




种群结构是种群生态学研究的核心内容(江洪, 1992), 反映了种群内不同大小个体数量的分布和配置情况, 也反映了种群数量动态及其发展趋势(刘普幸, 2011)。年龄结构、静态生命表和存活曲线是进行种群统计和种群数量动态研究的重要工具(张志祥等, 2008), 可以反映种群目前的存活状态和植物与环境之间的适合度, 结合种群动态量化分析和时间序列预测还能够阐明种群的受干扰状况及未来的发展趋势(Armesto et al., 1992; Fuchs et al., 2000; 韩路等, 2014; 赵阳等, 2020)。因此, 研究种群结构及其动态对正确描述并判定种群动态变化规律, 揭示种群与环境的关系及其在群落中的地位和作用, 预测其演化趋势对种群及群落的更新和生态系统的恢复具有重要意义(王进等, 2020)。
青藏高原东北缘的祁连山区地处青藏、蒙新、黄土三大高原交汇地带, 是黑河、石羊河和疏勒河三大内陆河及黄河流域的主要水源涵养地和集水区, 是中国生物多样性保护的优先区域, 在维护中国西部生态安全方面有着举足轻重和不可替代的地位, 是西北地区重要的生态安全屏障(Gao et al., 2013; Zhu et al., 2017)。但同时, 该区作为对气候变化最为敏感的地区之一, 生态环境尤为脆弱, 因此研究该区物种的生长状态和适应规律对青藏高原气候和生态的变化有着高度的指示作用, 对深入研究和预测全球生态系统的动态变化具有深远的意义。近年来, 因受气候变化和人为干扰双重影响, 该区域森林退化面积逐年增大, 水源涵养能力不断下降。针对祁连山区水源涵养林的退化, 进行该区域天然林的保护是促进祁连山区生态恢复, 快速提高水源涵养区生态系统稳定性及可持续发展的有效手段。
西藏沙棘(Hippophae tibetana)是青藏高原高海拔地区特有的低矮灌木(拉琼等, 2010), 雌雄异株, 高4-60 cm, 根系发达, 具有很强的克隆繁殖能力(陈学林和廉永善, 2007), 能够在自然条件严酷的高寒区形成大面积密集的单优植物群落, 在改善自然景观和生态环境, 为鸟兽提供栖息和生存地及水土保持方面有重要作用(拉琼等, 2009; 温江波等, 2010)。西藏沙棘作为祁连山高寒区低矮灌丛的主要建群种, 多以斑块状分布于海拔2 700-3 300 m的山坡退化草地, 是祁连山区群落演替的先锋物种, 对其种群结构和动态的研究有助于理解西藏沙棘适应高寒生境的策略及其在高寒地区群落稳定性中的作用。海拔等生态因子是影响西藏沙棘生长适应性的最显著的因子之一(拉琼等, 2010; 贺晓慧等, 2021), 气候变化将对西藏沙棘的海拔分布产生显著影响, 但目前对西藏沙棘种群结构和数量动态随海拔梯度有何响应至今未见报道。鉴于此, 本研究通过对青藏高原东北缘祁连山区天祝藏族自治县不同海拔西藏沙棘自然种群的野外调查, 来了解不同海拔西藏沙棘种群结构和数量动态特征的差异, 掌握西藏沙棘在复杂气候及环境条件下的生长规律及其适宜生境的潜在分布, 以期为该区西藏沙棘自然种群的保护提供重要的理论依据, 并为预测未来气候变化条件下高寒植被的命运提供可靠数据支撑。
1 材料和方法
1.1 研究区概况
研究区位于甘肃省武威市天祝藏族自治县, 地处青藏高原东北缘祁连山区, 地理跨度103.7°- 104.7° E, 36.6°-37.6° N, 海拔高度2 040-4 873 m, 属大陆性高原季风气候, 年平均气温-0.2-1.3 ℃, ≥0 ℃积温1 327.7-1 738.2 ℃, 降水量266- 630 mm, 无四季之分, 仅分冷暖两季, 冷季长达7个月。土壤以山地黑钙土、栗钙土、亚高山草甸土、高山草甸土等为主。西藏沙棘在该研究区的分布海拔范围为2 700-3 300 m, 在以西藏沙棘为优势种的群落中, 主要植物为金露梅(Potentilla fruticosa)、中国黄花柳(Salix sinica)等灌木和珠芽蓼(Polygonum viviparum)、赖草(Leymus secalinus)、黄囊薹草(Carex korshinskyi)、黄花棘豆(Oxytropis ochrocephala)、老芒麦(Elymus sibiricus)等草本。
1.2 研究方法
1.2.1 样地设置与调查
在对研究区多次踏查的基础上, 在西藏沙棘分布相对集中的天祝藏族自治县抓喜秀龙镇沿海拔高度在抓喜秀龙景区(2 868 m)、红疙瘩村(3 012 m)、白水口村(3 244 m)设置样地。2020年7-8月将西藏沙棘分布的样地划分为低海拔、中海拔和高海拔3个生境, 在每个生境选择西藏沙棘分布相对集中、林分良好且无人为干扰的地段设置5 m × 5 m的样方各6个, 每个样方间距100 m以上。西藏沙棘具有强的萌蘖克隆能力, 可通过水平根萌蘖产生幼苗, 调查时难以判定基株, 因此, 实际调查时仅以地上部分为参考划分植株个体, 利用游标卡尺和钢卷尺对样方内西藏沙棘进行每木检尺, 记录每株西藏沙棘的基径、株高及冠幅等。同时, 测定样地的经度、纬度、海拔, 并记录灌木层和草本层的物种组成、高度和盖度。
1.2.2 种群龄级划分
