土壤养分在空间分布上多呈时空异质性或者斑块分布(Hutching & John, 2004 )。产生土壤养分空间异质性的原因很多, 其中包括有机物(凋落物、根系死亡以及动物尸体)的分解、无机养分在土壤中的扩散系数差异以及微地形因素影响等(Hodge, 2004 ; 徐星凯等, 2012 ; 刘双娥等, 2015 )。植物为了获得更多的养分, 其根系会增加在土壤养分富集区域的分布比例, 进而提高根系对养分的获取和利用能力(Hodge, 2004 ; Mommer et al ., 2012 )。近年来, 随着合理高效的供肥管理措施(局部施肥)在农业中广泛应用及其高效的生产力, 使其在林业上尤其是人工林经营管理中也开始得到广泛应用。因此, 了解肥料集约经营管理下的林木根系生长、形态分布以及养分吸收状况, 将有利于解释林木在生长上存在的差异, 同时为提高速生丰产林生产力提供更为完善和高效的技术指导。
氮和磷是植物生长最重要的两种营养元素, 其有效性和空间分布对植物的生长和根系发育具有重要影响(Hodge, 2004 ; Vance et al ., 2003 )。研究揭示, 局部供氮或供磷可显著促进大麦(Hordeum vulgare )、玉米(Zea mays )和小麦(Triticum aestivum )等植物侧根数量的增加以及总根长的提高(Drew, 1975 ; Officer et al ., 2009 )。Zhang和Forde (2000)将拟南芥(Arabidopsis thaliana )分别放在局部高氮和均匀高氮的琼脂培养基上培养时发现, 根系会在富氮斑块中增生, 而均匀供氮处理中的根系总量降低。可见根系在富养斑块中的增生可促进植物对养分供应的有效利用(Linkohr et al ., 2002 )。更重要的是, 相比于单独局部供氮或供磷, 在斑块中同时添加氮和磷更能促进根系在富养斑块中增生, 提高营养元素的吸收效率, 进而加快整株的生长(Jing et al ., 2010 )。这表明植物根系在局部供养斑块中的增生受到营养元素组成以及元素之间互作效应的共同影响。在木本植物研究中也发现类似的变化规律, 但物种间差异较大(Hodge, 2004 )。Einsmann等(1999)发现, 10种木本植物中有4种植物的生物量在养分异质性的供应条件下有显著提高, 但在对同种植物的不同研究中, Bliss等(2002)并没有发现这一趋势。虽然有研究认为养分斑块的存在能够对植物地上部分的生长产生积极的促进效应(Jackson & Caldwell, 1996 ), 但多年生植物绒毛草(Holcus lanatus )仅在短期内出现大量根系增生和促进地上部分生长的现象, 长期来看这种优势很快会消失, 甘松茅(Nardus stricta )则对养分斑块反应不敏感(Fransen & de Kroon, 2001 )。显然, 养分斑块或者局部养分供应对植物生长的影响受很多因素的影响, 除了养分的属性外, 植物对养分的敏感性及试验中植物生长时间的长短对结果都会产生较大影响(李洪波等, 2013 )。
马尾松(Pinus massoniana )是重要的材用、脂用和生态造林树种, 广泛分布于我国的亚热带地区(秦国峰和周志春, 2012 )。当前研究已揭示了氮添加对低磷胁迫下马尾松生长影响的生物学机制, 包括地上部分生长、根系参数和氮、磷效率的适应性变化等(Zhang et al ., 2013 ; 庞丽等, 2014 )。 然而, 相关研究未考虑氮素本身在空间分布上的不均一性。鉴于此, 本文拟通过人工控制氮素在土壤空间上的异质性, 研究低磷胁迫下马尾松不同家系在局部供氮条件下的生长表现和根系发育, 进一步阐明马尾松不同家系对土壤异质养分的响应差异及机制, 为马尾松营养高效新品种选育和苗木高效栽培方式提供理论依据以及实践指导。
1 材料和方法
1.1 供试材料
试验在浙江省淳安县富溪林场温室大棚内进行(119.13° E, 29.48° N), 其所处海拔为130 m, 年平均气温17.5 ℃, 年降水量为1β701.2 mm。以马尾松二代育种群体内控制授粉产生的5个全同胞子代1、25、49、52和57作为试验材料。盆栽容器采用定制的圆柱形无纺布植树袋, 直径为12 cm, 高28 cm。土壤基质为浙江省淳安县千岛湖的贫瘠酸性红壤, 其理化性质见表1 。
1.2 试验设计
1.2.1 试验装置
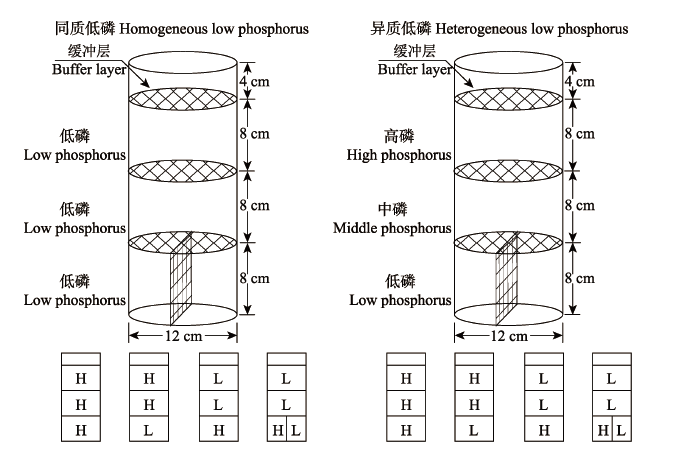
本试验中的马尾松分层隔网盆栽设计, 参考杨青等(2011) 的磷控释土培系统, 同时结合一年生马尾松苗木的根系形态特征进行分层处理(图1 )。盆栽容器自上而下依次分为表层、中间层和深层3个层次, 间距8 cm, 每层之间用孔径为3 mm的塑料纤维网隔离(主根、基根均能顺利穿过)。在试验装置上端填充4 cm厚的干净河沙作为缓冲层, 将经过0.2%- 0.4%的高锰酸钾溶液浸泡20 min的马尾松不同家系种子植入缓冲层中间, 每盆播种在3-5粒种子。
图1 不同磷环境下的局部供氮处理。HHH, 均匀供氮; HHL, 表层和中间层供氮; LLH, 底层供氮; LLH/L, 底层半侧供氮。
Fig. 1 The patterns of N added under two P deficiency conditions. HHH, homogeneously high N along the soil profile; HHL, high N-high N-low N; LLH, low N-low N-high N; LLH/L, low N-low N-on side with N addition and the other side without N supply.
1.2.2 磷素环境
低磷土壤环境设置参照Zhang等(2013)的浓度设计。基质土壤经过风干过筛后, 加入一定量的Ca(H2 PO4 )2 混匀。异质低磷盆栽基质的底层、中间层和表层依次填入添加0、0.10和1.00 g·kg-1 的Ca(H2 PO4 )2 , 相当于有效磷含量分别为0.99、15和 30 mg·kg-1 。盆栽基质由上而下有效磷含量逐渐降低的异质养分环境可模拟立地条件较好, 表层营养丰富的针阔混交林或阔叶林土壤环境(Hodge, 2004 ; Verma et al ., 2005 )。同质低磷盆栽基质的表层、中间层和底层皆均匀添加有效磷含量极低(0.99 mg·kg-1 )的贫瘠酸性红壤作为盆栽基质, 该处理可有效地模拟森林土壤中磷素极为匮乏的针叶林立地环境(Richaedson et al ., 2004 )。
1.2.3 局部供氮
土壤垂直方向上的氮素异质性设置及保持, 采用树脂膜控释氮肥(含氮量44%, 山东金正大公司生产)来调控。两种低磷环境下同时布设相同局部供氮处理, 氮素施入量参照庞丽等(2014) 研究方法。处理1 (HHH): 表层、中间层和底层均匀供氮, 其有效氮含量均为100 g·kg-1 ; 处理2 (HHL): 表层和中间层(0-16 cm)供氮, 有效氮含量均为100 g·kg-1 , 底层不添加氮; 处理3 (LLH): 底层(16-24 cm)供氮, 有效氮含量为100 g·kg-1 , 其他层次均不添加; 处理 4 (LLH/L): 底层一侧局部供氮的有效氮含量为 100 g·kg-1 。
1.2.4 栽培管理
盆栽试验在温室大棚内进行培养, 配置自动喷灌系统, 以保证马尾松不同家系苗木正常生长的水分需求。试验于2015年3月开始播种, 1个月后根据出苗及生长情况进行适当补植和剔除, 每盆保留一株正常生长的幼苗。每个处理每个家系设12次重复, 单株小区, 共计480株马尾松不同家系苗。经过一个生长季, 于12月收获。
1.3 收获和分析测定
1.3.1 生长指标测定
根据苗木生长情况, 剔除生长异常植株, 每处理选定10株测量株高(SH )和地径(SBD )。将根、茎和叶三部分别放在105 ℃的烘箱中杀青30 min, 然后79 ℃下至恒质量, 得到各部分的干物质量, 然后计算整株干物质量(SDW )、根冠比(R /S )。
1.3.2 根系形态参数测定
将根系与土壤分开, 用去离子水清洗干净, 擦干后用根系扫描仪(Expression 11000XL, EPSON, Nagano-ken, Japan)对根系进行扫描成像, 利用图像分析软件WinRHIZO Pro STD1600+ (Regent Instruments, Otlawa, Canada)进行分析, 以获得根长(RL )和根表面积(RSA )等数据。
1.3.3 氮、磷含量测定
分别称取0.1 g左右的根、茎和叶, 用H2 SO4 - H2 O2 消煮后, 取消煮液用钼锑抗比色法测定根系中的磷含量(PC ), 用FOSS定氮仪(Foss Sossanalytizal a-s., Ahlleroed, Denmark)测定氮含量(NC )。
1.4 数据分析
马尾松苗木的株高、地径、根干物质量、整株干物质量、根冠比、根长、根表面积以及氮、磷养分含量使用SAS的ANOVA程序进行两因素方差分析(N处理×家系)。在同一磷素环境、单个家系内, 用SAS的ANOVA程序进行不同局部供氮处理的单因素方差分析, 再用最小显著差异法(LSD )进行多重比较。最后结果用Origin 9.0绘图。
2 结果
2.1 局部供氮对不同磷环境下马尾松不同家系苗木生长的影响
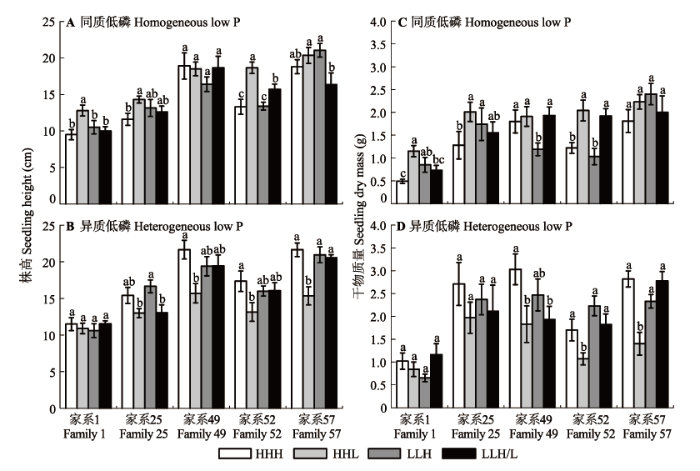
方差分析结果表明, 在两种低磷环境下, 局部供氮对马尾松苗木的株高、地径和干物质积累量均有显著影响(p < 0.001)(表2 )。与同质低磷相比, 异质低磷下的株高、地径和干物质积累量分别增加了5.1%、11.6%和22.0%, 其中干物质积累量之间的差异达到显著水平(p < 0.05), 但在HHL处理下的生长表现例外(表2 )。与均匀供氮(HHH)相比, HHL处理显著增加了同质低磷下马尾松苗木的株高、地径和干物质积累量, 相反, HHL处理却不利于异质低磷下马尾松苗木的生长, 其干物质积累量分别在同质低磷下增加了41.7%, 异质低磷下降了36.9% (表2 )。LLH和LLH/L处理后, 马尾松苗木的株高、地径和干物质积累量与HHH处理后的生长表现无显著差异, 说明在底层或底层半侧较少氮素投入的情况下, 马尾松苗木可获得与均匀供氮同样的生长表现。另外, 同质低磷下的根冠比小于异质低磷水平, 且在局部供氮处理后进一步降低, 表明局部供氮对同质低磷下马尾松苗木地上部分的生长更为有利(表2 )。
不同家系对局部供氮的响应差异显著(p < 0.05) (表2 )。如图2 所示, 在两种低磷环境下, 家系1在各氮处理后的株高和干物质积累量均显著低于其他供试家系。家系25在同质低磷、HHL处理后, 其株高和干物质积累量较HHH处理显著高出23.3%和57.0%; 但在异质低磷下, LLH和LLH/L处理更有利于该家系的生长。家系49和52的干物质积累量在同质低磷、LLH处理下较HHH处理分别降低了33.9%和15.6%, 但在异质低磷、LLH处理下与HHH处理之间无显著差异。家系57的株高和干物质积累量的变化与家系49以及52的变化规律类似, 且较其他家系具有更强的生长势, 尤其在同质低磷环境下更为突出。
图2 局部供氮对低磷胁迫下马尾松不同家系苗木生长的影响(平均值±标准误差)。HHH, 均匀供氮; HHL, 表层和中间层供氮; LLH, 底层供氮; LLH/L, 底层半侧供氮。不同小写字母表示不同处理之间差异显著(p < 0.05)。
Fig. 2 The seedling growth traits of Pinus massoniana families in different local N supply treatments (mean ± SE ). HHH, homogeneously high N along the soil profile; HHL, high N-high N-low N; LLH, low N-low N-high N; LLH/L, low N-low N-on side with N addition and the other side without N supply. Different lower letters indicate significant differences among treatments (p < 0.05).
2.2 局部供氮对不同磷环境下马尾松不同家系苗木根系参数的影响
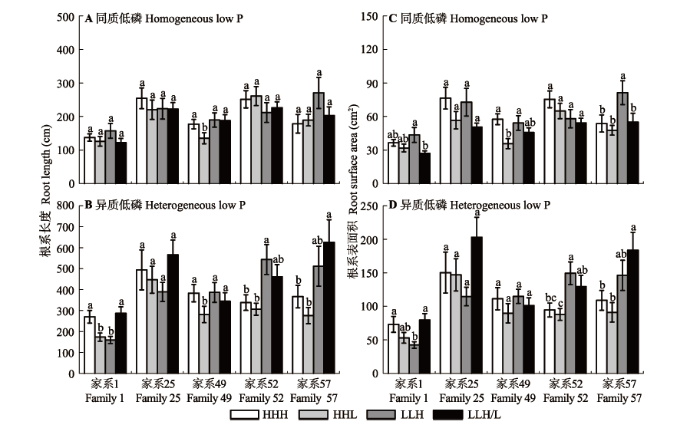
马尾松苗木的根系参数对局部供氮的响应在两种低磷环境下类似, 底层供氮皆有利于苗木根系的生长。与HHH相比, LLH和LLH/L处理促进了马尾松苗木根长和根表面积的增加, 尤其在异质低磷、LLH/L处理后, 其根长和根表面积显著增加了29.2%和32.3%; 相反, HHL处理降低了马尾松苗木的根长和根表面积(表2 )。异质低磷下的根长和根表面积约是同质低磷下的1.95倍和2.11倍, 说明异质低磷较同质低磷显著促进了根系的生长发育(p < 0.01) (表2 )。
不同家系的根系参数对局部供氮的响应程度有所差异。由图3 可知, 在两种低磷环境下, 家系25的根长和根表面积在不同供氮处理后均无明显变化, 但在同质低磷、HHL处理后, 该家系的根长和根表面积分别降低了13.4%和25.8% (p = 0.07)。家系49、52和57在同质低磷下, 经LLH和LLH/L处理后, 其根长和根表面积较HHH处理有所下降, 但未达到显著水平; 相反, 在异质低磷下, LLH和LLH/L处理后的根长和根表面积明显高于HHH处理水平, HHL处理则不利于根系的生长发育。家系1的根长和根表面积的变化与生长性状的变化规律相似, 均显著小于其他家系。
图3 局部供氮对低磷胁迫下马尾松不同家系苗木根系参数的影响(平均值±标准误差)。HHH, 均匀供氮; HHL, 表层和中间层供氮; LLH, 底层供氮; LLH/L, 底层半侧供氮。不同小写字母表示不同处理之间差异显著(p < 0.05)。
Fig. 3 The root parameters of Pinus massoniana families in different local N supply treatments (mean ± SE ). HHH, homogeneously high N along the soil profile; HHL, high N-high N-low N; LLH, low N-low N-high N; LLH/L, low N-low N-on side with N addition and the other side without N supply. Different lower letters indicate significant differences among treatments (p < 0.05).
