生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 )。目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 )。种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 )。灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究。在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究。
灌丛在水源涵养、碳固定、土壤保持和防风固沙等方面发挥重要的作用, 尤其在西南地区具有重要的生态价值(胡雄蛟等, 2021 )。云南省共有27种灌丛群系, 分属于河谷灌丛、山地灌丛、亚高山灌丛、高山灌丛4种植被型, 其中山地灌丛中的南烛(Vaccinium bracteatum )灌丛和云南杨梅(Myrica nana )灌丛分布较广, 两者面积共达到49.04 × 103 km2 (谢宗强等, 2019 )。云南杨梅抗污染能力强, 与根瘤菌共生, 具有杰出的水土保持和肥地功能, 同时具备较强的耐火、阻火能力, 因此可作优质的绿化树种及恢复植被的先锋树种(成宇, 2019 )。云南杨梅是滇中地区山地灌丛的优势种, 研究云南杨梅灌丛主要木本植物的生态位与种间联结不仅有利于更好地认识和发挥云南杨梅灌丛的生态功能, 而且可为揭示云南杨梅灌丛特征和维持云南杨梅灌丛的生物多样性提供科学依据。目前对云南杨梅灌丛的研究甚少, 仅包括对云南杨梅灌丛碳密度(郭子豪等, 2020 )、生物量(巩合德等, 2012 )和土壤有机碳(陈路红等, 2017 )等的研究。对云南杨梅灌丛群落的生态位和种间联结的研究目前还未有报道。本研究采用了Levins生态位宽度(B L )、Shannon生态位宽度(B S )、Schoener重叠指数(Cik )、Pianka重叠指数(Oik )、方差比率(VR)、χ2 检验、Pearson相关检验和Spearman秩相关检验进行数据分析与处理, 以说明下面几个科学问题: 1)云南杨梅灌丛主要木本植物的生态位宽度和生态位重叠特征; 2)云南杨梅灌丛主要木本植物的种间联结关系; 3)云南杨梅灌丛主要木本植物的生态位重叠和种间联结之间的联系。
1 材料和方法
1.1 研究区概况
研究样地位于云南省滇中地区昆明市、曲靖市和楚雄彝族自治州, 地理坐标为101.01°-104.37° E, 24.17°-25.48° N, 海拔1 936-2 737 m。滇中地处滇东高原, 地形以山地和山间盆地为主, 地势起伏和缓(吴志杰和何云玲, 2015 ); 土壤类型以红壤为主; 属低纬度亚热带高原季风气候, 2020年的年平均气温为17.72 ℃, 降水量为881.31 mm。滇中地区植被类型主要为河谷灌丛和山地灌丛, 河谷灌丛以余甘子(Phyllanthus emblica )为建群种, 山地灌丛以南烛、云南杨梅为建群种(陈路红等, 2017 )。样地中植被类型主要为山地灌丛, 主要物种包括云南杨梅、西南金丝梅(Hypericum henryi )、樟叶越桔(Vaccinium dunalianum )、乌鸦果(Vaccinium fragile )、铁仔(Myrsine africana )、碎米花(Rhododendron spiciferum )、茅莓(Rubus parvifolius )、云南松(Pinus yunnanensis )、滇青冈(Cyclobalanopsis glaucoides )、野丁香(Leptodermis potaninii )、粉叶小檗(Berberis pruinosa )、芳香白珠(Gaultheria fragrantissima )、钝叶橘红悬钩子(Rubus aurantiacus var. obtusifolius )、川梨(Pyrus pashia )、元江锥(Castanopsis orthacantha )、火棘(Pyracantha fortuneana )、亮毛杜鹃(Rhododendron microphyton )、高山锥(Castanopsis delavayi )、南烛、怒江红山茶(Camellia saluenensis )等。其中云南杨梅为优势种。
1.2 样地设置与调查
在滇中地区受人为干扰较轻的云南杨梅群落分布区设置了11个样地, 样地位于云南省安宁市、牟定县、师宗县、宜良县、昆明市五华区、富民县、姚安县、南华县、寻甸回族彝族自治县9个县(区), 其中富民县、姚安县设置2个样地, 其余县(区)各设置1个样地。每个样地设置3个样方, 共建立了33个5 m × 5 m样方, 调查并记录灌丛内灌木个体的种名、株数、高度、基径、冠幅等指标。
1.3 数据处理
(1) 物种重要值(IV) = (相对多度+相对频度)/2
利用B L (Levins, 1968 )和B S (Shannon & Weiner, 1949 )计算生态位宽度, 公式如下:
(2) ${{B}_{\text{L}}}={1}/{\sum\limits_{j=1}^{N}{{{({{P}_{ij}})}^{2}}}}\;$
(3) ${{B}_{\text{S}}}=-\sum\limits_{j=1}^{N}{{{P}_{ij}}}\ln {{P}_{ij}}$
式中, Pij 为物种i 在资源位j 上的个体数占物种i 在所有资源位上的个体数总和的比例。N 为样方总数, 以每个样方代表一个资源位。
利用Cik (Schoener, 1974 )和Oik (Pianka, 1973 )计算生态位重叠程度, 公式如下:
(4) ${{C}_{ik}}=1-\frac{1}{2}\sum\limits_{j=1}^{N}{|{{P}_{ij}}-{{P}_{kj}}|}$
(5) ${{O}_{ik}}={\sum\limits_{j=1}^{N}{{{P}_{ij}}}{{P}_{kj}}}/{\sqrt{{{\left( \sum\limits_{j=1}^{N}{{{P}_{ij}}} \right)}^{2}}{{\left( \sum\limits_{j=1}^{N}{{{P}_{kj}}} \right)}^{2}}}}\;$
式中, Cik 和Oik 为物种i 的资源利用曲线与种j 的资源利用曲线的重叠指数, 值域为[01], 其值越大表示生态位重叠程度越高; Pij 和Pkj 分别是物种i 和k 在资源位j 上的多度。
使用VR (Schluter, 1984 )检验总体联结性, 以统计量W检验联结性的显著度。
(6) $\delta _{f}^{2}=\sum\limits_{i=1}^{S}{{{P}_{i}}}(1-{{P}_{i}})$
(7) $\delta _{S}^{2}=\frac{1}{N}\sum\limits_{i=1}^{N}{{{({{S}_{j}}-{{S}_{\text{m}}})}^{2}}}$
(8) $\text{VR}=\delta _{S}^{2}/\delta _{f}^{2}$
(9) $\text{W}=\text{VR}\times N$
式中, Pi = ni /N , ni 为物种i 出现的样方数, N 为样方总数, S 为总物种数, Sj 为样方j 内出现的物种总数, S m 为所有样方中出现的物种平均数, δS 为所有样方物种数的方差, δf 为所有物种出现频度的方差。VR = 1表明种间彼此独立, VR > 1表明种间正关联, VR < 1表明种间负关联。W可检验VR偏离1是否显著, W服从χ 2 分布, 若W > χ 2 0.05 (N )或W < χ 2 0.95 (N ), 则物种间总体关联显著(p < 0.05), 若W值落入χ 2 分布的90%置信区间χ 2 0.95 (N ) < W < χ 2 0.05 (N )内, 则物种间总体关联不显著(p > 0.05), χ 2 0.05 (N )和χ 2 0.95 (N )为χ 2 临界值。
采用χ 2 检验、相关系数进行种间关联分析。鉴于本研究为非连续性取样, 种间关联的显著程度采用Yates的连续校正公式(张金屯, 2018 )进行χ2 检验, 公式如下:
(10) ${{\chi }^{2}}=\frac{N{{(|ad-bc|-0.5N)}^{2}}}{(a+b)(b+d)(c+d)(a+c)}$
式中, N 为总样方数, a 为2个物种同时出现的样方数, b 、c 分别为2个物种单独出现的样方数, d 为2个物种都不出现的样方数。当ad = bc 时, 两个种相互独立; 当ad > bc 时, 两个种之间呈正联结关系; 当ad < bc 时, 两个种之间呈负联结关系。若χ 2 < 3.841 (p > 0.05), 表明种对间呈不显著联结关系, 种对间基本独立; 若3.841 ≤ χ 2 ≤ 6.635 (0.01 ≤ p ≤ 0.05), 表明种对间呈显著联结关系; 若χ 2 > 6.635 (p < 0.01), 表明种对间呈极显著联结关系(宋永昌, 2001 )。
由于样地内云南杨梅出现的频率为100%, 导致χ 2 检验公式中的分母为0, 不能准确反映云南杨梅和其他物种之间的种间联结关系, 因此给该种一个加权值, 令其在公式中的c 、d 值均为1 (王伯荪和彭少麟, 1985 )。
(11) ${{r}_{\text{p}}}(i,k)=\frac{\sum\limits_{j=1}^{N}{({{x}_{ij}}-{{{\bar{x}}}_{i}})({{x}_{kj}}-{{{\bar{x}}}_{k}})}}{\sqrt{\sum\limits_{j=1}^{N}{{{({{x}_{ij}}-{{{\bar{x}}}_{i}})}^{2}}\sum\limits_{j=1}^{N}{{{({{x}_{kj}}-{{{\bar{x}}}_{k}})}^{2}}}}}}$
(12) ${{r}_{\text{s}}}(i,k)=1-\frac{6\sum\limits_{j=1}^{N}{{{({{x}_{ij}}-{{{\bar{x}}}_{i}})}^{2}}{{({{x}_{kj}}-{{{\bar{x}}}_{k}})}^{2}}}}{{{N}^{3}}-N}$
式中, N 为样方总数, xij 和xkj 分别是物种i 和物种k 在样方j 中的个体数; ${{\bar{x}}_{i}}$ ${{\bar{x}}_{k}}$ j 个样方中物种i 和物种k 个体数的平均值。r p (i,k )和r s (i,k )的值域为[–1, 1], 正值为正相关, 负值为负相关。
1.4 数据分析
生态位重叠指数和Pearson相关系数、Spearman秩相关系数运用Origin 9进行线性回归分析。重要值、生态位和种间联结采用Excel 2019和R 4.0.4软件(“spaa”程序包)进行数据计算。
2 结果和分析
2.1 重要值特征
经统计云南杨梅灌丛共有木本植物51种, 隶属于22科39属。如表1 所示, 前20位物种的重要值之和占全部物种重要值之和的83.30%, 说明这20个物种是群落的重要组成部分, 可作为研究该群落物种相互关系的主要种, 因此选择重要值前20位的物种进行生态位和种间联结分析。云南杨梅的重要值为15.13%, 是群落中重要值最大的物种, 为群落的优势种。
2.2 生态位宽度特征
如表1 所示, 滇中云南杨梅灌丛木本植物主要物种的B L 和B S 的变化范围分别为1.85-27.36和0.65-3.38。群落中云南杨梅的B L 和B S 分别为27.36和3.38, 生态位宽度最大。西南金丝梅、樟叶越桔、铁仔、乌鸦果、云南松这5个物种的生态位宽度较大。芳香白珠、怒江红山茶、钝叶橘红悬钩子这3个物种的生态位宽度较小。钝叶橘红悬钩子的B L 和B S 分别为1.85和0.65, 生态位宽度最小。多数物种的B L 和B S 生态位宽度分别在1.00-12.00和0.50-4.00之间。
2.3 生态位重叠特征
如表2 、表3 所示, Cik 和Oik 分别介于0-0.750和0-0.920之间。Cik 平均值为0.127, Cik ≥ 0.500的有4个种对(占总种对数的2.11%), Cik 最大的是粉叶小檗和钝叶橘红悬钩子, 为0.750, 处于[0.200, 0.500)的有51个种对(占总种对数的26.84%), 介于(0, 0.200)的有59个种对(占总种对数的31.05%), 为0的有76个种对(占总种对数的40.00%)。粉叶小檗和钝叶橘红悬钩子的生态位重叠值高达0.750。而西南金丝梅和亮毛杜鹃, 西南金丝梅和南烛, 樟叶越桔和钝叶橘红悬钩子等76个种对Cik 均为0。Oik 平均值为0.167, Oik ≥ 0.500的有15个种对(占总种对数的7.89%), Oik 最大的是元江锥和高山锥, 为0.920, 处于[0.200, 0.500)的有54个种对(占总种对数的28.42%), 介于(0, 0.200)的种对有45对(占总种对数的23.68%), 未产生生态位重叠(Oik = 0)的种对有76对(占总种对数的40.00%)。
2.4 总体联结性分析
总体联结性的VR = 1.70 > 1, 表明主要物种间总体呈正联结关系。χ 2 临界值(χ 2 0.95(33), χ 2 0.05(33) )分别为26.51和47.40。W = 55.96 > χ 2 0.05 (33), 表明物种间的总体联结显著(p < 0.05)。因此, 滇中云南杨梅灌丛主要木本植物间的总体联结性表现为显著正联结。
2.5 种对间的联结性分析
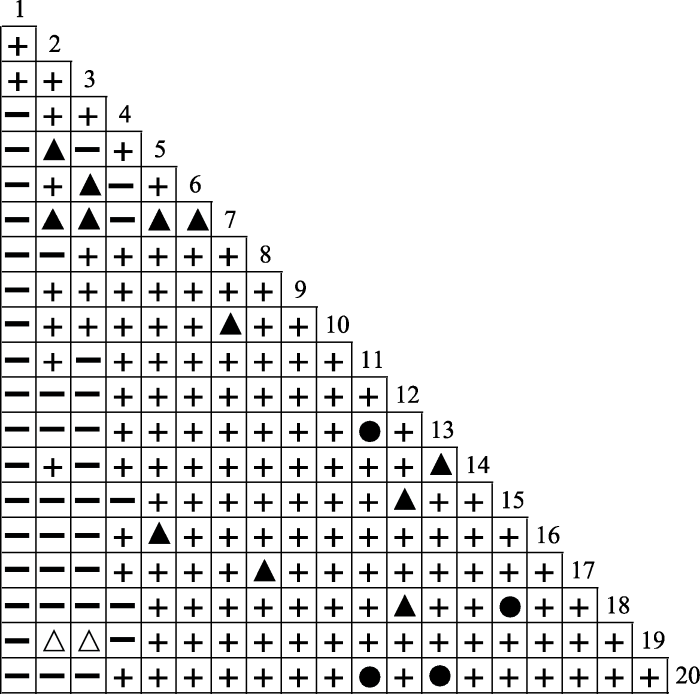
由表4 和图1 可看出, 20个物种组成的190个种对中, χ 2 检验结果表明呈正联结关系的种对有148对, 占总种对数的77.89%, 其中呈极显著正联结关系的有4对(2.11%), 呈显著正联结关系的有12对(6.32%), 呈不显著正联结关系的有132对(69.47%)。负联结的种对有42对, 占总种对数的22.11%, 其中呈极显著负联结关系的有0对, 呈显著负联结关系的有2对(1.05%), 呈不显著负联结关系的有40对(21.05%)。正联结的种对数是负联结种对数的3.52倍。显著联结的种对有18对(占总种对数的9.47%); 不显著联结的种对有172对(占总种对数的90.53%)。群落中云南杨梅与其他物种呈正联结关系的种对有2对, 负联结的种对有17对, 且均为不显著联结关系。
图1
图1
滇中云南杨梅灌丛主要木本植物的χ2 检验值半矩阵图。●, 极显著正联结(0 < p < 0.01); ▲, 显著正联结(0.01 ≤ p ≤ 0.05); ╂, 不显著正联结(p > 0.05); △, 显著负联结(0.01≤ p ≤ 0.05); ━, 不显著负联结(p > 0.05)。1-20, 物种编号同表1。
Fig. 1
Semi-matrix diagram of computed value of chi-square test of main woody plants in Myrica nana shrubland in central Yunnan. ●, extremely significant positive correlation (0 < p < 0.01); ▲, significant positive correlation (0.01 ≤ p ≤ 0.05); ╂, not significant positive correlation (p > 0.05); △, significant negative correlation (0.01≤ p ≤ 0.05); ━, not significant negative correlation (p > 0.05). 1-20, species No. see Table 1.
