生物入侵作为全球变化的一个重要方面, 严重威胁了土著生物群落的生物多样性和生态系统功能(Davis, 2003 ; Hooper et al ., 2005 )。作为我国中亚热带和南亚热带过渡区最具代表性的河口湿地之一的闽江河口湿地(刘剑秋等, 2006 ), 自2002年以来逐渐遭到互花米草(Spartina alterniflora )的大规模入侵。互花米草以其极强的适应力和繁殖力迅速占据空生态位, 侵占本地种短叶茳芏(Cyperus malaccensis var. brevifolius )的生境, 在潮间带形成了短叶茳芏单种群落、互花米草单种群落和互花米草-短叶茳芏混生群落共存的竞争格局。入侵的互花米草通过影响土著生态系统的结构, 进而影响生态系统的养分循环, 生态系统养分的可利用性改变反过来又影响植物的生长状况, 从而可能改变入侵种与土著种之间的竞争关系。养分元素氮(N)和磷(P)是湿地生态系统植物生长的主要限制资源, 其可利用性及生态化学计量关系是植物生长和生态系统物质循环的重要调节因子(熊汉锋和王运华, 2005 )。植物能在外界养分元素组成改变的情况下, 一定程度上维持自身各元素之间的相对比例。这种由生物在长期进化过程中为适应外界环境变化而形成的维持自身化学组成相对稳定的能力就叫作生态化学计量内稳性(简称内稳性)(Kooijman, 1995 )。
国外内稳性研究的对象由最初的真菌和细菌(Levi & Cowling, 1969 ; Goldman et al ., 1987 )发展到藻类、浮游动物和草本植物(Rhee, 1978 ; Andersen & Hessen, 1991 ; Güsewell et al ., 2005 )。由于高等植物体内元素的内稳性和化学计量特征复杂, 对高等植物的研究甚少(Cernusak et al ., 2009 ; Matzek & Vitousek, 2009 ; Yu et al ., 2010 )。生态系统水平上对内稳性的研究多集中于海洋生态系统和陆地生态系统(Güsewell, 2004 ; 苏强, 2012 ), 对湿地生态系统的研究较少(Wang et al ., 2014 )。国内内稳性的研究还处于起步阶段。庾强(2009)以内蒙古草原为研究对象, 通过1 200 km的横断范围的样带试验和内蒙古草原生态系统定位研究站27年的监测数据发现, 内稳性高的物种具有较高的优势度和生物量稳定性, 内稳性高的生态系统则具有较高的生产力和稳定性。因此, 内稳性是衡量物种竞争力的一个重要指标。已有研究表明内稳性的高低受多种因子的影响, 其中包括植物器官(庾强, 2009 )、养分供应(严正兵等, 2013 )、光强以及施肥(Sterner & Elser, 2002 )等, 影响因子的不确定性使得不同研究中内稳性对环境的响应不一致。目前有关互花米草入侵对本地生态系统的影响多集中在物质循环以及生态化学计量学, 而物种水平上内稳性的研究甚少(张仁懿等, 2015 )。已有研究发现, 闽江河口湿地生长季阶段的入侵种互花米草和本地种短叶茳芏均具有一定的内稳性, 互花米草N和N:P的内稳性高于短叶茳芏N和N:P的内稳性, 而短叶茳芏P的内稳性高于互花米草P的内稳性(蒋利玲等, 2014 )。
为了进一步研究湿地生态系统植物内稳性与生态系统功能的关系, 我们以互花米草入侵为切入点, 以短叶茳芏单种群落、互花米草单种群落以及互花米草-短叶茳芏混生群落为研究对象, 对其土壤和植物的养分动态以及植物的内稳性进行了研究, 旨在解决以下问题: 1)入侵过程如何影响外来种和本地种的限制性养分以及植物内稳性?2)元素类型、器官类型和物种类型等不同因子对物种植物内稳性有什么影响?
1 材料和方法
1.1 试验设计
本试验在福建省闽江河口鳝鱼滩湿地(26.01°- 26.06° N, 119.57°-119.68° E, 图1 )进行。该湿地为闽江河口区面积最大的砂泥质洲滩天然湿地。该区域气候暖热湿润, 年平均气温为19.3 ℃, 年降水量为 1 346 mm。土壤以滨海盐土、潮土以及水稻土等人工土壤为主。潮汐为正规半日潮。湿地内有维管植物109科294属408种(含变种), 本地优势种主要有短叶茳芏、芦苇(Phragmites australis )和藨草(Scirpus triqueter )。自2002年互花米草入侵以来, 其入侵斑块由零星分布发展到超过200万m2 的大面积分布, 已在潮间带和部分潮上带形成了明显的大面积入侵斑块(郑彩红等, 2006 )。
图1 采样点位置图。
Fig. 1 Location of the sampling site.
选择潮间带生境的互花米草单种群落、互花米草-短叶茳芏混生群落和短叶茳芏单种群落沼泽湿地为研究对象, 对沼泽湿地的土壤和植物进行取样。各群落斑块面积约为30 m × 30 m, 在每个斑块的样方内随机选取4个面积大小为25 cm × 25 cm的小样方作为重复。在2012年4月至2013年2月, 每2个月采样一次, 采用齐地面收割方式获得植物的地上部分, 采用挖掘法获取0-30 cm深度的地下所有根系和土壤。土壤无机氮的测定采用KCl浸提-连续流动分析仪法(李玉英等, 2009 ), 土壤有效磷的测定采用NaHCO3 浸提-钼锑抗比色法① (① Olsen SR, Cole CV, Watanabe FS, Dean LAC (1954). Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate. US Department of Agriculture, Washington, DC, USA. 939.); 植物全氮的测定采用元素分析仪法(江伟等, 2006 ), 植物全磷的测定采用浓硫酸-高氯酸消煮-连续流动分析仪法(鲍士旦, 2005 )。
1.2 样品的采集与处理
在样方内, 齐地面剪下植物地上部分装入自封袋内, 清除土壤表层的残落物和杂质后, 采集0- 30 cm深度的地下所有根系和土壤。所有样品带回实验室后进行清洗, 并对植物器官(根、茎、叶、鞘)进行分类, 由于短叶茳芏叶和鞘在生长过程中已退化, 故不做该分类。待植物器官分类后, 进行烘干, 保留测定。植物地上、地下部分在105 ℃下杀青0.5 h后, 于70 ℃烘箱中烘干至恒质量, 再粉碎过100目孔筛待用。土壤样品于室内自然风干后挑去根系等杂质, 过60目孔筛, 用于后续的测定。
1.3 数据统计分析
1.3.1 内稳性特征的研究原理
根据理论推导与研究, Sterner和Elser (2002) 提出生物生态化学计量内稳性模型。该模型指数公 式为:
y = cx 1 H (1)
其中, x 为土壤无机N或有效P含量(mg·kg-1 )或N:P值, y 为植物体器官对应的全氮(N)或全磷(P)含量(mg·kg-1 )或N:P值,c 为常数, H 为植物内稳性指数。H 使用年数据进行拟合计算得到, 其中植物器官的内稳性为各器官在N、P以及N:P值水平上的拟合, 物种的内稳性为所有器官内稳性的平均值, 生态系统N、P以及N:P值的内稳性为该生态系统所有植物整体在相应N、P和N:P值水平上的拟合。H 的类型可根据H > 4为稳态型、2 < H < 4为弱稳态型、4/3 < H < 2为弱敏感型, 以及H < 4/3为敏感型来界定(Jonas et al ., 2010 )。
1.3.2 数据处理与分析
数据分析前均进行正态性和方差齐性检验。采用Spearman相关分析方法分析土壤N、P含量与植物器官对应N、P含量之间的相关性。方差分析均在数据分析前先进行正态性和方差齐性检验, 若通过, 则进行单因素方差分析, 若通不过, 则将数据进行log10 对数转换, 然后检验转换后的数据是否符合正态性和方差齐性, 若符合, 则对转换后的数据进行方差分析, 若数据转换后仍不符合前提假设, 则采用非参数方法Kruskall-Wallis检验进行数据分析。对N、P含量在各群落沼泽湿地土壤、植物器官间以及物种间的差异进行分析, 分析内稳性指数在不同元素、器官以及物种间的差异。实验数据在Excel 2013统计软件中进行数据整理; 用SPSS 17.0统计软件进行数据分析; 用SigmaPlot 10.0绘图软件进行数据做图。
2 结果和分析
2.1 湿地土壤与植物N、P含量以及N:P值动态
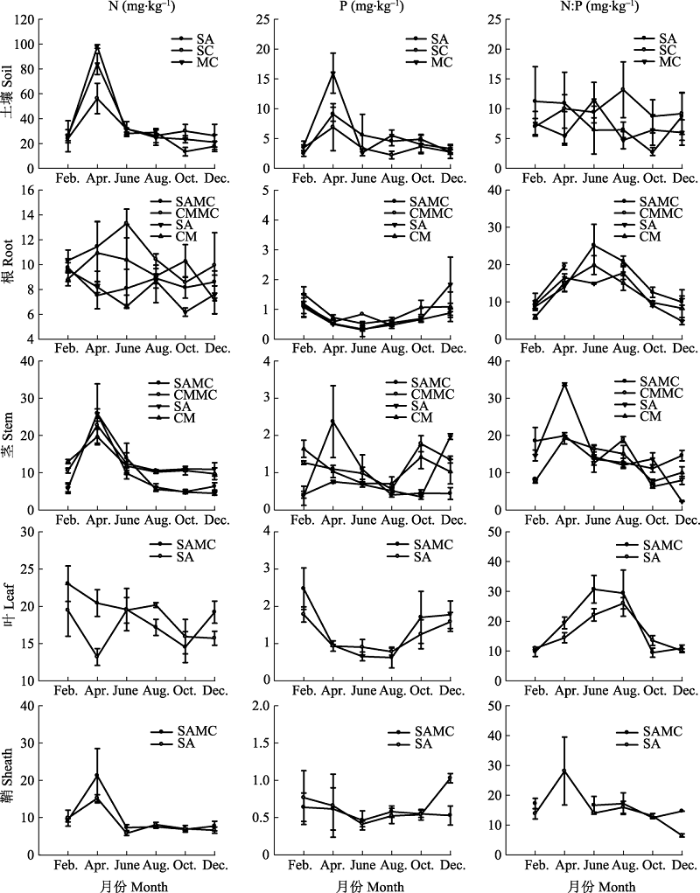
互花米草单种群落、互花米草-短叶茳芏混生群落和短叶茳芏单种群落沼泽湿地土壤N、P含量均在4月份达到峰值, 表现出先增后减的变化趋势, 且N、P含量之间具有显著的相关性(p < 0.05或p < 0.01); 此外, 3种群落土壤N:P值随季节变化的规律不明显(图2 )。互花米草单种群落、短叶茳芏单种群落及其混生群落土壤N含量平均值大小分别为33.91、33.21和31.54 mg·kg-1 (F 2,61 = 0.064, p > 0.05), 土壤P含量平均值大小分别为2.78、3.15和2.39 mg·kg-1 (F 2,62 = 1.131, p > 0.05), 土壤N:P值平均值大小分别为7.68、7.00和9.48, 且混生群落N:P值显著高于短叶茳芏单种群落N:P值(F 2,61 = 2.302, p < 0.05)。
图2 3种群落类型植物器官及土壤的N、P含量和N:P值动态变化(平均值±标准误差, n = 4)。CM, 短叶茳芏单种群落; CMMC, 混生群落中的短叶茳芏; MC, 互花米草-短叶茳芏混生群落; SA, 互花米草单种群落; SAMC, 混生群落中的互花米草。
Fig. 2 Dynamic of N, P and N:P in plant organs and soils across the three community types (mean ± SE, n = 4). CM, Cyperus malaccensis var. brevifolius community; CMMC, C. malaccensis var. brevifolius in mixed community; MC, S. alterniflora-C. malaccensis var. brevifolius mixture community; SA, Spartina alterniflora community; SAMC, S. alterniflora in mixed community.
