



群落构建研究一直是生态学研究的热点, 准确理解和预测驱动群落构建的关键因子并采取针对性的行动可以缓解全球气候变化所带来的问题(Naeem & Wright, 2003; McGill et al., 2006)。群落构建机制主要分为两类: 侧重物种生态策略差异的生态位过程和侧重物种相同适合度的中性过程(牛克昌等, 2009)。前者包括竞争作用和促进作用等; 后者为随机过程。近年来发现受非生物环境胁迫(如低温、土壤养分缺乏、干旱等)的影响, 促进作用和竞争作用能同时影响同一群落物种共存, 且影响大小随环境胁迫强弱而变化(Kraft et al., 2008; McIntire & Fajardo, 2014; Long et al., 2015a, 2015b)。非生物环境不仅可以作为环境筛影响植物形态结构、生理功能和系统发育关系等, 决定群落中共存物种的组成特征; 也可以通过给植物体提供不同数量和质量的资源而影响其生态位, 促进群落物种共存(He et al., 2009)。
早期的群落构建研究主要是从物种多样性变化角度, 把所有物种视为独立进化的生态等价物, 但不能解释物种进化历史和功能性状信息, 不能准确地揭示群落构建的成因(Webb et al., 2002; Swenson et al., 2012)。在近十余年里, 生态学家们选择功能多样性(functional diversity)和谱系多样性(phylogenetic diversity)来探究群落构建机制(Webb et al., 2002; Swenson et al., 2012; Swenson, 2013)。他们发现群落物种的功能多样性越高, 物种性状的趋异性越强, 物种间因生存策略高度差异使其在群落中生态位高度分化而稳定共存(Spasojevic & Suding, 2012); 而群落谱系结构研究既可以从进化角度分析群落物种组成现状和成因, 又能检验到影响群落物种共存的生态学过程(Cavender-Bares et al., 2009)。群落的组成还受到生物地理和进化历史过程的影响(Cornell, 1985; Ricklefs, 1987), Harrison和Cornell (2008)提出把区域物种库加入到生态学分析中, 可以很好地避免这些干扰。从广义上来说, 区域物种库为对于某个特定群落, 已经在该群落分布以及所有可能拓殖到该群落但目前并没有分布的潜在物种集合(Srivastava, 1999)。区域物种库概念为物种形成、灭绝和扩散对当地生物多样性的影响提供了新的见解(Eriksson, 1993; Cornell & Harrison, 2014; Zobel & Scheiner, 2016)。因此, 在探究群落的生态学进程(物种间相互作用或者环境筛)对群落的物种、功能性状和系统发育关系的影响时, 区域物种库是一个必不可少的因素(Cornell & Harrison, 2014)。
很多生态学家把热带森林选作验证物种共存机制的理想场所。从空间格局、功能性状、谱系结构等角度阐明了物种多样性的维持机制(Kraft et al., 2008; Paine et al., 2011; Pearse et al., 2013)。但这些研究多数局限于低海拔森林, 对高海拔森林(如热带云雾林)较少涉及。国内外热带云雾林研究集中于群落结构、生物多样性、植物对水分利用和养分循环等方面(Goldsmith et al., 2013; Eller et al., 2015), 部分学者开展了功能性状特征和系统发育的研究(Luna-Vega & Magallón, 2010; Long et al., 2011b, 2015a; 王茜茜等, 2016; Kang et al., 2017)。