生态学模式和生态学过程都具有明显的尺度效应(Crawley & Harral, 2001 ; He et al ., 2002 ; Rahbek, 2005 ; Wang et al ., 2008 )。单独一个尺度并不能满足所有科学问题的研究(Ray & Hastings, 1996 ), 例如较小的局部近邻尺度较有利于邻体竞争的研究, 而研究气候对物种多样性的影响时就需要扩展到更大的区域尺度。近些年来, 在生物多样性方面关于尺度效应的研究主要集中在5个方面: (1)不同研究区域内, 最佳取样单元大小、形状以及取样范围的选取(Mouillot & Leprêtre, 1999 ); (2)生物多样性热点地区(biodiversity hotspot)的研究(Bartha et al ., 1998 ; Fortin et al ., 1999 ; Hurlbert & Jetz, 2007 ); (3)生物多样性估算及多样性格局的尺度效应研究(He & Legendre, 1996 ; Wilson et al ., 1999 ; Harte et al ., 2009 ; Colwell et al ., 2012 ; Xu et al ., 2012 ); (4)不同尺度下物种多样性格局形成的机理及相应生态学过程的探究(程佳佳等, 2011 ); (5)物种、生态系统以及景观不同层次多样性之间尺度效应的关系(彭羽等, 2015 )。这些问题的研究对探讨物种多样性的形成和维持机制具有重要意义, 同时也可为生物多样性的保护和有效管理提供科学依据(Willis & Whittaker, 2002 ; Rahbek, 2005 ; Storch et al ., 2007 ; Giladi et al ., 2011 )。
关于物种多样性空间分布格局的尺度效应, 国内外已在热带雨林(He et al ., 2002 ; Kallimanis et al ., 2008 )、温带针阔混交林(Wang et al ., 2008 )、中亚热带喀斯特森林(张忠华等, 2012 )以及南亚热带常绿阔叶林(王志高等, 2008 )中开展过一些研究, 而针对我国热带森林物种多样性的空间分布及其与尺度间关系的研究比较缺乏。海南岛地处热带亚洲北缘, 丰富的热带雨林物种组成使其成为全球生物多样性研究的热点地区之一。本文以海南岛西南部尖峰岭国家级自然保护区内建成的60 hm2 热带山地雨林动态监测样地为数据来源, 探讨了不同空间尺度下各物种多样性指标的空间分布特征, 以期为进一步揭示海南尖峰岭地区物种多样性的形成和维持机制提供科学依据。
1 材料和方法
1.1 研究地概况
尖峰岭地区位于海南省西南部乐东黎族自治县和东方市交界处(18.33°-18.95° N, 108.68°-109.20° E), 总面积约640 km2 。尖峰岭处于低纬度热带岛屿季风气候区, 干湿两季明显, 雨季从5月至10月, 旱季从11月至翌年4月(李意德等, 2012 )。该区年平均降水量2 449 mm, 年平均气温19.8 ℃, 最冷月和最热月平均温度分别为10.8 ℃和27.5 ℃ (蒋有绪和卢俊培, 1991 )。其中热带山地雨林主要分布于海拔600-1200 m的山地丘陵, 砖黄壤-黄壤是主要的土壤类型。热带山地雨林也是尖峰岭地区分布面积最大的植被类型(蒋有绪和卢俊培, 1991 ; 李意德等, 2002 )。
1.2 样地设置及调查
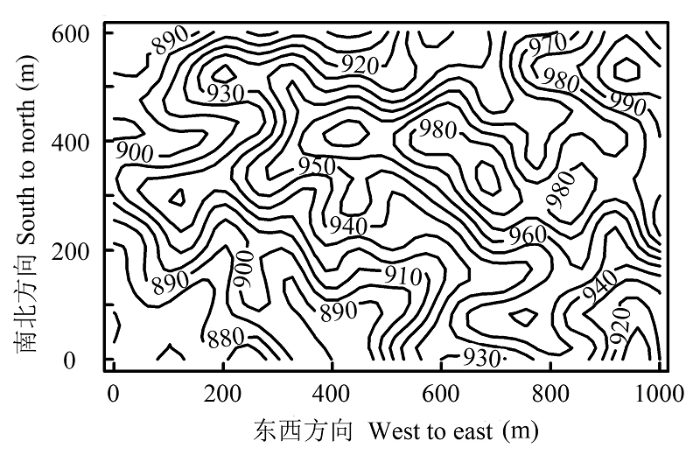
2012年底建设完成的60 hm2 森林生物多样性动态监测样地位于海南尖峰岭五分区的热带山地雨林原始林内, 整个样地严格参照美国史密森热带研究所(Smithsonian Institution)热带森林研究中心(Center for Tropical Forest Science, CTFS)的调查技术规范进行建设(Condit, 1998 )。样地东西长1 000 m, 南北宽600 m (图1 ), 共划分为1 500个20 m × 20 m的样方进行调查, 每个样方四个角均用全站仪测定了其在样方中的坐标位置, 并用水泥桩作了永久标记。样地调查以每个20 m × 20 m样方为单元, 记录了其中所有胸径(DBH )≥1.0 cm的木本植物种名、DBH 和坐标值, 以后每隔5年复查一次。首次调查结果表明, 样地内共记录到439 676株存活的DBH ≥1.0 cm的乔灌木植株, 包括391 686个独立植株、29 103个萌条和18 887个分枝。它们当中除61个植株未确定种名外, 其他分属于62科155属290种(许涵等, 2015 )。
图1 海南尖峰岭60 hm2 森林动态监测样地的地形图。
Fig. 1 Topography of the 60 hm2 Jianfengling Forest Dynamics Plot on Hainan Island.
1.3 数据处理与分析
本研究中, 我们通过对物种丰富度、多度、物种多样性指数(Shannon-Wiener指数、Simpson指数)以及Pielou均匀度指数共5种多样性指标进行分析, 来探讨尖峰岭热带山地雨林物种多样性的空间分布特征及其尺度效应。
整个60 hm2 样地划分为以下6个空间尺度: 5 m × 5 m (24 000个样方)、10 m × 10 m (6 000个样方)、20 m × 20 m (1 500个样方)、40 m × 40 m (375个样方)、100 m × 100 m (60个样方)、200 m × 200 m (15个样方), 分别计算每个尺度下各个样方水平的5种多样性指标。
根据计算结果, 绘制5种物种多样性测度指标的空间分布图, 以对整个样地不同尺度下的物种多样性分布特点及空间变化进行分析。计算每一尺度物种丰富度和多度的Spearman相关系数, 探讨物种丰富度和多度的空间格局随尺度变化的分异。方差和变异系数常常被用来寻找其中的拐点或峰值, 有研究认为这样的拐点或峰值对于确定相应尺度下某种潜在的物理或生态过程甚至群落的特征区域都是很有作用的(Juhász-Nagy & Podani, 1983 ; Horne & Schneider, 1995 )。同时利用多样性指标的方差和变异系数对取样空间尺度(取样面积)的自然对数进行做图, 以探讨尺度变化对物种多样性格局的影响。
各物种多样性指数(Hill, 1973 )以及方差和变异系数的计算公式如下:
Pielou均匀度指数: E = H / ln (S )
方差: δ 2 = (Ʃn j = 1 (Xj - µ )2 ) / (n - 1)
变异系数: CV = (sd /µ ) × 100%
式中Pi 为单个样方内第i 个物种的多度占所有物种多度之和的比例, S 为单个样方内的物种数, n 为样方总数, Xj 为计算的第j 个样方物种多样性指标的数值, µ 为平均值, sd 为标准差。
数据通过R 2.15.3软件(https://cran.usthb.dz/)进行分析。
2 结果
2.1 物种丰富度和多度的空间分布格局
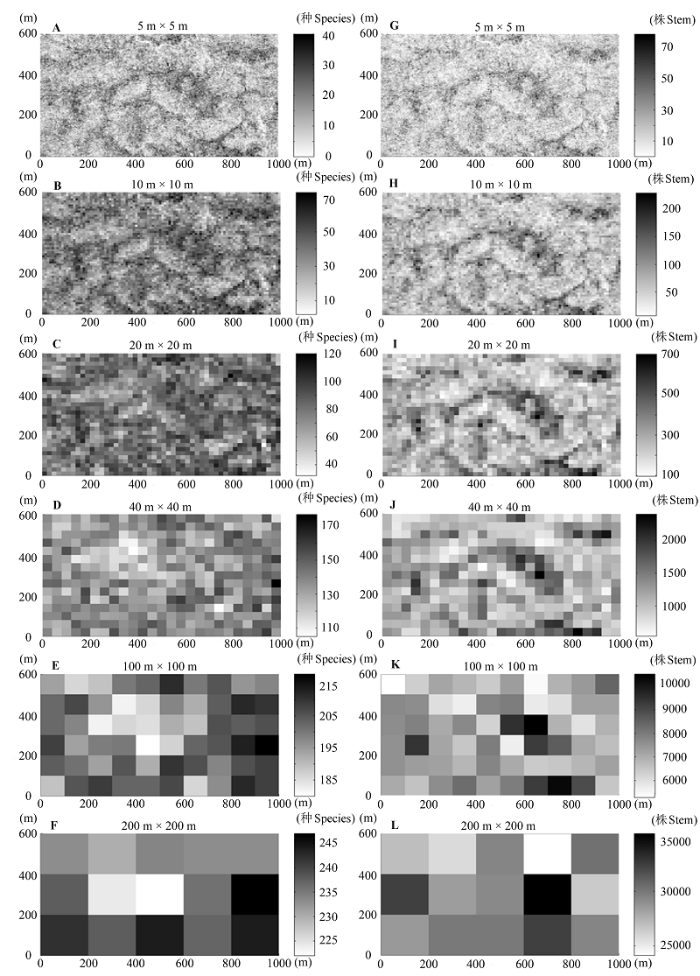
随着取样空间尺度的增加, 物种丰富度和多度也随之增大(图2 )。在小尺度上(小于20 m × 20 m), 两者的空间分布格局表现出相当的一致性, 即两者数值较高的样方均出现在山脊及其附近区域(图1 , 图2 ); 中等尺度上(40 m × 40 m), 这种区域分布的一致性不再明显; 在较大尺度上(大于100 m × 100 m), 两者数值大小趋势上的不一致性在灰度图上表现得更为直观, 例如在200 m × 200 m尺度, 物种丰富度最大的样方, 对应的物种多度却相对较小。此外, 物种丰富度和多度的秩相关分析结果也表明, 在小于40 m × 40 m尺度上, 物种丰富度和多度呈现出极显著(p < 0.001)的正相关关系, 并且相关系数在大于20 m × 20 m尺度时有明显减弱。当尺度超过40 m × 40 m时, 两者的相关性则不再显著(表1 )。
图2 物种丰富度(A-F)和多度(G-L)在不同尺度下的空间分布格局。标尺代表不同样方物种丰富度或多度的数值。
Fig. 2 Spatial distribution patterns of species richness (A-F) and abundance (G-L) at different scales. The sidebars represented the values of species richness or abundance at different plots, respectively.