由于植物的大小和年龄密切相关, 同一生境下相同物种的生长状况和年龄对环境具有相似的响应规律(Su et al., 2021), 因此利用植株大小代替年龄是种群年龄结构研究中常用的方法。刘明等(2009)用中国沙棘(Hippophae rhamnoides subsp. sinensis)的株高来代替年龄, 与之不同的是, 西藏沙棘植株矮化(拉琼等, 2010), 其基径与年龄的关系更为密切。对此, 本研究采用基径代替年龄的方法来研究西藏沙棘种群的年龄结构。本研究中西藏沙棘雌株最小基径约0.3 cm, 结合西藏沙棘的基径大小分布, 将基径(BD)以0.3 cm为一个等级对其进行划分, 可划分为: I (≤0.3 cm), II (0.3-0.6 cm), III (0.6- 0.9 cm), IV (0.9-1.2 cm), V (1.2-1.5 cm), VI (1.5- 1.8 cm), VII (>1.8 cm)共7个龄级。根据西藏沙棘的生长特性及调查到的基径大小分布特征将以上龄级划分为3个生长期: 幼龄期(I), 中龄期(II-III)和老龄期(IV-VII)(刘明等, 2009; 康佳鹏等, 2019; 赵家豪等, 2022)。调查各海拔全部样方内各龄级西藏沙棘的植株数, 并以不同龄级及各龄级植株数占各种群植株总数的百分比绘制不同海拔西藏沙棘种群的年龄结构图。
1.2.3 静态生命表编制
静态生命表包含的参数及关系如下: x为龄级; ax为x龄级的实际存活数量, a0为ax的初始值; lx为x龄级的标准化存活数, 以1 000为基数, lx = ax/a0 × 1000; dx表示从x龄级到x + 1龄级期间标准化死亡数, dx = lx - lx+1; qx为从x龄级到x + 1龄级期间死亡率, qx = dx/lx; Lx表示从x龄级到x + 1龄级间隔期的平均存活个体数, Lx = (lx + lx+1)/2; Tx表示从x龄级到超过x龄级的个体总数, Tx = ΣLx; ex表示x龄级的生命期望值, ex = Tx/lx; Kx为消失率, Kx = lnlx - lnlx+1。
以不同龄级为横坐标、不同海拔种群各龄级的标准存活数对数(lnlx)为纵坐标, 绘制种群存活曲线。为了检验西藏沙棘种群存活曲线是符合Deevey-II型还是Deevey-III型, 本研究采用Hett和Loucks (1976)提出的数学模型对种群存活曲线进行检验, 依据决定系数、F检验值来判定模型的拟合效果, 如果指数方程Nx = N0e-bx拟合效果好, 则存活曲线为Deevey-II型; 如果幂函数Nx = N0x-b拟合效果好, 存活曲线则为Deevey-III型。其中, 式中Nx、N0分别代表x龄级内存活数和种群形成初期的个体数, b为死亡率(Hett & Loucks, 1976)。
1.2.4 种群动态量化方法
式中, Sx与Sx+1分别为第x与第x + 1龄级的个体数, k表示种群龄级数量, Vx∈[-1, 1], Vx取正、负、0时分别反映种群相邻龄级个体数增长、衰退、稳定的结构动态关系。因种群的年龄结构在相邻龄级之间是以个体数量进行比较的, 所以Vpi可由Vx对Sx进行加权分配而得, Vpi仅适用于不考虑未来外部环境干扰的种群结构动态。当考虑未来外部干扰时, V′pi还与k及Sx有关, k与Sx对未来的外部干扰存在着“稀释效应”, 据此种群承担的风险概率(P)由k与Sx根据条件概率法则计算可得:$P=\frac{1}{k\times {S_{x}}}$, x = 1, 2, 3,…, k, 只有当P最大时才对Vpi构成最大影响, 将最大的P代入后, Vpi计算公式修正为:
式中, V′pi也可作为衡量种群结构动态对随机干扰的敏感性指标。Vpi、V′pi取正、负、0时反映的动态关系与Vx一致。
1.2.5 时间序列预测模型
采用时间序列分析中的一次平均推移法(谢衷洁, 1990)对西藏沙棘种群未来2、4、6个龄级的时间动态进行预测, 模型为:
式中, t为需要预测的未来时间年限; x为龄级;$M_{t}^{(1)}$表示种群生长到未来t时间(龄级)时, 第x龄级的种群存活数, 是t个观测值在x时刻的平均值, 称为第t周期的移动平均; Xk为当前k龄级种群存活数。
1.2.6 数据处理
采用SPSS 20.0软件对不同海拔间种群个体形态特征的差异显著性(p < 0.05)进行单因素方差分析, 并进行存活曲线的拟合, 利用Origin 2018和Excel 2016绘图。
2 结果和分析
2.1 种群基本特征
不同海拔西藏沙棘种群的株高、基径和冠幅均存在显著差异(图1), 西藏沙棘的株高、基径和冠幅均随海拔的升高而逐渐降低。
图1
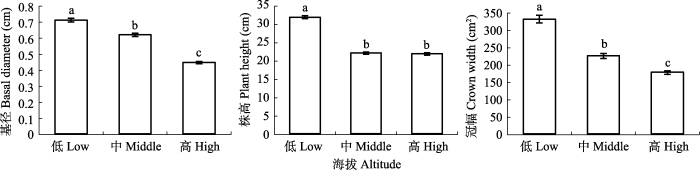
图1
祁连山区不同海拔西藏沙棘种群形态特征(平均值±标准误)。不同小写字母代表差异显著(p < 0.05)。
Fig. 1
Morphological characteristics of Hippophae tibetana populations at different altitudes in the Qilian Mountains (mean ± SE). Different lowercase letters indicate significant difference (p < 0.05).
