



叶片是植物进行光合作用的主要器官, 负责陆地生态系统的碳同化。叶片大小不仅反映了植物对光的拦截效率和碳获取能力(Parkhurst & Loucks, 1972; Givnish & Vermeij, 1976), 直接影响植物对环境的适应能力, 而且反映了植物碳同化和水分利用效率之间的权衡, 对不同气候条件下的叶片温度调节至关重要(Michaletz et al., 2014; Fauset et al., 2018; Li et al., 2020), 是植物生态策略中的一个关键性状。在全球范围内, 叶片大小的变化幅度达到了6个数量级(Milla & Reich, 2007), 且受诸多因素的影响。例如, 叶片大小随海拔的升高而降低, 随着年平均气温及年降水量的降低而减小(McDonald et al., 2003; 杨冬梅等, 2012); 在湿润或者阴蔽的环境下, 大叶由于具有更强的资源获取能力, 使光合收益最大化而占有优势(Givnish, 1979; 杨冬梅等, 2012); 而在高温、干燥和高光环境下, 小叶因具有较高的边界层导度, 可通过对流快速进行热交换降温而较有优势(Ackerly et al., 2002; Bragg & Westoby, 2002; McDonald et al., 2003)。
叶脉是叶片中主要的物质传输结构, 具有供应水分、传输养分和糖以及为叶片提供生物机械支撑的功能(Tsukaya, 2003)。叶脉网络结构决定了叶片内部水分运输系统的结构, 为整个叶片提供水分, 通过影响叶片的水分供应状况直接影响叶片的碳收支。在其他条件相同的情况下, 更高的叶脉密度意味着单位叶面积上具有更多平行的木质部水流途径, 可以缩短水分在叶肉中的运输距离(Niklas, 1999), 提高叶片的水分导度, 进而使叶片具有更高的气孔导度和光合速率(Sack & Frole, 2006)。理论上, 构建一个木质部导管直接插入到叶片蒸腾位点的叶脉网络可以达到一个最大的水分供应能力。但叶脉木质部的主要结构组分木质素是一种碳含量较高的物质(每克纤维素含6.5 mmol葡萄糖, 每克木质素含11.8 mmol葡萄糖, Lambers & Poorter, 2004), 要构建这样一个叶脉网络需要有很高的碳投资, 其构建和替换成本将总会高于光合作用所带来的收益(Brodribb & Feild, 2000)。很明显, 这样奢侈的木质部投资在植物中从来不会出现。
与叶片大小类似, 叶脉结构的变化也与众多环境因素密切相关。在同一植株个体上, 叶脉密度随着叶片所处冠层高度的增加而增加, 阴生叶的叶脉密度比阳生叶更低; 在土壤水分有效性和空气湿度较低、光照较强、强风以及养分缺乏等生境中, 叶脉密度往往较高(Roth-Nebelsick et al., 2001)。另外, 叶片越大, 对中脉的生物量投资越多(Niinemets et al., 2007), 单位叶面积的主脉体积也随叶片大小的增加而显著增加(Sack et al., 2012, 2013)。上述叶片大小以及叶脉结构等与环境因素的相关性表明, 叶片大小的地理变异可能与叶脉结构变化密切相关, 在植物长期进化过程中两者有可能是协同变化的。
然而, 到目前为止, 关于叶脉结构特征与叶片大小之间关系的研究仍相对较少, 且结论不一致。在一些物种数较少(10种以下)的研究中, 普遍发现主脉密度与叶片大小负相关, 细脉密度与叶片大小不相关(Sack et al., 2008; Dunbar-Co et al., 2009; Scoffoni et al., 2011)。而Price等(2012)采用分辨率较低的图像自动分析技术对史密森研究所自然历史博物馆中收集的339个物种的叶脉进行扫描, 测定整个叶片的叶脉网络结构, 发现叶脉密度与叶片大小不相关。但是该研究只测定了叶脉系统的一部分, 并且没有区分叶脉等级(Price et al., 2012)。随后, Sack等(2012)采用一种新的高清叶脉系统测定方法并结合文献中公布的相关数据, 对分布于全球的485个物种的不同等级叶脉特征参数与叶片大小之间的关系进行分析, 发现主脉直径与叶片大小成正比, 而主脉密度与叶片大小成反比, 细脉密度或直径都与叶片大小之间没有明显的相关关系。由于细脉通常占叶脉总长度的绝大部分(平均为84%), 导致总叶脉密度也与叶片大小无关(Sack et al., 2012)。Sack等(2012)虽然证明了上述关系在全球范围内普遍存在, 但没有进一步深入分析这些关系存在的原因, 只是将主脉密度和细脉密度与叶片大小之间的不同关系归因于主、细脉分别形成于叶片发育的不同阶段。但是, 叶脉作为一种碳成本较高的结构, 植物选择构建什么样的叶脉网络, 必然要受到叶脉构建成本的影响。虽然主脉的长度仅占叶脉总长度的一小部分(<20%), 但由于其直径要比细脉大得多, 主脉体积占单位叶面积上叶脉总体积的绝大部分(平均值±标准误, 82% ± 1.3%), 导致其质量在叶脉系统总质量中的占比也显著高于细脉(Sack et al., 2013)。关于叶脉构建成本与叶片大小及叶脉密度之间的变化关系, 目前还鲜有人报道, 叶片大小与叶脉结构之间的关系是否也会受到单位叶脉长度的构建成本(如叶脉细胞壁干质量)的调控目前还缺乏强有力的实验证据。
因此, 本研究在浙江省天童国家森林公园选取38种生长环境相对一致的阔叶木本植物(常绿26种, 落叶12种)为研究对象, 以叶片面积、叶片干质量、叶片周长表征叶片大小; 以主脉密度(单位叶面积上1°脉和2°脉的总长度)和细脉密度(单位叶面积上3°及以上叶脉的总长度)、总叶脉密度(单位叶面积上所有叶脉的总长度)表征叶脉结构特征; 同时测定了各级叶脉木质部的细胞壁干质量, 并以相应脉级的单位叶脉长度的细胞壁干质量作为叶脉的构建成本(即叶脉细胞壁干质量/叶脉长度)。采用标准化主轴估计(SMA)方法在物种水平上分析叶片大小特征参数、叶脉密度以及单位叶脉长度的细胞壁干质量之间的关系, 拟从叶片自身的内部结构和资源分配策略的角度深入探讨叶片大小与叶脉结构之间的关系及其原因, 为理解叶片大小的适应进化及植物对环境的生态适应策略提供参考。
1 材料和方法
1.1 研究区概况
本研究样地位于浙江宁波天童国家森林公园(29.80° N, 121.78° E)。该地属浙东低山丘陵, 土质为红黄壤, 成土母质为中生代的沉积岩、部分酸性火成岩以及石英砂岩和花岗岩的残积风化物。土壤厚薄不一, 一般在l m以下。气候属于亚热带季风气候, 雨量充沛, 年平均气温16.2 ℃, 年降水量1 389.7 mm, 年无霜期230-240天。主要植被类型为常绿阔叶林, 其中木荷(Schima superba)林分布面积最多(赵琦琳等, 2020)。园区内自然植被保存较好, 共有种子植物149科1 064种。该区域反映了亚热带东部低海拔森林生态系统的典型特征, 也是我国东部植被的典型类型, 更是代表了浙江东部丘陵地区地带性植被类型。
1.2 植物采样和性状测定
本研究于2019年7-8月, 当枝条生长和叶片展叶完成时, 选取区域内相对丰富度较高、优势度较大, 且叶片大小差异较大的38种阔叶木本植物为研究对象(常绿26种, 落叶12种), 属于17科27属, 物种间的亲缘关系及其进化过程和模式见系统发育树(图1; 附录I)。从系统发育树可以看出, 本研究中包括进化枝长较长的原始基部被子植物樟科和木兰科的多个物种。
图1

图1
天童国家森林公园38种阔叶木本植物系统发育树。
Fig. 1
Phylogeny tree of 38 broad-leaved woody species in Tiantong National Forest Park.
