叶片凋落物分解是陆地生态系统中一个重要的生态过程, 其分解速率对养分循环和生产力有着至关重要的作用(Parton et al ., 2007 ; Zhou et al ., 2008 ; Purahong et al ., 2014 )。凋落物分解将光合产物转变为无机养分和土壤有机质(Parton et al ., 2007 ), 提高了土壤肥力, 并提供植物生长所需的养分(王相娥等, 2009 ; Shi et al ., 2015 )。凋落物的分解是一个复杂的过程, 包括短暂的淋溶和长时间的生物分解阶段(Petersen & Cummins, 1974 )。通常情况下, 凋落物分解速率受凋落物质量、环境因子(如温度和湿度)和生物活动(如真菌和细菌)等多方面因素的综合影响(Couteaux et al ., 1995 ; Aerts, 1997 ; Wardle et al ., 2003 ; Ayres et al ., 2006 ; Zhou et al ., 2008 ; 武启骞等, 2013 ; 邓长春等, 2015 )。有研究表明, 凋落物的质量, 如初始N含量、木质素含量和C:N值等不但影响凋落物的分解速率(Aponte et al ., 2012 ; 邓长春等, 2015 ), 而且在很大程度上决定了其养分的固定和矿化(Parton et al ., 2007 ; Osono et al ., 2014 )。此外, 与叶片经济型谱密切联系的叶片功能性状(如叶片厚度与比叶面积)对其凋落物的分解过程也有着重要影响(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 )。
在水生生态系统中, 叶片凋落物作为主要的外源物质为其提供了大量的营养物质(Leroy & Marks, 2006 ), 且叶片的种类将影响着水体微生物及底栖无脊椎动物等的种群结构和数量(Graca, 2001 ; Hieber & Gessner, 2002 )。叶片凋落物在水体中的分解主要受其自身的品质和水体特征的影响。比如, 叶片大小、吸水性和可溶性物质含量将影响分解初期的淋溶速率(Ibrahima et al ., 2008 ); 叶片凋落物初始养分含量将影响其分解速率和养分的释放模式(Pascoal et al ., 2001 ; 刘昕等, 2008 ); 水温和流速也在很大程度上影响着凋落物的分解(Leroy & Marks, 2006 ; Martínez et al ., 2014 )。此外, 水生生态系统周围的植被构成及其季节变化将直接影响进入水体的叶片凋落物类型, 从而影响水体中生物种群变化及物质和能量循环(迟国梁和童晓立, 2010 )。与陆地生态系统相比, 由于水体的浸泡和流动冲击作用, 凋落物在水生生态系统中具有较快的分解速率(Petersen & Cummins, 1974 ; 张川等, 2015 ), 同时, 微生物和食碎屑动物的种类和数量在两种生态系统中的差异也在很大程度上影响着凋落物的分解过程(Leroy & Marks, 2006 ; Tiegs et al ., 2013 ; Handa et al ., 2014 )。目前, 关于叶片凋落物在水体中的分解速率及养分释放特征研究还相对较少。因此, 加强水体叶片凋落物分解特征研究, 尤其针对重要的水生生态系统开展相关研究具有重要意义。
九寨沟作为我国重要的自然保护区, 具有丰富的森林植被类型和众多的高山湖泊, 是长江流域上游一个重要的水源涵养地(刘少英等, 2007 )。我们在九寨沟自然保护区内选择了4个典型树种的叶片凋落物进行陆地和水体凋落物分解特征研究, 旨在解决以下科学问题: 1)不同树种叶片凋落物在林下的分解速率和养分释放特征如何?2)不同树种叶片凋落物在高山湖泊中具有何种分解和养分释放特征?3)不同树种叶片凋落物在林下与高山湖泊中分解速率和养分释放特征存在何种差异?4)凋落物分解速率与各初始养分含量间是否存在相关关系?本文通过对以上科学问题的探讨, 以期为探究九寨沟国家自然保护区内森林和高山湖泊生态系统间物质循环提供一定的科学依据, 并为保护区内森林和湖泊的管理及更好地发挥其水源涵养地的功能提供参考。
1 材料和方法
1.1 研究区域概括
九寨沟自然保护区位于青藏高原东缘的岷山山系南段(100.50°-104.45° E, 30.58°-34.32° N, 海拔1996-4764 m), 属川西高原气候区。该区域受高原季风和热带海洋季风影响, 年平均气温7.3 ℃, 年降水量622 mm, 降水集中在5-9月; 地势南高北低, 相对高差较大, 有高山山地、高山坡地和高山河谷地貌。九寨沟的水体景观具有极高的观赏价值, 有高山湖泊114个、瀑布群17个、滩流5处、泉水47眼、湍流11段。此外, 保护区内植被丰富, 包括寒温性针叶树如红杉(Larix potaninii )和黄果冷杉(Abies ernestii ), 温性针叶树如油松(Pinus tabulaeformis )和华山松(Pinus armandii ), 落叶阔叶树如红桦(Betula albo -sinensis )和辽东栎(Quercus wutaishanica ), 以及落叶阔叶灌丛如高山柳(又叫杯腺柳, Salix cupularis )和绣线菊(Spiraea salicifolia )等(刘少英等, 2007 )。样地概况详见表1 。
1.2 试验设计
2014年9月15日, 分别于黄果冷杉林、油松林、红桦林和高山柳灌丛下收集当年叶片凋落物, 放入烘箱65 ℃烘干至恒质量, 然后用电子天平准确称取5.00 g, 装入宽15 cm、长20 cm、孔径为1 mm的尼龙网袋中, 每一物种装取72袋。采用重铬酸钾-硫酸氧化法、H2 SO4 -H2 O2 -扩散法和钼锑钪比色法分别测定叶片凋落物初始全碳、全氮和全磷含量(鲁如坤, 2000 )。
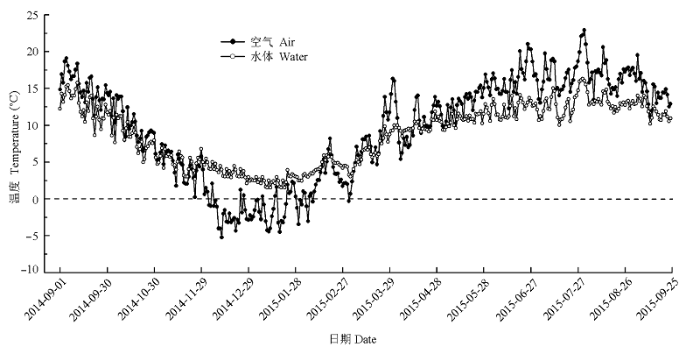
2014年9月25日, 将各物种凋落物分解袋放入其取样样地内, 各样地设置3个区组, 间隔5 m, 每一区组放置12袋。放置时将土壤表面凋落物去掉, 让分解袋贴近土壤表面, 尽量模拟自然状态下的凋落物分解。此外, 在陆地凋落物分解样地附近选择卧龙海(103.90° E, 33.20° N; 海拔2260 m)作为水体凋落物分解试验区。沿湖岸选择3个自然凋落物集中的位置作为凋落物袋放置点, 各样点每一物种放置12袋, 且每一分解袋用细绳连接固定于距岸边2-3 m的湖面上。此外, 在各样点放置一枚纽扣温度计(DS1921G-F5, Maxim Integrated Products, Dallas Semiconductor, Sunnyvale, USA)于空分解袋中, 每隔1 h测定一次湖水温度, 并计算其日平均温度(图1 )。空气日平均温度来自九寨沟扎如沟(103.93° E, 33.26° N; 海拔2064 m)气象站记录数据。
图1 空气及水体日平均温度动态(2014-09-01至2015-09-25)。
Fig. 1 Changes in daily mean air and water temperatures from 2014-09-01 to 2015-09-25.
分别于2014年12月25日、2015年3月25日、6月25日和9月25日, 每隔3个月在每一区组随机收集各物种3袋分解样品, 合计9袋。取回后仔细洗净剩余 凋落物表面的泥沙, 剔除长入袋内的根系, 于65 ℃烘干至恒质量, 在测定剩余凋落物质量后, 粉碎,分别测定其全氮和全磷含量。
1.3 分解模型拟合与计算
本文采用Olson (1963) 提出的负指数衰减模型进行叶片凋落物分解模式拟合, 即y = a e- kt y 为凋落物的剩余率(%), a 为拟合参数, k 为凋落物的分解系数, t 为分解时间(a)。
凋落物分解质量剩余率(MR ) (%) = Mt /M 0 × 100%, 其中M 0 为凋落物初始质量(g), Mt 为时间t 后剩余凋落物质量(g)。
凋落物氮素剩余率(NR ) (%) = (Nt × Mt ) / (N 0 × M 0 ) × 100%, 其中N 0 与Nt 分别为初始与时间t 的凋落物N含量(mg·g-1 ), M 0 与Mt 分别为初始与时间t 的凋落物质量(g)。凋落物P元素剩余率(PR )与NR 的计算方法相同。
1.4 统计分析
本文采用三因素方差分析探讨了物种、分解时间和环境类型对叶片凋落物MR、NR 和PR 的影响, 并分析各因素两两间的交互作用。然后, 采用单因素方差分析法分析凋落物MR、NR 和PR 在不同物种间的差异, 并采用Duncan法进行多重比较。采用Pearson相关分析法分析了叶片凋落物分解1年后MR、NR 和PR 与凋落物初始养分含量之间的相关关系。以上统计分析使用SAS 9.2统计软件进行处理,多重比较显著性水平α = 0.01 (SAS Institute, 2008 )。
2 结果
2.1 凋落物初始养分含量
叶片凋落物初始养分含量在各树种间差异极显著(p < 0.01; 表2 )。油松和红桦叶片凋落物C含量分别为530.59和529.91 mg·g-1 , 且两者差异不显著(p > 0.05), 但它们都极显著(p < 0.01)高于高山柳(505.12 mg·g-1 )和黄果冷杉(489.32 mg·g-1 )。高山柳N含量(16.69 mg·g-1 )极显著(p < 0.01)高于其他树种, 红桦(15.39 mg·g-1 )和黄果冷杉(15.45 mg·g-1 )之间差异不显著(p > 0.05), 而油松叶片N含量(9.42 mg·g-1 )最低。黄果冷杉和高山柳P含量差异不显著(p > 0.05), 其值分别为1.44和1.39 mg·g-1 , 但极显著(p < 0.01)高于红桦(0.88 mg·g-1 )和油松(0.65 mg·g-1 )。此外, 油松凋落物C:N值和C:P值最高, 分别达到56.37和812.60, 而黄果冷杉凋落物N:P值最低, 为10.71。
2.2 凋落物分解特征
2.2.1 凋落物分解质量剩余率(MR )
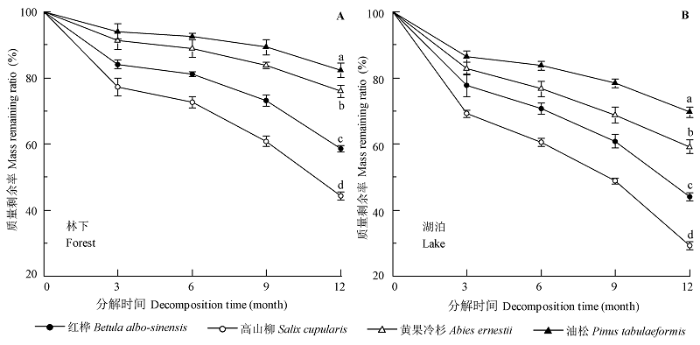
不同树种间叶片MR 差异极显著(p < 0.01), 且林下和湖泊间差异极显著(p < 0.01) (表3 )。MR 随分解时间增加而极显著减小(表3 ; p < 0.01)。各树种叶片凋落物在林下和水体中分解均表现出前3个月质量损失最快, 3-6月质量损失最慢, 6-12月质量损失逐渐增加。凋落物分解1年后, MR 在陆地各树种间大小排序为: 油松(82.34%) >黄果冷杉(75.93%) >红桦(58.54%) >高山柳(44.26%); 在水体中MR 极显著(p < 0.01)低于陆地, 各树种间大小排序为:油松(69.61%) >黄果冷杉(59.14%) >红桦(43.94 %) >高山柳(29.15%) (图2 )。
图2 叶片凋落物在林下(A)及湖泊(B)中分解的质量剩余率(平均值 ± 标准误差)。不同小写字母表示同一分解时间不同树种间叶片凋落物质量剩余率差异极显著(p < 0.01), 且各分解时间的多重比较结果一致。
Fig. 2 Mass remaining ratio of leaf litter decomposition in the forest (A) and lake (B) (mean ± SE ). The lowercase letters denote highly significant differences in mass remaining ratios among the species for the same decomposition time (p < 0.01), with each decomposition time holds the same multiple comparison results.
2.2.2 凋落物分解模式
采用负指数衰减模型对不同树种叶片凋落物在陆地和水体间的分解模式进行拟合的结果均较理想(r > 0.93, p < 0.01; 表4 ), 高山柳和红桦凋落物分解系数k 均高于油松和黄果冷杉, 且水体中各树种凋落物分解k 值均高于林下。水体中各树种凋落物分解k 值分别为: 高山柳(0.97) >红桦(0.68) >黄果冷杉(0.43) >油松(0.28), 而林下其k 值为: 高山柳(0.67) >红桦(0.44) >黄果冷杉(0.24) >油松(0.17)。各树种凋落物在陆地上完全分解(99%)所需的时间为: 高山柳(6.80 a) <红桦(10.34 a) <黄果冷杉(18.88 a) <油松(27.21 a), 且分别为水体中凋落物完全分解时间的1.48倍、1.55倍、1.80倍和1.65倍。
2.3 凋落物养分释放特征
不同树种叶片凋落物NR 和PR 在不同分解时期差异极显著(p < 0.01), PR 在陆地和水体中分解差异极显著(p < 0.01), 而NR 差异不显著(p > 0.05; 表3 )。
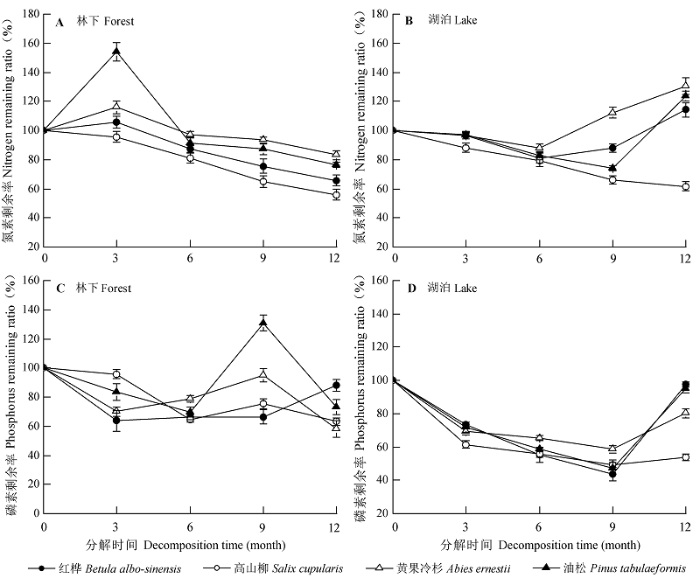
凋落物养分释放模式在不同树种和水陆环境间均存在差异(图3 )。在陆地凋落物分解中, 油松、黄果冷杉和红桦凋落物N含量在前3个月均存在明显的富集过程, 尤其对于油松, NR 可达到154.16%, 然后进入N释放过程; 而高山柳N并未经过富集过程便直接释放。凋落物在水体中分解时, 高山柳N释放模式同其在陆地上分解相似, 而油松、黄果冷杉和红桦N释放模式为: 释放—富集—释放, 而在分解1年后可能还未达到其富集阈值。
图3 叶片凋落物在林下(A和C)和湖泊(B和D)中分解的养分剩余率动态(平均值±标准误差)。
Fig. 3 Dynamics of nutrient remaining ratios of the leaf litter decomposition in the forest (A and C) and lake (B and D) (mean ± SE ).