2.2 种群年龄结构
中海拔幼、中和老龄期个体占种群的比例依次为18.04%、65.67%和16.29%; 低海拔种群幼龄期个体比例最低(2.02%), 老龄期次之(22.55%), 中龄期最高(75.43%); 高海拔种群老龄期比例最低(1.15%), 幼龄期次之(22.38%), 中龄期占比最高(76.47%)。总体来看, 低、中、高3个海拔西藏沙棘种群的年龄结构相似, 均呈中龄期比例最高, 幼龄期和老龄期比例次之的纺锤型结构(图2)。
图2
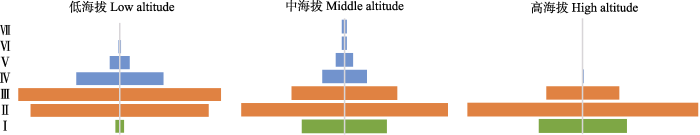
图2
祁连山区不同海拔西藏沙棘种群年龄结构。I, BD ≤ 0.3 cm; II, BD 0.3-0.6 cm; III, BD 0.6-0.9 cm; IV, BD 0.9-1.2 cm; V, BD 1.2-1.5 cm; VI, BD 1.5-1.8 cm; VII, BD > 1.8 cm; BD, 基径。
Fig. 2
Age structure of Hippophae tibetana populations at different altitudes in the Qilian Mountains. I, BD ≤ 0.3 cm; II, BD 0.3-0.6 cm; III, BD 0.6-0.9 cm; IV, BD 0.9-1.2 cm; V, BD 1.2-1.5 cm; VI, BD 1.5-1.8 cm; VII, BD > 1.8 cm; BD, basal diameter.
2.3 生命表、存活曲线和死亡率曲线
2.3.1 静态生命表
低、中、高3个海拔种群不同龄级植株存活数不同, 但均表现为随龄级的增长先上升后下降的趋势, 低海拔种群III龄级植株数最多, 有258株, 占种群总数的40.12%, 中、高海拔种群在II龄级存活数达到最大, 分别为522和404株, 占各种群的43.39%、57.96% (表1)。高海拔III龄级植株129株, 占种群总数的比例自II龄级的57.96%降低到18.51%, 表明高海拔种群在II龄级生长过程中有大量植株死亡。3个海拔种群中均有负的死亡率的存在, 表明3个海拔种群均有干扰存在。各海拔种群幼龄期死亡率均为负值, 种群均缺乏幼苗, 且3个海拔种群各龄级死亡率均较高(>0.5), 预测3个海拔种群未来均将趋于不同程度的衰退; 死亡率呈现高海拔>低海拔>中海拔的趋势(表1)。
表1 祁连山区不同海拔西藏沙棘种群的静态生命表
Table 1
| 海拔 Altitude | x | ax | lx | dx | qx | Lx | Tx | ex | lnlx | Kx |
|---|---|---|---|---|---|---|---|---|---|---|
| 低 Low | I | 13 | 1 000 | -16 462 | -16.46 | 9 231 | 48 923 | 48.92 | 6.91 | -2.86 |
| II | 227 | 17 462 | -2 385 | -0.14 | 18 654 | 39 692 | 2.27 | 9.77 | -0.13 | |
| III | 258 | 19 846 | 11 231 | 0.57 | 14 231 | 21 038 | 1.06 | 9.90 | 0.83 | |
| IV | 112 | 8 615 | 6 538 | 0.76 | 5 346 | 6 808 | 0.79 | 9.06 | 1.42 | |
| V | 27 | 2 077 | 1 692 | 0.81 | 1 231 | 1 462 | 0.70 | 7.64 | 1.69 | |
| VI | 5 | 385 | 308 | 0.80 | 231 | 231 | 0.60 | 5.95 | 1.61 | |
| VII | 1 | 77 | - | - | - | - | - | 4.34 | - | |
| 中 Middle | I | 217 | 1 000 | -1 406 | -1.41 | 1 703 | 5 005 | 5.00 | 6.91 | -0.88 |
| II | 522 | 2 406 | 1 171 | 0.49 | 1 820 | 3 302 | 1.37 | 7.79 | 0.67 | |
| III | 268 | 1 235 | 700 | 0.57 | 885 | 1 482 | 1.20 | 7.12 | 0.84 | |
| IV | 116 | 535 | 318 | 0.59 | 376 | 597 | 1.12 | 6.28 | 0.90 | |
| V | 47 | 217 | 143 | 0.66 | 145 | 221 | 1.02 | 5.38 | 1.08 | |
| VI | 16 | 74 | -5 | -0.06 | 76 | 76 | 1.03 | 4.30 | -0.06 | |
| VII | 17 | 78 | - | - | - | - | - | 4.36 | - | |
| 高 High | I | 156 | 1 000 | -1 590 | -1.59 | 1 795 | 3 967 | 3.97 | 6.91 | -0.95 |
| II | 404 | 2 590 | 1 763 | 0.68 | 1 708 | 2 172 | 0.84 | 7.86 | 1.14 | |
| III | 129 | 827 | 788 | 0.95 | 433 | 464 | 0.56 | 6.72 | 3.07 | |
| IV | 6 | 38 | 32 | 0.83 | 22 | 31 | 0.81 | 3.65 | 1.79 | |
| V | 1 | 6 | 0 | 0.00 | 6 | 9 | 1.40 | 1.86 | 0.00 | |
| VI | 1 | 6 | 6 | 1.00 | 3 | 3 | 0.47 | 1.86 | - | |
| VII | 0 | 0 | - | - | - | - | - | - | - |
ax, 实际存活数; dx, 标准化死亡数; ex, 生命期望值; Kx, 消失率; lx, 标准化存活数; Lx, x到x + 1龄级间隔期平均存活的个体数; qx, 死亡率; Tx, x到超过x龄级的个体总数; x, 龄级, 同
ax, actual survival number; dx, standardized deaths; ex, life expectancy value; Kx, disappearance rate; lx, standardized survival number; Lx, average number of survived individuals during the interval from x to x + 1 age class; qx, mortality rate; Tx, the total number of individuals from x to over x age class; x, age class, see