2.2 物种多样性指数和均匀度指数的空间分布格局
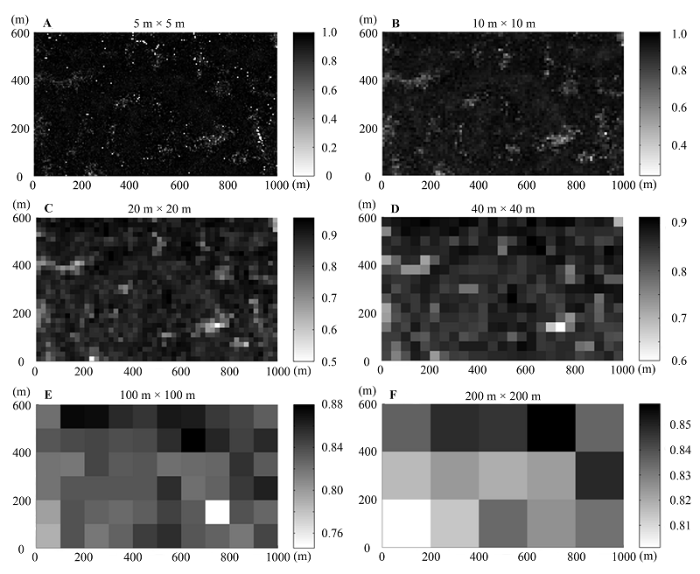
在不同取样空间尺度下, Shannon-Wiener指数也表现出较高的空间异质性, 并且在小于10 m × 10 m尺度上, 其数值较高的样方也多出现在山脊及其附近区域, 随着尺度的增加, 这种现象也变得不再明显(图1 , 图3 )。而Simpson指数和Pielou均匀度指数的空间分布特征表现得较为相似(图3 , 图4 ), 在较小的空间尺度上(小于10 m × 10 m), 各个样方下两者的数值都很高, 且十分相近, 甚至很难从灰度图中区分出数值较高的样方。
图3 Shannon-Wiener指数(A-F)和Simpson指数(G-L)在不同尺度下的空间分布格局。标尺代表不同样方Shannon-Wiener指数或Simpson指数的数值。
Fig. 3 Spatial distribution patterns of Shannon-Wiener (A-F) and Simpson indices (G-L) at different scales. The sidebars represented the values of Shannon-Wiener or Simpson indices at different plots, respectively.
图4 Pielou均匀度指数在不同尺度下的空间分布格局。标尺代表不同样方Pielou均匀度指数的数值。
Fig. 4 Spatial distribution patterns of Pielou’s evenness index at different scales. The sidebars represented the values of Pielou’s evenness index at different plots.
2.3 物种丰富度和多度空间变异随尺度的变化规律
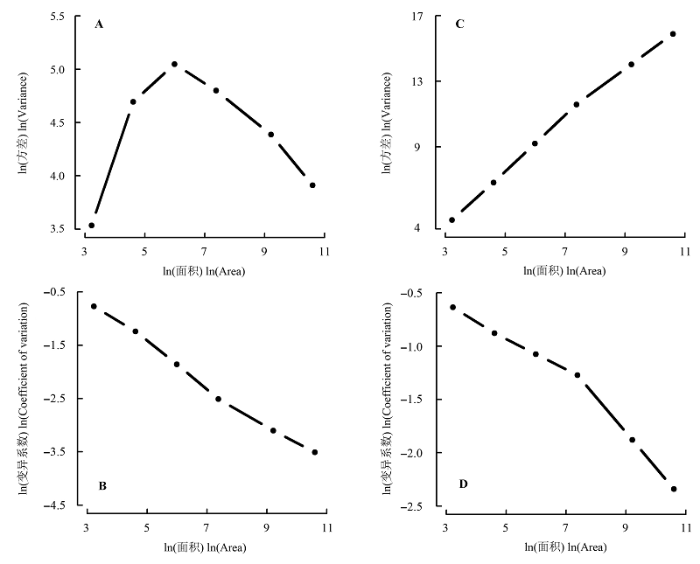
随着取样尺度的增加, 物种丰富度和多度的方差有着明显不同的变化趋势(图5 )。物种丰富度随着尺度的增加表现出单峰型的变化特征, 且在20 m × 20 m尺度上达到最大值, 而物种多度随取样尺度的增加逐渐增大, 呈现出明显的线性关系。物种丰富度和多度的变异系数均随着尺度的增加而下降, 其中物种丰富度的变异系数随着尺度的增加呈现出线性减小的趋势; 而多度在小于40 m × 40 m尺度上线性平缓减小, 之后则急剧减小。
图5 物种丰富度(A, B)和多度(C, D)在不同尺度下的空间变异。
Fig. 5 Spatial variation of species richness (A, B) and abundance (C, D) at different scales.
2.4 物种多样性指数和均匀度指数空间变异随尺度的变化规律
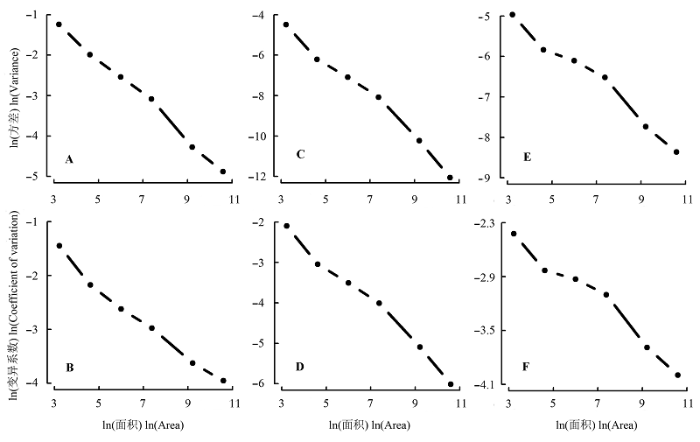
Shannon-Wiener指数、Simpson指数和Pielou均匀度指数的方差和变异系数均呈现出随着取样空间尺度的增加而减小的趋势(图6 )。尽管如此, 相比Shannon-Wiener指数而言, 各取样尺度下的Simpson指数和Pielou均匀度指数的方差要小得多; 而变异系数为标准差与平均数的比值, 三者的差异并不 明显。
图6 Shannon-Wiener指数(A, B)、Simpson指数(C, D)以及Pielou均匀度指数(E, F)在不同尺度下的空间变异。
Fig. 6 Spatial variation of Shannon-Wiener (A, B), Simpson (C, D) and Pielou’s evenness indices (E, F) at different scales.
3 讨论
3.1 物种丰富度和多度格局随空间尺度的变化
物种丰富度和多度的分布格局都是基于空间尺度存在的。在小尺度上, 两者的空间分布表现出相当的一致性, 即两者数值较高的样方均出现在山脊及其附近区域, 随着尺度的增加, 这种一致性则不再明显或消失。秩相关分析结果也表明, 在小于40 m × 40 m尺度上, 物种丰富度和多度呈现出极显著(p < 0.001)的正相关关系, 并且相关系数在大于20 m × 20 m尺度时有明显减弱, 当尺度超过40 m × 40 m时, 两者的相关性则不再显著, 这可能反映了决定生物多样性格局的机制随尺度增加而发生了变化。小尺度物种丰富度主要与样方内包含的木本植物个体数量有关(Preston, 1962 ), 随着尺度增加, 生境的异质性增加, 物种丰富度随面积的增加主要是由于新生境的出现(Turner & Tjørve, 2005 ), 因而物种丰富度和多度的相关性逐渐变得不显著。以往研究发现, 尖峰岭热带山地雨林60 hm2 大样地内, 不同深度土层的13项土壤化学性质(全氮、全磷、全钾、有效氮、有效磷和速效钾等)以及8项土壤物理性质(土壤容重、土壤含水量和土壤总孔隙等)空间自相关变程大多都在20-40 m之间(时雷雷, 2012 )。这反映了土壤属性的空间自相关范围, 当尺度较小, 处于变程内时, 各样方间的空间变异很小, 所提供的生境类型在一个比较稳定的水平, 物种丰富度和多度表现出了极显著的正相关, 当尺度超过变程时, 土壤性质的分布则更多体现为一种随机性, 此时两者的相关性逐渐减弱甚至消失。
3.2 物种丰富度和多度方差与变异系数的比较
尖峰岭60 hm2 热带山地雨林样地中, 物种丰富度和多度的方差随着空间尺度的变化表现出不同的趋势。物种丰富度的方差随取样尺度增加呈现出单峰分布特征, 并且在20 m × 20 m尺度上出现了峰值, 而产生这一峰值的尺度与物种丰富度和多度格局的分离尺度相一致, 这也在一定程度上印证了生态学家通过方差分析中产生的拐点来确定相应尺度下潜在的生态过程的合理性。类似的峰型曲线同样出现在其他地区的热带森林(He et al ., 2002 )、温带森林(Wang et al ., 2008 )以及中亚热带的喀斯特森林 (张忠华等, 2012 )中。而物种多度的方差则是随取样尺度的增加呈线性增加趋势, 意味着不同尺度下影响物种多度空间分布的生态过程变化不大。与方差分析不同, 物种丰富度和多度的变异系数均随取样尺度的增加而逐渐减小, 这主要是因为方差反映了多样性测度的绝对变异程度, 而变异系数则反映了多样性测度的相对变异程度。这些研究结果表明多样性的绝对变异更能反映出生态过程对生物多样性格局的影响。
3.3 取样尺度对物种多样性指数和均匀度指数空间分布的影响
Shannon-Wiener指数、Simpson指数和Pielou均匀度指数同样存在尺度效应, 这种尺度效应因研究区域不同而有所差异, 但在同一研究区域内较为一致。本研究中, 随着取样尺度的增加, 3种多样性指数的方差和变异系数均呈现出减小的趋势。而在南亚热带鼎湖山20 hm2 常绿阔叶林内开展的类似研究中, 3种多样性指数的方差随着取样尺度的增加都表现出单峰型的变化特征(王志高等, 2008 ), 这可能与两个样地内不同的稀有种组成有很大关系。尖峰岭样地内的稀有物种数量为60种, 占总物种数的20.7% (许涵等, 2015 ), 而鼎湖山样地内的稀有种数量为110种, 占总物种数的52.4% (叶万辉等, 2008 )。位于马来西亚的Pasoh热带雨林50 hm2 样地, 其稀有种数量为301种, 占总种数的36.5%。He等(1994)在该区域的研究发现, 相对较小以及较大尺度而言, 稀有种对Shannon-Wiener指数尺度效应的影响仅在10 m × 10 m到30 m × 30 m的尺度区间才表现明显。鼎湖山样地以及Pasoh样地中稀有种的组成较尖峰岭样地要高很多, 这很可能是导致物种多样性指数空间分布格局产生差异的原因之一。此外本研究中, 不同取样空间尺度下, Shannon- Wiener指数具有较大的空间变异, 而Simpson指数与Pielou均匀度指数的变异均较小。这与Shannon- Wiener指数和Simpson指数自身的性质有关, Shannon-Wiener指数与物种丰富度的关系最为密切, 而Simpson指数则对均匀度更为敏感(马克平等, 1995 ; 许晴等, 2011 )。
总的来说, 本文从空间尺度的层面上探讨了物种多样性的空间分布格局, 但是对于这种变化过程的内在机制有待进行更为深入的分析, 比如生境异质性是如何以及在多大程度上影响物种的空间分布。海南尖峰岭样地地形较为复杂, 整个样地20 m × 20 m样方水平上的海拔和坡度分别在866.3-1 016.7 m和1.7°-49.3°间变化, 许涵等(2015) 在该样地的研究发现有些物种的分布明显受地形影响, 例如变色山槟榔(Pinanga baviensis )和柏拉木(Blastus cochichinensis )经常在沟谷地区聚集分布, 而有些种则与土壤养分有紧密关系, 例如豆科的长脐红豆(Ormosia balansae ), 有些物种似乎受其他因素包括微生物等影响, 例如海南蕈树(Altingia obovata )。物种的生境偏好直接影响着物种多样性的空间分布格局, 未来对这些物种的研究, 有助于进一步揭示尖峰岭地区物种多样性的形成和维持机制。
致谢 感谢尖峰岭国家级森林生态站运行费资助本研究。感谢中国林业科学研究院的蒋有绪院士、台湾东海大学的孙义方教授、中山大学的李步杭博士和加拿大阿尔伯塔大学的何芳良教授在样地建设过程中给予的帮助, 感谢尖峰岭国家级自然保护区和尖峰岭林业局的莫锦华局长、蒋忠亮主任等的大力支持, 感谢北京大学马素辉博士在论文做图方面提出的宝贵意见, 感谢余传文、彭儒明、黄凤林等在大样地调查中付出的辛勤工作。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Bartha S Czárán T Podani J 1998 ). Exploring plant community dynamics in abstract coenostate spaces
.Abstracta Botanica , 22 , 49 -66 .
