张建华 , 唐志尧
, 唐志尧
ZHANG Jian-Hua, TANG Zhi-Yao
通讯作者:
责任编辑:
收稿日期: 2016-03-9
接受日期: 2016-09-21
网络出版日期: 2017-01-10
版权声明: 2017 植物生态学报编辑部 本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。
基金资助:
展开
摘要
土壤呼吸是陆地生态系统碳收支的重要组成部分。与森林相比, 自然或半自然的灌丛主要分布在养分贫瘠的地区, 通常认为它们对环境变化较为敏感。外源氮输入可能会显著影响灌丛的土壤呼吸。迄今为止, 人们对大气氮沉降对灌丛土壤呼吸的影响知之甚少。该文通过氮添加试验, 研究了北京东灵山荆条(Vitex negundo var. heterophylla)和绣线菊(Spiraea salicifolia)灌丛土壤呼吸及其对不同氮添加水平(对照(0)、低氮(20 kg N·hm-2·a-1)、中氮(50 kg N·hm-2·a-1)和高氮(100 kg N·hm-2·a-1))的响应。结果表明: 自然条件下, 荆条和绣线菊灌丛的土壤总呼吸年通量为5.91和4.23 t C·hm-2·a-1, 异养呼吸通量为5.76和3.53 t C·hm-2·a-1, 荆条和绣线菊灌丛的总呼吸和异养呼吸均与土壤温度呈显著的指数关系。荆条和绣线菊灌丛土壤总呼吸温度敏感性系数(Q10)的变化范围分别为1.44-1.58和1.43-1.98, 异养呼吸Q10的变化范围分别为1.38-2.11和1.49-1.88。短期氮添加抑制了荆条灌丛的自养呼吸, 而对土壤总呼吸和异养呼吸影响不明显; 氮添加促进了绣线菊灌丛的异养呼吸, 而对土壤总呼吸和自养呼吸均无显著影响; 氮添加对两种灌丛土壤呼吸年通量及土壤总呼吸Q10均无显著影响。
关键词:
Abstract
Aims Soil respiration from terrestrial ecosystems is an important component of terrestrial carbon budgets. Compared to forests, natural or semi-natural shrublands are mostly distributed in nutrient-poor sites, and usually considered to be relatively vulnerable to environmental changes. Increased nitrogen (N) input to ecosystems may remarkably influence soil respiration in shrublands. So far the effects of N deposition on shrubland soil respiration are poorly understood. The aim of this study is to investigate the soil respiration of Vitex negundo var. heterophylla and Spiraea salicifolia shrublands and their response to N deposition.
Methods We carried out a N enrichment experiment in V. negundo var. heterophylla and S. salicifolia shrublands in Mt. Dongling, Beijing, with four N addition levels (N0, control, 0; N1, low N, 20 kg N·hm-2·a-1; N2, medium N, 50 kg N·hm-2·a-1 and N3, high N, 100 kg N·hm-2·a-1). Respiration was measured from 2012-2013 within all treatments.
Important findings Under natural conditions, annual total and heterotrophic respiration were 5.91 and 4.23, 5.76 and 3.53 t C·hm-2·a-1 for the V. negundo var. heterophylla and S. salicifolia shrublands, respectively and both were not affected by short-term N addition. In both shrubland types, soil respiration rate exhibited significant exponential relationships with soil temperature. Temperature sensitivity (Q10) of total soil respiration in V. negundo var. heterophylla and S. salicifolia shrublands ranged from 1.44 to 1.58 and 1.43 to 1.98, and Q10 of heterotrophic soil respiration ranged from 1.38 to 2.11 and 1.49 to 1.88, respectively. Short-term N addition decreased only autotrophic respiration rate during the growing season, but had no significant effects on total and heterotrophic soil respiration in V. negundo var. heterophylla shrubland. In contrast, N addition enhanced the heterotrophic soil respiration rate and did not influence autotrophic and total soil respiration in S. salicifolia shrubland.
Keywords:
土壤呼吸是陆地生态系统碳循环的重要过程, 也是土壤碳库向大气输出碳的主要途径和大气CO2的重要来源(刘绍辉和方精云, 1997)。全球土壤呼吸年通量为75-120 Pg C, 是化石燃料燃烧排放量的10倍以上(Raich & Polter, 1995), 因此土壤呼吸的强弱在很大程度上决定了全球气候变化与碳循环间的反馈关系(杨庆朋等, 2011; 吴迪等, 2015)。在全球变化背景下, 土壤呼吸速率的微小变化就可能改变大气中CO2浓度和土壤碳素的周转速率(Bronson et al., 2008; 姚辉等, 2015), 进而延缓或加剧气候变化。土壤呼吸是一个复杂的生物化学过程, 在不同的生态系统中土壤呼吸受众多因素的综合影响(Bowden et al., 1993; Sitaula et al., 1995; Burton et al., 1998; Fisk & Fahey, 2001; Savage & Davidson, 2001; Hibbard et al., 2005; 张东秋等, 2005; Davidson & Janssens, 2006; Chen et al., 2010; 全权等, 2015)。
