



植被生物量在纬度梯度上的变化规律及其成因是揭示植被结构特征与环境相互作用的重要途径。纬度梯度上气候、土壤和植被等非生物、生物因素及其相互作用都会发生不同程度的变化(Scolforo et al., 2015; Zhang et al., 2015)。例如全球森林和我国西北干旱区草地(31.70°-39.58° N)群落生物量随着纬度升高呈显著增加趋势(韩彬等, 2006; 邓蕾和上官周平, 2012; Xu et al., 2018; Qi et al., 2019)。相反地, 在南美洲森林、稀树草原、我国北方温带森林和内蒙古草原等植被地上碳储量、叶生物量以及地上生物量随纬度升高而降低(Scolforo et al., 2015; Zhang et al., 2015; 张俊怡等, 2020); 在我国黄土高原(35.95°-38.36° N), 草本植物生物量随纬度的升高先增加后减少(李婷等, 2015); 在欧洲东北部温带森林细根生物量随纬度变化趋势不明显(Finér et al., 2019)等。这些研究多关注森林和草地生态系统, 并且由于他们涉及的生态系统类型、研究尺度及取样对象不同, 导致植被生物量纬度格局及其成因复杂而多样。谢宗强等(2019)通过对我国西南山地灌丛生态系统调查及模型估算, 发现其生物量与纬度无显著相关性。灌丛作为陆地生态系统的重要组成部分, 在干旱、半干旱以及高寒等资源贫瘠的地区广泛分布, 在植被结构和功能及其对环境的适应性方面具有独特性。研究典型灌丛群落及其组分生物量的纬度格局和成因可充分认识陆地生态系统植被结构和功能, 并且有助于解释慢速生长的木本植物对环境变化的生态适应性。然而, 现有研究对灌丛植被生物量及其纬度格局认识不足。
环境因子(气候和土壤)、植被的物种组成、结构变化和人为干扰均可对生物量产生影响。植被结构变化及其起主导作用的环境因子驱动了自然植被生物量纬度格局的产生。一般而言, 在大尺度上植被组成及其生物量主要受气候因素的影响(马安娜等, 2014)。例如, 研究表明, 纬度梯度上温带草地生物量与降雨量之间显著正相关(Ma et al., 2008)。纬度梯度上温度变化通常显著, 可能对植被产生重要影响。但是, 研究表明, 纬度梯度上植被生物量与温度之间的关联强度及方向并不一致, 例如高寒灌丛生物量与年平均气温之间呈显著正相关关系(聂秀青等, 2016); 温带森林根系生物量与年平均气温之间呈负相关关系(Ma et al., 2008); 而温带草地地上生物量与年平均气温无显著相关性(Sala et al., 1988; Epstein et al., 1997)。植被生物量与土壤因子密切相关。通常纬度梯度上根系生物量与土壤密度、土层厚度呈显著负相关关系, 与土壤养分(全碳、全氮、全磷)含量呈显著正相关关系(刘顺等, 2018; 唐立涛等, 2019; 杜美琪等, 2020)。此外, 群落数量特征等生物因素变化(物种更替、结构和多样性改变等)对群落生物量具有多方面的影响(Cuesta et al., 2010)。例如物种丰富度与生物量之间存在显著的正相关关系(Zhang et al., 2017; Guo et al., 2019)。
我国西南干旱河谷区自然植被多以灌丛为主(刘伦辉, 1989), 植被发育缓慢、结构单一、地表覆盖度较低、水土流失严重, 是典型的生态脆弱区。不同流域的植被差异较大, 其中干热河谷多以稀树灌草丛和肉质灌丛为主。干旱河谷部分地段如元江流域主要以黄茅(Heteropogon contortus)和拟金茅(Eulaliopsis binata)为优势种形成草地植被, 草本生物量大于灌木生物量(金振洲等, 1987); 干暖河谷主要是暖性落叶阔叶灌丛、常绿硬叶林、常绿阔叶灌丛, 也伴有耐旱、硬叶的乔木和灌丛(刘晔等, 2016a); 干温河谷主要是小叶灌丛和成丛散生的阔叶灌丛, 草本植物稀少(刘晔等, 2016b; 沈泽昊, 2016)。现有研究调查了干旱河谷部分流域植被生物量的数量特征。例如, 岷江上游地区群落地上生物量最大的物种为对节刺(Horaninovia ulicina)(11.55 t·hm-2), 最小的为滇紫草(Onosma paniculatum)(1.41 t·hm-2), 其余灌丛类型介于二者之间(刘国华等, 2003)。岷江河谷阴、阳坡平均群落生物量分别为3.02和1.49 t·hm-2, 其灌丛总生物量、各层片生物量以及各器官生物量均表现为随海拔升高而增加的趋势(陈泓等, 2007)。长期以来, 研究大多描述性地介绍干旱河谷主要灌丛类型及其特征, 而对干旱河谷植被组成、结构和生物量整体特征、空间差异及其驱动因素等缺乏系统研究, 尚不能解释干旱河谷现有植被生态适应及其机理。缺乏对群落特征与气候、土壤等环境因子之间关系的定量分析, 难以解释植被生物量空间格局及其成因。这些问题不仅限制了该区域生态系统服务功能的正确评价, 也影响生态系统的有效保护与持续利用。
本研究以西南干旱河谷植被作为研究对象,系统调查群落及其组分的生物量, 重点关注两方面的科学问题: 1)干旱河谷不同纬度植物群落及组分生物量如何变化? 2)纬度梯度上植物群落及组分生物量变化与环境因子之间如何关联? 主导生态因子的作用机制是什么? 本研究的结果可提升对干旱河谷植被空间差异性及形成机制的理论认识, 为区域脆弱生态系统恢复与保护提供理论依据。
1 材料和方法
1.1 研究区概况