[本文引用: 1]
[2]
Cheng JJ Mi XC Ma KP Zhang JT 2011 ). Responses of species-abundance distribution to varying sampling scales in a subtropical broad-leaved forest
.Biodiversity Science , 19 , 168 -177 . (in Chinese with English abstract)[程佳佳 , 米湘成 , 马克平 , 张金屯 (2011 ). 亚热带常绿阔叶林群落物种多度分布格局
. 生物多样性 , 19 , 168 -177 .]
[本文引用: 1]
[3]
Colwell RK Chao A Gotelli NJ Lin SY Mao CX Chazdon RL Longino JT 2012 ). Models and estimators linking individual-based and sample-based rarefaction, extrapolation, and comparison of assemblages
.Journal of Plant Ecology , 5 , 3 -21 .
[本文引用: 1]
[4]
Condit R 1998 ). Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and a Comparison with Other Plots
. Springer, Berlin.
[本文引用: 1]
[5]
Crawley MJ Harral JE 2001 ). Scale dependence in plant biodiversity
.Science , 291 , 864 -868 .
[本文引用: 1]
[6]
Fortin MJ Payette S Marineau K 1999 ). Spatial vegetation diversity index along a postfire successional gradient in the northern boreal forest
.Ecoscience , 6 , 204 -213 .
[本文引用: 1]
[7]
Giladi I Ziv Y May F Jeltsch F 2011 ). Scale-dependent determinants of plant species richness in a semi-arid fragmented agro-ecosystem
.Journal of Vegetation Science , 22 , 983 -996 .
[本文引用: 1]
[8]
Harte J Smith AB Storch D 2009 ). Biodiversity scales from plots to biomes with a universal species-area curve
.Ecology Letters , 12 , 789 -797 .
[本文引用: 1]
[9]
He FL LaFrankie JV Song B 2002 ). Scale dependence of tree abundance and richness in a tropical rain forest, Malaysia
.Landscape Ecology , 17 , 559 -568 .
[本文引用: 3]
[10]
He FL Legendre P 1996 ). On species-area relations
.The American Naturalist , 148 , 719 -737 .
[本文引用: 1]
[11]
He FL Legendre P Bellehumeur C LaFrankie JV 1994 ). Diversity pattern and spatial scale: A study of a tropical rain forest of Malaysia
.Environmental and Ecological Statistics , 1 , 265 -286 .
[12]
Hill MO 1973 ). Diversity and evenness: A unifying notation and its consequences
.Ecology , 54 , 427 -432 .
[本文引用: 1]
[13]
Horne JK Schneider DC 1995 ). Spatial variance in ecology
.Oikos , 74 , 18 -26 .
[本文引用: 1]
[14]
Hurlbert AH Jetz W 2007 ). Species richness, hotspots, and the scale dependence of range maps in ecology and conserva- tion
.Proceedings of the National Academy of Sciences of the United States of America , 14 , 13384 -13389 .
[本文引用: 1]
[15]
Jiang YX L JP (1991 ). Tropical Forest Ecosystem of Jian- fengling Mountain in Hainan Island, China . Science Press , Beijing . (in Chinese)[蒋有绪 , 卢俊培 (1991 ). 中国海南岛尖峰岭热带林生态系统 . 科学出版社 , 北京 .]
[本文引用: 2]
[16]
Juhász-Nagy P Podani J 1983 ). Information theory methods for the study of spatial processes and succession
.Plant Ecology , 51 , 129 -140 .
[本文引用: 1]
[17]
Kallimanis AS Halley JM Vokou D Sgardelis SP 2008 ). The scale of analysis determines the spatial pattern of woody species diversity in the Mediterranean environment
.Plant Ecology , 196 , 143 -151 .
[本文引用: 1]
[18]
Li YD Chen BF Zhou GY Wu ZM Zeng QB Luo TS Huang SN Xie MD Huang Q 2002 ). Research and Conservation of Tropical Forest and the Biodiversity: A Special Reference to Hainan Island, China . China Forestry Publishing House , Beijing . 47 -62 . (in Chinese)[李意德 , 陈步峰 , 周光益 , 吴仲民 , 曾庆波 , 骆土寿 , 黄世能 , 谢明东 , 黄全 (2002 ). 中国海南岛热带森林及其生物多样性保护研究 . 中国林业出版社 , 北京 . 47 -62 .]
[本文引用: 2]
[19]
Li YD Xu H Luo TS Chen DX Lin MX 2012 ). Permanent Monitoring and Research Dataset of Chinese Ecosystem: Forest Ecosystem: Jianfengling Station (Bio-species Checklist) . China Agriculture Press , Beijing . 10 -35 . (in Chinese)[李意德 , 许涵 , 骆土寿 , 陈德祥 , 林明献 (2012 ). 中国生态系统定位观测与研究数据集: 森林生态系统卷: 海南尖峰岭站(生物物种数据集) . 中国农业出版社 , 北京 . 10 -35 .]
[20]
Ma KP Huang JH Yu SL Chen LZ 1995 ). Plant community diversity in Dongling Mountain, Beijing, China: II, species richness, evenness and species diversities
.Acta Ecologica Sinica , 15 , 268 -277 . (in Chinese with English abstract)[马克平 , 黄建辉 , 于顺利 , 陈灵芝 (1995 ). 北京东灵山地区植物群落多样性的研究: II丰富度、均匀度和物种多样性
. 生态学报 , 15 , 268 -277 .]
[本文引用: 1]
[21]
Mouillot D Leprêtre A 1999 ). A comparison of species diversity estimators
.Researches on Population Ecology , 41 , 203 -215 .
[本文引用: 1]
[22]
Peng Y Qing FT Mi K Xue DY 2015 ). Study progress on spatial scale effects and coupling relationships of different levels in biodiversity
.Acta Ecologica Sinica , 35 , 577 -583 . (in Chinese with English abstract)[彭羽 , 卿凤婷 , 米凯 , 薛达元 (2015 ). 生物多样性不同层次尺度效应及其耦合关系研究进展
. 生态学报 , 35 , 577 -583 .]
[本文引用: 1]
[23]
Preston FW 1962 ). The canonical distribution of commonness and rarity: Part I.Ecology , 43 , 185 -215 .
[本文引用: 1]
[24]
Rahbek C 2005 ). The role of spatial scale and the perception of large-scale species-richness patterns
.Ecology Letters , 8 , 224 -239 .
[本文引用: 2]
[25]
Ray C Hastings A 1996 ). Density dependence: Are we searching at the wrong spatial scale?
Journal of Animal Ecology , 65 , 556 -566 .
[本文引用: 1]
[26]
Shi LL 2012 ). Study on the Spatial Heterogeneity of Soil Physical and Chemical Properties of Primary Tropical Montane Rainforest in Jianfengling, Hainan Island
. Master degree dissertation, Chinese Academy of Forestry , Beijing . 26 -41 . (in Chinese with English abstract)[时雷雷 (2012 ). 海南尖峰岭热带山地雨林原始林土壤理化性质空间异质性研究
. 硕士学位论文, 中国林业科学研究院, 北京 . 26 -41 .]
[本文引用: 1]
[27]
Storch D Marquet PA Brown JH 2007 ). Scaling Biodiversity . Cambridge University Press , Cambridge , UK . 347 -375 .
[本文引用: 1]
[28]
Turner WR Tjørve E 2005 ). Scale-dependence in species-area relationships
.Ecography , 28 , 721 -730 .
[本文引用: 1]
[29]
Wang XG Hao ZQ Ye J Li BH Yao XL 2008 ). Spatial variation of species diversity across scale in an old-growth temperate forest of China
.Ecological Research , 23 , 709 -717 .
[本文引用: 3]
[30]
Wang ZG Ye WH Cao HL Lian JY 2008 ). Spatial distribu- tion of species diversity indices in a monsoon evergreen broadleaved forest at Dinghushan Mountain
.Biodiversity Science , 16 , 454 -461 . (in Chinese with English abstract)[王志高 , 叶万辉 , 曹洪麟 , 练琚愉 (2008 ). 鼎湖山季风常绿阔叶林物种多样性指数空间分布特征
. 生物多样性 , 16 , 454 -461 .]
[本文引用: 2]
[31]
Willis KJ Whittaker RJ 2002 ). Species diversity-scale matters
.Science , 295 , 1245 -1248 .
[本文引用: 1]
[32]
Wilson JB Steel JB King WM Gitay H 1999 ). The effect of spatial scale on evenness
.Journal of Vegetation Science , 10 , 463 -468 .
[本文引用: 1]
[33]
Xu H Li YD Luo TS Chen DX Lin MX Wu JH Li YP Yang H Zhou Z 2015 ). Jianfengling Tropical Mountain Rain Forest Dynamic Plot: Community Characteristic, Tree Species and Their Distribution Patterns . China Forestry Publishing House , Beijing . 34 -48 . (in Chinese)[许涵 , 李意德 , 骆土寿 , 陈德祥 , 林明献 , 吴建辉 , 李艳朋 , 杨怀 , 周璋 (2015 ). 海南尖峰岭热带山地雨林——群落特征、树种及其分布格局 . 中国林业出版社 , 北京 . 34 -48 .]
[本文引用: 3]
[34]
Xu H Liu SR Li YD Zang RG He FL 2012 ). Assessing non-parametric and area-based methods for estimating regional species richness
.Journal of Vegetation Science , 23 , 1006 -1012 .
[本文引用: 1]
[35]
Xu Q Zhang F Xu ZQ Jia YL You JM 2011 ). Some characteristics of Simpson index and the Shannon-Wiener index and their dilution effect
.Pratacultural Science , 4 , 527 -531 . (in Chinese with English abstract)[许晴 , 张放 , 许中旗 , 贾彦龙 , 尤建民 (2011 ). Simpson指数和Shannon-Wiener指数若干特征的分析及“稀释效应”
. 草业科学 , 4 , 527 -531 .]
[本文引用: 1]
[36]
Ye WH Cao HL Huang ZL Lian JY Wang ZG Li L Wei SG Wang ZM 2008 ). Community structure of a 20 hm2 lower subtropical evergreen broadleaved forest plot in Dinghushan, China. Journal of Plant Ecology (Chinese Version)
, 32 , 274 -286 . (in Chinese with English abstract)[叶万辉 , 曹洪麟 , 黄忠良 , 练琚愉 , 王志高 , 李林 , 魏识广 , 王章明 (2008 ). 鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究
. 植物生态学报 , 32 , 274 -286 .]
[本文引用: 1]
[37]
Zhang ZH Hu G Zhu JD Ni J 2012 ). Scale-dependent spatial variation of species abundance and richness in two mixed evergreen-deciduous broad-leaved karst forests, Southwest China
.Acta Ecologica Sinica , 32 , 5663 -5672 . (in Chinese with English abstract)[张忠华 , 胡刚 , 祝介东 , 倪健 (2012 ). 喀斯特常绿落叶阔叶混交林物种多度与丰富度空间分布的尺度效应
. 生态学报 , 32 , 5663 -5672 .]