近几十年来, 人类活动排放到大气中的活性氮迅速增加, 其总量超过了所有陆地生态系统自然产生的活性氮排放, 这种趋势在未来的数十年内还将持续下去(Galloway et al., 2004; 2008)。氮沉降的持续增加已经对各生态系统的特征和过程产生了很大影响(Rabalais, 2002)。大气氮沉降引起森林的土壤酸化、影响树木生长及生物多样性, 甚至严重威胁森林生态系统的结构与功能(Matson et al., 1999; 李德军等, 2003; 莫江明等, 2005; 李秋玲等, 2013)。同样, 氮沉降会降低草原物种多样性(Stevens et al., 2004)、净氮矿化(Liu et al., 2015)及活性有机碳含量(郑娇娇等, 2012)。作为全球碳循环流通的一个关键环节, 土壤呼吸也受到大气氮沉降增加的影响, 有关氮沉降对土壤呼吸影响的研究多集中于森林和草地(Han et al., 2012; 李伟斌等, 2014; Qi et al., 2014; 吴迪等, 2015)。但在不同生态系统, 土壤呼吸对氮沉降的响应并不一致, 例如, 邓琦等(2009)发现高氮处理对鼎湖山南亚热带人工森林生态系统土壤呼吸的影响与季节的降雨量紧密相关。不同研究发现氮沉降可能促进(Diemer, 1997; Cleveland & Town-send, 2006; Ammann et al., 2007; 涂利华等, 2010; 张宇和红梅, 2014)或减缓土壤呼吸速率(贾淑霞等, 2007; 彭勇等, 2015), 但也有研究发现氮沉降对土壤呼吸无显著影响(Allison et al., 2008)。
灌丛具有种类繁多、生命力强、萌生力强、适生范围宽等特点, 不仅在群落演替过程中扮演着极其重要的角色, 而且在区域生态环境保护和替代能源方面也起着非常重要的作用(刘存琦, 1994; 胡会峰等, 2006)。然而人类的乱砍滥伐引起植被逆行演替, 大面积森林退化形成灌丛, 这种现象在华北地区尤为显著。灌丛通常为氮限制生态系统(Gorissen et al., 2004; Wessel et al., 2004)。在大气氮沉降显著增加的情况下, 灌丛碳循环一旦发生变化, 就会对全球气候变化产生深刻的影响。目前, 有关大气氮沉降对灌丛影响的研究十分缺乏, 氮沉降对灌丛的影响机制尚不清楚。荆条(Vitex negundo var. heterophylla)和绣线菊(Spiraea salicifolia)灌丛为暖温带分布面积广、类型典型的灌丛类型(谢华辉等, 2006; 张金瑞等, 2013)。本研究选择北京东灵山地区常见的荆条和绣线菊灌丛为对象, 通过氮添加实验, 研究东灵山灌丛土壤呼吸的变化特征, 以探讨土壤呼吸各组分对短期氮添加的初期响应特征, 并探讨其响应机制, 以期为预测该地区在大气氮沉降持续增加的情况下灌丛土壤的碳动态提供参考。
东灵山(40.00°-40.03° N,115.43°-115.50° E)为小五台山余脉, 位于北京市西郊的门头沟区, 最高峰海拔2303 m。该区域属于暖温带半湿润大陆季风气候, 寒冷期长, 积温低, 年平均气温5-10 ℃, 无霜期约195天, 年降水量500-650 mm, 多集中于夏季(6-8月), 春旱严重(冯云等, 2007)。地带性土壤类型为肥沃褐色土和棕色森林土。地带性植被类型为暖温带落叶阔叶林为主。但灌丛是低海拔地区最主要的植被类型, 主要分布于海拔1100 m以下, 其中荆条和绣线菊是当地的两种典型灌丛, 是森林植被遭受严重破坏后形成的次生群落(胡淑萍等, 2010)。研究区域背景氮沉降水平仅为14.5 kg N·hm-2·a-1, 基本没有受到污染(张建华, 2015)。荆条和绣线菊灌丛的样地情况见表1。
表1 试验样地地形、土壤和植被特征(平均值±标准误差, n = 3)
Table 1 Topography, soil and vegetation characteristics of the experimental sites (mean ± SE, n = 3)
| 项目 Item | 荆条灌丛 Vitex negundo var. heterophylla shrubland | 绣线菊灌丛 Spiraea salicifolia shrubland |
|---|---|---|
| 地形和气候 Topography and climate | ||
| 海拔 Elevation (m) | 791 | 1 170 |
| 坡向 Aspect | 南 South | 南 South |
| 坡度 Slope (°) | 28 | 25 |
| 年平均气温 Mean annual temperature (℃) | 12.3 | 9.2 |
| 表层土壤特征 Top soil property | ||
| 土壤pH值 Soil pH value | 8.7 | 8.9 |
| 总碳 Total carbon (mg·g-1) | 28.88 (±2.10) | 39.50 (±5.03) |
| 总氮 Total nitrogen (mg·g-1) | 2.72 (±0.32) | 2.29 (±0.36) |
| 总磷 Total phosphorous (mg·g-1) | 0.47 (±0.04) | 0.48 (±0.03) |
| 无机氮 Inorganic nitrogen (mg·kg-1) | 6.01 (±2.38) | 2.51 (±2.88) |
| 速效磷 Available phosphorous (mg·kg-1) | 1.03 (±0.09) | 1.38 (±0.77) |
| 群落特征 Community characteristics | ||
| 灌木高度 Shrub height (cm) | 78.1 (±12.37) | 79.8 (±7.43) |
| 平均基径 Average base diameter (cm) | 0.77 (±1.77) | 0.56 (±0.04) |
| 灌木密度 Shrub density (stems·hm-2) | 1.6 × 105 | 3.6 × 105 |
| 灌木层优势种 Dominant species of shrub layer | 荆条 Vitex negundo var. heterophylla, 河蒴荛花 Wikstroemia chamaedaphne | 绣线菊 Spiraea salicifolia |
| 草本层优势种 Dominant species of herb layer | 细叶薹草 Carex duriuscula subsp. stenophylloides | 细叶薹草 Carex duriuscula subsp. stenophylloides |
| 干扰程度 Levels of disturbance | 轻度干扰 Light disturbance | 轻度干扰 Light disturbance |
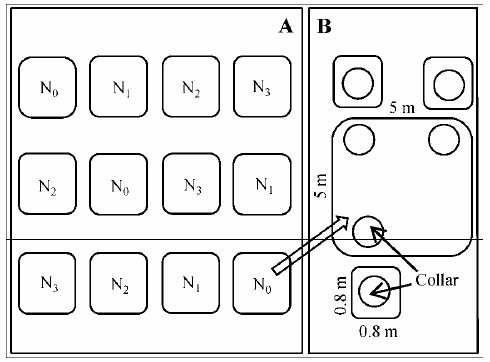
2012年5月, 在荆条和绣线菊灌丛内分别选择具有代表性、立地条件基本一致的地段, 按照随机区组试验设计方法设置实验。在每种灌丛类型内, 分别设置12个5 m × 5 m的实验样方, 相邻样方之间设置5-10 m的缓冲带(图1A)。在每个5 m × 5 m样方中随机放置3个直径20 cm的Collar环, 用于测量土壤总呼吸(Rs)。通过壕沟法切断根系区分自养呼吸和异养呼吸: 在每个样方四周挖掘1 m深(植物根系分布层以下)的壕沟后, 用厚塑料布贴在壕沟周围后将土回填, 隔离小区周围的根系, 以阻止根系向小区内生长(杜恩在, 2013)。再除去样方内所有活体植物, 设置3个Collar环, 用于土壤微生物呼吸的测定(图1B), 土壤总呼吸与土壤微生物呼吸之差为根系呼吸。
图1 北京东灵山地区灌丛氮添加试验设计。A, 样方示意图。B, Collar环示意图。N0、N1、N2、N3分别表示对照(0 kg N·hm-2·a-1)、低氮(20 kg N·hm-2·a-1)、中氮(50 kg N·hm-2·a-1)和高氮(100 kg N·hm-2·a-1)处理。
Fig. 1 Experimental design in shrublands on Mt. Dongling, Beijing, northern China. A, Schematic diagram of Quadrats. B, Schematic diagram of Collar rings. N0, N1, N2 and N3 denote control (0 kg N·hm-2·a-1), low (20 kg N·hm-2·a-1), medium (50 kg N·hm-2·a-1), and high (100 kg N·hm-2·a-1) nitrogen addition, respectively.