不同树种凋落物P释放模式在陆地和水体中分解比较相似, 即释放—富集—释放, 但在水体分解中P释放时间较陆地长, 且其在分解1年后可能仍未达其阈值。油松、黄果冷杉和高山柳PR 在陆地分解过程中第9个月达到最大值, 而红桦在第12个月可能仍未达到其阈值。此外, 油松P富集量较其他树种大, 其PR 在陆地和水体中分别为130.74%和95.18%, 而高山柳P富集量相对较少, 其PR 分别为75.48%和53.76%。
2.4 凋落物质量损失和养分释放与初始养分含量之间的关系
如表5 所示, 叶片凋落物分解1年后, MR 与初始C:N值具有极显著的正相关关系(r = 0.57, p < 0.01), 而与初始N含量具有极显著负相关关系(r = - 0.63, p < 0.01); NR 与MR 具有相似特征, 分别与初始C:N值和N含量存在极显著正相关和负相关关系(r = 0.58, r = - 0.60, p < 0.01), 而PR 与凋落物各初始养分含量之间不存在显著的相关关系(p > 0.05)。
3 讨论
3.1 不同树种叶片凋落物分解特征
本文通过研究九寨沟国家自然保护区内4种常见林型代表性树种, 即寒温性针叶林(黄果冷杉)、温性针叶林(油松)、落叶阔叶林(红桦)和落叶阔叶灌丛(高山柳)的叶片凋落物分解特征, 得出以下结果: 各树种间凋落物分解系数k 高山柳>红桦>黄果冷杉>油松, 且各物种间凋落物MR 差异极显著(p < 0.01, 图2 )。结果表明, 叶片凋落物分解速率存在明显的物种差异, 且阔叶树种叶片分解速率显著大于针叶树种, 这与郭忠玲等(2006)对长白山各植被带主要树种凋落物分解速率的研究结果一致。此外, 各树种叶片凋落物在前3个月质量损失最快, 而后迅速减慢, 第6-12月又逐渐增大(图2 )。这与Zhou等(2008)和Aponte等(2012)的研究结果一致, 凋落物分解初期可溶性物质会迅速通过淋溶作用进入到生态系统中, 而后进入较长且缓慢的生物分解过程(Petersen & Cummins, 1974 )。在本文中, 凋落物质量损失变化与外界环境温度的变化趋势一致, 且在降水丰沛的季节(5-9月)较高。这一结果很好地印证了温度和降水量作为主要的环境影响因子控制着凋落物的分解速率(Moore et al ., 1999 ; Hobbie et al ., 2006 ; Parton et al ., 2007 )。
本文结果显示, 叶片凋落物分解1年后MR 同其初始C:N值存在正相关关系, 而与初始N含量成负相关关系(表5 ), 表明凋落物初始养分含量, 尤其是N含量在很大程度上影响着凋落物的分解速率, 这与杨玉盛等(2003)和王希华等(2004)的研究结果一致。值得注意的是, 虽然黄果冷杉叶片凋落物初始N含量较高, 且具有较小的C:N值, 但其凋落物分解速率仍较低。因此, 除初始养分含量外的其他因子在凋落分解过程中起着重要的作用。Hättenschwiler 和Jørgensen (2010)对热带雨林叶片凋落物分解特征研究发现, 凋落物中碳的质量对分解速率的影响比其化学计量特征(C、N和P含量等)更大。此外, 王希华等(2004)和Santiago (2007)研究发现凋落物比叶面积与凋落物分解速率之间存在显著的正相关关系, 较大的比叶面积意味着单位质量凋落物具有更大的环境接触面积, 一方面加大了雨水的淋溶作用, 另一方面有利于微生物定居和土壤动物的碎裂作用, 从而大大提高凋落分解速率(郭忠玲等, 2006 )。也有研究表明, 叶片厚度与凋落物分解速率存在明显的负相关关系(Cornwell et al ., 2008 )。因此, 与叶片经济型谱存在密切联系的叶片功能特征(如比叶面积、叶片氮素含量和叶片厚度等)在叶片凋落物分解过程中起着重要作用(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 )。综上所述, 叶片凋落物分解是一个受其自身性质和外界环境因素共同作用的复杂过程, 它对陆地生态系统的养分循环有着深远影响(Kay et al ., 2008 ; Makkonen et al ., 2012 )。
3.2 不同树种叶片凋落物养分释放特征
在1年的分解期内, 4个树种叶片凋落物的NR 和PR 差异极显著(p < 0.01, 表3 ), 且NR 与初始N含量和C:N值分别存在极显著负相关和正相关关系(表5 ), 这表明凋落物初始养分含量在很大程度上影响着分解过程中的养分释放。在林下分解时, 高山柳叶片凋落物N并未经过富集过程便直接开始释放, 而红桦、黄果冷杉和油松要先经过富集过程才开始释放; 4个树种凋落物P在分解过程中均表现出释放—富集—释放模式。此外, 凋落物初始N和P含量低的物种(如油松)在分解过程中需富集更多的养分才开始出现N和P的释放, 这与王瑾和黄建辉(2001) 的研究结果一致, 而王希华等(2004) 认为营养元素的释放和富集与凋落叶初始营养状况无明显的相关性。因此, 凋落物N和P的富集和释放与初始养分条件的关系存在明显的物种差异。
凋落物自身往往缺乏大量元素, 如N、P和S等(Berg, 2000 ), 从而难以满足微生物的分解作用, 故需其在外界环境中固定相应的养分, 并达到某一阈值时才分解释放凋落物中的养分(Moore et al ., 2006 ; Parton et al ., 2007 ; Aponte et al ., 2012 )。Parnas (1975)认为凋落物C:N值是发生N固持和矿化的关键值, 当C:N值>30时发生N的固持, 而当C:N值<30时发生N的矿化。本文中高山柳凋落物初始C:N值(30.26)十分接近此值, 故在分解初期便出现N的释放, 而其他树种C:N值均大于30, 故先经历富集过程才进行N的释放。此外, 4个树种凋落物P在分解前期均存在明显的释放过程, 这一阶段主要受可溶性P淋溶作用的影响(彭少麟和刘强, 2002 ), 而在春季和夏季发生P的富集则与微生物活动加强有关(王希华等, 2004 )。
3.3 叶片凋落物水陆分解差异
4个树种叶片凋落物分解速率在高山湖泊中的分解速率均极显著高于其在林下的分解速率(p < 0.01, 表3 ; 图2 ), 且各物种凋落物在林下完全分解(99%)所需的时间是其在水体中的1.5倍左右, 这与陈书秀和江明喜(2006)对三峡地区香溪河流域不同树种叶片凋落物分解的研究结果一致。湖泊中凋落物分解速率较快的一个重要原因是凋落物长时间浸泡在水体中, 较强的淋溶作用会加速可溶性物质在分解初期的快速分解, 加之水体的流动将进一步加快这一过程的进行(Petersen & Cummins, 1974 ; 张川等, 2015 )。其次, 水体微生物和无脊椎动物在凋落物分解过程中也起着至关重要的作用(Leroy & Marks, 2006 ; Tiegs et al ., 2013 ), 相对稳定的水体温度(冬季湖泊中平均水温仍保持在2 ℃以上, 图1 )为凋落物分解过程中的生物活动提供了良好的环境保障。本文结果显示, 冬季水体凋落物的失重率保持在一个较高水平。此外, 不同树种间凋落物在水体中的分解速率差异与其在林下分解的规律一致, 即阔叶树种叶片分解速率明显大于针叶树, 一个重要的原因是针叶树种叶片较小且表面革质化大大降低了分解初期的淋溶速率(Ibrahima et al ., 2008 ; 迟国梁和童晓立, 2010 )。
本文研究发现, 高山柳凋落物N动态在水体和林下变化趋势一致, 即直接释放; 而其他3个树种凋落物N动态在水体与林下分解过程中存在差异, 即林下凋落物N释放模式为: 富集—释放, 而水体中表现为: 释放—富集—释放。这与张川等(2015)的研究结果一致, 表明与林下凋落物分解相比, 在分解初期将有更多的可溶性物质会在水体的淋溶作用下分解和流失, 而需生物分解的部分则相应减少。然而, 凋落物P的释放模式在水体和陆地分解中比较一致, 均表现为: 释放-富集-释放模式。不过, 水体凋落物P释放的时间和量均大于林下, 而富集量却小于林下。这表明, 叶片凋落物尤其是阔叶树种叶片凋落物在湖泊中分解时, 大量可溶性P将在较短的时间内进入到水体中, 长期而言水体中凋落物的快速分解也将导致湖岸生态系统中P元素的大量亏缺。
九寨沟自然保护区森林资源丰富, 总面积达248 km2 (刘少英等, 2007 ), 每年有大量叶片凋落物进入到数量众多的高山湖泊中, 而较快的水体凋落物分解速率将使大量N和P等养分元素在较短的时间内汇集到湖泊生态系统中。这势必对高山湖泊中水生生物种群和结构产生影响, 从而对保护区内现有的水体景观产生潜在影响。因此, 加强保护区内高山湖泊叶片凋落物的管理是一项值得关注的工作。
致谢 感谢中国科学院成都生物研究所周祥玉、九寨沟国家自然保护区管理局肖维阳和任锦海在野外试验布置和数据采集等工作中给予的帮助; 感谢生态系统恢复与生物多样性保育四川省重点实验室在样品分析过程中提供的帮助。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Aerts R 1997 ). Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems: A triangular relationship
. Oikos , 79 , 439 -449 .
[本文引用: 1]
[2]
Aponte C Garcia LV Maranon T 2012 ). Tree species effect on litter decomposition and nutrient release in Mediterranean oak forests changes over time
.Ecosystems , 15 , 1204 -1218 .
[本文引用: 2]
[3]
Ayres E Dromph KM Bardgett RD 2006 ). Do plant species encourage soil biota that specialise in the rapid decomposition of their litter?
Soil Biology & Biochemistry , 38 , 183 -186 .
[本文引用: 1]
[4]
Berg B 2000 ). Litter decomposition and organic matter turnover in northern forest soils
.Forest Ecology and Management , 133 , 13 -22 .
[本文引用: 1]
[5]
Chen SX Jiang MX 2006 ). Leaf litter decomposition dynamics of different tree species in Xiangxi River watershed, the Three Gorges Region, China
.Acta Ecologica Sinica , 26 , 2905 -2912 . (in Chinese with English abstract)[陈书秀 , 江明喜 (2006 ). 三峡地区香溪河流域不同树种叶片凋落物的分解
. 生态学报 , 26 , 2905 -2912 .]
[6]
Chi GL Tong XL 2010 ). Leaching process of leaf litter in running water and lentic water in subtropical China
.Ecological Science , 29 , 50 -55 . (in Chinese with English abstract)[迟国梁 , 童晓立 (2010 ). 亚热带地区树叶凋落物在流水和静水环境中的淋溶规律
. 生态科学 , 29 , 50 -55 .]
[本文引用: 2]
[7]
Couteaux MM Bottner P Berg B 1995 ). Litter decomposition, climate and litter quality
.Trends in Ecology Evolution , 10 , 63 -66 .
[本文引用: 1]
[8]
Cornwell WK Cornelissen JHC Amatangelo K Dorrepaal E Eviner VT Godoy O Hobbie SE Hoorens B Kurokawa H Pérez-Harguindeguy N Quested HM Santiago LS Wardle D Wright IJ Aerts R Allison SD van Bodegom P Brovkin V Chatain A Callaghan TV Díaz S Garnier E Gurvich DE Kazakou E Klein JA Read J Reich PB Soudzilovskaia NA Vaieretti MV Westoby M 2008 ). Plant species traits are the predominant control on litter decomposition rates within biomes worldwide
.Ecology Letters , 11 , 1065 -1071 .
[本文引用: 3]
[9]
Deng CC Jiang XM Liu Y Zhang J Chen YM He RL 2015 ). Litter decomposition of Rhododendron lapponicum in alpine timberline ecotone
.Acta Ecologica Sinica , 35 , 1769 -1778 . (in Chinese with English abstract)[邓长春 , 蒋先敏 , 刘洋 , 张健 , 陈亚梅 , 和润莲 (2015 ). 高山林线交错带高山杜鹃的凋落物分解
. 生态学报 , 35 , 1769 -1778 .]
[本文引用: 2]
[10]
Graca MAS 2001 ). The role of invertebrates on leaf litter decomposition in streams—A review
.International Review of Hydrobiology , 86 , 383 -393 .
[本文引用: 1]
[11]
Guo ZL Zheng JP Ma YD Li QK Yu GR Han SJ Fan CN Liu WD 2006 ). Researches on litterfall decomposition rates and model simulating of main species in various forest vegetations of Changbai Mountains, China
.Acta Ecologica Sinica , 26 , 1037 -1046 . (in Chinese with English abstract)[郭忠玲 , 郑金萍 , 马元丹 , 李庆康 , 于贵瑞 , 韩士杰 , 范春楠 , 刘万德 (2006 ). 长白山各植被带主要树种凋落物分解速率及模型模拟的试验研究
. 生态学报 , 26 , 1037 -1046 .]
[本文引用: 1]
[12]
Handa T Aerts R Berendse F Berg MP Bruder A But- enschoen O Chauvet E Gessner MO Jabiol J Makkonen M McKie BG Malmqvist B Peeters ETHM Scheu S Schmid B van Ruijven J Vos VCA Hättenschwiler S 2014 ). Consequences of biodiversity loss for litter decomposition across biomes
.Nature , 59 , 218 -221 .
[本文引用: 1]
[13]
Hättenschwiler S Jørgensen HB 2010 ). Carbon quality rather than stoichiometry controls litter decomposition in a tropical rain forest
.Journal of Ecology , 98 , 754 -763 .
[14]
Hieber M Gessner MO 2002 ). Contribution of stream detrivores, fungi, and bacteria to leaf breakdown based on biomass estimates
.Ecology , 83 , 1026 .
[本文引用: 1]
[15]
Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P 2006 ). Tree species effects on decomposition and forest floor dynamics in a common garden
.Ecology , 87 , 2288 -2297 .
[本文引用: 1]
[16]
Ibrahima A Biyanzi P Halima M 2008 ). Changes in organic compounds during leaf litter leaching: Laboratory experiment on eight plant species of the Sudano-guinea Savannas of Ngaoundere, Cameroon. Iforest-
Biogeo- sciences and Forestry , 1 , 27 -33 .
[本文引用: 2]
[17]
Kay AD Mankowski J Hobbie SE 2008 ). Long-term burning interacts with herbivory to slow decomposition
.Ecology , 89 , 1188 -1194 .
[本文引用: 1]
[18]
Leroy CJ Marks JC 2006 ). Litter quality, stream character- istics and litter diversity influence decomposition rates and macroinvertebrates
.Freshwater Biology , 51 , 605 -617 .
[本文引用: 4]
[19]
Liu SY Zhang XP Zeng ZY 2007 ). Biodiversity of the Jiuzhaigou National Nature Reserve . Sichuan Science and Technology Press , Chengdu . (in Chinese)[刘少英 , 章小平 , 曾宗永 (2007 ). 九寨沟自然保护区的生物多样性 . 四川科学科技出版社 , 成都 .]
