


随着全球气候变化的加剧, 维持生态系统稳定性至关重要, 那么了解维持不同群落稳定性的机制则十分必要。目前已被提出的稳定性机制包括物种多样性、林分结构多样性、优势物种稳定性以及一些非生物因素影响等。
超产效应和物种异步性是建议用来解释物种多样性对生态系统过程时间稳定性的两个主要机制(Hector et al., 2010)。超产效应指的是物种丰富度通过物种之间的功能互补或选择更具生产力的物种来增加群落生物量(Thibaut & Connolly, 2013), 通过增加生物量平均值(µ)多于增加生物量标准差(σ)来增加生态系统稳定性(Hector et al., 2010)。物种异步性是物种间对波动环境的异步动态, 是生态位分化的结果(Loreau & de Mazancourt, 2008), 最近, 一些研究还表明, 物种对波动环境的异步反应驱动了物种多样性的稳定, 这些反应的种内变异引起了物种多样性的稳定(Aussenac et al., 2017)。一个相反的观点是, 优势物种, 而非物种多样性, 会提高群落的稳定性, 正如Grime (1998)的质量比假说所预测的那样。该假说认为, 包括稳定性在内的生态系统功能在短期内很大程度上受优势物种控制, 而次优势物种和稀有物种等次要植被成分可能通过影响优势物种的补充来调节长期稳定性(Grime, 1998), 一些研究(Sasaki & Lauenroth, 2011)支持了该假说。
以往对草地的研究表明, 物种丰富度、系统发育多样性、物种异步性、优势物种稳定性和环境变化对群落时间稳定性有显著影响(Hillebrand et al., 2008; Xu et al., 2015; Duffy et al., 2017; Huang et al., 2018; Aussenac et al., 2019)。森林生态系统不仅为大量的物种提供了主要栖息地, 而且为人类社会提供各种生态系统服务(Isbell, 2017; Ferreira et al., 2018)。生态系统过程通过响应环境波动和干扰而产生时间变化(García-Palacios et al., 2018; Wayne Polley & Wilsey, 2018)。在这种情况下, 研究维持森林生态系统稳定性的机制有利于维持生态系统服务和应对全球变化(Millar & Stephenson, 2015; Wayne Polley & Wilsey, 2018)。
然而, 很少有研究致力于了解这些决定因素实际上如何有助于森林群落稳定, 鉴于群落组成的变化在森林中发生得非常缓慢, 很可能在更大范围内影响群落的时间稳定性。有证据表明物种多样性增强了森林稳定性和生态系统的运作与服务(Hautier et al., 2014; Yuan et al., 2018), 物种多样性对稳定性的影响已有广泛报道, 而结构多样性的研究相对较少(Soares et al., 2016; Forrester, 2019)。结构多样性的度量, 如基尼系数(Weiner & Solbrig, 1984), 胸径的标准差和变异系数(Soares et al., 2016; Schnabel et al., 2019)已被发现能够影响森林生产力(Zhang & Chen, 2015)。此外, 物种异步性和优势物种稳定性对森林群落稳定性影响的研究还相对较少。
因此, 本研究致力于探索超产效应、林分结构、物种异步性及优势物种稳定性对森林群落稳定性的影响, 以明确维持温带天然针阔混交林中群落稳定性的主要机制。其中森林群落稳定性大多指生产力时间稳定性, 但由于有限的森林资源调查数据, 我们使用地上生物量的时间稳定性代替(以下简称群落生物量稳定性), 构建3个结构方程模型, 解释变量为超产效应(用物种丰富度来量化)、林分结构(用胸径变异系数来量化)、物种异步性和优势物种稳定性, 响应变量分别为群落生物量稳定性、µ和σ, 以比较各变量间直接与间接效应的相对大小。
