Biochemistry of hexose and pentose transformations in soil analyzed by position-specific labeling and 13C-PLFA
1
2015
... 13C同位素示踪法技术可以很好地研究外源C对SOM周转的影响.例如, 将13C标记的有机物加入土壤进行田间或室内培养实验, 通过测定13CO2可以区分外源有机物和原有SOM来源的碳, 量化外源有机物添加SOM分解的影响(Bernoux et al., 1998; Blagodatskaya & Kuzyakov, 2008).高丰度13C标记的有机物结合13C-PLFA-SIP和13C-DNA-SIP技术, 可以准确示踪微生物在土壤有机碳周转过程中的参与机制(Williams et al., 2006; Apostel et al., 2015; Wang et al., 2016).而13C-微生物残留物对于量化和评估微生物对SOM积累的贡献起到了关键作用(Tian et al., 2014; Zhang et al., 2015; Liang et al., 2017). ...
Carbon isotope discrimination as indicator of water-use efficiency of spring wheat as affected by salinity and gypsum addition
1
1999
... 基于C3/C4植物δ13C的不同, 碳同位素组成可以用于区分植物的C3和C4光合途径以及不同植物功能群结构变化(Garten Jr. & Taylor Jr., 1992; Peñuelas et al., 1999).通过测定不同碳组分(可溶性碳水化合物和结构性碳库)的δ13C值, 从而估计植物水分利用效率和光合作用在不同时间尺度上的变化(Ebdon et al., 1998; Arslan et al., 1999; 陈世苹等, 2002).而在一个复杂的生态系统(如热带雨林)中, 水分利用效率的高低可能影响植物种间竞争的结果, 最终导致生态系统内部群落结构的差异, 因此δ13C为指示生 ...
Rice rhizodeposition and carbon stabilisation in paddy soil are regulated via drying- rewetting cycles and nitrogen fertilisation
1
2017
... 采用13CO2脉冲标记技术, 对生长在不同SOM含量水稻土上的水稻进行标记, 发现高、低SOM水稻土壤对水稻光合碳的转化和周转具有显著差异, 揭示了水稻光合碳向地下传输过程的“马太效应”(Zhu et al., 2017b).在水稻拔节期和灌浆期内, 通过水稻的根际沉积作用有4%-6%的光合碳进入土壤有机碳库, 土壤可溶性有机碳的2%-4%和土壤微生物生物量碳的9%-18%来源于这部分新碳(Lu et al., 2002a, 2002b; 祝贞科等, 2016).此外, 利用13CO2脉冲标记结合生物标志物(磷脂脂肪酸)的稳定碳同位素探针技术(13C-PLFA-SIP), 发现了碳从水稻向土壤微生物快速转移的现象, 且随着水稻生长期的变化根际碳的微生物同化量显著增加, 真菌及革兰氏阴性菌是主要的同化微生物(Yuan et al., 2016; Liu et al., 2019a).采用13CO2连续标记技术研究了水稻光合碳的传输和转化数量特征及其对氮肥施用的响应, 发现了高氮肥施用条件下水稻光合碳在土壤中的传输和积累比率非线性骤升的规律(Bradford et al., 2008; Ge et al., 2017).结合13C-PLFA-SIP技术发现, 外源C (羧甲基纤维素)和氮肥的联合施用降低了革兰氏阳性菌(G+)和革兰氏阴性菌(G-)的比值, 降低了G+和真菌的丰度, 单施氮肥则刺激了G-和放线菌的增长, 说明可利用碳氮的不同施用策略可显著影响土壤微生物的群落结构(Zhao et al., 2019).Atere等(2017)利用13CO2连续标记技术和土壤有机质的粒径组分分离技术联用, 发现水稻光合碳的固定效率在氮肥施用结合干湿交替条件下有显著提高.连续标记法有助于定量全部生长期或某一生长期全部投入C的净分配, 但是由于成本高以及技术困难, 运用受到限制. ...
Spatial patterns of soil δ 13C reveal grassland-to-woodland successional processes
1
2012
... 碳同位素原子质量的微小差异, 使得含有不同同位素的物质具有不同的理化性质, 导致不同的动力学和热力学效应, 在物理、化学和生物等反应过程中出现同位素分馏现象.碳循环过程中, 同位素交换反应使无机盐(如碳酸盐)富集重同位素(如13C); 而碳的有机循环, 如光合作用的动力分馏效应导致生物成因碳(有机物)中富集轻同位素(如12C), 从而导致陆地生态系统中有机体和大气CO2之间的碳循环过程中, 出现明显的碳同位素分馏现象(Tieszen & Boutton, 1989).植物在光合作用过程中固定CO2途径的差异, 导致不同光合型植物选择吸收13C的比例不同, 从而使C3和C4植株体内的稳定碳同位素比值(δ13C)存在差异, 因此δ13C可以作为表征植物株生理生态特性差异的指标.土壤有机碳(SOC)经过微生物分解矿化为CO2的过程中13C分馏作用较小, 仅为1‰-3‰, 而SOC在厌氧条件下产生甲烷的过程中13C的分馏作用相对较大, 为10‰-70‰ (Conrad et al., 2010; 林光辉, 2013; 袁红朝等, 2014).但SOC主要来源于陆生高等植物, 与植物光合作用固定CO2时的分馏作用相比, SOC分解过程中的分馏作用几乎可以忽略不计, 因此SOC与该土壤上种植植物的δ13C基本一致(Fernandez et al., 2003; Crow et al., 2006; 张林等, 2010; Bai et al., 2012).所以, 通过测定各个碳库中碳同位素比值的变化, 可以量化“大气-植物-土壤”生态系统中碳的来源、分配及其周转规律. ...
Measurement of soil organic matter turnover using 13C natural abundance
2
1996
... 随着碳同位素示踪技术的广泛应用和迅速发展, 多种碳同位素示踪技术及其与其他光谱和微生物探针技术联用, 对于揭示从微观到宏观尺度的碳循环过程起着重要促进作用(图1)(Radajewski et al., 2000; Herrmann et al., 2007).目前自然丰度法、与13C贫化示踪技术结合的自由空气中气体浓度增加(Free-Air Concentration Enrichment, FACE)实验、脉冲与连续标记法以及碳同位素高丰度底物富集标记法是研究陆地生态系统中碳循环过程常用的碳同位素示踪方法(Balesdent & Mariotti, 1996; Bernoux et al., 1998; Conley et al., 2001; Epron et al., 2012; Ge et al., 2017; 张蕊等, 2017).同时, 随着氢、氮(N)、氧等元素同位素示踪技术的发展与应用, 那些单独用碳同位素无法解决的问题在与其他同位素示踪技术、光谱和分子生物学技术的联用下得以解决(陈世苹等, 2002).例如, Kušlienë等(2014)利用13C/15N双标记法, 结合生物标志物分析, 研究了13C/15N标记的不同类型的根系分泌物对土壤碳氮动态的贡献, 这些技术的联用进一步促进了碳同位素示踪技术在生态学研究中的广泛应用.本文重点从稳定碳同位素示踪技术的原理、方法与应用进展等方面综述其在陆地生态系统碳循环过程中的研究进展. ...
... 态系统的功能群多样性以及区分不同功能群植物提供了一条重要途径(Garten Jr & Taylor Jr, 1992; 陈世苹等, 2002).任书杰和于贵瑞(2011)整合了中国区域的数据发现水分条件越好, C3植物叶片δ13C越低, 因此C3植物叶片的δ13C可以作为指示植物的水分利用效率的指标.C3/C4植物交互种植时会改变土壤有机质的δ13C, 因此, 以未交互种植作物土壤为对照, 通过土壤有机碳δ13C的变化可以研究土壤有机碳的周转速度, 以及不同植物来源碳占土壤各碳库和呼吸的比例以及激发效应的方向和强度(Kuzyakov, 2010; Werth & Kuzyakov, 2010; Blagodatskaya et al., 2011a), 也可以通过土壤碳自然丰度δ13C的变化来研究土壤碳库中活性组分碳(微生物生物量碳(MBC))的周转速率和稳定性(Blagodatskaya et al., 2011b).研究结果发现, SOC和MBC的周转时间分别为16.8 a和29-30 d, 且随种植时间增加, 周转时间延长.新碳和老碳对土壤各碳库和呼吸的贡献不同, MBC中20%的碳来源于老碳(来自C3作物), CO2气体中60%的碳来源于老碳(来自C3作物), 由于土壤微生物偏好矿化老碳而同化新碳, SOC中新碳(来自C4作物)的贡献将逐年增加.此外, 长期种植C3作物(小麦(Triticum aestivum))的土壤在连续耕种C4作物(玉米(zea mays)) 13年后, 土壤有机碳库更新了22%, 且SOC的周转速率在不同粒径团聚体中各不相同, 在>50 µm与<2 µm粒径的团聚体中包含更多新碳, 而黏粒中有机碳的更新速度最慢(Balesdent & Mariotti, 1996; 袁红朝等, 2014).13C自然丰度法为明确土壤新老碳库对SOC、MBC和CO2排放的贡献, 解析陆地生态系统δ13C的变化及其在全球碳平衡中的应用提供了重要依据(Cheng, 2009).随着测试精度的提高, 采用碳同位素配对标记实验, 只要SOC和其他一种或是两种碳源的碳同位素比值间有较为明显的差别, 也可以解析多碳源相互作用及其对土壤呼吸的贡献, 这对于研究复杂生态系统中碳通量和净碳平衡具有重要的应用价值(Whitman & Lehmann, 2015). ...
The use of stable carbon isotopes for estimating soil organic matter turnover rates
2
1998
... 随着碳同位素示踪技术的广泛应用和迅速发展, 多种碳同位素示踪技术及其与其他光谱和微生物探针技术联用, 对于揭示从微观到宏观尺度的碳循环过程起着重要促进作用(图1)(Radajewski et al., 2000; Herrmann et al., 2007).目前自然丰度法、与13C贫化示踪技术结合的自由空气中气体浓度增加(Free-Air Concentration Enrichment, FACE)实验、脉冲与连续标记法以及碳同位素高丰度底物富集标记法是研究陆地生态系统中碳循环过程常用的碳同位素示踪方法(Balesdent & Mariotti, 1996; Bernoux et al., 1998; Conley et al., 2001; Epron et al., 2012; Ge et al., 2017; 张蕊等, 2017).同时, 随着氢、氮(N)、氧等元素同位素示踪技术的发展与应用, 那些单独用碳同位素无法解决的问题在与其他同位素示踪技术、光谱和分子生物学技术的联用下得以解决(陈世苹等, 2002).例如, Kušlienë等(2014)利用13C/15N双标记法, 结合生物标志物分析, 研究了13C/15N标记的不同类型的根系分泌物对土壤碳氮动态的贡献, 这些技术的联用进一步促进了碳同位素示踪技术在生态学研究中的广泛应用.本文重点从稳定碳同位素示踪技术的原理、方法与应用进展等方面综述其在陆地生态系统碳循环过程中的研究进展. ...
... 13C同位素示踪法技术可以很好地研究外源C对SOM周转的影响.例如, 将13C标记的有机物加入土壤进行田间或室内培养实验, 通过测定13CO2可以区分外源有机物和原有SOM来源的碳, 量化外源有机物添加SOM分解的影响(Bernoux et al., 1998; Blagodatskaya & Kuzyakov, 2008).高丰度13C标记的有机物结合13C-PLFA-SIP和13C-DNA-SIP技术, 可以准确示踪微生物在土壤有机碳周转过程中的参与机制(Williams et al., 2006; Apostel et al., 2015; Wang et al., 2016).而13C-微生物残留物对于量化和评估微生物对SOM积累的贡献起到了关键作用(Tian et al., 2014; Zhang et al., 2015; Liang et al., 2017). ...
Host-compound foraging by intestinal microbiota revealed by single-cell stable isotope probing
1
2013
... 纳米二次离子质谱(NanoSIMS)能够完成对元素空间分布的超高分辨率显微镜成像, 与同位素示踪技术相结合, 在生态系统微观机制研究领域中显示出了巨大的潜力(Herrmann et al., 2007; Musat et al., 2016).该技术具有较高的灵敏度和准确度, 可应用于碳、氮、硫等元素生物地球化学循环的微生物驱动机制和土壤-微生物-植物相关作用关系的研究, 对于认识多元素耦合的生物地球化学循环规律具有重要意义(图1)(Lechene et al., 2007; Berry et al., 2013; Rumpel et al., 2015).目前, 稳定碳同位素探针-纳米二次离子质谱技术(NanoSIP)已在植物/动物-微生物共生关系(Lechene et al., 2007; Vidal et al., 2016), 微生物群落中碳的食物链传递(Musat et al., 2016), 厌氧甲烷氧化的碳氮耦合机制(Dekas et al., 2009)等研究中取得重要突破.在土壤有机碳周转的研究中, NanoSIP直接呈现了外源碳与土壤矿物的结合形态与动态过程, 有力推动了SOC的稳定机制的研究(Rumpel et al., 2015). ...
Three-source-partitioning of microbial biomass and of CO2 efflux from soil to evaluate mechanisms of priming effects
1
2011a
... 态系统的功能群多样性以及区分不同功能群植物提供了一条重要途径(Garten Jr & Taylor Jr, 1992; 陈世苹等, 2002).任书杰和于贵瑞(2011)整合了中国区域的数据发现水分条件越好, C3植物叶片δ13C越低, 因此C3植物叶片的δ13C可以作为指示植物的水分利用效率的指标.C3/C4植物交互种植时会改变土壤有机质的δ13C, 因此, 以未交互种植作物土壤为对照, 通过土壤有机碳δ13C的变化可以研究土壤有机碳的周转速度, 以及不同植物来源碳占土壤各碳库和呼吸的比例以及激发效应的方向和强度(Kuzyakov, 2010; Werth & Kuzyakov, 2010; Blagodatskaya et al., 2011a), 也可以通过土壤碳自然丰度δ13C的变化来研究土壤碳库中活性组分碳(微生物生物量碳(MBC))的周转速率和稳定性(Blagodatskaya et al., 2011b).研究结果发现, SOC和MBC的周转时间分别为16.8 a和29-30 d, 且随种植时间增加, 周转时间延长.新碳和老碳对土壤各碳库和呼吸的贡献不同, MBC中20%的碳来源于老碳(来自C3作物), CO2气体中60%的碳来源于老碳(来自C3作物), 由于土壤微生物偏好矿化老碳而同化新碳, SOC中新碳(来自C4作物)的贡献将逐年增加.此外, 长期种植C3作物(小麦(Triticum aestivum))的土壤在连续耕种C4作物(玉米(zea mays)) 13年后, 土壤有机碳库更新了22%, 且SOC的周转速率在不同粒径团聚体中各不相同, 在>50 µm与<2 µm粒径的团聚体中包含更多新碳, 而黏粒中有机碳的更新速度最慢(Balesdent & Mariotti, 1996; 袁红朝等, 2014).13C自然丰度法为明确土壤新老碳库对SOC、MBC和CO2排放的贡献, 解析陆地生态系统δ13C的变化及其在全球碳平衡中的应用提供了重要依据(Cheng, 2009).随着测试精度的提高, 采用碳同位素配对标记实验, 只要SOC和其他一种或是两种碳源的碳同位素比值间有较为明显的差别, 也可以解析多碳源相互作用及其对土壤呼吸的贡献, 这对于研究复杂生态系统中碳通量和净碳平衡具有重要的应用价值(Whitman & Lehmann, 2015). ...
Turnover of soil organic matter and of microbial biomass under C3-C4 vegetation change: consideration of 13C fractionation and preferential substrate utilization
1
2011b
... 态系统的功能群多样性以及区分不同功能群植物提供了一条重要途径(Garten Jr & Taylor Jr, 1992; 陈世苹等, 2002).任书杰和于贵瑞(2011)整合了中国区域的数据发现水分条件越好, C3植物叶片δ13C越低, 因此C3植物叶片的δ13C可以作为指示植物的水分利用效率的指标.C3/C4植物交互种植时会改变土壤有机质的δ13C, 因此, 以未交互种植作物土壤为对照, 通过土壤有机碳δ13C的变化可以研究土壤有机碳的周转速度, 以及不同植物来源碳占土壤各碳库和呼吸的比例以及激发效应的方向和强度(Kuzyakov, 2010; Werth & Kuzyakov, 2010; Blagodatskaya et al., 2011a), 也可以通过土壤碳自然丰度δ13C的变化来研究土壤碳库中活性组分碳(微生物生物量碳(MBC))的周转速率和稳定性(Blagodatskaya et al., 2011b).研究结果发现, SOC和MBC的周转时间分别为16.8 a和29-30 d, 且随种植时间增加, 周转时间延长.新碳和老碳对土壤各碳库和呼吸的贡献不同, MBC中20%的碳来源于老碳(来自C3作物), CO2气体中60%的碳来源于老碳(来自C3作物), 由于土壤微生物偏好矿化老碳而同化新碳, SOC中新碳(来自C4作物)的贡献将逐年增加.此外, 长期种植C3作物(小麦(Triticum aestivum))的土壤在连续耕种C4作物(玉米(zea mays)) 13年后, 土壤有机碳库更新了22%, 且SOC的周转速率在不同粒径团聚体中各不相同, 在>50 µm与<2 µm粒径的团聚体中包含更多新碳, 而黏粒中有机碳的更新速度最慢(Balesdent & Mariotti, 1996; 袁红朝等, 2014).13C自然丰度法为明确土壤新老碳库对SOC、MBC和CO2排放的贡献, 解析陆地生态系统δ13C的变化及其在全球碳平衡中的应用提供了重要依据(Cheng, 2009).随着测试精度的提高, 采用碳同位素配对标记实验, 只要SOC和其他一种或是两种碳源的碳同位素比值间有较为明显的差别, 也可以解析多碳源相互作用及其对土壤呼吸的贡献, 这对于研究复杂生态系统中碳通量和净碳平衡具有重要的应用价值(Whitman & Lehmann, 2015). ...
Mechanisms of real and apparent priming effects and their dependence on soil microbial biomass and community structure: critical review
1
2008
... 13C同位素示踪法技术可以很好地研究外源C对SOM周转的影响.例如, 将13C标记的有机物加入土壤进行田间或室内培养实验, 通过测定13CO2可以区分外源有机物和原有SOM来源的碳, 量化外源有机物添加SOM分解的影响(Bernoux et al., 1998; Blagodatskaya & Kuzyakov, 2008).高丰度13C标记的有机物结合13C-PLFA-SIP和13C-DNA-SIP技术, 可以准确示踪微生物在土壤有机碳周转过程中的参与机制(Williams et al., 2006; Apostel et al., 2015; Wang et al., 2016).而13C-微生物残留物对于量化和评估微生物对SOM积累的贡献起到了关键作用(Tian et al., 2014; Zhang et al., 2015; Liang et al., 2017). ...
Stable carbon isotope fractionation as tracer of carbon cycling in anoxic soil ecosystems
1
2016
... 自然界中已知的碳(C)同位素有7种(10C、11C、12C、13C、14C、15C和16C), 其中两种(12C和13C)为稳定碳同位素.不同同位素之间物理化学性质上的差异使反应前和反应后的物质在同位素组成上存在差异, 这种现象称为同位素分馏效应(陈世苹等, 2002; 林光辉, 2013).基于此, 碳同位素示踪技术是研究陆地生态系统碳氮循环过程的有效手段, 也是揭示全球气候变化成因、控制温室效应的重要工具(葛源等, 2006; Blaser & Conrad, 2016).碳同位素技术在研究土壤有机质的来源、动态转化与稳定性等方面具有高度的专一性和灵敏度, 且标记方法已比较成熟, 被广泛地应用到生态学研究中, 由原来在叶片、种群尺度的研究提高到了冠层、群落、生态系统乃至生物圈的水平, 从新的角度研究生物及其周围环境的关系(陈世苹等, 2002).13C稳定同位素作为示踪物, 与放射性碳同位素相比具有安全、无污染和容易控制等优点, 被广泛应用于陆地生态系统碳周转的生物地球化学过程研究. ...
Nonlinear root-derived carbon sequestration across a gradient of nitrogen and phosphorous deposition in experimental mesocosms
1
2008
... 采用13CO2脉冲标记技术, 对生长在不同SOM含量水稻土上的水稻进行标记, 发现高、低SOM水稻土壤对水稻光合碳的转化和周转具有显著差异, 揭示了水稻光合碳向地下传输过程的“马太效应”(Zhu et al., 2017b).在水稻拔节期和灌浆期内, 通过水稻的根际沉积作用有4%-6%的光合碳进入土壤有机碳库, 土壤可溶性有机碳的2%-4%和土壤微生物生物量碳的9%-18%来源于这部分新碳(Lu et al., 2002a, 2002b; 祝贞科等, 2016).此外, 利用13CO2脉冲标记结合生物标志物(磷脂脂肪酸)的稳定碳同位素探针技术(13C-PLFA-SIP), 发现了碳从水稻向土壤微生物快速转移的现象, 且随着水稻生长期的变化根际碳的微生物同化量显著增加, 真菌及革兰氏阴性菌是主要的同化微生物(Yuan et al., 2016; Liu et al., 2019a).采用13CO2连续标记技术研究了水稻光合碳的传输和转化数量特征及其对氮肥施用的响应, 发现了高氮肥施用条件下水稻光合碳在土壤中的传输和积累比率非线性骤升的规律(Bradford et al., 2008; Ge et al., 2017).结合13C-PLFA-SIP技术发现, 外源C (羧甲基纤维素)和氮肥的联合施用降低了革兰氏阳性菌(G+)和革兰氏阴性菌(G-)的比值, 降低了G+和真菌的丰度, 单施氮肥则刺激了G-和放线菌的增长, 说明可利用碳氮的不同施用策略可显著影响土壤微生物的群落结构(Zhao et al., 2019).Atere等(2017)利用13CO2连续标记技术和土壤有机质的粒径组分分离技术联用, 发现水稻光合碳的固定效率在氮肥施用结合干湿交替条件下有显著提高.连续标记法有助于定量全部生长期或某一生长期全部投入C的净分配, 但是由于成本高以及技术困难, 运用受到限制. ...
The effects of elevated [CO2] on plant-soil carbon below-ground: a summary and synthesis
1
1995
... 13C贫化示踪技术结合的FACE实验是研究CO2浓度增加的微域环境中SOC周转的常用方法.大气CO2的δ13C值约为-8‰, 在13C贫化示踪技术结合的FACE实验(13C-FACE)中供给13C贫化的CO2的δ13C值远低于大气, 其范围是-45‰- -18‰ (张蕊等, 2017).FACE系统是一个开放的模拟未来CO2增加的微域环境, 系统内部的气候条件如水分、光照、温度等非常接近自然生态环境.因此, 在这样的系统内进行模拟CO2升高的实验更接近于真实状况(McLeod & Long, 1999).目前全球已建立十余个涉及森林(Hagedorn et al., 2003), 农田(Leavitt et al., 1994)和草地(Six et al., 2001)等多个生态系统和多种植被类型的13C-FACE平台, 阐释了大气CO2浓度升高条件下, 不同生态系统植物生理与光合效率、养分和水分利用、土壤呼吸、土壤碳的输入量及周转率和微生物响应等过程机制(Leavitt et al., 1994; Canadell et al., 1995; Kiem et al., 2000). ...
大气浓度下N2O、CH4和CO2中氮、碳和氧稳定同位素比值的质谱测定
1
2008
... 质谱法、核磁共振法和光谱法是测定稳定碳同位素常用的方法, 质谱法是稳定碳同位素分析中最通用、最精确的方法(Paul et al., 2007; Eiler et al., 2012; 袁红朝等, 2014).质谱法的原理是先将制备的样品转化成含所测元素的纯气体(CO2、H2、N2、SO2等), 经离子源电离, 形成的离子束在电场、磁场的作用下, 导致具有不同质荷比的离子束分开从而检测同位素丰度.普通质谱仪在计算机辅助下直接给出同位素比值, 更先进的仪器如美国热电公司的Thermo Scientific MAT253, 不仅具有更高的灵敏度和更宽的测量范围, 而且可以与同位素数据系统整合, 自动化分析样品的同位素比值.核磁共振法是在磁场恒定条件下, 基于共振时的射频电场频率来测定不同基团上的同位素丰度, 与碳同位素示踪技术相结合可以检测底物(如葡萄糖)在土壤碳循环过程中的化学组分动态(Webster et al., 1997).光谱法是根据组成分子的不同同位素质量数不同从而使红外光谱谱线发生位移的现象来测定碳同位素丰度.近年来, 随着元素生物地球化学循环研究的进一步发展, 以及元素分析仪-同位素质谱(EA-IRMS)和热分析-傅里叶变换红外光谱-气相色谱-质谱(STA-FTIR-GC-MS)等联用技术的兴起, 极大促进了碳同位素分析测试技术的发展(曹亚澄等, 2008; 刘春波等, 2014). ...
大气浓度下N2O、CH4和CO2中氮、碳和氧稳定同位素比值的质谱测定
1
2008
... 质谱法、核磁共振法和光谱法是测定稳定碳同位素常用的方法, 质谱法是稳定碳同位素分析中最通用、最精确的方法(Paul et al., 2007; Eiler et al., 2012; 袁红朝等, 2014).质谱法的原理是先将制备的样品转化成含所测元素的纯气体(CO2、H2、N2、SO2等), 经离子源电离, 形成的离子束在电场、磁场的作用下, 导致具有不同质荷比的离子束分开从而检测同位素丰度.普通质谱仪在计算机辅助下直接给出同位素比值, 更先进的仪器如美国热电公司的Thermo Scientific MAT253, 不仅具有更高的灵敏度和更宽的测量范围, 而且可以与同位素数据系统整合, 自动化分析样品的同位素比值.核磁共振法是在磁场恒定条件下, 基于共振时的射频电场频率来测定不同基团上的同位素丰度, 与碳同位素示踪技术相结合可以检测底物(如葡萄糖)在土壤碳循环过程中的化学组分动态(Webster et al., 1997).光谱法是根据组成分子的不同同位素质量数不同从而使红外光谱谱线发生位移的现象来测定碳同位素丰度.近年来, 随着元素生物地球化学循环研究的进一步发展, 以及元素分析仪-同位素质谱(EA-IRMS)和热分析-傅里叶变换红外光谱-气相色谱-质谱(STA-FTIR-GC-MS)等联用技术的兴起, 极大促进了碳同位素分析测试技术的发展(曹亚澄等, 2008; 刘春波等, 2014). ...
稳定性碳同位素技术在生态学研究中的应用
5
2002
... 自然界中已知的碳(C)同位素有7种(10C、11C、12C、13C、14C、15C和16C), 其中两种(12C和13C)为稳定碳同位素.不同同位素之间物理化学性质上的差异使反应前和反应后的物质在同位素组成上存在差异, 这种现象称为同位素分馏效应(陈世苹等, 2002; 林光辉, 2013).基于此, 碳同位素示踪技术是研究陆地生态系统碳氮循环过程的有效手段, 也是揭示全球气候变化成因、控制温室效应的重要工具(葛源等, 2006; Blaser & Conrad, 2016).碳同位素技术在研究土壤有机质的来源、动态转化与稳定性等方面具有高度的专一性和灵敏度, 且标记方法已比较成熟, 被广泛地应用到生态学研究中, 由原来在叶片、种群尺度的研究提高到了冠层、群落、生态系统乃至生物圈的水平, 从新的角度研究生物及其周围环境的关系(陈世苹等, 2002).13C稳定同位素作为示踪物, 与放射性碳同位素相比具有安全、无污染和容易控制等优点, 被广泛应用于陆地生态系统碳周转的生物地球化学过程研究. ...
... ).碳同位素技术在研究土壤有机质的来源、动态转化与稳定性等方面具有高度的专一性和灵敏度, 且标记方法已比较成熟, 被广泛地应用到生态学研究中, 由原来在叶片、种群尺度的研究提高到了冠层、群落、生态系统乃至生物圈的水平, 从新的角度研究生物及其周围环境的关系(陈世苹等, 2002).13C稳定同位素作为示踪物, 与放射性碳同位素相比具有安全、无污染和容易控制等优点, 被广泛应用于陆地生态系统碳周转的生物地球化学过程研究. ...
... 随着碳同位素示踪技术的广泛应用和迅速发展, 多种碳同位素示踪技术及其与其他光谱和微生物探针技术联用, 对于揭示从微观到宏观尺度的碳循环过程起着重要促进作用(图1)(Radajewski et al., 2000; Herrmann et al., 2007).目前自然丰度法、与13C贫化示踪技术结合的自由空气中气体浓度增加(Free-Air Concentration Enrichment, FACE)实验、脉冲与连续标记法以及碳同位素高丰度底物富集标记法是研究陆地生态系统中碳循环过程常用的碳同位素示踪方法(Balesdent & Mariotti, 1996; Bernoux et al., 1998; Conley et al., 2001; Epron et al., 2012; Ge et al., 2017; 张蕊等, 2017).同时, 随着氢、氮(N)、氧等元素同位素示踪技术的发展与应用, 那些单独用碳同位素无法解决的问题在与其他同位素示踪技术、光谱和分子生物学技术的联用下得以解决(陈世苹等, 2002).例如, Kušlienë等(2014)利用13C/15N双标记法, 结合生物标志物分析, 研究了13C/15N标记的不同类型的根系分泌物对土壤碳氮动态的贡献, 这些技术的联用进一步促进了碳同位素示踪技术在生态学研究中的广泛应用.本文重点从稳定碳同位素示踪技术的原理、方法与应用进展等方面综述其在陆地生态系统碳循环过程中的研究进展. ...