2.3 局部供氮对不同磷环境下马尾松不同家系苗木氮、磷含量的影响
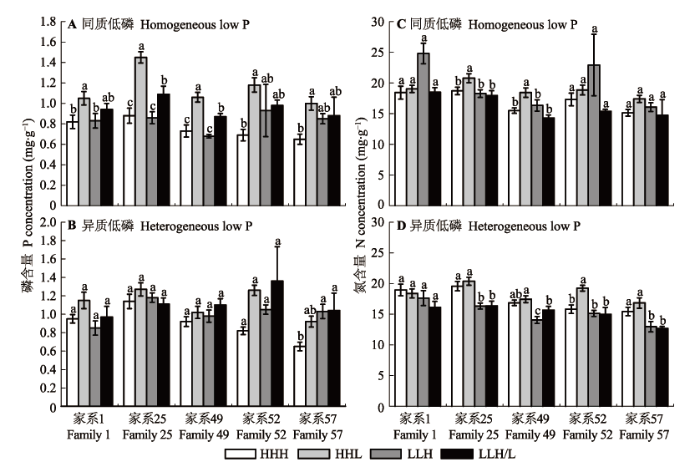
马尾松苗木植株的全氮含量受局部供氮处理影响显著, 全磷含量无显著差异, 但在异质低磷下的全磷含量显著高于同质低磷水平(表2 )。如图4 显示, 在异质低磷下, 马尾松苗木的全磷含量平均高于同质低磷水平13.5%, 全氮含量却平均下降了10.3%。与HHH处理相比, 在两种低磷环境下, 局部供氮(HHL、LLH和LLH/L)处理后, 马尾松苗木植株的全磷含量均有不同程度的增加, 全氮含量则因不同的供氮方式差异较大, 其中, HHL处理显著提高了马尾松苗木植株的全氮含量 (同质低磷下较HHH增加了27.4%; 异质低磷下增加了6.5%), LLH和LLH/L处理则不利于苗木植株全氮含量的增加, 甚至显著降低了异质低磷下苗木的全氮含量。
图4 局部供氮对两种磷环境下马尾松不同家系氮、磷含量的影响(平均值±标准误差)。HHH, 均匀供氮; HHL, 表层和中间层供氮; LLH, 底层供氮; LLH/L, 底层半侧供氮。不同小写字母表示不同处理之间差异显著(p < 0.05)。不同小写字母表示不同处理之间差异显著(p < 0.05)。
Fig. 4 N concentration and P concentration of P. massoniana families in different local N supply treatments (mean ± SE ). HHH, homogeneously high N along the soil profile; HHL, high N-high N-low N; LLH, low N-low N-high N; LLH/L, low N-low N-on side with N addition and the other side without N supply. Different lower letters indicate significant differences among treatments (p < 0.05).
局部供氮方式对不同家系全氮和全磷含量的影响差异也较大。由图4 可知, 家系25的全氮和全磷含量在不同供氮处理之间的变化规律与株高、干物质积累量的变化相似, 其在同质低磷、HHL处理下的全氮和全磷含量分别较HHH处理显著高出11.1%和64.8% (图4 A、4C)。家系49、52和57在异质低磷、LLH和LLH/L处理后, 其全氮含量较HHH处理有所增加, 但降低了植株的全磷含量。家系1的全氮和全磷含量在不同供氮处理之间差异不显著(图4 B、4D)。
3 讨论和结论
近年来, 土壤中养分元素的分布方式对植株生长的影响越来越受到关注(Hodge, 2004 ; 吕超群等, 2007 ; 徐星凯等, 2012 )。本研究中的异质低磷环境较同质低磷更有利于马尾松苗木的生长, 与前人的研究结果(杨青等, 2011 ; 庞丽等, 2014 )一致。已有研究表明, 外源氮素添加(100 g·kg-1 )会使土壤有效氮含量增加, 改变植株生物量的分配进而促进整株的生长(段洪浪等, 2009 ; 庞丽等, 2014 ; 刘双娥等, 2015 )。在本实验中, 局部供氮处理的斑块氮浓度与均匀供氮处理的氮含量相同, 但总施肥量要少于均匀供氮(相当于HHH处理施氮量的1/6-2/3)。研究结果显示, 不同的供氮方式对马尾松苗木生长的影响差异较大, 且受到土壤磷素环境影响。在同质低磷下, HHL显著促进了马尾松苗木的生长, 根冠比降低, 较其他供氮方式更有利于苗木地上部分的生长; 其次, LLH和LLH/L两种方式之间, LLH/L对马尾松整株的生长相对更有利, 这表明在同质低磷下, 局部供氮增强了土壤中养分的异质性, 促进了马尾松苗木整株的生长。类似的结果在桉树(Eucalyptus grandis )、油松(Pinus tabulaeformis )、火炬松(Pinus taeda )等植物中也被发现(Mou et al ., 1997 ; Zhou & Shangguan, 2007 ; Graciano et al ., 2009 )。然而, 值得注意的是, Fransen和de Kroon (2001)在研究多年生植物绒毛草和甘松茅时, 发现绒毛草仅在短期内能通过根系增生来获取斑块内大量养分并促进地上部分生长, 从长期生长来看这种优势则会消失。因此, 本文的实验结果对生产实践中的苗木培育具有一定的指导意义, 但是否在长期生产造林和人工林抚育中仍有指导性需要进一步研究证实。与同质低磷相反, 在异质低磷下, 与HHH相比, HHL不利于马尾松株高、地径和干物质量的提高, 这可能是由于异质低磷土壤中的表层磷含量较高, 导致土壤(酸性土壤) pH值偏高(Shen et al ., 2005 ), 植物在局部供氮后更倾向于对NH4+ -N的吸收, 而过多吸收NH4+ -N会对植物产生一定的毒害作用(Kronzucker et al ., 2001 ), 抑制其整株的生长。然而在有效磷较低的底层土壤中, LLH和LLH/L刺激后, 植株吸收的磷含量显著提高, 且整株干物质量达到均匀供氮处理水平。这表明底层(局部)供氮可促进马尾松苗木对土壤中磷素的有效吸收利用, 保证其快速生长的情况下减少对化学肥料的依赖。
局部供氮对植物根系形态和分布的影响较大(Li et al ., 2014 )。研究结果表明, 在两种低磷环境下, 与HHH相比, LLH/L对马尾松苗木根系的生长发育有明显的促进作用, 尤其在异质低磷土壤环境下的促进作用更为突出, 其次是LLH, HHL反而不利于根系的生长发育。分析认为, 底层半侧供氮有利于根系增生发育的原因可能在于: 植物根系感知一侧氮信号的缺乏, 诱导了根原基和初级根的形成, 而富氮斑块中充足的氮供应则加速了根系的伸长(He et al ., 2003 ; Li et al ., 2014 ), 进而促进了植物整体根长和根表面积的增加。如在异质低磷下, LLH/L处理中的总根长是LLH处理的1.30倍。相似的规律在水稻感知局部磷胁迫的研究中也被发现(He et al ., 2003 )。另一方面, HHL对根长和根表面积有一定的抑制效应, 推测这可能是由于: 首先, 马尾松本身属于深根型植物, 根的向地性促使其向土壤深处觅取更多的养分, 而HHL处理中的底层并没有充足的氮素供应, 限制了根系的生长发育。这一结果与前人对禾本科植物的研究结果有所差异, 如小麦、玉米和水稻(Oryza sativa )等植物可通过增加根系在土壤表层富养区域的分布来响应自然界土壤中的异质养分环境(Weligama et al ., 2008 ; Jing et al ., 2010 ; Li et al ., 2014 )。马尾松作为木本植物, 其对土壤养分异质性分布的响应可能因树种和土壤养分组成的不同而差异较大(Einsmann et al ., 1999 ; Guo et al ., 2002 ; 梅莉等, 2006 ; 杨青等, 2011 ); 其次, 该实验的种子播种于土壤表层, 其生长初期的根系增生和扩张及时获得了营养的补充, 使得植物在养分分配时, 将更多的营养元素输送到了其他器官(茎、叶), 降低了对根系的养分投入, 限制了根系的增生发育(段洪浪等, 2009 ; 吴茜等, 2011 )。结合苗期马尾松的株高、地径等地上部分的生长状况也印证了这一推测。
不同物种或不同基因型植物的氮素利用特性不同, 对不同氮供应方式的适应能力也存在差异。研究结果表明, 同质低磷下, 不同马尾松不同家系地上部分生长对局部供氮的响应差异较大, 而在异质低磷下, 地下部分的根系差异则更为明显。推测认为, 根系形态对土壤磷素异质分布的可塑性较强, 而对局部氮浓度的感知则较为迟钝(Tinker & Nye, 2000 ), 异质低磷环境中的磷素异质性增强了局部氮信号对马尾松苗木根系发育的影响, 进而产生了叠加效应(Jing et al ., 2010 )。在两种低磷环境下, 参试的5个马尾松不同家系中, 除了1号家系对氮添加不敏感外, 其他家系对局部供氮的响应因磷素环境和氮供应方式差异明显。如家系25的干物质积累量与植株氮、磷含量的变化有协同效应, 但与根系生长发育的变化相反, 这表明氮、磷元素在该家系内的快速转运以及利用可能对其整株生长贡献较大 (Kanno et al ., 2016 )。在异质低磷下, HHL限制了家系49、52和57的根系发育, 整株生长受到抑制; 而LLH和LLH/L促进了根系生长和干物质量的积累。这意味着在异质低磷环境下, 家系49、52和57主要通过改变根系的生长发育来响应局部氮素的变化。许多研究证实, 植物对土壤局部营养环境的感知及响应能力受营养元素分布、营养元素组成以及植株自身生长的环境影响(Robinson, 1994 ; Hodge, 2004 )。1号家系对局部供氮的响应较迟钝甚至有抑制效应, 分析认为, 这可能是因为家系1本身的基因型决定了其生长缓慢, 使得外界氮素的变化对植株根系发育的影响较小。本实验中局部养分供应对植物生长的促进效应可获得更多可利用的优良家系, 达到速生增产的目的, 但土壤中的养分异质性不仅是空间分布上的异质性, 也包括时间尺度上的异质性, 在长期的养分异质性环境下, 林木是否依然会保持旺盛的生长, 仍需更多的实验研究和长期观察, 以进一步客观地揭示异质养分土壤环境下, 局部氮素和磷素供应对马尾松不同家系苗木生长发育的影响。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Bilss KM Jones RH Mitchell RJ Mou PP 2002 ). Are competitive interactions influenced by spatial nutrient heterogeneity and root foraging behavior?
New Phytologist, 154 , 409 -417 .
[2]
Drew MC 1975 ). Comparison of effects of a localized supply of phosphate, nitrate, ammonium and potassium on growth of seminal root system and shoot in barley.
New Phytologist, 75 , 479 -490 .
[本文引用: 1]
[3]
Duan HL Liu JX Deng Q Chen XM Zhang DQ 2009 ). Effects of elevated CO2 and N deposition on plant biomass accumulation and allocation in subtropical forest ecosystems: A mesocosm study.
Chinese Journal of Plant Ecology ,33, 570-579.(in Chinese with English abstract) )[段洪浪 , 刘菊秀 , 邓琦 , 陈小梅 , 张德强 (2009 ). CO2 浓度升高与氮沉降对南亚热带森林生态系统植物生物量积累及分配格局的影响
.植物生态学报 , 33, 570-579.]
[本文引用: 2]
[4]
Einsmann JC Jones RH Pu M Mitchell RJ 1999 ). Nutrient forging traits in 10 co-occurring plant species of contrasting life forms.
Journal of Ecology, 87 , 609 -619 .
[本文引用: 1]
[5]
Fransen B de Kroon H 2001 ). Long-term disadvantages of selective root placement: Root proliferation and shoot biomass of two perennial grass species in a 2-year experiment.
Journal of Ecology, 89 , 711 -722 .
[本文引用: 1]
[6]
Graciano C Tanbussi EA Castan E Guiamet JJ 2009 ). Dry mass partitioning and nitrogen uptake by
Eucalyptus grandis plants in response to localized or mixed application of phosphorus. Plant and Soil, 319 , 175 -184 .
[本文引用: 1]
[7]
Guo DL Mou P Jones RH Mitchell RJ 2002 ). Temporal changes in spatial patterns of soil moisture following disturbance: An experimental approach.
Journal of Ecology, 90 , 338 -347 .
[本文引用: 1]
[8]
He Y Liao H Yan XL 2003 ). Localized supply of phosphorus induces root morphological and architectural changes of rice in split and stratified soil cultures.
Plant and Soil, 248 , 247 -256 .
[本文引用: 2]
[9]
Hodge A 2004 ). The plastic plant: Root responses to heterogeneous supplies of nutrients.
New Phytologist, 162 , 9 -24 .
[本文引用: 7]
[10]
Hutchings MJ John EA 2004 ). The effects of environmental heterogeneity on root growth and root/shoot partitioning.
Annals of Botany, 94 , 1 -8 .
[本文引用: 1]
[11]
Jackson RB Caldwell MM 1996 ). Integrating resource heterogeneity and plant plasticity: Modelling nitrate and phosphate uptake in a patchy soil environment.
Journal of Ecologist, 84 , 891 -904 .
[本文引用: 1]
[12]
Jing JY Rui YK Zhang FD Rengel Z Shen JB 2010 ). Localized application of phosphorus and ammonium improves growth of maize seedlings by stimulating root proliferation and rhizosphere acidification.
Field Crops Research, 119 , 355 -364 .
[本文引用: 3]
[13]
Kanno S Arrighi JF Chiarenza S Bayle V Berthom R Peret B Javot H Delannoy E Marin E Nakanishi TM Thibaud MC Nussanme L 2016 ). A novel role for the root cap in phosphate uptake and homeostasis.
eLife, 5 , e14577 . doi: 10.7554/eLife.14577 .
[本文引用: 1]
[14]
Kronzucker HJ Britto DT Davenport RJ Tester M 2001 ). Ammonium toxicity and the real cost of transport.
Trends in Plant Science, 6 , 335 -337 .
[本文引用: 1]
[15]
Li HB Ma QH Li HG Zhang FS Rengel Z Shen JB 2014 ). Root morphological responses to localized nutrient supply differ among crop species with contrasting root traits.
Plant and Soil, 376 , 151 -163 .
[本文引用: 3]
[16]
Li HB Xue MY Lin YR Shen JB 2013 ). Spatial heterogeneity of soil nutrients and root forging: Form individual to community.
Journal of Plant Nutrition and Fertilizer, 19 , 995 -1004 . (in Chinese with English abstract) )[李洪波 , 薛慕瑶 , 林雅茹 , 申建波 (2013 ). 土壤养分空间异质性与根系觅食作用: 从个体到群落
. 植物营养与肥料学报 , 19 , 995 -1004 .]
[本文引用: 1]
[17]
Linkohr BI Williamson LC Fitter AH Leyser HM 2002 ). Nitrate and phosphate availability and distribution have different effects on root system architecture of
Arabidopsis. Plant Journal for Cell & Molecular Biology, 29 , 751 -760 .
[本文引用: 1]
[18]
Liu SE Li YY Fang X Huang WJ Long FL Liu JX 2015 ). Effects of the level and regime of nitrogen addition on seedling growth of four major tree species in subtropical China.
Chinese Journal of Plant Ecology, 39 , 950 -961 . (in Chinese with English abstract) )[刘双娥 , 李义勇 , 方熊 , 黄文娟 , 龙凤玲 , 刘菊秀 (2015 ). 不同氮添加量和添加方式对南亚热带4个主要树种幼苗生长的影响
. 植物生态学报 , 39 , 950 -961 .]
[本文引用: 2]
[19]
Lü CQ Tian HQ Huang Y 2007 ). Ecological effects of increased nitrogen deposition in terrestrial ecosystems.
Journal of Plant Ecology (Chinese Version )31 , 205 -218 . (in Chinese with English abstract) )[吕超群 , 田汉勤 , 黄耀 (2007 ). 陆地生态系统氮沉降增加的生态效应
. 植物生态学报 , 31 , 205 -218 .]
[本文引用: 1]
[20]
Mei L Wang ZQ Han YZ Gu JC Wang XR Cheng YH Zhang XJ 2006 ). Distribution patterns of Fraxinus mandshurica root biomass, specific root length and root length density.
Chinese Journal of Applied Ecology, 17 , 1 -4 . (in Chinese with English abstract) )[梅莉 , 王政权 , 韩有志 , 谷加存 , 王向荣 , 程云环 , 张秀娟 (2006 ). 水曲柳根系生物量、比根长和根长密度的分布格局
. 应用生态学报 , 17 , 1 -4 .
[本文引用: 1]
[21]
Mommer L van Ruijven J Jansen C van de Steeg HM de Kroon H 2012 ). Interactive effects of nutrient heterogeneity and competition: Implications for root foraging theory?
Functional Ecology, 26 , 66 -73 .
[本文引用: 1]
[22]
Mou P Mitchell RJ Jones RH 1997 ). Root distribution of two tree species under a heterogeneous nutrient environment.
Journal Applied Ecology, 34 , 645 -656 .
[本文引用: 1]
[23]
Officer SJ Dunbabin VM Armstrong RD Norton RM Kearney GA 2009 ). Wheat roots proliferate in response to nitrogen and phosphorus fertilisers in Sodosol and Vertosol soils of south-eastern Australia.
Australian Journal of Soil Research, 47 , 91 -102 .
[本文引用: 1]
[24]
Pang L Zhang Y Zhou ZC Feng ZP Chu DY 2014 ). Effects of simulated nitrogen deposition on growth and phosphorus efficiency of Pinus massoniana under low phosphorus stress.
Chinese Journal of Applied Ecology, 25 , 1275 -1282 . (in Chinese with English abstract) )[庞丽 , 张一 , 周志春 , 丰忠平 , 储徳裕 (2014 ). 模拟氮沉降对低磷胁迫下马尾松生长和磷效率的影响
. 应用生态学报 , 25 , 1275 -1282 .]
[本文引用: 4]
[25]
Qin GF Zhou ZC 2012). Germplasm Resources of Chinese Masson Pine . China Forestry Publishing House, Beijing . (in Chinese) )[秦国峰 , 周志春 (2012). 中国马尾松优良种质资源 . 中国林业出版社 , 北京 .]
[本文引用: 1]
[26]
Richardson SJ Peltzer DA Allen RB McGlone MS Parfitt RL 2004 ). Rapid development of phosphorus limitation in temperate rainforest along the Franz Josef soil chronosequence.
Oecologia, 139 , 267 -276 .
[本文引用: 1]
[27]
Robinson D 1994 ). The response of plants to non-uniform supplies of nutrients.
New Phytologist, 127 , 635 -674 .
[本文引用: 1]
[28]
Shen J Li H Neumann G Zhang F 2005 ). Nutrient uptake, cluster root formation and exudation of protons and citrate in Lupinus albus as affected by localized supply of phosphorus in a split-root system.
Plant Science, 168 , 837 -845 .
[本文引用: 1]
[29]
Tinker PB Nye PH Solute Movement in the Rhizosphere . Oxford University Press, Oxford .
[本文引用: 1]
[30]
Vance CP Uhde-stone C Allan DL 2003 ). Phosphorus acquisition and use: Critical adaptations by plants for securing a nonrenewable resource.
New Phytologist, 57 , 423 -427 .