[本文引用: 1]
[20]
Liu X Jiang MX Deng HB 2008 ). Dynamics of nitrogen and phosphorus content during leaf litter decomposition in Xiangxi River watershed, the Three Gorges region
.Journal of Wuhan Botanical Research , 26 , 613 -619 . (in Chinese with English abstract)[刘昕 , 江明喜 , 邓红兵 (2008 ). 三峡地区香溪河流域叶片凋落物分解过程中N、P含量动态研究
. 武汉植物学研究 , 26 , 613 -619 .]
[本文引用: 1]
[21]
Lu RK 2000 ). Soil and Agro-chemical Analytical Methods . China Agricultural Science and Technology Press , Beijing . 302 -315 .[鲁如坤 (2000 ). 土壤农业化学分析方法 . 中国农业科技出版社 , 北京 . 302 -315 .]
[本文引用: 1]
[22]
Makkonen M Berg MP Handa T Hättenschwiler S van Ruijven J van Bodegom PM Aerts R 2012 ). Highly consistent effects of plant litter identity and functional traits on decomposition across a latitudinal gradient
.Ecology Letters , 15 , 1033 -1041 .
[本文引用: 3]
[23]
Martínez A Larrañaga A Pérez J Descals E Pozo J 2014 ). Temperature affects leaf litter decomposition in low-order forest streams: Field and microcosm approaches
.FEMS Microbiology Ecology , 87 , 257 -267 .
[本文引用: 1]
[24]
Moore TR Trofymow JA Prescott CE Fyles J Titus BD 2006 ). Patterns of carbon, nitrogen and phosphorus dynamics in decomposing foliar litter in Canadian forests
.Ecosystems , 9 , 46 -62 .
[本文引用: 1]
[25]
Moore TR Trofymow JA Taylor B Prescott C Camire C Duschene L Fyles J Kozak L Kranabetter M Morrison I Siltanen M Smith S Titus B Visser S Wein R Zoltai S 1999 ). Litter decomposition rates in Canadian forests
.Global Change Biology , 5 , 75 -82 .
[本文引用: 1]
[26]
Olson JS 1963 ). Energy storage and the balance of producers and decomposers in ecological systems
.Ecology , 44 , 322 -331 .
[本文引用: 1]
[27]
Osono T Azuma J Hirose D 2014 ). Plant species effect on the decomposition and chemical changes of leaf litter in grassland and pine and oak forest soils
.Plant and Soil , 376 , 411 -421 .
[本文引用: 1]
[28]
Parnas H 1975 ). Model for decomposition of organic material by microorganisms
.Soil Biology & Biochemistry , 7 , 161 -169 .
[29]
Parton W Silver WL Burke IC Grassens L Harmon ME Currie WS King JY Adair EC Brandt LA Hart SC Fasth B 2007 ). Global-scale similarities in nitrogen release patterns during long-term decomposition
.Science , 315 , 361 -364 .
[本文引用: 5]
[30]
Pascoal C Cassio F Gomes P 2001 ). Leaf breakdown rates: A measure of water quality?
International Review of Hydrobiology , 86 , 407 -416 .
[本文引用: 1]
[31]
Peng SL Liu Q 2002 ). The dynamics of forest litter and its responses to global warming
.Acta Ecologica Sinica , 22 , 1534 -1544 . (in Chinese with English abstract)[彭少麟 , 刘强 (2002 ). 森林凋落物动态及其对全球变暖的响应
. 生态学报 , 22 , 1534 -1544 .]
[本文引用: 1]
[32]
Petersen RC Cummins KW 1974 ). Leaf processing in a woodland stream
.Freshwater Biology , 4 , 343 -368 .
[本文引用: 4]
[33]
Purahong W Kapturska D Pecyna MJ Schulz E Schloter M Buscot F Hofrichter M Kruger D 2014 ). Influence of different forest system management practices on leaf litter decomposition rates, nutrient dynamics and the activity of ligninolytic enzymes: A case study from central European forests
.PLOS ONE , 9 , 1 -11 .
[本文引用: 1]
[34]
Santiago LS 2007 ). Extending the leaf economics spectrum to decomposition: Evidence from a tropical forest
.Ecology , 88 , 126 -1131 .
[本文引用: 2]
[35]
SAS Institute 2008 ). SAS 9.2 User's Guide. Carolina, USA.
[本文引用: 1]
[36]
Shi L Fan SH Jiang ZH Qi LH Liu GL 2015 ). Mixed leaf litter decomposition and N, P release with a focus on Phy- llostachys edulis (Carriere) J. Houz. forest in subtropical southeastern China
.Acta Societatis Botanicorum Poloniae , 84 , 207 -214 .
[本文引用: 1]
[37]
Tiegs SD Entrekin SA Reeves GH Kuntzsch D Merritt RW 2013 ). Litter decomposition, and associated invertebrate communities, in wetland ponds of the Copper River Delta, Alaska (USA)
.Wetlands , 33 , 1151 -1163 .
[本文引用: 2]
[38]
Wang J Huang JH 2001 ). Comparison of major nutrient release patterns in leaf litter decomposition in warm temperate zone of China
.Acta Phytoecologica Sinica , 25 , 375 -380 . (in Chinese with English abstract)[王瑾 , 黄建辉 (2001 ). 暖温带地区主要树种叶片凋落物分解过程中主要元素释放的比较
. 植物生态学报 , 25 , 375 -380 .]
[本文引用: 1]
[39]
Wang XE Xue L Xie TF 2009 ). A review on litter dec- omposition
.Chinese Journal of Soil Science , 40 , 1473 -1478 . (in Chinese with English abstract)[王相娥 , 薛立 , 谢腾芳 (2009 ). 凋落物分解研究综述
. 土壤通报 , 40 , 1473 -1478 .]
[本文引用: 1]
[40]
Wang XH Huang JJ Yan ER 2004 ). Leaf litter decomposition of common trees in Tiantong
.Acta Phytoecologica Sinica , 28 , 457 -467 . (in Chinese with English abstract)[王希华 , 黄建军 , 闫恩荣 (2004 ). 天童国家森林公园常见植物凋落叶分解的研究
. 植物生态学报 , 28 , 457 -467 .]
[本文引用: 2]
[41]
Wardle DA Nilsson MC Zackrisson O Gallet C 2003 ). Determinants of litter mixing effects in a Swedish boreal forest
.Soil Biology & Biochemistry , 35 , 827 -835 .
[本文引用: 1]
[42]
Wu QQ Wu FZ Yang WQ Xu ZF He W He M Zhao YY Zhu JX 2013 ). Effect of seasonal snow cover on litter decomposition in alpine forest
.Chinese Journal of Plant Ecology , 37 , 296 -305 . (in Chinese with English abstract)[武启骞 , 吴福忠 , 杨万勤 , 徐振锋 , 何伟 , 何敏 , 赵野逸 , 朱剑霄 (2013 ). 季节性雪被对高山森林凋落物分解的影响
. 植物生态学报 , 37 , 296 -305 .]
[本文引用: 1]
[43]
Yang YS Lin P Guo JF Lin RY Chen GS He ZM Xie JS 2003 ). Litter production, nutrient return and leaf-litter decomposition in natural and monoculture plantation forests of Castanopsis kawakamii in subtropical China
.Acta Ecologica Sinica , 23 , 1278 -1289 . (in Chinese with English abstract)[杨玉盛 , 林鹏 , 郭剑芬 , 林瑞余 , 陈光水 , 何宗明 , 谢锦升 (2003 ). 格氏栲天然林与人工林凋落物数量、养分归还及凋落叶分解
. 生态学报 , 23 , 1278 -1289 .]
[44]
Zhang C Yang WQ Yue K Huang CP Peng Y Wu FZ 2015 ). Soluble nitrogen and soluble phosphorus dynamics during foliar litter decomposition in winter in alpine forest streams
.Chinese Journal of Applied Ecology , 26 , 1601 -1608 . (in Chinese with English abstract)[张川 , 杨万勤 , 岳楷 , 黄春萍 , 彭艳 , 吴福忠 (2015 ). 高山森林溪流冬季不同时期凋落物分解中水溶性氮和磷的动态特征
. 应用生态学报 , 26 , 1601 -1608 .]
[本文引用: 2]
[45]
Zhou GY Guan LL Wei XH Tang XL Liu SG Liu JX Zhang DQ Yan JH 2008 ). Factors influencing leaf litter decomposition: An intersite decomposition experiment across China
.Plant and Soil , 311 , 61 -72 .
[本文引用: 4]
Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems: A triangular relationship
1
1997
... 叶片凋落物分解是陆地生态系统中一个重要的生态过程, 其分解速率对养分循环和生产力有着至关重要的作用(Parton et al ., 2007 ; Zhou et al ., 2008 ; Purahong et al ., 2014 ).凋落物分解将光合产物转变为无机养分和土壤有机质(Parton et al ., 2007 ), 提高了土壤肥力, 并提供植物生长所需的养分(王相娥等, 2009 ; Shi et al ., 2015 ).凋落物的分解是一个复杂的过程, 包括短暂的淋溶和长时间的生物分解阶段(Petersen & Cummins, 1974 ).通常情况下, 凋落物分解速率受凋落物质量、环境因子(如温度和湿度)和生物活动(如真菌和细菌)等多方面因素的综合影响(Couteaux et al ., 1995 ; Aerts, 1997 ; Wardle et al ., 2003 ; Ayres et al ., 2006 ; Zhou et al ., 2008 ; 武启骞等, 2013 ; 邓长春等, 2015 ).有研究表明, 凋落物的质量, 如初始N含量、木质素含量和C:N值等不但影响凋落物的分解速率(Aponte et al ., 2012 ; 邓长春等, 2015 ), 而且在很大程度上决定了其养分的固定和矿化(Parton et al ., 2007 ; Osono et al ., 2014 ).此外, 与叶片经济型谱密切联系的叶片功能性状(如叶片厚度与比叶面积)对其凋落物的分解过程也有着重要影响(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 ). ...
Tree species effect on litter decomposition and nutrient release in Mediterranean oak forests changes over time
2
2012
... 叶片凋落物分解是陆地生态系统中一个重要的生态过程, 其分解速率对养分循环和生产力有着至关重要的作用(Parton et al ., 2007 ; Zhou et al ., 2008 ; Purahong et al ., 2014 ).凋落物分解将光合产物转变为无机养分和土壤有机质(Parton et al ., 2007 ), 提高了土壤肥力, 并提供植物生长所需的养分(王相娥等, 2009 ; Shi et al ., 2015 ).凋落物的分解是一个复杂的过程, 包括短暂的淋溶和长时间的生物分解阶段(Petersen & Cummins, 1974 ).通常情况下, 凋落物分解速率受凋落物质量、环境因子(如温度和湿度)和生物活动(如真菌和细菌)等多方面因素的综合影响(Couteaux et al ., 1995 ; Aerts, 1997 ; Wardle et al ., 2003 ; Ayres et al ., 2006 ; Zhou et al ., 2008 ; 武启骞等, 2013 ; 邓长春等, 2015 ).有研究表明, 凋落物的质量, 如初始N含量、木质素含量和C:N值等不但影响凋落物的分解速率(Aponte et al ., 2012 ; 邓长春等, 2015 ), 而且在很大程度上决定了其养分的固定和矿化(Parton et al ., 2007 ; Osono et al ., 2014 ).此外, 与叶片经济型谱密切联系的叶片功能性状(如叶片厚度与比叶面积)对其凋落物的分解过程也有着重要影响(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 ). ...
... 凋落物自身往往缺乏大量元素, 如N、P和S等(Berg, 2000 ), 从而难以满足微生物的分解作用, 故需其在外界环境中固定相应的养分, 并达到某一阈值时才分解释放凋落物中的养分(Moore et al ., 2006 ; Parton et al ., 2007 ; Aponte et al ., 2012 ).Parnas (1975)认为凋落物C:N值是发生N固持和矿化的关键值, 当C:N值>30时发生N的固持, 而当C:N值<30时发生N的矿化.本文中高山柳凋落物初始C:N值(30.26)十分接近此值, 故在分解初期便出现N的释放, 而其他树种C:N值均大于30, 故先经历富集过程才进行N的释放.此外, 4个树种凋落物P在分解前期均存在明显的释放过程, 这一阶段主要受可溶性P淋溶作用的影响(彭少麟和刘强, 2002 ), 而在春季和夏季发生P的富集则与微生物活动加强有关(王希华等, 2004 ). ...
Do plant species encourage soil biota that specialise in the rapid decomposition of their litter?
1
2006
... 叶片凋落物分解是陆地生态系统中一个重要的生态过程, 其分解速率对养分循环和生产力有着至关重要的作用(Parton et al ., 2007 ; Zhou et al ., 2008 ; Purahong et al ., 2014 ).凋落物分解将光合产物转变为无机养分和土壤有机质(Parton et al ., 2007 ), 提高了土壤肥力, 并提供植物生长所需的养分(王相娥等, 2009 ; Shi et al ., 2015 ).凋落物的分解是一个复杂的过程, 包括短暂的淋溶和长时间的生物分解阶段(Petersen & Cummins, 1974 ).通常情况下, 凋落物分解速率受凋落物质量、环境因子(如温度和湿度)和生物活动(如真菌和细菌)等多方面因素的综合影响(Couteaux et al ., 1995 ; Aerts, 1997 ; Wardle et al ., 2003 ; Ayres et al ., 2006 ; Zhou et al ., 2008 ; 武启骞等, 2013 ; 邓长春等, 2015 ).有研究表明, 凋落物的质量, 如初始N含量、木质素含量和C:N值等不但影响凋落物的分解速率(Aponte et al ., 2012 ; 邓长春等, 2015 ), 而且在很大程度上决定了其养分的固定和矿化(Parton et al ., 2007 ; Osono et al ., 2014 ).此外, 与叶片经济型谱密切联系的叶片功能性状(如叶片厚度与比叶面积)对其凋落物的分解过程也有着重要影响(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 ). ...
Litter decomposition and organic matter turnover in northern forest soils
1
2000
... 凋落物自身往往缺乏大量元素, 如N、P和S等(Berg, 2000 ), 从而难以满足微生物的分解作用, 故需其在外界环境中固定相应的养分, 并达到某一阈值时才分解释放凋落物中的养分(Moore et al ., 2006 ; Parton et al ., 2007 ; Aponte et al ., 2012 ).Parnas (1975)认为凋落物C:N值是发生N固持和矿化的关键值, 当C:N值>30时发生N的固持, 而当C:N值<30时发生N的矿化.本文中高山柳凋落物初始C:N值(30.26)十分接近此值, 故在分解初期便出现N的释放, 而其他树种C:N值均大于30, 故先经历富集过程才进行N的释放.此外, 4个树种凋落物P在分解前期均存在明显的释放过程, 这一阶段主要受可溶性P淋溶作用的影响(彭少麟和刘强, 2002 ), 而在春季和夏季发生P的富集则与微生物活动加强有关(王希华等, 2004 ). ...
三峡地区香溪河流域不同树种叶片凋落物的分解
2006
亚热带地区树叶凋落物在流水和静水环境中的淋溶规律
2
2010
... 在水生生态系统中, 叶片凋落物作为主要的外源物质为其提供了大量的营养物质(Leroy & Marks, 2006 ), 且叶片的种类将影响着水体微生物及底栖无脊椎动物等的种群结构和数量(Graca, 2001 ; Hieber & Gessner, 2002 ).叶片凋落物在水体中的分解主要受其自身的品质和水体特征的影响.比如, 叶片大小、吸水性和可溶性物质含量将影响分解初期的淋溶速率(Ibrahima et al ., 2008 ); 叶片凋落物初始养分含量将影响其分解速率和养分的释放模式(Pascoal et al ., 2001 ; 刘昕等, 2008 ); 水温和流速也在很大程度上影响着凋落物的分解(Leroy & Marks, 2006 ; Martínez et al ., 2014 ).此外, 水生生态系统周围的植被构成及其季节变化将直接影响进入水体的叶片凋落物类型, 从而影响水体中生物种群变化及物质和能量循环(迟国梁和童晓立, 2010 ).与陆地生态系统相比, 由于水体的浸泡和流动冲击作用, 凋落物在水生生态系统中具有较快的分解速率(Petersen & Cummins, 1974 ; 张川等, 2015 ), 同时, 微生物和食碎屑动物的种类和数量在两种生态系统中的差异也在很大程度上影响着凋落物的分解过程(Leroy & Marks, 2006 ; Tiegs et al ., 2013 ; Handa et al ., 2014 ).目前, 关于叶片凋落物在水体中的分解速率及养分释放特征研究还相对较少.因此, 加强水体叶片凋落物分解特征研究, 尤其针对重要的水生生态系统开展相关研究具有重要意义. ...