2.3.2 生存现状
种群存活曲线可分为3种基本类型(Deevey, 1947): I型是凸型曲线, 属于该型的种群绝大多数个体均能实现其平均生理寿命, 幼体死亡率较低, 但当活到平均生理年龄时, 短期内全部死亡; II型是直线型, 属于该型的种群各年龄死亡率基本相同; III型是凹型曲线, 属于该型的种群幼体死亡率高, 一旦存活到一定年龄, 死亡率低而稳定, 只有极少数个体能活到生理寿命(牛翠娟等, 2015)。从西藏沙棘的存活曲线(图3)可以看出, 3个海拔种群的存活能力表现为低海拔>中海拔>高海拔。生命期望(ex)是对x龄级内植株个体平均生存能力的反映, 种群生命期望值最大时, 表明种群对环境适应能力最强, 生理活动达到旺盛期(金慧等, 2017)。各海拔种群的Ι龄级个体数均较少, ex的峰值均出现在I龄级, 随龄级的增加ex降低(图3), 中龄期个体均充足, ex自幼龄期到中龄期早期呈急剧下降趋势, 从中龄期到老龄期则趋于稳定; 西藏沙棘中、幼龄期ex呈低海拔>中海拔>高海拔的趋势, 老龄期ex则呈中海拔>低海拔>高海拔(图3)。
图3
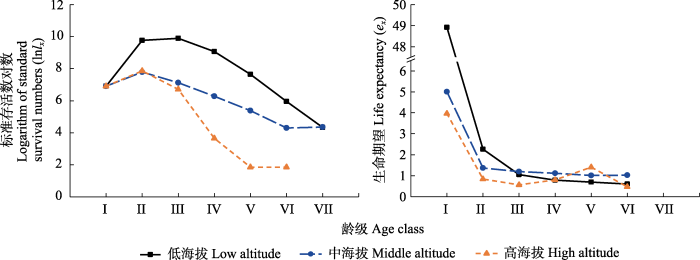
图3
祁连山区不同海拔西藏沙棘种群的存活曲线和生命期望曲线。龄级同
Fig. 3
Survivorship and life expectancy curves of Hippophae tibetana populations at different altitudes in the Qilian Mountains. See
以龄级为横坐标, 各龄级个体的死亡率(qx)及消失率(Kx)为纵坐标做死亡率和消失率曲线(图4), 3个海拔西藏沙棘的死亡率与消失率曲线变化趋势分别一致, 均发生一定的波动。低、中海拔种群的死亡率在V龄级达到峰值, 但高海拔种群III龄级死亡率高达0.95, 表明高海拔种群III龄级个体遇到了环境筛, 导致其种群生长受到的阻碍增大。高海拔种群的消失率曲线较陡峭, 也表明高海拔种群受到的环境阻力最大。
图4
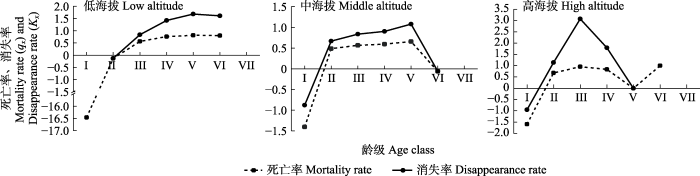
图4
祁连山区不同海拔西藏沙棘种群的死亡率和消失率曲线。龄级同
Fig. 4
Mortality and disappearance curves of Hippophae tibetana populations at different altitudes in the Qilian Mountains. See
2.3.3 存活曲线拟合
表2 祁连山区不同海拔西藏沙棘种群存活曲线拟合方程
Table 2
| 海拔 Altitude | 方程 Equation | R2 | F | p |
|---|---|---|---|---|
| 低 Low | y = 13.712e-0.125x | 0.807 | 17.724 | 0.024 |
| y = 14.454x-0.420 | 0.636 | 7.986 | 0.066 | |
| 中 Middle | y = 10.867e-0.147x | 0.961 | 99.555 | 0.002 |
| y = 11.881x-0.514 | 0.861 | 25.819 | 0.015 | |
| 高 High | y = 19.451e-0.417x | 0.905 | 39.069 | 0.008 |
| y = 26.450x-1.501 | 0.874 | 28.852 | 0.013 |
2.4 种群结构动态量化分析与时间序列预测
2.4.1 种群动态量化分析
种群数量动态是种群个体生存能力与环境相互作用的结果。3个海拔西藏沙棘种群相邻龄级间个体数量动态变化指数(Vx)均有波动: 3个海拔种群的VI均小于0, VIII-VV均大于0, 中海拔的VVI小于0 (表3), 即各海拔种群幼龄期均呈衰退趋势, 中、老龄期基本呈增长趋势, 这与各种群年龄结构相一致(图2)。3个海拔梯度种群总体数量动态变化指数Vpi和V′pi的结果显示, Vpi (高海拔>低海拔>中海拔) > V′pi (高海拔>低海拔>中海拔) > 0, 表明3个海拔种群在有无干扰的影响下均表现为增长趋势, 外部干扰的存在会减缓种群的增长趋势; 高海拔种群的增长潜力最大, 高海拔种群II龄级比例最高, 导致其增长潜力高于环境比其优越的低、中海拔的。但低、中、高3个海拔种群的V′pi分别为0.048、0.003、0.063, 均接近于0, 表明3个海拔种群在干扰影响下缓慢增长, 均逐渐趋于稳定, 这与3个海拔种群的存活曲线拟合结果(表2)相一致, 中海拔种群最稳定; 随机干扰风险概率极大值(Pmax)呈现出高海拔=低海拔>中海拔, 其中, 中海拔的为0.009, 表明各海拔种群对随机干扰均具有一定的敏感性, 但中海拔种群受干扰影响的概率最小, 中海拔种群的稳定性最好。
表3 祁连山区不同海拔西藏沙棘种群年龄结构动态指数
Table 3
| 海拔 Altitude | 动态指数级 Dynamic index level | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| VI | VII | VIII | VIV | VV | VVI | Vpi | V′pi | Pmax | |
| 低 Low | -0.943 | -0.120 | 0.566 | 0.759 | 0.815 | 0.800 | 0.339 | 0.048 | 0.143 |
| 中 Middle | -0.584 | 0.487 | 0.567 | 0.595 | 0.660 | -0.059 | 0.319 | 0.003 | 0.009 |
| 高 High | -0.614 | 0.681 | 0.954 | 0.833 | 0 | 1.000 | 0.442 | 0.063 | 0.143 |
Pmax, 随机干扰风险概率极大值; Vpi, 种群动态变化指数; V′pi, 受外界干扰时种群动态变化指数; VI-VVI, 相邻龄级数量动态变化指数, 龄级同
Pmax, the maximum of probability in random disturbance; Vpi, quantitative dynamics index of population; V′pi, quantitative dynamics index of population by external interference; VI-VVI, quantitative dynamics index of adjacent age class, see
