


种子萌发受到外界环境条件, 主要是光照、温度、水分等的综合影响(Kołodziejek et al., 2017; Carón et al., 2020)。种子在适宜的环境条件下萌发是确保植物存活、竞争以及繁殖等的重要机制(张佳宁和刘坤, 2014)。受到林窗、林缘、林层高度和时间(不同时刻、不同季节)的影响, 森林群落中生境条件存在高度的异质性(Ritter et al., 2005)。空间尺度上, 不同大小林窗和林窗中的不同位置均会引起林下光照强度和温度的差异(管云云等, 2016); 时间尺度上, 随着一天以及一年四季太阳高度角的变化, 森林生态系统中同一位置的光照和温度也发生剧烈的变化(陈圣宾等, 2005), 为林下种子的萌发和幼苗更新提供了不同的微生境。种子萌发也受到种子自身遗传特性的影响。植物在分布范围内的不同区域, 由于地理气候条件存在差异, 植物对其响应不同, 在自然选择过程中会逐渐产生地理变异(徐化成, 1992)。研究发现, 不同地理种源的树木种子呈现不同的萌发特性(Fredrick et al., 2017; 宿昊等, 2021), 对环境因子的响应不同。研究不同种源种子在环境梯度下的萌发特征有利于揭示物种种源的遗传规律和变异程度。掌握种子在萌发要求上的变异规律, 能够完善对种群生态属性的认识, 为退化种群的恢复和建植提供科学依据, 促进天然更新过程。
红松(Pinus koraiensis)种群在我国主要分布于东北东部地区, 包括辽宁、吉林和黑龙江省等种源地(马建路等, 1992)。红松和阔叶树种紫椴(Tilia amurensis)、蒙古栎(Quercus mongolica)、水曲柳(Fraxinus mandshurica)等构成了我国东北东部山区的地带性顶极群落——阔叶红松林。由于人为和自然灾害的干扰作用, 红松建群种大量缺失, 造成大约60%的阔叶红松林转变为次生林、农田、人类居住地、道路和其他土地利用类型(Li et al., 2012)。天然林保护工程实施以来, 部分次生林已经进入了正向演替的更新和恢复阶段(朱教君, 2002; 谷瑞升等, 2005), 但其森林生态功能还远远不如原始阔叶红松林(于立忠等, 2005; 邹莉等, 2015)。提高建群种红松的天然更新能力是恢复阔叶红松林的关键措施。前期调查发现, 次生林生态系统中红松天然更新差, 林下可见实生幼苗数量不多, 且不同林冠开阔度下红松幼苗更新差异显著(Zhang et al., 2012)。因此, 在森林经营中, 通过人为干预提高野外林分下红松种子的萌发率和萌发速率, 能够促进红松种群的天然更新。红松种子具深休眠性, 且常受种源地及其光、温等生境的影响, 但种源与光、温耦合对红松种子萌发影响的研究相对薄弱。基于此, 本研究以辽宁清原、吉林长白山和黑龙江伊春种源地的红松种子为研究对象, 分别设置林下不同光温生境和室内光温控制实验, 观测红松种子的萌发特征, 旨在: (1)揭示天然次生林中不同季节和林分生境下红松种群的更新格局和规律; (2)明确红松不同种源对环境梯度的响应差异; (3)阐明光、温在红松种子萌发过程中的作用, 进而找出林下红松天然更新差的原因。研究结果能够为促进次生林下红松种群的天然更新和引导东北次生林向阔叶红松林恢复及正向演替提供科学依据。
1 材料和方法
1.1 种子采集
2019年9-10月, 分别从辽宁清原(41.85° N, 124.94° E)、吉林长白山(42.40° N, 128.47° E)和黑龙江伊春(48.15° N, 129.24° E) 3个种源地采集新鲜的红松种子(其中, 清原和长白山种源为人工林, 伊春种源为天然林)。每个种源地种子均采自10株以上的母树(年龄>50 a), 并将采集的种子充分混合以确保遗传异质性。3个种源地红松种子的百粒质量分别为清原: (55.83 ± 1.09) g, 长白山: (52.89 ± 0.33) g, 伊春: (64.87 ± 0.29) g。新采集的红松种子在室温下阴干1个月, 后装袋分别贴上种源地标签, 置于-20 ℃冰箱内保存, 用于室内控制实验。
1.2 实验设计与处理
3个种源地的红松种子在自然室温下阴干1个月后, 于10月底分别置于不同透光林分下进行越冬埋藏。选取林窗(12%林冠开阔度)和郁闭林分(5%林冠开阔度)作为2个不同透光处理, 每个透光处理重复3次。每个生境埋藏红松种子1 800粒(600粒/种源× 3种源)。为防止啮齿类动物取食, 每100粒红松种子放入1个尼龙网袋, 每个重复共计18袋(6袋/种源× 3种源), 每4个尼龙网袋一起放入铁丝编织的网袋内。分别于次年的5月(春)、7月(夏)、9月(秋)分3次取出, 每次每个生境和种源取出2袋(200粒), 调查并记录种子萌发数, 每个季节未萌发的种子均带回实验室, 在光照培养箱内继续培养, 萌发光强为200 μmol·m-2·s-1, 昼夜比为12/12 h, 昼夜温度为25/15 ℃。以可见白色针状物为萌发标准, 连续7天不再有种子萌发视为萌发结束(Argyris et al., 2008; Zhu et al., 2008)。
