




植物间相互作用包括竞争作用和促进作用, 前者也叫负相互作用, 后者则为正相互作用。竞争作用是指在共享有限的生存空间和资源时, 种内或种间个体对同一资源的争夺现象, 通常是双向的关系。促进作用是指在植物种间或种内, 施利者(护卫植物)通过改善恶劣的生存环境、增加资源的可利用性、消除潜在竞争者或抵御取食者等方式, 直接或间接地促进受益者的生存、生长或者增加其丰富度, 促进作用可能是单向或双向的, 包括互利共生(+/+)和偏利共生(+/0和+/-)(Bertness & Callaway, 1994; Callaway, 1995, 2007; Bruno et al., 2003; 张炜平等, 2013; Liancourt & Dolezal, 2021)。
竞争作用和促进作用同时存在于自然界的植物群落, 其净效应取决于二者的相对强度, 而它们的相对强弱与环境密切相关, 胁迫梯度假说(SGH)指出, 随环境胁迫程度的增加, 竞争作用通常减弱, 而促进作用增强, 前者在低胁迫环境中起主导作用, 后者则在高胁迫环境发挥更重要的作用(Bertness & Callaway, 1994)。自SGH提出以来, 围绕其在各类生态系统中普适性的验证成为了植物间促进作用研究的热点, 大量的野外观测、控制实验、meta分析被用来验证和完善SGH (Callaway et al., 2002; Callaway, 2007; Maestre et al., 2009b; Malkinson & Tielbörger, 2010; He et al., 2013; Soliveres et al., 2015; Michalet & Pugnaire, 2016)。为准确预测促进作用随环境胁迫度的变化规律, 促进作用发生机制的研究随之成为植物间相互作用研究的另一重要议题, 护卫植物通过直接或间接的作用改善周围环境, 从而影响受益者的生存、生长和繁殖(Maestre et al., 2003; Armas & Pugnaire, 2005; Michalet et al., 2015; Tirado et al., 2015; Michalet & Pugnaire, 2016; Noumi et al., 2016)。随着研究的不断深入, 促进作用逐渐融入到生态学的重要理论中, 例如, 促进作用被证明是胁迫环境下物种多样性形成与维持的重要机制(Hooper et al., 2005; Kikvidze et al., 2005; Butterfield et al., 2013; Cavieres et al., 2014, 2016; Soliveres & Maestre, 2014; Ballantyne & Pickering, 2015; Bråthen & Lortie, 2016; Michalet & Pugnaire, 2016; Pistón et al., 2016; Parajuli et al., 2021)。促进作用拓宽了一些物种的分布范围, 相关研究完善了一直以来基于竞争关系建立的生态位理论(Choler et al., 2001; Bruno et al., 2003)。对促进作用的研究修正了前期基于竞争关系提出的个体生物量与种群密度关系理论(个体生物量随种群密度的增加线性降低), 在考虑促进作用后, 个体生物量与种群密度的关系随环境胁迫的增强由单调递减变为先增后减的单峰格局, 即个体生物量最高出现在中等密度条件(Chu et al., 2008)。此外, 植物间促进作用还被广泛用于退化生态系统的植被恢复(Maestre et al., 2001; Gómez-Aparicio et al., 2004; Padilla & Pugnaire, 2006; Gómez-Aparicio, 2009; 刘泽正等, 2020)。
对植物间促进作用的系统研究, 最早始于盐沼地生态系统(Bertness, 1991; Bertness & Shumway, 1993; Bertness & Hacker, 1994), 后来逐步扩展到以垫状植物作为护卫植物的高山生态系统(Cavieres et al., 2006, 2016; Butterfield et al., 2013; Pugnaire et al., 2015)和以灌木作为护卫植物的干旱半干旱生态系统(Pugnaire et al., 1996; Moro et al., 1997a; Michalet et al., 2015; Noumi et al., 2016; Zhang et al., 2018a)。在气候变化和人类活动的影响下, 干旱半干旱区生态系统正发生着剧烈变化, 灌丛作为干旱半干旱区重要的植被类型, 其促进作用对生态系统-气候变化响应关系及生态系统功能的体现具有重要影响。然而, 干旱半干旱区灌丛促进作用的发生机制, 尤其是灌丛对草本植物的促进作用沿水分梯度的变化, 目前还没有较为统一的结论。此外, 灌丛在植被恢复中的作用也尚未得到全面的评估和运用。本文从灌丛对草本植物的促进作用随水分的变化规律、灌丛促进作用的发生机制以及灌丛促进作用在植被恢复中的应用等方面总结了最近30年来的主要研究进展, 并在此基础上提出了未来研究中需深入探索的问题。
1 灌丛促进作用随水分的变化规律
植物间竞争和促进作用的强度与环境密切相关, 作为植物间相互作用随环境胁迫程度变化研究中影响最深远的预测模型, SGH得到了大量野外数据的验证(Pugnaire & Luque, 2001; Callaway et al., 2002; Holzapfel et al., 2006; Armas et al., 2011; Ziffer- Berger et al., 2014; Lortie et al., 2016)。然而越来越多的研究发现植物间促进作用随环境胁迫增强并不总是线性增强, 这种不确定性在干旱半干旱生态系统尤为明显(Tielbörger & Kadmon, 2000; Maestre & Cortina, 2004; Maestre et al., 2005, 2006, 2009b; Lortie & Callaway, 2006; Michalet et al., 2006; Holmgren & Scheffer, 2010; Liancourt et al., 2017; O’Brien et al., 2017; Zhang et al., 2018b; Berdugo et al., 2019)。例如, 在以色列Negev沙漠, 灌木对一年生草本植物密度和种子数的促进作用随降水量的减少而减弱, 原因是在极度干旱的年份(年降水量38 mm), 灌木冠层的截流作用导致其下草本植物遭受严重水分胁迫, 而在湿润年份, 水分不是主要限制因子, 灌木对微气候的改善发挥了较强的促进作用(Tielbörger & Kadmon, 2000)。在西班牙半干旱区, 随降水量的减少, 豆科灌木Retama sphaerocarpa对其冠层下草本植物生物量和物种丰富度的影响由促进作用转为竞争作用(O’Brien et al., 2017)。在中国巴丹吉林沙漠, 沙拐枣(Calligonum mongolicum)对其冠层下草本植物的促进作用随夏季降水量的减少转为中性作用, 原因是灌丛对土壤水分的促进作用随夏季降水量的减少而减弱(Zhang et al., 2018a)。
促进作用发挥的关键是护卫植物能改善受益者存活或生长的限制因子, 水分作为干旱半干旱区植物存活和生长的主要限制因子(Liancourt et al., 2005; Báez et al., 2013), 灌丛对水分的影响复杂多变, 这可能是造成灌丛与草本植物间相互作用沿水分梯度较难预测的重要原因。干旱半干旱区灌木能通过根系提水作用将深层土壤水分转移到浅层土壤, 提高浅层土壤的水分含量(Richards & Caldwell, 1987; Prieto et al., 2011)。灌木遮阴能减弱蒸散发, 进而提高土壤含水量(Liu et al., 2021)。然而, 灌木的截流和竞争等作用可能会对土壤水分产生负作用(Tielbörger & Kadmon, 2000; Darrouzet-Nardi et al., 2006; Hamerlynck et al., 2011)。Butterfield等(2016)模拟了美国西部灌丛内外土壤含水量的动态变化, 基于多年平均值时, 灌丛对土壤水分的促进作用随干旱增强而减弱, 而在极端干旱或湿润年份, 灌丛对土壤水分的影响随干旱增强呈单峰格局, 研究指出灌丛对土壤水分的影响与降水、蒸发以及蒸腾速率的相对强弱密切相关。