2.4.2 时间序列预测
时间序列分析是研究种群未来年龄结构和发展趋势的理想手段, 可对植物种群动态进行预测(吴承祯和洪伟, 1999)。由表4可知, 在未来2、4、6个龄级时间后, 各种群大龄级个体数不同程度增加。未来2个龄级时间后, 低海拔种群II、III龄级个体数各减少47.14%、5.81%, IV龄级个体数逐渐增长, 中、高海拔种群II龄级个体数各减少29.11%、30.69%, III龄级个体数逐渐增加; 未来4个龄级时间后, 低海拔种群IV龄级个体数减少, V龄级个体数有明显增加的趋势, 中、高海拔种群IV龄级个体数明显增加; 未来6个龄级后, 各海拔VI、VII龄级个体数明显增加。由于各种群均存在相对丰富的中龄期个体, 随着种群的生长, 老龄期个体逐渐得到补充, 若幼苗无法得到及时补充, 各种群将逐渐进入衰退状态。
表4 祁连山区不同海拔西藏沙棘种群数量动态时间序列预测
Table 4
| 龄级 Age class | 低海拔 Low altitude | 中海拔 Middle altitude | 高海拔 High altitude | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M0 | M2(1) | M4(1) | M6(1) | M0 | M2(1) | M4(1) | M6(1) | M0 | M2(1) | M4(1) | M6(1) | |
| I | 13 | 217 | 156 | |||||||||
| II | 227 | 120 | 522 | 370 | 404 | 280 | ||||||
| III | 258 | 243 | 268 | 395 | 129 | 267 | ||||||
| IV | 112 | 185 | 153 | 116 | 192 | 281 | 6 | 68 | 174 | |||
| V | 27 | 70 | 156 | 47 | 82 | 238 | 1 | 4 | 135 | |||
| VI | 5 | 16 | 101 | 107 | 16 | 32 | 112 | 198 | 1 | 1 | 34 | 116 |
| VII | 1 | 3 | 36 | 105 | 17 | 17 | 49 | 164 | 0 | 1 | 2 | 90 |
M0, 实际存活数; M2(1)、M4(1)、M6(1), 未来2、4、6个龄级后各龄级个体数。龄级同
M0, number of original survival; M2(1), M4(1), M6(1), number of every age class after 2, 4, 6 age class time. See
3 讨论
海拔是影响植物种群数量和动态的重要因素(程子敏等, 2018)。本研究中, 西藏沙棘的种群结构及动态在海拔间既有相似性, 又受海拔影响存在不同。一方面, 3个海拔西藏沙棘种群年龄结构存在着相似性。首先, 种群在3个海拔均呈幼龄期和老龄期个体较少、中龄期个体充足的纺锤型年龄结构, 3个海拔种群存活曲线均为II型曲线。其次, 对3个海拔种群动态的研究结果显示, 种群动态指数Vpi和V′pi均大于0, 且趋近于0, 证明了西藏沙棘种群趋于稳定, 这与黄土丘壑区13年林龄的沙棘人工林的年龄结构相似(张文辉等, 2005)。另一方面, 西藏沙棘种群的年龄结构在不同海拔间也存在差异。首先, 低海拔种群III龄级个体占比最高, 而中、高海拔II龄级比例最高; 其次, 西藏沙棘的形态特征(基径、株高、冠幅)也呈低海拔>中海拔>高海拔的趋势。以上表明海拔的升高会阻碍西藏沙棘植株的生长, 究其原因, 可能是随海拔的升高, 低温、低氧、强紫外辐射等胁迫的增强会限制种群的生长(韩发等, 1997), 这与Li等(2007)发现海拔2 800 m以上, 海拔的升高不利于卧龙自然保护区沙棘生长的研究结果一致。另外, 3个海拔种群存活能力呈低海拔>中海拔>高海拔的趋势, 死亡率趋势与之相反, 表现为高海拔>低海拔>中海拔。高海拔种群III龄级遇到环境筛, 死亡率急剧升高, 低、中海拔种群在V龄级死亡率最大, 一方面证实高海拔更为恶劣的环境会导致西藏沙棘种群生长和存活受到阻碍; 同时也表明老龄期西藏沙棘死亡率较高, 存活能力较低, 这主要是西藏沙棘的生理衰老所致(张文辉等, 2005)。
对西藏沙棘生命期望曲线的研究结果表明祁连山区西藏沙棘更适宜在3 000 m左右的中海拔生长。首先, 西藏沙棘老龄期ex表现为中海拔>低海拔>高海拔, 由于ex越高表明种群的适应能力越强, 存活能力较低的老龄期个体在中海拔却表现出最强的适应能力; 其次, 对西藏沙棘种群动态的研究表明, 中海拔种群的随机干扰敏感性指数(Pmax = 0.009)接近于0, 表明中海拔种群的抗干扰能力最强; 再者, 中海拔种群各龄级的死亡率最小, 预测其未来衰退速度最慢。以上皆表明西藏沙棘在3 000 m左右的中海拔具有最优的种群生存能力, 这是西藏沙棘适应高海拔环境的结果, 因此最低分布线的低海拔(2 800 m)并不是其最优的生长环境, 环境随海拔的变化而发生变化, 在高海拔地区, 低温、强风和生长季短等不利于植物的生长, 故中海拔是该研究区西藏沙棘生存最为适宜的海拔高度, 这也是符合“中度干扰假说”: 分布于中间海拔的植物生长优于低或高海拔植物(Kiełtyk, 2018)。另外, 需要强调的是, 3个海拔种群的ex均在I龄级达到峰值, 随龄级的增长, ex降低, 自幼龄期至中龄期, ex急剧下降, 这与西藏沙棘的生物学特征和生活史特征相一致, 西藏沙棘具有较强的克隆生长能力, 可通过水平根的连接进行相邻个体间资源共享来削弱种间竞争的压力, 但随龄级的增加, 个体数量和植株大小亦不断生长, 相邻植株对空间、营养、水分和光照资源的需求相对增大, 使得种群内部发生激烈竞争而产生自疏作用, 致使生命期望值随龄级的增加逐渐降低, 且随着植株的生长, 西藏沙棘逐渐进入生理衰老阶段, ex也随之降低。