海南岛的热带云雾林分布在霸王岭、五指山、尖峰岭、黎母山、吊罗山等山顶或山脊, 群落空气温度低, 云雾出现频率高, 风力强劲, 土壤含水量常处于饱和状态; 树木高度和径级小, 树干常弯曲, 小叶革质, 单叶比例高(Long et al., 2011b)。在前期研究中, 发现比叶面积小、高度大的物种在热带云雾林有生存优势(Long et al., 2015a), 在热带云雾林低温和低磷环境中, 竞争作用和促进作用同时影响物种共存; 而基于物种比叶面积功能性状数据时发现生境过滤在较大空间尺度上主导了热带云雾林群落构建过程(Long et al., 2015a, 2015b)。但是, 功能多样性能根据植物在资源获取、更新繁衍、环境耐受等多个功能性状轴信息, 检验多个生态过程共同作用于群落构建(Spasojevic & Suding, 2012); 而谱系多样性能够利用物种的系统发育状况推测历史因素对现有群落的影响(Webb et al., 2002; Cadotte et al., 2008)。因此通过功能性状和谱系多样性探究热带云雾林群落构建机制, 可能会有新的研究发现。在海南霸王岭热带云雾林设置固定样地, 测定物种的功能性状、DNA条形码和群落环境因子, 分析功能多样性和谱系多样性对群落构建的影响。我们假设: 受环境因子影响, 热带云雾林的功能多样性和谱系多样性呈聚集状态。
1 材料和方法
1.1 研究样地概况
图1
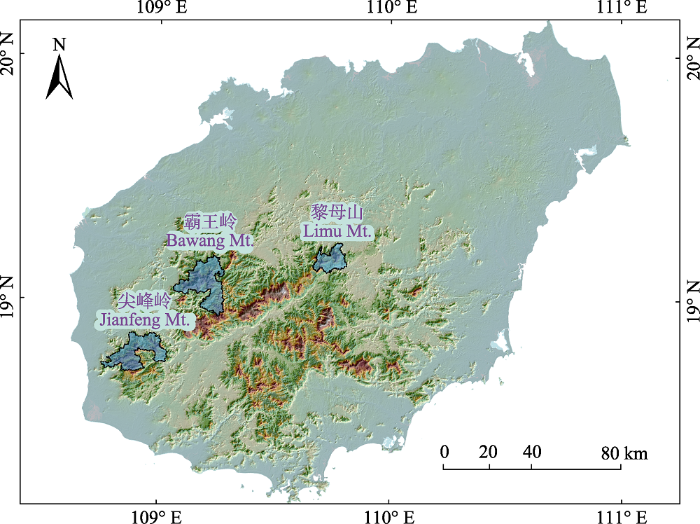
图1
海南岛热带云雾林样地分布图。
Fig. 1
Distribution of the tropical cloud forest study sites in Hainan Island.
表1 海南岛云雾林样地概况
Table 1
| 研究样地 Study site | 海拔 Elevation (m) | 经度 Longitude (E) | 纬度 Latitude (N) | 坡度 Slope (°) | 样方数 No. of plots | 样地面积 Plot area (m2) | 优势种 Dominant species |
|---|---|---|---|---|---|---|---|
| 尖峰岭 Jianfeng Mt. | 1 187.17- 1 397.19 | 108.87° | 18.72° | 10-65 | 12 | 4 800 | 罗浮锥、丛花厚壳桂、美丽新木姜子、黄叶树 Castanopsis faberi, Cryptocarya densiflora, Neolitsea pulchella, Xanthophyllum hainanense |
| 霸王岭 Bawang Mt. | 1 313.24- 1 385.24 | 109.21° | 19.08° | 2-45 | 21 | 8 400 | 蚊母树、赤楠、九节、黄杞 Distylium racemosum, Syzygium buxifolium, Psychotria rubra, Engelhardtia roxburghiana |
| 黎母山 Limu Mt. | 1 363.73- 1 403.32 | 109.76° | 19.18° | 3-42 | 15 | 6 000 | 普洱茶、岭南青冈、罗浮锥、细枝柃 Camellia sinensis var. assamica, Cyclobalanopsis championii, Castanopsis faberi, Eurya loquaiana |
The area for each plot is 400 m2.