... 4个树种叶片凋落物分解速率在高山湖泊中的分解速率均极显著高于其在林下的分解速率(p < 0.01, 表3 ; 图2 ), 且各物种凋落物在林下完全分解(99%)所需的时间是其在水体中的1.5倍左右, 这与陈书秀和江明喜(2006)对三峡地区香溪河流域不同树种叶片凋落物分解的研究结果一致.湖泊中凋落物分解速率较快的一个重要原因是凋落物长时间浸泡在水体中, 较强的淋溶作用会加速可溶性物质在分解初期的快速分解, 加之水体的流动将进一步加快这一过程的进行(Petersen & Cummins, 1974 ; 张川等, 2015 ).其次, 水体微生物和无脊椎动物在凋落物分解过程中也起着至关重要的作用(Leroy & Marks, 2006 ; Tiegs et al ., 2013 ), 相对稳定的水体温度(冬季湖泊中平均水温仍保持在2 ℃以上, 图1 )为凋落物分解过程中的生物活动提供了良好的环境保障.本文结果显示, 冬季水体凋落物的失重率保持在一个较高水平.此外, 不同树种间凋落物在水体中的分解速率差异与其在林下分解的规律一致, 即阔叶树种叶片分解速率明显大于针叶树, 一个重要的原因是针叶树种叶片较小且表面革质化大大降低了分解初期的淋溶速率(Ibrahima et al ., 2008 ; 迟国梁和童晓立, 2010 ). ...
Litter decomposition, climate and litter quality
1
1995
... 叶片凋落物分解是陆地生态系统中一个重要的生态过程, 其分解速率对养分循环和生产力有着至关重要的作用(Parton et al ., 2007 ; Zhou et al ., 2008 ; Purahong et al ., 2014 ).凋落物分解将光合产物转变为无机养分和土壤有机质(Parton et al ., 2007 ), 提高了土壤肥力, 并提供植物生长所需的养分(王相娥等, 2009 ; Shi et al ., 2015 ).凋落物的分解是一个复杂的过程, 包括短暂的淋溶和长时间的生物分解阶段(Petersen & Cummins, 1974 ).通常情况下, 凋落物分解速率受凋落物质量、环境因子(如温度和湿度)和生物活动(如真菌和细菌)等多方面因素的综合影响(Couteaux et al ., 1995 ; Aerts, 1997 ; Wardle et al ., 2003 ; Ayres et al ., 2006 ; Zhou et al ., 2008 ; 武启骞等, 2013 ; 邓长春等, 2015 ).有研究表明, 凋落物的质量, 如初始N含量、木质素含量和C:N值等不但影响凋落物的分解速率(Aponte et al ., 2012 ; 邓长春等, 2015 ), 而且在很大程度上决定了其养分的固定和矿化(Parton et al ., 2007 ; Osono et al ., 2014 ).此外, 与叶片经济型谱密切联系的叶片功能性状(如叶片厚度与比叶面积)对其凋落物的分解过程也有着重要影响(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 ). ...
Plant species traits are the predominant control on litter decomposition rates within biomes worldwide
3
2008
... 叶片凋落物分解是陆地生态系统中一个重要的生态过程, 其分解速率对养分循环和生产力有着至关重要的作用(Parton et al ., 2007 ; Zhou et al ., 2008 ; Purahong et al ., 2014 ).凋落物分解将光合产物转变为无机养分和土壤有机质(Parton et al ., 2007 ), 提高了土壤肥力, 并提供植物生长所需的养分(王相娥等, 2009 ; Shi et al ., 2015 ).凋落物的分解是一个复杂的过程, 包括短暂的淋溶和长时间的生物分解阶段(Petersen & Cummins, 1974 ).通常情况下, 凋落物分解速率受凋落物质量、环境因子(如温度和湿度)和生物活动(如真菌和细菌)等多方面因素的综合影响(Couteaux et al ., 1995 ; Aerts, 1997 ; Wardle et al ., 2003 ; Ayres et al ., 2006 ; Zhou et al ., 2008 ; 武启骞等, 2013 ; 邓长春等, 2015 ).有研究表明, 凋落物的质量, 如初始N含量、木质素含量和C:N值等不但影响凋落物的分解速率(Aponte et al ., 2012 ; 邓长春等, 2015 ), 而且在很大程度上决定了其养分的固定和矿化(Parton et al ., 2007 ; Osono et al ., 2014 ).此外, 与叶片经济型谱密切联系的叶片功能性状(如叶片厚度与比叶面积)对其凋落物的分解过程也有着重要影响(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 ). ...
... 本文结果显示, 叶片凋落物分解1年后MR 同其初始C:N值存在正相关关系, 而与初始N含量成负相关关系(表5 ), 表明凋落物初始养分含量, 尤其是N含量在很大程度上影响着凋落物的分解速率, 这与杨玉盛等(2003)和王希华等(2004)的研究结果一致.值得注意的是, 虽然黄果冷杉叶片凋落物初始N含量较高, 且具有较小的C:N值, 但其凋落物分解速率仍较低.因此, 除初始养分含量外的其他因子在凋落分解过程中起着重要的作用.Hättenschwiler 和Jørgensen (2010)对热带雨林叶片凋落物分解特征研究发现, 凋落物中碳的质量对分解速率的影响比其化学计量特征(C、N和P含量等)更大.此外, 王希华等(2004)和Santiago (2007)研究发现凋落物比叶面积与凋落物分解速率之间存在显著的正相关关系, 较大的比叶面积意味着单位质量凋落物具有更大的环境接触面积, 一方面加大了雨水的淋溶作用, 另一方面有利于微生物定居和土壤动物的碎裂作用, 从而大大提高凋落分解速率(郭忠玲等, 2006 ).也有研究表明, 叶片厚度与凋落物分解速率存在明显的负相关关系(Cornwell et al ., 2008 ).因此, 与叶片经济型谱存在密切联系的叶片功能特征(如比叶面积、叶片氮素含量和叶片厚度等)在叶片凋落物分解过程中起着重要作用(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 ).综上所述, 叶片凋落物分解是一个受其自身性质和外界环境因素共同作用的复杂过程, 它对陆地生态系统的养分循环有着深远影响(Kay et al ., 2008 ; Makkonen et al ., 2012 ). ...
... ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 ).综上所述, 叶片凋落物分解是一个受其自身性质和外界环境因素共同作用的复杂过程, 它对陆地生态系统的养分循环有着深远影响(Kay et al ., 2008 ; Makkonen et al ., 2012 ). ...
高山林线交错带高山杜鹃的凋落物分解
2
2015
... 叶片凋落物分解是陆地生态系统中一个重要的生态过程, 其分解速率对养分循环和生产力有着至关重要的作用(Parton et al ., 2007 ; Zhou et al ., 2008 ; Purahong et al ., 2014 ).凋落物分解将光合产物转变为无机养分和土壤有机质(Parton et al ., 2007 ), 提高了土壤肥力, 并提供植物生长所需的养分(王相娥等, 2009 ; Shi et al ., 2015 ).凋落物的分解是一个复杂的过程, 包括短暂的淋溶和长时间的生物分解阶段(Petersen & Cummins, 1974 ).通常情况下, 凋落物分解速率受凋落物质量、环境因子(如温度和湿度)和生物活动(如真菌和细菌)等多方面因素的综合影响(Couteaux et al ., 1995 ; Aerts, 1997 ; Wardle et al ., 2003 ; Ayres et al ., 2006 ; Zhou et al ., 2008 ; 武启骞等, 2013 ; 邓长春等, 2015 ).有研究表明, 凋落物的质量, 如初始N含量、木质素含量和C:N值等不但影响凋落物的分解速率(Aponte et al ., 2012 ; 邓长春等, 2015 ), 而且在很大程度上决定了其养分的固定和矿化(Parton et al ., 2007 ; Osono et al ., 2014 ).此外, 与叶片经济型谱密切联系的叶片功能性状(如叶片厚度与比叶面积)对其凋落物的分解过程也有着重要影响(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 ). ...
... ; 邓长春等, 2015 ), 而且在很大程度上决定了其养分的固定和矿化(Parton et al ., 2007 ; Osono et al ., 2014 ).此外, 与叶片经济型谱密切联系的叶片功能性状(如叶片厚度与比叶面积)对其凋落物的分解过程也有着重要影响(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 ). ...
The role of invertebrates on leaf litter decomposition in streams—A review
1
2001
... 在水生生态系统中, 叶片凋落物作为主要的外源物质为其提供了大量的营养物质(Leroy & Marks, 2006 ), 且叶片的种类将影响着水体微生物及底栖无脊椎动物等的种群结构和数量(Graca, 2001 ; Hieber & Gessner, 2002 ).叶片凋落物在水体中的分解主要受其自身的品质和水体特征的影响.比如, 叶片大小、吸水性和可溶性物质含量将影响分解初期的淋溶速率(Ibrahima et al ., 2008 ); 叶片凋落物初始养分含量将影响其分解速率和养分的释放模式(Pascoal et al ., 2001 ; 刘昕等, 2008 ); 水温和流速也在很大程度上影响着凋落物的分解(Leroy & Marks, 2006 ; Martínez et al ., 2014 ).此外, 水生生态系统周围的植被构成及其季节变化将直接影响进入水体的叶片凋落物类型, 从而影响水体中生物种群变化及物质和能量循环(迟国梁和童晓立, 2010 ).与陆地生态系统相比, 由于水体的浸泡和流动冲击作用, 凋落物在水生生态系统中具有较快的分解速率(Petersen & Cummins, 1974 ; 张川等, 2015 ), 同时, 微生物和食碎屑动物的种类和数量在两种生态系统中的差异也在很大程度上影响着凋落物的分解过程(Leroy & Marks, 2006 ; Tiegs et al ., 2013 ; Handa et al ., 2014 ).目前, 关于叶片凋落物在水体中的分解速率及养分释放特征研究还相对较少.因此, 加强水体叶片凋落物分解特征研究, 尤其针对重要的水生生态系统开展相关研究具有重要意义. ...
长白山各植被带主要树种凋落物分解速率及模型模拟的试验研究
1
2006
... 本文结果显示, 叶片凋落物分解1年后MR 同其初始C:N值存在正相关关系, 而与初始N含量成负相关关系(表5 ), 表明凋落物初始养分含量, 尤其是N含量在很大程度上影响着凋落物的分解速率, 这与杨玉盛等(2003)和王希华等(2004)的研究结果一致.值得注意的是, 虽然黄果冷杉叶片凋落物初始N含量较高, 且具有较小的C:N值, 但其凋落物分解速率仍较低.因此, 除初始养分含量外的其他因子在凋落分解过程中起着重要的作用.Hättenschwiler 和Jørgensen (2010)对热带雨林叶片凋落物分解特征研究发现, 凋落物中碳的质量对分解速率的影响比其化学计量特征(C、N和P含量等)更大.此外, 王希华等(2004)和Santiago (2007)研究发现凋落物比叶面积与凋落物分解速率之间存在显著的正相关关系, 较大的比叶面积意味着单位质量凋落物具有更大的环境接触面积, 一方面加大了雨水的淋溶作用, 另一方面有利于微生物定居和土壤动物的碎裂作用, 从而大大提高凋落分解速率(郭忠玲等, 2006 ).也有研究表明, 叶片厚度与凋落物分解速率存在明显的负相关关系(Cornwell et al ., 2008 ).因此, 与叶片经济型谱存在密切联系的叶片功能特征(如比叶面积、叶片氮素含量和叶片厚度等)在叶片凋落物分解过程中起着重要作用(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 ).综上所述, 叶片凋落物分解是一个受其自身性质和外界环境因素共同作用的复杂过程, 它对陆地生态系统的养分循环有着深远影响(Kay et al ., 2008 ; Makkonen et al ., 2012 ). ...
Consequences of biodiversity loss for litter decomposition across biomes
1
2014
... 在水生生态系统中, 叶片凋落物作为主要的外源物质为其提供了大量的营养物质(Leroy & Marks, 2006 ), 且叶片的种类将影响着水体微生物及底栖无脊椎动物等的种群结构和数量(Graca, 2001 ; Hieber & Gessner, 2002 ).叶片凋落物在水体中的分解主要受其自身的品质和水体特征的影响.比如, 叶片大小、吸水性和可溶性物质含量将影响分解初期的淋溶速率(Ibrahima et al ., 2008 ); 叶片凋落物初始养分含量将影响其分解速率和养分的释放模式(Pascoal et al ., 2001 ; 刘昕等, 2008 ); 水温和流速也在很大程度上影响着凋落物的分解(Leroy & Marks, 2006 ; Martínez et al ., 2014 ).此外, 水生生态系统周围的植被构成及其季节变化将直接影响进入水体的叶片凋落物类型, 从而影响水体中生物种群变化及物质和能量循环(迟国梁和童晓立, 2010 ).与陆地生态系统相比, 由于水体的浸泡和流动冲击作用, 凋落物在水生生态系统中具有较快的分解速率(Petersen & Cummins, 1974 ; 张川等, 2015 ), 同时, 微生物和食碎屑动物的种类和数量在两种生态系统中的差异也在很大程度上影响着凋落物的分解过程(Leroy & Marks, 2006 ; Tiegs et al ., 2013 ; Handa et al ., 2014 ).目前, 关于叶片凋落物在水体中的分解速率及养分释放特征研究还相对较少.因此, 加强水体叶片凋落物分解特征研究, 尤其针对重要的水生生态系统开展相关研究具有重要意义. ...
Carbon quality rather than stoichiometry controls litter decomposition in a tropical rain forest
2010
Contribution of stream detrivores, fungi, and bacteria to leaf breakdown based on biomass estimates
1
2002
... 在水生生态系统中, 叶片凋落物作为主要的外源物质为其提供了大量的营养物质(Leroy & Marks, 2006 ), 且叶片的种类将影响着水体微生物及底栖无脊椎动物等的种群结构和数量(Graca, 2001 ; Hieber & Gessner, 2002 ).叶片凋落物在水体中的分解主要受其自身的品质和水体特征的影响.比如, 叶片大小、吸水性和可溶性物质含量将影响分解初期的淋溶速率(Ibrahima et al ., 2008 ); 叶片凋落物初始养分含量将影响其分解速率和养分的释放模式(Pascoal et al ., 2001 ; 刘昕等, 2008 ); 水温和流速也在很大程度上影响着凋落物的分解(Leroy & Marks, 2006 ; Martínez et al ., 2014 ).此外, 水生生态系统周围的植被构成及其季节变化将直接影响进入水体的叶片凋落物类型, 从而影响水体中生物种群变化及物质和能量循环(迟国梁和童晓立, 2010 ).与陆地生态系统相比, 由于水体的浸泡和流动冲击作用, 凋落物在水生生态系统中具有较快的分解速率(Petersen & Cummins, 1974 ; 张川等, 2015 ), 同时, 微生物和食碎屑动物的种类和数量在两种生态系统中的差异也在很大程度上影响着凋落物的分解过程(Leroy & Marks, 2006 ; Tiegs et al ., 2013 ; Handa et al ., 2014 ).目前, 关于叶片凋落物在水体中的分解速率及养分释放特征研究还相对较少.因此, 加强水体叶片凋落物分解特征研究, 尤其针对重要的水生生态系统开展相关研究具有重要意义. ...