[本文引用: 2]
Exploring plant community dynamics in abstract coenostate spaces
1
1998
... 生态学模式和生态学过程都具有明显的尺度效应(Crawley & Harral, 2001 ; He et al ., 2002 ; Rahbek, 2005 ; Wang et al ., 2008 ).单独一个尺度并不能满足所有科学问题的研究(Ray & Hastings, 1996 ), 例如较小的局部近邻尺度较有利于邻体竞争的研究, 而研究气候对物种多样性的影响时就需要扩展到更大的区域尺度.近些年来, 在生物多样性方面关于尺度效应的研究主要集中在5个方面: (1)不同研究区域内, 最佳取样单元大小、形状以及取样范围的选取(Mouillot & Leprêtre, 1999 ); (2)生物多样性热点地区(biodiversity hotspot)的研究(Bartha et al ., 1998 ; Fortin et al ., 1999 ; Hurlbert & Jetz, 2007 ); (3)生物多样性估算及多样性格局的尺度效应研究(He & Legendre, 1996 ; Wilson et al ., 1999 ; Harte et al ., 2009 ; Colwell et al ., 2012 ; Xu et al ., 2012 ); (4)不同尺度下物种多样性格局形成的机理及相应生态学过程的探究(程佳佳等, 2011 ); (5)物种、生态系统以及景观不同层次多样性之间尺度效应的关系(彭羽等, 2015 ).这些问题的研究对探讨物种多样性的形成和维持机制具有重要意义, 同时也可为生物多样性的保护和有效管理提供科学依据(Willis & Whittaker, 2002 ; Rahbek, 2005 ; Storch et al ., 2007 ; Giladi et al ., 2011 ). ...
亚热带常绿阔叶林群落物种多度分布格局
1
2011
... 生态学模式和生态学过程都具有明显的尺度效应(Crawley & Harral, 2001 ; He et al ., 2002 ; Rahbek, 2005 ; Wang et al ., 2008 ).单独一个尺度并不能满足所有科学问题的研究(Ray & Hastings, 1996 ), 例如较小的局部近邻尺度较有利于邻体竞争的研究, 而研究气候对物种多样性的影响时就需要扩展到更大的区域尺度.近些年来, 在生物多样性方面关于尺度效应的研究主要集中在5个方面: (1)不同研究区域内, 最佳取样单元大小、形状以及取样范围的选取(Mouillot & Leprêtre, 1999 ); (2)生物多样性热点地区(biodiversity hotspot)的研究(Bartha et al ., 1998 ; Fortin et al ., 1999 ; Hurlbert & Jetz, 2007 ); (3)生物多样性估算及多样性格局的尺度效应研究(He & Legendre, 1996 ; Wilson et al ., 1999 ; Harte et al ., 2009 ; Colwell et al ., 2012 ; Xu et al ., 2012 ); (4)不同尺度下物种多样性格局形成的机理及相应生态学过程的探究(程佳佳等, 2011 ); (5)物种、生态系统以及景观不同层次多样性之间尺度效应的关系(彭羽等, 2015 ).这些问题的研究对探讨物种多样性的形成和维持机制具有重要意义, 同时也可为生物多样性的保护和有效管理提供科学依据(Willis & Whittaker, 2002 ; Rahbek, 2005 ; Storch et al ., 2007 ; Giladi et al ., 2011 ). ...
Models and estimators linking individual-based and sample-based rarefaction, extrapolation, and comparison of assemblages
1
2012
... 生态学模式和生态学过程都具有明显的尺度效应(Crawley & Harral, 2001 ; He et al ., 2002 ; Rahbek, 2005 ; Wang et al ., 2008 ).单独一个尺度并不能满足所有科学问题的研究(Ray & Hastings, 1996 ), 例如较小的局部近邻尺度较有利于邻体竞争的研究, 而研究气候对物种多样性的影响时就需要扩展到更大的区域尺度.近些年来, 在生物多样性方面关于尺度效应的研究主要集中在5个方面: (1)不同研究区域内, 最佳取样单元大小、形状以及取样范围的选取(Mouillot & Leprêtre, 1999 ); (2)生物多样性热点地区(biodiversity hotspot)的研究(Bartha et al ., 1998 ; Fortin et al ., 1999 ; Hurlbert & Jetz, 2007 ); (3)生物多样性估算及多样性格局的尺度效应研究(He & Legendre, 1996 ; Wilson et al ., 1999 ; Harte et al ., 2009 ; Colwell et al ., 2012 ; Xu et al ., 2012 ); (4)不同尺度下物种多样性格局形成的机理及相应生态学过程的探究(程佳佳等, 2011 ); (5)物种、生态系统以及景观不同层次多样性之间尺度效应的关系(彭羽等, 2015 ).这些问题的研究对探讨物种多样性的形成和维持机制具有重要意义, 同时也可为生物多样性的保护和有效管理提供科学依据(Willis & Whittaker, 2002 ; Rahbek, 2005 ; Storch et al ., 2007 ; Giladi et al ., 2011 ). ...
Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and a Comparison with Other Plots
1
1998
... 2012年底建设完成的60 hm2 森林生物多样性动态监测样地位于海南尖峰岭五分区的热带山地雨林原始林内, 整个样地严格参照美国史密森热带研究所(Smithsonian Institution)热带森林研究中心(Center for Tropical Forest Science, CTFS)的调查技术规范进行建设(Condit, 1998 ).样地东西长1 000 m, 南北宽600 m (图1 ), 共划分为1 500个20 m × 20 m的样方进行调查, 每个样方四个角均用全站仪测定了其在样方中的坐标位置, 并用水泥桩作了永久标记.样地调查以每个20 m × 20 m样方为单元, 记录了其中所有胸径(DBH )≥1.0 cm的木本植物种名、DBH 和坐标值, 以后每隔5年复查一次.首次调查结果表明, 样地内共记录到439 676株存活的DBH ≥1.0 cm的乔灌木植株, 包括391 686个独立植株、29 103个萌条和18 887个分枝.它们当中除61个植株未确定种名外, 其他分属于62科155属290种(许涵等, 2015 ). ...
Scale dependence in plant biodiversity
1
2001
... 生态学模式和生态学过程都具有明显的尺度效应(Crawley & Harral, 2001 ; He et al ., 2002 ; Rahbek, 2005 ; Wang et al ., 2008 ).单独一个尺度并不能满足所有科学问题的研究(Ray & Hastings, 1996 ), 例如较小的局部近邻尺度较有利于邻体竞争的研究, 而研究气候对物种多样性的影响时就需要扩展到更大的区域尺度.近些年来, 在生物多样性方面关于尺度效应的研究主要集中在5个方面: (1)不同研究区域内, 最佳取样单元大小、形状以及取样范围的选取(Mouillot & Leprêtre, 1999 ); (2)生物多样性热点地区(biodiversity hotspot)的研究(Bartha et al ., 1998 ; Fortin et al ., 1999 ; Hurlbert & Jetz, 2007 ); (3)生物多样性估算及多样性格局的尺度效应研究(He & Legendre, 1996 ; Wilson et al ., 1999 ; Harte et al ., 2009 ; Colwell et al ., 2012 ; Xu et al ., 2012 ); (4)不同尺度下物种多样性格局形成的机理及相应生态学过程的探究(程佳佳等, 2011 ); (5)物种、生态系统以及景观不同层次多样性之间尺度效应的关系(彭羽等, 2015 ).这些问题的研究对探讨物种多样性的形成和维持机制具有重要意义, 同时也可为生物多样性的保护和有效管理提供科学依据(Willis & Whittaker, 2002 ; Rahbek, 2005 ; Storch et al ., 2007 ; Giladi et al ., 2011 ). ...
Spatial vegetation diversity index along a postfire successional gradient in the northern boreal forest
1
1999
... 生态学模式和生态学过程都具有明显的尺度效应(Crawley & Harral, 2001 ; He et al ., 2002 ; Rahbek, 2005 ; Wang et al ., 2008 ).单独一个尺度并不能满足所有科学问题的研究(Ray & Hastings, 1996 ), 例如较小的局部近邻尺度较有利于邻体竞争的研究, 而研究气候对物种多样性的影响时就需要扩展到更大的区域尺度.近些年来, 在生物多样性方面关于尺度效应的研究主要集中在5个方面: (1)不同研究区域内, 最佳取样单元大小、形状以及取样范围的选取(Mouillot & Leprêtre, 1999 ); (2)生物多样性热点地区(biodiversity hotspot)的研究(Bartha et al ., 1998 ; Fortin et al ., 1999 ; Hurlbert & Jetz, 2007 ); (3)生物多样性估算及多样性格局的尺度效应研究(He & Legendre, 1996 ; Wilson et al ., 1999 ; Harte et al ., 2009 ; Colwell et al ., 2012 ; Xu et al ., 2012 ); (4)不同尺度下物种多样性格局形成的机理及相应生态学过程的探究(程佳佳等, 2011 ); (5)物种、生态系统以及景观不同层次多样性之间尺度效应的关系(彭羽等, 2015 ).这些问题的研究对探讨物种多样性的形成和维持机制具有重要意义, 同时也可为生物多样性的保护和有效管理提供科学依据(Willis & Whittaker, 2002 ; Rahbek, 2005 ; Storch et al ., 2007 ; Giladi et al ., 2011 ). ...
Scale-dependent determinants of plant species richness in a semi-arid fragmented agro-ecosystem
1
2011
... 生态学模式和生态学过程都具有明显的尺度效应(Crawley & Harral, 2001 ; He et al ., 2002 ; Rahbek, 2005 ; Wang et al ., 2008 ).单独一个尺度并不能满足所有科学问题的研究(Ray & Hastings, 1996 ), 例如较小的局部近邻尺度较有利于邻体竞争的研究, 而研究气候对物种多样性的影响时就需要扩展到更大的区域尺度.近些年来, 在生物多样性方面关于尺度效应的研究主要集中在5个方面: (1)不同研究区域内, 最佳取样单元大小、形状以及取样范围的选取(Mouillot & Leprêtre, 1999 ); (2)生物多样性热点地区(biodiversity hotspot)的研究(Bartha et al ., 1998 ; Fortin et al ., 1999 ; Hurlbert & Jetz, 2007 ); (3)生物多样性估算及多样性格局的尺度效应研究(He & Legendre, 1996 ; Wilson et al ., 1999 ; Harte et al ., 2009 ; Colwell et al ., 2012 ; Xu et al ., 2012 ); (4)不同尺度下物种多样性格局形成的机理及相应生态学过程的探究(程佳佳等, 2011 ); (5)物种、生态系统以及景观不同层次多样性之间尺度效应的关系(彭羽等, 2015 ).这些问题的研究对探讨物种多样性的形成和维持机制具有重要意义, 同时也可为生物多样性的保护和有效管理提供科学依据(Willis & Whittaker, 2002 ; Rahbek, 2005 ; Storch et al ., 2007 ; Giladi et al ., 2011 ). ...
Biodiversity scales from plots to biomes with a universal species-area curve
1
2009
... 生态学模式和生态学过程都具有明显的尺度效应(Crawley & Harral, 2001 ; He et al ., 2002 ; Rahbek, 2005 ; Wang et al ., 2008 ).单独一个尺度并不能满足所有科学问题的研究(Ray & Hastings, 1996 ), 例如较小的局部近邻尺度较有利于邻体竞争的研究, 而研究气候对物种多样性的影响时就需要扩展到更大的区域尺度.近些年来, 在生物多样性方面关于尺度效应的研究主要集中在5个方面: (1)不同研究区域内, 最佳取样单元大小、形状以及取样范围的选取(Mouillot & Leprêtre, 1999 ); (2)生物多样性热点地区(biodiversity hotspot)的研究(Bartha et al ., 1998 ; Fortin et al ., 1999 ; Hurlbert & Jetz, 2007 ); (3)生物多样性估算及多样性格局的尺度效应研究(He & Legendre, 1996 ; Wilson et al ., 1999 ; Harte et al ., 2009 ; Colwell et al ., 2012 ; Xu et al ., 2012 ); (4)不同尺度下物种多样性格局形成的机理及相应生态学过程的探究(程佳佳等, 2011 ); (5)物种、生态系统以及景观不同层次多样性之间尺度效应的关系(彭羽等, 2015 ).这些问题的研究对探讨物种多样性的形成和维持机制具有重要意义, 同时也可为生物多样性的保护和有效管理提供科学依据(Willis & Whittaker, 2002 ; Rahbek, 2005 ; Storch et al ., 2007 ; Giladi et al ., 2011 ). ...