干旱河谷是横断山区高山峡谷区特有的景观类型, 具有明显的峡谷深切地貌特征。该地区气候干湿季差异大, 冬季干冷, 夏季暖湿(包维楷等, 2012), 焚风效应明显, 年降水量在662-1 350 mm之间, 年平均气温在6.42-24.10 ℃之间。根据不同流域的气候和植被等特征差异, 通常分为干热河谷、干暖河谷、干温河谷等类型(张荣祖, 1992), 也体现了明显的从南向北的纬度变化格局。干旱河谷地区自然植被类型主要是稀树灌(草)丛、肉质灌丛和小叶刺灌丛等(刘伦辉, 1989)。土壤以褐红壤为主, 总体上干旱、贫瘠, 空间差异明显, 易受人为干扰影响(张荣祖, 1992)。该地区人口相对密集, 是典型的自然-社会-经济综合生态系统(包维楷等, 2012)。研究区域位于川滇两省境内的典型干旱河谷地区(23.23°-32.26° N, 101.16°-103.52° E)(图1)。本研究调查取样点的分布从南至北跨越9个纬度, 根据范建容等(2020)最新界定的我国西南干旱河谷纬度范围(22.99°-34.08° N, 97.42°-105.16° E), 取样点覆盖了干旱河谷地区所在纬度范围的75%左右。从南至北海拔高度由323 m (云南元江)上升至2 688 m (四川松潘)。
图1
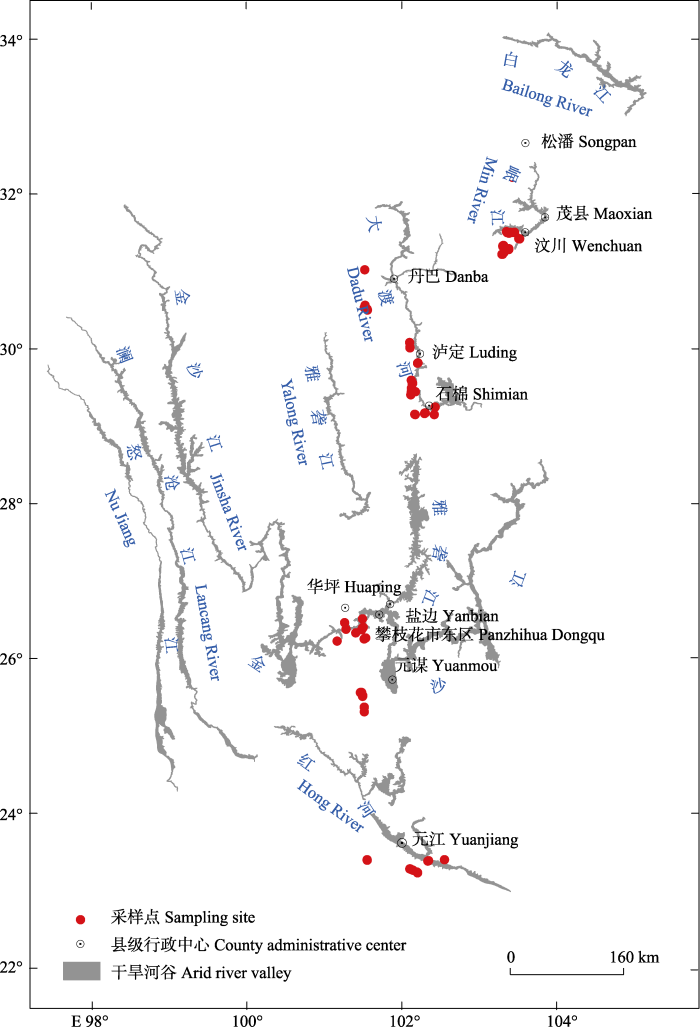
图1
西南干旱河谷不同纬度植物群落生物量取样地点。
Fig. 1
Biomass sampling sites of plant communities along latitudes in the arid valley, southwestern China.
1.2 不同纬度样地选择
由于本研究重点关注自然植被生物量随纬度梯度的变化格局, 因此所有样方的经度控制在2°以内(101.40°-103.52° E)。基于此原则, 于2019年7-8月和2020年7-8月植物生长旺盛期, 由南至北沿元江、金沙江、雅砻江、大渡河、岷江河谷流域跨越9个纬度(23.23°-32.26° N), 进行野外样地调查与取样。依据纬度、海拔、坡向、土壤类型等差异, 在每个流域干旱河谷地区选取自然植被类型, 布置3-5个点, 每个点布设3-6个面积为4 m × 6 m的群落样方, 总共获得有效样方101个(图1)。北纬27°-28°之间的地区, 自然植被受人为活动破坏严重, 未取样。
1.3 群落调查和生物量取样
1.3.1 样方布设与群落调查
群落数量特征采用典型群落调查法(方精云等, 2009)进行。本研究共调查获得101个4 m × 6 m的植物群落样方, 记录各样方的环境因子(经纬度、海拔、坡向、坡度、坡位等)、群落数量参数(群落类型、盖度、凋落物盖度等)和人为干扰情况等。群落调查时, 首先将上述每个面积4 m × 6 m的样方细分为6个2 m × 2 m的灌木样方, 记录灌木各物种高度、盖度、多度等参数。然后, 在每个植物分布较均匀的灌木样方中, 对角线布置2个l m × l m的草本样方, 记录草本植物各物种的高度、盖度、多度等参数。根据《中国植物志》鉴定标本。参照方精云等(2009)方法, 分别计算草本和灌木的Shannon-Wiener指数和灌木Margalef指数。
1.3.2 生物量测定
群落调查后, 每个上述面积4 m × 6 m样地中选取1个植株密度相对均匀的灌木样方, 采用收割法分不同物种进行灌木生物量取样。首先, 收割灌木地上部分, 并将其花果、茎、叶器官分开, 装入信封, 编号。然后, 挖取样方内每个物种的全部根系, 并以直径2 cm为分界, 将每个物种的粗根与细根分开, 装入信封, 编号。对于个体较大的灌木, 用电子秤称量上述样品鲜质量后, 均匀取200 g左右, 装入信封, 编号带回室内, 烘干, 通过含水率推算这些物种的生物量。样方内若有不能分辨的物种根系, 将其分为粗根和细根, 测定干质量, 计入样方总生物量。草本植物生物量和凋落物生物量在面积1 m × 1 m小样方内进行。在上述每个面积4 m × 6 m样地中, 采用对角线机械布点, 选3个1 m × 1 m小样方。这3个小样方包括1个灌木生物量测定对应的样方和其余2个草本生长相对均匀的小样方。首先, 收集每个小样方中所有凋落物, 然后混合取样后编号, 带回实验室。然后, 采用收割法对草本植物进行取样, 测定其生物量。每个小样方中草本地上、地下部分分开收割, 然后将3个样方混合取样, 编号, 带回实验室。所有新鲜植物样品在105 ℃杀青2 h, 然后在65 ℃下烘干至恒质量, 称量。根据公式(1)计算含水率, 并根据样品干质量, 用公式(2)换算全株生物量。公式如下:
式中, P为含水率; mF为样品鲜质量; mD为样品干质量。
群落生物量(CB)为样方内所有灌木、草本和凋落物生物量总和; 灌木生物量(SB)为样方内所有灌木茎、叶、粗根及细根生物量之和; 草本生物量(HB)为样方内所有草本地上与地下生物量之和。凋落物生物量(LB)为样方内凋落物、草本枯死物和灌木枯死枝生物量的总和。植被地上生物量(VAGB)包括灌木地上和草本地上生物量; 植被地下生物量(VBGB)包括灌木地下和草本地下生物量; 灌木地上生物量(SAGB)包括灌木茎和叶生物量; 灌木地下生物量(SBGB)包括灌木粗根和细根生物量。所有生物量单位用t·hm-2表示。
1.4 土壤理化性质测定
采用土钻法在上述每个灌木样方沿对角线进行0-10 cm土层土壤样品采集, 每个样地共取18钻, 混合成一个土壤样品。土壤样品挑出砾石和根系, 过 2 mm筛后分成两份, 一份4 ℃冷藏保存用于速效养分测定, 另一份置于阴凉干燥处风干用于常量养分测定。土壤pH采用电位法测定(水土体积比= 2.5:1); 土壤总碳(TC)和总氮(TN)含量采用元素分析仪(Vario MACRO cube, Elementar, Langenselbold, Germany)测定; 土壤总磷(TP)含量采用浓硫酸-高氯酸酸溶、钼锑钪比色法测定; 土壤NO3--N和NH4+-N采用2 mol·L-1 KCl提取、连续流动分析仪(AA3, SEAL, Norderstedt, Germany)测定含量, 总无机氮(TIN)含量为NO3--N和NH4+-N含量之和; 土壤阳离子交换量(CEC)用乙酸钠-火焰光度法测定; 土壤钙离子(Ca2+)和镁离子(Mg2+)含量用乙二胺四乙酸(EDTA)络合滴定法测得。