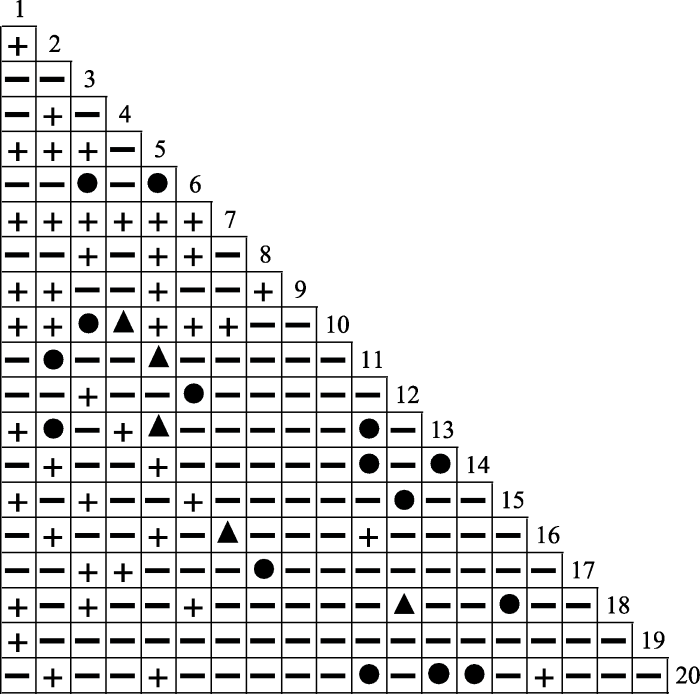
Pearson相关检验的结果如表4 和图2 所示, 显示正相关关系的种对有63对, 占总种对数的33.16%,其中呈极显著正相关关系的有15对(7.89%), 呈显著正相关关系的有5对(2.63%), 呈不显著正相关关系的有43对(22.63%)。负相关种对有127对, 占总种对数的66.84%, 其中呈极显著负相关关系的有0对, 呈显著负相关关系的有0对, 呈不显著负相关关系的有127对(66.84%)。正相关种对数与负相关种对数的比值为0.50, 显著相关种对有20对, 检验显著率10.53%; 不显著相关种对有170对, 占总种对数的89.47%, 绝大部分种对呈不显著相关关系, 这同χ 2 检验的结果一致。群落中云南杨梅与其他物种呈正联结关系的种对有9对, 呈负联结关系的种对有10对, 且均为不显著联结关系。
图2
图2
滇中云南杨梅灌丛主要木本植物的Pearson相关检验半矩阵图。●, 极显著正联结(0 < p < 0.01); ▲, 显著正联结(0.01 ≤ p ≤ 0.05); ╂, 不显著正联结(p > 0.05); ━, 不显著负联结(p > 0.05)。1-20, 物种编号同表1。
Fig. 2
Semi-matrix diagram of Pearson correlation test of main woody plants in Myrica nana shrubland in central Yunnan. ●, extremely significant positive correlation (0 < p < 0.01); ▲, significant positive correlation (0.01 ≤ p ≤ 0.05); ╂, not significant positive correlation (p > 0.05); ━, not significant negative correlation (p > 0.05). 1-20, species No. see Table 1.
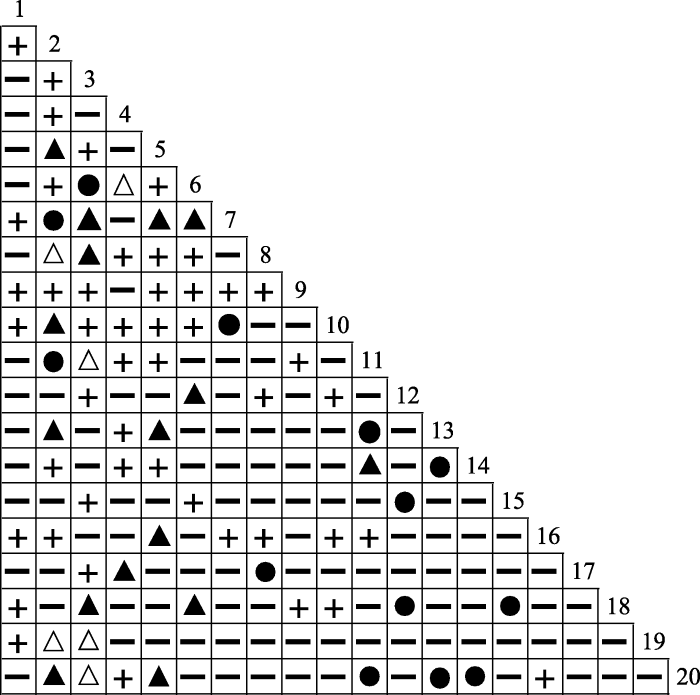
Spearman秩相关检验的结果如表4 和图3 所示, 显示正相关关系的种对有76对, 占总种对数的40%, 其中呈极显著正相关关系的有13对(6.84%), 呈显著正相关关系的有16对(8.42%), 呈不显著正相关关系的有47对(24.74%)。负相关种对有114对, 占总种对数的60.00%, 其中呈极显著负相关关系的有0对, 呈显著负相关关系的有6对(3.16%), 呈不显著负相关关系的有108对(56.84%)。正负相关种对的比值为0.67, 显著相关种对有35对, 检验显著率为18.42%; 不显著相关种对有155对, 占总种对数的81.58%, 绝大部分种对呈不显著相关关系, 这同χ2 检验的结果一致。群落中云南杨梅与其他物种呈正联结关系的种对有7对, 呈负联结关系的种对有12对, 且均为不显著联结关系。
图3
图3
滇中云南杨梅灌丛主要木本植物的Spearman秩相关检验半矩阵图。●, 极显著正联结(0 < p < 0.01); ▲, 显著正联结(0.01 ≤ p ≤ 0.05); ╂, 不显著正联结(p > 0.05); △, 显著负联结(0.01≤ p ≤ 0.05); ━, 不显著负联结(p > 0.05)。1-20, 物种编号同表1。
Fig. 3
Semi-matrix diagram of Spearman rank correlation test of main woody plants in Myrica nana shrubland in central Yunnan. ●, extremely significant positive correlation (0 < p < 0.01); ▲, significant positive correlation (0.01 ≤ p ≤ 0.05); ╂, not significant positive correlation (p > 0.05); △, significant negative correlation (0.01≤ p ≤ 0.05); ━, not significant negative correlation (p > 0.05). 1-20, species No. see Table 1.
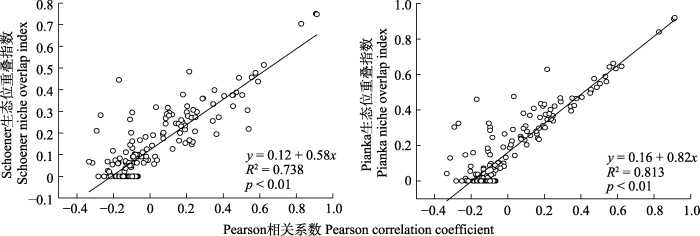
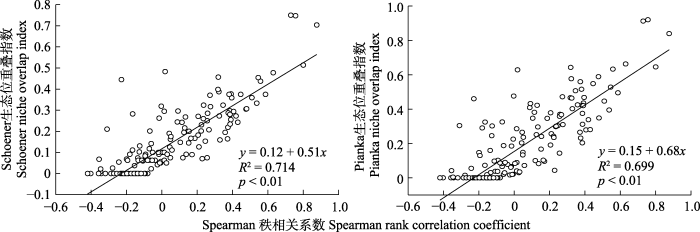
2.6 种间联结与生态位重叠的回归分析
回归分析的结果如图4 、图5 所示。滇中云南杨梅灌丛木本植物主要物种的Pearson相关系数、Spearman秩相关系数与生态位重叠指数之间的p < 0.01, 且图中斜率均大于0, 表明它们之间均存在极显著的线性正相关关系, 即生态位重叠程度随着Pearson相关系数、Spearman秩相关系数的变大(变小)而变大(变小)。
图4
图4
滇中云南杨梅灌丛主要木本植物的Pearson相关系数和生态位重叠指数的回归分析。
Fig. 4
Regression analysis between Pearson correlation coefficient and niche overlap index of main woody plants in Myrica nana shrubland in central Yunnan.
图5
图5
滇中云南杨梅灌丛主要木本植物的Spearman秩相关系数和生态位重叠指数的回归分析。
Fig. 5
Regression analysis between Spearman rank correlation coefficient and niche overlap index of main woody plants in Myrica nana shrubland in central Yunnan.