Scale dependence of tree abundance and richness in a tropical rain forest, Malaysia
3
2002
... 生态学模式和生态学过程都具有明显的尺度效应(Crawley & Harral, 2001 ; He et al ., 2002 ; Rahbek, 2005 ; Wang et al ., 2008 ).单独一个尺度并不能满足所有科学问题的研究(Ray & Hastings, 1996 ), 例如较小的局部近邻尺度较有利于邻体竞争的研究, 而研究气候对物种多样性的影响时就需要扩展到更大的区域尺度.近些年来, 在生物多样性方面关于尺度效应的研究主要集中在5个方面: (1)不同研究区域内, 最佳取样单元大小、形状以及取样范围的选取(Mouillot & Leprêtre, 1999 ); (2)生物多样性热点地区(biodiversity hotspot)的研究(Bartha et al ., 1998 ; Fortin et al ., 1999 ; Hurlbert & Jetz, 2007 ); (3)生物多样性估算及多样性格局的尺度效应研究(He & Legendre, 1996 ; Wilson et al ., 1999 ; Harte et al ., 2009 ; Colwell et al ., 2012 ; Xu et al ., 2012 ); (4)不同尺度下物种多样性格局形成的机理及相应生态学过程的探究(程佳佳等, 2011 ); (5)物种、生态系统以及景观不同层次多样性之间尺度效应的关系(彭羽等, 2015 ).这些问题的研究对探讨物种多样性的形成和维持机制具有重要意义, 同时也可为生物多样性的保护和有效管理提供科学依据(Willis & Whittaker, 2002 ; Rahbek, 2005 ; Storch et al ., 2007 ; Giladi et al ., 2011 ). ...
... 关于物种多样性空间分布格局的尺度效应, 国内外已在热带雨林(He et al ., 2002 ; Kallimanis et al ., 2008 )、温带针阔混交林(Wang et al ., 2008 )、中亚热带喀斯特森林(张忠华等, 2012 )以及南亚热带常绿阔叶林(王志高等, 2008 )中开展过一些研究, 而针对我国热带森林物种多样性的空间分布及其与尺度间关系的研究比较缺乏.海南岛地处热带亚洲北缘, 丰富的热带雨林物种组成使其成为全球生物多样性研究的热点地区之一.本文以海南岛西南部尖峰岭国家级自然保护区内建成的60 hm2 热带山地雨林动态监测样地为数据来源, 探讨了不同空间尺度下各物种多样性指标的空间分布特征, 以期为进一步揭示海南尖峰岭地区物种多样性的形成和维持机制提供科学依据. ...
... 尖峰岭60 hm2 热带山地雨林样地中, 物种丰富度和多度的方差随着空间尺度的变化表现出不同的趋势.物种丰富度的方差随取样尺度增加呈现出单峰分布特征, 并且在20 m × 20 m尺度上出现了峰值, 而产生这一峰值的尺度与物种丰富度和多度格局的分离尺度相一致, 这也在一定程度上印证了生态学家通过方差分析中产生的拐点来确定相应尺度下潜在的生态过程的合理性.类似的峰型曲线同样出现在其他地区的热带森林(He et al ., 2002 )、温带森林(Wang et al ., 2008 )以及中亚热带的喀斯特森林 (张忠华等, 2012 )中.而物种多度的方差则是随取样尺度的增加呈线性增加趋势, 意味着不同尺度下影响物种多度空间分布的生态过程变化不大.与方差分析不同, 物种丰富度和多度的变异系数均随取样尺度的增加而逐渐减小, 这主要是因为方差反映了多样性测度的绝对变异程度, 而变异系数则反映了多样性测度的相对变异程度.这些研究结果表明多样性的绝对变异更能反映出生态过程对生物多样性格局的影响. ...
On species-area relations
1
1996
... 生态学模式和生态学过程都具有明显的尺度效应(Crawley & Harral, 2001 ; He et al ., 2002 ; Rahbek, 2005 ; Wang et al ., 2008 ).单独一个尺度并不能满足所有科学问题的研究(Ray & Hastings, 1996 ), 例如较小的局部近邻尺度较有利于邻体竞争的研究, 而研究气候对物种多样性的影响时就需要扩展到更大的区域尺度.近些年来, 在生物多样性方面关于尺度效应的研究主要集中在5个方面: (1)不同研究区域内, 最佳取样单元大小、形状以及取样范围的选取(Mouillot & Leprêtre, 1999 ); (2)生物多样性热点地区(biodiversity hotspot)的研究(Bartha et al ., 1998 ; Fortin et al ., 1999 ; Hurlbert & Jetz, 2007 ); (3)生物多样性估算及多样性格局的尺度效应研究(He & Legendre, 1996 ; Wilson et al ., 1999 ; Harte et al ., 2009 ; Colwell et al ., 2012 ; Xu et al ., 2012 ); (4)不同尺度下物种多样性格局形成的机理及相应生态学过程的探究(程佳佳等, 2011 ); (5)物种、生态系统以及景观不同层次多样性之间尺度效应的关系(彭羽等, 2015 ).这些问题的研究对探讨物种多样性的形成和维持机制具有重要意义, 同时也可为生物多样性的保护和有效管理提供科学依据(Willis & Whittaker, 2002 ; Rahbek, 2005 ; Storch et al ., 2007 ; Giladi et al ., 2011 ). ...
Diversity pattern and spatial scale: A study of a tropical rain forest of Malaysia
1994
Diversity and evenness: A unifying notation and its consequences
1
1973
... 各物种多样性指数(Hill, 1973 )以及方差和变异系数的计算公式如下: ...
Spatial variance in ecology
1
1995
... 根据计算结果, 绘制5种物种多样性测度指标的空间分布图, 以对整个样地不同尺度下的物种多样性分布特点及空间变化进行分析.计算每一尺度物种丰富度和多度的Spearman相关系数, 探讨物种丰富度和多度的空间格局随尺度变化的分异.方差和变异系数常常被用来寻找其中的拐点或峰值, 有研究认为这样的拐点或峰值对于确定相应尺度下某种潜在的物理或生态过程甚至群落的特征区域都是很有作用的(Juhász-Nagy & Podani, 1983 ; Horne & Schneider, 1995 ).同时利用多样性指标的方差和变异系数对取样空间尺度(取样面积)的自然对数进行做图, 以探讨尺度变化对物种多样性格局的影响. ...
Species richness, hotspots, and the scale dependence of range maps in ecology and conserva- tion
1
2007
... 生态学模式和生态学过程都具有明显的尺度效应(Crawley & Harral, 2001 ; He et al ., 2002 ; Rahbek, 2005 ; Wang et al ., 2008 ).单独一个尺度并不能满足所有科学问题的研究(Ray & Hastings, 1996 ), 例如较小的局部近邻尺度较有利于邻体竞争的研究, 而研究气候对物种多样性的影响时就需要扩展到更大的区域尺度.近些年来, 在生物多样性方面关于尺度效应的研究主要集中在5个方面: (1)不同研究区域内, 最佳取样单元大小、形状以及取样范围的选取(Mouillot & Leprêtre, 1999 ); (2)生物多样性热点地区(biodiversity hotspot)的研究(Bartha et al ., 1998 ; Fortin et al ., 1999 ; Hurlbert & Jetz, 2007 ); (3)生物多样性估算及多样性格局的尺度效应研究(He & Legendre, 1996 ; Wilson et al ., 1999 ; Harte et al ., 2009 ; Colwell et al ., 2012 ; Xu et al ., 2012 ); (4)不同尺度下物种多样性格局形成的机理及相应生态学过程的探究(程佳佳等, 2011 ); (5)物种、生态系统以及景观不同层次多样性之间尺度效应的关系(彭羽等, 2015 ).这些问题的研究对探讨物种多样性的形成和维持机制具有重要意义, 同时也可为生物多样性的保护和有效管理提供科学依据(Willis & Whittaker, 2002 ; Rahbek, 2005 ; Storch et al ., 2007 ; Giladi et al ., 2011 ). ...
2
1991
... 尖峰岭地区位于海南省西南部乐东黎族自治县和东方市交界处(18.33°-18.95° N, 108.68°-109.20° E), 总面积约640 km2 .尖峰岭处于低纬度热带岛屿季风气候区, 干湿两季明显, 雨季从5月至10月, 旱季从11月至翌年4月(李意德等, 2012 ).该区年平均降水量2 449 mm, 年平均气温19.8 ℃, 最冷月和最热月平均温度分别为10.8 ℃和27.5 ℃ (蒋有绪和卢俊培, 1991 ).其中热带山地雨林主要分布于海拔600-1200 m的山地丘陵, 砖黄壤-黄壤是主要的土壤类型.热带山地雨林也是尖峰岭地区分布面积最大的植被类型(蒋有绪和卢俊培, 1991 ; 李意德等, 2002 ). ...
... ).其中热带山地雨林主要分布于海拔600-1200 m的山地丘陵, 砖黄壤-黄壤是主要的土壤类型.热带山地雨林也是尖峰岭地区分布面积最大的植被类型(蒋有绪和卢俊培, 1991 ; 李意德等, 2002 ). ...
Information theory methods for the study of spatial processes and succession
1
1983
... 根据计算结果, 绘制5种物种多样性测度指标的空间分布图, 以对整个样地不同尺度下的物种多样性分布特点及空间变化进行分析.计算每一尺度物种丰富度和多度的Spearman相关系数, 探讨物种丰富度和多度的空间格局随尺度变化的分异.方差和变异系数常常被用来寻找其中的拐点或峰值, 有研究认为这样的拐点或峰值对于确定相应尺度下某种潜在的物理或生态过程甚至群落的特征区域都是很有作用的(Juhász-Nagy & Podani, 1983 ; Horne & Schneider, 1995 ).同时利用多样性指标的方差和变异系数对取样空间尺度(取样面积)的自然对数进行做图, 以探讨尺度变化对物种多样性格局的影响. ...
The scale of analysis determines the spatial pattern of woody species diversity in the Mediterranean environment
1
2008
... 关于物种多样性空间分布格局的尺度效应, 国内外已在热带雨林(He et al ., 2002 ; Kallimanis et al ., 2008 )、温带针阔混交林(Wang et al ., 2008 )、中亚热带喀斯特森林(张忠华等, 2012 )以及南亚热带常绿阔叶林(王志高等, 2008 )中开展过一些研究, 而针对我国热带森林物种多样性的空间分布及其与尺度间关系的研究比较缺乏.海南岛地处热带亚洲北缘, 丰富的热带雨林物种组成使其成为全球生物多样性研究的热点地区之一.本文以海南岛西南部尖峰岭国家级自然保护区内建成的60 hm2 热带山地雨林动态监测样地为数据来源, 探讨了不同空间尺度下各物种多样性指标的空间分布特征, 以期为进一步揭示海南尖峰岭地区物种多样性的形成和维持机制提供科学依据. ...
2
2002
... 尖峰岭地区位于海南省西南部乐东黎族自治县和东方市交界处(18.33°-18.95° N, 108.68°-109.20° E), 总面积约640 km2 .尖峰岭处于低纬度热带岛屿季风气候区, 干湿两季明显, 雨季从5月至10月, 旱季从11月至翌年4月(李意德等, 2012 ).该区年平均降水量2 449 mm, 年平均气温19.8 ℃, 最冷月和最热月平均温度分别为10.8 ℃和27.5 ℃ (蒋有绪和卢俊培, 1991 ).其中热带山地雨林主要分布于海拔600-1200 m的山地丘陵, 砖黄壤-黄壤是主要的土壤类型.热带山地雨林也是尖峰岭地区分布面积最大的植被类型(蒋有绪和卢俊培, 1991 ; 李意德等, 2002 ). ...
... ; 李意德等, 2002 ). ...
