沈佳, 李亚鹤, 张琳, 孙雪
宁波大学海洋学院, 浙江省海洋生物工程重点实验室, 浙江宁波 315211
SHEN Jia, LI Ya-He, ZHANG Lin, SUN Xue
通讯作者:
责任编辑:
收稿日期: 2015-07-8
接受日期: 2016-04-25
网络出版日期: 2016-09-10
版权声明: 2016 植物生态学报编辑部 本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。
基金资助:
展开
摘要
为了探讨光照强度和CO2浓度对蛋白核小球藻(Chlorella pyrenoidosa)生长、无机碳利用的复合效应, 丰富绿藻中无机碳浓缩机制的资料, 该文设置两种光照强度(40和120 µmol photons•m-2•s-1)和两种CO2浓度(0.04%和0.16%)组合成4种条件, 比较了蛋白核小球藻生长、无机碳浓度、pH补偿点、光合放氧速率、碳酸酐酶(CA)活性和α-CA基因转录表达对这4种培养条件的响应。结果发现: 蛋白核小球藻在高光强高CO2浓度组生长最快; 低光强高CO2浓度组培养体系中总无机碳浓度为1163.3 µmol•L-1, 显著高于其他3组; 高光强低CO2浓度组藻的pH补偿点最高(9.8), 而低光强高CO2浓度组藻的pH补偿点最低(8.6); 低光强高CO2浓度组藻的最大光合速率(Vmax)和最大光合速率一半时的无机碳浓度(K0.5)最高, 分别是其他3组的1.28-1.91倍和1.61-2.00倍; 高光强低CO2浓度组藻的胞外CA活性最高; 而低光强低CO2浓度组藻的胞外α-CA基因表达量显著高于其他3组。以上结果表明低CO2浓度可促进蛋白核小球藻的pH补偿点和无机碳亲和力的提高, 诱导胞外CA活性及α-CA基因的表达; 该藻主要以HCO3-为无机碳源, 其对无机碳的利用受光照的调节。
关键词:
Abstract
AimsCarbon concentrating mechanism (CCM) is one of the important contents in algal physiology and ecology. Numerous studies have been carried out in eukaryotic and prokaryotic algae, but the information on economic microalga Chlorella pyrenoidosa (Chlorophyta) is limited. Our purpose is to explore the composite effect of light and CO2 on growth, inorganic carbon utilization in C. pyrenoidosa, and enrich the data on CCM in green algae.
Methods Two light intensities (40 and 120 µmol photons•m-2•s-1) and two CO2 concentrations (0.04% and 0.16%) were combined into four treatments, and then the algal growth, inorganic carbon concentration, pH compensation point, photosynthetic oxygen evolution rate, carbonic anhydrase (CA) activity and α-CA gene expression were investigated.
Important findingsChlorella pyrenoidosa grew fastest under the high-light and high-CO2 condition. The total inorganic carbon concentration under low-light and high-CO2 group was 1163.3 µmol·L-1, which was significantly higher than that of other three groups. The alga had the maximal pH compensation point of 9.8 under the high-light and low-CO2 condition, and the minimal pH compensation point of 8.6 under the low-light and high-CO2 condition. The maximum photosynthetic rate (Vmax) and inorganic carbon concentration in half maximum photosynthetic rate (K0.5) in the low-light and high-CO2 group were the highest, which were 1.28-1.91 times and 1.61-2.00 times of that in other three groups, respectively. The highest activity of extracellular CA was detected in the high-light and lower-CO2 group. However, α-CA gene expression reached the maximum under the low-light and low-CO2 condition. The results indicated that the low CO2 level could increase the algal pH compensation point, photosynthetic inorganic carbon affinity, and induce the external CA activity and α-CA gene expression in C. pyrenoidosa. HCO3- was used as the primary inorganic carbon source, and the inorganic carbon utilization was also regulated by light in C. pyrenoidosa.