西藏沙棘作为青藏高原特有的物种, 是高海拔区水土保持、荒山造林的演替先锋阳性树种(贺晓慧等, 2021), 其种群动态变化遵循阳性树种“增长—稳定—衰退”的种群动态趋势(刘明等, 2009)。我们对西藏沙棘种群结构的研究也证实了这一规律: 目前3个海拔种群II、III龄级植株数较多, 即各种群的中龄期个体充足, 表明3个海拔种群目前可以依靠种群丰富的中龄期个体来维持种群短期的发展, 但由于3个海拔种群幼龄期个体均较少, 且幼龄期种群动态表现为衰退状态(VI均小于0), 表明各种群均存在幼苗补充不及时的风险, 随着各种群的中龄期个体向老龄期个体的生长, 这将促使3个海拔种群走向衰退; 加之3个海拔种群各龄级的死亡率均较高, 其中高海拔种群因面临更恶劣的环境其死亡率最高, 这会导致高海拔种群提前进入衰退期, 衰退速度加快; 同时, 对3个海拔种群的时间预测分析亦表明未来2、4、6个龄级的时间后, 各种群幼龄期个体均逐渐减少, 中、老龄期个体逐渐增多, 种群向老龄期发展并逐渐进入衰退状态, 以上皆表明3个海拔的西藏沙棘种群均处于由稳定型向衰退型发展的阶段。其中, 幼苗更新能力弱, 种内、种间竞争压力造成的种群数量减少及高海拔恶劣的环境造成的高死亡率是导致种群衰退的主要原因。本研究对西藏沙棘种群年龄结构及动态的预测结果与对中国沙棘的研究结果(Li et al., 2007; 刘明等, 2014)一致。
当前, 全球变化正导致许多种群、群落和生态系统地理分布的改变(方精云等, 2018), 贺晓慧等(2021)模拟升温情景的分析表明西藏沙棘对剧烈升温的气候较为敏感, 且随气候的变暖西藏沙棘的适宜生境面积减小。西藏沙棘作为该区生态建设的优良物种, 其种群对该区生态环境的保护及生物多样性的维持具有重要作用, 因此掌握西藏沙棘在复杂气候及生态环境下的生长规律, 不仅可为该区西藏沙棘自然种群的保护提供重要的理论依据, 亦可对预测全球气候变暖下高寒植物海拔分布的变化提供可靠数据支撑。值得一提的是, 西藏沙棘具有较强的克隆繁殖能力, 可以通过克隆生长进行种群的补充, 因此未来还需结合西藏沙棘的克隆生长和克隆繁殖能力对环境因子和气候条件的响应策略来探讨西藏沙棘种群及群落的发展动态及趋势。
参考文献
Age structure and dynamics of Patagonian beech forests in Torres del Paine National Park, Chile
DOI:10.1007/BF00031633 URL [本文引用: 1]
A study on the method of quantitative analysis for plant population and community structural dynamics
植物种群与群落结构动态量化分析方法研究
Distribution pattern and cause of formation of the Genus Hippophae
沙棘属植物的分布格局及其成因
Structure and dynamic of natural Armeniaca sibirica populations at different altitudes in mountain areas of northern Hebei
冀北山地不同海拔天然山杏种群的结构与动态
Life tables for natural populations of animals
DOI:10.1086/395888 URL [本文引用: 1]
The response of ecosystems to global warming
生态系统对全球变暖的响应
Factors affecting emergence and first-year survival of seedlings of Garry oaks (Quercus garryana) in British Columbia, Canada
DOI:10.1016/S0378-1127(99)00329-1 URL [本文引用: 1]
Climate-growth analysis of Qilian juniper across an altitudinal gradient in the central Qilian Mountains, northwest China
DOI:10.1007/s00468-012-0776-6 URL [本文引用: 1]
Contents of protein, fat and starch of Kobresia humilis plants grown at different altitudes in Qinghai-Xizang Plateau
青藏高原不同海拔矮嵩草蛋白质、脂肪和淀粉含量的变异
生长在青藏高原4个不同海拔地区的矮嵩草(Kobresia humilis),其蛋白质、脂肪和碳水化合物的含量有明显差异。其中海拔最高的大坂山(4,000m)植株中蛋白质含量平均比海拔依次较低的金羊岭(3,800m),海北站(3,200m)和西宁地区(2,200m)的分别高出9.9%,17.3%和39.4%;脂肪含量平均分别高出7.1%,77.8%和106.0%;淀粉含量也平均分别高出2.4%,21.6%和32.7%。同时表明,海拔高度对上述若干生化成分含量的季节性动态变化均有明显的影响。从而显示,高海拔地区的植物所以具有较强的抗寒性和适应逆境胁迫的能力,是与体内蛋白质、脂肪和碳水化合物(淀粉)的大量积累增加密切相关。
The population structure and dynamics of Populus euphratica at the upper reaches of the Tarim River
塔里木河上游胡杨种群结构与动态
Age structure models of balsam fir and eastern hemlock
DOI:10.2307/2258822 URL [本文引用: 3]
Potential distribution of Hippophae thibetana and its predicted responses to climate change
西藏沙棘(Hippophae thibetana)潜在地理分布及其对未来气候变化的响应模拟
Quantitative characteristics and dynamic analysis of the endangered species Rhododendron chrysanthum population in Changbai Mountain
长白山濒危植物牛皮杜鹃(Rhododendron chrysanthum)种群数量特征与动态分析
Dynamic changes of spatial pattern and structure of the Tamarix ramosissima population at the desert-oasis ecotone of the Tarim Basin
荒漠绿洲过渡带柽柳种群结构与空间格局动态
Variation of vegetative and floral traits in the alpine plant Solidago minuta: evidence for local optimum along an elevational gradient