5-9月期间, 每个月选择无太阳直射条件下拍摄6个不同透光林分样地的鱼眼镜头照片, 同时记录样地的经纬度和坡度数据, 利用GLA软件对照片进行处理, 计算每个样地的林冠开阔度, 以表征林下光照条件的变化。从11月至次年10月, 在各样地埋设WDS自动温湿度记录仪(DS1923, 上海沃第森电子科技有限公司, 上海), 实时自动监测6个样地的土壤温湿度数据。
1.3 光照培养
为了阐明光、温以及二者交互作用对3个种源红松种子萌发的影响, 利用光照培养箱进行光温控制萌发实验。萌发实验开始前, 将每个种源地种子取出, 置于蒸馏水中吸胀7天, 每隔2天换一次水, 7天后除去漂浮的不合格种子, 以下沉的饱满红松种子为供试种子。吸胀种子在5 ℃恒温箱中混沙湿藏30天以解除休眠。萌发实验在光照培养箱中进行, 分别设置3个光照强度: 200、10、0 μmol·m-2·s-1 (记为L200、L20和L0), 2个温度水平: 25、15 ℃, 共6个光温处理, 每个处理每个种源种子数50粒, 每个处理重复4次, 昼夜比为12/12 h。采用培养皿滤纸萌发法, 每个培养皿中放置2张滤纸, 种子置于滤纸上, 每隔2天浇水1次并记录萌发种子数, 每隔4天更换一次滤纸。萌发标准同1.2。
1.4 萌发指标
采用萌发率(GP)、平均萌发时间(MGT)和萌发指数(GV)来评估种子萌发情况。GP、MGT和GV的计算公式如下:
GP = n/N
式中, n为萌发的种子数, N为用于萌发实验的种子总数。
$MGT= \sum (n_d × d)/N$
GV=PV×MDG
式中, t为从萌发开始到结束的总天数, PV为平均每天萌发率的最大值(Han et al., 2018)。
1.5 数据处理
对不符合正态分布和方差齐性要求的数据, 进行反正弦平方根或对数转换。对不同种源、季节和透光率下的萌发率, 不同种源、取出季节和透光率下的室内萌发率, 不同种源、光照和温度条件下的萌发率、平均萌发时间和萌发指数进行三因素方差分析。为探讨不同种源、季节、取出季节及控制光照条件的单因子效应, 利用单因素方差分析比较不同水平之间的差异。方差齐性情况下, 采用最小显著差异(LSD)法进行多重比较; 方差不齐的情况下, 采用Dunnett's T3检验进行多重比较。为探讨林窗和林下之间, 室内两个温度之间的单因子效应, 利用t检验比较两水平间的差异。显著性水平为α = 0.05。利用SPSS 23.0软件进行数据分析和处理, SigmaPlot 14.0软件作图。
2 结果和分析
2.1 林窗和林下光温季节变化
图1
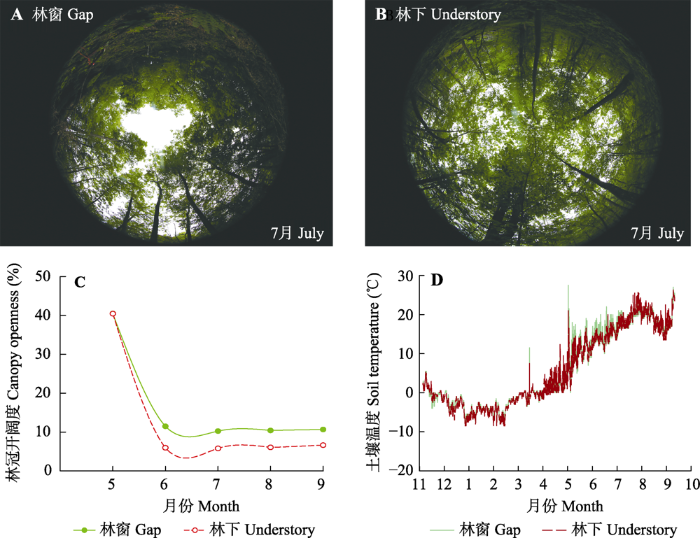
图1
不同透光林分鱼眼镜头照片(A、B)、林冠开阔度(C)和土壤温度(D)的季节变化。
Fig. 1
Fisheye photographs (A, B), seasonal variations of canopy openness (C), and daily soil temperature (D) in forest stands with different transmittances.
2.2 不同季节野外条件下红松种子的萌发特性
由表1可见, 季节、种源、透光率、季节与种源交互和季节与透光率交互作用均显著影响林下红松种子的萌发率, 3个因素之间均不存在交互作用。
表1 不同透光林分下红松种子萌发率的季节、种源和透光率三因素方差分析
Table 1
| 源 Source | III类平方和 Type III sum of squares | 自由度 df | F | p |
|---|---|---|---|---|
| 模型 Model | 6 942.93a | 17 | 20.66 | 0.00 |
| 季节 Season | 2 066.04 | 1 | 195.69 | 0.00 |
| 种源 Provenance | 2 859.23 | 2 | 72.34 | 0.00 |
| 透光率 Light | 106.96 | 1 | 5.41 | 0.03 |
| 季节×种源 Season × Provenance | 1 475.41 | 4 | 18.66 | 0.00 |
| 季节×透光率 Season × Light | 222.48 | 2 | 5.63 | 0.01 |
| 种源×透光率 Provenance × Light | 98.84 | 2 | 2.50 | 0.10 |
| 种源×透光率×季节 Provenance × Light × Season | 113.96 | 4 | 1.44 | 0.24 |
a, 决定系数R2 = 0.907 (调整后R2 = 0.863)。加粗表示差异显著(p ≤ 0.05)。
a, coefficient of determination R2 = 0.907 (adjusted R2 = 0.863). Significant differences (p ≤ 0.05) are shown in bold.