3 讨论
3.1 物种生态位宽度和生态位重叠
本研究中云南杨梅的生态位宽度最大, 其竞争能力和对资源的利用能力强, 环境适应性较好, 在此群落中会向着泛化种的方向发展。钝叶橘红悬钩子的生态位宽度最小, 其竞争能力和对资源的利用能力弱, 环境的适应性较差, 倾向于发展为特化种。B L 和B S 测定结果基本一致, 但在排序上有微小差异, 如B L 中, 西南金丝梅的生态位宽度大于樟叶越桔, 芳香白珠的生态位宽度大于怒江红山茶; B S 中, 西南金丝梅的生态位宽度小于樟叶越桔, 芳香白珠的生态位宽度小于怒江红山茶。生态位宽度和重要值没有呈现出一致联系, 说明群落的资源还有待充分利用, 群落正在朝着结构复杂化, 功能完善化的方向发展。云南松、川梨和南烛的生态位宽度排序较重要值排序靠前, 表明这些物种具有较大的发展潜力。
生态位重叠可以解释物种间竞争或共存关系, 通常认为重叠值大于0.500, 则种间具有较大的生态和资源利用相似性, 竞争比较严重(沈雪梨等, 2020 )。生态位重叠值大于0.500的种对数均较少, 且为0的种对数较多, 表明云南杨梅灌丛主要木本植物之间对环境资源需求的相似性不大, 种间竞争较弱, 种间关系趋于稳定。云南杨梅与西南金丝梅、樟叶越桔、铁仔的生态位重叠程度较大, 可能因为这些物种具有较大的生态位宽度值和重要值; 与亮毛杜鹃、芳香白珠、怒江红山茶的生态位重叠程度较小, 可能因为它们对资源需求的相似性低, 出现了不同程度的生态位分化, 且这些物种在样方中数量分布较为集中, 相对频度较低。
生态位宽度较大(较小)的物种与其他物种产生生态位重叠的概率也较大(较小)(陈玉凯等, 2014 )。本研究中云南杨梅出现在每个样方中, 生态位宽度最大, 与20个主要物种产生生态位重叠的概率为100%。亮毛杜鹃的生态位宽度较小, 与20个主要物种间的生态位重叠概率也较小。生态位宽度较小的物种间也出现生态位重叠值较高的情况, 如粉叶小檗和钝叶橘红悬钩子、粉叶小檗和怒江红山茶、钝叶橘红悬钩子和怒江红山茶, 这与张东梅等(2018 )的研究结论一致。
3.2 种间联结性分析
滇中云南杨梅灌丛主要木本植物的总体联结性表现为显著正联结, 表明群落各物种之间可以协调共存, 对资源的利用较为充分, 群落结构趋于稳定, 可能是因为绝大多数样地中云南杨梅群落受到的人为干扰程度轻微。检验显著率18.42% (Spearman秩相关检验) > 10.53% (Pearson相关检验) > 9.47% (χ 2 检验); Spearman秩相关检验中呈显著关联关系的种对数比Pearson相关检验更多, 说明Spearman秩相关检验有更高的灵敏度, 可以更为准确地反映种间关系。这与朱利君等(2005 )的研究结果一致。以Spearman秩相关检验结果为依据, 即种对间正负联结比小于1, 判断云南杨梅群落可能处于演替的前期或中期阶段。综合3种检验结果, 不显著种对数均大于显著种对数, 说明群落物种间的联结性较弱, 呈独立分布格局。粉叶小檗-钝叶橘红悬钩子、粉叶小檗-怒江红山茶、钝叶橘红悬钩子-怒江红山茶和元江锥-高山锥均呈现极显著正联结关系, 说明这些物种之间相互依存或资源需求上互补。
云南杨梅的生态习性是中等喜光, 不耐强烈的日照, 喜温暖湿润气候, 喜空气湿度大, 宜排水良好的酸性土壤, 稍耐贫瘠(欧晓昆等, 2015 )。云南杨梅在χ2 检验下与其他物种多呈不显著负联结关系, 在Pearson相关检验和Spearman秩相关检验中与半数物种呈不显著正联结关系, 包括西南金丝梅、茅莓、滇青冈、野丁香、高山锥等, 这些物种的共同特性是喜光, 较耐阴, 在一般土壤上均能够较好地生长, 随着群落的演化, 这些物种会发展为伴生种, 因此需要加大对这些物种的保护; 半数呈不显著负联结关系, 包括樟叶越桔、乌鸦果、碎米花、云南松等, 可能因为这些物种和云南杨梅均喜酸性土壤和光, 它们和云南杨梅产生了竞争, 因而形成负联结关系。当环境容量不够时, 这些物种和云南杨梅的竞争会加剧, 在保护云南杨梅群落的过程中, 需要适当协调这些物种和云南杨梅之间的关系。
3.3 生态位重叠与种间联结的相关性
多数研究认为: 种间正联结关系越强, 其生态位重叠值就越大; 反之, 种间负联结关系越强, 其生态位重叠值就越小(郭志华等, 1997 ; 霍红等, 2013 )。本研究显示, 滇中云南杨梅灌丛木本植物主要物种的Pearson相关系数、Spearman秩相关系数与生态位重叠指数之间均存在极显著的正相关关系。如元江锥和高山锥的Pearson相关系数和Spearman秩相关系数值较大, 种对呈极显著正联结关系, 生态位重叠值也较大。但也有研究认为种间负关联关系则比较复杂, 由生境差异导致的负关联关系生态位重叠值低, 而由资源竞争引起的负关联关系生态位重叠值高(刘润红等, 2020 )。如本研究中西南金丝梅和南烛, 西南金丝梅和亮毛杜鹃的Pearson相关系数和Spearman秩相关系数值为负值, 种间关系为负联结, 其种对间的生态位重叠值为0, 说明这两个种对间的负联结关系是由生境差异引起的。
3.4 生物多样性保护策略
生态位和种间联结关系的研究结果表明云南杨梅灌丛群落主要木本植物物种之间的联系较弱, 群落处于较为稳定的发展时期。对于群落的保护, 一是需要保护云南杨梅群落所在的原生环境, 减少人为干扰, 二是需要加强保护和云南杨梅呈正联结关系的物种, 如西南金丝梅和滇青冈。樟叶越桔、乌鸦果和云南杨梅呈负联结关系且生态位重叠程度较大, 在群落发展中要及时关注这些物种的状况。
参考文献
View Option
[1]
Bai HH (2018 ). Multi-dimension Nutrition Ecological Niche and Interspecific Association Characteristics Research of Dominant Wood Plants of Pinus taiwanensis Communities. Master degree dissertation, Chinese Academy of Forestry, Beijing.
[本文引用: 1]
[ 白欢欢 (2018 ). 黄山松群落主要木本植物营养生态位和种间联结特征研究. 硕士学位论文, 中国林业科学研究院, 北京.]
[本文引用: 1]
[2]
Chen L Xin JN Su Y Li YF Song NP Wang L Yang XG Bian YY Tian N (2019 ). Effects of heterogeneous habitats on community composition and niche characteristics of different plant populations in the desert steppe of China
Acta Ecologica Sinica , 39 , 6187 -6205 .
[本文引用: 1]
[ 陈林 , 辛佳宁 , 苏莹 , 李月飞 , 宋乃平 , 王磊 , 杨新国 , 卞莹莹 , 田娜 (2019 ). 异质生境对荒漠草原植物群落组成和种群生态位的影响
生态学报 , 39 , 6187 -6205 .]
[本文引用: 1]
[3]
Chen LH Su KW Zheng W Yin HJ Gong HD (2017 ). Distribution characteristics and influencing factors of soil organic carbon in 2 dominant shrub soil in Yunnan Province
Journal of Southwest Forestry University (Natural Science ), 37 (5 ), 106 -113 .
[本文引用: 2]
[ 陈路红 , 苏凯文 , 郑伟 , 尹华军 , 巩合德 (2017 ). 云南2种主要灌丛土壤有机碳分布特征及其影响因子研究
西南林业大学学报(自然科学) , 37 (5 ), 106 -113 .]
[本文引用: 2]
[4]
Chen YK Yang Q Mo YN Yang XB Li DH Hong XJ (2014 ). A study on the niches of the state’s key protected plants in Bawangling, Hainan Island
Chinese Journal of Plant Ecology , 38 , 576 -584 .
DOI:10.3724/SP.J.1258.2014.00053
URL
[本文引用: 1]
[ 陈玉凯 , 杨琦 , 莫燕妮 , 杨小波 , 李东海 , 洪小江 (2014 ). 海南岛霸王岭国家重点保护植物的生态位研究
植物生态学报 , 38 , 576 -584 .]
DOI:10.3724/SP.J.1258.2014.00053
[本文引用: 1]
为进一步了解国家重点野生保护植物的生存群落及不同种群利用资源和占据生态空间的能力, 为制定国家重点保护植物种群的保护措施提供依据, 2009-2011年, 对霸王岭有国家重点保护植物分布的70个样地进行群落调查, 研究了霸王岭保护区内12种国家重点保护植物的生态位宽度以及物种间的生态位重叠。结果表明: (1)生态位宽度值从大到小依次为: 土沉香(Aquilaria sinensis)、海南韶子(Nephelium topengii)、普洱茶(Camellia sinensis var. assamica)、油丹(Alseodaphne hainanensis)、海南油杉(Keteleeria hainanensis)、鸡毛松(Podocarpus imbricatus)、海南梧桐(Firmiana hainanensis)、乐东拟单性木兰(Parakmeria lotungensis)、海南粗榧(Cephalotaxus mannii)、香子含笑(Michelia hedyosperma)、粘木(Ixonanthes reticulata)和陆均松(Dacrydium pierrei); (2)物种生态位宽度值的大小与其自身的地理分布范围密切相关, 如: 陆均松、乐东拟单性木兰等生态位宽度较小的原因与其分布的海拔范围狭窄有关。生态位宽度较大的种群对资源的利用能力较强, 与其他物种的生态位重叠一般较大, 如: 生态位宽度较大的普洱茶与海南油杉的生态位重叠值最大, 为0.872, 而一些生态位宽度较小的物种生态位重叠值较高, 如: 香子含笑的生态位宽度较小, 但与海南梧桐的生态位重叠较大, 为0.693。因此, 种群间的生态位重叠值与它们对环境资源利用的相似程度以及物种自身的生物学特性密切相关; (3)香子含笑、粘木等的生态位宽度较小, 而且个体数量较少, 适宜其生存的群落和生境极少, 应该给予更多的关注和优先保护; (4)未来对濒危植物的保护不仅要考虑各物种自身的生物学特性, 还应从植物群落的角度考虑。只有保护好这些濒危植物的生存群落, 如山地雨林等, 才能实现对濒危植物的有效保护。
[5]
Cheng Y (2019 ). Phylogeography of Myrica nana, Endemic Plant Species Distributed in Yunnan-Guizhou Plateau. Master degree dissertation, Guizhou University, Guiyang.
[本文引用: 1]
[ 成宇 (2019 ). 云贵高原特有植物矮杨梅的谱系地理学研究. 硕士学位论文, 贵州大学, 贵阳.]
[本文引用: 1]
[6]
Dong X Xin ZM Duan RB Huang YR Jian KM Ma HF (2020 ). Species diversity of typical shrubs and niches of dominant shrub species in the Ulanbuh Desert
Arid Zone Research , 37 , 1009 -1017 .
[本文引用: 1]
[ 董雪 , 辛智鸣 , 段瑞兵 , 黄雅茹 , 菅凯敏 , 马海峰 (2020 ). 乌兰布和沙漠典型灌木群落多样性及其生态位
干旱区研究 , 37 , 1009 -1017 .]
[本文引用: 1]
[7]
Gao YH Zeng XY Zhou GY Wang GX (2011 ). Interspecific relationships of dominant populations in the alpine wetlands in the source region of Yangtze River
Wetland Science , 9 , 1 -7 .
[本文引用: 1]
[ 高永恒 , 曾晓阳 , 周国英 , 王根绪 (2011 ). 长江源区高寒湿地植物群落主要种群种间关系分析
湿地科学 , 9 , 1 -7 .]
[本文引用: 1]
[8]
Gong HD Cheng XP Ma YW (2012 ). Characteristics of biomass distribution in Myrica nana
Nonwood Forest Research , 30 (4 ), 106 -108 .
[本文引用: 1]
[ 巩合德 , 程希平 , 马月伟 (2012 ). 云南杨梅灌丛生物量的分配特征
经济林研究 , 30 (4 ), 106 -108 .]
[本文引用: 1]
[9]
Guo ZH Li YK Gong HD (2020 ). Carbon density and its spatial distribution dominated pattern of the Myrica nana cheval shrub in central Yunnan
Journal of West China Forestry Science , 49 (1 ), 149 -155 .
[本文引用: 1]
[ 郭子豪 , 李瑶珂 , 巩合德 (2020 ). 滇中地区矮杨梅灌丛碳密度分配格局研究
西部林业科学 , 49 (1 ), 149 -155 .]
[本文引用: 1]
[10]
Guo ZH Zhuo ZD Chen J Wu MF (1997 ). Interspecific association of trees in mixed evergreen and deciduous broadleaved forest in Lushan Mountain
Acta Phytoecologica Sinica , 21 , 424 -432 .
[本文引用: 1]
[ 郭志华 , 卓正大 , 陈洁 , 吴梅凤 (1997 ). 庐山常绿阔叶、落叶阔叶混交林乔木种群种间联结性研究
植物生态学报 , 21 , 424 -432 .]
[本文引用: 1]
[11]
Hu XJ Song CS Fan XY Xiao Y Xu WH Ouyang ZY (2021 ). Main regulation services and value assessment of shrub ecosystem in China
Journal of Beijing Forestry University (Social Sciences )20 (3 ), 58 -64 .
[本文引用: 1]
[ 胡雄蛟 , 宋昌素 , 范馨悦 , 肖燚 , 徐卫华 , 欧阳志云 (2021 ). 中国灌丛生态系统主要调节服务及价值评估
北京林业大学学报(社会科学版) , 20 (3 ), 58 -64 .]
[本文引用: 1]
[12]
Huo H Feng Q Su YH Si JH Xi HY Yu TF (2013 ). Interspecific relationship and niche analysis on phytocoenosium in the Ejina Oasis
Journal of Desert Research , 33 , 1027 -1033 .
[本文引用: 2]
[ 霍红 , 冯起 , 苏永红 , 司建华 , 席海洋 , 鱼腾飞 (2013 ). 额济纳绿洲植物群落种间关系和生态位研究
中国沙漠 , 33 , 1027 -1033 .]
[本文引用: 2]
[13]
Levins R (1968 ). Evolution in Changing Environments: Some Theoretical Explorations . Princeton University Press , Princeton .
[本文引用: 1]
[14]
Li B Li SQ Zhang JT (2010 ). A study on niches of dominant species of subalpine meadow in the Yunding Mountain
Acta Prataculturae Sinica , 19 (1 ), 6 -13 .
[本文引用: 1]
[ 李斌 , 李素清 , 张金屯 (2010 ). 云顶山亚高山草甸优势种群生态位研究
草业学报 , 19 (1 ), 6 -13 .]
[本文引用: 1]
[15]
Li JL Zhang JT Yuan JY (2004 ). Interspecific relationships of dominant species of sub-alpine scrub community and sub-alpine meadow community in the Guandi Mountains
Acta Agrestia Sinica , 12 , 113 -119 .
[本文引用: 1]
[ 李军玲 , 张金屯 , 袁建英 (2004 ). 关帝山亚高山灌丛群落和草甸群落优势种的种间关系
草地学报 , 12 , 113 -119 .]