随着研究的不断深入, 学者们普遍认识到灌丛与草本植物间相互作用不仅受水分的影响, 还与物种特性、植物生活史阶段以及环境胁迫类型等密切相关(He et al., 2013; Soliveres et al., 2014; Zhang et al., 2016; Losapio et al., 2018; Chaieb et al., 2021; Parajuli et al., 2021), 例如, 在巴西东北部半干旱地区, 矮灌木Poincianella microphylla对冠层下草本物种始终表现为促进作用, 而Cnidoscolus quercifolius和Mimosa tenuiflora的促进作用随草本物种的生长逐渐转变为中性或竞争作用(Paterno et al., 2016)。在印度北部地区, 变色锦鸡儿(Caragana versicolor)对禾草类和直立生长的物种发挥促进作用, 而对匍匐生长的物种更多地表现为竞争作用(Iyengar et al., 2017)。在西班牙东南部, 岩蔷薇属灌木Cistus clusii对针茅属植物Stipa tenacissima的促进作用随针茅的成熟而减弱, 原因是成熟针茅获取限制性资源即水分的能力增强(Armas & Pugnaire, 2005)。此外, 研究方法、研究水平和指标选取的不同, 也会影响灌丛与草本植物间相互作用的结果(Maestre et al., 2009b; Soliveres et al., 2015; Iyengar et al., 2017), 例如, Maestre等(2005)对1970-2004年间干旱半干旱区植物间相互作用研究案例进行了综合分析, 发现护卫植物对受益者的存活和生长普遍表现出中性作用, 而对受益者密度和繁殖而言, 在干旱胁迫较低的生境中表现为促进作用, 在干旱胁迫较高的生境则表现为竞争作用。Liancourt等(2017)在印度西北部拉达克地区的研究发现, 随海拔降低(干旱增强), 变色锦鸡儿对草本植物的促进作用在群落水平呈现单峰格局, 而对优势伴生种低株披碱草(Elymus jacquemontii)和驼绒藜属植物Krascheninnikovia pungens的促进作用随海拔降低而增强。Metz和Tielbörger (2016)利用空间梯度法(沿降水梯度设置调查样点)、时间法(研究样点连续9年监测)和控制实验法(通过截留和增水设置干旱梯度)比较了以色列蔷薇科灌木Sarcopoterium spinosum对其冠层下草本植物的影响, 发现不同研究方法的结果并不一致: 模拟增雨和干旱处理条件下, 灌木对草本植物生物量和存活率无显著影响; 空间梯度法的结果表明灌木对草本植物生物量的促进作用随干旱增强而增强, 但对存活率无显著影响; 时间法结果表明随干旱增强, 灌木对草本植物存活率和生物量的竞争作用增强。Maestre等(2009b)给出了一系列环境胁迫类型(包括资源胁迫如水、光照和养分, 以及非资源胁迫如温度、风和盐碱度)和物种特性(胁迫忍耐种和竞争性物种)组合下, 植物间相互作用随环境胁迫程度变化的趋势, 例如当护卫植物和受益者分别是竞争性物种和胁迫忍耐种时, 更容易出现SGH预测的结果; 而当相互作用的物种具有相似的竞争性或胁迫忍耐生活史性状, 且胁迫梯度受资源因子驱动时, 相互作用的两个物种在胁迫梯度的两端都会出现竞争作用。
在植物间相互作用研究中, 定量描述环境胁迫程度是准确预测相互作用随环境胁迫变化的关键(Callaway, 2007)。在实际研究中, 环境胁迫因子并不总是显而易见的, 物理环境的胁迫并不等于植物的生理胁迫, 因为植物往往能通过调节物候期、生物量分配、器官性状等的方式来适应干旱环境, 其对干旱的响应也并非是线性的(Padilla & Pugnaire, 2007; de Dios Miranda et al., 2009; Padilla et al., 2009a; Malkinson & Tielbörger, 2010; Soliveres et al., 2015; Xie et al., 2015; Wei et al., 2016)。例如, Wei等(2016)发现内蒙古毛乌素沙地黑沙蒿(Artemisia ordosica)通过改变冠层水平氮分配和叶片结构性状的策略来适应更干旱的环境。促进作用只有在物理环境偏离物种生态幅时才会体现(Liancourt et al., 2005, 2017; Wang et al., 2008; He et al., 2016; Liancourt & Dolezal, 2021), 因此, 在定义和度量胁迫程度时, 要充分考虑护卫植物和受益者对胁迫环境的进化适应性, 这是因为, 在护卫植物分布的边界, 物种可能因不适应当地环境而受到较强的胁迫, 此时护卫植物对环境改善作用会减弱, 促进作用就可能在极度胁迫环境下失效或转为竞争作用(Michalet et al., 2006; Maestre et al., 2009b; Schöb et al., 2013)。相反, 如果护卫植物能很好地适应当地气候环境, 促进作用就可能随环境胁迫增强而线性增加(Holzapfel et al., 2006; Armas et al., 2011)。一项基于喜马拉雅干旱山区垫状植物与草本植物间相互作用的研究发现, 水分可利用性是驱动该地区植物间相互作用的主要因子, 叶碳稳定同位素比值(δ13C)和基于卫星遥感数据的归一化植被指数(NDVI)能很好地指示植物间相互作用的变化, 提出了利用生物因子指示环境胁迫程度和种间关系变化的新思路(Ale et al., 2018a, 2018b)。
2 灌丛促进作用的发生机制
干旱半干旱区灌丛促进作用影响着其冠层下草本植物的物种丰富度和生长, 灌丛促进作用是干旱半干旱生态系统物种多样性形成与维持的重要驱动因子, 主要有如下3个机制。首先, 由于灌木对微环境的改善作用, 其冠层下物种丰富度往往高于冠层外(Holzapfel et al., 2006; Pistón et al., 2016; Iyengar et al., 2017; Zhang et al., 2018a; Parajuli et al., 2021)。其次, 由于灌木提供的异质环境, 使得灌木冠层内外物种组成存在差异, 一些物种只在灌木冠层下或冠层外出现, 进而提高了群落水平的物种丰富度(Armas et al., 2011; Ballantyne & Pickering, 2015; Madrigal-González et al., 2016; Liancourt et al., 2017; Han et al., 2021)。另一方面, 共存的灌木物种塑造了不同的微环境, 进而庇护不同物种在其冠层下存活(Paterno et al., 2016; Pistón et al., 2016)。护卫植物提高群落水平物种丰富度这一结论, 在以垫状植物为护卫植物的高山生态系统中也得到了证实(Cavieres et al., 2014, 2016)。此外, 灌木对其冠层下物种的种子萌发、幼苗存活及生长同样发挥着促进作用(Valiente-Banuet & Ezcurra, 1991; Armas & Pugnaire, 2005; Barchuk et al., 2005; Armas et al., 2011; Zhang et al., 2018a, 2018b)。受益者对护卫植物的反馈作用是评估共存物种长期进化方向和群落动态的重要依据(Armas & Pugnaire, 2005; Michalet et al., 2016), 目前有关草本植物对灌木的反馈作用鲜有报道。Armas和Pugnaire (2005)在西班牙东南部开展的岩蔷薇属灌木Cistus clusii和针茅属植物Stipa tenacissima的去除实验发现, 针茅幼苗对护卫灌木表现为中性作用, 针茅成熟以后, 其获取限制性资源的能力增强, 对护卫灌木表现为竞争作用。Iyengar等(2017)对印度北部豆科灌木变色锦鸡儿开展的去除冠层下草本植物的研究发现, 草本植物的去除提高了灌木下一年的开花数。
干旱半干旱区灌木往往通过增加其冠层下土壤水分、提高土壤养分、减弱辐射、降低温度和抵御动物取食等方式发挥促进作用(图1)。干旱半干旱区灌丛能通过遮阴减弱辐射和降低温度, 有效保护冠层下物种免受强辐射和极端温度的伤害(Valiente-Banuet & Ezcurra, 1991; Maestre et al., 2001; Holmgren et al., 2012; Pistón et al., 2016)。灌丛还能抵御捕食者, 尤其是带刺的灌木物种, 能有效地保护草本物种免受或少受食草动物的取食(Rebollo et al., 2002; Tirado et al., 2015; Lortie et al., 2016; Xie et al., 2017)。例如, 在喜马拉雅山区的Langtang河谷, 研究发现有棱小檗(Berberis angulosa)因其对牲畜取食的抵御, 灌丛内物种丰富度显著高于灌丛外, 一些适口性较好且不耐取食的物种如多星韭(Allium wallichii)、野燕麦(Avena fatua)和云梅花草(Parnassia nubicola)等仅在有棱小檗冠层下出现(Parajuli et al., 2021)。
图1
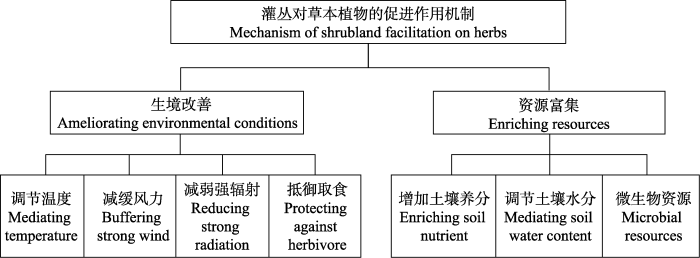
图1
干旱半干旱区灌丛对草本植物的促进作用机制框架图。
Fig. 1
Frame diagram dipicting the machenism of shrubland facilitation on herbs in arid and semi-arid regions.
