光是植物光合作用和生长发育的重要影响因子, 光强和光质的变化直接影响着植物的生长发育, 植物对光强、光质的变化也会做出灵活的响应(许大全, 2013 )。植物在长期的进化过程中, 由于不同植物接收的太阳辐射量存在着明显差异, 因此形成了自身特有的需光特性(张聪颖等, 2011 )。植物经过遮阴后, 叶片形态、光合生理参数会发生变化, 表现为耐阴植物通常拥有较薄的颜色较深的叶片、较低的暗呼吸速率、较高的光合效率、较高的光系统II (PSII)效率以及较低的PSI/PSII (Wu et al ., 2013 )。C3 植物在弱光下均通过减少分枝、增加株高, 向高空发展以截获更多光能(Poorther & Perez-Soba, 2000 )。弱光条件下叶绿素含量增多是为了捕获更多的光能, 而植物的色素变化对光照的响应是不一样的, 绝对的阴生植物其色素含量和构成都是不随光照条件变化而变化的, 阳生植物通过增加叶绿素含量来适应弱光条件, 中间型植物则是通过既改变叶绿素含量也改变叶绿素构成来适应光照变化(Murchie & Horton, 1998 )。当光照强度超过植物的光饱和点时, 植物会发生光抑制或光破坏, 在中等光(稍高于2倍生长光强, 生长光强: 300-350 μmol·m-2 ·s-1 )下, 大豆(Glycine max )叶片的光抑制主要是由于PSII的可逆失活或下调, 与捕光天线LHCII的可逆脱离有关; 在强光(4倍或7倍于生长光强)下, 会造成大豆叶片光破坏, 主要是PSII的不可逆破坏, 表现为叶绿素膜蛋白(DI)发生净损失(Cai & Xu, 2002 )。在没有其他环境胁迫因素存在的自然条件下, 即使太阳光足够强, 人们也不大容易看到植物发生不可逆光破坏的情况, 之所以如此, 是因为植物在长期的进化过程中已经形成一系列防御光破坏的机制, 例如, 捕光天线脱离反应中心复合体和状态转换(Hong & Xu, 1999 ), 加强对过剩光能的热耗散, 加强光合作用对光能的利用, 叶片和叶绿体的避光运动(Kagawa et al ., 2001 ; Kasahara et al ., 2002 ), 加强光呼吸等耗能代谢, 加强对活性氧的清除和DI蛋白的周转等(Xu & Sheng, 1999 )。在正常情况下, 这些机制足以保护光合机构免于过量光能可能引起的不可逆破坏。
叶绿素荧光与光合作用中各个反应过程紧密相关, 逆境对光合作用各过程产生的影响都可通过植物体内叶绿素荧光诱导动力学变化反映出来(崔晓伟等, 2011 ), 同时叶绿素荧光参数更反映植物“内在性”的特点, 而且它具有快速、灵敏、非破坏性测量等优点, 因而被视为研究植物光合作用与环境关系的内在探针(Nedbal et al ., 2000 ), 把叶片的荧光分析与气体交换测定技术结合起来, 不仅可以从生理学水平上更迅速、全面地了解光合机构的实际运转状况, 而且可以间接地从生物化学水平上了解电子传递速率及其与不同代谢过程的关系。
绣球(Hydrangea macrophylla )又名八仙花、粉团花, 是虎耳草科绣球属植物, 高1-4 m, 常于基部发出多数放射枝而形成一圆形灌丛, 叶纸质或近革质, 倒卵形或阔椭圆形, 花期6-8月, 属暖温带半耐寒性落叶小灌木(黄淑美等, 1995 )。开花时花团锦簇, 颜色丰富艳丽。花色会随着土壤pH值的变化而变成红色或蓝色。由于绣球观赏效果较好且耐阴, 既可种植于庭院、公园、绿地, 又能作盆花栽培, 在园林绿化、美化中应用广泛(殷丽青等, 2010 )。目前对绣球的研究已经涉及栽培管理、花色花期调控、快繁技术、抗性等方面, 但针对绣球的耐阴性研究鲜有报道。本文以绣球品种‘无尽夏新娘’为试验材料, 采用模拟自然光照环境的方法, 从光合特性及叶绿素荧光参数来探究绣球对弱光的适应程度, 旨在为绣球的栽培和园林应用提供理论依据。
1 材料和方法
1.1 试验材料
试验于2014-2015年在浙江农林大学试验大棚内进行, 以生长健壮、长势一致的绣球植株为材料, 为保证绣球基因型一致, 2014年9月选取生长整齐一致的插穗, 扦插于河沙中至生根, 10月选取60株长势良好且一致的生根苗移栽于上口径15.3 cm、下口径12.3 cm、高17 cm的容器, 单株定植, 栽培基质一致。容器苗置于连栋大棚内, 进行常规栽培管理。生长至2015年7月选用黑色遮阳网进行遮阴处理: 分别为遮光率50%、遮光率75%以及不遮光。本试验采用随机区组试验, 既: 将受试材料, 分别随机栽培于3个实验小区, 共3个光照处理, 每处理小区5株, 株行距各为20 cm, 重复3次, 共计9个小区, 肥水管理一致。60天后进行光合特性和叶绿素荧光参数的测定。
1.2 试验方法
1.2.1 光响应曲线的测定
测定时间为2015年10月上旬晴天条件下, 应用LI-6400便携式光合作用测定仪(LI-COR, Lincoln, USA)进行光响应曲线测定。CO2 浓度设定为400 μmol·mol-1 , 由系统CO2 注入系统自动控制; 叶室温度控制在(25 ± 0.5) ℃; 叶室相对湿度控制在(40 ± 5)%; 光强梯度分别为2 000、1 500、1 200、1 000、800、600、400、200、100、50、20、0 μmol·m-2 ·s-1 。选择中部同向新生叶片作为测定叶。首先对叶片进行15 min光诱导, 光强设置为1 000 μmol·m-2 ·s-1 。然后开启光响应自动测量程序, 每个不同光强值设定数据采集时间3 min后, 进行测量。
1.2.2 叶片气体交换参数的测定
测定时间同上, 应用LI-6400便携式光合作用测定仪进行气体交换参数测定。各参数设置与光响应曲线一致。光强设为1 000 μmol·m-2 ·s-1 , 每个处理选择绣球各3株, 每株分别选择3个中部同向新生叶片作为测定叶, 测定参数包括单叶净光合速率(P n )、蒸腾速率(T r )、气孔导度(G s )、胞间CO2 浓度(C i )、叶片温度(T l ); 主要环境影响因子, 包括水汽压亏缺(VPD )、大气CO2 浓度(C a )、光合有效辐射(PAR )、气温(T a )、相对湿度(RH )等参数。水分利用效率(WUE ) = P n /T r 。
1.2.3 叶绿素荧光参数的测定
用LI-6400便携式光合作用测定仪测定初始荧光(F o )、最大荧光(F m ), PSII最大光化学效率(F v /F m ), 光下最大荧光(F° m )和光下最小荧光(F° o )。测定F v /F m 前叶片充分暗适应30 min。选择中部同向新生叶片作为测定叶。每个处理选择绣球各3株, 每株测定3个叶片, 每叶片重复3次。计算吸收光能用于天线热耗散(D ) = (1 - F° v /F° m ) × 100%, 光化学耗散(P ) = q P × F° v /F° m × 100%, PSII反应中心非光化学能量耗散(E ) = (1 - q P ) × F° v /F° m × 100% (Ehlert & Hinchaok, 2008 ; 崔晓伟等, 2011 ), q P 为光化学淬灭系数。
1.3 数据分析
根据刘建锋等(2011) 的方法拟合光响应曲线, 模型表达式为:
式中, θ 为光响应曲线的曲角; AQY 为表观量子效率(mmol CO2 ·mol-1 photos); P nmax 为最大净光合速率(μmol CO2 ·m-2 ·s-1 ); R d 为暗呼吸速率(μmol CO2 · m-2 ·s-1 )。光补偿点(LCP, μmol·m-2 ·s-1 )和光饱和点(LSP, μmol·m-2 ·s-1 )是利用低光条件下(PAR ≤ 200 μmol·m-2 ·s-1 )的P n 与PAR 构建线性方程(P n = mPAR + n, LCP = - n /m , LSP = (P nmax - n )/m )所得。
利用Microsoft Excel 2007整理原始数据。用SPSS 22.0软件进行方差分析, 采用Duncan氏新复极差测验法进行多重比较。用光合计算4.0.1.软件辅助计算光响应曲线拟合值, 利用Origin 8.0作图。
2 结果
2.1 遮阴对光合-光响应曲线的影响
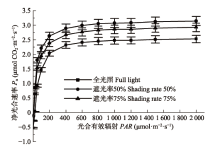
光响应曲线可反映植物P n 随PAR 增减的变化规律(李金鹏和董然, 2011 ), 由图1 可知, 3种光照处理下的绣球光响应曲线变化规律相似, 当PAR 为0时, P n 为负值, 随着PAR 的增强, P n 逐渐增加, 由负值转为正值, 当PAR 为400 μmol·m-2 ·s-1 时, P n 上升趋势减缓并趋向平稳, 光响应曲线结果表明, 遮阴提高了绣球的AQY 和P nmax , 并降低了LCP 、LSP 和R d 。由表1 可知, 50%遮光率条件下的绣球叶R d 比全光照下降低了31.4%, LCP 降低了57.4%, LSP 降低了9.5%, 这表明在遮阴环境下, 绣球可通过降低LCP 、LSP 来提高对弱光的利用能力, 并通过降低呼吸消耗以维持植株正常生长, 从而表现出了极强的适应能力。
图1 不同遮阴处理下绣球叶片的光响应曲线(平均值±标准偏差)。
Fig. 1 Light response curves of net photosynthetic rate (P n ) in Hydrangea macrophylla under different light treatments (mean ± SD ).