DOI:10.11733/j.issn.1007-0435.2004.02.008
[本文引用: 1]
运用2×2列联表、χ<sup>2</sup>检验方法对关帝山亚高山草甸群落的28个优势种及亚高山灌丛群落的26个优势种进行总体相关性和种对间的联结性检验,利用Ochiai指数测定种对间的关联度,应用Pearson相关系数和Spearman秩相关系数刻画种对间的数量变化关系。结果表明:群落优势种间多呈不显著关联,草甸群落中显著相关的种对数仅占总对数的17.9%,灌丛群落为25.9%;对生境要求相同或相似的种对呈显著的正关联;种间关系因群落类型不同存在明显差异。
[16]
Liu Q Bao WK Qiao YK Qian NB (1996 ). Studies on the interspecific relationship among dominant species of the semi-arid valley scrubs in Maoxian on the upper reaches of the Minjiang River
Chinese Journal of Applied and Environmental Biology , 2 , 36 -42 .
[本文引用: 1]
[ 刘庆 , 包维楷 , 乔永康 , 钱能斌 (1996 ). 岷江上游茂县半干旱河谷灌丛优势种间关系的研究
应用与环境生物学报 , 2 , 36 -42 .]
[本文引用: 1]
[17]
Liu RH Chen L Tu HR Liang SC Jiang Y Li YJ Huang DL Nong JL (2020 ). Niche and interspecific association of main species in shrub layer of Cyclobalanopsis glauca community in karst hills of Guilin, southwest China
Acta Ecologica Sinica , 40 , 2057 -2071 .
[本文引用: 2]
[ 刘润红 , 陈乐 , 涂洪润 , 梁士楚 , 姜勇 , 李月娟 , 黄冬柳 , 农娟丽 (2020 ). 桂林岩溶石山青冈群落灌木层主要物种生态位与种间联结
生态学报 , 40 , 2057 -2071 .]
[本文引用: 2]
[18]
Liu Y Zheng YY Ao YY (2019 ). Niche and interspecific association of dominant bryophytes on different substrates
Acta Ecologica Sinica , 39 , 286 -293 .
[本文引用: 1]
[ 刘艳 , 郑越月 , 敖艳艳 (2019 ). 不同生长基质的苔藓植物优势种生态位与种间联结
生态学报 , 39 , 286 -293 .]
[本文引用: 1]
[19]
Ou XK Song D Liu M (2015 ). Native Plants for Garden in Yunnan, China. Yunnan Science and Technology Press , Kunming .
[本文引用: 1]
[ 欧晓昆 , 宋鼎 , 刘敏 (2015 ). 中国云南乡土园林植物 . 云南科技出版社 , 昆明 .]
[本文引用: 1]
[21]
Schluter D (1984 ). A variance test for detecting species associations, with some example applications
Ecology , 65 , 998 -1005 .
DOI:10.2307/1938071
URL
[本文引用: 1]
[22]
Schoener TW (1974 ). Resource partitioning in ecological communities
Science , 185 , 27 -39 .
DOI:10.1126/science.185.4145.27
PMID:17779277
[本文引用: 1]
To understand resource partitioning, essentially a community phenomenon, we require a holistic theory that draws upon models at the individual and population level. Yet some investigators are still content mainly to document differences between species, a procedure of only limited interest. Therefore, it may be useful to conclude with a list of questions appropriate for studies of resource partitioning, questions this article has related to the theory in a preliminary way. 1) What is the mechanism of competition? What is the relative importance of predation? Are differences likely to be caused by pressures toward reproductive isolation? 2) Are niches (utilizations) regularly spaced along a single dimension? 3) How many dimensions are important, and is there a tendency for more dimensions to be added as species number increases? 4) Is dimensional separation complementary? 5) Which dimensions are utilized, how do they rank in importance, and why? How do particular dimensions change in rank as species nuimber increases? 6) What is the relation of dimensional separation to difference in phenotypic indicators? To what extent does the functional relation of phenotype to resource characteristics constrain partitioning? 7) What is the distance between mean position of niches, what is the niche standard deviation, and what is the ratio of the two? What is the niche shape?
[23]
Shannon CE Weiner W (1949 ). The Mathematical Theory of Communication: Unknown Distance Function. University of Illinois Press, Urbana , USA.
[本文引用: 1]
[24]
Shen XL Wu YM Zhu YX Tong L Hu QL Jiang WJ Fan YQ Xia XF Mu XY Zhang ZX (2020 ). Community niche characteristics and interspecific associations of critically endangered species, Vitis baihuashannensis M. S. Kang et D. Z. Lu.
Plant Science Journal , 38 , 195 -204 .
[本文引用: 1]
[ 沈雪梨 , 吴远密 , 朱艺璇 , 童玲 , 胡巧立 , 蒋万杰 , 范雅倩 , 夏晓飞 , 沐先运 , 张志翔 (2020 ). 濒危植物百花山葡萄群落物种生态位特征与种间联结研究
植物科学学报 , 38 , 195 -204 .]
[本文引用: 1]
[25]
Shi FX Zhao CZ Gao FY Ren H Sheng YP Shi LL (2012 ). Dynamics of species diversity and interspecific association in shrub layer of naturally restored woodland on northern slope of Qilian Mountains, Northwest China
Chinese Journal of Ecology , 31 , 2177 -2183 .
[本文引用: 1]
[ 石福习 , 赵成章 , 高福元 , 任珩 , 盛亚萍 , 史丽丽 (2012 ). 祁连山北坡自然恢复林地灌木层物种多样性及种间关联动态
生态学杂志 , 31 , 2177 -2183 .]
[本文引用: 1]
[26]
Song YC (2001 ). Vegetation Ecology . East China Normal University Press , Shanghai .
[本文引用: 2]
[ 宋永昌 (2001 ). 植被生态学 . 华东师范大学出版社 , 上海 .]
[本文引用: 2]
[27]
Wang BS Peng SL (1985 ). Studies on the measuring techniques of interspecific association of lower-subtropical evergreen-broadleaved forests I. The exploration and the revision on the measuring formulas of interspecific association
Acta Phytoecologica et Geobotanica Sinica , 9 , 274 -285 .
[本文引用: 1]
[ 王伯荪 , 彭少麟 (1985 ). 南亚热带常绿阔叶林种间联结测定技术研究I. 种间联结测式的探讨与修正
植物生态学与地植物学丛刊 , 9 , 274 -285 .]
[本文引用: 1]
[28]
Wang WW Yang HL He KN Hu XB Wang H Dong M Xie ZW (2012 ). Niche and ecological response of herb layer in spruce plantation of Qilian Mountains
Acta Agrestia Sinica , 20 , 626 -630 .
[本文引用: 1]
[ 王伟伟 , 杨海龙 , 贺康宁 , 胡兴波 , 王辉 , 董梅 , 谢增武 (2012 ). 祁连山青海云杉人工林草本层生态位及生态响应研究
草地学报 , 20 , 626 -630 .]
DOI:10.11733/j.issn.1007-0435.2012.04.005
[本文引用: 1]
对祁连山区青海云杉(Picea crassifolia)人工林草本层植物群落的60个样方进行调查,运用Levins和Shannon-Wiener生态位宽度指数及Pianka 生态位重叠度指数,对该人工林下草本层各植物种群的生态位宽度及生态位重叠度进行定量研究。结果表明:珠芽蓼(Polygonum viviparum)和东方草莓(Fragaria orientalis)的生态位宽度最大,是青海云杉人工林下草本层的优势种,表明这2种物种在该人工林下具有广泛的适应性。生态位重叠指数表明:在青海云杉林下生态位宽度较大或较小的草本种群,他们之间的生态位重叠既有较大的,也有较小的,草本种群的生态位宽度和生态位重叠程度无相关性,可能是由于该区草本物种分布的斑块性和环境资源的高度空间异质性造成的。总体上看,青海云杉人工林下草本层的生态位重叠相对较低,对资源利用的相似度较低;从生态响应速率来看,高山老鹳草(Geranium pylzowianum)和马先蒿(Pedicularis)的发展性最强,在今后的演替过程中将会发展为青海云杉人工林下草本层的优势种群。
[29]
Wang ZT Bao Y Li Y (2006 ). Interspecific relationships in Salix shrub community on alpine grassland of east Qilian Mountains
Chinese Journal of Ecology , 25 , 1177 -1180 .
[本文引用: 1]
[ 王志泰 , 包玉 , 李毅 (2006 ). 东祁连山高寒草地柳灌丛群落种间关联性
生态学杂志 , 25 , 1177 -1180 .]
[本文引用: 1]
[30]
Wu ZJ He YL (2015 ). Spatial-temporal variation of surface air temperature and its influence by urbanization in central Yunnan plateau
Areal Research and Development , 34 , 137 -142 .
[本文引用: 1]
[ 吴志杰 , 何云玲 (2015 ). 云南中部区域气温变化特征及其受城市化影响程度分析
地域研究与开发 , 34 , 137 -142 .]
[本文引用: 1]
[31]
Xiao YM Xie JY Yao YP Liang SC Gao LN Zhang H Mo SZ (2021 ). Niche of dominant species in arbor layer of evergreen deciduous broad-leaved mixed forest in karst hills of Guilin, southwestern China
Acta Ecologica Sinica , 41 , 8159 -8170 .
[本文引用: 1]
[ 肖艳梅 , 解婧媛 , 姚义鹏 , 梁士楚 , 高丽娜 , 张惠 , 莫适祯 (2021 ). 桂林岩溶石山常绿落叶阔叶混交林乔木层优势物种生态位研究
生态学报 , 41 , 8159 -8170 .]
[本文引用: 1]
[32]
Xie ZQ Tang ZY Liu Q Xu WT (2019 ). Study on Carbon Budget of Shrub Ecosystems in China . Science Press , Beijing .
[本文引用: 1]
[ 谢宗强 , 唐志尧 , 刘庆 , 徐文婷 (2019 ). 中国碳丛生态系统碳收支研究 . 科学出版社 , 北京 .]
[本文引用: 1]
[33]
Xu MH Liu M Zhai DT Liu T (2016 ). A review of contents and methods used to analyze various aspects of plant interspecific associations
Acta Ecologica Sinica , 36 , 8224 -8233 .
[本文引用: 1]
[ 徐满厚 , 刘敏 , 翟大彤 , 刘彤 (2016 ). 植物种间联结研究内容与方法评述
生态学报 , 36 , 8224 -8233 .]
[本文引用: 1]
[34]
Yang WH Shen H Zhou ML Li WP Zhang S (2020 ). Seasonal variation analysis of the niche and interspecific association with respect to the dominant phytoplankton species in Nanhai Lake
China Environmental Science , 40 , 383 -391 .
[本文引用: 1]
[ 杨文焕 , 申涵 , 周明利 , 李卫平 , 张生 (2020 ). 包头南海湖浮游植物优势种生态位及种间联结性季节分析
中国环境科学 , 40 , 383 -391 .]
[本文引用: 1]
[35]
Yu XY Yu R Huang J Chen J Yuan ZY Yang F (2017 ). Niche characteristics of dominant plant populations of Loropetalum subcordatum community in Maolan karst forest, Guizhou
Chinese Journal of Ecology , 36 , 3470 -3478 .
[本文引用: 1]
[ 俞筱押 , 余瑞 , 黄娟 , 陈菊 , 袁子勇 , 杨芳 (2017 ). 贵州茂兰喀斯特森林四药门花群落优势种群生态位特征
生态学杂志 , 36 , 3470 -3478 .]
[本文引用: 1]
[36]
Zhang DM Zhao WZ Luo WC (2018 ). Niche and interspecific association of dominant plant species in saline-alkaline soils of desert steppe zone
Chinese Journal of Ecology , 37 , 1307 -1315 .
[本文引用: 1]
[ 张东梅 , 赵文智 , 罗维成 (2018 ). 荒漠草原带盐碱地优势植物生态位与种间联结
生态学杂志 , 37 , 1307 -1315 .]
[本文引用: 1]
[37]
Zhang JT (2018 ). Quantitative Ecology . 3rd ed. Science Press , Beijing .
[本文引用: 1]
[ 张金屯 (2018 ). 数量生态学 . 3版 . 科学出版社 , 北京 .]
[本文引用: 1]
[38]
Zhu LJ Su ZX Wang WW Zhang YS (2005 ). Numerical analysis on interspecific relationships in Davidia involucrata community in Sanjiang Wolong Nature Reserve
Chinese Journal of Ecology , 24 , 1167 -1171 .
[本文引用: 1]
[ 朱利君 , 苏智先 , 王伟伟 , 张亚爽 (2005 ). 卧龙自然保护区三江珙桐群落种间关系的数量分析
生态学杂志 , 24 , 1167 -1171 .]