Tree species effects on decomposition and forest floor dynamics in a common garden
1
2006
... 本文通过研究九寨沟国家自然保护区内4种常见林型代表性树种, 即寒温性针叶林(黄果冷杉)、温性针叶林(油松)、落叶阔叶林(红桦)和落叶阔叶灌丛(高山柳)的叶片凋落物分解特征, 得出以下结果: 各树种间凋落物分解系数k 高山柳>红桦>黄果冷杉>油松, 且各物种间凋落物MR 差异极显著(p < 0.01, 图2 ).结果表明, 叶片凋落物分解速率存在明显的物种差异, 且阔叶树种叶片分解速率显著大于针叶树种, 这与郭忠玲等(2006)对长白山各植被带主要树种凋落物分解速率的研究结果一致.此外, 各树种叶片凋落物在前3个月质量损失最快, 而后迅速减慢, 第6-12月又逐渐增大(图2 ).这与Zhou等(2008)和Aponte等(2012)的研究结果一致, 凋落物分解初期可溶性物质会迅速通过淋溶作用进入到生态系统中, 而后进入较长且缓慢的生物分解过程(Petersen & Cummins, 1974 ).在本文中, 凋落物质量损失变化与外界环境温度的变化趋势一致, 且在降水丰沛的季节(5-9月)较高.这一结果很好地印证了温度和降水量作为主要的环境影响因子控制着凋落物的分解速率(Moore et al ., 1999 ; Hobbie et al ., 2006 ; Parton et al ., 2007 ). ...
Changes in organic compounds during leaf litter leaching: Laboratory experiment on eight plant species of the Sudano-guinea Savannas of Ngaoundere, Cameroon. Iforest-
2
2008
... 在水生生态系统中, 叶片凋落物作为主要的外源物质为其提供了大量的营养物质(Leroy & Marks, 2006 ), 且叶片的种类将影响着水体微生物及底栖无脊椎动物等的种群结构和数量(Graca, 2001 ; Hieber & Gessner, 2002 ).叶片凋落物在水体中的分解主要受其自身的品质和水体特征的影响.比如, 叶片大小、吸水性和可溶性物质含量将影响分解初期的淋溶速率(Ibrahima et al ., 2008 ); 叶片凋落物初始养分含量将影响其分解速率和养分的释放模式(Pascoal et al ., 2001 ; 刘昕等, 2008 ); 水温和流速也在很大程度上影响着凋落物的分解(Leroy & Marks, 2006 ; Martínez et al ., 2014 ).此外, 水生生态系统周围的植被构成及其季节变化将直接影响进入水体的叶片凋落物类型, 从而影响水体中生物种群变化及物质和能量循环(迟国梁和童晓立, 2010 ).与陆地生态系统相比, 由于水体的浸泡和流动冲击作用, 凋落物在水生生态系统中具有较快的分解速率(Petersen & Cummins, 1974 ; 张川等, 2015 ), 同时, 微生物和食碎屑动物的种类和数量在两种生态系统中的差异也在很大程度上影响着凋落物的分解过程(Leroy & Marks, 2006 ; Tiegs et al ., 2013 ; Handa et al ., 2014 ).目前, 关于叶片凋落物在水体中的分解速率及养分释放特征研究还相对较少.因此, 加强水体叶片凋落物分解特征研究, 尤其针对重要的水生生态系统开展相关研究具有重要意义. ...
... 4个树种叶片凋落物分解速率在高山湖泊中的分解速率均极显著高于其在林下的分解速率(p < 0.01, 表3 ; 图2 ), 且各物种凋落物在林下完全分解(99%)所需的时间是其在水体中的1.5倍左右, 这与陈书秀和江明喜(2006)对三峡地区香溪河流域不同树种叶片凋落物分解的研究结果一致.湖泊中凋落物分解速率较快的一个重要原因是凋落物长时间浸泡在水体中, 较强的淋溶作用会加速可溶性物质在分解初期的快速分解, 加之水体的流动将进一步加快这一过程的进行(Petersen & Cummins, 1974 ; 张川等, 2015 ).其次, 水体微生物和无脊椎动物在凋落物分解过程中也起着至关重要的作用(Leroy & Marks, 2006 ; Tiegs et al ., 2013 ), 相对稳定的水体温度(冬季湖泊中平均水温仍保持在2 ℃以上, 图1 )为凋落物分解过程中的生物活动提供了良好的环境保障.本文结果显示, 冬季水体凋落物的失重率保持在一个较高水平.此外, 不同树种间凋落物在水体中的分解速率差异与其在林下分解的规律一致, 即阔叶树种叶片分解速率明显大于针叶树, 一个重要的原因是针叶树种叶片较小且表面革质化大大降低了分解初期的淋溶速率(Ibrahima et al ., 2008 ; 迟国梁和童晓立, 2010 ). ...
Long-term burning interacts with herbivory to slow decomposition
1
2008
... 本文结果显示, 叶片凋落物分解1年后MR 同其初始C:N值存在正相关关系, 而与初始N含量成负相关关系(表5 ), 表明凋落物初始养分含量, 尤其是N含量在很大程度上影响着凋落物的分解速率, 这与杨玉盛等(2003)和王希华等(2004)的研究结果一致.值得注意的是, 虽然黄果冷杉叶片凋落物初始N含量较高, 且具有较小的C:N值, 但其凋落物分解速率仍较低.因此, 除初始养分含量外的其他因子在凋落分解过程中起着重要的作用.Hättenschwiler 和Jørgensen (2010)对热带雨林叶片凋落物分解特征研究发现, 凋落物中碳的质量对分解速率的影响比其化学计量特征(C、N和P含量等)更大.此外, 王希华等(2004)和Santiago (2007)研究发现凋落物比叶面积与凋落物分解速率之间存在显著的正相关关系, 较大的比叶面积意味着单位质量凋落物具有更大的环境接触面积, 一方面加大了雨水的淋溶作用, 另一方面有利于微生物定居和土壤动物的碎裂作用, 从而大大提高凋落分解速率(郭忠玲等, 2006 ).也有研究表明, 叶片厚度与凋落物分解速率存在明显的负相关关系(Cornwell et al ., 2008 ).因此, 与叶片经济型谱存在密切联系的叶片功能特征(如比叶面积、叶片氮素含量和叶片厚度等)在叶片凋落物分解过程中起着重要作用(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 ).综上所述, 叶片凋落物分解是一个受其自身性质和外界环境因素共同作用的复杂过程, 它对陆地生态系统的养分循环有着深远影响(Kay et al ., 2008 ; Makkonen et al ., 2012 ). ...
Litter quality, stream character- istics and litter diversity influence decomposition rates and macroinvertebrates
4
2006
... 在水生生态系统中, 叶片凋落物作为主要的外源物质为其提供了大量的营养物质(Leroy & Marks, 2006 ), 且叶片的种类将影响着水体微生物及底栖无脊椎动物等的种群结构和数量(Graca, 2001 ; Hieber & Gessner, 2002 ).叶片凋落物在水体中的分解主要受其自身的品质和水体特征的影响.比如, 叶片大小、吸水性和可溶性物质含量将影响分解初期的淋溶速率(Ibrahima et al ., 2008 ); 叶片凋落物初始养分含量将影响其分解速率和养分的释放模式(Pascoal et al ., 2001 ; 刘昕等, 2008 ); 水温和流速也在很大程度上影响着凋落物的分解(Leroy & Marks, 2006 ; Martínez et al ., 2014 ).此外, 水生生态系统周围的植被构成及其季节变化将直接影响进入水体的叶片凋落物类型, 从而影响水体中生物种群变化及物质和能量循环(迟国梁和童晓立, 2010 ).与陆地生态系统相比, 由于水体的浸泡和流动冲击作用, 凋落物在水生生态系统中具有较快的分解速率(Petersen & Cummins, 1974 ; 张川等, 2015 ), 同时, 微生物和食碎屑动物的种类和数量在两种生态系统中的差异也在很大程度上影响着凋落物的分解过程(Leroy & Marks, 2006 ; Tiegs et al ., 2013 ; Handa et al ., 2014 ).目前, 关于叶片凋落物在水体中的分解速率及养分释放特征研究还相对较少.因此, 加强水体叶片凋落物分解特征研究, 尤其针对重要的水生生态系统开展相关研究具有重要意义. ...
... ); 水温和流速也在很大程度上影响着凋落物的分解(Leroy & Marks, 2006 ; Martínez et al ., 2014 ).此外, 水生生态系统周围的植被构成及其季节变化将直接影响进入水体的叶片凋落物类型, 从而影响水体中生物种群变化及物质和能量循环(迟国梁和童晓立, 2010 ).与陆地生态系统相比, 由于水体的浸泡和流动冲击作用, 凋落物在水生生态系统中具有较快的分解速率(Petersen & Cummins, 1974 ; 张川等, 2015 ), 同时, 微生物和食碎屑动物的种类和数量在两种生态系统中的差异也在很大程度上影响着凋落物的分解过程(Leroy & Marks, 2006 ; Tiegs et al ., 2013 ; Handa et al ., 2014 ).目前, 关于叶片凋落物在水体中的分解速率及养分释放特征研究还相对较少.因此, 加强水体叶片凋落物分解特征研究, 尤其针对重要的水生生态系统开展相关研究具有重要意义. ...
... ), 同时, 微生物和食碎屑动物的种类和数量在两种生态系统中的差异也在很大程度上影响着凋落物的分解过程(Leroy & Marks, 2006 ; Tiegs et al ., 2013 ; Handa et al ., 2014 ).目前, 关于叶片凋落物在水体中的分解速率及养分释放特征研究还相对较少.因此, 加强水体叶片凋落物分解特征研究, 尤其针对重要的水生生态系统开展相关研究具有重要意义. ...
... 4个树种叶片凋落物分解速率在高山湖泊中的分解速率均极显著高于其在林下的分解速率(p < 0.01, 表3 ; 图2 ), 且各物种凋落物在林下完全分解(99%)所需的时间是其在水体中的1.5倍左右, 这与陈书秀和江明喜(2006)对三峡地区香溪河流域不同树种叶片凋落物分解的研究结果一致.湖泊中凋落物分解速率较快的一个重要原因是凋落物长时间浸泡在水体中, 较强的淋溶作用会加速可溶性物质在分解初期的快速分解, 加之水体的流动将进一步加快这一过程的进行(Petersen & Cummins, 1974 ; 张川等, 2015 ).其次, 水体微生物和无脊椎动物在凋落物分解过程中也起着至关重要的作用(Leroy & Marks, 2006 ; Tiegs et al ., 2013 ), 相对稳定的水体温度(冬季湖泊中平均水温仍保持在2 ℃以上, 图1 )为凋落物分解过程中的生物活动提供了良好的环境保障.本文结果显示, 冬季水体凋落物的失重率保持在一个较高水平.此外, 不同树种间凋落物在水体中的分解速率差异与其在林下分解的规律一致, 即阔叶树种叶片分解速率明显大于针叶树, 一个重要的原因是针叶树种叶片较小且表面革质化大大降低了分解初期的淋溶速率(Ibrahima et al ., 2008 ; 迟国梁和童晓立, 2010 ). ...
1
2007
... 九寨沟自然保护区森林资源丰富, 总面积达248 km2 (刘少英等, 2007 ), 每年有大量叶片凋落物进入到数量众多的高山湖泊中, 而较快的水体凋落物分解速率将使大量N和P等养分元素在较短的时间内汇集到湖泊生态系统中.这势必对高山湖泊中水生生物种群和结构产生影响, 从而对保护区内现有的水体景观产生潜在影响.因此, 加强保护区内高山湖泊叶片凋落物的管理是一项值得关注的工作. ...
三峡地区香溪河流域叶片凋落物分解过程中N、P含量动态研究
1
2008
... 在水生生态系统中, 叶片凋落物作为主要的外源物质为其提供了大量的营养物质(Leroy & Marks, 2006 ), 且叶片的种类将影响着水体微生物及底栖无脊椎动物等的种群结构和数量(Graca, 2001 ; Hieber & Gessner, 2002 ).叶片凋落物在水体中的分解主要受其自身的品质和水体特征的影响.比如, 叶片大小、吸水性和可溶性物质含量将影响分解初期的淋溶速率(Ibrahima et al ., 2008 ); 叶片凋落物初始养分含量将影响其分解速率和养分的释放模式(Pascoal et al ., 2001 ; 刘昕等, 2008 ); 水温和流速也在很大程度上影响着凋落物的分解(Leroy & Marks, 2006 ; Martínez et al ., 2014 ).此外, 水生生态系统周围的植被构成及其季节变化将直接影响进入水体的叶片凋落物类型, 从而影响水体中生物种群变化及物质和能量循环(迟国梁和童晓立, 2010 ).与陆地生态系统相比, 由于水体的浸泡和流动冲击作用, 凋落物在水生生态系统中具有较快的分解速率(Petersen & Cummins, 1974 ; 张川等, 2015 ), 同时, 微生物和食碎屑动物的种类和数量在两种生态系统中的差异也在很大程度上影响着凋落物的分解过程(Leroy & Marks, 2006 ; Tiegs et al ., 2013 ; Handa et al ., 2014 ).目前, 关于叶片凋落物在水体中的分解速率及养分释放特征研究还相对较少.因此, 加强水体叶片凋落物分解特征研究, 尤其针对重要的水生生态系统开展相关研究具有重要意义. ...
1
2000
... 2014年9月15日, 分别于黄果冷杉林、油松林、红桦林和高山柳灌丛下收集当年叶片凋落物, 放入烘箱65 ℃烘干至恒质量, 然后用电子天平准确称取5.00 g, 装入宽15 cm、长20 cm、孔径为1 mm的尼龙网袋中, 每一物种装取72袋.采用重铬酸钾-硫酸氧化法、H2 SO4 -H2 O2 -扩散法和钼锑钪比色法分别测定叶片凋落物初始全碳、全氮和全磷含量(鲁如坤, 2000 ). ...
Highly consistent effects of plant litter identity and functional traits on decomposition across a latitudinal gradient
3
2012
... 叶片凋落物分解是陆地生态系统中一个重要的生态过程, 其分解速率对养分循环和生产力有着至关重要的作用(Parton et al ., 2007 ; Zhou et al ., 2008 ; Purahong et al ., 2014 ).凋落物分解将光合产物转变为无机养分和土壤有机质(Parton et al ., 2007 ), 提高了土壤肥力, 并提供植物生长所需的养分(王相娥等, 2009 ; Shi et al ., 2015 ).凋落物的分解是一个复杂的过程, 包括短暂的淋溶和长时间的生物分解阶段(Petersen & Cummins, 1974 ).通常情况下, 凋落物分解速率受凋落物质量、环境因子(如温度和湿度)和生物活动(如真菌和细菌)等多方面因素的综合影响(Couteaux et al ., 1995 ; Aerts, 1997 ; Wardle et al ., 2003 ; Ayres et al ., 2006 ; Zhou et al ., 2008 ; 武启骞等, 2013 ; 邓长春等, 2015 ).有研究表明, 凋落物的质量, 如初始N含量、木质素含量和C:N值等不但影响凋落物的分解速率(Aponte et al ., 2012 ; 邓长春等, 2015 ), 而且在很大程度上决定了其养分的固定和矿化(Parton et al ., 2007 ; Osono et al ., 2014 ).此外, 与叶片经济型谱密切联系的叶片功能性状(如叶片厚度与比叶面积)对其凋落物的分解过程也有着重要影响(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 ). ...