施肥梯度主要参照中国森林养分添加实验网络(NEECF) (Du et al., 2013)。添加氮素为尿素, 共设置4个水平, 分别为对照(N0)、低氮(N1, 相当于20 kg N·hm-2·a-1)、中氮(N2, 相当于50 kg N·hm-2·a-1)和高氮(N3, 相当于100 kg N·hm-2·a-1)。从2012年5月开始, 一直到2013年9月在生长季(5-9月)每月月初进行施肥。施肥主要通过肩背式喷雾器方式进行: 施肥前将每个样地所需尿素溶于2 L清水中(Du et al., 2013), 在样地灌丛下人工均匀地进行喷洒; 每次施肥时, 对照样地(N0)喷洒等量的清水。
使用LI-8100A土壤碳通量自动测量系统(LI-COR, Lincoln, USA)观测土壤呼吸。于2012年7月开始, 每月上旬选取天气晴朗(不包括降雨后)的9:00-11:00或13:00-16:00测定1次土壤呼吸, 同时用LI-8100自带的土壤温度和湿度传感器分别测定5 cm深度的土壤体积含水量和土壤温度。每次测量时按照不同顺序从而避免系统误差(杜恩在, 2013)。从2012年4月开始, 在每种灌丛类型地下5 cm各放置一个Stow Away Tidbit Temp Logger土壤温度记录仪(Onset Computer, Bourne, USA), 连续记录土壤温度, 采样间隔为1 h。
土壤呼吸速率(R) (μmol CO2·m-2·s-1)和5 cm土壤温度(T)以及0-5 cm土壤体积含水率(VWC)的拟合关系如下:
R = aeKT (1)
R = a VWC +b (2)
并基于上述公式(1)推导出全年土壤呼吸温度敏感性系数(Q10):
Q10= e10K (3)
a、b、K为拟合参数。
根据土壤呼吸速率与土壤温度的指数关系(公式(1)), 通过样地内放置的温度自动记录仪记录的土壤温度, 分别计算各样地年平均土壤呼吸速率和单位面积土壤CO2通量为:

Ri、Rd和Ry分别为土壤呼吸速率(μmol CO2·m-2·s-1)、土壤呼吸日通量(t C·m-2·d-1)和年通量(t C·hm-2·a-1)。
采用重复单因素方差分析比较不同氮添加水平对土壤呼吸的影响。
方差分析在软件SPSS 17.0中完成, 图表绘制均在Word 2003、Excel 2003和SigmaPlot 12.0内完成。
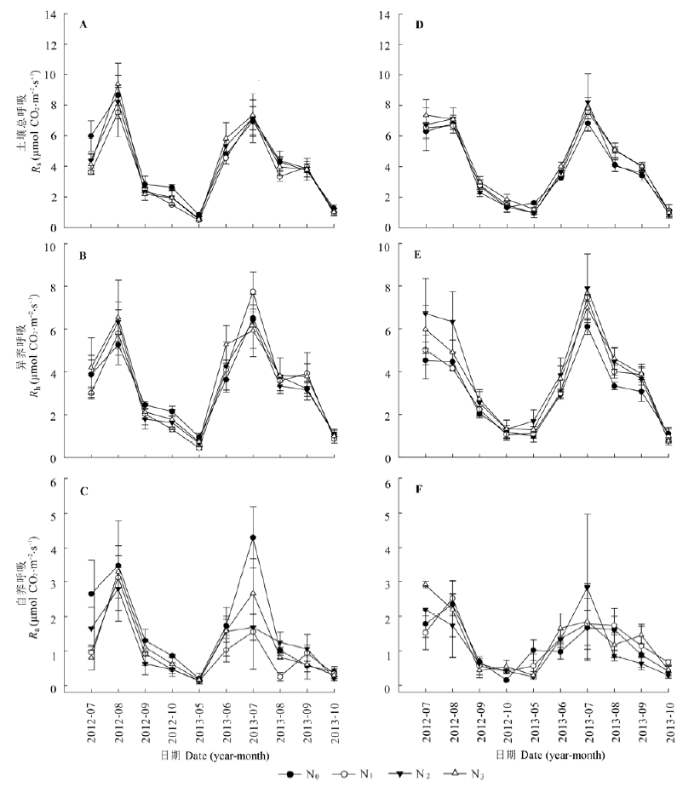
荆条和绣线菊灌丛的土壤总呼吸存在明显的季节格局(表1, p < 0.001), 总体呈单峰型。2012年的峰值均出现在8月, 2013年的峰值出现在7月(图2A, 2D)。重复测量方差分析结果显示, 在生长季, 施肥对荆条灌丛和绣线菊灌丛土壤总呼吸均无显著影响(p > 0.05) (表2)。
图2 不同氮添加对荆条(左)和绣线菊(右)灌丛土壤总呼吸(Rs)、自养呼吸(Ra)及异养呼吸(Rh)的影响(平均值±标准误差)。N0、N1、N2、N3分别表示对照(0 kg N·hm-2·a-1)、低氮(20 kg N·hm-2·a-1)、中氮(50 kg N·hm-2·a-1)和高氮(100 kg N·hm-2·a-1)处理。
Fig. 2 Influence of different nitrogen addition on the soil total respiration (Rs), autotrophic respiration (Ra) and heterotrophic respiration (Rh) of Vitex negundo var. heterophylla (left) and Spiraea salicifolia (right) shrublands (mean ± SE). N0, N1, N2 and N3 denote control (0 kg N·hm-2·a-1), low (20 kg N·hm-2·a-1), medium (50 kg N·hm-2·a-1), and high (100 kg N·hm-2·a-1) nitrogen addition, respectively.