互花米草和短叶茳芏植物器官中的N、P含量和N:P值对土壤的响应模式不同(图2 )。N元素方面, 单种群落和混生群落中的互花米草叶N含量均显著高于对应单种群落和混生群落中的互花米草根、茎和鞘N含量(F 3,80 = 20.207, p < 0.05), 单种群落中的短叶茳芏茎N含量显著高于对应单种群落中的短叶茳芏根N含量(F 3,78 = 5.062, p < 0.05); 在混生群落中, 短叶茳芏根N含量显著高于对应混生群落中的互花米草根N含量(p < 0.05); 在单种群落中, 短叶茳芏根N含量显著高于对应单种群落中的互花米草根N含量(F 3,78 = 12.168, p < 0.05)。P元素方面, 单种群落和混生群落中的互花米草叶P含量均显著高于对应单种群落和混生群落中的互花米草根、茎和鞘P含量(F 3,81 = 8.867, p < 0.05), 混生群落中的短叶茳芏茎P含量显著高于对应混生群落中的短叶茳芏根P含量(F 3,69 = 3.169, p < 0.05); 在混生群落中, 短叶茳芏茎P含量显著高于对应混生群落中的互花米草茎P含量(p < 0.05); 在单种群落中, 短叶茳芏茎P含量显著高于对应单种群落中的互花米草茎P含量(F 3,74 = 3.265, p < 0.05)。植物器官间的N:P值无显著差异(F 11,227 = 0.593, p > 0.05)。混生群落中的互花米草和短叶茳芏N:P值范围分别为7.54-30.71和6.26- 19.54, 平均值分别为16.17和12.87; 单种群落中的互花米草和短叶茳芏N:P值范围分别为2.31-33.75和5.45-19.86, 平均值分别为14.64和12.19; 所有植物的根、茎、叶和鞘N:P值范围分别为4.84-25.15、2.31-33.75、9.45-30.71和6.49-28.09, 平均值分别为13.43、13.49、17.22和15.76。混生群落中的互花米草叶和鞘N:P值均显著高于单种群落中的互花米草根N:P值和单种群落中的短叶茳芏根N:P值, 显著 高于单种群落和混生群落中的短叶茳芏茎N:P值 (F 11,227 = 2.358, p < 0.05)。
2.2 土壤与植物养分相关性分析
对土壤和植物器官之间养分相关性进行分析的结果(表1 )表明: 单种群落和混生群落中的短叶茳芏茎N含量均与对应的短叶茳芏单种群落和短叶茳芏混生群落的土壤N含量显著正相关(p < 0.05); 单种群落中的互花米草茎N含量与对应互花米草单种群落土壤N含量极显著正相关(p < 0.01), 单种群落中的互花米草鞘N含量与对应互花米草单种群落土壤N含量显著正相关 (p < 0.05); 单种群落中的互花米草根和叶P含量与对应互花米草单种群落土壤P含量显著负相关(p < 0.05); 混生群落中的互花米草茎N和茎P含量均与对应混生群落土壤N和土壤P含量显著正相关(p < 0.05)。
单种群落中的互花米草茎N含量与其土壤N含量的显著相关程度大于对应单种群落中的短叶茳芏茎N含量与其土壤N含量的相关程度; 混生群落中的短叶茳芏茎N含量与其土壤N含量的显著相关程度大于对应混生群落中的互花米草茎N含量与其土壤N含量的相关程度。互花米草入侵后, 单种群落中的互花米草鞘N含量与其土壤N含量间的相关性由显著正相关转变为混生群落中的不显著正相关。同时, 单种群落中的互花米草根P含量与其土壤P含量的相关性由显著负相关转变为混生群落中的不显著正相关; 单种群落中的互花米草茎P含量与其土壤P含量的相关性由不显著正相关转变为混生群落中的显著正相关; 单种群落中的互花米草叶P含量与其土壤P含量的相关性由显著负相关转变为混生群落中的不显著正相关。相比互花米草, 单种群落和混生群落中的短叶茳芏茎N含量与其土壤N含量的相关性均显著, 且混生群落中的相关程度较大;单种群落中的短叶茳芏茎P含量与其土壤P含量的相关性由负相关转变为混生群落中的正相关, 且两者的相关关系均不显著。
2.3 植物化学计量内稳性特征
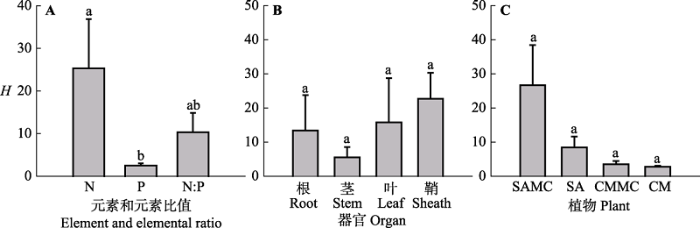
不同元素的内稳性不同。该湿地生态系统N、P和N:P值的内稳性指数(H N 、H P 和H N:P )平均值分别为25.31、2.5和10.33, 其中H N 显著高于H P (F 2,35 = 2.632, p < 0.05)(图3A )。根据内稳性大小的界定, H P 为弱稳态型, H N 和H N:P 为稳态型。对互花米草而言, 混生群落中的互花米草根和叶H N 显著高于对应混生群落中的互花米草根和叶H P 以及根和叶H N:P , 混生群落中的互花米草鞘H N 和鞘H N:P 显著高于对应混生群落中的互花米草鞘H P (p < 0.05); 单种群落中的互花米草茎H N:P 显著高于对应单种群落中的互花米草茎H N 和茎H P , 单种群落中的互花米草鞘H N 显著高于对应单种群落中的互花米草鞘H P (p < 0.05)(图4 )。 对短叶茳芏而言, 无论单种群落还是混生群落, 短叶茳芏H N 、H P 和H N:P 之间均无显著差异(p > 0.05) (图4 )。
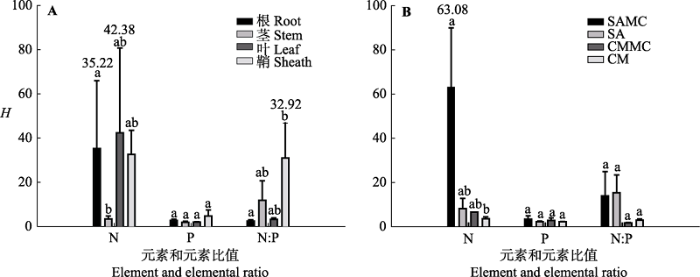
图3 不同元素、器官以及植物类型的内稳性指数(H )(平均值±标准误差, n = 4)。不同小写字母表示不同因子之间差异显著。缩写见图2 。
Fig. 3 The homeostatic index (H ) in different element, organ and plant types (mean ± SE, n = 4). Different lowercase letters denote significant differences among different factors. The abbreviations are the same as in Fig. 2 .
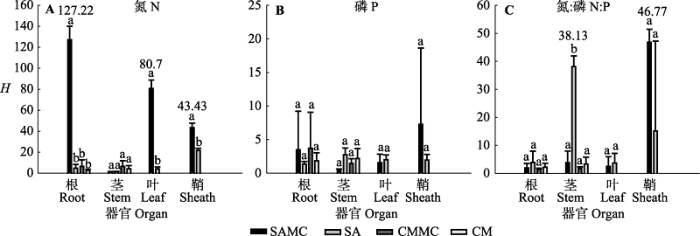
图4 3种群落类型植物器官的N、P和N:P值的内稳性指数(H )(平均值±标准误差, n = 4)。不同小写字母表示不同因子之间差异显著。缩写见图2 。
Fig. 4 The homeostatic index (H ) of N, P and N:P in plant organs across the three community types (mean ± SE, n = 4). Different lowercase letters denote significant differences among different factors. The abbreviations are the same as in Fig. 2 .
植物内稳性的大小在不同器官中存在差异。生态系统根、茎、叶和鞘的内稳性指数平均值分别为13.37、5.55、15.75和22.7 (F 3,35 = 0.614, p > 0.05) (图3B )。对互花米草而言, 对比混生群落中的互花米草器官间的H N 大小发现, 根H N > 叶H N > 鞘H N > 茎H N , 各器官间差异显著(p < 0.05); 混生群落中的互花米草鞘H N:P 显著高于对应混生群落中的互花米草根、茎和叶H N:P ; 单种群落中的互花米草鞘H N 显著高于对应单种群落中的互花米草根、茎和叶H N , 单种群落中的互花米草茎H N:P 显著高于对应单种群落中的互花米草根H N:P 和叶H N:P (图4 )。对短叶茳芏而言, 无论单种群落还是混生群落, 植物器官间的内稳性均无显著差异(p > 0.05)(图4 )。所有植物根、茎、叶和鞘H N 分别为35.22、3.24、42.38和32.56, 且根H N 显著高于茎H N (F 3,11 = 2.566, p < 0.05); 器官间H P 差异不显著(F 3,11 = 1.577, p > 0.05), 鞘H N:P 显著高于根H N:P (F 3,11 = 2.256, p < 0.05)(图5A )。
图5 不同器官和植物类型N、P和N:P值的内稳性指数(H )(平均值±标准误差, n = 4)。不同小写字母表示不同因子之间差异显著。缩写见图2 。
Fig. 5 The homeostatic index (H ) for different organs and plants in N, P and N:P (mean ± SE, n = 4). Different lowercase letters denote significant differences among different factors. The abbreviations are the same as in Fig. 2 .
不同植物类型的内稳性也存在差异。混生群落中的互花米草和短叶茳芏植物内稳性指数平均值分别为26.66和3.48; 单种群落中的互花米草和短叶茳芏植物内稳性指数平均值分别为8.39和2.71 (图3C )。混生群落中的互花米草根H N (127.22)显著高于对应混生群落中的短叶茳芏根H N (6.35)(p < 0.05) (图4A ), 混生群落中的互花米草和短叶茳芏根H P 平均值分别为3.49和3.7 (p > 0.05), 茎H P 平均值分别为0.46和1.45; 单种群落的互花米草和短叶茳芏根H P 平均值分别为1.37和1.85 (p > 0.05), 茎H P 平均值分别为2.74和2.18 (p > 0.05)(图4B ); 单种群落中的互花米草茎H N:P (38.13)显著高于对应单种群落中的短叶茳芏茎H N:P (3.27) (p < 0.05)(图4C )。单种群落和混生群落中的互花米草与对应单种群落和混生群落中的短叶茳芏间的N、P和N:P值的内稳性均无显著差异(p > 0.05), 混生群落中的互花米草 H N 显著高于单种群落中的短叶茳芏H N (p < 0.05)(图5B )。
3 讨论
3.1 植物生长限制性养分分析
根据Garnier理论, 植物与土壤对应养分含量存在正相关关系则说明植物生长受到养分限制(Garnier, 1998 )。根据该理论, 福建省闽江河口鳝鱼滩湿地生态系统中短叶茳芏茎受N限制, 单种群落中的互花米草茎和鞘受N限制, 混生群落中的互花米草茎受N和P共同限制。根据Koerselman理论, 植物体N:P < 14表明其相对受N限制, N:P > 16相对受P限制, 14 < N:P < 16则受N和P共同限制(Koerselman & Meuleman, 1996 ; Güsewell & Koerselman, 2002 )。由于不同研究区域、不同生态系统以及不同植被类型的N:P临界阈值均会发生改变, 精确的N:P临界阈值需要通过严格的施肥试验来确定(Zhang et al ., 2004 ; Güsewell, 2004 )。胡伟芳等(2014)对中国主要湿地649种植物的N、P化学计量特征进行了整合分析, 结果表明不同湿地生境的植物受限制状况不尽相同, 其中N限制(N:P < 14)比例最高, 占59%。针对湿地生态系统的大多数研究也多用14和16为阈值来判断养分受限制情况(Güsewell & Koerselman, 2002 ; Tessier & Raynal, 2003 ; Bott et al ., 2008 )。因此, 若本研究沿用该临界阈值进行分析, 则单种群落和混生群落中的互花米草茎均受N、P共同限制, 其叶均受P限制, 单种群落和混生群落中的短叶茳芏茎均受N限制。本研究中, 互花米草各器官N、P含量平均值均低于短叶茳芏, 但N:P值平均值高于短叶茳芏, 这表明互花米草在生长过程中对养分的需求量相对较低, 但对N的利用效率高于对P的利用效率。
Garnier理论和Koerselman理论是分析植物限制性养分的两种常用方法, Koerselman理论是基于生态化学计量对系统能力平衡与元素平衡相互影响的分析, 已在水生和湿地生态系统中得到广泛应用(Güsewell & Koerselman, 2002 ), 但因受气候特征、物种类型等影响而具有局限性。Garnier理论是特定环境中土壤与植物间相互作用的结果, 是植物需求的内在属性、土壤养分提供以及气候等外因的综 合表现, 具有较强的耦联性, 但结果也受外因影响(樊晓勇, 2012 )。本研究中, Garnier理论和Koerselman理论均表明单种群落和混生群落中的短叶茳芏茎的生长均受N限制, 混生群落中的互花米草茎的生长受N和P共同限制, 其他群落中的部分植物器官生长的受限结果存在差异。因此, 本研究同时采用这两种方法进行养分限制性分析, 由两种方法一致得到的结果以及由此获得的结论, 我们认为更可靠。
3.2 入侵对植物内稳性的影响
入侵对植物内稳性产生的影响主要表现在N元素上。混生群落中的互花米草根、叶和鞘的H N 分别显著高于对应单种群落中的互花米草根、茎和鞘的H N 。混生群落中的互花米草根的H N 比单种群落中的互花米草根的H N 高25.7倍, 混生群落中的短叶茳芏根的H N 比单种群落中的短叶茳芏根的H N 增大了1.48倍, 两者之间的差异可能与互花米草根际微生物的固氮作用有关, 这使得互花米草获得更多的N, 进而缓解了N的限制(Bagwell & Lovell, 2000 ; Moisander et al ., 2005 ; 曾艳等, 2011 )。入侵初期, 单种群落的植物生长受N限制, 植物体内N的含量很大程度上取决于土壤中可利用N的含量, 故植物内稳性较低。随着入侵过程的进行, 互花米草的根际固氮作用使植物体内N含量的获取对土壤中N的依赖性降低, 从而使混生群落中的植物表现出较高的内稳性。相比短叶茳芏根的养分利用性, 互花米草根内稳性的调节能力使其能够在多变的环境中保持较好的生长, 进而具备更强的竞争力和适应力。已有的研究表明, 生态系统的稳定性与根生物量正相关, 根在变化的环境中可通过回收利用和贮存养分来调节生长, 从而提高物种和群落水平的生产力和稳定性(Tilman et al ., 2006 )。因此, 根内稳性与群落生产力和稳定性的关系研究值得进一步探索。
此外, 在入侵过程中, 混生群落中的互花米草根的H N 和根的H P 较单种群落中的互花米草根的H N 和根的H P 增大, 茎的H N 和茎的H P 相应减小; 根的H N:P 减小, 鞘的H N:P 相应增大。这种地上-地下内稳性变化趋势相反的现象正是植物为适应特定环境而表现出的调节作用。如庾强(2009)研究表明, 贫瘠的典型草原中, 高内稳性的根对应低内稳性的叶, 这是为保持叶片元素组成的稳定, 根的元素组成做出了较大程度的调节, 从而适应该特定环境。
3.3 植物内稳性与生态功能
Yu等(2015)研究发现, 物种水平的内稳性与物种优势度和稳定性正相关, 而且在多数情况下, 群落水平的内稳性与生态系统功能以及稳定性也正相关。因此, 内稳性可作为衡量物种竞争力的重要指标。本研究结果表明, 入侵使得混生群落中的互花米草根、叶和鞘的H N 均显著高于单种群落中的互花米草根、茎和鞘的H N , 但由于内稳性是植物在长期进化过程中适应环境变化的结果(Elser et al ., 2010 ), 是植物所固有的特性。因此, 无论单种群落还是混生群落中, 混生群落中的互花米草根的H N 显著高于对应混生群落中的短叶茳芏根的H N (p < 0.05), 单种群落中的互花米草茎的H N:P 显著高于对应单种群落中的短叶茳芏茎的H N:P (p < 0.05), 没有发现短叶茳芏显著高于互花米草的研究结果。因此, 入侵过程对植物内稳性的改变不影响物种间内稳性的相对大小, 互花米草较高的内稳性可能是其入侵成功的一个原因。
目前, 互花米草的防治措施主要有物理控制(Frid et al. , 1999 )、化学控制(Patten, 2002 )和生物控制(Wu et al ., 1999 , Grevstad et al ., 2003 )这3种方法, 其中以收割、碎根水淹等物理方法居多。研究表明, 翻耕和碎根处理方法仅在当年有一定的控制效果, 不具有长期有效性, 且割除碎根也需要在最佳季节进行(李贺鹏和张利权, 2007 )。已有的研究认为: 闽江河口湿地生态系统在7月份时互花米草内稳性最低, 是治理互花米草的最佳时期(蒋利玲等, 2014 )。充分考虑植物内稳性特征的影响, 建议采用物理-化学综合方法进行互花米草的控制, 在7月份对互花米草进行碎根, 再通过施加N缓解短叶茳芏根和茎的养分限制, 从而增大短叶茳芏群落在湿地生态系统中的内稳性。
4 结论
1)互花米草入侵闽江河口湿地生态系统后, 其生长由受N和P共同限制转变为受P限制, 短叶茳芏均受N限制。
2)该湿地生态系统中, N的内稳性显著高于P的内稳性; 根的H N 显著高于茎的H N , 鞘的H N:P 显著高于根的H N:P 。
3)互花米草物种内稳性高于短叶茳芏物种内稳性, 这可能是互花米草成功入侵的一个原因。
致谢 感谢福建师范大学翁少峰、颜远烽、吴丽凤、何诗、叶小霖、谢志煌、蒋米亮和柯梦凡等同学在野外采样与室内分析工作中给予的帮助。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Andersen T Hessen DO 1991 ). Carbon, nitrogen, and phosphorus content of freshwater zooplankton
.Limnology and Oceanography 36 , 807 -814 .