[本文引用: 1]
1
2018
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
1
2018
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
异质生境对荒漠草原植物群落组成和种群生态位的影响
1
2019
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
异质生境对荒漠草原植物群落组成和种群生态位的影响
1
2019
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
云南2种主要灌丛土壤有机碳分布特征及其影响因子研究
2
2017
... 灌丛在水源涵养、碳固定、土壤保持和防风固沙等方面发挥重要的作用, 尤其在西南地区具有重要的生态价值(胡雄蛟等, 2021 ).云南省共有27种灌丛群系, 分属于河谷灌丛、山地灌丛、亚高山灌丛、高山灌丛4种植被型, 其中山地灌丛中的南烛(Vaccinium bracteatum )灌丛和云南杨梅(Myrica nana )灌丛分布较广, 两者面积共达到49.04 × 103 km2 (谢宗强等, 2019 ).云南杨梅抗污染能力强, 与根瘤菌共生, 具有杰出的水土保持和肥地功能, 同时具备较强的耐火、阻火能力, 因此可作优质的绿化树种及恢复植被的先锋树种(成宇, 2019 ).云南杨梅是滇中地区山地灌丛的优势种, 研究云南杨梅灌丛主要木本植物的生态位与种间联结不仅有利于更好地认识和发挥云南杨梅灌丛的生态功能, 而且可为揭示云南杨梅灌丛特征和维持云南杨梅灌丛的生物多样性提供科学依据.目前对云南杨梅灌丛的研究甚少, 仅包括对云南杨梅灌丛碳密度(郭子豪等, 2020 )、生物量(巩合德等, 2012 )和土壤有机碳(陈路红等, 2017 )等的研究.对云南杨梅灌丛群落的生态位和种间联结的研究目前还未有报道.本研究采用了Levins生态位宽度(B L )、Shannon生态位宽度(B S )、Schoener重叠指数(Cik )、Pianka重叠指数(Oik )、方差比率(VR)、χ2 检验、Pearson相关检验和Spearman秩相关检验进行数据分析与处理, 以说明下面几个科学问题: 1)云南杨梅灌丛主要木本植物的生态位宽度和生态位重叠特征; 2)云南杨梅灌丛主要木本植物的种间联结关系; 3)云南杨梅灌丛主要木本植物的生态位重叠和种间联结之间的联系. ...
... 研究样地位于云南省滇中地区昆明市、曲靖市和楚雄彝族自治州, 地理坐标为101.01°-104.37° E, 24.17°-25.48° N, 海拔1 936-2 737 m.滇中地处滇东高原, 地形以山地和山间盆地为主, 地势起伏和缓(吴志杰和何云玲, 2015 ); 土壤类型以红壤为主; 属低纬度亚热带高原季风气候, 2020年的年平均气温为17.72 ℃, 降水量为881.31 mm.滇中地区植被类型主要为河谷灌丛和山地灌丛, 河谷灌丛以余甘子(Phyllanthus emblica )为建群种, 山地灌丛以南烛、云南杨梅为建群种(陈路红等, 2017 ).样地中植被类型主要为山地灌丛, 主要物种包括云南杨梅、西南金丝梅(Hypericum henryi )、樟叶越桔(Vaccinium dunalianum )、乌鸦果(Vaccinium fragile )、铁仔(Myrsine africana )、碎米花(Rhododendron spiciferum )、茅莓(Rubus parvifolius )、云南松(Pinus yunnanensis )、滇青冈(Cyclobalanopsis glaucoides )、野丁香(Leptodermis potaninii )、粉叶小檗(Berberis pruinosa )、芳香白珠(Gaultheria fragrantissima )、钝叶橘红悬钩子(Rubus aurantiacus var. obtusifolius )、川梨(Pyrus pashia )、元江锥(Castanopsis orthacantha )、火棘(Pyracantha fortuneana )、亮毛杜鹃(Rhododendron microphyton )、高山锥(Castanopsis delavayi )、南烛、怒江红山茶(Camellia saluenensis )等.其中云南杨梅为优势种. ...
云南2种主要灌丛土壤有机碳分布特征及其影响因子研究
2
2017
... 灌丛在水源涵养、碳固定、土壤保持和防风固沙等方面发挥重要的作用, 尤其在西南地区具有重要的生态价值(胡雄蛟等, 2021 ).云南省共有27种灌丛群系, 分属于河谷灌丛、山地灌丛、亚高山灌丛、高山灌丛4种植被型, 其中山地灌丛中的南烛(Vaccinium bracteatum )灌丛和云南杨梅(Myrica nana )灌丛分布较广, 两者面积共达到49.04 × 103 km2 (谢宗强等, 2019 ).云南杨梅抗污染能力强, 与根瘤菌共生, 具有杰出的水土保持和肥地功能, 同时具备较强的耐火、阻火能力, 因此可作优质的绿化树种及恢复植被的先锋树种(成宇, 2019 ).云南杨梅是滇中地区山地灌丛的优势种, 研究云南杨梅灌丛主要木本植物的生态位与种间联结不仅有利于更好地认识和发挥云南杨梅灌丛的生态功能, 而且可为揭示云南杨梅灌丛特征和维持云南杨梅灌丛的生物多样性提供科学依据.目前对云南杨梅灌丛的研究甚少, 仅包括对云南杨梅灌丛碳密度(郭子豪等, 2020 )、生物量(巩合德等, 2012 )和土壤有机碳(陈路红等, 2017 )等的研究.对云南杨梅灌丛群落的生态位和种间联结的研究目前还未有报道.本研究采用了Levins生态位宽度(B L )、Shannon生态位宽度(B S )、Schoener重叠指数(Cik )、Pianka重叠指数(Oik )、方差比率(VR)、χ2 检验、Pearson相关检验和Spearman秩相关检验进行数据分析与处理, 以说明下面几个科学问题: 1)云南杨梅灌丛主要木本植物的生态位宽度和生态位重叠特征; 2)云南杨梅灌丛主要木本植物的种间联结关系; 3)云南杨梅灌丛主要木本植物的生态位重叠和种间联结之间的联系. ...
... 研究样地位于云南省滇中地区昆明市、曲靖市和楚雄彝族自治州, 地理坐标为101.01°-104.37° E, 24.17°-25.48° N, 海拔1 936-2 737 m.滇中地处滇东高原, 地形以山地和山间盆地为主, 地势起伏和缓(吴志杰和何云玲, 2015 ); 土壤类型以红壤为主; 属低纬度亚热带高原季风气候, 2020年的年平均气温为17.72 ℃, 降水量为881.31 mm.滇中地区植被类型主要为河谷灌丛和山地灌丛, 河谷灌丛以余甘子(Phyllanthus emblica )为建群种, 山地灌丛以南烛、云南杨梅为建群种(陈路红等, 2017 ).样地中植被类型主要为山地灌丛, 主要物种包括云南杨梅、西南金丝梅(Hypericum henryi )、樟叶越桔(Vaccinium dunalianum )、乌鸦果(Vaccinium fragile )、铁仔(Myrsine africana )、碎米花(Rhododendron spiciferum )、茅莓(Rubus parvifolius )、云南松(Pinus yunnanensis )、滇青冈(Cyclobalanopsis glaucoides )、野丁香(Leptodermis potaninii )、粉叶小檗(Berberis pruinosa )、芳香白珠(Gaultheria fragrantissima )、钝叶橘红悬钩子(Rubus aurantiacus var. obtusifolius )、川梨(Pyrus pashia )、元江锥(Castanopsis orthacantha )、火棘(Pyracantha fortuneana )、亮毛杜鹃(Rhododendron microphyton )、高山锥(Castanopsis delavayi )、南烛、怒江红山茶(Camellia saluenensis )等.其中云南杨梅为优势种. ...
海南岛霸王岭国家重点保护植物的生态位研究
1
2014
... 生态位宽度较大(较小)的物种与其他物种产生生态位重叠的概率也较大(较小)(陈玉凯等, 2014 ).本研究中云南杨梅出现在每个样方中, 生态位宽度最大, 与20个主要物种产生生态位重叠的概率为100%.亮毛杜鹃的生态位宽度较小, 与20个主要物种间的生态位重叠概率也较小.生态位宽度较小的物种间也出现生态位重叠值较高的情况, 如粉叶小檗和钝叶橘红悬钩子、粉叶小檗和怒江红山茶、钝叶橘红悬钩子和怒江红山茶, 这与张东梅等(2018 )的研究结论一致. ...
海南岛霸王岭国家重点保护植物的生态位研究
1
2014
... 生态位宽度较大(较小)的物种与其他物种产生生态位重叠的概率也较大(较小)(陈玉凯等, 2014 ).本研究中云南杨梅出现在每个样方中, 生态位宽度最大, 与20个主要物种产生生态位重叠的概率为100%.亮毛杜鹃的生态位宽度较小, 与20个主要物种间的生态位重叠概率也较小.生态位宽度较小的物种间也出现生态位重叠值较高的情况, 如粉叶小檗和钝叶橘红悬钩子、粉叶小檗和怒江红山茶、钝叶橘红悬钩子和怒江红山茶, 这与张东梅等(2018 )的研究结论一致. ...
1
2019
... 灌丛在水源涵养、碳固定、土壤保持和防风固沙等方面发挥重要的作用, 尤其在西南地区具有重要的生态价值(胡雄蛟等, 2021 ).云南省共有27种灌丛群系, 分属于河谷灌丛、山地灌丛、亚高山灌丛、高山灌丛4种植被型, 其中山地灌丛中的南烛(Vaccinium bracteatum )灌丛和云南杨梅(Myrica nana )灌丛分布较广, 两者面积共达到49.04 × 103 km2 (谢宗强等, 2019 ).云南杨梅抗污染能力强, 与根瘤菌共生, 具有杰出的水土保持和肥地功能, 同时具备较强的耐火、阻火能力, 因此可作优质的绿化树种及恢复植被的先锋树种(成宇, 2019 ).云南杨梅是滇中地区山地灌丛的优势种, 研究云南杨梅灌丛主要木本植物的生态位与种间联结不仅有利于更好地认识和发挥云南杨梅灌丛的生态功能, 而且可为揭示云南杨梅灌丛特征和维持云南杨梅灌丛的生物多样性提供科学依据.目前对云南杨梅灌丛的研究甚少, 仅包括对云南杨梅灌丛碳密度(郭子豪等, 2020 )、生物量(巩合德等, 2012 )和土壤有机碳(陈路红等, 2017 )等的研究.对云南杨梅灌丛群落的生态位和种间联结的研究目前还未有报道.本研究采用了Levins生态位宽度(B L )、Shannon生态位宽度(B S )、Schoener重叠指数(Cik )、Pianka重叠指数(Oik )、方差比率(VR)、χ2 检验、Pearson相关检验和Spearman秩相关检验进行数据分析与处理, 以说明下面几个科学问题: 1)云南杨梅灌丛主要木本植物的生态位宽度和生态位重叠特征; 2)云南杨梅灌丛主要木本植物的种间联结关系; 3)云南杨梅灌丛主要木本植物的生态位重叠和种间联结之间的联系. ...
1
2019
... 灌丛在水源涵养、碳固定、土壤保持和防风固沙等方面发挥重要的作用, 尤其在西南地区具有重要的生态价值(胡雄蛟等, 2021 ).云南省共有27种灌丛群系, 分属于河谷灌丛、山地灌丛、亚高山灌丛、高山灌丛4种植被型, 其中山地灌丛中的南烛(Vaccinium bracteatum )灌丛和云南杨梅(Myrica nana )灌丛分布较广, 两者面积共达到49.04 × 103 km2 (谢宗强等, 2019 ).云南杨梅抗污染能力强, 与根瘤菌共生, 具有杰出的水土保持和肥地功能, 同时具备较强的耐火、阻火能力, 因此可作优质的绿化树种及恢复植被的先锋树种(成宇, 2019 ).云南杨梅是滇中地区山地灌丛的优势种, 研究云南杨梅灌丛主要木本植物的生态位与种间联结不仅有利于更好地认识和发挥云南杨梅灌丛的生态功能, 而且可为揭示云南杨梅灌丛特征和维持云南杨梅灌丛的生物多样性提供科学依据.目前对云南杨梅灌丛的研究甚少, 仅包括对云南杨梅灌丛碳密度(郭子豪等, 2020 )、生物量(巩合德等, 2012 )和土壤有机碳(陈路红等, 2017 )等的研究.对云南杨梅灌丛群落的生态位和种间联结的研究目前还未有报道.本研究采用了Levins生态位宽度(B L )、Shannon生态位宽度(B S )、Schoener重叠指数(Cik )、Pianka重叠指数(Oik )、方差比率(VR)、χ2 检验、Pearson相关检验和Spearman秩相关检验进行数据分析与处理, 以说明下面几个科学问题: 1)云南杨梅灌丛主要木本植物的生态位宽度和生态位重叠特征; 2)云南杨梅灌丛主要木本植物的种间联结关系; 3)云南杨梅灌丛主要木本植物的生态位重叠和种间联结之间的联系. ...