Keywords:
水生藻类获取CO2存在两个障碍, 一是光合固碳反应的关键酶——1,5-二磷酸核酮糖羧化酶/加氧酶(Rubisco)对CO2的亲和力很低; 二是CO2在水中的扩散速度极慢(Moroney & Ynalvez, 2007)。藻类为适应水中这种低CO2浓度环境并保持较高的光合作用速率, 会形成一种CO2浓缩机制(CO2 concentrating mechanism, CCM)。该CCM使藻类在细胞内部浓缩无机碳(Ci), 导致细胞对Ci亲和力增加和Rubisco附近Ci浓度升高, 以利于藻类光合作用的进行(Badger, 1980)。碳酸酐酶(CA)是CCM的重要组成部分, 负责催化CO2和HCO3-之间的相互转化。除了CA, 参与CCM的还有一系列转录调控因子, 如CCM1/CIA5和LCR1等(Miura et al., 2002; Yoshioka et al., 2004)。
多种环境因素如光照、温度、CO2浓度、pH值等都能对CCM产生影响。其中CO2浓度是影响藻类CCM及对Ci利用的重要因素。藻类在不同CO2浓度环境下培养会表现出不同的生理状态(Wang & Spalding, 2006)。低CO2浓度是CCM的诱导因素, 如0.04% CO2能激活莱茵衣藻(Chlamydomonas reinh- ardtii)胞外CA编码基因Cah1的表达, 而5% CO2会抑制其表达(Kucho et al., 2003)。低CO2浓度还可以激活莱茵衣藻中CCM转录调控因子CIA5的表达, CIA5又可以控制各CCM相关基因的表达(Wang et al., 2005)。
光照强度可以直接影响藻类对Ci的亲和力, 从而影响藻类的光合作用和生长。藻类细胞中Ci的转运和积累需要能量驱动, Ci转运所需ATP与光合作用中光系统I的电子流有关(夏建荣和高坤山, 2002)。光照强度与藻类对Ci的亲和力有密切关系, 在限制光照条件下杜氏藻(Dunaliella tertioleca)对Ci亲和力显著降低(Goyal et al., 1992)。光照强度也是参与调控藻类CCM的另一重要环境因素。高光强不仅能增加光合作用的速率, 而且能迅速降低细胞外环境中的Ci浓度, 从而增强CCM。
为了保证CO2高效地转化成碳水化合物, 光吸收和Ci的摄取之间需要很好地平衡。在低光照强度下, 光子通量密度的增加与光合固碳能力的增强相关(Elrad et al., 2002)。但过多的光能量会引起细胞内Ci的相对耗竭。相比之下, 暴露在限制Ci条件下, 又会引起过多光能的吸收(Yamano et al., 2008)。因此, 研究光照强度和CO2浓度的复合效应对于理解藻类对Ci的利用和CCM具有重要意义。
CCM是藻类生理和生态学研究的重要内容之一。目前CCM已在原核蓝藻和真核绿藻的模式藻——莱茵衣藻中开展了大量研究, 但是具有重要营养和经济价值的另一种绿藻——蛋白核小球藻(Chlorella pyrenoidosa)中相关研究还比较少。小球藻属中CCM研究主要集中在CO2浓度对Ci转运和光合参数的影响, 以及CA的鉴定等方面(Bozzo et al., 2000; Ochiai et al., 2007; Li et al., 2012), 而光照强度和CO2浓度对小球藻复合效应的报道极少。本文以蛋白核小球藻为实验材料, 研究了不同光照强度和CO2浓度对该藻的生长、pH补偿点、无机碳亲和力、碳酸酐酶活性等的影响, 以探讨蛋白核小球藻生长和CCM对光照强度和CO2浓度复合效应的响应, 了解蛋白核小球藻的CCM机制, 为提高蛋白核小球藻对无机碳的利用和提高其生物量等提供资料。
本实验所用藻种为‘蛋白核小球藻820’, 来自宁波大学海洋生物工程重点实验室藻种室。使用添加f/2的人工海水培养基(Guillard & Ryther, 1962), 盐度为1.5%, 培养温度为(25 ± 1) ℃, 光暗周期为12 h/12 h (light/dark)。
设置两种光照强度: 低光强(40 µmol photons•m-2•s-1)和高光强(120 µmol photons•m-2•s-1); 两种CO2浓度: 空气CO2浓度(0.04%, 低CO2浓度)和4倍空气CO2浓度(0.16%, 高CO2浓度)。CO2浓度控制通过CO2光照培养箱来完成, 在培养过程中CO2浓度变化小于0.005%。将藻细胞分别置于低光强低CO2浓度、低光强高CO2浓度、高光强低CO2浓度、高光强高CO2浓度4种条件(分别简记为LL、LH、HL和HH)下进行适应性培养4-5天(6代以上)后, 再转接到新鲜培养基中继续培养1-2天, 细胞密度为(6.0-8.0) × 106 cell•mL-1, 用于后续实验。
将对数期的蛋白核小球藻按照20%的比例转接到新鲜培养基中, 每天定时取样, 连续5天测定藻液在440 nm波长下的吸光度, 再根据本实验室做的藻细胞密度与吸光值的标准曲线公式来计算蛋白核小球藻的密度。每个处理条件设3个重复, 结果取其平均值。以培养时间为横坐标, 藻密度为纵坐标来绘制藻的生长曲线。然后通过公式µ = (lnx2 - lnx1)/(t2 - t1)计算比生长速率(µ), 其中x1、x2分别代表在t1、t2时间的藻细胞密度。
总碱度(TA)的测定方法采用酸式滴定法(王淑刚等, 2013)。参考Millero (2006)的计算方法, 在已测溶液总磷含量、总硅含量、pH值、盐度、TA和温度的基础上, 使用CO2SYS软件计算培养液中碳酸盐系统各参数: 溶解的无机碳(DIC)、CO2、HCO3-、CO32-浓度。