样品采集时, 每物种选取5-6株生长良好、健康成熟、生境类似的个体, 用高枝剪(部分较高植株借助伸缩梯或攀爬)从每个个体树冠2/3以上的部位选取3个阳生当年生末端小枝, 做好标志后迅速放入装有湿润纸球的大黑色塑料袋中带回实验室备用。主要性状测定如下:
(1)叶片大小相关的形态指标的测定。在每个小枝从顶端往下第3-4叶位处选取1片发育成熟、完全展叶且没有明显叶片面积损伤的叶片(即每物种共约15个叶片), 轻轻擦拭表面污渍后用扫描仪对叶片进行扫描, 设置分辨率为300 dpi, 用ImageJ 18.0测定叶片面积和周长。然后将叶片装入信封中, 70 ℃烘48 h至恒质量, 称干质量。
细脉的测定参考秦茜等(2017)的方法, 将已扫描过叶片面积的叶片置于5% NaOH溶液中在恒温水浴锅中煮沸3-20 min, 直至叶肉被明显清除, 叶脉逐渐清晰甚至透明。用去离子水多次清洗叶片, 避开1°脉与2°脉, 从叶片基部、中部、尖部对称位置各取1 cm2样品(即一个叶片包含6个样品)。样品于5% NaClO溶液中漂白, 后经3%甲苯胺蓝酒精溶液染色, 滴加封片剂于玻片, 使用压片法制片保存, 待野外实验结束后带回实验室进行拍照并测定分析(秦茜等, 2017)。样品用显微镜放大40倍后拍照, 每个样本尽量避开1°、2°脉拍摄3个不重复图片(即一个个体共3片叶片× 6个样品× 3个视野= 54张图片)。根据Bühler等(2015)的研究方法, 运用MeVisLab软件加载phenoVein工具并进行手工校正, 最终得到叶脉基本性状(Bühler et al., 2015)。
(3)叶脉木质部细胞壁干质量的测定。叶脉木质部细胞壁干质量的测定参考Wang等(2017)的测定方法。叶片采集与上述叶片大小采样方法相同, 用扫描仪扫描后, 放入1 mol·L-1 NaOH溶液的烧杯中在水浴锅中煮沸20-30 min,然后置于清水中, 用小刷子轻轻将叶肉去除。随后, 样品用去离子水清洗干净, 吸水纸吸干表面水分。参考测定叶脉性状时的叶脉分级方法, 用剪刀和镊子将叶片1°脉、2°脉和叶柄分开, 用精度为0.000 1 g的天平称取组织鲜质量后放入75%乙醇的培养皿中备用。将组织样品转移到离心管中, 加入少量75%乙醇(漫过样品表面), 放置小钢珠, 于植物组织研磨仪中70 Hz研磨120 s至匀浆。研磨完成后, 将离心管内部和小钢珠表面残留匀浆用75%的乙醇逐次转移至50 mL的离心管中, 冰浴静置20 min后使用离心机1 000 g下离心10 min, 小心弃去上清液, 并依次用1:7 (组织鲜质量/体积)预冷丙酮、甲醇-氯仿混合物(体积比1:1)和甲醇对沉淀进行洗涤, 弃去每次洗涤的上清液。最后一次去除上清液后, 将沉淀置于70 ℃烘箱内烘干至恒质量即得叶脉木质部细胞壁干质量(Wang et al., 2017)。
1.3 数据分析
对于叶片大小和叶脉相关性状依次在个体水平、物种水平上计算其平均值, 然后在分析前进行对数转换, 使之符合正态分布用于种间比较。各对性状间的关系采用方程y = bxa来描述, 线性转换为lg(y) = lg(b) + alg(x), 其中x和y分别表示两个性状变量, b表示性状关系的截距, a表示斜率, 即异速生长指数(a = 1时为等速生长, a > 1或a < 1时为异速生长)(Harvey & Pagel, 1991)。采用SMA方法估计异速生长方程的参数(Warton et al., 2006), 由(S)MATR Version 2.0软件计算完成。按照Pitman (1939)记载的方法计算回归斜率的置信区间。
采用系统发育独立比较(PIC)方法分析不同功能性状间的相互关系是否随进化分歧而改变, 即运用PIC方法去除系统发育对功能性状关系的影响。系统发育树是基于Phylomatic 3 (
2 结果
2.1 叶片面积与叶片干质量及叶片周长的关系
在本研究中, 物种水平的平均单叶面积变化范围7.69-196.01 cm2; 单叶干质量0.08-1.64 g; 叶片周长14.77-57.08 cm。叶片面积和干质量的种间差异均达到2个数量级, 表明叶片大小在所采集物种间具有较大的变化范围。本研究发现叶片面积与干质量及周长均呈极显著的正相关关系, 其与叶片干质量关系的斜率为1.102 (95%置信区间: 0.953, 1.274), 呈现与1无显著差异的等速生长关系(图2A; 表1); 而与叶片周长关系的斜率为2.029 (95%置信区间: 1.821, 2.260), 呈现与2无显著差异的异速生长关系(图2B; 表1)。系统发育独立比较分析结果显示, 在剔除系统发育的影响后, 叶片面积与干质量及周长之间的显著正相关关系仍然存在(表2), 表明叶片大小性状之间在物种水平上具有明显的趋同进化现象。
图2
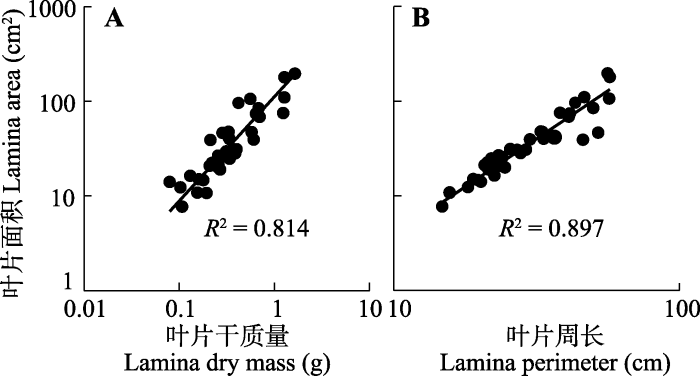
图2
天童国家森林公园38个阔叶木本植物叶片面积与叶片干质量(A)、叶片周长(B)的关系。
Fig. 2
Relationships between lamina area and lamina dry mass (A), lamina area and lamina perimeter (B) of 38 broad-leaved woody species in Tiantong National Forest Park.