... 基于C3/C4植物δ13C的不同, 碳同位素组成可以用于区分植物的C3和C4光合途径以及不同植物功能群结构变化(Garten Jr. & Taylor Jr., 1992; Peñuelas et al., 1999).通过测定不同碳组分(可溶性碳水化合物和结构性碳库)的δ13C值, 从而估计植物水分利用效率和光合作用在不同时间尺度上的变化(Ebdon et al., 1998; Arslan et al., 1999; 陈世苹等, 2002).而在一个复杂的生态系统(如热带雨林)中, 水分利用效率的高低可能影响植物种间竞争的结果, 最终导致生态系统内部群落结构的差异, 因此δ13C为指示生 ...
... 态系统的功能群多样性以及区分不同功能群植物提供了一条重要途径(Garten Jr & Taylor Jr, 1992; 陈世苹等, 2002).任书杰和于贵瑞(2011)整合了中国区域的数据发现水分条件越好, C3植物叶片δ13C越低, 因此C3植物叶片的δ13C可以作为指示植物的水分利用效率的指标.C3/C4植物交互种植时会改变土壤有机质的δ13C, 因此, 以未交互种植作物土壤为对照, 通过土壤有机碳δ13C的变化可以研究土壤有机碳的周转速度, 以及不同植物来源碳占土壤各碳库和呼吸的比例以及激发效应的方向和强度(Kuzyakov, 2010; Werth & Kuzyakov, 2010; Blagodatskaya et al., 2011a), 也可以通过土壤碳自然丰度δ13C的变化来研究土壤碳库中活性组分碳(微生物生物量碳(MBC))的周转速率和稳定性(Blagodatskaya et al., 2011b).研究结果发现, SOC和MBC的周转时间分别为16.8 a和29-30 d, 且随种植时间增加, 周转时间延长.新碳和老碳对土壤各碳库和呼吸的贡献不同, MBC中20%的碳来源于老碳(来自C3作物), CO2气体中60%的碳来源于老碳(来自C3作物), 由于土壤微生物偏好矿化老碳而同化新碳, SOC中新碳(来自C4作物)的贡献将逐年增加.此外, 长期种植C3作物(小麦(Triticum aestivum))的土壤在连续耕种C4作物(玉米(zea mays)) 13年后, 土壤有机碳库更新了22%, 且SOC的周转速率在不同粒径团聚体中各不相同, 在>50 µm与<2 µm粒径的团聚体中包含更多新碳, 而黏粒中有机碳的更新速度最慢(Balesdent & Mariotti, 1996; 袁红朝等, 2014).13C自然丰度法为明确土壤新老碳库对SOC、MBC和CO2排放的贡献, 解析陆地生态系统δ13C的变化及其在全球碳平衡中的应用提供了重要依据(Cheng, 2009).随着测试精度的提高, 采用碳同位素配对标记实验, 只要SOC和其他一种或是两种碳源的碳同位素比值间有较为明显的差别, 也可以解析多碳源相互作用及其对土壤呼吸的贡献, 这对于研究复杂生态系统中碳通量和净碳平衡具有重要的应用价值(Whitman & Lehmann, 2015). ...
稳定性碳同位素技术在生态学研究中的应用
5
2002
... 自然界中已知的碳(C)同位素有7种(10C、11C、12C、13C、14C、15C和16C), 其中两种(12C和13C)为稳定碳同位素.不同同位素之间物理化学性质上的差异使反应前和反应后的物质在同位素组成上存在差异, 这种现象称为同位素分馏效应(陈世苹等, 2002; 林光辉, 2013).基于此, 碳同位素示踪技术是研究陆地生态系统碳氮循环过程的有效手段, 也是揭示全球气候变化成因、控制温室效应的重要工具(葛源等, 2006; Blaser & Conrad, 2016).碳同位素技术在研究土壤有机质的来源、动态转化与稳定性等方面具有高度的专一性和灵敏度, 且标记方法已比较成熟, 被广泛地应用到生态学研究中, 由原来在叶片、种群尺度的研究提高到了冠层、群落、生态系统乃至生物圈的水平, 从新的角度研究生物及其周围环境的关系(陈世苹等, 2002).13C稳定同位素作为示踪物, 与放射性碳同位素相比具有安全、无污染和容易控制等优点, 被广泛应用于陆地生态系统碳周转的生物地球化学过程研究. ...
... ).碳同位素技术在研究土壤有机质的来源、动态转化与稳定性等方面具有高度的专一性和灵敏度, 且标记方法已比较成熟, 被广泛地应用到生态学研究中, 由原来在叶片、种群尺度的研究提高到了冠层、群落、生态系统乃至生物圈的水平, 从新的角度研究生物及其周围环境的关系(陈世苹等, 2002).13C稳定同位素作为示踪物, 与放射性碳同位素相比具有安全、无污染和容易控制等优点, 被广泛应用于陆地生态系统碳周转的生物地球化学过程研究. ...
... 随着碳同位素示踪技术的广泛应用和迅速发展, 多种碳同位素示踪技术及其与其他光谱和微生物探针技术联用, 对于揭示从微观到宏观尺度的碳循环过程起着重要促进作用(图1)(Radajewski et al., 2000; Herrmann et al., 2007).目前自然丰度法、与13C贫化示踪技术结合的自由空气中气体浓度增加(Free-Air Concentration Enrichment, FACE)实验、脉冲与连续标记法以及碳同位素高丰度底物富集标记法是研究陆地生态系统中碳循环过程常用的碳同位素示踪方法(Balesdent & Mariotti, 1996; Bernoux et al., 1998; Conley et al., 2001; Epron et al., 2012; Ge et al., 2017; 张蕊等, 2017).同时, 随着氢、氮(N)、氧等元素同位素示踪技术的发展与应用, 那些单独用碳同位素无法解决的问题在与其他同位素示踪技术、光谱和分子生物学技术的联用下得以解决(陈世苹等, 2002).例如, Kušlienë等(2014)利用13C/15N双标记法, 结合生物标志物分析, 研究了13C/15N标记的不同类型的根系分泌物对土壤碳氮动态的贡献, 这些技术的联用进一步促进了碳同位素示踪技术在生态学研究中的广泛应用.本文重点从稳定碳同位素示踪技术的原理、方法与应用进展等方面综述其在陆地生态系统碳循环过程中的研究进展. ...
... 基于C3/C4植物δ13C的不同, 碳同位素组成可以用于区分植物的C3和C4光合途径以及不同植物功能群结构变化(Garten Jr. & Taylor Jr., 1992; Peñuelas et al., 1999).通过测定不同碳组分(可溶性碳水化合物和结构性碳库)的δ13C值, 从而估计植物水分利用效率和光合作用在不同时间尺度上的变化(Ebdon et al., 1998; Arslan et al., 1999; 陈世苹等, 2002).而在一个复杂的生态系统(如热带雨林)中, 水分利用效率的高低可能影响植物种间竞争的结果, 最终导致生态系统内部群落结构的差异, 因此δ13C为指示生 ...
... 态系统的功能群多样性以及区分不同功能群植物提供了一条重要途径(Garten Jr & Taylor Jr, 1992; 陈世苹等, 2002).任书杰和于贵瑞(2011)整合了中国区域的数据发现水分条件越好, C3植物叶片δ13C越低, 因此C3植物叶片的δ13C可以作为指示植物的水分利用效率的指标.C3/C4植物交互种植时会改变土壤有机质的δ13C, 因此, 以未交互种植作物土壤为对照, 通过土壤有机碳δ13C的变化可以研究土壤有机碳的周转速度, 以及不同植物来源碳占土壤各碳库和呼吸的比例以及激发效应的方向和强度(Kuzyakov, 2010; Werth & Kuzyakov, 2010; Blagodatskaya et al., 2011a), 也可以通过土壤碳自然丰度δ13C的变化来研究土壤碳库中活性组分碳(微生物生物量碳(MBC))的周转速率和稳定性(Blagodatskaya et al., 2011b).研究结果发现, SOC和MBC的周转时间分别为16.8 a和29-30 d, 且随种植时间增加, 周转时间延长.新碳和老碳对土壤各碳库和呼吸的贡献不同, MBC中20%的碳来源于老碳(来自C3作物), CO2气体中60%的碳来源于老碳(来自C3作物), 由于土壤微生物偏好矿化老碳而同化新碳, SOC中新碳(来自C4作物)的贡献将逐年增加.此外, 长期种植C3作物(小麦(Triticum aestivum))的土壤在连续耕种C4作物(玉米(zea mays)) 13年后, 土壤有机碳库更新了22%, 且SOC的周转速率在不同粒径团聚体中各不相同, 在>50 µm与<2 µm粒径的团聚体中包含更多新碳, 而黏粒中有机碳的更新速度最慢(Balesdent & Mariotti, 1996; 袁红朝等, 2014).13C自然丰度法为明确土壤新老碳库对SOC、MBC和CO2排放的贡献, 解析陆地生态系统δ13C的变化及其在全球碳平衡中的应用提供了重要依据(Cheng, 2009).随着测试精度的提高, 采用碳同位素配对标记实验, 只要SOC和其他一种或是两种碳源的碳同位素比值间有较为明显的差别, 也可以解析多碳源相互作用及其对土壤呼吸的贡献, 这对于研究复杂生态系统中碳通量和净碳平衡具有重要的应用价值(Whitman & Lehmann, 2015). ...
Rhizosphere priming effect: its functional relationships with microbial turnover, evapotranspiration, and C-N budgets
1
2009
... 态系统的功能群多样性以及区分不同功能群植物提供了一条重要途径(Garten Jr & Taylor Jr, 1992; 陈世苹等, 2002).任书杰和于贵瑞(2011)整合了中国区域的数据发现水分条件越好, C3植物叶片δ13C越低, 因此C3植物叶片的δ13C可以作为指示植物的水分利用效率的指标.C3/C4植物交互种植时会改变土壤有机质的δ13C, 因此, 以未交互种植作物土壤为对照, 通过土壤有机碳δ13C的变化可以研究土壤有机碳的周转速度, 以及不同植物来源碳占土壤各碳库和呼吸的比例以及激发效应的方向和强度(Kuzyakov, 2010; Werth & Kuzyakov, 2010; Blagodatskaya et al., 2011a), 也可以通过土壤碳自然丰度δ13C的变化来研究土壤碳库中活性组分碳(微生物生物量碳(MBC))的周转速率和稳定性(Blagodatskaya et al., 2011b).研究结果发现, SOC和MBC的周转时间分别为16.8 a和29-30 d, 且随种植时间增加, 周转时间延长.新碳和老碳对土壤各碳库和呼吸的贡献不同, MBC中20%的碳来源于老碳(来自C3作物), CO2气体中60%的碳来源于老碳(来自C3作物), 由于土壤微生物偏好矿化老碳而同化新碳, SOC中新碳(来自C4作物)的贡献将逐年增加.此外, 长期种植C3作物(小麦(Triticum aestivum))的土壤在连续耕种C4作物(玉米(zea mays)) 13年后, 土壤有机碳库更新了22%, 且SOC的周转速率在不同粒径团聚体中各不相同, 在>50 µm与<2 µm粒径的团聚体中包含更多新碳, 而黏粒中有机碳的更新速度最慢(Balesdent & Mariotti, 1996; 袁红朝等, 2014).13C自然丰度法为明确土壤新老碳库对SOC、MBC和CO2排放的贡献, 解析陆地生态系统δ13C的变化及其在全球碳平衡中的应用提供了重要依据(Cheng, 2009).随着测试精度的提高, 采用碳同位素配对标记实验, 只要SOC和其他一种或是两种碳源的碳同位素比值间有较为明显的差别, 也可以解析多碳源相互作用及其对土壤呼吸的贡献, 这对于研究复杂生态系统中碳通量和净碳平衡具有重要的应用价值(Whitman & Lehmann, 2015). ...
CO2 enrichment increases water-use efficiency in sorghum
1
2001
... 随着碳同位素示踪技术的广泛应用和迅速发展, 多种碳同位素示踪技术及其与其他光谱和微生物探针技术联用, 对于揭示从微观到宏观尺度的碳循环过程起着重要促进作用(图1)(Radajewski et al., 2000; Herrmann et al., 2007).目前自然丰度法、与13C贫化示踪技术结合的自由空气中气体浓度增加(Free-Air Concentration Enrichment, FACE)实验、脉冲与连续标记法以及碳同位素高丰度底物富集标记法是研究陆地生态系统中碳循环过程常用的碳同位素示踪方法(Balesdent & Mariotti, 1996; Bernoux et al., 1998; Conley et al., 2001; Epron et al., 2012; Ge et al., 2017; 张蕊等, 2017).同时, 随着氢、氮(N)、氧等元素同位素示踪技术的发展与应用, 那些单独用碳同位素无法解决的问题在与其他同位素示踪技术、光谱和分子生物学技术的联用下得以解决(陈世苹等, 2002).例如, Kušlienë等(2014)利用13C/15N双标记法, 结合生物标志物分析, 研究了13C/15N标记的不同类型的根系分泌物对土壤碳氮动态的贡献, 这些技术的联用进一步促进了碳同位素示踪技术在生态学研究中的广泛应用.本文重点从稳定碳同位素示踪技术的原理、方法与应用进展等方面综述其在陆地生态系统碳循环过程中的研究进展. ...
Methanogenic pathway, 13C isotope fractionation, and archaeal community composition in the sediment of two clear- water lakes of Amazonia
1
2010
... 碳同位素原子质量的微小差异, 使得含有不同同位素的物质具有不同的理化性质, 导致不同的动力学和热力学效应, 在物理、化学和生物等反应过程中出现同位素分馏现象.碳循环过程中, 同位素交换反应使无机盐(如碳酸盐)富集重同位素(如13C); 而碳的有机循环, 如光合作用的动力分馏效应导致生物成因碳(有机物)中富集轻同位素(如12C), 从而导致陆地生态系统中有机体和大气CO2之间的碳循环过程中, 出现明显的碳同位素分馏现象(Tieszen & Boutton, 1989).植物在光合作用过程中固定CO2途径的差异, 导致不同光合型植物选择吸收13C的比例不同, 从而使C3和C4植株体内的稳定碳同位素比值(δ13C)存在差异, 因此δ13C可以作为表征植物株生理生态特性差异的指标.土壤有机碳(SOC)经过微生物分解矿化为CO2的过程中13C分馏作用较小, 仅为1‰-3‰, 而SOC在厌氧条件下产生甲烷的过程中13C的分馏作用相对较大, 为10‰-70‰ (Conrad et al., 2010; 林光辉, 2013; 袁红朝等, 2014).但SOC主要来源于陆生高等植物, 与植物光合作用固定CO2时的分馏作用相比, SOC分解过程中的分馏作用几乎可以忽略不计, 因此SOC与该土壤上种植植物的δ13C基本一致(Fernandez et al., 2003; Crow et al., 2006; 张林等, 2010; Bai et al., 2012).所以, 通过测定各个碳库中碳同位素比值的变化, 可以量化“大气-植物-土壤”生态系统中碳的来源、分配及其周转规律. ...
Stable carbon isotope fractionation, carbon flux partitioning and priming effects in anoxic soils during methanogenic degradation of straw and soil organic matter
1
2012
... 13CO2同位素标记法, 是指以高丰度13CO2为示踪物, 在植物通过光合作用固定这些示踪物后, 通过在特定的时间内测定植物-土壤系统中各碳库示踪物量的变化, 可定量研究该时间段内13C在植物-土壤系统的转移和分布, 阐明植物某一生长期的碳分配特征, 定量植物碳向土壤的输入以及土壤碳周转, 从而明确植物光合碳对SOM的来源、稳定化过程的影响及其微生物驱动机制(Hanson et al., 2000; Kuzyakov & Cheng, 2001; Kuzyakov, 2006; Yuan et al., 2016; 张蕊等, 2017; Liu et al., 2019b).脉冲标记法是一次或分几次加入13CO2, 标记数小时后在特定时间内测定13C在土壤-植物系统各碳库的分配比例(Kuzyakov & Gavrichkova, 2010)(表1).脉冲标记法只能提供植物生长关键时期光合碳的分配特征, 但是在植物生长期内标记13CO2, 就可以估计植物来源碳输入土壤各组分的量(Lu et al., 2002a; Kimura et al., 2004; Xiao et al., 2019a, 2019b; Zang et al., 2019).连续标记法是指在植物生长某个生长期(或全生长期), 在可控气室等封闭的空间内对植物进行不间断标记, 可以定量全部生长期投入碳的净分配(表1).例如, 在水稻(Oryza satira)拔节期和灌浆期进行13CO2连续标记, 可以定量研究生长过程中水稻光合碳对SOC各组分的贡献(Kuzyakov, 2005; Conrad et al., 2012; Wang et al., 2016), 以及对土壤原有有机质的激发效应(Zhu et al., 2018a). ...
Formation of soil organic matter via biochemical and physical pathways of litter mass loss
1
2015
... SOC周转是一个连续动态变化的过程, 利用碳同位素技术能够示踪新碳在土壤中的转化过程和赋存状态, 揭示外源碳在土壤-微生物之间的周转过程和机理(金鑫鑫等, 2017).利用13C富集标记小分子有机物或是植物秸秆, 结合生物标志物技术等, 可以表征不同来源的有机质在土壤中的相对丰度和降解状况, 以及SOC的动态变化和储量的转化和迁移, 也可以定量评价新老SOC对碳储量的相对贡献, 可有效评估陆地生态系统碳平衡过程及其微观过程机制(Kuzyakov & Domanski, 2000; Kemmitt et al., 2008; Ge et al., 2017; Zhu et al., 2017a).外源C输入土壤后可以通过物理和生物化学途径形成新的SOM (Cotrufo et al., 2013, 2015), 从而增加SOC库; 另一方面, 外源C的输入也会促进土壤中原有SOM降解(Guenet et al., 2018; Zhu et al., 2018b; Perveen et al., 2019), 从而减少SOC库.因此, 这两个过程之间的平衡对SOC库变化的方向和程度起到至关重要的作用(Liang et al., 2018). ...
The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter?
1
2013
... SOC周转是一个连续动态变化的过程, 利用碳同位素技术能够示踪新碳在土壤中的转化过程和赋存状态, 揭示外源碳在土壤-微生物之间的周转过程和机理(金鑫鑫等, 2017).利用13C富集标记小分子有机物或是植物秸秆, 结合生物标志物技术等, 可以表征不同来源的有机质在土壤中的相对丰度和降解状况, 以及SOC的动态变化和储量的转化和迁移, 也可以定量评价新老SOC对碳储量的相对贡献, 可有效评估陆地生态系统碳平衡过程及其微观过程机制(Kuzyakov & Domanski, 2000; Kemmitt et al., 2008; Ge et al., 2017; Zhu et al., 2017a).外源C输入土壤后可以通过物理和生物化学途径形成新的SOM (Cotrufo et al., 2013, 2015), 从而增加SOC库; 另一方面, 外源C的输入也会促进土壤中原有SOM降解(Guenet et al., 2018; Zhu et al., 2018b; Perveen et al., 2019), 从而减少SOC库.因此, 这两个过程之间的平衡对SOC库变化的方向和程度起到至关重要的作用(Liang et al., 2018). ...
Isotopic analysis of respired CO2 during decomposition of separated soil organic matter pools
1
2006
... 碳同位素原子质量的微小差异, 使得含有不同同位素的物质具有不同的理化性质, 导致不同的动力学和热力学效应, 在物理、化学和生物等反应过程中出现同位素分馏现象.碳循环过程中, 同位素交换反应使无机盐(如碳酸盐)富集重同位素(如13C); 而碳的有机循环, 如光合作用的动力分馏效应导致生物成因碳(有机物)中富集轻同位素(如12C), 从而导致陆地生态系统中有机体和大气CO2之间的碳循环过程中, 出现明显的碳同位素分馏现象(Tieszen & Boutton, 1989).植物在光合作用过程中固定CO2途径的差异, 导致不同光合型植物选择吸收13C的比例不同, 从而使C3和C4植株体内的稳定碳同位素比值(δ13C)存在差异, 因此δ13C可以作为表征植物株生理生态特性差异的指标.土壤有机碳(SOC)经过微生物分解矿化为CO2的过程中13C分馏作用较小, 仅为1‰-3‰, 而SOC在厌氧条件下产生甲烷的过程中13C的分馏作用相对较大, 为10‰-70‰ (Conrad et al., 2010; 林光辉, 2013; 袁红朝等, 2014).但SOC主要来源于陆生高等植物, 与植物光合作用固定CO2时的分馏作用相比, SOC分解过程中的分馏作用几乎可以忽略不计, 因此SOC与该土壤上种植植物的δ13C基本一致(Fernandez et al., 2003; Crow et al., 2006; 张林等, 2010; Bai et al., 2012).所以, 通过测定各个碳库中碳同位素比值的变化, 可以量化“大气-植物-土壤”生态系统中碳的来源、分配及其周转规律. ...
Deep-sea archaea fix and share nitrogen in methane-consuming microbial consortia
1
2009
... 纳米二次离子质谱(NanoSIMS)能够完成对元素空间分布的超高分辨率显微镜成像, 与同位素示踪技术相结合, 在生态系统微观机制研究领域中显示出了巨大的潜力(Herrmann et al., 2007; Musat et al., 2016).该技术具有较高的灵敏度和准确度, 可应用于碳、氮、硫等元素生物地球化学循环的微生物驱动机制和土壤-微生物-植物相关作用关系的研究, 对于认识多元素耦合的生物地球化学循环规律具有重要意义(图1)(Lechene et al., 2007; Berry et al., 2013; Rumpel et al., 2015).目前, 稳定碳同位素探针-纳米二次离子质谱技术(NanoSIP)已在植物/动物-微生物共生关系(Lechene et al., 2007; Vidal et al., 2016), 微生物群落中碳的食物链传递(Musat et al., 2016), 厌氧甲烷氧化的碳氮耦合机制(Dekas et al., 2009)等研究中取得重要突破.在土壤有机碳周转的研究中, NanoSIP直接呈现了外源碳与土壤矿物的结合形态与动态过程, 有力推动了SOC的稳定机制的研究(Rumpel et al., 2015). ...
C and N in soil organic matter density fractions under elevated atmospheric CO2: turnover vs. stabilization
1
2011
... 在草地生态系统中, FACE实验研究发现CO2浓度升高, 黑麦草(Lolium perenne)草地0-10 cm 土层土壤团聚体中的颗粒有机碳增加了40%, 表明大气CO2浓度升高可促进草地生态系统的碳周转(Six et al., 2001).在森林生态系统中, Griepentrog等(2015)也发现, CO2浓度升高增加了C3植物向土壤碳的输入量, 提高了土壤碳的周转率.新碳在SOM中的积累程度受有机质密度的影响, 有机质密度增加, 新碳的积累程度减小, 大部分的新碳进入到颗粒态有机质中(Dorodnikov et al., 2011; Hofmockel et al., 2011).同时, 13C-FACE结合PLFA技术, 进一步揭示了CO2浓度升高条件下革兰氏阴性菌和真菌可矿化更多的植物来源碳, 促进了植物新合成的光合碳的分解, 也加快了SOM的分解速率(Pendall et al., 2003; Hagedorn et al., 2013). ...
Identification of a complete methane monooxygenase operon from soil by combining stable isotope probing and metagenomic analysis
1
2006
... 尽管PFLA-SIP、DNA/RNA-SIP技术有力推动了微生物群落动态与生态系统功能的关联研究, 但生物地球化学循环微生物驱动机制的深入揭示仍需要稳定碳同位素示踪技术与多组学技术的结合.稳定碳同位素探针-宏基因组/宏转录组技术能够揭示特定生态过程中活性微生物群落组成、功能多样性、生态网络关系等, 实现对碳氮等元素微生物周转过程的代谢通路预测和重构(Dumont et al., 2006; Neufeld et al., 2008).宏基因组和宏转录组整合分析了代谢过程的可能性, 蛋白质组学、代谢组学技术则为特定底物的代谢通路的建立提供直接依据, 是生态系统中生物地球化学循环过程完整解析的必经之路. ...
Relationship between carbon isotope discrimination, water use efficiency, and evapotranspiration in Kentucky bluegrass
1
1998
... 基于C3/C4植物δ13C的不同, 碳同位素组成可以用于区分植物的C3和C4光合途径以及不同植物功能群结构变化(Garten Jr. & Taylor Jr., 1992; Peñuelas et al., 1999).通过测定不同碳组分(可溶性碳水化合物和结构性碳库)的δ13C值, 从而估计植物水分利用效率和光合作用在不同时间尺度上的变化(Ebdon et al., 1998; Arslan et al., 1999; 陈世苹等, 2002).而在一个复杂的生态系统(如热带雨林)中, 水分利用效率的高低可能影响植物种间竞争的结果, 最终导致生态系统内部群落结构的差异, 因此δ13C为指示生 ...
Variation in quantum yield for CO2 uptake among C3 and C4 plants
1
1983
... 不同光合型植物光合作用过程中固定CO2途径的差异, 导致其对13C选择吸收的比例不同, 从而使不同植物的δ13C存在差异.大气CO2经植物气孔向叶片内扩散, 使进入叶肉细胞间的CO2比叶外大气CO2具有更低的δ13C值; 而CO2溶于水的过程中, 植物优先从叶胞间空气吸收12CO2使之溶于细胞质中, 使空气中CO2浓度越高分馏效应越大; 此外, 光合作用过程中, 光合羧化酶同化CO2, 使细胞质中的12CO2通过酶的作用优先结合到磷酸甘油酸中, 合成的光合产物贫化13C丰度.所以, 由于不同植被类型光合作用过程中的碳同位素分馏过程不同, 而具有不同的δ13C值, 通常C3植物(光合作用中同化CO2的最初产物是三碳化合物3-磷酸甘油酸的植物) δ13C的变化范围为-20‰-35‰, C4植物(CO2同化的最初产物是四碳化合物苹果酸或天门冬氨酸的植物) δ13C的变化范围为-17‰- -9‰ (Ehleringer & Pearcy, 1983).碳同位素自然丰度比值法在植物光合作用途径、C3/C4植被变化、土壤碳循环、全球碳平衡等研究中得到日益广泛的应用. ...
A high-resolution gas-source isotope ratio mass spectrometer
1
2012
... 质谱法、核磁共振法和光谱法是测定稳定碳同位素常用的方法, 质谱法是稳定碳同位素分析中最通用、最精确的方法(Paul et al., 2007; Eiler et al., 2012; 袁红朝等, 2014).质谱法的原理是先将制备的样品转化成含所测元素的纯气体(CO2、H2、N2、SO2等), 经离子源电离, 形成的离子束在电场、磁场的作用下, 导致具有不同质荷比的离子束分开从而检测同位素丰度.普通质谱仪在计算机辅助下直接给出同位素比值, 更先进的仪器如美国热电公司的Thermo Scientific MAT253, 不仅具有更高的灵敏度和更宽的测量范围, 而且可以与同位素数据系统整合, 自动化分析样品的同位素比值.核磁共振法是在磁场恒定条件下, 基于共振时的射频电场频率来测定不同基团上的同位素丰度, 与碳同位素示踪技术相结合可以检测底物(如葡萄糖)在土壤碳循环过程中的化学组分动态(Webster et al., 1997).光谱法是根据组成分子的不同同位素质量数不同从而使红外光谱谱线发生位移的现象来测定碳同位素丰度.近年来, 随着元素生物地球化学循环研究的进一步发展, 以及元素分析仪-同位素质谱(EA-IRMS)和热分析-傅里叶变换红外光谱-气相色谱-质谱(STA-FTIR-GC-MS)等联用技术的兴起, 极大促进了碳同位素分析测试技术的发展(曹亚澄等, 2008; 刘春波等, 2014). ...
