Logging residue harvest may decrease enzymatic activity of boreal forest soils
1
2015
... 森林更新演替的恢复过程中, 林分结构、物种组成、凋落物量以及林下微气候发生了不同程度的变化, 导致其林下土壤碳氮源不同而引起土壤有机质分解速率、土壤微生物多样性及其代谢活动发生变化(Wardle et al., 2004; Lucas-Borja & Delgado- Baquerizo, 2019), 进而影响到土壤酶活性(Adamczyk et al., 2015; Xu et al., 2017).此外, 年平均气温、降水量、土壤酸碱性和物种多样性也是影响土壤酶活性的主要因素(于洋等, 2014; Zhou et al., 2020).本研究中, 天然次生林更新早期演替(<60 a)不同阶段的表层土壤水解酶(βG、CBH和NAG)活性显著低于岷江冷杉原始林(表4), 且随自然更新演替进程呈显著降低趋势(80-NSF > 70-NSF > 60-NSF), 不同于川西亚高山平武县白桦林水解酶活性高于冷杉林(熊浩仲等, 2004)及温带白桦(Betula platyphylla)林随演替进程表现为先增加后降低的变化趋势(胡嵩等, 2013), 一方面说明岷江冷杉原始林的土壤养分循环速率相对较高, 对土壤养分供给更快; 另一方面说明次生林自然更新演替过程中, 地上凋落物增加导致土壤碳氮及其活性组分含量的下降(Liu et al., 2017; Lyu et al., 2019), 使其土壤微生物活性降低(辜翔等, 2018), 进而导致土壤水解酶活性下降.此外, 80-NSF、70-NSF和60-NSF土壤中氧化酶活性变化不显著, 但PHO的活性显著低于CK, 而PEO的活性与CK差异不显著(表4), 其原因可能是岷江冷杉原始林中含有较为丰富的木质素、角质和蜡质等难降解化合物, 促进了降解木质素的微生物群落多样性或种类数量增加, 且随次生林更新演替阶段的进行, 根系随着林上植被的演替而发生变化, 导致土壤的pH增加, 致使PHO活性增加(Fujii et al., 2013). ...
Plant roots increase both decomposition and stable organic matter formation in boreal forest soil
1
2019
... 森林自然更新使其树种组成、凋落物数量以及植物根系发生变化而影响到土壤碳氮循环过程(范跃新等, 2013; Dhillon & van Rees, 2017).有研究认为, 凋落物输入量会促进部分新输入土壤的碳分解释放而产生“激发效应”, 加速土壤中原有的有机质分解(Saiya-Cork et al., 2002; Qiao et al., 2019), 随着森林更新演替的进行, 输入到土壤的凋落物量渐增(张远东等, 2019), 从而促使土壤原有储存的部分有机质分解, 且植物根系含量的增加提高了土壤有机质分解速率(Adamczyk et al., 2019), 使得土壤有机碳氮储量随森林更新演替呈显著下降趋势, 主要因为土壤有机碳氮储量受到植物生长以及凋落物种类组成的影响(del Valle et al., 2020).本研究中, 原始冷杉林(CK)中SOC和TN含量显著高于3种天然次生林(p < 0.05), 而在3种天然次生林中, SOC含量表现为80-NSF > 70-NSF > 60-NSF和TN为70-NSF > 80-NSF > 60-NSF, 其中80-NSF和70-NSF差异不显著(p > 0.05), 一方面由于采伐迹地自然更新演替次生林的早期阶段, 土壤微生境的变化加速了土壤中累积的有机质分解速率, 以及阔叶树种中的碳和木质素多低于针叶树种且分解速率较快(谌贤等, 2017); 另一方面天然次生林演替至60年, 外来物种进入难度增加致使物种多样性出现降低趋势(图3), 导致DON含量减少而影响到土壤SOC和TN含量.综上, 在川西亚高山高海拔特殊生境条件及其生态脆弱环境背景下, 如森林遭受破坏后, 其土壤肥力难以恢复.本研究仅对60年内不同演替阶段的次生林进行了观察, 对土壤碳氮动态变化机制的理解有限, 但研究结果显示, 如果要恢复到采伐前原始林的土壤肥力水平(表3)需要更长的时间. ...
中亚热带3种典型森林土壤碳氮含量和酶活性的关系
2018
中亚热带3种典型森林土壤碳氮含量和酶活性的关系
2018
Soil enzymes in a changing environment: current knowledge and future directions
1
2013
... 土壤活性有机碳氮是土壤可溶性有机质的主要组分, 主要来源于新近枯枝落叶、腐殖质、根系分泌物以及微生物残体(Kalbitz et al., 2000; 鲍勇等, 2018).森林不同自然更新恢复阶段, 其林分土壤肥力难以恢复及物种组成的差异, 导致地表凋落物质量和数量、土壤温湿度、微生物组成等发生不同程度变化, 从而影响到土壤活性有机碳氮含量的变化(张雪等, 2016; Hagedorn et al., 2019).本研究中, DOC、LFOC和DON含量均随次生林演替呈显著下降趋势(p < 0.05), 且在70-NSF和80-NSF中差异不显著(表3), 而演替到岷江冷杉原始林阶段, 其土壤活性碳氮含量显著增加(p < 0.05), 这与该区域人工林演替过程中表层(0-20 cm)土壤活性碳氮含量的研究结果相似(姜发艳等, 2009), 但不同于热带(张哲等, 2019)和亚热带(范跃新等, 2013)森林演替的研究结果, 说明不同温湿条件及植被林分结构对土壤活性有机碳氮组分含量的影响表现出不同的变化特征(Li et al., 2014; Taylor et al., 2019).究其原因, 一方面随着森林更新演替, 土壤碳氮矿化速率增加(Mylliemngap et al., 2016), 且阔叶林的矿化速率高于针叶林(Côté et al., 2000); 另一方面由于表层土壤密度随演替增加, 而土壤团聚度、结构系数、总孔隙及毛管孔隙等土壤物理性质则呈现降低趋势(庞学勇等, 2003), 土壤有机碳、氮缺少土壤团聚体的物理性保护, 导致土壤活性有机碳和DON含量降低.此外, 岷江冷杉原始林的土壤活性碳氮组分含量高于天然次生林(表3), 可能与该林分主要物种构成(针叶+阔叶树种)有一定关联, 源于该区域针阔树种凋落叶融合在一起易于分解(黄玉梅等, 2015), 其释放的养分便于微生物分解利用, 导致土壤活性有机碳含量增加.如要明晰上述影响因素对土壤活性有机碳氮的驱动机理, 仍需进一步深入研究.相关分析表明, 不同天然更新演替阶段林地表层土壤SOC和TN含量与活性有机碳各组分含量及DON含量均呈极显著正相关关系(p < 0.01), 说明土壤活性有机碳和氮含量与土壤SOC、TN含量之间关系极为密切, 这与已有研究(辜翔等, 2016; 宫立等, 2017) 一致, 说明天然次生林土壤氮储量是调控其碳固持的重要参数(Li et al., 2012; Smal et al., 2019). ...
Impact of forest vegetation on soil characteristics: a correlation between soil biological and physico-chemical properties
1
2016
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
Costimulation of soil glycosidase activity and soil respiration by nitrogen addition
1
2017
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
川西亚高山针叶林土壤酶及其化学计量比对模拟氮沉降的响应
2019
川西亚高山针叶林土壤酶及其化学计量比对模拟氮沉降的响应
2019
Dynamics of carbon and nitrogen mineralization in relation to stand type, stand age and soil texture in the boreal mixedwood
2000
Temporal and vertical variation of selected extracellular enzyme activities on tree litter degradation of a subtropical forest
1
2019
... 在川西亚高山森林土壤氮相对缺乏背景下, 增加土壤氮含量可提高NAG和PEO活性(陈倩妹等, 2019).RDA分析进一步发现, TN、LFOC和DOC含量与土壤酶活性显著正相关(图2), 且分别解释了土壤酶活性变化的69.2%、13.9%和3.2%, 说明在森林土壤氮限制背景下, 天然次生林演替早期阶段表层土壤氧化酶和水解酶的酶活性变化主要受控于TN、LFOC和DOC含量, 表明在次生林天然更新演替过程中, 土壤TN含量及土壤碳有效性降低可能导致酶促反应底物的减少, 进而抑制土壤微生物活性, 使其与碳氮循环相关的土壤酶活性降低. ...
The influence of time, storage temperature, and substrate age on potential soil enzyme activity in acidic forest soils using MUB-linked substrates and l-DOPA
1
2009
... 土壤活性有机碳氮是土壤可溶性有机质的主要组分, 主要来源于新近枯枝落叶、腐殖质、根系分泌物以及微生物残体(Kalbitz et al., 2000; 鲍勇等, 2018).森林不同自然更新恢复阶段, 其林分土壤肥力难以恢复及物种组成的差异, 导致地表凋落物质量和数量、土壤温湿度、微生物组成等发生不同程度变化, 从而影响到土壤活性有机碳氮含量的变化(张雪等, 2016; Hagedorn et al., 2019).本研究中, DOC、LFOC和DON含量均随次生林演替呈显著下降趋势(p < 0.05), 且在70-NSF和80-NSF中差异不显著(表3), 而演替到岷江冷杉原始林阶段, 其土壤活性碳氮含量显著增加(p < 0.05), 这与该区域人工林演替过程中表层(0-20 cm)土壤活性碳氮含量的研究结果相似(姜发艳等, 2009), 但不同于热带(张哲等, 2019)和亚热带(范跃新等, 2013)森林演替的研究结果, 说明不同温湿条件及植被林分结构对土壤活性有机碳氮组分含量的影响表现出不同的变化特征(Li et al., 2014; Taylor et al., 2019).究其原因, 一方面随着森林更新演替, 土壤碳氮矿化速率增加(Mylliemngap et al., 2016), 且阔叶林的矿化速率高于针叶林(Côté et al., 2000); 另一方面由于表层土壤密度随演替增加, 而土壤团聚度、结构系数、总孔隙及毛管孔隙等土壤物理性质则呈现降低趋势(庞学勇等, 2003), 土壤有机碳、氮缺少土壤团聚体的物理性保护, 导致土壤活性有机碳和DON含量降低.此外, 岷江冷杉原始林的土壤活性碳氮组分含量高于天然次生林(表3), 可能与该林分主要物种构成(针叶+阔叶树种)有一定关联, 源于该区域针阔树种凋落叶融合在一起易于分解(黄玉梅等, 2015), 其释放的养分便于微生物分解利用, 导致土壤活性有机碳含量增加.如要明晰上述影响因素对土壤活性有机碳氮的驱动机理, 仍需进一步深入研究.相关分析表明, 不同天然更新演替阶段林地表层土壤SOC和TN含量与活性有机碳各组分含量及DON含量均呈极显著正相关关系(p < 0.01), 说明土壤活性有机碳和氮含量与土壤SOC、TN含量之间关系极为密切, 这与已有研究(辜翔等, 2016; 宫立等, 2017) 一致, 说明天然次生林土壤氮储量是调控其碳固持的重要参数(Li et al., 2012; Smal et al., 2019). ...
Soil organic matter attenuates the efficacy of flavonoid-based plant-microbe communication
1
2020
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
Soil organic carbon storage capacity positively related to forest succession on the Loess Plateau, China
1
2013
... 土壤酶活性参照DeForest (2009)和Chen等(2017)的方法: 取新鲜土壤1 g, 加蒸馏水125 mL, 振荡2 h (25 ℃, 180 r·min-1), 吸取1 mL的悬浊液, 用4-甲基伞形酮做标准曲线, 用4-MUB-β-D-乙酰基氨基葡萄糖苷和4-MUB-β-D-葡萄糖苷为底物标示酶活性, 置于25 ℃下避光培养后, 用多功能酶标仪(M200PRO, TECAN, 广州)测定β-4-N-乙酰基氨基葡萄糖苷酶和β-4-葡萄糖苷酶; 过氧化物酶和多酚氧化酶采用酶联免疫微量法: 分别取过50目的风干土样0.200 g, 加入底物(邻苯三酚) 120 mL和100 μL 1%邻苯三酚溶液和20 μL过氧化氢溶液, 混匀后置于30 ℃恒温培育后, 用紫色没食子素作为标准曲线, 分别加入50 mL和50 μL PBS及430 mL乙醚, 振荡并静置后, 用多功能酶标仪测定多酚氧化酶和过氧化物酶; 纤维素水解酶采用3,5-二硝基水杨酸比色法测定: 取10 g土壤, 加入1.5 mL甲苯, 摇匀放置15 min后, 用葡萄糖作出标准曲线, 加5 mL 1%羧甲基纤维素溶液和5 mL pH为5.5的醋酸盐缓冲液, 37 ℃恒温箱培养后, 过滤用岛津-UV2450-紫外分光光度计进行比色测定.5种土壤酶的名称、缩写、功能和标定底物见表2.酶活性均通过预实验确定获得最大酶活性所需要的底物浓度和培养时间. ...
Distribution of soil organic carbon in the light and heavy fractions for six shelterbelt species and their adjacent agricultural fields in Saskatchewan
2
2017
... DOC和DON含量测定: 取10 g风干土样, 按土水体积比1:5加入蒸馏水, 室温下振荡后过0.45 μm滤膜后, 用岛津SOC-VcPH分析仪(岛津, 广州)测定DOC含量和紫外分光光度计(UV-2450, 岛津, 广州)测定可溶性全氮(DTN)含量, 可溶性无机氮(DIN) 含量用AA3-连续流动分析仪测定, DON含量= DTN含量- DIN含量; LFOC含量测定参考Dhillon和van Rees (2017)的方法, 取过2 mm筛的风干土样20.0 g, 加入1.7 g·cm-3 NaI溶液50 mL, 振荡(≥30 min)离 心后, 取轻质部分过滤(0.45 μm纤维滤膜), 再用0.01 mol·L-1 CaCl2和蒸馏水抽滤冲洗, 烘干(60 ℃)过0.15 mm筛, 用岛津SOC-VcPH分析仪测定. ...