表1 天童国家森林公园38个阔叶木本植物叶片大小与叶脉结构的标准化主轴估计回归关系
Table 1
| 指标 (y轴-x轴) Index (y axis-x axis) | 决定系数 Coefficient of determination (R2) | 斜率(95%置信区间) Slope (95% confidence interval) | p |
|---|---|---|---|
| 叶片面积-叶片干质量 Lamina area-lamina dry mass | 0.081 | 1.102 (0.953, 1.274) | <0.001 |
| 叶片面积-叶片周长 Lamina area-lamina perimeter | 0.897 | 2.029 (1.821, 2.260) | <0.001 |
| 主脉密度-叶片面积 Major vein density-lamina area | 0.893 | -0.503 (-0.562, -0.451) | <0.001 |
| 主脉密度-叶片周长 Major vein density-lamina perimeter | 0.705 | -1.021 (-1.226, -0.851) | <0.001 |
| 主脉密度-叶片干质量 Major vein density-lamina dry mass | 0.712 | -0.555 (-0.664, -0.463) | <0.001 |
| 主脉密度-主脉单位长度的细胞壁干质量 Major vein density-cell wall dry mass per unit length of major vein | 0.615 | -0.699 (-0.861, -0.567) | <0.001 |
| 主脉单位长度的细胞壁干质量-叶片面积 Cell wall dry mass per unit length of major vein-lamina area | 0.554 | 0.721 (0.576, 0.901) | <0.001 |
| 主脉单位长度的细胞壁干质量-叶片周长 Cell wall dry mass per unit length of major vein-lamina perimeter | 0.445 | 1.462 (1.142, 1.872) | <0.001 |
| 主脉单位长度的细胞壁干质量-叶片干质量 Cell wall dry mass per unit length of major vein-lamina dry mass | 0.412 | 0.794 (0.614, 1.026) | <0.001 |
表2 天童国家森林公园38种阔叶木本植物叶片大小与叶脉性状间的系统发育独立性比较及最小二乘法回归分析
Table 2
| 指标(y轴-x轴) Index (y axis-x axis) | 斜率 Slope | 决定系数 Coefficient of determination (R2) |
|---|---|---|
| 叶片面积-叶片干质量 Lamina area-lamina dry mass | 1.164 | 0.952 |
| 叶片面积-叶片周长 Lamina area-lamina perimeter | 2.046 | 0.989 |
| 主脉密度-叶片面积 Major vein density-lamina area | -0.440 | 0.985 |
| 主脉密度-叶片周长 Major vein density-lamina perimeter | -0.898 | 0.971 |
| 主脉密度-叶片干质量 Major vein density-lamina dry mass | -0.508 | 0.924 |
| 主脉密度-主脉单位长度的细胞壁干质量 Major vein density-Cell wall dry mass per unit length of major vein | -0.817 | 0.923 |
| 主脉单位长度的细胞壁干质量-叶片面积 Cell wall dry mass per unit length of major vein-lamina area | 0.493 | 0.897 |
| 主脉单位长度的细胞壁干质量-叶片周长 Cell wall dry mass per unit length of major vein-lamina perimeter | 1.010 | 0.889 |
| 主脉单位长度的细胞壁干质量-叶片干质量 Cell wall dry mass per unit length of major vein-lamina dry mass | 0.562 | 0.819 |
回归关系均达极显著水平(p < 0.001)。
All scaling relationships were significant (p < 0.001).
2.2 叶片大小与叶脉密度的关系
在本研究的38个物种中, 各级叶脉的平均值为: 主脉密度(2.35 ± 0.14) cm·cm-2, 细脉密度(76.79 ± 3.39) mm·mm-2, 总叶脉密度(79.68 ± 3.59) cm·cm-2。各级叶脉(包括主脉、细脉和总叶脉)长度与叶片大小各特征参数(叶片面积、干质量以及周长)之间都存在极显著的正相关关系(p < 0.01), 表明叶片越大, 叶片内各级叶脉的长度越长。各级叶脉长度以单位叶面积进行标准化后(即叶脉密度), 则与叶片大小各性状之间的关系略有不同。主脉密度与3个叶片大小性状之间均呈极显著的负相关关系(p < 0.001)(图3; 表1), 其中与叶片面积、干质量的斜率分别为-0.503 (95%置信区间: -0.562, -0.451)和-0.555 (95%置信区间: -0.664, -0.463), 均呈大于-1的异速生长关系, 但与-0.5无显著差异(图3A、3B; 表1); 与叶片周长的关系斜率为-1.021 (95%置信区间: -1.226, -0.851), 呈现与-1无显著差异的负等速生长关系(图3C; 表1)。
图3
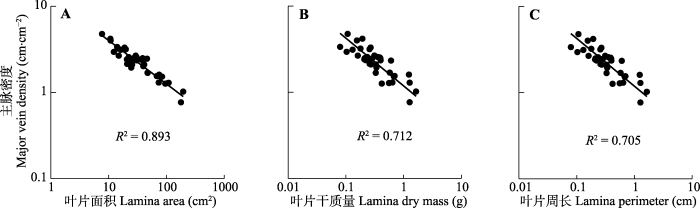
图3
天童国家森林公园38种阔叶木本植物叶片主脉密度与叶片面积(A)、叶片干质量(B)和叶片周长(C)的关系。
Fig. 3
Relationships between the major vein density and lamina area (A), lamina dry mass (B), and lamina perimeter (C) of 38 broad-leaved woody species in Tiantong National Forest Park.