将装有100 mL藻液的三角瓶, 放置于4组不同光照和CO2条件下进行培养, 每隔6 h检测pH值变化, 直至pH值趋于稳定, 此时的pH值即为pH补偿点(Maberly, 1990)。
采用Clark型溶氧微电极(Unisense FertiliTech A/S, Aarhus, Denmark)测定光合放氧速率对无机碳浓度的响应曲线。在25 ℃和400 µmol photons•m-2•s-1的光照强度下, 测定重悬浮于无“CO2”的海水缓冲液(25 mmol·L-1 HEPES缓冲液, pH 8.0)中的藻细胞在不同HCO3-浓度下的光合作用速度。根据Michaelis-Menten方程: V = Vmax × [S]/(K0.5(DIC) + [S])来拟合求得K0.5(DIC)。式中, V为给定无机碳浓度下的净光合放氧速率; Vmax为饱和无机碳浓度下的净光合放氧速率; [S]为DIC浓度; K0.5(DIC)为达到最大光合速率(Vmax)一半时的DIC浓度。K0.5(CO2)和K0.5(HCO3-)根据K0.5(DIC)计算得出: K0.5(CO2) = K0.5(DIC)/44.2, K0.5(HCO3-) = K0.5(DIC) × 43.2/44.2 (高坤山, 1999)。
先8000 r•min-1离心10 min收集培养24 h的藻细胞, 再采用pH电极法测定CA活性, 在Wilbur和 Anderson (1948)方法的基础上稍加改进。胞外CA活性测定: 向重悬浮于巴比妥缓冲液(pH 8.4)的离心后藻细胞中加入0 ℃的饱和CO2蒸馏水, 用pH计监测反应体系pH值的变化, 记录pH值下降一个单位所需的时间, 以不加藻体的同样体系作空白对照。根据CA活性(U)计算公式: U = 10 × (T0/T - 1)来计算CA活性, 其中T0和T分别是反应体系中未加藻和加藻细胞时pH值下降一个单位所需的时间。总CA活性测定: 藻细胞在500 W功率超声波中进行破碎, 以破碎8 s, 间歇9.9 s为一个周期, 间歇超声破碎1.7 min后, 再按照以上胞外CA活性测定方法来进行。胞内CA活性=总CA活性-胞外CA活性。
将在空气CO2浓度下培养4-5天的蛋白核小球藻分别转移到4组不同光照强度和CO2浓度条件的培养箱中培养, 分别在0、2、6、12 h离心收集不同条件下的培养藻。用Trizol试剂(invitrogen)分别提取4组样品的总RNA, 再利用PrimeScript RT reagent Kit with gDNA Eraser逆转录试剂盒(TaKaRa)将RNA反转录成cDNA。以18S rDNA作为内参基因, α-CA基因为目的基因, 两者的荧光定量PCR引物序列参照王玮蔚等(2014)。按照SYBR Premix Ex Taq (TaKaRa)说明书进行荧光定量PCR反应, 反应体系中SYBR Premix Ex Taq 10 µL, 上下游引物各0.4 µL, cDNA模板2 µL, 加DEPC-水补足总体积到20 µL。荧光定量PCR反应条件为: 95 ℃预变性2 min后, 按照95 ℃ 15 s, 58 ℃ 15 s, 72 ℃ 20 s进行40次循环。荧光定量PCR数据采用2-△△Ct法进行分析(Livak & Schmittgen, 2001)。
采用Excel工作表进行数据处理和作图, 用SPSS 19.0软件进行方差分析和多重比较(LSD)统计分析, 其中p < 0.05表示差异显著; p < 0.01表示差异极显著。
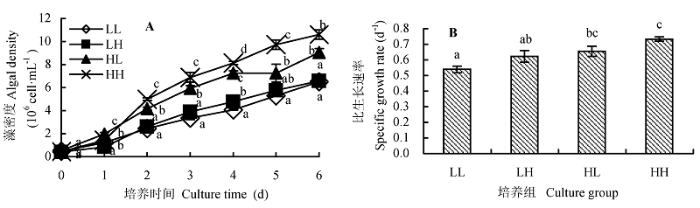
光照强度和CO2浓度对蛋白核小球藻生长影响显著(图1)。藻生长最快的是HH组, 其次是HL组, 而LH和LL组藻生长最慢(图1A)。可见高光强和高CO2浓度有利于该蛋白核小球藻的生长。
图1 不同光照强度和CO2浓度对蛋白核小球藻生长的影响(平均值±标准偏差)。A, 生长曲线。B, 比生长速率。LL、LH、HL和HH分别表示低光强低CO2浓度、低光强高CO2浓度、高光强低CO2浓度、高光强高CO2浓度条件。不同小写字母表示差异显著(p < 0.05)。
Fig. 1 Effect of different light intensity and CO2 concentration conditions on the growth of Chlorella pyrenoidosa (mean ± SD). A, Growth curve. B, Specific growth rate. LL, LH, HL and HH represent low-light intensity and low-CO2 concentration, low-light intensity and high-CO2 concentration, high-light intensity and low-CO2 concentration, high-light intensity and high-CO2 concentration conditions, respectively. Different lowercase letters indicate significant difference (p < 0.05).