... 本文结果显示, 叶片凋落物分解1年后MR 同其初始C:N值存在正相关关系, 而与初始N含量成负相关关系(表5 ), 表明凋落物初始养分含量, 尤其是N含量在很大程度上影响着凋落物的分解速率, 这与杨玉盛等(2003)和王希华等(2004)的研究结果一致.值得注意的是, 虽然黄果冷杉叶片凋落物初始N含量较高, 且具有较小的C:N值, 但其凋落物分解速率仍较低.因此, 除初始养分含量外的其他因子在凋落分解过程中起着重要的作用.Hättenschwiler 和Jørgensen (2010)对热带雨林叶片凋落物分解特征研究发现, 凋落物中碳的质量对分解速率的影响比其化学计量特征(C、N和P含量等)更大.此外, 王希华等(2004)和Santiago (2007)研究发现凋落物比叶面积与凋落物分解速率之间存在显著的正相关关系, 较大的比叶面积意味着单位质量凋落物具有更大的环境接触面积, 一方面加大了雨水的淋溶作用, 另一方面有利于微生物定居和土壤动物的碎裂作用, 从而大大提高凋落分解速率(郭忠玲等, 2006 ).也有研究表明, 叶片厚度与凋落物分解速率存在明显的负相关关系(Cornwell et al ., 2008 ).因此, 与叶片经济型谱存在密切联系的叶片功能特征(如比叶面积、叶片氮素含量和叶片厚度等)在叶片凋落物分解过程中起着重要作用(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 ).综上所述, 叶片凋落物分解是一个受其自身性质和外界环境因素共同作用的复杂过程, 它对陆地生态系统的养分循环有着深远影响(Kay et al ., 2008 ; Makkonen et al ., 2012 ). ...
... ; Makkonen et al ., 2012 ). ...
Temperature affects leaf litter decomposition in low-order forest streams: Field and microcosm approaches
1
2014
... 在水生生态系统中, 叶片凋落物作为主要的外源物质为其提供了大量的营养物质(Leroy & Marks, 2006 ), 且叶片的种类将影响着水体微生物及底栖无脊椎动物等的种群结构和数量(Graca, 2001 ; Hieber & Gessner, 2002 ).叶片凋落物在水体中的分解主要受其自身的品质和水体特征的影响.比如, 叶片大小、吸水性和可溶性物质含量将影响分解初期的淋溶速率(Ibrahima et al ., 2008 ); 叶片凋落物初始养分含量将影响其分解速率和养分的释放模式(Pascoal et al ., 2001 ; 刘昕等, 2008 ); 水温和流速也在很大程度上影响着凋落物的分解(Leroy & Marks, 2006 ; Martínez et al ., 2014 ).此外, 水生生态系统周围的植被构成及其季节变化将直接影响进入水体的叶片凋落物类型, 从而影响水体中生物种群变化及物质和能量循环(迟国梁和童晓立, 2010 ).与陆地生态系统相比, 由于水体的浸泡和流动冲击作用, 凋落物在水生生态系统中具有较快的分解速率(Petersen & Cummins, 1974 ; 张川等, 2015 ), 同时, 微生物和食碎屑动物的种类和数量在两种生态系统中的差异也在很大程度上影响着凋落物的分解过程(Leroy & Marks, 2006 ; Tiegs et al ., 2013 ; Handa et al ., 2014 ).目前, 关于叶片凋落物在水体中的分解速率及养分释放特征研究还相对较少.因此, 加强水体叶片凋落物分解特征研究, 尤其针对重要的水生生态系统开展相关研究具有重要意义. ...
Patterns of carbon, nitrogen and phosphorus dynamics in decomposing foliar litter in Canadian forests
1
2006
... 凋落物自身往往缺乏大量元素, 如N、P和S等(Berg, 2000 ), 从而难以满足微生物的分解作用, 故需其在外界环境中固定相应的养分, 并达到某一阈值时才分解释放凋落物中的养分(Moore et al ., 2006 ; Parton et al ., 2007 ; Aponte et al ., 2012 ).Parnas (1975)认为凋落物C:N值是发生N固持和矿化的关键值, 当C:N值>30时发生N的固持, 而当C:N值<30时发生N的矿化.本文中高山柳凋落物初始C:N值(30.26)十分接近此值, 故在分解初期便出现N的释放, 而其他树种C:N值均大于30, 故先经历富集过程才进行N的释放.此外, 4个树种凋落物P在分解前期均存在明显的释放过程, 这一阶段主要受可溶性P淋溶作用的影响(彭少麟和刘强, 2002 ), 而在春季和夏季发生P的富集则与微生物活动加强有关(王希华等, 2004 ). ...
Litter decomposition rates in Canadian forests
1
1999
... 本文通过研究九寨沟国家自然保护区内4种常见林型代表性树种, 即寒温性针叶林(黄果冷杉)、温性针叶林(油松)、落叶阔叶林(红桦)和落叶阔叶灌丛(高山柳)的叶片凋落物分解特征, 得出以下结果: 各树种间凋落物分解系数k 高山柳>红桦>黄果冷杉>油松, 且各物种间凋落物MR 差异极显著(p < 0.01, 图2 ).结果表明, 叶片凋落物分解速率存在明显的物种差异, 且阔叶树种叶片分解速率显著大于针叶树种, 这与郭忠玲等(2006)对长白山各植被带主要树种凋落物分解速率的研究结果一致.此外, 各树种叶片凋落物在前3个月质量损失最快, 而后迅速减慢, 第6-12月又逐渐增大(图2 ).这与Zhou等(2008)和Aponte等(2012)的研究结果一致, 凋落物分解初期可溶性物质会迅速通过淋溶作用进入到生态系统中, 而后进入较长且缓慢的生物分解过程(Petersen & Cummins, 1974 ).在本文中, 凋落物质量损失变化与外界环境温度的变化趋势一致, 且在降水丰沛的季节(5-9月)较高.这一结果很好地印证了温度和降水量作为主要的环境影响因子控制着凋落物的分解速率(Moore et al ., 1999 ; Hobbie et al ., 2006 ; Parton et al ., 2007 ). ...
Energy storage and the balance of producers and decomposers in ecological systems
1
1963
... 本文采用Olson (1963) 提出的负指数衰减模型进行叶片凋落物分解模式拟合, 即y = a e- kt y 为凋落物的剩余率(%), a 为拟合参数, k 为凋落物的分解系数, t 为分解时间(a). ...
Plant species effect on the decomposition and chemical changes of leaf litter in grassland and pine and oak forest soils
1
2014
... 叶片凋落物分解是陆地生态系统中一个重要的生态过程, 其分解速率对养分循环和生产力有着至关重要的作用(Parton et al ., 2007 ; Zhou et al ., 2008 ; Purahong et al ., 2014 ).凋落物分解将光合产物转变为无机养分和土壤有机质(Parton et al ., 2007 ), 提高了土壤肥力, 并提供植物生长所需的养分(王相娥等, 2009 ; Shi et al ., 2015 ).凋落物的分解是一个复杂的过程, 包括短暂的淋溶和长时间的生物分解阶段(Petersen & Cummins, 1974 ).通常情况下, 凋落物分解速率受凋落物质量、环境因子(如温度和湿度)和生物活动(如真菌和细菌)等多方面因素的综合影响(Couteaux et al ., 1995 ; Aerts, 1997 ; Wardle et al ., 2003 ; Ayres et al ., 2006 ; Zhou et al ., 2008 ; 武启骞等, 2013 ; 邓长春等, 2015 ).有研究表明, 凋落物的质量, 如初始N含量、木质素含量和C:N值等不但影响凋落物的分解速率(Aponte et al ., 2012 ; 邓长春等, 2015 ), 而且在很大程度上决定了其养分的固定和矿化(Parton et al ., 2007 ; Osono et al ., 2014 ).此外, 与叶片经济型谱密切联系的叶片功能性状(如叶片厚度与比叶面积)对其凋落物的分解过程也有着重要影响(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 ). ...
Model for decomposition of organic material by microorganisms
1975
Global-scale similarities in nitrogen release patterns during long-term decomposition
5
2007
... 叶片凋落物分解是陆地生态系统中一个重要的生态过程, 其分解速率对养分循环和生产力有着至关重要的作用(Parton et al ., 2007 ; Zhou et al ., 2008 ; Purahong et al ., 2014 ).凋落物分解将光合产物转变为无机养分和土壤有机质(Parton et al ., 2007 ), 提高了土壤肥力, 并提供植物生长所需的养分(王相娥等, 2009 ; Shi et al ., 2015 ).凋落物的分解是一个复杂的过程, 包括短暂的淋溶和长时间的生物分解阶段(Petersen & Cummins, 1974 ).通常情况下, 凋落物分解速率受凋落物质量、环境因子(如温度和湿度)和生物活动(如真菌和细菌)等多方面因素的综合影响(Couteaux et al ., 1995 ; Aerts, 1997 ; Wardle et al ., 2003 ; Ayres et al ., 2006 ; Zhou et al ., 2008 ; 武启骞等, 2013 ; 邓长春等, 2015 ).有研究表明, 凋落物的质量, 如初始N含量、木质素含量和C:N值等不但影响凋落物的分解速率(Aponte et al ., 2012 ; 邓长春等, 2015 ), 而且在很大程度上决定了其养分的固定和矿化(Parton et al ., 2007 ; Osono et al ., 2014 ).此外, 与叶片经济型谱密切联系的叶片功能性状(如叶片厚度与比叶面积)对其凋落物的分解过程也有着重要影响(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 ). ...
... ).凋落物分解将光合产物转变为无机养分和土壤有机质(Parton et al ., 2007 ), 提高了土壤肥力, 并提供植物生长所需的养分(王相娥等, 2009 ; Shi et al ., 2015 ).凋落物的分解是一个复杂的过程, 包括短暂的淋溶和长时间的生物分解阶段(Petersen & Cummins, 1974 ).通常情况下, 凋落物分解速率受凋落物质量、环境因子(如温度和湿度)和生物活动(如真菌和细菌)等多方面因素的综合影响(Couteaux et al ., 1995 ; Aerts, 1997 ; Wardle et al ., 2003 ; Ayres et al ., 2006 ; Zhou et al ., 2008 ; 武启骞等, 2013 ; 邓长春等, 2015 ).有研究表明, 凋落物的质量, 如初始N含量、木质素含量和C:N值等不但影响凋落物的分解速率(Aponte et al ., 2012 ; 邓长春等, 2015 ), 而且在很大程度上决定了其养分的固定和矿化(Parton et al ., 2007 ; Osono et al ., 2014 ).此外, 与叶片经济型谱密切联系的叶片功能性状(如叶片厚度与比叶面积)对其凋落物的分解过程也有着重要影响(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 ). ...
... ), 而且在很大程度上决定了其养分的固定和矿化(Parton et al ., 2007 ; Osono et al ., 2014 ).此外, 与叶片经济型谱密切联系的叶片功能性状(如叶片厚度与比叶面积)对其凋落物的分解过程也有着重要影响(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 ). ...
... 本文通过研究九寨沟国家自然保护区内4种常见林型代表性树种, 即寒温性针叶林(黄果冷杉)、温性针叶林(油松)、落叶阔叶林(红桦)和落叶阔叶灌丛(高山柳)的叶片凋落物分解特征, 得出以下结果: 各树种间凋落物分解系数k 高山柳>红桦>黄果冷杉>油松, 且各物种间凋落物MR 差异极显著(p < 0.01, 图2 ).结果表明, 叶片凋落物分解速率存在明显的物种差异, 且阔叶树种叶片分解速率显著大于针叶树种, 这与郭忠玲等(2006)对长白山各植被带主要树种凋落物分解速率的研究结果一致.此外, 各树种叶片凋落物在前3个月质量损失最快, 而后迅速减慢, 第6-12月又逐渐增大(图2 ).这与Zhou等(2008)和Aponte等(2012)的研究结果一致, 凋落物分解初期可溶性物质会迅速通过淋溶作用进入到生态系统中, 而后进入较长且缓慢的生物分解过程(Petersen & Cummins, 1974 ).在本文中, 凋落物质量损失变化与外界环境温度的变化趋势一致, 且在降水丰沛的季节(5-9月)较高.这一结果很好地印证了温度和降水量作为主要的环境影响因子控制着凋落物的分解速率(Moore et al ., 1999 ; Hobbie et al ., 2006 ; Parton et al ., 2007 ). ...
... 凋落物自身往往缺乏大量元素, 如N、P和S等(Berg, 2000 ), 从而难以满足微生物的分解作用, 故需其在外界环境中固定相应的养分, 并达到某一阈值时才分解释放凋落物中的养分(Moore et al ., 2006 ; Parton et al ., 2007 ; Aponte et al ., 2012 ).Parnas (1975)认为凋落物C:N值是发生N固持和矿化的关键值, 当C:N值>30时发生N的固持, 而当C:N值<30时发生N的矿化.本文中高山柳凋落物初始C:N值(30.26)十分接近此值, 故在分解初期便出现N的释放, 而其他树种C:N值均大于30, 故先经历富集过程才进行N的释放.此外, 4个树种凋落物P在分解前期均存在明显的释放过程, 这一阶段主要受可溶性P淋溶作用的影响(彭少麟和刘强, 2002 ), 而在春季和夏季发生P的富集则与微生物活动加强有关(王希华等, 2004 ). ...
Leaf breakdown rates: A measure of water quality?
1
2001
... 在水生生态系统中, 叶片凋落物作为主要的外源物质为其提供了大量的营养物质(Leroy & Marks, 2006 ), 且叶片的种类将影响着水体微生物及底栖无脊椎动物等的种群结构和数量(Graca, 2001 ; Hieber & Gessner, 2002 ).叶片凋落物在水体中的分解主要受其自身的品质和水体特征的影响.比如, 叶片大小、吸水性和可溶性物质含量将影响分解初期的淋溶速率(Ibrahima et al ., 2008 ); 叶片凋落物初始养分含量将影响其分解速率和养分的释放模式(Pascoal et al ., 2001 ; 刘昕等, 2008 ); 水温和流速也在很大程度上影响着凋落物的分解(Leroy & Marks, 2006 ; Martínez et al ., 2014 ).此外, 水生生态系统周围的植被构成及其季节变化将直接影响进入水体的叶片凋落物类型, 从而影响水体中生物种群变化及物质和能量循环(迟国梁和童晓立, 2010 ).与陆地生态系统相比, 由于水体的浸泡和流动冲击作用, 凋落物在水生生态系统中具有较快的分解速率(Petersen & Cummins, 1974 ; 张川等, 2015 ), 同时, 微生物和食碎屑动物的种类和数量在两种生态系统中的差异也在很大程度上影响着凋落物的分解过程(Leroy & Marks, 2006 ; Tiegs et al ., 2013 ; Handa et al ., 2014 ).目前, 关于叶片凋落物在水体中的分解速率及养分释放特征研究还相对较少.因此, 加强水体叶片凋落物分解特征研究, 尤其针对重要的水生生态系统开展相关研究具有重要意义. ...
森林凋落物动态及其对全球变暖的响应
1
2002
... 凋落物自身往往缺乏大量元素, 如N、P和S等(Berg, 2000 ), 从而难以满足微生物的分解作用, 故需其在外界环境中固定相应的养分, 并达到某一阈值时才分解释放凋落物中的养分(Moore et al ., 2006 ; Parton et al ., 2007 ; Aponte et al ., 2012 ).Parnas (1975)认为凋落物C:N值是发生N固持和矿化的关键值, 当C:N值>30时发生N的固持, 而当C:N值<30时发生N的矿化.本文中高山柳凋落物初始C:N值(30.26)十分接近此值, 故在分解初期便出现N的释放, 而其他树种C:N值均大于30, 故先经历富集过程才进行N的释放.此外, 4个树种凋落物P在分解前期均存在明显的释放过程, 这一阶段主要受可溶性P淋溶作用的影响(彭少麟和刘强, 2002 ), 而在春季和夏季发生P的富集则与微生物活动加强有关(王希华等, 2004 ). ...