Pulse-labelling trees to study carbon allocation dynamics: a review of methods, current knowledge and future prospects
1
2012
... 随着碳同位素示踪技术的广泛应用和迅速发展, 多种碳同位素示踪技术及其与其他光谱和微生物探针技术联用, 对于揭示从微观到宏观尺度的碳循环过程起着重要促进作用(图1)(Radajewski et al., 2000; Herrmann et al., 2007).目前自然丰度法、与13C贫化示踪技术结合的自由空气中气体浓度增加(Free-Air Concentration Enrichment, FACE)实验、脉冲与连续标记法以及碳同位素高丰度底物富集标记法是研究陆地生态系统中碳循环过程常用的碳同位素示踪方法(Balesdent & Mariotti, 1996; Bernoux et al., 1998; Conley et al., 2001; Epron et al., 2012; Ge et al., 2017; 张蕊等, 2017).同时, 随着氢、氮(N)、氧等元素同位素示踪技术的发展与应用, 那些单独用碳同位素无法解决的问题在与其他同位素示踪技术、光谱和分子生物学技术的联用下得以解决(陈世苹等, 2002).例如, Kušlienë等(2014)利用13C/15N双标记法, 结合生物标志物分析, 研究了13C/15N标记的不同类型的根系分泌物对土壤碳氮动态的贡献, 这些技术的联用进一步促进了碳同位素示踪技术在生态学研究中的广泛应用.本文重点从稳定碳同位素示踪技术的原理、方法与应用进展等方面综述其在陆地生态系统碳循环过程中的研究进展. ...
Probing potential microbial coupling of carbon and nitrogen cycling during decomposition of maize residue by 13C-DNA-SIP
1
2014
... 将稳定碳同位素示踪技术与分子生物学方法相结合, 在表征以微生物为主要研究对象的土壤生物群落功能过程研究中发挥了重要作用(Lu & Conrad, 2005; Ruyters et al., 2013; Fan et al., 2014).13C同位素探针-磷脂脂肪酸(PLFA-SIP)是在土壤碳循环微生物驱动机制的研究中广泛应用的技术.该方法灵敏度高, 常用于指示活体微生物生物量和群落结构, 能够定量检测微生物对标记底物的同化利用.近年来, 以氨基糖为主要成分的微生物残留物的相对稳定性及其对SOC的贡献得到广泛关注(Liang et al., 2017), 将PLFA-SIP与氨基糖生物标志物相结合, 建立外源碳-微生物-土壤碳库的碳周转连续体模型, 有助于揭示SOC的累积机制.PLFA-SIP对物种组成的分辨率较低, 难以判断功能微生物, 且无法检测古菌.以DNA/RNA测序为基础的DNA/RNA-SIP技术可以很好地解决上述问题, 只要选择合适的底物和目的基因, DNA/RNA-SIP技术能准确解析关键微生物在土壤中的群落结构和功能变化, 特异性地将微生物群体与生态过程、生理特性相关联, 揭示微生物驱动下的土壤元素循环过程及其调控机制.近年来, Hungate等(2015)将13C与18O标记相结合, 建立了定量DNA-SIP技术(DNA-qSIP), 揭示了参与土壤中新输入的有机碳(新碳)和土壤原有有机碳(老 碳)周转过程的微生物群落差异(Morrissey et al., 2017). ...
Carbon isotopic fractionation during decomposition of plant materials of different quality
1
2003
... 碳同位素原子质量的微小差异, 使得含有不同同位素的物质具有不同的理化性质, 导致不同的动力学和热力学效应, 在物理、化学和生物等反应过程中出现同位素分馏现象.碳循环过程中, 同位素交换反应使无机盐(如碳酸盐)富集重同位素(如13C); 而碳的有机循环, 如光合作用的动力分馏效应导致生物成因碳(有机物)中富集轻同位素(如12C), 从而导致陆地生态系统中有机体和大气CO2之间的碳循环过程中, 出现明显的碳同位素分馏现象(Tieszen & Boutton, 1989).植物在光合作用过程中固定CO2途径的差异, 导致不同光合型植物选择吸收13C的比例不同, 从而使C3和C4植株体内的稳定碳同位素比值(δ13C)存在差异, 因此δ13C可以作为表征植物株生理生态特性差异的指标.土壤有机碳(SOC)经过微生物分解矿化为CO2的过程中13C分馏作用较小, 仅为1‰-3‰, 而SOC在厌氧条件下产生甲烷的过程中13C的分馏作用相对较大, 为10‰-70‰ (Conrad et al., 2010; 林光辉, 2013; 袁红朝等, 2014).但SOC主要来源于陆生高等植物, 与植物光合作用固定CO2时的分馏作用相比, SOC分解过程中的分馏作用几乎可以忽略不计, 因此SOC与该土壤上种植植物的δ13C基本一致(Fernandez et al., 2003; Crow et al., 2006; 张林等, 2010; Bai et al., 2012).所以, 通过测定各个碳库中碳同位素比值的变化, 可以量化“大气-植物-土壤”生态系统中碳的来源、分配及其周转规律. ...
Foliar δ13C within a temperate deciduous forest: spatial, temporal, and species sources of variation
2
1992
... 基于C3/C4植物δ13C的不同, 碳同位素组成可以用于区分植物的C3和C4光合途径以及不同植物功能群结构变化(Garten Jr. & Taylor Jr., 1992; Peñuelas et al., 1999).通过测定不同碳组分(可溶性碳水化合物和结构性碳库)的δ13C值, 从而估计植物水分利用效率和光合作用在不同时间尺度上的变化(Ebdon et al., 1998; Arslan et al., 1999; 陈世苹等, 2002).而在一个复杂的生态系统(如热带雨林)中, 水分利用效率的高低可能影响植物种间竞争的结果, 最终导致生态系统内部群落结构的差异, 因此δ13C为指示生 ...
... 态系统的功能群多样性以及区分不同功能群植物提供了一条重要途径(Garten Jr & Taylor Jr, 1992; 陈世苹等, 2002).任书杰和于贵瑞(2011)整合了中国区域的数据发现水分条件越好, C3植物叶片δ13C越低, 因此C3植物叶片的δ13C可以作为指示植物的水分利用效率的指标.C3/C4植物交互种植时会改变土壤有机质的δ13C, 因此, 以未交互种植作物土壤为对照, 通过土壤有机碳δ13C的变化可以研究土壤有机碳的周转速度, 以及不同植物来源碳占土壤各碳库和呼吸的比例以及激发效应的方向和强度(Kuzyakov, 2010; Werth & Kuzyakov, 2010; Blagodatskaya et al., 2011a), 也可以通过土壤碳自然丰度δ13C的变化来研究土壤碳库中活性组分碳(微生物生物量碳(MBC))的周转速率和稳定性(Blagodatskaya et al., 2011b).研究结果发现, SOC和MBC的周转时间分别为16.8 a和29-30 d, 且随种植时间增加, 周转时间延长.新碳和老碳对土壤各碳库和呼吸的贡献不同, MBC中20%的碳来源于老碳(来自C3作物), CO2气体中60%的碳来源于老碳(来自C3作物), 由于土壤微生物偏好矿化老碳而同化新碳, SOC中新碳(来自C4作物)的贡献将逐年增加.此外, 长期种植C3作物(小麦(Triticum aestivum))的土壤在连续耕种C4作物(玉米(zea mays)) 13年后, 土壤有机碳库更新了22%, 且SOC的周转速率在不同粒径团聚体中各不相同, 在>50 µm与<2 µm粒径的团聚体中包含更多新碳, 而黏粒中有机碳的更新速度最慢(Balesdent & Mariotti, 1996; 袁红朝等, 2014).13C自然丰度法为明确土壤新老碳库对SOC、MBC和CO2排放的贡献, 解析陆地生态系统δ13C的变化及其在全球碳平衡中的应用提供了重要依据(Cheng, 2009).随着测试精度的提高, 采用碳同位素配对标记实验, 只要SOC和其他一种或是两种碳源的碳同位素比值间有较为明显的差别, 也可以解析多碳源相互作用及其对土壤呼吸的贡献, 这对于研究复杂生态系统中碳通量和净碳平衡具有重要的应用价值(Whitman & Lehmann, 2015). ...
Rice rhizodeposition and its utilization by microbial groups depends on N fertilization
3
2017
... 随着碳同位素示踪技术的广泛应用和迅速发展, 多种碳同位素示踪技术及其与其他光谱和微生物探针技术联用, 对于揭示从微观到宏观尺度的碳循环过程起着重要促进作用(图1)(Radajewski et al., 2000; Herrmann et al., 2007).目前自然丰度法、与13C贫化示踪技术结合的自由空气中气体浓度增加(Free-Air Concentration Enrichment, FACE)实验、脉冲与连续标记法以及碳同位素高丰度底物富集标记法是研究陆地生态系统中碳循环过程常用的碳同位素示踪方法(Balesdent & Mariotti, 1996; Bernoux et al., 1998; Conley et al., 2001; Epron et al., 2012; Ge et al., 2017; 张蕊等, 2017).同时, 随着氢、氮(N)、氧等元素同位素示踪技术的发展与应用, 那些单独用碳同位素无法解决的问题在与其他同位素示踪技术、光谱和分子生物学技术的联用下得以解决(陈世苹等, 2002).例如, Kušlienë等(2014)利用13C/15N双标记法, 结合生物标志物分析, 研究了13C/15N标记的不同类型的根系分泌物对土壤碳氮动态的贡献, 这些技术的联用进一步促进了碳同位素示踪技术在生态学研究中的广泛应用.本文重点从稳定碳同位素示踪技术的原理、方法与应用进展等方面综述其在陆地生态系统碳循环过程中的研究进展. ...
... 采用13CO2脉冲标记技术, 对生长在不同SOM含量水稻土上的水稻进行标记, 发现高、低SOM水稻土壤对水稻光合碳的转化和周转具有显著差异, 揭示了水稻光合碳向地下传输过程的“马太效应”(Zhu et al., 2017b).在水稻拔节期和灌浆期内, 通过水稻的根际沉积作用有4%-6%的光合碳进入土壤有机碳库, 土壤可溶性有机碳的2%-4%和土壤微生物生物量碳的9%-18%来源于这部分新碳(Lu et al., 2002a, 2002b; 祝贞科等, 2016).此外, 利用13CO2脉冲标记结合生物标志物(磷脂脂肪酸)的稳定碳同位素探针技术(13C-PLFA-SIP), 发现了碳从水稻向土壤微生物快速转移的现象, 且随着水稻生长期的变化根际碳的微生物同化量显著增加, 真菌及革兰氏阴性菌是主要的同化微生物(Yuan et al., 2016; Liu et al., 2019a).采用13CO2连续标记技术研究了水稻光合碳的传输和转化数量特征及其对氮肥施用的响应, 发现了高氮肥施用条件下水稻光合碳在土壤中的传输和积累比率非线性骤升的规律(Bradford et al., 2008; Ge et al., 2017).结合13C-PLFA-SIP技术发现, 外源C (羧甲基纤维素)和氮肥的联合施用降低了革兰氏阳性菌(G+)和革兰氏阴性菌(G-)的比值, 降低了G+和真菌的丰度, 单施氮肥则刺激了G-和放线菌的增长, 说明可利用碳氮的不同施用策略可显著影响土壤微生物的群落结构(Zhao et al., 2019).Atere等(2017)利用13CO2连续标记技术和土壤有机质的粒径组分分离技术联用, 发现水稻光合碳的固定效率在氮肥施用结合干湿交替条件下有显著提高.连续标记法有助于定量全部生长期或某一生长期全部投入C的净分配, 但是由于成本高以及技术困难, 运用受到限制. ...
... SOC周转是一个连续动态变化的过程, 利用碳同位素技术能够示踪新碳在土壤中的转化过程和赋存状态, 揭示外源碳在土壤-微生物之间的周转过程和机理(金鑫鑫等, 2017).利用13C富集标记小分子有机物或是植物秸秆, 结合生物标志物技术等, 可以表征不同来源的有机质在土壤中的相对丰度和降解状况, 以及SOC的动态变化和储量的转化和迁移, 也可以定量评价新老SOC对碳储量的相对贡献, 可有效评估陆地生态系统碳平衡过程及其微观过程机制(Kuzyakov & Domanski, 2000; Kemmitt et al., 2008; Ge et al., 2017; Zhu et al., 2017a).外源C输入土壤后可以通过物理和生物化学途径形成新的SOM (Cotrufo et al., 2013, 2015), 从而增加SOC库; 另一方面, 外源C的输入也会促进土壤中原有SOM降解(Guenet et al., 2018; Zhu et al., 2018b; Perveen et al., 2019), 从而减少SOC库.因此, 这两个过程之间的平衡对SOC库变化的方向和程度起到至关重要的作用(Liang et al., 2018). ...
稳定性同位素探测技术在微生物生态学研究中的应用
1
2006
... 自然界中已知的碳(C)同位素有7种(10C、11C、12C、13C、14C、15C和16C), 其中两种(12C和13C)为稳定碳同位素.不同同位素之间物理化学性质上的差异使反应前和反应后的物质在同位素组成上存在差异, 这种现象称为同位素分馏效应(陈世苹等, 2002; 林光辉, 2013).基于此, 碳同位素示踪技术是研究陆地生态系统碳氮循环过程的有效手段, 也是揭示全球气候变化成因、控制温室效应的重要工具(葛源等, 2006; Blaser & Conrad, 2016).碳同位素技术在研究土壤有机质的来源、动态转化与稳定性等方面具有高度的专一性和灵敏度, 且标记方法已比较成熟, 被广泛地应用到生态学研究中, 由原来在叶片、种群尺度的研究提高到了冠层、群落、生态系统乃至生物圈的水平, 从新的角度研究生物及其周围环境的关系(陈世苹等, 2002).13C稳定同位素作为示踪物, 与放射性碳同位素相比具有安全、无污染和容易控制等优点, 被广泛应用于陆地生态系统碳周转的生物地球化学过程研究. ...
稳定性同位素探测技术在微生物生态学研究中的应用
1
2006
... 自然界中已知的碳(C)同位素有7种(10C、11C、12C、13C、14C、15C和16C), 其中两种(12C和13C)为稳定碳同位素.不同同位素之间物理化学性质上的差异使反应前和反应后的物质在同位素组成上存在差异, 这种现象称为同位素分馏效应(陈世苹等, 2002; 林光辉, 2013).基于此, 碳同位素示踪技术是研究陆地生态系统碳氮循环过程的有效手段, 也是揭示全球气候变化成因、控制温室效应的重要工具(葛源等, 2006; Blaser & Conrad, 2016).碳同位素技术在研究土壤有机质的来源、动态转化与稳定性等方面具有高度的专一性和灵敏度, 且标记方法已比较成熟, 被广泛地应用到生态学研究中, 由原来在叶片、种群尺度的研究提高到了冠层、群落、生态系统乃至生物圈的水平, 从新的角度研究生物及其周围环境的关系(陈世苹等, 2002).13C稳定同位素作为示踪物, 与放射性碳同位素相比具有安全、无污染和容易控制等优点, 被广泛应用于陆地生态系统碳周转的生物地球化学过程研究. ...
Interactive effects of elevated CO2 and nitrogen deposition on fatty acid molecular and isotope composition of above- and belowground tree biomass and forest soil fractions
1
2015
... 在草地生态系统中, FACE实验研究发现CO2浓度升高, 黑麦草(Lolium perenne)草地0-10 cm 土层土壤团聚体中的颗粒有机碳增加了40%, 表明大气CO2浓度升高可促进草地生态系统的碳周转(Six et al., 2001).在森林生态系统中, Griepentrog等(2015)也发现, CO2浓度升高增加了C3植物向土壤碳的输入量, 提高了土壤碳的周转率.新碳在SOM中的积累程度受有机质密度的影响, 有机质密度增加, 新碳的积累程度减小, 大部分的新碳进入到颗粒态有机质中(Dorodnikov et al., 2011; Hofmockel et al., 2011).同时, 13C-FACE结合PLFA技术, 进一步揭示了CO2浓度升高条件下革兰氏阴性菌和真菌可矿化更多的植物来源碳, 促进了植物新合成的光合碳的分解, 也加快了SOM的分解速率(Pendall et al., 2003; Hagedorn et al., 2013). ...
Impact of priming on global soil carbon stocks
1
2018
... SOC周转是一个连续动态变化的过程, 利用碳同位素技术能够示踪新碳在土壤中的转化过程和赋存状态, 揭示外源碳在土壤-微生物之间的周转过程和机理(金鑫鑫等, 2017).利用13C富集标记小分子有机物或是植物秸秆, 结合生物标志物技术等, 可以表征不同来源的有机质在土壤中的相对丰度和降解状况, 以及SOC的动态变化和储量的转化和迁移, 也可以定量评价新老SOC对碳储量的相对贡献, 可有效评估陆地生态系统碳平衡过程及其微观过程机制(Kuzyakov & Domanski, 2000; Kemmitt et al., 2008; Ge et al., 2017; Zhu et al., 2017a).外源C输入土壤后可以通过物理和生物化学途径形成新的SOM (Cotrufo et al., 2013, 2015), 从而增加SOC库; 另一方面, 外源C的输入也会促进土壤中原有SOM降解(Guenet et al., 2018; Zhu et al., 2018b; Perveen et al., 2019), 从而减少SOC库.因此, 这两个过程之间的平衡对SOC库变化的方向和程度起到至关重要的作用(Liang et al., 2018). ...
Nine years of CO2 enrichment at the alpine treeline stimulates soil respiration but does not alter soil microbial communities
1
2013
... 在草地生态系统中, FACE实验研究发现CO2浓度升高, 黑麦草(Lolium perenne)草地0-10 cm 土层土壤团聚体中的颗粒有机碳增加了40%, 表明大气CO2浓度升高可促进草地生态系统的碳周转(Six et al., 2001).在森林生态系统中, Griepentrog等(2015)也发现, CO2浓度升高增加了C3植物向土壤碳的输入量, 提高了土壤碳的周转率.新碳在SOM中的积累程度受有机质密度的影响, 有机质密度增加, 新碳的积累程度减小, 大部分的新碳进入到颗粒态有机质中(Dorodnikov et al., 2011; Hofmockel et al., 2011).同时, 13C-FACE结合PLFA技术, 进一步揭示了CO2浓度升高条件下革兰氏阴性菌和真菌可矿化更多的植物来源碳, 促进了植物新合成的光合碳的分解, 也加快了SOM的分解速率(Pendall et al., 2003; Hagedorn et al., 2013). ...
Increased N deposition retards mineralization of old soil organic matter
1
2003
... 13C贫化示踪技术结合的FACE实验是研究CO2浓度增加的微域环境中SOC周转的常用方法.大气CO2的δ13C值约为-8‰, 在13C贫化示踪技术结合的FACE实验(13C-FACE)中供给13C贫化的CO2的δ13C值远低于大气, 其范围是-45‰- -18‰ (张蕊等, 2017).FACE系统是一个开放的模拟未来CO2增加的微域环境, 系统内部的气候条件如水分、光照、温度等非常接近自然生态环境.因此, 在这样的系统内进行模拟CO2升高的实验更接近于真实状况(McLeod & Long, 1999).目前全球已建立十余个涉及森林(Hagedorn et al., 2003), 农田(Leavitt et al., 1994)和草地(Six et al., 2001)等多个生态系统和多种植被类型的13C-FACE平台, 阐释了大气CO2浓度升高条件下, 不同生态系统植物生理与光合效率、养分和水分利用、土壤呼吸、土壤碳的输入量及周转率和微生物响应等过程机制(Leavitt et al., 1994; Canadell et al., 1995; Kiem et al., 2000). ...
Separating root and soil microbial contributions to soil respiration: a review of methods and observations
1
2000
... 13CO2同位素标记法, 是指以高丰度13CO2为示踪物, 在植物通过光合作用固定这些示踪物后, 通过在特定的时间内测定植物-土壤系统中各碳库示踪物量的变化, 可定量研究该时间段内13C在植物-土壤系统的转移和分布, 阐明植物某一生长期的碳分配特征, 定量植物碳向土壤的输入以及土壤碳周转, 从而明确植物光合碳对SOM的来源、稳定化过程的影响及其微生物驱动机制(Hanson et al., 2000; Kuzyakov & Cheng, 2001; Kuzyakov, 2006; Yuan et al., 2016; 张蕊等, 2017; Liu et al., 2019b).脉冲标记法是一次或分几次加入13CO2, 标记数小时后在特定时间内测定13C在土壤-植物系统各碳库的分配比例(Kuzyakov & Gavrichkova, 2010)(表1).脉冲标记法只能提供植物生长关键时期光合碳的分配特征, 但是在植物生长期内标记13CO2, 就可以估计植物来源碳输入土壤各组分的量(Lu et al., 2002a; Kimura et al., 2004; Xiao et al., 2019a, 2019b; Zang et al., 2019).连续标记法是指在植物生长某个生长期(或全生长期), 在可控气室等封闭的空间内对植物进行不间断标记, 可以定量全部生长期投入碳的净分配(表1).例如, 在水稻(Oryza satira)拔节期和灌浆期进行13CO2连续标记, 可以定量研究生长过程中水稻光合碳对SOC各组分的贡献(Kuzyakov, 2005; Conrad et al., 2012; Wang et al., 2016), 以及对土壤原有有机质的激发效应(Zhu et al., 2018a). ...
1
2016
... 目前稳定碳同位素示踪耦合原位检测技术在生态学的应用中还面临诸多问题, 特别是在土壤系统中, 由于土壤的复杂性, 多种原位检测技术的性能均受到限制, 稳定碳同位素示踪与原位检测技术耦合性能的提升是其推广应用的前提.由于土壤颗粒的强荧光背景以及土壤孔隙气体的干扰, 拉曼光谱成像对土壤样品的检测难度很大, 目前主要通过类土壤介质和土壤有机体分离间接实现对土壤体系的研究, 干扰掩蔽和矫正技术的研发对稳定碳同位素耦合拉曼光谱成像技术在植物-土壤系统研究中的应用至关重要(何勇, 2016). ...
1
2016
... 目前稳定碳同位素示踪耦合原位检测技术在生态学的应用中还面临诸多问题, 特别是在土壤系统中, 由于土壤的复杂性, 多种原位检测技术的性能均受到限制, 稳定碳同位素示踪与原位检测技术耦合性能的提升是其推广应用的前提.由于土壤颗粒的强荧光背景以及土壤孔隙气体的干扰, 拉曼光谱成像对土壤样品的检测难度很大, 目前主要通过类土壤介质和土壤有机体分离间接实现对土壤体系的研究, 干扰掩蔽和矫正技术的研发对稳定碳同位素耦合拉曼光谱成像技术在植物-土壤系统研究中的应用至关重要(何勇, 2016). ...
Nano-scale secondary ion mass spectrometry—A new analytical tool in biogeochemistry and soil ecology: a review article
2
2007
... 随着碳同位素示踪技术的广泛应用和迅速发展, 多种碳同位素示踪技术及其与其他光谱和微生物探针技术联用, 对于揭示从微观到宏观尺度的碳循环过程起着重要促进作用(图1)(Radajewski et al., 2000; Herrmann et al., 2007).目前自然丰度法、与13C贫化示踪技术结合的自由空气中气体浓度增加(Free-Air Concentration Enrichment, FACE)实验、脉冲与连续标记法以及碳同位素高丰度底物富集标记法是研究陆地生态系统中碳循环过程常用的碳同位素示踪方法(Balesdent & Mariotti, 1996; Bernoux et al., 1998; Conley et al., 2001; Epron et al., 2012; Ge et al., 2017; 张蕊等, 2017).同时, 随着氢、氮(N)、氧等元素同位素示踪技术的发展与应用, 那些单独用碳同位素无法解决的问题在与其他同位素示踪技术、光谱和分子生物学技术的联用下得以解决(陈世苹等, 2002).例如, Kušlienë等(2014)利用13C/15N双标记法, 结合生物标志物分析, 研究了13C/15N标记的不同类型的根系分泌物对土壤碳氮动态的贡献, 这些技术的联用进一步促进了碳同位素示踪技术在生态学研究中的广泛应用.本文重点从稳定碳同位素示踪技术的原理、方法与应用进展等方面综述其在陆地生态系统碳循环过程中的研究进展. ...
... 纳米二次离子质谱(NanoSIMS)能够完成对元素空间分布的超高分辨率显微镜成像, 与同位素示踪技术相结合, 在生态系统微观机制研究领域中显示出了巨大的潜力(Herrmann et al., 2007; Musat et al., 2016).该技术具有较高的灵敏度和准确度, 可应用于碳、氮、硫等元素生物地球化学循环的微生物驱动机制和土壤-微生物-植物相关作用关系的研究, 对于认识多元素耦合的生物地球化学循环规律具有重要意义(图1)(Lechene et al., 2007; Berry et al., 2013; Rumpel et al., 2015).目前, 稳定碳同位素探针-纳米二次离子质谱技术(NanoSIP)已在植物/动物-微生物共生关系(Lechene et al., 2007; Vidal et al., 2016), 微生物群落中碳的食物链传递(Musat et al., 2016), 厌氧甲烷氧化的碳氮耦合机制(Dekas et al., 2009)等研究中取得重要突破.在土壤有机碳周转的研究中, NanoSIP直接呈现了外源碳与土壤矿物的结合形态与动态过程, 有力推动了SOC的稳定机制的研究(Rumpel et al., 2015). ...
Changes in forest soil organic matter pools after a decade of elevated CO2 and O3
1
2011
... 在草地生态系统中, FACE实验研究发现CO2浓度升高, 黑麦草(Lolium perenne)草地0-10 cm 土层土壤团聚体中的颗粒有机碳增加了40%, 表明大气CO2浓度升高可促进草地生态系统的碳周转(Six et al., 2001).在森林生态系统中, Griepentrog等(2015)也发现, CO2浓度升高增加了C3植物向土壤碳的输入量, 提高了土壤碳的周转率.新碳在SOM中的积累程度受有机质密度的影响, 有机质密度增加, 新碳的积累程度减小, 大部分的新碳进入到颗粒态有机质中(Dorodnikov et al., 2011; Hofmockel et al., 2011).同时, 13C-FACE结合PLFA技术, 进一步揭示了CO2浓度升高条件下革兰氏阴性菌和真菌可矿化更多的植物来源碳, 促进了植物新合成的光合碳的分解, 也加快了SOM的分解速率(Pendall et al., 2003; Hagedorn et al., 2013). ...
Visualization and quantification of root exudation using 14C imaging: challenges and uncertainties
1
2019
... 尽管NanoSIP能够完成对土壤稳定碳同位素空间分布的超高分辨率成像, 但成本太高, 且测量范围在纳米尺度(Rumpel et al., 2015).将14C同位素示踪与放射自显影相结合, 能够直观呈现植物、土壤碳的分布与传输过程, 为准确解析碳元素源-库关系和分配动态提供了有力工具(Pausch & Kuzyakov, 2011).在利用放射自显影技术研究14C在土壤中的空间分布时, 土壤颗粒对β-粒子的激发、电离和轫致辐射等过程可强烈削弱14C信号, 通过基质转移(Holz et al., 2018)、衰减系数核算(Holz et al., 2019)等手段有望校正测定偏差, 提高该技术在土壤样品检测中的可信度.磷光体成像的应用提高了放射自显影技术的安全性和灵敏度, 为其在生态学的推广应用提供了良好的机遇.同位素示踪与超高分辨率成像技术相结合, 能在微观尺度上研究碳周转中植物-微生物-土壤的互相作用过程及碳与其他元素的耦合关系, 对全球碳循环的机理揭示有重要意义. ...
Root hairs increase rhizosphere extension and carbon input to soil
1
2018
... 尽管NanoSIP能够完成对土壤稳定碳同位素空间分布的超高分辨率成像, 但成本太高, 且测量范围在纳米尺度(Rumpel et al., 2015).将14C同位素示踪与放射自显影相结合, 能够直观呈现植物、土壤碳的分布与传输过程, 为准确解析碳元素源-库关系和分配动态提供了有力工具(Pausch & Kuzyakov, 2011).在利用放射自显影技术研究14C在土壤中的空间分布时, 土壤颗粒对β-粒子的激发、电离和轫致辐射等过程可强烈削弱14C信号, 通过基质转移(Holz et al., 2018)、衰减系数核算(Holz et al., 2019)等手段有望校正测定偏差, 提高该技术在土壤样品检测中的可信度.磷光体成像的应用提高了放射自显影技术的安全性和灵敏度, 为其在生态学的推广应用提供了良好的机遇.同位素示踪与超高分辨率成像技术相结合, 能在微观尺度上研究碳周转中植物-微生物-土壤的互相作用过程及碳与其他元素的耦合关系, 对全球碳循环的机理揭示有重要意义. ...