... 森林自然更新使其树种组成、凋落物数量以及植物根系发生变化而影响到土壤碳氮循环过程(范跃新等, 2013; Dhillon & van Rees, 2017).有研究认为, 凋落物输入量会促进部分新输入土壤的碳分解释放而产生“激发效应”, 加速土壤中原有的有机质分解(Saiya-Cork et al., 2002; Qiao et al., 2019), 随着森林更新演替的进行, 输入到土壤的凋落物量渐增(张远东等, 2019), 从而促使土壤原有储存的部分有机质分解, 且植物根系含量的增加提高了土壤有机质分解速率(Adamczyk et al., 2019), 使得土壤有机碳氮储量随森林更新演替呈显著下降趋势, 主要因为土壤有机碳氮储量受到植物生长以及凋落物种类组成的影响(del Valle et al., 2020).本研究中, 原始冷杉林(CK)中SOC和TN含量显著高于3种天然次生林(p < 0.05), 而在3种天然次生林中, SOC含量表现为80-NSF > 70-NSF > 60-NSF和TN为70-NSF > 80-NSF > 60-NSF, 其中80-NSF和70-NSF差异不显著(p > 0.05), 一方面由于采伐迹地自然更新演替次生林的早期阶段, 土壤微生境的变化加速了土壤中累积的有机质分解速率, 以及阔叶树种中的碳和木质素多低于针叶树种且分解速率较快(谌贤等, 2017); 另一方面天然次生林演替至60年, 外来物种进入难度增加致使物种多样性出现降低趋势(图3), 导致DON含量减少而影响到土壤SOC和TN含量.综上, 在川西亚高山高海拔特殊生境条件及其生态脆弱环境背景下, 如森林遭受破坏后, 其土壤肥力难以恢复.本研究仅对60年内不同演替阶段的次生林进行了观察, 对土壤碳氮动态变化机制的理解有限, 但研究结果显示, 如果要恢复到采伐前原始林的土壤肥力水平(表3)需要更长的时间. ...
Carbon control on terrestrial ecosystem function across contrasting site productivities: the carbon connection revisited
1
2019
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
中亚热带常绿阔叶林不同演替阶段土壤活性有机碳含量及季节动态
1
2013
... 森林自然更新使其树种组成、凋落物数量以及植物根系发生变化而影响到土壤碳氮循环过程(范跃新等, 2013; Dhillon & van Rees, 2017).有研究认为, 凋落物输入量会促进部分新输入土壤的碳分解释放而产生“激发效应”, 加速土壤中原有的有机质分解(Saiya-Cork et al., 2002; Qiao et al., 2019), 随着森林更新演替的进行, 输入到土壤的凋落物量渐增(张远东等, 2019), 从而促使土壤原有储存的部分有机质分解, 且植物根系含量的增加提高了土壤有机质分解速率(Adamczyk et al., 2019), 使得土壤有机碳氮储量随森林更新演替呈显著下降趋势, 主要因为土壤有机碳氮储量受到植物生长以及凋落物种类组成的影响(del Valle et al., 2020).本研究中, 原始冷杉林(CK)中SOC和TN含量显著高于3种天然次生林(p < 0.05), 而在3种天然次生林中, SOC含量表现为80-NSF > 70-NSF > 60-NSF和TN为70-NSF > 80-NSF > 60-NSF, 其中80-NSF和70-NSF差异不显著(p > 0.05), 一方面由于采伐迹地自然更新演替次生林的早期阶段, 土壤微生境的变化加速了土壤中累积的有机质分解速率, 以及阔叶树种中的碳和木质素多低于针叶树种且分解速率较快(谌贤等, 2017); 另一方面天然次生林演替至60年, 外来物种进入难度增加致使物种多样性出现降低趋势(图3), 导致DON含量减少而影响到土壤SOC和TN含量.综上, 在川西亚高山高海拔特殊生境条件及其生态脆弱环境背景下, 如森林遭受破坏后, 其土壤肥力难以恢复.本研究仅对60年内不同演替阶段的次生林进行了观察, 对土壤碳氮动态变化机制的理解有限, 但研究结果显示, 如果要恢复到采伐前原始林的土壤肥力水平(表3)需要更长的时间. ...
中亚热带常绿阔叶林不同演替阶段土壤活性有机碳含量及季节动态
1
2013
... 森林自然更新使其树种组成、凋落物数量以及植物根系发生变化而影响到土壤碳氮循环过程(范跃新等, 2013; Dhillon & van Rees, 2017).有研究认为, 凋落物输入量会促进部分新输入土壤的碳分解释放而产生“激发效应”, 加速土壤中原有的有机质分解(Saiya-Cork et al., 2002; Qiao et al., 2019), 随着森林更新演替的进行, 输入到土壤的凋落物量渐增(张远东等, 2019), 从而促使土壤原有储存的部分有机质分解, 且植物根系含量的增加提高了土壤有机质分解速率(Adamczyk et al., 2019), 使得土壤有机碳氮储量随森林更新演替呈显著下降趋势, 主要因为土壤有机碳氮储量受到植物生长以及凋落物种类组成的影响(del Valle et al., 2020).本研究中, 原始冷杉林(CK)中SOC和TN含量显著高于3种天然次生林(p < 0.05), 而在3种天然次生林中, SOC含量表现为80-NSF > 70-NSF > 60-NSF和TN为70-NSF > 80-NSF > 60-NSF, 其中80-NSF和70-NSF差异不显著(p > 0.05), 一方面由于采伐迹地自然更新演替次生林的早期阶段, 土壤微生境的变化加速了土壤中累积的有机质分解速率, 以及阔叶树种中的碳和木质素多低于针叶树种且分解速率较快(谌贤等, 2017); 另一方面天然次生林演替至60年, 外来物种进入难度增加致使物种多样性出现降低趋势(图3), 导致DON含量减少而影响到土壤SOC和TN含量.综上, 在川西亚高山高海拔特殊生境条件及其生态脆弱环境背景下, 如森林遭受破坏后, 其土壤肥力难以恢复.本研究仅对60年内不同演替阶段的次生林进行了观察, 对土壤碳氮动态变化机制的理解有限, 但研究结果显示, 如果要恢复到采伐前原始林的土壤肥力水平(表3)需要更长的时间. ...
Soil enzyme activities increase following restoration of degraded subtropical forests
1
2019
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
Environmental control of lignin peroxidase, manganese peroxidase, and laccase activities in forest floor layers in humid Asia
2013
川西卧龙岷江冷杉林土壤有机碳组分与氮素关系随海拔梯度的变化特征
2
2017
... 森林自然更新使其树种组成、凋落物数量以及植物根系发生变化而影响到土壤碳氮循环过程(范跃新等, 2013; Dhillon & van Rees, 2017).有研究认为, 凋落物输入量会促进部分新输入土壤的碳分解释放而产生“激发效应”, 加速土壤中原有的有机质分解(Saiya-Cork et al., 2002; Qiao et al., 2019), 随着森林更新演替的进行, 输入到土壤的凋落物量渐增(张远东等, 2019), 从而促使土壤原有储存的部分有机质分解, 且植物根系含量的增加提高了土壤有机质分解速率(Adamczyk et al., 2019), 使得土壤有机碳氮储量随森林更新演替呈显著下降趋势, 主要因为土壤有机碳氮储量受到植物生长以及凋落物种类组成的影响(del Valle et al., 2020).本研究中, 原始冷杉林(CK)中SOC和TN含量显著高于3种天然次生林(p < 0.05), 而在3种天然次生林中, SOC含量表现为80-NSF > 70-NSF > 60-NSF和TN为70-NSF > 80-NSF > 60-NSF, 其中80-NSF和70-NSF差异不显著(p > 0.05), 一方面由于采伐迹地自然更新演替次生林的早期阶段, 土壤微生境的变化加速了土壤中累积的有机质分解速率, 以及阔叶树种中的碳和木质素多低于针叶树种且分解速率较快(谌贤等, 2017); 另一方面天然次生林演替至60年, 外来物种进入难度增加致使物种多样性出现降低趋势(图3), 导致DON含量减少而影响到土壤SOC和TN含量.综上, 在川西亚高山高海拔特殊生境条件及其生态脆弱环境背景下, 如森林遭受破坏后, 其土壤肥力难以恢复.本研究仅对60年内不同演替阶段的次生林进行了观察, 对土壤碳氮动态变化机制的理解有限, 但研究结果显示, 如果要恢复到采伐前原始林的土壤肥力水平(表3)需要更长的时间. ...
... 土壤活性有机碳氮是土壤可溶性有机质的主要组分, 主要来源于新近枯枝落叶、腐殖质、根系分泌物以及微生物残体(Kalbitz et al., 2000; 鲍勇等, 2018).森林不同自然更新恢复阶段, 其林分土壤肥力难以恢复及物种组成的差异, 导致地表凋落物质量和数量、土壤温湿度、微生物组成等发生不同程度变化, 从而影响到土壤活性有机碳氮含量的变化(张雪等, 2016; Hagedorn et al., 2019).本研究中, DOC、LFOC和DON含量均随次生林演替呈显著下降趋势(p < 0.05), 且在70-NSF和80-NSF中差异不显著(表3), 而演替到岷江冷杉原始林阶段, 其土壤活性碳氮含量显著增加(p < 0.05), 这与该区域人工林演替过程中表层(0-20 cm)土壤活性碳氮含量的研究结果相似(姜发艳等, 2009), 但不同于热带(张哲等, 2019)和亚热带(范跃新等, 2013)森林演替的研究结果, 说明不同温湿条件及植被林分结构对土壤活性有机碳氮组分含量的影响表现出不同的变化特征(Li et al., 2014; Taylor et al., 2019).究其原因, 一方面随着森林更新演替, 土壤碳氮矿化速率增加(Mylliemngap et al., 2016), 且阔叶林的矿化速率高于针叶林(Côté et al., 2000); 另一方面由于表层土壤密度随演替增加, 而土壤团聚度、结构系数、总孔隙及毛管孔隙等土壤物理性质则呈现降低趋势(庞学勇等, 2003), 土壤有机碳、氮缺少土壤团聚体的物理性保护, 导致土壤活性有机碳和DON含量降低.此外, 岷江冷杉原始林的土壤活性碳氮组分含量高于天然次生林(表3), 可能与该林分主要物种构成(针叶+阔叶树种)有一定关联, 源于该区域针阔树种凋落叶融合在一起易于分解(黄玉梅等, 2015), 其释放的养分便于微生物分解利用, 导致土壤活性有机碳含量增加.如要明晰上述影响因素对土壤活性有机碳氮的驱动机理, 仍需进一步深入研究.相关分析表明, 不同天然更新演替阶段林地表层土壤SOC和TN含量与活性有机碳各组分含量及DON含量均呈极显著正相关关系(p < 0.01), 说明土壤活性有机碳和氮含量与土壤SOC、TN含量之间关系极为密切, 这与已有研究(辜翔等, 2016; 宫立等, 2017) 一致, 说明天然次生林土壤氮储量是调控其碳固持的重要参数(Li et al., 2012; Smal et al., 2019). ...
川西卧龙岷江冷杉林土壤有机碳组分与氮素关系随海拔梯度的变化特征
2
2017
... 森林自然更新使其树种组成、凋落物数量以及植物根系发生变化而影响到土壤碳氮循环过程(范跃新等, 2013; Dhillon & van Rees, 2017).有研究认为, 凋落物输入量会促进部分新输入土壤的碳分解释放而产生“激发效应”, 加速土壤中原有的有机质分解(Saiya-Cork et al., 2002; Qiao et al., 2019), 随着森林更新演替的进行, 输入到土壤的凋落物量渐增(张远东等, 2019), 从而促使土壤原有储存的部分有机质分解, 且植物根系含量的增加提高了土壤有机质分解速率(Adamczyk et al., 2019), 使得土壤有机碳氮储量随森林更新演替呈显著下降趋势, 主要因为土壤有机碳氮储量受到植物生长以及凋落物种类组成的影响(del Valle et al., 2020).本研究中, 原始冷杉林(CK)中SOC和TN含量显著高于3种天然次生林(p < 0.05), 而在3种天然次生林中, SOC含量表现为80-NSF > 70-NSF > 60-NSF和TN为70-NSF > 80-NSF > 60-NSF, 其中80-NSF和70-NSF差异不显著(p > 0.05), 一方面由于采伐迹地自然更新演替次生林的早期阶段, 土壤微生境的变化加速了土壤中累积的有机质分解速率, 以及阔叶树种中的碳和木质素多低于针叶树种且分解速率较快(谌贤等, 2017); 另一方面天然次生林演替至60年, 外来物种进入难度增加致使物种多样性出现降低趋势(图3), 导致DON含量减少而影响到土壤SOC和TN含量.综上, 在川西亚高山高海拔特殊生境条件及其生态脆弱环境背景下, 如森林遭受破坏后, 其土壤肥力难以恢复.本研究仅对60年内不同演替阶段的次生林进行了观察, 对土壤碳氮动态变化机制的理解有限, 但研究结果显示, 如果要恢复到采伐前原始林的土壤肥力水平(表3)需要更长的时间. ...