[本文引用: 1]
[2]
Bagwell CE Lovell CR 2000 ). Microdiversity of culturable diazotrophs from the rhizoplanes of the salt marsh grasses Spartina alterniflora and Juncus roemerianus
. Microbial Ecology 39 , 128 -136 .
[本文引用: 1]
[3]
Bao SD 2005 ). Agricultural Soil Analysis. 3rd edn . China Agricultural Press , Beijing . 141 -149 . (in Chinese)[鲍士旦 (2005 ). 土壤农化分析. 第三版 . 中国农业出版社 , 北京 . 141 -149 .]
[本文引用: 1]
[4]
Bott T Meyer GA Young EB 2008 ). Nutrient limitation and morphological plasticity of the carnivorous pitcher plant Sarracenia purpurea in contrasting wetland environments
. New Phytologist , 180 , 631 -641 .
[本文引用: 1]
[5]
Cernusak LA Winter K Turner BL 2009 ). Leaf nitrogen to phosphorus ratios of tropical trees: Experimental assessment of physiological and environmental controls
.New Phytologist 185 , 770 -779 .
[本文引用: 1]
[6]
Davis MA 2003 ). Biotic globalization: Does competition from introduced species threaten biodiversity?
Bioscience 53 , 481 -489 .
[本文引用: 1]
[7]
Elser JJ Fagan WF Kerkhoff AJ Swenson NG Enquist BJ 2010 ). Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change
.New Phytologist 186 , 593 -608 .
[本文引用: 1]
[8]
Fan XY 2012 ). Spatial Variation in Nutrient of Dominant Plant and Ecological Stoichiometry from Laohu Gou of Qilian Mountain
. Master degree dissertation, Lanzhou University, Lanzhou . (in Chinese with English abstract)[樊晓勇 (2012 ). 祁连山老虎沟优势植物的养分空间变化与生态化学计量学研究
. 硕士学位论文, 兰州大学, 兰州 .]
[本文引用: 1]
[9]
Frid CLJ Chandrasekara WU Davey P 1999 ). The restoration of mud flats invaded by common cord-grass (Spartina anglica, CE Hubbard) using mechanical disturbance and its effects on the macrobenthic fauna
. Aquatic Conservation Marine & Freshwater Ecosystems 9 , 47 -61 .
[本文引用: 1]
[10]
Garnier E 1998 ). Interspecific Variation in Plasticity of Grasses in Response to Nitrogen Supply
. Cambridge University Press , Cambridge, UK . 155 -181 .
[本文引用: 1]
[11]
Goldman JC Caron DA Dennett MR 1987 ). Regulations of gross growth efficiency and ammonium regeneration in bacteria by substrate C:N ratio
.Limnology and Oceanography 32 , 1239 -1252 .
[本文引用: 1]
[12]
Grevstad FS Strong DR Garcia-Rossi D Switzer RW Wecker MS 2003 ). Biological control of Spartina alterniflora in Willapa Bay, Washington using the planthopper Prokelisia marginata: Agent specificity and early results
. Biological Control 27 , 32 -42 .
[本文引用: 1]
[13]
Güsewell S 2004 ). N:P ratios in terrestrial plants: Variation and functional significance
.New Phytologist 164 , 243 -266 .
[本文引用: 2]
[14]
Güsewell S Jewell PL Edwards PJ 2005 ). Effects of heterogeneous habitat use by cattle on nutrient availability and litter decomposition in soils of an Alpine pasture
.Plant and Soil 268 , 135 -149 .
[本文引用: 1]
[15]
Güsewell S Koerselman W 2002 ). Variation in nitrogen and phosphorus concentrations of wetland plants
.Perspectives in Plant Ecology Evolution and Systematics 5 , 37 -61 .
[本文引用: 3]
[16]
Hooper DU Chapin FS Ewel JJ Hector A Inchausti P Lavorel S Lawton JH Lodge DM Loreau M Naeem S Schmid B Setälä H Symstad AJ Vandermeer J Wardle DA 2005 ). Effects of biodiversity on ecosystem functioning: A consensus of current knowledge
.Ecological Monographs 75 , 3 -35 .
[本文引用: 1]
[17]
Hu WF Zhang WL Zhang LH Chen XY Lin W Zeng CS Tong C 2014 ). Stoichiometric characteristics of nitrogen and phosphorus in major wetland vegetation of China
. Chinese Journal of Plant Ecology 38 , 1041 -1052 . (in Chinese with English abstract)[胡伟芳 , 章文龙 , 张林海 , 陈晓艳 , 林伟 , 曾从盛 , 仝川 (2014 ). 中国主要湿地植被氮和磷生态化学计量学特征
. 植物生态学报 , 38 , 1041 -1052 .]
[18]
Jiang LL He S Wu LF Yan YF Weng SF Liu J Wang WQ Zeng CS 2014 ). Characteristics of stoichiometric homeostasis of three plant species in wetlands in Minjiang estuary
.Wetland Science 3 , 293 -298 . (in Chinese with English abstract)[蒋利玲 , 何诗 , 吴丽凤 , 颜远烽 , 翁少峰 , 刘静 , 王维奇 , 曾从盛 (2014 ). 闽江河口湿地3种植物化学计量内稳性特征
. 湿地科学 , 3 , 293 -298 .]
[本文引用: 2]
[19]
Jiang W Li XQ Jiang Q Huang DK Cheng HG 2006 ). Kjeldahl method and the elemental analyzer method in measurement of total nitrogen in sediments: Comparison and its significance
.Geochimica 35 , 221 -226 . (in Chinese with English abstract)[江伟 , 李心清 , 蒋倩 , 黄代宽 , 程红光 (2006 ). 凯氏蒸馏法和元素分析仪法测定沉积物中全氮含量的异同及其意义
. 地球化学 , 35 , 221 -226 .]
[本文引用: 1]
[20]
Jonas P Patrick F Akira G James MH Jayne J Satoshi K 2010 ). To be or not to be what you eat: Regulation of stoichiometric homeostasis among autotrophs and heterotrophs
.Oikos 119 , 741 -751 .
[本文引用: 1]
[21]
Koerselman W Meuleman AMF 1996 ). The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation
.Journal of Applied Ecology 33 , 1441 -1450 .
[本文引用: 1]
[22]
Kooijman SALM 1995 ). The stoichiometry of animal energetics
.Journal of Theoretical Biology 177 , 139 -149 .
[本文引用: 1]
[23]
Levi MP Cowling EB 1969 ). Role of nitrogen in wood deterioration. VII. Physiological adaptation of wood-destroying and other fungi to substrates deficient in nitrogen
.Phytopathology 59 , 460 -468 .
[本文引用: 1]
[24]
Li HP Zhang LQ 2007 ). Experimental study on physical controls of an exotic plant Spartina alterniflora in Shanghai
. Journal of East China Normal University (Natural Science ) , 6 , 44 -55 . (in Chinese with English abstract)[李贺鹏 , 张利权 (2007 ). 外来植物互花米草的物理控制实验研究
. 华东师范大学学报(自然科学版) , 6 , 44 -55 .]
[本文引用: 1]
[25]
Li YY Sun JH Yu CB Cheng X Zhang FS Li L 2009 ). Effects of nitrogen fertilization application and faba bean/maize intercropping on the spatial and temporal distribution of soil inorganic nitrogen
.Plant Nutrition and Fertilizer Science 15 , 815 -823 . (in Chinese with English abstract)[李玉英 , 孙建好 , 余常兵 , 程序 , 张福锁 , 李隆 (2009 ). 施氮量和蚕豆/玉米间作对土壤无机氮时空分布的影响
. 植物营养与肥料学报 , 15 , 815 -823 .]
[本文引用: 1]
[26]
Liu JQ Zeng CS Chen N 2006 ). Study on the Wetlands of Minjiang River Estuary . Science Press , Beijing . 21 -47 . (in Chinese)[刘剑秋 , 曾从盛 , 陈宁 (2006 ). 闽江河口湿地研究 . 科学出版社 , 北京 . 21 -47 .]
[本文引用: 1]
[27]
Matzek V Vitousek PM 2009 ). N:P stoichiometry and protein: RNA ratios in vascular plants: An evaluation of the growth-rate hypothesis
.Ecology Letters 12 , 765 -771 .
[本文引用: 1]
[28]
Moisander PH Piehler MF Paerl HW 2005 ). Diversity and activity of epiphytic nitrogen fixers on standing dead stems of the salt marsh grass Spartina alterniflora
. Aquatic Microbial Ecology , 39 , 271 -279 .
[本文引用: 1]
[29]
Patten K 2002 ). Smooth cordgrass (Spartina alterniflora ) control with Imazapyr
. Weed Technology , 16 , 826 -832 .
[本文引用: 1]
[30]
Rhee GY 1978 ). Effects of N:P atomic ratios and nitrate limitation on algal growth, cell composition and nitrate uptake
.Limnology and Oceanography 23 , 10 -25 .
[本文引用: 1]
[31]
Sterner RW Elser JJ 2002 ). Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere . Princeton University Press , New Jersey . 226 -226 .
[本文引用: 2]
[32]
Su Q 2012 ). The framework of stoichiometry homeostasis in zooplankton elemental composition
.Acta Ecologica Sinica 32 , 7213 -7219 . (in Chinese with English abstract)[苏强 (2012 ). 浮游动物化学计量学稳态性特征研究进展
. 生态学报 , 32 , 7213 -7219 .]
[本文引用: 1]
[33]
Tessier JT Raynal DJ 2003 ). Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation
.Journal of Applied Ecology 40 , 523 -534 .
[本文引用: 1]
[34]
Tilman D Reich PB Knops JMH 2006 ). Biodiversity and ecosystem stability in a decade-long grassland experiment
.Nature 441 , 629 -632 .
[本文引用: 1]
[35]
Wang H Li HY Zhang ZJ Muehlbauer JD He Q Xu XH Yue CL Jiang DQ 2014 ). Linking stoichiometric homeostasis of microorganisms with soil phosphorus dynamics in wetlands subjected to microcosm warming
.PLOS ONE 9 , e85575 -e85575 . doi: 10.1371/journal.pone.0085575 .
[本文引用: 1]
[36]
Wu MY Hacker S Ayres D Strong DR 1999 ). Potential of Prokelisia spp. as biological control agents of english cordgrass, Spartina anglica
. Biological Control , 16 , 267 -273 .
[本文引用: 1]
[37]
Xiong HF Wang YH 2005 ). Advances in researches on biogeochemical circulation of C, N and P in wetlands
.Chinese Journal of Soil Science 36 , 240 -243 . (in Chinese with English abstract)[熊汉锋 , 王运华 (2005 ). 湿地碳氮磷的生物地球化学循环研究进展
. 土壤通报 , 36 , 240 -243 .]
[本文引用: 1]
[38]
Yan ZB Kim NY Han TS Fang JY Han WX 2013 ). Effects of nitrogen and phosphorus fertilization on leaf carbon, nitrogen and phosphorus stoichiometry of Arabidopsis thaliana
. Chinese Journal of Plant Ecology , 37 , 551 -557 . (in Chinese with English abstract)[严正兵 , 金南瑛 , 韩廷申 , 方精云 , 韩文轩 (2013 ). 氮磷施肥对拟南芥叶片碳氮磷化学计量特征的影响
. 植物生态学报 , 37 , 551 -557 .]
[本文引用: 1]
[39]
Yu Q 2009 ). Ecological Stoichiometric Study on Vascular Plants in the Inner Mongolia Steppe
. PhD dissertation, Institute of Botany,Chinese Academy of Science , Beijing . 56 -67 . (in Chinese with English abstract)[庾强 (2009 ). 内蒙古草原植物化学计量生态学研究
. 博士学位论文, 中国科学院植物研究所, 北京 . 56 -67 .]
[本文引用: 1]
[40]
Yu Q Chen QS Elser JJ He NP Wu HH Zhang GM Wu JG Bai YF Han XG 2010 ). Linking stoichiometric homeostasis with ecosystem structure, functioning, and stability
.Ecology Letters 13 , 1390 -1399 .
[本文引用: 1]
[41]
Yu Q Wilcox K Pierre KL Knapp AK Han XG Smith MD 2015 ). Stoichiometric homeostasis predicts plant species dominance, temporal stability, and responses to global change
.Ecology 96 , 2328 -2335 .
[42]
Zeng Y Tian GH Chen LY Li J An D Lei ZS Tang H Peng SL 2011 ). Influence ofSpartina alterniflora invasion on soil ecosystem: A review
. Chinese Journal of Ecology , 30 , 2080 -2087 . (in Chinese with English abstract)[曾艳 , 田广红 , 陈蕾伊 , 李静 , 安东 , 雷振胜 , 唐虹 , 彭少麟 (2011 ). 互花米草入侵对土壤生态系统的影响
. 生态学杂志 , 30 , 2080 -2087 .]
[本文引用: 1]
[43]
Zhang LX Bai YF Han XG 2004 ). Differential responses of N:P stoichiometry of Leymus chinensis and Carex korshinskyi to N additions in a steppe ecosystem in Nei Mongol
. Acta Botanica Sinica , 46 , 259 -270 .
[本文引用: 1]
[44]
Zhang RY Shi XM Li WJ Guo R Wang G 2015 ). Response of species homeostasis and biomass on a sub-alpine grassland
.Pratacultural Science 32 , 1539 -1547 . (in Chinese with English abstract)[张仁懿 , 史小明 , 李文金 , 郭睿 , 王刚 (2015 ). 亚高寒草甸物种内稳性与生物量变化模式
. 草业科学 , 32 , 1539 -1547 .]
[本文引用: 1]
[45]
Zheng CH Zeng CS Chen ZQ Lin MC 2006 ). A study on the changes of landscape pattern of estuary wetlands of the Minjiang River
.Wetland Science 4 , 29 -35 . (in Chinese with English abstract)[郑彩红 , 曾从盛 , 陈志强 , 林茂昌 (2006 ). 闽江河口区湿地景观格局演变研究
. 湿地科学 , 4 , 29 -35 .]