每个样方面积为400 m2。
1.2 数据收集
1.2.1 物种库的建立
调查5 m × 5 m小样方内所有胸径≥1 cm的植株。根据《中国植物志》确定个体物种名, 现场无法确认的物种做成标本请专家鉴定。尖峰岭群落共调查植株6 879株, 235种; 霸王岭9 714株, 117种; 黎母山3 809株, 148种。三个样地共采集126个物种, 汇总功能性状数据建立物种库。
1.2.2 功能性状测定
功能性状选择原则是: 充分体现植物对胁迫响应、对干扰适应及对环境资源的获取能力; 兼顾反应性状和效应性状。选取13个功能性状, 包括主茎特征: 木材密度(WD)、茎干可溶性糖含量(SS)、茎干氮含量(SN)、茎干磷含量(SP); 叶片特征: 叶片厚度(LTh)、比叶面积(SLA)、叶绿素含量(Chl)、叶片可溶性糖含量(LS)、叶片氮含量(LN)、叶片磷含量(LP); 根部特征: 根可溶性糖含量(RS)、根氮含量(RN)、根磷含量(RP)。其中根茎叶的可溶性糖含量与植物对环境胁迫的响应能力有关, 比叶面积和叶绿素含量与光合作用有关, 木材密度与植物水分利用及固碳能力有关。根茎叶的氮磷含量与植物的养分利用与分配有关。
每个物种采集3株个体, 每株个体采集2-3片成熟的、受太阳光直射的、健康的冠层叶片(Cornelissen et al., 2003), 叶片厚度用数显游标卡尺(SF2000, 广陆, 广西桂林)测量; 叶绿素含量用便携式叶绿素仪(SPAD-502 Plus, Konica Minolta, Tokyo, Japan)测量; 叶面积用叶面积仪(LI-COR 3100C Area Meter, LI-COR, Lincoln, USA)测定。然后将叶片装入信封放于80 ℃烘箱烘干至恒质量, 用电子天平(AR2140, Ohaus, Parsippany, USA)称量干质量。比叶面积由叶面积与干质量的比值算得。在采集叶片样品的同时, 从每个植株上截取4根直径为2-3 cm、长约10 cm的枝条并采取根部样品, 去皮后用排水法测量枝条体积。然后将枝条装入信封放于80 ℃烘箱烘干至恒质量, 称量其干质量, 根据枝条干质量与体积之比计算木材密度。研究发现热带森林树木枝条密度与木材密度显著相关(硬木: R2 = 0.95, p < 0.001; 软木: R2 = 0.93, p < 0.001; Bu et al., 2014), 所以可用植物枝条密度表示木材密度, 如此可避免用生长锥钻取年轮条对树木的伤害。氮含量用凯氏定氮法测定, 磷含量用HClO4-H2SO4消化法分解样品, 然后用钼蓝比色法测量, 可溶性糖含量用蒽酮比色法测定。
1.2.3 DNA条形码的测定
采集3个样地的乔木树种, 每个乔木树种选择1-2株胸径≥5 cm的个体, 采集树木新鲜完整叶片, 用硅胶干燥。DNA的提取使用成都福际生物技术有限公司的植物总DNA提取试剂盒。选择ITS、rbcL、matK 3个片段作为研究片段。参照相关的PCR反应体系(CBOL Plant Working Group, 2009), 做出筛选和优化。扩增产物送至北京六合华大基因科技有限公司(BGI)广州分公司进行测序工作。
1.2.4 环境因子的测定
环境因子的测定均以5 m × 5 m小样方为单位测定, 然后取平均值得到20 m × 20 m样方的平均值。
(1)林冠开阔度测定: 在每个5 m × 5 m小样方中, 将数码相机(D5100, Nikon, Tokyo, Japan)和鱼眼镜头(Sigma F2.8 HSM, Tokyo, Japan)放置在离地面约1.3 m的三脚架上采集图像。对图像的处理先用Sidelook软件确定阈值, 然后用Gap Light Analyzer Version 2.0 (GLA )分析处理图像计算林冠开阔度。