Quantitative microbial ecology through stable isotope probing
1
2015
... 将稳定碳同位素示踪技术与分子生物学方法相结合, 在表征以微生物为主要研究对象的土壤生物群落功能过程研究中发挥了重要作用(Lu & Conrad, 2005; Ruyters et al., 2013; Fan et al., 2014).13C同位素探针-磷脂脂肪酸(PLFA-SIP)是在土壤碳循环微生物驱动机制的研究中广泛应用的技术.该方法灵敏度高, 常用于指示活体微生物生物量和群落结构, 能够定量检测微生物对标记底物的同化利用.近年来, 以氨基糖为主要成分的微生物残留物的相对稳定性及其对SOC的贡献得到广泛关注(Liang et al., 2017), 将PLFA-SIP与氨基糖生物标志物相结合, 建立外源碳-微生物-土壤碳库的碳周转连续体模型, 有助于揭示SOC的累积机制.PLFA-SIP对物种组成的分辨率较低, 难以判断功能微生物, 且无法检测古菌.以DNA/RNA测序为基础的DNA/RNA-SIP技术可以很好地解决上述问题, 只要选择合适的底物和目的基因, DNA/RNA-SIP技术能准确解析关键微生物在土壤中的群落结构和功能变化, 特异性地将微生物群体与生态过程、生理特性相关联, 揭示微生物驱动下的土壤元素循环过程及其调控机制.近年来, Hungate等(2015)将13C与18O标记相结合, 建立了定量DNA-SIP技术(DNA-qSIP), 揭示了参与土壤中新输入的有机碳(新碳)和土壤原有有机碳(老 碳)周转过程的微生物群落差异(Morrissey et al., 2017). ...
稳定 13C同位素示踪技术在农田土壤碳循环和团聚体固碳研究中的应用进展
1
2017
... SOC周转是一个连续动态变化的过程, 利用碳同位素技术能够示踪新碳在土壤中的转化过程和赋存状态, 揭示外源碳在土壤-微生物之间的周转过程和机理(金鑫鑫等, 2017).利用13C富集标记小分子有机物或是植物秸秆, 结合生物标志物技术等, 可以表征不同来源的有机质在土壤中的相对丰度和降解状况, 以及SOC的动态变化和储量的转化和迁移, 也可以定量评价新老SOC对碳储量的相对贡献, 可有效评估陆地生态系统碳平衡过程及其微观过程机制(Kuzyakov & Domanski, 2000; Kemmitt et al., 2008; Ge et al., 2017; Zhu et al., 2017a).外源C输入土壤后可以通过物理和生物化学途径形成新的SOM (Cotrufo et al., 2013, 2015), 从而增加SOC库; 另一方面, 外源C的输入也会促进土壤中原有SOM降解(Guenet et al., 2018; Zhu et al., 2018b; Perveen et al., 2019), 从而减少SOC库.因此, 这两个过程之间的平衡对SOC库变化的方向和程度起到至关重要的作用(Liang et al., 2018). ...
稳定 13C同位素示踪技术在农田土壤碳循环和团聚体固碳研究中的应用进展
1
2017
... SOC周转是一个连续动态变化的过程, 利用碳同位素技术能够示踪新碳在土壤中的转化过程和赋存状态, 揭示外源碳在土壤-微生物之间的周转过程和机理(金鑫鑫等, 2017).利用13C富集标记小分子有机物或是植物秸秆, 结合生物标志物技术等, 可以表征不同来源的有机质在土壤中的相对丰度和降解状况, 以及SOC的动态变化和储量的转化和迁移, 也可以定量评价新老SOC对碳储量的相对贡献, 可有效评估陆地生态系统碳平衡过程及其微观过程机制(Kuzyakov & Domanski, 2000; Kemmitt et al., 2008; Ge et al., 2017; Zhu et al., 2017a).外源C输入土壤后可以通过物理和生物化学途径形成新的SOM (Cotrufo et al., 2013, 2015), 从而增加SOC库; 另一方面, 外源C的输入也会促进土壤中原有SOM降解(Guenet et al., 2018; Zhu et al., 2018b; Perveen et al., 2019), 从而减少SOC库.因此, 这两个过程之间的平衡对SOC库变化的方向和程度起到至关重要的作用(Liang et al., 2018). ...
Mineralization of native soil organic matter is not regulated by the size, activity or composition of the soil microbial biomass—A new perspective
1
2008
... SOC周转是一个连续动态变化的过程, 利用碳同位素技术能够示踪新碳在土壤中的转化过程和赋存状态, 揭示外源碳在土壤-微生物之间的周转过程和机理(金鑫鑫等, 2017).利用13C富集标记小分子有机物或是植物秸秆, 结合生物标志物技术等, 可以表征不同来源的有机质在土壤中的相对丰度和降解状况, 以及SOC的动态变化和储量的转化和迁移, 也可以定量评价新老SOC对碳储量的相对贡献, 可有效评估陆地生态系统碳平衡过程及其微观过程机制(Kuzyakov & Domanski, 2000; Kemmitt et al., 2008; Ge et al., 2017; Zhu et al., 2017a).外源C输入土壤后可以通过物理和生物化学途径形成新的SOM (Cotrufo et al., 2013, 2015), 从而增加SOC库; 另一方面, 外源C的输入也会促进土壤中原有SOM降解(Guenet et al., 2018; Zhu et al., 2018b; Perveen et al., 2019), 从而减少SOC库.因此, 这两个过程之间的平衡对SOC库变化的方向和程度起到至关重要的作用(Liang et al., 2018). ...
Refractory organic carbon in C-depleted arable soils, as studied by 13C NMR spectroscopy and carbohydrate analysis
1
2000
... 13C贫化示踪技术结合的FACE实验是研究CO2浓度增加的微域环境中SOC周转的常用方法.大气CO2的δ13C值约为-8‰, 在13C贫化示踪技术结合的FACE实验(13C-FACE)中供给13C贫化的CO2的δ13C值远低于大气, 其范围是-45‰- -18‰ (张蕊等, 2017).FACE系统是一个开放的模拟未来CO2增加的微域环境, 系统内部的气候条件如水分、光照、温度等非常接近自然生态环境.因此, 在这样的系统内进行模拟CO2升高的实验更接近于真实状况(McLeod & Long, 1999).目前全球已建立十余个涉及森林(Hagedorn et al., 2003), 农田(Leavitt et al., 1994)和草地(Six et al., 2001)等多个生态系统和多种植被类型的13C-FACE平台, 阐释了大气CO2浓度升高条件下, 不同生态系统植物生理与光合效率、养分和水分利用、土壤呼吸、土壤碳的输入量及周转率和微生物响应等过程机制(Leavitt et al., 1994; Canadell et al., 1995; Kiem et al., 2000). ...
Carbon cycling in rice field ecosystems in the context of input, decomposition and translocation of organic materials and the fates of their end products (CO2 and CH4)
1
2004
... 13CO2同位素标记法, 是指以高丰度13CO2为示踪物, 在植物通过光合作用固定这些示踪物后, 通过在特定的时间内测定植物-土壤系统中各碳库示踪物量的变化, 可定量研究该时间段内13C在植物-土壤系统的转移和分布, 阐明植物某一生长期的碳分配特征, 定量植物碳向土壤的输入以及土壤碳周转, 从而明确植物光合碳对SOM的来源、稳定化过程的影响及其微生物驱动机制(Hanson et al., 2000; Kuzyakov & Cheng, 2001; Kuzyakov, 2006; Yuan et al., 2016; 张蕊等, 2017; Liu et al., 2019b).脉冲标记法是一次或分几次加入13CO2, 标记数小时后在特定时间内测定13C在土壤-植物系统各碳库的分配比例(Kuzyakov & Gavrichkova, 2010)(表1).脉冲标记法只能提供植物生长关键时期光合碳的分配特征, 但是在植物生长期内标记13CO2, 就可以估计植物来源碳输入土壤各组分的量(Lu et al., 2002a; Kimura et al., 2004; Xiao et al., 2019a, 2019b; Zang et al., 2019).连续标记法是指在植物生长某个生长期(或全生长期), 在可控气室等封闭的空间内对植物进行不间断标记, 可以定量全部生长期投入碳的净分配(表1).例如, 在水稻(Oryza satira)拔节期和灌浆期进行13CO2连续标记, 可以定量研究生长过程中水稻光合碳对SOC各组分的贡献(Kuzyakov, 2005; Conrad et al., 2012; Wang et al., 2016), 以及对土壤原有有机质的激发效应(Zhu et al., 2018a). ...
Isotopic evidence for neogene hominid paleoenvironments in the Kenya rift valley
1
1994
... 13C自然丰度法是基于C3/C4植物具有不同的δ13C, 将C3 (C4)植物种植在具有C4 (C3)植物种植历史的土壤, 植物来源与原有SOM来源的δ13C存在差异, 可以以此来区分土壤源呼吸和根源呼吸的C (Kingston et al., 1994).该方法成本低且容易操作, 但是需要注意的是, 选取的C4 (C3)土壤需要长期一直种植C4 (C3)植物, 且该方法灵敏度相对较低, 在植物生长季后期可靠性下降(Rochette et al., 1999). ...
Medium-term response of microbial community to rhizodeposits of white clover and ryegrass and tracing of active processes induced by 13C and 15N labelled exudates
1
2014
... 随着碳同位素示踪技术的广泛应用和迅速发展, 多种碳同位素示踪技术及其与其他光谱和微生物探针技术联用, 对于揭示从微观到宏观尺度的碳循环过程起着重要促进作用(图1)(Radajewski et al., 2000; Herrmann et al., 2007).目前自然丰度法、与13C贫化示踪技术结合的自由空气中气体浓度增加(Free-Air Concentration Enrichment, FACE)实验、脉冲与连续标记法以及碳同位素高丰度底物富集标记法是研究陆地生态系统中碳循环过程常用的碳同位素示踪方法(Balesdent & Mariotti, 1996; Bernoux et al., 1998; Conley et al., 2001; Epron et al., 2012; Ge et al., 2017; 张蕊等, 2017).同时, 随着氢、氮(N)、氧等元素同位素示踪技术的发展与应用, 那些单独用碳同位素无法解决的问题在与其他同位素示踪技术、光谱和分子生物学技术的联用下得以解决(陈世苹等, 2002).例如, Kušlienë等(2014)利用13C/15N双标记法, 结合生物标志物分析, 研究了13C/15N标记的不同类型的根系分泌物对土壤碳氮动态的贡献, 这些技术的联用进一步促进了碳同位素示踪技术在生态学研究中的广泛应用.本文重点从稳定碳同位素示踪技术的原理、方法与应用进展等方面综述其在陆地生态系统碳循环过程中的研究进展. ...
Theoretical background for partitioning of root and rhizomicrobial respiration by δ13C of microbial biomass
1
2005
... 13CO2同位素标记法, 是指以高丰度13CO2为示踪物, 在植物通过光合作用固定这些示踪物后, 通过在特定的时间内测定植物-土壤系统中各碳库示踪物量的变化, 可定量研究该时间段内13C在植物-土壤系统的转移和分布, 阐明植物某一生长期的碳分配特征, 定量植物碳向土壤的输入以及土壤碳周转, 从而明确植物光合碳对SOM的来源、稳定化过程的影响及其微生物驱动机制(Hanson et al., 2000; Kuzyakov & Cheng, 2001; Kuzyakov, 2006; Yuan et al., 2016; 张蕊等, 2017; Liu et al., 2019b).脉冲标记法是一次或分几次加入13CO2, 标记数小时后在特定时间内测定13C在土壤-植物系统各碳库的分配比例(Kuzyakov & Gavrichkova, 2010)(表1).脉冲标记法只能提供植物生长关键时期光合碳的分配特征, 但是在植物生长期内标记13CO2, 就可以估计植物来源碳输入土壤各组分的量(Lu et al., 2002a; Kimura et al., 2004; Xiao et al., 2019a, 2019b; Zang et al., 2019).连续标记法是指在植物生长某个生长期(或全生长期), 在可控气室等封闭的空间内对植物进行不间断标记, 可以定量全部生长期投入碳的净分配(表1).例如, 在水稻(Oryza satira)拔节期和灌浆期进行13CO2连续标记, 可以定量研究生长过程中水稻光合碳对SOC各组分的贡献(Kuzyakov, 2005; Conrad et al., 2012; Wang et al., 2016), 以及对土壤原有有机质的激发效应(Zhu et al., 2018a). ...
Sources of CO2 efflux from soil and review of partitioning methods
2
2006
... 13CO2同位素标记法, 是指以高丰度13CO2为示踪物, 在植物通过光合作用固定这些示踪物后, 通过在特定的时间内测定植物-土壤系统中各碳库示踪物量的变化, 可定量研究该时间段内13C在植物-土壤系统的转移和分布, 阐明植物某一生长期的碳分配特征, 定量植物碳向土壤的输入以及土壤碳周转, 从而明确植物光合碳对SOM的来源、稳定化过程的影响及其微生物驱动机制(Hanson et al., 2000; Kuzyakov & Cheng, 2001; Kuzyakov, 2006; Yuan et al., 2016; 张蕊等, 2017; Liu et al., 2019b).脉冲标记法是一次或分几次加入13CO2, 标记数小时后在特定时间内测定13C在土壤-植物系统各碳库的分配比例(Kuzyakov & Gavrichkova, 2010)(表1).脉冲标记法只能提供植物生长关键时期光合碳的分配特征, 但是在植物生长期内标记13CO2, 就可以估计植物来源碳输入土壤各组分的量(Lu et al., 2002a; Kimura et al., 2004; Xiao et al., 2019a, 2019b; Zang et al., 2019).连续标记法是指在植物生长某个生长期(或全生长期), 在可控气室等封闭的空间内对植物进行不间断标记, 可以定量全部生长期投入碳的净分配(表1).例如, 在水稻(Oryza satira)拔节期和灌浆期进行13CO2连续标记, 可以定量研究生长过程中水稻光合碳对SOC各组分的贡献(Kuzyakov, 2005; Conrad et al., 2012; Wang et al., 2016), 以及对土壤原有有机质的激发效应(Zhu et al., 2018a). ...
... 目前应用碳同位素示踪技术对C3/C4植被演替、土壤有机质源汇和各组分的周转及其微生物响应机制等方面进行了大量研究并日趋成熟(Six et al., 2001; Kuzyakov, 2006; Werth & Kuzyakov, 2010; Wang et al., 2016; Liu et al., 2019b), 而对于植物-土壤碳循环中植物、微生物以及土壤动物(如原生动物、线虫和节肢动物等)所扮演的角色及其循环机理研究较少.尽管碳同位素示踪在植物生理生态研究中已发挥重要作用, 但许多物理、化学和生物过程还不十分清楚; 放牧、施肥、耕作等人类活动等多种因素可直接影响或与气候协同作用影响植被变化以及导致土壤有机质变化的综合研究尚需深入; 还缺乏碳与其他养分元素的化学计量学方面的研究, 以及碳、氮、磷、硫、氧等多种同位素联合探测技术在生态学领域的综合开发及应用.鉴于此, 我们提出以下四点展望: ...
Priming effects: interactions between living and dead organic matter
2
2010
... 态系统的功能群多样性以及区分不同功能群植物提供了一条重要途径(Garten Jr & Taylor Jr, 1992; 陈世苹等, 2002).任书杰和于贵瑞(2011)整合了中国区域的数据发现水分条件越好, C3植物叶片δ13C越低, 因此C3植物叶片的δ13C可以作为指示植物的水分利用效率的指标.C3/C4植物交互种植时会改变土壤有机质的δ13C, 因此, 以未交互种植作物土壤为对照, 通过土壤有机碳δ13C的变化可以研究土壤有机碳的周转速度, 以及不同植物来源碳占土壤各碳库和呼吸的比例以及激发效应的方向和强度(Kuzyakov, 2010; Werth & Kuzyakov, 2010; Blagodatskaya et al., 2011a), 也可以通过土壤碳自然丰度δ13C的变化来研究土壤碳库中活性组分碳(微生物生物量碳(MBC))的周转速率和稳定性(Blagodatskaya et al., 2011b).研究结果发现, SOC和MBC的周转时间分别为16.8 a和29-30 d, 且随种植时间增加, 周转时间延长.新碳和老碳对土壤各碳库和呼吸的贡献不同, MBC中20%的碳来源于老碳(来自C3作物), CO2气体中60%的碳来源于老碳(来自C3作物), 由于土壤微生物偏好矿化老碳而同化新碳, SOC中新碳(来自C4作物)的贡献将逐年增加.此外, 长期种植C3作物(小麦(Triticum aestivum))的土壤在连续耕种C4作物(玉米(zea mays)) 13年后, 土壤有机碳库更新了22%, 且SOC的周转速率在不同粒径团聚体中各不相同, 在>50 µm与<2 µm粒径的团聚体中包含更多新碳, 而黏粒中有机碳的更新速度最慢(Balesdent & Mariotti, 1996; 袁红朝等, 2014).13C自然丰度法为明确土壤新老碳库对SOC、MBC和CO2排放的贡献, 解析陆地生态系统δ13C的变化及其在全球碳平衡中的应用提供了重要依据(Cheng, 2009).随着测试精度的提高, 采用碳同位素配对标记实验, 只要SOC和其他一种或是两种碳源的碳同位素比值间有较为明显的差别, 也可以解析多碳源相互作用及其对土壤呼吸的贡献, 这对于研究复杂生态系统中碳通量和净碳平衡具有重要的应用价值(Whitman & Lehmann, 2015). ...
... ; Werth & Kuzyakov, 2010; Blagodatskaya et al., 2011a), 也可以通过土壤碳自然丰度δ13C的变化来研究土壤碳库中活性组分碳(微生物生物量碳(MBC))的周转速率和稳定性(Blagodatskaya et al., 2011b).研究结果发现, SOC和MBC的周转时间分别为16.8 a和29-30 d, 且随种植时间增加, 周转时间延长.新碳和老碳对土壤各碳库和呼吸的贡献不同, MBC中20%的碳来源于老碳(来自C3作物), CO2气体中60%的碳来源于老碳(来自C3作物), 由于土壤微生物偏好矿化老碳而同化新碳, SOC中新碳(来自C4作物)的贡献将逐年增加.此外, 长期种植C3作物(小麦(Triticum aestivum))的土壤在连续耕种C4作物(玉米(zea mays)) 13年后, 土壤有机碳库更新了22%, 且SOC的周转速率在不同粒径团聚体中各不相同, 在>50 µm与<2 µm粒径的团聚体中包含更多新碳, 而黏粒中有机碳的更新速度最慢(Balesdent & Mariotti, 1996; 袁红朝等, 2014).13C自然丰度法为明确土壤新老碳库对SOC、MBC和CO2排放的贡献, 解析陆地生态系统δ13C的变化及其在全球碳平衡中的应用提供了重要依据(Cheng, 2009).随着测试精度的提高, 采用碳同位素配对标记实验, 只要SOC和其他一种或是两种碳源的碳同位素比值间有较为明显的差别, 也可以解析多碳源相互作用及其对土壤呼吸的贡献, 这对于研究复杂生态系统中碳通量和净碳平衡具有重要的应用价值(Whitman & Lehmann, 2015). ...
Photosynthesis controls of rhizosphere respiration and organic matter decomposition
1
2001
... 13CO2同位素标记法, 是指以高丰度13CO2为示踪物, 在植物通过光合作用固定这些示踪物后, 通过在特定的时间内测定植物-土壤系统中各碳库示踪物量的变化, 可定量研究该时间段内13C在植物-土壤系统的转移和分布, 阐明植物某一生长期的碳分配特征, 定量植物碳向土壤的输入以及土壤碳周转, 从而明确植物光合碳对SOM的来源、稳定化过程的影响及其微生物驱动机制(Hanson et al., 2000; Kuzyakov & Cheng, 2001; Kuzyakov, 2006; Yuan et al., 2016; 张蕊等, 2017; Liu et al., 2019b).脉冲标记法是一次或分几次加入13CO2, 标记数小时后在特定时间内测定13C在土壤-植物系统各碳库的分配比例(Kuzyakov & Gavrichkova, 2010)(表1).脉冲标记法只能提供植物生长关键时期光合碳的分配特征, 但是在植物生长期内标记13CO2, 就可以估计植物来源碳输入土壤各组分的量(Lu et al., 2002a; Kimura et al., 2004; Xiao et al., 2019a, 2019b; Zang et al., 2019).连续标记法是指在植物生长某个生长期(或全生长期), 在可控气室等封闭的空间内对植物进行不间断标记, 可以定量全部生长期投入碳的净分配(表1).例如, 在水稻(Oryza satira)拔节期和灌浆期进行13CO2连续标记, 可以定量研究生长过程中水稻光合碳对SOC各组分的贡献(Kuzyakov, 2005; Conrad et al., 2012; Wang et al., 2016), 以及对土壤原有有机质的激发效应(Zhu et al., 2018a). ...
Carbon input by plants into the soil. Review
1
2000
... SOC周转是一个连续动态变化的过程, 利用碳同位素技术能够示踪新碳在土壤中的转化过程和赋存状态, 揭示外源碳在土壤-微生物之间的周转过程和机理(金鑫鑫等, 2017).利用13C富集标记小分子有机物或是植物秸秆, 结合生物标志物技术等, 可以表征不同来源的有机质在土壤中的相对丰度和降解状况, 以及SOC的动态变化和储量的转化和迁移, 也可以定量评价新老SOC对碳储量的相对贡献, 可有效评估陆地生态系统碳平衡过程及其微观过程机制(Kuzyakov & Domanski, 2000; Kemmitt et al., 2008; Ge et al., 2017; Zhu et al., 2017a).外源C输入土壤后可以通过物理和生物化学途径形成新的SOM (Cotrufo et al., 2013, 2015), 从而增加SOC库; 另一方面, 外源C的输入也会促进土壤中原有SOM降解(Guenet et al., 2018; Zhu et al., 2018b; Perveen et al., 2019), 从而减少SOC库.因此, 这两个过程之间的平衡对SOC库变化的方向和程度起到至关重要的作用(Liang et al., 2018). ...
Time lag between photosynthesis and carbon dioxide efflux from soil: a review of mechanisms and controls
2
2010
... 13CO2同位素标记法, 是指以高丰度13CO2为示踪物, 在植物通过光合作用固定这些示踪物后, 通过在特定的时间内测定植物-土壤系统中各碳库示踪物量的变化, 可定量研究该时间段内13C在植物-土壤系统的转移和分布, 阐明植物某一生长期的碳分配特征, 定量植物碳向土壤的输入以及土壤碳周转, 从而明确植物光合碳对SOM的来源、稳定化过程的影响及其微生物驱动机制(Hanson et al., 2000; Kuzyakov & Cheng, 2001; Kuzyakov, 2006; Yuan et al., 2016; 张蕊等, 2017; Liu et al., 2019b).脉冲标记法是一次或分几次加入13CO2, 标记数小时后在特定时间内测定13C在土壤-植物系统各碳库的分配比例(Kuzyakov & Gavrichkova, 2010)(表1).脉冲标记法只能提供植物生长关键时期光合碳的分配特征, 但是在植物生长期内标记13CO2, 就可以估计植物来源碳输入土壤各组分的量(Lu et al., 2002a; Kimura et al., 2004; Xiao et al., 2019a, 2019b; Zang et al., 2019).连续标记法是指在植物生长某个生长期(或全生长期), 在可控气室等封闭的空间内对植物进行不间断标记, 可以定量全部生长期投入碳的净分配(表1).例如, 在水稻(Oryza satira)拔节期和灌浆期进行13CO2连续标记, 可以定量研究生长过程中水稻光合碳对SOC各组分的贡献(Kuzyakov, 2005; Conrad et al., 2012; Wang et al., 2016), 以及对土壤原有有机质的激发效应(Zhu et al., 2018a). ...
... 13CO2脉冲标记在密闭的植物生长箱中进行, 当植物生长到一定时期, 在光照充足时通入13CO2或通过化学反应(如NaH13CO3和H2SO4)获得13CO2, 标记数个小时后移出, 在标记期间由CO2发生器和检测系统实时监测并控制CO2浓度, 生长箱内温湿度需与对照处理(未标记13CO2的植物)保持一致(Kuzyakov & Gavrichkova, 2010); 为避免13CO2污染, 对照处理要距离标记的植物至少5 m以上(Lu et al., 2003).13CO2连续标记的方法与脉冲标记基本一致, 但是并不是短时间标记, 而是在植物生长某个生长期, 甚至整个生长期内, 对植物进行不间断标记. ...
Carbon isotope dynamics of free-air CO2-enriched cotton and soils
3
1994
... 13C贫化示踪技术结合的FACE实验是研究CO2浓度增加的微域环境中SOC周转的常用方法.大气CO2的δ13C值约为-8‰, 在13C贫化示踪技术结合的FACE实验(13C-FACE)中供给13C贫化的CO2的δ13C值远低于大气, 其范围是-45‰- -18‰ (张蕊等, 2017).FACE系统是一个开放的模拟未来CO2增加的微域环境, 系统内部的气候条件如水分、光照、温度等非常接近自然生态环境.因此, 在这样的系统内进行模拟CO2升高的实验更接近于真实状况(McLeod & Long, 1999).目前全球已建立十余个涉及森林(Hagedorn et al., 2003), 农田(Leavitt et al., 1994)和草地(Six et al., 2001)等多个生态系统和多种植被类型的13C-FACE平台, 阐释了大气CO2浓度升高条件下, 不同生态系统植物生理与光合效率、养分和水分利用、土壤呼吸、土壤碳的输入量及周转率和微生物响应等过程机制(Leavitt et al., 1994; Canadell et al., 1995; Kiem et al., 2000). ...
... 浓度升高条件下, 不同生态系统植物生理与光合效率、养分和水分利用、土壤呼吸、土壤碳的输入量及周转率和微生物响应等过程机制(Leavitt et al., 1994; Canadell et al., 1995; Kiem et al., 2000). ...
... FACE实验系统一般由FACE圈(CO2递增圈)和Ambient圈(常规对照圈)构成(Leavitt et al., 1994).采用人为添加的方式(如通过围绕在FACE圈的纯CO2气体管道向圈中心喷散CO2)提高FACE圈CO2气体浓度, 由远程控制器实时监测并控制FACE圈内CO2浓度, 使其始终保持在目标浓度.实验过程中FACE圈和Ambient圈的大小和降水量、温度等自然条件要保持一致, 且相邻的两圈之间设置缓冲区, 以避免CO2气体增加对其他圈的影响. ...
Quantitative imaging of nitrogen fixation by individual bacteria within animal cells
2
2007
... 纳米二次离子质谱(NanoSIMS)能够完成对元素空间分布的超高分辨率显微镜成像, 与同位素示踪技术相结合, 在生态系统微观机制研究领域中显示出了巨大的潜力(Herrmann et al., 2007; Musat et al., 2016).该技术具有较高的灵敏度和准确度, 可应用于碳、氮、硫等元素生物地球化学循环的微生物驱动机制和土壤-微生物-植物相关作用关系的研究, 对于认识多元素耦合的生物地球化学循环规律具有重要意义(图1)(Lechene et al., 2007; Berry et al., 2013; Rumpel et al., 2015).目前, 稳定碳同位素探针-纳米二次离子质谱技术(NanoSIP)已在植物/动物-微生物共生关系(Lechene et al., 2007; Vidal et al., 2016), 微生物群落中碳的食物链传递(Musat et al., 2016), 厌氧甲烷氧化的碳氮耦合机制(Dekas et al., 2009)等研究中取得重要突破.在土壤有机碳周转的研究中, NanoSIP直接呈现了外源碳与土壤矿物的结合形态与动态过程, 有力推动了SOC的稳定机制的研究(Rumpel et al., 2015). ...
... ).目前, 稳定碳同位素探针-纳米二次离子质谱技术(NanoSIP)已在植物/动物-微生物共生关系(Lechene et al., 2007; Vidal et al., 2016), 微生物群落中碳的食物链传递(Musat et al., 2016), 厌氧甲烷氧化的碳氮耦合机制(Dekas et al., 2009)等研究中取得重要突破.在土壤有机碳周转的研究中, NanoSIP直接呈现了外源碳与土壤矿物的结合形态与动态过程, 有力推动了SOC的稳定机制的研究(Rumpel et al., 2015). ...
Rapid resonance Raman microspectroscopy to probe carbon dioxide fixation by single cells in microbial communities
1
2012
... 拉曼光谱成像能高分辨率捕捉不同化学成分在样品中的空间分布特征, 具有样本需求量小、无需前处理、非侵入性、无损伤、对样品物理状态要求低等诸多优势, 自开发以来得到广泛应用.13C稳定同位素探针-拉曼显微光谱技术(SIRM)利用重同位素的光谱红移, 可原位连续检测含碳化合物在生态系统和生物体内的合成、分解、运输、分配等过程, 不仅能够在单细胞水平上提供微生物的生理生态特征信息, 还能够检测到自然环境丰度较低但能发挥重要作用的微生物类群, 对深入理解生态系统碳循环过程有重要意义(图1)(Li et al., 2012; Wiesheu et al., 2018).近年发展起来的表面增强拉曼光谱成像技术通过强化拉曼散射信号, 有效提高了拉曼光谱的灵敏性(Zong et al., 2018). ...