土壤水分和养分是干旱半干旱区植物存活和生长的主要限制因子, 二者深刻影响着灌丛与草本植物间的相互作用。干旱灌丛通常能发挥“肥岛效应”, 提高冠层下土壤养分含量, 从而促进其下植物的生长(Pugnaire et al., 1996; Wezel et al., 2000; Maestre et al., 2001; Hortal et al., 2013; Noumi et al., 2016; Zhang et al., 2018b; Cai et al., 2020)。灌丛对土壤养分的改善作用在冠层下的不同位置可能存在差异, 研究发现西班牙东南部豆科灌木Retama sphaerocarpa冠层中心的土壤有机质和氮含量高于冠层边缘, 而介于冠层中心与边缘地带之间的土壤矿化速率和有机质分解速率最高(Moro et al., 1997a, 1997b)。作为干旱半干旱区典型的优势灌木物种, 豆科灌木对土壤养分和草本植物的促进作用还与根瘤菌的共生固氮作用密切相关(Ouyang et al., 2016; Zhang et al., 2016; Dovrat et al., 2018; Dovrat & Sheffer, 2019), 豆科灌木能显著提高其冠层下土壤氮含量(Pugnaire et al., 1996; Zhang et al., 2011; Saixiyala et al., 2017)。土壤微生物在灌丛促进作用中的重要性近年备受关注(Wang et al., 2018; Pugnaire et al., 2019; Yu et al., 2021), 例如, Hortal等(2013)发现西班牙干旱区豆科植物Retama sphaerocarpa灌层内外土壤微生物多样性无显著差异, 但灌木冠层下土壤微生物活性、生物量以及与高碳利用效率和固氮等功能相关的类群数量明显高于冠层外。Xie等(2021)发现内蒙古狭叶锦鸡儿(Caragana stenophylla)灌层下土壤线虫以及细菌、真菌群落多样性高于灌层外, 灌木的存在直接提高其下土壤线虫和细菌群落多样性, 而灌木通过提高土壤养分的方式间接促进土壤真菌生长。总的来说, 干旱半干旱区灌丛通过影响其冠层下土壤微生物的丰度、组成和活性, 进而增强土壤养分矿化速率, 改善土壤物理结构, 释放化学信号影响植物物候和功能性状, 增强植物防御取食和病原体的能力, 发挥菌根真菌的补偿作用等, 最终影响灌木冠层下的草本植物(Maestre et al., 2009a; Hortal et al., 2013; Rodríguez-Echeverría et al., 2016)。
灌木与其冠层下草本植物间的相互作用通常是灌木对微环境改善后的综合体现, 学者们普遍采用比较冠层内外各环境要素差异的方法来探究灌木发挥促进作用的机制(Ziffer-Berger et al., 2014; Zhang et al., 2018a), 与此相比, 去除实验能定量比较灌木地上(冠层)和地下(土壤)部分对草本植物的相对影响(Maestre et al., 2003; Armas & Pugnaire, 2005; Michalet et al., 2015; Tirado et al., 2015; Wang et al., 2017)。在青藏高原东北部开展的金露梅(Potentilla fruticosa)去除实验发现, 灌木地上和地下部分对草本物种丰富度的影响分别表现为竞争和中性作用, 净效应表现为竞争作用, 并且不同物种受灌木地上和地下及净效应的影响不同, 进而导致灌木下物种组成存在差异(Wang et al., 2017)。在突尼斯干旱半干旱区, 灌木去除实验的结果发现, 灌木地下部分对草本植物表现为促进作用, 冠层表现为竞争或中性作用, 且随干旱增强, 地下部分的促进作用减弱, 冠层的竞争作用则增强, 因为随干旱增强, 灌木对土壤水分的影响逐渐减弱(Noumi et al., 2016; Chaieb et al., 2021)。Lozano等(2020)通过土块互移实验发现西班牙半干旱区豆科灌木Retama sphaerocarpa冠幅大小和土壤性质对草本植物群落结构的影响是相互独立的, 土壤性质(土壤水分、温度、氮含量和团聚体结构)决定冠层下草本植物的物种组成, 而冠幅大小造成的微气候差异深刻影响着灌层内外的草本植物群落地上生物量和物种丰富度。由此可见, 地下部分(包括土壤养分和土壤微生物)在灌丛与草本植物的相互作用中发挥着重要作用。然而, 目前开展的地下部分对灌丛促进作用沿水分梯度变化的影响的研究还较少, 这可能是今后探索灌丛促进作用沿干旱梯度变化规律研究中值得关注的重点。
3 灌丛促进作用在植被恢复中的应用
退化生态系统的植被恢复是生态修复中的重要措施, 在干旱半干旱区的退化生态系统, 植被恢复常常受到水分可利用性、极端温度和土壤养分等的限制(Hoover et al., 2020; Shackelford et al., 2021)。灌木能改善其冠层下土壤和微气候等条件, 促进相邻物种的存活和生长, 因此被广泛用于干旱半干旱区退化生态系统的植被恢复(Maestre et al., 2001; Young et al., 2005; Padilla & Pugnaire, 2006; Gómez- Aparicio, 2009; Michalet & Pugnaire, 2016; Ibáñez & Rodríguez, 2020; 刘泽正等, 2020)。例如, 在摩洛哥半干旱地区, 先锋灌木Lavandula stoechas改变了土壤微生物群落组成并促成丛枝菌根真菌群落建成, 进而促进Cupressus atlantica幼苗的生长(Duponnois et al., 2011)。在内蒙古高原, 成熟的狭叶锦鸡儿通过改善微生境和防止食草动物取食的方式促进种内幼苗建成(Xie et al., 2017)。在塞浦路斯地中海气候的干旱半干旱区, 深根系灌木Ziziphus lotus能显著提高在其附近生长的Thymbra capitata种群密度(Constantinou et al., 2021)。
值得注意的是, 植被恢复成功与否受到环境胁迫程度、护卫植物和目标恢复物种的物种特性和植物生活史阶段以及土地利用等的综合影响。例如, 在西班牙东南部, Gómez-Aparicio等(2004)比较了16种护卫灌木对目标恢复物种幼苗存活和生长随海拔和坡向的变化, 发现促进作用的强度因护卫灌木和目标恢复物种的物种差异而有所不同, 并且表现出在高海拔强于低海拔、在阴坡强于阳坡的生境差异。Padilla等(2009b)比较了用于干旱生态系统植被恢复的14种灌木在浇水和对照处理下的幼苗成活率, 结果发现: 处于演替中期的物种, 如无叶植物Ephedra fragilisand, C4植物Salsola oppositifolia和豆科植物Coronilla juncea、Genista umbellat和Retama sphaerocarpa的存活率远远高于处于演替后期的物种, 且在不浇水处理下, 演替中期的物种存活率高于演替后期的物种, 研究指出处于演替中期的物种更有利于干旱区植被恢复。Padilla和Pugnaire (2009)在西班牙干旱半干旱区的研究发现, 豆科灌木Retama sphaerocarpa对其冠层下Olea europaea幼树成活率表现为促进作用, 而对Pistacia lentiscus和Ziziphus lotus幼树成活率分别表现为中性和竞争作用。植物生活史阶段对植被恢复的影响主要表现为: 在植被恢复前期, 护卫植物对目标物种发挥促进作用, 随着目标物种种群的建立, 对资源的需求随之增多, 护卫植物和目标物种的关系就可能从促进转为中性或竞争, 导致植被恢复的失败(Miriti, 2006; Suding, 2011; Yang et al., 2014)。放牧是干旱半干旱区灌丛化草原最主要的土地利用方式, 研究发现内蒙古小叶锦鸡儿(Caragana microphylla)灌丛对土壤养分的促进作用随放牧强度增强呈单峰格局, 即在轻度放牧条件下灌丛对土壤养分促进作用最强, 促进作用在过度放牧条件下失效(Cai et al., 2020)。因此, 在植被恢复过程中, 要充分考虑促进和竞争作用的潜在影响, 准确评估促进作用在植被恢复过程中的贡献, 所选择的护卫植物要最大程度避免与目标物种竞争有限的资源, 从而有效促进目标物种的建立和生长(Padilla & Pugnaire, 2006; Ibáñez & Rodríguez, 2020)。
4 展望
干旱半干旱区占全球陆地面积的1/4, 而自20世纪70年代以来, 全球干旱区的面积扩大了2倍以上(Dai et al., 2004; Dai, 2011)。近百年来, 干旱半干旱区气温升高幅度明显高于其他地区(Huang et al., 2012), 气温的持续升高导致干旱半干旱区气候呈现明显的暖干化趋势(IPCC, 2013)。在气候变化和人类活动的共同影响下, 全球大部分干旱半干旱区都发生了灌木密度、盖度或分布面积增加的灌丛化现象(Knapp et al., 2008; Maestre et al., 2009a; Eldridge et al., 2011; Loydi et al., 2013), 研究表明, 过去150年里全球草原大约10%-20%的地区发生了灌丛化(van Auken, 2009), 豆科灌木是灌丛化物种中占比最高的类群(Eldridge et al., 2011)。干旱半干旱区发生的气候变化和灌丛化无疑对区域生态系统结构和功能产生重要影响, 因此, 准确评估灌丛在干旱生态系统中的作用是预测干旱半干旱区物种和生态系统对气候变化的响应, 以及制定干旱生态系统管理和实施生态安全屏障建设工程的关键。我国干旱半干旱区分布面积广, 灌丛和草原是干旱半干旱区重要的生态系统类型, 近年来, 学者们针对我国内蒙古高原以锦鸡儿属灌木为主的灌丛(Zhang et al., 2011, 2018a, 2018b; Xie et al., 2017, 2021; Liu et al., 2021)以及青藏高原东部金露梅灌丛(Xu et al., 2010; Wang et al., 2017, 2018, 2019, 2021)的促进作用开展了较为详细的研究。青藏高原中西部是典型的高寒干旱半干旱区, 灌丛化草原广泛分布, 生态环境脆弱, 目前尚未见到该地区有关灌丛促进作用的相关报道, 因此, 加强推进灌丛促进作用相关研究, 对于青藏高原生态安全屏障建设以及区域植被恢复无疑具有重要的理论和实践意义。
经过近30年的研究, 植物间促进作用已成为生态学研究的重要组成部分。基于现有认识, 我们对未来植物间促进作用, 尤其是干旱半干旱区灌丛促进作用提出以下值得关注的研究内容。
4.1 环境胁迫程度的量化
4.2 明确地下部分对灌丛促进作用的相对贡献
4.3 探索受益者对护卫植物的反馈作用机制
目前, 大量研究已证实护卫植物对其相邻植物的存活和生长普遍发挥着促进作用, 而有关受益物种对护卫植物的反馈研究鲜有报道, 受益物种对护卫植物的影响可能表现为竞争或促进作用。研究发现禾草类植物对高山地区的垫状护卫植物(Michalet et al., 2016)以及干旱半干旱区的护卫灌木(Armas & Pugnaire, 2005)都表现出竞争作用, 原因是与其他类群植物相比, 禾草类对根的投资更多, 且从土壤获取资源的能力更强。在祁连山地区的研究发现杂类草和豆科植物对护卫植物囊种草(Thylacospermum caespitosum)果实的形成有促进作用, 这可能与杂类草能吸引更多传粉者有关(Michalet et al., 2016)。由此可见, 受益者对护卫植物的反馈作用可能受不同植物功能群的调节, 加强相关研究进而明确植物间相互作用及其机制是评估物种间长期进化关系的重要依据。
4.4 综合评估促进作用在植被恢复中的贡献
护卫植物对微环境和相邻植物群落有明显促进作用, 因此, 人为种植护卫植物进而发挥其促进作用被认为是潜在的用于开展退化生态系统植被恢复的有效手段。然而, 护卫植物的促进作用还与环境胁迫程度、物种特性以及土地利用方式等密切相关, 在实际的应用中应综合考虑各方面影响因素, 预先评估促进作用在植被恢复中的贡献。
4.5 加强模拟增水实验和长期定位观测研究
4.6 促进作用与其他研究领域的结合
参考文献
Leaf δ13C as an indicator of water availability along elevation gradients in the dry Himalayas
DOI:10.1016/j.ecolind.2018.07.002 URL [本文引用: 2]
Water shortage drives interactions between cushion and beneficiary species along elevation gradients in dry Himalayas
DOI:10.1002/2017JG004365 URL [本文引用: 2]
Facilitation among plants in alpine environments in the face of climate change
DOI:10.3389/ fpls.2014.00387 [本文引用: 1]
Plant interactions govern population dynamics in a semi-arid plant community
DOI:10.1111/j.1365-2745.2005.01033.x URL [本文引用: 7]
A field test of the stress-gradient hypothesis along an aridity gradient
DOI:10.1111/j.1654-1103.2011.01301.x URL [本文引用: 4]
Effects of experimental rainfall manipulations on Chihuahuan Desert grassland and shrubland plant communities
DOI:10.1007/s00442-012-2552-0
PMID:23263528
[本文引用: 1]

Aridland ecosystems are predicted to be responsive to both increases and decreases in precipitation. In addition, chronic droughts may contribute to encroachment of native C3 shrubs into C4-dominated grasslands. We conducted a long-term rainfall manipulation experiment in native grassland, shrubland and the grass-shrub ecotone in the northern Chihuahuan Desert, USA. We evaluated the effects of 5 years of experimental drought and 4 years of water addition on plant community structure and dynamics. We assessed the effects of altered rainfall regimes on the abundance of dominant species as well as on species richness and subdominant grasses, forbs and shrubs. Nonmetric multidimensional scaling and MANOVA were used to quantify changes in species composition in response to chronic addition or reduction of rainfall. We found that drought consistently and strongly decreased cover of Bouteloua eriopoda, the dominant C4 grass in this system, whereas water addition slightly increased cover, with little variation between years. In contrast, neither chronic drought nor increased rainfall had consistent effects on the cover of Larrea tridentata, the dominant C3 shrub. Species richness declined in shrub-dominated vegetation in response to drought whereas richness increased or was unaffected by water addition or drought in mixed- and grass-dominated vegetation. Cover of subdominant shrubs, grasses and forbs changed significantly over time, primarily in response to interannual rainfall variability more so than to our experimental rainfall treatments. Nevertheless, drought and water addition shifted the species composition of plant communities in all three vegetation types. Overall, we found that B. eriopoda responded strongly to drought and less so to irrigation, whereas L. tridentata showed limited response to either treatment. The strong decline in grass cover and the resistance of shrub cover to rainfall reduction suggest that chronic drought may be a key factor promoting shrub dominance during encroachment into desert grassland.
