碳(C)、氮(N)和磷(P)是植物基本营养元素, 在植物生长和各种生理调节机能中发挥着重要作用。C是构成植物体内干物质的最主要元素, 而N和P则分别与植物的光合作用和细胞分裂等重要生理活动有关(Niklas et al ., 2005 )。N:C和P:C与植物的相对生长率关系密切(Ågren, 2004 ), N:P则可以反映植物生长受N或P的限制情况(Koerselman & Meuleman, 1996 ; Elser et al ., 2000 ; Güsewell, 2004 )。植物不同组织器官营养元素水平与变化是调整它们的生长速率以适应环境变化的一种策略(Marschner, 1995 ), 如干旱区植物相对于湿润区植物通常拥有更高的单位面积(或质量)叶N、P含量(Wright et al ., 2001 ; 李玉霖等, 2010 ), 植物地下根系的N、P含量低于地上部分的叶片, 但二者具有相似的N:P化学计量比(Yuan et al ., 2011 )。当前, 区域C、N、P化学计量格局及其驱动因素仍然是生态化学计量学的一个重要研究领域, 其结果不仅有助于揭示物种间化学计量的趋同与分异, 对于预测生态系统功能、过程对全球变化的响应亦具有重要意义(贺金生和韩兴国, 2010 ; Elser et al ., 2010 )。近年来, 国内学者在不同时空尺度, 针对森林(刘万德等, 2010 ; 崔高阳等, 2015 )、草地(Yu et al ., 2010 ; 雒明伟等, 2015 )、荒漠(李玉霖等, 2010 ; 黄菊莹和余海龙, 2016 )、湿地(Wang et al ., 2015 ; 聂兰琴等, 2016 )等生态系统类型开展了大量的研究。然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al ., 2010 )。同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014 )。
土地沙化长期以来一直是我国最为严重的生态环境问题(董光荣等, 1999 ), 不仅出现在我国北方干旱、半干旱地区, 亦出现于南方湖滨及河流下游冲积平原地区(朱震达和崔书红, 1996 )。以鄱阳湖区为例, 沙化土地面积多达3.89万hm2 (左长青, 1986 )。鄱阳湖沙化土地虽处于亚热带湿润区, 但水热不同期, 特别是8-10月的高温、少雨与地表强蒸发, 以及冬春季节固定的风向与强劲的风速是影响植被恢复的重要因素。近几十年来, 马鞭草科落叶小灌木单叶蔓荆(Vitex rotundifolia )被成功地应用于鄱阳湖沙化土地的植被恢复, 并且在不同沙化程度区形成稳定的群落。研究表明生物有机体具有极高的化学计量内稳性(Elser et al ., 2010 ), 并且通过调节生物对环境变化的响应, 化学计量内稳性成为生态系统结构、功能和稳定性维持的重要机制(Yu et al ., 2010 )。单叶蔓荆的养分及其化学计量关系在不同沙化程度区会发生怎样的变化?其N、P功能性状之间的关系是否因沙化梯度而发生策略位移?本研究通过测定鄱阳湖典型沙山重度、中度、轻度沙化区单叶蔓荆花、叶、枝条、匍匐茎、细根等部位的C、N、P含量, 分析各器官养分元素分配格局、化学计量比的差异与关联, 以及随沙化程度的变化, 试图探讨上述两个科学问题, 为进一步揭示单叶蔓荆的沙地适应机制提供科学依据, 为鄱阳湖沙化区植被恢复提供参考。
1 材料和方法
1.1 研究区概况
研究区设置在鄱阳湖沙化土地的典型分布区——江西省都昌县多宝乡鄱阳湖湖滨沙山, 地理位置29.36°-29.46° N, 116.05°-116.13° E。该区属于亚热带湿润性季风气候, 年降水量1β310 mm, 年蒸发量1β883 mm, 日照充足。多宝沙山主要是由冬季风吹蚀湖滨沙地堆积于近湖滨的一级阶地而形成的。依据野外实地调查, 参考沙丘分布形态、地表裸露程度、植被覆盖度、土壤有机质、氮磷养分等指标, 研究区沙山可以划分为3个沙化梯度, 其中湖滨 800 m范围内为重度沙化区, 800-2β000 m为中度沙化区, 2β000 m之外为轻度沙化区。重度沙化区主要由流动沙丘、半流动沙丘组成, 呈面状、条带状分布;中度沙化区地表裸露呈斑块状, 主要为半固定沙丘;轻度沙化区以固定沙丘为主, 地表裸露呈斑点状零散分布。3种不同沙化程度区土壤(0-10 cm)、植被基本特征见表1 。
1.2 样品采集与分析
2014年7月自湖滨至沙山边缘, 沿沙化梯度设置长度约3β000 m宽度约200 m的样带, 样带起点为湖岸线水陆交界处。沿样带利用手持GPS测距, 在离湖岸线100 m处的湖滨沙地设置了第一个采样点, 然后每隔400 m (误差15 m左右)增加一个采样点, 共计8个采样点。在每个采样点设置4个采样小区, 于每个采样区随机选择8-10株未发现病虫害、生长良好且正值花期的单叶蔓荆, 在各采样植株处用小铁锹挖掘至0-40 cm土壤深度, 然后采集保持不同器官的完整植株, 所采植株混合在一起作为1个重复, 每个采样点采集4个重复。同时, 在上述植物采样点以多点混合的方法, 采集土壤0-10 cm、10-30 cm样品, 以分析土壤有机碳、全N、全P等养分。将取出的完整植株迅速运回实验室, 分离花、叶片、枝条、匍匐茎、细根(直径<2 mm)。用恒温烘箱在65 ℃下烘至恒质量, 称质量并磨碎, 以四分法取其中一部分进行全C、全N、全P的测定。其中全C、全N采用EA3000元素分析仪(EuroVector, Redavalle, Italy)测定, 全P采用钼锑抗比色法测定。土壤样品风干磨细后, 有机碳采用高锰酸钾氧化外加热法测定, 全N、全P测定方法同植物。上述测试结果均以单位质量的养分含量表示(mg·g-1 )。
1.3 数据处理与分析
采用Excel 2003和Origin 9.0进行数据处理与制图, 利用SPSS 19.0软件中单因素方差分析方法对3种沙化程度区的土壤、植被特征, 以及单叶蔓荆各器官养分元素含量的差异进行比较。采用GLM模型中的双因素方差方法分析沙化程度和不同器官对植物C、N、P含量及其化学计量比的影响。采用协方差分析方法比较3种沙化程度区植物N mass -P mass 线性关系的斜率与截距的差异, 上述分析均采用LSD 多重比较不同组的差异;利用Pearson相关系数及线性回归模型分析各器官间C、N、P含量及化学计量比之间的关系, 以及与土壤养分之间的关系。文中显著性水平设置为α = 0.05。
2 结果和分析
2.1 单叶蔓荆各器官C、N、P含量及化学计量比
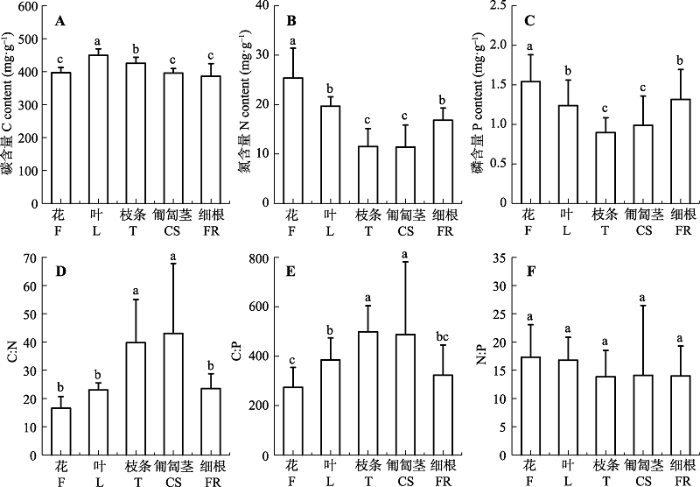
C在单叶蔓荆各器官中的变化范围为386.28- 449.47 mg·g-1 。N、P含量变化范围则分别为11.40-25.37 mg·g-1 、0.89-1.54 mg·g-1 。3种元素中以N在各器官间的变异最大, 变异系数达到30.4%。C在各器官间的变异系数最小, 仅为5.7%。单叶蔓荆不同器官间的C、N、P含量均存在显著差异(图1A -1C), 其中花的N、P含量显著高于其他器官;匍匐茎和枝条的N、P含量显著低于其他器官。总体而言, 单叶蔓荆各器官N、P平均含量表现为花>叶>细根>匍匐茎>枝条。各器官C:N、C:P、N:P值分别在16.56- 43.05、273.85-498.20、13.84-17.31之间波动, 其中C:N、C:P在各器官间差异显著(p < 0.05), C:N平均值匍匐茎>枝条>细根>叶>花, C:P平均值枝条>匍匐茎> 叶>细根>花。N:P则保持相对稳定, 在各器官间未达到显著性差异(图1D -1F)。
图1 单叶蔓荆各器官碳、氮、磷含量及化学计量比(平均值±标准偏差)。不同小写字母表示各器官间差异显著(p < 0.05)。
Fig. 1 Carbon (C), nitrogen (N) and phosphorus (P) contents and their stoichiometry in various organs of Vitex rotundifolia (mean ± SD ). F, flower; L, leaf; T, twig; CS, creeping stem; FR, fine root. Different lowercase letters indicate significant differences between organs at p < 0.05 levels.
2.2 单叶蔓荆各器官C、N、P含量及化学计量比相关性
单叶蔓荆各器官C、N、P含量相关性多数未达到显著性水平(表2 )。相关性达到显著水平的元素表现为: 叶与花的C含量为极显著正相关(p < 0.01), 花和叶与枝条C之间相关性一致, 均为极显著负相关(p < 0.01), 匍匐茎和枝条C亦为显著负相关(p < 0.05)。花与枝条的N呈极显著正相关(p < 0.01), 而花和枝条与匍匐茎N相关性一致, 均为极显著负相关(p < 0.01)。匍匐茎与枝条的P含量为极显著正相关(p < 0.01), 而细根与枝条的P含量显著负相关(p < 0.05)。
各器官间N:P化学计量比的相关性均未达到显著性水平。C:N化学计量比在花与枝条之间表现为极显著正相关(p < 0.01), 而花、枝条与匍匐茎之间则表现为显著负相关(p < 0.05)。叶、枝条与匍匐茎的C:P化学计量比均为显著正相关(p < 0.05), 细根与枝条C:P则为显著负相关(p < 0.05)。各器官中以枝条与匍匐茎之间的关系最为密切, 二者之间的C、N、P含量及C:N、C:P化学计量比均表现为显著性相关。
2.3 单叶蔓荆各器官C、N、P含量随沙化梯度的变化
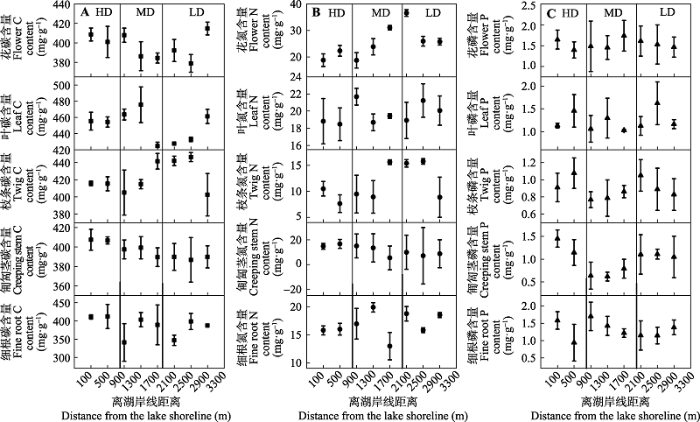
单叶蔓荆各器官C含量随着沙化程度的变化发生较大的波动, 在5种器官中, 除了匍匐茎表现出显著的下降趋势外, 其他器官的波动没有表现出一致的规律(图2A )。多数器官N含量随着沙化程度的降低表现出增加趋势, 且最低值多出现于湖岸线边缘500 m范围内的重度沙化区, N含量的高值则多分布于离湖岸线2β000 m之外的轻度沙化区。匍匐茎N含量与其他器官相比表现出下降的趋势(图2B )。各器官中P含量随沙化程度的变化没有明显规律, 变异系数范围为7.4%-28.9% (图2C )。
图2 单叶蔓荆各器官碳(C)、氮(N)、磷(P)含量随沙化程度变化(平均值±标准偏差)。HD、MD和LD同表1 。
Fig. 2 Changes of carbon(C), nitrogen(N) and phosphorus(P) contents in various organs of Vitex rotundifolia along desertification gradient (mean ± SD ). HD, MD and LD see Table 1 .
2.4 单叶蔓荆N mass -P mass 关系沿沙化梯度的变化
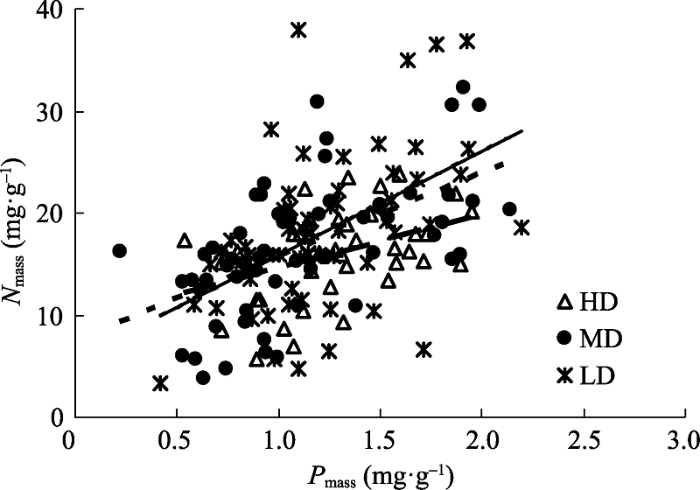
N mass -P mass 间呈极显著正相关关系(r = 0.49, p < 0.001, n = 160), 且重度、中度、轻度沙化区单叶蔓荆N、P含量之间均呈显著线性关系(p < 0.01)(图3 )。N mass -P mass 之间一元线性关系的斜率表现为轻度沙化区高于中度与重度沙化区, 截距则反之, 表现为重度沙化区高于中度与轻度沙化区。协方差分析表明3种沙化程度区N mass -P mass 线性关系的斜率与截距均未达到显著性差异(表3 )。
图3 不同沙化程度区单叶蔓荆基于质量的氮含量(N mass )和磷含量(P mass )之间的关系。HD、MD和LD同表1 。
Fig. 3 Relationship between mass-based nitrogen content (N mass ) and mass-based phosphorus content (P mass ) of Vitex rotundifolia at different desertification zones. HD, MD and LD see Table 1 .
