mass confounds leaf density and thickness.
... 目前研究表明, 具有不同形态和生境的植物种间或种内的gm差异很大(Flexas et al., 2008; Warren, 2008; Niinemets et al., 2009a, 2011), 而且特定种类或基因型的植物, 其最大gm与其叶片的解剖学特征有很大的关系(Niinemets et al., 2009a; Tosens et al., 2012).叶片内的CO2的扩散路径主要由两种相组成, 一种是细胞间的气相, 另一种是细胞的液相(主要由水和脂类组成) (Niinemets & Reichstein, 2003; Evans et al., 2009).细胞间气相的变化对CO2的扩散能力影响不大, 因此对整个光合能力没有显著的影响(Parkhurst & Mott, 1990; Evans et al., 2009).细胞相主要由细胞壁、质膜、胞液、叶绿体被膜、基质组成, 其中细胞壁和叶绿体被膜对gm的限制最大(Terashima et al., 2011).据目前的研究估算, 细胞壁厚度(Tcw)对gm的影响占25%-50%, 两者呈负相关关系(Evans et al., 2009; Terashima et al., 2011; Tosens et al., 2012).LMA高的植物, 总是伴随细胞壁的增厚, gm下降, CO2的同化速率降低, 最终导致光合能力下降(Niinemets, 1999; Terashima et al., 2006; Niinemets et al., 2007; Hassiotou et al., 2009).关于这一观点, 目前已在很多硬叶木本植物中得到广泛的证实, 如澳大利亚49种山龙眼科(Proteaceae)的植物(Hassiotou et al., 2010), Abies alba和Abies pinsapo (Peguero-Pina et al., 2012), 冬青栎(Quercus ilex)、柑橘(Citrus reticulate)、橡皮树(Ficus elastica)、海桐(Pittosporum tobira)、华盛顿棕榈(Washingtonia filifera) (Tomás et al., 2013).而且在冠层内, LMA随树木高度增加而增加的过程中所伴随的最大光合效率的下降的主要因素也是CO2的扩散阻力增加, gm下降(Han, 2011). ...
Responses of leaf structure and photosynthetic properties to intra-canopy light gradients: A common garden test with four broadleaf deciduous angiosperm and seven evergreen conifer tree species.
1
2012
... 树木冠层内的LMA的差异也是森林生态系统研究的重要内容.LMA与叶片在冠层中的位置有关, 一般处于冠层顶部的叶片LMA值比冠层底部的高(Koch et al., 2004; Lichtenthaler et al., 2007; Poorter et al., 2009; Weerasinghe et al., 2014).据Koch等(2004)报道100 m高的红杉(Sequoia sempervirens)的冠层底部LMA为150 g·m-2, 而冠层顶部LMA值达800 g·m-2, 这是比较极端的例子.一般来说, 不管是木本植物还是草本植物, 其冠层LMA大约是底层LMA的两倍(Poorter et al., 2009).这种底层和冠层之间的差异在一定程度上是由物种的特异性决定的(Lichtenthaler et al., 2007).此外, LMA还随叶寿命的不同表现出较大的差异.叶寿命是植物在长期适应环境的过程中为获得最大光合生产所形成的适应策略(Reich et al., 1991; 张林和罗天祥, 2004).LMA反映了植物单位叶面积的构建成本, 通常与叶寿命呈正相关关系.LMA低的植物, 其光合效率高, 生长速度快, 环境胁迫应答策略表现为逃避, 因而用于防御投资的物质较少, 叶寿命较短, 如草本植物、落叶乔灌木; 而LMA高的植物, 光合效率慢, 生长速度慢, 环境胁迫应答的策略表现为抵御和耐受, 需要不断积累更多的物质构建保卫构造, 因此表现出较长的叶寿命, 如常绿木本植物(Reich et al., 1991; Wright et al., 2004; Wyka et al., 2012; 王瑞雪等, 2014).常绿木本植物的LMA通常随叶龄的增长而逐渐增加, LMA的增加主要与叶肉细胞的细胞壁变厚、木质素含量增加、叶片变厚、叶片的机械组织含量增加有关(Castro-Díez et al., 2000; Hanba et al., 2002; Wright et al., 2004).Niinemets等(2005, 2006)报道地中海3种阔叶常绿树种月桂(Laurus nobilis)、油橄榄(Olea europea)、冬青栎(Quercus ilex)的LMA随叶龄的增长而增加.而且叶龄对LMA的影响还与叶片在冠层中的位置有关, Han等(2008)报道赤松冠层底部LMA随叶龄的增加而增加的幅度为27%, 但叶龄对冠层顶部LMA无影响.此外, 在常绿木本植物中, 叶寿命不同的植物, 其LMA也有较大的差异, Wyka等(2012)报道松柏类比阔叶类植物的叶寿命长, 其LMA也显著高于阔叶类植物. ...