[本文引用: 1]
[31]
Verma S Subehia SK Sharma SP 2005 ). Phosphorus fractions in an acid soil continuously fertilized with mineral and organic fertilizers.
Biology and Fertility of Soils, 41 , 295 -300 .
[本文引用: 1]
[32]
Weligama C Tang C Sale PWG Conyers MK Liu DL 2008 ). Localized nitrate and phosphate application enhances root proliferation by wheat and maximises rhizosphere alkalization in acid subsoil.
Plant and Soil, 312 , 101 -115 .
[本文引用: 1]
[33]
Wu Q Ding J Yan H Zhang SR Fang T Ma KP 2011 ). Effects of simulated precipitation and nitrogen addition on seedling growth and biomass in five tree species in Gutian Mountain, Zhejiang Province, China.
Chinese Journal of Plant Ecology, 35 , 256 -267 . (in Chinese with English abstract) )[吴茜 , 丁佳 , 闫慧 , 张守仁 , 方腾 , 马克平 (2011 ). 模拟降水变化和土壤施氮对浙江古田山5个树种幼苗生长和生物量的影响
. 植物生态学报 , 35 , 256 -267 .
[本文引用: 1]
[34]
Xu XK Han L Luo XB 2012 ). Effects of stimulated nitrogen deposition on soil net nitrogen mineralization under a temperate Korean pine and broadleaf mixed forest.
Climatic and Environmental Research, 17 , 628 -638 . (in Chinese with English abstract) )[徐星凯 , 韩林 , 罗献宝 (2012 ). 模拟氮沉降对温带阔叶红松林地氮素净矿化量的影响
. 气候与环境研究 , 17 , 628 -638 .]
[本文引用: 2]
[35]
Yang Q Zhang Y Zhou ZC Ma XH Liu WH Feng ZP 2011 ). Genetic variation in root architecture and phosphorus efficiency in response to heterogeneous phosphorus deficiency in Pinus massoniana families.
Chinese Journal of Plant Ecology, 35 , 1226 -1235 . (in Chinese with English abstract) )[杨青 , 张一 , 周志春 , 马雪红 , 刘伟宏 , 丰忠平 (2011 ). 异质低磷胁迫下马尾松家系根构型和磷效率的遗传变异
. 植物生态学报 , 35 , 1226 -1235 .]
[本文引用: 3]
[36]
Zhang H Forde BG 2000 ). Regulation of Arabidopsis root development by nitrate availability.
Journal of Experimental Botany, 51 , 51 -59 .
[37]
Zhang Y Zhou ZC Yang Q 2013 ). Genetic variations in root morphology and phosphorus efficiency of Pinus massoniana under heterogeneous and homogeneous low phosphorus conditions.
Plant and Soil, 364 , 93 -104 .
[本文引用: 1]
[38]
Zhou ZC Shangguan ZP 2007 ). Vertical distribution of fine roots in relation to soil factors in Pinus tabulaeformis Carr. forest of the Loess Plateau of China.
Plant and Soil, 291 , 119 -129 .
[本文引用: 1]
Are competitive interactions influenced by spatial nutrient heterogeneity and root foraging behavior?
2002
Comparison of effects of a localized supply of phosphate, nitrate, ammonium and potassium on growth of seminal root system and shoot in barley.
1
1975
... 氮和磷是植物生长最重要的两种营养元素, 其有效性和空间分布对植物的生长和根系发育具有重要影响(Hodge, 2004 ; Vance et al ., 2003 ).研究揭示, 局部供氮或供磷可显著促进大麦(Hordeum vulgare )、玉米(Zea mays )和小麦(Triticum aestivum )等植物侧根数量的增加以及总根长的提高(Drew, 1975 ; Officer et al ., 2009 ).Zhang和Forde (2000)将拟南芥(Arabidopsis thaliana )分别放在局部高氮和均匀高氮的琼脂培养基上培养时发现, 根系会在富氮斑块中增生, 而均匀供氮处理中的根系总量降低.可见根系在富养斑块中的增生可促进植物对养分供应的有效利用(Linkohr et al ., 2002 ).更重要的是, 相比于单独局部供氮或供磷, 在斑块中同时添加氮和磷更能促进根系在富养斑块中增生, 提高营养元素的吸收效率, 进而加快整株的生长(Jing et al ., 2010 ).这表明植物根系在局部供养斑块中的增生受到营养元素组成以及元素之间互作效应的共同影响.在木本植物研究中也发现类似的变化规律, 但物种间差异较大(Hodge, 2004 ).Einsmann等(1999)发现, 10种木本植物中有4种植物的生物量在养分异质性的供应条件下有显著提高, 但在对同种植物的不同研究中, Bliss等(2002)并没有发现这一趋势.虽然有研究认为养分斑块的存在能够对植物地上部分的生长产生积极的促进效应(Jackson & Caldwell, 1996 ), 但多年生植物绒毛草(Holcus lanatus )仅在短期内出现大量根系增生和促进地上部分生长的现象, 长期来看这种优势很快会消失, 甘松茅(Nardus stricta )则对养分斑块反应不敏感(Fransen & de Kroon, 2001 ).显然, 养分斑块或者局部养分供应对植物生长的影响受很多因素的影响, 除了养分的属性外, 植物对养分的敏感性及试验中植物生长时间的长短对结果都会产生较大影响(李洪波等, 2013 ). ...
CO2 浓度升高与氮沉降对南亚热带森林生态系统植物生物量积累及分配格局的影响
2
2009
... 近年来, 土壤中养分元素的分布方式对植株生长的影响越来越受到关注(Hodge, 2004 ; 吕超群等, 2007 ; 徐星凯等, 2012 ).本研究中的异质低磷环境较同质低磷更有利于马尾松苗木的生长, 与前人的研究结果(杨青等, 2011 ; 庞丽等, 2014 )一致.已有研究表明, 外源氮素添加(100 g·kg-1 )会使土壤有效氮含量增加, 改变植株生物量的分配进而促进整株的生长(段洪浪等, 2009 ; 庞丽等, 2014 ; 刘双娥等, 2015 ).在本实验中, 局部供氮处理的斑块氮浓度与均匀供氮处理的氮含量相同, 但总施肥量要少于均匀供氮(相当于HHH处理施氮量的1/6-2/3).研究结果显示, 不同的供氮方式对马尾松苗木生长的影响差异较大, 且受到土壤磷素环境影响.在同质低磷下, HHL显著促进了马尾松苗木的生长, 根冠比降低, 较其他供氮方式更有利于苗木地上部分的生长; 其次, LLH和LLH/L两种方式之间, LLH/L对马尾松整株的生长相对更有利, 这表明在同质低磷下, 局部供氮增强了土壤中养分的异质性, 促进了马尾松苗木整株的生长.类似的结果在桉树(Eucalyptus grandis )、油松(Pinus tabulaeformis )、火炬松(Pinus taeda )等植物中也被发现(Mou et al ., 1997 ; Zhou & Shangguan, 2007 ; Graciano et al ., 2009 ).然而, 值得注意的是, Fransen和de Kroon (2001)在研究多年生植物绒毛草和甘松茅时, 发现绒毛草仅在短期内能通过根系增生来获取斑块内大量养分并促进地上部分生长, 从长期生长来看这种优势则会消失.因此, 本文的实验结果对生产实践中的苗木培育具有一定的指导意义, 但是否在长期生产造林和人工林抚育中仍有指导性需要进一步研究证实.与同质低磷相反, 在异质低磷下, 与HHH相比, HHL不利于马尾松株高、地径和干物质量的提高, 这可能是由于异质低磷土壤中的表层磷含量较高, 导致土壤(酸性土壤) pH值偏高(Shen et al ., 2005 ), 植物在局部供氮后更倾向于对NH4+ -N的吸收, 而过多吸收NH4+ -N会对植物产生一定的毒害作用(Kronzucker et al ., 2001 ), 抑制其整株的生长.然而在有效磷较低的底层土壤中, LLH和LLH/L刺激后, 植株吸收的磷含量显著提高, 且整株干物质量达到均匀供氮处理水平.这表明底层(局部)供氮可促进马尾松苗木对土壤中磷素的有效吸收利用, 保证其快速生长的情况下减少对化学肥料的依赖. ...
... 局部供氮对植物根系形态和分布的影响较大(Li et al ., 2014 ).研究结果表明, 在两种低磷环境下, 与HHH相比, LLH/L对马尾松苗木根系的生长发育有明显的促进作用, 尤其在异质低磷土壤环境下的促进作用更为突出, 其次是LLH, HHL反而不利于根系的生长发育.分析认为, 底层半侧供氮有利于根系增生发育的原因可能在于: 植物根系感知一侧氮信号的缺乏, 诱导了根原基和初级根的形成, 而富氮斑块中充足的氮供应则加速了根系的伸长(He et al ., 2003 ; Li et al ., 2014 ), 进而促进了植物整体根长和根表面积的增加.如在异质低磷下, LLH/L处理中的总根长是LLH处理的1.30倍.相似的规律在水稻感知局部磷胁迫的研究中也被发现(He et al ., 2003 ).另一方面, HHL对根长和根表面积有一定的抑制效应, 推测这可能是由于: 首先, 马尾松本身属于深根型植物, 根的向地性促使其向土壤深处觅取更多的养分, 而HHL处理中的底层并没有充足的氮素供应, 限制了根系的生长发育.这一结果与前人对禾本科植物的研究结果有所差异, 如小麦、玉米和水稻(Oryza sativa )等植物可通过增加根系在土壤表层富养区域的分布来响应自然界土壤中的异质养分环境(Weligama et al ., 2008 ; Jing et al ., 2010 ; Li et al ., 2014 ).马尾松作为木本植物, 其对土壤养分异质性分布的响应可能因树种和土壤养分组成的不同而差异较大(Einsmann et al ., 1999 ; Guo et al ., 2002 ; 梅莉等, 2006 ; 杨青等, 2011 ); 其次, 该实验的种子播种于土壤表层, 其生长初期的根系增生和扩张及时获得了营养的补充, 使得植物在养分分配时, 将更多的营养元素输送到了其他器官(茎、叶), 降低了对根系的养分投入, 限制了根系的增生发育(段洪浪等, 2009 ; 吴茜等, 2011 ).结合苗期马尾松的株高、地径等地上部分的生长状况也印证了这一推测. ...
Nutrient forging traits in 10 co-occurring plant species of contrasting life forms.
1
1999
... 局部供氮对植物根系形态和分布的影响较大(Li et al ., 2014 ).研究结果表明, 在两种低磷环境下, 与HHH相比, LLH/L对马尾松苗木根系的生长发育有明显的促进作用, 尤其在异质低磷土壤环境下的促进作用更为突出, 其次是LLH, HHL反而不利于根系的生长发育.分析认为, 底层半侧供氮有利于根系增生发育的原因可能在于: 植物根系感知一侧氮信号的缺乏, 诱导了根原基和初级根的形成, 而富氮斑块中充足的氮供应则加速了根系的伸长(He et al ., 2003 ; Li et al ., 2014 ), 进而促进了植物整体根长和根表面积的增加.如在异质低磷下, LLH/L处理中的总根长是LLH处理的1.30倍.相似的规律在水稻感知局部磷胁迫的研究中也被发现(He et al ., 2003 ).另一方面, HHL对根长和根表面积有一定的抑制效应, 推测这可能是由于: 首先, 马尾松本身属于深根型植物, 根的向地性促使其向土壤深处觅取更多的养分, 而HHL处理中的底层并没有充足的氮素供应, 限制了根系的生长发育.这一结果与前人对禾本科植物的研究结果有所差异, 如小麦、玉米和水稻(Oryza sativa )等植物可通过增加根系在土壤表层富养区域的分布来响应自然界土壤中的异质养分环境(Weligama et al ., 2008 ; Jing et al ., 2010 ; Li et al ., 2014 ).马尾松作为木本植物, 其对土壤养分异质性分布的响应可能因树种和土壤养分组成的不同而差异较大(Einsmann et al ., 1999 ; Guo et al ., 2002 ; 梅莉等, 2006 ; 杨青等, 2011 ); 其次, 该实验的种子播种于土壤表层, 其生长初期的根系增生和扩张及时获得了营养的补充, 使得植物在养分分配时, 将更多的营养元素输送到了其他器官(茎、叶), 降低了对根系的养分投入, 限制了根系的增生发育(段洪浪等, 2009 ; 吴茜等, 2011 ).结合苗期马尾松的株高、地径等地上部分的生长状况也印证了这一推测. ...
Long-term disadvantages of selective root placement: Root proliferation and shoot biomass of two perennial grass species in a 2-year experiment.
1
2001
... 氮和磷是植物生长最重要的两种营养元素, 其有效性和空间分布对植物的生长和根系发育具有重要影响(Hodge, 2004 ; Vance et al ., 2003 ).研究揭示, 局部供氮或供磷可显著促进大麦(Hordeum vulgare )、玉米(Zea mays )和小麦(Triticum aestivum )等植物侧根数量的增加以及总根长的提高(Drew, 1975 ; Officer et al ., 2009 ).Zhang和Forde (2000)将拟南芥(Arabidopsis thaliana )分别放在局部高氮和均匀高氮的琼脂培养基上培养时发现, 根系会在富氮斑块中增生, 而均匀供氮处理中的根系总量降低.可见根系在富养斑块中的增生可促进植物对养分供应的有效利用(Linkohr et al ., 2002 ).更重要的是, 相比于单独局部供氮或供磷, 在斑块中同时添加氮和磷更能促进根系在富养斑块中增生, 提高营养元素的吸收效率, 进而加快整株的生长(Jing et al ., 2010 ).这表明植物根系在局部供养斑块中的增生受到营养元素组成以及元素之间互作效应的共同影响.在木本植物研究中也发现类似的变化规律, 但物种间差异较大(Hodge, 2004 ).Einsmann等(1999)发现, 10种木本植物中有4种植物的生物量在养分异质性的供应条件下有显著提高, 但在对同种植物的不同研究中, Bliss等(2002)并没有发现这一趋势.虽然有研究认为养分斑块的存在能够对植物地上部分的生长产生积极的促进效应(Jackson & Caldwell, 1996 ), 但多年生植物绒毛草(Holcus lanatus )仅在短期内出现大量根系增生和促进地上部分生长的现象, 长期来看这种优势很快会消失, 甘松茅(Nardus stricta )则对养分斑块反应不敏感(Fransen & de Kroon, 2001 ).显然, 养分斑块或者局部养分供应对植物生长的影响受很多因素的影响, 除了养分的属性外, 植物对养分的敏感性及试验中植物生长时间的长短对结果都会产生较大影响(李洪波等, 2013 ). ...
Dry mass partitioning and nitrogen uptake by
1
2009
... 近年来, 土壤中养分元素的分布方式对植株生长的影响越来越受到关注(Hodge, 2004 ; 吕超群等, 2007 ; 徐星凯等, 2012 ).本研究中的异质低磷环境较同质低磷更有利于马尾松苗木的生长, 与前人的研究结果(杨青等, 2011 ; 庞丽等, 2014 )一致.已有研究表明, 外源氮素添加(100 g·kg-1 )会使土壤有效氮含量增加, 改变植株生物量的分配进而促进整株的生长(段洪浪等, 2009 ; 庞丽等, 2014 ; 刘双娥等, 2015 ).在本实验中, 局部供氮处理的斑块氮浓度与均匀供氮处理的氮含量相同, 但总施肥量要少于均匀供氮(相当于HHH处理施氮量的1/6-2/3).研究结果显示, 不同的供氮方式对马尾松苗木生长的影响差异较大, 且受到土壤磷素环境影响.在同质低磷下, HHL显著促进了马尾松苗木的生长, 根冠比降低, 较其他供氮方式更有利于苗木地上部分的生长; 其次, LLH和LLH/L两种方式之间, LLH/L对马尾松整株的生长相对更有利, 这表明在同质低磷下, 局部供氮增强了土壤中养分的异质性, 促进了马尾松苗木整株的生长.类似的结果在桉树(Eucalyptus grandis )、油松(Pinus tabulaeformis )、火炬松(Pinus taeda )等植物中也被发现(Mou et al ., 1997 ; Zhou & Shangguan, 2007 ; Graciano et al ., 2009 ).然而, 值得注意的是, Fransen和de Kroon (2001)在研究多年生植物绒毛草和甘松茅时, 发现绒毛草仅在短期内能通过根系增生来获取斑块内大量养分并促进地上部分生长, 从长期生长来看这种优势则会消失.因此, 本文的实验结果对生产实践中的苗木培育具有一定的指导意义, 但是否在长期生产造林和人工林抚育中仍有指导性需要进一步研究证实.与同质低磷相反, 在异质低磷下, 与HHH相比, HHL不利于马尾松株高、地径和干物质量的提高, 这可能是由于异质低磷土壤中的表层磷含量较高, 导致土壤(酸性土壤) pH值偏高(Shen et al ., 2005 ), 植物在局部供氮后更倾向于对NH4+ -N的吸收, 而过多吸收NH4+ -N会对植物产生一定的毒害作用(Kronzucker et al ., 2001 ), 抑制其整株的生长.然而在有效磷较低的底层土壤中, LLH和LLH/L刺激后, 植株吸收的磷含量显著提高, 且整株干物质量达到均匀供氮处理水平.这表明底层(局部)供氮可促进马尾松苗木对土壤中磷素的有效吸收利用, 保证其快速生长的情况下减少对化学肥料的依赖. ...
Temporal changes in spatial patterns of soil moisture following disturbance: An experimental approach.
