



+++110年前, 德国化学家弗里茨·哈伯(Fritz Haber)和他的助手申请了由氮气合成氨的专利(Haber & Le Rossignol, 1909), 后经卡尔·博施(Carl Bosch)的工业化改进得以大规模应用, 他们也因此分别获得了1918和1931年度的诺贝尔化学奖。此后, 人类进入了一个高度依赖哈伯-博施(Haber-Bosch)氮转化技术的世界。一个多世纪以来, 这项技术满足了数十亿人口不断增长的食物需求。据估算, 目前人类身体中接近50%的氮来源于哈伯-博施氮转化工程(Erisman et al., 2008)。然而, 由于人类对氮的不合理利用引发了一连串生态环境问题, 包括水和大气的污染、土壤的酸化和板结、大气中温室气体浓度的升高, 以及生物多样性的丧失。
大气氮沉降主要由人类活动产生的过量活性氮化合物的排放及其大气迁移过程引起, 它可以通过干、湿沉降两种方式将局部氮循环的改变扩展到区域和全球尺度, 从而影响全球生态系统(Holland et al., 2005; Fowler et al., 2013), 目前已成为全球变化的重要议题。活性氮是指地球大气和生物圈层中所有具有生物活性、化学活性和辐射活性的氮化合物, 主要包括无机还原型氮化合物(如NH3和NH4+等)、无机氧化型氮化合物(如NOx、HNO3、N2O、NO3-等)和有机氮化合物(如尿素、有机胺类和蛋白质等)(Galloway et al., 2004)。除N2O之外, 所有排放到大气中的活性氮化合物在经过大气运输之后, 都将以干、湿沉降的方式重新回到地球表面。活性氮的干沉降是指气态的NH3和HNO3以不同形式掺入大气颗粒中, 继而沉降到地面和水体的过程, 包括它们相互反应形成细颗粒的硝酸铵, 或者NH3与硫酸反应生成细颗粒的硫酸铵, 以及HNO3与土壤粉尘或海盐反应生成粗颗粒的硝酸盐等(Lee et al., 2008); 活性氮的湿沉降是指在气象条件允许的情况下, 硝酸、氨、颗粒状硝酸盐和铵盐与空气中的水蒸气结合, 最终以降水的形式携带铵态氮和硝态氮沉降到地面和水体的过程(Zhu et al., 2015)。
大气活性氮的输送范围从数十千米到数千千米, 而随后的氮沉降过程则会把活性氮化合物广泛输入到不同生态系统中。在陆地生态系统中, 过量氮沉降会增加土壤氮的有效性, 导致土壤酸化(Lu et al., 2014; Tian & Niu, 2015), 影响地上植物多样性(Bobbink et al., 1998, 2010; Payne et al., 2017)和净初级生产力(LeBauer & Treseder, 2008), 进而影响到碳、氮、磷的生物地球化学循环(Galloway et al., 2008; Liu & Greaver, 2010; Thomas et al., 2010; Peñuelas et al., 2013)。对于土壤而言, 氮有效性的增加和土壤酸化会降低土壤微生物多样性并改变其群落结构(Demoling et al., 2008; Wang et al., 2018a), 影响土壤微生物的胞外酶活性和土壤呼吸作用(Janssens et al., 2010; Zhou et al., 2014; Jian et al., 2016), 并最终影响到土壤微生物的生态功能(Zhang et al., 2018)(图1)。大气氮沉降对陆地生态系统的深刻影响引发了研究者的广泛研究兴趣, 已成为全球变化生态学的研究热点。
图1
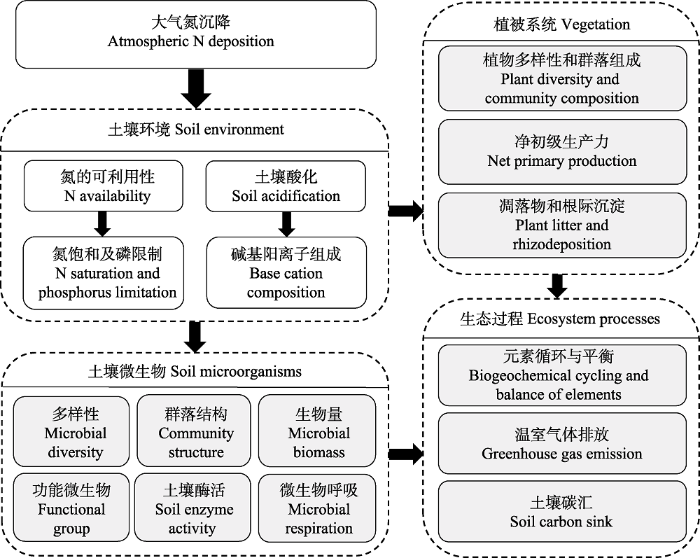
图1
陆地生态系统氮沉降生态效应示意图。
Fig. 1
Schematic diagram of the ecological impacts of nitrogen (N) deposition on terrestrial ecosystems.
据测算, 在不考虑人为活动影响的情况下, 自然氮沉降速率一般在0.5 kg·hm-2·a-1左右(Erisman et al., 2008), 而当前全球很多地区的平均氮沉降速率已超过10 kg·hm-2·a-1 (Dentener et al., 2006)。如果以目前的态势发展下去, 到2050年全球氮沉降速率将会翻倍, 有些地区可能会高达50 kg·hm-2·a-1 (Galloway et al., 2004)。欧洲和北美地区由于较早进行了工业革命, 氮沉降在20世纪80年代达到峰值, 平均为20-50 kg·hm-2·a-1, 最高达到115 kg·hm-2·a-1 (如欧洲西北部和中部以及北美东北部地区)(Jefferies & Maron, 1997)。在我国, 伴随着社会经济的快速发展, 氮肥生产和施用量及化石能源需求量的急剧增加, 致使我国成为继西欧和北美之后的第三个全球氮沉降热点区域(Dentener et al., 2006)。从1980年到2000年的20年间, 我国的平均氮沉降速率从13.2 kg·hm-2·a-1快速增长至21.1 kg·hm-2·a-1, 而部分热点地区的氮沉降速率更是达到甚至超过了欧洲20世纪80年代氮沉降速率的峰值, 导致自然和人工生态系统中植物叶片氮浓度显著上升(Liu et al., 2011, 2013)。近年来, 随着经济结构的调整和环境保护措施的实施, 我国的氮沉降总量及沉降模式也表现出了新的变化趋势, Yu等(2019)通过对我国氮沉降动态变化和空间格局的研究, 发现目前我国大气氮沉降已经停止增加, 并表现出三个重要特征: (1)全国氮沉降量快速增长的趋势得到遏制, 氮沉降量逐渐趋于稳定, NH4+湿沉降量有所降低; (2)氮沉降模式发生转变, 由以往的以湿沉降为主逐步转型为干、湿沉降并重的模式; (3)氮沉降形态发生转变, 沉降物中NH4+/NO3-比值逐渐降低, NO3-比重持续升高。总体上, 当前我国氮沉降量依然处于较高水平, 形势不容乐观, 因此仍然需要高度关注。
1 陆地生态系统大气氮沉降生态效应的研究历史
大气氮沉降研究最早始于英国, 洛桑实验站自1853年开始测定雨水中氮的含量, 发现氮的湿沉降量从1855年的4 kg·hm-2·a-1开始缓慢升高, 到1980年达到最高峰18 kg·hm-2·a-1, 之后开始下降(Goulding et al., 1998)。世界范围内对大气氮沉降的系统研究始于20世纪80年代, 相关研究论文的数量在20世纪90年代初开始快速增长(图2)。我国的氮沉降研究虽然起步较晚, 但发展速度较快, 模拟氮沉降研究基本上从2000年开始起步, 如2002年中国科学院华南植物园建立了包括季风常绿阔叶林、针叶阔叶混交林和马尾松(Pinus massoniana)针叶林在内的鼎湖山森林生态系统长期氮沉降试验研究样地(李德军等, 2005; 鲁显楷等, 2019), 为国内森林生态系统相关研究做出了开创性的贡献。经过十几年的努力, 我国现已建成包括森林(Mo et al., 2008; Zhao et al., 2018; 鲁显楷等, 2019), 草地(Bai et al., 2010; Zhang et al., 2014)和农田(Liu et al., 2013)在内的多个长期定位实验站点, 相关研究论文的数量在2005年之后开始快速增长(图2), 并有不少研究成果发表在Nature、Ecology Letters、Nature Geoscience 和Global Change Biology等国际知名学术期刊上。不过, 相较于欧美国家, 现阶段我国氮沉降生态效应的研究仍较为离散, 研究平台建设还相对薄弱。例如, 我们还缺乏长期的、覆盖不同生态系统类型的大气氮沉降联网实验平台, 还不能对区域尺度的氮沉降生态效应进行整体预测和评估。因此, 在这一领域我们还需要开展大量基础性的研究工作, 并在推进研究的深度和广度的同时, 将研究成果推广应用到生态环境保护中去。
图2
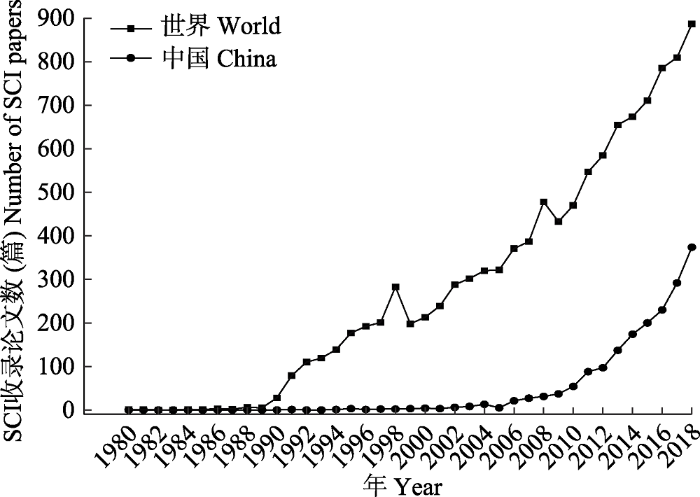
图2
1980-2018年中国与全世界氮沉降相关研究论文数量的变化情况。基于Web of Science数据资源进行统计, 检索式详见附录Ⅰ。
Fig. 2
Increase in the number of SCI papers related to nitrogen deposition from China and world from 1980 to 2018. The statistics are based on Web of Science data resources. Check Supplementary I for details.