Shrub facilitation is an important driver of alpine plant community diversity and functional composition
DOI:10.1007/s10531-015-0910-z URL [本文引用: 2]
Effect of shrubs and seasonal variability of rainfall on the establishment of Aspidosperma quebracho-blanco in two edaphically contrasting environments
DOI:10.1111/j.1442-9993.2005.01511.x URL [本文引用: 1]
Aridity preferences alter the relative importance of abiotic and biotic drivers on plant species abundance in global drylands
DOI:10.1111/1365-2745.13006
[本文引用: 1]
1. Disentangling the interplay between species-specific environmental preferences and micro- and macroscale determinants of species abundance within plant communities remains challenging. Most existing studies addressing this issue either lack empirical data regarding species interactions and local abundances or cover a narrow range of environmental conditions. 2. We merged species distribution models and local spatial patterns to investigate the relative importance of key macro- (aridity) and micro(facilitation and competition)scale determinants of plant species abundance along aridity gradients in drylands world-wide. We used information derived from the environmental niches of species to evaluate how species-specific aridity preferences modulate the importance of such factors to drive species relative abundance. 3. Facilitation and aridity preferences were more important than competition to explain species local abundances in global drylands. The specialization of communities (i.e. their compositional shifts from species with a large range of aridity preferences towards only aridity specialists) also modulated the effect of aridity and plant-plant interactions on species abundances. The importance of facilitation to drive species abundances decreased with aridity, as species preferred arid conditions and did not need neighbours to thrive. Instead, competition showed stronger relationships with species abundances under high levels of aridity. As composition became dominated by aridity specialists, the importance of aridity in shaping dryland plant communities did not increase further from moderate to high aridity levels. 4. Synthesis. Our results showed that: (a) the degree of community specialization to aridity mediates the relative importance of plant-plant interactions in determining species abundances and (b) facilitation and competition were more strongly related to species abundance in communities dominated by generalists and specialists, respectively. We observed a shift from facilitation to competition as drivers of species abundances as aridity increases in global drylands. Our findings also pave the way to develop more robust predictions about the consequences of ongoing climate change on the assemblage of plant communities in drylands, the largest terrestrial biome.
Interspecific interactions among high marsh perennials in a New England salt marsh
DOI:10.2307/1938908 URL [本文引用: 1]
Positive interactions in communities
DOI:10.1016/0169-5347(94)90088-4 URL [本文引用: 2]
Physical stress and positive associations among marsh plants
DOI:10.1086/285681 URL [本文引用: 1]
Competition and facilitation in marsh plants
DOI:10.1086/285567 URL [本文引用: 1]
A portfolio effect of shrub canopy height on species richness in both stressful and competitive environments
DOI:10.1111/1365-2435.12458 URL [本文引用: 1]
Plant-plant interactions and environmental change
Natural systems are being subjected to unprecedented rates of change and unique pressures from a combination of anthropogenic environmental change drivers. Plant-plant interactions are an important part of the mechanisms governing the response of plant species and communities to these drivers. For example, competition plays a central role in mediating the impacts of atmospheric nitrogen deposition, increased atmospheric carbon dioxide concentrations, climate change and invasive nonnative species. Other plant-plant interaction processes are also being recognized as important factors in determining the impacts of environmental change, including facilitation and evolutionary processes associated with plant-plant interactions. However, plant-plant interactions are not the only factors determining the response of species and communities to environmental change drivers - their activity must be placed within the context of the wide range of factors that regulate species, communities and ecosystems. A major research challenge is to understand when plant-plant interactions play a key role in regulating the impact of environmental change drivers, and the type of role that plant-plant interactions play. Although this is a considerable challenge, some areas of current research may provide the starting point to achieving these goals, and should be pursued through large-scale, integrated, multisite experiments.
Facilitation in plant communities: the past, the present, and the future
Inclusion of facilitation into ecological theory
DOI:10.1016/S0169-5347(02)00045-9 URL [本文引用: 2]
Does the stress-gradient hypothesis hold water? Disentangling spatial and temporal variation in plant effects on soil moisture in dryland systems
DOI:10.1111/1365-2435.12592 URL [本文引用: 1]
Alpine cushion plants inhibit the loss of phylogenetic diversity in severe environments
DOI:10.1111/ele.12070
PMID:23346919
[本文引用: 2]
Biotic interactions can shape phylogenetic community structure (PCS). However, we do not know how the asymmetric effects of foundation species on communities extend to effects on PCS. We assessed PCS of alpine plant communities around the world, both within cushion plant foundation species and adjacent open ground, and compared the effects of foundation species and climate on alpha (within-microsite), beta (between open and cushion) and gamma (open and cushion combined) PCS. In the open, alpha PCS shifted from highly related to distantly related with increasing potential productivity. However, we found no relationship between gamma PCS and climate, due to divergence in phylogenetic composition between cushion and open sub-communities in severe environments, as demonstrated by increasing phylo-beta diversity. Thus, foundation species functioned as micro-refugia by facilitating less stress-tolerant lineages in severe environments, erasing a global productivity - phylogenetic diversity relationship that would go undetected without accounting for this important biotic interaction.© 2013 Blackwell Publishing Ltd/CNRS.
The fertile island effect collapses under extreme overgrazing: evidence from a shrub-encroached grassland
DOI:10.1007/s11104-020-04426-2 URL [本文引用: 2]
Positive interactions among plants
DOI:10.1007/BF02912621 URL [本文引用: 1]
Positive interactions among alpine plants increase with stress
DOI:10.1038/nature00812 URL [本文引用: 2]
Positive interactions between alpine plant species and the nurse cushion plant Laretia acaulis do not increase with elevation in the Andes of central Chile
DOI:10.1111/j.1469-8137.2005.01573.x URL [本文引用: 1]
Facilitative plant interactions and climate simultaneously drive alpine plant diversity
DOI:10.1111/ele.12217
PMID:24238015
[本文引用: 2]
Interactions among species determine local-scale diversity, but local interactions are thought to have minor effects at larger scales. However, quantitative comparisons of the importance of biotic interactions relative to other drivers are rarely made at larger scales. Using a data set spanning 78 sites and five continents, we assessed the relative importance of biotic interactions and climate in determining plant diversity in alpine ecosystems dominated by nurse-plant cushion species. Climate variables related with water balance showed the highest correlation with richness at the global scale. Strikingly, although the effect of cushion species on diversity was lower than that of climate, its contribution was still substantial. In particular, cushion species enhanced species richness more in systems with inherently impoverished local diversity. Nurse species appear to act as a 'safety net' sustaining diversity under harsh conditions, demonstrating that climate and species interactions should be integrated when predicting future biodiversity effects of climate change. © 2013 John Wiley & Sons Ltd/CNRS.