Contrasting leaf trait scaling relationships in tropical andtemperate species.
1
2013
... 自然界中的每一种植物都有自己独特的叶片形态和结构, 这种特异性并不是偶然的, 而是自然界选择进化的结果.每一种叶片的形态和组织结构都有与之相协调的生理过程紧密联系.因此, 从叶片形态和结构入手揭示植物的生态策略和生理过程是一种行之有效的研究手段.在植物的叶片解剖研究过程中, 大量研究资料显示比叶质量(LMA)是一个非常重要的反应植物功能的结构参数.LMA指叶片干质量与面积之间的比率(g·m-2), 是叶片众多解剖结构特征的综合体, 例如叶肉组织厚度、细胞壁厚度、细胞形状、细胞内含物等(Muir et al., 2014).在大量的叶片形态解剖结构参数中, LMA是最容易定量测定的参数之一, 它能定量测定植物在单位叶面积的投入成本, 因而被认为是反映植物生态策略的重要指标(Westoby et al., 2002; Wright et al., 2004).LMA的高低反映了植物对资源的利用效率以及对环境的应对策略.总的来讲, 低LMA的植物光合效率高, 单位叶面积的投入成本低, 显示出较快的资源流动速度及回报效率; 而高LMA的植物光合效率低, 单位叶面积的投入成本高, 资源的流动速度及回报效率低(Niinemets et al., 2007; Hikosaka & Shigeno, 2009). ...
Leaf structural characteristics are less important than leaf chemical properties in determining the response of leaf mass per area and photosynthesis of Eucalyptus saligna to industrial-age changes in [CO2] and temperature.
2
2012
... 从目前的研究结果来看, 木本植物比草本植物的LMA高, 在同一环境控制下, 两者之间的差异大约为35%; 在野外环境下, 两者之间的差异约83% (Poorter et al., 2009).LMA在叶片水平可分解为叶片密度和厚度.根据目前的研究文献报道, 密度和厚度都有可能是决定木本植物LMA高低的原因(Castro-Díez et al., 2000; Wright & Cannon, 2001; Villar et al., 2013).LMA高的木本植物, 其硬化组织占整个叶片的比例高, 细胞小而紧凑、叶片含水量和氮含量较低(Wright & Cannon, 2001).常绿木本植物普遍比落叶木本植物的LMA高(Castro-Díez et al., 2000; Villar & Merino, 2001; Wright et al., 2005; Li et al., 2013; Villar et al., 2013).Castro-Díez (2000)和Villar等(2013)认为常绿和落叶木本植物的叶片密度之间没有显著的差异, 常绿木本植物较高的LMA主要与叶片厚度有关, 同科属中的常绿木本植物的叶片通常比落叶木本植物的叶片厚, 而叶肉组织增厚是导致叶片变厚的主要原因.然而, 不同科属的常绿和落叶植物之间的LMA也存在交差和重叠, 这主要是由植物的系统发育决定的; 也就是不同科属之间植物的LMA是有差异的, 而在同一科属中, 常绿植物比落叶植物的LMA高(Poorter et al., 2009; Villar et al., 2013). ...
... 高的木本植物, 其硬化组织占整个叶片的比例高, 细胞小而紧凑、叶片含水量和氮含量较低(Wright & Cannon, 2001).常绿木本植物普遍比落叶木本植物的LMA高(Castro-Díez et al., 2000; Villar & Merino, 2001; Wright et al., 2005; Li et al., 2013; Villar et al., 2013).Castro-Díez (2000)和Villar等(2013)认为常绿和落叶木本植物的叶片密度之间没有显著的差异, 常绿木本植物较高的LMA主要与叶片厚度有关, 同科属中的常绿木本植物的叶片通常比落叶木本植物的叶片厚, 而叶肉组织增厚是导致叶片变厚的主要原因.然而, 不同科属的常绿和落叶植物之间的LMA也存在交差和重叠, 这主要是由植物的系统发育决定的; 也就是不同科属之间植物的LMA是有差异的, 而在同一科属中, 常绿植物比落叶植物的LMA高(Poorter et al., 2009; Villar et al., 2013). ...