2.2 遮阴对叶片气体交换参数的影响
由表2 可知, 50%遮光率处理下的绣球叶片的P n 和T r 分别高于全光照和75%遮光率处理, 而C i 和WUE 低于全光照。50%遮光率处理叶的P n 、C i 、T r 和WUE 与全光照叶差异显著, 其G s 与后者差异不显著。说明P n 与T r 的变化一致, 遮阴增加了叶片的P n , 较高的P n 伴随着较高的T r 。50%遮光处理后, 叶片G s 没有发生变化, 但是WUE 降低了18.18%, 说明遮阴下的叶片需要消耗更多的水分以维持其高的净光合效率, 而C i 降低说明叶片固定C的能力提高。75%遮光率下的G s 与50%遮光率和全光照下的差异显著, 下降了50%, 说明过度遮阴影响绣球叶片气体交换能力, 从而引起叶片的P n 下降。由此得出, 绣球在适度遮阴后, 叶片会通过增加T r 和提高CO2 同化力来提高P n , 但过度遮阴会使叶片P n 下降, 原因是G s 降低使得叶片气体交换能力下降。
2.3 遮阴对叶绿素荧光参数的影响
叶绿素荧光是光合作用的探针, 通过荧光参数分析可以了解光合机构内部一系列重要的调节过程, F v /F m 是PSII光能转化率的指标(缴丽莉等, 2007 )。由表3 看出, 遮阴导致F v /F m 增加, 3种光照处理间差异显著, 全光照下F v /F m 低于50%遮光率处理, F o 高于50%遮光率处理, 根据张守仁(1999)的观点, F o 是判断PSII反应中心运转情况的重要指标, F o 上升表明PSII反应中心受到破坏或失活, F v /F m 下降是光抑制的重要特征, 因此推测全光照下的绣球叶片发生了光抑制。NPQ 的变化反映了非光化学耗散的能量, 其中包括了类囊体膜耗散热能, 是植物光合机构的自我保护机制(何炎红等, 2006 )。随着生长光照的减弱, NPQ 值在降低, 说明遮光处理降低了PSII天线色素吸收光能以热的形式耗散的比例, 提高了PSII吸收光能的利用效率。
2.4 叶片吸收光能的分配
植物叶片吸收的光能可分为3部分: 一是天线热耗散的能量(D ), 二是用于光化学反应的部分(P ), 三是反应中心有非光化学反应耗散的能量(E ) (Depuydt et al ., 2009 )。分析吸收光能的分配将有利于了解植物叶片对吸收光能的分配策略(胡文海等, 2015 )。由表4 可知, P 表现为: 50%遮光率 > 全光照> 75%遮光率。E 表现为: 75%遮光率 > 50%遮光率 > 全光照。D 表现为: 全光照和遮光率75%的D 值高于遮光率50%, 由此看出, 遮光率50%的绣球把吸收的光能较多的分配于光化学反应部分, 而用于天线热耗散的能量较少。说明在此处理下, 绣球叶片的光化学反应能量利用率最大, 这与NQP 变化一致。遮光率75%条件下, 绣球由于光照不足, 叶片吸收的光能用于参与光化学反应耗散的份额降低。而较高的E 值表明PSII光化学反应和保护性调节机制没有发挥作用, 积累了大量的激发能, 这有可能会对PSII反应中心产生破坏作用。全光照下, 绣球叶片D 值较高, 说明叶片此时具有较高的可调控生理代谢, 能够将过剩的光能较多地分配给热耗散途径。而热耗散比率的增加也反映了此时光能捕获效率降低, 推测植物有可能处于光抑制状态下(黄秋娴等, 2015 )。因此, 绣球处于饱和光环境时, 绣球叶片吸收的能量约70%用于热耗散, 约20%用于非光化学反应, 仅有4%的能量用于光化学反应, 绣球的主要光保护机制为PSII天线色素吸收光能通过热的形式耗散, 以此削弱反应中心过量的激发能。
3 讨论
LSP 和LCP 分别代表植物光合作用的光照强度上限和下限, 同时也代表植物叶片对强光和弱光的利用能力。耐阴植物的LCP 小于20 μmol·m-2 ·s-1 , LSP 为50-1β000μmol·m-2 ·s-1 或更低(Boardman, 1977 )。绣球的LCP 为6.71-15.75 μmol·m-2 ·s-1 , LSP 为278.14-307.42 μmol·m-2 ·s-1 。因此, 绣球属于耐阴植物, 遮阴处理下的绣球R d 低于全光照下, 这说明遮阴叶比全光照叶呼吸强度小, 绣球在遮阴环境下能够通过降低LCP 和LSP 来增强对弱光的利用能力, 与王凯等(2009)的研究结果类似。AQY 反映了叶片在弱光环境下的光合能力, 其值越大, 表明植物吸收与转换光能的色素蛋白复合体越多, 植物利用弱光的能力越强(Richardson & Berlyn, 2002 )。绣球在弱光环境下的AQY 较全光照环境下高, 说明绣球对弱光有较强的适应性。
正常情况下, 没有遭受环境胁迫并经过充分暗适应的植物叶片F v /F m 是比较恒定的, 一般为0.80- 0.85 (许大全, 2013 ), 但是本实验3种光照处理下绣球的F v /F m 平均值仅为0.63, 显著低于正常值, P n 值也偏低, 分析原因可能是: 在试验中, 无法准确知道绣球光诱导期的时间, 使得弱光下生长的植株较长时间暴露在全光照下, 导致光合机构的过剩激发能大幅增加, 最大光化学效率迅速降低, 因而出现光抑制现象(李志真等, 2014 )。由F° v /F° m 、F v /F m 和F o 的降低, 可判断此时绣球的热耗散机制是依赖叶黄素循环的能量耗散(许大全, 2013 )。另一方面, 由表3 和表4 看出, 遮光率75%的绣球E 值、F o 最大, 说明较
其他两种光照处理, 受到的光抑制更加严重。一个原因是E 值的增加会使单线态叶绿素(1 Chl)不能及时返回三线态叶绿素(3 Chl), 1 Chl的积累会将能量传递给O2 , 形成活性极强的单线态氧(1 O2 ), 这会引起光合机构的失活甚至破坏(Asada, 1999 )。F o 的增加说明植物叶片类囊体膜受到损害, PSII反应中心失活(黄秋娴等, 2015 )。另一个原因是其阴生叶片的结构和生理特征均缺少对强光的适应性, 所以遮阴下生长的绣球叶片过剩激发能多, 光抑制程度也比直接生长在全日照下的绣球要严重。这符合3种光照处理下非光化学反应耗散变化趋势, 也与杨兴洪等(2001)的研究结果一致。光合作用诱导期的长短差别很大,短的仅有几分钟, 长的可达1-2 h。光诱导的形成, 主要是来自光合碳同化酶的活化, 这些酶需要光照作催化剂(许大全, 2003 )。因此, 对如何控制光照时间和光合同化酶的活性等因素使绣球达到合适的光合诱导期有待继续探究。
综上所述, 绣球为耐半阴植物, 适度遮阴能够提高其F v /F m , 同时降低热耗散比例, 绣球在全光照条件下很容易导致PSII反应中心失活而出现光抑制, 光合作用效率大幅度降低。在强光下绣球会启动光保护机制, 是以热耗散途径为主的PSII调节性能量耗散, 遮阴75%处理限制了光合碳同化酶的活性, 绣球光合性能并非最大, 因此绣球的最佳光照环境为全光照的50%, 在郁闭度为0.5左右时, 光合作用捕获率最大, 有利于植物生长发育。
致谢 感谢浙江农林大学风景园林与建筑学院的韦孟琪、章毅在实验工作中给予的帮助。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Asada K 1999 ). The water-water cycle in chloroplasts: Scavenging of active oxygens and dissipation of excess photons.
Plant Biology, 50 , 601 -639 .
[本文引用: 1]
[2]
Boardman NK 1977 ). Comparative photosynthesis of sun and shade plants.
Plant Physiology, 28 , 355 -377 .
[本文引用: 1]
[3]
Cai SQ Xu DQ 2002 ). Light intensity-dependent reversible down-regulation and irreversible damage of PSII in soybean leaves.
Plant Science, 163 , 847 -853 .
[本文引用: 1]
[4]
Cui XW Gao J Zhang ZJ Yue XH Ma YJ 2011 ). Chlorophyll fluorescent characteristics of five dwarf bamboos.
Acta Agriculturae Universitatis Jiangxiensis, 33 , 726 -730 . (in Chinese with English abstract)[崔晓伟 , 高健 , 张志坚 , 岳祥华 , 马艳军 (2011 ). 5种地被竹叶绿素荧光特性研究
. 江西农业大学学报 , 33 , 726 -730 .]
[本文引用: 2]
[5]
Depuydt S Trenkamp S Fernie AR Elftieh S Renou JP Vuylsteke M Holsters M Vereecke D 2009 ). An integrated genomics approach to define niche establishment by
Rhodococcus fascians . Plant Physiology, 149 , 1366 -1386 .
[本文引用: 1]
[6]
Ehlert B Hinchaok DK 2008 ). Chlomphyll fluorescence imaging accurately quantifies freezing damage and cold acclimation responses in
Arabidaopsis leaves. Plant Methods, 4 , 1 -7 .
[本文引用: 1]
[7]
He YH Guo LS Tian YL 2006 ). Chlorophyll fluorescence quenching characteristics of seven coniferous and broadleaved species in different light intensities.