在LL条件下, 蛋白核小球藻的比生长速率(µ)最低, 为0.54•d-1, 则而LH、HL、HH组的µ分别是LL组的1.15倍、1.22倍和1.36倍(图1B)。其中低光强的两组(LL和LH)藻之间, 以及高光强的两组(HL和HH)藻之间的µ差异均不显著(p > 0.05), 说明在相同光照条件下, CO2浓度对蛋白核小球藻生长的影响不大。而低CO2浓度两组(LL和HL)之间, 以及高CO2浓度两组(LH和HH)之间都是高光强培养藻的µ大于低光强的(p < 0.05), 说明在相同CO2条件下, 较高光强有利于藻的生长。以上结果表明相对于CO2 浓度, 光照强度是影响蛋白核小球藻生长的主要因素。
不同光照强度和CO2浓度影响了蛋白核小球藻培养液中碳酸盐系统各参数(表1)。对4组不同条件培养体系中不同形式无机碳含量进行比较, 发现在高CO2浓度条件下水中可利用的无机碳——CO2和HCO3-浓度较高, 而不能被吸收的CO32-则含量较少(p < 0.01)。LH组的总DIC浓度(1163.3µmol•L-1)、CO2浓度(4.37 µmol•L-1)和HCO3-浓度(1022.1 µmol•L-1)都显著高于其他3组(p < 0.01), 而pH值和CO32-浓度低于其他3组(p < 0.05), 总碱度仅比HH组高16.63%, 与LL和HL组差异不显著。在HH组培养藻中DIC浓度(803.4 µmol•L-1)分别比LL、HL两组高出4.98%、20.67%, 比LH组低30.94% (p < 0.05), HCO3-浓度(574.9 µmol•L-1)分别比LL、HL两组高49.87%、84.44% (p < 0.05), pH值和CO32-浓度则低于LL、HL两组(p < 0.05), 而CO2浓度与LL、HL两组无显著差异。在低CO2浓度的两组(LL和HL)藻的培养体系之间, 以上所有参数及总碱度均无显著差异。总之, 高CO2浓度的两组(LH和HH)藻中的各碳酸盐参数均差异显著(p < 0.05); 而低CO2浓度的两组(LL和HL)藻中的各参数差异均不显著(p > 0.05)。以上结果说明不同光照强度对低CO2浓度培养藻中碳酸盐系统各参数影响不大, 而对高CO2浓度培养藻有显著影响。
表1 不同光照强度和CO2浓度条件下蛋白核小球藻培养体系中各碳酸盐参数的比较(平均值±标准偏差, n = 3)
Table 1 Comparison of the parameters of carbonate system under different light intensity and CO2 concentration conditions in Chlorella pyrenoidosa (mean ± SD, n = 3)
| 处理组 Treatment group | 总碱度 Total alkalinity (µmol•L-1) | pH | DIC (µmol•L-1) | CO2 (µmol•L-1) | HCO3- (µmol•L-1) | CO32- (µmol•L-1) |
|---|---|---|---|---|---|---|
| LL | 1 387.8 ± 55.1b | 9.35 ± 0.00c | 765.3 ± 36.8ab | 0.22 ± 0.18a | 383.6 ± 18.44a | 381.5 ± 18.3c |
| LH | 1 388.4 ± 57.3b | 8.48 ± 0.03a | 1 163.3 ± 58.8c | 4.37 ± 0.46b | 1 022.1 ± 57.4c | 136.8 ± 3.6a |
| HL | 1 275.0 ± 16.5ab | 9.41 ± 0.04c | 665.8 ± 22.6a | 0.16 ± 0.44a | 311.7 ± 24.2a | 354.0 ± 5.8c |
| HH | 1 190.1 ± 0.0a | 8.95 ± 0.04b | 803.4 ± 17.2b | 0.84 ± 0.12a | 574.9 ± 27.2b | 227.6 ± 10.2b |
低光强培养的两组(LL/LH)藻中除了总碱度之外, 其余各碳酸盐参数均差异显著; 而高光强两组(HL和HH)除了总碱度和CO2浓度外, 其余参数也均差异显著。可见, 相比较于光照强度, CO2浓度是影响藻类碳酸盐系统各参数的主要因素。除了总碱度外, LH组的碳酸盐系统各参数与其他3组存在明显差别, 因为在较低光照强度下, 高浓度CO2大量溶解在培养液中, 从而使培养液中的CO2浓度升高, HCO3-浓度和总DIC浓度也相应升高, 而大量CO2的溶解导致pH值降低, 降低的pH值又影响了CO32-浓度, 使其降低。
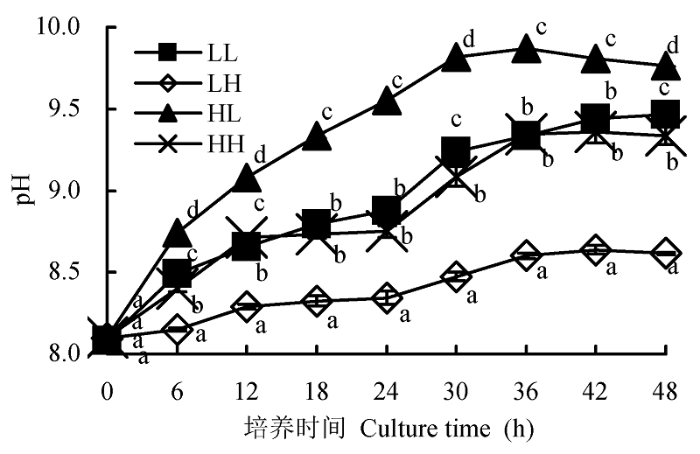
不同光照强度和CO2浓度条件下, 蛋白核小球培养体系中pH值随时间变化如图2所示。随着培养时间的延长, 培养体系中pH值逐渐升高, 直到达一个稳定值, 即pH补偿点。在4组不同条件中, HL组藻的pH补偿点最高, 为9.8; LL组、HH组和LH组的pH补偿点依次是9.5、9.3和8.6。低CO2浓度的两组(HL和LL)藻的pH补偿点高于高CO2浓度的两组(HH和LH), 高光强培养的两组藻的pH补偿点分别高于相应低光强的两组(HL > LL, HH > LH)。可见, 高CO2浓度可以降低蛋白核小球藻的pH补偿点, 而较高光照强度则可以升高其pH补偿点。
图2 不同光照强度和CO2浓度下蛋白核小球藻的pH补偿点(平均值±标准偏差)。LL、LH、HL和HH分别表示低光强低CO2浓度、低光强高CO2浓度、高光强低CO2浓度、高光强高CO2浓度条件。不同小写字母表示差异显著(p < 0.05)。
Fig. 2 Effect of different light intensity and CO2 concentration conditions on pH compensation point of Chlorella pyrenoidosa (mean ± SD). LL, LH, HL and HH represent low-light intensity and low-CO2 concentration, low-light intensity and high-CO2 concentration, high-light intensity and low-CO2 concentration, high-light intensity and high-CO2 concentration conditions, respectively. Different lowercase letters indicate significant difference (p < 0.05).