[本文引用: 1]
Carbon, nitrogen, and phosphorus content of freshwater zooplankton
1
1991
... 国外内稳性研究的对象由最初的真菌和细菌(Levi & Cowling, 1969 ; Goldman et al ., 1987 )发展到藻类、浮游动物和草本植物(Rhee, 1978 ; Andersen & Hessen, 1991 ; Güsewell et al ., 2005 ).由于高等植物体内元素的内稳性和化学计量特征复杂, 对高等植物的研究甚少(Cernusak et al ., 2009 ; Matzek & Vitousek, 2009 ; Yu et al ., 2010 ).生态系统水平上对内稳性的研究多集中于海洋生态系统和陆地生态系统(Güsewell, 2004 ; 苏强, 2012 ), 对湿地生态系统的研究较少(Wang et al ., 2014 ).国内内稳性的研究还处于起步阶段.庾强(2009)以内蒙古草原为研究对象, 通过1 200 km的横断范围的样带试验和内蒙古草原生态系统定位研究站27年的监测数据发现, 内稳性高的物种具有较高的优势度和生物量稳定性, 内稳性高的生态系统则具有较高的生产力和稳定性.因此, 内稳性是衡量物种竞争力的一个重要指标.已有研究表明内稳性的高低受多种因子的影响, 其中包括植物器官(庾强, 2009 )、养分供应(严正兵等, 2013 )、光强以及施肥(Sterner & Elser, 2002 )等, 影响因子的不确定性使得不同研究中内稳性对环境的响应不一致.目前有关互花米草入侵对本地生态系统的影响多集中在物质循环以及生态化学计量学, 而物种水平上内稳性的研究甚少(张仁懿等, 2015 ).已有研究发现, 闽江河口湿地生长季阶段的入侵种互花米草和本地种短叶茳芏均具有一定的内稳性, 互花米草N和N:P的内稳性高于短叶茳芏N和N:P的内稳性, 而短叶茳芏P的内稳性高于互花米草P的内稳性(蒋利玲等, 2014 ). ...
Microdiversity of culturable diazotrophs from the rhizoplanes of the salt marsh grasses Spartina alterniflora and Juncus roemerianus
1
2000
... 入侵对植物内稳性产生的影响主要表现在N元素上.混生群落中的互花米草根、叶和鞘的H N 分别显著高于对应单种群落中的互花米草根、茎和鞘的H N .混生群落中的互花米草根的H N 比单种群落中的互花米草根的H N 高25.7倍, 混生群落中的短叶茳芏根的H N 比单种群落中的短叶茳芏根的H N 增大了1.48倍, 两者之间的差异可能与互花米草根际微生物的固氮作用有关, 这使得互花米草获得更多的N, 进而缓解了N的限制(Bagwell & Lovell, 2000 ; Moisander et al ., 2005 ; 曾艳等, 2011 ).入侵初期, 单种群落的植物生长受N限制, 植物体内N的含量很大程度上取决于土壤中可利用N的含量, 故植物内稳性较低.随着入侵过程的进行, 互花米草的根际固氮作用使植物体内N含量的获取对土壤中N的依赖性降低, 从而使混生群落中的植物表现出较高的内稳性.相比短叶茳芏根的养分利用性, 互花米草根内稳性的调节能力使其能够在多变的环境中保持较好的生长, 进而具备更强的竞争力和适应力.已有的研究表明, 生态系统的稳定性与根生物量正相关, 根在变化的环境中可通过回收利用和贮存养分来调节生长, 从而提高物种和群落水平的生产力和稳定性(Tilman et al ., 2006 ).因此, 根内稳性与群落生产力和稳定性的关系研究值得进一步探索. ...
1
2005
... 选择潮间带生境的互花米草单种群落、互花米草-短叶茳芏混生群落和短叶茳芏单种群落沼泽湿地为研究对象, 对沼泽湿地的土壤和植物进行取样.各群落斑块面积约为30 m × 30 m, 在每个斑块的样方内随机选取4个面积大小为25 cm × 25 cm的小样方作为重复.在2012年4月至2013年2月, 每2个月采样一次, 采用齐地面收割方式获得植物的地上部分, 采用挖掘法获取0-30 cm深度的地下所有根系和土壤.土壤无机氮的测定采用KCl浸提-连续流动分析仪法(李玉英等, 2009 ), 土壤有效磷的测定采用NaHCO3 浸提-钼锑抗比色法① (① Olsen SR, Cole CV, Watanabe FS, Dean LAC (1954). Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate. US Department of Agriculture, Washington, DC, USA. 939.); 植物全氮的测定采用元素分析仪法(江伟等, 2006 ), 植物全磷的测定采用浓硫酸-高氯酸消煮-连续流动分析仪法(鲍士旦, 2005 ). ...
Nutrient limitation and morphological plasticity of the carnivorous pitcher plant Sarracenia purpurea in contrasting wetland environments
1
2008
... 根据Garnier理论, 植物与土壤对应养分含量存在正相关关系则说明植物生长受到养分限制(Garnier, 1998 ).根据该理论, 福建省闽江河口鳝鱼滩湿地生态系统中短叶茳芏茎受N限制, 单种群落中的互花米草茎和鞘受N限制, 混生群落中的互花米草茎受N和P共同限制.根据Koerselman理论, 植物体N:P < 14表明其相对受N限制, N:P > 16相对受P限制, 14 < N:P < 16则受N和P共同限制(Koerselman & Meuleman, 1996 ; Güsewell & Koerselman, 2002 ).由于不同研究区域、不同生态系统以及不同植被类型的N:P临界阈值均会发生改变, 精确的N:P临界阈值需要通过严格的施肥试验来确定(Zhang et al ., 2004 ; Güsewell, 2004 ).胡伟芳等(2014)对中国主要湿地649种植物的N、P化学计量特征进行了整合分析, 结果表明不同湿地生境的植物受限制状况不尽相同, 其中N限制(N:P < 14)比例最高, 占59%.针对湿地生态系统的大多数研究也多用14和16为阈值来判断养分受限制情况(Güsewell & Koerselman, 2002 ; Tessier & Raynal, 2003 ; Bott et al ., 2008 ).因此, 若本研究沿用该临界阈值进行分析, 则单种群落和混生群落中的互花米草茎均受N、P共同限制, 其叶均受P限制, 单种群落和混生群落中的短叶茳芏茎均受N限制.本研究中, 互花米草各器官N、P含量平均值均低于短叶茳芏, 但N:P值平均值高于短叶茳芏, 这表明互花米草在生长过程中对养分的需求量相对较低, 但对N的利用效率高于对P的利用效率. ...
Leaf nitrogen to phosphorus ratios of tropical trees: Experimental assessment of physiological and environmental controls
1
2009
... 国外内稳性研究的对象由最初的真菌和细菌(Levi & Cowling, 1969 ; Goldman et al ., 1987 )发展到藻类、浮游动物和草本植物(Rhee, 1978 ; Andersen & Hessen, 1991 ; Güsewell et al ., 2005 ).由于高等植物体内元素的内稳性和化学计量特征复杂, 对高等植物的研究甚少(Cernusak et al ., 2009 ; Matzek & Vitousek, 2009 ; Yu et al ., 2010 ).生态系统水平上对内稳性的研究多集中于海洋生态系统和陆地生态系统(Güsewell, 2004 ; 苏强, 2012 ), 对湿地生态系统的研究较少(Wang et al ., 2014 ).国内内稳性的研究还处于起步阶段.庾强(2009)以内蒙古草原为研究对象, 通过1 200 km的横断范围的样带试验和内蒙古草原生态系统定位研究站27年的监测数据发现, 内稳性高的物种具有较高的优势度和生物量稳定性, 内稳性高的生态系统则具有较高的生产力和稳定性.因此, 内稳性是衡量物种竞争力的一个重要指标.已有研究表明内稳性的高低受多种因子的影响, 其中包括植物器官(庾强, 2009 )、养分供应(严正兵等, 2013 )、光强以及施肥(Sterner & Elser, 2002 )等, 影响因子的不确定性使得不同研究中内稳性对环境的响应不一致.目前有关互花米草入侵对本地生态系统的影响多集中在物质循环以及生态化学计量学, 而物种水平上内稳性的研究甚少(张仁懿等, 2015 ).已有研究发现, 闽江河口湿地生长季阶段的入侵种互花米草和本地种短叶茳芏均具有一定的内稳性, 互花米草N和N:P的内稳性高于短叶茳芏N和N:P的内稳性, 而短叶茳芏P的内稳性高于互花米草P的内稳性(蒋利玲等, 2014 ). ...
Biotic globalization: Does competition from introduced species threaten biodiversity?
1
2003
... 生物入侵作为全球变化的一个重要方面, 严重威胁了土著生物群落的生物多样性和生态系统功能(Davis, 2003 ; Hooper et al ., 2005 ).作为我国中亚热带和南亚热带过渡区最具代表性的河口湿地之一的闽江河口湿地(刘剑秋等, 2006 ), 自2002年以来逐渐遭到互花米草(Spartina alterniflora )的大规模入侵.互花米草以其极强的适应力和繁殖力迅速占据空生态位, 侵占本地种短叶茳芏(Cyperus malaccensis var. brevifolius )的生境, 在潮间带形成了短叶茳芏单种群落、互花米草单种群落和互花米草-短叶茳芏混生群落共存的竞争格局.入侵的互花米草通过影响土著生态系统的结构, 进而影响生态系统的养分循环, 生态系统养分的可利用性改变反过来又影响植物的生长状况, 从而可能改变入侵种与土著种之间的竞争关系.养分元素氮(N)和磷(P)是湿地生态系统植物生长的主要限制资源, 其可利用性及生态化学计量关系是植物生长和生态系统物质循环的重要调节因子(熊汉锋和王运华, 2005 ).植物能在外界养分元素组成改变的情况下, 一定程度上维持自身各元素之间的相对比例.这种由生物在长期进化过程中为适应外界环境变化而形成的维持自身化学组成相对稳定的能力就叫作生态化学计量内稳性(简称内稳性)(Kooijman, 1995 ). ...
Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change
1
2010
... Yu等(2015)研究发现, 物种水平的内稳性与物种优势度和稳定性正相关, 而且在多数情况下, 群落水平的内稳性与生态系统功能以及稳定性也正相关.因此, 内稳性可作为衡量物种竞争力的重要指标.本研究结果表明, 入侵使得混生群落中的互花米草根、叶和鞘的H N 均显著高于单种群落中的互花米草根、茎和鞘的H N , 但由于内稳性是植物在长期进化过程中适应环境变化的结果(Elser et al ., 2010 ), 是植物所固有的特性.因此, 无论单种群落还是混生群落中, 混生群落中的互花米草根的H N 显著高于对应混生群落中的短叶茳芏根的H N (p < 0.05), 单种群落中的互花米草茎的H N:P 显著高于对应单种群落中的短叶茳芏茎的H N:P (p < 0.05), 没有发现短叶茳芏显著高于互花米草的研究结果.因此, 入侵过程对植物内稳性的改变不影响物种间内稳性的相对大小, 互花米草较高的内稳性可能是其入侵成功的一个原因. ...
祁连山老虎沟优势植物的养分空间变化与生态化学计量学研究
1
2012
... Garnier理论和Koerselman理论是分析植物限制性养分的两种常用方法, Koerselman理论是基于生态化学计量对系统能力平衡与元素平衡相互影响的分析, 已在水生和湿地生态系统中得到广泛应用(Güsewell & Koerselman, 2002 ), 但因受气候特征、物种类型等影响而具有局限性.Garnier理论是特定环境中土壤与植物间相互作用的结果, 是植物需求的内在属性、土壤养分提供以及气候等外因的综 合表现, 具有较强的耦联性, 但结果也受外因影响(樊晓勇, 2012 ).本研究中, Garnier理论和Koerselman理论均表明单种群落和混生群落中的短叶茳芏茎的生长均受N限制, 混生群落中的互花米草茎的生长受N和P共同限制, 其他群落中的部分植物器官生长的受限结果存在差异.因此, 本研究同时采用这两种方法进行养分限制性分析, 由两种方法一致得到的结果以及由此获得的结论, 我们认为更可靠. ...
The restoration of mud flats invaded by common cord-grass (Spartina anglica, CE Hubbard) using mechanical disturbance and its effects on the macrobenthic fauna
1
1999
... 目前, 互花米草的防治措施主要有物理控制(Frid et al. , 1999 )、化学控制(Patten, 2002 )和生物控制(Wu et al ., 1999 , Grevstad et al ., 2003 )这3种方法, 其中以收割、碎根水淹等物理方法居多.研究表明, 翻耕和碎根处理方法仅在当年有一定的控制效果, 不具有长期有效性, 且割除碎根也需要在最佳季节进行(李贺鹏和张利权, 2007 ).已有的研究认为: 闽江河口湿地生态系统在7月份时互花米草内稳性最低, 是治理互花米草的最佳时期(蒋利玲等, 2014 ).充分考虑植物内稳性特征的影响, 建议采用物理-化学综合方法进行互花米草的控制, 在7月份对互花米草进行碎根, 再通过施加N缓解短叶茳芏根和茎的养分限制, 从而增大短叶茳芏群落在湿地生态系统中的内稳性. ...
Interspecific Variation in Plasticity of Grasses in Response to Nitrogen Supply
1
1998
... 根据Garnier理论, 植物与土壤对应养分含量存在正相关关系则说明植物生长受到养分限制(Garnier, 1998 ).根据该理论, 福建省闽江河口鳝鱼滩湿地生态系统中短叶茳芏茎受N限制, 单种群落中的互花米草茎和鞘受N限制, 混生群落中的互花米草茎受N和P共同限制.根据Koerselman理论, 植物体N:P < 14表明其相对受N限制, N:P > 16相对受P限制, 14 < N:P < 16则受N和P共同限制(Koerselman & Meuleman, 1996 ; Güsewell & Koerselman, 2002 ).由于不同研究区域、不同生态系统以及不同植被类型的N:P临界阈值均会发生改变, 精确的N:P临界阈值需要通过严格的施肥试验来确定(Zhang et al ., 2004 ; Güsewell, 2004 ).胡伟芳等(2014)对中国主要湿地649种植物的N、P化学计量特征进行了整合分析, 结果表明不同湿地生境的植物受限制状况不尽相同, 其中N限制(N:P < 14)比例最高, 占59%.针对湿地生态系统的大多数研究也多用14和16为阈值来判断养分受限制情况(Güsewell & Koerselman, 2002 ; Tessier & Raynal, 2003 ; Bott et al ., 2008 ).因此, 若本研究沿用该临界阈值进行分析, 则单种群落和混生群落中的互花米草茎均受N、P共同限制, 其叶均受P限制, 单种群落和混生群落中的短叶茳芏茎均受N限制.本研究中, 互花米草各器官N、P含量平均值均低于短叶茳芏, 但N:P值平均值高于短叶茳芏, 这表明互花米草在生长过程中对养分的需求量相对较低, 但对N的利用效率高于对P的利用效率. ...