DOI:10.1007/s00035-017-0197-7 URL [本文引用: 1]
Habitat types and phenotypic variation of Hippophae tibetana along an altitudinal gradient in the Rongbu Valley of Mt. Everest, Tibet, China
DOI:10.3724/SP.J.1145.2010.00173 URL [本文引用: 3]
珠穆朗玛峰绒布沟西藏沙棘生境类型及海拔梯度下表型变异
A preliminary study on flowering date, flower morphology and flower number of Hippophae tibetana
西藏沙棘(Hippophae tibetana)的花期、花形态及其花朵数量的初步研究
Sex-related differences in leaf morphological and physiological responses in Hippophae rhamnoides along an altitudinal gradient
DOI:10.1093/treephys/27.3.399 URL [本文引用: 2]
Height-class structure dynamics of Hippophae rhamnoides ssp. sinensis clone population in the Loess Plateau
黄土高原中国沙棘克隆种群高度结构动态
A study on the population dynamics and its biomass adjustment mechanisms of clonal plant Hippophae rhamnoides L. ssp. inensis
克隆植物中国沙棘种群动态及其生物量调节机制
Study on population structure and dynamics of Populus euphratica in the middle and lower reaches of the Shule River Basin oasis, Hexi Corridor
疏勒河中下游绿洲胡杨种群结构与动态研究
Field and laboratory exercises in ecology
DOI:10.2307/2259667 URL [本文引用: 1]
Population structure and dynamics of an endangered desert shrub endemic to northwestern China
Quantitative dynamics of Picea crassifolia population in Dayekou basin of Qilian Mountains
祁连山大野口流域青海云杉种群数量动态
Structure and dynamic characteristics of Betula luminifera populations in different regions of Southwest Hubei Province, China
DOI:10.13287/j.1001-9332.202002.010
PMID:32476326
[本文引用: 2]

We investigated Betula luminifera populations in three regions (Mulinzi, Qizimei Mountains, and Jinzi Mountains) in the southwest Hubei Province, China. Population structure was divided by age classes and height classes. Population structure figures were drawn. The static life tables of B. luminifera populations in different regions were analyzed using the method of substitution of space for time. The survival curve, mortality rate curve and disappearance rate curve were created. Four functions of survival analysis were used to analyze the dynamics of B. luminifera population in different regions. The results showed that the B. luminifera populations in three regions were the increasing type. The height class structures were relatively complete. Some age classes were absent from the age structures of B. luminifera populations in Qizimei Mountains and Jinzi Mountains. Although the dynamic index of trees number V>0, but it was sensitive to external disturbance. The survival of B. luminifera of different age classes varied greatly in static life table, which gradually decreased with increasing age class, with Deevey-type 2 survival curve. The trend of mortality rate changed similarly to the disappearance rate, but fluctuated differently. All B. luminifera populations in different regions appeared to decrease in the early stage and keep dynamically stable in the medium-late stage.