进一步的PIC分析结果显示, 主脉密度与叶片大小之间的显著负相关关系在排除系统发育影响后仍然存在(p < 0.001, 表2), 即主脉密度与叶片大小性状间存在趋异进化。上述关系表明, 不管叶片大小用哪种参数表征, 主脉密度与叶片大小之间均呈极显著的负相关关系, 即小的叶片具有较高的主脉密度; 而细脉密度与叶片面积、周长或干质量的变化无显著相关性。
2.3 叶片大小与单位叶脉长度的细胞壁干质量的关系
在本研究的38个物种中, 各级叶脉的单位长度细胞壁干质量的平均值为: 总叶脉的单位长度细胞壁干质量(0.016 2 ± 0.001 5) mg·cm-1, 主脉的单位长度细胞壁干质量(0.178 8 ± 0.019 5) mg·cm-1, 细脉的单位长度细胞壁干质量(0.011 5 ± 0.001 2) mg·cm-1。SMA分析结果显示, 单位主脉长度的细胞壁干质量与3个叶片大小性状均呈显著正相关关系(图4; 表1), 其中与叶片面积的关系斜率为0.721 (95%置信区间:0.576, 0.901)(图4A; 表1), 呈现显著小于1的异速生长关系, 表明在叶片伸展过程中对主脉细胞壁的生物量投资增加的速率跟不上叶片面积增大的速率; 但与叶片干质量的关系斜率为与1无显著差异的等速生长关系(斜率=0.794, 95%置信区间: 0.614, 1.026)(图4B; 表1), 表明单位主脉长度的细胞壁干质量投资的增加可以获得等速的叶片干质量收益; 与叶片周长则是斜率与1.5无显著差异的异速生长关系(斜率=1.462, 95%置信区间: 1.142, 1.872)(图4C; 表1), 表明对主脉细胞壁的生物量投资将随着叶片周长的增加而急剧增加。而单位细脉和总脉长度的细胞壁干质量与所有叶片大小性状的相关性均不显著(p > 0.05)。
图4
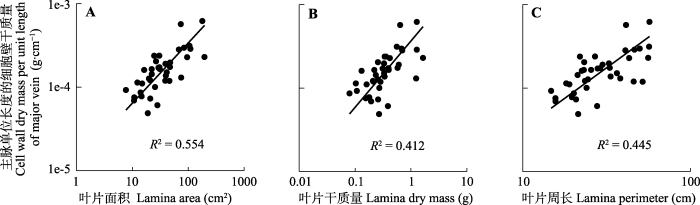
图4
天童国家森林公园38种阔叶木本植物叶片单位主脉长度的细胞壁干质量与叶片面积(A)、叶片干质量(B)和叶片周长(C)的关系。
Fig. 4
Relationships between the cell wall dry mass per unit length of major vein and lamina area (A), lamina dry mass (B), and lamina perimeter (C) of 38 broad-leaved woody species in Tiantong National Forest Park.
PIC分析结果与SMA结果一致, 在排除系统发育影响后, 单位主脉长度的细胞壁干质量与3个叶片大小性状之间的正相关关系仍然显著(表2), 表明对主脉细胞壁的生物量投资与叶片大小存在趋同进化。
2.4 叶脉密度与单位叶脉长度的细胞壁干质量的关系
图5
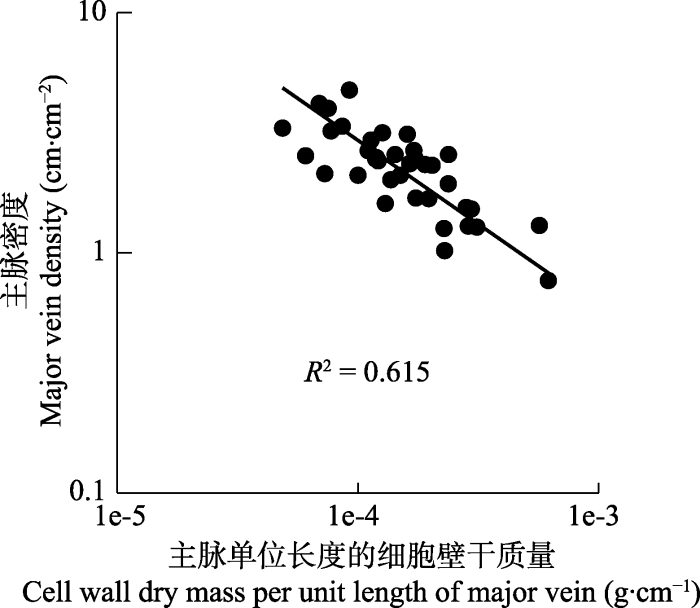
图5
天童国家森林公园38种阔叶木本植物主脉密度与单位主脉长度的细胞壁干质量的关系。
Fig. 5
Relationship between the major vein density and cell wall dry mass per unit length of major vein of 38 broad-leaved woody species in Tiantong National Forest Park.