长期模拟增温对岷江冷杉幼苗生长与生物量分配的影响
6
2010
... 自然界中的每一种植物都有自己独特的叶片形态和结构, 这种特异性并不是偶然的, 而是自然界选择进化的结果.每一种叶片的形态和组织结构都有与之相协调的生理过程紧密联系.因此, 从叶片形态和结构入手揭示植物的生态策略和生理过程是一种行之有效的研究手段.在植物的叶片解剖研究过程中, 大量研究资料显示比叶质量(LMA)是一个非常重要的反应植物功能的结构参数.LMA指叶片干质量与面积之间的比率(g·m-2), 是叶片众多解剖结构特征的综合体, 例如叶肉组织厚度、细胞壁厚度、细胞形状、细胞内含物等(Muir et al., 2014).在大量的叶片形态解剖结构参数中, LMA是最容易定量测定的参数之一, 它能定量测定植物在单位叶面积的投入成本, 因而被认为是反映植物生态策略的重要指标(Westoby et al., 2002; Wright et al., 2004).LMA的高低反映了植物对资源的利用效率以及对环境的应对策略.总的来讲, 低LMA的植物光合效率高, 单位叶面积的投入成本低, 显示出较快的资源流动速度及回报效率; 而高LMA的植物光合效率低, 单位叶面积的投入成本高, 资源的流动速度及回报效率低(Niinemets et al., 2007; Hikosaka & Shigeno, 2009). ...
... LMA是一个复合的结构参数, 与叶片的很多解剖结构有关, 因而对光合作用有潜在的影响(Niinemets et al., 2009a; Scafaro et al., 2011; Terashima et al., 2011).近年来, LMA对光合作用的影响引起了生理生态学家的广泛关注.全球叶片经济型谱的建立已明确LMA与叶片单位干物质的最大光合能力、叶龄、单位干物质的氮、磷含量等密切相关(Wright et al., 2004).LMA低的植物, 其叶片生命周期短、单位干物质营养元素含量高, 光合能力强、单位叶面积的经济投入较低, 导致植物资源的利用效率高而生长速度快; LMA高的植物, 其单位叶面积的投入较高, 尤其增加了叶片的维管组织和细胞壁的投入(Niinemets et al., 2007; Hikosaka & Shigeno, 2009), 虽然增强了植物耐贫瘠、耐干旱的能力, 但导致较低的光合能力和缓慢的生长速率(Niinemets, 2001; Wright et al., 2004).近年的研究进一步表明, LMA与植物的单位叶面积的光合能力(Aarea)没有明显的相关性; 但随LMA的增加, 单位叶干质量的最大光合能力(Amass)有明显的下降趋势(Niinemets & Sack, 2006; Hassiotou et al., 2010). ...
... ; Wright et al., 2004).近年的研究进一步表明, LMA与植物的单位叶面积的光合能力(Aarea)没有明显的相关性; 但随LMA的增加, 单位叶干质量的最大光合能力(Amass)有明显的下降趋势(Niinemets & Sack, 2006; Hassiotou et al., 2010). ...
... 其次, 高LMA植物叶片中氮浓度低可能也是植物光合能力(Amass)降低的原因.植物叶片中大部分的氮参与光合作用的运转, LMA高的植物叶片中的关键营养元素的浓度都比较低, 例如氮、磷(Wright et al., 2004).目前研究表明, 植物叶片中氮的浓度低会导致光合作用的氮利用效率以及分配到核酮糖-1,5-二磷酸羧化酶/加氧酶(Rubisco)和类囊体的氮显著下降, 从而引起光合能力的下降(Hikosaka, 2004; Hassiotou et al., 2010).Niinemets等(2004)研究表明冬青栎的LMA随叶龄的增长而增加, 而氮含量随LMA增加而下降, 氮含量的降低导致Rubisco的最大羧化活性以及光合作用的电子传递能力下降.Xu等(2012)的研究表明柳叶桉(Eucalyptus saligna)的叶氮含量随LMA的增加而降低, 单位质量的最大光合效率随氮含量降低而降低.但并非所有情况下氮含量都随LMA增加而下降, Han (2011)发现LMA随赤松(Pinus densiflora)树高的增加而增加的过程中, Nmass和Narea均表现出增加的趋势, 这可能是植物为了减少因高度引起的光合效率的下降而采取的补偿机制. ...