图2
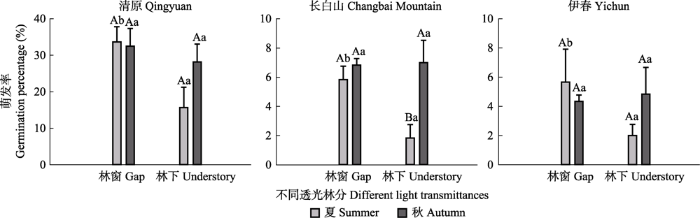
图2
三个种源红松种子在不同透光林分下不同季节的自然萌发率(平均值±标准误)。红松种子在春季均未萌发, 因此仅显示夏、秋季数据。不同大写字母表示同一林分不同季节间差异显著(p ≤ 0.05), 不同小写字母表示同一季节不同透光林分间差异显著(p ≤ 0.05)。
Fig. 2
Germination percentage of Pinus koraiensis seeds from three provenances under different light conditions between different seasons (mean ± SE). The seeds did not germinate in spring, thus only summer and autumn data were displayed. Different uppercase letters indicate significant differences between different seasons in the forest stands with the same light transmittance (p ≤ 0.05). Different lowercase letters indicate significant differences between forest stands with different light transmittances in the same season (p ≤ 0.05).
各季节林下未萌发的种子带回实验室, 在适宜温度(25/15 ℃)下测定其萌发率, 进一步分析种子未萌发或萌发少的原因。春季未萌发种子带回实验室培养, 萌发率最高的为清原红松种子, 为61%- 77%, 其次是伊春红松种子, 萌发率为56%-57%, 长白山红松种子萌发率最低, 仅为32%-34%。夏季未萌发种子在室内的萌发率在不同种源间差异不明显, 均在45%-62%之间。秋季未萌发种子取出在光照培养箱内培养, 发现萌发率极低, 均不高于2% (表2)。
表2 不同透光林分下越冬储藏的3个种源地红松种子室内萌发率(%) (平均值±标准误)
Table 2
| 种源地 Provenance | 种子取出季节 Seed-collection season | 林窗 Gap | 林下 Understory |
|---|---|---|---|
| 清原 Qingyuan | 春 Spring | 76.67 ± 1.45a | 61.00 ± 8.19a |
| 夏 Summer | 56.00 ± 5.29a | 58.33 ± 7.54a | |
| 秋 Autumn | 2.00 ± 1.00a | 0.67 ± 0.33a | |
| 长白山 Changbai Mountain | 春 Spring | 34.00 ± 8.50a | 32.00 ± 4.93a |
| 夏 Summer | 53.00 ± 1.15a | 45.00 ± 10.69a | |
| 秋 Autumn | 1.67 ± 0.89a | 0.33 ± 0.33a | |
| 伊春 Yichun | 春 Spring | 57.33 ± 5.90a | 56.00 ± 11.50a |
| 夏 Summer | 62.33 ± 5.04a | 50.00 ± 10.69a | |
| 秋 Autumn | 0.67 ± 0.33b | 0a |
不同小写字母表示不同透光林分间差异显著(p ≤ 0.05)。
Different lowercase letters indicate significant difference between forest stands with different light conditions (p ≤ 0.05).
2.3 室内光温控制下红松种子的萌发特性
三因素方差分析表明, 除了种源和光照的交互作用对MGT影响不显著(表3), 光照、温度、种源、光照和温度互作、光照和种源互作、种源和温度互作以及三因素互作对红松种子的GP、MGT和GV均产生显著影响。
表3 不同光照、温度和种源地对红松种子萌发率(GP)、平均萌发时间(MGT)、萌发指数(GV)的三因素方差分析
Table 3
| 指标 Index | 源 Source | III类平方和 Type III sum of squares | 自由度 df | F | p |
|---|---|---|---|---|---|
| GP | 模型 Model | 27 581.179a | 17 | 56.175 | 0.00 |
| 种源 Provenance | 6 191.542 | 2 | 107.189 | 0.00 | |
| 光 Light | 2 987.890 | 2 | 51.727 | 0.00 | |
| 温度 Temp | 12 454.163 | 1 | 431.217 | 0.00 | |
| 种源×光 Provenance × Light | 1 289.040 | 4 | 11.158 | 0.00 | |
| 种源×温度 Provenance × Temperature | 1 448.825 | 2 | 25.082 | 0.00 | |
| 光×温度 Light × Temperature | 877.690 | 2 | 15.195 | 0.00 | |
| 种源×光×温度 Provenance × Light × Temperature | 1 090.786 | 4 | 9.442 | 0.00 | |
| MGT | 模型 Model | 4 888.740b | 17 | 6.396 | 0.00 |
| 种源 Provenance | 686.604 | 2 | 7.636 | 0.00 | |
| 光 Light | 542.808 | 2 | 6.037 | 0.00 | |
| 温度 Temp | 1 355.676 | 1 | 30.154 | 0.00 | |
| 种源×光 Provenance × Light | 178.422 | 4 | 0.992 | 0.42 | |
| 种源×温度 Provenance × Temperature | 679.857 | 2 | 7.561 | 0.00 | |
| 光×温度 Light × Temperature | 371.673 | 2 | 4.133 | 0.02 | |
| 种源×光×温度 Provenance × Light × Temperature | 743.805 | 4 | 4.136 | 0.00 | |
| GV | 模型 Model | 57.775c | 17 | 47.783 | 0.00 |
| 种源 Provenance | 17.631 | 2 | 123.943 | 0.00 | |
| 光 Light | 0.556 | 2 | 3.911 | 0.03 | |
| 温度 Temperature | 28.159 | 1 | 395.914 | 0.00 | |
| 种源×光 Provenance × Light | 1.801 | 4 | 6.331 | 0.00 | |
| 种源×温度 Provenance × Temperature | 6.471 | 2 | 45.491 | 0.00 | |
| 光×温度 Light × Temperature | 0.869 | 2 | 6.106 | 0.00 | |
| 种源×光×温度 Provenance × Light × Temperature | 2.266 | 4 | 7.964 | 0.00 |
a, 决定系数(R2) = 0.943 (调整后R2 = 0.926); b, R2 = 0.664 (调整后R2 = 0.560); c, R2 = 0.937 (调整后R2 = 0.917)。加粗表示差异显著(p ≤ 0.05)。
a, coefficient of determination (R2) = 0.943 (adjusted R2 = 0.926); b, R2 = 0.664 (adjusted R2 = 0.560); c, R2 = 0.937 (adjusted R2 = 0.917). Significant differences (p ≤ 0.05) are shown in bold.