3 讨论
3.1 单叶蔓荆不同器官养分含量与化学计量比分布特征
单叶蔓荆作为鄱阳湖湖滨沙山植被恢复过程的先锋物种, 广布于沙山不同程度沙化区, 为进一步说明研究区单叶蔓荆不同器官的养分含量及其化学计量关系特征, 我们把单叶蔓荆地上部分的叶片、地下部分的细根与其他荒漠区植物, 以及全球、中国大样本研究中的植物养分平均值做了比较。本研究区单叶蔓荆叶N (19.66 mg·g-1 )、P (1.24 mg·g-1 )含量明显低于中国北方典型荒漠及荒漠化地区植物N (24.45 mg·g-1 )、P (1.74 mg·g-1 )含量(李玉霖等,2010 )。此外, 叶N含量亦低于Killingbeck和Whitford (1996)报道的全球5大洲11个干旱荒漠中78种灌木叶N含量平均值(22 mg·g-1 )。本研究区虽属于南方典型荒漠, 但地处亚热带湿润区, 年降水量远高于其他干旱区荒漠, 这可能是单叶蔓荆叶N、P含量低于其他干旱、半干旱区荒漠植物的重要原因, 因为湿润区植物相对于干旱区通常具有更低的叶N、P含量(Wright et al ., 2001 ; 李永华等, 2005 )。
Kattge等(2011)搜集整理了全球植物性状数据库, 发现全球植物叶片C、N、P含量的平均值分别为476.1 mg·g-1 、17.4 mg·g-1 、1.23 mg·g-1 , C:N、N:P则分别为23.4、12.8。与上述全球叶性状数据库相比较, 单叶蔓荆叶C (449.5 mg·g-1 )含量偏低, 叶N (19.66 mg·g-1 )含量略高, 叶P (1.24 mg·g-1 )含量则高度一致; 叶片C:N (23.1)基本一致, N:P (16.8)明显偏高。单叶蔓荆细根C (386.3 mg·g-1 )、N (16.8 mg·g-1 )、P (1.32 mg·g-1 )含量与Yuan等(2011)报道的全球植物细根养分含量及化学计量比相比, C含量与C:N明显偏低, N、P含量明显偏高, 但N:P化学计量比基本一致。同样, 与马玉珠等(2015)报道的中国植物细根的C、N、P含量相比, 我们也发现本研究中的单叶蔓荆细根C含量明显偏低, N、P含量明显偏高。与全球及中国大样本数据相比, 单叶蔓荆的叶片及细根的N、P含量, 特别是N含量明显偏高, 一方面可能与上述数据库中的多数样本来自于森林中的乔木有关, 同时还可能与本研究区单叶蔓荆特殊的荒漠生境相关。研究区虽不具有干旱区荒漠的长期水分限制, 但因为水热不同期, 特别是夏秋季节的高温、少雨与地表的强蒸发, 容易形成季节性的水分胁迫。同时, 沙地土壤养分匮乏, 单叶蔓荆在水分与养分均存在限制的情形下增加叶片N投入可能是一种有效的适应策略, 因为叶N增加能够提升叶片内部光合作用酶的数量, 进一步提高叶片光合速率(Chapin et al ., 1987 )。此外, 叶N含量的增加还可以增加叶片内部非光合器官或组织氮的投入, 从而提高细胞内部的渗透压, 增强对植物体内水分的保护, 是一种有效的节水方式(Wright et al ., 2001 )。
N、P养分元素在单叶蔓荆各器官中的分配存在显著差异(图1 ), 表现为花的N、P含量最高, 叶与细根次之, 枝条与匍匐茎之间没有显著差异且显著低于其他器官。植物不同器官营养元素的分布, 除受土壤养分元素的供给影响之外, 主要与相应器官的生理功能密切相关(Kerkhoff et al ., 2006 )。采样时单叶蔓荆正处于花期, 作为重要的繁殖器官, 花富含线粒体, 且吸收储存大量营养物质为果实形成做准备, 因此具有最高的N、P含量。叶片是植物进行光合作用的器官, 保持较高的N含量有利于提高光合速率。同时, 叶片中存储较多的N、P也是对特殊生境的一种适应策略。根系是吸收营养物质的主要器官, 根系特别是细根具有较高的N、P含量。匍匐茎和枝条主要起水分与养分输导及支撑植物的作用, 因此, 养分元素的含量相对较低。
近年来, 越来越多的研究尝试从植物功能性状或资源经济谱的角度来探讨不同器官之间的性状关联, 相关研究不仅有助于理解植物各性状之间相互作用的机制(Eviner & Chapin, 2003 )、植物生长过程中对资源的利用和分配(Kerkhoff et al ., 2006 ), 还对进一步预测植物对环境变化的响应具有重要意义。本研究发现单叶蔓荆各器官的养分元素及化学计量关系的相关性主要集中在地上部分, 即花、叶、枝条、匍匐茎之间, 其中尤以枝条与匍匐茎之间表现明显, 二者除了N:P没有达到显著相关之外, 其他的养分元素及其化学计量关系均达到了显著性相关(表2 ), 显示了二者之间在养分存储与运输之间的密切关系。在植物的所有器官中, 叶片与细根之间的性状关联最受关注。一些大尺度多物种间的研究表明叶片和细根的N含量(Craine & Lee, 2003 ; Tjoelker et al ., 2005 )、P含量(Kerkhoff et al ., 2006 ; Geng et al ., 2014 )均呈显著正相关。本研究未发现叶片与细根之间的3种元素及其化学计量关系显著相关, 一个可能的原因是叶片与细根性状的关联主要发生在种间, 小尺度上植物生境条件相对一致, 种内个体间功能性状因变异幅度较小而关联不明显(徐冰等, 2010 )。另一种可能的原因是本研究中的细根样品的采集是以直径<2 mm为标准的。由于根系在发育过程中形成明显的分枝结构, 具有高度的形态异质性(Pregitzer et al ., 2002 ; 王政权和郭大立, 2008 ), 直径<2 mm的细根并不是一个均质的单一组分, 而是由形态、寿命、生理差异巨大的不同个体根组成的高度异质的混合体(Pregitzer et al ., 2002; Guo et al ., 2008 ), 从而影响了与叶片之间相应元素的相关关系。
3.2 单叶蔓荆养分含量与化学计量比的影响因子
通过沙化程度和各器官对单叶蔓荆C、N、P含量及其化学计量关系影响的一般线性模型(GLM)分析发现: 沙化程度对单叶蔓荆的C、N含量影响显著, 对P含量的影响未达到显著性水平。相应的C:N和N:P受沙化程度的影响不显著, 但C:P受沙化程度影响显著。各器官这一因子除对N:P影响不显著外, 对其他元素及化学计量关系的影响都极为显著(p < 0.01)。沙化程度与各器官的交互作用对所有元素及其化学计量关系均有显著影响(表4 )。
植物体营养元素含量及化学计量关系不仅受植物的进化史、生活史、遗传、不同器官的生理功能等因素影响, 同时还受到土壤营养元素的供给及其在植物体内的运输、储藏、利用等过程的影响。本研究中除了匍匐茎N含量与土壤0-10 cm全N含量呈显著负相关之外, 未发现单叶蔓荆其他器官与土壤表层0-10 cm、10-30 cm之间相应养分元素之间的相关关系(表5 )。
上述GLM分析结果表明: 相对于沙化程度的变化所引起的植物生境条件, 如土壤水分、养分供给的变化, 不同器官的生理功能差异是影响单叶蔓荆C、N、P含量及化学计量比的主导因子。化学计量内稳性(stoichiometric homoeostasis)是生态化学计量学存在的基础(Sterner & Elser, 2002 )。Elser等(2010)认为尽管C、N、P化学计量比具有较大的时空变异, 但生物有机体仍保持极高的化学计量内稳性。化学计量内稳性通过调节生物对环境因子的响应, 成为生态系统结构、功能和稳定性维持的重要机制(Yu et al ., 2010 )。本研究中3种元素的化学计量关系中C:N、N:P在不同沙化程度区均未表现出显著差异。这一结果表明单叶蔓荆能够在沙山不同沙化区形成稳定的群落并保持较高的优势度, 化学计量内稳性很可能是其中非常重要的维持机制, 但这一机制仍然有待于进一步进行植物养分与土壤养分, 特别是速效养分之间关系的动态监测来加以验证。以往研究表明植物叶片与细根具有相似的N:P化学计量比(Yuan et al ., 2011 ), 同样, 本研究也发现N、P养分元素虽然在单叶蔓荆不同器官中的含量具有显著差异, 但是具有相似的N:P化学计量比, 保持了相对稳定性。
3.3 沙化程度对单叶蔓荆N mass -P mass 关系的影响
由于降水、土壤养分等环境梯度而引起的植物功能性状之间关系的策略位移现象不仅出现在种间水平上(Wright et al ., 2001 ; 李永华等, 2005 ; Wei et al ., 2011 ), 也出现在种内水平上(Wei et al ., 2011 )。但这一现象是否具有普遍性目前仍不清楚, 例如, 胡梦瑶等(2012)并没有发现紫花针茅(Stipa purpurea )在不同降水梯度下具有明显的策略位移现象。此外, 上述功能性状的位移现象多出现于较大尺度下的环境梯度中, 局地尺度上是否也存在类似的现象仍缺乏研究。本研究区尽管重度、中度与轻度沙化区土壤有机质、全氮、全磷等养分, 以及地表植被覆盖度等存在显著差异(表1 ), 但未发现单叶蔓荆N mass -P mass 线性关系的斜率与截距在3种沙化程度区发生显著变化(表3 ), 表明单叶蔓荆这一对功能性状关系在不同沙化区未发生明显的位移现象。成对植物功能性状关系发生策略位移主要是由于环境梯度对其中一个功能性状影响更大, 例如, Wei等(2011)发现低降水区的油蒿(Artemisia ordosica )种群N mass 高于较高降水区, 但比叶面积差异不大。本研究中N mass -P mass 在不同沙化程度区未出现明显的策略位移现象, 表明土壤沙化所引起的水、肥等限制因素可能共同作用于这两个性状, 使得植物N mass 与P mass 发生同步变化, 同时这也进一步佐证了N:P化学计量关系在不同沙化区的相对稳定。此外, 本研究结果也表明局地尺度种内水平上很可能因为环境梯度过小而不具备发生N mass -P mass 关系的位移现象。
4 结论
单叶蔓荆不同器官间C、N、P含量存在显著差异, 其中花的N、P含量显著高于其他器官。各器官中以枝条与匍匐茎之间的关系最为密切, 二者之间的C、N、P含量及C:N、C:P化学计量比均表现为显著性相关。沙化程度仅对C、N含量及C:P影响显著, 不同器官是影响单叶蔓荆C、N、P含量及化学计量比的主导因子。单叶蔓荆N:P化学计量比具有保守性特征, 没有随沙化程度与器官的改变而发生显著变化, N mass -P mass 关系在不同沙化程度区未发生位移现象。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Ågren GI 2004 ). The C:N:P stoichiometry of autotrophs-theory and observations
.Ecology Letters 7 , 185 -191 .
[本文引用: 1]
[2]
Chapin FS Bloom AJ Field CB Waring RH 1987 ). Plant responses to multiple environmental factors
.BioScience 37 , 49 -57 .
[本文引用: 1]
[3]
Craine JM Lee WG 2003 ). Covariation in leaf and root traits for native and non-native grasses along an altitudinal gradient in New Zealand
.Oecologia 134 , 471 -478 .
[本文引用: 1]
[4]
Cui GY Cao Y Chen YM 2015 ). Characteristics of nitrogen and phosphorus stoichiometry across components of forest ecosystem in Shaanxi Province
.Chinese Journal of Plant Ecology 39 , 1146 -1155 . (in Chinese with English abstract)[崔高阳 , 曹扬 , 陈云明 (2015 ). 陕西省森林各生态系统组分氮磷化学计量特征
. 植物生态学报 , 39 , 1146 -1155 .]
[本文引用: 1]
[5]
Dong GR Wu B Ci LJ Zhou HS Lu Q Luo B 1999 ). Present situation, causes and control way of desertification in China
.Journal of Desert Research 19 (4 ), 22 -36 . (in Chinese with English abstract)[董光荣 , 吴波 , 慈龙骏 , 周欢水 , 卢琦 , 罗斌 (1999 ). 我国荒漠化现状、成因与防治对策
. 中国沙漠 , 19 (4 ), 22 -36 .]
[本文引用: 1]
[6]
Elser JJ Fagan WF Denno RF Dobberfuhl DR Ayoola F Andrea H Sebastian I Kilham SS Edward M Schulz KL Siemann EH Sterner RW 2000 ). Nutritional constraints in terrestrial and freshwater food webs
.Nature 408 , 578 -580 .
[本文引用: 1]
[7]
Elser JJ Fagan WF Kerkhoff AJ Swenson NG Enquis BJ 2010 ). Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change
.New Phytologist 186 , 593 -608 .
[本文引用: 3]
[8]
Eviner VT Chapin FS III 2003 ). Functional matrix: A conceptual framework for predicting multiple plant effects on ecosystem processes
.Ecology, Evolution and Systematics 34 , 455 -485 .
[本文引用: 1]
[9]
Geng Y Wang L Jin DM Liu HY He JS 2014 ). Alpine climate alters the relationships between leaf and root morphological traits but not chemical traits
.Oecologia 175 , 445 -455 .
[本文引用: 1]
[10]
Guo DL Li H Mitchell RJ Han WX Hendricks JJ Fahey TJ Hendrick RL 2008 ). Heterogeneity by root branch order: Exploring the discrepancy in root longevity and turnover estimates between minirhizotron and C isotope methods
.New Phytologist 177 , 443 -456 .
[本文引用: 1]
[11]
Güsewell S 2004 ). N:P ratios in terrestrial plants: Variation and functional significance
.New Phytologist 164 , 243 -266 .
[本文引用: 1]
[12]
He JS Han XG 2010 ). Ecological stoichiometry: Searching for unifying principles from individuals to ecosystems
.Chinese Journal of Plant Ecology 34 , 2 -6 . (in Chinese with English abstract)[贺金生 , 韩兴国 (2010 ). 生态化学计量学: 探索从个体到生态系统的统一化理论
. 植物生态学报 , 34 , 2 -6 .]
[本文引用: 1]
[13]
Hu MY Zhang L Luo TX Shen W 2012 ). Variations in leaf functional traits of Stipa purpurea along a rainfall gradient in Xizang, China
. Chinese Journal of Plant Ecology , 36 , 136 -143 . (in Chinese with English abstract)[胡梦瑶 , 张林 , 罗天祥 , 沈维 (2012 ). 西藏紫花针茅叶功能性状沿降水梯度的变化
. 植物生态学报 , 36 , 136 -143 .]
[14]
Huang JY Yu HL 2016 ). Responses of growth of four desert species to different N addition levels
.Chinese Journal of Plant Ecology 40 , 165 -176 . (in Chinese with English abstract)[黄菊莹 , 余海龙 (2016 ). 四种荒漠草原植物的生长对不同氮添加水平的响应
. 植物生态学报 , 40 , 165 -176 .]