Leaf processing in a woodland stream
4
1974
... 叶片凋落物分解是陆地生态系统中一个重要的生态过程, 其分解速率对养分循环和生产力有着至关重要的作用(Parton et al ., 2007 ; Zhou et al ., 2008 ; Purahong et al ., 2014 ).凋落物分解将光合产物转变为无机养分和土壤有机质(Parton et al ., 2007 ), 提高了土壤肥力, 并提供植物生长所需的养分(王相娥等, 2009 ; Shi et al ., 2015 ).凋落物的分解是一个复杂的过程, 包括短暂的淋溶和长时间的生物分解阶段(Petersen & Cummins, 1974 ).通常情况下, 凋落物分解速率受凋落物质量、环境因子(如温度和湿度)和生物活动(如真菌和细菌)等多方面因素的综合影响(Couteaux et al ., 1995 ; Aerts, 1997 ; Wardle et al ., 2003 ; Ayres et al ., 2006 ; Zhou et al ., 2008 ; 武启骞等, 2013 ; 邓长春等, 2015 ).有研究表明, 凋落物的质量, 如初始N含量、木质素含量和C:N值等不但影响凋落物的分解速率(Aponte et al ., 2012 ; 邓长春等, 2015 ), 而且在很大程度上决定了其养分的固定和矿化(Parton et al ., 2007 ; Osono et al ., 2014 ).此外, 与叶片经济型谱密切联系的叶片功能性状(如叶片厚度与比叶面积)对其凋落物的分解过程也有着重要影响(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 ). ...
... 在水生生态系统中, 叶片凋落物作为主要的外源物质为其提供了大量的营养物质(Leroy & Marks, 2006 ), 且叶片的种类将影响着水体微生物及底栖无脊椎动物等的种群结构和数量(Graca, 2001 ; Hieber & Gessner, 2002 ).叶片凋落物在水体中的分解主要受其自身的品质和水体特征的影响.比如, 叶片大小、吸水性和可溶性物质含量将影响分解初期的淋溶速率(Ibrahima et al ., 2008 ); 叶片凋落物初始养分含量将影响其分解速率和养分的释放模式(Pascoal et al ., 2001 ; 刘昕等, 2008 ); 水温和流速也在很大程度上影响着凋落物的分解(Leroy & Marks, 2006 ; Martínez et al ., 2014 ).此外, 水生生态系统周围的植被构成及其季节变化将直接影响进入水体的叶片凋落物类型, 从而影响水体中生物种群变化及物质和能量循环(迟国梁和童晓立, 2010 ).与陆地生态系统相比, 由于水体的浸泡和流动冲击作用, 凋落物在水生生态系统中具有较快的分解速率(Petersen & Cummins, 1974 ; 张川等, 2015 ), 同时, 微生物和食碎屑动物的种类和数量在两种生态系统中的差异也在很大程度上影响着凋落物的分解过程(Leroy & Marks, 2006 ; Tiegs et al ., 2013 ; Handa et al ., 2014 ).目前, 关于叶片凋落物在水体中的分解速率及养分释放特征研究还相对较少.因此, 加强水体叶片凋落物分解特征研究, 尤其针对重要的水生生态系统开展相关研究具有重要意义. ...
... 本文通过研究九寨沟国家自然保护区内4种常见林型代表性树种, 即寒温性针叶林(黄果冷杉)、温性针叶林(油松)、落叶阔叶林(红桦)和落叶阔叶灌丛(高山柳)的叶片凋落物分解特征, 得出以下结果: 各树种间凋落物分解系数k 高山柳>红桦>黄果冷杉>油松, 且各物种间凋落物MR 差异极显著(p < 0.01, 图2 ).结果表明, 叶片凋落物分解速率存在明显的物种差异, 且阔叶树种叶片分解速率显著大于针叶树种, 这与郭忠玲等(2006)对长白山各植被带主要树种凋落物分解速率的研究结果一致.此外, 各树种叶片凋落物在前3个月质量损失最快, 而后迅速减慢, 第6-12月又逐渐增大(图2 ).这与Zhou等(2008)和Aponte等(2012)的研究结果一致, 凋落物分解初期可溶性物质会迅速通过淋溶作用进入到生态系统中, 而后进入较长且缓慢的生物分解过程(Petersen & Cummins, 1974 ).在本文中, 凋落物质量损失变化与外界环境温度的变化趋势一致, 且在降水丰沛的季节(5-9月)较高.这一结果很好地印证了温度和降水量作为主要的环境影响因子控制着凋落物的分解速率(Moore et al ., 1999 ; Hobbie et al ., 2006 ; Parton et al ., 2007 ). ...
... 4个树种叶片凋落物分解速率在高山湖泊中的分解速率均极显著高于其在林下的分解速率(p < 0.01, 表3 ; 图2 ), 且各物种凋落物在林下完全分解(99%)所需的时间是其在水体中的1.5倍左右, 这与陈书秀和江明喜(2006)对三峡地区香溪河流域不同树种叶片凋落物分解的研究结果一致.湖泊中凋落物分解速率较快的一个重要原因是凋落物长时间浸泡在水体中, 较强的淋溶作用会加速可溶性物质在分解初期的快速分解, 加之水体的流动将进一步加快这一过程的进行(Petersen & Cummins, 1974 ; 张川等, 2015 ).其次, 水体微生物和无脊椎动物在凋落物分解过程中也起着至关重要的作用(Leroy & Marks, 2006 ; Tiegs et al ., 2013 ), 相对稳定的水体温度(冬季湖泊中平均水温仍保持在2 ℃以上, 图1 )为凋落物分解过程中的生物活动提供了良好的环境保障.本文结果显示, 冬季水体凋落物的失重率保持在一个较高水平.此外, 不同树种间凋落物在水体中的分解速率差异与其在林下分解的规律一致, 即阔叶树种叶片分解速率明显大于针叶树, 一个重要的原因是针叶树种叶片较小且表面革质化大大降低了分解初期的淋溶速率(Ibrahima et al ., 2008 ; 迟国梁和童晓立, 2010 ). ...
Influence of different forest system management practices on leaf litter decomposition rates, nutrient dynamics and the activity of ligninolytic enzymes: A case study from central European forests
1
2014
... 叶片凋落物分解是陆地生态系统中一个重要的生态过程, 其分解速率对养分循环和生产力有着至关重要的作用(Parton et al ., 2007 ; Zhou et al ., 2008 ; Purahong et al ., 2014 ).凋落物分解将光合产物转变为无机养分和土壤有机质(Parton et al ., 2007 ), 提高了土壤肥力, 并提供植物生长所需的养分(王相娥等, 2009 ; Shi et al ., 2015 ).凋落物的分解是一个复杂的过程, 包括短暂的淋溶和长时间的生物分解阶段(Petersen & Cummins, 1974 ).通常情况下, 凋落物分解速率受凋落物质量、环境因子(如温度和湿度)和生物活动(如真菌和细菌)等多方面因素的综合影响(Couteaux et al ., 1995 ; Aerts, 1997 ; Wardle et al ., 2003 ; Ayres et al ., 2006 ; Zhou et al ., 2008 ; 武启骞等, 2013 ; 邓长春等, 2015 ).有研究表明, 凋落物的质量, 如初始N含量、木质素含量和C:N值等不但影响凋落物的分解速率(Aponte et al ., 2012 ; 邓长春等, 2015 ), 而且在很大程度上决定了其养分的固定和矿化(Parton et al ., 2007 ; Osono et al ., 2014 ).此外, 与叶片经济型谱密切联系的叶片功能性状(如叶片厚度与比叶面积)对其凋落物的分解过程也有着重要影响(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 ). ...
Extending the leaf economics spectrum to decomposition: Evidence from a tropical forest
2
2007
... 叶片凋落物分解是陆地生态系统中一个重要的生态过程, 其分解速率对养分循环和生产力有着至关重要的作用(Parton et al ., 2007 ; Zhou et al ., 2008 ; Purahong et al ., 2014 ).凋落物分解将光合产物转变为无机养分和土壤有机质(Parton et al ., 2007 ), 提高了土壤肥力, 并提供植物生长所需的养分(王相娥等, 2009 ; Shi et al ., 2015 ).凋落物的分解是一个复杂的过程, 包括短暂的淋溶和长时间的生物分解阶段(Petersen & Cummins, 1974 ).通常情况下, 凋落物分解速率受凋落物质量、环境因子(如温度和湿度)和生物活动(如真菌和细菌)等多方面因素的综合影响(Couteaux et al ., 1995 ; Aerts, 1997 ; Wardle et al ., 2003 ; Ayres et al ., 2006 ; Zhou et al ., 2008 ; 武启骞等, 2013 ; 邓长春等, 2015 ).有研究表明, 凋落物的质量, 如初始N含量、木质素含量和C:N值等不但影响凋落物的分解速率(Aponte et al ., 2012 ; 邓长春等, 2015 ), 而且在很大程度上决定了其养分的固定和矿化(Parton et al ., 2007 ; Osono et al ., 2014 ).此外, 与叶片经济型谱密切联系的叶片功能性状(如叶片厚度与比叶面积)对其凋落物的分解过程也有着重要影响(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 ). ...
... 本文结果显示, 叶片凋落物分解1年后MR 同其初始C:N值存在正相关关系, 而与初始N含量成负相关关系(表5 ), 表明凋落物初始养分含量, 尤其是N含量在很大程度上影响着凋落物的分解速率, 这与杨玉盛等(2003)和王希华等(2004)的研究结果一致.值得注意的是, 虽然黄果冷杉叶片凋落物初始N含量较高, 且具有较小的C:N值, 但其凋落物分解速率仍较低.因此, 除初始养分含量外的其他因子在凋落分解过程中起着重要的作用.Hättenschwiler 和Jørgensen (2010)对热带雨林叶片凋落物分解特征研究发现, 凋落物中碳的质量对分解速率的影响比其化学计量特征(C、N和P含量等)更大.此外, 王希华等(2004)和Santiago (2007)研究发现凋落物比叶面积与凋落物分解速率之间存在显著的正相关关系, 较大的比叶面积意味着单位质量凋落物具有更大的环境接触面积, 一方面加大了雨水的淋溶作用, 另一方面有利于微生物定居和土壤动物的碎裂作用, 从而大大提高凋落分解速率(郭忠玲等, 2006 ).也有研究表明, 叶片厚度与凋落物分解速率存在明显的负相关关系(Cornwell et al ., 2008 ).因此, 与叶片经济型谱存在密切联系的叶片功能特征(如比叶面积、叶片氮素含量和叶片厚度等)在叶片凋落物分解过程中起着重要作用(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 ).综上所述, 叶片凋落物分解是一个受其自身性质和外界环境因素共同作用的复杂过程, 它对陆地生态系统的养分循环有着深远影响(Kay et al ., 2008 ; Makkonen et al ., 2012 ). ...
1
2008
... 本文采用三因素方差分析探讨了物种、分解时间和环境类型对叶片凋落物MR、NR 和PR 的影响, 并分析各因素两两间的交互作用.然后, 采用单因素方差分析法分析凋落物MR、NR 和PR 在不同物种间的差异, 并采用Duncan法进行多重比较.采用Pearson相关分析法分析了叶片凋落物分解1年后MR、NR 和PR 与凋落物初始养分含量之间的相关关系.以上统计分析使用SAS 9.2统计软件进行处理,多重比较显著性水平α = 0.01 (SAS Institute, 2008 ). ...
Mixed leaf litter decomposition and N, P release with a focus on Phy- llostachys edulis (Carriere) J. Houz. forest in subtropical southeastern China
1
2015
... 叶片凋落物分解是陆地生态系统中一个重要的生态过程, 其分解速率对养分循环和生产力有着至关重要的作用(Parton et al ., 2007 ; Zhou et al ., 2008 ; Purahong et al ., 2014 ).凋落物分解将光合产物转变为无机养分和土壤有机质(Parton et al ., 2007 ), 提高了土壤肥力, 并提供植物生长所需的养分(王相娥等, 2009 ; Shi et al ., 2015 ).凋落物的分解是一个复杂的过程, 包括短暂的淋溶和长时间的生物分解阶段(Petersen & Cummins, 1974 ).通常情况下, 凋落物分解速率受凋落物质量、环境因子(如温度和湿度)和生物活动(如真菌和细菌)等多方面因素的综合影响(Couteaux et al ., 1995 ; Aerts, 1997 ; Wardle et al ., 2003 ; Ayres et al ., 2006 ; Zhou et al ., 2008 ; 武启骞等, 2013 ; 邓长春等, 2015 ).有研究表明, 凋落物的质量, 如初始N含量、木质素含量和C:N值等不但影响凋落物的分解速率(Aponte et al ., 2012 ; 邓长春等, 2015 ), 而且在很大程度上决定了其养分的固定和矿化(Parton et al ., 2007 ; Osono et al ., 2014 ).此外, 与叶片经济型谱密切联系的叶片功能性状(如叶片厚度与比叶面积)对其凋落物的分解过程也有着重要影响(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 ). ...
Litter decomposition, and associated invertebrate communities, in wetland ponds of the Copper River Delta, Alaska (USA)
2
2013
... 在水生生态系统中, 叶片凋落物作为主要的外源物质为其提供了大量的营养物质(Leroy & Marks, 2006 ), 且叶片的种类将影响着水体微生物及底栖无脊椎动物等的种群结构和数量(Graca, 2001 ; Hieber & Gessner, 2002 ).叶片凋落物在水体中的分解主要受其自身的品质和水体特征的影响.比如, 叶片大小、吸水性和可溶性物质含量将影响分解初期的淋溶速率(Ibrahima et al ., 2008 ); 叶片凋落物初始养分含量将影响其分解速率和养分的释放模式(Pascoal et al ., 2001 ; 刘昕等, 2008 ); 水温和流速也在很大程度上影响着凋落物的分解(Leroy & Marks, 2006 ; Martínez et al ., 2014 ).此外, 水生生态系统周围的植被构成及其季节变化将直接影响进入水体的叶片凋落物类型, 从而影响水体中生物种群变化及物质和能量循环(迟国梁和童晓立, 2010 ).与陆地生态系统相比, 由于水体的浸泡和流动冲击作用, 凋落物在水生生态系统中具有较快的分解速率(Petersen & Cummins, 1974 ; 张川等, 2015 ), 同时, 微生物和食碎屑动物的种类和数量在两种生态系统中的差异也在很大程度上影响着凋落物的分解过程(Leroy & Marks, 2006 ; Tiegs et al ., 2013 ; Handa et al ., 2014 ).目前, 关于叶片凋落物在水体中的分解速率及养分释放特征研究还相对较少.因此, 加强水体叶片凋落物分解特征研究, 尤其针对重要的水生生态系统开展相关研究具有重要意义. ...