3 讨论
本研究在对浙江天童38种阔叶木本植物的叶片大小与叶脉结构及单位叶脉长度的细胞壁干质量之间关系的研究中, 物种水平上的叶片大小平均值差异达到2个数量级, 能够代表较大的叶大小变化范围。叶片大小无论是用叶片面积、干质量还是周长来表征, 主脉密度均随着叶片大小的增加而逐渐降低(图2; 表1)。与此同时, 单位主脉长度的细胞壁干质量随着叶片大小的增加而增加(图4; 表1), 主脉密度则随着单位主脉长度的细胞壁干质量的增加而下降(图5; 表1)。这些结果表明, 主脉密度与叶片大小之间的负相关关系是受单位主脉长度的细胞壁构建成本调控的结果, 即随着叶片增大, 由于单位主脉长度的细胞壁干质量的增加导致主脉密度急剧下降。而细脉和总叶脉密度与叶片大小的相关性不显著, 也是由于单位细脉和总叶脉长度的细胞壁干质量与叶片大小及各自的叶脉密度之间的相关性均不显著所导致的。并且, PIC分析结果显示, 在排除系统发育的影响后, 上述关系依然成立(表2)。由此我们认为, 叶片大小与主、细脉密度之间不同的变化关系, 除了叶片发育对主脉和细脉的调控效应不同的原因外, 也与单位叶脉长度的细胞壁干质量密切相关。这一研究有助于我们更好地理解叶片大小的适应进化及植物对环境的生态适应策略。
3.1 叶片大小与叶脉结构的变化关系
在叶片大小各表征性状与叶脉结构之间关系的分析中, 叶片大小与各级叶脉长度(即实际测得的叶脉长度值)均存在极显著的正相关关系(p < 0.01), 表明叶片越大, 需要的各级叶脉长度越长。为了使不同大小叶片的叶脉长度具有可比性, 将叶脉长度以单位叶面积进行标准化, 得到各级叶脉的密度, 并按Bühler等(2015)的方法将1°和2°脉统称为主脉, 将3°及以上叶脉统称为细脉, 再分析它们与叶片大小特征性状之间的关系。结果显示, 主脉密度与叶片面积、周长和干质量都极显著负相关, 与其他学者在不同研究尺度上发现的叶片大小与主脉密度都显著负相关的研究结果(Sack et al., 2008; Dunbar- Co et al., 2009; Scoffoni et al., 2011)一致。值得一提的是, 本研究中叶片面积和干质量与主脉密度之间的关系斜率分别为-0.503和-0.555 (图3A、3B; 表1), 与Sack等(2012)在全球尺度上分析的1°和2°脉密度与叶片大小的关系斜率(分别为-0.40和-0.53, 95%置信区间: -0.57, -0.38)基本一致, 都与-0.5无显著差异。细脉密度与叶片面积、周长和干质量均无显著相关关系, 与Sack等(2012)研究中细脉密度与叶片大小不相关的结果一致。
此外, 本研究中主脉密度与3个叶片大小性状之间关系的指数不完全相同, 其主要原因是3个叶片大小性状具有不同的几何维度, 使得三者间具有不同的函数关系。叶片面积(LA)与叶片周长(Perimeter)之间关系为斜率与2无显著差异的异速生长关系(LA∝Perimeter2), 主脉密度(VD)与叶片面积之间关系为斜率与-0.5无显著差异的异速生长关系(VD∝LA-0.5), 导致主脉密度与叶片周长呈指数为-1的负等速的生长关系(VD∝Perimeter-1); 而叶片面积与叶片干质量间存在等速的生长关系(图2A; 表1)则导致了主脉密度和叶片干质量的关系斜率与主脉密度和叶片面积的斜率相同, 并且都是与-0.5无显著差异的异速生长关系。
3.2 叶片大小与单位叶脉长度的细胞壁干质量的变化关系
尽管本研究中各物种不同等级叶脉的细胞壁总干质量都随着叶片大小的增加而增加, 即对叶脉细胞壁的生物量分配随着叶片的增大而增加, 但单位叶脉长度的细胞壁干质量在叶片长大过程中并不是固定不变的, 且主、细脉表现出不同的变化规律。单位主脉长度的细胞壁干质量与各叶片大小性状之间均存在极显著正相关关系(图4; 表1), 而单位细脉长度的细胞壁干质量与所有叶片大小性状之间的相关性都不显著。由于细脉通常占叶脉总长度的绝大部分, 导致单位总叶脉长度的细胞壁干质量总体上也与叶片大小性状无显著相关性。主、细脉之间的这种差异是由叶片发育过程中主脉和细脉的直径生长变化不同所导致的。1°脉直径随叶片大小的增加呈几何级的增大(斜率=0.45, 95%置信区间: 0.43, 0.48), 在整个展叶期的大部分时间内始终保持增粗生长; 2°和3°脉直径随叶片大小增加而增大的速率则相对保守, 其斜率分别为0.37 (95%置信区间: 0.34, 0.39)和0.22 (95%置信区间: 0.20, 0.25); 而3°以上细脉在形成后其直径很快达到最大值, 所以与叶片大小不相关(Sack et al., 2012)。主脉直径的增粗导致单位叶面积上的主脉体积增加, 必然导致对主脉的细胞壁生物量投资增加(Niinemets et al., 2007; Niklas et al., 2007)。上述结果表明, 随着叶片增大, 延长主脉所需要付出的代价越来越大, 而延长细脉的代价则相对稳定。
本研究结果显示单位主脉长度的细胞壁干质量与叶片面积呈显著小于1的异速生长关系(图4A; 表1), 表明在展叶过程中对主脉的生物量分配增加的速率跟不上叶片面积增大的速率, 也就是说对主脉的生物量投资所能获得的叶片面积随着叶片大小的增加而增加, 即收益递增。而单位主脉长度的细胞壁干质量与叶片干质量间的关系也呈斜率与1无显著差异的等速生长关系, 即叶片的碳收益随着对主脉生物量投资的增加而等速增加。因此, 如果单纯从单位主脉长度的细胞壁干质量与叶片大小之间的关系来看, 增加对主脉的生物量投资可以获得等额比例甚至更高的叶片收益回报, 对植株生长更有利。然而, 事实上植物对叶片的主脉细胞壁投资不可能无限制的增大, 这主要是由于叶脉密度的相关制约, 详见下文讨论。
3.3 叶片大小与主脉密度显著负相关的主要原因及生态学意义
叶片大小与主脉密度显著负相关, 而与细脉及总脉密度之间关系不显著, 以前的相关研究认为其中的一个主要原因是由于主、细脉分别形成于叶片发育的不同阶段所导致的。在叶片发育过程中, 最先形成的是1°和2°脉, 然后是3°脉。这些低级主脉都是在叶片发育初期的“缓慢”展叶阶段(即细胞增殖阶段)开始形成, 然后随着叶片的伸展逐渐延长, 到原形成层形成时随着其发育信号的衰弱或消失达到最大值(Sack et al., 2012)。在叶片发育后期的“快速”展叶阶段(即细胞增大阶段), 这些叶脉随着叶片的增大而被逐渐推开, 叶脉之间的间距拉大, 导致单位叶面积上的叶脉长度(即叶脉密度)被“稀释”, 所以叶片大小与主脉密度显著负相关。与主脉不同, 3°以上的细脉则主要形成于叶片发育后期的“快速”展叶阶段, 伴随着叶片的“快速”伸展不断形成新的细脉, 细脉之间的间距比较稳定, 所以细脉密度与叶大小无关(Sack et al., 2012)。