1 材料和方法
1.1 研究地点与数据集
研究地点为吉林省蛟河林业实验区管理局, 位于蛟河市前进乡境内。该区域属于受季风影响的温带大陆性山地气候, 年平均气温3.8 ℃; 月平均气温最低在1月份(-18.6 ℃), 最高气温在7月份(21.7 ℃); 年降水量695.9 mm。土壤类型属于典型的山地暗棕色森林土, 土层深度在10-20 cm。研究区域由两个大样地组成: 第一个样地面积为21.12 hm2 (320 m × 660 m), 位于43.96°-43.97° N, 127.71°- 127.72° E, 该样地在2009年夏季进行初次植被调查, 并于2014和2019年夏季进行复测; 第二个样地面积为33.00 hm2 (500 m × 660 m), 位于43.96°-43.97° N, 127.7° E, 该样地在2010年夏季进行初次植被调查, 并于2015和2020年夏季进行复测。由于地理空间上比较接近, 两个样地具有类似的物种组成和环境条件, 并且都处于群落演替的中后期阶段(Wang et al., 2016)。植被类型为原始阔叶红松(Pinus koraiensis)林受到采伐干扰后形成的针阔混交林, 最后一次采伐树木活动是在50多年前进行的, 对实验样地中所有胸径(DBH) ≥1 cm的木本植物都进行了标记和测量, 并确定了物种(Zhang et al., 2012; Wang et al., 2016)。
为了探究超产效应、物种异步性和优势物种稳定性3种机制对群落生物量稳定性的影响, 我们在排除异常值后使用了样地中所有样方(20 m × 20 m)的数据来计算各项指标。
1.2 群落生物量稳定性
鉴于DBH < 1 cm的植株对林分生物量贡献极低, 研究对象为DBH ≥ 1 cm的乔木和大灌木。样地中共调查到116 611株个体, 隶属于23科38属72种。每个样方中的平均物种数为11 (范围为5-18)。
1.3 优势物种稳定性
优势物种稳定性使用生物量时间稳定性量化。将树种按照地上生物量总和由高到低进行排序, 前6名分别为五角枫(Acer pictum subsp. mono)、水曲柳(Fraxinus mandshurica)、胡桃楸(Juglans mandshurica)、红松、紫椴(Tilia amurensis)和蒙古栎(Quercus mongolica), 其生物量占比分别为16.23%、14.60%、13.38%、10.51%、8.70%和7.70%, 总和超过70%, 故将这6个树种作为优势物种。每个样方的优势物种生物量为样方内所有优势物种个体(包括随后死亡的个体)的生物量之和。使用该优势物种3次测量的生物量平均值(μd)与标准差(σd)比值计算每个样方的优势树种稳定性。
1.4 超产效应与物种异步性
超产效应是物种多样性作用机制之一, 主要通过物种丰富度对群落生物量稳定性产生影响, 因此用物种丰富度(样方中的物种数量)来量化超产效应。同时我们还计算了Faith系统发育多样性指数以增加解释性, 该指数基于物种的进化距离来量化生物多样性, 用特定面积样方中物种的分支长度总和表征。我们首先利用R软件的“phytools”包以及S.PhyloMaker函数构建包含样地中所有物种的系统发育树, 再利用“picante”包计算物种分支长度总和。
物种异步性是物种间对波动环境条件的异步动态, 使用公式(3)计算每个样方的物种异步性(Loreau & de Mazancourt, 2013)。
式中,
1.5 林分结构多样性指标
林分结构多样性主要由胸径多样性来量化。我们使用Schnabel等(2019)建议的方法, 计算了3个林分结构多样性指标: 胸径标准差(公式(4)), 胸径变异系数(公式(5))和胸径基尼系数(公式(6))。另使用每hm2的胸高断面积(m2·hm-2)来量化林分密度以代表树木在森林中的分布, 因天然林中树木大小变化十分显著, 以每hm2树木株数来衡量林分密度则会导致误差大大增加。