1
2002
... 局部供氮对植物根系形态和分布的影响较大(Li et al ., 2014 ).研究结果表明, 在两种低磷环境下, 与HHH相比, LLH/L对马尾松苗木根系的生长发育有明显的促进作用, 尤其在异质低磷土壤环境下的促进作用更为突出, 其次是LLH, HHL反而不利于根系的生长发育.分析认为, 底层半侧供氮有利于根系增生发育的原因可能在于: 植物根系感知一侧氮信号的缺乏, 诱导了根原基和初级根的形成, 而富氮斑块中充足的氮供应则加速了根系的伸长(He et al ., 2003 ; Li et al ., 2014 ), 进而促进了植物整体根长和根表面积的增加.如在异质低磷下, LLH/L处理中的总根长是LLH处理的1.30倍.相似的规律在水稻感知局部磷胁迫的研究中也被发现(He et al ., 2003 ).另一方面, HHL对根长和根表面积有一定的抑制效应, 推测这可能是由于: 首先, 马尾松本身属于深根型植物, 根的向地性促使其向土壤深处觅取更多的养分, 而HHL处理中的底层并没有充足的氮素供应, 限制了根系的生长发育.这一结果与前人对禾本科植物的研究结果有所差异, 如小麦、玉米和水稻(Oryza sativa )等植物可通过增加根系在土壤表层富养区域的分布来响应自然界土壤中的异质养分环境(Weligama et al ., 2008 ; Jing et al ., 2010 ; Li et al ., 2014 ).马尾松作为木本植物, 其对土壤养分异质性分布的响应可能因树种和土壤养分组成的不同而差异较大(Einsmann et al ., 1999 ; Guo et al ., 2002 ; 梅莉等, 2006 ; 杨青等, 2011 ); 其次, 该实验的种子播种于土壤表层, 其生长初期的根系增生和扩张及时获得了营养的补充, 使得植物在养分分配时, 将更多的营养元素输送到了其他器官(茎、叶), 降低了对根系的养分投入, 限制了根系的增生发育(段洪浪等, 2009 ; 吴茜等, 2011 ).结合苗期马尾松的株高、地径等地上部分的生长状况也印证了这一推测. ...
Localized supply of phosphorus induces root morphological and architectural changes of rice in split and stratified soil cultures.
2
2003
... 局部供氮对植物根系形态和分布的影响较大(Li et al ., 2014 ).研究结果表明, 在两种低磷环境下, 与HHH相比, LLH/L对马尾松苗木根系的生长发育有明显的促进作用, 尤其在异质低磷土壤环境下的促进作用更为突出, 其次是LLH, HHL反而不利于根系的生长发育.分析认为, 底层半侧供氮有利于根系增生发育的原因可能在于: 植物根系感知一侧氮信号的缺乏, 诱导了根原基和初级根的形成, 而富氮斑块中充足的氮供应则加速了根系的伸长(He et al ., 2003 ; Li et al ., 2014 ), 进而促进了植物整体根长和根表面积的增加.如在异质低磷下, LLH/L处理中的总根长是LLH处理的1.30倍.相似的规律在水稻感知局部磷胁迫的研究中也被发现(He et al ., 2003 ).另一方面, HHL对根长和根表面积有一定的抑制效应, 推测这可能是由于: 首先, 马尾松本身属于深根型植物, 根的向地性促使其向土壤深处觅取更多的养分, 而HHL处理中的底层并没有充足的氮素供应, 限制了根系的生长发育.这一结果与前人对禾本科植物的研究结果有所差异, 如小麦、玉米和水稻(Oryza sativa )等植物可通过增加根系在土壤表层富养区域的分布来响应自然界土壤中的异质养分环境(Weligama et al ., 2008 ; Jing et al ., 2010 ; Li et al ., 2014 ).马尾松作为木本植物, 其对土壤养分异质性分布的响应可能因树种和土壤养分组成的不同而差异较大(Einsmann et al ., 1999 ; Guo et al ., 2002 ; 梅莉等, 2006 ; 杨青等, 2011 ); 其次, 该实验的种子播种于土壤表层, 其生长初期的根系增生和扩张及时获得了营养的补充, 使得植物在养分分配时, 将更多的营养元素输送到了其他器官(茎、叶), 降低了对根系的养分投入, 限制了根系的增生发育(段洪浪等, 2009 ; 吴茜等, 2011 ).结合苗期马尾松的株高、地径等地上部分的生长状况也印证了这一推测. ...
... ), 进而促进了植物整体根长和根表面积的增加.如在异质低磷下, LLH/L处理中的总根长是LLH处理的1.30倍.相似的规律在水稻感知局部磷胁迫的研究中也被发现(He et al ., 2003 ).另一方面, HHL对根长和根表面积有一定的抑制效应, 推测这可能是由于: 首先, 马尾松本身属于深根型植物, 根的向地性促使其向土壤深处觅取更多的养分, 而HHL处理中的底层并没有充足的氮素供应, 限制了根系的生长发育.这一结果与前人对禾本科植物的研究结果有所差异, 如小麦、玉米和水稻(Oryza sativa )等植物可通过增加根系在土壤表层富养区域的分布来响应自然界土壤中的异质养分环境(Weligama et al ., 2008 ; Jing et al ., 2010 ; Li et al ., 2014 ).马尾松作为木本植物, 其对土壤养分异质性分布的响应可能因树种和土壤养分组成的不同而差异较大(Einsmann et al ., 1999 ; Guo et al ., 2002 ; 梅莉等, 2006 ; 杨青等, 2011 ); 其次, 该实验的种子播种于土壤表层, 其生长初期的根系增生和扩张及时获得了营养的补充, 使得植物在养分分配时, 将更多的营养元素输送到了其他器官(茎、叶), 降低了对根系的养分投入, 限制了根系的增生发育(段洪浪等, 2009 ; 吴茜等, 2011 ).结合苗期马尾松的株高、地径等地上部分的生长状况也印证了这一推测. ...
The plastic plant: Root responses to heterogeneous supplies of nutrients.
7
2004
... 土壤养分在空间分布上多呈时空异质性或者斑块分布(Hutching & John, 2004 ).产生土壤养分空间异质性的原因很多, 其中包括有机物(凋落物、根系死亡以及动物尸体)的分解、无机养分在土壤中的扩散系数差异以及微地形因素影响等(Hodge, 2004 ; 徐星凯等, 2012 ; 刘双娥等, 2015 ).植物为了获得更多的养分, 其根系会增加在土壤养分富集区域的分布比例, 进而提高根系对养分的获取和利用能力(Hodge, 2004 ; Mommer et al ., 2012 ).近年来, 随着合理高效的供肥管理措施(局部施肥)在农业中广泛应用及其高效的生产力, 使其在林业上尤其是人工林经营管理中也开始得到广泛应用.因此, 了解肥料集约经营管理下的林木根系生长、形态分布以及养分吸收状况, 将有利于解释林木在生长上存在的差异, 同时为提高速生丰产林生产力提供更为完善和高效的技术指导. ...
... ).植物为了获得更多的养分, 其根系会增加在土壤养分富集区域的分布比例, 进而提高根系对养分的获取和利用能力(Hodge, 2004 ; Mommer et al ., 2012 ).近年来, 随着合理高效的供肥管理措施(局部施肥)在农业中广泛应用及其高效的生产力, 使其在林业上尤其是人工林经营管理中也开始得到广泛应用.因此, 了解肥料集约经营管理下的林木根系生长、形态分布以及养分吸收状况, 将有利于解释林木在生长上存在的差异, 同时为提高速生丰产林生产力提供更为完善和高效的技术指导. ...
... 氮和磷是植物生长最重要的两种营养元素, 其有效性和空间分布对植物的生长和根系发育具有重要影响(Hodge, 2004 ; Vance et al ., 2003 ).研究揭示, 局部供氮或供磷可显著促进大麦(Hordeum vulgare )、玉米(Zea mays )和小麦(Triticum aestivum )等植物侧根数量的增加以及总根长的提高(Drew, 1975 ; Officer et al ., 2009 ).Zhang和Forde (2000)将拟南芥(Arabidopsis thaliana )分别放在局部高氮和均匀高氮的琼脂培养基上培养时发现, 根系会在富氮斑块中增生, 而均匀供氮处理中的根系总量降低.可见根系在富养斑块中的增生可促进植物对养分供应的有效利用(Linkohr et al ., 2002 ).更重要的是, 相比于单独局部供氮或供磷, 在斑块中同时添加氮和磷更能促进根系在富养斑块中增生, 提高营养元素的吸收效率, 进而加快整株的生长(Jing et al ., 2010 ).这表明植物根系在局部供养斑块中的增生受到营养元素组成以及元素之间互作效应的共同影响.在木本植物研究中也发现类似的变化规律, 但物种间差异较大(Hodge, 2004 ).Einsmann等(1999)发现, 10种木本植物中有4种植物的生物量在养分异质性的供应条件下有显著提高, 但在对同种植物的不同研究中, Bliss等(2002)并没有发现这一趋势.虽然有研究认为养分斑块的存在能够对植物地上部分的生长产生积极的促进效应(Jackson & Caldwell, 1996 ), 但多年生植物绒毛草(Holcus lanatus )仅在短期内出现大量根系增生和促进地上部分生长的现象, 长期来看这种优势很快会消失, 甘松茅(Nardus stricta )则对养分斑块反应不敏感(Fransen & de Kroon, 2001 ).显然, 养分斑块或者局部养分供应对植物生长的影响受很多因素的影响, 除了养分的属性外, 植物对养分的敏感性及试验中植物生长时间的长短对结果都会产生较大影响(李洪波等, 2013 ). ...
... ).这表明植物根系在局部供养斑块中的增生受到营养元素组成以及元素之间互作效应的共同影响.在木本植物研究中也发现类似的变化规律, 但物种间差异较大(Hodge, 2004 ).Einsmann等(1999)发现, 10种木本植物中有4种植物的生物量在养分异质性的供应条件下有显著提高, 但在对同种植物的不同研究中, Bliss等(2002)并没有发现这一趋势.虽然有研究认为养分斑块的存在能够对植物地上部分的生长产生积极的促进效应(Jackson & Caldwell, 1996 ), 但多年生植物绒毛草(Holcus lanatus )仅在短期内出现大量根系增生和促进地上部分生长的现象, 长期来看这种优势很快会消失, 甘松茅(Nardus stricta )则对养分斑块反应不敏感(Fransen & de Kroon, 2001 ).显然, 养分斑块或者局部养分供应对植物生长的影响受很多因素的影响, 除了养分的属性外, 植物对养分的敏感性及试验中植物生长时间的长短对结果都会产生较大影响(李洪波等, 2013 ). ...
... 低磷土壤环境设置参照Zhang等(2013)的浓度设计.基质土壤经过风干过筛后, 加入一定量的Ca(H2 PO4 )2 混匀.异质低磷盆栽基质的底层、中间层和表层依次填入添加0、0.10和1.00 g·kg-1 的Ca(H2 PO4 )2 , 相当于有效磷含量分别为0.99、15和 30 mg·kg-1 .盆栽基质由上而下有效磷含量逐渐降低的异质养分环境可模拟立地条件较好, 表层营养丰富的针阔混交林或阔叶林土壤环境(Hodge, 2004 ; Verma et al ., 2005 ).同质低磷盆栽基质的表层、中间层和底层皆均匀添加有效磷含量极低(0.99 mg·kg-1 )的贫瘠酸性红壤作为盆栽基质, 该处理可有效地模拟森林土壤中磷素极为匮乏的针叶林立地环境(Richaedson et al ., 2004 ). ...
... 近年来, 土壤中养分元素的分布方式对植株生长的影响越来越受到关注(Hodge, 2004 ; 吕超群等, 2007 ; 徐星凯等, 2012 ).本研究中的异质低磷环境较同质低磷更有利于马尾松苗木的生长, 与前人的研究结果(杨青等, 2011 ; 庞丽等, 2014 )一致.已有研究表明, 外源氮素添加(100 g·kg-1 )会使土壤有效氮含量增加, 改变植株生物量的分配进而促进整株的生长(段洪浪等, 2009 ; 庞丽等, 2014 ; 刘双娥等, 2015 ).在本实验中, 局部供氮处理的斑块氮浓度与均匀供氮处理的氮含量相同, 但总施肥量要少于均匀供氮(相当于HHH处理施氮量的1/6-2/3).研究结果显示, 不同的供氮方式对马尾松苗木生长的影响差异较大, 且受到土壤磷素环境影响.在同质低磷下, HHL显著促进了马尾松苗木的生长, 根冠比降低, 较其他供氮方式更有利于苗木地上部分的生长; 其次, LLH和LLH/L两种方式之间, LLH/L对马尾松整株的生长相对更有利, 这表明在同质低磷下, 局部供氮增强了土壤中养分的异质性, 促进了马尾松苗木整株的生长.类似的结果在桉树(Eucalyptus grandis )、油松(Pinus tabulaeformis )、火炬松(Pinus taeda )等植物中也被发现(Mou et al ., 1997 ; Zhou & Shangguan, 2007 ; Graciano et al ., 2009 ).然而, 值得注意的是, Fransen和de Kroon (2001)在研究多年生植物绒毛草和甘松茅时, 发现绒毛草仅在短期内能通过根系增生来获取斑块内大量养分并促进地上部分生长, 从长期生长来看这种优势则会消失.因此, 本文的实验结果对生产实践中的苗木培育具有一定的指导意义, 但是否在长期生产造林和人工林抚育中仍有指导性需要进一步研究证实.与同质低磷相反, 在异质低磷下, 与HHH相比, HHL不利于马尾松株高、地径和干物质量的提高, 这可能是由于异质低磷土壤中的表层磷含量较高, 导致土壤(酸性土壤) pH值偏高(Shen et al ., 2005 ), 植物在局部供氮后更倾向于对NH4+ -N的吸收, 而过多吸收NH4+ -N会对植物产生一定的毒害作用(Kronzucker et al ., 2001 ), 抑制其整株的生长.然而在有效磷较低的底层土壤中, LLH和LLH/L刺激后, 植株吸收的磷含量显著提高, 且整株干物质量达到均匀供氮处理水平.这表明底层(局部)供氮可促进马尾松苗木对土壤中磷素的有效吸收利用, 保证其快速生长的情况下减少对化学肥料的依赖. ...
... 不同物种或不同基因型植物的氮素利用特性不同, 对不同氮供应方式的适应能力也存在差异.研究结果表明, 同质低磷下, 不同马尾松不同家系地上部分生长对局部供氮的响应差异较大, 而在异质低磷下, 地下部分的根系差异则更为明显.推测认为, 根系形态对土壤磷素异质分布的可塑性较强, 而对局部氮浓度的感知则较为迟钝(Tinker & Nye, 2000 ), 异质低磷环境中的磷素异质性增强了局部氮信号对马尾松苗木根系发育的影响, 进而产生了叠加效应(Jing et al ., 2010 ).在两种低磷环境下, 参试的5个马尾松不同家系中, 除了1号家系对氮添加不敏感外, 其他家系对局部供氮的响应因磷素环境和氮供应方式差异明显.如家系25的干物质积累量与植株氮、磷含量的变化有协同效应, 但与根系生长发育的变化相反, 这表明氮、磷元素在该家系内的快速转运以及利用可能对其整株生长贡献较大 (Kanno et al ., 2016 ).在异质低磷下, HHL限制了家系49、52和57的根系发育, 整株生长受到抑制; 而LLH和LLH/L促进了根系生长和干物质量的积累.这意味着在异质低磷环境下, 家系49、52和57主要通过改变根系的生长发育来响应局部氮素的变化.许多研究证实, 植物对土壤局部营养环境的感知及响应能力受营养元素分布、营养元素组成以及植株自身生长的环境影响(Robinson, 1994 ; Hodge, 2004 ).1号家系对局部供氮的响应较迟钝甚至有抑制效应, 分析认为, 这可能是因为家系1本身的基因型决定了其生长缓慢, 使得外界氮素的变化对植株根系发育的影响较小.本实验中局部养分供应对植物生长的促进效应可获得更多可利用的优良家系, 达到速生增产的目的, 但土壤中的养分异质性不仅是空间分布上的异质性, 也包括时间尺度上的异质性, 在长期的养分异质性环境下, 林木是否依然会保持旺盛的生长, 仍需更多的实验研究和长期观察, 以进一步客观地揭示异质养分土壤环境下, 局部氮素和磷素供应对马尾松不同家系苗木生长发育的影响. ...
The effects of environmental heterogeneity on root growth and root/shoot partitioning.
1
2004
... 土壤养分在空间分布上多呈时空异质性或者斑块分布(Hutching & John, 2004 ).产生土壤养分空间异质性的原因很多, 其中包括有机物(凋落物、根系死亡以及动物尸体)的分解、无机养分在土壤中的扩散系数差异以及微地形因素影响等(Hodge, 2004 ; 徐星凯等, 2012 ; 刘双娥等, 2015 ).植物为了获得更多的养分, 其根系会增加在土壤养分富集区域的分布比例, 进而提高根系对养分的获取和利用能力(Hodge, 2004 ; Mommer et al ., 2012 ).近年来, 随着合理高效的供肥管理措施(局部施肥)在农业中广泛应用及其高效的生产力, 使其在林业上尤其是人工林经营管理中也开始得到广泛应用.因此, 了解肥料集约经营管理下的林木根系生长、形态分布以及养分吸收状况, 将有利于解释林木在生长上存在的差异, 同时为提高速生丰产林生产力提供更为完善和高效的技术指导. ...
Integrating resource heterogeneity and plant plasticity: Modelling nitrate and phosphate uptake in a patchy soil environment.
1
1996
... 氮和磷是植物生长最重要的两种营养元素, 其有效性和空间分布对植物的生长和根系发育具有重要影响(Hodge, 2004 ; Vance et al ., 2003 ).研究揭示, 局部供氮或供磷可显著促进大麦(Hordeum vulgare )、玉米(Zea mays )和小麦(Triticum aestivum )等植物侧根数量的增加以及总根长的提高(Drew, 1975 ; Officer et al ., 2009 ).Zhang和Forde (2000)将拟南芥(Arabidopsis thaliana )分别放在局部高氮和均匀高氮的琼脂培养基上培养时发现, 根系会在富氮斑块中增生, 而均匀供氮处理中的根系总量降低.可见根系在富养斑块中的增生可促进植物对养分供应的有效利用(Linkohr et al ., 2002 ).更重要的是, 相比于单独局部供氮或供磷, 在斑块中同时添加氮和磷更能促进根系在富养斑块中增生, 提高营养元素的吸收效率, 进而加快整株的生长(Jing et al ., 2010 ).这表明植物根系在局部供养斑块中的增生受到营养元素组成以及元素之间互作效应的共同影响.在木本植物研究中也发现类似的变化规律, 但物种间差异较大(Hodge, 2004 ).Einsmann等(1999)发现, 10种木本植物中有4种植物的生物量在养分异质性的供应条件下有显著提高, 但在对同种植物的不同研究中, Bliss等(2002)并没有发现这一趋势.虽然有研究认为养分斑块的存在能够对植物地上部分的生长产生积极的促进效应(Jackson & Caldwell, 1996 ), 但多年生植物绒毛草(Holcus lanatus )仅在短期内出现大量根系增生和促进地上部分生长的现象, 长期来看这种优势很快会消失, 甘松茅(Nardus stricta )则对养分斑块反应不敏感(Fransen & de Kroon, 2001 ).显然, 养分斑块或者局部养分供应对植物生长的影响受很多因素的影响, 除了养分的属性外, 植物对养分的敏感性及试验中植物生长时间的长短对结果都会产生较大影响(李洪波等, 2013 ). ...