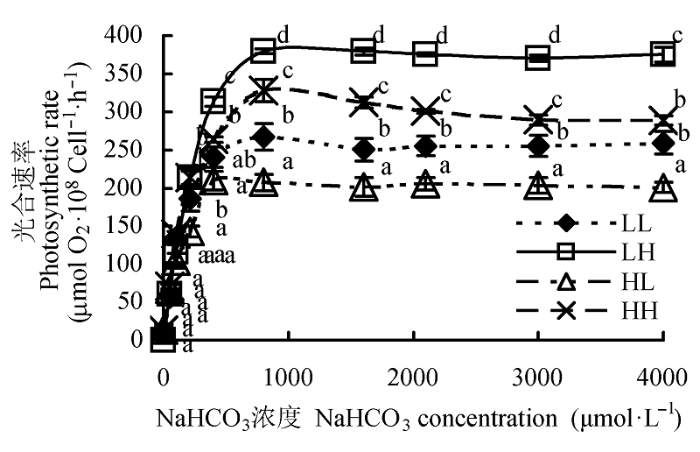
不同光照强度和CO2浓度培养条件下蛋白核小球藻对不同Ci浓度的响应不同。从光合放氧速率对DIC浓度的响应曲线(P-C)中可以发现, 随着培养介质中Ci浓度的升高, 各组藻的光合放氧速率均快速增加, 并迅速达到平衡(图3)。在4组条件下培养的藻, 其光合放氧速率存在差异, 表明不同培养条件下藻细胞对Ci亲和力不同。
图3 不同光照强度和CO2浓度下蛋白核小球藻的P-C曲线(平均值±标准偏差)。LL、LH、HL和HH分别表示低光强低CO2浓度、低光强高CO2浓度、高光强低CO2浓度、高光强高CO2浓度条件。不同小写字母表示差异显著(p < 0.05)。
Fig. 3 Effect of different light intensity and CO2 concentration conditions on P-C curve of Chlorella pyrenoidosa (mean ± SD). LL, LH, HL and HH represent low-light intensity and low-CO2 concentration, low-light intensity and high-CO2 concentration, high-light intensity and low-CO2 concentration, high-light intensity and high-CO2 concentration conditions, respectively. Different lowercase letters indicate significant difference (p < 0.05).
通过米氏方程进行非线性拟合计算后, 得到4组条件下藻的最大光合速率(Vmax)和最大光合速率一半时的Ci浓度(K0.5)。Vmax和K0.5是判断细胞对Ci亲和力的重要参数, Vmax和K0.5越小, 藻细胞对CO2的亲和力就越大(Badger, 1987; 高坤山, 1999)。
按照Vmax和K0.5的高低来排列, 4组不同培养条件的顺序都是LH > HH > LL > HL (表2)。在LH条件下, 藻细胞的Vmax是(415.44 ± 4.23) µmol O2•10-8 cell-1·h-1, 显著高于其他3组条件下藻的Vmax, 分别是HH组的1.28倍, LL的1.51倍和HL的1.91倍, 且各 组之间差异均显著(p < 0.05)。在LH条件下, 藻细胞的K0.5(DIC)也最高((188.33 ± 20.04) µmol•L-1), 分别是HH的1.61倍, LL的1.76倍和HL的2.00倍, 与其他3组差异显著; 但其他3组之间的K0.5 (DIC)差异均不显著。K0.5 (CO2)和K0.5 (HCO3-)的变化规律同K0.5 (DIC)一致。可见, 光照强度和CO2浓度都会影响藻类对Ci的亲和力, 其中光照强度越高, CO2浓度越低, 藻细胞对Ci的亲和力越高。
表2 不同光照强度和CO2浓度下蛋白核小球藻的Vmax和K0.5 (平均值±标准偏差, n = 3)
Table 2 Effect of different light intensity and CO2 concentration conditions on Vmax and K0.5 of Chlorella pyrenoidosa (mean ± SD, n = 3)
| 处理组 Treatmeat group | Vmax (µmol O2• 108cell·h-1) | K0.5 (µmol•L-1) | ||
|---|---|---|---|---|
| DIC | CO2 | HCO3- | ||
| LL | 275.93 ± 16.83b | 107.20 ± 2.49a | 2.43 ± 0.06a | 104.78 ± 2.43a |
| LH | 415.44 ± 4.23d | 188.33 ± 2.04b | 4.26 ± 0.45b | 184.07 ± 19.59b |
| HL | 218.03 ± 10.16a | 94.20 ± 7.03a | 2.13 ± 0.16a | 92.07 ± 6.87a |
| HH | 324.05 ± 1.34c | 116.83 ± 6.89a | 2.64 ± 0.16a | 114.19 ± 6.73a |
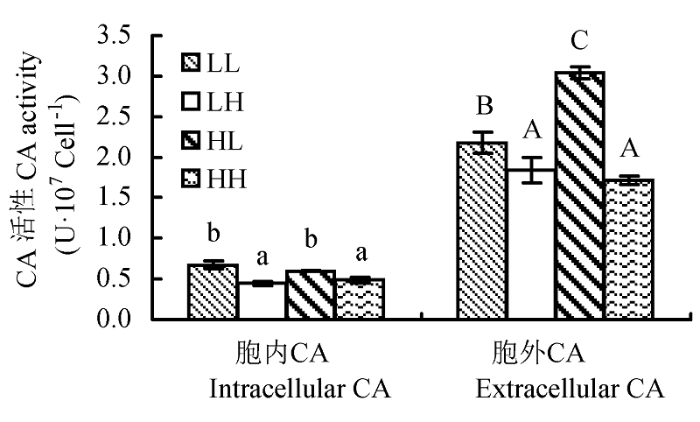
不同光照强度和CO2浓度对蛋白核小球藻CA活性的影响如图4所示。低CO2浓度的两组(LL和HL)藻的胞内CA活性显著高于高CO2浓度两组(LH和HH), 其中LL组藻的胞内CA活性是LH组的1.51倍, 而HL组的胞内CA活性是HH组的1.24倍, 均有显著差异(p < 0.05)。而LL和HL组、LH和HH组之间藻的胞内CA活性无显著差异。可见低CO2浓度诱导了胞内CA活性, 但光照强度对胞内CA活性则无影响。
图4 不同光照强度和CO2浓度下蛋白核小球藻的碳酸酐酶(CA)活性(平均值±标准偏差)。LL、LH、HL和HH分别表示低光强低CO2浓度、低光强高CO2浓度、高光强低CO2浓度、高光强高CO2浓度条件。不同字母表示差异显著(p < 0.05)。
Fig. 4 Effect of different light intensity and CO2 concentration conditions on carbonic anhydrase (CA) activity of Chlorella pyrenoidosa (mean ± SD). LL, LH, HL and HH represent low-light intensity and low-CO2 concentration, low-light intensity and high-CO2 concentration, high-light intensity and low-CO2 concentration, high-light intensity and high-CO2 concentration conditions, respectively. Different letters indicate significant difference (p < 0.05).