式中, SDD为胸径的标准差; VarD为胸径变异系数; GiniD为胸径基尼系数; N为样方内总个体数; DBHi为样方内第i个个体的胸径值;
1.6 统计分析
在进行统计分析之前, 首先对最终响应变量, 即群落生物量及其两个部分进行底数为10的对数转换, 以满足线性和正态性假设, 之后将所有变量的观测值做中心化处理, 以避免偏离正态性并使所有变量在相似范围内具有可比性。使用R软件中的“CAR”包计算方差膨胀因子(VIF), 以VIF < 5为标准来识别并去除多元回归模型中的多元共线性变量(Chen et al., 2019)。最终保留的指标为物种丰富度、胸径标准差、胸径变异系数、林分密度、物种异步性和优势物种稳定性。
使用结构方程模型检验上述变量对群落生物量稳定性的影响, 之后将群落生物量稳定性分为两个部分(μ和σ), 分别作为响应变量再次构建两个结构方程模型, 这代表了稳定性的基本机制(Craven et al., 2018)。对结构方程模型进行参数化处理, 并使用拟合优度指数(GFI), Chi-Square检验和标准化残差均方根(SRMR)测试了其拟合度。可接受模型的要求是Chi-Square检验不显著, 包括p > 0.05, GFI > 0.95和SRMR < 0.08 (Grace et al., 2016)。使用R软件 “Lavaan”包构建结构方程模型, 所有统计分析都在R 4.0.5软件中进行。结构方程模型去掉了一些拟合效果不好的变量, 最终解释变量包括物种丰富度、胸径变异系数、物种异步性和优势物种稳定性, 最后我们获得了标准化的直接、间接和总效应以及相关的p, 以便解释它们对群落生物量稳定性及其两个部分的相对影响。
为增加解释性和对比性, 构建3个多元回归模型, 响应变量仍分别为群落生物量稳定性及其两个部分, 但与结构方程模型相比, 解释变量增加了胸径标准差和林分密度。但由于多元回归模型难以考虑变量间的交互作用, 故最终结果以结构方程模型为主, 多元回归模型为辅。
2 结果
在多元线性回归中(图1), 物种丰富度与μ和σ均显著正相关, 且与前者相关性更大, 与生物量稳定性无显著相关性。林分结构中的胸径标准差也与μ和σ显著正相关, 而与生物量稳定性无显著关系; 胸径变异系数与生物量稳定性显著负相关, 与μ呈负相关关系, 与σ呈正相关关系, 但均不显著; 林分密度与生物量稳定性显著正相关, 与生物量稳定性的两部分也均呈显著正相关关系。物种异步性与生物量稳定性及其两个部分均呈显著正相关关系。优势物种稳定性与群落生物量稳定性显著正相关, 标准化后的系数最大(0.61), 且明显高于物种异步性(0.11); 与σ显著负相关, 标准化后的系数也为最大(-0.69)但与物种异步性(0.42)的差异相对较小; 与μ之间无显著关系, 且标准化后的系数较小(0.02), 明显低于物种异步性(0.61)。
图1
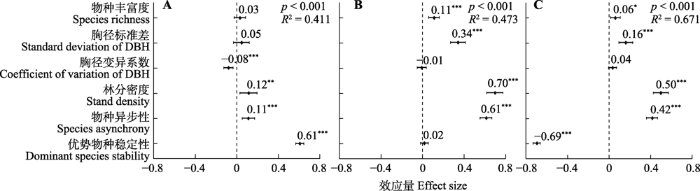
图1
多元回归模型中的预测变量对温带针阔混交林群落生物量稳定性(A)、生物量平均值(B)和生物量标准差(C)的影响(平均值±标准误)。*, p < 0.05; **, p < 0.01; ***, p < 0.001。
Fig. 1
Effect of the predictor variables on community biomass stability (A), mean biomass (B) and standard deviation of biomass (C) in a temperate coniferous and broadleaved mixed forest, from multiple regression models (mean ± SE). DBH, diameter at breast height. *, p < 0.05; **, p < 0.01; ***, p < 0.001.