Scientia Silvae Sinicae, 42 (2 ), 27 -31 . (in Chinese with English abstract)[何炎红 , 郭连生 , 田有亮 (2006 ). 7种针阔叶树种不同光照强度下叶绿素荧光猝灭特征
. 林业科学 , 42 (2 ), 27 -31 .]
[本文引用: 1]
[8]
Hong SS Xu DQ 1999 ). Reversible inactivation of PSII reaction centers and the dissociation of LHCII from PSII complex in soybean leaves.
Plant Science, 1147 , 111 -118 .
[本文引用: 1]
[9]
Hu WH Zhang SS Xiao YA Yan XH 2015 ). Physiological responses and photo-protective mechanisms of two
Rhododendron plants to natural sunlight after long term shading. Chinese Journal of Plant Ecology, 39 , 1093 -1100 . (in Chinese with English abstract)[胡文海 , 张斯斯 , 肖宜安 , 闫小红 (2015 ). 两种杜鹃花属植物对长期遮阴后全光照环境的生理响应及其光保护机制
. 植物生态学报 , 39 , 1093 -1100 .]
[本文引用: 1]
[10]
Huang QX Zhao S Liu CM Li YL 2015 ). Effects of shading treatments on chlorophyll fluorescence characteristics of
Sabina vulgaris seedlings grown in iron tailings media. Scientia Silvae Sinicae, 51 (6 ), 18 -25 . (in Chinese with English abstract)[黄秋娴 , 赵顺 , 刘春梅 , 李玉灵 (2015 ). 遮荫处理对铁尾矿基质臭柏实生苗快速叶绿素荧光特性的影响
. 林业科学 , 51 (6 ), 18 -25 .]
[本文引用: 2]
[11]
Huang SM Wei ZF Lu LD Pan JT 1995). Flora Republicae Popularis Sinicae . Science Press , Beijing . 226 -227 . (in Chinese with English abstract)[黄淑美 , 卫兆芬 , 陆玲娣 , 潘锦堂 (1995). 中国植物志. 科学出版社 , 北京 . 226 -227 .]
[本文引用: 1]
[12]
Ji LL Lu BS Zhou RJ Bai ZY Liang HY 2007 ). Effects of shading on photosynthetic parameters of leaf in David maple (
Acer davidii Franch). Acta Horticulturae Sinica, 34 (1 ), 173 -178 . (in Chinese with English abstract)[缴丽莉 , 路丙社 , 周如久 , 白志英 , 梁海永 (2007 ). 遮光对青榨槭光合速率及叶绿素荧光参数的影响
. 园艺学报 , 34 (1 ), 173 -178 .]
[本文引用: 1]
[13]
Kagawa T Sakai T Suetsugu N Oikawa K Lshiguro S 2001 ). Arabidopsis NPL1: A phototropin homolog controlling the chloroplast high-light avoidance response.
Science, 291 , 2138 -2141 .
[本文引用: 1]
[14]
Kasahara M Kagawa T Oikawa , Suetsugu N Miyao M 2002 ). Chloroplast avoidance movement reduces photodamage in plant.
Nature, 420 , 829 -832 .
[本文引用: 1]
[15]
Li JP Dong R 2011 ). Diurnal change of photosynthetic characteristics of three
Hosta cultivars. Journal of Northeast Forestry University, 39 (10 ), 56 -58 . (in Chinese with English abstract)[李金鹏 , 董然 (2011 ). 3种彩叶玉簪光合日变化
. 东北林业大学学报 , 39 (10 ), 56 -58 .]
[本文引用: 1]
[16]
Li ZZ Liu DH Zhao SW Jiang CD Shi L 2014 ). Mechanisms of photoinhibition induced by high light in
Hosta grown outdoors. Chinese Journal of Plant Ecology, 38 , 720 -728 . (in Chinese with English abstract)[李志真 , 刘东焕 , 赵世伟 , 姜闯道 , 石雷 (2014 ). 环境强光诱导玉簪叶片光抑制的机制
. 植物生态学报 , 38 , 720 -728 .]
[本文引用: 1]
[17]
Liu JF Yang WJ Jing ZP Guo QS Jing JQ Xue L 2011 ). Effects of shading on photosynthetic characteristics and chlorophyll fluorescence parameters in leaves of the endangered plant
Thuja sutchuenensis . Acta Ecologica Sinica, 31 , 5999 -6004 . (in Chinese with English abstract)[刘建锋 , 杨文娟 , 江泽平 , 郭泉水 , 金江群 , 薛亮 (2011 ). 遮荫对濒危植物崖柏光合作用和叶绿素荧光参数的影响
. 生态学报 , 31 , 5999 -6004 .]
[本文引用: 1]
[18]
Murchie EH Horton P 1998 ). Contrasting patterns of photo-synthetic acclimation to the light environment are dependent on the differential expression of the responses to altered irradiance and spectral quality.
Plant, Cell & Environment, 21 , 139 -148 .
[本文引用: 1]
[19]
Nedbal L Soukupova J Kaftan D Whitmarsh J Trtilek M 2000 ). Kinetic imaging lf chlorophyll fluorescence using modulated light.
Photosynthesis Research, 66 , 3 -12 .
[本文引用: 1]
[20]
Poorther H Perez-Soba M 2000 ). The growth response of plants to elevated CO2 under non-optimal environmental conditions.
Oecologia, 27 , 595 -607 .
[本文引用: 1]
[21]
Richardson AD Berlyn GP 2002 ). Spectral reflectance and photosynthetic properties of Betula papyrifera (Betulaceae) leaves along an elevational gradient on Mt.
Mansfield. American Journal of Botany, 9 , 88 -94 .
[本文引用: 1]
[22]
Wang K Zhu JJ Yu LZ Sun YR Chen GH 2009 ). Effects of shading on the photosynthetic characteristics and light use efficiency of
Phellodendron amurense seedlings. Chinese Journal of Plant Ecology, 33 , 1003 -1012 . (in Chinese with English abstract)[王凯 , 朱教君 , 于立忠 , 孙一荣 , 陈光华 (2009 ). 遮阴对黄波罗幼苗的光合特性及光能利用效率的影响
. 植物生态学报 , 33 , 1003 -1012 .]
[23]
Wu CW Ke TS Chang YJ Chang YS 2013 ). Chlorophyll fluorescence and leaf-air temperature difference as potential shade-tolerance indexes of ornamental plants.
Journal of Computational & Theoretical Nanoscience, 19 , 3063 -3066 .
[本文引用: 1]
[24]
Xu DQ 2003 ). Several problems in the research of plant light stress.
Plant Physiology Communications, 39 , 493 -495 . (in Chinese with English abstract)[许大全 (2003 ). 植物光胁迫研究中的几个问题
. 植物生理学通讯 , 39 , 493 -495 .]
[本文引用: 1]
[25]
Xu DQ The Science of Photosynthesis .. Science Press , Beijing . 94 -95 . (in Chinese)[许大全 (2013). 光合作用学 . 科学出版社 , 北京 . 94 -95 .]
[本文引用: 3]
[26]
Xu DQ Sheng YK Light stress: Photoinhibition of photosynthesis in plants under natural conditions . In: Pessarakli M ed. Handbook of Plant and Crop Stress. Marcel Dekker , New York .315 -336 .
[本文引用: 1]
[27]
Yang XH Zou Q Wang W 2001 ). Photoinhibition in shaded cotton leaves after exposing to high light and the time course of its restoration.
Acta Botanica Sinica, 43 , 1255 -1259 . (in Chinese with English abstract)[杨兴洪 , 邹琦 , 王玮 (2001 ). 遮荫棉花转入强光后光合作用的光抑制及其恢复
. 植物学报 , 43 , 1255 -1259 .]
[28]
Yin LQ Hu Y Tang GJ Huang WC Li SZ Wang XQ 2010 ). In vitro culture and rapid propagation of
Hydrangea serrata ‘Preziosa’. Acta Agriculturae Shanghai, 26 (1 ), 38 -41 . (in Chinese with English abstract)[殷丽青 , 胡永 , 汤桂钧 , 黄卫昌 , 李世忠 , 王新其 (2010 ). 优良八仙花品种(Hydrangea serrata
. 上海农业学报 , 26 (1 ), 38 -41 .]
[本文引用: 1]
[29]
Zhang CY Fang YM Ji HL Ma XT 2011 ). Effects of shading on photosynthesis characteristics of
Photinia × frasery and Aucuba japonica var. variegate . Chinese Journal of Applied Ecology, 22 , 1743 -1749 . (in Chinese with English abstract)[张聪颖 , 方炎明 , 姬红利 , 马戌涛 (2011 ). 遮荫处理对红叶石楠和洒金桃叶珊瑚光合特性的影响
. 应用生态学报 , 22 , 1743 -1749 .]
[本文引用: 1]
[30]
Zhang SR 1999 ). A discussion on chlorophyll fluorescence kinetics parameters and their significance.
Chinese Bulletin of Botany, 16 , 444 -448 . (in Chinese with English abstract)[张守仁 (1999 ). 叶绿素荧光动力学参数的意义及讨论
. 植物学通报 , 16 , 444 -448 .]
The water-water cycle in chloroplasts: Scavenging of active oxygens and dissipation of excess photons.