[本文引用: 1]
[15]
Kattge J Diaz S Lavorel S Prentice LC Leadley P Garnier E Westoby M Reich PB Wright IJ Cornelissen JHC Violle C Harrison SP Bodegom PMV Reichstein M Enquist BJ Soudzilovskaia NA Ackerly DD Anand M Atkin O Bahn M Baker TR Baldocchi D Bekker R Blanco CC Blonder B Bond WJ Bradstock R Bunker DE Casanoves F Cavender BJ Chambers JQ Chapin FS Chave J Coomes D Cornwell WK Craine JM Dobrin BH Duarte L Durka W Elser J Esser G Estiarte M Fagan WF Fang J Fidelis GT Fyllas NM Gallagher RV Green WA Gutierrez AG Hickler T Higgins SI Hodgson GJ Jalili A Jansen S Joly CA Kerkhoff AJ Kirkup D Kitajima K Kleyer M Klotz S Knops JMH Kramer K Kurokawa H Laughlin D Lee TD Leishman M Lens F Lenz T Lewis SL Lloyd J Louault F Ma S Mahecha MD Manning P Massad P Massad T Medlyn BE Messier J Moles AT Nadrowski K Naeem S Ogaya R Oleksyn J Onipchenko VG Onoda Y Overbeck G Ozinga WA Paula S Pausas JG Phillips OL Pillar V Poorter H Poorter L Poschlod P Prinzing A Proulx R Rammig A Reinsch S Reu B Sack L Salgado NB Sardans J Shiodera S Shipley B Siefert A Sosinski E Swaine E Swenson N Thompson K Thornton P Waldram M Weiher E White M Wright SJ Yguel B Zaehle S Zanne AE Wirth C 2011 ). TRY—A global database of plant traits
.Global Change Biology 17 , 2905 -2935 .
[16]
Kerkhoff AJ Fagan WF Elser JJ Enquist BJ 2006 ). Phylogenetic and growth form variation in the scaling of nitrogen and phosphorus in the seed plants
.The American Naturalist 168 , 103 -122 .
[本文引用: 3]
[17]
Killingbeck KT Whitford WG 1996 ). High foliar nitrogen in desert shrubs: An important ecosystem trait or defective desert doctrine
.Ecology 77 , 1728 -1737 .
[18]
Koerselman W Meuleman AFM 1996 ). The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation
.Journal of Applied Ecology 33 , 1441 -1450 .
[本文引用: 1]
[19]
Li YH Luo TX Lu Q Tian XY Wu B Yang HH 2005 ). Comparisons of leaf traits among 17 major plant species in Shazhuyu Sand Control Experimental Station of Qinghai Province
.Acta Ecology Sinica 25 , 994 -999 . (in Chinese with English abstract)[李永华 , 罗天祥 , 卢琦 , 田晓娅 , 吴波 , 杨恒华 (2005 ). 青海省沙珠玉治沙站17种主要植物叶性因子的比较
. 生态学报 , 25 , 994 -999 .]
[本文引用: 2]
[20]
Li YL Mao W Zhao XY Zhang TH 2010 ). Leaf nitrogen and phosphorus stoichiometry in typical desert and desertified regions, North China
.Environment Science 31 , 1716 -1725 . (in Chinese with English abstract)[李玉霖 , 毛伟 , 赵学勇 , 张铜会 (2010 ). 北方典型荒漠及荒漠化地区植物叶片氮磷化学计量特征研究
. 环境科学 , 31 , 1716 -1725 .]
[本文引用: 3]
[21]
Liu WD Su JR Li SF Zhang ZJ Li ZG 2010 ). Stoichiometry study of C, N and P in plant and soil at different successional stages of monsoon evergreen broad-leaved forest in Pu’er, Yunnan Province
.Acta Ecologica Sinica 30 , 6581 -6590 . (in Chinese with English abstract)[刘万德 , 苏建荣 , 李帅锋 , 张志钧 , 李忠文 (2010 ). 云南普洱季风常绿阔叶林演替系列植物和土壤C、N、P化学计量特征
. 生态学报 , 30 , 6581 -6590 .]
[本文引用: 1]
[22]
Luo MW Mao L Li QQ Zhao X Xiao Y Jia TT Guo ZG 2015 ). C, N, and P stoichiometry of plant and soil in the restorable plant communities distributed on the land used for Qinghai-Tibet Highway Construction in the Qinghai-Tibetan Plateau, China
.Acta Ecologica Sinica 35 , 7832 -7841 . (in Chinese with English abstract)[雒明伟 , 毛亮 , 李倩倩 , 赵旭 , 肖玉 , 贾婷婷 , 郭正刚 (2015 ). 青藏高原筑路取土迹地恢复植物群落与土壤的碳氮磷化学计量特征
. 生态学报 , 35 , 7832 -7841 .]
[本文引用: 1]
[23]
Ma YZ Zhong QL Jin BJ Lu HD Guo BQ Zheng Y Li M Cheng DL 2015 ). Spatial changes and influencing factors of fine root carbon, nitrogen and phosphorus stoichiometry of plants in China
.Chinese Journal of Plant Ecology 39 , 159 -166 . (in Chinese with English abstract)[马玉珠 , 钟全林 , 靳冰洁 , 卢宏典 , 郭炳桥 , 郑媛 , 李曼 , 程栋梁 (2015 ). 中国植物细根碳、氮、磷化学计量学的空间变化及其影响因子
. 植物生态学报 , 39 , 159 -166 .]
[24]
Marschner H 1995 ). Mineral Nutrition of Higher Plants . Academic Press , London .
[本文引用: 1]
[25]
Nie LQ Wu Q Yao B Fu S Hu QW 2016 ). Leaf litter and soil carbon, nitrogen, and phosphorus stoichiometry of dominant plant species in the Poyang Lake wetland
.Acta Eclogica Sinica 36 , 1898 -1906 . (in Chinese with English abstract)[聂兰琴 , 吴琴 , 尧波 , 付姗 , 胡启武 (2016 ). 鄱阳湖湿地优势植物叶片-凋落物-土壤碳氮磷化学计量特征
. 生态学报 , 36 , 1898 -1906 .]
[本文引用: 1]
[26]
Niklas KJ Owens T Reich PB Cobb ED 2005 ). Nitrogen phosphorus leaf stoichiometry and the scaling of plant growth
.Ecology Letters 8 , 636 -642 .
[本文引用: 1]
[27]
Pregitzer KS Deforest JL Burton AJ Allen ME Ruess RW Hendrick RL 2002 ). Fine root architecture of nine North American trees
. Ecological Monographs 72 , 293 -309 .
[本文引用: 1]
[28]
Reich PB 2014 ). The world-wide “fast-slow” plant economics spectrum: A traits manifesto
.Journal of Ecology 102 , 275 -301 .
[本文引用: 1]
[29]
Sterner RW Elser JJ 2002 ). Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere . Princeton University Press , Princeton, USA .
[本文引用: 1]
[30]
Tjoelker MG Craine JM Wedin D Reich PB Tilman D 2005 ). Linking leaf and root trait syndromes among 39 grassland and savannah species
.New Phytologist 167 , 493 -508 .
[本文引用: 1]
[31]
Wang WQ Wang C Sardans J Zeng CS Tong C Penuelas J 2015 ). Plant invasive success associated with higher N-use efficiency and stoichiometric shifts in the soil-plant system in the Minjiang River tidal estuarine wetlands of China
.Wetlands Ecology and Management 23 , 865 -880 .
[本文引用: 1]
[32]
Wang ZQ Guo DL 2008 ). Root ecology
. Journal of Plant Ecology (Chinese Version ) , 32 , 1213 -1216 . (in Chinese)[王政权 , 郭大立 (2008 ). 根系生态学
. 植物生态学报 , 32 , 1213 -1216 .]
[本文引用: 1]
[33]
Wei H Wu B Yang W Luo T 2011 ). Low rainfall-induced shift in leaf trait relationship within species along a semi-arid sandy land transect in northern China
.Plant Biology 13 , 85 -92 .
[本文引用: 2]
[34]
Wright IJ Reich PB Westoby M 2001 ). Strategy shifts in leaf physiology, structure and nutrient content between species of high -and low-rainfall and high -and low-nutrient habitats
.Functional Ecology 15 , 423 -434 .
[本文引用: 4]
[35]
Xu B Cheng YX Gan HJ Zhou WJ He JS 2010 ). Correlations between leaf and fine root traits among and within species of typical temperate grassland in Xilin River Basin, Inner Mongolia, China
.Chinese Journal of Plant Ecology 34 , 29 -38 . (in Chinese with English abstract)[徐冰 , 程雨曦 , 甘慧洁 , 周文嘉 , 贺金生 (2010 ). 内蒙古锡林河流域典型草原植物叶片与细根性状在种间及种内水平上的关联
. 植物生态学报 , 34 , 29 -38 .]
[本文引用: 1]
[36]
Yu Q Chen QS Elser JJ He NP Wu HH Zhang GM Wu JG Bai YF Han XG 2010 ). Linking stoichiometric homoeostasis with ecosystem structure, functioning and stability
.Ecology Letters 13 , 1390 -1399 .
[本文引用: 3]
[37]
Yuan ZY Chen HYH Reich PB 2011 ). Global-scale latitudinal patterns of plant fine-root nitrogen and phosphorus
.Nature Communications 2 , 1 -6 .
[本文引用: 2]
[38]
Zhu ZD Cui SH 1996 ). The problem of land desertification in South China
.Journal of Desert Research 16 , 331 -337 . (in Chinese with English abstract)[朱震达 , 崔书红 . 中国南方的土地荒漠化问题 (1996 ). 中国沙漠
, 16 , 331 -337 .]
[本文引用: 1]
[39]
Zuo CQ 1986 ). Analysis for the causes, utilization and governance in sand hill, Poyang Lake
.Soil and Water Conservation 4 ), 2 -5 . (in Chinese)[左长青 (1986 ). 鄱阳湖沙山成因及治理利用浅析
. 中国水土保持 , (4 ), 2 -5 .]
[本文引用: 1]
The C:N:P stoichiometry of autotrophs-theory and observations
1
2004
... 碳(C)、氮(N)和磷(P)是植物基本营养元素, 在植物生长和各种生理调节机能中发挥着重要作用.C是构成植物体内干物质的最主要元素, 而N和P则分别与植物的光合作用和细胞分裂等重要生理活动有关(Niklas et al ., 2005 ).N:C和P:C与植物的相对生长率关系密切(Ågren, 2004 ), N:P则可以反映植物生长受N或P的限制情况(Koerselman & Meuleman, 1996 ; Elser et al ., 2000 ; Güsewell, 2004 ).植物不同组织器官营养元素水平与变化是调整它们的生长速率以适应环境变化的一种策略(Marschner, 1995 ), 如干旱区植物相对于湿润区植物通常拥有更高的单位面积(或质量)叶N、P含量(Wright et al ., 2001 ; 李玉霖等, 2010 ), 植物地下根系的N、P含量低于地上部分的叶片, 但二者具有相似的N:P化学计量比(Yuan et al ., 2011 ).当前, 区域C、N、P化学计量格局及其驱动因素仍然是生态化学计量学的一个重要研究领域, 其结果不仅有助于揭示物种间化学计量的趋同与分异, 对于预测生态系统功能、过程对全球变化的响应亦具有重要意义(贺金生和韩兴国, 2010 ; Elser et al ., 2010 ).近年来, 国内学者在不同时空尺度, 针对森林(刘万德等, 2010 ; 崔高阳等, 2015 )、草地(Yu et al ., 2010 ; 雒明伟等, 2015 )、荒漠(李玉霖等, 2010 ; 黄菊莹和余海龙, 2016 )、湿地(Wang et al ., 2015 ; 聂兰琴等, 2016 )等生态系统类型开展了大量的研究.然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al ., 2010 ).同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014 ). ...
Plant responses to multiple environmental factors
1
1987
... Kattge等(2011)搜集整理了全球植物性状数据库, 发现全球植物叶片C、N、P含量的平均值分别为476.1 mg·g-1 、17.4 mg·g-1 、1.23 mg·g-1 , C:N、N:P则分别为23.4、12.8.与上述全球叶性状数据库相比较, 单叶蔓荆叶C (449.5 mg·g-1 )含量偏低, 叶N (19.66 mg·g-1 )含量略高, 叶P (1.24 mg·g-1 )含量则高度一致; 叶片C:N (23.1)基本一致, N:P (16.8)明显偏高.单叶蔓荆细根C (386.3 mg·g-1 )、N (16.8 mg·g-1 )、P (1.32 mg·g-1 )含量与Yuan等(2011)报道的全球植物细根养分含量及化学计量比相比, C含量与C:N明显偏低, N、P含量明显偏高, 但N:P化学计量比基本一致.同样, 与马玉珠等(2015)报道的中国植物细根的C、N、P含量相比, 我们也发现本研究中的单叶蔓荆细根C含量明显偏低, N、P含量明显偏高.与全球及中国大样本数据相比, 单叶蔓荆的叶片及细根的N、P含量, 特别是N含量明显偏高, 一方面可能与上述数据库中的多数样本来自于森林中的乔木有关, 同时还可能与本研究区单叶蔓荆特殊的荒漠生境相关.研究区虽不具有干旱区荒漠的长期水分限制, 但因为水热不同期, 特别是夏秋季节的高温、少雨与地表的强蒸发, 容易形成季节性的水分胁迫.同时, 沙地土壤养分匮乏, 单叶蔓荆在水分与养分均存在限制的情形下增加叶片N投入可能是一种有效的适应策略, 因为叶N增加能够提升叶片内部光合作用酶的数量, 进一步提高叶片光合速率(Chapin et al ., 1987 ).此外, 叶N含量的增加还可以增加叶片内部非光合器官或组织氮的投入, 从而提高细胞内部的渗透压, 增强对植物体内水分的保护, 是一种有效的节水方式(Wright et al ., 2001 ). ...
Covariation in leaf and root traits for native and non-native grasses along an altitudinal gradient in New Zealand
1
2003
... 近年来, 越来越多的研究尝试从植物功能性状或资源经济谱的角度来探讨不同器官之间的性状关联, 相关研究不仅有助于理解植物各性状之间相互作用的机制(Eviner & Chapin, 2003 )、植物生长过程中对资源的利用和分配(Kerkhoff et al ., 2006 ), 还对进一步预测植物对环境变化的响应具有重要意义.本研究发现单叶蔓荆各器官的养分元素及化学计量关系的相关性主要集中在地上部分, 即花、叶、枝条、匍匐茎之间, 其中尤以枝条与匍匐茎之间表现明显, 二者除了N:P没有达到显著相关之外, 其他的养分元素及其化学计量关系均达到了显著性相关(表2 ), 显示了二者之间在养分存储与运输之间的密切关系.在植物的所有器官中, 叶片与细根之间的性状关联最受关注.一些大尺度多物种间的研究表明叶片和细根的N含量(Craine & Lee, 2003 ; Tjoelker et al ., 2005 )、P含量(Kerkhoff et al ., 2006 ; Geng et al ., 2014 )均呈显著正相关.本研究未发现叶片与细根之间的3种元素及其化学计量关系显著相关, 一个可能的原因是叶片与细根性状的关联主要发生在种间, 小尺度上植物生境条件相对一致, 种内个体间功能性状因变异幅度较小而关联不明显(徐冰等, 2010 ).另一种可能的原因是本研究中的细根样品的采集是以直径<2 mm为标准的.由于根系在发育过程中形成明显的分枝结构, 具有高度的形态异质性(Pregitzer et al ., 2002 ; 王政权和郭大立, 2008 ), 直径<2 mm的细根并不是一个均质的单一组分, 而是由形态、寿命、生理差异巨大的不同个体根组成的高度异质的混合体(Pregitzer et al ., 2002; Guo et al ., 2008 ), 从而影响了与叶片之间相应元素的相关关系. ...