北京东灵山地区植物群落多样性的研究: II丰富度、均匀度和物种多样性
1
1995
... Shannon-Wiener指数、Simpson指数和Pielou均匀度指数同样存在尺度效应, 这种尺度效应因研究区域不同而有所差异, 但在同一研究区域内较为一致.本研究中, 随着取样尺度的增加, 3种多样性指数的方差和变异系数均呈现出减小的趋势.而在南亚热带鼎湖山20 hm2 常绿阔叶林内开展的类似研究中, 3种多样性指数的方差随着取样尺度的增加都表现出单峰型的变化特征(王志高等, 2008 ), 这可能与两个样地内不同的稀有种组成有很大关系.尖峰岭样地内的稀有物种数量为60种, 占总物种数的20.7% (许涵等, 2015 ), 而鼎湖山样地内的稀有种数量为110种, 占总物种数的52.4% (叶万辉等, 2008 ).位于马来西亚的Pasoh热带雨林50 hm2 样地, 其稀有种数量为301种, 占总种数的36.5%.He等(1994)在该区域的研究发现, 相对较小以及较大尺度而言, 稀有种对Shannon-Wiener指数尺度效应的影响仅在10 m × 10 m到30 m × 30 m的尺度区间才表现明显.鼎湖山样地以及Pasoh样地中稀有种的组成较尖峰岭样地要高很多, 这很可能是导致物种多样性指数空间分布格局产生差异的原因之一.此外本研究中, 不同取样空间尺度下, Shannon- Wiener指数具有较大的空间变异, 而Simpson指数与Pielou均匀度指数的变异均较小.这与Shannon- Wiener指数和Simpson指数自身的性质有关, Shannon-Wiener指数与物种丰富度的关系最为密切, 而Simpson指数则对均匀度更为敏感(马克平等, 1995 ; 许晴等, 2011 ). ...
A comparison of species diversity estimators
1
1999
... 生态学模式和生态学过程都具有明显的尺度效应(Crawley & Harral, 2001 ; He et al ., 2002 ; Rahbek, 2005 ; Wang et al ., 2008 ).单独一个尺度并不能满足所有科学问题的研究(Ray & Hastings, 1996 ), 例如较小的局部近邻尺度较有利于邻体竞争的研究, 而研究气候对物种多样性的影响时就需要扩展到更大的区域尺度.近些年来, 在生物多样性方面关于尺度效应的研究主要集中在5个方面: (1)不同研究区域内, 最佳取样单元大小、形状以及取样范围的选取(Mouillot & Leprêtre, 1999 ); (2)生物多样性热点地区(biodiversity hotspot)的研究(Bartha et al ., 1998 ; Fortin et al ., 1999 ; Hurlbert & Jetz, 2007 ); (3)生物多样性估算及多样性格局的尺度效应研究(He & Legendre, 1996 ; Wilson et al ., 1999 ; Harte et al ., 2009 ; Colwell et al ., 2012 ; Xu et al ., 2012 ); (4)不同尺度下物种多样性格局形成的机理及相应生态学过程的探究(程佳佳等, 2011 ); (5)物种、生态系统以及景观不同层次多样性之间尺度效应的关系(彭羽等, 2015 ).这些问题的研究对探讨物种多样性的形成和维持机制具有重要意义, 同时也可为生物多样性的保护和有效管理提供科学依据(Willis & Whittaker, 2002 ; Rahbek, 2005 ; Storch et al ., 2007 ; Giladi et al ., 2011 ). ...
生物多样性不同层次尺度效应及其耦合关系研究进展
1
2015
... 生态学模式和生态学过程都具有明显的尺度效应(Crawley & Harral, 2001 ; He et al ., 2002 ; Rahbek, 2005 ; Wang et al ., 2008 ).单独一个尺度并不能满足所有科学问题的研究(Ray & Hastings, 1996 ), 例如较小的局部近邻尺度较有利于邻体竞争的研究, 而研究气候对物种多样性的影响时就需要扩展到更大的区域尺度.近些年来, 在生物多样性方面关于尺度效应的研究主要集中在5个方面: (1)不同研究区域内, 最佳取样单元大小、形状以及取样范围的选取(Mouillot & Leprêtre, 1999 ); (2)生物多样性热点地区(biodiversity hotspot)的研究(Bartha et al ., 1998 ; Fortin et al ., 1999 ; Hurlbert & Jetz, 2007 ); (3)生物多样性估算及多样性格局的尺度效应研究(He & Legendre, 1996 ; Wilson et al ., 1999 ; Harte et al ., 2009 ; Colwell et al ., 2012 ; Xu et al ., 2012 ); (4)不同尺度下物种多样性格局形成的机理及相应生态学过程的探究(程佳佳等, 2011 ); (5)物种、生态系统以及景观不同层次多样性之间尺度效应的关系(彭羽等, 2015 ).这些问题的研究对探讨物种多样性的形成和维持机制具有重要意义, 同时也可为生物多样性的保护和有效管理提供科学依据(Willis & Whittaker, 2002 ; Rahbek, 2005 ; Storch et al ., 2007 ; Giladi et al ., 2011 ). ...
1
1962
... 物种丰富度和多度的分布格局都是基于空间尺度存在的.在小尺度上, 两者的空间分布表现出相当的一致性, 即两者数值较高的样方均出现在山脊及其附近区域, 随着尺度的增加, 这种一致性则不再明显或消失.秩相关分析结果也表明, 在小于40 m × 40 m尺度上, 物种丰富度和多度呈现出极显著(p < 0.001)的正相关关系, 并且相关系数在大于20 m × 20 m尺度时有明显减弱, 当尺度超过40 m × 40 m时, 两者的相关性则不再显著, 这可能反映了决定生物多样性格局的机制随尺度增加而发生了变化.小尺度物种丰富度主要与样方内包含的木本植物个体数量有关(Preston, 1962 ), 随着尺度增加, 生境的异质性增加, 物种丰富度随面积的增加主要是由于新生境的出现(Turner & Tjørve, 2005 ), 因而物种丰富度和多度的相关性逐渐变得不显著.以往研究发现, 尖峰岭热带山地雨林60 hm2 大样地内, 不同深度土层的13项土壤化学性质(全氮、全磷、全钾、有效氮、有效磷和速效钾等)以及8项土壤物理性质(土壤容重、土壤含水量和土壤总孔隙等)空间自相关变程大多都在20-40 m之间(时雷雷, 2012 ).这反映了土壤属性的空间自相关范围, 当尺度较小, 处于变程内时, 各样方间的空间变异很小, 所提供的生境类型在一个比较稳定的水平, 物种丰富度和多度表现出了极显著的正相关, 当尺度超过变程时, 土壤性质的分布则更多体现为一种随机性, 此时两者的相关性逐渐减弱甚至消失. ...
The role of spatial scale and the perception of large-scale species-richness patterns
2
2005
... 生态学模式和生态学过程都具有明显的尺度效应(Crawley & Harral, 2001 ; He et al ., 2002 ; Rahbek, 2005 ; Wang et al ., 2008 ).单独一个尺度并不能满足所有科学问题的研究(Ray & Hastings, 1996 ), 例如较小的局部近邻尺度较有利于邻体竞争的研究, 而研究气候对物种多样性的影响时就需要扩展到更大的区域尺度.近些年来, 在生物多样性方面关于尺度效应的研究主要集中在5个方面: (1)不同研究区域内, 最佳取样单元大小、形状以及取样范围的选取(Mouillot & Leprêtre, 1999 ); (2)生物多样性热点地区(biodiversity hotspot)的研究(Bartha et al ., 1998 ; Fortin et al ., 1999 ; Hurlbert & Jetz, 2007 ); (3)生物多样性估算及多样性格局的尺度效应研究(He & Legendre, 1996 ; Wilson et al ., 1999 ; Harte et al ., 2009 ; Colwell et al ., 2012 ; Xu et al ., 2012 ); (4)不同尺度下物种多样性格局形成的机理及相应生态学过程的探究(程佳佳等, 2011 ); (5)物种、生态系统以及景观不同层次多样性之间尺度效应的关系(彭羽等, 2015 ).这些问题的研究对探讨物种多样性的形成和维持机制具有重要意义, 同时也可为生物多样性的保护和有效管理提供科学依据(Willis & Whittaker, 2002 ; Rahbek, 2005 ; Storch et al ., 2007 ; Giladi et al ., 2011 ). ...
... ; Rahbek, 2005 ; Storch et al ., 2007 ; Giladi et al ., 2011 ). ...
Density dependence: Are we searching at the wrong spatial scale?
1
1996
... 生态学模式和生态学过程都具有明显的尺度效应(Crawley & Harral, 2001 ; He et al ., 2002 ; Rahbek, 2005 ; Wang et al ., 2008 ).单独一个尺度并不能满足所有科学问题的研究(Ray & Hastings, 1996 ), 例如较小的局部近邻尺度较有利于邻体竞争的研究, 而研究气候对物种多样性的影响时就需要扩展到更大的区域尺度.近些年来, 在生物多样性方面关于尺度效应的研究主要集中在5个方面: (1)不同研究区域内, 最佳取样单元大小、形状以及取样范围的选取(Mouillot & Leprêtre, 1999 ); (2)生物多样性热点地区(biodiversity hotspot)的研究(Bartha et al ., 1998 ; Fortin et al ., 1999 ; Hurlbert & Jetz, 2007 ); (3)生物多样性估算及多样性格局的尺度效应研究(He & Legendre, 1996 ; Wilson et al ., 1999 ; Harte et al ., 2009 ; Colwell et al ., 2012 ; Xu et al ., 2012 ); (4)不同尺度下物种多样性格局形成的机理及相应生态学过程的探究(程佳佳等, 2011 ); (5)物种、生态系统以及景观不同层次多样性之间尺度效应的关系(彭羽等, 2015 ).这些问题的研究对探讨物种多样性的形成和维持机制具有重要意义, 同时也可为生物多样性的保护和有效管理提供科学依据(Willis & Whittaker, 2002 ; Rahbek, 2005 ; Storch et al ., 2007 ; Giladi et al ., 2011 ). ...
海南尖峰岭热带山地雨林原始林土壤理化性质空间异质性研究
1
2012
... 物种丰富度和多度的分布格局都是基于空间尺度存在的.在小尺度上, 两者的空间分布表现出相当的一致性, 即两者数值较高的样方均出现在山脊及其附近区域, 随着尺度的增加, 这种一致性则不再明显或消失.秩相关分析结果也表明, 在小于40 m × 40 m尺度上, 物种丰富度和多度呈现出极显著(p < 0.001)的正相关关系, 并且相关系数在大于20 m × 20 m尺度时有明显减弱, 当尺度超过40 m × 40 m时, 两者的相关性则不再显著, 这可能反映了决定生物多样性格局的机制随尺度增加而发生了变化.小尺度物种丰富度主要与样方内包含的木本植物个体数量有关(Preston, 1962 ), 随着尺度增加, 生境的异质性增加, 物种丰富度随面积的增加主要是由于新生境的出现(Turner & Tjørve, 2005 ), 因而物种丰富度和多度的相关性逐渐变得不显著.以往研究发现, 尖峰岭热带山地雨林60 hm2 大样地内, 不同深度土层的13项土壤化学性质(全氮、全磷、全钾、有效氮、有效磷和速效钾等)以及8项土壤物理性质(土壤容重、土壤含水量和土壤总孔隙等)空间自相关变程大多都在20-40 m之间(时雷雷, 2012 ).这反映了土壤属性的空间自相关范围, 当尺度较小, 处于变程内时, 各样方间的空间变异很小, 所提供的生境类型在一个比较稳定的水平, 物种丰富度和多度表现出了极显著的正相关, 当尺度超过变程时, 土壤性质的分布则更多体现为一种随机性, 此时两者的相关性逐渐减弱甚至消失. ...