自20世纪90年代, 各国科学家就开始研究氮沉降对植物生态生理的影响(Morecroft et al., 1994; Goulding et al., 1998)。此后, 植物多样性和初级生产力响应逐渐成为本领域的研究重点, 大量研究工作围绕森林和草地生态系统展开(Hurd et al., 1998; Stevens et al., 2004; Clark & Tilman, 2008; Lu et al., 2010), 同时也涉及苔原和灌丛等多种陆地生态系统类型(Gordon et al., 2001), 研究范围也从局地尺度向区域和全球尺度拓展(Phoenix et al., 2006)。2000年前后, 由于发现了大气氮沉降对生态系统碳循环的重要影响(Mack et al., 2004; Bragazza et al., 2006; Magnani et al., 2007), 研究人员开始关注氮沉降背景下的元素生物地球化学循环, 包括氮沉降对土壤呼吸和生态系统磷循环的影响(Janssens et al., 2010; Peñuelas et al., 2013), 近年来则开始更多关注不同元素循环之间的耦联与平衡关系(Fleischer et al., 2019; Peng et al., 2019)。
与地上植被系统相比, 对土壤系统的研究起步相对较晚。这方面的研究始于对土壤酶活性、微生物生物量和土壤呼吸的观测(Carreiro et al., 2000; Waldrop & Zak, 2006; Mo et al., 2008; Treseder, 2008), 后来发展到对土壤微生物多样性和不同功能类群的研究(Campbell et al., 2010; Morrison et al., 2016)。随着各种组学技术的广泛应用, 土壤微生物组和生态功能响应的研究正在成为这一研究领域的前沿热点(Fierer et al., 2012; Wang et al., 2017; Kuypers et al., 2018)。
纵观近40年来陆地生态系统氮沉降生态效应的研究历史, 相关研究工作基本围绕着一条主线展开, 即氮沉降对生态系统的结构、功能和过程的影响, 主要研究方向可以归纳为: (1)氮沉降对地上植被系统结构和功能的影响; (2)氮沉降对土壤系统结构和功能的影响; (3)氮沉降对生源要素碳、氮、磷循环的影响。尽管近年来已有不少有关氮沉降生态效应的综述, 但这些综述各有侧重, 或关注特定生态系统(Humbert et al., 2016; 鲁显楷等, 2019), 针对某一特定地区(Liu et al., 2011; Tian et al., 2018), 或关注特定生物类群(Lilleskov et al., 2019), 或某一方面的生态效应(Cheng et al., 2019; 冯婵莹等, 2019), 尚缺乏对这一研究领域发展脉络和前沿进展的全面回顾和总结。基于此, 本文对国内外氮沉降生态效应相关研究进行系统梳理, 考虑不同陆地生态系统类型, 从生态系统结构、功能和过程响应三个方面归纳总结氮沉降生态效应的研究进展, 希望有助于相关研究者从总体上了解和把握本研究领域的重要进展、存在的问题及发展趋势, 从而全面提升我国氮沉降基础研究和生态风险管理水平。
2 氮沉降对地上植被系统的影响
大气氮沉降引起的活性氮在生态系统中的累积被认为是全球生物多样性的重要威胁之一(Simkin et al., 2016; Payne et al., 2017)。大量的氮添加实验和生态调查研究均显示, 氮沉降往往导致植物多样性的丧失(Clark & Tilman, 2008; Zhang et al., 2014; Midolo et al., 2018)。例如, 在经历过去半个多世纪大气氮沉降的持续作用之后, 欧洲和北美广大地区的植物群落组成已发生显著改变(Bobbink et al., 1998; Duprè et al., 2010)。究其原因, 一方面是因为活性氮化合物在生态系统中的累积改变了物种间的相互作用关系, 例如环境中氮的累积会引起喜氮物种在群落中的快速生长, 获得相对竞争优势(Bobbink et al., 1998; Gilliam, 2006); 另一方面氮沉降可以引起土壤酸化, 改变土壤碱基阳离子的组成, 进而影响地上植物群落组成(Lu et al., 2010; Tian & Niu, 2015; Tian et al., 2016)。总体上, 地上植被对大气氮沉降的响应主要由活性氮的积累驱动(de Schrijver et al., 2011), 其响应程度主要取决于三个方面: (1)氮沉降速率、持续时间和氮的输入形式(Humbert et al., 2016); (2)不同植物物种对氮的内在敏感性(Nordin et al., 2006; Britton & Fisher, 2010); (3)非生物环境条件(如气候条件和土壤肥力特征等)(Grman et al., 2010)。
2.1 氮沉降对森林生态系统植物多样性的影响
北方针叶林、温带森林和热带雨林是主要的森林生态系统类型。北方针叶林所在气候带温度低, 土壤养分供应相对不足, 植被生长季短, 理论上对大气氮沉降响应较为敏感; 温带森林因为靠近人类的活动区, 大气氮沉降量相对较高, 因此受影响的程度也较高; 热带雨林有着最高的植物多样性, 但其多为磷限制型生态系统, 理论上对氮沉降响应不敏感。与林下植被相比, 乔木对氮沉降的响应较慢, 因此目前森林生态系统响应大气氮沉降的研究多集中于林下植被, 但研究的结果却不尽相同。例如, 有研究表明长期施氮可导致温带森林和热带森林中林下植被的物种丢失(Lu et al., 2010; Walter et al., 2017), 但在其他大多数的研究中, 氮添加对林下植被的物种丰富度并没有显著影响, 只是导致了植物群落结构的变化(Hurd et al., 1998; Xing et al., 2019)。林下植被多样性响应的不一致性可能是由于植被类型(Bobbink et al., 2010), 氮沉降背景值(Hedwall et al., 2013)及观测持续时间(Gilliam et al., 2016)不同, 或是氮沉降引起的非生物环境条件的改变(Lu et al., 2010)所致。例如, Lu等(2010)在我国南方富氮热带森林中的研究发现, 植物物种丰富度的降低与土壤酸化及土壤可提取铝和钙浓度的升高有关。
不同植物种类对氮沉降的差异化响应是导致林下植被群落结构改变的主要原因。氮添加可以导致生长速度快的草本植物覆盖度增加, 生长缓慢的矮灌木覆盖度降低(Strengbom & Nordin, 2012; Xing et al., 2019), 而附生植物如苔藓等对大气氮沉降很敏感, 氮添加条件下, 其覆盖面积和生物量都呈现显著下降的趋势(de Schrijver et al., 2011; Du et al., 2014)。不同植物对氮沉降的差异化响应主要与植物的生物学特性(Dickson et al., 2014)和对氮添加的敏感性(Nordin et al., 2006)有关。例如, Nordin等(2006)在瑞典北部森林的氮添加实验发现, 矮灌木如欧洲越桔(Vaccinium myrtillus)的生长对不同氮形态无显著响应, 而草本植物如曲芒发草(Deschampsia flexuosa)的生长主要受硝态氮的影响, 苔藓植物则主要受铵态氮的影响, 且苔藓的相对丰度与体内氮浓度呈负相关关系。
尽管已有不少森林生态系统响应氮沉降的研究报道, 但由于森林生态系统的植被群落结构具有高度复杂性, 不同植物类群对氮沉降的响应差异明显, 且森林生态系统中环境异质性较高, 因此客观评估氮沉降对森林植被的影响可能需要更长的时间和更大的观测尺度。与此同时, 现阶段的研究多为现象描述性研究, 还需进一步加强响应机理的研究。
2.2 氮沉降对草地生态系统植物多样性的影响
全球草地面积约占陆地总面积的30%-40%, 草地土壤养分相对贫乏, 历史氮沉降量比较低, 加上放牧和刈割所造成的营养物质流失, 使得草地生态系统明显受到养分供给限制, 因此即使是较低水平的长期氮沉降, 也可能引起土壤的富营养化和酸化(Aber et al., 1998; Tian & Niu, 2015; Chen et al., 2016b), 对草地生态系统植物多样性带来负面影响。例如, 一项横跨英国草地的研究表明, 长期低剂量的大气氮沉降显著降低了植物物种的丰富度, 且物种丰富度随无机氮沉降速率升高呈线性下降趋势(Stevens et al., 2004)。英国的这一发现与北美高草草原的长期研究结果一致: 经过23年的慢性低剂量氮添加, 实验处理组(10 kg·hm-2·a-1)相比对照组(6 kg·hm-2·a-1)植物物种数量减少了17% (Clark & Tilman, 2008)。值得注意的是, 相较于自然氮沉降的缓慢施肥作用, 氮添加实验往往存在急性刺激的弊端。Zhang等(2014)在我国北方温带草原的研究发现, 植物物种丰富度在低频率、高剂量氮添加时下降速度更快, 说明氮沉降模拟实验中的施氮方式会对实验结果产生较大影响, 因此现有低频、高剂量氮添加模拟实验可能难以反映真实的氮沉降生态效应。
虽然在不同的实验中, 草地植物多样性对氮沉降表现出类似的响应模式, 但其内在的响应机理却不尽相同。大量活性氮的累积可以通过直接毒性、土壤酸化、营养失衡以及改变种间竞争等途径影响植物的多样性(Bobbink et al., 2010; Payne et al., 2017)。例如, Clark和Tilman (2008)将植物多样性对氮沉降的响应归因于氮输入的富营养化效应, 即活性氮的累积改变了植物的种间竞争关系, 在新的环境条件下失去竞争力的物种在群落中逐渐消失。然而, 另外一些研究发现, 土壤酸化作用活化的金属离子(如Al3+、Mn2+、Fe3+等)产生的毒性效应, 可能是植物多样性下降的主要原因(Tian et al., 2016)。
2.3 氮沉降对苔原生态系统植物多样性的影响
苔原生态系统分布于极地和高山生态系统中, 主要由苔藓、地衣、矮灌木和耐寒草本植物组成。苔原生态环境较为恶劣, 气候寒冷, 生长季短, 且因为处在多年冻土带, 植物根系生长受到限制。除了欧洲中南部的阿尔卑斯山地区氮沉降量相对较高(10-30 kg·hm-2·a-1)(Flückiger & Braun, 1998), 极地和高山生态系统因为远离人类活动区, 其氮沉降量相对较低。已有的少量研究表明, 氮添加可以影响苔原植物多样性。例如Gordon等(2001)在北极地区的斯瓦尔巴群岛开展的研究发现, 氮添加量为50 kg·hm-2·a-1时植物物种的丰富度显著降低。在我国, Zong等(2016)在长白山高山苔原生态系统开展了氮沉降模拟实验, 发现草本植物小叶章(Deyeuxia angustifolia)对氮添加非常敏感, 其生态位扩张可以显著影响苔原植被的群落均匀度和物种丰富度。
基于已有研究结果的梳理我们不难发现, 地上植被多样性对大气氮沉降的响应存在较大的不确定性, 与环境条件、植被类型及氮沉降模式等多种因素相关。为了客观评估大气氮沉降的生态风险, 需要充分考虑上述影响因素。离散的个案研究结果往往难以直接比较和整合, 最有效的方案是建立覆盖不同生态系统的大尺度长期氮沉降观测网络, 开展区域尺度联网研究, 在这方面我国与欧洲及北美等发达国家还存在客观差距。
2.4 氮沉降对陆地生态系统生产力的影响
大部分陆地生态系统受到氮可利用性的限制(Vitousek & Howarth, 1991; Vitousek et al., 2010), 使得地上净初级生产力对大气氮沉降表现出普遍的积极响应(LeBauer & Treseder, 2008; Stevens et al., 2015)。一项全球范围内的meta分析研究表明, 除了荒漠生态系统, 温带森林、温带草原、热带森林、热带草原, 以及湿地和苔原生态系统的地上净初级生产力对氮添加均表现出积极响应(LeBauer & Treseder, 2008)。相较于群落总体响应的一致性, 在物种水平植物对大气氮沉降的响应具有显著的物种特异性, 会受到物种属性(Hättenschwiler & Körner, 1997; Shen et al., 2019), 环境因子(如光照、温度和土壤酸度)(Tian & Niu, 2015), 以及菌根共生状况(Thomas et al., 2010)的影响而表现出相异的响应特征。例如, Thomas等(2010)对美国北部森林的研究发现, 大气氮沉降显著促进了24种常见树木中11种树木的生长, 抑制了3种树木的生长。进一步研究发现, 大气氮沉降对所有能够与丛枝菌根真菌共生的树种都表现出生长促进作用(Thomas et al., 2010), 这可能是因为丛枝菌根共生体系可以帮助植物改善磷营养, 从而有助于缓解氮沉降下植物生长的磷限制。
植物根系生长是生态系统净初级生产力的重要组成部分。据估计, 根系生长的贡献约占到全球年净初级生产的30%-60% (Jackson et al., 1997)。植物根系生长对氮添加的响应可能会受施氮量和持续时间(Wang et al., 2019b), 氮化学形态(Baldi et al., 2010), 施氮频率(Wang et al., 2019a), 植物物种(Bai et al., 2015), 以及气候因子(如年平均气温和年降水量)(Peng et al., 2017a)等的影响而表现出不同的响应特征。全球范围内的meta分析研究表明, 氮添加对根系生长和根系周转总体上具有显著的负作用(Peng et al., 2017a)。植物根系生长对氮添加的响应很大程度上受到植物自身的碳分配策略调控(Poorter et al., 2012)。氮沉降解除了植物的氮限制, 植物就会减少向地下部分的碳投资, 而向地上部分投资更多的碳以获取其他的限制性资源(如光照等), 因此根系生长与地上生物量积累往往对氮沉降表现出不同的响应特征。例如, Wang等(2019a)在我国内蒙古草原的模拟研究发现, 在较低的氮添加量下, 地上净初级生产力随着氮添加量的增加而升高, 而地下净初级生产力则随着氮添加量的增加而降低。
3 氮沉降对土壤微生物的影响
土壤微生物在有机质分解、生态系统养分循环, 维持地上植物多样性和生态系统功能方面发挥着重要作用(van der Heijden et al., 1998, 2008; Bardgett & van der Putten, 2014; Li et al., 2019), 其物种多样性、群落结构、胞外酶活性和呼吸作用等对大气氮沉降的响应会对全球碳、氮、磷循环和气候变化产生深刻的影响, 因而土壤微生物也成为氮沉降生态效应研究领域的重点关注对象。
3.1 氮沉降对土壤微生物多样性的影响
氮沉降可以通过改变土壤环境条件(如氮的有效性、土壤酸化、碱基阳离子组成等)直接影响土壤微生物多样性(Tian & Niu, 2015; Zhou et al., 2017; Chen et al., 2019), 也可以通过地上植被的生理和生态响应间接作用于土壤微生物(Högberg et al., 2010)。大量研究表明氮添加对土壤微生物的多样性和群落结构具有显著影响(Allison et al., 2007; Leff et al., 2015), 且影响程度随着氮添加量的升高和持续时间的延长而增大(Fierer et al., 2012; Morrison et al., 2016)。例如, 最近的一项meta分析研究表明, 氮添加显著降低了土壤微生物的α多样性(香农指数和Chao1指数), 且长期氮添加处理(>10年)对土壤微生物的影响程度要显著大于短期处理(Wang et al., 2018a)。
氮沉降在降低土壤微生物α多样性的同时, 也会改变土壤微生物的群落组成和结构。不同的土壤微生物营养需求和生理结构不同, 环境适应性存在显著差异, 对氮沉降的响应也不尽相同, 在群落水平即表现为群落组成和结构的变化。例如, 氮沉降能够促进富营养型微生物如变形菌门和拟杆菌门等的生长, 而降低贫营养型微生物如放线菌门等在群落中的丰度(Fierer et al., 2012); 类似地, 革兰氏阴性菌如疣微菌门和装甲菌门等对土壤酸化非常敏感(Ramirez et al., 2012; Yao et al., 2014), 因此氮沉降引起的土壤酸化会导致革兰氏阴性菌相对丰度的降低, 由此显著提高土壤中革兰氏阳性菌与阴性菌的比例(G+/G-)(Wang et al., 2018b; Zhang et al., 2018)。