1.5 气候数据
使用ArcGIS 10.2.2将气象数据按经纬度提取每个样方的年平均气温(MAT)和年降水量(MAP)数据, 作为气候因子WorldClim数据库(
1.6 数据分析
以样方为统计单元对植物群落、土壤、气候数据进行整理、汇总, 计算出群落及各组分生物量的最大值、最小值、平均值和标准误(表1)。数据分析前, 对群落及其各组分生物量进行对数转换使实测数值更接近于正态分布。采用一般线性模型模拟群落及其各组分生物量在纬度梯度上的变化格局。对于生物量与环境因子之间的关系, 首先采用Pearson相关系数初步分析了群落及其组分生物量与群落多样性指数、土壤因子和气候因子之间的相关性。然后以生物、土壤、气候因子3类因子为解释变量, 采用方差分解分析(VPA)检验不同类型环境因子及其组合对群落生物量和组分生物量在纬度梯度上变化的解释程度。最后以群落生物量、灌木生物量、草本生物量、凋落物生物量作为响应变量, 以灌木盖度、土壤总碳含量、年平均气温等因子作为解释变量对二者进行冗余分析(RDA), 进一步综合分析影响植被生物量纬度梯度格局的主导环境因子。上述统计分析和作图用Origin 2018、R 4.0.5软件中的“vegan”包、“ggplot2”包和SPSS 19.0完成。
表1 西南干旱河谷群落及组分生物量的基本特征描述(n = 101)
Table 1
| 变量 Variable | 最大值 Maximum | 最小值 Minimum | 平均值±标准误 Mean ± SE |
|---|---|---|---|
| 群落生物量 CB (t·hm-2) | 56.50 | 2.51 | 17.05 ± 1.09 |
| 灌木生物量 SB (t·hm-2) | 52.49 | 0.26 | 11.51 ± 1.03 |
| 草本生物量 HB (t·hm-2) | 9.68 | 0.03 | 2.11 ± 0.21 |
| 凋落物生物量 LB (t·hm-2) | 18.34 | 0.29 | 3.41 ± 0.34 |
| 植被地上生物量 VAGB (t·hm-2) | 34.37 | 0.12 | 7.32 ± 0.58 |
| 植被地下生物量 VAGB (t·hm-2) | 43.57 | 0.21 | 6.30 ± 0.59 |
| 灌木地上生物量 SAGB (t·hm-2) | 32.73 | 0.10 | 6.51 ± 0.58 |
| 灌木地下生物量 SAGB (t·hm-2) | 42.21 | 0.12 | 5.00 ± 0.59 |
| 灌木生物量百分比 SB/CB (%) | 96.53 | 1.62 | 60.18 ± 2.76 |
| 草本生物量百分比 HB/CB (%) | 69.90 | 0.57 | 15.57 ± 1.67 |
| 凋落物生物量百分比 LB/CB (%) | 86.67 | 1.57 | 24.14 ± 2.09 |
CB, community biomass; HB, herb biomass; LB, litter biomass; SB, shrub biomass; SAGB, shrub aboveground biomass; SBGB, shrub belowground biomass; VAGB, vegetation aboveground biomass; VBGB, vegetation belowground biomass.
2 研究结果
2.1 西南干旱河谷植被生物量及其在不同纬度的变化特点
干旱河谷纬度梯度(23.23°-32.26° N)上, 群落生物量变化范围在2.51-56.50 t·hm-2之间, 平均值是(17.05 ± 1.09) t·hm-2 (表1), 随着纬度增加, 群落生物量具有显著增加趋势(图2A)。灌木生物量的变化范围在0.26-52.49 t·hm-2之间, 平均值是(11.51 ± 1.03) t·hm-2 (表1), 随着纬度增加也具有显著增加趋势(图2B)。草本生物量的变化范围在0.03-9.68 t·hm-2之间, 平均值是(2.11 ± 0.21) t·hm-2 (表1 ), 纬度梯度上无明显变化(图2C)。凋落物生物量的变化范围在0.29-18.34 t·hm-2之间, 平均值是(3.41 ± 0.34) t·hm-2 (表1), 随着纬度增加具有显著的减小趋势(图2D)。
图2
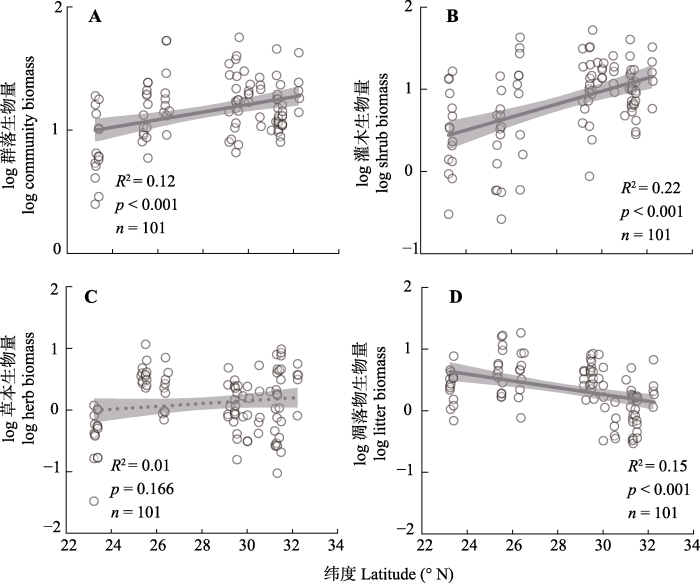
图2
西南干旱河谷纬度梯度上群落及其组分生物量的变化。
Fig. 2
Changes in biomass of plant communities and its components along latitude gradients in the arid valley, southwestern China.
2.2 西南干旱河谷植被各组分生物量比例在不同纬度的变化特点
图3
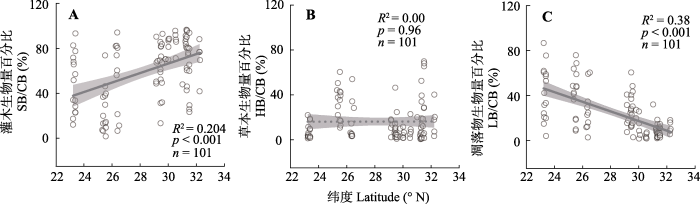
图3
西南干旱河谷植物群落各组分生物量比例在纬度梯度上的变化。
Fig. 3
Changes in proportions of biomass of community components along latitude gradients in the arid valley, southwestern China. CB, community biomass; HB, herb biomass; LB, litter biomass; SB, shrub biomass.