The importance of anabolism in microbial control over soil carbon storage
4
2017
... 将稳定碳同位素示踪技术与分子生物学方法相结合, 在表征以微生物为主要研究对象的土壤生物群落功能过程研究中发挥了重要作用(Lu & Conrad, 2005; Ruyters et al., 2013; Fan et al., 2014).13C同位素探针-磷脂脂肪酸(PLFA-SIP)是在土壤碳循环微生物驱动机制的研究中广泛应用的技术.该方法灵敏度高, 常用于指示活体微生物生物量和群落结构, 能够定量检测微生物对标记底物的同化利用.近年来, 以氨基糖为主要成分的微生物残留物的相对稳定性及其对SOC的贡献得到广泛关注(Liang et al., 2017), 将PLFA-SIP与氨基糖生物标志物相结合, 建立外源碳-微生物-土壤碳库的碳周转连续体模型, 有助于揭示SOC的累积机制.PLFA-SIP对物种组成的分辨率较低, 难以判断功能微生物, 且无法检测古菌.以DNA/RNA测序为基础的DNA/RNA-SIP技术可以很好地解决上述问题, 只要选择合适的底物和目的基因, DNA/RNA-SIP技术能准确解析关键微生物在土壤中的群落结构和功能变化, 特异性地将微生物群体与生态过程、生理特性相关联, 揭示微生物驱动下的土壤元素循环过程及其调控机制.近年来, Hungate等(2015)将13C与18O标记相结合, 建立了定量DNA-SIP技术(DNA-qSIP), 揭示了参与土壤中新输入的有机碳(新碳)和土壤原有有机碳(老 碳)周转过程的微生物群落差异(Morrissey et al., 2017). ...
... 13C富集底物标记技术一般用于室内培养实验, 13C富集底物可分为简单的小分子有机物(如葡萄糖、丙氨酸等)和复杂的大分子有机物(如纤维素、木质素、植物秸秆等).在实验土壤中加入13C标记的底物, 以未标记的底物为对照, 结合PLFA、氨基糖等生物标志物技术, 测定13C在土壤各组分有机碳的量及其来源, 有助于阐明外源碳对土壤碳储量的贡献(Yuan et al., 2016; Liang et al., 2017). ...
... 13C同位素示踪法技术可以很好地研究外源C对SOM周转的影响.例如, 将13C标记的有机物加入土壤进行田间或室内培养实验, 通过测定13CO2可以区分外源有机物和原有SOM来源的碳, 量化外源有机物添加SOM分解的影响(Bernoux et al., 1998; Blagodatskaya & Kuzyakov, 2008).高丰度13C标记的有机物结合13C-PLFA-SIP和13C-DNA-SIP技术, 可以准确示踪微生物在土壤有机碳周转过程中的参与机制(Williams et al., 2006; Apostel et al., 2015; Wang et al., 2016).而13C-微生物残留物对于量化和评估微生物对SOM积累的贡献起到了关键作用(Tian et al., 2014; Zhang et al., 2015; Liang et al., 2017). ...
... 基于质量平衡原理解析土壤-植物-微生物连续体系中有机物的二元或三元组成与贡献, 量化生态系统中小分子根系分泌物-植物残体-生物质炭-土壤有机质-微生物活体与残留物等碳源的相对比例, 揭示生态系统碳源流通过程与机理(Phillips & Gregg, 2003; Whitman & Lehmann, 2015; Liang et al., 2017).然而, 生态系统中有机物是多碳源组成的, 质量平衡并不能适应所有环境条件下有机物的源解析.因此, 需要基于同位素示踪基本原理, 构建合理的方程模型与原创性理论, 准确区分与量化复杂生态系统中有机碳的来源与周转通量, 揭示生态系统碳循环与调控机理. ...
More replenishment than priming loss of soil organic carbon with additional carbon input
1
2018
... SOC周转是一个连续动态变化的过程, 利用碳同位素技术能够示踪新碳在土壤中的转化过程和赋存状态, 揭示外源碳在土壤-微生物之间的周转过程和机理(金鑫鑫等, 2017).利用13C富集标记小分子有机物或是植物秸秆, 结合生物标志物技术等, 可以表征不同来源的有机质在土壤中的相对丰度和降解状况, 以及SOC的动态变化和储量的转化和迁移, 也可以定量评价新老SOC对碳储量的相对贡献, 可有效评估陆地生态系统碳平衡过程及其微观过程机制(Kuzyakov & Domanski, 2000; Kemmitt et al., 2008; Ge et al., 2017; Zhu et al., 2017a).外源C输入土壤后可以通过物理和生物化学途径形成新的SOM (Cotrufo et al., 2013, 2015), 从而增加SOC库; 另一方面, 外源C的输入也会促进土壤中原有SOM降解(Guenet et al., 2018; Zhu et al., 2018b; Perveen et al., 2019), 从而减少SOC库.因此, 这两个过程之间的平衡对SOC库变化的方向和程度起到至关重要的作用(Liang et al., 2018). ...
2
2013
... 自然界中已知的碳(C)同位素有7种(10C、11C、12C、13C、14C、15C和16C), 其中两种(12C和13C)为稳定碳同位素.不同同位素之间物理化学性质上的差异使反应前和反应后的物质在同位素组成上存在差异, 这种现象称为同位素分馏效应(陈世苹等, 2002; 林光辉, 2013).基于此, 碳同位素示踪技术是研究陆地生态系统碳氮循环过程的有效手段, 也是揭示全球气候变化成因、控制温室效应的重要工具(葛源等, 2006; Blaser & Conrad, 2016).碳同位素技术在研究土壤有机质的来源、动态转化与稳定性等方面具有高度的专一性和灵敏度, 且标记方法已比较成熟, 被广泛地应用到生态学研究中, 由原来在叶片、种群尺度的研究提高到了冠层、群落、生态系统乃至生物圈的水平, 从新的角度研究生物及其周围环境的关系(陈世苹等, 2002).13C稳定同位素作为示踪物, 与放射性碳同位素相比具有安全、无污染和容易控制等优点, 被广泛应用于陆地生态系统碳周转的生物地球化学过程研究. ...
... 碳同位素原子质量的微小差异, 使得含有不同同位素的物质具有不同的理化性质, 导致不同的动力学和热力学效应, 在物理、化学和生物等反应过程中出现同位素分馏现象.碳循环过程中, 同位素交换反应使无机盐(如碳酸盐)富集重同位素(如13C); 而碳的有机循环, 如光合作用的动力分馏效应导致生物成因碳(有机物)中富集轻同位素(如12C), 从而导致陆地生态系统中有机体和大气CO2之间的碳循环过程中, 出现明显的碳同位素分馏现象(Tieszen & Boutton, 1989).植物在光合作用过程中固定CO2途径的差异, 导致不同光合型植物选择吸收13C的比例不同, 从而使C3和C4植株体内的稳定碳同位素比值(δ13C)存在差异, 因此δ13C可以作为表征植物株生理生态特性差异的指标.土壤有机碳(SOC)经过微生物分解矿化为CO2的过程中13C分馏作用较小, 仅为1‰-3‰, 而SOC在厌氧条件下产生甲烷的过程中13C的分馏作用相对较大, 为10‰-70‰ (Conrad et al., 2010; 林光辉, 2013; 袁红朝等, 2014).但SOC主要来源于陆生高等植物, 与植物光合作用固定CO2时的分馏作用相比, SOC分解过程中的分馏作用几乎可以忽略不计, 因此SOC与该土壤上种植植物的δ13C基本一致(Fernandez et al., 2003; Crow et al., 2006; 张林等, 2010; Bai et al., 2012).所以, 通过测定各个碳库中碳同位素比值的变化, 可以量化“大气-植物-土壤”生态系统中碳的来源、分配及其周转规律. ...
2
2013
... 自然界中已知的碳(C)同位素有7种(10C、11C、12C、13C、14C、15C和16C), 其中两种(12C和13C)为稳定碳同位素.不同同位素之间物理化学性质上的差异使反应前和反应后的物质在同位素组成上存在差异, 这种现象称为同位素分馏效应(陈世苹等, 2002; 林光辉, 2013).基于此, 碳同位素示踪技术是研究陆地生态系统碳氮循环过程的有效手段, 也是揭示全球气候变化成因、控制温室效应的重要工具(葛源等, 2006; Blaser & Conrad, 2016).碳同位素技术在研究土壤有机质的来源、动态转化与稳定性等方面具有高度的专一性和灵敏度, 且标记方法已比较成熟, 被广泛地应用到生态学研究中, 由原来在叶片、种群尺度的研究提高到了冠层、群落、生态系统乃至生物圈的水平, 从新的角度研究生物及其周围环境的关系(陈世苹等, 2002).13C稳定同位素作为示踪物, 与放射性碳同位素相比具有安全、无污染和容易控制等优点, 被广泛应用于陆地生态系统碳周转的生物地球化学过程研究. ...
... 碳同位素原子质量的微小差异, 使得含有不同同位素的物质具有不同的理化性质, 导致不同的动力学和热力学效应, 在物理、化学和生物等反应过程中出现同位素分馏现象.碳循环过程中, 同位素交换反应使无机盐(如碳酸盐)富集重同位素(如13C); 而碳的有机循环, 如光合作用的动力分馏效应导致生物成因碳(有机物)中富集轻同位素(如12C), 从而导致陆地生态系统中有机体和大气CO2之间的碳循环过程中, 出现明显的碳同位素分馏现象(Tieszen & Boutton, 1989).植物在光合作用过程中固定CO2途径的差异, 导致不同光合型植物选择吸收13C的比例不同, 从而使C3和C4植株体内的稳定碳同位素比值(δ13C)存在差异, 因此δ13C可以作为表征植物株生理生态特性差异的指标.土壤有机碳(SOC)经过微生物分解矿化为CO2的过程中13C分馏作用较小, 仅为1‰-3‰, 而SOC在厌氧条件下产生甲烷的过程中13C的分馏作用相对较大, 为10‰-70‰ (Conrad et al., 2010; 林光辉, 2013; 袁红朝等, 2014).但SOC主要来源于陆生高等植物, 与植物光合作用固定CO2时的分馏作用相比, SOC分解过程中的分馏作用几乎可以忽略不计, 因此SOC与该土壤上种植植物的δ13C基本一致(Fernandez et al., 2003; Crow et al., 2006; 张林等, 2010; Bai et al., 2012).所以, 通过测定各个碳库中碳同位素比值的变化, 可以量化“大气-植物-土壤”生态系统中碳的来源、分配及其周转规律. ...
热分析-傅里叶变换红外光谱-气相色谱-质谱联用测定葡萄糖的热分解产物
1
2014
... 质谱法、核磁共振法和光谱法是测定稳定碳同位素常用的方法, 质谱法是稳定碳同位素分析中最通用、最精确的方法(Paul et al., 2007; Eiler et al., 2012; 袁红朝等, 2014).质谱法的原理是先将制备的样品转化成含所测元素的纯气体(CO2、H2、N2、SO2等), 经离子源电离, 形成的离子束在电场、磁场的作用下, 导致具有不同质荷比的离子束分开从而检测同位素丰度.普通质谱仪在计算机辅助下直接给出同位素比值, 更先进的仪器如美国热电公司的Thermo Scientific MAT253, 不仅具有更高的灵敏度和更宽的测量范围, 而且可以与同位素数据系统整合, 自动化分析样品的同位素比值.核磁共振法是在磁场恒定条件下, 基于共振时的射频电场频率来测定不同基团上的同位素丰度, 与碳同位素示踪技术相结合可以检测底物(如葡萄糖)在土壤碳循环过程中的化学组分动态(Webster et al., 1997).光谱法是根据组成分子的不同同位素质量数不同从而使红外光谱谱线发生位移的现象来测定碳同位素丰度.近年来, 随着元素生物地球化学循环研究的进一步发展, 以及元素分析仪-同位素质谱(EA-IRMS)和热分析-傅里叶变换红外光谱-气相色谱-质谱(STA-FTIR-GC-MS)等联用技术的兴起, 极大促进了碳同位素分析测试技术的发展(曹亚澄等, 2008; 刘春波等, 2014). ...
热分析-傅里叶变换红外光谱-气相色谱-质谱联用测定葡萄糖的热分解产物
1
2014
... 质谱法、核磁共振法和光谱法是测定稳定碳同位素常用的方法, 质谱法是稳定碳同位素分析中最通用、最精确的方法(Paul et al., 2007; Eiler et al., 2012; 袁红朝等, 2014).质谱法的原理是先将制备的样品转化成含所测元素的纯气体(CO2、H2、N2、SO2等), 经离子源电离, 形成的离子束在电场、磁场的作用下, 导致具有不同质荷比的离子束分开从而检测同位素丰度.普通质谱仪在计算机辅助下直接给出同位素比值, 更先进的仪器如美国热电公司的Thermo Scientific MAT253, 不仅具有更高的灵敏度和更宽的测量范围, 而且可以与同位素数据系统整合, 自动化分析样品的同位素比值.核磁共振法是在磁场恒定条件下, 基于共振时的射频电场频率来测定不同基团上的同位素丰度, 与碳同位素示踪技术相结合可以检测底物(如葡萄糖)在土壤碳循环过程中的化学组分动态(Webster et al., 1997).光谱法是根据组成分子的不同同位素质量数不同从而使红外光谱谱线发生位移的现象来测定碳同位素丰度.近年来, 随着元素生物地球化学循环研究的进一步发展, 以及元素分析仪-同位素质谱(EA-IRMS)和热分析-傅里叶变换红外光谱-气相色谱-质谱(STA-FTIR-GC-MS)等联用技术的兴起, 极大促进了碳同位素分析测试技术的发展(曹亚澄等, 2008; 刘春波等, 2014). ...
Initial utilization of rhizodeposits with rice growth in paddy soils: rhizosphere and N fertilization effects
1
2019a
... 采用13CO2脉冲标记技术, 对生长在不同SOM含量水稻土上的水稻进行标记, 发现高、低SOM水稻土壤对水稻光合碳的转化和周转具有显著差异, 揭示了水稻光合碳向地下传输过程的“马太效应”(Zhu et al., 2017b).在水稻拔节期和灌浆期内, 通过水稻的根际沉积作用有4%-6%的光合碳进入土壤有机碳库, 土壤可溶性有机碳的2%-4%和土壤微生物生物量碳的9%-18%来源于这部分新碳(Lu et al., 2002a, 2002b; 祝贞科等, 2016).此外, 利用13CO2脉冲标记结合生物标志物(磷脂脂肪酸)的稳定碳同位素探针技术(13C-PLFA-SIP), 发现了碳从水稻向土壤微生物快速转移的现象, 且随着水稻生长期的变化根际碳的微生物同化量显著增加, 真菌及革兰氏阴性菌是主要的同化微生物(Yuan et al., 2016; Liu et al., 2019a).采用13CO2连续标记技术研究了水稻光合碳的传输和转化数量特征及其对氮肥施用的响应, 发现了高氮肥施用条件下水稻光合碳在土壤中的传输和积累比率非线性骤升的规律(Bradford et al., 2008; Ge et al., 2017).结合13C-PLFA-SIP技术发现, 外源C (羧甲基纤维素)和氮肥的联合施用降低了革兰氏阳性菌(G+)和革兰氏阴性菌(G-)的比值, 降低了G+和真菌的丰度, 单施氮肥则刺激了G-和放线菌的增长, 说明可利用碳氮的不同施用策略可显著影响土壤微生物的群落结构(Zhao et al., 2019).Atere等(2017)利用13CO2连续标记技术和土壤有机质的粒径组分分离技术联用, 发现水稻光合碳的固定效率在氮肥施用结合干湿交替条件下有显著提高.连续标记法有助于定量全部生长期或某一生长期全部投入C的净分配, 但是由于成本高以及技术困难, 运用受到限制. ...
Carbon input and allocation by rice into paddy soils: a review
2
2019b
... 13CO2同位素标记法, 是指以高丰度13CO2为示踪物, 在植物通过光合作用固定这些示踪物后, 通过在特定的时间内测定植物-土壤系统中各碳库示踪物量的变化, 可定量研究该时间段内13C在植物-土壤系统的转移和分布, 阐明植物某一生长期的碳分配特征, 定量植物碳向土壤的输入以及土壤碳周转, 从而明确植物光合碳对SOM的来源、稳定化过程的影响及其微生物驱动机制(Hanson et al., 2000; Kuzyakov & Cheng, 2001; Kuzyakov, 2006; Yuan et al., 2016; 张蕊等, 2017; Liu et al., 2019b).脉冲标记法是一次或分几次加入13CO2, 标记数小时后在特定时间内测定13C在土壤-植物系统各碳库的分配比例(Kuzyakov & Gavrichkova, 2010)(表1).脉冲标记法只能提供植物生长关键时期光合碳的分配特征, 但是在植物生长期内标记13CO2, 就可以估计植物来源碳输入土壤各组分的量(Lu et al., 2002a; Kimura et al., 2004; Xiao et al., 2019a, 2019b; Zang et al., 2019).连续标记法是指在植物生长某个生长期(或全生长期), 在可控气室等封闭的空间内对植物进行不间断标记, 可以定量全部生长期投入碳的净分配(表1).例如, 在水稻(Oryza satira)拔节期和灌浆期进行13CO2连续标记, 可以定量研究生长过程中水稻光合碳对SOC各组分的贡献(Kuzyakov, 2005; Conrad et al., 2012; Wang et al., 2016), 以及对土壤原有有机质的激发效应(Zhu et al., 2018a). ...
... 目前应用碳同位素示踪技术对C3/C4植被演替、土壤有机质源汇和各组分的周转及其微生物响应机制等方面进行了大量研究并日趋成熟(Six et al., 2001; Kuzyakov, 2006; Werth & Kuzyakov, 2010; Wang et al., 2016; Liu et al., 2019b), 而对于植物-土壤碳循环中植物、微生物以及土壤动物(如原生动物、线虫和节肢动物等)所扮演的角色及其循环机理研究较少.尽管碳同位素示踪在植物生理生态研究中已发挥重要作用, 但许多物理、化学和生物过程还不十分清楚; 放牧、施肥、耕作等人类活动等多种因素可直接影响或与气候协同作用影响植被变化以及导致土壤有机质变化的综合研究尚需深入; 还缺乏碳与其他养分元素的化学计量学方面的研究, 以及碳、氮、磷、硫、氧等多种同位素联合探测技术在生态学领域的综合开发及应用.鉴于此, 我们提出以下四点展望: ...
In situ stable isotope probing of methanogenic archaea in the rice rhizosphere
1
2005
... 将稳定碳同位素示踪技术与分子生物学方法相结合, 在表征以微生物为主要研究对象的土壤生物群落功能过程研究中发挥了重要作用(Lu & Conrad, 2005; Ruyters et al., 2013; Fan et al., 2014).13C同位素探针-磷脂脂肪酸(PLFA-SIP)是在土壤碳循环微生物驱动机制的研究中广泛应用的技术.该方法灵敏度高, 常用于指示活体微生物生物量和群落结构, 能够定量检测微生物对标记底物的同化利用.近年来, 以氨基糖为主要成分的微生物残留物的相对稳定性及其对SOC的贡献得到广泛关注(Liang et al., 2017), 将PLFA-SIP与氨基糖生物标志物相结合, 建立外源碳-微生物-土壤碳库的碳周转连续体模型, 有助于揭示SOC的累积机制.PLFA-SIP对物种组成的分辨率较低, 难以判断功能微生物, 且无法检测古菌.以DNA/RNA测序为基础的DNA/RNA-SIP技术可以很好地解决上述问题, 只要选择合适的底物和目的基因, DNA/RNA-SIP技术能准确解析关键微生物在土壤中的群落结构和功能变化, 特异性地将微生物群体与生态过程、生理特性相关联, 揭示微生物驱动下的土壤元素循环过程及其调控机制.近年来, Hungate等(2015)将13C与18O标记相结合, 建立了定量DNA-SIP技术(DNA-qSIP), 揭示了参与土壤中新输入的有机碳(新碳)和土壤原有有机碳(老 碳)周转过程的微生物群落差异(Morrissey et al., 2017). ...
Contribution of plant-derived carbon to soil microbial biomass dynamics in a paddy rice microcosm
2
2002a
... 13CO2同位素标记法, 是指以高丰度13CO2为示踪物, 在植物通过光合作用固定这些示踪物后, 通过在特定的时间内测定植物-土壤系统中各碳库示踪物量的变化, 可定量研究该时间段内13C在植物-土壤系统的转移和分布, 阐明植物某一生长期的碳分配特征, 定量植物碳向土壤的输入以及土壤碳周转, 从而明确植物光合碳对SOM的来源、稳定化过程的影响及其微生物驱动机制(Hanson et al., 2000; Kuzyakov & Cheng, 2001; Kuzyakov, 2006; Yuan et al., 2016; 张蕊等, 2017; Liu et al., 2019b).脉冲标记法是一次或分几次加入13CO2, 标记数小时后在特定时间内测定13C在土壤-植物系统各碳库的分配比例(Kuzyakov & Gavrichkova, 2010)(表1).脉冲标记法只能提供植物生长关键时期光合碳的分配特征, 但是在植物生长期内标记13CO2, 就可以估计植物来源碳输入土壤各组分的量(Lu et al., 2002a; Kimura et al., 2004; Xiao et al., 2019a, 2019b; Zang et al., 2019).连续标记法是指在植物生长某个生长期(或全生长期), 在可控气室等封闭的空间内对植物进行不间断标记, 可以定量全部生长期投入碳的净分配(表1).例如, 在水稻(Oryza satira)拔节期和灌浆期进行13CO2连续标记, 可以定量研究生长过程中水稻光合碳对SOC各组分的贡献(Kuzyakov, 2005; Conrad et al., 2012; Wang et al., 2016), 以及对土壤原有有机质的激发效应(Zhu et al., 2018a). ...
... 采用13CO2脉冲标记技术, 对生长在不同SOM含量水稻土上的水稻进行标记, 发现高、低SOM水稻土壤对水稻光合碳的转化和周转具有显著差异, 揭示了水稻光合碳向地下传输过程的“马太效应”(Zhu et al., 2017b).在水稻拔节期和灌浆期内, 通过水稻的根际沉积作用有4%-6%的光合碳进入土壤有机碳库, 土壤可溶性有机碳的2%-4%和土壤微生物生物量碳的9%-18%来源于这部分新碳(Lu et al., 2002a, 2002b; 祝贞科等, 2016).此外, 利用13CO2脉冲标记结合生物标志物(磷脂脂肪酸)的稳定碳同位素探针技术(13C-PLFA-SIP), 发现了碳从水稻向土壤微生物快速转移的现象, 且随着水稻生长期的变化根际碳的微生物同化量显著增加, 真菌及革兰氏阴性菌是主要的同化微生物(Yuan et al., 2016; Liu et al., 2019a).采用13CO2连续标记技术研究了水稻光合碳的传输和转化数量特征及其对氮肥施用的响应, 发现了高氮肥施用条件下水稻光合碳在土壤中的传输和积累比率非线性骤升的规律(Bradford et al., 2008; Ge et al., 2017).结合13C-PLFA-SIP技术发现, 外源C (羧甲基纤维素)和氮肥的联合施用降低了革兰氏阳性菌(G+)和革兰氏阴性菌(G-)的比值, 降低了G+和真菌的丰度, 单施氮肥则刺激了G-和放线菌的增长, 说明可利用碳氮的不同施用策略可显著影响土壤微生物的群落结构(Zhao et al., 2019).Atere等(2017)利用13CO2连续标记技术和土壤有机质的粒径组分分离技术联用, 发现水稻光合碳的固定效率在氮肥施用结合干湿交替条件下有显著提高.连续标记法有助于定量全部生长期或某一生长期全部投入C的净分配, 但是由于成本高以及技术困难, 运用受到限制. ...
Input and distribution of photosynthesized carbon in a flooded rice soil
1
2002b
... 采用13CO2脉冲标记技术, 对生长在不同SOM含量水稻土上的水稻进行标记, 发现高、低SOM水稻土壤对水稻光合碳的转化和周转具有显著差异, 揭示了水稻光合碳向地下传输过程的“马太效应”(Zhu et al., 2017b).在水稻拔节期和灌浆期内, 通过水稻的根际沉积作用有4%-6%的光合碳进入土壤有机碳库, 土壤可溶性有机碳的2%-4%和土壤微生物生物量碳的9%-18%来源于这部分新碳(Lu et al., 2002a, 2002b; 祝贞科等, 2016).此外, 利用13CO2脉冲标记结合生物标志物(磷脂脂肪酸)的稳定碳同位素探针技术(13C-PLFA-SIP), 发现了碳从水稻向土壤微生物快速转移的现象, 且随着水稻生长期的变化根际碳的微生物同化量显著增加, 真菌及革兰氏阴性菌是主要的同化微生物(Yuan et al., 2016; Liu et al., 2019a).采用13CO2连续标记技术研究了水稻光合碳的传输和转化数量特征及其对氮肥施用的响应, 发现了高氮肥施用条件下水稻光合碳在土壤中的传输和积累比率非线性骤升的规律(Bradford et al., 2008; Ge et al., 2017).结合13C-PLFA-SIP技术发现, 外源C (羧甲基纤维素)和氮肥的联合施用降低了革兰氏阳性菌(G+)和革兰氏阴性菌(G-)的比值, 降低了G+和真菌的丰度, 单施氮肥则刺激了G-和放线菌的增长, 说明可利用碳氮的不同施用策略可显著影响土壤微生物的群落结构(Zhao et al., 2019).Atere等(2017)利用13CO2连续标记技术和土壤有机质的粒径组分分离技术联用, 发现水稻光合碳的固定效率在氮肥施用结合干湿交替条件下有显著提高.连续标记法有助于定量全部生长期或某一生长期全部投入C的净分配, 但是由于成本高以及技术困难, 运用受到限制. ...
Carbon dynamics of rhizodeposits, root- and shoot-residues in a rice soil
1
2003
... 13CO2脉冲标记在密闭的植物生长箱中进行, 当植物生长到一定时期, 在光照充足时通入13CO2或通过化学反应(如NaH13CO3和H2SO4)获得13CO2, 标记数个小时后移出, 在标记期间由CO2发生器和检测系统实时监测并控制CO2浓度, 生长箱内温湿度需与对照处理(未标记13CO2的植物)保持一致(Kuzyakov & Gavrichkova, 2010); 为避免13CO2污染, 对照处理要距离标记的植物至少5 m以上(Lu et al., 2003).13CO2连续标记的方法与脉冲标记基本一致, 但是并不是短时间标记, 而是在植物生长某个生长期, 甚至整个生长期内, 对植物进行不间断标记. ...
Free-air carbon dioxide enrichment (FACE) in global change research: a review
1
1999
... 13C贫化示踪技术结合的FACE实验是研究CO2浓度增加的微域环境中SOC周转的常用方法.大气CO2的δ13C值约为-8‰, 在13C贫化示踪技术结合的FACE实验(13C-FACE)中供给13C贫化的CO2的δ13C值远低于大气, 其范围是-45‰- -18‰ (张蕊等, 2017).FACE系统是一个开放的模拟未来CO2增加的微域环境, 系统内部的气候条件如水分、光照、温度等非常接近自然生态环境.因此, 在这样的系统内进行模拟CO2升高的实验更接近于真实状况(McLeod & Long, 1999).目前全球已建立十余个涉及森林(Hagedorn et al., 2003), 农田(Leavitt et al., 1994)和草地(Six et al., 2001)等多个生态系统和多种植被类型的13C-FACE平台, 阐释了大气CO2浓度升高条件下, 不同生态系统植物生理与光合效率、养分和水分利用、土壤呼吸、土壤碳的输入量及周转率和微生物响应等过程机制(Leavitt et al., 1994; Canadell et al., 1995; Kiem et al., 2000). ...
Bacterial carbon use plasticity, phylogenetic diversity and the priming of soil organic matter
1
2017
... 将稳定碳同位素示踪技术与分子生物学方法相结合, 在表征以微生物为主要研究对象的土壤生物群落功能过程研究中发挥了重要作用(Lu & Conrad, 2005; Ruyters et al., 2013; Fan et al., 2014).13C同位素探针-磷脂脂肪酸(PLFA-SIP)是在土壤碳循环微生物驱动机制的研究中广泛应用的技术.该方法灵敏度高, 常用于指示活体微生物生物量和群落结构, 能够定量检测微生物对标记底物的同化利用.近年来, 以氨基糖为主要成分的微生物残留物的相对稳定性及其对SOC的贡献得到广泛关注(Liang et al., 2017), 将PLFA-SIP与氨基糖生物标志物相结合, 建立外源碳-微生物-土壤碳库的碳周转连续体模型, 有助于揭示SOC的累积机制.PLFA-SIP对物种组成的分辨率较低, 难以判断功能微生物, 且无法检测古菌.以DNA/RNA测序为基础的DNA/RNA-SIP技术可以很好地解决上述问题, 只要选择合适的底物和目的基因, DNA/RNA-SIP技术能准确解析关键微生物在土壤中的群落结构和功能变化, 特异性地将微生物群体与生态过程、生理特性相关联, 揭示微生物驱动下的土壤元素循环过程及其调控机制.近年来, Hungate等(2015)将13C与18O标记相结合, 建立了定量DNA-SIP技术(DNA-qSIP), 揭示了参与土壤中新输入的有机碳(新碳)和土壤原有有机碳(老 碳)周转过程的微生物群落差异(Morrissey et al., 2017). ...