所有光温处理下, 清原、清原与伊春红松种子萌发率显著高于长白山红松种子萌发率。清原、长白山和伊春的红松种子萌发率均为25 ℃条件下显著高于15 ℃条件下(图3)。25 ℃条件下, 3个种源地红松种子萌发率均呈现随着光照强度的降低逐渐降低的趋势, L200条件下显著高于L0条件。15 ℃条件下, 清原红松种子萌发率随着光照强度的下降逐渐下降, 呈现L200条件下显著高于L20和L0条件; 长白山种子萌发率呈现随着光照强度的降低逐渐升高的趋势, 在L200条件下显著低于L20和L0; 伊春种子萌发率则呈现随着光照强度的下降先上升后下降趋势, 在L200和L0条件下显著低于L20条件。15 ℃温度下, 长白山和伊春的红松种子在3个光照强度下萌发率均低于20%。可见, 长白山和伊春种源红松种子萌发对温度的要求可能更高于清原红松种子。长白山和伊春红松种子随着温度降低, 其萌发对光的需求降低。
图3
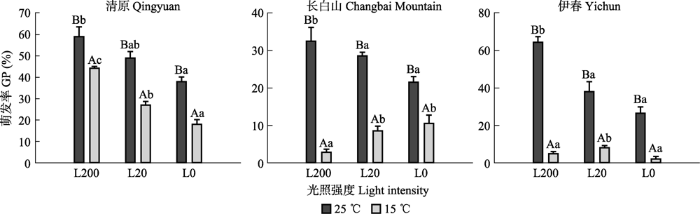
图3
光照培养箱不同光照和温度条件下红松种子的萌发率(平均值±标准误)。L0, L20, L200代表光照强度分别为0, 20, 200 μmol·m-2·s-1。不同大写字母表示相同光照下不同温度之间萌发率差异显著(p ≤ 0.05); 不同小写字母表示相同温度条件下不同光照之间萌发率差异显著(p ≤ 0.05)。
Fig. 3
Germination percentage (GP) of Pinus koraiensis seeds from three provenances under different light and temperature conditions in growth chamber (mean ± SE). L0, L2 and L200 represent 0, 20, 200 μmol·m-2·s-1 light intensities, respectively. Different uppercase letters indicate significant differences between different temperature conditions under the same light intensity (p ≤0.05); different lowercase letters indicate significant differences between different light intensities under the same temperature condition (p ≤ 0.05).
图4可见, 光照对两个温度下清原红松种子的平均萌发时间均无显著影响。25 ℃温度下, 长白山种子的平均萌发时间在L20显著高于L200和L0条件, 伊春种子的平均萌发时间在L20和L0显著高于L200条件。15 ℃温度下, 长白山种子的平均萌发时间在L200和L20显著高于L0条件, 伊春种子的平均萌发时间在3个光照强度间无显著差异。整体上, 长白山(L200和L20条件)和伊春红松种子(L200条件)在15 ℃条件下平均萌发时间显著高于25 ℃条件。
图4
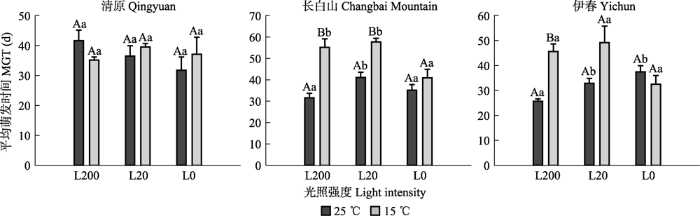
图4
不同光照和温度条件下红松的平均萌发时间(平均值±标准误)。L0、L20、L200代表光照强度分别为0、20、200 μmol·m-2·s-1。不同大写字母表示相同光照下不同温度之间萌发率差异显著(p ≤ 0.05); 不同小写字母表示相同温度条件下不同光照之间萌发率差异显著(p ≤ 0.05)。
Fig. 4
Mean germination time (MGT) of Pinus koraiensis seeds from three provenances under different light and temperature conditions (mean ± SE). L0, L2 and L200 represent 0, 20, 200 μmol·m-2·s-1 light intensities, respectively. Different uppercase letters indicate significant differences between different temperature conditions under the same light intensity (p ≤ 0.05); different lowercase letters indicate significant differences between different light intensities under the same temperature condition (p ≤ 0.05).