乌兰布和沙漠典型灌木群落多样性及其生态位
1
2020
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
乌兰布和沙漠典型灌木群落多样性及其生态位
1
2020
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
长江源区高寒湿地植物群落主要种群种间关系分析
1
2011
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
长江源区高寒湿地植物群落主要种群种间关系分析
1
2011
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
云南杨梅灌丛生物量的分配特征
1
2012
... 灌丛在水源涵养、碳固定、土壤保持和防风固沙等方面发挥重要的作用, 尤其在西南地区具有重要的生态价值(胡雄蛟等, 2021 ).云南省共有27种灌丛群系, 分属于河谷灌丛、山地灌丛、亚高山灌丛、高山灌丛4种植被型, 其中山地灌丛中的南烛(Vaccinium bracteatum )灌丛和云南杨梅(Myrica nana )灌丛分布较广, 两者面积共达到49.04 × 103 km2 (谢宗强等, 2019 ).云南杨梅抗污染能力强, 与根瘤菌共生, 具有杰出的水土保持和肥地功能, 同时具备较强的耐火、阻火能力, 因此可作优质的绿化树种及恢复植被的先锋树种(成宇, 2019 ).云南杨梅是滇中地区山地灌丛的优势种, 研究云南杨梅灌丛主要木本植物的生态位与种间联结不仅有利于更好地认识和发挥云南杨梅灌丛的生态功能, 而且可为揭示云南杨梅灌丛特征和维持云南杨梅灌丛的生物多样性提供科学依据.目前对云南杨梅灌丛的研究甚少, 仅包括对云南杨梅灌丛碳密度(郭子豪等, 2020 )、生物量(巩合德等, 2012 )和土壤有机碳(陈路红等, 2017 )等的研究.对云南杨梅灌丛群落的生态位和种间联结的研究目前还未有报道.本研究采用了Levins生态位宽度(B L )、Shannon生态位宽度(B S )、Schoener重叠指数(Cik )、Pianka重叠指数(Oik )、方差比率(VR)、χ2 检验、Pearson相关检验和Spearman秩相关检验进行数据分析与处理, 以说明下面几个科学问题: 1)云南杨梅灌丛主要木本植物的生态位宽度和生态位重叠特征; 2)云南杨梅灌丛主要木本植物的种间联结关系; 3)云南杨梅灌丛主要木本植物的生态位重叠和种间联结之间的联系. ...
云南杨梅灌丛生物量的分配特征
1
2012
... 灌丛在水源涵养、碳固定、土壤保持和防风固沙等方面发挥重要的作用, 尤其在西南地区具有重要的生态价值(胡雄蛟等, 2021 ).云南省共有27种灌丛群系, 分属于河谷灌丛、山地灌丛、亚高山灌丛、高山灌丛4种植被型, 其中山地灌丛中的南烛(Vaccinium bracteatum )灌丛和云南杨梅(Myrica nana )灌丛分布较广, 两者面积共达到49.04 × 103 km2 (谢宗强等, 2019 ).云南杨梅抗污染能力强, 与根瘤菌共生, 具有杰出的水土保持和肥地功能, 同时具备较强的耐火、阻火能力, 因此可作优质的绿化树种及恢复植被的先锋树种(成宇, 2019 ).云南杨梅是滇中地区山地灌丛的优势种, 研究云南杨梅灌丛主要木本植物的生态位与种间联结不仅有利于更好地认识和发挥云南杨梅灌丛的生态功能, 而且可为揭示云南杨梅灌丛特征和维持云南杨梅灌丛的生物多样性提供科学依据.目前对云南杨梅灌丛的研究甚少, 仅包括对云南杨梅灌丛碳密度(郭子豪等, 2020 )、生物量(巩合德等, 2012 )和土壤有机碳(陈路红等, 2017 )等的研究.对云南杨梅灌丛群落的生态位和种间联结的研究目前还未有报道.本研究采用了Levins生态位宽度(B L )、Shannon生态位宽度(B S )、Schoener重叠指数(Cik )、Pianka重叠指数(Oik )、方差比率(VR)、χ2 检验、Pearson相关检验和Spearman秩相关检验进行数据分析与处理, 以说明下面几个科学问题: 1)云南杨梅灌丛主要木本植物的生态位宽度和生态位重叠特征; 2)云南杨梅灌丛主要木本植物的种间联结关系; 3)云南杨梅灌丛主要木本植物的生态位重叠和种间联结之间的联系. ...
滇中地区矮杨梅灌丛碳密度分配格局研究
1
2020
... 灌丛在水源涵养、碳固定、土壤保持和防风固沙等方面发挥重要的作用, 尤其在西南地区具有重要的生态价值(胡雄蛟等, 2021 ).云南省共有27种灌丛群系, 分属于河谷灌丛、山地灌丛、亚高山灌丛、高山灌丛4种植被型, 其中山地灌丛中的南烛(Vaccinium bracteatum )灌丛和云南杨梅(Myrica nana )灌丛分布较广, 两者面积共达到49.04 × 103 km2 (谢宗强等, 2019 ).云南杨梅抗污染能力强, 与根瘤菌共生, 具有杰出的水土保持和肥地功能, 同时具备较强的耐火、阻火能力, 因此可作优质的绿化树种及恢复植被的先锋树种(成宇, 2019 ).云南杨梅是滇中地区山地灌丛的优势种, 研究云南杨梅灌丛主要木本植物的生态位与种间联结不仅有利于更好地认识和发挥云南杨梅灌丛的生态功能, 而且可为揭示云南杨梅灌丛特征和维持云南杨梅灌丛的生物多样性提供科学依据.目前对云南杨梅灌丛的研究甚少, 仅包括对云南杨梅灌丛碳密度(郭子豪等, 2020 )、生物量(巩合德等, 2012 )和土壤有机碳(陈路红等, 2017 )等的研究.对云南杨梅灌丛群落的生态位和种间联结的研究目前还未有报道.本研究采用了Levins生态位宽度(B L )、Shannon生态位宽度(B S )、Schoener重叠指数(Cik )、Pianka重叠指数(Oik )、方差比率(VR)、χ2 检验、Pearson相关检验和Spearman秩相关检验进行数据分析与处理, 以说明下面几个科学问题: 1)云南杨梅灌丛主要木本植物的生态位宽度和生态位重叠特征; 2)云南杨梅灌丛主要木本植物的种间联结关系; 3)云南杨梅灌丛主要木本植物的生态位重叠和种间联结之间的联系. ...
滇中地区矮杨梅灌丛碳密度分配格局研究
1
2020
... 灌丛在水源涵养、碳固定、土壤保持和防风固沙等方面发挥重要的作用, 尤其在西南地区具有重要的生态价值(胡雄蛟等, 2021 ).云南省共有27种灌丛群系, 分属于河谷灌丛、山地灌丛、亚高山灌丛、高山灌丛4种植被型, 其中山地灌丛中的南烛(Vaccinium bracteatum )灌丛和云南杨梅(Myrica nana )灌丛分布较广, 两者面积共达到49.04 × 103 km2 (谢宗强等, 2019 ).云南杨梅抗污染能力强, 与根瘤菌共生, 具有杰出的水土保持和肥地功能, 同时具备较强的耐火、阻火能力, 因此可作优质的绿化树种及恢复植被的先锋树种(成宇, 2019 ).云南杨梅是滇中地区山地灌丛的优势种, 研究云南杨梅灌丛主要木本植物的生态位与种间联结不仅有利于更好地认识和发挥云南杨梅灌丛的生态功能, 而且可为揭示云南杨梅灌丛特征和维持云南杨梅灌丛的生物多样性提供科学依据.目前对云南杨梅灌丛的研究甚少, 仅包括对云南杨梅灌丛碳密度(郭子豪等, 2020 )、生物量(巩合德等, 2012 )和土壤有机碳(陈路红等, 2017 )等的研究.对云南杨梅灌丛群落的生态位和种间联结的研究目前还未有报道.本研究采用了Levins生态位宽度(B L )、Shannon生态位宽度(B S )、Schoener重叠指数(Cik )、Pianka重叠指数(Oik )、方差比率(VR)、χ2 检验、Pearson相关检验和Spearman秩相关检验进行数据分析与处理, 以说明下面几个科学问题: 1)云南杨梅灌丛主要木本植物的生态位宽度和生态位重叠特征; 2)云南杨梅灌丛主要木本植物的种间联结关系; 3)云南杨梅灌丛主要木本植物的生态位重叠和种间联结之间的联系. ...
庐山常绿阔叶、落叶阔叶混交林乔木种群种间联结性研究
1
1997
... 多数研究认为: 种间正联结关系越强, 其生态位重叠值就越大; 反之, 种间负联结关系越强, 其生态位重叠值就越小(郭志华等, 1997 ; 霍红等, 2013 ).本研究显示, 滇中云南杨梅灌丛木本植物主要物种的Pearson相关系数、Spearman秩相关系数与生态位重叠指数之间均存在极显著的正相关关系.如元江锥和高山锥的Pearson相关系数和Spearman秩相关系数值较大, 种对呈极显著正联结关系, 生态位重叠值也较大.但也有研究认为种间负关联关系则比较复杂, 由生境差异导致的负关联关系生态位重叠值低, 而由资源竞争引起的负关联关系生态位重叠值高(刘润红等, 2020 ).如本研究中西南金丝梅和南烛, 西南金丝梅和亮毛杜鹃的Pearson相关系数和Spearman秩相关系数值为负值, 种间关系为负联结, 其种对间的生态位重叠值为0, 说明这两个种对间的负联结关系是由生境差异引起的. ...
庐山常绿阔叶、落叶阔叶混交林乔木种群种间联结性研究
1
1997
... 多数研究认为: 种间正联结关系越强, 其生态位重叠值就越大; 反之, 种间负联结关系越强, 其生态位重叠值就越小(郭志华等, 1997 ; 霍红等, 2013 ).本研究显示, 滇中云南杨梅灌丛木本植物主要物种的Pearson相关系数、Spearman秩相关系数与生态位重叠指数之间均存在极显著的正相关关系.如元江锥和高山锥的Pearson相关系数和Spearman秩相关系数值较大, 种对呈极显著正联结关系, 生态位重叠值也较大.但也有研究认为种间负关联关系则比较复杂, 由生境差异导致的负关联关系生态位重叠值低, 而由资源竞争引起的负关联关系生态位重叠值高(刘润红等, 2020 ).如本研究中西南金丝梅和南烛, 西南金丝梅和亮毛杜鹃的Pearson相关系数和Spearman秩相关系数值为负值, 种间关系为负联结, 其种对间的生态位重叠值为0, 说明这两个种对间的负联结关系是由生境差异引起的. ...
中国灌丛生态系统主要调节服务及价值评估
1
2021
... 灌丛在水源涵养、碳固定、土壤保持和防风固沙等方面发挥重要的作用, 尤其在西南地区具有重要的生态价值(胡雄蛟等, 2021 ).云南省共有27种灌丛群系, 分属于河谷灌丛、山地灌丛、亚高山灌丛、高山灌丛4种植被型, 其中山地灌丛中的南烛(Vaccinium bracteatum )灌丛和云南杨梅(Myrica nana )灌丛分布较广, 两者面积共达到49.04 × 103 km2 (谢宗强等, 2019 ).云南杨梅抗污染能力强, 与根瘤菌共生, 具有杰出的水土保持和肥地功能, 同时具备较强的耐火、阻火能力, 因此可作优质的绿化树种及恢复植被的先锋树种(成宇, 2019 ).云南杨梅是滇中地区山地灌丛的优势种, 研究云南杨梅灌丛主要木本植物的生态位与种间联结不仅有利于更好地认识和发挥云南杨梅灌丛的生态功能, 而且可为揭示云南杨梅灌丛特征和维持云南杨梅灌丛的生物多样性提供科学依据.目前对云南杨梅灌丛的研究甚少, 仅包括对云南杨梅灌丛碳密度(郭子豪等, 2020 )、生物量(巩合德等, 2012 )和土壤有机碳(陈路红等, 2017 )等的研究.对云南杨梅灌丛群落的生态位和种间联结的研究目前还未有报道.本研究采用了Levins生态位宽度(B L )、Shannon生态位宽度(B S )、Schoener重叠指数(Cik )、Pianka重叠指数(Oik )、方差比率(VR)、χ2 检验、Pearson相关检验和Spearman秩相关检验进行数据分析与处理, 以说明下面几个科学问题: 1)云南杨梅灌丛主要木本植物的生态位宽度和生态位重叠特征; 2)云南杨梅灌丛主要木本植物的种间联结关系; 3)云南杨梅灌丛主要木本植物的生态位重叠和种间联结之间的联系. ...
中国灌丛生态系统主要调节服务及价值评估
1
2021
... 灌丛在水源涵养、碳固定、土壤保持和防风固沙等方面发挥重要的作用, 尤其在西南地区具有重要的生态价值(胡雄蛟等, 2021 ).云南省共有27种灌丛群系, 分属于河谷灌丛、山地灌丛、亚高山灌丛、高山灌丛4种植被型, 其中山地灌丛中的南烛(Vaccinium bracteatum )灌丛和云南杨梅(Myrica nana )灌丛分布较广, 两者面积共达到49.04 × 103 km2 (谢宗强等, 2019 ).云南杨梅抗污染能力强, 与根瘤菌共生, 具有杰出的水土保持和肥地功能, 同时具备较强的耐火、阻火能力, 因此可作优质的绿化树种及恢复植被的先锋树种(成宇, 2019 ).云南杨梅是滇中地区山地灌丛的优势种, 研究云南杨梅灌丛主要木本植物的生态位与种间联结不仅有利于更好地认识和发挥云南杨梅灌丛的生态功能, 而且可为揭示云南杨梅灌丛特征和维持云南杨梅灌丛的生物多样性提供科学依据.目前对云南杨梅灌丛的研究甚少, 仅包括对云南杨梅灌丛碳密度(郭子豪等, 2020 )、生物量(巩合德等, 2012 )和土壤有机碳(陈路红等, 2017 )等的研究.对云南杨梅灌丛群落的生态位和种间联结的研究目前还未有报道.本研究采用了Levins生态位宽度(B L )、Shannon生态位宽度(B S )、Schoener重叠指数(Cik )、Pianka重叠指数(Oik )、方差比率(VR)、χ2 检验、Pearson相关检验和Spearman秩相关检验进行数据分析与处理, 以说明下面几个科学问题: 1)云南杨梅灌丛主要木本植物的生态位宽度和生态位重叠特征; 2)云南杨梅灌丛主要木本植物的种间联结关系; 3)云南杨梅灌丛主要木本植物的生态位重叠和种间联结之间的联系. ...