表2 时间和氮添加处理对荆条和绣线菊灌丛土壤总呼吸(Rs)、异养呼吸(Rh)及自养呼吸(Ra)影响的双因子方差分析
Table 2 Two-way ANOVA test results for the effects of time and nitrogen addition treatments on total soil respiration (Rs), heterotrophic respiration (Rh) and autotrophic respiration rate (Ra) in Vitex negundo var. heterophylla and Spiraea salicifolia shrublands
| 自由度 Degree of freedom | Rs | Rh | Ra | ||||||
|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | ||||
| 荆条灌丛 Vitex negundo var. heterophylla shrubland | |||||||||
| 处理 Treatment | 3 | 1.51 | 0.218 | 0.33 | 0.807 | 3.33 | 0.024 | ||
| 时间 Time1) | 9 | 52.49 | <0.001 | 33.18 | <0.001 | 14.60 | <0.001 | ||
| 处理×时间 Treatment × Time | 27 | 0.42 | 0.994 | 0.45 | 0.989 | 0.88 | 0.631 | ||
| 绣线菊灌丛 Spiraea salicifolia shrubland | |||||||||
| 处理 Treatment | 3 | 1.49 | 0.224 | 4.43 | 0.006 | 0.42 | 0.740 | ||
| 时间 Time | 9 | 88.65 | <0.001 | 42.90 | <0.001 | 7.25 | <0.001 | ||
| 处理×时间 Treatment × Time | 27 | 0.39 | 0.996 | 0.49 | 0.980 | 0.32 | 0.999 | ||
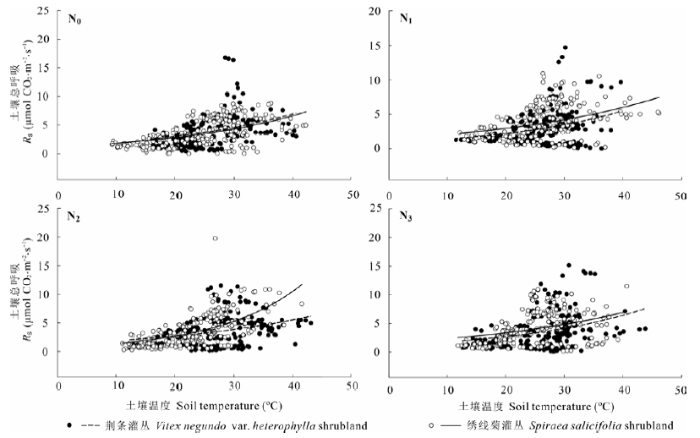
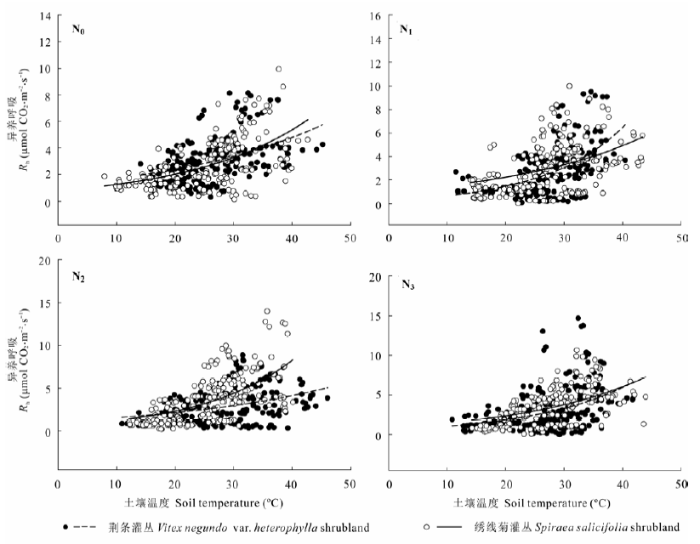
荆条和绣线菊灌丛土壤总呼吸与土壤温度呈显著的指数关系(图3)。两灌丛对氮添加的响应不同, 具体表现在, 低高氮(N1, N3)处理和中氮(N2)处理在一定程度上分别提高和降低了荆条灌丛土壤呼吸Q10, 而绣线菊灌丛在相应氮处理的土壤呼吸Q10的变化却表现出相反趋势。N0、N1、N2、N3处理下荆条和绣线菊灌丛土壤总呼吸的Q10分别为1.52、1.58、1.44、1.55和1.54、1.43、1.98、1.44 (表3)。施肥没有显著改变Q10。各处理水平的土壤总呼吸与土壤水分均显著正相关(图4)。
图3 不同氮添加处理的荆条和绣线菊灌丛土壤总呼吸(Rs)与土壤温度的关系。N0、N1、N2、N3分别表示对照(0 kg N·hm-2·a-1)、低氮(20 kg N·hm-2·a-1)、中氮(50 kg N·hm-2·a-1)和高氮(100 kg N·hm-2·a-1)处理。
Fig. 3 Relationship between total soil respiration (Rs) and soil temperature in Vitex negundo var. heterophylla and Spiraea salicifolia shrublands under different nitrogen addition treatments. N0, N1, N2 and N3 denote control (0 kg N·hm-2·a-1), low (20 kg N·hm-2·a-1), medium (50 kg N·hm-2·a-1), and high (100 kg N·hm-2·a-1) nitrogen addition, respectively.
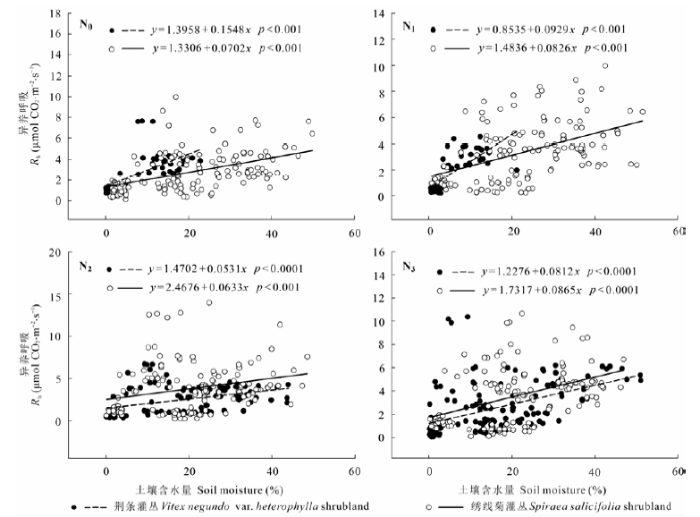
图4 不同处理的荆条和绣线菊灌丛土壤总呼吸(Rs)与土壤含水量的关系。N0、N1、N2、N3分别表示对照(0 kg N·hm-2·a-1)、低氮(20 kg N·hm-2·a-1)、中氮(50 kg N·hm-2·a-1)和高氮(100 kg N·hm-2·a-1)处理。
Fig. 4 Relationship between total soil respiration (Rs) and soil moisture in Vitex negundo var. heterophylla and Spiraea salicifolia shrublands under different nitrogen addition treatments. N0, N1, N2 and N3 denote control (0 kg N·hm-2·a-1), low (20 kg N·hm-2·a-1), medium (50 kg N·hm-2·a-1), and high (100 kg N·hm-2·a-1) nitrogen addition, respectively.