陕西省森林各生态系统组分氮磷化学计量特征
1
2015
... 碳(C)、氮(N)和磷(P)是植物基本营养元素, 在植物生长和各种生理调节机能中发挥着重要作用.C是构成植物体内干物质的最主要元素, 而N和P则分别与植物的光合作用和细胞分裂等重要生理活动有关(Niklas et al ., 2005 ).N:C和P:C与植物的相对生长率关系密切(Ågren, 2004 ), N:P则可以反映植物生长受N或P的限制情况(Koerselman & Meuleman, 1996 ; Elser et al ., 2000 ; Güsewell, 2004 ).植物不同组织器官营养元素水平与变化是调整它们的生长速率以适应环境变化的一种策略(Marschner, 1995 ), 如干旱区植物相对于湿润区植物通常拥有更高的单位面积(或质量)叶N、P含量(Wright et al ., 2001 ; 李玉霖等, 2010 ), 植物地下根系的N、P含量低于地上部分的叶片, 但二者具有相似的N:P化学计量比(Yuan et al ., 2011 ).当前, 区域C、N、P化学计量格局及其驱动因素仍然是生态化学计量学的一个重要研究领域, 其结果不仅有助于揭示物种间化学计量的趋同与分异, 对于预测生态系统功能、过程对全球变化的响应亦具有重要意义(贺金生和韩兴国, 2010 ; Elser et al ., 2010 ).近年来, 国内学者在不同时空尺度, 针对森林(刘万德等, 2010 ; 崔高阳等, 2015 )、草地(Yu et al ., 2010 ; 雒明伟等, 2015 )、荒漠(李玉霖等, 2010 ; 黄菊莹和余海龙, 2016 )、湿地(Wang et al ., 2015 ; 聂兰琴等, 2016 )等生态系统类型开展了大量的研究.然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al ., 2010 ).同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014 ). ...
我国荒漠化现状、成因与防治对策
1
1999
... 土地沙化长期以来一直是我国最为严重的生态环境问题(董光荣等, 1999 ), 不仅出现在我国北方干旱、半干旱地区, 亦出现于南方湖滨及河流下游冲积平原地区(朱震达和崔书红, 1996 ).以鄱阳湖区为例, 沙化土地面积多达3.89万hm2 (左长青, 1986 ).鄱阳湖沙化土地虽处于亚热带湿润区, 但水热不同期, 特别是8-10月的高温、少雨与地表强蒸发, 以及冬春季节固定的风向与强劲的风速是影响植被恢复的重要因素.近几十年来, 马鞭草科落叶小灌木单叶蔓荆(Vitex rotundifolia )被成功地应用于鄱阳湖沙化土地的植被恢复, 并且在不同沙化程度区形成稳定的群落.研究表明生物有机体具有极高的化学计量内稳性(Elser et al ., 2010 ), 并且通过调节生物对环境变化的响应, 化学计量内稳性成为生态系统结构、功能和稳定性维持的重要机制(Yu et al ., 2010 ).单叶蔓荆的养分及其化学计量关系在不同沙化程度区会发生怎样的变化?其N、P功能性状之间的关系是否因沙化梯度而发生策略位移?本研究通过测定鄱阳湖典型沙山重度、中度、轻度沙化区单叶蔓荆花、叶、枝条、匍匐茎、细根等部位的C、N、P含量, 分析各器官养分元素分配格局、化学计量比的差异与关联, 以及随沙化程度的变化, 试图探讨上述两个科学问题, 为进一步揭示单叶蔓荆的沙地适应机制提供科学依据, 为鄱阳湖沙化区植被恢复提供参考. ...
Nutritional constraints in terrestrial and freshwater food webs
1
2000
... 碳(C)、氮(N)和磷(P)是植物基本营养元素, 在植物生长和各种生理调节机能中发挥着重要作用.C是构成植物体内干物质的最主要元素, 而N和P则分别与植物的光合作用和细胞分裂等重要生理活动有关(Niklas et al ., 2005 ).N:C和P:C与植物的相对生长率关系密切(Ågren, 2004 ), N:P则可以反映植物生长受N或P的限制情况(Koerselman & Meuleman, 1996 ; Elser et al ., 2000 ; Güsewell, 2004 ).植物不同组织器官营养元素水平与变化是调整它们的生长速率以适应环境变化的一种策略(Marschner, 1995 ), 如干旱区植物相对于湿润区植物通常拥有更高的单位面积(或质量)叶N、P含量(Wright et al ., 2001 ; 李玉霖等, 2010 ), 植物地下根系的N、P含量低于地上部分的叶片, 但二者具有相似的N:P化学计量比(Yuan et al ., 2011 ).当前, 区域C、N、P化学计量格局及其驱动因素仍然是生态化学计量学的一个重要研究领域, 其结果不仅有助于揭示物种间化学计量的趋同与分异, 对于预测生态系统功能、过程对全球变化的响应亦具有重要意义(贺金生和韩兴国, 2010 ; Elser et al ., 2010 ).近年来, 国内学者在不同时空尺度, 针对森林(刘万德等, 2010 ; 崔高阳等, 2015 )、草地(Yu et al ., 2010 ; 雒明伟等, 2015 )、荒漠(李玉霖等, 2010 ; 黄菊莹和余海龙, 2016 )、湿地(Wang et al ., 2015 ; 聂兰琴等, 2016 )等生态系统类型开展了大量的研究.然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al ., 2010 ).同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014 ). ...
Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change
3
2010
... 碳(C)、氮(N)和磷(P)是植物基本营养元素, 在植物生长和各种生理调节机能中发挥着重要作用.C是构成植物体内干物质的最主要元素, 而N和P则分别与植物的光合作用和细胞分裂等重要生理活动有关(Niklas et al ., 2005 ).N:C和P:C与植物的相对生长率关系密切(Ågren, 2004 ), N:P则可以反映植物生长受N或P的限制情况(Koerselman & Meuleman, 1996 ; Elser et al ., 2000 ; Güsewell, 2004 ).植物不同组织器官营养元素水平与变化是调整它们的生长速率以适应环境变化的一种策略(Marschner, 1995 ), 如干旱区植物相对于湿润区植物通常拥有更高的单位面积(或质量)叶N、P含量(Wright et al ., 2001 ; 李玉霖等, 2010 ), 植物地下根系的N、P含量低于地上部分的叶片, 但二者具有相似的N:P化学计量比(Yuan et al ., 2011 ).当前, 区域C、N、P化学计量格局及其驱动因素仍然是生态化学计量学的一个重要研究领域, 其结果不仅有助于揭示物种间化学计量的趋同与分异, 对于预测生态系统功能、过程对全球变化的响应亦具有重要意义(贺金生和韩兴国, 2010 ; Elser et al ., 2010 ).近年来, 国内学者在不同时空尺度, 针对森林(刘万德等, 2010 ; 崔高阳等, 2015 )、草地(Yu et al ., 2010 ; 雒明伟等, 2015 )、荒漠(李玉霖等, 2010 ; 黄菊莹和余海龙, 2016 )、湿地(Wang et al ., 2015 ; 聂兰琴等, 2016 )等生态系统类型开展了大量的研究.然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al ., 2010 ).同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014 ). ...
... )等生态系统类型开展了大量的研究.然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al ., 2010 ).同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014 ). ...
... 土地沙化长期以来一直是我国最为严重的生态环境问题(董光荣等, 1999 ), 不仅出现在我国北方干旱、半干旱地区, 亦出现于南方湖滨及河流下游冲积平原地区(朱震达和崔书红, 1996 ).以鄱阳湖区为例, 沙化土地面积多达3.89万hm2 (左长青, 1986 ).鄱阳湖沙化土地虽处于亚热带湿润区, 但水热不同期, 特别是8-10月的高温、少雨与地表强蒸发, 以及冬春季节固定的风向与强劲的风速是影响植被恢复的重要因素.近几十年来, 马鞭草科落叶小灌木单叶蔓荆(Vitex rotundifolia )被成功地应用于鄱阳湖沙化土地的植被恢复, 并且在不同沙化程度区形成稳定的群落.研究表明生物有机体具有极高的化学计量内稳性(Elser et al ., 2010 ), 并且通过调节生物对环境变化的响应, 化学计量内稳性成为生态系统结构、功能和稳定性维持的重要机制(Yu et al ., 2010 ).单叶蔓荆的养分及其化学计量关系在不同沙化程度区会发生怎样的变化?其N、P功能性状之间的关系是否因沙化梯度而发生策略位移?本研究通过测定鄱阳湖典型沙山重度、中度、轻度沙化区单叶蔓荆花、叶、枝条、匍匐茎、细根等部位的C、N、P含量, 分析各器官养分元素分配格局、化学计量比的差异与关联, 以及随沙化程度的变化, 试图探讨上述两个科学问题, 为进一步揭示单叶蔓荆的沙地适应机制提供科学依据, 为鄱阳湖沙化区植被恢复提供参考. ...
Functional matrix: A conceptual framework for predicting multiple plant effects on ecosystem processes
1
2003
... 近年来, 越来越多的研究尝试从植物功能性状或资源经济谱的角度来探讨不同器官之间的性状关联, 相关研究不仅有助于理解植物各性状之间相互作用的机制(Eviner & Chapin, 2003 )、植物生长过程中对资源的利用和分配(Kerkhoff et al ., 2006 ), 还对进一步预测植物对环境变化的响应具有重要意义.本研究发现单叶蔓荆各器官的养分元素及化学计量关系的相关性主要集中在地上部分, 即花、叶、枝条、匍匐茎之间, 其中尤以枝条与匍匐茎之间表现明显, 二者除了N:P没有达到显著相关之外, 其他的养分元素及其化学计量关系均达到了显著性相关(表2 ), 显示了二者之间在养分存储与运输之间的密切关系.在植物的所有器官中, 叶片与细根之间的性状关联最受关注.一些大尺度多物种间的研究表明叶片和细根的N含量(Craine & Lee, 2003 ; Tjoelker et al ., 2005 )、P含量(Kerkhoff et al ., 2006 ; Geng et al ., 2014 )均呈显著正相关.本研究未发现叶片与细根之间的3种元素及其化学计量关系显著相关, 一个可能的原因是叶片与细根性状的关联主要发生在种间, 小尺度上植物生境条件相对一致, 种内个体间功能性状因变异幅度较小而关联不明显(徐冰等, 2010 ).另一种可能的原因是本研究中的细根样品的采集是以直径<2 mm为标准的.由于根系在发育过程中形成明显的分枝结构, 具有高度的形态异质性(Pregitzer et al ., 2002 ; 王政权和郭大立, 2008 ), 直径<2 mm的细根并不是一个均质的单一组分, 而是由形态、寿命、生理差异巨大的不同个体根组成的高度异质的混合体(Pregitzer et al ., 2002; Guo et al ., 2008 ), 从而影响了与叶片之间相应元素的相关关系. ...
Alpine climate alters the relationships between leaf and root morphological traits but not chemical traits
1
2014
... 近年来, 越来越多的研究尝试从植物功能性状或资源经济谱的角度来探讨不同器官之间的性状关联, 相关研究不仅有助于理解植物各性状之间相互作用的机制(Eviner & Chapin, 2003 )、植物生长过程中对资源的利用和分配(Kerkhoff et al ., 2006 ), 还对进一步预测植物对环境变化的响应具有重要意义.本研究发现单叶蔓荆各器官的养分元素及化学计量关系的相关性主要集中在地上部分, 即花、叶、枝条、匍匐茎之间, 其中尤以枝条与匍匐茎之间表现明显, 二者除了N:P没有达到显著相关之外, 其他的养分元素及其化学计量关系均达到了显著性相关(表2 ), 显示了二者之间在养分存储与运输之间的密切关系.在植物的所有器官中, 叶片与细根之间的性状关联最受关注.一些大尺度多物种间的研究表明叶片和细根的N含量(Craine & Lee, 2003 ; Tjoelker et al ., 2005 )、P含量(Kerkhoff et al ., 2006 ; Geng et al ., 2014 )均呈显著正相关.本研究未发现叶片与细根之间的3种元素及其化学计量关系显著相关, 一个可能的原因是叶片与细根性状的关联主要发生在种间, 小尺度上植物生境条件相对一致, 种内个体间功能性状因变异幅度较小而关联不明显(徐冰等, 2010 ).另一种可能的原因是本研究中的细根样品的采集是以直径<2 mm为标准的.由于根系在发育过程中形成明显的分枝结构, 具有高度的形态异质性(Pregitzer et al ., 2002 ; 王政权和郭大立, 2008 ), 直径<2 mm的细根并不是一个均质的单一组分, 而是由形态、寿命、生理差异巨大的不同个体根组成的高度异质的混合体(Pregitzer et al ., 2002; Guo et al ., 2008 ), 从而影响了与叶片之间相应元素的相关关系. ...
Heterogeneity by root branch order: Exploring the discrepancy in root longevity and turnover estimates between minirhizotron and C isotope methods
1
2008
... 近年来, 越来越多的研究尝试从植物功能性状或资源经济谱的角度来探讨不同器官之间的性状关联, 相关研究不仅有助于理解植物各性状之间相互作用的机制(Eviner & Chapin, 2003 )、植物生长过程中对资源的利用和分配(Kerkhoff et al ., 2006 ), 还对进一步预测植物对环境变化的响应具有重要意义.本研究发现单叶蔓荆各器官的养分元素及化学计量关系的相关性主要集中在地上部分, 即花、叶、枝条、匍匐茎之间, 其中尤以枝条与匍匐茎之间表现明显, 二者除了N:P没有达到显著相关之外, 其他的养分元素及其化学计量关系均达到了显著性相关(表2 ), 显示了二者之间在养分存储与运输之间的密切关系.在植物的所有器官中, 叶片与细根之间的性状关联最受关注.一些大尺度多物种间的研究表明叶片和细根的N含量(Craine & Lee, 2003 ; Tjoelker et al ., 2005 )、P含量(Kerkhoff et al ., 2006 ; Geng et al ., 2014 )均呈显著正相关.本研究未发现叶片与细根之间的3种元素及其化学计量关系显著相关, 一个可能的原因是叶片与细根性状的关联主要发生在种间, 小尺度上植物生境条件相对一致, 种内个体间功能性状因变异幅度较小而关联不明显(徐冰等, 2010 ).另一种可能的原因是本研究中的细根样品的采集是以直径<2 mm为标准的.由于根系在发育过程中形成明显的分枝结构, 具有高度的形态异质性(Pregitzer et al ., 2002 ; 王政权和郭大立, 2008 ), 直径<2 mm的细根并不是一个均质的单一组分, 而是由形态、寿命、生理差异巨大的不同个体根组成的高度异质的混合体(Pregitzer et al ., 2002; Guo et al ., 2008 ), 从而影响了与叶片之间相应元素的相关关系. ...