... 土壤活性有机碳氮是土壤可溶性有机质的主要组分, 主要来源于新近枯枝落叶、腐殖质、根系分泌物以及微生物残体(Kalbitz et al., 2000; 鲍勇等, 2018).森林不同自然更新恢复阶段, 其林分土壤肥力难以恢复及物种组成的差异, 导致地表凋落物质量和数量、土壤温湿度、微生物组成等发生不同程度变化, 从而影响到土壤活性有机碳氮含量的变化(张雪等, 2016; Hagedorn et al., 2019).本研究中, DOC、LFOC和DON含量均随次生林演替呈显著下降趋势(p < 0.05), 且在70-NSF和80-NSF中差异不显著(表3), 而演替到岷江冷杉原始林阶段, 其土壤活性碳氮含量显著增加(p < 0.05), 这与该区域人工林演替过程中表层(0-20 cm)土壤活性碳氮含量的研究结果相似(姜发艳等, 2009), 但不同于热带(张哲等, 2019)和亚热带(范跃新等, 2013)森林演替的研究结果, 说明不同温湿条件及植被林分结构对土壤活性有机碳氮组分含量的影响表现出不同的变化特征(Li et al., 2014; Taylor et al., 2019).究其原因, 一方面随着森林更新演替, 土壤碳氮矿化速率增加(Mylliemngap et al., 2016), 且阔叶林的矿化速率高于针叶林(Côté et al., 2000); 另一方面由于表层土壤密度随演替增加, 而土壤团聚度、结构系数、总孔隙及毛管孔隙等土壤物理性质则呈现降低趋势(庞学勇等, 2003), 土壤有机碳、氮缺少土壤团聚体的物理性保护, 导致土壤活性有机碳和DON含量降低.此外, 岷江冷杉原始林的土壤活性碳氮组分含量高于天然次生林(表3), 可能与该林分主要物种构成(针叶+阔叶树种)有一定关联, 源于该区域针阔树种凋落叶融合在一起易于分解(黄玉梅等, 2015), 其释放的养分便于微生物分解利用, 导致土壤活性有机碳含量增加.如要明晰上述影响因素对土壤活性有机碳氮的驱动机理, 仍需进一步深入研究.相关分析表明, 不同天然更新演替阶段林地表层土壤SOC和TN含量与活性有机碳各组分含量及DON含量均呈极显著正相关关系(p < 0.01), 说明土壤活性有机碳和氮含量与土壤SOC、TN含量之间关系极为密切, 这与已有研究(辜翔等, 2016; 宫立等, 2017) 一致, 说明天然次生林土壤氮储量是调控其碳固持的重要参数(Li et al., 2012; Smal et al., 2019). ...
中亚热带植被恢复对土壤有机碳含量、碳密度的影响
1
2018
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
中亚热带植被恢复对土壤有机碳含量、碳密度的影响
1
2018
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
中亚热带4种森林类型土壤活性有机碳的季节动态特征
1
2016
... 森林更新演替的恢复过程中, 林分结构、物种组成、凋落物量以及林下微气候发生了不同程度的变化, 导致其林下土壤碳氮源不同而引起土壤有机质分解速率、土壤微生物多样性及其代谢活动发生变化(Wardle et al., 2004; Lucas-Borja & Delgado- Baquerizo, 2019), 进而影响到土壤酶活性(Adamczyk et al., 2015; Xu et al., 2017).此外, 年平均气温、降水量、土壤酸碱性和物种多样性也是影响土壤酶活性的主要因素(于洋等, 2014; Zhou et al., 2020).本研究中, 天然次生林更新早期演替(<60 a)不同阶段的表层土壤水解酶(βG、CBH和NAG)活性显著低于岷江冷杉原始林(表4), 且随自然更新演替进程呈显著降低趋势(80-NSF > 70-NSF > 60-NSF), 不同于川西亚高山平武县白桦林水解酶活性高于冷杉林(熊浩仲等, 2004)及温带白桦(Betula platyphylla)林随演替进程表现为先增加后降低的变化趋势(胡嵩等, 2013), 一方面说明岷江冷杉原始林的土壤养分循环速率相对较高, 对土壤养分供给更快; 另一方面说明次生林自然更新演替过程中, 地上凋落物增加导致土壤碳氮及其活性组分含量的下降(Liu et al., 2017; Lyu et al., 2019), 使其土壤微生物活性降低(辜翔等, 2018), 进而导致土壤水解酶活性下降.此外, 80-NSF、70-NSF和60-NSF土壤中氧化酶活性变化不显著, 但PHO的活性显著低于CK, 而PEO的活性与CK差异不显著(表4), 其原因可能是岷江冷杉原始林中含有较为丰富的木质素、角质和蜡质等难降解化合物, 促进了降解木质素的微生物群落多样性或种类数量增加, 且随次生林更新演替阶段的进行, 根系随着林上植被的演替而发生变化, 导致土壤的pH增加, 致使PHO活性增加(Fujii et al., 2013). ...
中亚热带4种森林类型土壤活性有机碳的季节动态特征
1
2016
... 森林更新演替的恢复过程中, 林分结构、物种组成、凋落物量以及林下微气候发生了不同程度的变化, 导致其林下土壤碳氮源不同而引起土壤有机质分解速率、土壤微生物多样性及其代谢活动发生变化(Wardle et al., 2004; Lucas-Borja & Delgado- Baquerizo, 2019), 进而影响到土壤酶活性(Adamczyk et al., 2015; Xu et al., 2017).此外, 年平均气温、降水量、土壤酸碱性和物种多样性也是影响土壤酶活性的主要因素(于洋等, 2014; Zhou et al., 2020).本研究中, 天然次生林更新早期演替(<60 a)不同阶段的表层土壤水解酶(βG、CBH和NAG)活性显著低于岷江冷杉原始林(表4), 且随自然更新演替进程呈显著降低趋势(80-NSF > 70-NSF > 60-NSF), 不同于川西亚高山平武县白桦林水解酶活性高于冷杉林(熊浩仲等, 2004)及温带白桦(Betula platyphylla)林随演替进程表现为先增加后降低的变化趋势(胡嵩等, 2013), 一方面说明岷江冷杉原始林的土壤养分循环速率相对较高, 对土壤养分供给更快; 另一方面说明次生林自然更新演替过程中, 地上凋落物增加导致土壤碳氮及其活性组分含量的下降(Liu et al., 2017; Lyu et al., 2019), 使其土壤微生物活性降低(辜翔等, 2018), 进而导致土壤水解酶活性下降.此外, 80-NSF、70-NSF和60-NSF土壤中氧化酶活性变化不显著, 但PHO的活性显著低于CK, 而PEO的活性与CK差异不显著(表4), 其原因可能是岷江冷杉原始林中含有较为丰富的木质素、角质和蜡质等难降解化合物, 促进了降解木质素的微生物群落多样性或种类数量增加, 且随次生林更新演替阶段的进行, 根系随着林上植被的演替而发生变化, 导致土壤的pH增加, 致使PHO活性增加(Fujii et al., 2013). ...
Above- and belowground linkages shape responses of mountain vegetation to climate change
2019
长白山原始红松林次生演替过程中土壤微生物生物量和酶活性变化
1
2013
... 土壤活性有机碳氮是土壤可溶性有机质的主要组分, 主要来源于新近枯枝落叶、腐殖质、根系分泌物以及微生物残体(Kalbitz et al., 2000; 鲍勇等, 2018).森林不同自然更新恢复阶段, 其林分土壤肥力难以恢复及物种组成的差异, 导致地表凋落物质量和数量、土壤温湿度、微生物组成等发生不同程度变化, 从而影响到土壤活性有机碳氮含量的变化(张雪等, 2016; Hagedorn et al., 2019).本研究中, DOC、LFOC和DON含量均随次生林演替呈显著下降趋势(p < 0.05), 且在70-NSF和80-NSF中差异不显著(表3), 而演替到岷江冷杉原始林阶段, 其土壤活性碳氮含量显著增加(p < 0.05), 这与该区域人工林演替过程中表层(0-20 cm)土壤活性碳氮含量的研究结果相似(姜发艳等, 2009), 但不同于热带(张哲等, 2019)和亚热带(范跃新等, 2013)森林演替的研究结果, 说明不同温湿条件及植被林分结构对土壤活性有机碳氮组分含量的影响表现出不同的变化特征(Li et al., 2014; Taylor et al., 2019).究其原因, 一方面随着森林更新演替, 土壤碳氮矿化速率增加(Mylliemngap et al., 2016), 且阔叶林的矿化速率高于针叶林(Côté et al., 2000); 另一方面由于表层土壤密度随演替增加, 而土壤团聚度、结构系数、总孔隙及毛管孔隙等土壤物理性质则呈现降低趋势(庞学勇等, 2003), 土壤有机碳、氮缺少土壤团聚体的物理性保护, 导致土壤活性有机碳和DON含量降低.此外, 岷江冷杉原始林的土壤活性碳氮组分含量高于天然次生林(表3), 可能与该林分主要物种构成(针叶+阔叶树种)有一定关联, 源于该区域针阔树种凋落叶融合在一起易于分解(黄玉梅等, 2015), 其释放的养分便于微生物分解利用, 导致土壤活性有机碳含量增加.如要明晰上述影响因素对土壤活性有机碳氮的驱动机理, 仍需进一步深入研究.相关分析表明, 不同天然更新演替阶段林地表层土壤SOC和TN含量与活性有机碳各组分含量及DON含量均呈极显著正相关关系(p < 0.01), 说明土壤活性有机碳和氮含量与土壤SOC、TN含量之间关系极为密切, 这与已有研究(辜翔等, 2016; 宫立等, 2017) 一致, 说明天然次生林土壤氮储量是调控其碳固持的重要参数(Li et al., 2012; Smal et al., 2019). ...
长白山原始红松林次生演替过程中土壤微生物生物量和酶活性变化
1
2013
... 土壤活性有机碳氮是土壤可溶性有机质的主要组分, 主要来源于新近枯枝落叶、腐殖质、根系分泌物以及微生物残体(Kalbitz et al., 2000; 鲍勇等, 2018).森林不同自然更新恢复阶段, 其林分土壤肥力难以恢复及物种组成的差异, 导致地表凋落物质量和数量、土壤温湿度、微生物组成等发生不同程度变化, 从而影响到土壤活性有机碳氮含量的变化(张雪等, 2016; Hagedorn et al., 2019).本研究中, DOC、LFOC和DON含量均随次生林演替呈显著下降趋势(p < 0.05), 且在70-NSF和80-NSF中差异不显著(表3), 而演替到岷江冷杉原始林阶段, 其土壤活性碳氮含量显著增加(p < 0.05), 这与该区域人工林演替过程中表层(0-20 cm)土壤活性碳氮含量的研究结果相似(姜发艳等, 2009), 但不同于热带(张哲等, 2019)和亚热带(范跃新等, 2013)森林演替的研究结果, 说明不同温湿条件及植被林分结构对土壤活性有机碳氮组分含量的影响表现出不同的变化特征(Li et al., 2014; Taylor et al., 2019).究其原因, 一方面随着森林更新演替, 土壤碳氮矿化速率增加(Mylliemngap et al., 2016), 且阔叶林的矿化速率高于针叶林(Côté et al., 2000); 另一方面由于表层土壤密度随演替增加, 而土壤团聚度、结构系数、总孔隙及毛管孔隙等土壤物理性质则呈现降低趋势(庞学勇等, 2003), 土壤有机碳、氮缺少土壤团聚体的物理性保护, 导致土壤活性有机碳和DON含量降低.此外, 岷江冷杉原始林的土壤活性碳氮组分含量高于天然次生林(表3), 可能与该林分主要物种构成(针叶+阔叶树种)有一定关联, 源于该区域针阔树种凋落叶融合在一起易于分解(黄玉梅等, 2015), 其释放的养分便于微生物分解利用, 导致土壤活性有机碳含量增加.如要明晰上述影响因素对土壤活性有机碳氮的驱动机理, 仍需进一步深入研究.相关分析表明, 不同天然更新演替阶段林地表层土壤SOC和TN含量与活性有机碳各组分含量及DON含量均呈极显著正相关关系(p < 0.01), 说明土壤活性有机碳和氮含量与土壤SOC、TN含量之间关系极为密切, 这与已有研究(辜翔等, 2016; 宫立等, 2017) 一致, 说明天然次生林土壤氮储量是调控其碳固持的重要参数(Li et al., 2012; Smal et al., 2019). ...
川西亚高山云杉叶凋落物质量损失过程及土壤生物的作用
2015
川西亚高山云杉叶凋落物质量损失过程及土壤生物的作用
2015
川西亚高山云杉人工林恢复过程中表层土壤碳动态变化
1
2009
... 森林更新演替的恢复过程中, 林分结构、物种组成、凋落物量以及林下微气候发生了不同程度的变化, 导致其林下土壤碳氮源不同而引起土壤有机质分解速率、土壤微生物多样性及其代谢活动发生变化(Wardle et al., 2004; Lucas-Borja & Delgado- Baquerizo, 2019), 进而影响到土壤酶活性(Adamczyk et al., 2015; Xu et al., 2017).此外, 年平均气温、降水量、土壤酸碱性和物种多样性也是影响土壤酶活性的主要因素(于洋等, 2014; Zhou et al., 2020).本研究中, 天然次生林更新早期演替(<60 a)不同阶段的表层土壤水解酶(βG、CBH和NAG)活性显著低于岷江冷杉原始林(表4), 且随自然更新演替进程呈显著降低趋势(80-NSF > 70-NSF > 60-NSF), 不同于川西亚高山平武县白桦林水解酶活性高于冷杉林(熊浩仲等, 2004)及温带白桦(Betula platyphylla)林随演替进程表现为先增加后降低的变化趋势(胡嵩等, 2013), 一方面说明岷江冷杉原始林的土壤养分循环速率相对较高, 对土壤养分供给更快; 另一方面说明次生林自然更新演替过程中, 地上凋落物增加导致土壤碳氮及其活性组分含量的下降(Liu et al., 2017; Lyu et al., 2019), 使其土壤微生物活性降低(辜翔等, 2018), 进而导致土壤水解酶活性下降.此外, 80-NSF、70-NSF和60-NSF土壤中氧化酶活性变化不显著, 但PHO的活性显著低于CK, 而PEO的活性与CK差异不显著(表4), 其原因可能是岷江冷杉原始林中含有较为丰富的木质素、角质和蜡质等难降解化合物, 促进了降解木质素的微生物群落多样性或种类数量增加, 且随次生林更新演替阶段的进行, 根系随着林上植被的演替而发生变化, 导致土壤的pH增加, 致使PHO活性增加(Fujii et al., 2013). ...