1
2007
... 生态学模式和生态学过程都具有明显的尺度效应(Crawley & Harral, 2001 ; He et al ., 2002 ; Rahbek, 2005 ; Wang et al ., 2008 ).单独一个尺度并不能满足所有科学问题的研究(Ray & Hastings, 1996 ), 例如较小的局部近邻尺度较有利于邻体竞争的研究, 而研究气候对物种多样性的影响时就需要扩展到更大的区域尺度.近些年来, 在生物多样性方面关于尺度效应的研究主要集中在5个方面: (1)不同研究区域内, 最佳取样单元大小、形状以及取样范围的选取(Mouillot & Leprêtre, 1999 ); (2)生物多样性热点地区(biodiversity hotspot)的研究(Bartha et al ., 1998 ; Fortin et al ., 1999 ; Hurlbert & Jetz, 2007 ); (3)生物多样性估算及多样性格局的尺度效应研究(He & Legendre, 1996 ; Wilson et al ., 1999 ; Harte et al ., 2009 ; Colwell et al ., 2012 ; Xu et al ., 2012 ); (4)不同尺度下物种多样性格局形成的机理及相应生态学过程的探究(程佳佳等, 2011 ); (5)物种、生态系统以及景观不同层次多样性之间尺度效应的关系(彭羽等, 2015 ).这些问题的研究对探讨物种多样性的形成和维持机制具有重要意义, 同时也可为生物多样性的保护和有效管理提供科学依据(Willis & Whittaker, 2002 ; Rahbek, 2005 ; Storch et al ., 2007 ; Giladi et al ., 2011 ). ...
Scale-dependence in species-area relationships
1
2005
... 物种丰富度和多度的分布格局都是基于空间尺度存在的.在小尺度上, 两者的空间分布表现出相当的一致性, 即两者数值较高的样方均出现在山脊及其附近区域, 随着尺度的增加, 这种一致性则不再明显或消失.秩相关分析结果也表明, 在小于40 m × 40 m尺度上, 物种丰富度和多度呈现出极显著(p < 0.001)的正相关关系, 并且相关系数在大于20 m × 20 m尺度时有明显减弱, 当尺度超过40 m × 40 m时, 两者的相关性则不再显著, 这可能反映了决定生物多样性格局的机制随尺度增加而发生了变化.小尺度物种丰富度主要与样方内包含的木本植物个体数量有关(Preston, 1962 ), 随着尺度增加, 生境的异质性增加, 物种丰富度随面积的增加主要是由于新生境的出现(Turner & Tjørve, 2005 ), 因而物种丰富度和多度的相关性逐渐变得不显著.以往研究发现, 尖峰岭热带山地雨林60 hm2 大样地内, 不同深度土层的13项土壤化学性质(全氮、全磷、全钾、有效氮、有效磷和速效钾等)以及8项土壤物理性质(土壤容重、土壤含水量和土壤总孔隙等)空间自相关变程大多都在20-40 m之间(时雷雷, 2012 ).这反映了土壤属性的空间自相关范围, 当尺度较小, 处于变程内时, 各样方间的空间变异很小, 所提供的生境类型在一个比较稳定的水平, 物种丰富度和多度表现出了极显著的正相关, 当尺度超过变程时, 土壤性质的分布则更多体现为一种随机性, 此时两者的相关性逐渐减弱甚至消失. ...
Spatial variation of species diversity across scale in an old-growth temperate forest of China
3
2008
... 生态学模式和生态学过程都具有明显的尺度效应(Crawley & Harral, 2001 ; He et al ., 2002 ; Rahbek, 2005 ; Wang et al ., 2008 ).单独一个尺度并不能满足所有科学问题的研究(Ray & Hastings, 1996 ), 例如较小的局部近邻尺度较有利于邻体竞争的研究, 而研究气候对物种多样性的影响时就需要扩展到更大的区域尺度.近些年来, 在生物多样性方面关于尺度效应的研究主要集中在5个方面: (1)不同研究区域内, 最佳取样单元大小、形状以及取样范围的选取(Mouillot & Leprêtre, 1999 ); (2)生物多样性热点地区(biodiversity hotspot)的研究(Bartha et al ., 1998 ; Fortin et al ., 1999 ; Hurlbert & Jetz, 2007 ); (3)生物多样性估算及多样性格局的尺度效应研究(He & Legendre, 1996 ; Wilson et al ., 1999 ; Harte et al ., 2009 ; Colwell et al ., 2012 ; Xu et al ., 2012 ); (4)不同尺度下物种多样性格局形成的机理及相应生态学过程的探究(程佳佳等, 2011 ); (5)物种、生态系统以及景观不同层次多样性之间尺度效应的关系(彭羽等, 2015 ).这些问题的研究对探讨物种多样性的形成和维持机制具有重要意义, 同时也可为生物多样性的保护和有效管理提供科学依据(Willis & Whittaker, 2002 ; Rahbek, 2005 ; Storch et al ., 2007 ; Giladi et al ., 2011 ). ...
... 关于物种多样性空间分布格局的尺度效应, 国内外已在热带雨林(He et al ., 2002 ; Kallimanis et al ., 2008 )、温带针阔混交林(Wang et al ., 2008 )、中亚热带喀斯特森林(张忠华等, 2012 )以及南亚热带常绿阔叶林(王志高等, 2008 )中开展过一些研究, 而针对我国热带森林物种多样性的空间分布及其与尺度间关系的研究比较缺乏.海南岛地处热带亚洲北缘, 丰富的热带雨林物种组成使其成为全球生物多样性研究的热点地区之一.本文以海南岛西南部尖峰岭国家级自然保护区内建成的60 hm2 热带山地雨林动态监测样地为数据来源, 探讨了不同空间尺度下各物种多样性指标的空间分布特征, 以期为进一步揭示海南尖峰岭地区物种多样性的形成和维持机制提供科学依据. ...
... 尖峰岭60 hm2 热带山地雨林样地中, 物种丰富度和多度的方差随着空间尺度的变化表现出不同的趋势.物种丰富度的方差随取样尺度增加呈现出单峰分布特征, 并且在20 m × 20 m尺度上出现了峰值, 而产生这一峰值的尺度与物种丰富度和多度格局的分离尺度相一致, 这也在一定程度上印证了生态学家通过方差分析中产生的拐点来确定相应尺度下潜在的生态过程的合理性.类似的峰型曲线同样出现在其他地区的热带森林(He et al ., 2002 )、温带森林(Wang et al ., 2008 )以及中亚热带的喀斯特森林 (张忠华等, 2012 )中.而物种多度的方差则是随取样尺度的增加呈线性增加趋势, 意味着不同尺度下影响物种多度空间分布的生态过程变化不大.与方差分析不同, 物种丰富度和多度的变异系数均随取样尺度的增加而逐渐减小, 这主要是因为方差反映了多样性测度的绝对变异程度, 而变异系数则反映了多样性测度的相对变异程度.这些研究结果表明多样性的绝对变异更能反映出生态过程对生物多样性格局的影响. ...
鼎湖山季风常绿阔叶林物种多样性指数空间分布特征
2
2008
... 关于物种多样性空间分布格局的尺度效应, 国内外已在热带雨林(He et al ., 2002 ; Kallimanis et al ., 2008 )、温带针阔混交林(Wang et al ., 2008 )、中亚热带喀斯特森林(张忠华等, 2012 )以及南亚热带常绿阔叶林(王志高等, 2008 )中开展过一些研究, 而针对我国热带森林物种多样性的空间分布及其与尺度间关系的研究比较缺乏.海南岛地处热带亚洲北缘, 丰富的热带雨林物种组成使其成为全球生物多样性研究的热点地区之一.本文以海南岛西南部尖峰岭国家级自然保护区内建成的60 hm2 热带山地雨林动态监测样地为数据来源, 探讨了不同空间尺度下各物种多样性指标的空间分布特征, 以期为进一步揭示海南尖峰岭地区物种多样性的形成和维持机制提供科学依据. ...
... Shannon-Wiener指数、Simpson指数和Pielou均匀度指数同样存在尺度效应, 这种尺度效应因研究区域不同而有所差异, 但在同一研究区域内较为一致.本研究中, 随着取样尺度的增加, 3种多样性指数的方差和变异系数均呈现出减小的趋势.而在南亚热带鼎湖山20 hm2 常绿阔叶林内开展的类似研究中, 3种多样性指数的方差随着取样尺度的增加都表现出单峰型的变化特征(王志高等, 2008 ), 这可能与两个样地内不同的稀有种组成有很大关系.尖峰岭样地内的稀有物种数量为60种, 占总物种数的20.7% (许涵等, 2015 ), 而鼎湖山样地内的稀有种数量为110种, 占总物种数的52.4% (叶万辉等, 2008 ).位于马来西亚的Pasoh热带雨林50 hm2 样地, 其稀有种数量为301种, 占总种数的36.5%.He等(1994)在该区域的研究发现, 相对较小以及较大尺度而言, 稀有种对Shannon-Wiener指数尺度效应的影响仅在10 m × 10 m到30 m × 30 m的尺度区间才表现明显.鼎湖山样地以及Pasoh样地中稀有种的组成较尖峰岭样地要高很多, 这很可能是导致物种多样性指数空间分布格局产生差异的原因之一.此外本研究中, 不同取样空间尺度下, Shannon- Wiener指数具有较大的空间变异, 而Simpson指数与Pielou均匀度指数的变异均较小.这与Shannon- Wiener指数和Simpson指数自身的性质有关, Shannon-Wiener指数与物种丰富度的关系最为密切, 而Simpson指数则对均匀度更为敏感(马克平等, 1995 ; 许晴等, 2011 ). ...
Species diversity-scale matters
1
2002
... 生态学模式和生态学过程都具有明显的尺度效应(Crawley & Harral, 2001 ; He et al ., 2002 ; Rahbek, 2005 ; Wang et al ., 2008 ).单独一个尺度并不能满足所有科学问题的研究(Ray & Hastings, 1996 ), 例如较小的局部近邻尺度较有利于邻体竞争的研究, 而研究气候对物种多样性的影响时就需要扩展到更大的区域尺度.近些年来, 在生物多样性方面关于尺度效应的研究主要集中在5个方面: (1)不同研究区域内, 最佳取样单元大小、形状以及取样范围的选取(Mouillot & Leprêtre, 1999 ); (2)生物多样性热点地区(biodiversity hotspot)的研究(Bartha et al ., 1998 ; Fortin et al ., 1999 ; Hurlbert & Jetz, 2007 ); (3)生物多样性估算及多样性格局的尺度效应研究(He & Legendre, 1996 ; Wilson et al ., 1999 ; Harte et al ., 2009 ; Colwell et al ., 2012 ; Xu et al ., 2012 ); (4)不同尺度下物种多样性格局形成的机理及相应生态学过程的探究(程佳佳等, 2011 ); (5)物种、生态系统以及景观不同层次多样性之间尺度效应的关系(彭羽等, 2015 ).这些问题的研究对探讨物种多样性的形成和维持机制具有重要意义, 同时也可为生物多样性的保护和有效管理提供科学依据(Willis & Whittaker, 2002 ; Rahbek, 2005 ; Storch et al ., 2007 ; Giladi et al ., 2011 ). ...
The effect of spatial scale on evenness
1
1999
... 生态学模式和生态学过程都具有明显的尺度效应(Crawley & Harral, 2001 ; He et al ., 2002 ; Rahbek, 2005 ; Wang et al ., 2008 ).单独一个尺度并不能满足所有科学问题的研究(Ray & Hastings, 1996 ), 例如较小的局部近邻尺度较有利于邻体竞争的研究, 而研究气候对物种多样性的影响时就需要扩展到更大的区域尺度.近些年来, 在生物多样性方面关于尺度效应的研究主要集中在5个方面: (1)不同研究区域内, 最佳取样单元大小、形状以及取样范围的选取(Mouillot & Leprêtre, 1999 ); (2)生物多样性热点地区(biodiversity hotspot)的研究(Bartha et al ., 1998 ; Fortin et al ., 1999 ; Hurlbert & Jetz, 2007 ); (3)生物多样性估算及多样性格局的尺度效应研究(He & Legendre, 1996 ; Wilson et al ., 1999 ; Harte et al ., 2009 ; Colwell et al ., 2012 ; Xu et al ., 2012 ); (4)不同尺度下物种多样性格局形成的机理及相应生态学过程的探究(程佳佳等, 2011 ); (5)物种、生态系统以及景观不同层次多样性之间尺度效应的关系(彭羽等, 2015 ).这些问题的研究对探讨物种多样性的形成和维持机制具有重要意义, 同时也可为生物多样性的保护和有效管理提供科学依据(Willis & Whittaker, 2002 ; Rahbek, 2005 ; Storch et al ., 2007 ; Giladi et al ., 2011 ). ...