鄂西南不同区域亮叶桦种群结构与动态特征
Structure and dynamic characteristics of Gymnocarpos przewalskii in different habitats
DOI:10.17521/cjpe.2015.0095
[本文引用: 1]
<sec><title>Aims</title> <p><i>Gymnocarpos przewalskii</i> is a relict plant of the Tethys flora, which is also one of the constructive species in stony desert. Our objective was to determine the structure and dynamic characteristics of <i>G</i>.<i> przewalskii</i> in different habitats.</p></sec> <sec><title>Methods</title> <p>Based on the equal-area plot investigations in three habitats of <i>G</i>.<i> przewalskii,</i> the static life table was established, survival curves and the function curves derived from the survival analysis were also constructed to evaluate the population characteristics. The population dynamics were predicted by a time-sequence model. </p></sec> <sec><title><i>Important findings</i></title> <p>The survival curves of <i>G</i>.<i> przewalskii</i> appear to approach the Deevey-II type, which indicate that the mortality rate in different size classes was similar, and the population fit in with the growing type in all habitats, but sensitive to environment disturbances. The population size of <i>G</i>.<i> przewalskii</i> was largest in flooding lands, medium in gullies and smallest in the flat desert and the age structure showed that the flooding lands and gullies are better habitats for the growth of <i>G</i>.<i> przewalskii</i>. The survival analysis and the time-sequence model demonstrated that the lack of young individuals could be the major cause for population declines of <i>G</i>.<i> przewalskii</i> in all habitats; moreover, a decline is likely to occur in the flat desert earlier and quicker.</p></sec>
不同生境下濒危植物裸果木种群结构及动态特征
DOI:10.17521/cjpe.2015.0095
[本文引用: 1]
裸果木(Gymnocarpos przewalskii)为古地中海旱生植物区系孑遗种, 是砾质荒漠的主要建群种之一。通过对安西极旱荒漠国家级自然保护区内3种生境(水冲滩地、山间冲沟、平缓戈壁)条件下裸果木种群的样地调查, 编制不同生境裸果木种群的静态生命表, 绘制其存活曲线和生存分析函数的曲线, 并采用时间序列模型对种群的数量动态进行预测, 以揭示不同生境下裸果木的种群结构和动态特征。结果表明: 3种生境下裸果木种群均属增长型, 数量变化动态指数(V’ pi) > 0, 但对外界干扰比较敏感, 存活曲线趋于Deevey-II型, 表明种群各龄级的死亡率基本接近; 不同生境裸果木种群大小依次为水冲滩地>山间冲沟>平缓戈壁, 结合对种群年龄结构的分析, 进一步表明水分条件较好的水冲滩地和山间冲沟生境更加适宜裸果木生存; 生存分析和时间序列预测表明幼龄个体的缺乏是未来裸果木种群发生衰退的主要原因, 且平缓戈壁生境下的裸果木种群将先于另外两种生境下的种群发生更加快速的衰退。
Variation in fruit characteristics of Hippophae tibetana (Elaeagnaceae) in the eastern Qinghai-Tibet Plateau, China
青藏高原东缘西藏沙棘(Hippophae tibetana)果实性状变异
Multidimensional time series analysis on tree growth
林木生长的多维时间序列分析
Quantity dynamics of Hippophae rhamnoides population in different habitats standing in hilly loess regions
黄土丘陵区不同生境沙棘种群数量动态分析
Population quantitative characteristics and dynamics of rare and endangered Tsuga tchekiangensis in Jiulongshan Nature Reserve of China
九龙山珍稀濒危植物南方铁杉种群数量动态
DOI:10.3773/j.issn.1005-264x.2008.05.019
[本文引用: 1]
针对九龙山国家级自然保护区南方铁杉(Tsuga tchekiangensis)种群的分布特点, 设置了10个具有代表性的样地。以种群生命表及生存分析理论为基础, 编制了南方铁杉种群的静态生命表, 绘制了存活曲线、死亡率曲线、消失率曲线、生存率曲线、累计死亡率曲线、种群死亡密度曲线和危险率曲线, 分析了种群数量特征; 同时结合谱分析方法, 分析了南方铁杉种群数量的动态变化。结果表明: 1) 南方铁杉种群结构存在波动性, 幼年阶段的个体较丰富, 成年个体数量相对较少, 种群趋于Deevey Ⅱ型。2) 南方铁杉种群死亡率和消失率曲线变化趋势基本一致, 均出现两个高峰, 一个出现在第5龄级阶段, 另一个出现在第15龄级阶段。3) 南方铁杉种群的生存率单调下降, 累计死亡率单调上升, 生存率下降趋势前期高于后期, 累计死亡率则相反。4) 4个生存函数曲线表明, 南方铁杉具有前期稳定、中期锐减和后期衰退的特点。5) 种群动态的谱分析显示, 南方铁杉种群动态除受基波影响外, 还存在着明显的小周期波动, 谐波A3处周期的波动与南方铁杉的高生长有关; A6处周期的波动与外界环境变化有关; A8处周期的波动与南方铁杉进入生理衰退期有关。
Population dynamics and spatial distribution of the rare and endangered plant Tsuga chinensis var. tchekiangensis in Wuyishan, Jiangxi Province
江西武夷山珍稀濒危植物南方铁杉种群动态与空间分布
Population structure characteristics of Picea purpurea in the upstream of Taohe River
洮河上游紫果云杉种群结构特征
DOI:10.17521/cjpe.2019.0077
[本文引用: 1]
该研究以洮河上游尕海-则岔自然保护区、卡车林区和冶力关林区的紫果云杉(Picea purpurea)天然种群为研究对象, 通过样地调查和数据统计, 绘制种群结构图, 编制静态生命表, 拟合并分析存活曲线, 运用数量化方法研究种群动态, 揭示种群生存现状, 预测种群发展趋势, 以期为该物种的保护、管理及结构恢复提供理论依据。结果显示: 3个林区紫果云杉种群自然更新能力强, 幼苗、幼树储量丰富, 幼小龄期死亡率普遍偏高; 尕海-则岔种群存活曲线符合Deevey-III型, 种群稳定结构完整, 卡车林区和冶力关林区种群存活曲线均符合Deevey-II型, 且均出现了局部衰退; 3个林区种群动态指数均大于0, 说明种群均属于增长型, 增长潜力为尕海-则岔>卡车>冶力关; 受随机干扰时卡车林区紫果云杉最敏感, 冶力关次之, 尕海-则岔种群最稳定。该研究表明: 竞争和自疏作用是造成紫果云杉幼小龄级个体存活率偏低的普遍因素, 3个林区不同的生存状况反映了紫果云杉种群在不同生境及生活史下生存能力的差异, 保护幼苗生存环境并提高幼苗质量和存活率是种群更新和发展的关键。尕海-则岔紫果云杉生存良好, 种群生存状况主要受自身生物学特性和环境因子的影响; 卡车林区主要受人为影响, 种群结构遭到破坏; 冶力关林区受分布限制, 造成种群结构不稳定, 须采取一定的人工措施来促进种群更新与增长。
Temporal variability in soil moisture after thinning in semi-arid Picea crassifolia plantations in northwestern China
DOI:10.1016/j.foreco.2017.07.022 URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}