(2)土壤因子测定: 在每个5 m × 5 m小样方中, 去除土壤表层凋落物, 自上而下取20 cm混合土壤样品, 取好的土壤样品混合均匀做好标记带回实验室, 自然晾干后, 测定土壤有机质含量(SOM)、全氮含量(TN)、全磷含量(TP)、有效氮含量(AN)、有效磷含量(AP)。土壤有机质含量的测定用重铬酸钾容量法, 全磷含量的测定方法是用HClO4-H2SO4消化法分解样品, 然后用钼锑抗比色法测定, 全氮含量用凯氏定氮法测定, 有效磷含量用盐酸-氟化铵法测定, 有效氮含量用碱解扩散法测定(中国土壤学会农业化学专业委员会, 1983)。
(3)地形因子测定: 在每个5 m × 5 m小样方中, 用坡度坡向仪在每个样方的中心位置测量坡度。
1.3 数据分析
1.3.1 环境因子的筛选
为了筛选热带云雾林中的主要环境因子, 首先建立20 m × 20 m样方尺度上样方×环境因子矩阵, 然后用主成分分析(PCA)对环境因子进行分析。当累积解释方差比例在78.0%以上时, 取前面全部PCA轴, 并依照环境因子在PCA轴上的负荷评定其影响作用, 取负荷绝对值在0.50以上的环境因子。其次, 用Pearson相关性分析环境因子间的相关性, 观察PCA判定的主要环境因子与其他环境因子的相关性。
1.3.2 系统发育树的构建
DNA条形码测定后经过序列编辑、比对, 条形码的拼接, 然后进行系统发育树的构建, 主要选用软件Bioedit、Sequencematrix和Mega 6.0, 详细方法参考Kang等(2017)。
1.3.3 功能性状的系统发育信号检测
群落谱系关系是影响功能性状的重要因素。通常认为亲缘关系近的物种间性状差异较小, 亲缘关系较远的物种间性状差异较大。所以在物种间性状研究中需检验物种的功能性状是否表现出系统发育信号。采用Blomberg’s K (Blomberg et al., 2003)检验功能性状的系统发育信号强度。若K = 1, 则表明该功能性状表现出按布朗运动模型的方式进化, 这预示着某种程度的系统发育信号和性状保守性; 若K < 1, 则表示功能性状表现出的系统发育信号比按布朗运动模型进化弱, 若K接近于0, 则说明功能性状表现出的系统发育信号是随机或者发散的; 若K > 1, 则表示功能性状表现出强烈的系统发育信号, 预示功能性状表现出系统发育保守性。功能性状系统发育信号的显著性采用与零模型比较的方式来衡量, 若是实际值大于零模型值的次数达到950次以上, 就认为功能性状表现出显著的系统发育信号(p < 0.05)。
1.3.4 功能多样性对群落构建的影响
功能多样性指数选择Rao二次熵(RaoQ), 它同时包含物种的相对丰度和物种之间成对功能差异的度量(Mason et al., 2012), 可以很好地反映群落功能性状的变化, 在生境过滤主导的群落中, 相较于其他的功能多样性指数, RaoQ对群落构建机制有较好的检验能力(Botta-Dukát & Czúcz, 2016)。以物种库的功能性状和多度数据为基础, 保证物种数量和个体数目不变, 对物种多度数据随机分配9 999次, 得到随机的样方物种多度矩阵, 建立零模型, 得到群落RaoQ期望值, 用霸王岭样地的功能性状和多度数据计算RaoQ的观测值。用R语言中的randomizeMatrix和dbFD函数计算。运用Wilcoxon test检验期望值与观测值的差异性。
1.3.5 谱系多样性对群落构建的影响
选择平均成对谱系距离MPD谱系多样性指数, MPD指数可以预估在聚类树上所有成对分类单元之间的平均谱系距离(Webb et al., 2008), 可以说明进化树的整体聚类情况(Webb et al., 2002), 而且MPD指数常常是基于物种多度或者基面积加权, 这种特性可以更好地反映生物和非生物因素相互作用的相对重要性(Hardy, 2008; Vamosi et al., 2009)。以物种库数据为基础, 构建系统发育树, 保证物种数量和个体数目不变, 系统发育树末端的物种随机置换9 999次, 得到群落MPD的期望值; 用霸王岭样地的功能性状和多度数据计算MPD观测值; 基于R语言中的randomizeMatrix和MPD函数进行计算。运用Wilcoxon test检验期望值与观测值的差异性。
1.3.6 环境因子对热带云雾林群落构建的影响
利用RaoQ和MPD的预期值与观测值计算标准效应值(SES), 标准效应值计算过程如下:
SES.RaoQ = -(RaoQobs - RaoQnull)/sdRaoQnull
NRI = -(MPDobs - MPDnull)/sdMPDnull
RaoQobs为RaoQ观测值, RaoQnull为预期值的均值, sdRaoQnull为预期值的标准偏差; NRI为净种间亲缘关系指数; MPDobs为MPD观测值, MPDnull为预期值的均值, sdMPDnull为预期值的标准偏差。运用单元线性回归, 分析环境因子与标准效应值间相关性, 筛选环境筛。
2 结果
2.1 环境因子PCA及相关性分析
表2 热带云雾林环境因子的主成分分析
Table 2
| 环境变量 Environmental variable | PCA1 | PCA2 | PCA3 | PCA4 |
|---|---|---|---|---|
| 林冠开阔度 CO (%) | -0.33 | - | 0.89 | -0.24 |
| 土壤有机质含量 SOM (g·kg-1) | -0.35 | 0.47 | -0.23 | -0.34 |
| 全磷含量 TP (g·kg-1) | -0.50 | - | - | 0.31 |
| 全氮含量 TN (g·kg-1) | -0.47 | -0.11 | -0.32 | -0.15 |
| 有效氮含量 AN (mg·kg-1) | -0.47 | -0.19 | -0.19 | -0.19 |
| 有效磷含量 AP (mg·kg-1) | 0.24 | 0.59 | - | -0.48 |
| 坡度 SP (º) | 0.13 | -0.61 | - | -0.66 |
| 特征值 Characteristic value | 3.32 | 1.87 | 0.48 | 0.17 |
| 解释方差比例 Explained variance proportion | 0.52 | 0.27 | 0.10 | 0.07 |
| 累积解释方差比例 Cumulative explained variance proportion | 0.52 | 0.79 | 0.89 | 0.96 |
Data in bold letters indicate that the absolute loadings of environmental variables are over 0.5. CO, canopy openness; SOM, soil organic matter content; TP, total phosphorus content; TN, total nitrogen content; AN, available nitrogen content; AP, available phosphorus content; SP, slope.
加粗数字表示该变量的负荷绝对值大于0.5。
图2
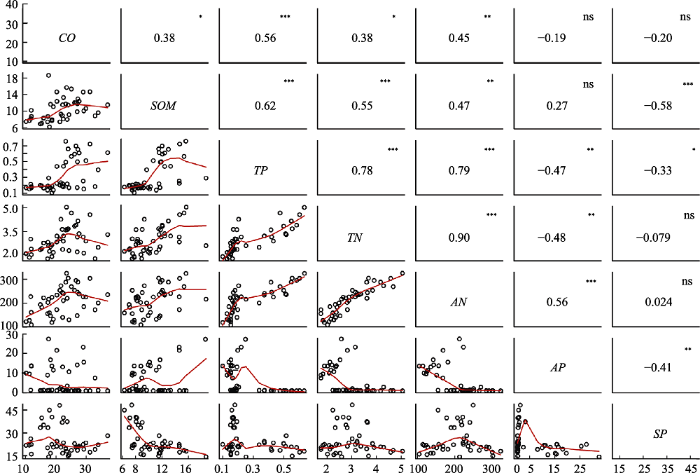
图2
海南岛热带云雾林群落环境因子相关性分析。CO, 林冠开阔度; SOM, 土壤有机质含量; TP, 全磷含量; TN, 全氮含量; AN, 有效氮含量; AP, 有效磷含量; SP, 坡度。*, p < 0.05; **, p < 0.01; ***, p < 0.001。
Fig. 2
Correlation analysis among environmental factors of Hainan tropical cloud forests. CO, canopy openness; SOM, soil organic matter content; TP, total phosphorus content; TN, total nitrogen content; AN, available nitrogen content; AP, available phosphorus content; SP, slope. *, p < 0.05; **, p < 0.01; ***, p < 0.001.