Regulations of gross growth efficiency and ammonium regeneration in bacteria by substrate C:N ratio
1
1987
... 国外内稳性研究的对象由最初的真菌和细菌(Levi & Cowling, 1969 ; Goldman et al ., 1987 )发展到藻类、浮游动物和草本植物(Rhee, 1978 ; Andersen & Hessen, 1991 ; Güsewell et al ., 2005 ).由于高等植物体内元素的内稳性和化学计量特征复杂, 对高等植物的研究甚少(Cernusak et al ., 2009 ; Matzek & Vitousek, 2009 ; Yu et al ., 2010 ).生态系统水平上对内稳性的研究多集中于海洋生态系统和陆地生态系统(Güsewell, 2004 ; 苏强, 2012 ), 对湿地生态系统的研究较少(Wang et al ., 2014 ).国内内稳性的研究还处于起步阶段.庾强(2009)以内蒙古草原为研究对象, 通过1 200 km的横断范围的样带试验和内蒙古草原生态系统定位研究站27年的监测数据发现, 内稳性高的物种具有较高的优势度和生物量稳定性, 内稳性高的生态系统则具有较高的生产力和稳定性.因此, 内稳性是衡量物种竞争力的一个重要指标.已有研究表明内稳性的高低受多种因子的影响, 其中包括植物器官(庾强, 2009 )、养分供应(严正兵等, 2013 )、光强以及施肥(Sterner & Elser, 2002 )等, 影响因子的不确定性使得不同研究中内稳性对环境的响应不一致.目前有关互花米草入侵对本地生态系统的影响多集中在物质循环以及生态化学计量学, 而物种水平上内稳性的研究甚少(张仁懿等, 2015 ).已有研究发现, 闽江河口湿地生长季阶段的入侵种互花米草和本地种短叶茳芏均具有一定的内稳性, 互花米草N和N:P的内稳性高于短叶茳芏N和N:P的内稳性, 而短叶茳芏P的内稳性高于互花米草P的内稳性(蒋利玲等, 2014 ). ...
Biological control of Spartina alterniflora in Willapa Bay, Washington using the planthopper Prokelisia marginata: Agent specificity and early results
1
2003
... 目前, 互花米草的防治措施主要有物理控制(Frid et al. , 1999 )、化学控制(Patten, 2002 )和生物控制(Wu et al ., 1999 , Grevstad et al ., 2003 )这3种方法, 其中以收割、碎根水淹等物理方法居多.研究表明, 翻耕和碎根处理方法仅在当年有一定的控制效果, 不具有长期有效性, 且割除碎根也需要在最佳季节进行(李贺鹏和张利权, 2007 ).已有的研究认为: 闽江河口湿地生态系统在7月份时互花米草内稳性最低, 是治理互花米草的最佳时期(蒋利玲等, 2014 ).充分考虑植物内稳性特征的影响, 建议采用物理-化学综合方法进行互花米草的控制, 在7月份对互花米草进行碎根, 再通过施加N缓解短叶茳芏根和茎的养分限制, 从而增大短叶茳芏群落在湿地生态系统中的内稳性. ...
N:P ratios in terrestrial plants: Variation and functional significance
2
2004
... 国外内稳性研究的对象由最初的真菌和细菌(Levi & Cowling, 1969 ; Goldman et al ., 1987 )发展到藻类、浮游动物和草本植物(Rhee, 1978 ; Andersen & Hessen, 1991 ; Güsewell et al ., 2005 ).由于高等植物体内元素的内稳性和化学计量特征复杂, 对高等植物的研究甚少(Cernusak et al ., 2009 ; Matzek & Vitousek, 2009 ; Yu et al ., 2010 ).生态系统水平上对内稳性的研究多集中于海洋生态系统和陆地生态系统(Güsewell, 2004 ; 苏强, 2012 ), 对湿地生态系统的研究较少(Wang et al ., 2014 ).国内内稳性的研究还处于起步阶段.庾强(2009)以内蒙古草原为研究对象, 通过1 200 km的横断范围的样带试验和内蒙古草原生态系统定位研究站27年的监测数据发现, 内稳性高的物种具有较高的优势度和生物量稳定性, 内稳性高的生态系统则具有较高的生产力和稳定性.因此, 内稳性是衡量物种竞争力的一个重要指标.已有研究表明内稳性的高低受多种因子的影响, 其中包括植物器官(庾强, 2009 )、养分供应(严正兵等, 2013 )、光强以及施肥(Sterner & Elser, 2002 )等, 影响因子的不确定性使得不同研究中内稳性对环境的响应不一致.目前有关互花米草入侵对本地生态系统的影响多集中在物质循环以及生态化学计量学, 而物种水平上内稳性的研究甚少(张仁懿等, 2015 ).已有研究发现, 闽江河口湿地生长季阶段的入侵种互花米草和本地种短叶茳芏均具有一定的内稳性, 互花米草N和N:P的内稳性高于短叶茳芏N和N:P的内稳性, 而短叶茳芏P的内稳性高于互花米草P的内稳性(蒋利玲等, 2014 ). ...
... 根据Garnier理论, 植物与土壤对应养分含量存在正相关关系则说明植物生长受到养分限制(Garnier, 1998 ).根据该理论, 福建省闽江河口鳝鱼滩湿地生态系统中短叶茳芏茎受N限制, 单种群落中的互花米草茎和鞘受N限制, 混生群落中的互花米草茎受N和P共同限制.根据Koerselman理论, 植物体N:P < 14表明其相对受N限制, N:P > 16相对受P限制, 14 < N:P < 16则受N和P共同限制(Koerselman & Meuleman, 1996 ; Güsewell & Koerselman, 2002 ).由于不同研究区域、不同生态系统以及不同植被类型的N:P临界阈值均会发生改变, 精确的N:P临界阈值需要通过严格的施肥试验来确定(Zhang et al ., 2004 ; Güsewell, 2004 ).胡伟芳等(2014)对中国主要湿地649种植物的N、P化学计量特征进行了整合分析, 结果表明不同湿地生境的植物受限制状况不尽相同, 其中N限制(N:P < 14)比例最高, 占59%.针对湿地生态系统的大多数研究也多用14和16为阈值来判断养分受限制情况(Güsewell & Koerselman, 2002 ; Tessier & Raynal, 2003 ; Bott et al ., 2008 ).因此, 若本研究沿用该临界阈值进行分析, 则单种群落和混生群落中的互花米草茎均受N、P共同限制, 其叶均受P限制, 单种群落和混生群落中的短叶茳芏茎均受N限制.本研究中, 互花米草各器官N、P含量平均值均低于短叶茳芏, 但N:P值平均值高于短叶茳芏, 这表明互花米草在生长过程中对养分的需求量相对较低, 但对N的利用效率高于对P的利用效率. ...
Effects of heterogeneous habitat use by cattle on nutrient availability and litter decomposition in soils of an Alpine pasture
1
2005
... 国外内稳性研究的对象由最初的真菌和细菌(Levi & Cowling, 1969 ; Goldman et al ., 1987 )发展到藻类、浮游动物和草本植物(Rhee, 1978 ; Andersen & Hessen, 1991 ; Güsewell et al ., 2005 ).由于高等植物体内元素的内稳性和化学计量特征复杂, 对高等植物的研究甚少(Cernusak et al ., 2009 ; Matzek & Vitousek, 2009 ; Yu et al ., 2010 ).生态系统水平上对内稳性的研究多集中于海洋生态系统和陆地生态系统(Güsewell, 2004 ; 苏强, 2012 ), 对湿地生态系统的研究较少(Wang et al ., 2014 ).国内内稳性的研究还处于起步阶段.庾强(2009)以内蒙古草原为研究对象, 通过1 200 km的横断范围的样带试验和内蒙古草原生态系统定位研究站27年的监测数据发现, 内稳性高的物种具有较高的优势度和生物量稳定性, 内稳性高的生态系统则具有较高的生产力和稳定性.因此, 内稳性是衡量物种竞争力的一个重要指标.已有研究表明内稳性的高低受多种因子的影响, 其中包括植物器官(庾强, 2009 )、养分供应(严正兵等, 2013 )、光强以及施肥(Sterner & Elser, 2002 )等, 影响因子的不确定性使得不同研究中内稳性对环境的响应不一致.目前有关互花米草入侵对本地生态系统的影响多集中在物质循环以及生态化学计量学, 而物种水平上内稳性的研究甚少(张仁懿等, 2015 ).已有研究发现, 闽江河口湿地生长季阶段的入侵种互花米草和本地种短叶茳芏均具有一定的内稳性, 互花米草N和N:P的内稳性高于短叶茳芏N和N:P的内稳性, 而短叶茳芏P的内稳性高于互花米草P的内稳性(蒋利玲等, 2014 ). ...
Variation in nitrogen and phosphorus concentrations of wetland plants
3
2002
... 根据Garnier理论, 植物与土壤对应养分含量存在正相关关系则说明植物生长受到养分限制(Garnier, 1998 ).根据该理论, 福建省闽江河口鳝鱼滩湿地生态系统中短叶茳芏茎受N限制, 单种群落中的互花米草茎和鞘受N限制, 混生群落中的互花米草茎受N和P共同限制.根据Koerselman理论, 植物体N:P < 14表明其相对受N限制, N:P > 16相对受P限制, 14 < N:P < 16则受N和P共同限制(Koerselman & Meuleman, 1996 ; Güsewell & Koerselman, 2002 ).由于不同研究区域、不同生态系统以及不同植被类型的N:P临界阈值均会发生改变, 精确的N:P临界阈值需要通过严格的施肥试验来确定(Zhang et al ., 2004 ; Güsewell, 2004 ).胡伟芳等(2014)对中国主要湿地649种植物的N、P化学计量特征进行了整合分析, 结果表明不同湿地生境的植物受限制状况不尽相同, 其中N限制(N:P < 14)比例最高, 占59%.针对湿地生态系统的大多数研究也多用14和16为阈值来判断养分受限制情况(Güsewell & Koerselman, 2002 ; Tessier & Raynal, 2003 ; Bott et al ., 2008 ).因此, 若本研究沿用该临界阈值进行分析, 则单种群落和混生群落中的互花米草茎均受N、P共同限制, 其叶均受P限制, 单种群落和混生群落中的短叶茳芏茎均受N限制.本研究中, 互花米草各器官N、P含量平均值均低于短叶茳芏, 但N:P值平均值高于短叶茳芏, 这表明互花米草在生长过程中对养分的需求量相对较低, 但对N的利用效率高于对P的利用效率. ...
... ).胡伟芳等(2014)对中国主要湿地649种植物的N、P化学计量特征进行了整合分析, 结果表明不同湿地生境的植物受限制状况不尽相同, 其中N限制(N:P < 14)比例最高, 占59%.针对湿地生态系统的大多数研究也多用14和16为阈值来判断养分受限制情况(Güsewell & Koerselman, 2002 ; Tessier & Raynal, 2003 ; Bott et al ., 2008 ).因此, 若本研究沿用该临界阈值进行分析, 则单种群落和混生群落中的互花米草茎均受N、P共同限制, 其叶均受P限制, 单种群落和混生群落中的短叶茳芏茎均受N限制.本研究中, 互花米草各器官N、P含量平均值均低于短叶茳芏, 但N:P值平均值高于短叶茳芏, 这表明互花米草在生长过程中对养分的需求量相对较低, 但对N的利用效率高于对P的利用效率. ...
... Garnier理论和Koerselman理论是分析植物限制性养分的两种常用方法, Koerselman理论是基于生态化学计量对系统能力平衡与元素平衡相互影响的分析, 已在水生和湿地生态系统中得到广泛应用(Güsewell & Koerselman, 2002 ), 但因受气候特征、物种类型等影响而具有局限性.Garnier理论是特定环境中土壤与植物间相互作用的结果, 是植物需求的内在属性、土壤养分提供以及气候等外因的综 合表现, 具有较强的耦联性, 但结果也受外因影响(樊晓勇, 2012 ).本研究中, Garnier理论和Koerselman理论均表明单种群落和混生群落中的短叶茳芏茎的生长均受N限制, 混生群落中的互花米草茎的生长受N和P共同限制, 其他群落中的部分植物器官生长的受限结果存在差异.因此, 本研究同时采用这两种方法进行养分限制性分析, 由两种方法一致得到的结果以及由此获得的结论, 我们认为更可靠. ...
Effects of biodiversity on ecosystem functioning: A consensus of current knowledge
1
2005
... 生物入侵作为全球变化的一个重要方面, 严重威胁了土著生物群落的生物多样性和生态系统功能(Davis, 2003 ; Hooper et al ., 2005 ).作为我国中亚热带和南亚热带过渡区最具代表性的河口湿地之一的闽江河口湿地(刘剑秋等, 2006 ), 自2002年以来逐渐遭到互花米草(Spartina alterniflora )的大规模入侵.互花米草以其极强的适应力和繁殖力迅速占据空生态位, 侵占本地种短叶茳芏(Cyperus malaccensis var. brevifolius )的生境, 在潮间带形成了短叶茳芏单种群落、互花米草单种群落和互花米草-短叶茳芏混生群落共存的竞争格局.入侵的互花米草通过影响土著生态系统的结构, 进而影响生态系统的养分循环, 生态系统养分的可利用性改变反过来又影响植物的生长状况, 从而可能改变入侵种与土著种之间的竞争关系.养分元素氮(N)和磷(P)是湿地生态系统植物生长的主要限制资源, 其可利用性及生态化学计量关系是植物生长和生态系统物质循环的重要调节因子(熊汉锋和王运华, 2005 ).植物能在外界养分元素组成改变的情况下, 一定程度上维持自身各元素之间的相对比例.这种由生物在长期进化过程中为适应外界环境变化而形成的维持自身化学组成相对稳定的能力就叫作生态化学计量内稳性(简称内稳性)(Kooijman, 1995 ). ...