2.3 西南干旱河谷纬度梯度上植物群落及其组分生物量变化的影响因子
环境因子对群落及组分生物量的RDA结果如图4所示。RDA1和RDA2轴解释了总变异的53.11%。解释变量中, 灌木盖度、草本盖度、凋落物盖度、草本Shannon-Wiener指数、灌木Shannon-Wiener指数和年平均气温主要驱动了RDA1的变化(图4; 表2); 群落平均高度、草本Shannon-Wiener指数和年平均降水量主要驱动RDA2的变化(图4; 表2)。响应变量灌木生物量和凋落物生物量对RDA1贡献最大; 草本生物量对RDA2贡献最大; 群落生物量在前两轴中无明显变化(图4; 表2)。RDA1轴从左到右, 草本盖度、凋落物盖度、草本Shannon-Wiener指数和年平均气温表现出增大趋势(图4); RDA2轴从下到上, 群落平均高度、草本Shannon-Wiener指数、年降水量表现出增大趋势(图4)。
图4
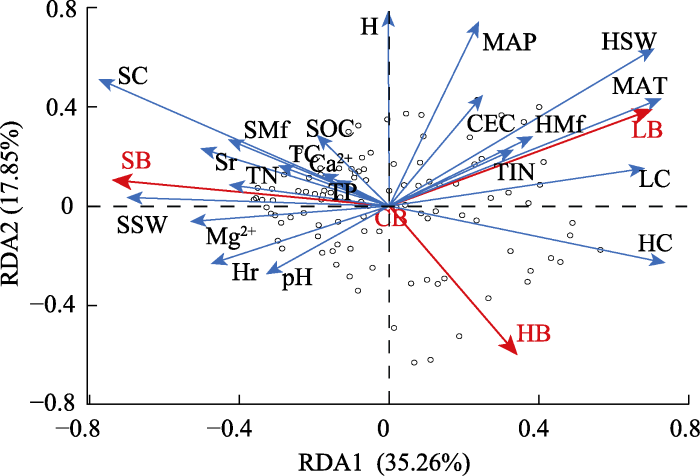
图4
西南干旱河谷植物群落及各组分生物量与环境因子的冗余分析(RDA)(n = 101)。Ca2+, 土壤钙离子交换量; CB, 群落生物量; CEC, 土壤阳离子交换量; H, 群落平均高度; HB, 草本生物量; HC, 草本盖度; HMf, 草本Margalef指数; Hr, 草本丰富度; HSW, 草本Shannon-Wiener指数; LB, 凋落物生物量; LC, 凋落物盖度; MAP, 年降水量; MAT, 年平均气温; Mg2+, 土壤镁离子交换量; SB, 灌木生物量; SC, 灌木盖度; SMf, 灌木Margalef指数; SOC, 土壤有机碳含量; Sr, 灌木丰富度; SSW, 灌木Shannon-Wiener指数; TC, 土壤总碳含量; TIN, 土壤总无机氮含量; TN, 土壤总氮含量; TP, 土壤总磷含量。
Fig. 4
Redundancy analysis (RDA) of community biomass and environmental factors in the arid valley, southwestern China (n = 101). Ca2+, soil calcium ion exchange capacity; CB, community biomass; CEC, soil cation exchange capacity; H, mean community height; HB, herb biomass; HC, herb coverage; HMf, herb Margalef index; Hr, herb richness; HSW, herb Shannon-Wiener index; LB, litter biomass; LC, litter coverage; MAP, mean annual precipitation; MAT, mean annual air temperature; Mg2+, soil magnesium ion exchange capacity; SB, shrub biomass; SC, shrub coverage; SMf, shrub Margalef index; SOC, soil organic carbon content; Sr, shrub richness; SSW, shrub Shannon-Wiener index; TC, soil total carbon content; TIN, soil total inorganic nitrogen content; TN, soil total nitrogen content; TP, soil total phosphorus content.
表2 西南干旱河谷植物群落及各组分生物量冗余分析(RDA)的前三轴特征值(n = 101)
Table 2
| 变量 Variable | 第一轴 RDA1 | 第二轴 RDA2 | 第三轴 RDA3 |
|---|---|---|---|
| 土壤阳离子交换量 Soil cation exchange capacity | 0.22 | 0.41 | 0.19 |
| 土壤总磷含量 Soil total phosphorus content | -0.11 | 0.09 | 0.15 |
| 土壤总氮含量 Soil total nitrogen content | -0.37 | 0.08 | -0.18 |
| 土壤总碳含量 Soil total carbon content | -0.25 | 0.15 | -0.21 |
| 土壤 pH Soil pH | -0.28 | -0.25 | -0.50 |
| 土壤有机碳含量 Soil organic carbon content | -0.16 | 0.26 | 0.03 |
| 土壤钙离子交换量 Soil calcium ion exchange capacity | -0.14 | 0.11 | -0.28 |
| 土壤镁离子交换量 Soil magnesium ion exchange capacity | -0.47 | -0.06 | 0.24 |
| 土壤总无机氮含量 Soil total inorganic nitrogen content | 0.28 | 0.21 | -0.03 |
| 群落平均高度 Mean community height | 0.00 | 0.77 | 0.31 |
| 灌木盖度 Shrub coverage | -0.70 | 0.49 | 0.11 |
| 草本盖度 Herb coverage | 0.66 | -0.22 | 0.33 |
| 凋落物盖度 Litter coverage | 0.61 | 0.15 | 0.42 |
| 灌木丰富度 Shrub richness | -0.44 | 0.22 | 0.19 |
| 草本丰富度 Herb richness | -0.42 | -0.22 | 0.31 |
| 草本Shannon-Wiener指数 Herb Shannon-Wiener index | 0.64 | 0.61 | -0.21 |
| 草本Margalef指数 Herb Margalef index | 0.33 | 0.26 | -0.41 |
| 灌木Shannon-Wiener指数 Shrub Shannon-Wiener index | -0.63 | 0.03 | 0.16 |
| 灌木Margalef指数 Shrub Margalef index | -0.38 | 0.25 | 0.21 |
| 年降水量 Mean annual precipitation | 0.21 | 0.70 | 0.06 |
| 年平均气温 Mean annual air temperature | 0.65 | 0.42 | 0.07 |
| 群落生物量 Community biomass | 0.00 | 0.00 | 0.00 |
| 灌木生物量 Shrub biomass | -0.65 | 0.10 | 0.09 |