Localized application of phosphorus and ammonium improves growth of maize seedlings by stimulating root proliferation and rhizosphere acidification.
3
2010
... 氮和磷是植物生长最重要的两种营养元素, 其有效性和空间分布对植物的生长和根系发育具有重要影响(Hodge, 2004 ; Vance et al ., 2003 ).研究揭示, 局部供氮或供磷可显著促进大麦(Hordeum vulgare )、玉米(Zea mays )和小麦(Triticum aestivum )等植物侧根数量的增加以及总根长的提高(Drew, 1975 ; Officer et al ., 2009 ).Zhang和Forde (2000)将拟南芥(Arabidopsis thaliana )分别放在局部高氮和均匀高氮的琼脂培养基上培养时发现, 根系会在富氮斑块中增生, 而均匀供氮处理中的根系总量降低.可见根系在富养斑块中的增生可促进植物对养分供应的有效利用(Linkohr et al ., 2002 ).更重要的是, 相比于单独局部供氮或供磷, 在斑块中同时添加氮和磷更能促进根系在富养斑块中增生, 提高营养元素的吸收效率, 进而加快整株的生长(Jing et al ., 2010 ).这表明植物根系在局部供养斑块中的增生受到营养元素组成以及元素之间互作效应的共同影响.在木本植物研究中也发现类似的变化规律, 但物种间差异较大(Hodge, 2004 ).Einsmann等(1999)发现, 10种木本植物中有4种植物的生物量在养分异质性的供应条件下有显著提高, 但在对同种植物的不同研究中, Bliss等(2002)并没有发现这一趋势.虽然有研究认为养分斑块的存在能够对植物地上部分的生长产生积极的促进效应(Jackson & Caldwell, 1996 ), 但多年生植物绒毛草(Holcus lanatus )仅在短期内出现大量根系增生和促进地上部分生长的现象, 长期来看这种优势很快会消失, 甘松茅(Nardus stricta )则对养分斑块反应不敏感(Fransen & de Kroon, 2001 ).显然, 养分斑块或者局部养分供应对植物生长的影响受很多因素的影响, 除了养分的属性外, 植物对养分的敏感性及试验中植物生长时间的长短对结果都会产生较大影响(李洪波等, 2013 ). ...
... 局部供氮对植物根系形态和分布的影响较大(Li et al ., 2014 ).研究结果表明, 在两种低磷环境下, 与HHH相比, LLH/L对马尾松苗木根系的生长发育有明显的促进作用, 尤其在异质低磷土壤环境下的促进作用更为突出, 其次是LLH, HHL反而不利于根系的生长发育.分析认为, 底层半侧供氮有利于根系增生发育的原因可能在于: 植物根系感知一侧氮信号的缺乏, 诱导了根原基和初级根的形成, 而富氮斑块中充足的氮供应则加速了根系的伸长(He et al ., 2003 ; Li et al ., 2014 ), 进而促进了植物整体根长和根表面积的增加.如在异质低磷下, LLH/L处理中的总根长是LLH处理的1.30倍.相似的规律在水稻感知局部磷胁迫的研究中也被发现(He et al ., 2003 ).另一方面, HHL对根长和根表面积有一定的抑制效应, 推测这可能是由于: 首先, 马尾松本身属于深根型植物, 根的向地性促使其向土壤深处觅取更多的养分, 而HHL处理中的底层并没有充足的氮素供应, 限制了根系的生长发育.这一结果与前人对禾本科植物的研究结果有所差异, 如小麦、玉米和水稻(Oryza sativa )等植物可通过增加根系在土壤表层富养区域的分布来响应自然界土壤中的异质养分环境(Weligama et al ., 2008 ; Jing et al ., 2010 ; Li et al ., 2014 ).马尾松作为木本植物, 其对土壤养分异质性分布的响应可能因树种和土壤养分组成的不同而差异较大(Einsmann et al ., 1999 ; Guo et al ., 2002 ; 梅莉等, 2006 ; 杨青等, 2011 ); 其次, 该实验的种子播种于土壤表层, 其生长初期的根系增生和扩张及时获得了营养的补充, 使得植物在养分分配时, 将更多的营养元素输送到了其他器官(茎、叶), 降低了对根系的养分投入, 限制了根系的增生发育(段洪浪等, 2009 ; 吴茜等, 2011 ).结合苗期马尾松的株高、地径等地上部分的生长状况也印证了这一推测. ...
... 不同物种或不同基因型植物的氮素利用特性不同, 对不同氮供应方式的适应能力也存在差异.研究结果表明, 同质低磷下, 不同马尾松不同家系地上部分生长对局部供氮的响应差异较大, 而在异质低磷下, 地下部分的根系差异则更为明显.推测认为, 根系形态对土壤磷素异质分布的可塑性较强, 而对局部氮浓度的感知则较为迟钝(Tinker & Nye, 2000 ), 异质低磷环境中的磷素异质性增强了局部氮信号对马尾松苗木根系发育的影响, 进而产生了叠加效应(Jing et al ., 2010 ).在两种低磷环境下, 参试的5个马尾松不同家系中, 除了1号家系对氮添加不敏感外, 其他家系对局部供氮的响应因磷素环境和氮供应方式差异明显.如家系25的干物质积累量与植株氮、磷含量的变化有协同效应, 但与根系生长发育的变化相反, 这表明氮、磷元素在该家系内的快速转运以及利用可能对其整株生长贡献较大 (Kanno et al ., 2016 ).在异质低磷下, HHL限制了家系49、52和57的根系发育, 整株生长受到抑制; 而LLH和LLH/L促进了根系生长和干物质量的积累.这意味着在异质低磷环境下, 家系49、52和57主要通过改变根系的生长发育来响应局部氮素的变化.许多研究证实, 植物对土壤局部营养环境的感知及响应能力受营养元素分布、营养元素组成以及植株自身生长的环境影响(Robinson, 1994 ; Hodge, 2004 ).1号家系对局部供氮的响应较迟钝甚至有抑制效应, 分析认为, 这可能是因为家系1本身的基因型决定了其生长缓慢, 使得外界氮素的变化对植株根系发育的影响较小.本实验中局部养分供应对植物生长的促进效应可获得更多可利用的优良家系, 达到速生增产的目的, 但土壤中的养分异质性不仅是空间分布上的异质性, 也包括时间尺度上的异质性, 在长期的养分异质性环境下, 林木是否依然会保持旺盛的生长, 仍需更多的实验研究和长期观察, 以进一步客观地揭示异质养分土壤环境下, 局部氮素和磷素供应对马尾松不同家系苗木生长发育的影响. ...
A novel role for the root cap in phosphate uptake and homeostasis.
1
2016
... 不同物种或不同基因型植物的氮素利用特性不同, 对不同氮供应方式的适应能力也存在差异.研究结果表明, 同质低磷下, 不同马尾松不同家系地上部分生长对局部供氮的响应差异较大, 而在异质低磷下, 地下部分的根系差异则更为明显.推测认为, 根系形态对土壤磷素异质分布的可塑性较强, 而对局部氮浓度的感知则较为迟钝(Tinker & Nye, 2000 ), 异质低磷环境中的磷素异质性增强了局部氮信号对马尾松苗木根系发育的影响, 进而产生了叠加效应(Jing et al ., 2010 ).在两种低磷环境下, 参试的5个马尾松不同家系中, 除了1号家系对氮添加不敏感外, 其他家系对局部供氮的响应因磷素环境和氮供应方式差异明显.如家系25的干物质积累量与植株氮、磷含量的变化有协同效应, 但与根系生长发育的变化相反, 这表明氮、磷元素在该家系内的快速转运以及利用可能对其整株生长贡献较大 (Kanno et al ., 2016 ).在异质低磷下, HHL限制了家系49、52和57的根系发育, 整株生长受到抑制; 而LLH和LLH/L促进了根系生长和干物质量的积累.这意味着在异质低磷环境下, 家系49、52和57主要通过改变根系的生长发育来响应局部氮素的变化.许多研究证实, 植物对土壤局部营养环境的感知及响应能力受营养元素分布、营养元素组成以及植株自身生长的环境影响(Robinson, 1994 ; Hodge, 2004 ).1号家系对局部供氮的响应较迟钝甚至有抑制效应, 分析认为, 这可能是因为家系1本身的基因型决定了其生长缓慢, 使得外界氮素的变化对植株根系发育的影响较小.本实验中局部养分供应对植物生长的促进效应可获得更多可利用的优良家系, 达到速生增产的目的, 但土壤中的养分异质性不仅是空间分布上的异质性, 也包括时间尺度上的异质性, 在长期的养分异质性环境下, 林木是否依然会保持旺盛的生长, 仍需更多的实验研究和长期观察, 以进一步客观地揭示异质养分土壤环境下, 局部氮素和磷素供应对马尾松不同家系苗木生长发育的影响. ...
Ammonium toxicity and the real cost of transport.
1
2001
... 近年来, 土壤中养分元素的分布方式对植株生长的影响越来越受到关注(Hodge, 2004 ; 吕超群等, 2007 ; 徐星凯等, 2012 ).本研究中的异质低磷环境较同质低磷更有利于马尾松苗木的生长, 与前人的研究结果(杨青等, 2011 ; 庞丽等, 2014 )一致.已有研究表明, 外源氮素添加(100 g·kg-1 )会使土壤有效氮含量增加, 改变植株生物量的分配进而促进整株的生长(段洪浪等, 2009 ; 庞丽等, 2014 ; 刘双娥等, 2015 ).在本实验中, 局部供氮处理的斑块氮浓度与均匀供氮处理的氮含量相同, 但总施肥量要少于均匀供氮(相当于HHH处理施氮量的1/6-2/3).研究结果显示, 不同的供氮方式对马尾松苗木生长的影响差异较大, 且受到土壤磷素环境影响.在同质低磷下, HHL显著促进了马尾松苗木的生长, 根冠比降低, 较其他供氮方式更有利于苗木地上部分的生长; 其次, LLH和LLH/L两种方式之间, LLH/L对马尾松整株的生长相对更有利, 这表明在同质低磷下, 局部供氮增强了土壤中养分的异质性, 促进了马尾松苗木整株的生长.类似的结果在桉树(Eucalyptus grandis )、油松(Pinus tabulaeformis )、火炬松(Pinus taeda )等植物中也被发现(Mou et al ., 1997 ; Zhou & Shangguan, 2007 ; Graciano et al ., 2009 ).然而, 值得注意的是, Fransen和de Kroon (2001)在研究多年生植物绒毛草和甘松茅时, 发现绒毛草仅在短期内能通过根系增生来获取斑块内大量养分并促进地上部分生长, 从长期生长来看这种优势则会消失.因此, 本文的实验结果对生产实践中的苗木培育具有一定的指导意义, 但是否在长期生产造林和人工林抚育中仍有指导性需要进一步研究证实.与同质低磷相反, 在异质低磷下, 与HHH相比, HHL不利于马尾松株高、地径和干物质量的提高, 这可能是由于异质低磷土壤中的表层磷含量较高, 导致土壤(酸性土壤) pH值偏高(Shen et al ., 2005 ), 植物在局部供氮后更倾向于对NH4+ -N的吸收, 而过多吸收NH4+ -N会对植物产生一定的毒害作用(Kronzucker et al ., 2001 ), 抑制其整株的生长.然而在有效磷较低的底层土壤中, LLH和LLH/L刺激后, 植株吸收的磷含量显著提高, 且整株干物质量达到均匀供氮处理水平.这表明底层(局部)供氮可促进马尾松苗木对土壤中磷素的有效吸收利用, 保证其快速生长的情况下减少对化学肥料的依赖. ...
Root morphological responses to localized nutrient supply differ among crop species with contrasting root traits.
3
2014
... 局部供氮对植物根系形态和分布的影响较大(Li et al ., 2014 ).研究结果表明, 在两种低磷环境下, 与HHH相比, LLH/L对马尾松苗木根系的生长发育有明显的促进作用, 尤其在异质低磷土壤环境下的促进作用更为突出, 其次是LLH, HHL反而不利于根系的生长发育.分析认为, 底层半侧供氮有利于根系增生发育的原因可能在于: 植物根系感知一侧氮信号的缺乏, 诱导了根原基和初级根的形成, 而富氮斑块中充足的氮供应则加速了根系的伸长(He et al ., 2003 ; Li et al ., 2014 ), 进而促进了植物整体根长和根表面积的增加.如在异质低磷下, LLH/L处理中的总根长是LLH处理的1.30倍.相似的规律在水稻感知局部磷胁迫的研究中也被发现(He et al ., 2003 ).另一方面, HHL对根长和根表面积有一定的抑制效应, 推测这可能是由于: 首先, 马尾松本身属于深根型植物, 根的向地性促使其向土壤深处觅取更多的养分, 而HHL处理中的底层并没有充足的氮素供应, 限制了根系的生长发育.这一结果与前人对禾本科植物的研究结果有所差异, 如小麦、玉米和水稻(Oryza sativa )等植物可通过增加根系在土壤表层富养区域的分布来响应自然界土壤中的异质养分环境(Weligama et al ., 2008 ; Jing et al ., 2010 ; Li et al ., 2014 ).马尾松作为木本植物, 其对土壤养分异质性分布的响应可能因树种和土壤养分组成的不同而差异较大(Einsmann et al ., 1999 ; Guo et al ., 2002 ; 梅莉等, 2006 ; 杨青等, 2011 ); 其次, 该实验的种子播种于土壤表层, 其生长初期的根系增生和扩张及时获得了营养的补充, 使得植物在养分分配时, 将更多的营养元素输送到了其他器官(茎、叶), 降低了对根系的养分投入, 限制了根系的增生发育(段洪浪等, 2009 ; 吴茜等, 2011 ).结合苗期马尾松的株高、地径等地上部分的生长状况也印证了这一推测. ...
... ; Li et al ., 2014 ), 进而促进了植物整体根长和根表面积的增加.如在异质低磷下, LLH/L处理中的总根长是LLH处理的1.30倍.相似的规律在水稻感知局部磷胁迫的研究中也被发现(He et al ., 2003 ).另一方面, HHL对根长和根表面积有一定的抑制效应, 推测这可能是由于: 首先, 马尾松本身属于深根型植物, 根的向地性促使其向土壤深处觅取更多的养分, 而HHL处理中的底层并没有充足的氮素供应, 限制了根系的生长发育.这一结果与前人对禾本科植物的研究结果有所差异, 如小麦、玉米和水稻(Oryza sativa )等植物可通过增加根系在土壤表层富养区域的分布来响应自然界土壤中的异质养分环境(Weligama et al ., 2008 ; Jing et al ., 2010 ; Li et al ., 2014 ).马尾松作为木本植物, 其对土壤养分异质性分布的响应可能因树种和土壤养分组成的不同而差异较大(Einsmann et al ., 1999 ; Guo et al ., 2002 ; 梅莉等, 2006 ; 杨青等, 2011 ); 其次, 该实验的种子播种于土壤表层, 其生长初期的根系增生和扩张及时获得了营养的补充, 使得植物在养分分配时, 将更多的营养元素输送到了其他器官(茎、叶), 降低了对根系的养分投入, 限制了根系的增生发育(段洪浪等, 2009 ; 吴茜等, 2011 ).结合苗期马尾松的株高、地径等地上部分的生长状况也印证了这一推测. ...
... ; Li et al ., 2014 ).马尾松作为木本植物, 其对土壤养分异质性分布的响应可能因树种和土壤养分组成的不同而差异较大(Einsmann et al ., 1999 ; Guo et al ., 2002 ; 梅莉等, 2006 ; 杨青等, 2011 ); 其次, 该实验的种子播种于土壤表层, 其生长初期的根系增生和扩张及时获得了营养的补充, 使得植物在养分分配时, 将更多的营养元素输送到了其他器官(茎、叶), 降低了对根系的养分投入, 限制了根系的增生发育(段洪浪等, 2009 ; 吴茜等, 2011 ).结合苗期马尾松的株高、地径等地上部分的生长状况也印证了这一推测. ...
土壤养分空间异质性与根系觅食作用: 从个体到群落
1
2013
... 氮和磷是植物生长最重要的两种营养元素, 其有效性和空间分布对植物的生长和根系发育具有重要影响(Hodge, 2004 ; Vance et al ., 2003 ).研究揭示, 局部供氮或供磷可显著促进大麦(Hordeum vulgare )、玉米(Zea mays )和小麦(Triticum aestivum )等植物侧根数量的增加以及总根长的提高(Drew, 1975 ; Officer et al ., 2009 ).Zhang和Forde (2000)将拟南芥(Arabidopsis thaliana )分别放在局部高氮和均匀高氮的琼脂培养基上培养时发现, 根系会在富氮斑块中增生, 而均匀供氮处理中的根系总量降低.可见根系在富养斑块中的增生可促进植物对养分供应的有效利用(Linkohr et al ., 2002 ).更重要的是, 相比于单独局部供氮或供磷, 在斑块中同时添加氮和磷更能促进根系在富养斑块中增生, 提高营养元素的吸收效率, 进而加快整株的生长(Jing et al ., 2010 ).这表明植物根系在局部供养斑块中的增生受到营养元素组成以及元素之间互作效应的共同影响.在木本植物研究中也发现类似的变化规律, 但物种间差异较大(Hodge, 2004 ).Einsmann等(1999)发现, 10种木本植物中有4种植物的生物量在养分异质性的供应条件下有显著提高, 但在对同种植物的不同研究中, Bliss等(2002)并没有发现这一趋势.虽然有研究认为养分斑块的存在能够对植物地上部分的生长产生积极的促进效应(Jackson & Caldwell, 1996 ), 但多年生植物绒毛草(Holcus lanatus )仅在短期内出现大量根系增生和促进地上部分生长的现象, 长期来看这种优势很快会消失, 甘松茅(Nardus stricta )则对养分斑块反应不敏感(Fransen & de Kroon, 2001 ).显然, 养分斑块或者局部养分供应对植物生长的影响受很多因素的影响, 除了养分的属性外, 植物对养分的敏感性及试验中植物生长时间的长短对结果都会产生较大影响(李洪波等, 2013 ). ...