Tracking microbial interactions with NanoSIMS
2
2016
... 纳米二次离子质谱(NanoSIMS)能够完成对元素空间分布的超高分辨率显微镜成像, 与同位素示踪技术相结合, 在生态系统微观机制研究领域中显示出了巨大的潜力(Herrmann et al., 2007; Musat et al., 2016).该技术具有较高的灵敏度和准确度, 可应用于碳、氮、硫等元素生物地球化学循环的微生物驱动机制和土壤-微生物-植物相关作用关系的研究, 对于认识多元素耦合的生物地球化学循环规律具有重要意义(图1)(Lechene et al., 2007; Berry et al., 2013; Rumpel et al., 2015).目前, 稳定碳同位素探针-纳米二次离子质谱技术(NanoSIP)已在植物/动物-微生物共生关系(Lechene et al., 2007; Vidal et al., 2016), 微生物群落中碳的食物链传递(Musat et al., 2016), 厌氧甲烷氧化的碳氮耦合机制(Dekas et al., 2009)等研究中取得重要突破.在土壤有机碳周转的研究中, NanoSIP直接呈现了外源碳与土壤矿物的结合形态与动态过程, 有力推动了SOC的稳定机制的研究(Rumpel et al., 2015). ...
... ), 微生物群落中碳的食物链传递(Musat et al., 2016), 厌氧甲烷氧化的碳氮耦合机制(Dekas et al., 2009)等研究中取得重要突破.在土壤有机碳周转的研究中, NanoSIP直接呈现了外源碳与土壤矿物的结合形态与动态过程, 有力推动了SOC的稳定机制的研究(Rumpel et al., 2015). ...
Marine methylotrophs revealed by stable-isotope probing, multiple displacement amplification and metagenomics
1
2008
... 尽管PFLA-SIP、DNA/RNA-SIP技术有力推动了微生物群落动态与生态系统功能的关联研究, 但生物地球化学循环微生物驱动机制的深入揭示仍需要稳定碳同位素示踪技术与多组学技术的结合.稳定碳同位素探针-宏基因组/宏转录组技术能够揭示特定生态过程中活性微生物群落组成、功能多样性、生态网络关系等, 实现对碳氮等元素微生物周转过程的代谢通路预测和重构(Dumont et al., 2006; Neufeld et al., 2008).宏基因组和宏转录组整合分析了代谢过程的可能性, 蛋白质组学、代谢组学技术则为特定底物的代谢通路的建立提供直接依据, 是生态系统中生物地球化学循环过程完整解析的必经之路. ...
Normalization of measured stable isotopic compositions to isotope reference scales—A review
1
2007
... 质谱法、核磁共振法和光谱法是测定稳定碳同位素常用的方法, 质谱法是稳定碳同位素分析中最通用、最精确的方法(Paul et al., 2007; Eiler et al., 2012; 袁红朝等, 2014).质谱法的原理是先将制备的样品转化成含所测元素的纯气体(CO2、H2、N2、SO2等), 经离子源电离, 形成的离子束在电场、磁场的作用下, 导致具有不同质荷比的离子束分开从而检测同位素丰度.普通质谱仪在计算机辅助下直接给出同位素比值, 更先进的仪器如美国热电公司的Thermo Scientific MAT253, 不仅具有更高的灵敏度和更宽的测量范围, 而且可以与同位素数据系统整合, 自动化分析样品的同位素比值.核磁共振法是在磁场恒定条件下, 基于共振时的射频电场频率来测定不同基团上的同位素丰度, 与碳同位素示踪技术相结合可以检测底物(如葡萄糖)在土壤碳循环过程中的化学组分动态(Webster et al., 1997).光谱法是根据组成分子的不同同位素质量数不同从而使红外光谱谱线发生位移的现象来测定碳同位素丰度.近年来, 随着元素生物地球化学循环研究的进一步发展, 以及元素分析仪-同位素质谱(EA-IRMS)和热分析-傅里叶变换红外光谱-气相色谱-质谱(STA-FTIR-GC-MS)等联用技术的兴起, 极大促进了碳同位素分析测试技术的发展(曹亚澄等, 2008; 刘春波等, 2014). ...
Photoassimilate allocation and dynamics of hotspots in roots visualized by 14C phosphor imaging
1
2011
... 尽管NanoSIP能够完成对土壤稳定碳同位素空间分布的超高分辨率成像, 但成本太高, 且测量范围在纳米尺度(Rumpel et al., 2015).将14C同位素示踪与放射自显影相结合, 能够直观呈现植物、土壤碳的分布与传输过程, 为准确解析碳元素源-库关系和分配动态提供了有力工具(Pausch & Kuzyakov, 2011).在利用放射自显影技术研究14C在土壤中的空间分布时, 土壤颗粒对β-粒子的激发、电离和轫致辐射等过程可强烈削弱14C信号, 通过基质转移(Holz et al., 2018)、衰减系数核算(Holz et al., 2019)等手段有望校正测定偏差, 提高该技术在土壤样品检测中的可信度.磷光体成像的应用提高了放射自显影技术的安全性和灵敏度, 为其在生态学的推广应用提供了良好的机遇.同位素示踪与超高分辨率成像技术相结合, 能在微观尺度上研究碳周转中植物-微生物-土壤的互相作用过程及碳与其他元素的耦合关系, 对全球碳循环的机理揭示有重要意义. ...
Elevated atmospheric CO2 effects and soil water feedbacks on soil respiration components in a Colorado grassland
1
2003
... 在草地生态系统中, FACE实验研究发现CO2浓度升高, 黑麦草(Lolium perenne)草地0-10 cm 土层土壤团聚体中的颗粒有机碳增加了40%, 表明大气CO2浓度升高可促进草地生态系统的碳周转(Six et al., 2001).在森林生态系统中, Griepentrog等(2015)也发现, CO2浓度升高增加了C3植物向土壤碳的输入量, 提高了土壤碳的周转率.新碳在SOM中的积累程度受有机质密度的影响, 有机质密度增加, 新碳的积累程度减小, 大部分的新碳进入到颗粒态有机质中(Dorodnikov et al., 2011; Hofmockel et al., 2011).同时, 13C-FACE结合PLFA技术, 进一步揭示了CO2浓度升高条件下革兰氏阴性菌和真菌可矿化更多的植物来源碳, 促进了植物新合成的光合碳的分解, 也加快了SOM的分解速率(Pendall et al., 2003; Hagedorn et al., 2013). ...
Variability of plant nitrogen and water use in a 100-m transect of a subdesertic depression of the Ebro valley (Spain) characterized by leaf δ13C and δ15N
1
1999
... 基于C3/C4植物δ13C的不同, 碳同位素组成可以用于区分植物的C3和C4光合途径以及不同植物功能群结构变化(Garten Jr. & Taylor Jr., 1992; Peñuelas et al., 1999).通过测定不同碳组分(可溶性碳水化合物和结构性碳库)的δ13C值, 从而估计植物水分利用效率和光合作用在不同时间尺度上的变化(Ebdon et al., 1998; Arslan et al., 1999; 陈世苹等, 2002).而在一个复杂的生态系统(如热带雨林)中, 水分利用效率的高低可能影响植物种间竞争的结果, 最终导致生态系统内部群落结构的差异, 因此δ13C为指示生 ...
Universality of priming effect: an analysis using thirty five soils with contrasted properties sampled from five continents
1
2019
... SOC周转是一个连续动态变化的过程, 利用碳同位素技术能够示踪新碳在土壤中的转化过程和赋存状态, 揭示外源碳在土壤-微生物之间的周转过程和机理(金鑫鑫等, 2017).利用13C富集标记小分子有机物或是植物秸秆, 结合生物标志物技术等, 可以表征不同来源的有机质在土壤中的相对丰度和降解状况, 以及SOC的动态变化和储量的转化和迁移, 也可以定量评价新老SOC对碳储量的相对贡献, 可有效评估陆地生态系统碳平衡过程及其微观过程机制(Kuzyakov & Domanski, 2000; Kemmitt et al., 2008; Ge et al., 2017; Zhu et al., 2017a).外源C输入土壤后可以通过物理和生物化学途径形成新的SOM (Cotrufo et al., 2013, 2015), 从而增加SOC库; 另一方面, 外源C的输入也会促进土壤中原有SOM降解(Guenet et al., 2018; Zhu et al., 2018b; Perveen et al., 2019), 从而减少SOC库.因此, 这两个过程之间的平衡对SOC库变化的方向和程度起到至关重要的作用(Liang et al., 2018). ...
Source partitioning using stable isotopes: coping with too many sources
1
2003
... 基于质量平衡原理解析土壤-植物-微生物连续体系中有机物的二元或三元组成与贡献, 量化生态系统中小分子根系分泌物-植物残体-生物质炭-土壤有机质-微生物活体与残留物等碳源的相对比例, 揭示生态系统碳源流通过程与机理(Phillips & Gregg, 2003; Whitman & Lehmann, 2015; Liang et al., 2017).然而, 生态系统中有机物是多碳源组成的, 质量平衡并不能适应所有环境条件下有机物的源解析.因此, 需要基于同位素示踪基本原理, 构建合理的方程模型与原创性理论, 准确区分与量化复杂生态系统中有机碳的来源与周转通量, 揭示生态系统碳循环与调控机理. ...
Stable-isotope probing as a tool in microbial ecology
1
2000
... 随着碳同位素示踪技术的广泛应用和迅速发展, 多种碳同位素示踪技术及其与其他光谱和微生物探针技术联用, 对于揭示从微观到宏观尺度的碳循环过程起着重要促进作用(图1)(Radajewski et al., 2000; Herrmann et al., 2007).目前自然丰度法、与13C贫化示踪技术结合的自由空气中气体浓度增加(Free-Air Concentration Enrichment, FACE)实验、脉冲与连续标记法以及碳同位素高丰度底物富集标记法是研究陆地生态系统中碳循环过程常用的碳同位素示踪方法(Balesdent & Mariotti, 1996; Bernoux et al., 1998; Conley et al., 2001; Epron et al., 2012; Ge et al., 2017; 张蕊等, 2017).同时, 随着氢、氮(N)、氧等元素同位素示踪技术的发展与应用, 那些单独用碳同位素无法解决的问题在与其他同位素示踪技术、光谱和分子生物学技术的联用下得以解决(陈世苹等, 2002).例如, Kušlienë等(2014)利用13C/15N双标记法, 结合生物标志物分析, 研究了13C/15N标记的不同类型的根系分泌物对土壤碳氮动态的贡献, 这些技术的联用进一步促进了碳同位素示踪技术在生态学研究中的广泛应用.本文重点从稳定碳同位素示踪技术的原理、方法与应用进展等方面综述其在陆地生态系统碳循环过程中的研究进展. ...
中国区域478种C3植物叶片碳稳定性同位素组成与水分利用效率
1
2011
... 态系统的功能群多样性以及区分不同功能群植物提供了一条重要途径(Garten Jr & Taylor Jr, 1992; 陈世苹等, 2002).任书杰和于贵瑞(2011)整合了中国区域的数据发现水分条件越好, C3植物叶片δ13C越低, 因此C3植物叶片的δ13C可以作为指示植物的水分利用效率的指标.C3/C4植物交互种植时会改变土壤有机质的δ13C, 因此, 以未交互种植作物土壤为对照, 通过土壤有机碳δ13C的变化可以研究土壤有机碳的周转速度, 以及不同植物来源碳占土壤各碳库和呼吸的比例以及激发效应的方向和强度(Kuzyakov, 2010; Werth & Kuzyakov, 2010; Blagodatskaya et al., 2011a), 也可以通过土壤碳自然丰度δ13C的变化来研究土壤碳库中活性组分碳(微生物生物量碳(MBC))的周转速率和稳定性(Blagodatskaya et al., 2011b).研究结果发现, SOC和MBC的周转时间分别为16.8 a和29-30 d, 且随种植时间增加, 周转时间延长.新碳和老碳对土壤各碳库和呼吸的贡献不同, MBC中20%的碳来源于老碳(来自C3作物), CO2气体中60%的碳来源于老碳(来自C3作物), 由于土壤微生物偏好矿化老碳而同化新碳, SOC中新碳(来自C4作物)的贡献将逐年增加.此外, 长期种植C3作物(小麦(Triticum aestivum))的土壤在连续耕种C4作物(玉米(zea mays)) 13年后, 土壤有机碳库更新了22%, 且SOC的周转速率在不同粒径团聚体中各不相同, 在>50 µm与<2 µm粒径的团聚体中包含更多新碳, 而黏粒中有机碳的更新速度最慢(Balesdent & Mariotti, 1996; 袁红朝等, 2014).13C自然丰度法为明确土壤新老碳库对SOC、MBC和CO2排放的贡献, 解析陆地生态系统δ13C的变化及其在全球碳平衡中的应用提供了重要依据(Cheng, 2009).随着测试精度的提高, 采用碳同位素配对标记实验, 只要SOC和其他一种或是两种碳源的碳同位素比值间有较为明显的差别, 也可以解析多碳源相互作用及其对土壤呼吸的贡献, 这对于研究复杂生态系统中碳通量和净碳平衡具有重要的应用价值(Whitman & Lehmann, 2015). ...
中国区域478种C3植物叶片碳稳定性同位素组成与水分利用效率
1
2011
... 态系统的功能群多样性以及区分不同功能群植物提供了一条重要途径(Garten Jr & Taylor Jr, 1992; 陈世苹等, 2002).任书杰和于贵瑞(2011)整合了中国区域的数据发现水分条件越好, C3植物叶片δ13C越低, 因此C3植物叶片的δ13C可以作为指示植物的水分利用效率的指标.C3/C4植物交互种植时会改变土壤有机质的δ13C, 因此, 以未交互种植作物土壤为对照, 通过土壤有机碳δ13C的变化可以研究土壤有机碳的周转速度, 以及不同植物来源碳占土壤各碳库和呼吸的比例以及激发效应的方向和强度(Kuzyakov, 2010; Werth & Kuzyakov, 2010; Blagodatskaya et al., 2011a), 也可以通过土壤碳自然丰度δ13C的变化来研究土壤碳库中活性组分碳(微生物生物量碳(MBC))的周转速率和稳定性(Blagodatskaya et al., 2011b).研究结果发现, SOC和MBC的周转时间分别为16.8 a和29-30 d, 且随种植时间增加, 周转时间延长.新碳和老碳对土壤各碳库和呼吸的贡献不同, MBC中20%的碳来源于老碳(来自C3作物), CO2气体中60%的碳来源于老碳(来自C3作物), 由于土壤微生物偏好矿化老碳而同化新碳, SOC中新碳(来自C4作物)的贡献将逐年增加.此外, 长期种植C3作物(小麦(Triticum aestivum))的土壤在连续耕种C4作物(玉米(zea mays)) 13年后, 土壤有机碳库更新了22%, 且SOC的周转速率在不同粒径团聚体中各不相同, 在>50 µm与<2 µm粒径的团聚体中包含更多新碳, 而黏粒中有机碳的更新速度最慢(Balesdent & Mariotti, 1996; 袁红朝等, 2014).13C自然丰度法为明确土壤新老碳库对SOC、MBC和CO2排放的贡献, 解析陆地生态系统δ13C的变化及其在全球碳平衡中的应用提供了重要依据(Cheng, 2009).随着测试精度的提高, 采用碳同位素配对标记实验, 只要SOC和其他一种或是两种碳源的碳同位素比值间有较为明显的差别, 也可以解析多碳源相互作用及其对土壤呼吸的贡献, 这对于研究复杂生态系统中碳通量和净碳平衡具有重要的应用价值(Whitman & Lehmann, 2015). ...
Separating soil respiration into plant and soil components using analyses of the natural abundance of carbon-13
1
1999
... 13C自然丰度法是基于C3/C4植物具有不同的δ13C, 将C3 (C4)植物种植在具有C4 (C3)植物种植历史的土壤, 植物来源与原有SOM来源的δ13C存在差异, 可以以此来区分土壤源呼吸和根源呼吸的C (Kingston et al., 1994).该方法成本低且容易操作, 但是需要注意的是, 选取的C4 (C3)土壤需要长期一直种植C4 (C3)植物, 且该方法灵敏度相对较低, 在植物生长季后期可靠性下降(Rochette et al., 1999). ...
Nanoscale evidence of contrasted processes for root-derived organic matter stabilization by mineral interactions depending on soil depth
3
2015
... 纳米二次离子质谱(NanoSIMS)能够完成对元素空间分布的超高分辨率显微镜成像, 与同位素示踪技术相结合, 在生态系统微观机制研究领域中显示出了巨大的潜力(Herrmann et al., 2007; Musat et al., 2016).该技术具有较高的灵敏度和准确度, 可应用于碳、氮、硫等元素生物地球化学循环的微生物驱动机制和土壤-微生物-植物相关作用关系的研究, 对于认识多元素耦合的生物地球化学循环规律具有重要意义(图1)(Lechene et al., 2007; Berry et al., 2013; Rumpel et al., 2015).目前, 稳定碳同位素探针-纳米二次离子质谱技术(NanoSIP)已在植物/动物-微生物共生关系(Lechene et al., 2007; Vidal et al., 2016), 微生物群落中碳的食物链传递(Musat et al., 2016), 厌氧甲烷氧化的碳氮耦合机制(Dekas et al., 2009)等研究中取得重要突破.在土壤有机碳周转的研究中, NanoSIP直接呈现了外源碳与土壤矿物的结合形态与动态过程, 有力推动了SOC的稳定机制的研究(Rumpel et al., 2015). ...
... )等研究中取得重要突破.在土壤有机碳周转的研究中, NanoSIP直接呈现了外源碳与土壤矿物的结合形态与动态过程, 有力推动了SOC的稳定机制的研究(Rumpel et al., 2015). ...
... 尽管NanoSIP能够完成对土壤稳定碳同位素空间分布的超高分辨率成像, 但成本太高, 且测量范围在纳米尺度(Rumpel et al., 2015).将14C同位素示踪与放射自显影相结合, 能够直观呈现植物、土壤碳的分布与传输过程, 为准确解析碳元素源-库关系和分配动态提供了有力工具(Pausch & Kuzyakov, 2011).在利用放射自显影技术研究14C在土壤中的空间分布时, 土壤颗粒对β-粒子的激发、电离和轫致辐射等过程可强烈削弱14C信号, 通过基质转移(Holz et al., 2018)、衰减系数核算(Holz et al., 2019)等手段有望校正测定偏差, 提高该技术在土壤样品检测中的可信度.磷光体成像的应用提高了放射自显影技术的安全性和灵敏度, 为其在生态学的推广应用提供了良好的机遇.同位素示踪与超高分辨率成像技术相结合, 能在微观尺度上研究碳周转中植物-微生物-土壤的互相作用过程及碳与其他元素的耦合关系, 对全球碳循环的机理揭示有重要意义. ...
Activity of the ammonia oxidising bacteria is responsible for zinc tolerance development of the ammonia oxidising community in soil: a stable isotope probing study
1
2013
... 将稳定碳同位素示踪技术与分子生物学方法相结合, 在表征以微生物为主要研究对象的土壤生物群落功能过程研究中发挥了重要作用(Lu & Conrad, 2005; Ruyters et al., 2013; Fan et al., 2014).13C同位素探针-磷脂脂肪酸(PLFA-SIP)是在土壤碳循环微生物驱动机制的研究中广泛应用的技术.该方法灵敏度高, 常用于指示活体微生物生物量和群落结构, 能够定量检测微生物对标记底物的同化利用.近年来, 以氨基糖为主要成分的微生物残留物的相对稳定性及其对SOC的贡献得到广泛关注(Liang et al., 2017), 将PLFA-SIP与氨基糖生物标志物相结合, 建立外源碳-微生物-土壤碳库的碳周转连续体模型, 有助于揭示SOC的累积机制.PLFA-SIP对物种组成的分辨率较低, 难以判断功能微生物, 且无法检测古菌.以DNA/RNA测序为基础的DNA/RNA-SIP技术可以很好地解决上述问题, 只要选择合适的底物和目的基因, DNA/RNA-SIP技术能准确解析关键微生物在土壤中的群落结构和功能变化, 特异性地将微生物群体与生态过程、生理特性相关联, 揭示微生物驱动下的土壤元素循环过程及其调控机制.近年来, Hungate等(2015)将13C与18O标记相结合, 建立了定量DNA-SIP技术(DNA-qSIP), 揭示了参与土壤中新输入的有机碳(新碳)和土壤原有有机碳(老 碳)周转过程的微生物群落差异(Morrissey et al., 2017). ...
Impact of elevated CO2 on soil organic matter dynamics as related to changes in aggregate turnover and residue quality
3
2001
... 13C贫化示踪技术结合的FACE实验是研究CO2浓度增加的微域环境中SOC周转的常用方法.大气CO2的δ13C值约为-8‰, 在13C贫化示踪技术结合的FACE实验(13C-FACE)中供给13C贫化的CO2的δ13C值远低于大气, 其范围是-45‰- -18‰ (张蕊等, 2017).FACE系统是一个开放的模拟未来CO2增加的微域环境, 系统内部的气候条件如水分、光照、温度等非常接近自然生态环境.因此, 在这样的系统内进行模拟CO2升高的实验更接近于真实状况(McLeod & Long, 1999).目前全球已建立十余个涉及森林(Hagedorn et al., 2003), 农田(Leavitt et al., 1994)和草地(Six et al., 2001)等多个生态系统和多种植被类型的13C-FACE平台, 阐释了大气CO2浓度升高条件下, 不同生态系统植物生理与光合效率、养分和水分利用、土壤呼吸、土壤碳的输入量及周转率和微生物响应等过程机制(Leavitt et al., 1994; Canadell et al., 1995; Kiem et al., 2000). ...
... 在草地生态系统中, FACE实验研究发现CO2浓度升高, 黑麦草(Lolium perenne)草地0-10 cm 土层土壤团聚体中的颗粒有机碳增加了40%, 表明大气CO2浓度升高可促进草地生态系统的碳周转(Six et al., 2001).在森林生态系统中, Griepentrog等(2015)也发现, CO2浓度升高增加了C3植物向土壤碳的输入量, 提高了土壤碳的周转率.新碳在SOM中的积累程度受有机质密度的影响, 有机质密度增加, 新碳的积累程度减小, 大部分的新碳进入到颗粒态有机质中(Dorodnikov et al., 2011; Hofmockel et al., 2011).同时, 13C-FACE结合PLFA技术, 进一步揭示了CO2浓度升高条件下革兰氏阴性菌和真菌可矿化更多的植物来源碳, 促进了植物新合成的光合碳的分解, 也加快了SOM的分解速率(Pendall et al., 2003; Hagedorn et al., 2013). ...
... 目前应用碳同位素示踪技术对C3/C4植被演替、土壤有机质源汇和各组分的周转及其微生物响应机制等方面进行了大量研究并日趋成熟(Six et al., 2001; Kuzyakov, 2006; Werth & Kuzyakov, 2010; Wang et al., 2016; Liu et al., 2019b), 而对于植物-土壤碳循环中植物、微生物以及土壤动物(如原生动物、线虫和节肢动物等)所扮演的角色及其循环机理研究较少.尽管碳同位素示踪在植物生理生态研究中已发挥重要作用, 但许多物理、化学和生物过程还不十分清楚; 放牧、施肥、耕作等人类活动等多种因素可直接影响或与气候协同作用影响植被变化以及导致土壤有机质变化的综合研究尚需深入; 还缺乏碳与其他养分元素的化学计量学方面的研究, 以及碳、氮、磷、硫、氧等多种同位素联合探测技术在生态学领域的综合开发及应用.鉴于此, 我们提出以下四点展望: ...
Pulse-dynamic and monotonic decline patterns of soil respiration in long term laboratory microcosms
1
2014
... 13C同位素示踪法技术可以很好地研究外源C对SOM周转的影响.例如, 将13C标记的有机物加入土壤进行田间或室内培养实验, 通过测定13CO2可以区分外源有机物和原有SOM来源的碳, 量化外源有机物添加SOM分解的影响(Bernoux et al., 1998; Blagodatskaya & Kuzyakov, 2008).高丰度13C标记的有机物结合13C-PLFA-SIP和13C-DNA-SIP技术, 可以准确示踪微生物在土壤有机碳周转过程中的参与机制(Williams et al., 2006; Apostel et al., 2015; Wang et al., 2016).而13C-微生物残留物对于量化和评估微生物对SOM积累的贡献起到了关键作用(Tian et al., 2014; Zhang et al., 2015; Liang et al., 2017). ...
Stable carbon isotopes in terrestrial ecosystem research
1
1989
... 碳同位素原子质量的微小差异, 使得含有不同同位素的物质具有不同的理化性质, 导致不同的动力学和热力学效应, 在物理、化学和生物等反应过程中出现同位素分馏现象.碳循环过程中, 同位素交换反应使无机盐(如碳酸盐)富集重同位素(如13C); 而碳的有机循环, 如光合作用的动力分馏效应导致生物成因碳(有机物)中富集轻同位素(如12C), 从而导致陆地生态系统中有机体和大气CO2之间的碳循环过程中, 出现明显的碳同位素分馏现象(Tieszen & Boutton, 1989).植物在光合作用过程中固定CO2途径的差异, 导致不同光合型植物选择吸收13C的比例不同, 从而使C3和C4植株体内的稳定碳同位素比值(δ13C)存在差异, 因此δ13C可以作为表征植物株生理生态特性差异的指标.土壤有机碳(SOC)经过微生物分解矿化为CO2的过程中13C分馏作用较小, 仅为1‰-3‰, 而SOC在厌氧条件下产生甲烷的过程中13C的分馏作用相对较大, 为10‰-70‰ (Conrad et al., 2010; 林光辉, 2013; 袁红朝等, 2014).但SOC主要来源于陆生高等植物, 与植物光合作用固定CO2时的分馏作用相比, SOC分解过程中的分馏作用几乎可以忽略不计, 因此SOC与该土壤上种植植物的δ13C基本一致(Fernandez et al., 2003; Crow et al., 2006; 张林等, 2010; Bai et al., 2012).所以, 通过测定各个碳库中碳同位素比值的变化, 可以量化“大气-植物-土壤”生态系统中碳的来源、分配及其周转规律. ...
Incorporation of 13C labelled shoot residues in Lumbricus terrestris casts: a combination of transmission electron microscopy and nanoscale secondary ion mass spectrometry
1
2016
... 纳米二次离子质谱(NanoSIMS)能够完成对元素空间分布的超高分辨率显微镜成像, 与同位素示踪技术相结合, 在生态系统微观机制研究领域中显示出了巨大的潜力(Herrmann et al., 2007; Musat et al., 2016).该技术具有较高的灵敏度和准确度, 可应用于碳、氮、硫等元素生物地球化学循环的微生物驱动机制和土壤-微生物-植物相关作用关系的研究, 对于认识多元素耦合的生物地球化学循环规律具有重要意义(图1)(Lechene et al., 2007; Berry et al., 2013; Rumpel et al., 2015).目前, 稳定碳同位素探针-纳米二次离子质谱技术(NanoSIP)已在植物/动物-微生物共生关系(Lechene et al., 2007; Vidal et al., 2016), 微生物群落中碳的食物链传递(Musat et al., 2016), 厌氧甲烷氧化的碳氮耦合机制(Dekas et al., 2009)等研究中取得重要突破.在土壤有机碳周转的研究中, NanoSIP直接呈现了外源碳与土壤矿物的结合形态与动态过程, 有力推动了SOC的稳定机制的研究(Rumpel et al., 2015). ...
Incorporation of 13C-labelled rice rhizodeposition into soil microbial communities under different fertilizer applications
3
2016
... 13CO2同位素标记法, 是指以高丰度13CO2为示踪物, 在植物通过光合作用固定这些示踪物后, 通过在特定的时间内测定植物-土壤系统中各碳库示踪物量的变化, 可定量研究该时间段内13C在植物-土壤系统的转移和分布, 阐明植物某一生长期的碳分配特征, 定量植物碳向土壤的输入以及土壤碳周转, 从而明确植物光合碳对SOM的来源、稳定化过程的影响及其微生物驱动机制(Hanson et al., 2000; Kuzyakov & Cheng, 2001; Kuzyakov, 2006; Yuan et al., 2016; 张蕊等, 2017; Liu et al., 2019b).脉冲标记法是一次或分几次加入13CO2, 标记数小时后在特定时间内测定13C在土壤-植物系统各碳库的分配比例(Kuzyakov & Gavrichkova, 2010)(表1).脉冲标记法只能提供植物生长关键时期光合碳的分配特征, 但是在植物生长期内标记13CO2, 就可以估计植物来源碳输入土壤各组分的量(Lu et al., 2002a; Kimura et al., 2004; Xiao et al., 2019a, 2019b; Zang et al., 2019).连续标记法是指在植物生长某个生长期(或全生长期), 在可控气室等封闭的空间内对植物进行不间断标记, 可以定量全部生长期投入碳的净分配(表1).例如, 在水稻(Oryza satira)拔节期和灌浆期进行13CO2连续标记, 可以定量研究生长过程中水稻光合碳对SOC各组分的贡献(Kuzyakov, 2005; Conrad et al., 2012; Wang et al., 2016), 以及对土壤原有有机质的激发效应(Zhu et al., 2018a). ...