| 草本生物量 Herb biomass | 0.30 | -0.56 | 0.06 |
| 凋落物生物量 Litter biomass | 0.62 | 0.37 | 0.07 |
群落生物量与年降水量以及土壤pH具有显著负相关关系(表3)。灌木生物量与灌木盖度、灌木丰富度、草本丰富度、灌木Shannon-Wiener指数、灌木Margalef指数呈现显著正相关关系(表3; 图4), 但是灌木生物量与草本Shannon-Wiener指数及年平均气温呈现显著负相关关系(表3; 图4)。草本生物量与草本盖度呈显著正相关关系(表3; 图4), 但与群落平均高度、草本Shannon-Wiener指数、灌木盖度、灌木丰富度、年降水量和土壤阳离子交换量呈显著负相关关系(表3; 图4)。凋落物生物量与群落平均高度、凋落物盖度、草本Shannon-Wiener指数、土壤阳离子交换量、年平均气温呈显著正相关关系(表4; 图4), 与灌木盖度、灌木Shannon-Wiener指数、草本丰富度、土壤pH、镁离子交换量呈显著负相关关系(表3; 图4)。
表3 西南干旱河谷植物群落及各组分生物量与环境因子之间的Pearson相关系数(n = 101)
Table 3
| 变量 Variable | 群落 Community | 灌木 Shrub | 草本 Herb | 凋落物 Litter |
|---|---|---|---|---|
| 土壤阳离子交换量 Soil cation exchange capacity | 0.028 | -0.011 | -0.202* | 0.244* |
| 土壤总磷含量 Soil total phosphorus content | -0.010 | 0.022 | -0.088 | -0.033 |
| 土壤总氮含量 Soil total nitrogen content | 0.094 | 0.183 | -0.074 | -0.182 |
| 土壤总碳含量 Soil total carbon content | 0.083 | 0.140 | -0.082 | -0.090 |
| 土壤pH Soil pH | -0.236* | -0.115 | 0.046 | -0.418** |
| 土壤有机碳含量 Soil organic carbon content | 0.045 | 0.084 | -0.128 | -0.016 |
| 土壤钙离子交换量 Soil calcium ion exchange capacity | -0.034 | 0.051 | -0.076 | -0.191 |
| 土壤镁离子交换量 Soil magnesium ion exchange capacity | 0.055 | 0.155 | -0.082 | -0.213* |
| 土壤总无机氮含量 Soil total inorganic nitrogen content | 0.013 | -0.046 | -0.053 | 0.201* |
| 群落均高 Mean community height | 0.053 | 0.072 | -0.414** | 0.220* |
| 灌木盖度 Shrub coverage | 0.014 | 0.212* | -0.483** | -0.244* |
| 草本盖度 Herb coverage | 0.090 | -0.100 | 0.282** | 0.364** |
| 凋落物盖度 Litter coverage | 0.077 | -0.092 | 0.074 | 0.436** |
| 灌木丰富度 Shrub richness | 0.157 | 0.250* | -0.206* | -0.097 |
| 草本丰富度 Herb richness | 0.159 | 0.239* | 0.045 | -0.219* |
| 草本Shannon-Wiener指数 Herb Shannon-Wiener index | -0.155 | -0.224* | -0.295** | 0.347** |
| 草本Margalef指数 Herb Margalef index | -0.143 | -0.147 | -0.151 | 0.082 |
| 灌木Shannon-Wiener指数 Shrub Shannon-Wiener index | 0.073 | 0.224* | -0.139 | -0.314** |
| 灌木Margalef指数 Shrub Margalef index | 0.144 | 0.211* | -0.185 | -0.041 |
| 年降水量 Mean annual precipitation | -0.262** | -0.189 | -0.449** | 0.034 |
| 年平均气温 Mean annual air temperature | -0.077 | -0.199* | -0.110 | 0.395** |
*, p < 0.05; **, p < 0.01.
方差分解结果显示, 环境对群落生物量的解释率大小依次是气候因子(17.5%) >生物因子(13.6%) >土壤因子(11.8%)(图5)。生物因子、土壤因子和气候因子单个因子解释率分别占1.0%、2.3%和7.8%。生物因子和土壤因子共同解释了6.2%, 生物因子和气候因子共同解释了6.4%, 气候因子和土壤因子共同解释了3.3% (图5)。环境对灌木生物量的解释率大小依次是生物因子(27.6%) >土壤因子(17.0%) >气候因子(11.2%), 单独贡献率生物因子、气候因子分别占11.4%和1.5%, 生物因子和土壤因子共同解释了9.2%, 生物因子和气候因子共同解释了1.9%, 气候因子和土壤因子共同解释了2.7%, 三者共同解释了5.1% (图5)。对草本生物量的解释率大小依次是生物因子(61.2%) >气候因子(30.9%) >土壤因子(2.9%), 单独贡献率生物因子和气候因子分别占27.9%和0.2%, 生物因子和土壤因子共同解释了2.6%, 生物因子和气候因子共同解释了30.4%, 三者共同解释了0.3% (图5)。环境对凋落物生物量的解释率大小依次是生物因子(46.9%) >土壤因子(35.8%) >气候因子(25.4%), 单独贡献率生物因子占10.6%, 土壤因子占1.6%, 生物因子和土壤因子共同解释了11.0%, 生物因子和气候因子共同解释了2.2%, 气候因子和土壤因子共同解释了0.1%, 三者共同解释了23.1% (图5)。
图5
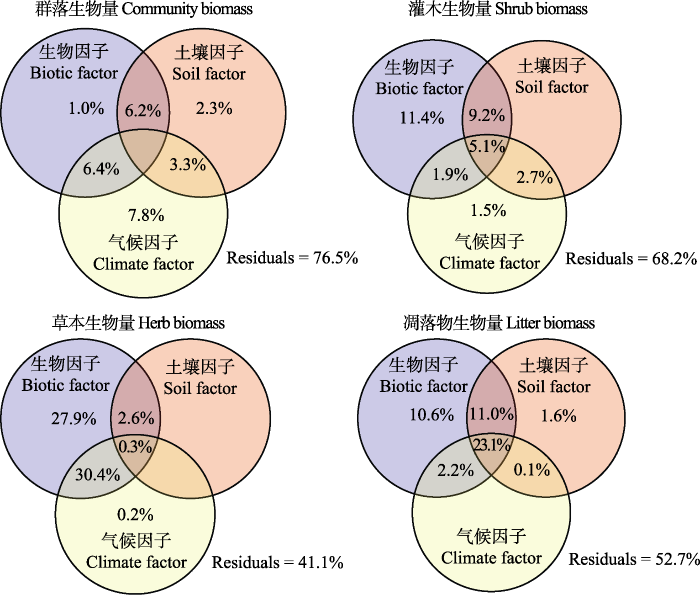
图5
西南干旱河谷植物群落及各组分生物量与环境因子的方差分解分析(n = 101)。Residuals, 残差。
Fig. 5
Variation partitioning analysis between biomass of community and its components and environmental factors in the arid valley, southwestern China (n = 101).