相对于胞内CA, 蛋白核小球藻的胞外CA更为活跃。该藻的胞外CA活性在HL培养条件下最高, 达到3.04 U•107 cell-1, 分别是LL、LH和HH的1.40倍、1.65倍和1.78倍, 与其他3组差异极显著(p < 0.01); 其次是LL组, 藻的胞外CA活性为2.18 U•107 cell-1。而高CO2浓度的两组(LH和HH)藻的胞外CA活性最低, 且两者之间差异不显著(p > 0.05)。可见低CO2浓度同样诱导了蛋白核小球藻胞外CA活性的升高, 而高光强则能促进低CO2浓度诱导的CA活性升高的幅度; 在高CO2浓度条件下, 不同光强对藻胞外CA活性无影响。
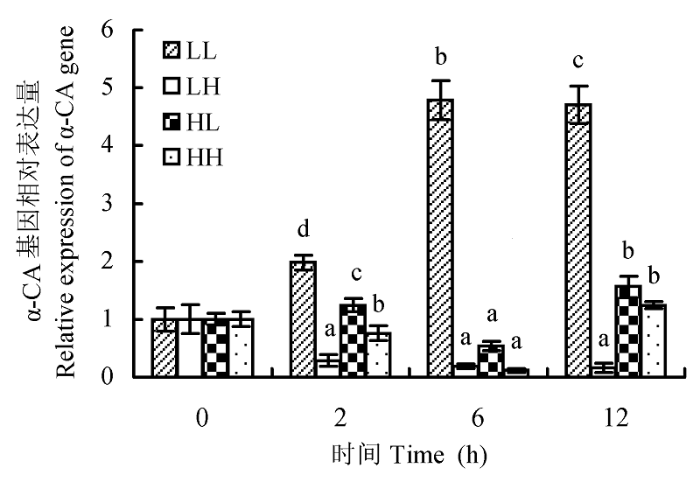
光照强度和CO2浓度还可以影响蛋白核小球藻一个胞外基因α-CA的表达(图5)。在低光强低CO2浓度条件下, 藻的α-CA基因表达量显著高于其他3组(p < 0.01), 在6 h其表达量最高, 为0 h表达量的4.79倍, 而LH、HL和HH组的α-CA基因表达量分别降为各自0 h表达量的19.11%、54.14%和11.55%。其次为高光强低CO2浓度条件, 藻的α-CA基因表达量在2 h和12 h比高CO2浓度的两组(LH和HH)高(p < 0.05)。而高CO2浓度的两组(HH和LH)藻, 在2 h和12 h时间点HH组的α-CA基因表达量要高于LH组(p < 0.05)。该结果说明低CO2浓度可以诱导α-CA基因的表达, 且低光照促进了低CO2浓度诱导的α-CA mRNA的积累。
图5 不同光照强度和CO2浓度对蛋白核小球藻α-CA基因表达的影响(平均值±标准偏差)。LL、LH、HL和HH分别表示低光强低CO2浓度、低光强高CO2浓度、高光强低CO2浓度、高光强高CO2浓度条件。不同小写字母表示差异显著(p < 0.05)。
Fig. 5 The effect of different light intensity and CO2 concentration conditions on α-CA gene expression of Chlorella pyrenoidosa (mean ± SD). LL, LH, HL and HH represent low-light intensity and low-CO2 concentration, low-light intensity and high-CO2 concentration, high-light intensity and low-CO2 concentration, high-light intensity and high-CO2 concentration conditions, respectively. Different lowercase letters indicate significant difference (p < 0.05).