除了上述原因, 我们在本研究中的实验结果表明叶脉的细胞壁构建成本(单位长度的细胞壁干质量)也是导致叶片大小与主脉密度显著负相关的一个重要原因。叶脉是一个悬臂结构, 叶内最大的机械应力位于叶片的纵轴上, 叶片的机械稳定性主要取决于低级主脉(Kull & Herbig, 1994; Roth-Nebelsick et al., 2001)。随着叶片的不断增大, 需要有更大的主脉为其提供更强有力的机械支撑, 再加上提高主脉的粗度进而提高其水分运输效率是提高整个叶片水分运输效率的前提(McKown et al., 2010)。因此, 随着叶片增大, 主脉直径需要不断增粗, 在对主脉的细胞壁生物量投资一定的情况下, 植物将首先选择增加主脉的粗度而非延长其长度, 导致单位主脉长度的体积增大, 进而导致单位主脉长度的细胞壁干质量随叶片大小增加而增加(图4)。更多的主脉生物量被用于粗度生长, 必然导致叶脉的长度生长受限, 所以主脉密度又随单位主脉长度的细胞壁干质量增加而显著下降(图5; 表1), 两者是相互权衡的关系。所以, 叶片大小、单位主脉长度的细胞壁干质量和主脉密度三者之间是密切相关、相互影响的, 单位主脉长度的细胞壁干质量是导致叶片大小与主脉密度显著负相关的直接原因。
单位主脉长度的细胞壁干质量和主脉密度相互权衡的关系对叶大小的优化具有重要的意义, 因为较粗的主脉内通常具有更长更粗的导管(Choat et al., 2005), 水分运输效率较高, 主脉密度的减小会导致水分在主脉内的运输距离缩短, 而在主脉外的运输距离增长, 直接影响到整个叶片的水分传输效率, 进而影响整个植株的光合收益和生产力。因此, 尽管增加对主脉的生物量投资可以获得的叶片增大收益是随叶大小的增加而递增的(图4A、4B), 貌似单位主脉长度的细胞壁投资越大越好, 但由于主脉的增粗生长会限制其长度生长, 导致主脉密度减小, 所以为了保持高效的水分运输效率, 叶片不可能通过无限的增加单位主脉长度的细胞壁投资来增加叶片大小, 而是处于一个最优的状态, 与主脉密度相权衡并获得合理的叶片大小。
然而, 与主脉不同, 细脉的直径在叶片的整个生长发育过程中没有显著的变化(Sack et al., 2012), 导致单位细脉长度的细胞壁干质量与叶片大小和细脉密度之间的相关性都不显著。细脉长度的增加不会导致其单位长度的细胞壁构建成本增加, 但却可以缩短水分在叶肉组织中的扩散距离, 极大地提高叶片的水分运输效率, 对叶片的光合生理和生长极为有利(Sack & Frole, 2006; Brodribb et al., 2007; Boyce et al., 2009; McKown et al., 2010)。换言之, 对细脉的生物量投资可以获得更好的收益回报, 所以植物会选择在展叶后期仍不断形成新的细脉, 从而有利于不同大小叶片的物种适应不同的环境。
叶片大小与各级叶脉密度之间不同的变化关系, 不管是叶片发育调控的结果还是受单位叶脉长度的细胞壁干质量介导调控的结果, 都为解释在干燥和更开阔的生境中小叶占优势这一最著名的全球植物生物地理分布模式(Givinish, 1987; Peppe et al., 2011)提供了直接的、水力学方面的证据, 具有重要的生态意义。在最优环境下叶片的最大水分运输效率与叶脉密度以及水分在叶肉组织中运输路径长度(即从叶脉末端到蒸腾位点之间的传输距离)密切相关, 而后两者本身也是彼此相关的(Sack & Frole, 2006; Brodribb et al., 2007; Noblin et al., 2008)。根据Brodribb等(2007)的研究结果显示, 在特定环境条件下, 叶肉组织的水分运输阻力在不同物种间的变化是比较保守的, 叶片水分导度的变化主要取决于叶脉密度。网状叶脉结构的理论模型研究显示, 只有在低级主脉的水分传输效率较高时, 较高的叶脉密度对叶片而言才是比较经济有利的(McKown et al., 2010), 所以主脉通常较粗并具有更长更宽的导管(Choat et al., 2005), 这使主脉在获得较高的水分传输效率的同时对干旱或低温胁迫诱导的气穴化栓塞也更脆弱。而叶片的水分导度(Kleaf)、气孔导度和光合作用都对大的主脉的损伤非常敏感(Nardini et al., 2001; Hüve et al., 2002; Nardini & Salleo, 2003; Sack et al., 2003; Salleo et al., 2003)。小叶中更高的主脉密度为叶内的水分运输提供了冗余的“高速公路”, 当部分主脉因干旱或低温胁迫而发生栓塞导致水分传输失败时, 在栓塞导管周围可以有更多的水流通道继续维持水分运输功能(Sack et al., 2008), 有助于提高叶片和植株的抗旱能力, 更好地保护水分运输系统免受叶脉损伤所造成的伤害(Sack et al., 2008; Scoffoni et al., 2011)。所以, 在高温、干燥和高光环境下, 叶片通常更小(Ackerly et al., 2002; Bragg & Westoby, 2002; McDonald et al., 2003), 并且叶大小随着年平均气温及年降水量的降低而减小(McDonald et al., 2003)。
总之, 本研究通过对亚热带常绿阔叶林相同群落下38种阔叶木本植物的叶片大小、不同等级叶脉的密度以及单位叶脉长度的细胞壁干质量等性状之间的关系进行研究, 验证了有关学者在全球尺度上发现的叶片大小与主脉密度显著负相关, 与细脉和总叶脉密度相关性不显著的变化规律在群落内部依然存在; 发现了单位主脉长度的细胞壁干质量与主脉密度是相互权衡的关系, 而单位细脉或总叶脉长度的细胞壁干质量与相应的细脉密度或总叶脉密度之间关系不显著, 由此证明了单位叶脉长度的细胞壁干质量是导致叶片大小与主、细脉密度之间不同变化关系的直接因素。本研究从叶片内部的主要输水系统生物量分配的角度探明了叶片大小与叶脉结构协同变化的主要原因, 对于我们理解全球范围内叶片大小变化的生物地理分布模式以及植物对环境的适应策略具有重要的意义。
责任编委: 何念鹏 编辑: 赵 航
附录
附录I 天童国家森林公园38个阔叶木本植物叶片大小与叶脉结构性状数据
Supplement I Lamina size and vein architecture traits of 38 broad-leaved woody species in Tiantong National Forest Park
致谢
在浙江天童国家森林公园野外工作期间, 得到了华东师范大学浙江天童森林生态系统国家野外科学观测研究站工作人员的大力支持和帮助, 浙江师范大学化学与生命科学学院本科生朱思丽、宋志雯、郑宁静、唐翔翔、徐佳禾几位同学也参与了部分实验研究工作, 在此一并表示感谢!