3 讨论和结论
西南干旱河谷在高山峡谷区的地形背景下植被结构、气候、土壤等呈现出较大的空间异质性以及相互作用的复杂性(张荣祖, 1992), 导致自然植被的群落及其组分生物量在空间尺度上变化很大, 表现在不同纬度之间以及相同纬度的不同微地形之间。怒江、澜沧江、金沙江和元江等流域气候特点与世界萨瓦纳地区气候特点较相似(金振洲和欧晓昆, 2000), 均表现出干旱、炎热、降水少等特点; 全年四季之分不明显, 但旱季和雨季特征十分明显, 且干旱期至少持续4个月(de Bie et al., 1998)。本研究在元江和金沙江河谷区的大多植被群落外貌中呈“稀树灌草丛” “稀树草丛” “灌草丛”状, 其群落类型和气候特征与萨瓦纳群落相似, 沈泽昊等(2016)认为这种干热河谷植被是全球萨瓦纳植被、地中海型马基植被在中国西南地区的残余和替代类型。因此, 我国西南干热河谷植被被称为“河谷型萨瓦纳植被(savanna of valley type)”。但是, 我们对元江等流域的萨瓦纳植物群落与大渡河、岷江等流域的灌丛群落作为一个整体分析, 发现植被结构和生物量有较大差异, 总体变化规律为: 随着区域内纬度增加, 自然植被的群落生物量及灌木生物量都具有逐渐增加的趋势, 草本植物生物量变化不明显, 凋落物生物量却明显减少(图2)。相关研究表明, 群落结构越复杂, 其生物量积累量越大; 群落中物种多样性减少会显著降低植被生物量和凋落物分解速率(Balvanera et al., 2006; Hooper et al., 2012)。另外, 群落内灌木、草本和凋落物等组分之间或者不同物种之间的相互作用将影响群落物种多样性, 进而影响生物量(Ma et al., 2010)。
研究结果显示, 在我国西南干旱河谷地区, 植物群落及组分生物量之间的关系复杂多样, 其中灌木生物量与群落高度、灌木丰富度、灌木盖度、草本丰富度、灌木Shannon-Wiener指数以及灌木Margalef指数之间呈显著正相关关系, 与草本Shannon-Wiener指数之间呈显著负相关关系(图4; 表3)。在干旱河谷地区, 灌木多样性和优势度都随着纬度升高而增大, 导致群落尤其是灌木生物量显著增大。此结果表明, 在干旱河谷地区, 不同纬度灌木的发育程度(高度、多样性和优势度等)是植被生物量形成纬度梯度格局的内在驱动因素。我们对群落内部不同组分之间的相关性分析发现, 草本生物量与群落高度、灌木盖度、灌木丰富度之间具有显著的负相关关系(图4; 表3)。这说明随着纬度增加灌木层发育对群落中草本植物个体生长和生物量积累产生一定的限制作用, 部分地支持Cuesta等(2010)的观点, 认为群落内灌木与草本植物存在相互抑制作用。然而, 我们也发现, 在干旱河谷地区草本丰富度与灌木生物量之间存在正相关关系(表3), 表明随着纬度增加, 尽管草本植物个体大小(用生物量表示)没有显著变化, 但草本丰富度增加, 可能有利于灌木生长。Maestre等(2003)在地中海半干旱地区的研究结果表明, 草本生物量与灌丛生物量之间存在正相关关系, 说明草本层与灌木层可能存在相互促进作用, 其原因是在这些干旱生态系统中, 群落下层草本植物覆盖能够改变群落微环境, 主要表现为有效降低土壤表面水分蒸发和改善土壤微生物环境, 从而促进灌木生长。在萨瓦纳稀树灌丛和其他干旱区灌丛中发现“灌丛肥岛”效应, 灌丛覆盖的区域比裸露地段更加有利于幼苗存活(胡慧等, 2020), 一方面, 灌木凋落物归还和遮阴可能提高灌丛土壤水分和养分; 另一方面, 深根系灌木能够将深层土壤的水分运输到表层, 提高土壤表层湿度, 促进群落内部浅根系植物生长(Pugnaire & Valladares, 2007)。研究结果表明在极端干旱环境中, 群落内部不同组分或者物种之间存在相互促进作用。
我们认为凋落物生物量随着纬度增加而呈明显下降趋势的原因主要在于两个方面: 一是从群落物种组成方面, 西南干旱河谷低纬度地区部分地段禾本科植物在草本层占据优势地位, 它们产生大量枯落物, 因此决定凋落物生物量的大小; 而在高纬度地区, 灌木优势度增加, 禾本科植物集中分布区域较少, 优势度不明显, 使得群落总凋落物生物量在低纬度地区明显大于高纬度地区; 二是纬度较高的地区如岷江流域等风速大, 冠层上部枯死部分通常随风迁移, 这对地表凋落物累积有一定的负面影响(包维楷等, 2012)。因此, 在干旱河谷纬度梯度上, 相比于土壤因子和气候因子, 生物因子对凋落物生物量的纬度变化解释率最高, 达46.9% (图5)。此外, 纬度梯度上凋落物生物量与凋落物自身的盖度、草本盖度、群落高度、草本Shannon-Wiener指数呈显著正相关关系, 而与草本丰富度、灌木盖度、灌木Shannon-Wiener指数呈显著负相关关系(图4; 表3), 这进一步证明干旱河谷地区草本植物个体生长量(而非多样性)是决定凋落物量的关键因素。
从外在环境因子来看, 气候因子对群落生物量的空间格局和生物多样性产生直接或者间接影响(Bai et al., 2008)。在全球尺度上纬度变化代表环境中热量的变化, 驱动了现有植被纬度梯度上的分布格局。在干旱河谷地区, 降水量和温度都随着纬度升高而降低, 干燥度变化不明显(范建容等, 2020)。研究结果显示, 在干旱河谷地区自然植物群落及其组分生物量纬度梯度上的变化格局是气候和土壤环境因子综合作用的结果。长期以来, 人们认为该地区植被受到水热综合作用的影响。其中, 水分短缺及其季节变化引起的干旱胁迫是影响植被组成和结构的关键生态因子, 而温度尤其是生长季节高温加剧了干旱胁迫对植物生长与分布的限制作用(李芳兰等, 2009; 胡慧等, 2020)。在西南干旱河谷地区, 虽然降雨量随着纬度增加而呈减少趋势, 但是植被生物量尤其是灌木生物量呈增加趋势, 因此群落生物量以及草本生物量与MAP存在显著负相关关系(表3), 该结果不支持Ma等(2008)的发现: 温带草地生态系统降水变化驱动植被生物量变化。这与干旱河谷地区自然环境的独特性有关。我们认为, 在位于干旱河谷低纬度的元江、金沙江等流域的干热河谷地区, 尽管年降水量较大, 但是旱季更长, 温度也更高, 干热环境对植物生长胁迫更加突出, 尤其限制灌木发育, 导致植被生物量偏低; 随着纬度升高, 如在岷江和大渡河流域, 干热环境胁迫有所缓解, 利于灌木发育, 从而促进植被生物量增加。虽然干旱胁迫是影响该地区植被发育的主要限制因子, 但是纬度梯度上植被生物量与年降水量之间不能完全正向匹配, 需要进一步细致分析蒸发量和土壤水分的变化特点及其植被生物量空间格局的影响因子去证实此推论。
环境温度与群落生物量变化之间关联性比较复杂, 当前不同研究涉及的生态系统类型和调查取样尺度差异导致所获得的结果不一致。Gruner等(2017)对全球森林、草原等生态系统的meta分析结果显示, 温度升高降低了物种丰富度, 从而降低了群落生物量。在西南干旱河谷纬度梯度上, 我们也发现灌木生物量变化与MAT呈显著负相关关系(图4; 表3), 表明在干旱河谷高纬度地区温度相对降低, 促进灌木发育而提高灌木生物量。纬度梯度上灌木物种丰富度及多样性指数与其生物量之间的正相关关系支持此结论(图4; 表3)。Zhang等(2008)发现, 在全球尺度上随纬度升高, 凋落物分解速率下降导致其生物量增加, 因此MAT与凋落物分解速率呈正相关关系, 而与凋落物生物量呈负相关关系, 这与本研究结果(图2D)不同。“碳质量-温度假说”为凋落物生物量与MAT之间的正相关关系提供了机理性解释, 其预测凋落物分解的温度敏感性与凋落物的有机碳质量呈负相关关系, 表明温度越高, 凋落物生物量越大(Bosatta & Ågren, 1999; Fierer et al., 2005)。温度变化通过调控根系呼吸和微生物的分解过程, 进而影响群落及凋落物生物量(Jassal et al., 2008; Wang et al., 2014)。理论上, 土壤养分空间变化是影响植物生长和分布的重要因子。研究已经发现, 土壤养分和植被根系生物量呈显著负相关关系(刘顺等, 2018; 唐立涛等, 2019)。然而, 在干旱河谷(表3)和冀西北(杜美琪等, 2020)的灌丛生态系统中, 土壤养分与群落及组分生物量空间变化并无显著相关性, 仅土壤pH与群落生物量存在显著负相关关系(表3)。Palpurina等(2017)的研究结果也显示, 土壤pH越高, 群落物种丰富度越低, 生物量减少。综合分析表明, 干旱河谷地区气候因子变化是自然植物群落生物量纬度格局形成的主要外在因素。
由此可见, 西南干旱河谷植被生物量在纬度梯度上具有明确的变化规律, 群落生物量及灌木生物量随纬度的升高而显著升高, 草本生物量随纬度的升高无明显变化, 凋落物生物量随纬度的升高而显著下降; 群落各组分生物量所占比例随纬度的变化与对应的组分生物量表现出相同的变化趋势。研究证明西南干旱河谷植被生物量随着纬度升高而增加主要体现在灌木生物量显著增大。群落组成和结构变化是物种及其群落与气候等环境因子相互作用的结果。但是, 由于高山峡谷地形和气候存在多样性和复杂性, 使得气候因子和土壤因子对纬度梯度上植被生物量变化的解释较低, MAP、MAT和土壤pH与群落生物量变化有一定的关系, 但解释率都相对较低。
参考文献
Primary production and rain use efficiency across a precipitation gradient on the Mongolia Plateau
DOI:10.1890/07-0992.1 URL [本文引用: 1]
Quantifying the evidence for biodiversity effects on ecosystem functioning and services
Concern is growing about the consequences of biodiversity loss for ecosystem functioning, for the provision of ecosystem services, and for human well being. Experimental evidence for a relationship between biodiversity and ecosystem process rates is compelling, but the issue remains contentious. Here, we present the first rigorous quantitative assessment of this relationship through meta-analysis of experimental work spanning 50 years to June 2004. We analysed 446 measures of biodiversity effects (252 in grasslands), 319 of which involved primary producer manipulations or measurements. Our analyses show that: biodiversity effects are weaker if biodiversity manipulations are less well controlled; effects of biodiversity change on processes are weaker at the ecosystem compared with the community level and are negative at the population level; productivity-related effects decline with increasing number of trophic links between those elements manipulated and those measured; biodiversity effects on stability measures ('insurance' effects) are not stronger than biodiversity effects on performance measures. For those ecosystem services which could be assessed here, there is clear evidence that biodiversity has positive effects on most. Whilst such patterns should be further confirmed, a precautionary approach to biodiversity management would seem prudent in the meantime.