1
1999
... 其他两种光照处理, 受到的光抑制更加严重.一个原因是E 值的增加会使单线态叶绿素(1 Chl)不能及时返回三线态叶绿素(3 Chl), 1 Chl的积累会将能量传递给O2 , 形成活性极强的单线态氧(1 O2 ), 这会引起光合机构的失活甚至破坏(Asada, 1999 ).F o 的增加说明植物叶片类囊体膜受到损害, PSII反应中心失活(黄秋娴等, 2015 ).另一个原因是其阴生叶片的结构和生理特征均缺少对强光的适应性, 所以遮阴下生长的绣球叶片过剩激发能多, 光抑制程度也比直接生长在全日照下的绣球要严重.这符合3种光照处理下非光化学反应耗散变化趋势, 也与杨兴洪等(2001)的研究结果一致.光合作用诱导期的长短差别很大,短的仅有几分钟, 长的可达1-2 h.光诱导的形成, 主要是来自光合碳同化酶的活化, 这些酶需要光照作催化剂(许大全, 2003 ).因此, 对如何控制光照时间和光合同化酶的活性等因素使绣球达到合适的光合诱导期有待继续探究. ...
Comparative photosynthesis of sun and shade plants.
1
1977
... LSP 和LCP 分别代表植物光合作用的光照强度上限和下限, 同时也代表植物叶片对强光和弱光的利用能力.耐阴植物的LCP 小于20 μmol·m-2 ·s-1 , LSP 为50-1β000μmol·m-2 ·s-1 或更低(Boardman, 1977 ).绣球的LCP 为6.71-15.75 μmol·m-2 ·s-1 , LSP 为278.14-307.42 μmol·m-2 ·s-1 .因此, 绣球属于耐阴植物, 遮阴处理下的绣球R d 低于全光照下, 这说明遮阴叶比全光照叶呼吸强度小, 绣球在遮阴环境下能够通过降低LCP 和LSP 来增强对弱光的利用能力, 与王凯等(2009)的研究结果类似.AQY 反映了叶片在弱光环境下的光合能力, 其值越大, 表明植物吸收与转换光能的色素蛋白复合体越多, 植物利用弱光的能力越强(Richardson & Berlyn, 2002 ).绣球在弱光环境下的AQY 较全光照环境下高, 说明绣球对弱光有较强的适应性. ...
Light intensity-dependent reversible down-regulation and irreversible damage of PSII in soybean leaves.
1
2002
... 光是植物光合作用和生长发育的重要影响因子, 光强和光质的变化直接影响着植物的生长发育, 植物对光强、光质的变化也会做出灵活的响应(许大全, 2013 ).植物在长期的进化过程中, 由于不同植物接收的太阳辐射量存在着明显差异, 因此形成了自身特有的需光特性(张聪颖等, 2011 ).植物经过遮阴后, 叶片形态、光合生理参数会发生变化, 表现为耐阴植物通常拥有较薄的颜色较深的叶片、较低的暗呼吸速率、较高的光合效率、较高的光系统II (PSII)效率以及较低的PSI/PSII (Wu et al ., 2013 ).C3 植物在弱光下均通过减少分枝、增加株高, 向高空发展以截获更多光能(Poorther & Perez-Soba, 2000 ).弱光条件下叶绿素含量增多是为了捕获更多的光能, 而植物的色素变化对光照的响应是不一样的, 绝对的阴生植物其色素含量和构成都是不随光照条件变化而变化的, 阳生植物通过增加叶绿素含量来适应弱光条件, 中间型植物则是通过既改变叶绿素含量也改变叶绿素构成来适应光照变化(Murchie & Horton, 1998 ).当光照强度超过植物的光饱和点时, 植物会发生光抑制或光破坏, 在中等光(稍高于2倍生长光强, 生长光强: 300-350 μmol·m-2 ·s-1 )下, 大豆(Glycine max )叶片的光抑制主要是由于PSII的可逆失活或下调, 与捕光天线LHCII的可逆脱离有关; 在强光(4倍或7倍于生长光强)下, 会造成大豆叶片光破坏, 主要是PSII的不可逆破坏, 表现为叶绿素膜蛋白(DI)发生净损失(Cai & Xu, 2002 ).在没有其他环境胁迫因素存在的自然条件下, 即使太阳光足够强, 人们也不大容易看到植物发生不可逆光破坏的情况, 之所以如此, 是因为植物在长期的进化过程中已经形成一系列防御光破坏的机制, 例如, 捕光天线脱离反应中心复合体和状态转换(Hong & Xu, 1999 ), 加强对过剩光能的热耗散, 加强光合作用对光能的利用, 叶片和叶绿体的避光运动(Kagawa et al ., 2001 ; Kasahara et al ., 2002 ), 加强光呼吸等耗能代谢, 加强对活性氧的清除和DI蛋白的周转等(Xu & Sheng, 1999 ).在正常情况下, 这些机制足以保护光合机构免于过量光能可能引起的不可逆破坏. ...
5种地被竹叶绿素荧光特性研究
2
2011
... 叶绿素荧光与光合作用中各个反应过程紧密相关, 逆境对光合作用各过程产生的影响都可通过植物体内叶绿素荧光诱导动力学变化反映出来(崔晓伟等, 2011 ), 同时叶绿素荧光参数更反映植物“内在性”的特点, 而且它具有快速、灵敏、非破坏性测量等优点, 因而被视为研究植物光合作用与环境关系的内在探针(Nedbal et al ., 2000 ), 把叶片的荧光分析与气体交换测定技术结合起来, 不仅可以从生理学水平上更迅速、全面地了解光合机构的实际运转状况, 而且可以间接地从生物化学水平上了解电子传递速率及其与不同代谢过程的关系. ...
... 用LI-6400便携式光合作用测定仪测定初始荧光(F o )、最大荧光(F m ), PSII最大光化学效率(F v /F m ), 光下最大荧光(F° m )和光下最小荧光(F° o ).测定F v /F m 前叶片充分暗适应30 min.选择中部同向新生叶片作为测定叶.每个处理选择绣球各3株, 每株测定3个叶片, 每叶片重复3次.计算吸收光能用于天线热耗散(D ) = (1 - F° v /F° m ) × 100%, 光化学耗散(P ) = q P × F° v /F° m × 100%, PSII反应中心非光化学能量耗散(E ) = (1 - q P ) × F° v /F° m × 100% (Ehlert & Hinchaok, 2008 ; 崔晓伟等, 2011 ), q P 为光化学淬灭系数. ...
An integrated genomics approach to define niche establishment by
1
2009
... 植物叶片吸收的光能可分为3部分: 一是天线热耗散的能量(D ), 二是用于光化学反应的部分(P ), 三是反应中心有非光化学反应耗散的能量(E ) (Depuydt et al ., 2009 ).分析吸收光能的分配将有利于了解植物叶片对吸收光能的分配策略(胡文海等, 2015 ).由表4 可知, P 表现为: 50%遮光率 > 全光照> 75%遮光率.E 表现为: 75%遮光率 > 50%遮光率 > 全光照.D 表现为: 全光照和遮光率75%的D 值高于遮光率50%, 由此看出, 遮光率50%的绣球把吸收的光能较多的分配于光化学反应部分, 而用于天线热耗散的能量较少.说明在此处理下, 绣球叶片的光化学反应能量利用率最大, 这与NQP 变化一致.遮光率75%条件下, 绣球由于光照不足, 叶片吸收的光能用于参与光化学反应耗散的份额降低.而较高的E 值表明PSII光化学反应和保护性调节机制没有发挥作用, 积累了大量的激发能, 这有可能会对PSII反应中心产生破坏作用.全光照下, 绣球叶片D 值较高, 说明叶片此时具有较高的可调控生理代谢, 能够将过剩的光能较多地分配给热耗散途径.而热耗散比率的增加也反映了此时光能捕获效率降低, 推测植物有可能处于光抑制状态下(黄秋娴等, 2015 ).因此, 绣球处于饱和光环境时, 绣球叶片吸收的能量约70%用于热耗散, 约20%用于非光化学反应, 仅有4%的能量用于光化学反应, 绣球的主要光保护机制为PSII天线色素吸收光能通过热的形式耗散, 以此削弱反应中心过量的激发能. ...
Chlomphyll fluorescence imaging accurately quantifies freezing damage and cold acclimation responses in
1
2008
... 用LI-6400便携式光合作用测定仪测定初始荧光(F o )、最大荧光(F m ), PSII最大光化学效率(F v /F m ), 光下最大荧光(F° m )和光下最小荧光(F° o ).测定F v /F m 前叶片充分暗适应30 min.选择中部同向新生叶片作为测定叶.每个处理选择绣球各3株, 每株测定3个叶片, 每叶片重复3次.计算吸收光能用于天线热耗散(D ) = (1 - F° v /F° m ) × 100%, 光化学耗散(P ) = q P × F° v /F° m × 100%, PSII反应中心非光化学能量耗散(E ) = (1 - q P ) × F° v /F° m × 100% (Ehlert & Hinchaok, 2008 ; 崔晓伟等, 2011 ), q P 为光化学淬灭系数. ...
7种针阔叶树种不同光照强度下叶绿素荧光猝灭特征
1
2006
... 叶绿素荧光是光合作用的探针, 通过荧光参数分析可以了解光合机构内部一系列重要的调节过程, F v /F m 是PSII光能转化率的指标(缴丽莉等, 2007 ).由表3 看出, 遮阴导致F v /F m 增加, 3种光照处理间差异显著, 全光照下F v /F m 低于50%遮光率处理, F o 高于50%遮光率处理, 根据张守仁(1999)的观点, F o 是判断PSII反应中心运转情况的重要指标, F o 上升表明PSII反应中心受到破坏或失活, F v /F m 下降是光抑制的重要特征, 因此推测全光照下的绣球叶片发生了光抑制.NPQ 的变化反映了非光化学耗散的能量, 其中包括了类囊体膜耗散热能, 是植物光合机构的自我保护机制(何炎红等, 2006 ).随着生长光照的减弱, NPQ 值在降低, 说明遮光处理降低了PSII天线色素吸收光能以热的形式耗散的比例, 提高了PSII吸收光能的利用效率. ...