中国主要湿地植被氮和磷生态化学计量学特征
2014
闽江河口湿地3种植物化学计量内稳性特征
2
2014
... 国外内稳性研究的对象由最初的真菌和细菌(Levi & Cowling, 1969 ; Goldman et al ., 1987 )发展到藻类、浮游动物和草本植物(Rhee, 1978 ; Andersen & Hessen, 1991 ; Güsewell et al ., 2005 ).由于高等植物体内元素的内稳性和化学计量特征复杂, 对高等植物的研究甚少(Cernusak et al ., 2009 ; Matzek & Vitousek, 2009 ; Yu et al ., 2010 ).生态系统水平上对内稳性的研究多集中于海洋生态系统和陆地生态系统(Güsewell, 2004 ; 苏强, 2012 ), 对湿地生态系统的研究较少(Wang et al ., 2014 ).国内内稳性的研究还处于起步阶段.庾强(2009)以内蒙古草原为研究对象, 通过1 200 km的横断范围的样带试验和内蒙古草原生态系统定位研究站27年的监测数据发现, 内稳性高的物种具有较高的优势度和生物量稳定性, 内稳性高的生态系统则具有较高的生产力和稳定性.因此, 内稳性是衡量物种竞争力的一个重要指标.已有研究表明内稳性的高低受多种因子的影响, 其中包括植物器官(庾强, 2009 )、养分供应(严正兵等, 2013 )、光强以及施肥(Sterner & Elser, 2002 )等, 影响因子的不确定性使得不同研究中内稳性对环境的响应不一致.目前有关互花米草入侵对本地生态系统的影响多集中在物质循环以及生态化学计量学, 而物种水平上内稳性的研究甚少(张仁懿等, 2015 ).已有研究发现, 闽江河口湿地生长季阶段的入侵种互花米草和本地种短叶茳芏均具有一定的内稳性, 互花米草N和N:P的内稳性高于短叶茳芏N和N:P的内稳性, 而短叶茳芏P的内稳性高于互花米草P的内稳性(蒋利玲等, 2014 ). ...
... 目前, 互花米草的防治措施主要有物理控制(Frid et al. , 1999 )、化学控制(Patten, 2002 )和生物控制(Wu et al ., 1999 , Grevstad et al ., 2003 )这3种方法, 其中以收割、碎根水淹等物理方法居多.研究表明, 翻耕和碎根处理方法仅在当年有一定的控制效果, 不具有长期有效性, 且割除碎根也需要在最佳季节进行(李贺鹏和张利权, 2007 ).已有的研究认为: 闽江河口湿地生态系统在7月份时互花米草内稳性最低, 是治理互花米草的最佳时期(蒋利玲等, 2014 ).充分考虑植物内稳性特征的影响, 建议采用物理-化学综合方法进行互花米草的控制, 在7月份对互花米草进行碎根, 再通过施加N缓解短叶茳芏根和茎的养分限制, 从而增大短叶茳芏群落在湿地生态系统中的内稳性. ...
凯氏蒸馏法和元素分析仪法测定沉积物中全氮含量的异同及其意义
1
2006
... 选择潮间带生境的互花米草单种群落、互花米草-短叶茳芏混生群落和短叶茳芏单种群落沼泽湿地为研究对象, 对沼泽湿地的土壤和植物进行取样.各群落斑块面积约为30 m × 30 m, 在每个斑块的样方内随机选取4个面积大小为25 cm × 25 cm的小样方作为重复.在2012年4月至2013年2月, 每2个月采样一次, 采用齐地面收割方式获得植物的地上部分, 采用挖掘法获取0-30 cm深度的地下所有根系和土壤.土壤无机氮的测定采用KCl浸提-连续流动分析仪法(李玉英等, 2009 ), 土壤有效磷的测定采用NaHCO3 浸提-钼锑抗比色法① (① Olsen SR, Cole CV, Watanabe FS, Dean LAC (1954). Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate. US Department of Agriculture, Washington, DC, USA. 939.); 植物全氮的测定采用元素分析仪法(江伟等, 2006 ), 植物全磷的测定采用浓硫酸-高氯酸消煮-连续流动分析仪法(鲍士旦, 2005 ). ...
To be or not to be what you eat: Regulation of stoichiometric homeostasis among autotrophs and heterotrophs
1
2010
... 其中, x 为土壤无机N或有效P含量(mg·kg-1 )或N:P值, y 为植物体器官对应的全氮(N)或全磷(P)含量(mg·kg-1 )或N:P值,c 为常数, H 为植物内稳性指数.H 使用年数据进行拟合计算得到, 其中植物器官的内稳性为各器官在N、P以及N:P值水平上的拟合, 物种的内稳性为所有器官内稳性的平均值, 生态系统N、P以及N:P值的内稳性为该生态系统所有植物整体在相应N、P和N:P值水平上的拟合.H 的类型可根据H > 4为稳态型、2 < H < 4为弱稳态型、4/3 < H < 2为弱敏感型, 以及H < 4/3为敏感型来界定(Jonas et al ., 2010 ). ...
The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation
1
1996
... 根据Garnier理论, 植物与土壤对应养分含量存在正相关关系则说明植物生长受到养分限制(Garnier, 1998 ).根据该理论, 福建省闽江河口鳝鱼滩湿地生态系统中短叶茳芏茎受N限制, 单种群落中的互花米草茎和鞘受N限制, 混生群落中的互花米草茎受N和P共同限制.根据Koerselman理论, 植物体N:P < 14表明其相对受N限制, N:P > 16相对受P限制, 14 < N:P < 16则受N和P共同限制(Koerselman & Meuleman, 1996 ; Güsewell & Koerselman, 2002 ).由于不同研究区域、不同生态系统以及不同植被类型的N:P临界阈值均会发生改变, 精确的N:P临界阈值需要通过严格的施肥试验来确定(Zhang et al ., 2004 ; Güsewell, 2004 ).胡伟芳等(2014)对中国主要湿地649种植物的N、P化学计量特征进行了整合分析, 结果表明不同湿地生境的植物受限制状况不尽相同, 其中N限制(N:P < 14)比例最高, 占59%.针对湿地生态系统的大多数研究也多用14和16为阈值来判断养分受限制情况(Güsewell & Koerselman, 2002 ; Tessier & Raynal, 2003 ; Bott et al ., 2008 ).因此, 若本研究沿用该临界阈值进行分析, 则单种群落和混生群落中的互花米草茎均受N、P共同限制, 其叶均受P限制, 单种群落和混生群落中的短叶茳芏茎均受N限制.本研究中, 互花米草各器官N、P含量平均值均低于短叶茳芏, 但N:P值平均值高于短叶茳芏, 这表明互花米草在生长过程中对养分的需求量相对较低, 但对N的利用效率高于对P的利用效率. ...
The stoichiometry of animal energetics
1
1995
... 生物入侵作为全球变化的一个重要方面, 严重威胁了土著生物群落的生物多样性和生态系统功能(Davis, 2003 ; Hooper et al ., 2005 ).作为我国中亚热带和南亚热带过渡区最具代表性的河口湿地之一的闽江河口湿地(刘剑秋等, 2006 ), 自2002年以来逐渐遭到互花米草(Spartina alterniflora )的大规模入侵.互花米草以其极强的适应力和繁殖力迅速占据空生态位, 侵占本地种短叶茳芏(Cyperus malaccensis var. brevifolius )的生境, 在潮间带形成了短叶茳芏单种群落、互花米草单种群落和互花米草-短叶茳芏混生群落共存的竞争格局.入侵的互花米草通过影响土著生态系统的结构, 进而影响生态系统的养分循环, 生态系统养分的可利用性改变反过来又影响植物的生长状况, 从而可能改变入侵种与土著种之间的竞争关系.养分元素氮(N)和磷(P)是湿地生态系统植物生长的主要限制资源, 其可利用性及生态化学计量关系是植物生长和生态系统物质循环的重要调节因子(熊汉锋和王运华, 2005 ).植物能在外界养分元素组成改变的情况下, 一定程度上维持自身各元素之间的相对比例.这种由生物在长期进化过程中为适应外界环境变化而形成的维持自身化学组成相对稳定的能力就叫作生态化学计量内稳性(简称内稳性)(Kooijman, 1995 ). ...
Role of nitrogen in wood deterioration. VII. Physiological adaptation of wood-destroying and other fungi to substrates deficient in nitrogen
1
1969
... 国外内稳性研究的对象由最初的真菌和细菌(Levi & Cowling, 1969 ; Goldman et al ., 1987 )发展到藻类、浮游动物和草本植物(Rhee, 1978 ; Andersen & Hessen, 1991 ; Güsewell et al ., 2005 ).由于高等植物体内元素的内稳性和化学计量特征复杂, 对高等植物的研究甚少(Cernusak et al ., 2009 ; Matzek & Vitousek, 2009 ; Yu et al ., 2010 ).生态系统水平上对内稳性的研究多集中于海洋生态系统和陆地生态系统(Güsewell, 2004 ; 苏强, 2012 ), 对湿地生态系统的研究较少(Wang et al ., 2014 ).国内内稳性的研究还处于起步阶段.庾强(2009)以内蒙古草原为研究对象, 通过1 200 km的横断范围的样带试验和内蒙古草原生态系统定位研究站27年的监测数据发现, 内稳性高的物种具有较高的优势度和生物量稳定性, 内稳性高的生态系统则具有较高的生产力和稳定性.因此, 内稳性是衡量物种竞争力的一个重要指标.已有研究表明内稳性的高低受多种因子的影响, 其中包括植物器官(庾强, 2009 )、养分供应(严正兵等, 2013 )、光强以及施肥(Sterner & Elser, 2002 )等, 影响因子的不确定性使得不同研究中内稳性对环境的响应不一致.目前有关互花米草入侵对本地生态系统的影响多集中在物质循环以及生态化学计量学, 而物种水平上内稳性的研究甚少(张仁懿等, 2015 ).已有研究发现, 闽江河口湿地生长季阶段的入侵种互花米草和本地种短叶茳芏均具有一定的内稳性, 互花米草N和N:P的内稳性高于短叶茳芏N和N:P的内稳性, 而短叶茳芏P的内稳性高于互花米草P的内稳性(蒋利玲等, 2014 ). ...
外来植物互花米草的物理控制实验研究
1
2007
... 目前, 互花米草的防治措施主要有物理控制(Frid et al. , 1999 )、化学控制(Patten, 2002 )和生物控制(Wu et al ., 1999 , Grevstad et al ., 2003 )这3种方法, 其中以收割、碎根水淹等物理方法居多.研究表明, 翻耕和碎根处理方法仅在当年有一定的控制效果, 不具有长期有效性, 且割除碎根也需要在最佳季节进行(李贺鹏和张利权, 2007 ).已有的研究认为: 闽江河口湿地生态系统在7月份时互花米草内稳性最低, 是治理互花米草的最佳时期(蒋利玲等, 2014 ).充分考虑植物内稳性特征的影响, 建议采用物理-化学综合方法进行互花米草的控制, 在7月份对互花米草进行碎根, 再通过施加N缓解短叶茳芏根和茎的养分限制, 从而增大短叶茳芏群落在湿地生态系统中的内稳性. ...
施氮量和蚕豆/玉米间作对土壤无机氮时空分布的影响
1
2009
... 选择潮间带生境的互花米草单种群落、互花米草-短叶茳芏混生群落和短叶茳芏单种群落沼泽湿地为研究对象, 对沼泽湿地的土壤和植物进行取样.各群落斑块面积约为30 m × 30 m, 在每个斑块的样方内随机选取4个面积大小为25 cm × 25 cm的小样方作为重复.在2012年4月至2013年2月, 每2个月采样一次, 采用齐地面收割方式获得植物的地上部分, 采用挖掘法获取0-30 cm深度的地下所有根系和土壤.土壤无机氮的测定采用KCl浸提-连续流动分析仪法(李玉英等, 2009 ), 土壤有效磷的测定采用NaHCO3 浸提-钼锑抗比色法① (① Olsen SR, Cole CV, Watanabe FS, Dean LAC (1954). Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate. US Department of Agriculture, Washington, DC, USA. 939.); 植物全氮的测定采用元素分析仪法(江伟等, 2006 ), 植物全磷的测定采用浓硫酸-高氯酸消煮-连续流动分析仪法(鲍士旦, 2005 ). ...
1
2006
... 生物入侵作为全球变化的一个重要方面, 严重威胁了土著生物群落的生物多样性和生态系统功能(Davis, 2003 ; Hooper et al ., 2005 ).作为我国中亚热带和南亚热带过渡区最具代表性的河口湿地之一的闽江河口湿地(刘剑秋等, 2006 ), 自2002年以来逐渐遭到互花米草(Spartina alterniflora )的大规模入侵.互花米草以其极强的适应力和繁殖力迅速占据空生态位, 侵占本地种短叶茳芏(Cyperus malaccensis var. brevifolius )的生境, 在潮间带形成了短叶茳芏单种群落、互花米草单种群落和互花米草-短叶茳芏混生群落共存的竞争格局.入侵的互花米草通过影响土著生态系统的结构, 进而影响生态系统的养分循环, 生态系统养分的可利用性改变反过来又影响植物的生长状况, 从而可能改变入侵种与土著种之间的竞争关系.养分元素氮(N)和磷(P)是湿地生态系统植物生长的主要限制资源, 其可利用性及生态化学计量关系是植物生长和生态系统物质循环的重要调节因子(熊汉锋和王运华, 2005 ).植物能在外界养分元素组成改变的情况下, 一定程度上维持自身各元素之间的相对比例.这种由生物在长期进化过程中为适应外界环境变化而形成的维持自身化学组成相对稳定的能力就叫作生态化学计量内稳性(简称内稳性)(Kooijman, 1995 ). ...
N:P stoichiometry and protein: RNA ratios in vascular plants: An evaluation of the growth-rate hypothesis
1
2009
... 国外内稳性研究的对象由最初的真菌和细菌(Levi & Cowling, 1969 ; Goldman et al ., 1987 )发展到藻类、浮游动物和草本植物(Rhee, 1978 ; Andersen & Hessen, 1991 ; Güsewell et al ., 2005 ).由于高等植物体内元素的内稳性和化学计量特征复杂, 对高等植物的研究甚少(Cernusak et al ., 2009 ; Matzek & Vitousek, 2009 ; Yu et al ., 2010 ).生态系统水平上对内稳性的研究多集中于海洋生态系统和陆地生态系统(Güsewell, 2004 ; 苏强, 2012 ), 对湿地生态系统的研究较少(Wang et al ., 2014 ).国内内稳性的研究还处于起步阶段.庾强(2009)以内蒙古草原为研究对象, 通过1 200 km的横断范围的样带试验和内蒙古草原生态系统定位研究站27年的监测数据发现, 内稳性高的物种具有较高的优势度和生物量稳定性, 内稳性高的生态系统则具有较高的生产力和稳定性.因此, 内稳性是衡量物种竞争力的一个重要指标.已有研究表明内稳性的高低受多种因子的影响, 其中包括植物器官(庾强, 2009 )、养分供应(严正兵等, 2013 )、光强以及施肥(Sterner & Elser, 2002 )等, 影响因子的不确定性使得不同研究中内稳性对环境的响应不一致.目前有关互花米草入侵对本地生态系统的影响多集中在物质循环以及生态化学计量学, 而物种水平上内稳性的研究甚少(张仁懿等, 2015 ).已有研究发现, 闽江河口湿地生长季阶段的入侵种互花米草和本地种短叶茳芏均具有一定的内稳性, 互花米草N和N:P的内稳性高于短叶茳芏N和N:P的内稳性, 而短叶茳芏P的内稳性高于互花米草P的内稳性(蒋利玲等, 2014 ). ...