表3 土壤呼吸(R, μmol CO2·m-2·s-1)和组分与5 cm土壤温度(T, °C))的指数关系模型(R = aeKt)
Table 3 Models (R = aeKt) for the relationship between total and component of soil respiration (R, μmol CO2·m-2·s-1) and soil temperature 5 cm under the surface
| 灌丛类型 Shrubland type | 呼吸组分 Respiration component | 处理 Treatment | n | R2 | 参数 Parameter | ||
|---|---|---|---|---|---|---|---|
| a | K | Q10 | |||||
| 荆条 Vitex negundo var. heterophylla | 土壤总呼吸 | N0 | 217 | 0.13 | 1.322 | 0.042 | 1.52 |
| Total soil respiration | N1 | 227 | 0.08 | 0.878 | 0.046 | 1.58 | |
| N2 | 231 | 0.13 | 1.296 | 0.036 | 1.44 | ||
| N3 | 234 | 0.12 | 1.109 | 0.044 | 1.55 | ||
| 异养呼吸 | N0 | 220 | 0.20 | 1.183 | 0.035 | 1.42 | |
| Heterotrophic respiration | N1 | 221 | 0.26 | 0.327 | 0.075 | 2.11 | |
| N2 | 224 | 0.14 | 1.137 | 0.032 | 1.38 | ||
| N3 | 242 | 0.18 | 0.594 | 0.058 | 1.78 | ||
| 绣线菊 Spiraea salicifolia | 土壤总呼吸 | N0 | 228 | 0.28 | 1.192 | 0.043 | 1.54 |
| Total soil respiration | N1 | 218 | 0.16 | 1.445 | 0.036 | 1.43 | |
| N2 | 232 | 0.39 | 0.678 | 0.069 | 1.98 | ||
| N3 | 216 | 0.15 | 1.609 | 0.037 | 1.44 | ||
| 异养呼吸 | N0 | 224 | 0.33 | 0.753 | 0.049 | 1.63 | |
| Heterotrophic respiration | N1 | 221 | 0.22 | 1.003 | 0.040 | 1.49 | |
| N2 | 234 | 0.39 | 0.666 | 0.063 | 1.88 | ||
| N3 | 213 | 0.31 | 0.942 | 0.047 | 1.59 | ||
荆条和绣线菊灌丛的异养呼吸存在明显的季节格局, 总体呈单峰型。荆条灌丛的土壤异养呼吸在2012和2013年的峰值分别出现在8月和7月(图2B, 2E), 而绣线菊灌丛的峰值均出现在7月。各处理对荆条灌丛的异养呼吸无显著影响(图2B), 在生长季, N2和N3处理显著地促进了绣线菊灌丛的异养呼吸, 而N1处理效果不显著(图2E; 表2)。
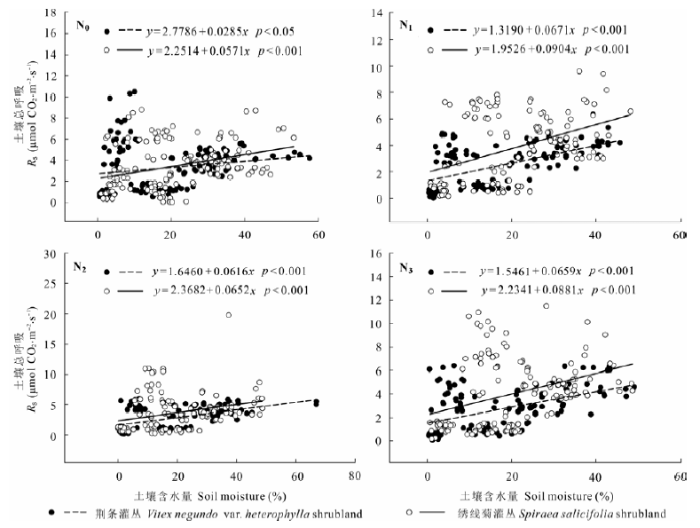
各氮处理水平下, 荆条和绣线菊灌丛土壤异养呼吸与土壤温度呈显著的指数关系(图5; 表3)。氮添加对两灌丛土壤异养呼吸温度敏感性的影响不同, N1和N3处理提高了荆条灌丛土壤异养呼吸温度敏感性, 而N2处理降低了其温度敏感性; N0、N1、N2、N3处理下的Q10分别为1.42、2.11、1.38、1.78 (图5; 表3)。N1和N3处理降低了绣线菊灌丛土壤异养呼吸温度敏感性, 而N2处理提高了其温度敏感性; N0、N1、N2、N3处理下的Q10分别为1.63、1.49、1.88、1.59。各氮处理水平下, 荆条和绣线菊灌丛的土壤异养呼吸与土壤水分均显著正相关(图6)。
图5 不同氮添加处理的荆条和绣线菊灌丛土壤异养呼吸(Rh)与土壤温度的关系。N0、N1、N2、N3分别表示对照(0 kg N·hm-2·a-1)、低氮(20 kg N·hm-2·a-1)、中氮(50 kg N·hm-2·a-1)和高氮(100 kg N·hm-2·a-1)处理。
Fig. 5 Relationship between soil heterotrophic respiration (Rh) and soil temperature in Vitex negundo var. heterophylla and Spiraea salicifolia shrublands under different nitrogen addition treatments. N0, N1, N2 and N3 denote control (0 kg N·hm-2·a-1), low (20 kg N·hm-2·a-1), medium (50 kg N·hm-2·a-1), and high (100 kg N·hm-2·a-1) nitrogen addition, respectively.
图6 不同氮处理的荆条和绣线菊灌丛土壤异养呼吸(Rh)与土壤含水量的关系。N0、N1、N2、N3分别表示对照(0 kg N·hm-2·a-1)、低氮(20 kg N·hm-2·a-1)、中氮(50 kg N·hm-2·a-1)和高氮(100 kg N·hm-2·a-1)处理。
Fig. 6 Relationship between soil heterotrophic respiration (Rh) and soil moisture in Vitex negundo var. heterophylla and Spiraea salicifolia shrublands under different nitrogen addition treatments. N0, N1, N2 and N3 denote control (0 kg N·hm-2·a-1), low (20 kg N·hm-2·a-1), medium (50 kg N·hm-2·a-1), and high (100 kg N·hm-2·a-1) nitrogen addition, respectively.