Soil organic matter quality interpreted thermodynamically
DOI:10.1016/S0038-0717(99)00105-4 URL [本文引用: 1]
Research on the correlations of shrub biomass with slope-aspect and altitude in dry valley of the upper reach of the Minjiang River
岷江上游干旱河谷灌丛生物量与坡向及海拔梯度相关性研究
Facilitation of Quercus ilex in Mediterranean shrubland is explained by both direct and indirect interactions mediated by herbs
DOI:10.1111/j.1365-2745.2010.01655.x URL [本文引用: 2]
Woody plant phenology in the West Africa savanna
DOI:10.1046/j.1365-2699.1998.00229.x URL [本文引用: 1]
Distribution of natural grassland biomass and its relationship with influencing factors in Shaanxi
陕西省天然草地生物量空间分布格局及其影响因素
Distribution of shrub-herb community biomass and its relationship with soil factors in middle and low mountainous areas of northwest Hebei Province
冀西北中低山区灌草群落生物量分配及其与土壤因子的关系
Effects of temperature and soil texture on ANPP in the U.S. Great Plains
DOI:10.1890/0012-9658(1997)078[2628:EOTAST]2.0.CO;2 URL [本文引用: 1]
Distribution scope and district statistical analysis of dry valleys in southwest China
西南地区干旱河谷分布范围及分区统计分析
Why are East Asian ecosystems important for carbon cycle research
DOI:10.1360/zc2010-40-7-561 URL [本文引用: 1]
碳循环研究: 东亚生态系统为什么重要
Methods and protocols for plant community inventory
DOI:10.3724/SP.J.1003.2009.09253 URL [本文引用: 2]
植物群落清查的主要内容、方法和技术规范
DOI:10.3724/SP.J.1003.2009.09253
[本文引用: 2]

植物群落是不同植物在长期环境变化中相互作用、相互适应而形成的组合。它提供着人类赖以生存的主要物质资源, 维系着地球生态系统的健康和功能, 也为各种动物和其他生物提供食物来源和栖息地, 是人类生存和发展不可或缺的物质基础, 具有不可替代的作用。我国植物群落类型多样, 在世界上首屈一指, 但我国至今尚没有一次全面和系统的植物群落清查, 不仅影响了人们对我国植物资源的了解、利用和保护, 也不利于我国生态学、环境科学和地理学等相关学科的发展。采用统一的方法体系和技术规范开展我国植物群落的清查工作势在必行, 并具有紧迫性。本文基于作者长期的野外工作实践和国内外的群落调查方法, 首先简要定义了与植物群落清查有关的重要概念, 在此基础上, 论述了调查样地的设置原则和体系、群落清查的技术指标和方法、主要优势种生态属性的测定方法和规范, 并介绍了大样地调查的主要步骤。通过本文的介绍、归纳和总结, 试图为制定我国植物群落清查的技术规范提供基础材料和技术储备。
WorldClim 2: new 1-km spatial resolution climate surfaces for global land areas
DOI:10.1002/joc.5086 URL [本文引用: 1]
Litter quality and the temperature sensitivity of decomposition
DOI:10.1890/04-1254 URL [本文引用: 1]
Variation in fine root biomass along a 1000 km long latitudinal climatic gradient in mixed boreal forests of North-East Europe
DOI:10.1016/j.foreco.2018.09.060 URL [本文引用: 1]
Effects of experimental warming on biodiversity depend on ecosystem type and local species composition
DOI:10.1111/oik.03688 URL [本文引用: 1]
Increasing water availability and facilitation weaken biodiversity-biomass relationships in shrublands
DOI:10.1002/ecy.2624 [本文引用: 1]
Grassland biomass of communities along gradients of the Inner Mongolia grassland transect
内蒙古草地样带植物群落生物量的梯度研究
DOI:10.17521/cjpe.2006.0073
[本文引用: 1]
采用样带法对内蒙古草地植物群落生物量沿水热梯度的变化特征进行了研究,并对几种回归方法进行了比较。一元回归结果表明:在本样带的限定范围内,生物量与年均温、≥ 0 ℃年积温、≥10 ℃年积温、年实际日照总时数等热量因子呈负相关(年均温的相关性最高),而与年降水量、年均相对湿度等水分因子呈正相关(年均相对湿度的相关性最高),其中年均温和年均相对湿度对生物量的影响最为显著,二者对生物量的空间变异起着互为消长的作用,而海拔高度的影响则不显著。多元回归结果表明,作为半干旱区植物生长的主要限制因子,年降水量在大尺度上对生物量产生影响的途径更为复杂,但其作用不可低估。生物量和地下地上生物量比值会因不同的气候区、不同的植被类型和物种组成,对环境因子的响应程度不同,在经向、纬向和草地类型梯度上的变化特征也不同。水热的配比关系要比单一的水分和温度与植物的生长具有更紧密的关系,地下地上生物量比随着水热配比关系的变化也会呈现出不同的变化规律,其驱动因子并不一定始终都是降水。也许可以认为:半干旱/干旱的划分界限是本研究所涉及的草地样带上生物量和地下地上生物量比值沿草地类型梯度变化的一个转折界限,在此界限前后,气候对生物量的主导因素和生物量对此关键因子的响应程度都有所变化。
A global synthesis reveals biodiversity loss as a major driver of ecosystem change
DOI:10.1038/nature11118 URL [本文引用: 1]
Effects of microhabitat changes on seedling establishment of native plants in a dry valley
DOI:10.17521/cjpe.2020.0216 URL [本文引用: 2]
干旱河谷微生境变化对乡土植物幼苗定植的影响
Effect of soil water stress on soil respiration and its temperature sensitivity in an 18-year-old temperate Douglas- fir stand
DOI:10.1111/j.1365-2486.2008.01573.x URL [本文引用: 1]
The general situation of natural vegetation in dry-hot river valley of Yuanmou, Yunnan Province
云南元谋干热河谷植被概况
Effects of environmental stress on seedlings root growth and nodulation of leguminous shrubs in the dry valley of Minjiang River
环境胁迫对岷江干旱河谷豆科灌木幼苗根系生长及其结瘤的影响
Latitude gradient changes on herbaceous biomass and leaf N and P stoichiometry characteristics in Loess Plateau
黄土高原纬度梯度下草本植物生物量的变化及其氮、磷化学计量学特征
Aboveground biomass of main shrubs in dry valley of Minjiang River
岷江干旱河谷主要灌丛类型地上生物量研究
Vegetational types of the arid valleys in the Hengduan mountainous region
横断山区干旱河谷植被类型