参考文献
Leaf size, specific leaf area and microhabitat distribution of chaparral woody plants: contrasting patterns in species level and community level analyses
DOI:10.1007/s004420100805
PMID:28547053
[本文引用: 2]

We examined variation in leaf size and specific leaf area (SLA) in relation to the distribution of 22 chaparral shrub species on small-scale gradients of aspect and elevation. Potential incident solar radiation (insolation) was estimated from a geographic information system to quantify microclimate affinities of these species across north- and south-facing slopes. At the community level, leaf size and SLA both declined with increasing insolation, based on average trait values for the species found in plots along the gradient. However, leaf size and SLA were not significantly correlated across species, suggesting that these two traits are decoupled and associated with different aspects of performance along this environmental gradient. For individual species, SLA was negatively correlated with species distributions along the insolation gradient, and was significantly lower in evergreen versus deciduous species. Leaf size exhibited a negative but non-significant trend in relation to insolation distribution of individual species. At the community level, variance in leaf size increased with increasing insolation. For individual species, there was a greater range of leaf size on south-facing slopes, while there was an absence of small-leaved species on north-facing slopes. These results demonstrate that analyses of plant functional traits along environmental gradients based on community level averages may obscure important aspects of trait variation and distribution among the constituent species.
Angiosperm leaf vein evolution was physiologically and environmentally transformative
Leaf size and foraging for light in a sclerophyll woodland
DOI:10.1046/j.1365-2435.2002.00661.x URL [本文引用: 2]
Stem hydraulic supply is linked to leaf photosynthetic capacity: evidence from New Caledonian and Tasmanian rainforests
Leaf maximum photosynthetic rate and venation are linked by hydraulics
Leaf veins are almost ubiquitous across the range of terrestrial plant diversity, yet their influence on leaf photosynthetic performance remains uncertain. We show here that specific physical attributes of the vascular plumbing network are key limiters of the hydraulic and photosynthetic proficiency of any leaf. Following the logic that leaf veins evolved to bypass inefficient water transport through living mesophyll tissue, we examined the hydraulic pathway beyond the distal ends of the vein system as a possible limiter of water transport in leaves. We tested a mechanistic hypothesis that the length of this final traverse, as water moves from veins across the mesophyll to where it evaporates from the leaf, governs the hydraulic efficiency and photosynthetic carbon assimilation of any leaf. Sampling 43 species across the breadth of plant diversity from mosses to flowering plants, we found that the post-vein traverse as determined by characters such as vein density, leaf thickness, and cell shape, was strongly correlated with the hydraulic conductivity and maximum photosynthetic rate of foliage. The shape of this correlation provided clear support for the a priori hypothesis that vein positioning limits photosynthesis via its influence on leaf hydraulic efficiency.
PhenoVein-A tool for leaf vein segmentation and analysis
DOI:10.1104/pp.15.00974
PMID:26468519
[本文引用: 4]
Precise measurements of leaf vein traits are an important aspect of plant phenotyping for ecological and genetic research. Here, we present a powerful and user-friendly image analysis tool named phenoVein. It is dedicated to automated segmenting and analyzing of leaf veins in images acquired with different imaging modalities (microscope, macrophotography, etc.), including options for comfortable manual correction. Advanced image filtering emphasizes veins from the background and compensates for local brightness inhomogeneities. The most important traits being calculated are total vein length, vein density, piecewise vein lengths and widths, areole area, and skeleton graph statistics, like the number of branching or ending points. For the determination of vein widths, a model-based vein edge estimation approach has been implemented. Validation was performed for the measurement of vein length, vein width, and vein density of Arabidopsis (Arabidopsis thaliana), proving the reliability of phenoVein. We demonstrate the power of phenoVein on a set of previously described vein structure mutants of Arabidopsis (hemivenata, ondulata3, and asymmetric leaves2-101) compared with wild-type accessions Columbia-0 and Landsberg erecta-0. phenoVein is freely available as open-source software. © 2015 American Society of Plant Biologists. All Rights Reserved.
The spatial pattern of air seeding thresholds in mature sugar maple trees
Leaf trait diversification and design in seven rare taxa of the Hawaiian Plantago radiation
DOI:10.1086/593111 URL [本文引用: 1]
Differences in leaf thermoregulation and water use strategies between three co-occurring Atlantic forest tree species
Comparative studies of leaf form: assessing the relative roles of selective pressures and phylogenetic constraints
DOI:10.1111/j.1469-8137.1987.tb04687.x URL [本文引用: 1]
On the adaptive significance of leaf form
//Solbrig OT, Jain S, Johnson GB, Raven PH.
Sizes and shapes of liane leaves
DOI:10.1086/283101 URL [本文引用: 1]
Water transport in impaired leaf vein systems
DOI:10.1055/s-2002-35429 URL [本文引用: 1]
Leaf venation patterns and principles of evolution
Inherent variation in growth rate between higher plants: a search for physiological causes and ecological consequences
Manual of Leaf Architecture-Morphological Description and Categorization of Dicotyledonous and Net-veined Monocotyledonous Angiosperms
Leaf size of woody dicots predicts ecosystem primary productivity
DOI:10.1111/ele.13503 URL [本文引用: 1]
Leaf-size divergence along rainfall and soil-nutrient gradients: Is the method of size reduction common among clades
DOI:10.1046/j.1365-2435.2003.00698.x URL [本文引用: 4]
Decoding leaf hydraulics with a spatially explicit model: principles of venation architecture and implications for its evolution
DOI:10.1086/650721 URL [本文引用: 3]
Convergence of terrestrial plant production across global climate gradients
DOI:10.1038/nature13470 URL [本文引用: 1]
The scaling of leaf area and mass: the cost of light interception increases with leaf size
DOI:10.1098/rspb.2007.0417 URL [本文引用: 1]
Effects of the experimental blockage of the major veins on hydraulics and gas exchange of Prunus laurocerasus L. leaves
The impact of leaf vein blockage on leaf hydraulic conductance (K(L)), gas exchange (g(L)) and water potential (Psi(L)) was studied in Prunus laurocerasus L., a broad-leaved evergreen. For this purpose, leaves were measured for the three variables above, either with an intact leaf blade (controls) or with the midrib cut a third of the way up (cut a), or with the midrib cut at three different points and the first-order veins cut through near their insertion to the midrib (cut b), or with the midrib cut at 2 mm from the leaf base (cut c). All the cut surfaces were sealed with cyanoacrylate. A serial decrease of K(L) was recorded from cut a to cut c with respect to that measured for the controls, i.e. a K(L) loss of about 37% (cut a), 57% (cut b) and 87% (cut c). A positive linear relationship appeared to exist between g(L) and K(L) with a high correlation coefficient (r(2)=0.99) and a high statistical significance (P <0.01). Even under a severe drop in K(L) (as that induced by cut c), leaf water potential remained approximately constant and not statistically different from Psi(L) measured for the controls. In fact, Psi(L) ranged between -0.83 and -0.98 MPa, i.e. within the cavitation threshold of leaves in terms of the critical Psi(L) inducing a significant production of ultrasound acoustic emissions which was -0.94+/-0.09 MPa. The conclusion was that stomata were very sensitive to changes in K(L) and that stomatal closure led to the homeostatic maintenance of Psi(L) and cavitation avoidance.