N:P ratios in terrestrial plants: Variation and functional significance
1
2004
... 碳(C)、氮(N)和磷(P)是植物基本营养元素, 在植物生长和各种生理调节机能中发挥着重要作用.C是构成植物体内干物质的最主要元素, 而N和P则分别与植物的光合作用和细胞分裂等重要生理活动有关(Niklas et al ., 2005 ).N:C和P:C与植物的相对生长率关系密切(Ågren, 2004 ), N:P则可以反映植物生长受N或P的限制情况(Koerselman & Meuleman, 1996 ; Elser et al ., 2000 ; Güsewell, 2004 ).植物不同组织器官营养元素水平与变化是调整它们的生长速率以适应环境变化的一种策略(Marschner, 1995 ), 如干旱区植物相对于湿润区植物通常拥有更高的单位面积(或质量)叶N、P含量(Wright et al ., 2001 ; 李玉霖等, 2010 ), 植物地下根系的N、P含量低于地上部分的叶片, 但二者具有相似的N:P化学计量比(Yuan et al ., 2011 ).当前, 区域C、N、P化学计量格局及其驱动因素仍然是生态化学计量学的一个重要研究领域, 其结果不仅有助于揭示物种间化学计量的趋同与分异, 对于预测生态系统功能、过程对全球变化的响应亦具有重要意义(贺金生和韩兴国, 2010 ; Elser et al ., 2010 ).近年来, 国内学者在不同时空尺度, 针对森林(刘万德等, 2010 ; 崔高阳等, 2015 )、草地(Yu et al ., 2010 ; 雒明伟等, 2015 )、荒漠(李玉霖等, 2010 ; 黄菊莹和余海龙, 2016 )、湿地(Wang et al ., 2015 ; 聂兰琴等, 2016 )等生态系统类型开展了大量的研究.然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al ., 2010 ).同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014 ). ...
生态化学计量学: 探索从个体到生态系统的统一化理论
1
2010
... 碳(C)、氮(N)和磷(P)是植物基本营养元素, 在植物生长和各种生理调节机能中发挥着重要作用.C是构成植物体内干物质的最主要元素, 而N和P则分别与植物的光合作用和细胞分裂等重要生理活动有关(Niklas et al ., 2005 ).N:C和P:C与植物的相对生长率关系密切(Ågren, 2004 ), N:P则可以反映植物生长受N或P的限制情况(Koerselman & Meuleman, 1996 ; Elser et al ., 2000 ; Güsewell, 2004 ).植物不同组织器官营养元素水平与变化是调整它们的生长速率以适应环境变化的一种策略(Marschner, 1995 ), 如干旱区植物相对于湿润区植物通常拥有更高的单位面积(或质量)叶N、P含量(Wright et al ., 2001 ; 李玉霖等, 2010 ), 植物地下根系的N、P含量低于地上部分的叶片, 但二者具有相似的N:P化学计量比(Yuan et al ., 2011 ).当前, 区域C、N、P化学计量格局及其驱动因素仍然是生态化学计量学的一个重要研究领域, 其结果不仅有助于揭示物种间化学计量的趋同与分异, 对于预测生态系统功能、过程对全球变化的响应亦具有重要意义(贺金生和韩兴国, 2010 ; Elser et al ., 2010 ).近年来, 国内学者在不同时空尺度, 针对森林(刘万德等, 2010 ; 崔高阳等, 2015 )、草地(Yu et al ., 2010 ; 雒明伟等, 2015 )、荒漠(李玉霖等, 2010 ; 黄菊莹和余海龙, 2016 )、湿地(Wang et al ., 2015 ; 聂兰琴等, 2016 )等生态系统类型开展了大量的研究.然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al ., 2010 ).同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014 ). ...
四种荒漠草原植物的生长对不同氮添加水平的响应
1
2016
... 碳(C)、氮(N)和磷(P)是植物基本营养元素, 在植物生长和各种生理调节机能中发挥着重要作用.C是构成植物体内干物质的最主要元素, 而N和P则分别与植物的光合作用和细胞分裂等重要生理活动有关(Niklas et al ., 2005 ).N:C和P:C与植物的相对生长率关系密切(Ågren, 2004 ), N:P则可以反映植物生长受N或P的限制情况(Koerselman & Meuleman, 1996 ; Elser et al ., 2000 ; Güsewell, 2004 ).植物不同组织器官营养元素水平与变化是调整它们的生长速率以适应环境变化的一种策略(Marschner, 1995 ), 如干旱区植物相对于湿润区植物通常拥有更高的单位面积(或质量)叶N、P含量(Wright et al ., 2001 ; 李玉霖等, 2010 ), 植物地下根系的N、P含量低于地上部分的叶片, 但二者具有相似的N:P化学计量比(Yuan et al ., 2011 ).当前, 区域C、N、P化学计量格局及其驱动因素仍然是生态化学计量学的一个重要研究领域, 其结果不仅有助于揭示物种间化学计量的趋同与分异, 对于预测生态系统功能、过程对全球变化的响应亦具有重要意义(贺金生和韩兴国, 2010 ; Elser et al ., 2010 ).近年来, 国内学者在不同时空尺度, 针对森林(刘万德等, 2010 ; 崔高阳等, 2015 )、草地(Yu et al ., 2010 ; 雒明伟等, 2015 )、荒漠(李玉霖等, 2010 ; 黄菊莹和余海龙, 2016 )、湿地(Wang et al ., 2015 ; 聂兰琴等, 2016 )等生态系统类型开展了大量的研究.然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al ., 2010 ).同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014 ). ...
TRY—A global database of plant traits
2011
Phylogenetic and growth form variation in the scaling of nitrogen and phosphorus in the seed plants
3
2006
... N、P养分元素在单叶蔓荆各器官中的分配存在显著差异(图1 ), 表现为花的N、P含量最高, 叶与细根次之, 枝条与匍匐茎之间没有显著差异且显著低于其他器官.植物不同器官营养元素的分布, 除受土壤养分元素的供给影响之外, 主要与相应器官的生理功能密切相关(Kerkhoff et al ., 2006 ).采样时单叶蔓荆正处于花期, 作为重要的繁殖器官, 花富含线粒体, 且吸收储存大量营养物质为果实形成做准备, 因此具有最高的N、P含量.叶片是植物进行光合作用的器官, 保持较高的N含量有利于提高光合速率.同时, 叶片中存储较多的N、P也是对特殊生境的一种适应策略.根系是吸收营养物质的主要器官, 根系特别是细根具有较高的N、P含量.匍匐茎和枝条主要起水分与养分输导及支撑植物的作用, 因此, 养分元素的含量相对较低. ...
... 近年来, 越来越多的研究尝试从植物功能性状或资源经济谱的角度来探讨不同器官之间的性状关联, 相关研究不仅有助于理解植物各性状之间相互作用的机制(Eviner & Chapin, 2003 )、植物生长过程中对资源的利用和分配(Kerkhoff et al ., 2006 ), 还对进一步预测植物对环境变化的响应具有重要意义.本研究发现单叶蔓荆各器官的养分元素及化学计量关系的相关性主要集中在地上部分, 即花、叶、枝条、匍匐茎之间, 其中尤以枝条与匍匐茎之间表现明显, 二者除了N:P没有达到显著相关之外, 其他的养分元素及其化学计量关系均达到了显著性相关(表2 ), 显示了二者之间在养分存储与运输之间的密切关系.在植物的所有器官中, 叶片与细根之间的性状关联最受关注.一些大尺度多物种间的研究表明叶片和细根的N含量(Craine & Lee, 2003 ; Tjoelker et al ., 2005 )、P含量(Kerkhoff et al ., 2006 ; Geng et al ., 2014 )均呈显著正相关.本研究未发现叶片与细根之间的3种元素及其化学计量关系显著相关, 一个可能的原因是叶片与细根性状的关联主要发生在种间, 小尺度上植物生境条件相对一致, 种内个体间功能性状因变异幅度较小而关联不明显(徐冰等, 2010 ).另一种可能的原因是本研究中的细根样品的采集是以直径<2 mm为标准的.由于根系在发育过程中形成明显的分枝结构, 具有高度的形态异质性(Pregitzer et al ., 2002 ; 王政权和郭大立, 2008 ), 直径<2 mm的细根并不是一个均质的单一组分, 而是由形态、寿命、生理差异巨大的不同个体根组成的高度异质的混合体(Pregitzer et al ., 2002; Guo et al ., 2008 ), 从而影响了与叶片之间相应元素的相关关系. ...
... )、P含量(Kerkhoff et al ., 2006 ; Geng et al ., 2014 )均呈显著正相关.本研究未发现叶片与细根之间的3种元素及其化学计量关系显著相关, 一个可能的原因是叶片与细根性状的关联主要发生在种间, 小尺度上植物生境条件相对一致, 种内个体间功能性状因变异幅度较小而关联不明显(徐冰等, 2010 ).另一种可能的原因是本研究中的细根样品的采集是以直径<2 mm为标准的.由于根系在发育过程中形成明显的分枝结构, 具有高度的形态异质性(Pregitzer et al ., 2002 ; 王政权和郭大立, 2008 ), 直径<2 mm的细根并不是一个均质的单一组分, 而是由形态、寿命、生理差异巨大的不同个体根组成的高度异质的混合体(Pregitzer et al ., 2002; Guo et al ., 2008 ), 从而影响了与叶片之间相应元素的相关关系. ...
High foliar nitrogen in desert shrubs: An important ecosystem trait or defective desert doctrine
1996
The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation
1
1996
... 碳(C)、氮(N)和磷(P)是植物基本营养元素, 在植物生长和各种生理调节机能中发挥着重要作用.C是构成植物体内干物质的最主要元素, 而N和P则分别与植物的光合作用和细胞分裂等重要生理活动有关(Niklas et al ., 2005 ).N:C和P:C与植物的相对生长率关系密切(Ågren, 2004 ), N:P则可以反映植物生长受N或P的限制情况(Koerselman & Meuleman, 1996 ; Elser et al ., 2000 ; Güsewell, 2004 ).植物不同组织器官营养元素水平与变化是调整它们的生长速率以适应环境变化的一种策略(Marschner, 1995 ), 如干旱区植物相对于湿润区植物通常拥有更高的单位面积(或质量)叶N、P含量(Wright et al ., 2001 ; 李玉霖等, 2010 ), 植物地下根系的N、P含量低于地上部分的叶片, 但二者具有相似的N:P化学计量比(Yuan et al ., 2011 ).当前, 区域C、N、P化学计量格局及其驱动因素仍然是生态化学计量学的一个重要研究领域, 其结果不仅有助于揭示物种间化学计量的趋同与分异, 对于预测生态系统功能、过程对全球变化的响应亦具有重要意义(贺金生和韩兴国, 2010 ; Elser et al ., 2010 ).近年来, 国内学者在不同时空尺度, 针对森林(刘万德等, 2010 ; 崔高阳等, 2015 )、草地(Yu et al ., 2010 ; 雒明伟等, 2015 )、荒漠(李玉霖等, 2010 ; 黄菊莹和余海龙, 2016 )、湿地(Wang et al ., 2015 ; 聂兰琴等, 2016 )等生态系统类型开展了大量的研究.然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al ., 2010 ).同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014 ). ...
青海省沙珠玉治沙站17种主要植物叶性因子的比较
2
2005
... 单叶蔓荆作为鄱阳湖湖滨沙山植被恢复过程的先锋物种, 广布于沙山不同程度沙化区, 为进一步说明研究区单叶蔓荆不同器官的养分含量及其化学计量关系特征, 我们把单叶蔓荆地上部分的叶片、地下部分的细根与其他荒漠区植物, 以及全球、中国大样本研究中的植物养分平均值做了比较.本研究区单叶蔓荆叶N (19.66 mg·g-1 )、P (1.24 mg·g-1 )含量明显低于中国北方典型荒漠及荒漠化地区植物N (24.45 mg·g-1 )、P (1.74 mg·g-1 )含量(李玉霖等,2010 ).此外, 叶N含量亦低于Killingbeck和Whitford (1996)报道的全球5大洲11个干旱荒漠中78种灌木叶N含量平均值(22 mg·g-1 ).本研究区虽属于南方典型荒漠, 但地处亚热带湿润区, 年降水量远高于其他干旱区荒漠, 这可能是单叶蔓荆叶N、P含量低于其他干旱、半干旱区荒漠植物的重要原因, 因为湿润区植物相对于干旱区通常具有更低的叶N、P含量(Wright et al ., 2001 ; 李永华等, 2005 ). ...
... 由于降水、土壤养分等环境梯度而引起的植物功能性状之间关系的策略位移现象不仅出现在种间水平上(Wright et al ., 2001 ; 李永华等, 2005 ; Wei et al ., 2011 ), 也出现在种内水平上(Wei et al ., 2011 ).但这一现象是否具有普遍性目前仍不清楚, 例如, 胡梦瑶等(2012)并没有发现紫花针茅(Stipa purpurea )在不同降水梯度下具有明显的策略位移现象.此外, 上述功能性状的位移现象多出现于较大尺度下的环境梯度中, 局地尺度上是否也存在类似的现象仍缺乏研究.本研究区尽管重度、中度与轻度沙化区土壤有机质、全氮、全磷等养分, 以及地表植被覆盖度等存在显著差异(表1 ), 但未发现单叶蔓荆N mass -P mass 线性关系的斜率与截距在3种沙化程度区发生显著变化(表3 ), 表明单叶蔓荆这一对功能性状关系在不同沙化区未发生明显的位移现象.成对植物功能性状关系发生策略位移主要是由于环境梯度对其中一个功能性状影响更大, 例如, Wei等(2011)发现低降水区的油蒿(Artemisia ordosica )种群N mass 高于较高降水区, 但比叶面积差异不大.本研究中N mass -P mass 在不同沙化程度区未出现明显的策略位移现象, 表明土壤沙化所引起的水、肥等限制因素可能共同作用于这两个性状, 使得植物N mass 与P mass 发生同步变化, 同时这也进一步佐证了N:P化学计量关系在不同沙化区的相对稳定.此外, 本研究结果也表明局地尺度种内水平上很可能因为环境梯度过小而不具备发生N mass -P mass 关系的位移现象. ...
北方典型荒漠及荒漠化地区植物叶片氮磷化学计量特征研究
3
2010
... 碳(C)、氮(N)和磷(P)是植物基本营养元素, 在植物生长和各种生理调节机能中发挥着重要作用.C是构成植物体内干物质的最主要元素, 而N和P则分别与植物的光合作用和细胞分裂等重要生理活动有关(Niklas et al ., 2005 ).N:C和P:C与植物的相对生长率关系密切(Ågren, 2004 ), N:P则可以反映植物生长受N或P的限制情况(Koerselman & Meuleman, 1996 ; Elser et al ., 2000 ; Güsewell, 2004 ).植物不同组织器官营养元素水平与变化是调整它们的生长速率以适应环境变化的一种策略(Marschner, 1995 ), 如干旱区植物相对于湿润区植物通常拥有更高的单位面积(或质量)叶N、P含量(Wright et al ., 2001 ; 李玉霖等, 2010 ), 植物地下根系的N、P含量低于地上部分的叶片, 但二者具有相似的N:P化学计量比(Yuan et al ., 2011 ).当前, 区域C、N、P化学计量格局及其驱动因素仍然是生态化学计量学的一个重要研究领域, 其结果不仅有助于揭示物种间化学计量的趋同与分异, 对于预测生态系统功能、过程对全球变化的响应亦具有重要意义(贺金生和韩兴国, 2010 ; Elser et al ., 2010 ).近年来, 国内学者在不同时空尺度, 针对森林(刘万德等, 2010 ; 崔高阳等, 2015 )、草地(Yu et al ., 2010 ; 雒明伟等, 2015 )、荒漠(李玉霖等, 2010 ; 黄菊莹和余海龙, 2016 )、湿地(Wang et al ., 2015 ; 聂兰琴等, 2016 )等生态系统类型开展了大量的研究.然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al ., 2010 ).同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014 ). ...