空气中CO2含量升高后, 水中溶解的CO2浓度也随之增加, 藻细胞可利用的无机碳增加, 因此藻类具有较高的光合速率和生长速率。如在0.08%和0.64% CO2浓度条件下, 铜绿微囊藻(Microcystis aeruginosa)的比生长速率比空气组(0.04%)分别提高了3.20%和1.59% (李娜等, 2011)。微拟球藻(Nannochloropsis sp.)在2800 μL CO2•L-1的加富CO2并充气培养获得了最高的生物量(Hu & Gao, 2003)。高CO2浓度促进藻类生长的原因, 与培养液中CO2分压和Ci浓度升高, 导致细胞内CO2/O2的比例增加, 从而抑制了光呼吸并促进Rubisco起羧化作用有关(Gao et al., 1993)。
光照强度是影响光合自养藻光合作用和生长的重要因素。而且光照强度和CO2浓度对藻类生长具有交互作用, 如在高光强下中肋骨条藻(Skelet- onema costatum)的生长明显高于低光强, 且低光强下其生长和光合作用受低CO2浓度的限制(陈雄文和高坤山, 2003)。在高光强条件下, 高CO2浓度能促进极大螺旋藻(Spirulina maxima)生长; 而低光强条件下, 高CO2浓度对其生长则无明显影响(夏建荣和高坤山, 2001)。本文研究结果也表明高光强高CO2浓度组蛋白核小球藻生长最快, 并且不论在高或低CO2浓度条件下, 都是较高光强培养的藻生长更快。
溶液中不同形式Ci所占比例与pH值相关, 在pH值小于6.4时溶液中主要Ci为CO2, pH值在6.4-10.3之间时溶液中主要Ci为HCO3- (Moroney et al., 2011)。藻类培养液中pH值的变化是研究藻类Ci利用的一个重要参数, 也是辨别藻类利用Ci方式的一项重要指标。在pH补偿点大于9.2时, 藻类具有HCO3-利用能力, 并且pH补偿点越高, 其利用能力就越强(Maberly, 1990)。本研究中除低光强高CO2浓度组之外, 其余3组条件培养的蛋白核小球藻的pH补偿点均高于9.2, 且高光强下pH补偿点比低光强下要高, 该结果表明蛋白核小球藻不仅能吸收CO2, 也具有利用HCO3-的能力, 且其Ci利用能力受高光强条件的促进。
在高浓度CO2条件下培养, 小新月菱形藻(Nitzschia closterium var. minutissima)对Ci的亲和力明显下降, CO2补偿点升高, HCO3-的利用受限(夏建荣和余锦兰, 2009)。中肋骨条藻对CO2的亲和力随着培养液中CO2浓度升高而下降, 且高光强条件下该藻对CO2的亲和力明显高于低光强条件(陈雄文和高坤山, 2003)。本研究发现蛋白核小球藻在高浓度CO2条件下培养, 其光合Ci亲和力也下降, 与以上研究结果一致。这也与在高浓度CO2下海洋浮游植物Ci亲和力往往会降低的结果一致, 而光合Ci亲和力的变化与藻类Ci浓缩机制的调节是密切相关的(Raven, 2010; Wu et al., 2010)。
光照强度和CO2浓度在藻类CCM中发挥着重要作用, 其中一个方式就是调节CA活性。如高光强能增强铜绿微囊藻的CA活性(王山杉等, 2006); 而高浓度CO2培养的小新月菱形藻胞外CA活性下降(夏建荣和余锦兰, 2009)。在12和31 μmol·L-1 CO2时, 中肋骨条藻中没有检测出其胞外CA活性, 但在4 μmol·L-1 CO2时则有明显活性, 并且在高光强条件下其CA活性是低光强条件下的2.5倍(陈雄文和高坤山, 2003)。
本文结果表明高光强低CO2浓度培养蛋白核小球藻的胞外CA活性最高, 低光强低CO2浓度次之, 与以上陈雄文和高坤山(2003)的研究结果一致。该结果也证明了低CO2浓度可以诱导藻的胞外CA活性, 并且高光强能促进低CO2浓度诱导的CA活性升高的幅度。这与在低浓度CO2下, 藻细胞为了得到足够CO2用于光合作用, 会通过提高胞外CA活性以补偿CO2的供应, 即CCM相关。而在低光强低CO2浓度条件下, 藻的胞外CA活性比高光强低CO2浓度条件要低, 则可能与低光强限制了藻细胞的能量供应, 因此胞外CA活性降低有关。
光照强度和CO2浓度不仅可以影响CA酶的活性, 同时也影响了CA基因的转录水平。低CO2浓度可以诱导胞外CA基因的表达, 而光照强度可以提高CA基因的表达强度。如三角褐指藻(Phaeod- actylum tricornutum)的β-CA基因(ptca1)只能在低CO2浓度或空气条件下才有转录表达活性, 且在光照条件下其基因表达量比黑暗条件下增加50% (Harada et al., 2005)。但也有例外, 某些类型的CA基因表达不受低CO2浓度诱导。如三角褐指藻中6个α-和γ-CA基因表达量在空气CO2浓度和5% CO2浓度条件下没有变化(Tachibana et al., 2011)。
本文研究结果也表明低CO2浓度能诱导蛋白核小球藻胞外α-CA基因的表达; 并且在不同CO2浓度条件下, 胞外α-CA基因表达受光照的影响不同。本文胞外CA活性和α-CA基因表达结果并不一致, 虽然低CO2浓度促进了两者的表达, 但光强对CA活性和α-CA mRNA累积的影响不同。这可能是因为CA活性测定的是总的胞外酶, 而CA基因表达用的只是胞外CA编码基因中的一种α-CA有关。
高光强和高CO2浓度水平有利于促进蛋白核小球藻生长和对Ci的利用。在低CO2浓度(空气CO2浓度)条件下, 不论光强高低, 蛋白核小球藻的pH补偿点均高于9.2, 说明该藻主要利用的Ci是HCO3-。高CO2浓度会导致蛋白核小球藻对Ci亲和力下降; 与低光强条件相比, 高光强培养的蛋白核小球藻对Ci的亲和力高。高光强低CO2浓度条件能提高藻的胞外CA活性, 而低光强低CO2浓度条件能诱导一种胞外α-CA基因大量表达。可见, 蛋白核小球藻对Ci的利用效率不仅依赖于CO2浓度还依赖于光照强度的变化。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
| [1] |
Internal inorganic carbon pool of Chlamydomonas reinhardtii: Evidence for a carbon dioxide- concentrating mechanism . |
| [2] |
|
| [3] |
Active transport of CO2 and bicarbonate is induced in response to external CO2 concentration in the green alga Chlorella kessleri . |
| [4] |
Effect of CO2 concentrations on the activity of photosynthetic CO2 fixation and extracellular carbonic anhydrase in the marine diatom Skeletonema costatum .CO2浓度对中肋骨条藻的光合无机碳吸收和胞外碳酸酐酶活性的影响 . |
| [5] |
A major light- harvesting polypeptide of photosystem II functions in thermal dissipation . |
| [6] |
Influence of enhanced CO2 on growth and photosynthesis of the red algae Gracilaria sp. and G. chilensis . |
| [7] |
Research techniques and methods in characterizing photosynthetic carbon fixation by algae .藻类光合固碳的研究技术与解析方法 . |
| [8] |
External and internal carbonic anhydrases in Dunaliella species . |
| [9] |