光照和温度对红松种子萌发指数的影响与萌发率相似(图5)。在所有光温处理中, 清原、清原和伊春红松种子的萌发指数均高于长白山种子。3个光照强度下, 所有种源地红松种子萌发指数均为25 ℃下显著高于15 ℃。25 ℃条件下, 3个种源地红松种子的萌发指数均呈现L200显著高于L0条件下。15 ℃条件下, 清原红松种子的萌发指数呈现L200显著大于L20和L0条件, 长白山种子的萌发指数呈现L200和L20显著小于L0光照强度, 伊春种子的萌发指数在3个光照强度间无显著差异。
图5
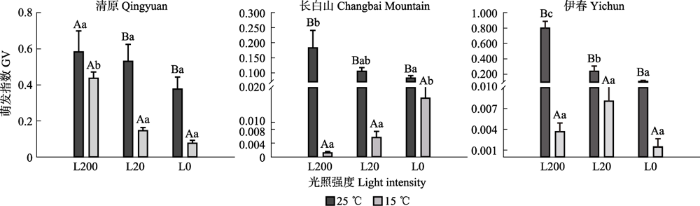
图5
不同光照和温度条件下红松种子萌发指数(平均值±标准误)。L0、L20、L200代表光照强度分别为0、20、200 μmol·m-2·s-1。不同大写字母表示相同光照下不同温度之间萌发率差异显著(p ≤ 0.05); 不同小写字母表示相同温度条件下不同光照之间萌发率差异显著(p ≤ 0.05)。
Fig. 5
Germination value (GV) of Pinus koraiensis seeds from three provenances under different light and temperature conditions (mean ± SE). L0, L2 and L200 represent 0, 20, 200 μmol·m-2·s-1 light intensities, respectively. Different uppercase letters indicate significant differences between different temperature conditions under the same light intensity (p ≤ 0.05); different lowercase letters indicate significant differences between different light intensities under the same temperature condition (p ≤ 0.05).
3 讨论
本研究发现, 林窗和林下清原红松种子萌发率均显著高于伊春和长白山种子, 这可能是因为林分实验在清原地区开展, 与不同种源地种子对分布区自然生境长期的适应有关(Bu et al., 2008; 谭美等, 2018)。种源地理位置差异引起的气候差异是造成不同种源地红松种子萌发对温度需求不同的原因。各个光温处理下, 清原红松种子萌发率均略高于或显著高于长白山和伊春红松种子, 且15 ℃条件下长白山和伊春红松种子萌发率均低于20%, 这表明长白山和伊春红松种子的萌发对温度的要求相比于清原可能更高。这与李阳等(2016)的研究结果相一致。本研究为保证立地条件一致, 并未在伊春和长白山地区同时设置不同种源的萌发试验, 而是统一设置在清原次生林下进行。研究结果能够揭示光、温对不同种源红松种子萌发的影响。对林下未萌发红松种子进行光温控制培养, 发现伊春和长白山红松种子的萌发高峰稍晚于清原红松种子(表2), 这也从侧面佐证了伊春和长白山红松种子完全解除休眠可能需要更高的积温条件。
温度对种子萌发的影响是一个持续积累的过程(Finch-Savage & Leubner-Metzger, 2006)。在适宜的温度条件下, 种胚才能长大实现形态后熟(Song et al., 2017), 只有同时完成形态后熟和生理后熟的红松种子才能萌发(陈彩霞等, 1997)。本研究发现, 林窗和林下3个种源红松种子春季均不萌发, 夏季和秋季仅少量萌发, 但春夏季未萌发种子给予适宜的光温条件(25/15 ℃, 200 μmol·m-2·s-1)后则大量萌发, 秋季未萌发种子在室内萌发率仍然很低。这表明林下越冬储藏的红松种子原生休眠已经打破, 春夏季萌发率低可能是因为春夏季林下的光、温未达到种子萌发的适宜条件, 而秋季种子萌发率低可能是由于红松种子启动了次生休眠(Song et al., 2020)。野外林分下, 从春季到秋季, 红松种子经历一个波动的逐渐升温(0-25 ℃)过程(图1), 研究证实该过程会诱导红松种子进入次生休眠(Song et al., 2020)。次生休眠能够避免植物种子在不适宜的条件下萌发增加死亡的风险(Baskin & Baskin, 2014)。夏季, 3个种源红松种子萌发率均呈现在林窗下显著高于郁闭林下的空间异质性。这是因为夏季林冠层乔木基本已完全展叶, 林窗的透光率显著高于郁闭林下, 满足了红松种子萌发的需光性。因此, 提高林下透光率能够促进红松种子萌发, 这与Yu等(2008)对箭毒木(Antiaris toxicaria)和红锥(Castanopsis hystrix)以及Kara和Topaçoglu (2018)对林下樟子松(Pinus sylvestris var. mongolica)的研究结论相似。
温度对种子萌发的影响因物种而异。所有光照强度下, 3个种源地红松种子的萌发率和萌发指数均为25 ℃条件下显著高于15 ℃条件下, 表明25 ℃为红松种子萌发的适宜温度且是红松种子大量萌发的必要前提。前人对其他针叶树种进行研究, 发现其种子萌发的适宜温度各不相同。例如, 李银科等(2011)研究发现20-25 ℃为樟子松和沙地云杉(白杄, Picea meyeri)发芽的最适温度; 郭聪聪等(2021)研究发现15和20 ℃温度条件最有利于白皮松(P. bungeana)种子的萌发。根据种子萌发对光照的响应, 种子可以划分为需光种子、忌光种子和光中性种子。光照促进需光树木种子的萌发, 但对忌光种子的萌发具有抑制作用(张敏等, 2012)。例如, 辽东栎(Quercus wutaishanica)种子在强光照(55.4%全光照)下萌发率最高(闫兴富等, 2011); 而Pinus pinea种子在光照或黑暗条件下萌发率均很高(Ganatsas & Tsakaldimi, 2007)。一些研究还发现, 种子萌发对光照和温度的响应存在耦合作用, 种子萌发对光照的响应随着温度条件的变化而变化(Guo et al., 2020)。本研究中, 25 ℃条件下, 3个种源地的红松种子萌发率均呈现随着光照强度的下降而下降的变化规律, 即红松种子在适宜温度下呈现萌发需光的特征。这与王玲玲等(2015)对侧柏(Platycladus orientalis)、Ghaderi-Far等(2021)对Lavandula stoechas等植物的研究结果相一致。