... )、荒漠(李玉霖等, 2010 ; 黄菊莹和余海龙, 2016 )、湿地(Wang et al ., 2015 ; 聂兰琴等, 2016 )等生态系统类型开展了大量的研究.然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al ., 2010 ).同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014 ). ...
... 单叶蔓荆作为鄱阳湖湖滨沙山植被恢复过程的先锋物种, 广布于沙山不同程度沙化区, 为进一步说明研究区单叶蔓荆不同器官的养分含量及其化学计量关系特征, 我们把单叶蔓荆地上部分的叶片、地下部分的细根与其他荒漠区植物, 以及全球、中国大样本研究中的植物养分平均值做了比较.本研究区单叶蔓荆叶N (19.66 mg·g-1 )、P (1.24 mg·g-1 )含量明显低于中国北方典型荒漠及荒漠化地区植物N (24.45 mg·g-1 )、P (1.74 mg·g-1 )含量(李玉霖等,2010 ).此外, 叶N含量亦低于Killingbeck和Whitford (1996)报道的全球5大洲11个干旱荒漠中78种灌木叶N含量平均值(22 mg·g-1 ).本研究区虽属于南方典型荒漠, 但地处亚热带湿润区, 年降水量远高于其他干旱区荒漠, 这可能是单叶蔓荆叶N、P含量低于其他干旱、半干旱区荒漠植物的重要原因, 因为湿润区植物相对于干旱区通常具有更低的叶N、P含量(Wright et al ., 2001 ; 李永华等, 2005 ). ...
云南普洱季风常绿阔叶林演替系列植物和土壤C、N、P化学计量特征
1
2010
... 碳(C)、氮(N)和磷(P)是植物基本营养元素, 在植物生长和各种生理调节机能中发挥着重要作用.C是构成植物体内干物质的最主要元素, 而N和P则分别与植物的光合作用和细胞分裂等重要生理活动有关(Niklas et al ., 2005 ).N:C和P:C与植物的相对生长率关系密切(Ågren, 2004 ), N:P则可以反映植物生长受N或P的限制情况(Koerselman & Meuleman, 1996 ; Elser et al ., 2000 ; Güsewell, 2004 ).植物不同组织器官营养元素水平与变化是调整它们的生长速率以适应环境变化的一种策略(Marschner, 1995 ), 如干旱区植物相对于湿润区植物通常拥有更高的单位面积(或质量)叶N、P含量(Wright et al ., 2001 ; 李玉霖等, 2010 ), 植物地下根系的N、P含量低于地上部分的叶片, 但二者具有相似的N:P化学计量比(Yuan et al ., 2011 ).当前, 区域C、N、P化学计量格局及其驱动因素仍然是生态化学计量学的一个重要研究领域, 其结果不仅有助于揭示物种间化学计量的趋同与分异, 对于预测生态系统功能、过程对全球变化的响应亦具有重要意义(贺金生和韩兴国, 2010 ; Elser et al ., 2010 ).近年来, 国内学者在不同时空尺度, 针对森林(刘万德等, 2010 ; 崔高阳等, 2015 )、草地(Yu et al ., 2010 ; 雒明伟等, 2015 )、荒漠(李玉霖等, 2010 ; 黄菊莹和余海龙, 2016 )、湿地(Wang et al ., 2015 ; 聂兰琴等, 2016 )等生态系统类型开展了大量的研究.然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al ., 2010 ).同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014 ). ...
青藏高原筑路取土迹地恢复植物群落与土壤的碳氮磷化学计量特征
1
2015
... 碳(C)、氮(N)和磷(P)是植物基本营养元素, 在植物生长和各种生理调节机能中发挥着重要作用.C是构成植物体内干物质的最主要元素, 而N和P则分别与植物的光合作用和细胞分裂等重要生理活动有关(Niklas et al ., 2005 ).N:C和P:C与植物的相对生长率关系密切(Ågren, 2004 ), N:P则可以反映植物生长受N或P的限制情况(Koerselman & Meuleman, 1996 ; Elser et al ., 2000 ; Güsewell, 2004 ).植物不同组织器官营养元素水平与变化是调整它们的生长速率以适应环境变化的一种策略(Marschner, 1995 ), 如干旱区植物相对于湿润区植物通常拥有更高的单位面积(或质量)叶N、P含量(Wright et al ., 2001 ; 李玉霖等, 2010 ), 植物地下根系的N、P含量低于地上部分的叶片, 但二者具有相似的N:P化学计量比(Yuan et al ., 2011 ).当前, 区域C、N、P化学计量格局及其驱动因素仍然是生态化学计量学的一个重要研究领域, 其结果不仅有助于揭示物种间化学计量的趋同与分异, 对于预测生态系统功能、过程对全球变化的响应亦具有重要意义(贺金生和韩兴国, 2010 ; Elser et al ., 2010 ).近年来, 国内学者在不同时空尺度, 针对森林(刘万德等, 2010 ; 崔高阳等, 2015 )、草地(Yu et al ., 2010 ; 雒明伟等, 2015 )、荒漠(李玉霖等, 2010 ; 黄菊莹和余海龙, 2016 )、湿地(Wang et al ., 2015 ; 聂兰琴等, 2016 )等生态系统类型开展了大量的研究.然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al ., 2010 ).同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014 ). ...
中国植物细根碳、氮、磷化学计量学的空间变化及其影响因子
2015
1
1995
... 碳(C)、氮(N)和磷(P)是植物基本营养元素, 在植物生长和各种生理调节机能中发挥着重要作用.C是构成植物体内干物质的最主要元素, 而N和P则分别与植物的光合作用和细胞分裂等重要生理活动有关(Niklas et al ., 2005 ).N:C和P:C与植物的相对生长率关系密切(Ågren, 2004 ), N:P则可以反映植物生长受N或P的限制情况(Koerselman & Meuleman, 1996 ; Elser et al ., 2000 ; Güsewell, 2004 ).植物不同组织器官营养元素水平与变化是调整它们的生长速率以适应环境变化的一种策略(Marschner, 1995 ), 如干旱区植物相对于湿润区植物通常拥有更高的单位面积(或质量)叶N、P含量(Wright et al ., 2001 ; 李玉霖等, 2010 ), 植物地下根系的N、P含量低于地上部分的叶片, 但二者具有相似的N:P化学计量比(Yuan et al ., 2011 ).当前, 区域C、N、P化学计量格局及其驱动因素仍然是生态化学计量学的一个重要研究领域, 其结果不仅有助于揭示物种间化学计量的趋同与分异, 对于预测生态系统功能、过程对全球变化的响应亦具有重要意义(贺金生和韩兴国, 2010 ; Elser et al ., 2010 ).近年来, 国内学者在不同时空尺度, 针对森林(刘万德等, 2010 ; 崔高阳等, 2015 )、草地(Yu et al ., 2010 ; 雒明伟等, 2015 )、荒漠(李玉霖等, 2010 ; 黄菊莹和余海龙, 2016 )、湿地(Wang et al ., 2015 ; 聂兰琴等, 2016 )等生态系统类型开展了大量的研究.然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al ., 2010 ).同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014 ). ...
鄱阳湖湿地优势植物叶片-凋落物-土壤碳氮磷化学计量特征
1
2016
... 碳(C)、氮(N)和磷(P)是植物基本营养元素, 在植物生长和各种生理调节机能中发挥着重要作用.C是构成植物体内干物质的最主要元素, 而N和P则分别与植物的光合作用和细胞分裂等重要生理活动有关(Niklas et al ., 2005 ).N:C和P:C与植物的相对生长率关系密切(Ågren, 2004 ), N:P则可以反映植物生长受N或P的限制情况(Koerselman & Meuleman, 1996 ; Elser et al ., 2000 ; Güsewell, 2004 ).植物不同组织器官营养元素水平与变化是调整它们的生长速率以适应环境变化的一种策略(Marschner, 1995 ), 如干旱区植物相对于湿润区植物通常拥有更高的单位面积(或质量)叶N、P含量(Wright et al ., 2001 ; 李玉霖等, 2010 ), 植物地下根系的N、P含量低于地上部分的叶片, 但二者具有相似的N:P化学计量比(Yuan et al ., 2011 ).当前, 区域C、N、P化学计量格局及其驱动因素仍然是生态化学计量学的一个重要研究领域, 其结果不仅有助于揭示物种间化学计量的趋同与分异, 对于预测生态系统功能、过程对全球变化的响应亦具有重要意义(贺金生和韩兴国, 2010 ; Elser et al ., 2010 ).近年来, 国内学者在不同时空尺度, 针对森林(刘万德等, 2010 ; 崔高阳等, 2015 )、草地(Yu et al ., 2010 ; 雒明伟等, 2015 )、荒漠(李玉霖等, 2010 ; 黄菊莹和余海龙, 2016 )、湿地(Wang et al ., 2015 ; 聂兰琴等, 2016 )等生态系统类型开展了大量的研究.然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al ., 2010 ).同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014 ). ...
Nitrogen phosphorus leaf stoichiometry and the scaling of plant growth
1
2005
... 碳(C)、氮(N)和磷(P)是植物基本营养元素, 在植物生长和各种生理调节机能中发挥着重要作用.C是构成植物体内干物质的最主要元素, 而N和P则分别与植物的光合作用和细胞分裂等重要生理活动有关(Niklas et al ., 2005 ).N:C和P:C与植物的相对生长率关系密切(Ågren, 2004 ), N:P则可以反映植物生长受N或P的限制情况(Koerselman & Meuleman, 1996 ; Elser et al ., 2000 ; Güsewell, 2004 ).植物不同组织器官营养元素水平与变化是调整它们的生长速率以适应环境变化的一种策略(Marschner, 1995 ), 如干旱区植物相对于湿润区植物通常拥有更高的单位面积(或质量)叶N、P含量(Wright et al ., 2001 ; 李玉霖等, 2010 ), 植物地下根系的N、P含量低于地上部分的叶片, 但二者具有相似的N:P化学计量比(Yuan et al ., 2011 ).当前, 区域C、N、P化学计量格局及其驱动因素仍然是生态化学计量学的一个重要研究领域, 其结果不仅有助于揭示物种间化学计量的趋同与分异, 对于预测生态系统功能、过程对全球变化的响应亦具有重要意义(贺金生和韩兴国, 2010 ; Elser et al ., 2010 ).近年来, 国内学者在不同时空尺度, 针对森林(刘万德等, 2010 ; 崔高阳等, 2015 )、草地(Yu et al ., 2010 ; 雒明伟等, 2015 )、荒漠(李玉霖等, 2010 ; 黄菊莹和余海龙, 2016 )、湿地(Wang et al ., 2015 ; 聂兰琴等, 2016 )等生态系统类型开展了大量的研究.然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al ., 2010 ).同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014 ). ...
Fine root architecture of nine North American trees
1
2002
... 近年来, 越来越多的研究尝试从植物功能性状或资源经济谱的角度来探讨不同器官之间的性状关联, 相关研究不仅有助于理解植物各性状之间相互作用的机制(Eviner & Chapin, 2003 )、植物生长过程中对资源的利用和分配(Kerkhoff et al ., 2006 ), 还对进一步预测植物对环境变化的响应具有重要意义.本研究发现单叶蔓荆各器官的养分元素及化学计量关系的相关性主要集中在地上部分, 即花、叶、枝条、匍匐茎之间, 其中尤以枝条与匍匐茎之间表现明显, 二者除了N:P没有达到显著相关之外, 其他的养分元素及其化学计量关系均达到了显著性相关(表2 ), 显示了二者之间在养分存储与运输之间的密切关系.在植物的所有器官中, 叶片与细根之间的性状关联最受关注.一些大尺度多物种间的研究表明叶片和细根的N含量(Craine & Lee, 2003 ; Tjoelker et al ., 2005 )、P含量(Kerkhoff et al ., 2006 ; Geng et al ., 2014 )均呈显著正相关.本研究未发现叶片与细根之间的3种元素及其化学计量关系显著相关, 一个可能的原因是叶片与细根性状的关联主要发生在种间, 小尺度上植物生境条件相对一致, 种内个体间功能性状因变异幅度较小而关联不明显(徐冰等, 2010 ).另一种可能的原因是本研究中的细根样品的采集是以直径<2 mm为标准的.由于根系在发育过程中形成明显的分枝结构, 具有高度的形态异质性(Pregitzer et al ., 2002 ; 王政权和郭大立, 2008 ), 直径<2 mm的细根并不是一个均质的单一组分, 而是由形态、寿命、生理差异巨大的不同个体根组成的高度异质的混合体(Pregitzer et al ., 2002; Guo et al ., 2008 ), 从而影响了与叶片之间相应元素的相关关系. ...
The world-wide “fast-slow” plant economics spectrum: A traits manifesto
1
2014
... 碳(C)、氮(N)和磷(P)是植物基本营养元素, 在植物生长和各种生理调节机能中发挥着重要作用.C是构成植物体内干物质的最主要元素, 而N和P则分别与植物的光合作用和细胞分裂等重要生理活动有关(Niklas et al ., 2005 ).N:C和P:C与植物的相对生长率关系密切(Ågren, 2004 ), N:P则可以反映植物生长受N或P的限制情况(Koerselman & Meuleman, 1996 ; Elser et al ., 2000 ; Güsewell, 2004 ).植物不同组织器官营养元素水平与变化是调整它们的生长速率以适应环境变化的一种策略(Marschner, 1995 ), 如干旱区植物相对于湿润区植物通常拥有更高的单位面积(或质量)叶N、P含量(Wright et al ., 2001 ; 李玉霖等, 2010 ), 植物地下根系的N、P含量低于地上部分的叶片, 但二者具有相似的N:P化学计量比(Yuan et al ., 2011 ).当前, 区域C、N、P化学计量格局及其驱动因素仍然是生态化学计量学的一个重要研究领域, 其结果不仅有助于揭示物种间化学计量的趋同与分异, 对于预测生态系统功能、过程对全球变化的响应亦具有重要意义(贺金生和韩兴国, 2010 ; Elser et al ., 2010 ).近年来, 国内学者在不同时空尺度, 针对森林(刘万德等, 2010 ; 崔高阳等, 2015 )、草地(Yu et al ., 2010 ; 雒明伟等, 2015 )、荒漠(李玉霖等, 2010 ; 黄菊莹和余海龙, 2016 )、湿地(Wang et al ., 2015 ; 聂兰琴等, 2016 )等生态系统类型开展了大量的研究.然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al ., 2010 ).同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014 ). ...