... 13C同位素示踪法技术可以很好地研究外源C对SOM周转的影响.例如, 将13C标记的有机物加入土壤进行田间或室内培养实验, 通过测定13CO2可以区分外源有机物和原有SOM来源的碳, 量化外源有机物添加SOM分解的影响(Bernoux et al., 1998; Blagodatskaya & Kuzyakov, 2008).高丰度13C标记的有机物结合13C-PLFA-SIP和13C-DNA-SIP技术, 可以准确示踪微生物在土壤有机碳周转过程中的参与机制(Williams et al., 2006; Apostel et al., 2015; Wang et al., 2016).而13C-微生物残留物对于量化和评估微生物对SOM积累的贡献起到了关键作用(Tian et al., 2014; Zhang et al., 2015; Liang et al., 2017). ...
... 目前应用碳同位素示踪技术对C3/C4植被演替、土壤有机质源汇和各组分的周转及其微生物响应机制等方面进行了大量研究并日趋成熟(Six et al., 2001; Kuzyakov, 2006; Werth & Kuzyakov, 2010; Wang et al., 2016; Liu et al., 2019b), 而对于植物-土壤碳循环中植物、微生物以及土壤动物(如原生动物、线虫和节肢动物等)所扮演的角色及其循环机理研究较少.尽管碳同位素示踪在植物生理生态研究中已发挥重要作用, 但许多物理、化学和生物过程还不十分清楚; 放牧、施肥、耕作等人类活动等多种因素可直接影响或与气候协同作用影响植被变化以及导致土壤有机质变化的综合研究尚需深入; 还缺乏碳与其他养分元素的化学计量学方面的研究, 以及碳、氮、磷、硫、氧等多种同位素联合探测技术在生态学领域的综合开发及应用.鉴于此, 我们提出以下四点展望: ...
Fates of 13C from enriched glucose and glycine in an organic soil determined by solid-state NMR
1
1997
... 质谱法、核磁共振法和光谱法是测定稳定碳同位素常用的方法, 质谱法是稳定碳同位素分析中最通用、最精确的方法(Paul et al., 2007; Eiler et al., 2012; 袁红朝等, 2014).质谱法的原理是先将制备的样品转化成含所测元素的纯气体(CO2、H2、N2、SO2等), 经离子源电离, 形成的离子束在电场、磁场的作用下, 导致具有不同质荷比的离子束分开从而检测同位素丰度.普通质谱仪在计算机辅助下直接给出同位素比值, 更先进的仪器如美国热电公司的Thermo Scientific MAT253, 不仅具有更高的灵敏度和更宽的测量范围, 而且可以与同位素数据系统整合, 自动化分析样品的同位素比值.核磁共振法是在磁场恒定条件下, 基于共振时的射频电场频率来测定不同基团上的同位素丰度, 与碳同位素示踪技术相结合可以检测底物(如葡萄糖)在土壤碳循环过程中的化学组分动态(Webster et al., 1997).光谱法是根据组成分子的不同同位素质量数不同从而使红外光谱谱线发生位移的现象来测定碳同位素丰度.近年来, 随着元素生物地球化学循环研究的进一步发展, 以及元素分析仪-同位素质谱(EA-IRMS)和热分析-傅里叶变换红外光谱-气相色谱-质谱(STA-FTIR-GC-MS)等联用技术的兴起, 极大促进了碳同位素分析测试技术的发展(曹亚澄等, 2008; 刘春波等, 2014). ...
13C fractionation at the root-microorganisms-soil interface: a review and outlook for partitioning studies
1
2010
... 目前应用碳同位素示踪技术对C3/C4植被演替、土壤有机质源汇和各组分的周转及其微生物响应机制等方面进行了大量研究并日趋成熟(Six et al., 2001; Kuzyakov, 2006; Werth & Kuzyakov, 2010; Wang et al., 2016; Liu et al., 2019b), 而对于植物-土壤碳循环中植物、微生物以及土壤动物(如原生动物、线虫和节肢动物等)所扮演的角色及其循环机理研究较少.尽管碳同位素示踪在植物生理生态研究中已发挥重要作用, 但许多物理、化学和生物过程还不十分清楚; 放牧、施肥、耕作等人类活动等多种因素可直接影响或与气候协同作用影响植被变化以及导致土壤有机质变化的综合研究尚需深入; 还缺乏碳与其他养分元素的化学计量学方面的研究, 以及碳、氮、磷、硫、氧等多种同位素联合探测技术在生态学领域的综合开发及应用.鉴于此, 我们提出以下四点展望: ...
A dualisotope approach to allow conclusive partitioning between three sources
2
2015
... 态系统的功能群多样性以及区分不同功能群植物提供了一条重要途径(Garten Jr & Taylor Jr, 1992; 陈世苹等, 2002).任书杰和于贵瑞(2011)整合了中国区域的数据发现水分条件越好, C3植物叶片δ13C越低, 因此C3植物叶片的δ13C可以作为指示植物的水分利用效率的指标.C3/C4植物交互种植时会改变土壤有机质的δ13C, 因此, 以未交互种植作物土壤为对照, 通过土壤有机碳δ13C的变化可以研究土壤有机碳的周转速度, 以及不同植物来源碳占土壤各碳库和呼吸的比例以及激发效应的方向和强度(Kuzyakov, 2010; Werth & Kuzyakov, 2010; Blagodatskaya et al., 2011a), 也可以通过土壤碳自然丰度δ13C的变化来研究土壤碳库中活性组分碳(微生物生物量碳(MBC))的周转速率和稳定性(Blagodatskaya et al., 2011b).研究结果发现, SOC和MBC的周转时间分别为16.8 a和29-30 d, 且随种植时间增加, 周转时间延长.新碳和老碳对土壤各碳库和呼吸的贡献不同, MBC中20%的碳来源于老碳(来自C3作物), CO2气体中60%的碳来源于老碳(来自C3作物), 由于土壤微生物偏好矿化老碳而同化新碳, SOC中新碳(来自C4作物)的贡献将逐年增加.此外, 长期种植C3作物(小麦(Triticum aestivum))的土壤在连续耕种C4作物(玉米(zea mays)) 13年后, 土壤有机碳库更新了22%, 且SOC的周转速率在不同粒径团聚体中各不相同, 在>50 µm与<2 µm粒径的团聚体中包含更多新碳, 而黏粒中有机碳的更新速度最慢(Balesdent & Mariotti, 1996; 袁红朝等, 2014).13C自然丰度法为明确土壤新老碳库对SOC、MBC和CO2排放的贡献, 解析陆地生态系统δ13C的变化及其在全球碳平衡中的应用提供了重要依据(Cheng, 2009).随着测试精度的提高, 采用碳同位素配对标记实验, 只要SOC和其他一种或是两种碳源的碳同位素比值间有较为明显的差别, 也可以解析多碳源相互作用及其对土壤呼吸的贡献, 这对于研究复杂生态系统中碳通量和净碳平衡具有重要的应用价值(Whitman & Lehmann, 2015). ...
... 基于质量平衡原理解析土壤-植物-微生物连续体系中有机物的二元或三元组成与贡献, 量化生态系统中小分子根系分泌物-植物残体-生物质炭-土壤有机质-微生物活体与残留物等碳源的相对比例, 揭示生态系统碳源流通过程与机理(Phillips & Gregg, 2003; Whitman & Lehmann, 2015; Liang et al., 2017).然而, 生态系统中有机物是多碳源组成的, 质量平衡并不能适应所有环境条件下有机物的源解析.因此, 需要基于同位素示踪基本原理, 构建合理的方程模型与原创性理论, 准确区分与量化复杂生态系统中有机碳的来源与周转通量, 揭示生态系统碳循环与调控机理. ...
Stable-isotope Raman microspectroscopy for the analysis of soil organic matter
1
2018
... 拉曼光谱成像能高分辨率捕捉不同化学成分在样品中的空间分布特征, 具有样本需求量小、无需前处理、非侵入性、无损伤、对样品物理状态要求低等诸多优势, 自开发以来得到广泛应用.13C稳定同位素探针-拉曼显微光谱技术(SIRM)利用重同位素的光谱红移, 可原位连续检测含碳化合物在生态系统和生物体内的合成、分解、运输、分配等过程, 不仅能够在单细胞水平上提供微生物的生理生态特征信息, 还能够检测到自然环境丰度较低但能发挥重要作用的微生物类群, 对深入理解生态系统碳循环过程有重要意义(图1)(Li et al., 2012; Wiesheu et al., 2018).近年发展起来的表面增强拉曼光谱成像技术通过强化拉曼散射信号, 有效提高了拉曼光谱的灵敏性(Zong et al., 2018). ...
Carbon flow from 13C-labeled straw and root residues into the phospholipid fatty acids of a soil microbial community under field conditions
1
2006
... 13C同位素示踪法技术可以很好地研究外源C对SOM周转的影响.例如, 将13C标记的有机物加入土壤进行田间或室内培养实验, 通过测定13CO2可以区分外源有机物和原有SOM来源的碳, 量化外源有机物添加SOM分解的影响(Bernoux et al., 1998; Blagodatskaya & Kuzyakov, 2008).高丰度13C标记的有机物结合13C-PLFA-SIP和13C-DNA-SIP技术, 可以准确示踪微生物在土壤有机碳周转过程中的参与机制(Williams et al., 2006; Apostel et al., 2015; Wang et al., 2016).而13C-微生物残留物对于量化和评估微生物对SOM积累的贡献起到了关键作用(Tian et al., 2014; Zhang et al., 2015; Liang et al., 2017). ...
Effect of nitrogen fertilizer on rice photosynthate allocation and carbon input in paddy soil
1
2019a
... 13CO2同位素标记法, 是指以高丰度13CO2为示踪物, 在植物通过光合作用固定这些示踪物后, 通过在特定的时间内测定植物-土壤系统中各碳库示踪物量的变化, 可定量研究该时间段内13C在植物-土壤系统的转移和分布, 阐明植物某一生长期的碳分配特征, 定量植物碳向土壤的输入以及土壤碳周转, 从而明确植物光合碳对SOM的来源、稳定化过程的影响及其微生物驱动机制(Hanson et al., 2000; Kuzyakov & Cheng, 2001; Kuzyakov, 2006; Yuan et al., 2016; 张蕊等, 2017; Liu et al., 2019b).脉冲标记法是一次或分几次加入13CO2, 标记数小时后在特定时间内测定13C在土壤-植物系统各碳库的分配比例(Kuzyakov & Gavrichkova, 2010)(表1).脉冲标记法只能提供植物生长关键时期光合碳的分配特征, 但是在植物生长期内标记13CO2, 就可以估计植物来源碳输入土壤各组分的量(Lu et al., 2002a; Kimura et al., 2004; Xiao et al., 2019a, 2019b; Zang et al., 2019).连续标记法是指在植物生长某个生长期(或全生长期), 在可控气室等封闭的空间内对植物进行不间断标记, 可以定量全部生长期投入碳的净分配(表1).例如, 在水稻(Oryza satira)拔节期和灌浆期进行13CO2连续标记, 可以定量研究生长过程中水稻光合碳对SOC各组分的贡献(Kuzyakov, 2005; Conrad et al., 2012; Wang et al., 2016), 以及对土壤原有有机质的激发效应(Zhu et al., 2018a). ...
Nitrogen fertilization alters the distribution and fates of photosynthesized carbon in rice-soil systems: a 13C-CO2 pulse labeling study
1
2019b
... 13CO2同位素标记法, 是指以高丰度13CO2为示踪物, 在植物通过光合作用固定这些示踪物后, 通过在特定的时间内测定植物-土壤系统中各碳库示踪物量的变化, 可定量研究该时间段内13C在植物-土壤系统的转移和分布, 阐明植物某一生长期的碳分配特征, 定量植物碳向土壤的输入以及土壤碳周转, 从而明确植物光合碳对SOM的来源、稳定化过程的影响及其微生物驱动机制(Hanson et al., 2000; Kuzyakov & Cheng, 2001; Kuzyakov, 2006; Yuan et al., 2016; 张蕊等, 2017; Liu et al., 2019b).脉冲标记法是一次或分几次加入13CO2, 标记数小时后在特定时间内测定13C在土壤-植物系统各碳库的分配比例(Kuzyakov & Gavrichkova, 2010)(表1).脉冲标记法只能提供植物生长关键时期光合碳的分配特征, 但是在植物生长期内标记13CO2, 就可以估计植物来源碳输入土壤各组分的量(Lu et al., 2002a; Kimura et al., 2004; Xiao et al., 2019a, 2019b; Zang et al., 2019).连续标记法是指在植物生长某个生长期(或全生长期), 在可控气室等封闭的空间内对植物进行不间断标记, 可以定量全部生长期投入碳的净分配(表1).例如, 在水稻(Oryza satira)拔节期和灌浆期进行13CO2连续标记, 可以定量研究生长过程中水稻光合碳对SOC各组分的贡献(Kuzyakov, 2005; Conrad et al., 2012; Wang et al., 2016), 以及对土壤原有有机质的激发效应(Zhu et al., 2018a). ...
稳定同位素分析技术在农田生态系统土壤碳循环中的应用
3
2014
... 碳同位素原子质量的微小差异, 使得含有不同同位素的物质具有不同的理化性质, 导致不同的动力学和热力学效应, 在物理、化学和生物等反应过程中出现同位素分馏现象.碳循环过程中, 同位素交换反应使无机盐(如碳酸盐)富集重同位素(如13C); 而碳的有机循环, 如光合作用的动力分馏效应导致生物成因碳(有机物)中富集轻同位素(如12C), 从而导致陆地生态系统中有机体和大气CO2之间的碳循环过程中, 出现明显的碳同位素分馏现象(Tieszen & Boutton, 1989).植物在光合作用过程中固定CO2途径的差异, 导致不同光合型植物选择吸收13C的比例不同, 从而使C3和C4植株体内的稳定碳同位素比值(δ13C)存在差异, 因此δ13C可以作为表征植物株生理生态特性差异的指标.土壤有机碳(SOC)经过微生物分解矿化为CO2的过程中13C分馏作用较小, 仅为1‰-3‰, 而SOC在厌氧条件下产生甲烷的过程中13C的分馏作用相对较大, 为10‰-70‰ (Conrad et al., 2010; 林光辉, 2013; 袁红朝等, 2014).但SOC主要来源于陆生高等植物, 与植物光合作用固定CO2时的分馏作用相比, SOC分解过程中的分馏作用几乎可以忽略不计, 因此SOC与该土壤上种植植物的δ13C基本一致(Fernandez et al., 2003; Crow et al., 2006; 张林等, 2010; Bai et al., 2012).所以, 通过测定各个碳库中碳同位素比值的变化, 可以量化“大气-植物-土壤”生态系统中碳的来源、分配及其周转规律. ...
... 质谱法、核磁共振法和光谱法是测定稳定碳同位素常用的方法, 质谱法是稳定碳同位素分析中最通用、最精确的方法(Paul et al., 2007; Eiler et al., 2012; 袁红朝等, 2014).质谱法的原理是先将制备的样品转化成含所测元素的纯气体(CO2、H2、N2、SO2等), 经离子源电离, 形成的离子束在电场、磁场的作用下, 导致具有不同质荷比的离子束分开从而检测同位素丰度.普通质谱仪在计算机辅助下直接给出同位素比值, 更先进的仪器如美国热电公司的Thermo Scientific MAT253, 不仅具有更高的灵敏度和更宽的测量范围, 而且可以与同位素数据系统整合, 自动化分析样品的同位素比值.核磁共振法是在磁场恒定条件下, 基于共振时的射频电场频率来测定不同基团上的同位素丰度, 与碳同位素示踪技术相结合可以检测底物(如葡萄糖)在土壤碳循环过程中的化学组分动态(Webster et al., 1997).光谱法是根据组成分子的不同同位素质量数不同从而使红外光谱谱线发生位移的现象来测定碳同位素丰度.近年来, 随着元素生物地球化学循环研究的进一步发展, 以及元素分析仪-同位素质谱(EA-IRMS)和热分析-傅里叶变换红外光谱-气相色谱-质谱(STA-FTIR-GC-MS)等联用技术的兴起, 极大促进了碳同位素分析测试技术的发展(曹亚澄等, 2008; 刘春波等, 2014). ...
... 态系统的功能群多样性以及区分不同功能群植物提供了一条重要途径(Garten Jr & Taylor Jr, 1992; 陈世苹等, 2002).任书杰和于贵瑞(2011)整合了中国区域的数据发现水分条件越好, C3植物叶片δ13C越低, 因此C3植物叶片的δ13C可以作为指示植物的水分利用效率的指标.C3/C4植物交互种植时会改变土壤有机质的δ13C, 因此, 以未交互种植作物土壤为对照, 通过土壤有机碳δ13C的变化可以研究土壤有机碳的周转速度, 以及不同植物来源碳占土壤各碳库和呼吸的比例以及激发效应的方向和强度(Kuzyakov, 2010; Werth & Kuzyakov, 2010; Blagodatskaya et al., 2011a), 也可以通过土壤碳自然丰度δ13C的变化来研究土壤碳库中活性组分碳(微生物生物量碳(MBC))的周转速率和稳定性(Blagodatskaya et al., 2011b).研究结果发现, SOC和MBC的周转时间分别为16.8 a和29-30 d, 且随种植时间增加, 周转时间延长.新碳和老碳对土壤各碳库和呼吸的贡献不同, MBC中20%的碳来源于老碳(来自C3作物), CO2气体中60%的碳来源于老碳(来自C3作物), 由于土壤微生物偏好矿化老碳而同化新碳, SOC中新碳(来自C4作物)的贡献将逐年增加.此外, 长期种植C3作物(小麦(Triticum aestivum))的土壤在连续耕种C4作物(玉米(zea mays)) 13年后, 土壤有机碳库更新了22%, 且SOC的周转速率在不同粒径团聚体中各不相同, 在>50 µm与<2 µm粒径的团聚体中包含更多新碳, 而黏粒中有机碳的更新速度最慢(Balesdent & Mariotti, 1996; 袁红朝等, 2014).13C自然丰度法为明确土壤新老碳库对SOC、MBC和CO2排放的贡献, 解析陆地生态系统δ13C的变化及其在全球碳平衡中的应用提供了重要依据(Cheng, 2009).随着测试精度的提高, 采用碳同位素配对标记实验, 只要SOC和其他一种或是两种碳源的碳同位素比值间有较为明显的差别, 也可以解析多碳源相互作用及其对土壤呼吸的贡献, 这对于研究复杂生态系统中碳通量和净碳平衡具有重要的应用价值(Whitman & Lehmann, 2015). ...
稳定同位素分析技术在农田生态系统土壤碳循环中的应用
3
2014
... 碳同位素原子质量的微小差异, 使得含有不同同位素的物质具有不同的理化性质, 导致不同的动力学和热力学效应, 在物理、化学和生物等反应过程中出现同位素分馏现象.碳循环过程中, 同位素交换反应使无机盐(如碳酸盐)富集重同位素(如13C); 而碳的有机循环, 如光合作用的动力分馏效应导致生物成因碳(有机物)中富集轻同位素(如12C), 从而导致陆地生态系统中有机体和大气CO2之间的碳循环过程中, 出现明显的碳同位素分馏现象(Tieszen & Boutton, 1989).植物在光合作用过程中固定CO2途径的差异, 导致不同光合型植物选择吸收13C的比例不同, 从而使C3和C4植株体内的稳定碳同位素比值(δ13C)存在差异, 因此δ13C可以作为表征植物株生理生态特性差异的指标.土壤有机碳(SOC)经过微生物分解矿化为CO2的过程中13C分馏作用较小, 仅为1‰-3‰, 而SOC在厌氧条件下产生甲烷的过程中13C的分馏作用相对较大, 为10‰-70‰ (Conrad et al., 2010; 林光辉, 2013; 袁红朝等, 2014).但SOC主要来源于陆生高等植物, 与植物光合作用固定CO2时的分馏作用相比, SOC分解过程中的分馏作用几乎可以忽略不计, 因此SOC与该土壤上种植植物的δ13C基本一致(Fernandez et al., 2003; Crow et al., 2006; 张林等, 2010; Bai et al., 2012).所以, 通过测定各个碳库中碳同位素比值的变化, 可以量化“大气-植物-土壤”生态系统中碳的来源、分配及其周转规律. ...
... 质谱法、核磁共振法和光谱法是测定稳定碳同位素常用的方法, 质谱法是稳定碳同位素分析中最通用、最精确的方法(Paul et al., 2007; Eiler et al., 2012; 袁红朝等, 2014).质谱法的原理是先将制备的样品转化成含所测元素的纯气体(CO2、H2、N2、SO2等), 经离子源电离, 形成的离子束在电场、磁场的作用下, 导致具有不同质荷比的离子束分开从而检测同位素丰度.普通质谱仪在计算机辅助下直接给出同位素比值, 更先进的仪器如美国热电公司的Thermo Scientific MAT253, 不仅具有更高的灵敏度和更宽的测量范围, 而且可以与同位素数据系统整合, 自动化分析样品的同位素比值.核磁共振法是在磁场恒定条件下, 基于共振时的射频电场频率来测定不同基团上的同位素丰度, 与碳同位素示踪技术相结合可以检测底物(如葡萄糖)在土壤碳循环过程中的化学组分动态(Webster et al., 1997).光谱法是根据组成分子的不同同位素质量数不同从而使红外光谱谱线发生位移的现象来测定碳同位素丰度.近年来, 随着元素生物地球化学循环研究的进一步发展, 以及元素分析仪-同位素质谱(EA-IRMS)和热分析-傅里叶变换红外光谱-气相色谱-质谱(STA-FTIR-GC-MS)等联用技术的兴起, 极大促进了碳同位素分析测试技术的发展(曹亚澄等, 2008; 刘春波等, 2014). ...
... 态系统的功能群多样性以及区分不同功能群植物提供了一条重要途径(Garten Jr & Taylor Jr, 1992; 陈世苹等, 2002).任书杰和于贵瑞(2011)整合了中国区域的数据发现水分条件越好, C3植物叶片δ13C越低, 因此C3植物叶片的δ13C可以作为指示植物的水分利用效率的指标.C3/C4植物交互种植时会改变土壤有机质的δ13C, 因此, 以未交互种植作物土壤为对照, 通过土壤有机碳δ13C的变化可以研究土壤有机碳的周转速度, 以及不同植物来源碳占土壤各碳库和呼吸的比例以及激发效应的方向和强度(Kuzyakov, 2010; Werth & Kuzyakov, 2010; Blagodatskaya et al., 2011a), 也可以通过土壤碳自然丰度δ13C的变化来研究土壤碳库中活性组分碳(微生物生物量碳(MBC))的周转速率和稳定性(Blagodatskaya et al., 2011b).研究结果发现, SOC和MBC的周转时间分别为16.8 a和29-30 d, 且随种植时间增加, 周转时间延长.新碳和老碳对土壤各碳库和呼吸的贡献不同, MBC中20%的碳来源于老碳(来自C3作物), CO2气体中60%的碳来源于老碳(来自C3作物), 由于土壤微生物偏好矿化老碳而同化新碳, SOC中新碳(来自C4作物)的贡献将逐年增加.此外, 长期种植C3作物(小麦(Triticum aestivum))的土壤在连续耕种C4作物(玉米(zea mays)) 13年后, 土壤有机碳库更新了22%, 且SOC的周转速率在不同粒径团聚体中各不相同, 在>50 µm与<2 µm粒径的团聚体中包含更多新碳, 而黏粒中有机碳的更新速度最慢(Balesdent & Mariotti, 1996; 袁红朝等, 2014).13C自然丰度法为明确土壤新老碳库对SOC、MBC和CO2排放的贡献, 解析陆地生态系统δ13C的变化及其在全球碳平衡中的应用提供了重要依据(Cheng, 2009).随着测试精度的提高, 采用碳同位素配对标记实验, 只要SOC和其他一种或是两种碳源的碳同位素比值间有较为明显的差别, 也可以解析多碳源相互作用及其对土壤呼吸的贡献, 这对于研究复杂生态系统中碳通量和净碳平衡具有重要的应用价值(Whitman & Lehmann, 2015). ...
Microbial utilizetion of rice root exudates: 13C labeling and PLFA composition
3
2016
... 13CO2同位素标记法, 是指以高丰度13CO2为示踪物, 在植物通过光合作用固定这些示踪物后, 通过在特定的时间内测定植物-土壤系统中各碳库示踪物量的变化, 可定量研究该时间段内13C在植物-土壤系统的转移和分布, 阐明植物某一生长期的碳分配特征, 定量植物碳向土壤的输入以及土壤碳周转, 从而明确植物光合碳对SOM的来源、稳定化过程的影响及其微生物驱动机制(Hanson et al., 2000; Kuzyakov & Cheng, 2001; Kuzyakov, 2006; Yuan et al., 2016; 张蕊等, 2017; Liu et al., 2019b).脉冲标记法是一次或分几次加入13CO2, 标记数小时后在特定时间内测定13C在土壤-植物系统各碳库的分配比例(Kuzyakov & Gavrichkova, 2010)(表1).脉冲标记法只能提供植物生长关键时期光合碳的分配特征, 但是在植物生长期内标记13CO2, 就可以估计植物来源碳输入土壤各组分的量(Lu et al., 2002a; Kimura et al., 2004; Xiao et al., 2019a, 2019b; Zang et al., 2019).连续标记法是指在植物生长某个生长期(或全生长期), 在可控气室等封闭的空间内对植物进行不间断标记, 可以定量全部生长期投入碳的净分配(表1).例如, 在水稻(Oryza satira)拔节期和灌浆期进行13CO2连续标记, 可以定量研究生长过程中水稻光合碳对SOC各组分的贡献(Kuzyakov, 2005; Conrad et al., 2012; Wang et al., 2016), 以及对土壤原有有机质的激发效应(Zhu et al., 2018a). ...
... 采用13CO2脉冲标记技术, 对生长在不同SOM含量水稻土上的水稻进行标记, 发现高、低SOM水稻土壤对水稻光合碳的转化和周转具有显著差异, 揭示了水稻光合碳向地下传输过程的“马太效应”(Zhu et al., 2017b).在水稻拔节期和灌浆期内, 通过水稻的根际沉积作用有4%-6%的光合碳进入土壤有机碳库, 土壤可溶性有机碳的2%-4%和土壤微生物生物量碳的9%-18%来源于这部分新碳(Lu et al., 2002a, 2002b; 祝贞科等, 2016).此外, 利用13CO2脉冲标记结合生物标志物(磷脂脂肪酸)的稳定碳同位素探针技术(13C-PLFA-SIP), 发现了碳从水稻向土壤微生物快速转移的现象, 且随着水稻生长期的变化根际碳的微生物同化量显著增加, 真菌及革兰氏阴性菌是主要的同化微生物(Yuan et al., 2016; Liu et al., 2019a).采用13CO2连续标记技术研究了水稻光合碳的传输和转化数量特征及其对氮肥施用的响应, 发现了高氮肥施用条件下水稻光合碳在土壤中的传输和积累比率非线性骤升的规律(Bradford et al., 2008; Ge et al., 2017).结合13C-PLFA-SIP技术发现, 外源C (羧甲基纤维素)和氮肥的联合施用降低了革兰氏阳性菌(G+)和革兰氏阴性菌(G-)的比值, 降低了G+和真菌的丰度, 单施氮肥则刺激了G-和放线菌的增长, 说明可利用碳氮的不同施用策略可显著影响土壤微生物的群落结构(Zhao et al., 2019).Atere等(2017)利用13CO2连续标记技术和土壤有机质的粒径组分分离技术联用, 发现水稻光合碳的固定效率在氮肥施用结合干湿交替条件下有显著提高.连续标记法有助于定量全部生长期或某一生长期全部投入C的净分配, 但是由于成本高以及技术困难, 运用受到限制. ...
... 13C富集底物标记技术一般用于室内培养实验, 13C富集底物可分为简单的小分子有机物(如葡萄糖、丙氨酸等)和复杂的大分子有机物(如纤维素、木质素、植物秸秆等).在实验土壤中加入13C标记的底物, 以未标记的底物为对照, 结合PLFA、氨基糖等生物标志物技术, 测定13C在土壤各组分有机碳的量及其来源, 有助于阐明外源碳对土壤碳储量的贡献(Yuan et al., 2016; Liang et al., 2017). ...