Facilitation among plants as an insurance policy for diversity in alpine communities
DOI:10.1111/1365-2435.12545 URL [本文引用: 3]
Shift from short-term competition to facilitation with drought stress is due to a decrease in long-term facilitation
DOI:10.1111/oik.07528 URL [本文引用: 3]
Facilitation and competition on gradients in alpine plant communities
DOI:10.1890/0012-9658(2001)082[3295:FACOGI]2.0.CO;2 URL [本文引用: 1]
Balance between facilitation and resource competition determines biomass-density relationships in plant populations
DOI:10.1111/j.1461-0248.2008.01228.x URL [本文引用: 1]
The possible role of Ziziphus lotus as an ecosystem engineer in semiarid landscapes
DOI:10.1016/j.jaridenv.2021.104614 [本文引用: 1]
Drought under global warming: a review
DOI:10.1002/wcc.81 URL [本文引用: 1]
A global dataset of Palmer Drought Severity Index for 1870-2002: relationship with soil moisture and effects of surface warming
DOI:10.1175/JHM-386.1 URL [本文引用: 1]
Depth of water acquisition by invading shrubs and resident herbs in a Sierra Nevada meadow
DOI:10.1007/s11104-005-4453-z URL [本文引用: 1]
Do changes in rainfall patterns affect semiarid annual plant communities?
DOI:10.1111/j.1654-1103.2009.05680.x URL [本文引用: 1]
Drought-adapted plants dramatically downregulate dinitrogen fixation: evidences from Mediterranean legume shrubs
DOI:10.1111/1365-2745.12940 URL [本文引用: 1]
Symbiotic dinitrogen fixation is seasonal and strongly regulated in water-limited environments
DOI:10.1111/nph.15526
PMID:30299536
[本文引用: 1]
Plants, especially perennials, growing in drylands and seasonally dry ecosystems are uniquely adapted to dry conditions. Legume shrubs and trees, capable of symbiotic dinitrogen (N ) fixation, often dominate in drylands. However, the strategies that allow symbiotic fixation in these ecosystems, and their influence on the nitrogen cycle, are largely unresolved. We evaluated the climatic, biogeochemical and ontogenetic factors influencing nitrogen fixation in an abundant Mediterranean legume shrub, Calicotome villosa. We measured nodulation, fixation rate, nitrogen allocation and soil biogeochemistry in three field sites over a full year. A controlled experiment evaluated differences in plant regulation of fixation as a function of soil nutrient availability and seedling and adult developmental stages. We found a strong seasonal pattern, shifting between high fixation rates during the rainy season at flowering and seed-set times to almost none in the rainless season. Under controlled conditions, plants downregulated fixation in response to soil nitrogen availability, but this response was stronger in seedlings than in adult shrubs. Finally, we did not find elevated soil nitrogen under N -fixing shrubs. We conclude that seasonal nitrogen fixation, regulation of fixation, and nitrogen conservation are key adaptations influencing the dominance of dryland legumes in the community, with broader consequences on the ecosystem nitrogen cycle.© 2018 The Authors. New Phytologist © 2018 New Phytologist Trust.
Nurse shrubs increased the early growth of Cupressus seedlings by enhancing belowground mutualism and soil microbial activity
Impacts of shrub encroachment on ecosystem structure and functioning: towards a global synthesis
DOI:10.1111/j.1461-0248.2011.01630.x
PMID:21592276
[本文引用: 2]
Encroachment of woody plants into grasslands has generated considerable interest among ecologists. Syntheses of encroachment effects on ecosystem processes have been limited in extent and confined largely to pastoral land uses or particular geographical regions. We used univariate analyses, meta-analysis and structural equation modelling to test the propositions that (1) shrub encroachment does not necessarily lead to declines in ecosystem functions and (2) shrub traits influence the functional outcome of encroachment. Analyses of 43 ecosystem attributes from 244 case studies worldwide showed that some attributes consistently increased with encroachment (e.g. soil C, N), and others declined (e.g. grass cover, pH), but most exhibited variable responses. Traits of shrubs were associated with significant, though weak, structural and functional outcomes of encroachment. Our review revealed that encroachment had mixed effects on ecosystem structure and functioning at global scales, and that shrub traits influence the functional outcome of encroachment. Thus, a simple designation of encroachment as a process leading to functionally, structurally or contextually degraded ecosystems is not supported by a critical analysis of existing literature. Our results highlight that the commonly established link between shrub encroachment and degradation is not universal.© 2011 Blackwell Publishing Ltd/CNRS and State Government of NSW.
The role of plant interactions in the restoration of degraded ecosystems: a meta-analysis across life-forms and ecosystems
DOI:10.1111/j.1365-2745.2009.01573.x URL [本文引用: 1]
Applying plant facilitation to forest restoration: a meta-analysis of the use of shrubs as nurse plants
DOI:10.1890/03-5084 URL [本文引用: 2]
Inter- and under-canopy soil water, leaf-level and whole-plant gas exchange dynamics of a semi-arid perennial C4 grass
Soil and shrub differentially determine understorey herbaceous plant richness and abundance in a semi-arid riparian meadow
DOI:10.1007/s42974-021-00041-y URL [本文引用: 1]
Deviation from niche optima affects the nature of plant-plant interactions along a soil acidity gradient
DOI:10.1098/rsbl.2015.0925 [本文引用: 1]
Global shifts towards positive species interactions with increasing environmental stress
DOI:10.1111/ele.12080
PMID:23363430
[本文引用: 2]
The study of positive species interactions is a rapidly evolving field in ecology. Despite decades of research, controversy has emerged as to whether positive and negative interactions predictably shift with increasing environmental stress as hypothesised by the stress-gradient hypothesis (SGH). Here, we provide a synthesis of 727 tests of the SGH in plant communities across the globe to examine its generality across a variety of ecological factors. Our results show that plant interactions change with stress through an outright shift to facilitation (survival) or a reduction in competition (growth and reproduction). In a limited number of cases, plant interactions do not respond to stress, but they never shift towards competition with stress. These findings are consistent across stress types, plant growth forms, life histories, origins (invasive vs. native), climates, ecosystems and methodologies, though the magnitude of the shifts towards facilitation with stress is dependent on these factors. We suggest that future studies should employ standardised definitions and protocols to test the SGH, take a multi-factorial approach that considers variables such as plant traits in addition to stress, and apply the SGH to better understand how species and communities will respond to environmental change.© 2013 Blackwell Publishing Ltd/CNRS.
Non-linear effects of drought under shade: reconciling physiological and ecological models in plant communities
DOI:10.1007/s00442-011-2196-5
PMID:22083284
[本文引用: 1]
The combined effects of shade and drought on plant performance and the implications for species interactions are highly debated in plant ecology. Empirical evidence for positive and negative effects of shade on the performance of plants under dry conditions supports two contrasting theoretical models about the role of shade under dry conditions: the trade-off and the facilitation hypotheses. We performed a meta-analysis of field and greenhouse studies evaluating the effects of drought at two or more irradiance levels on nine response variables describing plant physiological condition, growth, and survival. We explored differences in plant response across plant functional types, ecosystem types and methodological approaches. The data were best fit using quadratic models indicating a humped-back shape response to drought along an irradiance gradient for survival, whole plant biomass, maximum photosynthetic capacity, stomatal conductance and maximal photochemical efficiency. Drought effects were ameliorated at intermediate irradiance, becoming more severe at higher or lower light levels. This general pattern was maintained when controlling for potential variations in the strength of the drought treatment among light levels. Our quantitative meta-analysis indicates that dense shade ameliorates drought especially among drought-intolerant and shade-tolerant species. Wet tropical species showed larger negative effects of drought with increasing irradiance than semiarid and cold temperate species. Non-linear responses to irradiance were stronger under field conditions than under controlled greenhouse conditions. Non-linear responses to drought along the irradiance gradient reconciliate opposing views in plant ecology, indicating that facilitation is more likely within certain range of environmental conditions, fading under deep shade, especially for drought-tolerant species.
Strong facilitation in mild environments: the stress gradient hypothesis revisited
DOI:10.1111/j.1365-2745.2010.01709.x URL [本文引用: 1]
Annual plant-shrub interactions along an aridity gradient
DOI:10.1016/j.baae.2005.08.003 URL [本文引用: 3]
Effects of biodiversity on ecosystem functioning: a consensus of current knowledge
DOI:10.1890/04-0922 URL [本文引用: 1]
Traversing the wasteland: a framework for assessing ecological threats to drylands
DOI:10.1093/biosci/biz126 URL [本文引用: 1]
Soil microbial community under a nurse-plant species changes in composition, biomass and activity as the nurse grows
DOI:10.1016/j.soilbio.2013.04.018 URL [本文引用: 3]
Enhanced cold-season warming in semi-arid regions
Understanding neighborhood effects to increase restoration success of woody plant communities
DOI:10.1002/eap.2098 [本文引用: 2]
A dominant dwarf shrub increases diversity of herbaceous plant communities in a Trans-Himalayan rangeland
DOI:10.1007/s11258-017-0734-x URL [本文引用: 4]
Linking patterns and processes in alpine plant communities: a global study
DOI:10.1890/04-1926 URL [本文引用: 1]
Shrub encroachment in North American grasslands: shifts in growth form dominance rapidly alters control of ecosystem carbon inputs
DOI:10.1111/j.1365-2486.2007.01512.x URL [本文引用: 1]
Stress tolerance and competitive-response ability determine the outcome of biotic interactions
DOI:10.1890/04-1398 URL [本文引用: 2]
Community-scale effects and strain: facilitation beyond conspicuous patterns
DOI:10.1111/1365-2745.13458 URL [本文引用: 2]
SGH: stress or strain gradient hypothesis? Insights from an elevation gradient on the roof of the world
DOI:10.1093/aob/mcx037
PMID:28444363
[本文引用: 5]
The stress gradient hypothesis (SGH), the view that competition prevails in undisturbed and productive environments, and shifts to facilitation in disturbed or stressful environments, has become a central paradigm in ecology. However, an alternative view proposes that the relationship between biotic interactions and environmental severity should be unimodal instead of monotonic. Possible causes of discrepancies between these two views were examined in the high elevation desert of the arid Trans-Himalayas.A putative nurse species and its associated plant community was surveyed over its entire elevation range, spanning from alpine to desert vegetation belts. The results were analysed at the community level (vegetation cover and species richness), considering the distinction between the intensity and the importance of biotic interactions. Interactions at the species level (pairwise interactions) were also considered, i.e. the variation of biotic interactions within the niche of a species, for which the abundance (species cover) and probability of occurrence (presence/absence) for the most widespread species along the gradient were distinguished.Overall, facilitation was infrequent in our study system; however, it was observed for the two most widespread species. At the community level, the intensity and importance of biotic interactions showed a unimodal pattern. The departure from the prediction of the SGH happened abruptly where the nurse species entered the desert vegetation belt at the lowest elevation. This abrupt shift was attributed to the turnover of species with contrasting tolerances. At the species level, however, facilitation increased consistently as the level of stress increases and individuals deviate from their optimum (increasing strain).While the stress gradient hypothesis was not supported along our elevation gradient at the community level, the strain gradient hypothesis, considering how species perceive the ambient level of stress and deviate from their optimum, provided a parsimonious explanation for the outcome of plant-plant interactions at both scales.© The Author 2017. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved. For Permissions, please email: journals.permissions@oup.com
A nurse shrub species helps associated herbaceous plants by preventing shade-induced evaporation in a desert ecosystem
DOI:10.1002/ldr.3831 URL [本文引用: 2]
Advances and prospects of incorporating facilitation in ecological restoration
促进作用理论在生态修复中的应用进展与发展趋势
Re-analysis of meta-analysis: support for the stress-gradient hypothesis
DOI:10.1111/j.1365-2745.2005.01066.x URL [本文引用: 1]
Functional assessment of animal interactions with shrub-facilitation complexes: a formal synthesis and conceptual framework
DOI:10.1111/1365-2435.12530 URL [本文引用: 2]
Plant life history stage and nurse age change the development of ecological networks in an arid ecosystem
DOI:10.1111/oik.05199 URL [本文引用: 1]
Effects of litter on seedling establishment in natural and semi-natural grasslands: a meta-analysis
DOI:10.1111/1365-2745.12033 URL [本文引用: 1]
Complementarity in nurse plant systems: soil drives community composition while microclimate enhances productivity and diversity
DOI:10.1007/s11104-020-04503-6 URL [本文引用: 1]
Shrub-ephemeral plants interactions in semiarid north-central Chile: Is the nurse plant syndrome manifested at the community level?