1
2002
... 上述GLM分析结果表明: 相对于沙化程度的变化所引起的植物生境条件, 如土壤水分、养分供给的变化, 不同器官的生理功能差异是影响单叶蔓荆C、N、P含量及化学计量比的主导因子.化学计量内稳性(stoichiometric homoeostasis)是生态化学计量学存在的基础(Sterner & Elser, 2002 ).Elser等(2010)认为尽管C、N、P化学计量比具有较大的时空变异, 但生物有机体仍保持极高的化学计量内稳性.化学计量内稳性通过调节生物对环境因子的响应, 成为生态系统结构、功能和稳定性维持的重要机制(Yu et al ., 2010 ).本研究中3种元素的化学计量关系中C:N、N:P在不同沙化程度区均未表现出显著差异.这一结果表明单叶蔓荆能够在沙山不同沙化区形成稳定的群落并保持较高的优势度, 化学计量内稳性很可能是其中非常重要的维持机制, 但这一机制仍然有待于进一步进行植物养分与土壤养分, 特别是速效养分之间关系的动态监测来加以验证.以往研究表明植物叶片与细根具有相似的N:P化学计量比(Yuan et al ., 2011 ), 同样, 本研究也发现N、P养分元素虽然在单叶蔓荆不同器官中的含量具有显著差异, 但是具有相似的N:P化学计量比, 保持了相对稳定性. ...
Linking leaf and root trait syndromes among 39 grassland and savannah species
1
2005
... 近年来, 越来越多的研究尝试从植物功能性状或资源经济谱的角度来探讨不同器官之间的性状关联, 相关研究不仅有助于理解植物各性状之间相互作用的机制(Eviner & Chapin, 2003 )、植物生长过程中对资源的利用和分配(Kerkhoff et al ., 2006 ), 还对进一步预测植物对环境变化的响应具有重要意义.本研究发现单叶蔓荆各器官的养分元素及化学计量关系的相关性主要集中在地上部分, 即花、叶、枝条、匍匐茎之间, 其中尤以枝条与匍匐茎之间表现明显, 二者除了N:P没有达到显著相关之外, 其他的养分元素及其化学计量关系均达到了显著性相关(表2 ), 显示了二者之间在养分存储与运输之间的密切关系.在植物的所有器官中, 叶片与细根之间的性状关联最受关注.一些大尺度多物种间的研究表明叶片和细根的N含量(Craine & Lee, 2003 ; Tjoelker et al ., 2005 )、P含量(Kerkhoff et al ., 2006 ; Geng et al ., 2014 )均呈显著正相关.本研究未发现叶片与细根之间的3种元素及其化学计量关系显著相关, 一个可能的原因是叶片与细根性状的关联主要发生在种间, 小尺度上植物生境条件相对一致, 种内个体间功能性状因变异幅度较小而关联不明显(徐冰等, 2010 ).另一种可能的原因是本研究中的细根样品的采集是以直径<2 mm为标准的.由于根系在发育过程中形成明显的分枝结构, 具有高度的形态异质性(Pregitzer et al ., 2002 ; 王政权和郭大立, 2008 ), 直径<2 mm的细根并不是一个均质的单一组分, 而是由形态、寿命、生理差异巨大的不同个体根组成的高度异质的混合体(Pregitzer et al ., 2002; Guo et al ., 2008 ), 从而影响了与叶片之间相应元素的相关关系. ...
Plant invasive success associated with higher N-use efficiency and stoichiometric shifts in the soil-plant system in the Minjiang River tidal estuarine wetlands of China
1
2015
... 碳(C)、氮(N)和磷(P)是植物基本营养元素, 在植物生长和各种生理调节机能中发挥着重要作用.C是构成植物体内干物质的最主要元素, 而N和P则分别与植物的光合作用和细胞分裂等重要生理活动有关(Niklas et al ., 2005 ).N:C和P:C与植物的相对生长率关系密切(Ågren, 2004 ), N:P则可以反映植物生长受N或P的限制情况(Koerselman & Meuleman, 1996 ; Elser et al ., 2000 ; Güsewell, 2004 ).植物不同组织器官营养元素水平与变化是调整它们的生长速率以适应环境变化的一种策略(Marschner, 1995 ), 如干旱区植物相对于湿润区植物通常拥有更高的单位面积(或质量)叶N、P含量(Wright et al ., 2001 ; 李玉霖等, 2010 ), 植物地下根系的N、P含量低于地上部分的叶片, 但二者具有相似的N:P化学计量比(Yuan et al ., 2011 ).当前, 区域C、N、P化学计量格局及其驱动因素仍然是生态化学计量学的一个重要研究领域, 其结果不仅有助于揭示物种间化学计量的趋同与分异, 对于预测生态系统功能、过程对全球变化的响应亦具有重要意义(贺金生和韩兴国, 2010 ; Elser et al ., 2010 ).近年来, 国内学者在不同时空尺度, 针对森林(刘万德等, 2010 ; 崔高阳等, 2015 )、草地(Yu et al ., 2010 ; 雒明伟等, 2015 )、荒漠(李玉霖等, 2010 ; 黄菊莹和余海龙, 2016 )、湿地(Wang et al ., 2015 ; 聂兰琴等, 2016 )等生态系统类型开展了大量的研究.然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al ., 2010 ).同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014 ). ...
根系生态学
1
2008
... 近年来, 越来越多的研究尝试从植物功能性状或资源经济谱的角度来探讨不同器官之间的性状关联, 相关研究不仅有助于理解植物各性状之间相互作用的机制(Eviner & Chapin, 2003 )、植物生长过程中对资源的利用和分配(Kerkhoff et al ., 2006 ), 还对进一步预测植物对环境变化的响应具有重要意义.本研究发现单叶蔓荆各器官的养分元素及化学计量关系的相关性主要集中在地上部分, 即花、叶、枝条、匍匐茎之间, 其中尤以枝条与匍匐茎之间表现明显, 二者除了N:P没有达到显著相关之外, 其他的养分元素及其化学计量关系均达到了显著性相关(表2 ), 显示了二者之间在养分存储与运输之间的密切关系.在植物的所有器官中, 叶片与细根之间的性状关联最受关注.一些大尺度多物种间的研究表明叶片和细根的N含量(Craine & Lee, 2003 ; Tjoelker et al ., 2005 )、P含量(Kerkhoff et al ., 2006 ; Geng et al ., 2014 )均呈显著正相关.本研究未发现叶片与细根之间的3种元素及其化学计量关系显著相关, 一个可能的原因是叶片与细根性状的关联主要发生在种间, 小尺度上植物生境条件相对一致, 种内个体间功能性状因变异幅度较小而关联不明显(徐冰等, 2010 ).另一种可能的原因是本研究中的细根样品的采集是以直径<2 mm为标准的.由于根系在发育过程中形成明显的分枝结构, 具有高度的形态异质性(Pregitzer et al ., 2002 ; 王政权和郭大立, 2008 ), 直径<2 mm的细根并不是一个均质的单一组分, 而是由形态、寿命、生理差异巨大的不同个体根组成的高度异质的混合体(Pregitzer et al ., 2002; Guo et al ., 2008 ), 从而影响了与叶片之间相应元素的相关关系. ...
Low rainfall-induced shift in leaf trait relationship within species along a semi-arid sandy land transect in northern China
2
2011
... 由于降水、土壤养分等环境梯度而引起的植物功能性状之间关系的策略位移现象不仅出现在种间水平上(Wright et al ., 2001 ; 李永华等, 2005 ; Wei et al ., 2011 ), 也出现在种内水平上(Wei et al ., 2011 ).但这一现象是否具有普遍性目前仍不清楚, 例如, 胡梦瑶等(2012)并没有发现紫花针茅(Stipa purpurea )在不同降水梯度下具有明显的策略位移现象.此外, 上述功能性状的位移现象多出现于较大尺度下的环境梯度中, 局地尺度上是否也存在类似的现象仍缺乏研究.本研究区尽管重度、中度与轻度沙化区土壤有机质、全氮、全磷等养分, 以及地表植被覆盖度等存在显著差异(表1 ), 但未发现单叶蔓荆N mass -P mass 线性关系的斜率与截距在3种沙化程度区发生显著变化(表3 ), 表明单叶蔓荆这一对功能性状关系在不同沙化区未发生明显的位移现象.成对植物功能性状关系发生策略位移主要是由于环境梯度对其中一个功能性状影响更大, 例如, Wei等(2011)发现低降水区的油蒿(Artemisia ordosica )种群N mass 高于较高降水区, 但比叶面积差异不大.本研究中N mass -P mass 在不同沙化程度区未出现明显的策略位移现象, 表明土壤沙化所引起的水、肥等限制因素可能共同作用于这两个性状, 使得植物N mass 与P mass 发生同步变化, 同时这也进一步佐证了N:P化学计量关系在不同沙化区的相对稳定.此外, 本研究结果也表明局地尺度种内水平上很可能因为环境梯度过小而不具备发生N mass -P mass 关系的位移现象. ...
... ), 也出现在种内水平上(Wei et al ., 2011 ).但这一现象是否具有普遍性目前仍不清楚, 例如, 胡梦瑶等(2012)并没有发现紫花针茅(Stipa purpurea )在不同降水梯度下具有明显的策略位移现象.此外, 上述功能性状的位移现象多出现于较大尺度下的环境梯度中, 局地尺度上是否也存在类似的现象仍缺乏研究.本研究区尽管重度、中度与轻度沙化区土壤有机质、全氮、全磷等养分, 以及地表植被覆盖度等存在显著差异(表1 ), 但未发现单叶蔓荆N mass -P mass 线性关系的斜率与截距在3种沙化程度区发生显著变化(表3 ), 表明单叶蔓荆这一对功能性状关系在不同沙化区未发生明显的位移现象.成对植物功能性状关系发生策略位移主要是由于环境梯度对其中一个功能性状影响更大, 例如, Wei等(2011)发现低降水区的油蒿(Artemisia ordosica )种群N mass 高于较高降水区, 但比叶面积差异不大.本研究中N mass -P mass 在不同沙化程度区未出现明显的策略位移现象, 表明土壤沙化所引起的水、肥等限制因素可能共同作用于这两个性状, 使得植物N mass 与P mass 发生同步变化, 同时这也进一步佐证了N:P化学计量关系在不同沙化区的相对稳定.此外, 本研究结果也表明局地尺度种内水平上很可能因为环境梯度过小而不具备发生N mass -P mass 关系的位移现象. ...
Strategy shifts in leaf physiology, structure and nutrient content between species of high -and low-rainfall and high -and low-nutrient habitats
4
2001
... 碳(C)、氮(N)和磷(P)是植物基本营养元素, 在植物生长和各种生理调节机能中发挥着重要作用.C是构成植物体内干物质的最主要元素, 而N和P则分别与植物的光合作用和细胞分裂等重要生理活动有关(Niklas et al ., 2005 ).N:C和P:C与植物的相对生长率关系密切(Ågren, 2004 ), N:P则可以反映植物生长受N或P的限制情况(Koerselman & Meuleman, 1996 ; Elser et al ., 2000 ; Güsewell, 2004 ).植物不同组织器官营养元素水平与变化是调整它们的生长速率以适应环境变化的一种策略(Marschner, 1995 ), 如干旱区植物相对于湿润区植物通常拥有更高的单位面积(或质量)叶N、P含量(Wright et al ., 2001 ; 李玉霖等, 2010 ), 植物地下根系的N、P含量低于地上部分的叶片, 但二者具有相似的N:P化学计量比(Yuan et al ., 2011 ).当前, 区域C、N、P化学计量格局及其驱动因素仍然是生态化学计量学的一个重要研究领域, 其结果不仅有助于揭示物种间化学计量的趋同与分异, 对于预测生态系统功能、过程对全球变化的响应亦具有重要意义(贺金生和韩兴国, 2010 ; Elser et al ., 2010 ).近年来, 国内学者在不同时空尺度, 针对森林(刘万德等, 2010 ; 崔高阳等, 2015 )、草地(Yu et al ., 2010 ; 雒明伟等, 2015 )、荒漠(李玉霖等, 2010 ; 黄菊莹和余海龙, 2016 )、湿地(Wang et al ., 2015 ; 聂兰琴等, 2016 )等生态系统类型开展了大量的研究.然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al ., 2010 ).同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014 ). ...
... 单叶蔓荆作为鄱阳湖湖滨沙山植被恢复过程的先锋物种, 广布于沙山不同程度沙化区, 为进一步说明研究区单叶蔓荆不同器官的养分含量及其化学计量关系特征, 我们把单叶蔓荆地上部分的叶片、地下部分的细根与其他荒漠区植物, 以及全球、中国大样本研究中的植物养分平均值做了比较.本研究区单叶蔓荆叶N (19.66 mg·g-1 )、P (1.24 mg·g-1 )含量明显低于中国北方典型荒漠及荒漠化地区植物N (24.45 mg·g-1 )、P (1.74 mg·g-1 )含量(李玉霖等,2010 ).此外, 叶N含量亦低于Killingbeck和Whitford (1996)报道的全球5大洲11个干旱荒漠中78种灌木叶N含量平均值(22 mg·g-1 ).本研究区虽属于南方典型荒漠, 但地处亚热带湿润区, 年降水量远高于其他干旱区荒漠, 这可能是单叶蔓荆叶N、P含量低于其他干旱、半干旱区荒漠植物的重要原因, 因为湿润区植物相对于干旱区通常具有更低的叶N、P含量(Wright et al ., 2001 ; 李永华等, 2005 ). ...
... Kattge等(2011)搜集整理了全球植物性状数据库, 发现全球植物叶片C、N、P含量的平均值分别为476.1 mg·g-1 、17.4 mg·g-1 、1.23 mg·g-1 , C:N、N:P则分别为23.4、12.8.与上述全球叶性状数据库相比较, 单叶蔓荆叶C (449.5 mg·g-1 )含量偏低, 叶N (19.66 mg·g-1 )含量略高, 叶P (1.24 mg·g-1 )含量则高度一致; 叶片C:N (23.1)基本一致, N:P (16.8)明显偏高.单叶蔓荆细根C (386.3 mg·g-1 )、N (16.8 mg·g-1 )、P (1.32 mg·g-1 )含量与Yuan等(2011)报道的全球植物细根养分含量及化学计量比相比, C含量与C:N明显偏低, N、P含量明显偏高, 但N:P化学计量比基本一致.同样, 与马玉珠等(2015)报道的中国植物细根的C、N、P含量相比, 我们也发现本研究中的单叶蔓荆细根C含量明显偏低, N、P含量明显偏高.与全球及中国大样本数据相比, 单叶蔓荆的叶片及细根的N、P含量, 特别是N含量明显偏高, 一方面可能与上述数据库中的多数样本来自于森林中的乔木有关, 同时还可能与本研究区单叶蔓荆特殊的荒漠生境相关.研究区虽不具有干旱区荒漠的长期水分限制, 但因为水热不同期, 特别是夏秋季节的高温、少雨与地表的强蒸发, 容易形成季节性的水分胁迫.同时, 沙地土壤养分匮乏, 单叶蔓荆在水分与养分均存在限制的情形下增加叶片N投入可能是一种有效的适应策略, 因为叶N增加能够提升叶片内部光合作用酶的数量, 进一步提高叶片光合速率(Chapin et al ., 1987 ).此外, 叶N含量的增加还可以增加叶片内部非光合器官或组织氮的投入, 从而提高细胞内部的渗透压, 增强对植物体内水分的保护, 是一种有效的节水方式(Wright et al ., 2001 ). ...