真菌与细菌比(F:B)可以表征土壤微生物群落对环境变化的总体响应(Strickland & Rousk, 2010), 而土壤中F:B与土壤C:N存在正相关关系(Waring et al., 2013)。大气氮沉降可以增加土壤中氮的有效性, 降低土壤C:N, 从而降低土壤F:B。例如, Zhou等(2017)通过回归分析和主成分分析方法排除土壤酸化作用, 揭示了F:B降低的主要原因是氮添加提高了土壤中氮的有效性。此外, F:B的降低也可能是由土壤真菌和细菌的差异化响应导致的, 例如Demoling等(2008)研究发现, 虽然施氮导致了森林土壤中细菌和真菌生物量同时降低, 但真菌生物量的降低程度更大, 最终使得土壤F:B显著下降。
3.2 氮沉降对土壤功能微生物类群的影响
大气氮沉降不仅可以在总体上影响土壤微生物的多样性, 还会对微生物功能类群产生影响。相较于微生物总体多样性对氮沉降的一致性响应, 参与氮循环的不同微生物功能类群对氮沉降的响应存在明显差异。例如, 氮沉降引起的NH4+和NO3-等在环境中的积累, 一方面可以缓解以无机氮作为能量来源或电子受体的硝化和反硝化微生物的氮限制, 使得其丰富度增加(Redding et al., 2016); 另一方面因为环境中氮有效性的增加, 降低了植物对固氮微生物的依赖性, 会导致土壤中固氮微生物丰富度的下降(Berthrong et al., 2014)。
菌根真菌是一类重要的植物共生真菌, 可以改善植物矿质营养, 影响地上植物的多样性和群落结构(van der Heijden et al., 1998)。很多研究表明, 氮沉降会降低自然生态系统中共生真菌的生物量。例如, 在北方针叶林中氮沉降可以显著降低植物向外生菌根真菌的碳投入(Högberg et al., 2010), 从而引起菌根真菌的“碳饥饿”, 降低其生物量; 类似地, van Diepen等(2010)在北美阔叶林中开展的研究发现, 土壤中丛枝菌根真菌特征磷脂脂肪酸(PLFA-16:1ω5c)含量在长期氮添加处理下显著下降, 进一步表明氮添加对共生真菌生物量的负作用。与此同时, 长期氮添加还会导致菌根真菌丰度的下降和关键种的丢失(van Diepen et al., 2013; Mohan et al., 2014; Lilleskov et al., 2019), 影响其多样性、群落结构和功能。例如, Chen等(2014, 2017)在温带草地生态系统中开展的研究表明, 氮添加显著改变了植物根系和土壤中丛枝菌根真菌的多样性和群落组成, 并可以诱导群落谱系聚集。大尺度的生态调查研究显示, 氮沉降可以改变陆地植被的菌根类型, 导致植物群落从外生菌根型(Ectomycorrhizal, EcM)向丛枝菌根型(Arbuscular mycorrhizal, AM)转变(Averill et al., 2018; Jo et al., 2019)。植被菌根类型的转变可能与EcM真菌和AM真菌的生理功能差异有关, EcM真菌主要帮助植物吸收氮(Chalot & Brun, 1998), 而AM真菌主要帮助植物吸收磷(Smith & Read, 2008)。在氮沉降背景下EcM真菌帮助植物吸收土壤氮的生态功能被削弱, 而植物生长的磷限制被放大, 使得植物更倾向于与AM真菌共生以缓解自身生长的磷限制。
现阶段我们对微生物多样性及群落结构对氮沉降的响应已有不少认识, 但已有的研究很少关注微生物多样性变化对相关生态过程和功能的影响, 更少考虑不同微生物类群及土壤微生物组, 以及植物和土壤微生物对氮沉降的协同响应。今后可以通过应用新的组学方法、基因芯片和稳定性同位素示踪技术等加强相关研究。
3.3 氮沉降对土壤酶活性的影响
土壤微生物主要通过各种胞外酶直接参与土壤有机质的矿化和元素生物地球化学循环。土壤微生物对有机质的分解通常受到氮有效性和土壤酸化的影响。大气氮沉降一方面可以改变土壤C:N, 缓解微生物氮限制, 另一方面氮沉降引起的土壤酸化又会对微生物产生消极影响(Averill & Waring, 2018)。Carreiro等(2000)通过模拟氮沉降发现, 氮添加虽然提高了枫树(Acer rubrum)和橡树(Quercus rubra)凋落物中纤维素酶(cellulase)的活性, 但却显著降低了控制木质素降解的酚氧化酶(phenol oxidase)的活性, 最终抑制了森林凋落物的分解。在此后的十几年中, 研究人员围绕氮添加下土壤酶活性的响应开展了大量研究, 发现不同类型的土壤酶, 或者同一类型的土壤酶在不同实验条件下对氮添加的响应都存在较大差异(Burns et al., 2013; Jian et al., 2016)。例如, 葡萄糖苷酶(β-1,4-glucosidase)的活性在不同的研究中表现出了上升(Sinsabaugh et al., 2005), 无显著变化(Zeglin et al., 2007)或降低(Ramirez et al., 2012)的趋势; 在森林生态系统中, 乙酰葡糖胺糖苷酶(β-1,4-N-acetyl-glucosaminidase)的活性可以在土壤中显著增加(Saiya-Cork et al., 2002), 也可以显著降低(Billings & Ziegler, 2008); 而酸性磷酸酶(acid phosphatase)的活性却在不同研究中表现出一致的上升趋势(Marklein & Houlton, 2012)。最近的一项meta分析研究表明, 氮添加显著提高了土壤中水解酶的活性, 而降低了土壤氧化酶的活性, 且水解酶的响应比与微生物碳和微生物生物量之间具有显著的正相关关系(Jian et al., 2016)。基于已有的大量研究结果, 我们不难推断土壤酶活性对氮沉降的响应与微生物群落结构、生物量及生理响应等具有非常密切的关系, 但目前我们仍然很难将土壤酶活性的响应与微生物多样性响应相关联, 缺乏对土壤生态系统响应氮沉降的整体认识。
3.4 氮沉降对土壤呼吸的影响
土壤呼吸由微生物的异养呼吸和植物根系的自养呼吸两部分组成。氮沉降引起的土壤微生物和植物根系的生理生态变化, 最终会影响到土壤呼吸作用。Janssens等(2010)对温带森林相关研究进行meta分析发现, 氮添加显著降低了土壤微生物和植物根系的呼吸作用。不过, 氮添加对土壤呼吸的影响往往存在较大的不确定性, 受到生态系统类型、环境条件和实验体系等因素的影响(Moscatelli et al., 2008; Xu & Wan, 2008)。例如Zhou等(2014)通过meta分析考察了生态系统类型如何影响土壤呼吸作用对氮添加的响应, 结果表明氮添加对森林生态系统的土壤呼吸具有消极影响(-1.4%), 但却增强了草地(7.84%)和农田(12.4%)生态系统的土壤呼吸。
氮沉降主要通过改变土壤氮的有效性和土壤酸化作用影响土壤呼吸。在氮限制型生态系统(如我国北方草地生态系统)中, 氮添加往往可以解除植物和微生物的氮限制, 增强土壤呼吸作用(Han et al., 2012); 但对于富氮生态系统(如我国南方热带森林生态系统), 氮添加可能会通过降低植物向地下的碳分配和改变土壤非生物环境, 减少细根生物量和降低微生物活性, 从而抑制土壤呼吸作用(Mo et al., 2008)。土壤呼吸对氮沉降的响应同样受到土壤酸化作用的显著影响, 氮沉降引起的土壤酸化一方面提高了土壤中H+的浓度, 另一方面使得碱基阳离子Ca2+、Mg2+和K+流失, Al3+和Mn2+浓度升高(Tian & Niu, 2015), 增加了植物根系和土壤微生物的环境压力, 进而对土壤呼吸产生抑制作用。值得注意的是, 植物根系与土壤微生物对环境变化的响应往往并不相同, 因而根系呼吸和土壤微生物呼吸对氮添加的响应也会呈现相异的响应模式。例如, Chen等(2016a)在氮限制的内蒙古草地生态系统开展的研究表明, 氮添加显著提高了植物根系的生物量, 增强了植物根系的呼吸作用, 但另一方面却显著降低了土壤微生物的生物量, 并改变了其群落结构, 致使土壤微生物呼吸作用显著降低。
4 氮沉降对生态系统元素(碳、氮、磷)循环的影响
生态系统碳、氮、磷元素的生物地球化学循环紧密关联, 因此大气氮沉降对生态系统元素循环的影响并非孤立过程。大量研究表明活性氮化合物在生态系统中的积累和转化, 可以改变生态系统C:N:P的化学计量平衡, 深刻影响碳、氮、磷的生物地球化学循环过程, 乃至最终反馈调节温室气体(CO2、N2O、CH4)排放和全球变化。
4.1 氮沉降对生态系统碳循环的影响
陆地生态系统是重要的全球碳汇(Pan et al., 2011; Le Quéré et al., 2015), 吸收了人类活动排放的大量CO2。活性氮化合物的累积深刻影响到陆地生态系统地上净初级生产力和土壤有机质的分解过程(Carreiro et al., 2000; Magnani et al., 2007; Ye et al., 2018), 因而也影响到生态系统的碳循环和土壤碳汇。陆地生态系统大部分是氮限制型生态系统(Vitousek & Howarth, 1991), 氮沉降可以促进植物对大气CO2的吸收和同化(Ciais et al., 2008; LeBauer & Treseder, 2008; Thomas et al., 2010, 2013)。例如, Schulte-Uebbing和de Vries (2018)对全球范围森林生态系统的meta分析研究表明, 氮沉降总体上导致森林地上和地下生物量增加约177 (112-243) Tg·a-1, 约占森林生物量碳汇(1 400 Tg·a-1)的12%, 特别是在寒带森林和温带森林中, 平均每增加1 kg氮可分别增加大约14和13 kg的地上植被生物量碳, 因而氮沉降对于缓解大气CO2浓度升高引起的全球气候变化具有一定的积极作用。
除了植物的作用, 微生物活动也是土壤碳库的关键影响因素。氮沉降可以缓解土壤微生物的氮限制, 提高微生物活性和土壤有机质的分解速率, 可能会导致土壤碳的损失。例如Mack等(2004)的研究表明, 长期氮施加降低了北极苔原生态系统的土壤碳储量。与此相反, 大量研究表明氮施加可以显著降低土壤微生物的生物量和代谢活性(Treseder, 2008; Zhang et al., 2018), 减缓土壤呼吸作用(Janssens et al., 2010; Ramirez et al., 2010), 降低土壤酶活性(Jian et al., 2016; Carrara et al., 2018), 最终降低土壤有机质的分解速率(Riggs & Hobbie, 2016; Averill & Waring, 2018), 增加土壤碳汇。事实上, 除了氮的有效性, 其他因素如土壤酸化、水分条件以及土壤矿物组成等因素对土壤碳库响应氮沉降也具有强烈的调控作用。Averill和Waring (2018)提出了碳、酸度和矿物保护假说, 以解释土壤碳库对氮添加的不同响应。
氮沉降对土壤微生物的影响不仅直接影响到土壤呼吸和土壤碳汇, 还可能影响其他温室气体如CH4释放(Liu & Greaver, 2009; Gomez-Casanovas et al., 2016)。一项全球meta分析表明, 氮沉降引起CH4和N2O释放增加, 在很大程度上抵消了其在减少CO2释放方面的积极作用(Liu & Greaver, 2009)。
氮沉降对生态系统碳循环的影响与气候变化紧密关联, 受到了研究人员的重点关注, 但是我们也不能忽视其他气候变化因子, 如全球变暖、降水格局改变和极端气候事件等与氮沉降的交互作用。越来越多的证据表明, 极端干旱或风暴等极端气候事件可能导致区域生态系统碳储量的减少(Reichstein et al., 2013), 因此很有可能抵消氮沉降引起的陆地生态系统碳吸收, 从而影响未来的全球气候。然而, 我们对多因子的交互作用研究甚少。
4.2 氮沉降对生态系统氮循环的影响
氮是地球上最为丰富的元素之一, 是生物体蛋白质和核酸的重要组分。自然生态系统中大部分的氮以惰性氮气的形式存在, 难以被生物体直接同化利用(Gruber & Galloway, 2008; Canfield et al., 2010), 因此陆地生态系统氮循环的关键步骤是微生物将氮气还原为氨, 使其成为生物有效态氮。然而, 由于人类活动导致的活性氮化合物的大量输入, 打破了生态系统原有的氮平衡, 很大程度上改变了生态系统的氮循环(Niu et al., 2016)。氮在土壤中的形态转化, 包括有机氮矿化、硝化作用和反硝化作用等, 主要是由微生物驱动的(Li et al., 2019)。土壤中与氮循环相关的微生物功能基因对氮添加非常敏感(Levy- Booth et al., 2014), 氨单加氧酶基因(amoA)、亚硝酸盐还原酶基因(nirS)、氧化亚氮还原酶基因(nosZ)等的表达量会在氮添加情况下显著升高(Ning et al., 2015)。考虑到相关功能基因在氨氧化和反硝化过程中的关键作用, 可推测即使微小的土壤氮含量变化也可能显著影响微生物参与的土壤氮循环过程。
有机氮矿化受环境条件的影响会对氮添加表现出不同的响应模式(Brenner et al., 2005; Kowaljow & Mazzarino, 2007), 但总体上氮添加会通过降低土壤C:N促进土壤中有机质的矿化, 增加土壤中无机氮的积累(Lu et al., 2011)。一项全球范围内的meta分析研究表明, 氮添加显著提高了土壤中可溶性有机氮含量(21%), 促进了微生物的硝化作用, 使土壤NO3-浓度大幅度升高了429% (Lu et al., 2011)。土壤无机氮和有机氮的积累, 一方面可以缓解土壤微生物的氮限制, 促进土壤生态系统的硝化和反硝化过程, 提高土壤硝态氮浓度; 另一方面土壤中过量的NO3--N和NH4+-N无法快速地被微生物和植物吸收同化, 使得大量的含氮化合物通过淋溶作用从土壤中流失。例如, 在Lu等(2011)的meta分析研究中, 氮添加下无机氮的平均淋溶量增加了461%; Templer等(2012)通过整合大量15N同位素示踪研究, 发现在氮添加下氮在土壤中的滞留时间显著减少, 进一步表明氮沉降加剧了土壤中氮的淋溶。
土壤氮流失的另一个重要途径是经由微生物的硝化和反硝化作用以气态形式流失(Bouwman et al., 2002)。很多研究表明, 氮循环功能微生物对氮添加非常敏感(Levy-Booth et al., 2014), 氮添加往往导致硝化及反硝化作用增强。例如, Shcherbak等(2014)的meta分析研究发现, 大多数农田的N2O排放量随着施氮量的增加呈指数增长。因此, 在全球尺度上大气氮沉降会加速土壤中的氮转化过程, 使得大气N2O浓度持续升高, 有数据表明过去30年全球大气N2O的浓度正以每年(0.73 ± 0.03) μg·L-1·a-1的速度稳定增长(Pachauri et al., 2014)。作为一种温室气体, N2O的温室效应可能会在一定程度上抵消氮沉降在减缓全球气候变化(尤其是CO2浓度升高)方面的积极作用(Zaehle et al., 2011)。
4.3 氮沉降对生态系统磷循环的影响
磷和氮一样是生物生长发育的限制性营养元素, 是核酸的重要组分。磷在生态系统中的循环包括磷的输入、输出及其在生态系统内部的迁移转化。大气氮沉降持续增加的同时, 大气磷沉降并没有相应增加, 据估算全球大气磷沉降只有3-4 Tg·a-1 (Mahowald et al., 2008), 这使得陆地生态系统中氮、磷沉降的摩尔比(44-47)达到陆地植物N:P摩尔比(22- 30)的2倍左右(Peñuelas et al., 2012); 与此同时, 土壤磷主要以磷酸盐矿物和有机磷化合物的形式存在, 使得磷的淋溶输出受限。因此, 在陆地生态系统中氮沉降对磷循环的影响主要源于氮、磷输入的不平衡。
大气氮沉降引起的活性氮化合物的积累在缓解了植物氮限制的同时也增加了植物对磷的需求, 加之磷在生态系统中的循环相对较慢, 使得生态系统磷限制进一步加剧(Elser et al., 2009), 植物体内氮磷比升高(Yuan & Chen, 2015)。Deng等(2017)通过meta分析研究表明, 虽然氮添加下植物生物量及凋落物量提高了34%和15%, 但植物和凋落物中磷的浓度却分别降低了8%和11%。为了应对自身氮磷比的失衡, 维持正常的生理活动, 植物倾向于投资大量的碳和氮以提高土壤中磷酸酶的活性, 加速土壤磷的矿化, 缓解自身的磷限制(Marklein & Houlton, 2012)。然而, 土壤磷的活化对氮沉降的响应并不一定是正向的, 还会受到其他土壤生态过程的影响。氮沉降引起的土壤酸化是主要的影响因素之一, 土壤酸化一方面可以增加铁、铝等金属阳离子的有效性, 促进了可溶性磷酸根离子的络合(Tian & Niu, 2015), 另一方面会降低土壤中解磷菌和菌根真菌等相关功能微生物的活性(冯婵莹等, 2019), 减缓土壤磷活化, 降低土壤磷的有效性。