川西亚高山云杉人工林恢复过程中表层土壤碳动态变化
1
2009
... 森林更新演替的恢复过程中, 林分结构、物种组成、凋落物量以及林下微气候发生了不同程度的变化, 导致其林下土壤碳氮源不同而引起土壤有机质分解速率、土壤微生物多样性及其代谢活动发生变化(Wardle et al., 2004; Lucas-Borja & Delgado- Baquerizo, 2019), 进而影响到土壤酶活性(Adamczyk et al., 2015; Xu et al., 2017).此外, 年平均气温、降水量、土壤酸碱性和物种多样性也是影响土壤酶活性的主要因素(于洋等, 2014; Zhou et al., 2020).本研究中, 天然次生林更新早期演替(<60 a)不同阶段的表层土壤水解酶(βG、CBH和NAG)活性显著低于岷江冷杉原始林(表4), 且随自然更新演替进程呈显著降低趋势(80-NSF > 70-NSF > 60-NSF), 不同于川西亚高山平武县白桦林水解酶活性高于冷杉林(熊浩仲等, 2004)及温带白桦(Betula platyphylla)林随演替进程表现为先增加后降低的变化趋势(胡嵩等, 2013), 一方面说明岷江冷杉原始林的土壤养分循环速率相对较高, 对土壤养分供给更快; 另一方面说明次生林自然更新演替过程中, 地上凋落物增加导致土壤碳氮及其活性组分含量的下降(Liu et al., 2017; Lyu et al., 2019), 使其土壤微生物活性降低(辜翔等, 2018), 进而导致土壤水解酶活性下降.此外, 80-NSF、70-NSF和60-NSF土壤中氧化酶活性变化不显著, 但PHO的活性显著低于CK, 而PEO的活性与CK差异不显著(表4), 其原因可能是岷江冷杉原始林中含有较为丰富的木质素、角质和蜡质等难降解化合物, 促进了降解木质素的微生物群落多样性或种类数量增加, 且随次生林更新演替阶段的进行, 根系随着林上植被的演替而发生变化, 导致土壤的pH增加, 致使PHO活性增加(Fujii et al., 2013). ...
Controls on the dynamics of dissolved organic matter in soils: a review
2000
Variations of soil enzyme activities in a temperate forest soil
1
2009
... 土壤活性有机碳氮是土壤可溶性有机质的主要组分, 主要来源于新近枯枝落叶、腐殖质、根系分泌物以及微生物残体(Kalbitz et al., 2000; 鲍勇等, 2018).森林不同自然更新恢复阶段, 其林分土壤肥力难以恢复及物种组成的差异, 导致地表凋落物质量和数量、土壤温湿度、微生物组成等发生不同程度变化, 从而影响到土壤活性有机碳氮含量的变化(张雪等, 2016; Hagedorn et al., 2019).本研究中, DOC、LFOC和DON含量均随次生林演替呈显著下降趋势(p < 0.05), 且在70-NSF和80-NSF中差异不显著(表3), 而演替到岷江冷杉原始林阶段, 其土壤活性碳氮含量显著增加(p < 0.05), 这与该区域人工林演替过程中表层(0-20 cm)土壤活性碳氮含量的研究结果相似(姜发艳等, 2009), 但不同于热带(张哲等, 2019)和亚热带(范跃新等, 2013)森林演替的研究结果, 说明不同温湿条件及植被林分结构对土壤活性有机碳氮组分含量的影响表现出不同的变化特征(Li et al., 2014; Taylor et al., 2019).究其原因, 一方面随着森林更新演替, 土壤碳氮矿化速率增加(Mylliemngap et al., 2016), 且阔叶林的矿化速率高于针叶林(Côté et al., 2000); 另一方面由于表层土壤密度随演替增加, 而土壤团聚度、结构系数、总孔隙及毛管孔隙等土壤物理性质则呈现降低趋势(庞学勇等, 2003), 土壤有机碳、氮缺少土壤团聚体的物理性保护, 导致土壤活性有机碳和DON含量降低.此外, 岷江冷杉原始林的土壤活性碳氮组分含量高于天然次生林(表3), 可能与该林分主要物种构成(针叶+阔叶树种)有一定关联, 源于该区域针阔树种凋落叶融合在一起易于分解(黄玉梅等, 2015), 其释放的养分便于微生物分解利用, 导致土壤活性有机碳含量增加.如要明晰上述影响因素对土壤活性有机碳氮的驱动机理, 仍需进一步深入研究.相关分析表明, 不同天然更新演替阶段林地表层土壤SOC和TN含量与活性有机碳各组分含量及DON含量均呈极显著正相关关系(p < 0.01), 说明土壤活性有机碳和氮含量与土壤SOC、TN含量之间关系极为密切, 这与已有研究(辜翔等, 2016; 宫立等, 2017) 一致, 说明天然次生林土壤氮储量是调控其碳固持的重要参数(Li et al., 2012; Smal et al., 2019). ...
Returning forests analyzed with the forest identity
1
2006
... 土壤活性有机碳氮是土壤可溶性有机质的主要组分, 主要来源于新近枯枝落叶、腐殖质、根系分泌物以及微生物残体(Kalbitz et al., 2000; 鲍勇等, 2018).森林不同自然更新恢复阶段, 其林分土壤肥力难以恢复及物种组成的差异, 导致地表凋落物质量和数量、土壤温湿度、微生物组成等发生不同程度变化, 从而影响到土壤活性有机碳氮含量的变化(张雪等, 2016; Hagedorn et al., 2019).本研究中, DOC、LFOC和DON含量均随次生林演替呈显著下降趋势(p < 0.05), 且在70-NSF和80-NSF中差异不显著(表3), 而演替到岷江冷杉原始林阶段, 其土壤活性碳氮含量显著增加(p < 0.05), 这与该区域人工林演替过程中表层(0-20 cm)土壤活性碳氮含量的研究结果相似(姜发艳等, 2009), 但不同于热带(张哲等, 2019)和亚热带(范跃新等, 2013)森林演替的研究结果, 说明不同温湿条件及植被林分结构对土壤活性有机碳氮组分含量的影响表现出不同的变化特征(Li et al., 2014; Taylor et al., 2019).究其原因, 一方面随着森林更新演替, 土壤碳氮矿化速率增加(Mylliemngap et al., 2016), 且阔叶林的矿化速率高于针叶林(Côté et al., 2000); 另一方面由于表层土壤密度随演替增加, 而土壤团聚度、结构系数、总孔隙及毛管孔隙等土壤物理性质则呈现降低趋势(庞学勇等, 2003), 土壤有机碳、氮缺少土壤团聚体的物理性保护, 导致土壤活性有机碳和DON含量降低.此外, 岷江冷杉原始林的土壤活性碳氮组分含量高于天然次生林(表3), 可能与该林分主要物种构成(针叶+阔叶树种)有一定关联, 源于该区域针阔树种凋落叶融合在一起易于分解(黄玉梅等, 2015), 其释放的养分便于微生物分解利用, 导致土壤活性有机碳含量增加.如要明晰上述影响因素对土壤活性有机碳氮的驱动机理, 仍需进一步深入研究.相关分析表明, 不同天然更新演替阶段林地表层土壤SOC和TN含量与活性有机碳各组分含量及DON含量均呈极显著正相关关系(p < 0.01), 说明土壤活性有机碳和氮含量与土壤SOC、TN含量之间关系极为密切, 这与已有研究(辜翔等, 2016; 宫立等, 2017) 一致, 说明天然次生林土壤氮储量是调控其碳固持的重要参数(Li et al., 2012; Smal et al., 2019). ...
Increasing tree diversity enhances microbial and enzyme activities in temperate Iranian forests
2018
Global patterns of the dynamics of soil carbon and nitrogen stocks following afforestation: a meta-analysis
1
2012
... 森林更新演替的恢复过程中, 林分结构、物种组成、凋落物量以及林下微气候发生了不同程度的变化, 导致其林下土壤碳氮源不同而引起土壤有机质分解速率、土壤微生物多样性及其代谢活动发生变化(Wardle et al., 2004; Lucas-Borja & Delgado- Baquerizo, 2019), 进而影响到土壤酶活性(Adamczyk et al., 2015; Xu et al., 2017).此外, 年平均气温、降水量、土壤酸碱性和物种多样性也是影响土壤酶活性的主要因素(于洋等, 2014; Zhou et al., 2020).本研究中, 天然次生林更新早期演替(<60 a)不同阶段的表层土壤水解酶(βG、CBH和NAG)活性显著低于岷江冷杉原始林(表4), 且随自然更新演替进程呈显著降低趋势(80-NSF > 70-NSF > 60-NSF), 不同于川西亚高山平武县白桦林水解酶活性高于冷杉林(熊浩仲等, 2004)及温带白桦(Betula platyphylla)林随演替进程表现为先增加后降低的变化趋势(胡嵩等, 2013), 一方面说明岷江冷杉原始林的土壤养分循环速率相对较高, 对土壤养分供给更快; 另一方面说明次生林自然更新演替过程中, 地上凋落物增加导致土壤碳氮及其活性组分含量的下降(Liu et al., 2017; Lyu et al., 2019), 使其土壤微生物活性降低(辜翔等, 2018), 进而导致土壤水解酶活性下降.此外, 80-NSF、70-NSF和60-NSF土壤中氧化酶活性变化不显著, 但PHO的活性显著低于CK, 而PEO的活性与CK差异不显著(表4), 其原因可能是岷江冷杉原始林中含有较为丰富的木质素、角质和蜡质等难降解化合物, 促进了降解木质素的微生物群落多样性或种类数量增加, 且随次生林更新演替阶段的进行, 根系随着林上植被的演替而发生变化, 导致土壤的pH增加, 致使PHO活性增加(Fujii et al., 2013). ...
Effects of regenerating vegetation on soil enzyme activity and microbial structure in reclaimed soils on a surface coal mine site
2015
Soil organic carbon fractions and microbial community and functions under changes in vegetation: a case of vegetation succession in karst forest
1
2014
... 土壤活性有机碳氮是土壤可溶性有机质的主要组分, 主要来源于新近枯枝落叶、腐殖质、根系分泌物以及微生物残体(Kalbitz et al., 2000; 鲍勇等, 2018).森林不同自然更新恢复阶段, 其林分土壤肥力难以恢复及物种组成的差异, 导致地表凋落物质量和数量、土壤温湿度、微生物组成等发生不同程度变化, 从而影响到土壤活性有机碳氮含量的变化(张雪等, 2016; Hagedorn et al., 2019).本研究中, DOC、LFOC和DON含量均随次生林演替呈显著下降趋势(p < 0.05), 且在70-NSF和80-NSF中差异不显著(表3), 而演替到岷江冷杉原始林阶段, 其土壤活性碳氮含量显著增加(p < 0.05), 这与该区域人工林演替过程中表层(0-20 cm)土壤活性碳氮含量的研究结果相似(姜发艳等, 2009), 但不同于热带(张哲等, 2019)和亚热带(范跃新等, 2013)森林演替的研究结果, 说明不同温湿条件及植被林分结构对土壤活性有机碳氮组分含量的影响表现出不同的变化特征(Li et al., 2014; Taylor et al., 2019).究其原因, 一方面随着森林更新演替, 土壤碳氮矿化速率增加(Mylliemngap et al., 2016), 且阔叶林的矿化速率高于针叶林(Côté et al., 2000); 另一方面由于表层土壤密度随演替增加, 而土壤团聚度、结构系数、总孔隙及毛管孔隙等土壤物理性质则呈现降低趋势(庞学勇等, 2003), 土壤有机碳、氮缺少土壤团聚体的物理性保护, 导致土壤活性有机碳和DON含量降低.此外, 岷江冷杉原始林的土壤活性碳氮组分含量高于天然次生林(表3), 可能与该林分主要物种构成(针叶+阔叶树种)有一定关联, 源于该区域针阔树种凋落叶融合在一起易于分解(黄玉梅等, 2015), 其释放的养分便于微生物分解利用, 导致土壤活性有机碳含量增加.如要明晰上述影响因素对土壤活性有机碳氮的驱动机理, 仍需进一步深入研究.相关分析表明, 不同天然更新演替阶段林地表层土壤SOC和TN含量与活性有机碳各组分含量及DON含量均呈极显著正相关关系(p < 0.01), 说明土壤活性有机碳和氮含量与土壤SOC、TN含量之间关系极为密切, 这与已有研究(辜翔等, 2016; 宫立等, 2017) 一致, 说明天然次生林土壤氮储量是调控其碳固持的重要参数(Li et al., 2012; Smal et al., 2019). ...