DOI:10.1016/j.jaridenv.2015.08.001 URL [本文引用: 1]
Positive, negative, and net effects in grass-shrub interactions in Mediterranean semiarid grasslands
DOI:10.1890/02-0635 URL [本文引用: 2]
Potential for using facilitation by grasses to establish shrubs on a semiarid degraded steppe
DOI:10.1890/1051-0761(2001)011[1641:PFUFBG]2.0.CO;2 URL [本文引用: 4]
Shrub encroachment can reverse desertification in semi-arid Mediterranean grasslands
DOI:10.1111/j.1461-0248.2009.01352.x URL [本文引用: 2]
Refining the stress-gradient hypothesis for competition and facilitation in plant communities
DOI:10.1111/j.1365-2745.2008.01476.x URL [本文引用: 5]
Do positive interactions increase with abiotic stress? A test from a semi-arid steppe
Is the change of plant-plant interactions with abiotic stress predictable? A meta-analysis of field results in arid environments
DOI:10.1111/j.1365-2745.2005.01017.x URL [本文引用: 2]
The stress- gradient hypothesis does not fit all relationships between plant-plant interactions and abiotic stress: further insights from arid environments
DOI:10.1111/j.1365-2745.2005.01089.x URL [本文引用: 1]
What does the stress- gradient hypothesis predict? Resolving the discrepancies
DOI:10.1111/j.1600-0706.2010.18375.x URL [本文引用: 2]
Spatial and temporal aridity gradients provide poor proxies for plant-plant interactions under climate change: a large-scale experiment
DOI:10.1111/1365-2435.12599 URL [本文引用: 1]
Do biotic interactions shape both sides of the humped-back model of species richness in plant communities?
A humped-back relationship between species richness and community biomass has frequently been observed in plant communities, at both local and regional scales, although often improperly called a productivity-diversity relationship. Explanations for this relationship have emphasized the role of competitive exclusion, probably because at the time when the relationship was first examined, competition was considered to be the significant biotic filter structuring plant communities. However, over the last 15 years there has been a renewed interest in facilitation and this research has shown a clear link between the role of facilitation in structuring communities and both community biomass and the severity of the environment. Although facilitation may enlarge the realized niche of species and increase community richness in stressful environments, there has only been one previous attempt to revisit the humped-back model of species richness and to include facilitative processes. However, to date, no model has explored whether biotic interactions can potentially shape both sides of the humped-back model for species richness commonly detected in plant communities. Here, we propose a revision of Grime's original model that incorporates a new understanding of the role of facilitative interactions in plant communities. In this revised model, facilitation promotes diversity at medium to high environmental severity levels, by expanding the realized niche of stress-intolerant competitive species into harsh physical conditions. However, when environmental conditions become extremely severe the positive effects of the benefactors wane (as supported by recent research on facilitative interactions in extremely severe environments) and diversity is reduced. Conversely, with decreasing stress along the biomass gradient, facilitation decreases because stress-intolerant species become able to exist away from the canopy of the stress-tolerant species (as proposed by facilitation theory). At the same time competition increases for stress-tolerant species, reducing diversity in the most benign conditions (as proposed by models of competition theory). In this way our inclusion of facilitation into the classic model of plant species diversity and community biomass generates a more powerful and richer predictive framework for understanding the role of plant interactions in changing diversity. We then use our revised model to explain both the observed discrepancies between natural patterns of species richness and community biomass and the results of experimental studies of the impact of biodiversity on the productivity of herbaceous communities. It is clear that explicit consideration of concurrent changes in stress-tolerant and competitive species enhances our capacity to explain and interpret patterns in plant community diversity with respect to environmental severity.
Disentangling direct and indirect effects of a legume shrub on its understorey community
DOI:10.1111/oik.01819 URL [本文引用: 3]
Facilitation in communities: underlying mechanisms, community and ecosystem implications
DOI:10.1111/1365-2435.12602 URL [本文引用: 7]
Beneficiary feedback effects on alpine cushion benefactors become more negative with increasing cover of graminoids and in dry conditions
DOI:10.1111/1365-2435.12507 URL [本文引用: 2]
Ontogenetic shift from facilitation to competition in a desert shrub
DOI:10.1111/j.1365-2745.2006.01138.x URL [本文引用: 1]
Mechanisms of interaction between a leguminous shrub and its understorey in a semi-arid environment
DOI:10.1111/j.1600-0587.1997.tb00360.x URL [本文引用: 2]
Effect of the canopy of Retama sphaerocarpa on its understorey in a semiarid environment
DOI:10.1046/j.1365-2435.1997.00106.x URL [本文引用: 1]
The relative contribution of short-term versus long-term effects in shrub-understory species interactions under arid conditions
DOI:10.1007/s00442-015-3482-4
PMID:26527462
[本文引用: 4]
Plant-plant interactions (competition and facilitation) in terrestrial ecosystems include: (1) short-term effects, primarily quantified with experimental removals; and (2) long-term effects, mostly due to soil weathering processes, primarily quantified with observational methods. It has been argued that these effects are likely to vary in contrasting directions with increasing drought stress in arid systems. However, few studies have used appropriate methodology to assess both types of effects and their variation across nurse species and environmental conditions, in particular in arid systems. This knowledge is crucial for predicting variation in the mediating role of facilitation with climate change and assessing the importance of nurse effects in ecological restoration. In the arid climate of central-south Tunisia, understory species' biomass, abundance and composition and soil parameters were compared in shrub-control, shrub-removed and open areas for three shrub species and in two habitats with contrasting soil moisture conditions. Long-term effects were dominant, positive for all three shrub species and associated with increasing nutrient content in shrub patches. Short-term effects, mainly related to water consumption, were weaker, mostly negative and dependent on shrub species. Additionally, long-term effects were less positive and short-term effects more negative in the dry habitat than in the wet habitat. Our study provides evidence of the primary influence of positive (facilitative) long-term effects in this arid system. However, the net effects of shrubs could be less beneficial for other species with increasing aridity under climate change, due to both a decrease in positive long-term effects and an increase in negative short-term effects.
The shift from plant-plant facilitation to competition under severe water deficit is spatially explicit
DOI:10.1002/ece3.2875
PMID:28405307
[本文引用: 3]
The stress-gradient hypothesis predicts a higher frequency of facilitative interactions as resource limitation increases. Under severe resource limitation, it has been suggested that facilitation may revert to competition, and identifying the presence as well as determining the magnitude of this shift is important for predicting the effect of climate change on biodiversity and plant community dynamics. In this study, we perform a meta-analysis to compare temporal differences of species diversity and productivity under a nurse plant () with varying annual rainfall quantity to test the effect of water limitation on facilitation. Furthermore, we assess spatial differences in the herbaceous community under nurse plants during a year with below-average rainfall. We found evidence that severe rainfall deficit reduced species diversity and plant productivity under nurse plants relative to open areas. Our results indicate that the switch from facilitation to competition in response to rainfall quantity is nonlinear. The magnitude of this switch depended on the aspect around the nurse plant. Hotter south aspects under nurse plants resulted in negative effects on beneficiary species, while the north aspect still showed facilitation. Combined, these results emphasize the importance of spatial heterogeneity under nurse plants for mediating species loss under reduced precipitation, as predicted by future climate change scenarios. However, the decreased water availability expected under climate change will likely reduce overall facilitation and limit the role of nurse plants as refugia, amplifying biodiversity loss.