额济纳绿洲植物群落种间关系和生态位研究
2
2013
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
... 多数研究认为: 种间正联结关系越强, 其生态位重叠值就越大; 反之, 种间负联结关系越强, 其生态位重叠值就越小(郭志华等, 1997 ; 霍红等, 2013 ).本研究显示, 滇中云南杨梅灌丛木本植物主要物种的Pearson相关系数、Spearman秩相关系数与生态位重叠指数之间均存在极显著的正相关关系.如元江锥和高山锥的Pearson相关系数和Spearman秩相关系数值较大, 种对呈极显著正联结关系, 生态位重叠值也较大.但也有研究认为种间负关联关系则比较复杂, 由生境差异导致的负关联关系生态位重叠值低, 而由资源竞争引起的负关联关系生态位重叠值高(刘润红等, 2020 ).如本研究中西南金丝梅和南烛, 西南金丝梅和亮毛杜鹃的Pearson相关系数和Spearman秩相关系数值为负值, 种间关系为负联结, 其种对间的生态位重叠值为0, 说明这两个种对间的负联结关系是由生境差异引起的. ...
额济纳绿洲植物群落种间关系和生态位研究
2
2013
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
... 多数研究认为: 种间正联结关系越强, 其生态位重叠值就越大; 反之, 种间负联结关系越强, 其生态位重叠值就越小(郭志华等, 1997 ; 霍红等, 2013 ).本研究显示, 滇中云南杨梅灌丛木本植物主要物种的Pearson相关系数、Spearman秩相关系数与生态位重叠指数之间均存在极显著的正相关关系.如元江锥和高山锥的Pearson相关系数和Spearman秩相关系数值较大, 种对呈极显著正联结关系, 生态位重叠值也较大.但也有研究认为种间负关联关系则比较复杂, 由生境差异导致的负关联关系生态位重叠值低, 而由资源竞争引起的负关联关系生态位重叠值高(刘润红等, 2020 ).如本研究中西南金丝梅和南烛, 西南金丝梅和亮毛杜鹃的Pearson相关系数和Spearman秩相关系数值为负值, 种间关系为负联结, 其种对间的生态位重叠值为0, 说明这两个种对间的负联结关系是由生境差异引起的. ...
1
1968
... 利用B L (Levins, 1968 )和B S (Shannon & Weiner, 1949 )计算生态位宽度, 公式如下: ...
云顶山亚高山草甸优势种群生态位研究
1
2010
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
云顶山亚高山草甸优势种群生态位研究
1
2010
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
关帝山亚高山灌丛群落和草甸群落优势种的种间关系
1
2004
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
关帝山亚高山灌丛群落和草甸群落优势种的种间关系
1
2004
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
岷江上游茂县半干旱河谷灌丛优势种间关系的研究
1
1996
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
岷江上游茂县半干旱河谷灌丛优势种间关系的研究
1
1996
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
桂林岩溶石山青冈群落灌木层主要物种生态位与种间联结
2
2020
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
... 多数研究认为: 种间正联结关系越强, 其生态位重叠值就越大; 反之, 种间负联结关系越强, 其生态位重叠值就越小(郭志华等, 1997 ; 霍红等, 2013 ).本研究显示, 滇中云南杨梅灌丛木本植物主要物种的Pearson相关系数、Spearman秩相关系数与生态位重叠指数之间均存在极显著的正相关关系.如元江锥和高山锥的Pearson相关系数和Spearman秩相关系数值较大, 种对呈极显著正联结关系, 生态位重叠值也较大.但也有研究认为种间负关联关系则比较复杂, 由生境差异导致的负关联关系生态位重叠值低, 而由资源竞争引起的负关联关系生态位重叠值高(刘润红等, 2020 ).如本研究中西南金丝梅和南烛, 西南金丝梅和亮毛杜鹃的Pearson相关系数和Spearman秩相关系数值为负值, 种间关系为负联结, 其种对间的生态位重叠值为0, 说明这两个种对间的负联结关系是由生境差异引起的. ...
桂林岩溶石山青冈群落灌木层主要物种生态位与种间联结
2
2020
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
... 多数研究认为: 种间正联结关系越强, 其生态位重叠值就越大; 反之, 种间负联结关系越强, 其生态位重叠值就越小(郭志华等, 1997 ; 霍红等, 2013 ).本研究显示, 滇中云南杨梅灌丛木本植物主要物种的Pearson相关系数、Spearman秩相关系数与生态位重叠指数之间均存在极显著的正相关关系.如元江锥和高山锥的Pearson相关系数和Spearman秩相关系数值较大, 种对呈极显著正联结关系, 生态位重叠值也较大.但也有研究认为种间负关联关系则比较复杂, 由生境差异导致的负关联关系生态位重叠值低, 而由资源竞争引起的负关联关系生态位重叠值高(刘润红等, 2020 ).如本研究中西南金丝梅和南烛, 西南金丝梅和亮毛杜鹃的Pearson相关系数和Spearman秩相关系数值为负值, 种间关系为负联结, 其种对间的生态位重叠值为0, 说明这两个种对间的负联结关系是由生境差异引起的. ...
不同生长基质的苔藓植物优势种生态位与种间联结
1
2019
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
不同生长基质的苔藓植物优势种生态位与种间联结
1
2019
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
1
2015
... 云南杨梅的生态习性是中等喜光, 不耐强烈的日照, 喜温暖湿润气候, 喜空气湿度大, 宜排水良好的酸性土壤, 稍耐贫瘠(欧晓昆等, 2015 ).云南杨梅在χ2 检验下与其他物种多呈不显著负联结关系, 在Pearson相关检验和Spearman秩相关检验中与半数物种呈不显著正联结关系, 包括西南金丝梅、茅莓、滇青冈、野丁香、高山锥等, 这些物种的共同特性是喜光, 较耐阴, 在一般土壤上均能够较好地生长, 随着群落的演化, 这些物种会发展为伴生种, 因此需要加大对这些物种的保护; 半数呈不显著负联结关系, 包括樟叶越桔、乌鸦果、碎米花、云南松等, 可能因为这些物种和云南杨梅均喜酸性土壤和光, 它们和云南杨梅产生了竞争, 因而形成负联结关系.当环境容量不够时, 这些物种和云南杨梅的竞争会加剧, 在保护云南杨梅群落的过程中, 需要适当协调这些物种和云南杨梅之间的关系. ...
1
2015
... 云南杨梅的生态习性是中等喜光, 不耐强烈的日照, 喜温暖湿润气候, 喜空气湿度大, 宜排水良好的酸性土壤, 稍耐贫瘠(欧晓昆等, 2015 ).云南杨梅在χ2 检验下与其他物种多呈不显著负联结关系, 在Pearson相关检验和Spearman秩相关检验中与半数物种呈不显著正联结关系, 包括西南金丝梅、茅莓、滇青冈、野丁香、高山锥等, 这些物种的共同特性是喜光, 较耐阴, 在一般土壤上均能够较好地生长, 随着群落的演化, 这些物种会发展为伴生种, 因此需要加大对这些物种的保护; 半数呈不显著负联结关系, 包括樟叶越桔、乌鸦果、碎米花、云南松等, 可能因为这些物种和云南杨梅均喜酸性土壤和光, 它们和云南杨梅产生了竞争, 因而形成负联结关系.当环境容量不够时, 这些物种和云南杨梅的竞争会加剧, 在保护云南杨梅群落的过程中, 需要适当协调这些物种和云南杨梅之间的关系. ...
The structure of lizard communities
1
1973
... 利用Cik (Schoener, 1974 )和Oik (Pianka, 1973 )计算生态位重叠程度, 公式如下: ...
A variance test for detecting species associations, with some example applications
1
1984
... 使用VR (Schluter, 1984 )检验总体联结性, 以统计量W检验联结性的显著度. ...
Resource partitioning in ecological communities
1
1974
... 利用Cik (Schoener, 1974 )和Oik (Pianka, 1973 )计算生态位重叠程度, 公式如下: ...
1
1949
... 利用B L (Levins, 1968 )和B S (Shannon & Weiner, 1949 )计算生态位宽度, 公式如下: ...
濒危植物百花山葡萄群落物种生态位特征与种间联结研究
1
2020
... 生态位重叠可以解释物种间竞争或共存关系, 通常认为重叠值大于0.500, 则种间具有较大的生态和资源利用相似性, 竞争比较严重(沈雪梨等, 2020 ).生态位重叠值大于0.500的种对数均较少, 且为0的种对数较多, 表明云南杨梅灌丛主要木本植物之间对环境资源需求的相似性不大, 种间竞争较弱, 种间关系趋于稳定.云南杨梅与西南金丝梅、樟叶越桔、铁仔的生态位重叠程度较大, 可能因为这些物种具有较大的生态位宽度值和重要值; 与亮毛杜鹃、芳香白珠、怒江红山茶的生态位重叠程度较小, 可能因为它们对资源需求的相似性低, 出现了不同程度的生态位分化, 且这些物种在样方中数量分布较为集中, 相对频度较低. ...
濒危植物百花山葡萄群落物种生态位特征与种间联结研究
1
2020
... 生态位重叠可以解释物种间竞争或共存关系, 通常认为重叠值大于0.500, 则种间具有较大的生态和资源利用相似性, 竞争比较严重(沈雪梨等, 2020 ).生态位重叠值大于0.500的种对数均较少, 且为0的种对数较多, 表明云南杨梅灌丛主要木本植物之间对环境资源需求的相似性不大, 种间竞争较弱, 种间关系趋于稳定.云南杨梅与西南金丝梅、樟叶越桔、铁仔的生态位重叠程度较大, 可能因为这些物种具有较大的生态位宽度值和重要值; 与亮毛杜鹃、芳香白珠、怒江红山茶的生态位重叠程度较小, 可能因为它们对资源需求的相似性低, 出现了不同程度的生态位分化, 且这些物种在样方中数量分布较为集中, 相对频度较低. ...
祁连山北坡自然恢复林地灌木层物种多样性及种间关联动态
1
2012
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
祁连山北坡自然恢复林地灌木层物种多样性及种间关联动态
1
2012
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
2
2001
... 重要值(宋永昌, 2001 )计算公式如下: ...
... 式中, N 为总样方数, a 为2个物种同时出现的样方数, b 、c 分别为2个物种单独出现的样方数, d 为2个物种都不出现的样方数.当ad = bc 时, 两个种相互独立; 当ad > bc 时, 两个种之间呈正联结关系; 当ad < bc 时, 两个种之间呈负联结关系.若χ 2 < 3.841 (p > 0.05), 表明种对间呈不显著联结关系, 种对间基本独立; 若3.841 ≤ χ 2 ≤ 6.635 (0.01 ≤ p ≤ 0.05), 表明种对间呈显著联结关系; 若χ 2 > 6.635 (p < 0.01), 表明种对间呈极显著联结关系(宋永昌, 2001 ). ...
2
2001
... 重要值(宋永昌, 2001 )计算公式如下: ...
... 式中, N 为总样方数, a 为2个物种同时出现的样方数, b 、c 分别为2个物种单独出现的样方数, d 为2个物种都不出现的样方数.当ad = bc 时, 两个种相互独立; 当ad > bc 时, 两个种之间呈正联结关系; 当ad < bc 时, 两个种之间呈负联结关系.若χ 2 < 3.841 (p > 0.05), 表明种对间呈不显著联结关系, 种对间基本独立; 若3.841 ≤ χ 2 ≤ 6.635 (0.01 ≤ p ≤ 0.05), 表明种对间呈显著联结关系; 若χ 2 > 6.635 (p < 0.01), 表明种对间呈极显著联结关系(宋永昌, 2001 ). ...
南亚热带常绿阔叶林种间联结测定技术研究I. 种间联结测式的探讨与修正
1
1985
... 由于样地内云南杨梅出现的频率为100%, 导致χ 2 检验公式中的分母为0, 不能准确反映云南杨梅和其他物种之间的种间联结关系, 因此给该种一个加权值, 令其在公式中的c 、d 值均为1 (王伯荪和彭少麟, 1985 ). ...
南亚热带常绿阔叶林种间联结测定技术研究I. 种间联结测式的探讨与修正
1
1985
... 由于样地内云南杨梅出现的频率为100%, 导致χ 2 检验公式中的分母为0, 不能准确反映云南杨梅和其他物种之间的种间联结关系, 因此给该种一个加权值, 令其在公式中的c 、d 值均为1 (王伯荪和彭少麟, 1985 ). ...