4.4 氮沉降对植物和土壤碳氮磷生态化学计量特征的影响
生物体在养分供应及周围环境条件发生变化时, 能够通过特定的适应调节机制, 保持自身元素化学计量学稳定, 这种化学计量内稳性是生物体适应环境变化的一种普遍性机制, 也影响着生态系统结构和功能的稳定(Elser et al., 1996; Sterner & Elser, 2002)。生物体C:N:P与许多重要的生态学过程和功能相关, 如凋落物和有机质的分解(Güsewell & Gessner, 2009; Zhu et al., 2018)、植物生长(Fan et al., 2015)、碳氮循环(Buchkowski et al., 2015), 以及生态系统结构稳定性(Yu et al., 2010)等。Yu等(2010)在我国内蒙古草原生态系统的研究发现, 植物物种的化学计量内稳性与物种在群落中的优势度和植物群落稳定性呈正相关关系。不过, 植物的化学计量内稳性的调节作用也是有限的, 氮沉降已被广泛证实可以引起植物体内N:P的升高(Peñuelas et al., 2012; Yuan & Chen, 2015), 使得生态系统从氮限制型向磷限制型转化(Peng et al., 2019), 并影响到生态系统的碳循环。例如, Peng等(2017b)在高山草甸开展的相关研究表明, 氮添加引起的植物N:P的变化对地上净初级生产力、生态系统呼吸及净碳交换响应起到主导作用。生态化学计量学不仅可以影响地上植被系统, 对土壤物质循环的作用也不可忽视。例如, 氮、磷添加引起的环境C:N:P化学计量学特征的改变, 可以影响水稻土中酶活性和微生物的激发效应, 从而影响土壤有机质的分解过程(Zhu et al., 2018)。
生态系统C:N:P化学计量学特征在受到氮沉降影响的同时, 也受到大气CO2浓度升高、干旱和气候变暖等全球变化因子的影响(Sardans et al., 2012, 2017; Yue et al., 2017)。例如, Yue等(2017)研究了氮沉降、增温和CO2浓度升高对C:N:P化学计量比的影响, 发现氮沉降和CO2浓度升高对陆地生态系统C:N:P的影响强于增温的影响, 多因子复合作用通常表现为加和效应。因此, 客观评估氮沉降生态效应还需要考量氮沉降和其他全球变化因子之间的相互作用。此外, 虽然我们对氮沉降背景下地上植被和土壤微生物的化学计量学响应已有一定的认识, 但对生态系统地上部分与地下部分耦联的认知还比较有限, 因此还难以从生态系统总体水平评估氮沉降的生态化学计量学效应。未来的研究需要更多地考虑多因子复合作用对生态系统整体结构和功能稳定性的影响。
5 研究展望
随着我国社会经济的快速发展, 大气氮沉降将会给自然和农业生态系统带来更加严峻的负面影响。妥善应对大气氮沉降问题, 离不开科学技术的支撑, 因此结合观测和实验数据构建氮沉降的剂量-效应模型, 确定不同生态系统类型的氮沉降饱和阈值, 建立科学的氮沉降生态风险评价体系, 成为现阶段的当务之急。纵观陆地生态系统氮沉降生态效应的研究历史和前沿进展, 不难发现活性氮化合物累积导致的物种间相互作用和土壤环境的改变, 是生态系统结构、功能和生态过程响应氮沉降的主要驱动力, 同时生态系统本身的生物和非生物条件, 以及研究的时空尺度决定了响应的性质和程度。考虑到生态系统响应氮沉降的环境和尺度依赖性, 兼顾不同生态类型的多尺度长期综合监测和联网研究势在必行。与此同时, 今后氮沉降生态效应研究应加强生态系统结构、功能和生态过程的协同和整体响应机制研究, 关注氮沉降格局变化和气候变化因子之间的耦合和交互作用, 并通过采用新方法和新技术拓展研究的广度和深度。具体而言, 我们认为今后的研究工作应注重和加强以下几个方面。
5.1 建设氮沉降联网观测研究平台, 构建科学合理的氮沉降生态风险评价体系
很多研究表明, 大气氮沉降生态效应具有高度的环境和尺度依赖性。虽然过去30年间科研工作者围绕氮沉降生态效应开展了大量研究, 但在区域尺度甚至更大尺度上的系统研究仍相当匮乏。开展区域和全球尺度的联网研究将有助于全面揭示氮沉降生态效应的环境和尺度依赖性, 从而更客观地认识和评估我国乃至全球尺度氮沉降的生态效应和生态风险。
进行氮沉降生态风险评价和管理, 急需确定适合我国国情的氮沉降临界负荷和生态风险阈值。氮沉降临界负荷可以量化特定生态系统对大气氮沉降的敏感性(Reynolds et al., 1998; Fenn et al., 2010), 有助于合理保护敏感生态系统和推行氮排放削减措施(Bobbink et al., 2010)。建立一个覆盖不同生态系统类型的全国性大气氮沉降长期观测研究网络, 加强氮沉降剂量-效应关系研究, 可为不同区域和不同生态系统制定科学合理的氮沉降风险阈值, 进而为制定环境管理政策及治理方案提供科学依据。
5.2 加强氮沉降背景下生态系统不同组分及不同生态过程的协同和整体响应研究
现阶段, 我们对大气氮沉降生态效应的研究相对离散, 多数研究仅关注生态系统某些组分(比如植物或土壤微生物)或某些属性, 不能很好地将生态系统结构、功能和过程的响应有效连接起来, 难以形成对生态系统整体响应的认识。例如, 氮沉降引起的碳、氮、磷循环过程的改变必然是植物和微生物群落结构和功能协同响应的结果, 但已有的研究却很少关注氮沉降引起的生态系统不同组分及不同生态过程之间的关联和互馈响应。因此, 建议今后的研究应更加注重从生态系统整体出发, 更多关注植物和土壤微生物协同响应氮沉降的机制, 解析相关生态过程和生态功能变化之间的耦联关系。
生态系统功能和服务对氮沉降的响应是生态系统整体响应的直接体现, 而生态系统多功能性可作为反映这种整体响应的重要指标。在概念上, 生态系统的多功能性是指生态系统可以同时提供和维持多种生态功能和服务的能力(Manning et al., 2018), 可以有效整合生态系统的功能和过程对氮沉降的响应。采用生态系统的多功能性指标来评价生态系统对氮沉降的响应, 有助于我们从整体水平认识氮沉降的生态效应, 提高生态风险评价与管理的合理性。最近的研究表明, 植物和微生物的多样性对维持和驱动生态系统的多功能性具有重要的作用(Maestre et al., 2012; Delgado-Baquerizo et al., 2016; Le Bagousse-Pinguet et al., 2019), 而大气氮沉降对植物和微生物的多样性普遍具有消极影响, 因此也可以间接影响生态系统的多功能性, 但目前从生态系统多功能性角度评估氮沉降生态效应的研究还很少, 需要在今后切实加强。
5.3 考虑全球变化实际情形和演化趋势, 加强不同氮沉降情景以及多因子复合效应研究
在全面推进氮沉降生态效应研究的深度和广度的同时, 还应充分考虑随着社会经济发展和技术进步氮沉降模式和格局的变化。已有研究表明随着工农业的发展和排放削减措施的施行, 大气氮沉降的类型和沉降速率都会随之发生改变(Li et al., 2016; Ackerman et al., 2019; Yu et al., 2019)。例如, 当前我国氮沉降量快速增长的趋势已得到遏制, 氮沉降格局也发生了转变, NH4+/NO3–比值逐渐降低, NO3–比重持续增加(Yu et al., 2019)。针对氮沉降模式的改变, 在加强监测的同时, 更应关注未来氮沉降格局下生态系统会如何响应和反馈, 尤其是温室气体的排放和土壤有机碳的动态变化, 加强模型预测研究。
与此同时, 客观评估氮沉降生态效应还需要考虑氮沉降和其他全球气候变化因子之间的耦合和交互作用, 例如CO2浓度升高、全球变暖、降水格局变化, 以及极端气候之间的交互作用等。尽管目前已有一些包括氮沉降在内的多因子定位实验, 但结合未来全球变化情形开展不同因子的耦合和交互作用研究仍然是非常必要的。
5.4 注重新方法新技术的应用, 深入揭示氮沉降生态效应机制
系统深入地研究生态系统响应氮沉降的机制, 离不开新方法和新技术的应用。各种组学方法和同位素示踪技术的快速发展, 也使我们有机会深入研究生物系统及生态过程对氮沉降的响应。例如, 微生物组学和代谢组学技术的联用可以帮助我们深入探究微生物的多样性、群落结构、功能基因和代谢过程对氮沉降的关联响应。利用新技术新方法建立生态系统不同组分之间的网络互作关系, 有助于我们发现关键环节和调控途径, 可以帮助我们实现从生态系统整体水平认识和调控氮沉降的生态效应。在今后的研究中, 应充分利用各种新方法和新技术, 加强深层次机理研究。
附录I 文献计量分析检索标准
Supplementary I Literature search criteria for bibliometric analysis
附录I 文献计量分析检索标准
Supplementary I
| 时间跨度 Time span | 1980-2018 |
|---|---|
| 数据库 Database | Science Citation Index Expanded |
| 国家/地区 Country/Region | 中国/世界 China/World |
| 研究领域 Research domains | 环境科学, 生态学, 土壤学, 应用微生物技术, 湖沼学, 地学综合, 植物科学, 微生物学, 多学科科学, 生物学, 农学, 环境工程, 林学, 水资源, 生物多样性保护, 遥感, 海洋学, 海洋与淡水生物学, 生物化学分子生物学, 昆虫学, 进化生物学, 农业综合 Environmental Sciences, Ecology, Soil Science, Biotechnology Applied Microbiology, Limnology, Geosciences Multidisciplinary, Plant Sciences, Microbiology, Multidisciplinary Sciences, Biology, Agronomy, Engineering Environmental, Forestry, Water Resources, Biodiversity Conservation, Remote Sensing, Oceanography, Marine Freshwater Biology, Biochemistry Molecular Biology, Entomology, Evolutionary Biology, Agriculture Multidisciplinary |
| 排除领域 Excluded domains | 化学, 物理学, 材料学, 气象学 Chemistry, Physics, Materials, Meteorology |
| 检索式 Search sets | TS = Nitrogen Deposition AND CU = China AND WC = (Environmental Sciences OR Ecology OR Soil Science OR Biotechnology Applied Microbiology OR Limnology OR Geosciences Multidisciplinary OR Plant Sciences OR Microbiology OR Multidisciplinary Sciences OR Biology OR Agronomy OR Engineering Environmental OR Forestry OR Water Resources OR Biodiversity Conservation OR Remote Sensing OR Oceanography OR Marine Freshwater Biology OR Biochemistry Molecular Biology OR Entomology OR Evolutionary Biology OR Agriculture Multidisciplinary) NOT WC = (Chemistry OR Physics OR Materials OR Meteorology) |
| 检索时间 Search time | 2019-11-10 |

参考文献
Nitrogen saturation in temperate forest ecosystems
Global estimates of inorganic nitrogen deposition across four decades
Nitrogen fertilization reduces diversity and alters community structure of active fungi in boreal ecosystems
Continental-scale nitrogen pollution is shifting forest mycorrhizal associations and soil carbon stocks
Nitrogen limitation of decomposition and decay: How can it occur?
Differential responses of grasses and forbs led to marked reduction in below-ground productivity in temperate steppe following chronic N deposition
Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: evidence from Inner Mongolia Grasslands
Organic fertilization leads to increased peach root production and lifespan
Belowground biodiversity and ecosystem functioning
Nitrogen fertilization has a stronger effect on soil nitrogen-fixing bacterial communities than elevated atmospheric CO2
DOI:10.1128/AEM.04034-13
URL
PMID:24610855
[本文引用: 1]

Biological nitrogen fixation is the primary supply of N to most ecosystems, yet there is considerable uncertainty about how N-fixing bacteria will respond to global change factors such as increasing atmospheric CO2 and N deposition. Using the nifH gene as a molecular marker, we studied how the community structure of N-fixing soil bacteria from temperate pine, aspen, and sweet gum stands and a brackish tidal marsh responded to multiyear elevated CO2 conditions. We also examined how N availability, specifically, N fertilization, interacted with elevated CO2 to affect these communities in the temperate pine forest. Based on data from Sanger sequencing and quantitative PCR, the soil nifH composition in the three forest systems was dominated by species in the Geobacteraceae and, to a lesser extent, Alphaproteobacteria. The N-fixing-bacterial-community structure was subtly altered after 10 or more years of elevated atmospheric CO2, and the observed shifts differed in each biome. In the pine forest, N fertilization had a stronger effect on nifH community structure than elevated CO2 and suppressed the diversity and abundance of N-fixing bacteria under elevated atmospheric CO2 conditions. These results indicate that N-fixing bacteria have complex, interacting responses that will be important for understanding ecosystem productivity in a changing climate.