荆条和绣线菊灌丛的土壤自养呼吸均存在明显的季节格局, 且变化趋势相似, 总体呈单峰型。2012年的峰值均出现在8月, 2013年的峰值出现在7月(图2C、2F)。施氮降低了荆条灌丛土壤的自养呼吸, 其中N1处理对荆条灌丛土壤自养呼吸表现为显著的抑制效应。在生长季, 氮添加对绣线菊灌丛的自养呼吸无明显的影响(图2C、2F)。
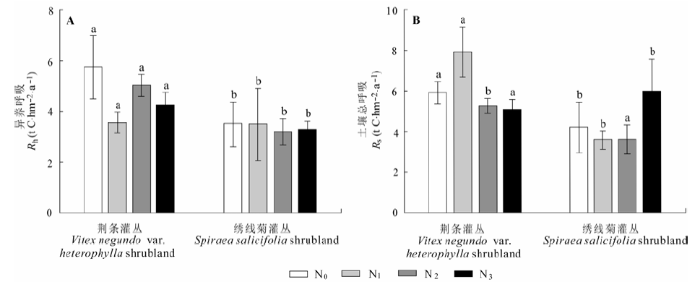
在荆条灌丛样地, N0、N1、N2和N3处理的土壤总呼吸年碳排放量分别为: 5.91、7.93、5.30和5.09 t C·hm-2·a-1, 低氮处理的排放比对照高出34.1%, 中氮和高氮处理的排放量几乎相同, 排放量均低于对照, 约降低了10.4%; 氮添加降低了异养呼吸, 各氮处理的异养呼吸年排放量分别为: 5.76、3.56、5.04和4.25 t C·hm-2·a-1 (图7A、7B)。方差分析结果表明, 氮添加对荆条灌丛土壤呼吸、异养呼吸年通量均无显著影响。
图7 氮添加对荆条和绣线菊灌丛异养呼吸(Rh)和土壤总呼吸(Rs)年通量的影响(平均值±标准误差)。相同字母a和b上标表示各处理间无显著差异(p > 0.05)。N0、N1、N2、N3分别表示对照(0 kg N·hm-2·a-1)、低氮(20 kg N·hm-2·a-1)、中氮(50 kg N·hm-2·a-1)和高氮(100 kg N·hm-2·a-1)处理。
Fig. 7 Influence of nitrogen addition on soil heterotrophic (Rh) and total (Rs) respiration in Vitex negundo var. heterophylla and Spiraea salicifolia shrublands (mean ± SE). The same letter a and b indicate no significant (p > 0.05) among treatments. N0, N1, N2 and N3 denote control (0 kg N·hm-2·a-1), low (20 kg N·hm-2·a-1), medium (50 kg N·hm-2·a-1), and high (100 kg N·hm-2·a-1) nitrogen addition, respectively.
在绣线菊样地, 各氮添加样地的土壤总呼吸年碳排放量分别为: 4.23、3.60、3.60和5.99 t C·hm-2·a-1; 高氮处理的排放比对照高出41.7%; 中氮和低氮处理的排放量相同, 均低于对照, 约降低了14.7%; 各氮添加样地的异养呼吸同对照组很接近(图7A、7B)。氮添加对绣线菊灌丛土壤总呼吸、异养呼吸年通量均无显著的影响。
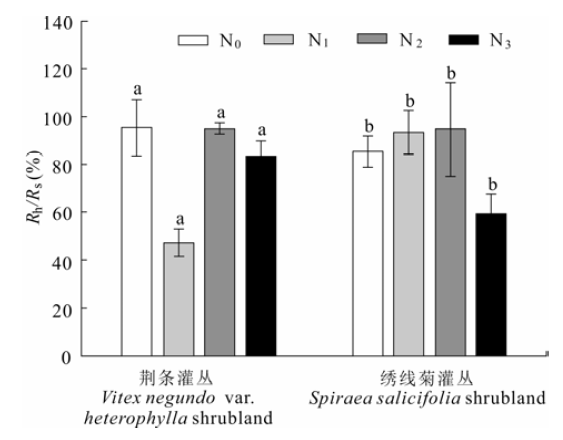
从图8和表4可知, 荆条样地对照组中异养呼吸占总呼吸比例最高(95.0%); 中氮处理(N2)下异养呼吸所占比例与对照组的接近; 低氮处理(N1)的异养呼吸占总呼吸的比例最低(47.0%)。在绣线菊样地, 中氮(N2)处理样地的异养呼吸占总呼吸的比例最高(95.0%), 高氮(N3)处理的异养呼吸贡献最低(54.5%)。
图8 氮添加对荆条和绣线菊灌丛异养呼吸组分贡献率(Rh/Rs)的影响(平均值±标准误差)。图中相同字母表示各处理间差异不显著(p > 0.05)。N0、N1、N2、N3分别表示对照(0 kg N·hm-2·a-1)、低氮(20 kg N·hm-2·a-1)、中氮(50 kg N·hm-2·a-1)和高氮(100 kg N·hm-2·a-1)处理。
Fig. 8 Influence of nitrogen addition on contributions of heterotrophic respiration to total soil respiration (Rh/Rs) in Vitex negundo var. heterophylla and Spiraea salicifolia shrublands (mean ± SE). The same letter a and b indicate no significant (p > 0.05) among treatments. N0, N1, N2 and N3 denote control (0 kg N·hm-2·a-1), low (20 kg N·hm-2·a-1), medium (50 kg N·hm-2·a-1), and high (100 kg N·hm-2·a-1) nitrogen addition, respectively.
表4 2013年不同处理下土壤总呼吸和异养呼吸的年通量(平均值±标准误差, n = 3)
Table 4 Annual flux of soil total (Rs) and heterotrophic (Rh) respiration (t C·hm-2·a-1) in 2013 under different nitrogen addition treatments (mean ± SE, n = 3)
 |
土壤呼吸在不同气候带和植被类型存在很大差异, 并与净初级生产力密切相关(涂丽华等, 2011)。本研究结果表明, 东灵山荆条和绣线菊灌丛土壤总呼吸的年通量分别为5.91和4.23 t C·hm-2·a-1, 均显著低于南亚热带鼎湖山季风常绿阔叶林(10.80 t C·hm-2·a-1)、北亚热带-南暖温带过渡区锐齿栎(Quercus aliena var. acuteserrata)老林(7.79 t C·hm-2·a-1)、北亚和热带北缘锐齿栎林(7.71 t C·hm-2·a-1)(胡正华等, 2010), 但与当地白桦(Betula platyphylla)林(5.74 t C·hm-2·a-1)、辽东栎(Quercus mongolica)林(4.55 t C·hm-2·a-1)和油松(Pinus tabuliformis)林(4.14 t C·hm-2·a-1) (姚辉等, 2015)的土壤总呼吸相差不大。说明不同植被类型土壤呼吸与纬度地带性的温度变化有一定的相关性, 土壤呼吸的差异可能与气候状况、植被类型、树龄和立地条件等因素有关(胡正华等, 2010)。
土壤呼吸受到土壤温度、湿度等因子的综合影响, 其过程十分复杂(彭勇等, 2015)。国内外许多研究表明, 森林土壤呼吸与土壤温度有良好的相关性(Jassal et al., 2007; 刘盛梅等, 2010)。在热带、亚热带森林和北温带森林中均发现土壤呼吸与地表温度之间具有极显著的指数关系(Jassal et al., 2007; 刘盛梅等, 2010)。本研究利用温度单因素指数模型拟合呼吸对温度的响应函数, 相关性极显著, 表明随着温度的升高, 土壤呼吸呈指数增长, 与国内外许多研究结果(Elberling & Brandt, 2003; 陈宝玉等, 2007; 王小国等, 2007)相同。对众多植被的研究结果表明, 土壤呼吸与土壤湿度之间也存在正相关关系(Sotta et al., 2004)。本研究中土壤呼吸与土壤温度和含水量表现出较强的相关性, 土壤含水量会影响生物的代谢, 使得土壤水分成为土壤呼吸的限制因子, 而温度通过直接影响植物、微生物活动和间接地改变土壤湿度、输入土壤有机物的数量和质量对土壤呼吸造成影响(Chapin, 2002), 成为东灵山灌丛土壤呼吸的另一主导影响因子。在各种生态系统中, Q10在时间和空间上的变化可能非常大(Xu & Qi, 2001; Curiel et al., 2004), 本研究中Q10的值在1.4- 2.1之间, 与全球Q10值范围(1.3-3.3) (Raich & Potter, 1995)相一致。