以往对松属植物的研究中发现, 一些小粒种子表现出萌发需光性, 例如矮松(Pinus virginiana)(Toole et al., 1961)、海岸松(P. pinaster) (Ruano et al., 2009)和樟子松(Gaudio et al., 2011)。本研究的红松种子单粒质量大于0.5 g, 相较于前人的研究为大粒种子。可见不同于Metcalfe和Grubb (1995)和Milberg等(2000)提出的种子萌发需光性随种子大小的增加而逐渐减弱的结论, 一些大粒种子也可能表现出萌发需光性。另外, 红松种子萌发对光照的响应还表现出种源特异性: 清原红松种子在2个温度条件下均表现出萌发率随着光的减弱而降低, 而长白山和伊春的红松种子只有在25 ℃条件下呈现萌发率随光强的减弱而降低。该结果可能归因于不同种源种子萌发对温度的需求不同, 而光照对红松种子萌发的影响受到温度的制约。根据Finch-Savage和Leubner-Metzger (2006)的研究, 相比于温度, 光照影响种子萌发常常是即时的, 并且是最后起作用的因子。15 ℃条件下, 不同于清原和伊春种源, 长白山红松种子在黑暗条件下萌发率反而更高。这与不同种群Lavandula stoechas的研究结果相类似(Ghaderi-Far et al., 2021), 可能是受到种子发育和成熟过程中母树所处的环境条件、种群对当地环境的适应或其他未考虑因素的影响(Pérez-García et al., 2013)。
4 结论
(1)相同生境条件下, 清原红松种子萌发率均显著高于长白山和伊春, 长白山和伊春红松种子需要更高的积温以解除休眠。
(2)野外林分下, 红松种子春、夏季不萌发或萌发率低, 不适宜种子萌发; 3个种源种子夏季在林窗下萌发率均显著高于林下, 红松种子萌发具有需光性。
(3) 25 ℃为红松种子萌发的适宜温度, 适宜温度下红松种子呈现出明显的萌发需光性。
因此, 野外林分下应适时(春、夏季)对林分进行透光处理, 改善林下光环境, 可促进林下红松种子萌发, 完成种群天然更新。
参考文献
Genetic variation for lettuce seed thermoinhibition is associated with temperature-sensitive expression of abscisic acid, gibberellin and ethylene biosynthesis, metabolism and response genes
DOI:10.1104/pp.108.125807
PMID:18753282
[本文引用: 1]

Lettuce (Lactuca sativa 'Salinas') seeds fail to germinate when imbibed at temperatures above 25 degrees C to 30 degrees C (termed thermoinhibition). However, seeds of an accession of Lactuca serriola (UC96US23) do not exhibit thermoinhibition up to 37 degrees C in the light. Comparative genetics, physiology, and gene expression were analyzed in these genotypes to determine the mechanisms governing the regulation of seed germination by temperature. Germination of the two genotypes was differentially sensitive to abscisic acid (ABA) and gibberellin (GA) at elevated temperatures. Quantitative trait loci associated with these phenotypes colocated with a major quantitative trait locus (Htg6.1) from UC96US23 conferring germination thermotolerance. ABA contents were elevated in Salinas seeds that exhibited thermoinhibition, consistent with the ability of fluridone (an ABA biosynthesis inhibitor) to improve germination at high temperatures. Expression of many genes involved in ABA, GA, and ethylene biosynthesis, metabolism, and response was differentially affected by high temperature and light in the two genotypes. In general, ABA-related genes were more highly expressed when germination was inhibited, and GA- and ethylene-related genes were more highly expressed when germination was permitted. In particular, LsNCED4, a gene encoding an enzyme in the ABA biosynthetic pathway, was up-regulated by high temperature only in Salinas seeds and also colocated with Htg6.1. The temperature sensitivity of expression of LsNCED4 may determine the upper temperature limit for lettuce seed germination and may indirectly influence other regulatory pathways via interconnected effects of increased ABA biosynthesis.