Nitrate and phosphate availability and distribution have different effects on root system architecture of
1
2002
... 氮和磷是植物生长最重要的两种营养元素, 其有效性和空间分布对植物的生长和根系发育具有重要影响(Hodge, 2004 ; Vance et al ., 2003 ).研究揭示, 局部供氮或供磷可显著促进大麦(Hordeum vulgare )、玉米(Zea mays )和小麦(Triticum aestivum )等植物侧根数量的增加以及总根长的提高(Drew, 1975 ; Officer et al ., 2009 ).Zhang和Forde (2000)将拟南芥(Arabidopsis thaliana )分别放在局部高氮和均匀高氮的琼脂培养基上培养时发现, 根系会在富氮斑块中增生, 而均匀供氮处理中的根系总量降低.可见根系在富养斑块中的增生可促进植物对养分供应的有效利用(Linkohr et al ., 2002 ).更重要的是, 相比于单独局部供氮或供磷, 在斑块中同时添加氮和磷更能促进根系在富养斑块中增生, 提高营养元素的吸收效率, 进而加快整株的生长(Jing et al ., 2010 ).这表明植物根系在局部供养斑块中的增生受到营养元素组成以及元素之间互作效应的共同影响.在木本植物研究中也发现类似的变化规律, 但物种间差异较大(Hodge, 2004 ).Einsmann等(1999)发现, 10种木本植物中有4种植物的生物量在养分异质性的供应条件下有显著提高, 但在对同种植物的不同研究中, Bliss等(2002)并没有发现这一趋势.虽然有研究认为养分斑块的存在能够对植物地上部分的生长产生积极的促进效应(Jackson & Caldwell, 1996 ), 但多年生植物绒毛草(Holcus lanatus )仅在短期内出现大量根系增生和促进地上部分生长的现象, 长期来看这种优势很快会消失, 甘松茅(Nardus stricta )则对养分斑块反应不敏感(Fransen & de Kroon, 2001 ).显然, 养分斑块或者局部养分供应对植物生长的影响受很多因素的影响, 除了养分的属性外, 植物对养分的敏感性及试验中植物生长时间的长短对结果都会产生较大影响(李洪波等, 2013 ). ...
不同氮添加量和添加方式对南亚热带4个主要树种幼苗生长的影响
2
2015
... 土壤养分在空间分布上多呈时空异质性或者斑块分布(Hutching & John, 2004 ).产生土壤养分空间异质性的原因很多, 其中包括有机物(凋落物、根系死亡以及动物尸体)的分解、无机养分在土壤中的扩散系数差异以及微地形因素影响等(Hodge, 2004 ; 徐星凯等, 2012 ; 刘双娥等, 2015 ).植物为了获得更多的养分, 其根系会增加在土壤养分富集区域的分布比例, 进而提高根系对养分的获取和利用能力(Hodge, 2004 ; Mommer et al ., 2012 ).近年来, 随着合理高效的供肥管理措施(局部施肥)在农业中广泛应用及其高效的生产力, 使其在林业上尤其是人工林经营管理中也开始得到广泛应用.因此, 了解肥料集约经营管理下的林木根系生长、形态分布以及养分吸收状况, 将有利于解释林木在生长上存在的差异, 同时为提高速生丰产林生产力提供更为完善和高效的技术指导. ...
... 近年来, 土壤中养分元素的分布方式对植株生长的影响越来越受到关注(Hodge, 2004 ; 吕超群等, 2007 ; 徐星凯等, 2012 ).本研究中的异质低磷环境较同质低磷更有利于马尾松苗木的生长, 与前人的研究结果(杨青等, 2011 ; 庞丽等, 2014 )一致.已有研究表明, 外源氮素添加(100 g·kg-1 )会使土壤有效氮含量增加, 改变植株生物量的分配进而促进整株的生长(段洪浪等, 2009 ; 庞丽等, 2014 ; 刘双娥等, 2015 ).在本实验中, 局部供氮处理的斑块氮浓度与均匀供氮处理的氮含量相同, 但总施肥量要少于均匀供氮(相当于HHH处理施氮量的1/6-2/3).研究结果显示, 不同的供氮方式对马尾松苗木生长的影响差异较大, 且受到土壤磷素环境影响.在同质低磷下, HHL显著促进了马尾松苗木的生长, 根冠比降低, 较其他供氮方式更有利于苗木地上部分的生长; 其次, LLH和LLH/L两种方式之间, LLH/L对马尾松整株的生长相对更有利, 这表明在同质低磷下, 局部供氮增强了土壤中养分的异质性, 促进了马尾松苗木整株的生长.类似的结果在桉树(Eucalyptus grandis )、油松(Pinus tabulaeformis )、火炬松(Pinus taeda )等植物中也被发现(Mou et al ., 1997 ; Zhou & Shangguan, 2007 ; Graciano et al ., 2009 ).然而, 值得注意的是, Fransen和de Kroon (2001)在研究多年生植物绒毛草和甘松茅时, 发现绒毛草仅在短期内能通过根系增生来获取斑块内大量养分并促进地上部分生长, 从长期生长来看这种优势则会消失.因此, 本文的实验结果对生产实践中的苗木培育具有一定的指导意义, 但是否在长期生产造林和人工林抚育中仍有指导性需要进一步研究证实.与同质低磷相反, 在异质低磷下, 与HHH相比, HHL不利于马尾松株高、地径和干物质量的提高, 这可能是由于异质低磷土壤中的表层磷含量较高, 导致土壤(酸性土壤) pH值偏高(Shen et al ., 2005 ), 植物在局部供氮后更倾向于对NH4+ -N的吸收, 而过多吸收NH4+ -N会对植物产生一定的毒害作用(Kronzucker et al ., 2001 ), 抑制其整株的生长.然而在有效磷较低的底层土壤中, LLH和LLH/L刺激后, 植株吸收的磷含量显著提高, 且整株干物质量达到均匀供氮处理水平.这表明底层(局部)供氮可促进马尾松苗木对土壤中磷素的有效吸收利用, 保证其快速生长的情况下减少对化学肥料的依赖. ...
陆地生态系统氮沉降增加的生态效应
1
2007
... 近年来, 土壤中养分元素的分布方式对植株生长的影响越来越受到关注(Hodge, 2004 ; 吕超群等, 2007 ; 徐星凯等, 2012 ).本研究中的异质低磷环境较同质低磷更有利于马尾松苗木的生长, 与前人的研究结果(杨青等, 2011 ; 庞丽等, 2014 )一致.已有研究表明, 外源氮素添加(100 g·kg-1 )会使土壤有效氮含量增加, 改变植株生物量的分配进而促进整株的生长(段洪浪等, 2009 ; 庞丽等, 2014 ; 刘双娥等, 2015 ).在本实验中, 局部供氮处理的斑块氮浓度与均匀供氮处理的氮含量相同, 但总施肥量要少于均匀供氮(相当于HHH处理施氮量的1/6-2/3).研究结果显示, 不同的供氮方式对马尾松苗木生长的影响差异较大, 且受到土壤磷素环境影响.在同质低磷下, HHL显著促进了马尾松苗木的生长, 根冠比降低, 较其他供氮方式更有利于苗木地上部分的生长; 其次, LLH和LLH/L两种方式之间, LLH/L对马尾松整株的生长相对更有利, 这表明在同质低磷下, 局部供氮增强了土壤中养分的异质性, 促进了马尾松苗木整株的生长.类似的结果在桉树(Eucalyptus grandis )、油松(Pinus tabulaeformis )、火炬松(Pinus taeda )等植物中也被发现(Mou et al ., 1997 ; Zhou & Shangguan, 2007 ; Graciano et al ., 2009 ).然而, 值得注意的是, Fransen和de Kroon (2001)在研究多年生植物绒毛草和甘松茅时, 发现绒毛草仅在短期内能通过根系增生来获取斑块内大量养分并促进地上部分生长, 从长期生长来看这种优势则会消失.因此, 本文的实验结果对生产实践中的苗木培育具有一定的指导意义, 但是否在长期生产造林和人工林抚育中仍有指导性需要进一步研究证实.与同质低磷相反, 在异质低磷下, 与HHH相比, HHL不利于马尾松株高、地径和干物质量的提高, 这可能是由于异质低磷土壤中的表层磷含量较高, 导致土壤(酸性土壤) pH值偏高(Shen et al ., 2005 ), 植物在局部供氮后更倾向于对NH4+ -N的吸收, 而过多吸收NH4+ -N会对植物产生一定的毒害作用(Kronzucker et al ., 2001 ), 抑制其整株的生长.然而在有效磷较低的底层土壤中, LLH和LLH/L刺激后, 植株吸收的磷含量显著提高, 且整株干物质量达到均匀供氮处理水平.这表明底层(局部)供氮可促进马尾松苗木对土壤中磷素的有效吸收利用, 保证其快速生长的情况下减少对化学肥料的依赖. ...
水曲柳根系生物量、比根长和根长密度的分布格局
1
2006
... 局部供氮对植物根系形态和分布的影响较大(Li et al ., 2014 ).研究结果表明, 在两种低磷环境下, 与HHH相比, LLH/L对马尾松苗木根系的生长发育有明显的促进作用, 尤其在异质低磷土壤环境下的促进作用更为突出, 其次是LLH, HHL反而不利于根系的生长发育.分析认为, 底层半侧供氮有利于根系增生发育的原因可能在于: 植物根系感知一侧氮信号的缺乏, 诱导了根原基和初级根的形成, 而富氮斑块中充足的氮供应则加速了根系的伸长(He et al ., 2003 ; Li et al ., 2014 ), 进而促进了植物整体根长和根表面积的增加.如在异质低磷下, LLH/L处理中的总根长是LLH处理的1.30倍.相似的规律在水稻感知局部磷胁迫的研究中也被发现(He et al ., 2003 ).另一方面, HHL对根长和根表面积有一定的抑制效应, 推测这可能是由于: 首先, 马尾松本身属于深根型植物, 根的向地性促使其向土壤深处觅取更多的养分, 而HHL处理中的底层并没有充足的氮素供应, 限制了根系的生长发育.这一结果与前人对禾本科植物的研究结果有所差异, 如小麦、玉米和水稻(Oryza sativa )等植物可通过增加根系在土壤表层富养区域的分布来响应自然界土壤中的异质养分环境(Weligama et al ., 2008 ; Jing et al ., 2010 ; Li et al ., 2014 ).马尾松作为木本植物, 其对土壤养分异质性分布的响应可能因树种和土壤养分组成的不同而差异较大(Einsmann et al ., 1999 ; Guo et al ., 2002 ; 梅莉等, 2006 ; 杨青等, 2011 ); 其次, 该实验的种子播种于土壤表层, 其生长初期的根系增生和扩张及时获得了营养的补充, 使得植物在养分分配时, 将更多的营养元素输送到了其他器官(茎、叶), 降低了对根系的养分投入, 限制了根系的增生发育(段洪浪等, 2009 ; 吴茜等, 2011 ).结合苗期马尾松的株高、地径等地上部分的生长状况也印证了这一推测. ...
Interactive effects of nutrient heterogeneity and competition: Implications for root foraging theory?
1
2012
... 土壤养分在空间分布上多呈时空异质性或者斑块分布(Hutching & John, 2004 ).产生土壤养分空间异质性的原因很多, 其中包括有机物(凋落物、根系死亡以及动物尸体)的分解、无机养分在土壤中的扩散系数差异以及微地形因素影响等(Hodge, 2004 ; 徐星凯等, 2012 ; 刘双娥等, 2015 ).植物为了获得更多的养分, 其根系会增加在土壤养分富集区域的分布比例, 进而提高根系对养分的获取和利用能力(Hodge, 2004 ; Mommer et al ., 2012 ).近年来, 随着合理高效的供肥管理措施(局部施肥)在农业中广泛应用及其高效的生产力, 使其在林业上尤其是人工林经营管理中也开始得到广泛应用.因此, 了解肥料集约经营管理下的林木根系生长、形态分布以及养分吸收状况, 将有利于解释林木在生长上存在的差异, 同时为提高速生丰产林生产力提供更为完善和高效的技术指导. ...
Root distribution of two tree species under a heterogeneous nutrient environment.
1
1997
... 近年来, 土壤中养分元素的分布方式对植株生长的影响越来越受到关注(Hodge, 2004 ; 吕超群等, 2007 ; 徐星凯等, 2012 ).本研究中的异质低磷环境较同质低磷更有利于马尾松苗木的生长, 与前人的研究结果(杨青等, 2011 ; 庞丽等, 2014 )一致.已有研究表明, 外源氮素添加(100 g·kg-1 )会使土壤有效氮含量增加, 改变植株生物量的分配进而促进整株的生长(段洪浪等, 2009 ; 庞丽等, 2014 ; 刘双娥等, 2015 ).在本实验中, 局部供氮处理的斑块氮浓度与均匀供氮处理的氮含量相同, 但总施肥量要少于均匀供氮(相当于HHH处理施氮量的1/6-2/3).研究结果显示, 不同的供氮方式对马尾松苗木生长的影响差异较大, 且受到土壤磷素环境影响.在同质低磷下, HHL显著促进了马尾松苗木的生长, 根冠比降低, 较其他供氮方式更有利于苗木地上部分的生长; 其次, LLH和LLH/L两种方式之间, LLH/L对马尾松整株的生长相对更有利, 这表明在同质低磷下, 局部供氮增强了土壤中养分的异质性, 促进了马尾松苗木整株的生长.类似的结果在桉树(Eucalyptus grandis )、油松(Pinus tabulaeformis )、火炬松(Pinus taeda )等植物中也被发现(Mou et al ., 1997 ; Zhou & Shangguan, 2007 ; Graciano et al ., 2009 ).然而, 值得注意的是, Fransen和de Kroon (2001)在研究多年生植物绒毛草和甘松茅时, 发现绒毛草仅在短期内能通过根系增生来获取斑块内大量养分并促进地上部分生长, 从长期生长来看这种优势则会消失.因此, 本文的实验结果对生产实践中的苗木培育具有一定的指导意义, 但是否在长期生产造林和人工林抚育中仍有指导性需要进一步研究证实.与同质低磷相反, 在异质低磷下, 与HHH相比, HHL不利于马尾松株高、地径和干物质量的提高, 这可能是由于异质低磷土壤中的表层磷含量较高, 导致土壤(酸性土壤) pH值偏高(Shen et al ., 2005 ), 植物在局部供氮后更倾向于对NH4+ -N的吸收, 而过多吸收NH4+ -N会对植物产生一定的毒害作用(Kronzucker et al ., 2001 ), 抑制其整株的生长.然而在有效磷较低的底层土壤中, LLH和LLH/L刺激后, 植株吸收的磷含量显著提高, 且整株干物质量达到均匀供氮处理水平.这表明底层(局部)供氮可促进马尾松苗木对土壤中磷素的有效吸收利用, 保证其快速生长的情况下减少对化学肥料的依赖. ...
Wheat roots proliferate in response to nitrogen and phosphorus fertilisers in Sodosol and Vertosol soils of south-eastern Australia.
1
2009
... 氮和磷是植物生长最重要的两种营养元素, 其有效性和空间分布对植物的生长和根系发育具有重要影响(Hodge, 2004 ; Vance et al ., 2003 ).研究揭示, 局部供氮或供磷可显著促进大麦(Hordeum vulgare )、玉米(Zea mays )和小麦(Triticum aestivum )等植物侧根数量的增加以及总根长的提高(Drew, 1975 ; Officer et al ., 2009 ).Zhang和Forde (2000)将拟南芥(Arabidopsis thaliana )分别放在局部高氮和均匀高氮的琼脂培养基上培养时发现, 根系会在富氮斑块中增生, 而均匀供氮处理中的根系总量降低.可见根系在富养斑块中的增生可促进植物对养分供应的有效利用(Linkohr et al ., 2002 ).更重要的是, 相比于单独局部供氮或供磷, 在斑块中同时添加氮和磷更能促进根系在富养斑块中增生, 提高营养元素的吸收效率, 进而加快整株的生长(Jing et al ., 2010 ).这表明植物根系在局部供养斑块中的增生受到营养元素组成以及元素之间互作效应的共同影响.在木本植物研究中也发现类似的变化规律, 但物种间差异较大(Hodge, 2004 ).Einsmann等(1999)发现, 10种木本植物中有4种植物的生物量在养分异质性的供应条件下有显著提高, 但在对同种植物的不同研究中, Bliss等(2002)并没有发现这一趋势.虽然有研究认为养分斑块的存在能够对植物地上部分的生长产生积极的促进效应(Jackson & Caldwell, 1996 ), 但多年生植物绒毛草(Holcus lanatus )仅在短期内出现大量根系增生和促进地上部分生长的现象, 长期来看这种优势很快会消失, 甘松茅(Nardus stricta )则对养分斑块反应不敏感(Fransen & de Kroon, 2001 ).显然, 养分斑块或者局部养分供应对植物生长的影响受很多因素的影响, 除了养分的属性外, 植物对养分的敏感性及试验中植物生长时间的长短对结果都会产生较大影响(李洪波等, 2013 ). ...
模拟氮沉降对低磷胁迫下马尾松生长和磷效率的影响
4
2014
... 马尾松(Pinus massoniana )是重要的材用、脂用和生态造林树种, 广泛分布于我国的亚热带地区(秦国峰和周志春, 2012 ).当前研究已揭示了氮添加对低磷胁迫下马尾松生长影响的生物学机制, 包括地上部分生长、根系参数和氮、磷效率的适应性变化等(Zhang et al ., 2013 ; 庞丽等, 2014 ). 然而, 相关研究未考虑氮素本身在空间分布上的不均一性.鉴于此, 本文拟通过人工控制氮素在土壤空间上的异质性, 研究低磷胁迫下马尾松不同家系在局部供氮条件下的生长表现和根系发育, 进一步阐明马尾松不同家系对土壤异质养分的响应差异及机制, 为马尾松营养高效新品种选育和苗木高效栽培方式提供理论依据以及实践指导. ...