3
2015
... 2012年底建设完成的60 hm2 森林生物多样性动态监测样地位于海南尖峰岭五分区的热带山地雨林原始林内, 整个样地严格参照美国史密森热带研究所(Smithsonian Institution)热带森林研究中心(Center for Tropical Forest Science, CTFS)的调查技术规范进行建设(Condit, 1998 ).样地东西长1 000 m, 南北宽600 m (图1 ), 共划分为1 500个20 m × 20 m的样方进行调查, 每个样方四个角均用全站仪测定了其在样方中的坐标位置, 并用水泥桩作了永久标记.样地调查以每个20 m × 20 m样方为单元, 记录了其中所有胸径(DBH )≥1.0 cm的木本植物种名、DBH 和坐标值, 以后每隔5年复查一次.首次调查结果表明, 样地内共记录到439 676株存活的DBH ≥1.0 cm的乔灌木植株, 包括391 686个独立植株、29 103个萌条和18 887个分枝.它们当中除61个植株未确定种名外, 其他分属于62科155属290种(许涵等, 2015 ). ...
... Shannon-Wiener指数、Simpson指数和Pielou均匀度指数同样存在尺度效应, 这种尺度效应因研究区域不同而有所差异, 但在同一研究区域内较为一致.本研究中, 随着取样尺度的增加, 3种多样性指数的方差和变异系数均呈现出减小的趋势.而在南亚热带鼎湖山20 hm2 常绿阔叶林内开展的类似研究中, 3种多样性指数的方差随着取样尺度的增加都表现出单峰型的变化特征(王志高等, 2008 ), 这可能与两个样地内不同的稀有种组成有很大关系.尖峰岭样地内的稀有物种数量为60种, 占总物种数的20.7% (许涵等, 2015 ), 而鼎湖山样地内的稀有种数量为110种, 占总物种数的52.4% (叶万辉等, 2008 ).位于马来西亚的Pasoh热带雨林50 hm2 样地, 其稀有种数量为301种, 占总种数的36.5%.He等(1994)在该区域的研究发现, 相对较小以及较大尺度而言, 稀有种对Shannon-Wiener指数尺度效应的影响仅在10 m × 10 m到30 m × 30 m的尺度区间才表现明显.鼎湖山样地以及Pasoh样地中稀有种的组成较尖峰岭样地要高很多, 这很可能是导致物种多样性指数空间分布格局产生差异的原因之一.此外本研究中, 不同取样空间尺度下, Shannon- Wiener指数具有较大的空间变异, 而Simpson指数与Pielou均匀度指数的变异均较小.这与Shannon- Wiener指数和Simpson指数自身的性质有关, Shannon-Wiener指数与物种丰富度的关系最为密切, 而Simpson指数则对均匀度更为敏感(马克平等, 1995 ; 许晴等, 2011 ). ...
... 总的来说, 本文从空间尺度的层面上探讨了物种多样性的空间分布格局, 但是对于这种变化过程的内在机制有待进行更为深入的分析, 比如生境异质性是如何以及在多大程度上影响物种的空间分布.海南尖峰岭样地地形较为复杂, 整个样地20 m × 20 m样方水平上的海拔和坡度分别在866.3-1 016.7 m和1.7°-49.3°间变化, 许涵等(2015) 在该样地的研究发现有些物种的分布明显受地形影响, 例如变色山槟榔(Pinanga baviensis )和柏拉木(Blastus cochichinensis )经常在沟谷地区聚集分布, 而有些种则与土壤养分有紧密关系, 例如豆科的长脐红豆(Ormosia balansae ), 有些物种似乎受其他因素包括微生物等影响, 例如海南蕈树(Altingia obovata ).物种的生境偏好直接影响着物种多样性的空间分布格局, 未来对这些物种的研究, 有助于进一步揭示尖峰岭地区物种多样性的形成和维持机制. ...
Assessing non-parametric and area-based methods for estimating regional species richness
1
2012
... 生态学模式和生态学过程都具有明显的尺度效应(Crawley & Harral, 2001 ; He et al ., 2002 ; Rahbek, 2005 ; Wang et al ., 2008 ).单独一个尺度并不能满足所有科学问题的研究(Ray & Hastings, 1996 ), 例如较小的局部近邻尺度较有利于邻体竞争的研究, 而研究气候对物种多样性的影响时就需要扩展到更大的区域尺度.近些年来, 在生物多样性方面关于尺度效应的研究主要集中在5个方面: (1)不同研究区域内, 最佳取样单元大小、形状以及取样范围的选取(Mouillot & Leprêtre, 1999 ); (2)生物多样性热点地区(biodiversity hotspot)的研究(Bartha et al ., 1998 ; Fortin et al ., 1999 ; Hurlbert & Jetz, 2007 ); (3)生物多样性估算及多样性格局的尺度效应研究(He & Legendre, 1996 ; Wilson et al ., 1999 ; Harte et al ., 2009 ; Colwell et al ., 2012 ; Xu et al ., 2012 ); (4)不同尺度下物种多样性格局形成的机理及相应生态学过程的探究(程佳佳等, 2011 ); (5)物种、生态系统以及景观不同层次多样性之间尺度效应的关系(彭羽等, 2015 ).这些问题的研究对探讨物种多样性的形成和维持机制具有重要意义, 同时也可为生物多样性的保护和有效管理提供科学依据(Willis & Whittaker, 2002 ; Rahbek, 2005 ; Storch et al ., 2007 ; Giladi et al ., 2011 ). ...
Simpson指数和Shannon-Wiener指数若干特征的分析及“稀释效应”
1
2011
... Shannon-Wiener指数、Simpson指数和Pielou均匀度指数同样存在尺度效应, 这种尺度效应因研究区域不同而有所差异, 但在同一研究区域内较为一致.本研究中, 随着取样尺度的增加, 3种多样性指数的方差和变异系数均呈现出减小的趋势.而在南亚热带鼎湖山20 hm2 常绿阔叶林内开展的类似研究中, 3种多样性指数的方差随着取样尺度的增加都表现出单峰型的变化特征(王志高等, 2008 ), 这可能与两个样地内不同的稀有种组成有很大关系.尖峰岭样地内的稀有物种数量为60种, 占总物种数的20.7% (许涵等, 2015 ), 而鼎湖山样地内的稀有种数量为110种, 占总物种数的52.4% (叶万辉等, 2008 ).位于马来西亚的Pasoh热带雨林50 hm2 样地, 其稀有种数量为301种, 占总种数的36.5%.He等(1994)在该区域的研究发现, 相对较小以及较大尺度而言, 稀有种对Shannon-Wiener指数尺度效应的影响仅在10 m × 10 m到30 m × 30 m的尺度区间才表现明显.鼎湖山样地以及Pasoh样地中稀有种的组成较尖峰岭样地要高很多, 这很可能是导致物种多样性指数空间分布格局产生差异的原因之一.此外本研究中, 不同取样空间尺度下, Shannon- Wiener指数具有较大的空间变异, 而Simpson指数与Pielou均匀度指数的变异均较小.这与Shannon- Wiener指数和Simpson指数自身的性质有关, Shannon-Wiener指数与物种丰富度的关系最为密切, 而Simpson指数则对均匀度更为敏感(马克平等, 1995 ; 许晴等, 2011 ). ...
鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究
1
2008
... Shannon-Wiener指数、Simpson指数和Pielou均匀度指数同样存在尺度效应, 这种尺度效应因研究区域不同而有所差异, 但在同一研究区域内较为一致.本研究中, 随着取样尺度的增加, 3种多样性指数的方差和变异系数均呈现出减小的趋势.而在南亚热带鼎湖山20 hm2 常绿阔叶林内开展的类似研究中, 3种多样性指数的方差随着取样尺度的增加都表现出单峰型的变化特征(王志高等, 2008 ), 这可能与两个样地内不同的稀有种组成有很大关系.尖峰岭样地内的稀有物种数量为60种, 占总物种数的20.7% (许涵等, 2015 ), 而鼎湖山样地内的稀有种数量为110种, 占总物种数的52.4% (叶万辉等, 2008 ).位于马来西亚的Pasoh热带雨林50 hm2 样地, 其稀有种数量为301种, 占总种数的36.5%.He等(1994)在该区域的研究发现, 相对较小以及较大尺度而言, 稀有种对Shannon-Wiener指数尺度效应的影响仅在10 m × 10 m到30 m × 30 m的尺度区间才表现明显.鼎湖山样地以及Pasoh样地中稀有种的组成较尖峰岭样地要高很多, 这很可能是导致物种多样性指数空间分布格局产生差异的原因之一.此外本研究中, 不同取样空间尺度下, Shannon- Wiener指数具有较大的空间变异, 而Simpson指数与Pielou均匀度指数的变异均较小.这与Shannon- Wiener指数和Simpson指数自身的性质有关, Shannon-Wiener指数与物种丰富度的关系最为密切, 而Simpson指数则对均匀度更为敏感(马克平等, 1995 ; 许晴等, 2011 ). ...
喀斯特常绿落叶阔叶混交林物种多度与丰富度空间分布的尺度效应
2
2012
... 关于物种多样性空间分布格局的尺度效应, 国内外已在热带雨林(He et al ., 2002 ; Kallimanis et al ., 2008 )、温带针阔混交林(Wang et al ., 2008 )、中亚热带喀斯特森林(张忠华等, 2012 )以及南亚热带常绿阔叶林(王志高等, 2008 )中开展过一些研究, 而针对我国热带森林物种多样性的空间分布及其与尺度间关系的研究比较缺乏.海南岛地处热带亚洲北缘, 丰富的热带雨林物种组成使其成为全球生物多样性研究的热点地区之一.本文以海南岛西南部尖峰岭国家级自然保护区内建成的60 hm2 热带山地雨林动态监测样地为数据来源, 探讨了不同空间尺度下各物种多样性指标的空间分布特征, 以期为进一步揭示海南尖峰岭地区物种多样性的形成和维持机制提供科学依据. ...
... 尖峰岭60 hm2 热带山地雨林样地中, 物种丰富度和多度的方差随着空间尺度的变化表现出不同的趋势.物种丰富度的方差随取样尺度增加呈现出单峰分布特征, 并且在20 m × 20 m尺度上出现了峰值, 而产生这一峰值的尺度与物种丰富度和多度格局的分离尺度相一致, 这也在一定程度上印证了生态学家通过方差分析中产生的拐点来确定相应尺度下潜在的生态过程的合理性.类似的峰型曲线同样出现在其他地区的热带森林(He et al ., 2002 )、温带森林(Wang et al ., 2008 )以及中亚热带的喀斯特森林 (张忠华等, 2012 )中.而物种多度的方差则是随取样尺度的增加呈线性增加趋势, 意味着不同尺度下影响物种多度空间分布的生态过程变化不大.与方差分析不同, 物种丰富度和多度的变异系数均随取样尺度的增加而逐渐减小, 这主要是因为方差反映了多样性测度的绝对变异程度, 而变异系数则反映了多样性测度的相对变异程度.这些研究结果表明多样性的绝对变异更能反映出生态过程对生物多样性格局的影响. ...
 , 李意德
, 李意德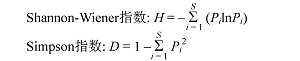



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}