Altered patterns of soil carbon substrate usage and heterotrophic respiration in a pine forest with elevated CO2 and N fertilization
Global assessment of nitrogen deposition effects on terrestrial plant diversity: a synthesis
The effects of air-borne nitrogen pollutants on species diversity in natural and semi-natural European vegetation
Emissions of N2O and NO from fertilized fields: summary of available measurement data
Atmospheric nitrogen deposition promotes carbon loss from peat bogs
Nitrogen additions to pristine, high-latitude, forest ecosystems: consequences for soil nitrogen transformations and retention in mid and late succession
Terricolous alpine lichens are sensitive to both load and concentration of applied nitrogen and have potential as bioindicators of nitrogen deposition
Microbial stoichiometry overrides biomass as a regulator of soil carbon and nitrogen cycling
Soil enzymes in a changing environment: current knowledge and future directions
The effect of nutrient deposition on bacterial communities in Arctic tundra soil
The evolution and future of earth’s nitrogen cycle
Interactions among plants, bacteria, and fungi reduce extracellular enzyme activities under long- term N fertilization
Microbial enzyme shifts explain litter decay responses to simulated nitrogen deposition
Physiology of organic nitrogen acquisition by ectomycorrhizal fungi and ectomycorrhizas
Soil acidification exerts a greater control on soil respiration than soil nitrogen availability in grasslands subjected to long-term nitrogen enrichment
Direct and indirect effects of nitrogen enrichment on soil organisms and carbon and nitrogen mineralization in a semi-arid grassland
Nitrogen saturation in humid tropical forests after 6 years of nitrogen and phosphorus addition: hypothesis testing
DOI:10.1111/fec.2016.30.issue-2 URL [本文引用: 1]
Nitrogen deposition and precipitation induced phylogenetic clustering of arbuscular mycorrhizal fungal communities
Six-year fertilization modifies the biodiversity of arbuscular mycorrhizal fungi in a temperate steppe in Inner Mongolia
Nitrogen deposition affects both net and gross soil nitrogen transformations in forest ecosystems: a review
Carbon accumulation in European forests
Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands
DOI:10.1038/nature06503
URL
PMID:18256670
[本文引用: 4]
Rates of atmospheric deposition of biologically active nitrogen (N) are two to seven times the pre-industrial rates in many developed nations because of combustion of fossil fuels and agricultural fertilization. They are expected to increase similarly over the next 50 years in industrializing nations of Asia and South America. Although the environmental impacts of high rates of nitrogen addition have been well studied, this is not so for the lower, chronic rates that characterize much of the globe. Here we present results of the first multi-decadal experiment to examine the impacts of chronic, experimental nitrogen addition as low as 10 kg N ha(-1) yr(-1) above ambient atmospheric nitrogen deposition (6 kg N ha(-1) yr(-1) at our site). This total input rate is comparable to terrestrial nitrogen deposition in many industrialized nations. We found that this chronic low-level nitrogen addition rate reduced plant species numbers by 17% relative to controls receiving ambient N deposition. Moreover, species numbers were reduced more per unit of added nitrogen at lower addition rates, suggesting that chronic but low-level nitrogen deposition may have a greater impact on diversity than previously thought. A second experiment showed that a decade after cessation of nitrogen addition, relative plant species number, although not species abundances, had recovered, demonstrating that some effects of nitrogen addition are reversible.
Cumulative nitrogen input drives species loss in terrestrial ecosystems
Microbial diversity drives multifunctionality in terrestrial ecosystems
Bacterial and fungal response to nitrogen fertilization in three coniferous forest soils
Responses of terrestrial ecosystem phosphorus cycling to nitrogen addition: a meta-analysis
Nitrogen and sulfur deposition on regional and global scales: a multimodel evaluation
Height and clonality traits determine plant community responses to fertilization
Effects of nitrogen additions on biomass, stoichiometry and nutrient pools of moss Rhytidium rugosum in a boreal forest in Northeast China
Changes in species richness and composition in European acidic grasslands over the past 70 years: the contribution of cumulative atmospheric nitrogen deposition
Shifts in lake N:P stoichiometry and nutrient limitation driven by atmospheric nitrogen deposition
Organism size, life history, and N:P stoichiometry: toward a unified view of cellular and ecosystem processes
How a century of ammonia synthesis changed the world
Linkages of plant and soil C:N:P stoichiometry and their relationships to forest growth in subtropical plantations
Impacts of nitrogen addition on plant phosphorus content in forest ecosystems and the underlying mechanisms
氮添加对森林植物磷含量的影响及其机制
Nitrogen critical loads and management alternatives for N-impacted ecosystems in California
Comparative metagenomic, phylogenetic and physiological analyses of soil microbial communities across nitrogen gradients
Nitrogen deposition maintains a positive effect on terrestrial carbon sequestration in the 21st century despite growing phosphorus limitation at regional scales
Nitrogen deposition in Swiss forests and its possible relevance for leaf nutrient status, parasite attacks and soil acidification
The global nitrogen cycle in the twenty-first century
Nitrogen cycles: past, present, and future
Transformation of the nitrogen cycle: recent trends, questions, and potential solutions
DOI:10.1126/science.1136674
URL
PMID:18487183
[本文引用: 1]
Humans continue to transform the global nitrogen cycle at a record pace, reflecting an increased combustion of fossil fuels, growing demand for nitrogen in agriculture and industry, and pervasive inefficiencies in its use. Much anthropogenic nitrogen is lost to air, water, and land to cause a cascade of environmental and human health problems. Simultaneously, food production in some parts of the world is nitrogen-deficient, highlighting inequities in the distribution of nitrogen-containing fertilizers. Optimizing the need for a key human resource while minimizing its negative consequences requires an integrated interdisciplinary approach and the development of strategies to decrease nitrogen-containing waste.