... 土壤垂直方向上的氮素异质性设置及保持, 采用树脂膜控释氮肥(含氮量44%, 山东金正大公司生产)来调控.两种低磷环境下同时布设相同局部供氮处理, 氮素施入量参照庞丽等(2014) 研究方法.处理1 (HHH): 表层、中间层和底层均匀供氮, 其有效氮含量均为100 g·kg-1 ; 处理2 (HHL): 表层和中间层(0-16 cm)供氮, 有效氮含量均为100 g·kg-1 , 底层不添加氮; 处理3 (LLH): 底层(16-24 cm)供氮, 有效氮含量为100 g·kg-1 , 其他层次均不添加; 处理 4 (LLH/L): 底层一侧局部供氮的有效氮含量为 100 g·kg-1 . ...
... 近年来, 土壤中养分元素的分布方式对植株生长的影响越来越受到关注(Hodge, 2004 ; 吕超群等, 2007 ; 徐星凯等, 2012 ).本研究中的异质低磷环境较同质低磷更有利于马尾松苗木的生长, 与前人的研究结果(杨青等, 2011 ; 庞丽等, 2014 )一致.已有研究表明, 外源氮素添加(100 g·kg-1 )会使土壤有效氮含量增加, 改变植株生物量的分配进而促进整株的生长(段洪浪等, 2009 ; 庞丽等, 2014 ; 刘双娥等, 2015 ).在本实验中, 局部供氮处理的斑块氮浓度与均匀供氮处理的氮含量相同, 但总施肥量要少于均匀供氮(相当于HHH处理施氮量的1/6-2/3).研究结果显示, 不同的供氮方式对马尾松苗木生长的影响差异较大, 且受到土壤磷素环境影响.在同质低磷下, HHL显著促进了马尾松苗木的生长, 根冠比降低, 较其他供氮方式更有利于苗木地上部分的生长; 其次, LLH和LLH/L两种方式之间, LLH/L对马尾松整株的生长相对更有利, 这表明在同质低磷下, 局部供氮增强了土壤中养分的异质性, 促进了马尾松苗木整株的生长.类似的结果在桉树(Eucalyptus grandis )、油松(Pinus tabulaeformis )、火炬松(Pinus taeda )等植物中也被发现(Mou et al ., 1997 ; Zhou & Shangguan, 2007 ; Graciano et al ., 2009 ).然而, 值得注意的是, Fransen和de Kroon (2001)在研究多年生植物绒毛草和甘松茅时, 发现绒毛草仅在短期内能通过根系增生来获取斑块内大量养分并促进地上部分生长, 从长期生长来看这种优势则会消失.因此, 本文的实验结果对生产实践中的苗木培育具有一定的指导意义, 但是否在长期生产造林和人工林抚育中仍有指导性需要进一步研究证实.与同质低磷相反, 在异质低磷下, 与HHH相比, HHL不利于马尾松株高、地径和干物质量的提高, 这可能是由于异质低磷土壤中的表层磷含量较高, 导致土壤(酸性土壤) pH值偏高(Shen et al ., 2005 ), 植物在局部供氮后更倾向于对NH4+ -N的吸收, 而过多吸收NH4+ -N会对植物产生一定的毒害作用(Kronzucker et al ., 2001 ), 抑制其整株的生长.然而在有效磷较低的底层土壤中, LLH和LLH/L刺激后, 植株吸收的磷含量显著提高, 且整株干物质量达到均匀供氮处理水平.这表明底层(局部)供氮可促进马尾松苗木对土壤中磷素的有效吸收利用, 保证其快速生长的情况下减少对化学肥料的依赖. ...
... ; 庞丽等, 2014 ; 刘双娥等, 2015 ).在本实验中, 局部供氮处理的斑块氮浓度与均匀供氮处理的氮含量相同, 但总施肥量要少于均匀供氮(相当于HHH处理施氮量的1/6-2/3).研究结果显示, 不同的供氮方式对马尾松苗木生长的影响差异较大, 且受到土壤磷素环境影响.在同质低磷下, HHL显著促进了马尾松苗木的生长, 根冠比降低, 较其他供氮方式更有利于苗木地上部分的生长; 其次, LLH和LLH/L两种方式之间, LLH/L对马尾松整株的生长相对更有利, 这表明在同质低磷下, 局部供氮增强了土壤中养分的异质性, 促进了马尾松苗木整株的生长.类似的结果在桉树(Eucalyptus grandis )、油松(Pinus tabulaeformis )、火炬松(Pinus taeda )等植物中也被发现(Mou et al ., 1997 ; Zhou & Shangguan, 2007 ; Graciano et al ., 2009 ).然而, 值得注意的是, Fransen和de Kroon (2001)在研究多年生植物绒毛草和甘松茅时, 发现绒毛草仅在短期内能通过根系增生来获取斑块内大量养分并促进地上部分生长, 从长期生长来看这种优势则会消失.因此, 本文的实验结果对生产实践中的苗木培育具有一定的指导意义, 但是否在长期生产造林和人工林抚育中仍有指导性需要进一步研究证实.与同质低磷相反, 在异质低磷下, 与HHH相比, HHL不利于马尾松株高、地径和干物质量的提高, 这可能是由于异质低磷土壤中的表层磷含量较高, 导致土壤(酸性土壤) pH值偏高(Shen et al ., 2005 ), 植物在局部供氮后更倾向于对NH4+ -N的吸收, 而过多吸收NH4+ -N会对植物产生一定的毒害作用(Kronzucker et al ., 2001 ), 抑制其整株的生长.然而在有效磷较低的底层土壤中, LLH和LLH/L刺激后, 植株吸收的磷含量显著提高, 且整株干物质量达到均匀供氮处理水平.这表明底层(局部)供氮可促进马尾松苗木对土壤中磷素的有效吸收利用, 保证其快速生长的情况下减少对化学肥料的依赖. ...
1
... 马尾松(Pinus massoniana )是重要的材用、脂用和生态造林树种, 广泛分布于我国的亚热带地区(秦国峰和周志春, 2012 ).当前研究已揭示了氮添加对低磷胁迫下马尾松生长影响的生物学机制, 包括地上部分生长、根系参数和氮、磷效率的适应性变化等(Zhang et al ., 2013 ; 庞丽等, 2014 ). 然而, 相关研究未考虑氮素本身在空间分布上的不均一性.鉴于此, 本文拟通过人工控制氮素在土壤空间上的异质性, 研究低磷胁迫下马尾松不同家系在局部供氮条件下的生长表现和根系发育, 进一步阐明马尾松不同家系对土壤异质养分的响应差异及机制, 为马尾松营养高效新品种选育和苗木高效栽培方式提供理论依据以及实践指导. ...
Rapid development of phosphorus limitation in temperate rainforest along the Franz Josef soil chronosequence.
1
2004
... 低磷土壤环境设置参照Zhang等(2013)的浓度设计.基质土壤经过风干过筛后, 加入一定量的Ca(H2 PO4 )2 混匀.异质低磷盆栽基质的底层、中间层和表层依次填入添加0、0.10和1.00 g·kg-1 的Ca(H2 PO4 )2 , 相当于有效磷含量分别为0.99、15和 30 mg·kg-1 .盆栽基质由上而下有效磷含量逐渐降低的异质养分环境可模拟立地条件较好, 表层营养丰富的针阔混交林或阔叶林土壤环境(Hodge, 2004 ; Verma et al ., 2005 ).同质低磷盆栽基质的表层、中间层和底层皆均匀添加有效磷含量极低(0.99 mg·kg-1 )的贫瘠酸性红壤作为盆栽基质, 该处理可有效地模拟森林土壤中磷素极为匮乏的针叶林立地环境(Richaedson et al ., 2004 ). ...
The response of plants to non-uniform supplies of nutrients.
1
1994
... 不同物种或不同基因型植物的氮素利用特性不同, 对不同氮供应方式的适应能力也存在差异.研究结果表明, 同质低磷下, 不同马尾松不同家系地上部分生长对局部供氮的响应差异较大, 而在异质低磷下, 地下部分的根系差异则更为明显.推测认为, 根系形态对土壤磷素异质分布的可塑性较强, 而对局部氮浓度的感知则较为迟钝(Tinker & Nye, 2000 ), 异质低磷环境中的磷素异质性增强了局部氮信号对马尾松苗木根系发育的影响, 进而产生了叠加效应(Jing et al ., 2010 ).在两种低磷环境下, 参试的5个马尾松不同家系中, 除了1号家系对氮添加不敏感外, 其他家系对局部供氮的响应因磷素环境和氮供应方式差异明显.如家系25的干物质积累量与植株氮、磷含量的变化有协同效应, 但与根系生长发育的变化相反, 这表明氮、磷元素在该家系内的快速转运以及利用可能对其整株生长贡献较大 (Kanno et al ., 2016 ).在异质低磷下, HHL限制了家系49、52和57的根系发育, 整株生长受到抑制; 而LLH和LLH/L促进了根系生长和干物质量的积累.这意味着在异质低磷环境下, 家系49、52和57主要通过改变根系的生长发育来响应局部氮素的变化.许多研究证实, 植物对土壤局部营养环境的感知及响应能力受营养元素分布、营养元素组成以及植株自身生长的环境影响(Robinson, 1994 ; Hodge, 2004 ).1号家系对局部供氮的响应较迟钝甚至有抑制效应, 分析认为, 这可能是因为家系1本身的基因型决定了其生长缓慢, 使得外界氮素的变化对植株根系发育的影响较小.本实验中局部养分供应对植物生长的促进效应可获得更多可利用的优良家系, 达到速生增产的目的, 但土壤中的养分异质性不仅是空间分布上的异质性, 也包括时间尺度上的异质性, 在长期的养分异质性环境下, 林木是否依然会保持旺盛的生长, 仍需更多的实验研究和长期观察, 以进一步客观地揭示异质养分土壤环境下, 局部氮素和磷素供应对马尾松不同家系苗木生长发育的影响. ...
Nutrient uptake, cluster root formation and exudation of protons and citrate in Lupinus albus as affected by localized supply of phosphorus in a split-root system.
1
2005
... 近年来, 土壤中养分元素的分布方式对植株生长的影响越来越受到关注(Hodge, 2004 ; 吕超群等, 2007 ; 徐星凯等, 2012 ).本研究中的异质低磷环境较同质低磷更有利于马尾松苗木的生长, 与前人的研究结果(杨青等, 2011 ; 庞丽等, 2014 )一致.已有研究表明, 外源氮素添加(100 g·kg-1 )会使土壤有效氮含量增加, 改变植株生物量的分配进而促进整株的生长(段洪浪等, 2009 ; 庞丽等, 2014 ; 刘双娥等, 2015 ).在本实验中, 局部供氮处理的斑块氮浓度与均匀供氮处理的氮含量相同, 但总施肥量要少于均匀供氮(相当于HHH处理施氮量的1/6-2/3).研究结果显示, 不同的供氮方式对马尾松苗木生长的影响差异较大, 且受到土壤磷素环境影响.在同质低磷下, HHL显著促进了马尾松苗木的生长, 根冠比降低, 较其他供氮方式更有利于苗木地上部分的生长; 其次, LLH和LLH/L两种方式之间, LLH/L对马尾松整株的生长相对更有利, 这表明在同质低磷下, 局部供氮增强了土壤中养分的异质性, 促进了马尾松苗木整株的生长.类似的结果在桉树(Eucalyptus grandis )、油松(Pinus tabulaeformis )、火炬松(Pinus taeda )等植物中也被发现(Mou et al ., 1997 ; Zhou & Shangguan, 2007 ; Graciano et al ., 2009 ).然而, 值得注意的是, Fransen和de Kroon (2001)在研究多年生植物绒毛草和甘松茅时, 发现绒毛草仅在短期内能通过根系增生来获取斑块内大量养分并促进地上部分生长, 从长期生长来看这种优势则会消失.因此, 本文的实验结果对生产实践中的苗木培育具有一定的指导意义, 但是否在长期生产造林和人工林抚育中仍有指导性需要进一步研究证实.与同质低磷相反, 在异质低磷下, 与HHH相比, HHL不利于马尾松株高、地径和干物质量的提高, 这可能是由于异质低磷土壤中的表层磷含量较高, 导致土壤(酸性土壤) pH值偏高(Shen et al ., 2005 ), 植物在局部供氮后更倾向于对NH4+ -N的吸收, 而过多吸收NH4+ -N会对植物产生一定的毒害作用(Kronzucker et al ., 2001 ), 抑制其整株的生长.然而在有效磷较低的底层土壤中, LLH和LLH/L刺激后, 植株吸收的磷含量显著提高, 且整株干物质量达到均匀供氮处理水平.这表明底层(局部)供氮可促进马尾松苗木对土壤中磷素的有效吸收利用, 保证其快速生长的情况下减少对化学肥料的依赖. ...
1
... 不同物种或不同基因型植物的氮素利用特性不同, 对不同氮供应方式的适应能力也存在差异.研究结果表明, 同质低磷下, 不同马尾松不同家系地上部分生长对局部供氮的响应差异较大, 而在异质低磷下, 地下部分的根系差异则更为明显.推测认为, 根系形态对土壤磷素异质分布的可塑性较强, 而对局部氮浓度的感知则较为迟钝(Tinker & Nye, 2000 ), 异质低磷环境中的磷素异质性增强了局部氮信号对马尾松苗木根系发育的影响, 进而产生了叠加效应(Jing et al ., 2010 ).在两种低磷环境下, 参试的5个马尾松不同家系中, 除了1号家系对氮添加不敏感外, 其他家系对局部供氮的响应因磷素环境和氮供应方式差异明显.如家系25的干物质积累量与植株氮、磷含量的变化有协同效应, 但与根系生长发育的变化相反, 这表明氮、磷元素在该家系内的快速转运以及利用可能对其整株生长贡献较大 (Kanno et al ., 2016 ).在异质低磷下, HHL限制了家系49、52和57的根系发育, 整株生长受到抑制; 而LLH和LLH/L促进了根系生长和干物质量的积累.这意味着在异质低磷环境下, 家系49、52和57主要通过改变根系的生长发育来响应局部氮素的变化.许多研究证实, 植物对土壤局部营养环境的感知及响应能力受营养元素分布、营养元素组成以及植株自身生长的环境影响(Robinson, 1994 ; Hodge, 2004 ).1号家系对局部供氮的响应较迟钝甚至有抑制效应, 分析认为, 这可能是因为家系1本身的基因型决定了其生长缓慢, 使得外界氮素的变化对植株根系发育的影响较小.本实验中局部养分供应对植物生长的促进效应可获得更多可利用的优良家系, 达到速生增产的目的, 但土壤中的养分异质性不仅是空间分布上的异质性, 也包括时间尺度上的异质性, 在长期的养分异质性环境下, 林木是否依然会保持旺盛的生长, 仍需更多的实验研究和长期观察, 以进一步客观地揭示异质养分土壤环境下, 局部氮素和磷素供应对马尾松不同家系苗木生长发育的影响. ...
Phosphorus acquisition and use: Critical adaptations by plants for securing a nonrenewable resource.
1
2003
... 氮和磷是植物生长最重要的两种营养元素, 其有效性和空间分布对植物的生长和根系发育具有重要影响(Hodge, 2004 ; Vance et al ., 2003 ).研究揭示, 局部供氮或供磷可显著促进大麦(Hordeum vulgare )、玉米(Zea mays )和小麦(Triticum aestivum )等植物侧根数量的增加以及总根长的提高(Drew, 1975 ; Officer et al ., 2009 ).Zhang和Forde (2000)将拟南芥(Arabidopsis thaliana )分别放在局部高氮和均匀高氮的琼脂培养基上培养时发现, 根系会在富氮斑块中增生, 而均匀供氮处理中的根系总量降低.可见根系在富养斑块中的增生可促进植物对养分供应的有效利用(Linkohr et al ., 2002 ).更重要的是, 相比于单独局部供氮或供磷, 在斑块中同时添加氮和磷更能促进根系在富养斑块中增生, 提高营养元素的吸收效率, 进而加快整株的生长(Jing et al ., 2010 ).这表明植物根系在局部供养斑块中的增生受到营养元素组成以及元素之间互作效应的共同影响.在木本植物研究中也发现类似的变化规律, 但物种间差异较大(Hodge, 2004 ).Einsmann等(1999)发现, 10种木本植物中有4种植物的生物量在养分异质性的供应条件下有显著提高, 但在对同种植物的不同研究中, Bliss等(2002)并没有发现这一趋势.虽然有研究认为养分斑块的存在能够对植物地上部分的生长产生积极的促进效应(Jackson & Caldwell, 1996 ), 但多年生植物绒毛草(Holcus lanatus )仅在短期内出现大量根系增生和促进地上部分生长的现象, 长期来看这种优势很快会消失, 甘松茅(Nardus stricta )则对养分斑块反应不敏感(Fransen & de Kroon, 2001 ).显然, 养分斑块或者局部养分供应对植物生长的影响受很多因素的影响, 除了养分的属性外, 植物对养分的敏感性及试验中植物生长时间的长短对结果都会产生较大影响(李洪波等, 2013 ). ...
Phosphorus fractions in an acid soil continuously fertilized with mineral and organic fertilizers.
1
2005
... 低磷土壤环境设置参照Zhang等(2013)的浓度设计.基质土壤经过风干过筛后, 加入一定量的Ca(H2 PO4 )2 混匀.异质低磷盆栽基质的底层、中间层和表层依次填入添加0、0.10和1.00 g·kg-1 的Ca(H2 PO4 )2 , 相当于有效磷含量分别为0.99、15和 30 mg·kg-1 .盆栽基质由上而下有效磷含量逐渐降低的异质养分环境可模拟立地条件较好, 表层营养丰富的针阔混交林或阔叶林土壤环境(Hodge, 2004 ; Verma et al ., 2005 ).同质低磷盆栽基质的表层、中间层和底层皆均匀添加有效磷含量极低(0.99 mg·kg-1 )的贫瘠酸性红壤作为盆栽基质, 该处理可有效地模拟森林土壤中磷素极为匮乏的针叶林立地环境(Richaedson et al ., 2004 ). ...