... 由于降水、土壤养分等环境梯度而引起的植物功能性状之间关系的策略位移现象不仅出现在种间水平上(Wright et al ., 2001 ; 李永华等, 2005 ; Wei et al ., 2011 ), 也出现在种内水平上(Wei et al ., 2011 ).但这一现象是否具有普遍性目前仍不清楚, 例如, 胡梦瑶等(2012)并没有发现紫花针茅(Stipa purpurea )在不同降水梯度下具有明显的策略位移现象.此外, 上述功能性状的位移现象多出现于较大尺度下的环境梯度中, 局地尺度上是否也存在类似的现象仍缺乏研究.本研究区尽管重度、中度与轻度沙化区土壤有机质、全氮、全磷等养分, 以及地表植被覆盖度等存在显著差异(表1 ), 但未发现单叶蔓荆N mass -P mass 线性关系的斜率与截距在3种沙化程度区发生显著变化(表3 ), 表明单叶蔓荆这一对功能性状关系在不同沙化区未发生明显的位移现象.成对植物功能性状关系发生策略位移主要是由于环境梯度对其中一个功能性状影响更大, 例如, Wei等(2011)发现低降水区的油蒿(Artemisia ordosica )种群N mass 高于较高降水区, 但比叶面积差异不大.本研究中N mass -P mass 在不同沙化程度区未出现明显的策略位移现象, 表明土壤沙化所引起的水、肥等限制因素可能共同作用于这两个性状, 使得植物N mass 与P mass 发生同步变化, 同时这也进一步佐证了N:P化学计量关系在不同沙化区的相对稳定.此外, 本研究结果也表明局地尺度种内水平上很可能因为环境梯度过小而不具备发生N mass -P mass 关系的位移现象. ...
内蒙古锡林河流域典型草原植物叶片与细根性状在种间及种内水平上的关联
1
2010
... 近年来, 越来越多的研究尝试从植物功能性状或资源经济谱的角度来探讨不同器官之间的性状关联, 相关研究不仅有助于理解植物各性状之间相互作用的机制(Eviner & Chapin, 2003 )、植物生长过程中对资源的利用和分配(Kerkhoff et al ., 2006 ), 还对进一步预测植物对环境变化的响应具有重要意义.本研究发现单叶蔓荆各器官的养分元素及化学计量关系的相关性主要集中在地上部分, 即花、叶、枝条、匍匐茎之间, 其中尤以枝条与匍匐茎之间表现明显, 二者除了N:P没有达到显著相关之外, 其他的养分元素及其化学计量关系均达到了显著性相关(表2 ), 显示了二者之间在养分存储与运输之间的密切关系.在植物的所有器官中, 叶片与细根之间的性状关联最受关注.一些大尺度多物种间的研究表明叶片和细根的N含量(Craine & Lee, 2003 ; Tjoelker et al ., 2005 )、P含量(Kerkhoff et al ., 2006 ; Geng et al ., 2014 )均呈显著正相关.本研究未发现叶片与细根之间的3种元素及其化学计量关系显著相关, 一个可能的原因是叶片与细根性状的关联主要发生在种间, 小尺度上植物生境条件相对一致, 种内个体间功能性状因变异幅度较小而关联不明显(徐冰等, 2010 ).另一种可能的原因是本研究中的细根样品的采集是以直径<2 mm为标准的.由于根系在发育过程中形成明显的分枝结构, 具有高度的形态异质性(Pregitzer et al ., 2002 ; 王政权和郭大立, 2008 ), 直径<2 mm的细根并不是一个均质的单一组分, 而是由形态、寿命、生理差异巨大的不同个体根组成的高度异质的混合体(Pregitzer et al ., 2002; Guo et al ., 2008 ), 从而影响了与叶片之间相应元素的相关关系. ...
Linking stoichiometric homoeostasis with ecosystem structure, functioning and stability
3
2010
... 碳(C)、氮(N)和磷(P)是植物基本营养元素, 在植物生长和各种生理调节机能中发挥着重要作用.C是构成植物体内干物质的最主要元素, 而N和P则分别与植物的光合作用和细胞分裂等重要生理活动有关(Niklas et al ., 2005 ).N:C和P:C与植物的相对生长率关系密切(Ågren, 2004 ), N:P则可以反映植物生长受N或P的限制情况(Koerselman & Meuleman, 1996 ; Elser et al ., 2000 ; Güsewell, 2004 ).植物不同组织器官营养元素水平与变化是调整它们的生长速率以适应环境变化的一种策略(Marschner, 1995 ), 如干旱区植物相对于湿润区植物通常拥有更高的单位面积(或质量)叶N、P含量(Wright et al ., 2001 ; 李玉霖等, 2010 ), 植物地下根系的N、P含量低于地上部分的叶片, 但二者具有相似的N:P化学计量比(Yuan et al ., 2011 ).当前, 区域C、N、P化学计量格局及其驱动因素仍然是生态化学计量学的一个重要研究领域, 其结果不仅有助于揭示物种间化学计量的趋同与分异, 对于预测生态系统功能、过程对全球变化的响应亦具有重要意义(贺金生和韩兴国, 2010 ; Elser et al ., 2010 ).近年来, 国内学者在不同时空尺度, 针对森林(刘万德等, 2010 ; 崔高阳等, 2015 )、草地(Yu et al ., 2010 ; 雒明伟等, 2015 )、荒漠(李玉霖等, 2010 ; 黄菊莹和余海龙, 2016 )、湿地(Wang et al ., 2015 ; 聂兰琴等, 2016 )等生态系统类型开展了大量的研究.然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al ., 2010 ).同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014 ). ...
... 土地沙化长期以来一直是我国最为严重的生态环境问题(董光荣等, 1999 ), 不仅出现在我国北方干旱、半干旱地区, 亦出现于南方湖滨及河流下游冲积平原地区(朱震达和崔书红, 1996 ).以鄱阳湖区为例, 沙化土地面积多达3.89万hm2 (左长青, 1986 ).鄱阳湖沙化土地虽处于亚热带湿润区, 但水热不同期, 特别是8-10月的高温、少雨与地表强蒸发, 以及冬春季节固定的风向与强劲的风速是影响植被恢复的重要因素.近几十年来, 马鞭草科落叶小灌木单叶蔓荆(Vitex rotundifolia )被成功地应用于鄱阳湖沙化土地的植被恢复, 并且在不同沙化程度区形成稳定的群落.研究表明生物有机体具有极高的化学计量内稳性(Elser et al ., 2010 ), 并且通过调节生物对环境变化的响应, 化学计量内稳性成为生态系统结构、功能和稳定性维持的重要机制(Yu et al ., 2010 ).单叶蔓荆的养分及其化学计量关系在不同沙化程度区会发生怎样的变化?其N、P功能性状之间的关系是否因沙化梯度而发生策略位移?本研究通过测定鄱阳湖典型沙山重度、中度、轻度沙化区单叶蔓荆花、叶、枝条、匍匐茎、细根等部位的C、N、P含量, 分析各器官养分元素分配格局、化学计量比的差异与关联, 以及随沙化程度的变化, 试图探讨上述两个科学问题, 为进一步揭示单叶蔓荆的沙地适应机制提供科学依据, 为鄱阳湖沙化区植被恢复提供参考. ...
... 上述GLM分析结果表明: 相对于沙化程度的变化所引起的植物生境条件, 如土壤水分、养分供给的变化, 不同器官的生理功能差异是影响单叶蔓荆C、N、P含量及化学计量比的主导因子.化学计量内稳性(stoichiometric homoeostasis)是生态化学计量学存在的基础(Sterner & Elser, 2002 ).Elser等(2010)认为尽管C、N、P化学计量比具有较大的时空变异, 但生物有机体仍保持极高的化学计量内稳性.化学计量内稳性通过调节生物对环境因子的响应, 成为生态系统结构、功能和稳定性维持的重要机制(Yu et al ., 2010 ).本研究中3种元素的化学计量关系中C:N、N:P在不同沙化程度区均未表现出显著差异.这一结果表明单叶蔓荆能够在沙山不同沙化区形成稳定的群落并保持较高的优势度, 化学计量内稳性很可能是其中非常重要的维持机制, 但这一机制仍然有待于进一步进行植物养分与土壤养分, 特别是速效养分之间关系的动态监测来加以验证.以往研究表明植物叶片与细根具有相似的N:P化学计量比(Yuan et al ., 2011 ), 同样, 本研究也发现N、P养分元素虽然在单叶蔓荆不同器官中的含量具有显著差异, 但是具有相似的N:P化学计量比, 保持了相对稳定性. ...
Global-scale latitudinal patterns of plant fine-root nitrogen and phosphorus
2
2011
... 碳(C)、氮(N)和磷(P)是植物基本营养元素, 在植物生长和各种生理调节机能中发挥着重要作用.C是构成植物体内干物质的最主要元素, 而N和P则分别与植物的光合作用和细胞分裂等重要生理活动有关(Niklas et al ., 2005 ).N:C和P:C与植物的相对生长率关系密切(Ågren, 2004 ), N:P则可以反映植物生长受N或P的限制情况(Koerselman & Meuleman, 1996 ; Elser et al ., 2000 ; Güsewell, 2004 ).植物不同组织器官营养元素水平与变化是调整它们的生长速率以适应环境变化的一种策略(Marschner, 1995 ), 如干旱区植物相对于湿润区植物通常拥有更高的单位面积(或质量)叶N、P含量(Wright et al ., 2001 ; 李玉霖等, 2010 ), 植物地下根系的N、P含量低于地上部分的叶片, 但二者具有相似的N:P化学计量比(Yuan et al ., 2011 ).当前, 区域C、N、P化学计量格局及其驱动因素仍然是生态化学计量学的一个重要研究领域, 其结果不仅有助于揭示物种间化学计量的趋同与分异, 对于预测生态系统功能、过程对全球变化的响应亦具有重要意义(贺金生和韩兴国, 2010 ; Elser et al ., 2010 ).近年来, 国内学者在不同时空尺度, 针对森林(刘万德等, 2010 ; 崔高阳等, 2015 )、草地(Yu et al ., 2010 ; 雒明伟等, 2015 )、荒漠(李玉霖等, 2010 ; 黄菊莹和余海龙, 2016 )、湿地(Wang et al ., 2015 ; 聂兰琴等, 2016 )等生态系统类型开展了大量的研究.然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al ., 2010 ).同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014 ). ...
... 上述GLM分析结果表明: 相对于沙化程度的变化所引起的植物生境条件, 如土壤水分、养分供给的变化, 不同器官的生理功能差异是影响单叶蔓荆C、N、P含量及化学计量比的主导因子.化学计量内稳性(stoichiometric homoeostasis)是生态化学计量学存在的基础(Sterner & Elser, 2002 ).Elser等(2010)认为尽管C、N、P化学计量比具有较大的时空变异, 但生物有机体仍保持极高的化学计量内稳性.化学计量内稳性通过调节生物对环境因子的响应, 成为生态系统结构、功能和稳定性维持的重要机制(Yu et al ., 2010 ).本研究中3种元素的化学计量关系中C:N、N:P在不同沙化程度区均未表现出显著差异.这一结果表明单叶蔓荆能够在沙山不同沙化区形成稳定的群落并保持较高的优势度, 化学计量内稳性很可能是其中非常重要的维持机制, 但这一机制仍然有待于进一步进行植物养分与土壤养分, 特别是速效养分之间关系的动态监测来加以验证.以往研究表明植物叶片与细根具有相似的N:P化学计量比(Yuan et al ., 2011 ), 同样, 本研究也发现N、P养分元素虽然在单叶蔓荆不同器官中的含量具有显著差异, 但是具有相似的N:P化学计量比, 保持了相对稳定性. ...
中国沙漠
1
1996
... 土地沙化长期以来一直是我国最为严重的生态环境问题(董光荣等, 1999 ), 不仅出现在我国北方干旱、半干旱地区, 亦出现于南方湖滨及河流下游冲积平原地区(朱震达和崔书红, 1996 ).以鄱阳湖区为例, 沙化土地面积多达3.89万hm2 (左长青, 1986 ).鄱阳湖沙化土地虽处于亚热带湿润区, 但水热不同期, 特别是8-10月的高温、少雨与地表强蒸发, 以及冬春季节固定的风向与强劲的风速是影响植被恢复的重要因素.近几十年来, 马鞭草科落叶小灌木单叶蔓荆(Vitex rotundifolia )被成功地应用于鄱阳湖沙化土地的植被恢复, 并且在不同沙化程度区形成稳定的群落.研究表明生物有机体具有极高的化学计量内稳性(Elser et al ., 2010 ), 并且通过调节生物对环境变化的响应, 化学计量内稳性成为生态系统结构、功能和稳定性维持的重要机制(Yu et al ., 2010 ).单叶蔓荆的养分及其化学计量关系在不同沙化程度区会发生怎样的变化?其N、P功能性状之间的关系是否因沙化梯度而发生策略位移?本研究通过测定鄱阳湖典型沙山重度、中度、轻度沙化区单叶蔓荆花、叶、枝条、匍匐茎、细根等部位的C、N、P含量, 分析各器官养分元素分配格局、化学计量比的差异与关联, 以及随沙化程度的变化, 试图探讨上述两个科学问题, 为进一步揭示单叶蔓荆的沙地适应机制提供科学依据, 为鄱阳湖沙化区植被恢复提供参考. ...
鄱阳湖沙山成因及治理利用浅析
1
1986
... 土地沙化长期以来一直是我国最为严重的生态环境问题(董光荣等, 1999 ), 不仅出现在我国北方干旱、半干旱地区, 亦出现于南方湖滨及河流下游冲积平原地区(朱震达和崔书红, 1996 ).以鄱阳湖区为例, 沙化土地面积多达3.89万hm2 (左长青, 1986 ).鄱阳湖沙化土地虽处于亚热带湿润区, 但水热不同期, 特别是8-10月的高温、少雨与地表强蒸发, 以及冬春季节固定的风向与强劲的风速是影响植被恢复的重要因素.近几十年来, 马鞭草科落叶小灌木单叶蔓荆(Vitex rotundifolia )被成功地应用于鄱阳湖沙化土地的植被恢复, 并且在不同沙化程度区形成稳定的群落.研究表明生物有机体具有极高的化学计量内稳性(Elser et al ., 2010 ), 并且通过调节生物对环境变化的响应, 化学计量内稳性成为生态系统结构、功能和稳定性维持的重要机制(Yu et al ., 2010 ).单叶蔓荆的养分及其化学计量关系在不同沙化程度区会发生怎样的变化?其N、P功能性状之间的关系是否因沙化梯度而发生策略位移?本研究通过测定鄱阳湖典型沙山重度、中度、轻度沙化区单叶蔓荆花、叶、枝条、匍匐茎、细根等部位的C、N、P含量, 分析各器官养分元素分配格局、化学计量比的差异与关联, 以及随沙化程度的变化, 试图探讨上述两个科学问题, 为进一步揭示单叶蔓荆的沙地适应机制提供科学依据, 为鄱阳湖沙化区植被恢复提供参考. ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}