... 树木冠层内的LMA的差异也是森林生态系统研究的重要内容.LMA与叶片在冠层中的位置有关, 一般处于冠层顶部的叶片LMA值比冠层底部的高(Koch et al., 2004; Lichtenthaler et al., 2007; Poorter et al., 2009; Weerasinghe et al., 2014).据Koch等(2004)报道100 m高的红杉(Sequoia sempervirens)的冠层底部LMA为150 g·m-2, 而冠层顶部LMA值达800 g·m-2, 这是比较极端的例子.一般来说, 不管是木本植物还是草本植物, 其冠层LMA大约是底层LMA的两倍(Poorter et al., 2009).这种底层和冠层之间的差异在一定程度上是由物种的特异性决定的(Lichtenthaler et al., 2007).此外, LMA还随叶寿命的不同表现出较大的差异.叶寿命是植物在长期适应环境的过程中为获得最大光合生产所形成的适应策略(Reich et al., 1991; 张林和罗天祥, 2004).LMA反映了植物单位叶面积的构建成本, 通常与叶寿命呈正相关关系.LMA低的植物, 其光合效率高, 生长速度快, 环境胁迫应答策略表现为逃避, 因而用于防御投资的物质较少, 叶寿命较短, 如草本植物、落叶乔灌木; 而LMA高的植物, 光合效率慢, 生长速度慢, 环境胁迫应答的策略表现为抵御和耐受, 需要不断积累更多的物质构建保卫构造, 因此表现出较长的叶寿命, 如常绿木本植物(Reich et al., 1991; Wright et al., 2004; Wyka et al., 2012; 王瑞雪等, 2014).常绿木本植物的LMA通常随叶龄的增长而逐渐增加, LMA的增加主要与叶肉细胞的细胞壁变厚、木质素含量增加、叶片变厚、叶片的机械组织含量增加有关(Castro-Díez et al., 2000; Hanba et al., 2002; Wright et al., 2004).Niinemets等(2005, 2006)报道地中海3种阔叶常绿树种月桂(Laurus nobilis)、油橄榄(Olea europea)、冬青栎(Quercus ilex)的LMA随叶龄的增长而增加.而且叶龄对LMA的影响还与叶片在冠层中的位置有关, Han等(2008)报道赤松冠层底部LMA随叶龄的增加而增加的幅度为27%, 但叶龄对冠层顶部LMA无影响.此外, 在常绿木本植物中, 叶寿命不同的植物, 其LMA也有较大的差异, Wyka等(2012)报道松柏类比阔叶类植物的叶寿命长, 其LMA也显著高于阔叶类植物. ...
Higher thermal acclimation potential of respiration but not photosynthesis in two alpine Picea taxa in contrast to two lowland congeners.
1
2015
... 从目前的研究结果来看, 木本植物比草本植物的LMA高, 在同一环境控制下, 两者之间的差异大约为35%; 在野外环境下, 两者之间的差异约83% (Poorter et al., 2009).LMA在叶片水平可分解为叶片密度和厚度.根据目前的研究文献报道, 密度和厚度都有可能是决定木本植物LMA高低的原因(Castro-Díez et al., 2000; Wright & Cannon, 2001; Villar et al., 2013).LMA高的木本植物, 其硬化组织占整个叶片的比例高, 细胞小而紧凑、叶片含水量和氮含量较低(Wright & Cannon, 2001).常绿木本植物普遍比落叶木本植物的LMA高(Castro-Díez et al., 2000; Villar & Merino, 2001; Wright et al., 2005; Li et al., 2013; Villar et al., 2013).Castro-Díez (2000)和Villar等(2013)认为常绿和落叶木本植物的叶片密度之间没有显著的差异, 常绿木本植物较高的LMA主要与叶片厚度有关, 同科属中的常绿木本植物的叶片通常比落叶木本植物的叶片厚, 而叶肉组织增厚是导致叶片变厚的主要原因.然而, 不同科属的常绿和落叶植物之间的LMA也存在交差和重叠, 这主要是由植物的系统发育决定的; 也就是不同科属之间植物的LMA是有差异的, 而在同一科属中, 常绿植物比落叶植物的LMA高(Poorter et al., 2009; Villar et al., 2013). ...