子午岭植被自然恢复过程中植物多样性的变化
2004
子午岭植被自然恢复过程中植物多样性的变化
2004
Increased litter in subtropical forests boosts soil respiration in natural forests but not plantations of
2
2017
... 土壤酶活性参照DeForest (2009)和Chen等(2017)的方法: 取新鲜土壤1 g, 加蒸馏水125 mL, 振荡2 h (25 ℃, 180 r·min-1), 吸取1 mL的悬浊液, 用4-甲基伞形酮做标准曲线, 用4-MUB-β-D-乙酰基氨基葡萄糖苷和4-MUB-β-D-葡萄糖苷为底物标示酶活性, 置于25 ℃下避光培养后, 用多功能酶标仪(M200PRO, TECAN, 广州)测定β-4-N-乙酰基氨基葡萄糖苷酶和β-4-葡萄糖苷酶; 过氧化物酶和多酚氧化酶采用酶联免疫微量法: 分别取过50目的风干土样0.200 g, 加入底物(邻苯三酚) 120 mL和100 μL 1%邻苯三酚溶液和20 μL过氧化氢溶液, 混匀后置于30 ℃恒温培育后, 用紫色没食子素作为标准曲线, 分别加入50 mL和50 μL PBS及430 mL乙醚, 振荡并静置后, 用多功能酶标仪测定多酚氧化酶和过氧化物酶; 纤维素水解酶采用3,5-二硝基水杨酸比色法测定: 取10 g土壤, 加入1.5 mL甲苯, 摇匀放置15 min后, 用葡萄糖作出标准曲线, 加5 mL 1%羧甲基纤维素溶液和5 mL pH为5.5的醋酸盐缓冲液, 37 ℃恒温箱培养后, 过滤用岛津-UV2450-紫外分光光度计进行比色测定.5种土壤酶的名称、缩写、功能和标定底物见表2.酶活性均通过预实验确定获得最大酶活性所需要的底物浓度和培养时间. ...
... 土壤活性有机碳氮是土壤可溶性有机质的主要组分, 主要来源于新近枯枝落叶、腐殖质、根系分泌物以及微生物残体(Kalbitz et al., 2000; 鲍勇等, 2018).森林不同自然更新恢复阶段, 其林分土壤肥力难以恢复及物种组成的差异, 导致地表凋落物质量和数量、土壤温湿度、微生物组成等发生不同程度变化, 从而影响到土壤活性有机碳氮含量的变化(张雪等, 2016; Hagedorn et al., 2019).本研究中, DOC、LFOC和DON含量均随次生林演替呈显著下降趋势(p < 0.05), 且在70-NSF和80-NSF中差异不显著(表3), 而演替到岷江冷杉原始林阶段, 其土壤活性碳氮含量显著增加(p < 0.05), 这与该区域人工林演替过程中表层(0-20 cm)土壤活性碳氮含量的研究结果相似(姜发艳等, 2009), 但不同于热带(张哲等, 2019)和亚热带(范跃新等, 2013)森林演替的研究结果, 说明不同温湿条件及植被林分结构对土壤活性有机碳氮组分含量的影响表现出不同的变化特征(Li et al., 2014; Taylor et al., 2019).究其原因, 一方面随着森林更新演替, 土壤碳氮矿化速率增加(Mylliemngap et al., 2016), 且阔叶林的矿化速率高于针叶林(Côté et al., 2000); 另一方面由于表层土壤密度随演替增加, 而土壤团聚度、结构系数、总孔隙及毛管孔隙等土壤物理性质则呈现降低趋势(庞学勇等, 2003), 土壤有机碳、氮缺少土壤团聚体的物理性保护, 导致土壤活性有机碳和DON含量降低.此外, 岷江冷杉原始林的土壤活性碳氮组分含量高于天然次生林(表3), 可能与该林分主要物种构成(针叶+阔叶树种)有一定关联, 源于该区域针阔树种凋落叶融合在一起易于分解(黄玉梅等, 2015), 其释放的养分便于微生物分解利用, 导致土壤活性有机碳含量增加.如要明晰上述影响因素对土壤活性有机碳氮的驱动机理, 仍需进一步深入研究.相关分析表明, 不同天然更新演替阶段林地表层土壤SOC和TN含量与活性有机碳各组分含量及DON含量均呈极显著正相关关系(p < 0.01), 说明土壤活性有机碳和氮含量与土壤SOC、TN含量之间关系极为密切, 这与已有研究(辜翔等, 2016; 宫立等, 2017) 一致, 说明天然次生林土壤氮储量是调控其碳固持的重要参数(Li et al., 2012; Smal et al., 2019). ...
Long-term forest succession improves plant diversity and soil quality but not significantly increase soil microbial diversity: evidence from the Loess Plateau
1
2020
... 土壤活性有机碳氮是土壤可溶性有机质的主要组分, 主要来源于新近枯枝落叶、腐殖质、根系分泌物以及微生物残体(Kalbitz et al., 2000; 鲍勇等, 2018).森林不同自然更新恢复阶段, 其林分土壤肥力难以恢复及物种组成的差异, 导致地表凋落物质量和数量、土壤温湿度、微生物组成等发生不同程度变化, 从而影响到土壤活性有机碳氮含量的变化(张雪等, 2016; Hagedorn et al., 2019).本研究中, DOC、LFOC和DON含量均随次生林演替呈显著下降趋势(p < 0.05), 且在70-NSF和80-NSF中差异不显著(表3), 而演替到岷江冷杉原始林阶段, 其土壤活性碳氮含量显著增加(p < 0.05), 这与该区域人工林演替过程中表层(0-20 cm)土壤活性碳氮含量的研究结果相似(姜发艳等, 2009), 但不同于热带(张哲等, 2019)和亚热带(范跃新等, 2013)森林演替的研究结果, 说明不同温湿条件及植被林分结构对土壤活性有机碳氮组分含量的影响表现出不同的变化特征(Li et al., 2014; Taylor et al., 2019).究其原因, 一方面随着森林更新演替, 土壤碳氮矿化速率增加(Mylliemngap et al., 2016), 且阔叶林的矿化速率高于针叶林(Côté et al., 2000); 另一方面由于表层土壤密度随演替增加, 而土壤团聚度、结构系数、总孔隙及毛管孔隙等土壤物理性质则呈现降低趋势(庞学勇等, 2003), 土壤有机碳、氮缺少土壤团聚体的物理性保护, 导致土壤活性有机碳和DON含量降低.此外, 岷江冷杉原始林的土壤活性碳氮组分含量高于天然次生林(表3), 可能与该林分主要物种构成(针叶+阔叶树种)有一定关联, 源于该区域针阔树种凋落叶融合在一起易于分解(黄玉梅等, 2015), 其释放的养分便于微生物分解利用, 导致土壤活性有机碳含量增加.如要明晰上述影响因素对土壤活性有机碳氮的驱动机理, 仍需进一步深入研究.相关分析表明, 不同天然更新演替阶段林地表层土壤SOC和TN含量与活性有机碳各组分含量及DON含量均呈极显著正相关关系(p < 0.01), 说明土壤活性有机碳和氮含量与土壤SOC、TN含量之间关系极为密切, 这与已有研究(辜翔等, 2016; 宫立等, 2017) 一致, 说明天然次生林土壤氮储量是调控其碳固持的重要参数(Li et al., 2012; Smal et al., 2019). ...
2
2002
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
... 土壤基本理化性质测定参照《土壤农业化学分析方法》(鲁如坤, 2002).土壤含水率用烘干法 (105 ℃烘干24 h)测定; 土壤pH按土水质量比1:2.5混合搅拌后用pH计测定; SOC含量测定采用重铬酸钾外加热法; TN含量测定采用硫酸-催化剂消解-流动分析仪(AA3-连续流动分析仪)(Auto Analyzer 3, SEAL, 广州)法. ...
2
2002
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
... 土壤基本理化性质测定参照《土壤农业化学分析方法》(鲁如坤, 2002).土壤含水率用烘干法 (105 ℃烘干24 h)测定; 土壤pH按土水质量比1:2.5混合搅拌后用pH计测定; SOC含量测定采用重铬酸钾外加热法; TN含量测定采用硫酸-催化剂消解-流动分析仪(AA3-连续流动分析仪)(Auto Analyzer 3, SEAL, 广州)法. ...
Plant diversity and soil stoichiometry regulates the changes in multifunctionality during pine temperate forest secondary succession
1
2019
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
Effects of forest conversion on carbon-degrading enzyme activities in subtropical China
1
2019
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
Root-microbial interaction accelerates soil nitrogen depletion but not soil carbon after increasing litter inputs to a coniferous forest
1
2019
... 土壤活性有机碳氮是土壤可溶性有机质的主要组分, 主要来源于新近枯枝落叶、腐殖质、根系分泌物以及微生物残体(Kalbitz et al., 2000; 鲍勇等, 2018).森林不同自然更新恢复阶段, 其林分土壤肥力难以恢复及物种组成的差异, 导致地表凋落物质量和数量、土壤温湿度、微生物组成等发生不同程度变化, 从而影响到土壤活性有机碳氮含量的变化(张雪等, 2016; Hagedorn et al., 2019).本研究中, DOC、LFOC和DON含量均随次生林演替呈显著下降趋势(p < 0.05), 且在70-NSF和80-NSF中差异不显著(表3), 而演替到岷江冷杉原始林阶段, 其土壤活性碳氮含量显著增加(p < 0.05), 这与该区域人工林演替过程中表层(0-20 cm)土壤活性碳氮含量的研究结果相似(姜发艳等, 2009), 但不同于热带(张哲等, 2019)和亚热带(范跃新等, 2013)森林演替的研究结果, 说明不同温湿条件及植被林分结构对土壤活性有机碳氮组分含量的影响表现出不同的变化特征(Li et al., 2014; Taylor et al., 2019).究其原因, 一方面随着森林更新演替, 土壤碳氮矿化速率增加(Mylliemngap et al., 2016), 且阔叶林的矿化速率高于针叶林(Côté et al., 2000); 另一方面由于表层土壤密度随演替增加, 而土壤团聚度、结构系数、总孔隙及毛管孔隙等土壤物理性质则呈现降低趋势(庞学勇等, 2003), 土壤有机碳、氮缺少土壤团聚体的物理性保护, 导致土壤活性有机碳和DON含量降低.此外, 岷江冷杉原始林的土壤活性碳氮组分含量高于天然次生林(表3), 可能与该林分主要物种构成(针叶+阔叶树种)有一定关联, 源于该区域针阔树种凋落叶融合在一起易于分解(黄玉梅等, 2015), 其释放的养分便于微生物分解利用, 导致土壤活性有机碳含量增加.如要明晰上述影响因素对土壤活性有机碳氮的驱动机理, 仍需进一步深入研究.相关分析表明, 不同天然更新演替阶段林地表层土壤SOC和TN含量与活性有机碳各组分含量及DON含量均呈极显著正相关关系(p < 0.01), 说明土壤活性有机碳和氮含量与土壤SOC、TN含量之间关系极为密切, 这与已有研究(辜翔等, 2016; 宫立等, 2017) 一致, 说明天然次生林土壤氮储量是调控其碳固持的重要参数(Li et al., 2012; Smal et al., 2019). ...
不同水氮水平对川西亚高山林地土壤酶活性的影响
1
2019
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
不同水氮水平对川西亚高山林地土壤酶活性的影响
1
2019
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
Tamm Review: Influence of forest management activities on soil organic carbon stocks: a knowledge synthesis
1
2020
... 土壤活性有机碳氮是土壤可溶性有机质的主要组分, 主要来源于新近枯枝落叶、腐殖质、根系分泌物以及微生物残体(Kalbitz et al., 2000; 鲍勇等, 2018).森林不同自然更新恢复阶段, 其林分土壤肥力难以恢复及物种组成的差异, 导致地表凋落物质量和数量、土壤温湿度、微生物组成等发生不同程度变化, 从而影响到土壤活性有机碳氮含量的变化(张雪等, 2016; Hagedorn et al., 2019).本研究中, DOC、LFOC和DON含量均随次生林演替呈显著下降趋势(p < 0.05), 且在70-NSF和80-NSF中差异不显著(表3), 而演替到岷江冷杉原始林阶段, 其土壤活性碳氮含量显著增加(p < 0.05), 这与该区域人工林演替过程中表层(0-20 cm)土壤活性碳氮含量的研究结果相似(姜发艳等, 2009), 但不同于热带(张哲等, 2019)和亚热带(范跃新等, 2013)森林演替的研究结果, 说明不同温湿条件及植被林分结构对土壤活性有机碳氮组分含量的影响表现出不同的变化特征(Li et al., 2014; Taylor et al., 2019).究其原因, 一方面随着森林更新演替, 土壤碳氮矿化速率增加(Mylliemngap et al., 2016), 且阔叶林的矿化速率高于针叶林(Côté et al., 2000); 另一方面由于表层土壤密度随演替增加, 而土壤团聚度、结构系数、总孔隙及毛管孔隙等土壤物理性质则呈现降低趋势(庞学勇等, 2003), 土壤有机碳、氮缺少土壤团聚体的物理性保护, 导致土壤活性有机碳和DON含量降低.此外, 岷江冷杉原始林的土壤活性碳氮组分含量高于天然次生林(表3), 可能与该林分主要物种构成(针叶+阔叶树种)有一定关联, 源于该区域针阔树种凋落叶融合在一起易于分解(黄玉梅等, 2015), 其释放的养分便于微生物分解利用, 导致土壤活性有机碳含量增加.如要明晰上述影响因素对土壤活性有机碳氮的驱动机理, 仍需进一步深入研究.相关分析表明, 不同天然更新演替阶段林地表层土壤SOC和TN含量与活性有机碳各组分含量及DON含量均呈极显著正相关关系(p < 0.01), 说明土壤活性有机碳和氮含量与土壤SOC、TN含量之间关系极为密切, 这与已有研究(辜翔等, 2016; 宫立等, 2017) 一致, 说明天然次生林土壤氮储量是调控其碳固持的重要参数(Li et al., 2012; Smal et al., 2019). ...