... 4个树种叶片凋落物分解速率在高山湖泊中的分解速率均极显著高于其在林下的分解速率(p < 0.01, 表3 ; 图2 ), 且各物种凋落物在林下完全分解(99%)所需的时间是其在水体中的1.5倍左右, 这与陈书秀和江明喜(2006)对三峡地区香溪河流域不同树种叶片凋落物分解的研究结果一致.湖泊中凋落物分解速率较快的一个重要原因是凋落物长时间浸泡在水体中, 较强的淋溶作用会加速可溶性物质在分解初期的快速分解, 加之水体的流动将进一步加快这一过程的进行(Petersen & Cummins, 1974 ; 张川等, 2015 ).其次, 水体微生物和无脊椎动物在凋落物分解过程中也起着至关重要的作用(Leroy & Marks, 2006 ; Tiegs et al ., 2013 ), 相对稳定的水体温度(冬季湖泊中平均水温仍保持在2 ℃以上, 图1 )为凋落物分解过程中的生物活动提供了良好的环境保障.本文结果显示, 冬季水体凋落物的失重率保持在一个较高水平.此外, 不同树种间凋落物在水体中的分解速率差异与其在林下分解的规律一致, 即阔叶树种叶片分解速率明显大于针叶树, 一个重要的原因是针叶树种叶片较小且表面革质化大大降低了分解初期的淋溶速率(Ibrahima et al ., 2008 ; 迟国梁和童晓立, 2010 ). ...
暖温带地区主要树种叶片凋落物分解过程中主要元素释放的比较
1
2001
... 在1年的分解期内, 4个树种叶片凋落物的NR 和PR 差异极显著(p < 0.01, 表3 ), 且NR 与初始N含量和C:N值分别存在极显著负相关和正相关关系(表5 ), 这表明凋落物初始养分含量在很大程度上影响着分解过程中的养分释放.在林下分解时, 高山柳叶片凋落物N并未经过富集过程便直接开始释放, 而红桦、黄果冷杉和油松要先经过富集过程才开始释放; 4个树种凋落物P在分解过程中均表现出释放—富集—释放模式.此外, 凋落物初始N和P含量低的物种(如油松)在分解过程中需富集更多的养分才开始出现N和P的释放, 这与王瑾和黄建辉(2001) 的研究结果一致, 而王希华等(2004) 认为营养元素的释放和富集与凋落叶初始营养状况无明显的相关性.因此, 凋落物N和P的富集和释放与初始养分条件的关系存在明显的物种差异. ...
凋落物分解研究综述
1
2009
... 叶片凋落物分解是陆地生态系统中一个重要的生态过程, 其分解速率对养分循环和生产力有着至关重要的作用(Parton et al ., 2007 ; Zhou et al ., 2008 ; Purahong et al ., 2014 ).凋落物分解将光合产物转变为无机养分和土壤有机质(Parton et al ., 2007 ), 提高了土壤肥力, 并提供植物生长所需的养分(王相娥等, 2009 ; Shi et al ., 2015 ).凋落物的分解是一个复杂的过程, 包括短暂的淋溶和长时间的生物分解阶段(Petersen & Cummins, 1974 ).通常情况下, 凋落物分解速率受凋落物质量、环境因子(如温度和湿度)和生物活动(如真菌和细菌)等多方面因素的综合影响(Couteaux et al ., 1995 ; Aerts, 1997 ; Wardle et al ., 2003 ; Ayres et al ., 2006 ; Zhou et al ., 2008 ; 武启骞等, 2013 ; 邓长春等, 2015 ).有研究表明, 凋落物的质量, 如初始N含量、木质素含量和C:N值等不但影响凋落物的分解速率(Aponte et al ., 2012 ; 邓长春等, 2015 ), 而且在很大程度上决定了其养分的固定和矿化(Parton et al ., 2007 ; Osono et al ., 2014 ).此外, 与叶片经济型谱密切联系的叶片功能性状(如叶片厚度与比叶面积)对其凋落物的分解过程也有着重要影响(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 ). ...
天童国家森林公园常见植物凋落叶分解的研究
2
2004
... 在1年的分解期内, 4个树种叶片凋落物的NR 和PR 差异极显著(p < 0.01, 表3 ), 且NR 与初始N含量和C:N值分别存在极显著负相关和正相关关系(表5 ), 这表明凋落物初始养分含量在很大程度上影响着分解过程中的养分释放.在林下分解时, 高山柳叶片凋落物N并未经过富集过程便直接开始释放, 而红桦、黄果冷杉和油松要先经过富集过程才开始释放; 4个树种凋落物P在分解过程中均表现出释放—富集—释放模式.此外, 凋落物初始N和P含量低的物种(如油松)在分解过程中需富集更多的养分才开始出现N和P的释放, 这与王瑾和黄建辉(2001) 的研究结果一致, 而王希华等(2004) 认为营养元素的释放和富集与凋落叶初始营养状况无明显的相关性.因此, 凋落物N和P的富集和释放与初始养分条件的关系存在明显的物种差异. ...
... 凋落物自身往往缺乏大量元素, 如N、P和S等(Berg, 2000 ), 从而难以满足微生物的分解作用, 故需其在外界环境中固定相应的养分, 并达到某一阈值时才分解释放凋落物中的养分(Moore et al ., 2006 ; Parton et al ., 2007 ; Aponte et al ., 2012 ).Parnas (1975)认为凋落物C:N值是发生N固持和矿化的关键值, 当C:N值>30时发生N的固持, 而当C:N值<30时发生N的矿化.本文中高山柳凋落物初始C:N值(30.26)十分接近此值, 故在分解初期便出现N的释放, 而其他树种C:N值均大于30, 故先经历富集过程才进行N的释放.此外, 4个树种凋落物P在分解前期均存在明显的释放过程, 这一阶段主要受可溶性P淋溶作用的影响(彭少麟和刘强, 2002 ), 而在春季和夏季发生P的富集则与微生物活动加强有关(王希华等, 2004 ). ...
Determinants of litter mixing effects in a Swedish boreal forest
1
2003
... 叶片凋落物分解是陆地生态系统中一个重要的生态过程, 其分解速率对养分循环和生产力有着至关重要的作用(Parton et al ., 2007 ; Zhou et al ., 2008 ; Purahong et al ., 2014 ).凋落物分解将光合产物转变为无机养分和土壤有机质(Parton et al ., 2007 ), 提高了土壤肥力, 并提供植物生长所需的养分(王相娥等, 2009 ; Shi et al ., 2015 ).凋落物的分解是一个复杂的过程, 包括短暂的淋溶和长时间的生物分解阶段(Petersen & Cummins, 1974 ).通常情况下, 凋落物分解速率受凋落物质量、环境因子(如温度和湿度)和生物活动(如真菌和细菌)等多方面因素的综合影响(Couteaux et al ., 1995 ; Aerts, 1997 ; Wardle et al ., 2003 ; Ayres et al ., 2006 ; Zhou et al ., 2008 ; 武启骞等, 2013 ; 邓长春等, 2015 ).有研究表明, 凋落物的质量, 如初始N含量、木质素含量和C:N值等不但影响凋落物的分解速率(Aponte et al ., 2012 ; 邓长春等, 2015 ), 而且在很大程度上决定了其养分的固定和矿化(Parton et al ., 2007 ; Osono et al ., 2014 ).此外, 与叶片经济型谱密切联系的叶片功能性状(如叶片厚度与比叶面积)对其凋落物的分解过程也有着重要影响(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 ). ...
季节性雪被对高山森林凋落物分解的影响
1
2013
... 叶片凋落物分解是陆地生态系统中一个重要的生态过程, 其分解速率对养分循环和生产力有着至关重要的作用(Parton et al ., 2007 ; Zhou et al ., 2008 ; Purahong et al ., 2014 ).凋落物分解将光合产物转变为无机养分和土壤有机质(Parton et al ., 2007 ), 提高了土壤肥力, 并提供植物生长所需的养分(王相娥等, 2009 ; Shi et al ., 2015 ).凋落物的分解是一个复杂的过程, 包括短暂的淋溶和长时间的生物分解阶段(Petersen & Cummins, 1974 ).通常情况下, 凋落物分解速率受凋落物质量、环境因子(如温度和湿度)和生物活动(如真菌和细菌)等多方面因素的综合影响(Couteaux et al ., 1995 ; Aerts, 1997 ; Wardle et al ., 2003 ; Ayres et al ., 2006 ; Zhou et al ., 2008 ; 武启骞等, 2013 ; 邓长春等, 2015 ).有研究表明, 凋落物的质量, 如初始N含量、木质素含量和C:N值等不但影响凋落物的分解速率(Aponte et al ., 2012 ; 邓长春等, 2015 ), 而且在很大程度上决定了其养分的固定和矿化(Parton et al ., 2007 ; Osono et al ., 2014 ).此外, 与叶片经济型谱密切联系的叶片功能性状(如叶片厚度与比叶面积)对其凋落物的分解过程也有着重要影响(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 ). ...
格氏栲天然林与人工林凋落物数量、养分归还及凋落叶分解
2003
高山森林溪流冬季不同时期凋落物分解中水溶性氮和磷的动态特征
2
2015
... 在水生生态系统中, 叶片凋落物作为主要的外源物质为其提供了大量的营养物质(Leroy & Marks, 2006 ), 且叶片的种类将影响着水体微生物及底栖无脊椎动物等的种群结构和数量(Graca, 2001 ; Hieber & Gessner, 2002 ).叶片凋落物在水体中的分解主要受其自身的品质和水体特征的影响.比如, 叶片大小、吸水性和可溶性物质含量将影响分解初期的淋溶速率(Ibrahima et al ., 2008 ); 叶片凋落物初始养分含量将影响其分解速率和养分的释放模式(Pascoal et al ., 2001 ; 刘昕等, 2008 ); 水温和流速也在很大程度上影响着凋落物的分解(Leroy & Marks, 2006 ; Martínez et al ., 2014 ).此外, 水生生态系统周围的植被构成及其季节变化将直接影响进入水体的叶片凋落物类型, 从而影响水体中生物种群变化及物质和能量循环(迟国梁和童晓立, 2010 ).与陆地生态系统相比, 由于水体的浸泡和流动冲击作用, 凋落物在水生生态系统中具有较快的分解速率(Petersen & Cummins, 1974 ; 张川等, 2015 ), 同时, 微生物和食碎屑动物的种类和数量在两种生态系统中的差异也在很大程度上影响着凋落物的分解过程(Leroy & Marks, 2006 ; Tiegs et al ., 2013 ; Handa et al ., 2014 ).目前, 关于叶片凋落物在水体中的分解速率及养分释放特征研究还相对较少.因此, 加强水体叶片凋落物分解特征研究, 尤其针对重要的水生生态系统开展相关研究具有重要意义. ...
... 4个树种叶片凋落物分解速率在高山湖泊中的分解速率均极显著高于其在林下的分解速率(p < 0.01, 表3 ; 图2 ), 且各物种凋落物在林下完全分解(99%)所需的时间是其在水体中的1.5倍左右, 这与陈书秀和江明喜(2006)对三峡地区香溪河流域不同树种叶片凋落物分解的研究结果一致.湖泊中凋落物分解速率较快的一个重要原因是凋落物长时间浸泡在水体中, 较强的淋溶作用会加速可溶性物质在分解初期的快速分解, 加之水体的流动将进一步加快这一过程的进行(Petersen & Cummins, 1974 ; 张川等, 2015 ).其次, 水体微生物和无脊椎动物在凋落物分解过程中也起着至关重要的作用(Leroy & Marks, 2006 ; Tiegs et al ., 2013 ), 相对稳定的水体温度(冬季湖泊中平均水温仍保持在2 ℃以上, 图1 )为凋落物分解过程中的生物活动提供了良好的环境保障.本文结果显示, 冬季水体凋落物的失重率保持在一个较高水平.此外, 不同树种间凋落物在水体中的分解速率差异与其在林下分解的规律一致, 即阔叶树种叶片分解速率明显大于针叶树, 一个重要的原因是针叶树种叶片较小且表面革质化大大降低了分解初期的淋溶速率(Ibrahima et al ., 2008 ; 迟国梁和童晓立, 2010 ). ...
Factors influencing leaf litter decomposition: An intersite decomposition experiment across China
4
2008
... 叶片凋落物分解是陆地生态系统中一个重要的生态过程, 其分解速率对养分循环和生产力有着至关重要的作用(Parton et al ., 2007 ; Zhou et al ., 2008 ; Purahong et al ., 2014 ).凋落物分解将光合产物转变为无机养分和土壤有机质(Parton et al ., 2007 ), 提高了土壤肥力, 并提供植物生长所需的养分(王相娥等, 2009 ; Shi et al ., 2015 ).凋落物的分解是一个复杂的过程, 包括短暂的淋溶和长时间的生物分解阶段(Petersen & Cummins, 1974 ).通常情况下, 凋落物分解速率受凋落物质量、环境因子(如温度和湿度)和生物活动(如真菌和细菌)等多方面因素的综合影响(Couteaux et al ., 1995 ; Aerts, 1997 ; Wardle et al ., 2003 ; Ayres et al ., 2006 ; Zhou et al ., 2008 ; 武启骞等, 2013 ; 邓长春等, 2015 ).有研究表明, 凋落物的质量, 如初始N含量、木质素含量和C:N值等不但影响凋落物的分解速率(Aponte et al ., 2012 ; 邓长春等, 2015 ), 而且在很大程度上决定了其养分的固定和矿化(Parton et al ., 2007 ; Osono et al ., 2014 ).此外, 与叶片经济型谱密切联系的叶片功能性状(如叶片厚度与比叶面积)对其凋落物的分解过程也有着重要影响(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 ). ...
... ; Zhou et al ., 2008 ; 武启骞等, 2013 ; 邓长春等, 2015 ).有研究表明, 凋落物的质量, 如初始N含量、木质素含量和C:N值等不但影响凋落物的分解速率(Aponte et al ., 2012 ; 邓长春等, 2015 ), 而且在很大程度上决定了其养分的固定和矿化(Parton et al ., 2007 ; Osono et al ., 2014 ).此外, 与叶片经济型谱密切联系的叶片功能性状(如叶片厚度与比叶面积)对其凋落物的分解过程也有着重要影响(Santiago, 2007 ; Cornwell et al ., 2008 ; Makkonen et al ., 2012 ). ...
... 九寨沟作为我国重要的自然保护区, 具有丰富的森林植被类型和众多的高山湖泊, 是长江流域上游一个重要的水源涵养地(刘少英等, 2007 ).我们在九寨沟自然保护区内选择了4个典型树种的叶片凋落物进行陆地和水体凋落物分解特征研究, 旨在解决以下科学问题: 1)不同树种叶片凋落物在林下的分解速率和养分释放特征如何?2)不同树种叶片凋落物在高山湖泊中具有何种分解和养分释放特征?3)不同树种叶片凋落物在林下与高山湖泊中分解速率和养分释放特征存在何种差异?4)凋落物分解速率与各初始养分含量间是否存在相关关系?本文通过对以上科学问题的探讨, 以期为探究九寨沟国家自然保护区内森林和高山湖泊生态系统间物质循环提供一定的科学依据, 并为保护区内森林和湖泊的管理及更好地发挥其水源涵养地的功能提供参考. ...
... 九寨沟自然保护区位于青藏高原东缘的岷山山系南段(100.50°-104.45° E, 30.58°-34.32° N, 海拔1996-4764 m), 属川西高原气候区.该区域受高原季风和热带海洋季风影响, 年平均气温7.3 ℃, 年降水量622 mm, 降水集中在5-9月; 地势南高北低, 相对高差较大, 有高山山地、高山坡地和高山河谷地貌.九寨沟的水体景观具有极高的观赏价值, 有高山湖泊114个、瀑布群17个、滩流5处、泉水47眼、湍流11段.此外, 保护区内植被丰富, 包括寒温性针叶树如红杉(Larix potaninii )和黄果冷杉(Abies ernestii ), 温性针叶树如油松(Pinus tabulaeformis )和华山松(Pinus armandii ), 落叶阔叶树如红桦(Betula albo -sinensis )和辽东栎(Quercus wutaishanica ), 以及落叶阔叶灌丛如高山柳(又叫杯腺柳, Salix cupularis )和绣线菊(Spiraea salicifolia )等(刘少英等, 2007 ).样地概况详见表1 . ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}