Symbiotic nitrogen fixation and interspecific transfer by Caragana microphylla in a temperate grassland with 15N dilution technique
DOI:10.1016/j.apsoil.2016.08.011 URL [本文引用: 1]
Variability in amount and frequency of water supply affects roots but not growth of arid shrubs
DOI:10.1007/s11258-009-9589-0 URL [本文引用: 1]
Rethinking species selection for restoration of arid shrublands
DOI:10.1016/j.baae.2009.03.003 URL [本文引用: 1]
The role of nurse plants in the restoration of degraded environments
DOI:10.1890/1540-9295(2006)004[0196:TRONPI]2.0.CO;2 URL [本文引用: 3]
Rooting depth and soil moisture control Mediterranean woody seedling survival during drought
DOI:10.1111/j.1365-2435.2007.01267.x URL [本文引用: 1]
Species identity and water availability determine establishment success under the canopy of Retama sphaerocarpa shrubs in a dry environment
DOI:10.1111/j.1526-100X.2008.00460.x URL [本文引用: 1]
Facilitation by a dwarf shrub enhances plant diversity of human-valued species at high elevations in the Himalayas of Nepal
DOI:10.1016/j.baae.2021.04.004 URL [本文引用: 4]
Species- specific facilitation, ontogenetic shifts and consequences for plant community succession
DOI:10.1111/jvs.12382 URL [本文引用: 2]
Contribution of co-occurring shrub species to community richness and phylogenetic diversity along an environmental gradient
DOI:10.1016/j.ppees.2016.02.002 URL [本文引用: 4]
The role of hydraulic lift on seedling establishment under a nurse plant species in a semi-arid environment
DOI:10.1016/j.ppees.2011.05.002 URL [本文引用: 1]
Facilitation and succession under the canopy of a leguminous shrub, Retama sphaerocarpa, in a semi-arid environment in south-east Spain
DOI:10.2307/3546339 URL [本文引用: 3]
Species interactions involving cushion plants in high-elevation environments under a changing climate
DOI:10.7818/ECOS.2186 [本文引用: 1]
Changes in plant interactions along a gradient of environmental stress
DOI:10.1034/j.1600-0706.2001.930104.x URL [本文引用: 1]
Climate change effects on plant-soil feedbacks and consequences for biodiversity and functioning of terrestrial ecosystems
DOI:10.1126/sciadv. aaz1834 [本文引用: 1]
No evidence of facilitation collapse in the Tibetan Plateau
DOI:10.1111/jvs.12233 URL [本文引用: 1]
The role of a spiny plant refuge in structuring grazed shortgrass steppe plant communities
DOI:10.1034/j.1600-0706.2002.980106.x URL [本文引用: 1]
Hydraulic lift: substantial nocturnal water transport between soil layers by Artemisia tridentata roots
DOI:10.1007/BF00379405
PMID:28311963
[本文引用: 1]
Diel soil water potential fluctuations reflected daytime depletion and nocturnal resupply of water in upper soil layers. Transpiration suppression experiments demonstrated that water absorption by roots caused the daytime depletion. The soil water potential data and experimental results suggest that at night water absorbed from moist soil by deeper roots is transported to and lost from roots into drier upper soil layers. The deeper roots appear to absorb and transport water both day and night. Implications for the efficiency of deep roots and water storage, nutrient uptake and water parasitism in upper soil layers are discussed.
Influence of soil microbiota in nurse plant systems
DOI:10.1111/1365-2435.12594 URL [本文引用: 1]
Facilitation by a spiny shrub on a rhizomatous clonal herbaceous in thicketization-grassland in Northern China: increased soil resources or shelter from herbivores
DOI:10.3389/fpls.2017.00809 [本文引用: 1]
Variability in functional traits mediates plant interactions along stress gradients
DOI:10.1111/1365-2745.12062 URL [本文引用: 1]
Drivers of seedling establishment success in dryland restoration efforts
Plant-plant interactions, environmental gradients and plant diversity: a global synthesis of community-level studies
Previous syntheses on the effects of environmental conditions on the outcome of plant-plant interactions summarize results from pairwise studies. However, the upscaling to the community-level of such studies is problematic because of the existence of multiple species assemblages and species-specific responses to both the environmental conditions and the presence of neighbors. We conducted the first global synthesis of community-level studies from harsh environments, which included data from 71 alpine and 137 dryland communities. Here we: i) test how important are facilitative interactions as a driver of community structure, ii) evaluate whether the frequency of positive plant-plant interactions across differing environmental conditions and habitats is predictable, and iii) assess whether thresholds in the response of plant-plant interactions to environmental gradients exists between "moderate" and "extreme" stress levels. We also used those community-level studies performed across gradients of at least three points to evaluate how the average environmental conditions, the length of the gradient studied, and the number of points sampled across such gradient affect the form and strength of the facilitation-environment relationship. Over 25% of the species present were more spatially associated to nurse plants than expected by chance in both alpine and dryland areas, illustrating the high importance of positive plant-plant interactions for the maintenance of plant diversity. Facilitative interactions were more frequent, and more related to environmental conditions, in alpine than in dryland areas, perhaps because drylands are generally characterized by a larger variety of environmental stress factors and plant functional traits. The frequency of facilitative interactions in alpine communities peaked at 1000 mm of annual rainfall, and globally decreased with elevation. The frequency of positive interactions in dryland communities decreased globally with water scarcity or temperature annual range. Positive facilitation-drought stress relationships are more likely in shorter regional gradients, but these relationships are obscured in regions with a greater species turnover or with complex environmental gradients. By showing the different climatic drivers and behaviors of plant-plant interactions in dryland and alpine areas, our results will improve predictions regarding the effect of facilitation on the assembly of plant communities and their response to changes in environmental conditions.
Functional traits determine plant co-occurrence more than environment or evolutionary relatedness in global drylands
Plant-plant interactions are driven by environmental conditions, evolutionary relationships (ER) and the functional traits of the plants involved. However, studies addressing the relative importance of these drivers are rare, but crucial to improve our predictions of the effects of plant-plant interactions on plant communities and of how they respond to differing environmental conditions. To analyze the relative importance of -and interrelationships among- these factors as drivers of plant-plant interactions, we analyzed perennial plant co-occurrence at 106 dryland plant communities established across rainfall gradients in nine countries. We used structural equation modeling to disentangle the relationships between environmental conditions (aridity and soil fertility), functional traits extracted from the literature, and ER, and to assess their relative importance as drivers of the 929 pairwise plant-plant co-occurrence levels measured. Functional traits, specifically facilitated plants' height and nurse growth form, were of primary importance, and modulated the effect of the environment and ER on plant-plant interactions. Environmental conditions and ER were important mainly for those interactions involving woody and graminoid nurses, respectively. The relative importance of different plant-plant interaction drivers (ER, functional traits, and the environment) varied depending on the region considered, illustrating the difficulty of predicting the outcome of plant-plant interactions at broader spatial scales. In our global-scale study on drylands, plant-plant interactions were more strongly related to functional traits of the species involved than to the environmental variables considered. Thus, moving to a trait-based facilitation/competition approach help to predict that: 1) positive plant-plant interactions are more likely to occur for taller facilitated species in drylands, and 2) plant-plant interactions within woody-dominated ecosystems might be more sensitive to changing environmental conditions than those within grasslands. By providing insights on which species are likely to better perform beneath a given neighbour, our results will also help to succeed in restoration practices involving the use of nurse plants.
Moving forward on facilitation research: response to changing environments and effects on the diversity, functioning and evolution of plant communities
DOI:10.1111/brv.12110 URL [本文引用: 4]
Toward an era of restoration in ecology: successes, failures, and opportunities ahead
DOI:10.1146/annurev-ecolsys-102710-145115 URL [本文引用: 1]
Temporal environmental variation tips the balance between facilitation and interference in desert plants
DOI:10.1890/0012-9658(2000)081[1544:TEVTTB]2.0.CO;2 URL [本文引用: 3]
Mutual positive effects between shrubs in an arid ecosystem
DOI:10.1038/srep14710 [本文引用: 3]
Shade as a cause of the association between the cactus Neobuxbaumia tetetzo and the nurse plant Mimosa luisana in the Tehuacan Valley, Mexico
DOI:10.2307/2261091 URL [本文引用: 2]
Causes and consequences of woody plant encroachment into western North American grasslands
DOI:10.1016/j.jenvman.2009.04.023
PMID:19501450
[本文引用: 1]
As woody plants encroach into grasslands, grass biomass, density and cover decline as wood plant biomass, density and cover increase. There is also a shift in location of the biomass from mostly belowground in the grasslands to aboveground in the woodlands. In addition, species richness and diversity change as herbaceous species are replaced by woody species. This is not a new phenomenon, but has been going on continually as the climate of the Planet has changed. However, in the past 160 years the changes have been unparalleled. The process is encroachment not invasion because woody species that have been increasing in density are native species and have been present in these communities for thousands of years. These indigenous or native woody species have increased in density, cover and biomass because of changes in one or more abiotic or biotic factors or conditions. Woody species that have increased in density and cover are not the cause of the encroachment, but the result of changes of other factors. Globally, the orbit of the Earth is becoming more circular and less elliptical, causing moderation of the climate. Additional global climate changing factors including elevated levels of CO2 and parallel increases in temperature are background factors and probably not the principal causes directing the current wave of encroachment. There is probably not a single reason for encroachment, but a combination of factors that are difficult to disentangle. The prime cause of the current and recent encroachment appears to be high and constant levels of grass herbivory by domestic animals. This herbivory reduces fine fuel with a concomitant reduction in fire frequency or in some cases a complete elimination of fire from these communities. Conditions would now favor the woody plants over the grasses. Reduced grass competition, woody plant seed dispersal and changes in animal populations seem to modify the rate of encroachment rather than being the cause. High concentrations of atmospheric CO2 are not required to explain current woody plant encroachment. Changes in these grassland communities will continue into the future but the specifics are difficult to predict. Density, cover and species composition will fluctuate and will probably continue to change. Increased levels of anthropogenic soil nitrogen suggest replacement of many legumes by other woody species. Modification and perhaps reversal of the changes in these former grassland communities will be an arduous, continuing and perhaps impossible management task.