Identification of general patterns of nutrient and labile carbon control on soil carbon dynamics across a successional gradient
2011
Changes in vegetation and nitrogen mineralization during recovery of a montane subtropical broadleaved forest in North-eastern India following anthropogenic disturbance
1
2016
... 统计分析利用SPSS 26.0软件对数据进行处理, 采用单因素方差分析和最小显著差异(LSD)法检验不同更新演替阶段天然次生林以及与原始冷杉林之间的土壤理化性质、活性有机碳组分含量和酶活性的差异显著性(水平为p < 0.05); 物种多样性选取Margalef丰富度指数(E)、Shannon-Wiener指数(H)、Simpson指数(D)和Pielou均匀度指数(J)等指标表示3种天然次生林及岷江冷杉原始林间的物种组成结构(李裕元和邵明安, 2004); 采用Pearson相关分析土壤酶活性与土壤养分指标及物种多样性的相关性.用Canoco Software 5.0软件, 以土壤酶活性为响应变量, 同时以土壤养分为解释变量做冗余分析(RDA).作图在Origin 14.0软件内完成. ...
Carbon cycling and net ecosystem production at an early stage of secondary succession in an abandoned coppice forest
1
2010
... 森林更新演替的恢复过程中, 林分结构、物种组成、凋落物量以及林下微气候发生了不同程度的变化, 导致其林下土壤碳氮源不同而引起土壤有机质分解速率、土壤微生物多样性及其代谢活动发生变化(Wardle et al., 2004; Lucas-Borja & Delgado- Baquerizo, 2019), 进而影响到土壤酶活性(Adamczyk et al., 2015; Xu et al., 2017).此外, 年平均气温、降水量、土壤酸碱性和物种多样性也是影响土壤酶活性的主要因素(于洋等, 2014; Zhou et al., 2020).本研究中, 天然次生林更新早期演替(<60 a)不同阶段的表层土壤水解酶(βG、CBH和NAG)活性显著低于岷江冷杉原始林(表4), 且随自然更新演替进程呈显著降低趋势(80-NSF > 70-NSF > 60-NSF), 不同于川西亚高山平武县白桦林水解酶活性高于冷杉林(熊浩仲等, 2004)及温带白桦(Betula platyphylla)林随演替进程表现为先增加后降低的变化趋势(胡嵩等, 2013), 一方面说明岷江冷杉原始林的土壤养分循环速率相对较高, 对土壤养分供给更快; 另一方面说明次生林自然更新演替过程中, 地上凋落物增加导致土壤碳氮及其活性组分含量的下降(Liu et al., 2017; Lyu et al., 2019), 使其土壤微生物活性降低(辜翔等, 2018), 进而导致土壤水解酶活性下降.此外, 80-NSF、70-NSF和60-NSF土壤中氧化酶活性变化不显著, 但PHO的活性显著低于CK, 而PEO的活性与CK差异不显著(表4), 其原因可能是岷江冷杉原始林中含有较为丰富的木质素、角质和蜡质等难降解化合物, 促进了降解木质素的微生物群落多样性或种类数量增加, 且随次生林更新演替阶段的进行, 根系随着林上植被的演替而发生变化, 导致土壤的pH增加, 致使PHO活性增加(Fujii et al., 2013). ...
Responses of soil labile organic carbon fractions and stocks to different vegetation restoration strategies in degraded karst ecosystems of southwest China
1
2019
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
川西亚高山针叶林植物群落演替对土壤性质的影响
2003
川西亚高山针叶林植物群落演替对土壤性质的影响
2003
The relation among soil microorganism, enzyme activity and soil nutrients under subalpine coniferous forest in Western Sichuan
2009
Soil carbon sequestration and land-use change: processes and potential
1
2000
... 森林更新演替的恢复过程中, 林分结构、物种组成、凋落物量以及林下微气候发生了不同程度的变化, 导致其林下土壤碳氮源不同而引起土壤有机质分解速率、土壤微生物多样性及其代谢活动发生变化(Wardle et al., 2004; Lucas-Borja & Delgado- Baquerizo, 2019), 进而影响到土壤酶活性(Adamczyk et al., 2015; Xu et al., 2017).此外, 年平均气温、降水量、土壤酸碱性和物种多样性也是影响土壤酶活性的主要因素(于洋等, 2014; Zhou et al., 2020).本研究中, 天然次生林更新早期演替(<60 a)不同阶段的表层土壤水解酶(βG、CBH和NAG)活性显著低于岷江冷杉原始林(表4), 且随自然更新演替进程呈显著降低趋势(80-NSF > 70-NSF > 60-NSF), 不同于川西亚高山平武县白桦林水解酶活性高于冷杉林(熊浩仲等, 2004)及温带白桦(Betula platyphylla)林随演替进程表现为先增加后降低的变化趋势(胡嵩等, 2013), 一方面说明岷江冷杉原始林的土壤养分循环速率相对较高, 对土壤养分供给更快; 另一方面说明次生林自然更新演替过程中, 地上凋落物增加导致土壤碳氮及其活性组分含量的下降(Liu et al., 2017; Lyu et al., 2019), 使其土壤微生物活性降低(辜翔等, 2018), 进而导致土壤水解酶活性下降.此外, 80-NSF、70-NSF和60-NSF土壤中氧化酶活性变化不显著, 但PHO的活性显著低于CK, 而PEO的活性与CK差异不显著(表4), 其原因可能是岷江冷杉原始林中含有较为丰富的木质素、角质和蜡质等难降解化合物, 促进了降解木质素的微生物群落多样性或种类数量增加, 且随次生林更新演替阶段的进行, 根系随着林上植被的演替而发生变化, 导致土壤的pH增加, 致使PHO活性增加(Fujii et al., 2013). ...
Chemical composition of soil organic matter and potential enzyme activity in the topsoil along a moisture gradient in the High Arctic (Svalbard)
1
2020
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
Priming alters soil carbon dynamics during forest succession
1
2019
... 森林更新演替的恢复过程中, 林分结构、物种组成、凋落物量以及林下微气候发生了不同程度的变化, 导致其林下土壤碳氮源不同而引起土壤有机质分解速率、土壤微生物多样性及其代谢活动发生变化(Wardle et al., 2004; Lucas-Borja & Delgado- Baquerizo, 2019), 进而影响到土壤酶活性(Adamczyk et al., 2015; Xu et al., 2017).此外, 年平均气温、降水量、土壤酸碱性和物种多样性也是影响土壤酶活性的主要因素(于洋等, 2014; Zhou et al., 2020).本研究中, 天然次生林更新早期演替(<60 a)不同阶段的表层土壤水解酶(βG、CBH和NAG)活性显著低于岷江冷杉原始林(表4), 且随自然更新演替进程呈显著降低趋势(80-NSF > 70-NSF > 60-NSF), 不同于川西亚高山平武县白桦林水解酶活性高于冷杉林(熊浩仲等, 2004)及温带白桦(Betula platyphylla)林随演替进程表现为先增加后降低的变化趋势(胡嵩等, 2013), 一方面说明岷江冷杉原始林的土壤养分循环速率相对较高, 对土壤养分供给更快; 另一方面说明次生林自然更新演替过程中, 地上凋落物增加导致土壤碳氮及其活性组分含量的下降(Liu et al., 2017; Lyu et al., 2019), 使其土壤微生物活性降低(辜翔等, 2018), 进而导致土壤水解酶活性下降.此外, 80-NSF、70-NSF和60-NSF土壤中氧化酶活性变化不显著, 但PHO的活性显著低于CK, 而PEO的活性与CK差异不显著(表4), 其原因可能是岷江冷杉原始林中含有较为丰富的木质素、角质和蜡质等难降解化合物, 促进了降解木质素的微生物群落多样性或种类数量增加, 且随次生林更新演替阶段的进行, 根系随着林上植被的演替而发生变化, 导致土壤的pH增加, 致使PHO活性增加(Fujii et al., 2013). ...
The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil
2002
Soil carbon dynamics in a chronosequence of secondary forests in northeastern Costa Rica
1
2008
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
川西亚高山森林凋落物不同分解阶段基质质量特征
1
2017
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
川西亚高山森林凋落物不同分解阶段基质质量特征
1
2017
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
Phenol oxidase, peroxidase and organic matter dynamics of soil
1
2010
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
Extracellular enzyme kinetics scale with resource availability
1
2014
... 土壤活性有机碳氮是土壤可溶性有机质的主要组分, 主要来源于新近枯枝落叶、腐殖质、根系分泌物以及微生物残体(Kalbitz et al., 2000; 鲍勇等, 2018).森林不同自然更新恢复阶段, 其林分土壤肥力难以恢复及物种组成的差异, 导致地表凋落物质量和数量、土壤温湿度、微生物组成等发生不同程度变化, 从而影响到土壤活性有机碳氮含量的变化(张雪等, 2016; Hagedorn et al., 2019).本研究中, DOC、LFOC和DON含量均随次生林演替呈显著下降趋势(p < 0.05), 且在70-NSF和80-NSF中差异不显著(表3), 而演替到岷江冷杉原始林阶段, 其土壤活性碳氮含量显著增加(p < 0.05), 这与该区域人工林演替过程中表层(0-20 cm)土壤活性碳氮含量的研究结果相似(姜发艳等, 2009), 但不同于热带(张哲等, 2019)和亚热带(范跃新等, 2013)森林演替的研究结果, 说明不同温湿条件及植被林分结构对土壤活性有机碳氮组分含量的影响表现出不同的变化特征(Li et al., 2014; Taylor et al., 2019).究其原因, 一方面随着森林更新演替, 土壤碳氮矿化速率增加(Mylliemngap et al., 2016), 且阔叶林的矿化速率高于针叶林(Côté et al., 2000); 另一方面由于表层土壤密度随演替增加, 而土壤团聚度、结构系数、总孔隙及毛管孔隙等土壤物理性质则呈现降低趋势(庞学勇等, 2003), 土壤有机碳、氮缺少土壤团聚体的物理性保护, 导致土壤活性有机碳和DON含量降低.此外, 岷江冷杉原始林的土壤活性碳氮组分含量高于天然次生林(表3), 可能与该林分主要物种构成(针叶+阔叶树种)有一定关联, 源于该区域针阔树种凋落叶融合在一起易于分解(黄玉梅等, 2015), 其释放的养分便于微生物分解利用, 导致土壤活性有机碳含量增加.如要明晰上述影响因素对土壤活性有机碳氮的驱动机理, 仍需进一步深入研究.相关分析表明, 不同天然更新演替阶段林地表层土壤SOC和TN含量与活性有机碳各组分含量及DON含量均呈极显著正相关关系(p < 0.01), 说明土壤活性有机碳和氮含量与土壤SOC、TN含量之间关系极为密切, 这与已有研究(辜翔等, 2016; 宫立等, 2017) 一致, 说明天然次生林土壤氮储量是调控其碳固持的重要参数(Li et al., 2012; Smal et al., 2019). ...
Changes in the stocks of soil organic carbon, total nitrogen and phosphorus following afforestation of post-arable soils: a chronosequence study
1
2019
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
Soil carbon fluxes and stocks in a Great Lakes forest chronosequence
1
2009
... 20世纪50-80年代, 川西米亚罗林区原始云冷杉暗针叶林遭受大规模采伐, 至1998年停采封育.采伐后的森林主要以人工更新为主, 自然更新演替为辅, 已形成原始林+人工林+次生林的植被镶嵌分布格局.森林植被次生演替过程中, 其类型和生境条件发生动态变化, 势必会影响森林生态系统的土壤有机碳氮储量以及酶活性变化.有研究显示, 土壤肥力随云杉(Picea asperata)人工林演替呈显著降低趋势(Pang et al., 2009), 森林的自然更新恢复更有益于土壤有机碳和氮的固存(Pang et al., 2019).虽然人们对该区域不同森林类型或演替阶段植被类型土壤有机碳氮储量变化进行了大量研究(Zhang et al., 2007; 周义贵等, 2014), 但对于天然次生林不同演替阶段的土壤碳氮含量及其与酶活性的关系仍然了解不够, 限制了对川西亚高山天然次生林恢复与保护对土壤肥力演变及酶活性的影响机制的认识.为此, 本研究采用空间代替时间的方法, 选取环境条件基本一致, 处于不同演替阶段的天然次生林及原始林作为一个自然更新恢复演替序列, 旨在分析土壤碳氮含量和酶活性的变化趋势, 结合相关分析及冗余分析(RDA)方法揭示土壤碳氮含量及影响土壤胞外酶活性变化的关键环境因子, 以验证以下假设: (1)森林自然更新演替提高了表层土壤碳氮含量与酶活性; (2)天然次生林土壤碳氮含量与酶活性低于原始林; (3)土壤理化性质的变化及物种多样性对土壤酶活性产生显著影响.本研究可为该区域天然次生林土壤碳氮固持潜力评价提供基础数据, 为天然次生林林分结构的合理调控及未来森林更新方式的选择提供科学依据. ...