Allocation of assimilated carbon in paddies depending on rice age, chase period and N fertilization: experiment with 13CO2 labelling and literature synthesis
1
2019
... 13CO2同位素标记法, 是指以高丰度13CO2为示踪物, 在植物通过光合作用固定这些示踪物后, 通过在特定的时间内测定植物-土壤系统中各碳库示踪物量的变化, 可定量研究该时间段内13C在植物-土壤系统的转移和分布, 阐明植物某一生长期的碳分配特征, 定量植物碳向土壤的输入以及土壤碳周转, 从而明确植物光合碳对SOM的来源、稳定化过程的影响及其微生物驱动机制(Hanson et al., 2000; Kuzyakov & Cheng, 2001; Kuzyakov, 2006; Yuan et al., 2016; 张蕊等, 2017; Liu et al., 2019b).脉冲标记法是一次或分几次加入13CO2, 标记数小时后在特定时间内测定13C在土壤-植物系统各碳库的分配比例(Kuzyakov & Gavrichkova, 2010)(表1).脉冲标记法只能提供植物生长关键时期光合碳的分配特征, 但是在植物生长期内标记13CO2, 就可以估计植物来源碳输入土壤各组分的量(Lu et al., 2002a; Kimura et al., 2004; Xiao et al., 2019a, 2019b; Zang et al., 2019).连续标记法是指在植物生长某个生长期(或全生长期), 在可控气室等封闭的空间内对植物进行不间断标记, 可以定量全部生长期投入碳的净分配(表1).例如, 在水稻(Oryza satira)拔节期和灌浆期进行13CO2连续标记, 可以定量研究生长过程中水稻光合碳对SOC各组分的贡献(Kuzyakov, 2005; Conrad et al., 2012; Wang et al., 2016), 以及对土壤原有有机质的激发效应(Zhu et al., 2018a). ...
稳定同位素比例质谱仪在土壤碳循环研究中的应用
1
2010
... 碳同位素原子质量的微小差异, 使得含有不同同位素的物质具有不同的理化性质, 导致不同的动力学和热力学效应, 在物理、化学和生物等反应过程中出现同位素分馏现象.碳循环过程中, 同位素交换反应使无机盐(如碳酸盐)富集重同位素(如13C); 而碳的有机循环, 如光合作用的动力分馏效应导致生物成因碳(有机物)中富集轻同位素(如12C), 从而导致陆地生态系统中有机体和大气CO2之间的碳循环过程中, 出现明显的碳同位素分馏现象(Tieszen & Boutton, 1989).植物在光合作用过程中固定CO2途径的差异, 导致不同光合型植物选择吸收13C的比例不同, 从而使C3和C4植株体内的稳定碳同位素比值(δ13C)存在差异, 因此δ13C可以作为表征植物株生理生态特性差异的指标.土壤有机碳(SOC)经过微生物分解矿化为CO2的过程中13C分馏作用较小, 仅为1‰-3‰, 而SOC在厌氧条件下产生甲烷的过程中13C的分馏作用相对较大, 为10‰-70‰ (Conrad et al., 2010; 林光辉, 2013; 袁红朝等, 2014).但SOC主要来源于陆生高等植物, 与植物光合作用固定CO2时的分馏作用相比, SOC分解过程中的分馏作用几乎可以忽略不计, 因此SOC与该土壤上种植植物的δ13C基本一致(Fernandez et al., 2003; Crow et al., 2006; 张林等, 2010; Bai et al., 2012).所以, 通过测定各个碳库中碳同位素比值的变化, 可以量化“大气-植物-土壤”生态系统中碳的来源、分配及其周转规律. ...
稳定同位素比例质谱仪在土壤碳循环研究中的应用
1
2010
... 碳同位素原子质量的微小差异, 使得含有不同同位素的物质具有不同的理化性质, 导致不同的动力学和热力学效应, 在物理、化学和生物等反应过程中出现同位素分馏现象.碳循环过程中, 同位素交换反应使无机盐(如碳酸盐)富集重同位素(如13C); 而碳的有机循环, 如光合作用的动力分馏效应导致生物成因碳(有机物)中富集轻同位素(如12C), 从而导致陆地生态系统中有机体和大气CO2之间的碳循环过程中, 出现明显的碳同位素分馏现象(Tieszen & Boutton, 1989).植物在光合作用过程中固定CO2途径的差异, 导致不同光合型植物选择吸收13C的比例不同, 从而使C3和C4植株体内的稳定碳同位素比值(δ13C)存在差异, 因此δ13C可以作为表征植物株生理生态特性差异的指标.土壤有机碳(SOC)经过微生物分解矿化为CO2的过程中13C分馏作用较小, 仅为1‰-3‰, 而SOC在厌氧条件下产生甲烷的过程中13C的分馏作用相对较大, 为10‰-70‰ (Conrad et al., 2010; 林光辉, 2013; 袁红朝等, 2014).但SOC主要来源于陆生高等植物, 与植物光合作用固定CO2时的分馏作用相比, SOC分解过程中的分馏作用几乎可以忽略不计, 因此SOC与该土壤上种植植物的δ13C基本一致(Fernandez et al., 2003; Crow et al., 2006; 张林等, 2010; Bai et al., 2012).所以, 通过测定各个碳库中碳同位素比值的变化, 可以量化“大气-植物-土壤”生态系统中碳的来源、分配及其周转规律. ...
基于稳定碳同位素技术研究大气CO2浓度升高对植物-土壤系统碳循环的影响
3
2017
... 随着碳同位素示踪技术的广泛应用和迅速发展, 多种碳同位素示踪技术及其与其他光谱和微生物探针技术联用, 对于揭示从微观到宏观尺度的碳循环过程起着重要促进作用(图1)(Radajewski et al., 2000; Herrmann et al., 2007).目前自然丰度法、与13C贫化示踪技术结合的自由空气中气体浓度增加(Free-Air Concentration Enrichment, FACE)实验、脉冲与连续标记法以及碳同位素高丰度底物富集标记法是研究陆地生态系统中碳循环过程常用的碳同位素示踪方法(Balesdent & Mariotti, 1996; Bernoux et al., 1998; Conley et al., 2001; Epron et al., 2012; Ge et al., 2017; 张蕊等, 2017).同时, 随着氢、氮(N)、氧等元素同位素示踪技术的发展与应用, 那些单独用碳同位素无法解决的问题在与其他同位素示踪技术、光谱和分子生物学技术的联用下得以解决(陈世苹等, 2002).例如, Kušlienë等(2014)利用13C/15N双标记法, 结合生物标志物分析, 研究了13C/15N标记的不同类型的根系分泌物对土壤碳氮动态的贡献, 这些技术的联用进一步促进了碳同位素示踪技术在生态学研究中的广泛应用.本文重点从稳定碳同位素示踪技术的原理、方法与应用进展等方面综述其在陆地生态系统碳循环过程中的研究进展. ...
... 13C贫化示踪技术结合的FACE实验是研究CO2浓度增加的微域环境中SOC周转的常用方法.大气CO2的δ13C值约为-8‰, 在13C贫化示踪技术结合的FACE实验(13C-FACE)中供给13C贫化的CO2的δ13C值远低于大气, 其范围是-45‰- -18‰ (张蕊等, 2017).FACE系统是一个开放的模拟未来CO2增加的微域环境, 系统内部的气候条件如水分、光照、温度等非常接近自然生态环境.因此, 在这样的系统内进行模拟CO2升高的实验更接近于真实状况(McLeod & Long, 1999).目前全球已建立十余个涉及森林(Hagedorn et al., 2003), 农田(Leavitt et al., 1994)和草地(Six et al., 2001)等多个生态系统和多种植被类型的13C-FACE平台, 阐释了大气CO2浓度升高条件下, 不同生态系统植物生理与光合效率、养分和水分利用、土壤呼吸、土壤碳的输入量及周转率和微生物响应等过程机制(Leavitt et al., 1994; Canadell et al., 1995; Kiem et al., 2000). ...
... 13CO2同位素标记法, 是指以高丰度13CO2为示踪物, 在植物通过光合作用固定这些示踪物后, 通过在特定的时间内测定植物-土壤系统中各碳库示踪物量的变化, 可定量研究该时间段内13C在植物-土壤系统的转移和分布, 阐明植物某一生长期的碳分配特征, 定量植物碳向土壤的输入以及土壤碳周转, 从而明确植物光合碳对SOM的来源、稳定化过程的影响及其微生物驱动机制(Hanson et al., 2000; Kuzyakov & Cheng, 2001; Kuzyakov, 2006; Yuan et al., 2016; 张蕊等, 2017; Liu et al., 2019b).脉冲标记法是一次或分几次加入13CO2, 标记数小时后在特定时间内测定13C在土壤-植物系统各碳库的分配比例(Kuzyakov & Gavrichkova, 2010)(表1).脉冲标记法只能提供植物生长关键时期光合碳的分配特征, 但是在植物生长期内标记13CO2, 就可以估计植物来源碳输入土壤各组分的量(Lu et al., 2002a; Kimura et al., 2004; Xiao et al., 2019a, 2019b; Zang et al., 2019).连续标记法是指在植物生长某个生长期(或全生长期), 在可控气室等封闭的空间内对植物进行不间断标记, 可以定量全部生长期投入碳的净分配(表1).例如, 在水稻(Oryza satira)拔节期和灌浆期进行13CO2连续标记, 可以定量研究生长过程中水稻光合碳对SOC各组分的贡献(Kuzyakov, 2005; Conrad et al., 2012; Wang et al., 2016), 以及对土壤原有有机质的激发效应(Zhu et al., 2018a). ...
基于稳定碳同位素技术研究大气CO2浓度升高对植物-土壤系统碳循环的影响
3
2017
... 随着碳同位素示踪技术的广泛应用和迅速发展, 多种碳同位素示踪技术及其与其他光谱和微生物探针技术联用, 对于揭示从微观到宏观尺度的碳循环过程起着重要促进作用(图1)(Radajewski et al., 2000; Herrmann et al., 2007).目前自然丰度法、与13C贫化示踪技术结合的自由空气中气体浓度增加(Free-Air Concentration Enrichment, FACE)实验、脉冲与连续标记法以及碳同位素高丰度底物富集标记法是研究陆地生态系统中碳循环过程常用的碳同位素示踪方法(Balesdent & Mariotti, 1996; Bernoux et al., 1998; Conley et al., 2001; Epron et al., 2012; Ge et al., 2017; 张蕊等, 2017).同时, 随着氢、氮(N)、氧等元素同位素示踪技术的发展与应用, 那些单独用碳同位素无法解决的问题在与其他同位素示踪技术、光谱和分子生物学技术的联用下得以解决(陈世苹等, 2002).例如, Kušlienë等(2014)利用13C/15N双标记法, 结合生物标志物分析, 研究了13C/15N标记的不同类型的根系分泌物对土壤碳氮动态的贡献, 这些技术的联用进一步促进了碳同位素示踪技术在生态学研究中的广泛应用.本文重点从稳定碳同位素示踪技术的原理、方法与应用进展等方面综述其在陆地生态系统碳循环过程中的研究进展. ...
... 13C贫化示踪技术结合的FACE实验是研究CO2浓度增加的微域环境中SOC周转的常用方法.大气CO2的δ13C值约为-8‰, 在13C贫化示踪技术结合的FACE实验(13C-FACE)中供给13C贫化的CO2的δ13C值远低于大气, 其范围是-45‰- -18‰ (张蕊等, 2017).FACE系统是一个开放的模拟未来CO2增加的微域环境, 系统内部的气候条件如水分、光照、温度等非常接近自然生态环境.因此, 在这样的系统内进行模拟CO2升高的实验更接近于真实状况(McLeod & Long, 1999).目前全球已建立十余个涉及森林(Hagedorn et al., 2003), 农田(Leavitt et al., 1994)和草地(Six et al., 2001)等多个生态系统和多种植被类型的13C-FACE平台, 阐释了大气CO2浓度升高条件下, 不同生态系统植物生理与光合效率、养分和水分利用、土壤呼吸、土壤碳的输入量及周转率和微生物响应等过程机制(Leavitt et al., 1994; Canadell et al., 1995; Kiem et al., 2000). ...
... 13CO2同位素标记法, 是指以高丰度13CO2为示踪物, 在植物通过光合作用固定这些示踪物后, 通过在特定的时间内测定植物-土壤系统中各碳库示踪物量的变化, 可定量研究该时间段内13C在植物-土壤系统的转移和分布, 阐明植物某一生长期的碳分配特征, 定量植物碳向土壤的输入以及土壤碳周转, 从而明确植物光合碳对SOM的来源、稳定化过程的影响及其微生物驱动机制(Hanson et al., 2000; Kuzyakov & Cheng, 2001; Kuzyakov, 2006; Yuan et al., 2016; 张蕊等, 2017; Liu et al., 2019b).脉冲标记法是一次或分几次加入13CO2, 标记数小时后在特定时间内测定13C在土壤-植物系统各碳库的分配比例(Kuzyakov & Gavrichkova, 2010)(表1).脉冲标记法只能提供植物生长关键时期光合碳的分配特征, 但是在植物生长期内标记13CO2, 就可以估计植物来源碳输入土壤各组分的量(Lu et al., 2002a; Kimura et al., 2004; Xiao et al., 2019a, 2019b; Zang et al., 2019).连续标记法是指在植物生长某个生长期(或全生长期), 在可控气室等封闭的空间内对植物进行不间断标记, 可以定量全部生长期投入碳的净分配(表1).例如, 在水稻(Oryza satira)拔节期和灌浆期进行13CO2连续标记, 可以定量研究生长过程中水稻光合碳对SOC各组分的贡献(Kuzyakov, 2005; Conrad et al., 2012; Wang et al., 2016), 以及对土壤原有有机质的激发效应(Zhu et al., 2018a). ...
Differentiating the mineralization dynamics of the originally present and newly synthesized amino acids in soil amended with available carbon and nitrogen substrates
1
2015
... 13C同位素示踪法技术可以很好地研究外源C对SOM周转的影响.例如, 将13C标记的有机物加入土壤进行田间或室内培养实验, 通过测定13CO2可以区分外源有机物和原有SOM来源的碳, 量化外源有机物添加SOM分解的影响(Bernoux et al., 1998; Blagodatskaya & Kuzyakov, 2008).高丰度13C标记的有机物结合13C-PLFA-SIP和13C-DNA-SIP技术, 可以准确示踪微生物在土壤有机碳周转过程中的参与机制(Williams et al., 2006; Apostel et al., 2015; Wang et al., 2016).而13C-微生物残留物对于量化和评估微生物对SOM积累的贡献起到了关键作用(Tian et al., 2014; Zhang et al., 2015; Liang et al., 2017). ...
Carbon and nitrogen availability in paddy soil affects rice photosynthate allocation, microbial community composition, and priming: combining continuous 13C labeling with PLFA analysis
1
2019
... 采用13CO2脉冲标记技术, 对生长在不同SOM含量水稻土上的水稻进行标记, 发现高、低SOM水稻土壤对水稻光合碳的转化和周转具有显著差异, 揭示了水稻光合碳向地下传输过程的“马太效应”(Zhu et al., 2017b).在水稻拔节期和灌浆期内, 通过水稻的根际沉积作用有4%-6%的光合碳进入土壤有机碳库, 土壤可溶性有机碳的2%-4%和土壤微生物生物量碳的9%-18%来源于这部分新碳(Lu et al., 2002a, 2002b; 祝贞科等, 2016).此外, 利用13CO2脉冲标记结合生物标志物(磷脂脂肪酸)的稳定碳同位素探针技术(13C-PLFA-SIP), 发现了碳从水稻向土壤微生物快速转移的现象, 且随着水稻生长期的变化根际碳的微生物同化量显著增加, 真菌及革兰氏阴性菌是主要的同化微生物(Yuan et al., 2016; Liu et al., 2019a).采用13CO2连续标记技术研究了水稻光合碳的传输和转化数量特征及其对氮肥施用的响应, 发现了高氮肥施用条件下水稻光合碳在土壤中的传输和积累比率非线性骤升的规律(Bradford et al., 2008; Ge et al., 2017).结合13C-PLFA-SIP技术发现, 外源C (羧甲基纤维素)和氮肥的联合施用降低了革兰氏阳性菌(G+)和革兰氏阴性菌(G-)的比值, 降低了G+和真菌的丰度, 单施氮肥则刺激了G-和放线菌的增长, 说明可利用碳氮的不同施用策略可显著影响土壤微生物的群落结构(Zhao et al., 2019).Atere等(2017)利用13CO2连续标记技术和土壤有机质的粒径组分分离技术联用, 发现水稻光合碳的固定效率在氮肥施用结合干湿交替条件下有显著提高.连续标记法有助于定量全部生长期或某一生长期全部投入C的净分配, 但是由于成本高以及技术困难, 运用受到限制. ...
Fate of rice shoot and root residues, rhizodeposits, and microbial assimilated carbon in paddy soil-part 2: turnover and microbial utilization
1
2017a
... SOC周转是一个连续动态变化的过程, 利用碳同位素技术能够示踪新碳在土壤中的转化过程和赋存状态, 揭示外源碳在土壤-微生物之间的周转过程和机理(金鑫鑫等, 2017).利用13C富集标记小分子有机物或是植物秸秆, 结合生物标志物技术等, 可以表征不同来源的有机质在土壤中的相对丰度和降解状况, 以及SOC的动态变化和储量的转化和迁移, 也可以定量评价新老SOC对碳储量的相对贡献, 可有效评估陆地生态系统碳平衡过程及其微观过程机制(Kuzyakov & Domanski, 2000; Kemmitt et al., 2008; Ge et al., 2017; Zhu et al., 2017a).外源C输入土壤后可以通过物理和生物化学途径形成新的SOM (Cotrufo et al., 2013, 2015), 从而增加SOC库; 另一方面, 外源C的输入也会促进土壤中原有SOM降解(Guenet et al., 2018; Zhu et al., 2018b; Perveen et al., 2019), 从而减少SOC库.因此, 这两个过程之间的平衡对SOC库变化的方向和程度起到至关重要的作用(Liang et al., 2018). ...
Rice rhizodeposits affect organic matter priming in paddy soil: the role of N fertilization and plant growth for enzyme activities, CO2 and CH4 emissions
1
2018a
... 13CO2同位素标记法, 是指以高丰度13CO2为示踪物, 在植物通过光合作用固定这些示踪物后, 通过在特定的时间内测定植物-土壤系统中各碳库示踪物量的变化, 可定量研究该时间段内13C在植物-土壤系统的转移和分布, 阐明植物某一生长期的碳分配特征, 定量植物碳向土壤的输入以及土壤碳周转, 从而明确植物光合碳对SOM的来源、稳定化过程的影响及其微生物驱动机制(Hanson et al., 2000; Kuzyakov & Cheng, 2001; Kuzyakov, 2006; Yuan et al., 2016; 张蕊等, 2017; Liu et al., 2019b).脉冲标记法是一次或分几次加入13CO2, 标记数小时后在特定时间内测定13C在土壤-植物系统各碳库的分配比例(Kuzyakov & Gavrichkova, 2010)(表1).脉冲标记法只能提供植物生长关键时期光合碳的分配特征, 但是在植物生长期内标记13CO2, 就可以估计植物来源碳输入土壤各组分的量(Lu et al., 2002a; Kimura et al., 2004; Xiao et al., 2019a, 2019b; Zang et al., 2019).连续标记法是指在植物生长某个生长期(或全生长期), 在可控气室等封闭的空间内对植物进行不间断标记, 可以定量全部生长期投入碳的净分配(表1).例如, 在水稻(Oryza satira)拔节期和灌浆期进行13CO2连续标记, 可以定量研究生长过程中水稻光合碳对SOC各组分的贡献(Kuzyakov, 2005; Conrad et al., 2012; Wang et al., 2016), 以及对土壤原有有机质的激发效应(Zhu et al., 2018a). ...
Microbial stoichiometric flexibility regulates rice straw mineralizetion and its priming effect in paddy soil
1
2018b
... SOC周转是一个连续动态变化的过程, 利用碳同位素技术能够示踪新碳在土壤中的转化过程和赋存状态, 揭示外源碳在土壤-微生物之间的周转过程和机理(金鑫鑫等, 2017).利用13C富集标记小分子有机物或是植物秸秆, 结合生物标志物技术等, 可以表征不同来源的有机质在土壤中的相对丰度和降解状况, 以及SOC的动态变化和储量的转化和迁移, 也可以定量评价新老SOC对碳储量的相对贡献, 可有效评估陆地生态系统碳平衡过程及其微观过程机制(Kuzyakov & Domanski, 2000; Kemmitt et al., 2008; Ge et al., 2017; Zhu et al., 2017a).外源C输入土壤后可以通过物理和生物化学途径形成新的SOM (Cotrufo et al., 2013, 2015), 从而增加SOC库; 另一方面, 外源C的输入也会促进土壤中原有SOM降解(Guenet et al., 2018; Zhu et al., 2018b; Perveen et al., 2019), 从而减少SOC库.因此, 这两个过程之间的平衡对SOC库变化的方向和程度起到至关重要的作用(Liang et al., 2018). ...
Belowground carbon allocation and dynamics under rice cultivation depends on soil organic matter content
1
2017b
... 采用13CO2脉冲标记技术, 对生长在不同SOM含量水稻土上的水稻进行标记, 发现高、低SOM水稻土壤对水稻光合碳的转化和周转具有显著差异, 揭示了水稻光合碳向地下传输过程的“马太效应”(Zhu et al., 2017b).在水稻拔节期和灌浆期内, 通过水稻的根际沉积作用有4%-6%的光合碳进入土壤有机碳库, 土壤可溶性有机碳的2%-4%和土壤微生物生物量碳的9%-18%来源于这部分新碳(Lu et al., 2002a, 2002b; 祝贞科等, 2016).此外, 利用13CO2脉冲标记结合生物标志物(磷脂脂肪酸)的稳定碳同位素探针技术(13C-PLFA-SIP), 发现了碳从水稻向土壤微生物快速转移的现象, 且随着水稻生长期的变化根际碳的微生物同化量显著增加, 真菌及革兰氏阴性菌是主要的同化微生物(Yuan et al., 2016; Liu et al., 2019a).采用13CO2连续标记技术研究了水稻光合碳的传输和转化数量特征及其对氮肥施用的响应, 发现了高氮肥施用条件下水稻光合碳在土壤中的传输和积累比率非线性骤升的规律(Bradford et al., 2008; Ge et al., 2017).结合13C-PLFA-SIP技术发现, 外源C (羧甲基纤维素)和氮肥的联合施用降低了革兰氏阳性菌(G+)和革兰氏阴性菌(G-)的比值, 降低了G+和真菌的丰度, 单施氮肥则刺激了G-和放线菌的增长, 说明可利用碳氮的不同施用策略可显著影响土壤微生物的群落结构(Zhao et al., 2019).Atere等(2017)利用13CO2连续标记技术和土壤有机质的粒径组分分离技术联用, 发现水稻光合碳的固定效率在氮肥施用结合干湿交替条件下有显著提高.连续标记法有助于定量全部生长期或某一生长期全部投入C的净分配, 但是由于成本高以及技术困难, 运用受到限制. ...
农田作物同化碳输入与周转的生物地球化学过程
1
2016
... 采用13CO2脉冲标记技术, 对生长在不同SOM含量水稻土上的水稻进行标记, 发现高、低SOM水稻土壤对水稻光合碳的转化和周转具有显著差异, 揭示了水稻光合碳向地下传输过程的“马太效应”(Zhu et al., 2017b).在水稻拔节期和灌浆期内, 通过水稻的根际沉积作用有4%-6%的光合碳进入土壤有机碳库, 土壤可溶性有机碳的2%-4%和土壤微生物生物量碳的9%-18%来源于这部分新碳(Lu et al., 2002a, 2002b; 祝贞科等, 2016).此外, 利用13CO2脉冲标记结合生物标志物(磷脂脂肪酸)的稳定碳同位素探针技术(13C-PLFA-SIP), 发现了碳从水稻向土壤微生物快速转移的现象, 且随着水稻生长期的变化根际碳的微生物同化量显著增加, 真菌及革兰氏阴性菌是主要的同化微生物(Yuan et al., 2016; Liu et al., 2019a).采用13CO2连续标记技术研究了水稻光合碳的传输和转化数量特征及其对氮肥施用的响应, 发现了高氮肥施用条件下水稻光合碳在土壤中的传输和积累比率非线性骤升的规律(Bradford et al., 2008; Ge et al., 2017).结合13C-PLFA-SIP技术发现, 外源C (羧甲基纤维素)和氮肥的联合施用降低了革兰氏阳性菌(G+)和革兰氏阴性菌(G-)的比值, 降低了G+和真菌的丰度, 单施氮肥则刺激了G-和放线菌的增长, 说明可利用碳氮的不同施用策略可显著影响土壤微生物的群落结构(Zhao et al., 2019).Atere等(2017)利用13CO2连续标记技术和土壤有机质的粒径组分分离技术联用, 发现水稻光合碳的固定效率在氮肥施用结合干湿交替条件下有显著提高.连续标记法有助于定量全部生长期或某一生长期全部投入C的净分配, 但是由于成本高以及技术困难, 运用受到限制. ...
农田作物同化碳输入与周转的生物地球化学过程
1
2016
... 采用13CO2脉冲标记技术, 对生长在不同SOM含量水稻土上的水稻进行标记, 发现高、低SOM水稻土壤对水稻光合碳的转化和周转具有显著差异, 揭示了水稻光合碳向地下传输过程的“马太效应”(Zhu et al., 2017b).在水稻拔节期和灌浆期内, 通过水稻的根际沉积作用有4%-6%的光合碳进入土壤有机碳库, 土壤可溶性有机碳的2%-4%和土壤微生物生物量碳的9%-18%来源于这部分新碳(Lu et al., 2002a, 2002b; 祝贞科等, 2016).此外, 利用13CO2脉冲标记结合生物标志物(磷脂脂肪酸)的稳定碳同位素探针技术(13C-PLFA-SIP), 发现了碳从水稻向土壤微生物快速转移的现象, 且随着水稻生长期的变化根际碳的微生物同化量显著增加, 真菌及革兰氏阴性菌是主要的同化微生物(Yuan et al., 2016; Liu et al., 2019a).采用13CO2连续标记技术研究了水稻光合碳的传输和转化数量特征及其对氮肥施用的响应, 发现了高氮肥施用条件下水稻光合碳在土壤中的传输和积累比率非线性骤升的规律(Bradford et al., 2008; Ge et al., 2017).结合13C-PLFA-SIP技术发现, 外源C (羧甲基纤维素)和氮肥的联合施用降低了革兰氏阳性菌(G+)和革兰氏阴性菌(G-)的比值, 降低了G+和真菌的丰度, 单施氮肥则刺激了G-和放线菌的增长, 说明可利用碳氮的不同施用策略可显著影响土壤微生物的群落结构(Zhao et al., 2019).Atere等(2017)利用13CO2连续标记技术和土壤有机质的粒径组分分离技术联用, 发现水稻光合碳的固定效率在氮肥施用结合干湿交替条件下有显著提高.连续标记法有助于定量全部生长期或某一生长期全部投入C的净分配, 但是由于成本高以及技术困难, 运用受到限制. ...
Surface-enhanced raman spectroscopy for bioanalysis: reliability and challenges
1
2018
... 拉曼光谱成像能高分辨率捕捉不同化学成分在样品中的空间分布特征, 具有样本需求量小、无需前处理、非侵入性、无损伤、对样品物理状态要求低等诸多优势, 自开发以来得到广泛应用.13C稳定同位素探针-拉曼显微光谱技术(SIRM)利用重同位素的光谱红移, 可原位连续检测含碳化合物在生态系统和生物体内的合成、分解、运输、分配等过程, 不仅能够在单细胞水平上提供微生物的生理生态特征信息, 还能够检测到自然环境丰度较低但能发挥重要作用的微生物类群, 对深入理解生态系统碳循环过程有重要意义(图1)(Li et al., 2012; Wiesheu et al., 2018).近年发展起来的表面增强拉曼光谱成像技术通过强化拉曼散射信号, 有效提高了拉曼光谱的灵敏性(Zong et al., 2018). ...



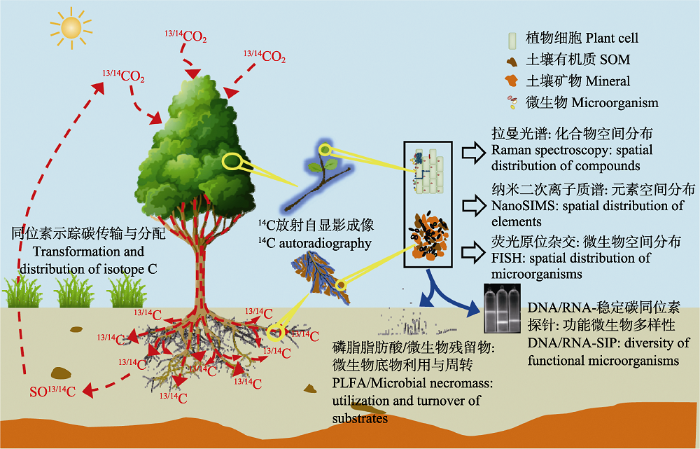



{kind=link}
{kind=link}