2.2 系统发育信号
茎秆磷含量的K值大于1 (表3), 表现出较强的系统发育保守性; 其他12个功能性状的K值均小于1, 没有强的系统发育保守性。叶片氮含量和茎秆磷含量表现出显著的系统发育信号(p < 0.05), 而其他11个功能性状均未检测到明显的系统发育信号(p > 0.05), 表明叶片氮含量和茎秆磷含量受物种的进化历史影响较大。
表3 热带云雾林植物功能性状的系统发育信号
Table 3
| 功能性状 Functional trait | K | p |
|---|---|---|
| 根部氮含量 Root nitrogen content (RN) | 0.258 | 0.639 |
| 根部磷含量 Root phosphorus content (RP) | 0.405 | 0.193 |
| 根部可溶性糖含量 Root soluble sugar content (RS) | 0.210 | 0.820 |
| 叶片氮含量 Leaf nitrogen content (LN) | 0.677 | 0.027 |
| 叶片磷含量 Leaf phosphorus content (LP) | 0.579 | 0.055 |
| 叶片可溶性糖含量 Leaf soluble sugar content (LS) | 0.190 | 0.857 |
| 茎干氮含量 Stem nitrogen content (SN) | 0.448 | 0.195 |
| 茎干磷含量 Stem phosphorus content (SP) | 4.258 | 0.005 |
| 茎干可溶性糖含量 Stem soluble sugar content (SS) | 0.314 | 0.429 |
| 比叶面积 Specific leaf area (SLA) | 0.373 | 0.413 |
| 叶绿素含量 Chlorophyll content (Chl) | 0.459 | 0.062 |
| 叶片厚度 Leaf thickness (LTh) | 0.282 | 0.589 |
| 木材密度 Wood density (WD) | 0.294 | 0.527 |
2.3 功能多样性和谱系多样性对群落构建的影响
系统发育树平均节点支持率为72.22% ± 27.79%, 可以用于谱系分析(附录I)。热带云雾林群落的功能多样性和谱系多样性指数都显著低于零模型期望值(图3A、3B; p < 0.05), 表明群落功能多样性和谱系多样性都呈现聚集状态。
图3
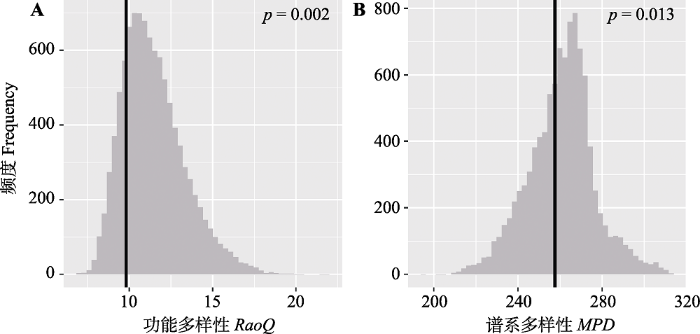
图3
热带云雾林功能多样性和谱系多样性期望值与观测值比较。预期值, 灰色直方图; 观测值, 黑色垂直线。CO, 林冠开阔度; TP, 全磷含量; SP, 坡度。
Fig. 3
Comparison of functional diversity and phylogenetic diversity expectations with observed values in tropical cloud forest. The null distribution is the grey histogram and the observed values are the black vertical bars. MPD, mean pairwise distance; RaoQ, Rao’s quadratic entropy.
2.4 环境因子对群落构建的影响
图4
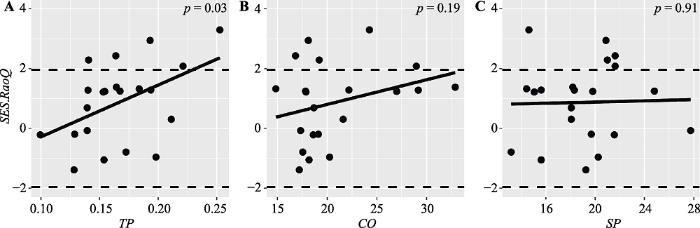
图4
热带云雾林Rao二次熵指数的标准效应值(SES.RaoQ)沿环境梯度的变化规律。CO, 林冠开阔度; TP, 全磷含量; SP, 坡度。
Fig. 4
Patterns of standard effect size of Rao’s quadratic entropy (SES.RaoQ) along environmental gradients in tropical cloud forest. CO, canopy openness; TP, total phosphorus content; SP, slope.