Contrasting understorey species responses to the canopy and root effects of a dominant shrub drive community composition
DOI:10.1111/jvs.12565 URL [本文引用: 3]
Stature of dependent forbs is more related to the direct and indirect above- and below-ground effects of a subalpine shrub than are foliage traits
DOI:10.1111/jvs.12739 URL [本文引用: 1]
Direct and indirect facilitation affect community productivity through changes in functional diversity in an alpine system
DOI:10.1093/aob/mcaa170
PMID:32940643
[本文引用: 2]
Facilitation is an important ecological process for plant community structure and functional composition. Although direct facilitation has accrued most of the evidence so far, indirect facilitation is ubiquitous in nature and it has an enormous potential to explain community structuring. In this study, we assess the effect of direct and indirect facilitation on community productivity via taxonomic and functional diversity.In an alpine community on the Tibetan Plateau, we manipulated the presence of the shrub Dasiphora fruticosa and graminoids in a fenced meadow and a grazed meadow to quantify the effects of direct and indirect facilitation. We measured four plant traits: height, lateral spread, specific leaf area (SLA) and leaf dry matter content (LDMC) of forbs; calculated two metrics of functional diversity [range of trait and community-weighted mean (CWM) of trait]; and assessed the responses of functional diversity to shrub facilitation. We used structural equation modelling to explore how shrubs directly and indirectly drove community productivity via taxonomic diversity and functional diversity.We found stronger effects from herbivore-mediated indirect facilitation than direct facilitation on productivity and taxonomic diversity, regardless of the presence of graminoids. For functional diversity, the range and CWM of height and SLA, rather than lateral spread and LDMC, generally increased due to direct and indirect facilitation. Moreover, we found that the range of traits played a primary role over taxonomic diversity and CWM of traits in terms of shrub effects on community productivity.Our study reveals that the mechanism of shrub direct and indirect facilitation of community productivity in this alpine community is expanding the realized niche (i.e. expanding range of traits). Our findings indicate that facilitators might increase trait dispersion in the local community, which could alleviate the effect of environmental filters on trait values in harsh environments, thereby contributing to ecosystem functioning.© The Author(s) 2020. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Grazing induces direct and indirect shrub effects on soil nematode communities
DOI:10.1016/j.soilbio.2018.03.007 URL [本文引用: 2]
On the relevance of facilitation in alpine meadow communities: an experimental assessment with multiple species differing in their ecological optimum
DOI:10.1016/j.actao.2007.10.002 URL [本文引用: 1]
Optimal balance of water use efficiency and leaf construction cost with a link to the drought threshold of the desert steppe ecotone in northern China
DOI:10.1093/aob/mcw127
PMID:27443298
[本文引用: 2]
In arid environments, a high nitrogen content per leaf area (Narea) induced by drought can enhance water use efficiency (WUE) of photosynthesis, but may also lead to high leaf construction cost (CC). Our aim was to investigate how maximizing Narea could balance WUE and CC in an arid-adapted, widespread species along a rainfall gradient, and how such a process may be related to the drought threshold of the desert-steppe ecotone in northern China.Along rainfall gradients with a moisture index (MI) of 0·17-0·41 in northern China and the northern Tibetan Plateau, we measured leaf traits and stand variables including specific leaf area (SLA), nitrogen content relative to leaf mass and area (Nmass, Narea) and construction cost (CCmass, CCarea), δ(13)C (indicator of WUE), leaf area index (LAI) and foliage N-pool across populations of Artemisia ordosicaIn samples from northern China, a continuous increase of Narea with decreasing MI was achieved by a higher Nmass and constant SLA (reduced LAI and constant N-pool) in high-rainfall areas (MI > 0·29), but by a lower SLA and Nmass (reduced LAI and N-pool) in low-rainfall areas (MI ≤ 0·29). While δ(13)C, CCmass and CCarea continuously increased with decreasing MI, the low-rainfall group had higher Narea and δ(13)C at a given CCarea, compared with the high-rainfall group. Similar patterns were also found in additional data for the same species in the northern Tibetan Plateau. The observed drought threshold where MI = 0·29 corresponded well to the zonal boundary between typical and desert steppes in northern China.Our data indicated that below a climatic drought threshold, drought-resistant plants tend to maximize their intrinsic WUE through increased Narea at a given CCarea, which suggests a linkage between leaf functional traits and arid vegetation zonation.© The Author 2016. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Influence of shrubs on soil characteristics and their function in Sahelian agro- ecosystems in semi-arid Niger
DOI:10.1006/jare.1999.0609 URL [本文引用: 1]
Changes in spatial patterns of Caragana stenophylla along a climatic drought gradient on the Inner Mongolian Plateau
DOI:10.1371/journal.pone.0121234 [本文引用: 1]
Shrubs facilitate recruitment of Caragana stenophylla Pojark: microhabitat amelioration and protection against herbivory
DOI:10.1007/s13595-016-0599-5 URL [本文引用: 3]
Woody species have stronger facilitative effects on soil biota than on plants along an aridity gradient
DOI:10.1111/jvs.13034 [本文引用: 2]
Assessing facilitative responses to a nurse shrub at the community level: the example of Potentilla fruticosa in a sub-alpine grassland of northwest China
DOI:10.1111/j.1438-8677.2009.00271.x
PMID:20701701
[本文引用: 1]
Biotic interaction studies have revealed a large discrepancy among experiments in target responses to the effects of neighbours, which may in part be due to both high species-specificity of plant responses and low number of target species used in experiments. Our aim was to assess facilitative responses at the community level and the role of both functional groups and ecological attributes of target species. In a sub-alpine grassland on the eastern Tibet plateau, we assessed growth responses of all species in the community to removal of a dominant shrub. We also measured changes in the main environmental variables. Species responses were analysed by functional group and in relation to their mean regional altitudinal distribution. All significant interactions were positive and affected one-third of the total species richness of the community. All functional groups were facilitated but forbs were less strongly facilitated than in the two other groups. High-alpine species were less strongly facilitated than low-sub-alpine species, but the strength of this relationship was weaker than that reported in previous work. There was evidence of a decrease in extreme temperatures below the canopy of the shrub but no variations in soil moisture. We conclude that the highly stressful conditions induced by the dry continental climate of the eastern Tibet plateau are a main driver of the exclusive dominance of positive interactions. Assessing interactive responses at the community level is likely to provide a useful tool to better understand the role of biotic interactions in community responses to environmental changes.
Response of temporal variation of soil moisture to vegetation restoration in semi-arid Loess Plateau, China
DOI:10.1016/j.catena.2013.12.005 URL [本文引用: 1]
The ecology of restoration: historical links, emerging issues and unexplored realms
DOI:10.1111/j.1461-0248.2005.00764.x URL [本文引用: 1]
Belowground facilitation and trait matching: two or three to tango?
DOI:10.1016/j.tplants.2021.07.014 URL [本文引用: 1]
Extreme drought stress shifts net facilitation to neutral interactions between shrubs and sub-canopy plants in an arid desert
DOI:10.1111/oik.04630 URL [本文引用: 6]
Facilitation by leguminous shrubs increases along a precipitation gradient
DOI:10.1111/1365-2435.12941 URL [本文引用: 5]
Impacts of leguminous shrub encroachment on neighboring grasses include transfer of fixed nitrogen
DOI:10.1007/s00442-015-3538-5 URL [本文引用: 2]
Effect of Caragana tibetica nebkhas on sand entrapment and fertile islands in steppe-desert ecotones on the Inner Mongolia Plateau, China
DOI:10.1007/s11104-011-0813-z URL [本文引用: 2]
Effects of positive plant interactions on population dynamics and community structures: a review based on individual-based simulation models
DOI:10.3724/SP.J.1258.2013.00059 URL [本文引用: 1]
植物间正相互作用对种群动态和群落结构的影响: 基于个体模型的研究进展
DOI:10.3724/SP.J.1258.2013.00059
[本文引用: 1]
植物间的相互作用对种群动态和群落结构有着重要的影响。大量的野外实验已经揭示了正相互作用(互利)在群落中的普遍存在及其重要性。为了弥补野外实验方法的不足, 模型方法被越来越多地应用于正相互作用及其生态学效应的研究中。该文基于个体模型研究, 探讨了植物间正相互作用对种群动态和群落结构的影响。介绍了植物间正相互作用的定义和发生机制、植物间相互作用与环境梯度的关系。正相互作用是指发生在相邻的植物个体之间, 至少对其中一个个体有益的相互作用。植物通过直接(生境改善或资源富集)或间接(协同防御等)作用使局部环境有利于邻体而发生正相互作用。胁迫梯度假说认为互利的强度或重要性随着环境胁迫度的增加而增加, 但是越来越多的经验研究认为胁迫梯度假说需要改进。以网格模型和影响域模型为例, 介绍了基于个体的植物间相互作用模型方法。基于个体模型, 对近年来国内外正相互作用对种群时间动态(如生物量-密度关系)、空间分布格局和群落结构(如群落生物量-物种丰富度关系)影响的研究进行了总结。指出未来的研究应集中在对正相互作用概念和机制的理解, 新的模型, 新的种群、群落, 甚至生态系统问题, 以及在全球变化背景下进行相关的研究。
Positive interactions in plant communities
植物邻体间的正相互作用
Spatial patterns provide support for the stress-gradient hypothesis over a range-wide aridity gradient
DOI:10.1016/j.jaridenv.2013.11.006 URL [本文引用: 1]

{kind=link}
{kind=link}