Fine root biomass, productivity and turnover of Abies faxoniana primary forest in subalpine region of western Sichuan, China
川西亚高山岷江冷杉原始林细根生物量、生产力和周转
Quantitative classification and ordination for plant communities in dry valleys of southwest China
DOI:10.17520/biods.2015241 URL [本文引用: 1]
中国西南干旱河谷植物群落的数量分类和排序分析
Quantitative classification and environmental interpretations for the structural differentiation of the plant communities in the dry valley of Jinshajiang River
DOI:10.17520/biods.2015353 URL [本文引用: 1]
金沙江干旱河谷植物群落的数量分类及其结构分异的环境解释
Progress on the research of shrub biomass estimation
灌木生物量测定的研究进展
Above- and below-ground biomass relationships in Chinaʼs grassland vegetation
中国草地植被地上和地下生物量的关系分析
Environmental factors covary with plant diversity-productivity relationships among Chinese grassland sites
DOI:10.1111/j.1466-8238.2009.00508.x URL [本文引用: 1]
Above- and belowground biomass in relation to environmental factors in temperate grasslands, Inner Mongolia
Positive, negative, and net effects in grass-shrub interactions in Mediterranean semiarid grasslands
DOI:10.1890/02-0635 URL [本文引用: 1]
Patterns of biomass partitioning across alpine shrubs in the Three-river Source Region
三江源地区高寒灌丛生物量空间分布格局
The relationship between plant species richness and soil pH vanishes with increasing aridity across Eurasian dry grasslands
DOI:10.1111/geb.12549 URL [本文引用: 1]
Plant root-shoot biomass allocation over diverse biomes: a global synthesis
DOI:10.1016/j.gecco.2019 URL [本文引用: 1]
Primary production of the central grassland region of the United States
DOI:10.2307/1943158 URL [本文引用: 1]
Spatial distribution of aboveground carbon stock of the arboreal vegetation in Brazilian biomes of Savanna, Atlantic Forest and Semi-arid woodland
DOI:10.1371/journal.pone.0128781 [本文引用: 2]
Plant diversity in the dry valleys of Southwest China: spatial deviation and determinants for flora and plant communities
DOI:10.17520/biods.2016049 URL [本文引用: 1]
中国西南干旱河谷的植物多样性: 区系和群落结构的空间分异与成因
DOI:10.17520/biods.2016049 [本文引用: 1]
Protection and utilization of plant biodiversity resources in dry valleys of Southwest China
DOI:10.17520/biods.2016022 URL [本文引用: 1]
西南干旱河谷植物多样性资源的保护与利用
DOI:10.17520/biods.2016022
[本文引用: 1]
西南干旱河谷是分布于我国亚热带横断山区的隐域性河谷景观, 其环境受季风气候、岭-谷地形环境和人类开发活动的共同影响。适应于季节性干旱和垂直气候分异, 干旱河谷具有独特的植被和丰富的物种多样性资源。在过去几十年中, 干旱河谷的气候变化以增温为一致特征, 同时经历着广泛的人类活动干扰, 特别是道路建设、水利工程、矿山开发和城镇化驱动的土地利用变化, 造成了普遍的植被破坏、生境退化和生物入侵, 给乡土植物多样性资源和生态系统稳定性带来了严重威胁, 然而至今对干旱河谷生物多样性的保护仍基本属于空白。因此, 建议及时建立干旱河谷的生物多样性保护体系; 因地制宜采取退化植被修复、立体农业规划、特色植物资源利用和风景旅游开发等措施, 以维持干旱河谷生态系统的可持续性; 落实生态文明建设的相关政策, 以保障西南生态屏障的生态安全和生态服务能力。
Forest biomass and its influencing factors in Qinghai Province
青海省森林细根生物量及其影响因子
Soil moisture modifies the response of soil respiration to temperature in a desert shrub ecosystem
DOI:10.5194/bg-11-259-2014 URL [本文引用: 1]
Carbon stock of Moso bamboo (Phyllostachys pubescens) forests along a latitude gradient in the subtropical region of China
DOI:10.1371/journal.pone.0193024 [本文引用: 1]
Rates of litter decomposition in terrestrial ecosystems: global patterns and controlling factors
DOI:10.1093/jpe/rtn002 URL [本文引用: 1]
Biogeographical patterns of biomass allocation in leaves, stems, and roots in China’s forests
DOI:10.1038/srep15997
PMID:26525117
[本文引用: 2]
To test whether there are general patterns in biomass partitioning in relation to environmental variation when stand biomass is considered, we investigated biomass allocation in leaves, stems, and roots in China's forests using both the national forest inventory data (2004-2008) and our field measurements (2011-2012). Distribution patterns of leaf, stem, and root biomass showed significantly different trends according to latitude, longitude, and altitude, and were positively and significantly correlated with stand age and mean annual precipitation. Trade-offs among leaves, stems, and roots varied with forest type and origin and were mainly explained by stand biomass. Based on the constraints of stand biomass, biomass allocation was also influenced by forest type, origin, stand age, stand density, mean annual temperature, precipitation, and maximum temperature in the growing season. Therefore, after stand biomass was accounted for, the residual variation in biomass allocation could be partially explained by stand characteristics and environmental factors, which may aid in quantifying carbon cycling in forest ecosystems and assessing the impacts of climate change on forest carbon dynamics in China.
Temporal and spatial distribution of aboveground biomass of vegetation and quantitative analysis of impact factors in semi-arid grassland basin
半干旱草原型流域植被地上生物量时空分布特征及其影响因子
Positive species diversity and above-ground biomass relationships are ubiquitous across forest strata despite interference from overstorey trees
DOI:10.1111/1365-2435.12699 URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}