本研究发现氮添加对荆条灌丛生长季的土壤总呼吸、异养呼吸无显著影响(图2; 表2)。这一结果与长期实验所发现的施氮促进了土壤呼吸的结果 (Bowden et al., 2004; Micks et al., 2004; 莫江明等, 2005)并不一致。这可能与施肥时间短、实验期间降水量少等因素导致土壤养分利用效率很低有关, 有关机制还需要更长时间的施肥处理才能验证。N2和N3处理促进了绣线菊灌丛生长季土壤的异养呼吸, 这可能与氮添加导致绣线菊细根生物量和代谢强度增加、增加土壤有机质进而增强微生物活性等有关(Zak et al., 2000; Bowden et al., 2004; 涂利华等, 2010)。氮添加对荆条和绣线菊灌丛土壤呼吸的年通量均无显著的影响, 可能原因在于东灵山灌丛地处阳坡, 白天蒸发量大, 再加上在实验期间雨量较少, 导致灌丛土壤含水量极低; 而土壤水分对土壤养分的溶解转移、微生物活动、矿物分化等都有重要影响, 土壤水分的缺乏导致上述过程受阻, 从而减弱了施肥对呼吸的影响。研究发现, 氮添加会改变土壤微生物群落结构, 而这种改变在不同类型森林会存在差异, 这会导致不同类型森林土壤呼吸对氮添加的响应不同。此外, 氮素的添加效应还取决于氮添加量、土壤氮素水平和土壤可利用性氮含量(Demoling et al., 2008)。通常情况下, 低氮添加可以促进植物生长、增加凋落物产量, 使较多的碳分配至地下部分等, 最终增强土壤呼吸(Bowden et al., 2004), 而高氮添加会导致土壤可利用性氮含量增加, 引起植物地下部分的碳素分配减少(Litton et al., 2007), 最终抑制根际的自养呼吸(Treseder, 2004)。氮添加还会降低微生物生物量和活性, 抑制土壤有机质分解, 使土壤呼吸降低(Bowden et al., 2004)。
本研究同时发现, 氮添加对荆条和绣线菊灌丛土壤呼吸的温度敏感性均无显著影响, 可能是因为施氮没有改变根系组织氮含量, 从而保持了较恒定的细胞呼吸(贾淑霞等, 2007)。但两种灌丛对氮添加的响应存在差异, 具体表现在: N1和N3在一定程度上提高了荆条灌丛土壤呼吸的Q10, 而N2降低了其土壤呼吸的Q10; 绣线菊灌丛在相应氮处理下Q10的变化趋势与荆条灌丛刚好相反, 这可能与氮添加改变了荆条和绣线菊灌丛土壤微生物数量和群落结构, 而不同的微生物类群对温度敏感性不同有关(彭勇等, 2015)。此外, 与森林相比, 灌丛所处的环境异质性较大, 样地用于测呼吸的Collar环所在的微环境可能会非常不同, 会干扰氮添加的影响效应, 使短期氮添加对灌丛土壤呼吸Q10的影响趋势不明了。由于施肥时间短, 氮沉降对土壤呼吸的影响可能存在阶段性, 在长期持续氮沉降条件下, 灌丛土壤呼吸的温度敏感性是如何变化的, 还需进行深入研究。
综上所述, 自然条件下, 北京东灵山灌丛的土壤呼吸主要受温度和土壤含水量的影响; 荆条和绣线菊灌丛土壤呼吸的年通量和土壤呼吸温度敏感性对短期氮添加的响应均不显著, 但变化趋势不同, 可能与两灌丛的群落组成结构和生态过程不同、试验时间短、样地异质性等因素有关。灌丛土壤呼吸对未来持续增加的氮沉降会如何响应还有待进一步研究。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
| [1] |
Microbial activity and soil respiration under nitrogen addition in Alaskan boreal forest . |
| [2] |
The carbon budget of newly established temperate grassland depends on management intensity . |
| [3] |
Chronic nitrogen additions reduce total soil respiration and microbial respiration in temperate forest soils at the Harvard Forest . |
| [4] |
Contributions of aboveground litter, below- ground litter, and root respiration to total soil respiration in a temperate mixed hardwood forest . |
| [5] |
Response of soil surface CO2 flux in a boreal forest to ecosystem warming . |
| [6] |
Drought reduces root respiration in sugar maple forests . |
| [7] |
Principles of Terrestrial Ecosystem Ecology . |
| [8] |
The relationship between soil respiration and the temperature at different soil depths in subalpine coniferous forest of western Sichuan Province .川西亚高山针叶林土壤呼吸速率与不同土层温度的关系 . |
| [9] |
Modeling interannual variability of global soil respiration from climate and soil properties . |
| [10] |
Nutrient additions to a tropical rain forest drive substantial soil carbon dioxide losses to the atmosphere . |
| [11] |
Annual Ql0 of soil respiration reflects plant phenological patterns as well as temperature sensitivity . |
| [12] |
Temperature sensitivity of soil carbon decomposition and feedbacks to climate change . |
| [13] |
Bacterial and fungal response to nitrogen fertilization in three coniferous forest soils . |
| [14] |
Effects of CO2 enrichment, high nitrogen deposition and high precipitation on a model forest ecosystem in southern China .CO2浓度倍增、高氮沉降和高降雨对南亚热带人工模拟森林生态系统土壤呼吸的影响 .
|
| [15] |
Effects of elevated CO2 on gas exchange characteristics of alpine grassland . |
| [16] |
Impacts of Nitrogen Enrichment on Carbon Cycling in an Old-growth Larch (Larix gmelinii Rupr.) Forest .氮添加对兴安落叶松原始林碳收支主要过程的影响 . |
| [17] |
NEECF: A project of nutrient enrichment experiments in China’s forests .