Reversible inactivation of PSII reaction centers and the dissociation of LHCII from PSII complex in soybean leaves.
1
1999
... 光是植物光合作用和生长发育的重要影响因子, 光强和光质的变化直接影响着植物的生长发育, 植物对光强、光质的变化也会做出灵活的响应(许大全, 2013 ).植物在长期的进化过程中, 由于不同植物接收的太阳辐射量存在着明显差异, 因此形成了自身特有的需光特性(张聪颖等, 2011 ).植物经过遮阴后, 叶片形态、光合生理参数会发生变化, 表现为耐阴植物通常拥有较薄的颜色较深的叶片、较低的暗呼吸速率、较高的光合效率、较高的光系统II (PSII)效率以及较低的PSI/PSII (Wu et al ., 2013 ).C3 植物在弱光下均通过减少分枝、增加株高, 向高空发展以截获更多光能(Poorther & Perez-Soba, 2000 ).弱光条件下叶绿素含量增多是为了捕获更多的光能, 而植物的色素变化对光照的响应是不一样的, 绝对的阴生植物其色素含量和构成都是不随光照条件变化而变化的, 阳生植物通过增加叶绿素含量来适应弱光条件, 中间型植物则是通过既改变叶绿素含量也改变叶绿素构成来适应光照变化(Murchie & Horton, 1998 ).当光照强度超过植物的光饱和点时, 植物会发生光抑制或光破坏, 在中等光(稍高于2倍生长光强, 生长光强: 300-350 μmol·m-2 ·s-1 )下, 大豆(Glycine max )叶片的光抑制主要是由于PSII的可逆失活或下调, 与捕光天线LHCII的可逆脱离有关; 在强光(4倍或7倍于生长光强)下, 会造成大豆叶片光破坏, 主要是PSII的不可逆破坏, 表现为叶绿素膜蛋白(DI)发生净损失(Cai & Xu, 2002 ).在没有其他环境胁迫因素存在的自然条件下, 即使太阳光足够强, 人们也不大容易看到植物发生不可逆光破坏的情况, 之所以如此, 是因为植物在长期的进化过程中已经形成一系列防御光破坏的机制, 例如, 捕光天线脱离反应中心复合体和状态转换(Hong & Xu, 1999 ), 加强对过剩光能的热耗散, 加强光合作用对光能的利用, 叶片和叶绿体的避光运动(Kagawa et al ., 2001 ; Kasahara et al ., 2002 ), 加强光呼吸等耗能代谢, 加强对活性氧的清除和DI蛋白的周转等(Xu & Sheng, 1999 ).在正常情况下, 这些机制足以保护光合机构免于过量光能可能引起的不可逆破坏. ...
两种杜鹃花属植物对长期遮阴后全光照环境的生理响应及其光保护机制
1
2015
... 植物叶片吸收的光能可分为3部分: 一是天线热耗散的能量(D ), 二是用于光化学反应的部分(P ), 三是反应中心有非光化学反应耗散的能量(E ) (Depuydt et al ., 2009 ).分析吸收光能的分配将有利于了解植物叶片对吸收光能的分配策略(胡文海等, 2015 ).由表4 可知, P 表现为: 50%遮光率 > 全光照> 75%遮光率.E 表现为: 75%遮光率 > 50%遮光率 > 全光照.D 表现为: 全光照和遮光率75%的D 值高于遮光率50%, 由此看出, 遮光率50%的绣球把吸收的光能较多的分配于光化学反应部分, 而用于天线热耗散的能量较少.说明在此处理下, 绣球叶片的光化学反应能量利用率最大, 这与NQP 变化一致.遮光率75%条件下, 绣球由于光照不足, 叶片吸收的光能用于参与光化学反应耗散的份额降低.而较高的E 值表明PSII光化学反应和保护性调节机制没有发挥作用, 积累了大量的激发能, 这有可能会对PSII反应中心产生破坏作用.全光照下, 绣球叶片D 值较高, 说明叶片此时具有较高的可调控生理代谢, 能够将过剩的光能较多地分配给热耗散途径.而热耗散比率的增加也反映了此时光能捕获效率降低, 推测植物有可能处于光抑制状态下(黄秋娴等, 2015 ).因此, 绣球处于饱和光环境时, 绣球叶片吸收的能量约70%用于热耗散, 约20%用于非光化学反应, 仅有4%的能量用于光化学反应, 绣球的主要光保护机制为PSII天线色素吸收光能通过热的形式耗散, 以此削弱反应中心过量的激发能. ...
遮荫处理对铁尾矿基质臭柏实生苗快速叶绿素荧光特性的影响
2
2015
... 植物叶片吸收的光能可分为3部分: 一是天线热耗散的能量(D ), 二是用于光化学反应的部分(P ), 三是反应中心有非光化学反应耗散的能量(E ) (Depuydt et al ., 2009 ).分析吸收光能的分配将有利于了解植物叶片对吸收光能的分配策略(胡文海等, 2015 ).由表4 可知, P 表现为: 50%遮光率 > 全光照> 75%遮光率.E 表现为: 75%遮光率 > 50%遮光率 > 全光照.D 表现为: 全光照和遮光率75%的D 值高于遮光率50%, 由此看出, 遮光率50%的绣球把吸收的光能较多的分配于光化学反应部分, 而用于天线热耗散的能量较少.说明在此处理下, 绣球叶片的光化学反应能量利用率最大, 这与NQP 变化一致.遮光率75%条件下, 绣球由于光照不足, 叶片吸收的光能用于参与光化学反应耗散的份额降低.而较高的E 值表明PSII光化学反应和保护性调节机制没有发挥作用, 积累了大量的激发能, 这有可能会对PSII反应中心产生破坏作用.全光照下, 绣球叶片D 值较高, 说明叶片此时具有较高的可调控生理代谢, 能够将过剩的光能较多地分配给热耗散途径.而热耗散比率的增加也反映了此时光能捕获效率降低, 推测植物有可能处于光抑制状态下(黄秋娴等, 2015 ).因此, 绣球处于饱和光环境时, 绣球叶片吸收的能量约70%用于热耗散, 约20%用于非光化学反应, 仅有4%的能量用于光化学反应, 绣球的主要光保护机制为PSII天线色素吸收光能通过热的形式耗散, 以此削弱反应中心过量的激发能. ...
... 其他两种光照处理, 受到的光抑制更加严重.一个原因是E 值的增加会使单线态叶绿素(1 Chl)不能及时返回三线态叶绿素(3 Chl), 1 Chl的积累会将能量传递给O2 , 形成活性极强的单线态氧(1 O2 ), 这会引起光合机构的失活甚至破坏(Asada, 1999 ).F o 的增加说明植物叶片类囊体膜受到损害, PSII反应中心失活(黄秋娴等, 2015 ).另一个原因是其阴生叶片的结构和生理特征均缺少对强光的适应性, 所以遮阴下生长的绣球叶片过剩激发能多, 光抑制程度也比直接生长在全日照下的绣球要严重.这符合3种光照处理下非光化学反应耗散变化趋势, 也与杨兴洪等(2001)的研究结果一致.光合作用诱导期的长短差别很大,短的仅有几分钟, 长的可达1-2 h.光诱导的形成, 主要是来自光合碳同化酶的活化, 这些酶需要光照作催化剂(许大全, 2003 ).因此, 对如何控制光照时间和光合同化酶的活性等因素使绣球达到合适的光合诱导期有待继续探究. ...
1
... 绣球(Hydrangea macrophylla )又名八仙花、粉团花, 是虎耳草科绣球属植物, 高1-4 m, 常于基部发出多数放射枝而形成一圆形灌丛, 叶纸质或近革质, 倒卵形或阔椭圆形, 花期6-8月, 属暖温带半耐寒性落叶小灌木(黄淑美等, 1995 ).开花时花团锦簇, 颜色丰富艳丽.花色会随着土壤pH值的变化而变成红色或蓝色.由于绣球观赏效果较好且耐阴, 既可种植于庭院、公园、绿地, 又能作盆花栽培, 在园林绿化、美化中应用广泛(殷丽青等, 2010 ).目前对绣球的研究已经涉及栽培管理、花色花期调控、快繁技术、抗性等方面, 但针对绣球的耐阴性研究鲜有报道.本文以绣球品种‘无尽夏新娘’为试验材料, 采用模拟自然光照环境的方法, 从光合特性及叶绿素荧光参数来探究绣球对弱光的适应程度, 旨在为绣球的栽培和园林应用提供理论依据. ...
遮光对青榨槭光合速率及叶绿素荧光参数的影响
1
2007
... 叶绿素荧光是光合作用的探针, 通过荧光参数分析可以了解光合机构内部一系列重要的调节过程, F v /F m 是PSII光能转化率的指标(缴丽莉等, 2007 ).由表3 看出, 遮阴导致F v /F m 增加, 3种光照处理间差异显著, 全光照下F v /F m 低于50%遮光率处理, F o 高于50%遮光率处理, 根据张守仁(1999)的观点, F o 是判断PSII反应中心运转情况的重要指标, F o 上升表明PSII反应中心受到破坏或失活, F v /F m 下降是光抑制的重要特征, 因此推测全光照下的绣球叶片发生了光抑制.NPQ 的变化反映了非光化学耗散的能量, 其中包括了类囊体膜耗散热能, 是植物光合机构的自我保护机制(何炎红等, 2006 ).随着生长光照的减弱, NPQ 值在降低, 说明遮光处理降低了PSII天线色素吸收光能以热的形式耗散的比例, 提高了PSII吸收光能的利用效率. ...