Localized nitrate and phosphate application enhances root proliferation by wheat and maximises rhizosphere alkalization in acid subsoil.
1
2008
... 局部供氮对植物根系形态和分布的影响较大(Li et al ., 2014 ).研究结果表明, 在两种低磷环境下, 与HHH相比, LLH/L对马尾松苗木根系的生长发育有明显的促进作用, 尤其在异质低磷土壤环境下的促进作用更为突出, 其次是LLH, HHL反而不利于根系的生长发育.分析认为, 底层半侧供氮有利于根系增生发育的原因可能在于: 植物根系感知一侧氮信号的缺乏, 诱导了根原基和初级根的形成, 而富氮斑块中充足的氮供应则加速了根系的伸长(He et al ., 2003 ; Li et al ., 2014 ), 进而促进了植物整体根长和根表面积的增加.如在异质低磷下, LLH/L处理中的总根长是LLH处理的1.30倍.相似的规律在水稻感知局部磷胁迫的研究中也被发现(He et al ., 2003 ).另一方面, HHL对根长和根表面积有一定的抑制效应, 推测这可能是由于: 首先, 马尾松本身属于深根型植物, 根的向地性促使其向土壤深处觅取更多的养分, 而HHL处理中的底层并没有充足的氮素供应, 限制了根系的生长发育.这一结果与前人对禾本科植物的研究结果有所差异, 如小麦、玉米和水稻(Oryza sativa )等植物可通过增加根系在土壤表层富养区域的分布来响应自然界土壤中的异质养分环境(Weligama et al ., 2008 ; Jing et al ., 2010 ; Li et al ., 2014 ).马尾松作为木本植物, 其对土壤养分异质性分布的响应可能因树种和土壤养分组成的不同而差异较大(Einsmann et al ., 1999 ; Guo et al ., 2002 ; 梅莉等, 2006 ; 杨青等, 2011 ); 其次, 该实验的种子播种于土壤表层, 其生长初期的根系增生和扩张及时获得了营养的补充, 使得植物在养分分配时, 将更多的营养元素输送到了其他器官(茎、叶), 降低了对根系的养分投入, 限制了根系的增生发育(段洪浪等, 2009 ; 吴茜等, 2011 ).结合苗期马尾松的株高、地径等地上部分的生长状况也印证了这一推测. ...
模拟降水变化和土壤施氮对浙江古田山5个树种幼苗生长和生物量的影响
1
2011
... 局部供氮对植物根系形态和分布的影响较大(Li et al ., 2014 ).研究结果表明, 在两种低磷环境下, 与HHH相比, LLH/L对马尾松苗木根系的生长发育有明显的促进作用, 尤其在异质低磷土壤环境下的促进作用更为突出, 其次是LLH, HHL反而不利于根系的生长发育.分析认为, 底层半侧供氮有利于根系增生发育的原因可能在于: 植物根系感知一侧氮信号的缺乏, 诱导了根原基和初级根的形成, 而富氮斑块中充足的氮供应则加速了根系的伸长(He et al ., 2003 ; Li et al ., 2014 ), 进而促进了植物整体根长和根表面积的增加.如在异质低磷下, LLH/L处理中的总根长是LLH处理的1.30倍.相似的规律在水稻感知局部磷胁迫的研究中也被发现(He et al ., 2003 ).另一方面, HHL对根长和根表面积有一定的抑制效应, 推测这可能是由于: 首先, 马尾松本身属于深根型植物, 根的向地性促使其向土壤深处觅取更多的养分, 而HHL处理中的底层并没有充足的氮素供应, 限制了根系的生长发育.这一结果与前人对禾本科植物的研究结果有所差异, 如小麦、玉米和水稻(Oryza sativa )等植物可通过增加根系在土壤表层富养区域的分布来响应自然界土壤中的异质养分环境(Weligama et al ., 2008 ; Jing et al ., 2010 ; Li et al ., 2014 ).马尾松作为木本植物, 其对土壤养分异质性分布的响应可能因树种和土壤养分组成的不同而差异较大(Einsmann et al ., 1999 ; Guo et al ., 2002 ; 梅莉等, 2006 ; 杨青等, 2011 ); 其次, 该实验的种子播种于土壤表层, 其生长初期的根系增生和扩张及时获得了营养的补充, 使得植物在养分分配时, 将更多的营养元素输送到了其他器官(茎、叶), 降低了对根系的养分投入, 限制了根系的增生发育(段洪浪等, 2009 ; 吴茜等, 2011 ).结合苗期马尾松的株高、地径等地上部分的生长状况也印证了这一推测. ...
模拟氮沉降对温带阔叶红松林地氮素净矿化量的影响
2
2012
... 土壤养分在空间分布上多呈时空异质性或者斑块分布(Hutching & John, 2004 ).产生土壤养分空间异质性的原因很多, 其中包括有机物(凋落物、根系死亡以及动物尸体)的分解、无机养分在土壤中的扩散系数差异以及微地形因素影响等(Hodge, 2004 ; 徐星凯等, 2012 ; 刘双娥等, 2015 ).植物为了获得更多的养分, 其根系会增加在土壤养分富集区域的分布比例, 进而提高根系对养分的获取和利用能力(Hodge, 2004 ; Mommer et al ., 2012 ).近年来, 随着合理高效的供肥管理措施(局部施肥)在农业中广泛应用及其高效的生产力, 使其在林业上尤其是人工林经营管理中也开始得到广泛应用.因此, 了解肥料集约经营管理下的林木根系生长、形态分布以及养分吸收状况, 将有利于解释林木在生长上存在的差异, 同时为提高速生丰产林生产力提供更为完善和高效的技术指导. ...
... 近年来, 土壤中养分元素的分布方式对植株生长的影响越来越受到关注(Hodge, 2004 ; 吕超群等, 2007 ; 徐星凯等, 2012 ).本研究中的异质低磷环境较同质低磷更有利于马尾松苗木的生长, 与前人的研究结果(杨青等, 2011 ; 庞丽等, 2014 )一致.已有研究表明, 外源氮素添加(100 g·kg-1 )会使土壤有效氮含量增加, 改变植株生物量的分配进而促进整株的生长(段洪浪等, 2009 ; 庞丽等, 2014 ; 刘双娥等, 2015 ).在本实验中, 局部供氮处理的斑块氮浓度与均匀供氮处理的氮含量相同, 但总施肥量要少于均匀供氮(相当于HHH处理施氮量的1/6-2/3).研究结果显示, 不同的供氮方式对马尾松苗木生长的影响差异较大, 且受到土壤磷素环境影响.在同质低磷下, HHL显著促进了马尾松苗木的生长, 根冠比降低, 较其他供氮方式更有利于苗木地上部分的生长; 其次, LLH和LLH/L两种方式之间, LLH/L对马尾松整株的生长相对更有利, 这表明在同质低磷下, 局部供氮增强了土壤中养分的异质性, 促进了马尾松苗木整株的生长.类似的结果在桉树(Eucalyptus grandis )、油松(Pinus tabulaeformis )、火炬松(Pinus taeda )等植物中也被发现(Mou et al ., 1997 ; Zhou & Shangguan, 2007 ; Graciano et al ., 2009 ).然而, 值得注意的是, Fransen和de Kroon (2001)在研究多年生植物绒毛草和甘松茅时, 发现绒毛草仅在短期内能通过根系增生来获取斑块内大量养分并促进地上部分生长, 从长期生长来看这种优势则会消失.因此, 本文的实验结果对生产实践中的苗木培育具有一定的指导意义, 但是否在长期生产造林和人工林抚育中仍有指导性需要进一步研究证实.与同质低磷相反, 在异质低磷下, 与HHH相比, HHL不利于马尾松株高、地径和干物质量的提高, 这可能是由于异质低磷土壤中的表层磷含量较高, 导致土壤(酸性土壤) pH值偏高(Shen et al ., 2005 ), 植物在局部供氮后更倾向于对NH4+ -N的吸收, 而过多吸收NH4+ -N会对植物产生一定的毒害作用(Kronzucker et al ., 2001 ), 抑制其整株的生长.然而在有效磷较低的底层土壤中, LLH和LLH/L刺激后, 植株吸收的磷含量显著提高, 且整株干物质量达到均匀供氮处理水平.这表明底层(局部)供氮可促进马尾松苗木对土壤中磷素的有效吸收利用, 保证其快速生长的情况下减少对化学肥料的依赖. ...
异质低磷胁迫下马尾松家系根构型和磷效率的遗传变异
3
2011
... 本试验中的马尾松分层隔网盆栽设计, 参考杨青等(2011) 的磷控释土培系统, 同时结合一年生马尾松苗木的根系形态特征进行分层处理(图1 ).盆栽容器自上而下依次分为表层、中间层和深层3个层次, 间距8 cm, 每层之间用孔径为3 mm的塑料纤维网隔离(主根、基根均能顺利穿过).在试验装置上端填充4 cm厚的干净河沙作为缓冲层, 将经过0.2%- 0.4%的高锰酸钾溶液浸泡20 min的马尾松不同家系种子植入缓冲层中间, 每盆播种在3-5粒种子. ...
... 近年来, 土壤中养分元素的分布方式对植株生长的影响越来越受到关注(Hodge, 2004 ; 吕超群等, 2007 ; 徐星凯等, 2012 ).本研究中的异质低磷环境较同质低磷更有利于马尾松苗木的生长, 与前人的研究结果(杨青等, 2011 ; 庞丽等, 2014 )一致.已有研究表明, 外源氮素添加(100 g·kg-1 )会使土壤有效氮含量增加, 改变植株生物量的分配进而促进整株的生长(段洪浪等, 2009 ; 庞丽等, 2014 ; 刘双娥等, 2015 ).在本实验中, 局部供氮处理的斑块氮浓度与均匀供氮处理的氮含量相同, 但总施肥量要少于均匀供氮(相当于HHH处理施氮量的1/6-2/3).研究结果显示, 不同的供氮方式对马尾松苗木生长的影响差异较大, 且受到土壤磷素环境影响.在同质低磷下, HHL显著促进了马尾松苗木的生长, 根冠比降低, 较其他供氮方式更有利于苗木地上部分的生长; 其次, LLH和LLH/L两种方式之间, LLH/L对马尾松整株的生长相对更有利, 这表明在同质低磷下, 局部供氮增强了土壤中养分的异质性, 促进了马尾松苗木整株的生长.类似的结果在桉树(Eucalyptus grandis )、油松(Pinus tabulaeformis )、火炬松(Pinus taeda )等植物中也被发现(Mou et al ., 1997 ; Zhou & Shangguan, 2007 ; Graciano et al ., 2009 ).然而, 值得注意的是, Fransen和de Kroon (2001)在研究多年生植物绒毛草和甘松茅时, 发现绒毛草仅在短期内能通过根系增生来获取斑块内大量养分并促进地上部分生长, 从长期生长来看这种优势则会消失.因此, 本文的实验结果对生产实践中的苗木培育具有一定的指导意义, 但是否在长期生产造林和人工林抚育中仍有指导性需要进一步研究证实.与同质低磷相反, 在异质低磷下, 与HHH相比, HHL不利于马尾松株高、地径和干物质量的提高, 这可能是由于异质低磷土壤中的表层磷含量较高, 导致土壤(酸性土壤) pH值偏高(Shen et al ., 2005 ), 植物在局部供氮后更倾向于对NH4+ -N的吸收, 而过多吸收NH4+ -N会对植物产生一定的毒害作用(Kronzucker et al ., 2001 ), 抑制其整株的生长.然而在有效磷较低的底层土壤中, LLH和LLH/L刺激后, 植株吸收的磷含量显著提高, 且整株干物质量达到均匀供氮处理水平.这表明底层(局部)供氮可促进马尾松苗木对土壤中磷素的有效吸收利用, 保证其快速生长的情况下减少对化学肥料的依赖. ...
... 局部供氮对植物根系形态和分布的影响较大(Li et al ., 2014 ).研究结果表明, 在两种低磷环境下, 与HHH相比, LLH/L对马尾松苗木根系的生长发育有明显的促进作用, 尤其在异质低磷土壤环境下的促进作用更为突出, 其次是LLH, HHL反而不利于根系的生长发育.分析认为, 底层半侧供氮有利于根系增生发育的原因可能在于: 植物根系感知一侧氮信号的缺乏, 诱导了根原基和初级根的形成, 而富氮斑块中充足的氮供应则加速了根系的伸长(He et al ., 2003 ; Li et al ., 2014 ), 进而促进了植物整体根长和根表面积的增加.如在异质低磷下, LLH/L处理中的总根长是LLH处理的1.30倍.相似的规律在水稻感知局部磷胁迫的研究中也被发现(He et al ., 2003 ).另一方面, HHL对根长和根表面积有一定的抑制效应, 推测这可能是由于: 首先, 马尾松本身属于深根型植物, 根的向地性促使其向土壤深处觅取更多的养分, 而HHL处理中的底层并没有充足的氮素供应, 限制了根系的生长发育.这一结果与前人对禾本科植物的研究结果有所差异, 如小麦、玉米和水稻(Oryza sativa )等植物可通过增加根系在土壤表层富养区域的分布来响应自然界土壤中的异质养分环境(Weligama et al ., 2008 ; Jing et al ., 2010 ; Li et al ., 2014 ).马尾松作为木本植物, 其对土壤养分异质性分布的响应可能因树种和土壤养分组成的不同而差异较大(Einsmann et al ., 1999 ; Guo et al ., 2002 ; 梅莉等, 2006 ; 杨青等, 2011 ); 其次, 该实验的种子播种于土壤表层, 其生长初期的根系增生和扩张及时获得了营养的补充, 使得植物在养分分配时, 将更多的营养元素输送到了其他器官(茎、叶), 降低了对根系的养分投入, 限制了根系的增生发育(段洪浪等, 2009 ; 吴茜等, 2011 ).结合苗期马尾松的株高、地径等地上部分的生长状况也印证了这一推测. ...
Regulation of Arabidopsis root development by nitrate availability.
2000
Genetic variations in root morphology and phosphorus efficiency of Pinus massoniana under heterogeneous and homogeneous low phosphorus conditions.
1
2013
... 马尾松(Pinus massoniana )是重要的材用、脂用和生态造林树种, 广泛分布于我国的亚热带地区(秦国峰和周志春, 2012 ).当前研究已揭示了氮添加对低磷胁迫下马尾松生长影响的生物学机制, 包括地上部分生长、根系参数和氮、磷效率的适应性变化等(Zhang et al ., 2013 ; 庞丽等, 2014 ). 然而, 相关研究未考虑氮素本身在空间分布上的不均一性.鉴于此, 本文拟通过人工控制氮素在土壤空间上的异质性, 研究低磷胁迫下马尾松不同家系在局部供氮条件下的生长表现和根系发育, 进一步阐明马尾松不同家系对土壤异质养分的响应差异及机制, 为马尾松营养高效新品种选育和苗木高效栽培方式提供理论依据以及实践指导. ...
Vertical distribution of fine roots in relation to soil factors in Pinus tabulaeformis Carr. forest of the Loess Plateau of China.
1
2007
... 近年来, 土壤中养分元素的分布方式对植株生长的影响越来越受到关注(Hodge, 2004 ; 吕超群等, 2007 ; 徐星凯等, 2012 ).本研究中的异质低磷环境较同质低磷更有利于马尾松苗木的生长, 与前人的研究结果(杨青等, 2011 ; 庞丽等, 2014 )一致.已有研究表明, 外源氮素添加(100 g·kg-1 )会使土壤有效氮含量增加, 改变植株生物量的分配进而促进整株的生长(段洪浪等, 2009 ; 庞丽等, 2014 ; 刘双娥等, 2015 ).在本实验中, 局部供氮处理的斑块氮浓度与均匀供氮处理的氮含量相同, 但总施肥量要少于均匀供氮(相当于HHH处理施氮量的1/6-2/3).研究结果显示, 不同的供氮方式对马尾松苗木生长的影响差异较大, 且受到土壤磷素环境影响.在同质低磷下, HHL显著促进了马尾松苗木的生长, 根冠比降低, 较其他供氮方式更有利于苗木地上部分的生长; 其次, LLH和LLH/L两种方式之间, LLH/L对马尾松整株的生长相对更有利, 这表明在同质低磷下, 局部供氮增强了土壤中养分的异质性, 促进了马尾松苗木整株的生长.类似的结果在桉树(Eucalyptus grandis )、油松(Pinus tabulaeformis )、火炬松(Pinus taeda )等植物中也被发现(Mou et al ., 1997 ; Zhou & Shangguan, 2007 ; Graciano et al ., 2009 ).然而, 值得注意的是, Fransen和de Kroon (2001)在研究多年生植物绒毛草和甘松茅时, 发现绒毛草仅在短期内能通过根系增生来获取斑块内大量养分并促进地上部分生长, 从长期生长来看这种优势则会消失.因此, 本文的实验结果对生产实践中的苗木培育具有一定的指导意义, 但是否在长期生产造林和人工林抚育中仍有指导性需要进一步研究证实.与同质低磷相反, 在异质低磷下, 与HHH相比, HHL不利于马尾松株高、地径和干物质量的提高, 这可能是由于异质低磷土壤中的表层磷含量较高, 导致土壤(酸性土壤) pH值偏高(Shen et al ., 2005 ), 植物在局部供氮后更倾向于对NH4+ -N的吸收, 而过多吸收NH4+ -N会对植物产生一定的毒害作用(Kronzucker et al ., 2001 ), 抑制其整株的生长.然而在有效磷较低的底层土壤中, LLH和LLH/L刺激后, 植株吸收的磷含量显著提高, 且整株干物质量达到均匀供氮处理水平.这表明底层(局部)供氮可促进马尾松苗木对土壤中磷素的有效吸收利用, 保证其快速生长的情况下减少对化学肥料的依赖. ...
 , 周志春
, 周志春

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}