Carbon storage in a chronosequence of red spruce (Picea rubens) forests in central Nova Scotia, Canada
2007
Successional dynamics of nitrogen fixation and forest growth in regenerating Costa Rican rainforests
1
2019
... 土壤活性有机碳氮是土壤可溶性有机质的主要组分, 主要来源于新近枯枝落叶、腐殖质、根系分泌物以及微生物残体(Kalbitz et al., 2000; 鲍勇等, 2018).森林不同自然更新恢复阶段, 其林分土壤肥力难以恢复及物种组成的差异, 导致地表凋落物质量和数量、土壤温湿度、微生物组成等发生不同程度变化, 从而影响到土壤活性有机碳氮含量的变化(张雪等, 2016; Hagedorn et al., 2019).本研究中, DOC、LFOC和DON含量均随次生林演替呈显著下降趋势(p < 0.05), 且在70-NSF和80-NSF中差异不显著(表3), 而演替到岷江冷杉原始林阶段, 其土壤活性碳氮含量显著增加(p < 0.05), 这与该区域人工林演替过程中表层(0-20 cm)土壤活性碳氮含量的研究结果相似(姜发艳等, 2009), 但不同于热带(张哲等, 2019)和亚热带(范跃新等, 2013)森林演替的研究结果, 说明不同温湿条件及植被林分结构对土壤活性有机碳氮组分含量的影响表现出不同的变化特征(Li et al., 2014; Taylor et al., 2019).究其原因, 一方面随着森林更新演替, 土壤碳氮矿化速率增加(Mylliemngap et al., 2016), 且阔叶林的矿化速率高于针叶林(Côté et al., 2000); 另一方面由于表层土壤密度随演替增加, 而土壤团聚度、结构系数、总孔隙及毛管孔隙等土壤物理性质则呈现降低趋势(庞学勇等, 2003), 土壤有机碳、氮缺少土壤团聚体的物理性保护, 导致土壤活性有机碳和DON含量降低.此外, 岷江冷杉原始林的土壤活性碳氮组分含量高于天然次生林(表3), 可能与该林分主要物种构成(针叶+阔叶树种)有一定关联, 源于该区域针阔树种凋落叶融合在一起易于分解(黄玉梅等, 2015), 其释放的养分便于微生物分解利用, 导致土壤活性有机碳含量增加.如要明晰上述影响因素对土壤活性有机碳氮的驱动机理, 仍需进一步深入研究.相关分析表明, 不同天然更新演替阶段林地表层土壤SOC和TN含量与活性有机碳各组分含量及DON含量均呈极显著正相关关系(p < 0.01), 说明土壤活性有机碳和氮含量与土壤SOC、TN含量之间关系极为密切, 这与已有研究(辜翔等, 2016; 宫立等, 2017) 一致, 说明天然次生林土壤氮储量是调控其碳固持的重要参数(Li et al., 2012; Smal et al., 2019). ...
Ecological linkages between aboveground and belowground Biota
1
2004
... 20世纪50-80年代, 川西米亚罗林区原始云冷杉暗针叶林遭受大规模采伐, 至1998年停采封育.采伐后的森林主要以人工更新为主, 自然更新演替为辅, 已形成原始林+人工林+次生林的植被镶嵌分布格局.森林植被次生演替过程中, 其类型和生境条件发生动态变化, 势必会影响森林生态系统的土壤有机碳氮储量以及酶活性变化.有研究显示, 土壤肥力随云杉(Picea asperata)人工林演替呈显著降低趋势(Pang et al., 2009), 森林的自然更新恢复更有益于土壤有机碳和氮的固存(Pang et al., 2019).虽然人们对该区域不同森林类型或演替阶段植被类型土壤有机碳氮储量变化进行了大量研究(Zhang et al., 2007; 周义贵等, 2014), 但对于天然次生林不同演替阶段的土壤碳氮含量及其与酶活性的关系仍然了解不够, 限制了对川西亚高山天然次生林恢复与保护对土壤肥力演变及酶活性的影响机制的认识.为此, 本研究采用空间代替时间的方法, 选取环境条件基本一致, 处于不同演替阶段的天然次生林及原始林作为一个自然更新恢复演替序列, 旨在分析土壤碳氮含量和酶活性的变化趋势, 结合相关分析及冗余分析(RDA)方法揭示土壤碳氮含量及影响土壤胞外酶活性变化的关键环境因子, 以验证以下假设: (1)森林自然更新演替提高了表层土壤碳氮含量与酶活性; (2)天然次生林土壤碳氮含量与酶活性低于原始林; (3)土壤理化性质的变化及物种多样性对土壤酶活性产生显著影响.本研究可为该区域天然次生林土壤碳氮固持潜力评价提供基础数据, 为天然次生林林分结构的合理调控及未来森林更新方式的选择提供科学依据. ...
The effects of tree rhizodeposition on soil exoenzyme activity, dissolved organic carbon, and nutrient availability in a subalpine forest ecosystem
1
2007
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
川西亚高山冷杉林和白桦林土壤酶活性季节动态
1
2004
... 相关分析发现, PHO活性与TN、SOC、DOC和LFOC含量显著相关, PEO活性则未受到土壤底物的显著影响(表6), 虽然土壤碳氮及活性有机碳组分含量随次生林更新演替呈显著降低趋势(表3), 但PHO和PEO活性并未发生明显变化(表4), 说明次生林更新演替早期土壤惰性碳库中难降解碳含量变化不明显; 另外, 在5种酶中, 只有CBH活性与H和D显著正相关, 说明多CBH活性受到物种多样性影响较大, 说明在森林早期的自然演替过程中, 物种多样性下降(图3), 供给土壤微生物分解利用物质的种类相对单一, 使得最易被微生物利用的DON含量减少而影响到土壤碳氮固存, 进而影响到其他酶活性.相关分析也显示, βG与NAG和PHO以及NAG与PHO活性之间均表现为显著正相关关系, 表明土壤微生物易于倾向利用树木凋落物形成的腐殖质分解产生的活性碳库, 说明次生林在更新演替进程中可能有利于土壤惰性碳库的积累; 土壤含水率显著影响βG、NAG和PHO酶活性, 且酶活性随土壤含水率增加而增加, 这与已有研究结果(Pushkareva et al., 2020; Zhou et al., 2020)相似.综上, 不同演替阶段的天然次生林, 其土壤酶活性的激发响应机理需做进一步深入研究. ...
川西亚高山冷杉林和白桦林土壤酶活性季节动态
1
2004
... 相关分析发现, PHO活性与TN、SOC、DOC和LFOC含量显著相关, PEO活性则未受到土壤底物的显著影响(表6), 虽然土壤碳氮及活性有机碳组分含量随次生林更新演替呈显著降低趋势(表3), 但PHO和PEO活性并未发生明显变化(表4), 说明次生林更新演替早期土壤惰性碳库中难降解碳含量变化不明显; 另外, 在5种酶中, 只有CBH活性与H和D显著正相关, 说明多CBH活性受到物种多样性影响较大, 说明在森林早期的自然演替过程中, 物种多样性下降(图3), 供给土壤微生物分解利用物质的种类相对单一, 使得最易被微生物利用的DON含量减少而影响到土壤碳氮固存, 进而影响到其他酶活性.相关分析也显示, βG与NAG和PHO以及NAG与PHO活性之间均表现为显著正相关关系, 表明土壤微生物易于倾向利用树木凋落物形成的腐殖质分解产生的活性碳库, 说明次生林在更新演替进程中可能有利于土壤惰性碳库的积累; 土壤含水率显著影响βG、NAG和PHO酶活性, 且酶活性随土壤含水率增加而增加, 这与已有研究结果(Pushkareva et al., 2020; Zhou et al., 2020)相似.综上, 不同演替阶段的天然次生林, 其土壤酶活性的激发响应机理需做进一步深入研究. ...
Soil enzyme activity and stoichiometry in forest ecosystems along the North-South Transect in eastern China (NSTEC)
1
2017
... 森林自然更新使其树种组成、凋落物数量以及植物根系发生变化而影响到土壤碳氮循环过程(范跃新等, 2013; Dhillon & van Rees, 2017).有研究认为, 凋落物输入量会促进部分新输入土壤的碳分解释放而产生“激发效应”, 加速土壤中原有的有机质分解(Saiya-Cork et al., 2002; Qiao et al., 2019), 随着森林更新演替的进行, 输入到土壤的凋落物量渐增(张远东等, 2019), 从而促使土壤原有储存的部分有机质分解, 且植物根系含量的增加提高了土壤有机质分解速率(Adamczyk et al., 2019), 使得土壤有机碳氮储量随森林更新演替呈显著下降趋势, 主要因为土壤有机碳氮储量受到植物生长以及凋落物种类组成的影响(del Valle et al., 2020).本研究中, 原始冷杉林(CK)中SOC和TN含量显著高于3种天然次生林(p < 0.05), 而在3种天然次生林中, SOC含量表现为80-NSF > 70-NSF > 60-NSF和TN为70-NSF > 80-NSF > 60-NSF, 其中80-NSF和70-NSF差异不显著(p > 0.05), 一方面由于采伐迹地自然更新演替次生林的早期阶段, 土壤微生境的变化加速了土壤中累积的有机质分解速率, 以及阔叶树种中的碳和木质素多低于针叶树种且分解速率较快(谌贤等, 2017); 另一方面天然次生林演替至60年, 外来物种进入难度增加致使物种多样性出现降低趋势(图3), 导致DON含量减少而影响到土壤SOC和TN含量.综上, 在川西亚高山高海拔特殊生境条件及其生态脆弱环境背景下, 如森林遭受破坏后, 其土壤肥力难以恢复.本研究仅对60年内不同演替阶段的次生林进行了观察, 对土壤碳氮动态变化机制的理解有限, 但研究结果显示, 如果要恢复到采伐前原始林的土壤肥力水平(表3)需要更长的时间. ...
Carbon and nitrogen dynamics during forest stand development: a global synthesis
1
2011
... 森林自然更新使其树种组成、凋落物数量以及植物根系发生变化而影响到土壤碳氮循环过程(范跃新等, 2013; Dhillon & van Rees, 2017).有研究认为, 凋落物输入量会促进部分新输入土壤的碳分解释放而产生“激发效应”, 加速土壤中原有的有机质分解(Saiya-Cork et al., 2002; Qiao et al., 2019), 随着森林更新演替的进行, 输入到土壤的凋落物量渐增(张远东等, 2019), 从而促使土壤原有储存的部分有机质分解, 且植物根系含量的增加提高了土壤有机质分解速率(Adamczyk et al., 2019), 使得土壤有机碳氮储量随森林更新演替呈显著下降趋势, 主要因为土壤有机碳氮储量受到植物生长以及凋落物种类组成的影响(del Valle et al., 2020).本研究中, 原始冷杉林(CK)中SOC和TN含量显著高于3种天然次生林(p < 0.05), 而在3种天然次生林中, SOC含量表现为80-NSF > 70-NSF > 60-NSF和TN为70-NSF > 80-NSF > 60-NSF, 其中80-NSF和70-NSF差异不显著(p > 0.05), 一方面由于采伐迹地自然更新演替次生林的早期阶段, 土壤微生境的变化加速了土壤中累积的有机质分解速率, 以及阔叶树种中的碳和木质素多低于针叶树种且分解速率较快(谌贤等, 2017); 另一方面天然次生林演替至60年, 外来物种进入难度增加致使物种多样性出现降低趋势(图3), 导致DON含量减少而影响到土壤SOC和TN含量.综上, 在川西亚高山高海拔特殊生境条件及其生态脆弱环境背景下, 如森林遭受破坏后, 其土壤肥力难以恢复.本研究仅对60年内不同演替阶段的次生林进行了观察, 对土壤碳氮动态变化机制的理解有限, 但研究结果显示, 如果要恢复到采伐前原始林的土壤肥力水平(表3)需要更长的时间. ...
天然红松林植被多样性与土壤功能多样性的关系
1
2014
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
天然红松林植被多样性与土壤功能多样性的关系
1
2014
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
Impacts of conversion from secondary forests to larch plantations on the structure and function of microbial communities
2017
长白山白桦林不同演替阶段土壤有机碳组分的变化
1
2016
... 森林自然更新使其树种组成、凋落物数量以及植物根系发生变化而影响到土壤碳氮循环过程(范跃新等, 2013; Dhillon & van Rees, 2017).有研究认为, 凋落物输入量会促进部分新输入土壤的碳分解释放而产生“激发效应”, 加速土壤中原有的有机质分解(Saiya-Cork et al., 2002; Qiao et al., 2019), 随着森林更新演替的进行, 输入到土壤的凋落物量渐增(张远东等, 2019), 从而促使土壤原有储存的部分有机质分解, 且植物根系含量的增加提高了土壤有机质分解速率(Adamczyk et al., 2019), 使得土壤有机碳氮储量随森林更新演替呈显著下降趋势, 主要因为土壤有机碳氮储量受到植物生长以及凋落物种类组成的影响(del Valle et al., 2020).本研究中, 原始冷杉林(CK)中SOC和TN含量显著高于3种天然次生林(p < 0.05), 而在3种天然次生林中, SOC含量表现为80-NSF > 70-NSF > 60-NSF和TN为70-NSF > 80-NSF > 60-NSF, 其中80-NSF和70-NSF差异不显著(p > 0.05), 一方面由于采伐迹地自然更新演替次生林的早期阶段, 土壤微生境的变化加速了土壤中累积的有机质分解速率, 以及阔叶树种中的碳和木质素多低于针叶树种且分解速率较快(谌贤等, 2017); 另一方面天然次生林演替至60年, 外来物种进入难度增加致使物种多样性出现降低趋势(图3), 导致DON含量减少而影响到土壤SOC和TN含量.综上, 在川西亚高山高海拔特殊生境条件及其生态脆弱环境背景下, 如森林遭受破坏后, 其土壤肥力难以恢复.本研究仅对60年内不同演替阶段的次生林进行了观察, 对土壤碳氮动态变化机制的理解有限, 但研究结果显示, 如果要恢复到采伐前原始林的土壤肥力水平(表3)需要更长的时间. ...