Arabidopsis NPL1: A phototropin homolog controlling the chloroplast high-light avoidance response.
1
2001
... 光是植物光合作用和生长发育的重要影响因子, 光强和光质的变化直接影响着植物的生长发育, 植物对光强、光质的变化也会做出灵活的响应(许大全, 2013 ).植物在长期的进化过程中, 由于不同植物接收的太阳辐射量存在着明显差异, 因此形成了自身特有的需光特性(张聪颖等, 2011 ).植物经过遮阴后, 叶片形态、光合生理参数会发生变化, 表现为耐阴植物通常拥有较薄的颜色较深的叶片、较低的暗呼吸速率、较高的光合效率、较高的光系统II (PSII)效率以及较低的PSI/PSII (Wu et al ., 2013 ).C3 植物在弱光下均通过减少分枝、增加株高, 向高空发展以截获更多光能(Poorther & Perez-Soba, 2000 ).弱光条件下叶绿素含量增多是为了捕获更多的光能, 而植物的色素变化对光照的响应是不一样的, 绝对的阴生植物其色素含量和构成都是不随光照条件变化而变化的, 阳生植物通过增加叶绿素含量来适应弱光条件, 中间型植物则是通过既改变叶绿素含量也改变叶绿素构成来适应光照变化(Murchie & Horton, 1998 ).当光照强度超过植物的光饱和点时, 植物会发生光抑制或光破坏, 在中等光(稍高于2倍生长光强, 生长光强: 300-350 μmol·m-2 ·s-1 )下, 大豆(Glycine max )叶片的光抑制主要是由于PSII的可逆失活或下调, 与捕光天线LHCII的可逆脱离有关; 在强光(4倍或7倍于生长光强)下, 会造成大豆叶片光破坏, 主要是PSII的不可逆破坏, 表现为叶绿素膜蛋白(DI)发生净损失(Cai & Xu, 2002 ).在没有其他环境胁迫因素存在的自然条件下, 即使太阳光足够强, 人们也不大容易看到植物发生不可逆光破坏的情况, 之所以如此, 是因为植物在长期的进化过程中已经形成一系列防御光破坏的机制, 例如, 捕光天线脱离反应中心复合体和状态转换(Hong & Xu, 1999 ), 加强对过剩光能的热耗散, 加强光合作用对光能的利用, 叶片和叶绿体的避光运动(Kagawa et al ., 2001 ; Kasahara et al ., 2002 ), 加强光呼吸等耗能代谢, 加强对活性氧的清除和DI蛋白的周转等(Xu & Sheng, 1999 ).在正常情况下, 这些机制足以保护光合机构免于过量光能可能引起的不可逆破坏. ...
Chloroplast avoidance movement reduces photodamage in plant.
1
2002
... 光是植物光合作用和生长发育的重要影响因子, 光强和光质的变化直接影响着植物的生长发育, 植物对光强、光质的变化也会做出灵活的响应(许大全, 2013 ).植物在长期的进化过程中, 由于不同植物接收的太阳辐射量存在着明显差异, 因此形成了自身特有的需光特性(张聪颖等, 2011 ).植物经过遮阴后, 叶片形态、光合生理参数会发生变化, 表现为耐阴植物通常拥有较薄的颜色较深的叶片、较低的暗呼吸速率、较高的光合效率、较高的光系统II (PSII)效率以及较低的PSI/PSII (Wu et al ., 2013 ).C3 植物在弱光下均通过减少分枝、增加株高, 向高空发展以截获更多光能(Poorther & Perez-Soba, 2000 ).弱光条件下叶绿素含量增多是为了捕获更多的光能, 而植物的色素变化对光照的响应是不一样的, 绝对的阴生植物其色素含量和构成都是不随光照条件变化而变化的, 阳生植物通过增加叶绿素含量来适应弱光条件, 中间型植物则是通过既改变叶绿素含量也改变叶绿素构成来适应光照变化(Murchie & Horton, 1998 ).当光照强度超过植物的光饱和点时, 植物会发生光抑制或光破坏, 在中等光(稍高于2倍生长光强, 生长光强: 300-350 μmol·m-2 ·s-1 )下, 大豆(Glycine max )叶片的光抑制主要是由于PSII的可逆失活或下调, 与捕光天线LHCII的可逆脱离有关; 在强光(4倍或7倍于生长光强)下, 会造成大豆叶片光破坏, 主要是PSII的不可逆破坏, 表现为叶绿素膜蛋白(DI)发生净损失(Cai & Xu, 2002 ).在没有其他环境胁迫因素存在的自然条件下, 即使太阳光足够强, 人们也不大容易看到植物发生不可逆光破坏的情况, 之所以如此, 是因为植物在长期的进化过程中已经形成一系列防御光破坏的机制, 例如, 捕光天线脱离反应中心复合体和状态转换(Hong & Xu, 1999 ), 加强对过剩光能的热耗散, 加强光合作用对光能的利用, 叶片和叶绿体的避光运动(Kagawa et al ., 2001 ; Kasahara et al ., 2002 ), 加强光呼吸等耗能代谢, 加强对活性氧的清除和DI蛋白的周转等(Xu & Sheng, 1999 ).在正常情况下, 这些机制足以保护光合机构免于过量光能可能引起的不可逆破坏. ...
3种彩叶玉簪光合日变化
1
2011
... 光响应曲线可反映植物P n 随PAR 增减的变化规律(李金鹏和董然, 2011 ), 由图1 可知, 3种光照处理下的绣球光响应曲线变化规律相似, 当PAR 为0时, P n 为负值, 随着PAR 的增强, P n 逐渐增加, 由负值转为正值, 当PAR 为400 μmol·m-2 ·s-1 时, P n 上升趋势减缓并趋向平稳, 光响应曲线结果表明, 遮阴提高了绣球的AQY 和P nmax , 并降低了LCP 、LSP 和R d .由表1 可知, 50%遮光率条件下的绣球叶R d 比全光照下降低了31.4%, LCP 降低了57.4%, LSP 降低了9.5%, 这表明在遮阴环境下, 绣球可通过降低LCP 、LSP 来提高对弱光的利用能力, 并通过降低呼吸消耗以维持植株正常生长, 从而表现出了极强的适应能力. ...
环境强光诱导玉簪叶片光抑制的机制
1
2014
... 正常情况下, 没有遭受环境胁迫并经过充分暗适应的植物叶片F v /F m 是比较恒定的, 一般为0.80- 0.85 (许大全, 2013 ), 但是本实验3种光照处理下绣球的F v /F m 平均值仅为0.63, 显著低于正常值, P n 值也偏低, 分析原因可能是: 在试验中, 无法准确知道绣球光诱导期的时间, 使得弱光下生长的植株较长时间暴露在全光照下, 导致光合机构的过剩激发能大幅增加, 最大光化学效率迅速降低, 因而出现光抑制现象(李志真等, 2014 ).由F° v /F° m 、F v /F m 和F o 的降低, 可判断此时绣球的热耗散机制是依赖叶黄素循环的能量耗散(许大全, 2013 ).另一方面, 由表3 和表4 看出, 遮光率75%的绣球E 值、F o 最大, 说明较 ...
遮荫对濒危植物崖柏光合作用和叶绿素荧光参数的影响
1
2011
... 根据刘建锋等(2011) 的方法拟合光响应曲线, 模型表达式为: ...
Contrasting patterns of photo-synthetic acclimation to the light environment are dependent on the differential expression of the responses to altered irradiance and spectral quality.
1
1998
... 光是植物光合作用和生长发育的重要影响因子, 光强和光质的变化直接影响着植物的生长发育, 植物对光强、光质的变化也会做出灵活的响应(许大全, 2013 ).植物在长期的进化过程中, 由于不同植物接收的太阳辐射量存在着明显差异, 因此形成了自身特有的需光特性(张聪颖等, 2011 ).植物经过遮阴后, 叶片形态、光合生理参数会发生变化, 表现为耐阴植物通常拥有较薄的颜色较深的叶片、较低的暗呼吸速率、较高的光合效率、较高的光系统II (PSII)效率以及较低的PSI/PSII (Wu et al ., 2013 ).C3 植物在弱光下均通过减少分枝、增加株高, 向高空发展以截获更多光能(Poorther & Perez-Soba, 2000 ).弱光条件下叶绿素含量增多是为了捕获更多的光能, 而植物的色素变化对光照的响应是不一样的, 绝对的阴生植物其色素含量和构成都是不随光照条件变化而变化的, 阳生植物通过增加叶绿素含量来适应弱光条件, 中间型植物则是通过既改变叶绿素含量也改变叶绿素构成来适应光照变化(Murchie & Horton, 1998 ).当光照强度超过植物的光饱和点时, 植物会发生光抑制或光破坏, 在中等光(稍高于2倍生长光强, 生长光强: 300-350 μmol·m-2 ·s-1 )下, 大豆(Glycine max )叶片的光抑制主要是由于PSII的可逆失活或下调, 与捕光天线LHCII的可逆脱离有关; 在强光(4倍或7倍于生长光强)下, 会造成大豆叶片光破坏, 主要是PSII的不可逆破坏, 表现为叶绿素膜蛋白(DI)发生净损失(Cai & Xu, 2002 ).在没有其他环境胁迫因素存在的自然条件下, 即使太阳光足够强, 人们也不大容易看到植物发生不可逆光破坏的情况, 之所以如此, 是因为植物在长期的进化过程中已经形成一系列防御光破坏的机制, 例如, 捕光天线脱离反应中心复合体和状态转换(Hong & Xu, 1999 ), 加强对过剩光能的热耗散, 加强光合作用对光能的利用, 叶片和叶绿体的避光运动(Kagawa et al ., 2001 ; Kasahara et al ., 2002 ), 加强光呼吸等耗能代谢, 加强对活性氧的清除和DI蛋白的周转等(Xu & Sheng, 1999 ).在正常情况下, 这些机制足以保护光合机构免于过量光能可能引起的不可逆破坏. ...