Xylem cavitation in the leaf of Prunus laurocerasus and its impact on leaf hydraulics
This paper reports how water stress correlates with changes in hydraulic conductivity of stems, leaf midrib, and whole leaves of Prunus laurocerasus. Water stress caused cavitation-induced dysfunction in vessels of P. laurocerasus. Cavitation was detected acoustically by counts of ultrasonic acoustic emissions and by the loss of hydraulic conductivity measured by a vacuum chamber method. Stems and midribs were approximately equally vulnerable to cavitations. Although midribs suffered a 70% loss of hydraulic conductance at leaf water potentials of -1.5 MPa, there was less than a 10% loss of hydraulic conductance in whole leaves. Cutting and sealing the midrib 20 mm from the leaf base caused only a 30% loss of conduction of the whole leaf. A high-pressure flow meter was used to measure conductance of whole leaves and as the leaf was progressively cut back from tip to base. These data were fitted to a model of hydraulic conductance of leaves that explained the above results, i.e. redundancy in hydraulic pathways whereby water can flow around embolized regions in the leaf, makes whole leaves relatively insensitive to significant changes in conductance of the midrib. The onset of cavitation events in P. laurocerasus leaves correlated with the onset of stomatal closure as found recently in studies of other species in our laboratory.
Leaf shape and venation pattern alter the support investments within leaf lamina in temperate species: a neglected source of leaf physiological differentiation
A mechanical perspective on foliage leaf form and function
DOI:10.1046/j.1469-8137.1999.00441.x URL [本文引用: 1]
“Diminishing returns” in the scaling of functional leaf traits across and within species groups
Optimal vein density in artificial and real leaves
DOI:10.1073/pnas.0709194105
PMID:18599446
[本文引用: 1]
The long evolution of vascular plants has resulted in a tremendous variety of natural networks responsible for the evaporatively driven transport of water. Nevertheless, little is known about the physical principles that constrain vascular architecture. Inspired by plant leaves, we used microfluidic devices consisting of simple parallel channel networks in a polymeric material layer, permeable to water, to study the mechanisms of and the limits to evaporation-driven flow. We show that the flow rate through our biomimetic leaves increases linearly with channel density (1/d) until the distance between channels (d) is comparable with the thickness of the polymer layer (delta), above which the flow rate saturates. A comparison with the plant vascular networks shows that the same optimization criterion can be used to describe the placement of veins in leaves. These scaling relations for evaporatively driven flow through simple networks reveal basic design principles for the engineering of evaporation-permeation-driven devices, and highlight the role of physical constraints on the biological design of leaves.
The relevance of specific language impairment in understanding the role of transfer in second language acquisition
Optimal leaf size in relation to environment
DOI:10.2307/2258359 URL [本文引用: 1]
Sensitivity of leaf size and shape to climate: global patterns and paleoclimatic applications
DOI:10.1111/j.1469-8137.2010.03615.x URL [本文引用: 1]
A note on normal correlation
DOI:10.1093/biomet/31.1-2.9 URL [本文引用: 1]
Scaling and structure of dicotyledonous leaf venation networks
DOI:10.1111/j.1461-0248.2011.01712.x URL [本文引用: 2]
The correlations of leaf anatomical characteristics with photosynthetic capacity and drought tolerance in seven sugarcane cultivars
DOI:10.1104/pp.53.5.705 URL [本文引用: 2]
七个甘蔗品种叶片解剖结构特征与光合能力和耐旱性的关联
Evolution and function of leaf venation architecture: a review
DOI:10.1006/anbo.2001.1391 URL [本文引用: 2]
The major veins of mesomorphic leaves revisited: tests for conductive overload in Acer saccharum (Aceraceae) and Quercus rubra (Fagaceae)
DOI:10.3732/ajb.90.1.32 URL [本文引用: 1]
Leaf palmate venation and vascular redundancy confer tolerance of hydraulic disruption
Leaf structural diversity is related to hydraulic capacity in tropical rain forest trees
DOI:10.1890/05-0710 URL [本文引用: 3]
How do leaf veins influence the worldwide leaf economic spectrum? Review and synthesis
DOI:10.1093/jxb/ert316 URL [本文引用: 2]
Developmentally based scaling of leaf venation architecture explains global ecological patterns
DOI:10.1038/ncomms1835 URL [本文引用: 10]
Axial-to-radial water permeability of leaf major veins: a possible determinant of the impact of vein embolism on leaf hydraulics
Decline of leaf hydraulic conductance with dehydration: relationship to leaf size and venation architecture
DOI:10.1104/pp.111.173856 URL [本文引用: 4]
Organ shape and size: a lesson from studies of leaf morphogenesis
DOI:10.1016/S1369526602000055 URL [本文引用: 1]
Aluminum toxicity in alfalfa (Medicago sativa) is alleviated by exogenous foliar IAA inducing reduction of Al accumulation in cell wall
DOI:10.1016/j.envexpbot.2017.03.018 URL [本文引用: 2]
Bivariate line-fitting methods for allometry
DOI:10.1017/S1464793106007007 URL [本文引用: 1]
Leaf and twig functional traits of woody plants and their relationships with environmental change: a review
木本植物茎叶功能性状及其关系随环境变化的研究进展

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}