Germination of Dracaena cinnabari Balf. f. seeds under controlled temperature conditions
DOI:10.3390/f11050521 URL [本文引用: 1]
The effect of light and seed mass on seed germination of common herbaceous species from the eastern Qinghai-Tibet Plateau
DOI:10.1111/1442-1984.12147 URL [本文引用: 1]
Seed germinating characteristics of 54 gramineous species in the alpine meadow on the eastern Qinghai-Tibet Plateau
DOI:10.1007/s11515-008-0027-9 URL [本文引用: 1]
Germination responses to light of four Neotropical forest tree species along an elevational gradient in the southern Central Andes
DOI:10.1111/1440-1703.12112 URL [本文引用: 1]
Research state of the dormancy and pregermination of Pinus koraiensis seed in China and aboard
国内外红松种子休眠及催芽问题研究动态
Research advance in re- sponse of forest seedling regeneration to light environmental heterogeneity
森林幼苗更新对光环境异质性的响应研究进展
在分析森林光环境异质性特点的基础上,从幼苗的光合、热耗散、生物量累积和分配、形态特征、萌发和种群动态等方面综述了国内外在森林幼苗更新对光环境异质性响应的研究进展.从森林物种多样性维持、森林演替和植被恢复角度探讨了幼苗更新对光环境异质性不同响应的生态学意义,并对今后该方面研究提出建议.
Differences in seed germination responses may promote coexistence of four sympatric Piper species
DOI:10.1046/j.1365-2435.2002.00615.x URL [本文引用: 1]
Seed dormancy and the control of germination
Seed dormancy is an innate seed property that defines the environmental conditions in which the seed is able to germinate. It is determined by genetics with a substantial environmental influence which is mediated, at least in part, by the plant hormones abscisic acid and gibberellins. Not only is the dormancy status influenced by the seed maturation environment, it is also continuously changing with time following shedding in a manner determined by the ambient environment. As dormancy is present throughout the higher plants in all major climatic regions, adaptation has resulted in divergent responses to the environment. Through this adaptation, germination is timed to avoid unfavourable weather for subsequent plant establishment and reproductive growth. In this review, we present an integrated view of the evolution, molecular genetics, physiology, biochemistry, ecology and modelling of seed dormancy mechanisms and their control of germination. We argue that adaptation has taken place on a theme rather than via fundamentally different paths and identify similarities underlying the extensive diversity in the dormancy response to the environment that controls germination.
Provenance and pretreatment effect on seed germination of six provenances of Faidherbia albida (Delile) A. Chev
DOI:10.1007/s10457-016-9974-3 URL [本文引用: 1]
Effect of light conditions and salinity on germination behaviour and early growth of umbrella pine (Pinus pinea L.) seed
DOI:10.1080/14620316.2007.11512280 URL [本文引用: 1]
Light-mediated influence of three understorey species (Calluna vulgaris, Pteridium aquilinum, Molinia caerulea) on the growth of Pinus sylvestris seedlings
DOI:10.1007/s10342-010-0403-2 URL [本文引用: 1]
Light and temperature requirements for germination in the Mediterranean shrub Lavandula stoechas (Lamiaceae)
DOI:10.1111/plb.13329
PMID:34516720
[本文引用: 2]
Available studies are far from giving a complete figure for the germination requirements of many Mediterranean Basin species. In this study, we investigated the germination properties of Lavandula stoechas L. (Lamiaceae) in response to different light and temperature regimes. We performed comprehensive experiments to test the effect of fixed and alternating temperatures, dark versus light conditions, and thermodormancy or thermoinhibition on several germination properties in three populations of L. stoechas from southwestern Turkey. Germination patterns showed a substantial variation among populations. Germination percentage was higher in the light conditions than in the dark at most temperatures, and alternating temperatures substituted light for the germination. The requirement of L. stoechas seeds to light for germination gradually increased through lower and higher temperatures than 15 °C. High temperature (30 °C) reduced the germination percentage to zero. However, in the presence of light, seeds were able to germinate after transferring to the optimum temperature (thermoinhibition), but this was not the case in darkness (thermodormancy). Seed germination in L. stoechas was stimulated by light, suggesting that germination has a phytochrome-mediated response. Our study makes an inference to the fire-environment interactions in the germination of Mediterranean seeder species by showing that both light and alternating temperatures stimulate germination in L. stoechas, a species well-known with its fire-related germination. Comprehensive germination studies are required for a better understanding of the early life-stage adaptations of plants to Mediterranean conditions, and for developing more robust strategies for conservation and ecosystem restoration.© 2021 German Society for Plant Sciences and The Royal Botanical Society of the Netherlands.
The status and development of Chinese forest basic research
我国林学基础研究及其发展
Advances in studies of forest gap ecology
林窗生态学研究进展
Response of respiration and hormones during germination of Pinus bungeana seeds to temperature changes
白皮松种子萌发过程中呼吸代谢和内源激素对温度变化的响应
Effect of temperature, light and storage time on the seed germination of Pinus bungeana Zucc. ex Endl.: the role of seed-covering layers and abscisic acid changes
DOI:10.3390/f11030300 URL [本文引用: 1]
Seed germination and initial seedling survival of the subalpine tree species, Picea jezoensis, on different forest floor substrates under elevated temperature
DOI:10.1016/j.foreco.2018.07.042 URL [本文引用: 1]
Influence of stand density and canopy structure on the germination and growth of Scots pine (Pinus sylvestris L.) seedlings
DOI:org/10.1007/s10661-018-7129-x URL [本文引用: 1]
Effect of light, gibberellic acid and nitrogen source on germination of eight taxa from dissapearing European temperate forest, Potentillo albae-Quercetum
DOI:10.1038/s41598-017-13101-z
PMID:29066749
[本文引用: 1]
Little is known about how light affects seed germination and revegetation of species of thermophilous oak forest. To reveal this relationship effects of white, red, far-red irradiations and dark incubation on germination of eight Potentillo albae-Quercetum taxa were examined. Attempts were also made to evaluate the influence of gibberellic acid and different nitrogen sources on the germination characteristics. Interaction between light and nitrogen was also studied. Freshly matured seeds of all taxa germinated very poorly, indicating presence of primary dormancy. Germination rates increased after wet-stratification treatment and were low in darkness. The highest concentration of the nitrogenous solutions that resulted in high germination level was 10 mM, whereas higher concentrations had a negative effect. Nitrate had the strongest influence which can be proved by a 'gap detection' mechanism for gaps in the vegetation. Far-red and red irradiation showed antagonistic effect on seed germination. There was a decrease in germination when far-red was followed by red and an improvement when red was followed by far-red treatment. Under red light, gibberellic acid enhanced germination of positively photoblastic taxa. It was concluded that light factor, associated with vegetation gaps, was the most important signal stimulating germination of the studied taxa.