图5
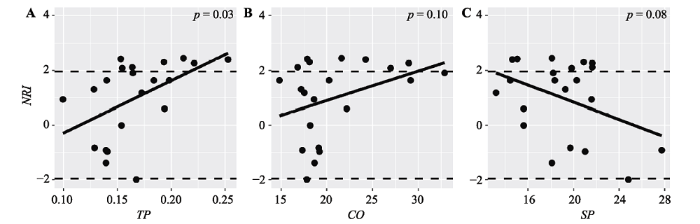
图5
热带云雾林净种间亲缘关系指数(NRI)沿环境梯度的变化规律。CO, 林冠开阔度; TP, 全磷含量; SP, 坡度。
Fig. 5
Patterns of net relatedness index (NRI) along environmental gradients in tropical cloud forest. CO, canopy openness; TP, total phosphorus content; SP, slope.
3 讨论
3.1 生境过滤影响热带云雾林的群落构建
在基于系统发育关系和功能性状的群落构建机制研究中, 有一个非常重要的假设: 系统发育关系更近的物种在生态学意义上总是更相似, 同一个属的物种往往比非同属的物种在性状上是更相似的, 物种之间的系统发育关系与其功能性状之间有很强的对应关系(Webb et al., 2002)。但是热带云雾林群落中, 功能性状并没有表现出系统发育保守性(表3, K < 1), 换言之, 功能性状格局和系统发育格局之间缺乏一致性, 两者之间并没有出现完美的匹配关系(Swenson & Enquist, 2009; Swenson, 2011)。与假设提出的关系不一致, 这种结果可能是很多原因造成的。首先, 系统发育-性状相似性的匹配关系往往出现在较大的分类学尺度上, 并不能在物种水平做到完美的一一对应的关系, 这样说明在不同的分类学尺度上这个假设并不是通用的。其次, Swenson等(2006)认为随机系统发育关系的产生是由于一些功能性状处于非随机的保守状态, 而另一部分却处于非随机的发散状态造成的。如果继续简单地把系统发育和功能性状相似性对应起来, 大量的功能性状信息就被忽略, 那么得到的结论就是错误的。最后, Swenson (2013)也提出系统发育关系只能是判断群落中物种性状相似的一个间接性指标, 而并不是绝对的一一对应关系, 它并不涵盖群落中物种所有功能性状的信息, 因此不能直接把物种的系统发育关系和其功能性状相似性直接匹配在一起。
正是因为基于系统发育的方法存在很大局限性, 很多的生态学家认为基于系统发育关系检验群落构建机制的方法并不合适(Losos, 2008), 也更倾向于使用基于功能性状的检验方法, 甚至提出在功能性状可以测定的条件下, 群落系统发育就不需要考虑了。但是基于功能性状的群落构建机制方法通常就是选择关于群落中物种生态策略或者生理过程相关的一些功能性状(例如形态和结构特征、营养成分等) (Yang et al., 2014), 包括本文也是利用一些营养元素和结构特征的功能性状。但是我们不可能把所有与植物生理生态相关的功能性状全部进行测定, 往往功能性状的选择只是根据一些常用的标准或者在其他研究中得到的结果, 但这些可能并不适用于所有的群落类型; 再者大多数的研究都主要集中于物种水平的功能性状变异, 往往忽略了种内变异情况。由于群落物种间的系统发育关系携带了大量群落历史进化信息, 这些不能由简简单单的功能性状表现出来(Cadotte et al., 2008)。因而研究群落构建机制的时候, 同时兼顾群落物种的功能性状和系统发育关系非常有必要。
我们的研究发现热带云雾林的群落构建主要受到生境过滤的作用, 土壤磷含量是影响热带云雾林群落构建的关键因子。这说明在热带云雾林的经营管理中要重视土壤条件的影响。但本文仅从单一尺度来探讨群落构建机制, 并未考虑到物种种内的功能性状变异, 在未来的研究中可以探究在不同尺度上基于种内和种间功能性状变化的群落构建规律。
附录
附录I 海南霸王岭热带云雾林系统发育树
Supplement I Phylogenetic tree of tropical cloud forest in Bawangling, Hainan
http://www.plant.ecology.com/fileup/PDF/cipe.2019.0003-s1.pdf

参考文献
Trait-based community assembly of understory palms along a soil nutrient gradient in a lower montane tropical forest
Testing for phylogenetic signal in comparative data: Behavioral traits are more labile
Testing the ability of functional diversity indices to detect trait convergence and divergence using individual-based simulation
Field observed relationships between biodiversity and ecosystem functioning during secondary succession in a tropical lowland rainforest
Evolutionary history and the effect of biodiversity on plant productivity
The merging of community ecology and phylogenetic biology
A DNA barcode for land plants
A handbook of protocols for standardised and easy measurement of plant functional traits worldwide
Local and regional richness of cynipine gall wasps on California oaks
What are species pools and when are they important?