Response of the herbaceous layer of forest ecosystems to excess nitrogen deposition
Twenty-five-year response of the herbaceous layer of a temperate hardwood forest to elevated nitrogen deposition
Nitrogen deposition and greenhouse gas emissions from grasslands: uncertainties and future directions
Impacts of increased nitrogen supply on high Arctic heath: the importance of bryophytes and phosphorus availability
Nitrogen deposition and its contribution to nitrogen cycling and associated soil processes
Mechanisms contributing to stability in ecosystem function depend on the environmental context
DOI:10.1111/j.1461-0248.2010.01533.x
URL
PMID:20849440
[本文引用: 1]
Stability in ecosystem function is an important but poorly understood phenomenon. Anthropogenic perturbations alter communities, but how they change stability and the strength of stabilizing mechanisms is not clear. We examined temporal stability (invariability) in aboveground productivity in replicated 18-year time series of experimentally perturbed grassland plant communities. We found that disturbed annual-dominated communities were more stable than undisturbed perennial communities, coincident with increases in the stabilizing effect of mean-variance scaling. We also found that nitrogen-fertilized communities maintained stability despite losses in species richness, probably because of increased compensatory dynamics and increased dominance by particularly stable dominant species. Among our communities, slight variation in diversity was not the strongest mechanism driving differences in stability. Instead, our study suggests that decreases in individual species variabilities and increases in the relative abundance of stable dominant species may help maintain stability in the functioning of ecosystems confronted with eutrophication, disturbance, and other global changes.
An Earth-system perspective of the global nitrogen cycle
N:P ratios influence litter decomposition and colonization by fungi and bacteria in microcosms
Effects of mowing and nitrogen addition on soil respiration in three patches in an old field grassland in Inner Mongolia
Biomass allocation and canopy development in spruce model ecosystems under elevated CO2 and increased N deposition
Does background nitrogen deposition affect the response of boreal vegetation to fertilization?
Quantification of effects of season and nitrogen supply on tree below-ground carbon transfer to ectomycorrhizal fungi and other soil organisms in a boreal pine forest
Nitrogen deposition onto the United States and Western Europe: synthesis of observations and models
Impacts of nitrogen addition on plant biodiversity in mountain grasslands depend on dose, application duration and climate: a systematic review
DOI:10.1111/gcb.12986
URL
PMID:26010833
[本文引用: 2]
Although the influence of nitrogen (N) addition on grassland plant communities has been widely studied, it is still unclear whether observed patterns and underlying mechanisms are constant across biomes. In this systematic review, we use meta-analysis and metaregression to investigate the influence of N addition (here referring mostly to fertilization) upon the biodiversity of temperate mountain grasslands (including montane, subalpine and alpine zones). Forty-two studies met our criteria of inclusion, resulting in 134 measures of effect size. The main general responses of mountain grasslands to N addition were increases in phytomass and reductions in plant species richness, as observed in lowland grasslands. More specifically, the analysis reveals that negative effects on species richness were exacerbated by dose (ha(-1) year(-1) ) and duration of N application (years) in an additive manner. Thus, sustained application of low to moderate levels of N over time had effects similar to short-term application of high N doses. The climatic context also played an important role: the overall effects of N addition on plant species richness and diversity (Shannon index) were less pronounced in mountain grasslands experiencing cool rather than warm summers. Furthermore, the relative negative effect of N addition on species richness was more pronounced in managed communities and was strongly negatively related to N-induced increases in phytomass, that is the greater the phytomass response to N addition, the greater the decline in richness. Altogether, this review not only establishes that plant biodiversity of mountain grasslands is negatively affected by N addition, but also demonstrates that several local management and abiotic factors interact with N addition to drive plant community changes. This synthesis yields essential information for a more sustainable management of mountain grasslands, emphasizing the importance of preserving and restoring grasslands with both low agricultural N application and limited exposure to N atmospheric deposition.
Response of understory vegetation of Adirondack forests to nitrogen additions
A global budget for fine root biomass, surface area, and nutrient contents
Reduction of forest soil respiration in response to nitrogen deposition
The embarrassment of riches: atmospheric deposition of nitrogen and community and ecosystem processes
Soil extracellular enzyme activities, soil carbon and nitrogen storage under nitrogen fertilization: a meta-analysis
Shifts in dominant tree mycorrhizal associations in response to anthropogenic impacts
Soil restoration in semiarid Patagonia: chemical and biological response to different compost quality
The microbial nitrogen-cycling network
Phylogenetic, functional, and taxonomic richness have both positive and negative effects on ecosystem multifunctionality
Global carbon budget 2015
Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed
DOI:10.1890/06-2057.1
URL
PMID:18409427
[本文引用: 4]
Our meta-analysis of 126 nitrogen addition experiments evaluated nitrogen (N) limitation of net primary production (NPP) in terrestrial ecosystems. We tested the hypothesis that N limitation is widespread among biomes and influenced by geography and climate. We used the response ratio (R approximately equal ANPP(N)/ANPP(ctrl)) of aboveground plant growth in fertilized to control plots and found that most ecosystems are nitrogen limited with an average 29% growth response to nitrogen (i.e., R = 1.29). The response ratio was significant within temperate forests (R = 1.19), tropical forests (R = 1.60), temperate grasslands (R = 1.53), tropical grasslands (R = 1.26), wetlands (R = 1.16), and tundra (R = 1.35), but not deserts. Eight tropical forest studies had been conducted on very young volcanic soils in Hawaii, and this subgroup was strongly N limited (R = 2.13), which resulted in a negative correlation between forest R and latitude. The degree of N limitation in the remainder of the tropical forest studies (R = 1.20) was comparable to that of temperate forests, and when the young Hawaiian subgroup was excluded, forest R did not vary with latitude. Grassland response increased with latitude, but was independent of temperature and precipitation. These results suggest that the global N and C cycles interact strongly and that geography can mediate ecosystem response to N within certain biome types.
Observations of fine and coarse particle nitrate at several rural locations in the United States
Consistent responses of soil microbial communities to elevated nutrient inputs in grasslands across the globe
Microbial functional genes involved in nitrogen fixation, nitrification and denitrification in forest ecosystems
Effects of simulated nitrogen deposition on biomass production and allocation in Schima superba and Cryptocarya concinna seedlings in subtropical China
模拟氮沉降对南亚热带两种乔木幼苗生物量及其分配的影响
Increasing importance of deposition of reduced nitrogen in the United States
Microbes drive global soil nitrogen mineralization and availability
Atmospheric nitrogen deposition impacts on the structure and function of forest mycorrhizal communities: a review
A review of nitrogen enrichment effects on three biogenic GHGs: the CO2 sink may be largely offset by stimulated N2O and CH4 emission
DOI:10.1111/j.1461-0248.2009.01351.x
URL
PMID:19694782
[本文引用: 2]
Anthropogenic nitrogen (N) enrichment of ecosystems, mainly from fuel combustion and fertilizer application, alters biogeochemical cycling of ecosystems in a way that leads to altered flux of biogenic greenhouse gases (GHGs). Our meta-analysis of 313 observations across 109 studies evaluated the effect of N addition on the flux of three major GHGs: CO(2), CH(4) and N(2)O. The objective was to quantitatively synthesize data from agricultural and non-agricultural terrestrial ecosystems across the globe and examine whether factors, such as ecosystem type, N addition level and chemical form of N addition influence the direction and magnitude of GHG fluxes. Results indicate that N addition increased ecosystem carbon content of forests by 6%, marginally increased soil organic carbon of agricultural systems by 2%, but had no significant effect on net ecosystem CO(2) exchange for non-forest natural ecosystems. Across all ecosystems, N addition increased CH(4) emission by 97%, reduced CH(4) uptake by 38% and increased N(2)O emission by 216%. The net effect of N on the global GHG budget is calculated and this topic is reviewed. Most often N addition is considered to increase forest C sequestration without consideration of N stimulation of GHG production in other ecosystems. However, our study indicated that although N addition increased the global terrestrial C sink, the CO(2) reduction could be largely offset (53-76%) by N stimulation of global CH(4) and N(2)O emission from multiple ecosystems.
A global perspective on belowground carbon dynamics under nitrogen enrichment
Nitrogen deposition and its ecological impact in China: an overview
DOI:10.1016/j.envpol.2010.08.002
URL
PMID:20828899
[本文引用: 2]
Nitrogen (N) deposition is an important component in the global N cycle that has induced large impacts on the health and services of terrestrial and aquatic ecosystems worldwide. Anthropogenic reactive N (N(r)) emissions to the atmosphere have increased dramatically in China due to rapid agricultural, industrial and urban development. Therefore increasing N deposition in China and its ecological impacts are of great concern since the 1980s. This paper synthesizes the data from various published papers to assess the status of the anthropogenic N(r) emissions and N deposition as well as their impacts on different ecosystems, including empirical critical loads for different ecosystems. Research challenges and policy implications on atmospheric N pollution and deposition are also discussed. China urgently needs to establish national networks for N deposition monitoring and cross-site N addition experiments in grasslands, forests and aquatic ecosystems. Critical loads and modeling tools will be further used in N(r) regulation.
Enhanced nitrogen deposition over China
DOI:10.1038/nature11917
URL
PMID:23426264
[本文引用: 2]
Responses of ecosystem nitrogen cycle to nitrogen addition: a meta-analysis
Nitrogen deposition contributes to soil acidification in tropical ecosystems
DOI:10.1111/gcb.12665
URL
PMID:24953639
[本文引用: 1]
Effects of experimental nitrogen additions on plant diversity in an old-growth tropical forest
Effects of simulated atmospheric nitrogen deposition on forest ecosystems in China: an overview
模拟大气氮沉降对中国森林生态系统影响的研究进展
Ecosystem carbon storage in arctic tundra reduced by long-term nutrient fertilization
Plant species richness and ecosystem multifunctionality in global drylands
DOI:10.1126/science.1215442
URL
PMID:22246775
[本文引用: 1]
Experiments suggest that biodiversity enhances the ability of ecosystems to maintain multiple functions, such as carbon storage, productivity, and the buildup of nutrient pools (multifunctionality). However, the relationship between biodiversity and multifunctionality has never been assessed globally in natural ecosystems. We report here on a global empirical study relating plant species richness and abiotic factors to multifunctionality in drylands, which collectively cover 41% of Earth's land surface and support over 38% of the human population. Multifunctionality was positively and significantly related to species richness. The best-fitting models accounted for over 55% of the variation in multifunctionality and always included species richness as a predictor variable. Our results suggest that the preservation of plant biodiversity is crucial to buffer negative effects of climate change and desertification in drylands.
The human footprint in the carbon cycle of temperate and boreal forests
Global distribution of atmospheric phosphorus sources, concentrations and deposition rates, and anthropogenic impacts
Redefining ecosystem multifunctionality
Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems
DOI:10.1111/j.1469-8137.2011.03967.x
URL
PMID:22122515
[本文引用: 2]
* Biologically essential elements--especially nitrogen (N) and phosphorus (P)--constrain plant growth and microbial functioning; however, human activities are drastically altering the magnitude and pattern of such nutrient limitations on land. Here we examine interactions between N and P cycles of P mineralizing enzyme activities (phosphatase enzymes) across a wide variety of terrestrial biomes. * We synthesized results from 34 separate studies and used meta-analysis to evaluate phosphatase activity with N, P, or NxP fertilization. * Our results show that N fertilization enhances phosphatase activity, from the tropics to the extra-tropics, both on plant roots and in bulk soils. By contrast, P fertilization strongly suppresses rates of phosphatase activity. * These results imply that phosphatase enzymes are strongly responsive to changes in local nutrient cycle conditions. We also show that plant phosphatases respond more strongly to fertilization than soil phosphatases. The tight coupling between N and P provides a mechanism for recent observations of N and P co-limitation on land. Moreover, our results suggest that terrestrial plants and microbes can allocate excess N to phosphatase enzymes, thus delaying the onset of single P limitation to plant productivity as can occur via human modifications to the global N cycle.