Diversity and activity of epiphytic nitrogen fixers on standing dead stems of the salt marsh grass Spartina alterniflora
1
2005
... 入侵对植物内稳性产生的影响主要表现在N元素上.混生群落中的互花米草根、叶和鞘的H N 分别显著高于对应单种群落中的互花米草根、茎和鞘的H N .混生群落中的互花米草根的H N 比单种群落中的互花米草根的H N 高25.7倍, 混生群落中的短叶茳芏根的H N 比单种群落中的短叶茳芏根的H N 增大了1.48倍, 两者之间的差异可能与互花米草根际微生物的固氮作用有关, 这使得互花米草获得更多的N, 进而缓解了N的限制(Bagwell & Lovell, 2000 ; Moisander et al ., 2005 ; 曾艳等, 2011 ).入侵初期, 单种群落的植物生长受N限制, 植物体内N的含量很大程度上取决于土壤中可利用N的含量, 故植物内稳性较低.随着入侵过程的进行, 互花米草的根际固氮作用使植物体内N含量的获取对土壤中N的依赖性降低, 从而使混生群落中的植物表现出较高的内稳性.相比短叶茳芏根的养分利用性, 互花米草根内稳性的调节能力使其能够在多变的环境中保持较好的生长, 进而具备更强的竞争力和适应力.已有的研究表明, 生态系统的稳定性与根生物量正相关, 根在变化的环境中可通过回收利用和贮存养分来调节生长, 从而提高物种和群落水平的生产力和稳定性(Tilman et al ., 2006 ).因此, 根内稳性与群落生产力和稳定性的关系研究值得进一步探索. ...
Smooth cordgrass (Spartina alterniflora ) control with Imazapyr
1
2002
... 目前, 互花米草的防治措施主要有物理控制(Frid et al. , 1999 )、化学控制(Patten, 2002 )和生物控制(Wu et al ., 1999 , Grevstad et al ., 2003 )这3种方法, 其中以收割、碎根水淹等物理方法居多.研究表明, 翻耕和碎根处理方法仅在当年有一定的控制效果, 不具有长期有效性, 且割除碎根也需要在最佳季节进行(李贺鹏和张利权, 2007 ).已有的研究认为: 闽江河口湿地生态系统在7月份时互花米草内稳性最低, 是治理互花米草的最佳时期(蒋利玲等, 2014 ).充分考虑植物内稳性特征的影响, 建议采用物理-化学综合方法进行互花米草的控制, 在7月份对互花米草进行碎根, 再通过施加N缓解短叶茳芏根和茎的养分限制, 从而增大短叶茳芏群落在湿地生态系统中的内稳性. ...
Effects of N:P atomic ratios and nitrate limitation on algal growth, cell composition and nitrate uptake
1
1978
... 国外内稳性研究的对象由最初的真菌和细菌(Levi & Cowling, 1969 ; Goldman et al ., 1987 )发展到藻类、浮游动物和草本植物(Rhee, 1978 ; Andersen & Hessen, 1991 ; Güsewell et al ., 2005 ).由于高等植物体内元素的内稳性和化学计量特征复杂, 对高等植物的研究甚少(Cernusak et al ., 2009 ; Matzek & Vitousek, 2009 ; Yu et al ., 2010 ).生态系统水平上对内稳性的研究多集中于海洋生态系统和陆地生态系统(Güsewell, 2004 ; 苏强, 2012 ), 对湿地生态系统的研究较少(Wang et al ., 2014 ).国内内稳性的研究还处于起步阶段.庾强(2009)以内蒙古草原为研究对象, 通过1 200 km的横断范围的样带试验和内蒙古草原生态系统定位研究站27年的监测数据发现, 内稳性高的物种具有较高的优势度和生物量稳定性, 内稳性高的生态系统则具有较高的生产力和稳定性.因此, 内稳性是衡量物种竞争力的一个重要指标.已有研究表明内稳性的高低受多种因子的影响, 其中包括植物器官(庾强, 2009 )、养分供应(严正兵等, 2013 )、光强以及施肥(Sterner & Elser, 2002 )等, 影响因子的不确定性使得不同研究中内稳性对环境的响应不一致.目前有关互花米草入侵对本地生态系统的影响多集中在物质循环以及生态化学计量学, 而物种水平上内稳性的研究甚少(张仁懿等, 2015 ).已有研究发现, 闽江河口湿地生长季阶段的入侵种互花米草和本地种短叶茳芏均具有一定的内稳性, 互花米草N和N:P的内稳性高于短叶茳芏N和N:P的内稳性, 而短叶茳芏P的内稳性高于互花米草P的内稳性(蒋利玲等, 2014 ). ...
2
2002
... 国外内稳性研究的对象由最初的真菌和细菌(Levi & Cowling, 1969 ; Goldman et al ., 1987 )发展到藻类、浮游动物和草本植物(Rhee, 1978 ; Andersen & Hessen, 1991 ; Güsewell et al ., 2005 ).由于高等植物体内元素的内稳性和化学计量特征复杂, 对高等植物的研究甚少(Cernusak et al ., 2009 ; Matzek & Vitousek, 2009 ; Yu et al ., 2010 ).生态系统水平上对内稳性的研究多集中于海洋生态系统和陆地生态系统(Güsewell, 2004 ; 苏强, 2012 ), 对湿地生态系统的研究较少(Wang et al ., 2014 ).国内内稳性的研究还处于起步阶段.庾强(2009)以内蒙古草原为研究对象, 通过1 200 km的横断范围的样带试验和内蒙古草原生态系统定位研究站27年的监测数据发现, 内稳性高的物种具有较高的优势度和生物量稳定性, 内稳性高的生态系统则具有较高的生产力和稳定性.因此, 内稳性是衡量物种竞争力的一个重要指标.已有研究表明内稳性的高低受多种因子的影响, 其中包括植物器官(庾强, 2009 )、养分供应(严正兵等, 2013 )、光强以及施肥(Sterner & Elser, 2002 )等, 影响因子的不确定性使得不同研究中内稳性对环境的响应不一致.目前有关互花米草入侵对本地生态系统的影响多集中在物质循环以及生态化学计量学, 而物种水平上内稳性的研究甚少(张仁懿等, 2015 ).已有研究发现, 闽江河口湿地生长季阶段的入侵种互花米草和本地种短叶茳芏均具有一定的内稳性, 互花米草N和N:P的内稳性高于短叶茳芏N和N:P的内稳性, 而短叶茳芏P的内稳性高于互花米草P的内稳性(蒋利玲等, 2014 ). ...
... 根据理论推导与研究, Sterner和Elser (2002) 提出生物生态化学计量内稳性模型.该模型指数公 式为: ...
浮游动物化学计量学稳态性特征研究进展
1
2012
... 国外内稳性研究的对象由最初的真菌和细菌(Levi & Cowling, 1969 ; Goldman et al ., 1987 )发展到藻类、浮游动物和草本植物(Rhee, 1978 ; Andersen & Hessen, 1991 ; Güsewell et al ., 2005 ).由于高等植物体内元素的内稳性和化学计量特征复杂, 对高等植物的研究甚少(Cernusak et al ., 2009 ; Matzek & Vitousek, 2009 ; Yu et al ., 2010 ).生态系统水平上对内稳性的研究多集中于海洋生态系统和陆地生态系统(Güsewell, 2004 ; 苏强, 2012 ), 对湿地生态系统的研究较少(Wang et al ., 2014 ).国内内稳性的研究还处于起步阶段.庾强(2009)以内蒙古草原为研究对象, 通过1 200 km的横断范围的样带试验和内蒙古草原生态系统定位研究站27年的监测数据发现, 内稳性高的物种具有较高的优势度和生物量稳定性, 内稳性高的生态系统则具有较高的生产力和稳定性.因此, 内稳性是衡量物种竞争力的一个重要指标.已有研究表明内稳性的高低受多种因子的影响, 其中包括植物器官(庾强, 2009 )、养分供应(严正兵等, 2013 )、光强以及施肥(Sterner & Elser, 2002 )等, 影响因子的不确定性使得不同研究中内稳性对环境的响应不一致.目前有关互花米草入侵对本地生态系统的影响多集中在物质循环以及生态化学计量学, 而物种水平上内稳性的研究甚少(张仁懿等, 2015 ).已有研究发现, 闽江河口湿地生长季阶段的入侵种互花米草和本地种短叶茳芏均具有一定的内稳性, 互花米草N和N:P的内稳性高于短叶茳芏N和N:P的内稳性, 而短叶茳芏P的内稳性高于互花米草P的内稳性(蒋利玲等, 2014 ). ...
Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation
1
2003
... 根据Garnier理论, 植物与土壤对应养分含量存在正相关关系则说明植物生长受到养分限制(Garnier, 1998 ).根据该理论, 福建省闽江河口鳝鱼滩湿地生态系统中短叶茳芏茎受N限制, 单种群落中的互花米草茎和鞘受N限制, 混生群落中的互花米草茎受N和P共同限制.根据Koerselman理论, 植物体N:P < 14表明其相对受N限制, N:P > 16相对受P限制, 14 < N:P < 16则受N和P共同限制(Koerselman & Meuleman, 1996 ; Güsewell & Koerselman, 2002 ).由于不同研究区域、不同生态系统以及不同植被类型的N:P临界阈值均会发生改变, 精确的N:P临界阈值需要通过严格的施肥试验来确定(Zhang et al ., 2004 ; Güsewell, 2004 ).胡伟芳等(2014)对中国主要湿地649种植物的N、P化学计量特征进行了整合分析, 结果表明不同湿地生境的植物受限制状况不尽相同, 其中N限制(N:P < 14)比例最高, 占59%.针对湿地生态系统的大多数研究也多用14和16为阈值来判断养分受限制情况(Güsewell & Koerselman, 2002 ; Tessier & Raynal, 2003 ; Bott et al ., 2008 ).因此, 若本研究沿用该临界阈值进行分析, 则单种群落和混生群落中的互花米草茎均受N、P共同限制, 其叶均受P限制, 单种群落和混生群落中的短叶茳芏茎均受N限制.本研究中, 互花米草各器官N、P含量平均值均低于短叶茳芏, 但N:P值平均值高于短叶茳芏, 这表明互花米草在生长过程中对养分的需求量相对较低, 但对N的利用效率高于对P的利用效率. ...
Biodiversity and ecosystem stability in a decade-long grassland experiment
1
2006
... 入侵对植物内稳性产生的影响主要表现在N元素上.混生群落中的互花米草根、叶和鞘的H N 分别显著高于对应单种群落中的互花米草根、茎和鞘的H N .混生群落中的互花米草根的H N 比单种群落中的互花米草根的H N 高25.7倍, 混生群落中的短叶茳芏根的H N 比单种群落中的短叶茳芏根的H N 增大了1.48倍, 两者之间的差异可能与互花米草根际微生物的固氮作用有关, 这使得互花米草获得更多的N, 进而缓解了N的限制(Bagwell & Lovell, 2000 ; Moisander et al ., 2005 ; 曾艳等, 2011 ).入侵初期, 单种群落的植物生长受N限制, 植物体内N的含量很大程度上取决于土壤中可利用N的含量, 故植物内稳性较低.随着入侵过程的进行, 互花米草的根际固氮作用使植物体内N含量的获取对土壤中N的依赖性降低, 从而使混生群落中的植物表现出较高的内稳性.相比短叶茳芏根的养分利用性, 互花米草根内稳性的调节能力使其能够在多变的环境中保持较好的生长, 进而具备更强的竞争力和适应力.已有的研究表明, 生态系统的稳定性与根生物量正相关, 根在变化的环境中可通过回收利用和贮存养分来调节生长, 从而提高物种和群落水平的生产力和稳定性(Tilman et al ., 2006 ).因此, 根内稳性与群落生产力和稳定性的关系研究值得进一步探索. ...
Linking stoichiometric homeostasis of microorganisms with soil phosphorus dynamics in wetlands subjected to microcosm warming
1
2014
... 国外内稳性研究的对象由最初的真菌和细菌(Levi & Cowling, 1969 ; Goldman et al ., 1987 )发展到藻类、浮游动物和草本植物(Rhee, 1978 ; Andersen & Hessen, 1991 ; Güsewell et al ., 2005 ).由于高等植物体内元素的内稳性和化学计量特征复杂, 对高等植物的研究甚少(Cernusak et al ., 2009 ; Matzek & Vitousek, 2009 ; Yu et al ., 2010 ).生态系统水平上对内稳性的研究多集中于海洋生态系统和陆地生态系统(Güsewell, 2004 ; 苏强, 2012 ), 对湿地生态系统的研究较少(Wang et al ., 2014 ).国内内稳性的研究还处于起步阶段.庾强(2009)以内蒙古草原为研究对象, 通过1 200 km的横断范围的样带试验和内蒙古草原生态系统定位研究站27年的监测数据发现, 内稳性高的物种具有较高的优势度和生物量稳定性, 内稳性高的生态系统则具有较高的生产力和稳定性.因此, 内稳性是衡量物种竞争力的一个重要指标.已有研究表明内稳性的高低受多种因子的影响, 其中包括植物器官(庾强, 2009 )、养分供应(严正兵等, 2013 )、光强以及施肥(Sterner & Elser, 2002 )等, 影响因子的不确定性使得不同研究中内稳性对环境的响应不一致.目前有关互花米草入侵对本地生态系统的影响多集中在物质循环以及生态化学计量学, 而物种水平上内稳性的研究甚少(张仁懿等, 2015 ).已有研究发现, 闽江河口湿地生长季阶段的入侵种互花米草和本地种短叶茳芏均具有一定的内稳性, 互花米草N和N:P的内稳性高于短叶茳芏N和N:P的内稳性, 而短叶茳芏P的内稳性高于互花米草P的内稳性(蒋利玲等, 2014 ). ...