长白山白桦林不同演替阶段土壤有机碳组分的变化
1
2016
... 森林自然更新使其树种组成、凋落物数量以及植物根系发生变化而影响到土壤碳氮循环过程(范跃新等, 2013; Dhillon & van Rees, 2017).有研究认为, 凋落物输入量会促进部分新输入土壤的碳分解释放而产生“激发效应”, 加速土壤中原有的有机质分解(Saiya-Cork et al., 2002; Qiao et al., 2019), 随着森林更新演替的进行, 输入到土壤的凋落物量渐增(张远东等, 2019), 从而促使土壤原有储存的部分有机质分解, 且植物根系含量的增加提高了土壤有机质分解速率(Adamczyk et al., 2019), 使得土壤有机碳氮储量随森林更新演替呈显著下降趋势, 主要因为土壤有机碳氮储量受到植物生长以及凋落物种类组成的影响(del Valle et al., 2020).本研究中, 原始冷杉林(CK)中SOC和TN含量显著高于3种天然次生林(p < 0.05), 而在3种天然次生林中, SOC含量表现为80-NSF > 70-NSF > 60-NSF和TN为70-NSF > 80-NSF > 60-NSF, 其中80-NSF和70-NSF差异不显著(p > 0.05), 一方面由于采伐迹地自然更新演替次生林的早期阶段, 土壤微生境的变化加速了土壤中累积的有机质分解速率, 以及阔叶树种中的碳和木质素多低于针叶树种且分解速率较快(谌贤等, 2017); 另一方面天然次生林演替至60年, 外来物种进入难度增加致使物种多样性出现降低趋势(图3), 导致DON含量减少而影响到土壤SOC和TN含量.综上, 在川西亚高山高海拔特殊生境条件及其生态脆弱环境背景下, 如森林遭受破坏后, 其土壤肥力难以恢复.本研究仅对60年内不同演替阶段的次生林进行了观察, 对土壤碳氮动态变化机制的理解有限, 但研究结果显示, 如果要恢复到采伐前原始林的土壤肥力水平(表3)需要更长的时间. ...
川西亚高山五种主要森林类型凋落物组成及动态
1
2019
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
川西亚高山五种主要森林类型凋落物组成及动态
1
2019
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
Microarray-based analysis of changes in diversity of microbial genes involved in organic carbon decomposition following land use/cover changes
1
2007
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
土壤易氧化有机碳对西双版纳热带森林群落演替的响应
1
2019
... 土壤活性有机碳氮是土壤可溶性有机质的主要组分, 主要来源于新近枯枝落叶、腐殖质、根系分泌物以及微生物残体(Kalbitz et al., 2000; 鲍勇等, 2018).森林不同自然更新恢复阶段, 其林分土壤肥力难以恢复及物种组成的差异, 导致地表凋落物质量和数量、土壤温湿度、微生物组成等发生不同程度变化, 从而影响到土壤活性有机碳氮含量的变化(张雪等, 2016; Hagedorn et al., 2019).本研究中, DOC、LFOC和DON含量均随次生林演替呈显著下降趋势(p < 0.05), 且在70-NSF和80-NSF中差异不显著(表3), 而演替到岷江冷杉原始林阶段, 其土壤活性碳氮含量显著增加(p < 0.05), 这与该区域人工林演替过程中表层(0-20 cm)土壤活性碳氮含量的研究结果相似(姜发艳等, 2009), 但不同于热带(张哲等, 2019)和亚热带(范跃新等, 2013)森林演替的研究结果, 说明不同温湿条件及植被林分结构对土壤活性有机碳氮组分含量的影响表现出不同的变化特征(Li et al., 2014; Taylor et al., 2019).究其原因, 一方面随着森林更新演替, 土壤碳氮矿化速率增加(Mylliemngap et al., 2016), 且阔叶林的矿化速率高于针叶林(Côté et al., 2000); 另一方面由于表层土壤密度随演替增加, 而土壤团聚度、结构系数、总孔隙及毛管孔隙等土壤物理性质则呈现降低趋势(庞学勇等, 2003), 土壤有机碳、氮缺少土壤团聚体的物理性保护, 导致土壤活性有机碳和DON含量降低.此外, 岷江冷杉原始林的土壤活性碳氮组分含量高于天然次生林(表3), 可能与该林分主要物种构成(针叶+阔叶树种)有一定关联, 源于该区域针阔树种凋落叶融合在一起易于分解(黄玉梅等, 2015), 其释放的养分便于微生物分解利用, 导致土壤活性有机碳含量增加.如要明晰上述影响因素对土壤活性有机碳氮的驱动机理, 仍需进一步深入研究.相关分析表明, 不同天然更新演替阶段林地表层土壤SOC和TN含量与活性有机碳各组分含量及DON含量均呈极显著正相关关系(p < 0.01), 说明土壤活性有机碳和氮含量与土壤SOC、TN含量之间关系极为密切, 这与已有研究(辜翔等, 2016; 宫立等, 2017) 一致, 说明天然次生林土壤氮储量是调控其碳固持的重要参数(Li et al., 2012; Smal et al., 2019). ...
土壤易氧化有机碳对西双版纳热带森林群落演替的响应
1
2019
... 土壤活性有机碳氮是土壤可溶性有机质的主要组分, 主要来源于新近枯枝落叶、腐殖质、根系分泌物以及微生物残体(Kalbitz et al., 2000; 鲍勇等, 2018).森林不同自然更新恢复阶段, 其林分土壤肥力难以恢复及物种组成的差异, 导致地表凋落物质量和数量、土壤温湿度、微生物组成等发生不同程度变化, 从而影响到土壤活性有机碳氮含量的变化(张雪等, 2016; Hagedorn et al., 2019).本研究中, DOC、LFOC和DON含量均随次生林演替呈显著下降趋势(p < 0.05), 且在70-NSF和80-NSF中差异不显著(表3), 而演替到岷江冷杉原始林阶段, 其土壤活性碳氮含量显著增加(p < 0.05), 这与该区域人工林演替过程中表层(0-20 cm)土壤活性碳氮含量的研究结果相似(姜发艳等, 2009), 但不同于热带(张哲等, 2019)和亚热带(范跃新等, 2013)森林演替的研究结果, 说明不同温湿条件及植被林分结构对土壤活性有机碳氮组分含量的影响表现出不同的变化特征(Li et al., 2014; Taylor et al., 2019).究其原因, 一方面随着森林更新演替, 土壤碳氮矿化速率增加(Mylliemngap et al., 2016), 且阔叶林的矿化速率高于针叶林(Côté et al., 2000); 另一方面由于表层土壤密度随演替增加, 而土壤团聚度、结构系数、总孔隙及毛管孔隙等土壤物理性质则呈现降低趋势(庞学勇等, 2003), 土壤有机碳、氮缺少土壤团聚体的物理性保护, 导致土壤活性有机碳和DON含量降低.此外, 岷江冷杉原始林的土壤活性碳氮组分含量高于天然次生林(表3), 可能与该林分主要物种构成(针叶+阔叶树种)有一定关联, 源于该区域针阔树种凋落叶融合在一起易于分解(黄玉梅等, 2015), 其释放的养分便于微生物分解利用, 导致土壤活性有机碳含量增加.如要明晰上述影响因素对土壤活性有机碳氮的驱动机理, 仍需进一步深入研究.相关分析表明, 不同天然更新演替阶段林地表层土壤SOC和TN含量与活性有机碳各组分含量及DON含量均呈极显著正相关关系(p < 0.01), 说明土壤活性有机碳和氮含量与土壤SOC、TN含量之间关系极为密切, 这与已有研究(辜翔等, 2016; 宫立等, 2017) 一致, 说明天然次生林土壤氮储量是调控其碳固持的重要参数(Li et al., 2012; Smal et al., 2019). ...
Soil extracellular enzyme activity and stoichiometry in China’s forests
1
2020
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
川西亚高山不同土地利用类型对土壤微生物量碳动态特征的影响
1
2014
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...
川西亚高山不同土地利用类型对土壤微生物量碳动态特征的影响
1
2014
... 森林土壤有机碳质量和数量是反映土壤质量和森林健康状况的重要指标, 直接或间接地影响到森林生物量和土壤肥力.研究显示, 森林土壤碳储量约为全球土壤碳储量的73% (Post & Kwon, 2000), 在调控全球碳平衡方面起着重要作用.次生林作为陆地植被生态系统的主要组分(Kauppi et al., 2006), 在维护生物多样性和土壤碳氮循环中扮演着重要角色(Yang et al., 2011).森林更新演替改变了植被群落结构、树种组成及其密度而影响到土壤微生境, 导致土壤有机碳、氮及其组分含量和微生物活性发生变化(Chandra et al., 2016; Feng et al., 2019).有研究认为, 森林植被次生演替早期阶段土壤中有机碳分解速率超过地上植被输入土壤中有机碳的固持速率, 使得其含量相对较低(Ohtsuka et al., 2010; Milcu et al., 2011), 也有研究表明, 土壤碳氮储量随着森林正向演替呈现出增加(Deng et al., 2013; 张雪等, 2016; Liu et al., 2020)、变化不明显(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)或先降后增(Tang et al., 2009)的变化趋势, 说明森林演替对土壤碳氮储量的影响较为复杂.土壤有机质降解受到微生物的显著影响(Sinsabaugh, 2010; Liu et al., 2020), 而微生物的生长活动也会受到土壤碳氮有效性的制约(Kooch et al., 2018; Dove et al., 2019).森林演替改变了土壤微生境, 微生物种类组成会随之发生变化, 进而改变了土壤微生物活性(Zhang et al., 2017), 其活性主要体现在来源于动植物残体分解过程中土壤微生物以及植物根系分泌物的胞外酶上.胞外酶是有机碳氮分解和周转的主要催化剂(Weintraub et al., 2007; Xu et al., 2017), 表明土壤有机碳氮的积累依赖于土壤酶的矿化分解作用(Sinsabaugh et al., 2014; Mayer et al., 2020).土壤有机质矿化速率受控于胞外酶活性, 其中水解酶主要参与活性碳氮分解转化(如糖类、淀粉、纤维素等), 包括β-葡萄糖苷酶(βG)、纤维素水解酶(CBH)、β-乙酰氨基葡萄糖苷酶(NAG)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化过程等, 包括多酚氧化酶(PHO)、过氧化物酶(PEO)等(Luo et al., 2019), 因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究显示, 土壤酶活性与土壤养分含量显著负相关(Burns et al., 2013; Das & Mondal, 2019)、显著正相关(Li et al., 2015; Feng et al., 2019).同时, 土壤底物的有效性也会影响有机碳和氮的分解酶活性, 使其凋落物及土壤有机质表现出不同的分解速率(Kang et al., 2009; Zhou et al., 2020).此外, 土壤酶活性与土壤温湿度及pH等因素有关(马伟伟等, 2019; Zhou et al., 2020), 说明土壤酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与土壤有机碳氮组分含量间的关系已被广泛关注, 但对于川西亚高山天然次生林土壤有机碳氮含量及土壤胞外酶活性特征认识不够深刻. ...



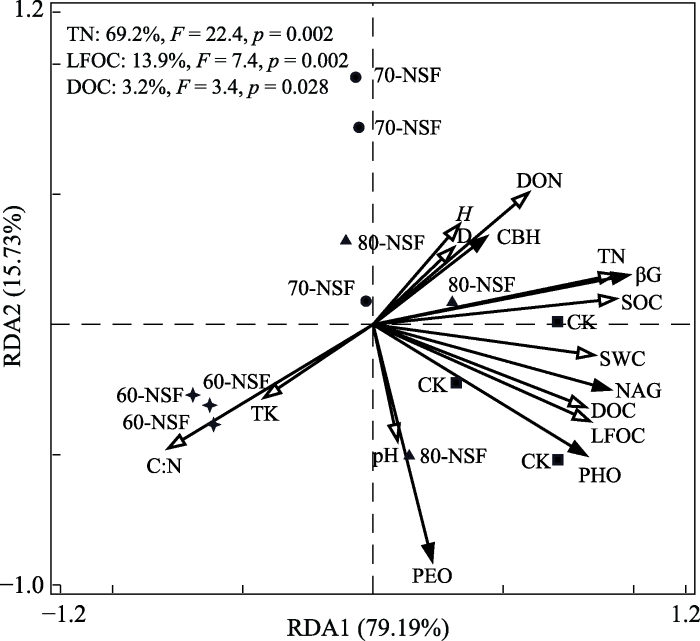
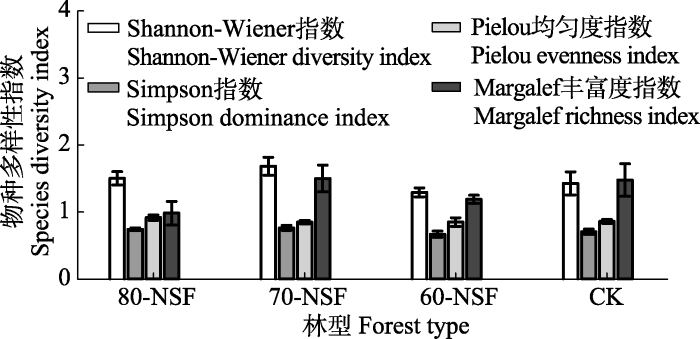


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}