Impacts of nitrogen addition on plant species richness and abundance: a global meta- analysis
Nitrogen addition reduces soil respiration in a mature tropical forest in southern China
Mycorrhizal fungi mediation of terrestrial ecosystem responses to global change: mini-review
An experimental investigation into the effects of atmospheric nitrogen deposition on two semi-natural grasslands
Chronic nitrogen additions fundamentally restructure the soil fungal community in a temperate forest
Short- and medium-term contrasting effects of nitrogen fertilization on C and N cycling in a poplar plantation soil
Effects of nitrogen deposition rates and frequencies on the abundance of soil nitrogen-related functional genes in temperate grassland of northern China
Global patterns and substrate-based mechanisms of the terrestrial nitrogen cycle
DOI:10.1111/ele.12591
URL
PMID:26932540
[本文引用: 1]
Nitrogen (N) deposition is impacting the services that ecosystems provide to humanity. However, the mechanisms determining impacts on the N cycle are not fully understood. To explore the mechanistic underpinnings of N impacts on N cycle processes, we reviewed and synthesised recent progress in ecosystem N research through empirical studies, conceptual analysis and model simulations. Experimental and observational studies have revealed that the stimulation of plant N uptake and soil retention generally diminishes as N loading increases, while dissolved and gaseous losses of N occur at low N availability but increase exponentially and become the dominant fate of N at high loading rates. The original N saturation hypothesis emphasises sequential N saturation from plant uptake to soil retention before N losses occur. However, biogeochemical models that simulate simultaneous competition for soil N substrates by multiple processes match the observed patterns of N losses better than models based on sequential competition. To enable better prediction of terrestrial N cycle responses to N loading, we recommend that future research identifies the response functions of different N processes to substrate availability using manipulative experiments, and incorporates the measured N saturation response functions into conceptual, theoretical and quantitative analyses.
Responses to ammonium and nitrate additions by boreal plants and their natural enemies
Climate Change 2014: Synthesis Report.Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change
A large and persistent carbon sink in the world’s forests
DOI:10.1126/science.1201609
URL
PMID:21764754
[本文引用: 1]
The terrestrial carbon sink has been large in recent decades, but its size and location remain uncertain. Using forest inventory data and long-term ecosystem carbon studies, we estimate a total forest sink of 2.4 +/- 0.4 petagrams of carbon per year (Pg C year(-1)) globally for 1990 to 2007. We also estimate a source of 1.3 +/- 0.7 Pg C year(-1) from tropical land-use change, consisting of a gross tropical deforestation emission of 2.9 +/- 0.5 Pg C year(-1) partially compensated by a carbon sink in tropical forest regrowth of 1.6 +/- 0.5 Pg C year(-1). Together, the fluxes comprise a net global forest sink of 1.1 +/- 0.8 Pg C year(-1), with tropical estimates having the largest uncertainties. Our total forest sink estimate is equivalent in magnitude to the terrestrial sink deduced from fossil fuel emissions and land-use change sources minus ocean and atmospheric sinks.
Nitrogen deposition and plant biodiversity: past, present, and future
Global patterns of root dynamics under nitrogen enrichment
Linkages of plant stoichiometry to ecosystem production and carbon fluxes with increasing nitrogen inputs in an alpine steppe
DOI:10.1111/gcb.13789
URL
PMID:28614594
[本文引用: 1]
Unprecedented levels of nitrogen (N) have entered terrestrial ecosystems over the past century, which substantially influences the carbon (C) exchange between the atmosphere and biosphere. Temperature and moisture are generally regarded as the major controllers over the N effects on ecosystem C uptake and release. N-phosphorous (P) stoichiometry regulates the growth and metabolisms of plants and soil organisms, thereby affecting many ecosystem C processes. However, it remains unclear how the N-induced shift in the plant N:P ratio affects ecosystem production and C fluxes and its relative importance. We conducted a field manipulative experiment with eight N addition levels in a Tibetan alpine steppe and assessed the influences of N on aboveground net primary production (ANPP), gross ecosystem productivity (GEP), ecosystem respiration (ER), and net ecosystem exchange (NEE); we used linear mixed-effects models to further determine the relative contributions of various factors to the N-induced changes in these parameters. Our results showed that the ANPP, GEP, ER, and NEE all exhibited nonlinear responses to increasing N additions. Further analysis demonstrated that the plant N:P ratio played a dominate role in shaping these C exchange processes. There was a positive relationship between the N-induced changes in ANPP (DeltaANPP) and the plant N:P ratio (DeltaN:P), whereas the DeltaGEP, DeltaER, and DeltaNEE exhibited quadratic correlations with the DeltaN:P. In contrast, soil temperature and moisture were only secondary predictors for the changes in ecosystem production and C fluxes along the N addition gradient. These findings highlight the importance of plant N:P ratio in regulating ecosystem C exchange, which is crucial for improving our understanding of C cycles under the scenarios of global N enrichment.
Effects of nitrogen-phosphorus imbalance on plant biomass production: a global perspective
Human-induced nitrogen-phosphorus imbalances alter natural and managed ecosystems across the globe
DOI:10.1038/ncomms3934
URL
PMID:24343268
[本文引用: 2]
The availability of carbon from rising atmospheric carbon dioxide levels and of nitrogen from various human-induced inputs to ecosystems is continuously increasing; however, these increases are not paralleled by a similar increase in phosphorus inputs. The inexorable change in the stoichiometry of carbon and nitrogen relative to phosphorus has no equivalent in Earth's history. Here we report the profound and yet uncertain consequences of the human imprint on the phosphorus cycle and nitrogen:phosphorus stoichiometry for the structure, functioning and diversity of terrestrial and aquatic organisms and ecosystems. A mass balance approach is used to show that limited phosphorus and nitrogen availability are likely to jointly reduce future carbon storage by natural ecosystems during this century. Further, if phosphorus fertilizers cannot be made increasingly accessible, the crop yields projections of the Millennium Ecosystem Assessment imply an increase of the nutrient deficit in developing regions.
The human-induced imbalance between C, N and P in earth’s life system
Atmospheric nitrogen deposition in world biodiversity hotspots: the need for a greater global perspective in assessing N deposition impacts
Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental control
DOI:10.1111/j.1469-8137.2011.03952.x
URL
PMID:22085245
[本文引用: 1]
We quantified the biomass allocation patterns to leaves, stems and roots in vegetative plants, and how this is influenced by the growth environment, plant size, evolutionary history and competition. Dose-response curves of allocation were constructed by means of a meta-analysis from a wide array of experimental data. They show that the fraction of whole-plant mass represented by leaves (LMF) increases most strongly with nutrients and decreases most strongly with light. Correction for size-induced allocation patterns diminishes the LMF-response to light, but makes the effect of temperature on LMF more apparent. There is a clear phylogenetic effect on allocation, as eudicots invest relatively more than monocots in leaves, as do gymnosperms compared with woody angiosperms. Plants grown at high densities show a clear increase in the stem fraction. However, in most comparisons across species groups or environmental factors, the variation in LMF is smaller than the variation in one of the other components of the growth analysis equation: the leaf area : leaf mass ratio (SLA). In competitive situations, the stem mass fraction increases to a smaller extent than the specific stem length (stem length : stem mass). Thus, we conclude that plants generally are less able to adjust allocation than to alter organ morphology.
Nitrogen fertilization inhibits soil microbial respiration regardless of the form of nitrogen applied
Consistent effects of nitrogen amendments on soil microbial communities and processes across biomes
Soil N availability, rather than N deposition, controls indirect N2O emissions
Climate extremes and the carbon cycle
DOI:10.1038/nature12350
URL
PMID:23955228
[本文引用: 1]
The terrestrial biosphere is a key component of the global carbon cycle and its carbon balance is strongly influenced by climate. Continuing environmental changes are thought to increase global terrestrial carbon uptake. But evidence is mounting that climate extremes such as droughts or storms can lead to a decrease in regional ecosystem carbon stocks and therefore have the potential to negate an expected increase in terrestrial carbon uptake. Here we explore the mechanisms and impacts of climate extremes on the terrestrial carbon cycle, and propose a pathway to improve our understanding of present and future impacts of climate extremes on the terrestrial carbon budget.
Evaluating critical loads of nutrient nitrogen and acidity for terrestrial systems using ecosystem-scale experiments (NITREX)
Mechanisms driving the soil organic matter decomposition response to nitrogen enrichment in grassland soils
The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil
Changes in nutrient concentrations of leaves and roots in response to global change factors
DOI:10.1111/gcb.13721
URL
PMID:28407324
[本文引用: 1]
Global change impacts on biogeochemical cycles have been widely studied, but our understanding of whether the responses of plant elemental composition to global change drivers differ between above- and belowground plant organs remains incomplete. We conducted a meta-analysis of 201 reports including 1,687 observations of studies that have analyzed simultaneously N and P concentrations changes in leaves and roots in the same plants in response to drought, elevated [CO2 ], and N and P fertilization around the world, and contrasted the results within those obtained with a general database (838 reports and 14,772 observations) that analyzed the changes in N and P concentrations in leaves and/or roots of plants submitted to the commented global change drivers. At global level, elevated [CO2 ] decreased N concentrations in leaves and roots and decreased N:P ratio in roots but no in leaves, but was not related to P concentration changes. However, the response differed among vegetation types. In temperate forests, elevated [CO2 ] was related with lower N concentrations in leaves but not in roots, whereas in crops, the contrary patterns were observed. Elevated [CO2 ] decreased N concentrations in leaves and roots in tundra plants, whereas not clear relationships were observed in temperate grasslands. However, when elevated [CO2 ] and N fertilization coincided, leaves had lower N concentrations, whereas root had higher N concentrations suggesting that more nutrients will be allocated to roots to improve uptake of the soil resources not directly provided by the global change drivers. N fertilization and drought increased foliar and root N concentrations while the effects on P concentrations were less clear. The changes in N and P allocation to leaves and root, especially those occurring in opposite direction between them have the capacity to differentially affect above- and belowground ecosystem functions, such as litter mineralization and above- and belowground food webs.
The C:N:P stoichiometry of organisms and ecosystems in a changing world: a review and perspectives
Global-scale impacts of nitrogen deposition on tree carbon sequestration in tropical, temperate, and boreal forests: a meta-analysis
Global meta-analysis of the nonlinear response of soil nitrous oxide (N2O) emissions to fertilizer nitrogen
DOI:10.1073/pnas.1322434111
URL
PMID:24927583
[本文引用: 1]
1.5% and soils with pH
Effects of simulated N deposition on photosynthesis and productivity of key plants from different functional groups of alpine meadow on Qinghai- Tibetan Plateau
Conditional vulnerability of plant diversity to atmospheric nitrogen deposition across the United States
DOI:10.1073/pnas.1515241113
URL
PMID:27035943
[本文引用: 1]
Atmospheric nitrogen (N) deposition has been shown to decrease plant species richness along regional deposition gradients in Europe and in experimental manipulations. However, the general response of species richness to N deposition across different vegetation types, soil conditions, and climates remains largely unknown even though responses may be contingent on these environmental factors. We assessed the effect of N deposition on herbaceous richness for 15,136 forest, woodland, shrubland, and grassland sites across the continental United States, to address how edaphic and climatic conditions altered vulnerability to this stressor. In our dataset, with N deposition ranging from 1 to 19 kg Nha(-1)y(-1), we found a unimodal relationship; richness increased at low deposition levels and decreased above 8.7 and 13.4 kg Nha(-1)y(-1) in open and closed-canopy vegetation, respectively. N deposition exceeded critical loads for loss of plant species richness in 24% of 15,136 sites examined nationwide. There were negative relationships between species richness and N deposition in 36% of 44 community gradients. Vulnerability to N deposition was consistently higher in more acidic soils whereas the moderating roles of temperature and precipitation varied across scales. We demonstrate here that negative relationships between N deposition and species richness are common, albeit not universal, and that fine-scale processes can moderate vegetation responses to N deposition. Our results highlight the importance of contingent factors when estimating ecosystem vulnerability to N deposition and suggest that N deposition is affecting species richness in forested and nonforested systems across much of the continental United States.