植物叶寿命及其相关叶性状的生态学研究进展
1
2004
... 20世纪90年代, 很多学者对不同植物的LMA做了大量研究, 并探讨了LVA和LD与LMA之间的关系.研究表明在同一可控环境条件下, 草本植物(Garnier & Laurent, 1994; van Arendonk & Poorter, 1994)和木本植物(Castro-Diez et al., 2000)的LMA主要受LD的影响, Poorter等(2009)对这些数据作了统计和分析, 表明LMA差异的80%由LD决定, 20%由LVA决定.然而在复杂的自然环境条件下, LD和LVA与LMA之间的关系是多样化的(Choong et al., 1992; Niinemets, 1999; Wilson et al., 1999).由此可见, 仅仅把LMA分解为LVA和LD并不能很好地解释导致植物种类之间LMA差异的原因. ...
The impact of long-term water stress on relative growth rate and morphology of needles and shoots of Metasequoia glyptostro- boides seedlings: Research toward identifying mechanistic models.
2
2011
... LMA不仅决定植物潜在的生长速率, 反映植物生态策略, 而且与许多生理过程息息相关(Antúnez et al., 2001; Villar et al., 2006).全球叶片经济型谱的研究明确了LMA与叶片单位干物质的最大光合能力(Amass)密切相关, Amass随LMA的增加有明显的下降趋势.LMA的大小主要由植物叶片的厚度和密度决定(Witkowski & Lamont, 1991), 两者的改变都会影响光的利用效率以及叶肉细胞导度(gm)的大小, 最终导致光合效率的改变(Muir et al., 2014).当受到各种选择压力时, 植物能通过调节LMA来改变光合效率, 使植物的投入和收益重新达到最佳的平衡点.研究表明, LMA是自然选择压力下的稳定遗传的结果, 反映了植物与环境共进化的过程, 具有遗传进化上的意义(Asner et al., 2011; Aspinwall et al., 2013; Flores et al., 2014; Tsujii et al., 2016).因此, 研究LMA具有生理、生态及进化的重要意义.从目前已发表的文章来看, 国内还缺乏关于LMA较深入的综合性报道.本文系统梳理了LMA在叶片整体、组织、细胞三个水平的结构解析和计算方法; 重点分析了LMA对光合作用的影响; 讨论了LMA的内在的遗传差异以及外部的环境胁迫因子(温度、水分、光照)对LMA的影响, 以期为今后更深入的研究提供借鉴和参考. ...
... 要进一步研究引起LMA变化的原因, 需要在组织水平进一步分解LMA.20世纪50年代以来, 很多学者在植物解剖结构方面开展了大量的研究, 表明不同的组织在整个叶片中的比例不一样, 同一组织在叶片中的比例受植物种类和环境的影响(Witkowski & Lamont, 1991; Garnier & Laurent, 1994; Poorter et al., 2009; Villar et al., 2013).在环境梯度上, 植物种间的LMA与其叶片组织特征(叶厚度、叶密度)的变化趋势、变化程度不一致(Fajardo & Piper, 2010; Richardson et al., 2013), 因此要想揭示种间LMA差异的原因, 必须分解LMA.在一些研究中, 常常将组织厚度当作组织体积, 这对于阔叶型的扁平状叶片的组织(叶肉组织或上表皮)是适当的, 但很多植物的叶片本身不规则(如松科、柏科), 而且叶片本身一些组织的形状也不规则, 例如维管组织, 因此将组织体积理解为组织厚度并不合适(Poorter et al., 2009).Roderick等(1999)提出LMA的计算公式中的体积应该是各部分组织的体积加上气室体积.Poorter等(2009)从组织水平提出LMA的计算公式: ...