Effects of temperature and seed size on seed germination in Potentilla aurea
温度和种子大小对黄花委陵菜种子萌发特征的影响
Evaluation of regeneration potential of Pinus koraiensis in mixed pine-hardwood forests in the Xiao Xing'an Mountains, China
DOI:10.1007/s11676-012-0294-9 URL [本文引用: 1]
Effects of temperature on germination characteristics of Pinus sylvesiris var. mongolica and Picea mongolica seed
温度对樟子松和沙地云杉种子萌发特征的影响
Geographic distribution of Pinus koraiensis in the world
红松的地理分布
Seed mass and light requirements for regeneration in Southeast Asian rain forest
DOI:10.1139/b95-090 URL [本文引用: 1]
Large-seeded species are less dependent on light for germination than small-seeded ones
DOI:10.1017/S0960258500000118 URL [本文引用: 1]
Interpopulation variation in seed germination of five Mediterranean Labiatae shrubby species
DOI:10.1560/2YAB-37UL-70QV-LMFT URL [本文引用: 1]
Light, temperature and soil moisture regimes following gap formation in a semi-natural beech-dominated forest in Denmark
DOI:10.1016/j.foreco.2004.08.011 URL [本文引用: 1]
How do light and water influence Pinus pinaster Ait. germination and early seedling development?
DOI:10.1016/j.foreco.2009.09.027 URL [本文引用: 1]
The temperature and length for the release of primary and induction of secondary physiological dormancy in Korean pine (Pinus koraiensis Sieb. et Zucc.) seeds
DOI:10.1007/s11056-019-09750-9 URL [本文引用: 2]
Korean pine seed: lingking changes in dormancy to germination in the 2 years following dispersal
DOI:10.1093/forestry/cpx037 URL [本文引用: 1]
Germination characteristics of Betula albo-sinensis seeds from different provenances
不同种源红桦种子的萌发特性
Study on seed germination and seedling growth of Houpoöa officinalis in different habitats
不同生境内厚朴种子萌发和幼苗生长研究
Responses of seeds of Pinus virginiana to light
DOI:10.1104/pp.36.3.285 PMID:16655506 [本文引用: 1]
Effects of temperatures and light on seed germination of Platycladus orientalis (Linn.) Franco
温度和光照对侧柏种子萌发的影响
Interaction of seed size with light quality and temperature regimes as germination cues in 10 temperate pioneer tree species
DOI:10.1111/1365-2435.12584 URL [本文引用: 1]
Effects of light intensity on Quercus liaotungensis seed germination and seedling growth
光照对辽东栎种子萌发和幼苗生长的影响
Higher plants species diversity in different types of artificial broad-leaved Korean pine forests
不同类型人工阔叶红松林高等植物物种多样性
Ecology of seed germination of eight non-pioneer tree species from a tropical seasonal rain forest in southwest China
DOI:10.1007/s11258-007-9355-0 URL [本文引用: 1]
Mechanisms for plants detecting the optimum time and place to germinate
植物调节萌发时间和萌发地点的机制
Review on influence mechanisms of light in seed germination
DOI:10.3724/SP.J.1258.2012.00899 URL [本文引用: 1]
光对种子萌发的影响机理研究进展
DOI:10.3724/SP.J.1258.2012.00899
[本文引用: 1]
种子萌发是植物成功实现天然更新的关键环节, 需要适宜的温度、水分或光照条件。对于需光性种子, 光照是决定其萌发与否或萌发率高低的主要因素。光对植物种子萌发的影响不仅是一个复杂的生理过程, 也是受到调控的信号传递和基因表达过程。该文系统总结了影响种子萌发的光照属性、光与水/热耦合作用和种子的光属性(光敏色素)与种子萌发的关系, 明确了光调控种子萌发的生态意义; 重点综述了种子内光敏色素调控种子萌发的生理反应模式和光敏色素的光信号转导途径。试图为全面评估光对种子萌发的影响和将来开展更深入的研究提供参考。
Seed germination of Pinus koraiensis Siebold & Zucc. in response to light regimes caused by shading and seed positions
DOI:10.5424/fs/2012213-02721 URL [本文引用: 1]
A review on fundamental studies of secondary forest management
次生林经营基础研究进展
次生林是中国森林的主体,在结构组成、林木生长、生产力、林分环境等诸多方面与原始林和人工林有着显著的不同.因此,经营好次生林对中国天然林保护等林业工程实施及国家生态安全建设具有重大意义.本文在广泛收集国内外有关次生林研究结果的基础上,探讨了次生林经营领域中基础研究所涉内容,揭示出次生林经营研究中所关注的基础问题,提出了次生林经营基础研究的主要内容与方向,对今后次生林经营研究和中国天然林保护等林业工程建设具有参考价值.
Effects of site preparation on emergence and early establishment of Larix olgensis in montane regions of northeastern China
DOI:10.1007/s11056-008-9097-4 URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}