Kinetic imaging lf chlorophyll fluorescence using modulated light.
1
2000
... 叶绿素荧光与光合作用中各个反应过程紧密相关, 逆境对光合作用各过程产生的影响都可通过植物体内叶绿素荧光诱导动力学变化反映出来(崔晓伟等, 2011 ), 同时叶绿素荧光参数更反映植物“内在性”的特点, 而且它具有快速、灵敏、非破坏性测量等优点, 因而被视为研究植物光合作用与环境关系的内在探针(Nedbal et al ., 2000 ), 把叶片的荧光分析与气体交换测定技术结合起来, 不仅可以从生理学水平上更迅速、全面地了解光合机构的实际运转状况, 而且可以间接地从生物化学水平上了解电子传递速率及其与不同代谢过程的关系. ...
The growth response of plants to elevated CO2 under non-optimal environmental conditions.
1
2000
... 光是植物光合作用和生长发育的重要影响因子, 光强和光质的变化直接影响着植物的生长发育, 植物对光强、光质的变化也会做出灵活的响应(许大全, 2013 ).植物在长期的进化过程中, 由于不同植物接收的太阳辐射量存在着明显差异, 因此形成了自身特有的需光特性(张聪颖等, 2011 ).植物经过遮阴后, 叶片形态、光合生理参数会发生变化, 表现为耐阴植物通常拥有较薄的颜色较深的叶片、较低的暗呼吸速率、较高的光合效率、较高的光系统II (PSII)效率以及较低的PSI/PSII (Wu et al ., 2013 ).C3 植物在弱光下均通过减少分枝、增加株高, 向高空发展以截获更多光能(Poorther & Perez-Soba, 2000 ).弱光条件下叶绿素含量增多是为了捕获更多的光能, 而植物的色素变化对光照的响应是不一样的, 绝对的阴生植物其色素含量和构成都是不随光照条件变化而变化的, 阳生植物通过增加叶绿素含量来适应弱光条件, 中间型植物则是通过既改变叶绿素含量也改变叶绿素构成来适应光照变化(Murchie & Horton, 1998 ).当光照强度超过植物的光饱和点时, 植物会发生光抑制或光破坏, 在中等光(稍高于2倍生长光强, 生长光强: 300-350 μmol·m-2 ·s-1 )下, 大豆(Glycine max )叶片的光抑制主要是由于PSII的可逆失活或下调, 与捕光天线LHCII的可逆脱离有关; 在强光(4倍或7倍于生长光强)下, 会造成大豆叶片光破坏, 主要是PSII的不可逆破坏, 表现为叶绿素膜蛋白(DI)发生净损失(Cai & Xu, 2002 ).在没有其他环境胁迫因素存在的自然条件下, 即使太阳光足够强, 人们也不大容易看到植物发生不可逆光破坏的情况, 之所以如此, 是因为植物在长期的进化过程中已经形成一系列防御光破坏的机制, 例如, 捕光天线脱离反应中心复合体和状态转换(Hong & Xu, 1999 ), 加强对过剩光能的热耗散, 加强光合作用对光能的利用, 叶片和叶绿体的避光运动(Kagawa et al ., 2001 ; Kasahara et al ., 2002 ), 加强光呼吸等耗能代谢, 加强对活性氧的清除和DI蛋白的周转等(Xu & Sheng, 1999 ).在正常情况下, 这些机制足以保护光合机构免于过量光能可能引起的不可逆破坏. ...
Spectral reflectance and photosynthetic properties of Betula papyrifera (Betulaceae) leaves along an elevational gradient on Mt.
1
2002
... LSP 和LCP 分别代表植物光合作用的光照强度上限和下限, 同时也代表植物叶片对强光和弱光的利用能力.耐阴植物的LCP 小于20 μmol·m-2 ·s-1 , LSP 为50-1β000μmol·m-2 ·s-1 或更低(Boardman, 1977 ).绣球的LCP 为6.71-15.75 μmol·m-2 ·s-1 , LSP 为278.14-307.42 μmol·m-2 ·s-1 .因此, 绣球属于耐阴植物, 遮阴处理下的绣球R d 低于全光照下, 这说明遮阴叶比全光照叶呼吸强度小, 绣球在遮阴环境下能够通过降低LCP 和LSP 来增强对弱光的利用能力, 与王凯等(2009)的研究结果类似.AQY 反映了叶片在弱光环境下的光合能力, 其值越大, 表明植物吸收与转换光能的色素蛋白复合体越多, 植物利用弱光的能力越强(Richardson & Berlyn, 2002 ).绣球在弱光环境下的AQY 较全光照环境下高, 说明绣球对弱光有较强的适应性. ...
遮阴对黄波罗幼苗的光合特性及光能利用效率的影响
2009
Chlorophyll fluorescence and leaf-air temperature difference as potential shade-tolerance indexes of ornamental plants.
1
2013
... 光是植物光合作用和生长发育的重要影响因子, 光强和光质的变化直接影响着植物的生长发育, 植物对光强、光质的变化也会做出灵活的响应(许大全, 2013 ).植物在长期的进化过程中, 由于不同植物接收的太阳辐射量存在着明显差异, 因此形成了自身特有的需光特性(张聪颖等, 2011 ).植物经过遮阴后, 叶片形态、光合生理参数会发生变化, 表现为耐阴植物通常拥有较薄的颜色较深的叶片、较低的暗呼吸速率、较高的光合效率、较高的光系统II (PSII)效率以及较低的PSI/PSII (Wu et al ., 2013 ).C3 植物在弱光下均通过减少分枝、增加株高, 向高空发展以截获更多光能(Poorther & Perez-Soba, 2000 ).弱光条件下叶绿素含量增多是为了捕获更多的光能, 而植物的色素变化对光照的响应是不一样的, 绝对的阴生植物其色素含量和构成都是不随光照条件变化而变化的, 阳生植物通过增加叶绿素含量来适应弱光条件, 中间型植物则是通过既改变叶绿素含量也改变叶绿素构成来适应光照变化(Murchie & Horton, 1998 ).当光照强度超过植物的光饱和点时, 植物会发生光抑制或光破坏, 在中等光(稍高于2倍生长光强, 生长光强: 300-350 μmol·m-2 ·s-1 )下, 大豆(Glycine max )叶片的光抑制主要是由于PSII的可逆失活或下调, 与捕光天线LHCII的可逆脱离有关; 在强光(4倍或7倍于生长光强)下, 会造成大豆叶片光破坏, 主要是PSII的不可逆破坏, 表现为叶绿素膜蛋白(DI)发生净损失(Cai & Xu, 2002 ).在没有其他环境胁迫因素存在的自然条件下, 即使太阳光足够强, 人们也不大容易看到植物发生不可逆光破坏的情况, 之所以如此, 是因为植物在长期的进化过程中已经形成一系列防御光破坏的机制, 例如, 捕光天线脱离反应中心复合体和状态转换(Hong & Xu, 1999 ), 加强对过剩光能的热耗散, 加强光合作用对光能的利用, 叶片和叶绿体的避光运动(Kagawa et al ., 2001 ; Kasahara et al ., 2002 ), 加强光呼吸等耗能代谢, 加强对活性氧的清除和DI蛋白的周转等(Xu & Sheng, 1999 ).在正常情况下, 这些机制足以保护光合机构免于过量光能可能引起的不可逆破坏. ...
植物光胁迫研究中的几个问题
1
2003
... 其他两种光照处理, 受到的光抑制更加严重.一个原因是E 值的增加会使单线态叶绿素(1 Chl)不能及时返回三线态叶绿素(3 Chl), 1 Chl的积累会将能量传递给O2 , 形成活性极强的单线态氧(1 O2 ), 这会引起光合机构的失活甚至破坏(Asada, 1999 ).F o 的增加说明植物叶片类囊体膜受到损害, PSII反应中心失活(黄秋娴等, 2015 ).另一个原因是其阴生叶片的结构和生理特征均缺少对强光的适应性, 所以遮阴下生长的绣球叶片过剩激发能多, 光抑制程度也比直接生长在全日照下的绣球要严重.这符合3种光照处理下非光化学反应耗散变化趋势, 也与杨兴洪等(2001)的研究结果一致.光合作用诱导期的长短差别很大,短的仅有几分钟, 长的可达1-2 h.光诱导的形成, 主要是来自光合碳同化酶的活化, 这些酶需要光照作催化剂(许大全, 2003 ).因此, 对如何控制光照时间和光合同化酶的活性等因素使绣球达到合适的光合诱导期有待继续探究. ...