Studies of marine planktonic diatoms: I. Cyclotella nana Hustedt, and Detonula confer- vacea (Cleve) Gran . |
| [10] |
Regulation of the expression of intracellular β-carbonic anhydrase in response to CO2 and light in the marine diatom Phaeodactylum tricornutum . |
| [11] |
Optimization of growth and fatty acid composition of a unicellular marine picoplankton, Nanno chloropsis sp., with enriched carbon sources . |
| [12] |
Cis-acting elements and DNA-binding proteins involved in CO2-responsive transcriptional activation of Cah1 encoding a periplasmic carbonic anhydrase in Chlamydomonas reinhardtii . |
| [13] |
Characterization of carbonic anhydrase II from Chlorella vulgaris in bio-CO2 capture . |
| [14] |
Effects of elevated CO2 concentration on growth of microcystis aeruginosa .大气CO2浓度变化对铜绿微囊藻生长的影响 . |
| [15] |
Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method . |
| [16] |
Exogenous sources of inorganic carbon for photosynthesis by marine macroalgae . |
| [17] |
Dissociation constants of carbonic acid in seawater as a function of salinity and temperature .
|
| [18] |
Regulation of a carbon concentrating mechanism through CCM1 in Chlamydomonas reinhardtii . |
| [19] |
Proposed carbon dioxide concentrating mechanism in Chlamydomonas reinhardtii . |
| [20] |
The car- bonic anhydrase isoforms of Chlamydomonas reinhardtii: Intracellular location, expression, and physiological roles . |
| [21] |
Acclimation of wild-type cells and CO2-insensitive mutants of the green alga Chlorella ellipsoidea to elevated [CO2] . |
| [22] |
Inorganic carbon acquisition by eukaryotic algae: Four current questions . |
| [23] |
Localization of putative carbonic anhy- drases in two marine diatoms, Phaeodactylum tricornutum and Thalassiosira pseudonana . |
| [24] |
Utilization of inorganic carbon in Pyropia haitanensis (Rhodophyta) under heat stress .高温胁迫下坛紫菜(Pyropia haitanensis)对无机碳的利用 . |
| [25] |
Modulation and adaptation of carbonic anhydrase activity in Microcystis spp. under different environmental factors .微囊藻碳酸酐酶活性在不同环境因素下的调节与适应 . |
| [26] |
Effects of salinity and inorganic carbon on the growth, extracellu- lar carbonic anhydrase activity and ca gene expression of Chlorella pyrenoidosa .盐度和无机碳对蛋白核小球藻生长、胞外碳酸酐酶活性及其基因表达的影响 .
|
| [27] |
An inorganic carbon transport system responsible for acclimation specific to air levels of CO2 in Chlamydomonas reinhardtii . |
| [28] |
Analyses of CIA5, the master regulator of the carbon-concentrating mechanism in Chlamydomonas reinhardtii, and its control of gene expression . |
| [29] |
Electrometric and colori- metric determination of carbonic anhydrase . |
| [30] |
CO2 induced seawater acidification affects physiological performance of the marine diatom Phaeodactylyum tricornutum . |
| [31] |
Effects of high CO2 concentration on growth and photosynthesis of Spirulina maxima .高浓度CO2对极大螺旋藻生长和光合作用的影响 . |
| [32] |
Advances in research on CO2 con- centrating mechanism of green algae .绿藻CO2浓缩机制的研究进展 . |
| [33] |
Effects of high CO2 concentration on ex- tracellular carbonic anhydrase activity and photosynthesis in Nitzschia closterium var. minutissima. Journal of Guangzhou University ( 高浓度CO2对小新月菱形藻胞外碳酸酐酶活性和光合作用的影响 . |
| [34] |
Expression analysis of genes associated with the induction of the carbon- concentrating mechanism in Chlamydomonas reinhardtii . |
| [35] |
The novel Myb transcription factor LCR1 regulates the CO2-responsive gene Cah1, encoding a periplasmic carbonic anhydrase in Chlamydomonas reinhardtii . |

/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}