Potential of Prokelisia spp. as biological control agents of english cordgrass, Spartina anglica
1
1999
... 目前, 互花米草的防治措施主要有物理控制(Frid et al. , 1999 )、化学控制(Patten, 2002 )和生物控制(Wu et al ., 1999 , Grevstad et al ., 2003 )这3种方法, 其中以收割、碎根水淹等物理方法居多.研究表明, 翻耕和碎根处理方法仅在当年有一定的控制效果, 不具有长期有效性, 且割除碎根也需要在最佳季节进行(李贺鹏和张利权, 2007 ).已有的研究认为: 闽江河口湿地生态系统在7月份时互花米草内稳性最低, 是治理互花米草的最佳时期(蒋利玲等, 2014 ).充分考虑植物内稳性特征的影响, 建议采用物理-化学综合方法进行互花米草的控制, 在7月份对互花米草进行碎根, 再通过施加N缓解短叶茳芏根和茎的养分限制, 从而增大短叶茳芏群落在湿地生态系统中的内稳性. ...
湿地碳氮磷的生物地球化学循环研究进展
1
2005
... 生物入侵作为全球变化的一个重要方面, 严重威胁了土著生物群落的生物多样性和生态系统功能(Davis, 2003 ; Hooper et al ., 2005 ).作为我国中亚热带和南亚热带过渡区最具代表性的河口湿地之一的闽江河口湿地(刘剑秋等, 2006 ), 自2002年以来逐渐遭到互花米草(Spartina alterniflora )的大规模入侵.互花米草以其极强的适应力和繁殖力迅速占据空生态位, 侵占本地种短叶茳芏(Cyperus malaccensis var. brevifolius )的生境, 在潮间带形成了短叶茳芏单种群落、互花米草单种群落和互花米草-短叶茳芏混生群落共存的竞争格局.入侵的互花米草通过影响土著生态系统的结构, 进而影响生态系统的养分循环, 生态系统养分的可利用性改变反过来又影响植物的生长状况, 从而可能改变入侵种与土著种之间的竞争关系.养分元素氮(N)和磷(P)是湿地生态系统植物生长的主要限制资源, 其可利用性及生态化学计量关系是植物生长和生态系统物质循环的重要调节因子(熊汉锋和王运华, 2005 ).植物能在外界养分元素组成改变的情况下, 一定程度上维持自身各元素之间的相对比例.这种由生物在长期进化过程中为适应外界环境变化而形成的维持自身化学组成相对稳定的能力就叫作生态化学计量内稳性(简称内稳性)(Kooijman, 1995 ). ...
氮磷施肥对拟南芥叶片碳氮磷化学计量特征的影响
1
2013
... 国外内稳性研究的对象由最初的真菌和细菌(Levi & Cowling, 1969 ; Goldman et al ., 1987 )发展到藻类、浮游动物和草本植物(Rhee, 1978 ; Andersen & Hessen, 1991 ; Güsewell et al ., 2005 ).由于高等植物体内元素的内稳性和化学计量特征复杂, 对高等植物的研究甚少(Cernusak et al ., 2009 ; Matzek & Vitousek, 2009 ; Yu et al ., 2010 ).生态系统水平上对内稳性的研究多集中于海洋生态系统和陆地生态系统(Güsewell, 2004 ; 苏强, 2012 ), 对湿地生态系统的研究较少(Wang et al ., 2014 ).国内内稳性的研究还处于起步阶段.庾强(2009)以内蒙古草原为研究对象, 通过1 200 km的横断范围的样带试验和内蒙古草原生态系统定位研究站27年的监测数据发现, 内稳性高的物种具有较高的优势度和生物量稳定性, 内稳性高的生态系统则具有较高的生产力和稳定性.因此, 内稳性是衡量物种竞争力的一个重要指标.已有研究表明内稳性的高低受多种因子的影响, 其中包括植物器官(庾强, 2009 )、养分供应(严正兵等, 2013 )、光强以及施肥(Sterner & Elser, 2002 )等, 影响因子的不确定性使得不同研究中内稳性对环境的响应不一致.目前有关互花米草入侵对本地生态系统的影响多集中在物质循环以及生态化学计量学, 而物种水平上内稳性的研究甚少(张仁懿等, 2015 ).已有研究发现, 闽江河口湿地生长季阶段的入侵种互花米草和本地种短叶茳芏均具有一定的内稳性, 互花米草N和N:P的内稳性高于短叶茳芏N和N:P的内稳性, 而短叶茳芏P的内稳性高于互花米草P的内稳性(蒋利玲等, 2014 ). ...
内蒙古草原植物化学计量生态学研究
1
2009
... 国外内稳性研究的对象由最初的真菌和细菌(Levi & Cowling, 1969 ; Goldman et al ., 1987 )发展到藻类、浮游动物和草本植物(Rhee, 1978 ; Andersen & Hessen, 1991 ; Güsewell et al ., 2005 ).由于高等植物体内元素的内稳性和化学计量特征复杂, 对高等植物的研究甚少(Cernusak et al ., 2009 ; Matzek & Vitousek, 2009 ; Yu et al ., 2010 ).生态系统水平上对内稳性的研究多集中于海洋生态系统和陆地生态系统(Güsewell, 2004 ; 苏强, 2012 ), 对湿地生态系统的研究较少(Wang et al ., 2014 ).国内内稳性的研究还处于起步阶段.庾强(2009)以内蒙古草原为研究对象, 通过1 200 km的横断范围的样带试验和内蒙古草原生态系统定位研究站27年的监测数据发现, 内稳性高的物种具有较高的优势度和生物量稳定性, 内稳性高的生态系统则具有较高的生产力和稳定性.因此, 内稳性是衡量物种竞争力的一个重要指标.已有研究表明内稳性的高低受多种因子的影响, 其中包括植物器官(庾强, 2009 )、养分供应(严正兵等, 2013 )、光强以及施肥(Sterner & Elser, 2002 )等, 影响因子的不确定性使得不同研究中内稳性对环境的响应不一致.目前有关互花米草入侵对本地生态系统的影响多集中在物质循环以及生态化学计量学, 而物种水平上内稳性的研究甚少(张仁懿等, 2015 ).已有研究发现, 闽江河口湿地生长季阶段的入侵种互花米草和本地种短叶茳芏均具有一定的内稳性, 互花米草N和N:P的内稳性高于短叶茳芏N和N:P的内稳性, 而短叶茳芏P的内稳性高于互花米草P的内稳性(蒋利玲等, 2014 ). ...
Linking stoichiometric homeostasis with ecosystem structure, functioning, and stability
1
2010
... 国外内稳性研究的对象由最初的真菌和细菌(Levi & Cowling, 1969 ; Goldman et al ., 1987 )发展到藻类、浮游动物和草本植物(Rhee, 1978 ; Andersen & Hessen, 1991 ; Güsewell et al ., 2005 ).由于高等植物体内元素的内稳性和化学计量特征复杂, 对高等植物的研究甚少(Cernusak et al ., 2009 ; Matzek & Vitousek, 2009 ; Yu et al ., 2010 ).生态系统水平上对内稳性的研究多集中于海洋生态系统和陆地生态系统(Güsewell, 2004 ; 苏强, 2012 ), 对湿地生态系统的研究较少(Wang et al ., 2014 ).国内内稳性的研究还处于起步阶段.庾强(2009)以内蒙古草原为研究对象, 通过1 200 km的横断范围的样带试验和内蒙古草原生态系统定位研究站27年的监测数据发现, 内稳性高的物种具有较高的优势度和生物量稳定性, 内稳性高的生态系统则具有较高的生产力和稳定性.因此, 内稳性是衡量物种竞争力的一个重要指标.已有研究表明内稳性的高低受多种因子的影响, 其中包括植物器官(庾强, 2009 )、养分供应(严正兵等, 2013 )、光强以及施肥(Sterner & Elser, 2002 )等, 影响因子的不确定性使得不同研究中内稳性对环境的响应不一致.目前有关互花米草入侵对本地生态系统的影响多集中在物质循环以及生态化学计量学, 而物种水平上内稳性的研究甚少(张仁懿等, 2015 ).已有研究发现, 闽江河口湿地生长季阶段的入侵种互花米草和本地种短叶茳芏均具有一定的内稳性, 互花米草N和N:P的内稳性高于短叶茳芏N和N:P的内稳性, 而短叶茳芏P的内稳性高于互花米草P的内稳性(蒋利玲等, 2014 ). ...
Stoichiometric homeostasis predicts plant species dominance, temporal stability, and responses to global change
2015
互花米草入侵对土壤生态系统的影响
1
2011
... 入侵对植物内稳性产生的影响主要表现在N元素上.混生群落中的互花米草根、叶和鞘的H N 分别显著高于对应单种群落中的互花米草根、茎和鞘的H N .混生群落中的互花米草根的H N 比单种群落中的互花米草根的H N 高25.7倍, 混生群落中的短叶茳芏根的H N 比单种群落中的短叶茳芏根的H N 增大了1.48倍, 两者之间的差异可能与互花米草根际微生物的固氮作用有关, 这使得互花米草获得更多的N, 进而缓解了N的限制(Bagwell & Lovell, 2000 ; Moisander et al ., 2005 ; 曾艳等, 2011 ).入侵初期, 单种群落的植物生长受N限制, 植物体内N的含量很大程度上取决于土壤中可利用N的含量, 故植物内稳性较低.随着入侵过程的进行, 互花米草的根际固氮作用使植物体内N含量的获取对土壤中N的依赖性降低, 从而使混生群落中的植物表现出较高的内稳性.相比短叶茳芏根的养分利用性, 互花米草根内稳性的调节能力使其能够在多变的环境中保持较好的生长, 进而具备更强的竞争力和适应力.已有的研究表明, 生态系统的稳定性与根生物量正相关, 根在变化的环境中可通过回收利用和贮存养分来调节生长, 从而提高物种和群落水平的生产力和稳定性(Tilman et al ., 2006 ).因此, 根内稳性与群落生产力和稳定性的关系研究值得进一步探索. ...
Differential responses of N:P stoichiometry of Leymus chinensis and Carex korshinskyi to N additions in a steppe ecosystem in Nei Mongol
1
2004
... 根据Garnier理论, 植物与土壤对应养分含量存在正相关关系则说明植物生长受到养分限制(Garnier, 1998 ).根据该理论, 福建省闽江河口鳝鱼滩湿地生态系统中短叶茳芏茎受N限制, 单种群落中的互花米草茎和鞘受N限制, 混生群落中的互花米草茎受N和P共同限制.根据Koerselman理论, 植物体N:P < 14表明其相对受N限制, N:P > 16相对受P限制, 14 < N:P < 16则受N和P共同限制(Koerselman & Meuleman, 1996 ; Güsewell & Koerselman, 2002 ).由于不同研究区域、不同生态系统以及不同植被类型的N:P临界阈值均会发生改变, 精确的N:P临界阈值需要通过严格的施肥试验来确定(Zhang et al ., 2004 ; Güsewell, 2004 ).胡伟芳等(2014)对中国主要湿地649种植物的N、P化学计量特征进行了整合分析, 结果表明不同湿地生境的植物受限制状况不尽相同, 其中N限制(N:P < 14)比例最高, 占59%.针对湿地生态系统的大多数研究也多用14和16为阈值来判断养分受限制情况(Güsewell & Koerselman, 2002 ; Tessier & Raynal, 2003 ; Bott et al ., 2008 ).因此, 若本研究沿用该临界阈值进行分析, 则单种群落和混生群落中的互花米草茎均受N、P共同限制, 其叶均受P限制, 单种群落和混生群落中的短叶茳芏茎均受N限制.本研究中, 互花米草各器官N、P含量平均值均低于短叶茳芏, 但N:P值平均值高于短叶茳芏, 这表明互花米草在生长过程中对养分的需求量相对较低, 但对N的利用效率高于对P的利用效率. ...
亚高寒草甸物种内稳性与生物量变化模式
1
2015
... 国外内稳性研究的对象由最初的真菌和细菌(Levi & Cowling, 1969 ; Goldman et al ., 1987 )发展到藻类、浮游动物和草本植物(Rhee, 1978 ; Andersen & Hessen, 1991 ; Güsewell et al ., 2005 ).由于高等植物体内元素的内稳性和化学计量特征复杂, 对高等植物的研究甚少(Cernusak et al ., 2009 ; Matzek & Vitousek, 2009 ; Yu et al ., 2010 ).生态系统水平上对内稳性的研究多集中于海洋生态系统和陆地生态系统(Güsewell, 2004 ; 苏强, 2012 ), 对湿地生态系统的研究较少(Wang et al ., 2014 ).国内内稳性的研究还处于起步阶段.庾强(2009)以内蒙古草原为研究对象, 通过1 200 km的横断范围的样带试验和内蒙古草原生态系统定位研究站27年的监测数据发现, 内稳性高的物种具有较高的优势度和生物量稳定性, 内稳性高的生态系统则具有较高的生产力和稳定性.因此, 内稳性是衡量物种竞争力的一个重要指标.已有研究表明内稳性的高低受多种因子的影响, 其中包括植物器官(庾强, 2009 )、养分供应(严正兵等, 2013 )、光强以及施肥(Sterner & Elser, 2002 )等, 影响因子的不确定性使得不同研究中内稳性对环境的响应不一致.目前有关互花米草入侵对本地生态系统的影响多集中在物质循环以及生态化学计量学, 而物种水平上内稳性的研究甚少(张仁懿等, 2015 ).已有研究发现, 闽江河口湿地生长季阶段的入侵种互花米草和本地种短叶茳芏均具有一定的内稳性, 互花米草N和N:P的内稳性高于短叶茳芏N和N:P的内稳性, 而短叶茳芏P的内稳性高于互花米草P的内稳性(蒋利玲等, 2014 ). ...
闽江河口区湿地景观格局演变研究
1
2006
... 本试验在福建省闽江河口鳝鱼滩湿地(26.01°- 26.06° N, 119.57°-119.68° E, 图1 )进行.该湿地为闽江河口区面积最大的砂泥质洲滩天然湿地.该区域气候暖热湿润, 年平均气温为19.3 ℃, 年降水量为 1 346 mm.土壤以滨海盐土、潮土以及水稻土等人工土壤为主.潮汐为正规半日潮.湿地内有维管植物109科294属408种(含变种), 本地优势种主要有短叶茳芏、芦苇(Phragmites australis )和藨草(Scirpus triqueter ).自2002年互花米草入侵以来, 其入侵斑块由零星分布发展到超过200万m2 的大面积分布, 已在潮间带和部分潮上带形成了明显的大面积入侵斑块(郑彩红等, 2006 ). ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}