3
... 光是植物光合作用和生长发育的重要影响因子, 光强和光质的变化直接影响着植物的生长发育, 植物对光强、光质的变化也会做出灵活的响应(许大全, 2013 ).植物在长期的进化过程中, 由于不同植物接收的太阳辐射量存在着明显差异, 因此形成了自身特有的需光特性(张聪颖等, 2011 ).植物经过遮阴后, 叶片形态、光合生理参数会发生变化, 表现为耐阴植物通常拥有较薄的颜色较深的叶片、较低的暗呼吸速率、较高的光合效率、较高的光系统II (PSII)效率以及较低的PSI/PSII (Wu et al ., 2013 ).C3 植物在弱光下均通过减少分枝、增加株高, 向高空发展以截获更多光能(Poorther & Perez-Soba, 2000 ).弱光条件下叶绿素含量增多是为了捕获更多的光能, 而植物的色素变化对光照的响应是不一样的, 绝对的阴生植物其色素含量和构成都是不随光照条件变化而变化的, 阳生植物通过增加叶绿素含量来适应弱光条件, 中间型植物则是通过既改变叶绿素含量也改变叶绿素构成来适应光照变化(Murchie & Horton, 1998 ).当光照强度超过植物的光饱和点时, 植物会发生光抑制或光破坏, 在中等光(稍高于2倍生长光强, 生长光强: 300-350 μmol·m-2 ·s-1 )下, 大豆(Glycine max )叶片的光抑制主要是由于PSII的可逆失活或下调, 与捕光天线LHCII的可逆脱离有关; 在强光(4倍或7倍于生长光强)下, 会造成大豆叶片光破坏, 主要是PSII的不可逆破坏, 表现为叶绿素膜蛋白(DI)发生净损失(Cai & Xu, 2002 ).在没有其他环境胁迫因素存在的自然条件下, 即使太阳光足够强, 人们也不大容易看到植物发生不可逆光破坏的情况, 之所以如此, 是因为植物在长期的进化过程中已经形成一系列防御光破坏的机制, 例如, 捕光天线脱离反应中心复合体和状态转换(Hong & Xu, 1999 ), 加强对过剩光能的热耗散, 加强光合作用对光能的利用, 叶片和叶绿体的避光运动(Kagawa et al ., 2001 ; Kasahara et al ., 2002 ), 加强光呼吸等耗能代谢, 加强对活性氧的清除和DI蛋白的周转等(Xu & Sheng, 1999 ).在正常情况下, 这些机制足以保护光合机构免于过量光能可能引起的不可逆破坏. ...
... 正常情况下, 没有遭受环境胁迫并经过充分暗适应的植物叶片F v /F m 是比较恒定的, 一般为0.80- 0.85 (许大全, 2013 ), 但是本实验3种光照处理下绣球的F v /F m 平均值仅为0.63, 显著低于正常值, P n 值也偏低, 分析原因可能是: 在试验中, 无法准确知道绣球光诱导期的时间, 使得弱光下生长的植株较长时间暴露在全光照下, 导致光合机构的过剩激发能大幅增加, 最大光化学效率迅速降低, 因而出现光抑制现象(李志真等, 2014 ).由F° v /F° m 、F v /F m 和F o 的降低, 可判断此时绣球的热耗散机制是依赖叶黄素循环的能量耗散(许大全, 2013 ).另一方面, 由表3 和表4 看出, 遮光率75%的绣球E 值、F o 最大, 说明较 ...
... 的降低, 可判断此时绣球的热耗散机制是依赖叶黄素循环的能量耗散(许大全, 2013 ).另一方面, 由表3 和表4 看出, 遮光率75%的绣球E 值、F o 最大, 说明较 ...
1
... 光是植物光合作用和生长发育的重要影响因子, 光强和光质的变化直接影响着植物的生长发育, 植物对光强、光质的变化也会做出灵活的响应(许大全, 2013 ).植物在长期的进化过程中, 由于不同植物接收的太阳辐射量存在着明显差异, 因此形成了自身特有的需光特性(张聪颖等, 2011 ).植物经过遮阴后, 叶片形态、光合生理参数会发生变化, 表现为耐阴植物通常拥有较薄的颜色较深的叶片、较低的暗呼吸速率、较高的光合效率、较高的光系统II (PSII)效率以及较低的PSI/PSII (Wu et al ., 2013 ).C3 植物在弱光下均通过减少分枝、增加株高, 向高空发展以截获更多光能(Poorther & Perez-Soba, 2000 ).弱光条件下叶绿素含量增多是为了捕获更多的光能, 而植物的色素变化对光照的响应是不一样的, 绝对的阴生植物其色素含量和构成都是不随光照条件变化而变化的, 阳生植物通过增加叶绿素含量来适应弱光条件, 中间型植物则是通过既改变叶绿素含量也改变叶绿素构成来适应光照变化(Murchie & Horton, 1998 ).当光照强度超过植物的光饱和点时, 植物会发生光抑制或光破坏, 在中等光(稍高于2倍生长光强, 生长光强: 300-350 μmol·m-2 ·s-1 )下, 大豆(Glycine max )叶片的光抑制主要是由于PSII的可逆失活或下调, 与捕光天线LHCII的可逆脱离有关; 在强光(4倍或7倍于生长光强)下, 会造成大豆叶片光破坏, 主要是PSII的不可逆破坏, 表现为叶绿素膜蛋白(DI)发生净损失(Cai & Xu, 2002 ).在没有其他环境胁迫因素存在的自然条件下, 即使太阳光足够强, 人们也不大容易看到植物发生不可逆光破坏的情况, 之所以如此, 是因为植物在长期的进化过程中已经形成一系列防御光破坏的机制, 例如, 捕光天线脱离反应中心复合体和状态转换(Hong & Xu, 1999 ), 加强对过剩光能的热耗散, 加强光合作用对光能的利用, 叶片和叶绿体的避光运动(Kagawa et al ., 2001 ; Kasahara et al ., 2002 ), 加强光呼吸等耗能代谢, 加强对活性氧的清除和DI蛋白的周转等(Xu & Sheng, 1999 ).在正常情况下, 这些机制足以保护光合机构免于过量光能可能引起的不可逆破坏. ...
遮荫棉花转入强光后光合作用的光抑制及其恢复
2001
优良八仙花品种(Hydrangea serrata
1
2010
... 绣球(Hydrangea macrophylla )又名八仙花、粉团花, 是虎耳草科绣球属植物, 高1-4 m, 常于基部发出多数放射枝而形成一圆形灌丛, 叶纸质或近革质, 倒卵形或阔椭圆形, 花期6-8月, 属暖温带半耐寒性落叶小灌木(黄淑美等, 1995 ).开花时花团锦簇, 颜色丰富艳丽.花色会随着土壤pH值的变化而变成红色或蓝色.由于绣球观赏效果较好且耐阴, 既可种植于庭院、公园、绿地, 又能作盆花栽培, 在园林绿化、美化中应用广泛(殷丽青等, 2010 ).目前对绣球的研究已经涉及栽培管理、花色花期调控、快繁技术、抗性等方面, 但针对绣球的耐阴性研究鲜有报道.本文以绣球品种‘无尽夏新娘’为试验材料, 采用模拟自然光照环境的方法, 从光合特性及叶绿素荧光参数来探究绣球对弱光的适应程度, 旨在为绣球的栽培和园林应用提供理论依据. ...
遮荫处理对红叶石楠和洒金桃叶珊瑚光合特性的影响
1
2011
... 光是植物光合作用和生长发育的重要影响因子, 光强和光质的变化直接影响着植物的生长发育, 植物对光强、光质的变化也会做出灵活的响应(许大全, 2013 ).植物在长期的进化过程中, 由于不同植物接收的太阳辐射量存在着明显差异, 因此形成了自身特有的需光特性(张聪颖等, 2011 ).植物经过遮阴后, 叶片形态、光合生理参数会发生变化, 表现为耐阴植物通常拥有较薄的颜色较深的叶片、较低的暗呼吸速率、较高的光合效率、较高的光系统II (PSII)效率以及较低的PSI/PSII (Wu et al ., 2013 ).C3 植物在弱光下均通过减少分枝、增加株高, 向高空发展以截获更多光能(Poorther & Perez-Soba, 2000 ).弱光条件下叶绿素含量增多是为了捕获更多的光能, 而植物的色素变化对光照的响应是不一样的, 绝对的阴生植物其色素含量和构成都是不随光照条件变化而变化的, 阳生植物通过增加叶绿素含量来适应弱光条件, 中间型植物则是通过既改变叶绿素含量也改变叶绿素构成来适应光照变化(Murchie & Horton, 1998 ).当光照强度超过植物的光饱和点时, 植物会发生光抑制或光破坏, 在中等光(稍高于2倍生长光强, 生长光强: 300-350 μmol·m-2 ·s-1 )下, 大豆(Glycine max )叶片的光抑制主要是由于PSII的可逆失活或下调, 与捕光天线LHCII的可逆脱离有关; 在强光(4倍或7倍于生长光强)下, 会造成大豆叶片光破坏, 主要是PSII的不可逆破坏, 表现为叶绿素膜蛋白(DI)发生净损失(Cai & Xu, 2002 ).在没有其他环境胁迫因素存在的自然条件下, 即使太阳光足够强, 人们也不大容易看到植物发生不可逆光破坏的情况, 之所以如此, 是因为植物在长期的进化过程中已经形成一系列防御光破坏的机制, 例如, 捕光天线脱离反应中心复合体和状态转换(Hong & Xu, 1999 ), 加强对过剩光能的热耗散, 加强光合作用对光能的利用, 叶片和叶绿体的避光运动(Kagawa et al ., 2001 ; Kasahara et al ., 2002 ), 加强光呼吸等耗能代谢, 加强对活性氧的清除和DI蛋白的周转等(Xu & Sheng, 1999 ).在正常情况下, 这些机制足以保护光合机构免于过量光能可能引起的不可逆破坏. ...
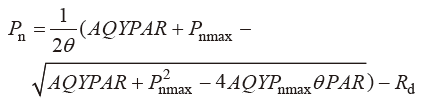


{kind=link}
{kind=link}