不同功能型植物光合特性及其与叶氮含量、比叶重的关系
1
2007
... 树木冠层内的LMA的差异也是森林生态系统研究的重要内容.LMA与叶片在冠层中的位置有关, 一般处于冠层顶部的叶片LMA值比冠层底部的高(Koch et al., 2004; Lichtenthaler et al., 2007; Poorter et al., 2009; Weerasinghe et al., 2014).据Koch等(2004)报道100 m高的红杉(Sequoia sempervirens)的冠层底部LMA为150 g·m-2, 而冠层顶部LMA值达800 g·m-2, 这是比较极端的例子.一般来说, 不管是木本植物还是草本植物, 其冠层LMA大约是底层LMA的两倍(Poorter et al., 2009).这种底层和冠层之间的差异在一定程度上是由物种的特异性决定的(Lichtenthaler et al., 2007).此外, LMA还随叶寿命的不同表现出较大的差异.叶寿命是植物在长期适应环境的过程中为获得最大光合生产所形成的适应策略(Reich et al., 1991; 张林和罗天祥, 2004).LMA反映了植物单位叶面积的构建成本, 通常与叶寿命呈正相关关系.LMA低的植物, 其光合效率高, 生长速度快, 环境胁迫应答策略表现为逃避, 因而用于防御投资的物质较少, 叶寿命较短, 如草本植物、落叶乔灌木; 而LMA高的植物, 光合效率慢, 生长速度慢, 环境胁迫应答的策略表现为抵御和耐受, 需要不断积累更多的物质构建保卫构造, 因此表现出较长的叶寿命, 如常绿木本植物(Reich et al., 1991; Wright et al., 2004; Wyka et al., 2012; 王瑞雪等, 2014).常绿木本植物的LMA通常随叶龄的增长而逐渐增加, LMA的增加主要与叶肉细胞的细胞壁变厚、木质素含量增加、叶片变厚、叶片的机械组织含量增加有关(Castro-Díez et al., 2000; Hanba et al., 2002; Wright et al., 2004).Niinemets等(2005, 2006)报道地中海3种阔叶常绿树种月桂(Laurus nobilis)、油橄榄(Olea europea)、冬青栎(Quercus ilex)的LMA随叶龄的增长而增加.而且叶龄对LMA的影响还与叶片在冠层中的位置有关, Han等(2008)报道赤松冠层底部LMA随叶龄的增加而增加的幅度为27%, 但叶龄对冠层顶部LMA无影响.此外, 在常绿木本植物中, 叶寿命不同的植物, 其LMA也有较大的差异, Wyka等(2012)报道松柏类比阔叶类植物的叶寿命长, 其LMA也显著高于阔叶类植物. ...



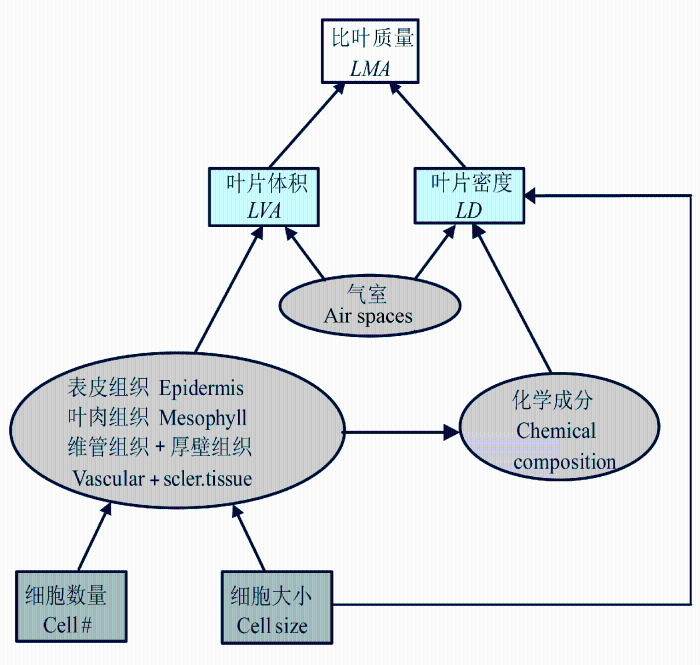
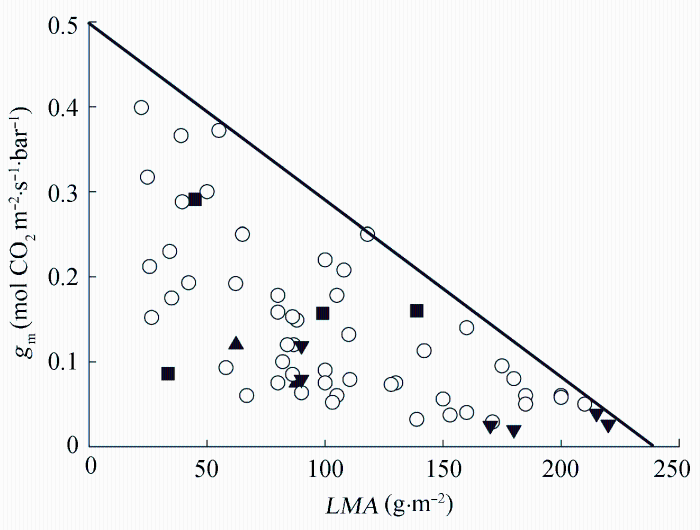
{kind=link}
{kind=link}
{kind=link}
{kind=link}