Functional species pool framework to test for biotic effects on community assembly
Environmental controls in the water use patterns of a tropical cloud forest tree species, Drimys brasiliensis (Winteraceae)
The species-pool hypothesis and plant community diversity
The incidence and implications of clouds for cloud forest plant water relations
Phylogenetic alpha and beta diversity in tropical tree assemblages along regional-scale environmental gradients in northwest South America
Testing the spatial phylogenetic structure of local communities: Statistical performances of different null models and test statistics on a locally neutral community
Toward a better understanding of the regional causes of local community richness
Taxonomic, phylogenetic, and environmental trade-offs between leaf productivity and persistence
DNA barcoding analysis and phylogenetic relationships of tree species in tropical cloud forests
Functional trait and phylogenetic tests of community assembly across spatial scales in an Amazonian forest
Functional traits and niche-based tree community assembly in an Amazonia forest
Community assembly in a tropical cloud forest related to specific leaf area and maximum species height
Changes in patterns of species co-occurrence across two tropical cloud forests differing in soil nutrients and air temperature
Air temperature and soil phosphorus availability correlate with trait differences between two types of tropical cloud forests
Within- and among-species variation in specific leaf area drive community assembly in a tropical cloud forest
Phylogenetic niche conservatism, phylogenetic signal and the relationship between phylogenetic relatedness and ecological similarity among species
Phylogenetic composition of angiosperm diversity in the cloud forests of Mexico
Changes in coexistence mechanisms along a long-term soil chronosequence revealed by functional trait diversity
Rebuilding community ecology from functional traits
Facilitation as a ubiquitous driver of biodiversity
Community assembly processes shape an altitudinal gradient of forest biodiversity
Disentangling biodiversity effects on ecosystem functioning: deriving solutions to a seemingly insurmountable problem
Community assemble: The relative important of neutral theory and niche theory
群落构建的中性理论和生态位理论
Functional traits of individual trees reveal ecological constraints on community assembly in tropical rain forests
Dark diversity: Shedding light on absent species
Barro Colorado Island’s phylogenetic assemblage structure across fine spatial scales and among clades of different ages
Community diversity: Relative roles of local and regional processes
Inferring community assembly mechanisms from functional diversity patterns: The importance of multiple assembly processes
Using local-regional richness plots to test for species saturation: Pitfalls and potentials
Phylogenetic beta diversity metrics, trait evolution and inferring the functional beta diversity of communities
The assembly of tropical tree communities-the advances and shortcomings of phylogenetic and functional trait analyses
Opposing assembly mechanisms in a Neotropical dry forest: Implications for phylogenetic and functional community ecology
The problem and promise of scale dependency in community phylogenetics
Phylogenetic and functional alpha and beta diversity in temperate and tropical tree communities
Emerging patterns in the comparative analysis of phylogenetic community structure
Patterns of plant diversity within and among three tropical cloud forest communities in Hainan Island
海南岛3个林区热带云雾林植物多样性变化
Phylocom: Software for the analysis of phylogenetic community structure and trait evolution
Phylogenies and community ecology
Community assembly rules, morphological dispersion, and the coexistence of plant species
Functional and phylogenetic assembly in a Chinese tropical tree community across size classes, spatial scales and habitats
The species pool concept as a framework for studying patterns of plant diversity: Official organ of the International Association for Vegetation Science

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}