Extracellular enzyme activities and soil organic matter dynamics for northern hardwood forests receiving simulated nitrogen deposition
Impact of nitrogen deposition on the species richness of grasslands
Anthropogenic nitrogen deposition predicts local grassland primary production worldwide
Physical disturbance determines effects from nitrogen addition on ground vegetation in boreal coniferous forests
Considering fungal:bacterial dominance in soils: methods, controls, and ecosystem implications
Sinks for nitrogen inputs in terrestrial ecosystems: a meta-analysis of 15N tracer field studies
Insights into mechanisms governing forest carbon response to nitrogen deposition: a model-data comparison using observed responses to nitrogen addition
Increased tree carbon storage in response to nitrogen deposition in the US
Responses of forest ecosystems to increasing N deposition in China: a critical review
A global analysis of soil acidification caused by nitrogen addition
A novel soil manganese mechanism drives plant species loss with increased nitrogen deposition in a temperate steppe
DOI:10.1890/15-0917.1
URL
PMID:27008776
[本文引用: 2]
Loss of plant diversity with increased anthropogenic nitrogen (N) deposition in grasslands has occurred globally. In most cases, competitive exclusion driven by preemption of light or space is invoked as a key mechanism. Here, we provide evidence from a 9-yr N-addition experiment for an alternative mechanism: differential sensitivity of forbs and grasses to increased soil manganese (Mn) levels. In Inner Mongolia steppes, increasing the N supply shifted plant community composition from grass-forb codominance (primarily Stipa krylovii and Artemisia frigida, respectively) to exclusive dominance by grass, with associated declines in overall species richness. Reduced abundance of forbs was linked to soil acidification that increased mobilization of soil Mn, with a 10-fold greater accumulation of Mn in forbs than in grasses. The enhanced accumulation of Mn in forbs was correlated with reduced photosynthetic rates and growth, and is consistent with the loss of forb species. Differential accumulation of Mn between forbs and grasses can be linked to fundamental differences between dicots and monocots in the biochemical pathways regulating metal transport. These findings provide a mechanistic explanation for N-induced species loss in temperate grasslands by linking metal mobilization in soil to differential metal acquisition and impacts on key functional groups in these ecosystems.
Nitrogen additions and microbial biomass: a meta-analysis of ecosystem studies
The unseen majority: soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems
Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity
Chronic nitrogen deposition and the composition of active arbuscular mycorrhizal fungi
Simulated nitrogen deposition causes a decline of intra- and extraradical abundance of arbuscular mycorrhizal fungi and changes in microbial community structure in northern hardwood forests
Nitrogen limitation on land and in the sea: How can it occur?
Terrestrial phosphorus limitation: mechanisms, implications, and nitrogen-phosphorus interactions
DOI:10.1890/08-0127.1
URL
PMID:20349827
[本文引用: 1]
Nutrient limitation to primary productivity and other biological processes is widespread in terrestrial ecosystems, and nitrogen (N) and phosphorus (P) are the most common limiting elements, both individually and in combination. Mechanisms that drive P limitation, and their interactions with the N cycle, have received less attention than mechanisms causing N limitation. We identify and discuss six mechanisms that could drive P limitation in terrestrial ecosystems. The best known of these is depletion-driven limitation, in which accumulated P losses during long-term soil and ecosystem development contribute to what Walker and Syers termed a
Response of oxidative enzyme activities to nitrogen deposition affects soil concentrations of dissolved organic carbon
Non-random species loss in a forest herbaceous layer following nitrogen addition
Decreasing soil microbial diversity is associated with decreasing microbial biomass under nitrogen addition
Responses of soil microbial community to continuous experimental nitrogen additions for 13 years in a nitrogen-rich tropical forest
Molecular mechanisms of water table lowering and nitrogen deposition in affecting greenhouse gas emissions from a Tibetan alpine wetland
DOI:10.1111/gcb.13467
URL
PMID:27536811
[本文引用: 1]
Rapid climate change and intensified human activities have resulted in water table lowering (WTL) and enhanced nitrogen (N) deposition in Tibetan alpine wetlands. These changes may alter the magnitude and direction of greenhouse gas (GHG) emissions, affecting the climate impact of these fragile ecosystems. We conducted a mesocosm experiment combined with a metagenomics approach (GeoChip 5.0) to elucidate the effects of WTL (-20 cm relative to control) and N deposition (30 kg N ha(-1) yr(-1) ) on carbon dioxide (CO2 ), methane (CH4 ) and nitrous oxide (N2 O) fluxes as well as the underlying mechanisms. Our results showed that WTL reduced CH4 emissions by 57.4% averaged over three growing seasons compared with no-WTL plots, but had no significant effect on net CO2 uptake or N2 O flux. N deposition increased net CO2 uptake by 25.2% in comparison with no-N deposition plots and turned the mesocosms from N2 O sinks to N2 O sources, but had little influence on CH4 emissions. The interactions between WTL and N deposition were not detected in all GHG emissions. As a result, WTL and N deposition both reduced the global warming potential (GWP) of growing season GHG budgets on a 100-year time horizon, but via different mechanisms. WTL reduced GWP from 337.3 to -480.1 g CO2 -eq m(-2) mostly because of decreased CH4 emissions, while N deposition reduced GWP from 21.0 to -163.8 g CO2 -eq m(-2) , mainly owing to increased net CO2 uptake. GeoChip analysis revealed that decreased CH4 production potential, rather than increased CH4 oxidation potential, may lead to the reduction in net CH4 emissions, and decreased nitrification potential and increased denitrification potential affected N2 O fluxes under WTL conditions. Our study highlights the importance of microbial mechanisms in regulating ecosystem-scale GHG responses to environmental changes.
Asymmetry in above- and belowground productivity responses to N addition in a semi-arid temperate steppe
Fine root dynamics responses to nitrogen addition depend on root order, soil layer, and experimental duration in a subtropical forest
Differences in fungal and bacterial physiology alter soil carbon and nitrogen cycling: insights from meta-analysis and theoretical models
DOI:10.1111/ele.12125
URL
PMID:23692657
[本文引用: 1]
Long term effect of nitrogen addition on understory community in a Chinese boreal forest
DOI:10.1016/j.scitotenv.2018.07.350
URL
PMID:30235651
[本文引用: 2]
Increasing atmospheric nitrogen (N) deposition is an important driver of biodiversity change. By conducting an eight-yearN addition experiment (0, 20, 50 and 100kgNha(-1)yr(-1)), we investigated the long-term effect of simulated N deposition on understory species composition and richness in a boreal forest, northeast China. We found that moss cover decreased significantly with increasing N addition. N addition had no significant effect on vascular plants species richness but changed the plant community composition. The relative coverage of evergreen shrubs decreased, while that of graminoids increased under high-level N addition (100kgNha(-1)yr(-1)). Under the high-level N treatment, Deyeuxia angustifolia cover increased significantly after 4years, while that of Vaccinium vitis-idaea decreased significantly after 3years and almost disappeared after 5years. The negative effect of N addition on mosses and evergreen shrubs accumulated over time, while the positive effect on graminoids increased during the first 4years and did not change significantly thereafter. Our results suggest that the effect of N deposition varies across functional groups and shifts over time.
Water- and plant-mediated responses of soil respiration to topography, fire, and nitrogen fertilization in a semiarid grassland in northern China
Rate-specific responses of prokaryotic diversity and structure to nitrogen deposition in the Leymus chinensis steppe
Reconciling multiple impacts of nitrogen enrichment on soil carbon: plant, microbial and geochemical controls
Stabilization of atmospheric nitrogen deposition in China over the past decade
Linking stoichiometric homoeostasis with ecosystem structure, functioning and stability
Decoupling of nitrogen and phosphorus in terrestrial plants associated with global changes
Effects of three global change drivers on terrestrial C:N:P stoichiometry: a global synthesis
DOI:10.1111/gcb.13569
URL
PMID:27859966
[本文引用: 2]
Over the last few decades, there has been an increasing number of controlled-manipulative experiments to investigate how plants and soils might respond to global change. These experiments typically examined the effects of each of three global change drivers [i.e., nitrogen (N) deposition, warming, and elevated CO2 ] on primary productivity and on the biogeochemistry of carbon (C), N, and phosphorus (P) across different terrestrial ecosystems. Here, we capitalize on this large amount of information by performing a comprehensive meta-analysis (>2000 case studies worldwide) to address how C:N:P stoichiometry of plants, soils, and soil microbial biomass might respond to individual vs. combined effects of the three global change drivers. Our results show that (i) individual effects of N addition and elevated CO2 on C:N:P stoichiometry are stronger than warming, (ii) combined effects of pairs of global change drivers (e.g., N addition + elevated CO2 , warming + elevated CO2 ) on C:N:P stoichiometry were generally weaker than the individual effects of each of these drivers, (iii) additive interactions (i.e., when combined effects are equal to or not significantly different from the sum of individual effects) were more common than synergistic or antagonistic interactions, (iv) C:N:P stoichiometry of soil and soil microbial biomass shows high homeostasis under global change manipulations, and (v) C:N:P responses to global change are strongly affected by ecosystem type, local climate, and experimental conditions. Our study is one of the first to compare individual vs. combined effects of the three global change drivers on terrestrial C:N:P ratios using a large set of data. To further improve our understanding of how ecosystems might respond to future global change, long-term ecosystem-scale studies testing multifactor effects on plants and soils are urgently required across different world regions.
Carbon benefits of anthropogenic reactive nitrogen offset by nitrous oxide emissions
Microbial responses to nitrogen addition in three contrasting grassland ecosystems
DOI:10.1007/s00442-007-0836-6
URL
PMID:17724617
[本文引用: 1]
Global negative effects of nitrogen deposition on soil microbes
DOI:10.1038/s41396-018-0096-y
URL
PMID:29588494
[本文引用: 3]
Soil microbes comprise a large portion of the genetic diversity on Earth and influence a large number of important ecosystem processes. Increasing atmospheric nitrogen (N) deposition represents a major global change driver; however, it is still debated whether the impacts of N deposition on soil microbial biomass and respiration are ecosystem-type dependent. Moreover, the extent of N deposition impacts on microbial composition remains unclear. Here we conduct a global meta-analysis using 1408 paired observations from 151 studies to evaluate the responses of soil microbial biomass, composition, and function to N addition. We show that nitrogen addition reduced total microbial biomass, bacterial biomass, fungal biomass, biomass carbon, and microbial respiration. Importantly, these negative effects increased with N application rate and experimental duration. Nitrogen addition reduced the fungi to bacteria ratio and the relative abundances of arbuscular mycorrhizal fungi and gram-negative bacteria and increased gram-positive bacteria. Our structural equation modeling showed that the negative effects of N application on soil microbial abundance and composition led to reduced microbial respiration. The effects of N addition were consistent across global terrestrial ecosystems. Our results suggest that atmospheric N deposition negatively affects soil microbial growth, composition, and function across all terrestrial ecosystems, with more pronounced effects with increasing N deposition rate and duration.
Rapid plant species loss at high rates and at low frequency of N addition in temperate steppe
DOI:10.1111/gcb.12611
URL
PMID:24753127
[本文引用: 3]
Humans are both intentionally (fertilization) and unintentionally (atmospheric nutrient deposition) adding nutrients worldwide. Increasing availability of biologically reactive nitrogen (N) is one of the major drivers of plant species loss. It remains unclear, however, whether plant diversity will be equally reduced by inputs of reactive N coming from either small and frequent N deposition events or large and infrequent N fertilization events. By independently manipulating the rate and frequency of reactive N inputs, our study teases apart these potentially contrasting effects. Plant species richness decreased more quickly at high rates and at low frequency of N addition, which suggests that previous fertilization studies have likely over-estimated the effects of N deposition on plant species loss. N-induced species loss resulted from both acidification and ammonium toxicity. Further study of small and frequent N additions will be necessary to project future rates of plant species loss under increasing aerial N deposition.
Influences of canopy nitrogen and water addition on AM fungal biodiversity and community composition in a mixed deciduous forest of China
Different responses of soil respiration and its components to nitrogen addition among biomes: a meta-analysis
DOI:10.1111/gcb.12490
URL
PMID:24323545
[本文引用: 2]
Patterns and mechanisms of responses by soil microbial communities to nitrogen addition
The composition, spatial patterns, and influencing factors of atmospheric wet nitrogen deposition in Chinese terrestrial ecosystems
DOI:10.1016/j.scitotenv.2014.12.038
URL
PMID:25617702
[本文引用: 1]
Microbial stoichiometric flexibility regulates rice straw mineralization and its priming effect in paddy soil
Nitrogen deposition but not climate warming promotes Deyeuxia angustifolia encroachment in alpine tundra of the Changbai Mountains, Northeast China

{kind=link}
{kind=link}
{kind=link}
{kind=link}