祁连山青海云杉人工林草本层生态位及生态响应研究
1
2012
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
祁连山青海云杉人工林草本层生态位及生态响应研究
1
2012
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
东祁连山高寒草地柳灌丛群落种间关联性
1
2006
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
东祁连山高寒草地柳灌丛群落种间关联性
1
2006
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
云南中部区域气温变化特征及其受城市化影响程度分析
1
2015
... 研究样地位于云南省滇中地区昆明市、曲靖市和楚雄彝族自治州, 地理坐标为101.01°-104.37° E, 24.17°-25.48° N, 海拔1 936-2 737 m.滇中地处滇东高原, 地形以山地和山间盆地为主, 地势起伏和缓(吴志杰和何云玲, 2015 ); 土壤类型以红壤为主; 属低纬度亚热带高原季风气候, 2020年的年平均气温为17.72 ℃, 降水量为881.31 mm.滇中地区植被类型主要为河谷灌丛和山地灌丛, 河谷灌丛以余甘子(Phyllanthus emblica )为建群种, 山地灌丛以南烛、云南杨梅为建群种(陈路红等, 2017 ).样地中植被类型主要为山地灌丛, 主要物种包括云南杨梅、西南金丝梅(Hypericum henryi )、樟叶越桔(Vaccinium dunalianum )、乌鸦果(Vaccinium fragile )、铁仔(Myrsine africana )、碎米花(Rhododendron spiciferum )、茅莓(Rubus parvifolius )、云南松(Pinus yunnanensis )、滇青冈(Cyclobalanopsis glaucoides )、野丁香(Leptodermis potaninii )、粉叶小檗(Berberis pruinosa )、芳香白珠(Gaultheria fragrantissima )、钝叶橘红悬钩子(Rubus aurantiacus var. obtusifolius )、川梨(Pyrus pashia )、元江锥(Castanopsis orthacantha )、火棘(Pyracantha fortuneana )、亮毛杜鹃(Rhododendron microphyton )、高山锥(Castanopsis delavayi )、南烛、怒江红山茶(Camellia saluenensis )等.其中云南杨梅为优势种. ...
云南中部区域气温变化特征及其受城市化影响程度分析
1
2015
... 研究样地位于云南省滇中地区昆明市、曲靖市和楚雄彝族自治州, 地理坐标为101.01°-104.37° E, 24.17°-25.48° N, 海拔1 936-2 737 m.滇中地处滇东高原, 地形以山地和山间盆地为主, 地势起伏和缓(吴志杰和何云玲, 2015 ); 土壤类型以红壤为主; 属低纬度亚热带高原季风气候, 2020年的年平均气温为17.72 ℃, 降水量为881.31 mm.滇中地区植被类型主要为河谷灌丛和山地灌丛, 河谷灌丛以余甘子(Phyllanthus emblica )为建群种, 山地灌丛以南烛、云南杨梅为建群种(陈路红等, 2017 ).样地中植被类型主要为山地灌丛, 主要物种包括云南杨梅、西南金丝梅(Hypericum henryi )、樟叶越桔(Vaccinium dunalianum )、乌鸦果(Vaccinium fragile )、铁仔(Myrsine africana )、碎米花(Rhododendron spiciferum )、茅莓(Rubus parvifolius )、云南松(Pinus yunnanensis )、滇青冈(Cyclobalanopsis glaucoides )、野丁香(Leptodermis potaninii )、粉叶小檗(Berberis pruinosa )、芳香白珠(Gaultheria fragrantissima )、钝叶橘红悬钩子(Rubus aurantiacus var. obtusifolius )、川梨(Pyrus pashia )、元江锥(Castanopsis orthacantha )、火棘(Pyracantha fortuneana )、亮毛杜鹃(Rhododendron microphyton )、高山锥(Castanopsis delavayi )、南烛、怒江红山茶(Camellia saluenensis )等.其中云南杨梅为优势种. ...
桂林岩溶石山常绿落叶阔叶混交林乔木层优势物种生态位研究
1
2021
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
桂林岩溶石山常绿落叶阔叶混交林乔木层优势物种生态位研究
1
2021
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
1
2019
... 灌丛在水源涵养、碳固定、土壤保持和防风固沙等方面发挥重要的作用, 尤其在西南地区具有重要的生态价值(胡雄蛟等, 2021 ).云南省共有27种灌丛群系, 分属于河谷灌丛、山地灌丛、亚高山灌丛、高山灌丛4种植被型, 其中山地灌丛中的南烛(Vaccinium bracteatum )灌丛和云南杨梅(Myrica nana )灌丛分布较广, 两者面积共达到49.04 × 103 km2 (谢宗强等, 2019 ).云南杨梅抗污染能力强, 与根瘤菌共生, 具有杰出的水土保持和肥地功能, 同时具备较强的耐火、阻火能力, 因此可作优质的绿化树种及恢复植被的先锋树种(成宇, 2019 ).云南杨梅是滇中地区山地灌丛的优势种, 研究云南杨梅灌丛主要木本植物的生态位与种间联结不仅有利于更好地认识和发挥云南杨梅灌丛的生态功能, 而且可为揭示云南杨梅灌丛特征和维持云南杨梅灌丛的生物多样性提供科学依据.目前对云南杨梅灌丛的研究甚少, 仅包括对云南杨梅灌丛碳密度(郭子豪等, 2020 )、生物量(巩合德等, 2012 )和土壤有机碳(陈路红等, 2017 )等的研究.对云南杨梅灌丛群落的生态位和种间联结的研究目前还未有报道.本研究采用了Levins生态位宽度(B L )、Shannon生态位宽度(B S )、Schoener重叠指数(Cik )、Pianka重叠指数(Oik )、方差比率(VR)、χ2 检验、Pearson相关检验和Spearman秩相关检验进行数据分析与处理, 以说明下面几个科学问题: 1)云南杨梅灌丛主要木本植物的生态位宽度和生态位重叠特征; 2)云南杨梅灌丛主要木本植物的种间联结关系; 3)云南杨梅灌丛主要木本植物的生态位重叠和种间联结之间的联系. ...
1
2019
... 灌丛在水源涵养、碳固定、土壤保持和防风固沙等方面发挥重要的作用, 尤其在西南地区具有重要的生态价值(胡雄蛟等, 2021 ).云南省共有27种灌丛群系, 分属于河谷灌丛、山地灌丛、亚高山灌丛、高山灌丛4种植被型, 其中山地灌丛中的南烛(Vaccinium bracteatum )灌丛和云南杨梅(Myrica nana )灌丛分布较广, 两者面积共达到49.04 × 103 km2 (谢宗强等, 2019 ).云南杨梅抗污染能力强, 与根瘤菌共生, 具有杰出的水土保持和肥地功能, 同时具备较强的耐火、阻火能力, 因此可作优质的绿化树种及恢复植被的先锋树种(成宇, 2019 ).云南杨梅是滇中地区山地灌丛的优势种, 研究云南杨梅灌丛主要木本植物的生态位与种间联结不仅有利于更好地认识和发挥云南杨梅灌丛的生态功能, 而且可为揭示云南杨梅灌丛特征和维持云南杨梅灌丛的生物多样性提供科学依据.目前对云南杨梅灌丛的研究甚少, 仅包括对云南杨梅灌丛碳密度(郭子豪等, 2020 )、生物量(巩合德等, 2012 )和土壤有机碳(陈路红等, 2017 )等的研究.对云南杨梅灌丛群落的生态位和种间联结的研究目前还未有报道.本研究采用了Levins生态位宽度(B L )、Shannon生态位宽度(B S )、Schoener重叠指数(Cik )、Pianka重叠指数(Oik )、方差比率(VR)、χ2 检验、Pearson相关检验和Spearman秩相关检验进行数据分析与处理, 以说明下面几个科学问题: 1)云南杨梅灌丛主要木本植物的生态位宽度和生态位重叠特征; 2)云南杨梅灌丛主要木本植物的种间联结关系; 3)云南杨梅灌丛主要木本植物的生态位重叠和种间联结之间的联系. ...
植物种间联结研究内容与方法评述
1
2016
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
植物种间联结研究内容与方法评述
1
2016
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
包头南海湖浮游植物优势种生态位及种间联结性季节分析
1
2020
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
包头南海湖浮游植物优势种生态位及种间联结性季节分析
1
2020
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
贵州茂兰喀斯特森林四药门花群落优势种群生态位特征
1
2017
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
贵州茂兰喀斯特森林四药门花群落优势种群生态位特征
1
2017
... 生态位和种间关系对于正确认识群落的结构、功能和分类有着重要的指导意义(高永恒等, 2011 ).目前生态位的研究内容主要包括以下几类, 一是对不同生活型植物的研究, 如苔藓植物(刘艳等, 2019 )、浮游植物(杨文焕等, 2020 )、木本植物(白欢欢, 2018 ); 二是对相异环境下植被的研究, 如荒漠草原(陈林等, 2019 )、高山草甸(李斌等, 2010 )、喀斯特地区植被(刘润红等, 2020 ); 三是对不同群落层片的研究, 如乔木层(肖艳梅等, 2021 )、灌木层(俞筱押等, 2017 )、草本层(王伟伟等, 2012 ); 四是对生态位与其他主题之间联系的研究, 如生态位与种间联结(霍红等, 2013 ), 生态位与生物多样性(董雪等, 2020 ).种间联结的研究内容主要涉及物种之间的利他作用、生物入侵、生态种组、不同演替阶段的种间关系、不同尺度格局的种间关系、生境异质性与种间关系等(徐满厚等, 2016 ).灌丛种间联结的研究内容在时间上分为静态时间点研究(王志泰等, 2006 )和用空间代替时间的研究(石福习等, 2012 ), 缺乏对同一群落在不同时间段的种间联结关系对比研究.在空间上分为河谷灌丛(刘庆等, 1996 )、亚高山灌丛(李军玲等, 2004 )等研究. ...
荒漠草原带盐碱地优势植物生态位与种间联结
1
2018
... 生态位宽度较大(较小)的物种与其他物种产生生态位重叠的概率也较大(较小)(陈玉凯等, 2014 ).本研究中云南杨梅出现在每个样方中, 生态位宽度最大, 与20个主要物种产生生态位重叠的概率为100%.亮毛杜鹃的生态位宽度较小, 与20个主要物种间的生态位重叠概率也较小.生态位宽度较小的物种间也出现生态位重叠值较高的情况, 如粉叶小檗和钝叶橘红悬钩子、粉叶小檗和怒江红山茶、钝叶橘红悬钩子和怒江红山茶, 这与张东梅等(2018 )的研究结论一致. ...
荒漠草原带盐碱地优势植物生态位与种间联结
1
2018
... 生态位宽度较大(较小)的物种与其他物种产生生态位重叠的概率也较大(较小)(陈玉凯等, 2014 ).本研究中云南杨梅出现在每个样方中, 生态位宽度最大, 与20个主要物种产生生态位重叠的概率为100%.亮毛杜鹃的生态位宽度较小, 与20个主要物种间的生态位重叠概率也较小.生态位宽度较小的物种间也出现生态位重叠值较高的情况, 如粉叶小檗和钝叶橘红悬钩子、粉叶小檗和怒江红山茶、钝叶橘红悬钩子和怒江红山茶, 这与张东梅等(2018 )的研究结论一致. ...
1
2018
... 采用χ 2 检验、相关系数进行种间关联分析.鉴于本研究为非连续性取样, 种间关联的显著程度采用Yates的连续校正公式(张金屯, 2018 )进行χ2 检验, 公式如下: ...
1
2018
... 采用χ 2 检验、相关系数进行种间关联分析.鉴于本研究为非连续性取样, 种间关联的显著程度采用Yates的连续校正公式(张金屯, 2018 )进行χ2 检验, 公式如下: ...
卧龙自然保护区三江珙桐群落种间关系的数量分析
1
2005
... 滇中云南杨梅灌丛主要木本植物的总体联结性表现为显著正联结, 表明群落各物种之间可以协调共存, 对资源的利用较为充分, 群落结构趋于稳定, 可能是因为绝大多数样地中云南杨梅群落受到的人为干扰程度轻微.检验显著率18.42% (Spearman秩相关检验) > 10.53% (Pearson相关检验) > 9.47% (χ 2 检验); Spearman秩相关检验中呈显著关联关系的种对数比Pearson相关检验更多, 说明Spearman秩相关检验有更高的灵敏度, 可以更为准确地反映种间关系.这与朱利君等(2005 )的研究结果一致.以Spearman秩相关检验结果为依据, 即种对间正负联结比小于1, 判断云南杨梅群落可能处于演替的前期或中期阶段.综合3种检验结果, 不显著种对数均大于显著种对数, 说明群落物种间的联结性较弱, 呈独立分布格局.粉叶小檗-钝叶橘红悬钩子、粉叶小檗-怒江红山茶、钝叶橘红悬钩子-怒江红山茶和元江锥-高山锥均呈现极显著正联结关系, 说明这些物种之间相互依存或资源需求上互补. ...
卧龙自然保护区三江珙桐群落种间关系的数量分析
1
2005
... 滇中云南杨梅灌丛主要木本植物的总体联结性表现为显著正联结, 表明群落各物种之间可以协调共存, 对资源的利用较为充分, 群落结构趋于稳定, 可能是因为绝大多数样地中云南杨梅群落受到的人为干扰程度轻微.检验显著率18.42% (Spearman秩相关检验) > 10.53% (Pearson相关检验) > 9.47% (χ 2 检验); Spearman秩相关检验中呈显著关联关系的种对数比Pearson相关检验更多, 说明Spearman秩相关检验有更高的灵敏度, 可以更为准确地反映种间关系.这与朱利君等(2005 )的研究结果一致.以Spearman秩相关检验结果为依据, 即种对间正负联结比小于1, 判断云南杨梅群落可能处于演替的前期或中期阶段.综合3种检验结果, 不显著种对数均大于显著种对数, 说明群落物种间的联结性较弱, 呈独立分布格局.粉叶小檗-钝叶橘红悬钩子、粉叶小檗-怒江红山茶、钝叶橘红悬钩子-怒江红山茶和元江锥-高山锥均呈现极显著正联结关系, 说明这些物种之间相互依存或资源需求上互补. ...






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}