|
| [18] |
Uncoupling of microbial CO2 production and release in frozen soil and its implications for field studies of arctic C cycling . |
| [19] |
Species abundance distribution of Quercus liaotungensis forest along altitudinal gradient in Dongling Mountain, Beijing .北京东灵山辽东栎(Quercus liaotungensis)林沿海拔梯度的物种多度分布 . |
| [20] |
Microbial biomass and nitrogen cycling responses to fertilization and litter removal in young northern hardwood forests . |
| [21] |
Nitrogen cycles: Past, present, and future . |
| [22] |
Transformation of the nitrogen cycle: Recent trends, questions, and potential solutions . |
| [23] |
Climate change affects carbon allocation to the soil in shrublands . |
| [24] |
Effects of mowing and nitrogen addition on soil respiration in three patches in an old field grassland in Inner Mongolia . |
| [25] |
An analysis of soil respiration across northern hemisphere temperate ecosystems . |
| [26] |
Vegetation carbon storage of major shrublands in China .中国主要灌丛植被碳储量 . |
| [27] |
Quantification analysis on site conditions of natural Vitex negundo community in Beijing mountainous area .北京山区天然荆条灌丛立地条件的数量化分析 . |
| [28] |
Effects of simulated nitrogen deposition on soil respiration in northern subtropical deciduous broad-leaved forest .模拟氮沉降对北亚热带落叶阔叶林土壤呼吸的影响 . |
| [29] |
Components of ecosystem respiration and an estimate of net primary productivity of an Intermediate-aged Douglas-fir stand . |
| [30] |
Effect of nitrogen fertilization on soil respiration in Larix gmelinii and Fraxinus mandshurica plantations in China .施肥对落叶松和水曲柳人工林土壤呼吸的影响 . |
| [31] |
Impact of nitrogen deposition on forest plants .氮沉降对森林植物的影响 . |
| [32] |
Effects of simulated nitrogen deposition on soil chemical properties of forests .模拟氮沉降对森林土壤化学性质的影响 . |
| [33] |
Response of soil respiration to enhanced nitrogen deposition in broadleaved Korean pine forest in Changbai Mountains .长白山阔叶红松林土壤呼吸对氮沉降增加的响应 . |
| [34] |
Carbon allocation in forest ecosystems . |
| [35] |
The study on techniques in determining shrub phytomass .灌木植物量测定技术的研究 . |
| [36] |
Effect factors of soil respiration and the temperature’s effects on soil respiration in the global scale .土壤呼吸的影响因素及全球尺度下温度的影响 . |
| [37] |
Response of soil respiration to simulated nitrogen deposition under the middle-aged Eucalyptus grandis plantation in Ya’an City .华西雨屏区巨桉中龄林土壤呼吸对模拟氮沉降的响应 . |
| [38] |
Effects of simul- ated nitrogen deposition on soil net nitrogen mineralization in the meadow steppe of Inner Mongolia, China . |
| [39] |
The globalization of N deposition: Ecosystem consequences in tropical environments . |
| [40] |
Short term soil respiration and nitrogen immobilization response to nitrogen applications in control and nitrogen enriched temperate forests . |
| [41] |
The short- term responses of soil CO2 emission and CH4 uptake to simulated N deposition in nursery and forests of Dinghushan in subtropical China .鼎湖山苗圃和主要森林土壤CO2排放和CH4吸收对模拟氮沉降的短期响应 . |
| [42] |
Effects of simulated nitrogen deposition on soil respiration in a secondary evergreen broad-leaved forest on Wawushan Mountain .模拟氮沉降对瓦屋山常绿阔叶次生林土壤呼吸的影响 . |
| [43] |
Differential responses of short-term soil respiration dynamics to the experimental addition of nitrogen and water in the temperate semi-arid steppe of Inner Mongolia, China . |
| [44] |
Short-term effect of nitrogen addition on soil respiration of three temperate forests in Dongling Mountain .短期氮添加对东灵山三种森林土壤呼吸的影响 . |
| [45] |
Nitrogen in aquatic ecosystems . |
| [46] |
Global patterns of carbon dioxide emissions from soils . |
| [47] |
Interannual variation of soil respiration in two New England forests . |
| [48] |
N-fertilization and soil acidification effects on N2O and CO2 emission from temperate pine forest soil . |
| [49] |
Soil CO2 efflux in a tropical forest in the central Amazon . |
| [50] |
Impact of nitrogen deposition on the species richness of grasslands . |
| [51] |
A meta-analysis of mycorrhizal responses to nitrogen, phosphorus, and atmospheric CO2 in field studies . |
| [52] |
Effects of simulated nitrogen deposition on soil respiration in aBambusa pervariabilis × Dendrocala mopsi plantation in rainy area of West China .模拟氮沉降对华西雨屏区撑绿杂交竹林土壤呼吸的影响 . |
| [53] |
Effects of simulated nitrogen deposition on the fine root characteristics and soil respiration in a Pleioblastus amarus plantation in rainy area of West China .模拟氮沉降对华西雨屏区苦竹林细根特性和土壤呼吸的影响 . |
| [54] |
Soil respiration and its sensitivity to temperature under different land use conditions .不同土地利用方式下土壤呼吸及其温度敏感性 . |
| [55] |
A qualitative ecosystem assessment for different shrublands in western Europe under impact of climate change . |
| [56] |
Effects of simulated nitrogen deposition on the each component of soil respiration in the Populus L. plantations in a riparian zone of the mid-lower Yangtze River .模拟氮沉降对长江中下游滩地杨树林土壤呼吸各组分的影响 . |
| [57] |
Resources of the Spiraeas and their application to landscaping .绣线菊属植物资源及其在园林中的应用前景 . |
| [58] |
Spatial and seasonal variations ofQ10 determined by soil respiration measurements at a Sierra Nevada forest . |
| [59] |
Impact factors and uncertainties of the temperature sensitivity of soil respiration .土壤呼吸温度敏感性的影响因素和不确定性 . |
| [60] |
Soil respiration and the 20-year change in three temperate forests in Mt. Dongling, Beijing .北京东灵山3种温带森林土壤呼吸及其20年的变化 . |
| [61] |
Atmospheric CO2, soil-N availability, and allocation of biomass and nitrogen byPopulus tremuloides . |
| [62] |
Some advance in the main factors controlling soil respiration .土壤呼吸主要影响因素的研究进展 . |
| [63] |
Effects of Nitrogen Addition on Carbon Cycling of Shrublands in Mt. Dongling, Beijing .氮添加对北京东灵山灌丛碳循环的影响 . |
| [64] |
Point pattern analysis for relationships of Vitex negundo var. heterophylla in three typical stands .3种典型立地荆条种群及种间分布的空间点格局 . |
| [65] |
Response of soil respiration to experimental warming and nitrogen addition in Inner Mongolia desert steppe .内蒙古荒漠草原土壤呼吸对模拟增温和氮素添加的响应 . |
| [66] |
Effects of N addition on soil organic carbon components in an alpine meadow on the eastern Qinghai-Tibetan Plateau .增氮对青藏高原东缘典型高寒草甸土壤有机碳组成的影响 . |

Copyright © 2026 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19
![]()
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}