结构方程模型具有良好的拟合效果, 并解释了群落生物量稳定性变化的40.6% (图2A), 当把群落生物量稳定性分别换成μ和σ后, 解释的百分比变为4.5% (图2B)和36.9% (图2C), 即后者的变化更能引起群落生物量稳定性的改变。物种异步性与群落生物量稳定性及其两个部分均呈显著正相关关系, 路径系数分别为0.055、0.085和0.055, 物种异步性主要通过增加μ来对群落生物量稳定性产生正向影响。结构方程模型的结果(图2)还表明, 物种丰富度、胸径变异系数与物种异步性均呈显著正相关关系, 路径系数分别为0.286和0.430, 并通过物种异步性间接影响群落生物量稳定性。优势物种稳定性与群落生物量稳定性呈显著正相关关系, 路径系数为0.623, 相对影响大于物种异步性; 与μ也呈显著正相关关系, 路径系数为0.148, 与物种异步性的相对影响差异不大; 与σ间显著负相关, 路径系数为-0.608, 相对影响显著大于物种异步性。总体上, 优势物种稳定性的相对影响显著大于物种异步性。此外, 物种丰富度、胸径变异系数与优势物种稳定性呈负相关关系, 但胸径变异系数与之关系显著, 路径系数为-0.065, 物种丰富度与之关系不显著, 路径系数为-0.054, 即林分结构通过优势物种稳定性对群落生物量稳定性产生了间接的负向影响。
图2
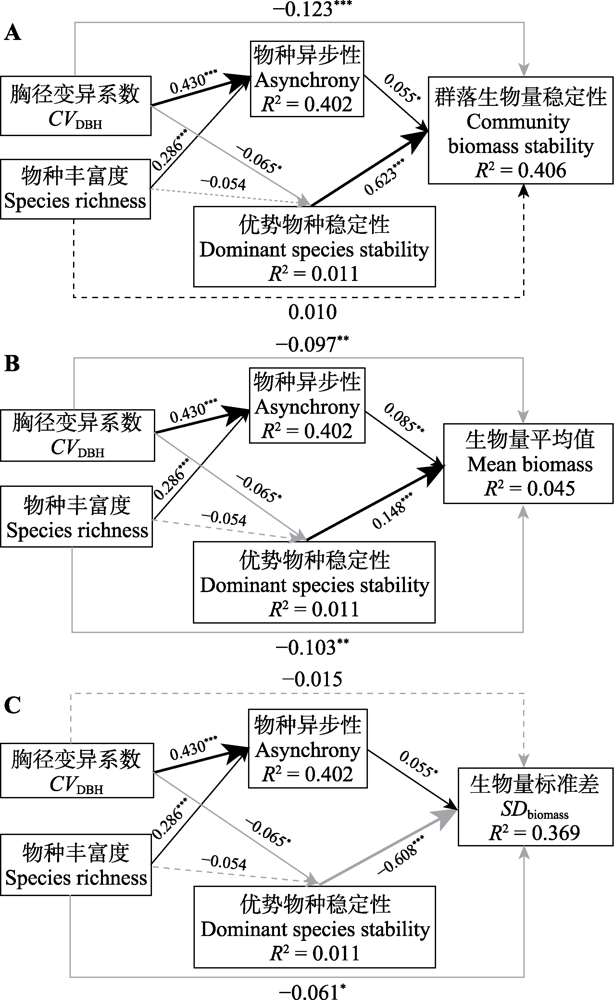
图2
用于测试物种丰富度、胸径变异系数、物种异步性和优势物种稳定性对温带针阔混交林群落生物量稳定性(A)、生物量平均值(B)和生物量标准差(C)相对影响的结构方程模型。检验参数: p = 0.692, 比较拟合指数(CFI) =1.000, 标准化均方根残差(SRMR) = 0.002。实线箭头表示显著路径, 虚线箭头表示不显著路径; 黑色为正, 灰色为负。对于每条路径, 都展示了标准化回归系数。箭头的粗细反映了标准化回归系数的大小。R2表示解释的方差比例。*, p < 0.05; **, p < 0.01; ***, p < 0.001。
Fig. 2
Structural equation models for testing the relative effects of species richness, coefficient of variation of diameter at breast height (CVDBH), species asynchrony and dominant species stability on community biomass stability (A), mean biomass (B) and standard deviation of biomass (SDbiomass)(C) in a temperate coniferous and broadleaved mixed forest. The test parameters were p = 0.692, comparative fit index (CFI) = 1.000, standardized root mean square residual (SRMR) = 0.002. Solid arrows represent significant paths and dashed arrows represent non-significant paths (black = positive; grey = negative). For each path, the standardized regression coefficient is shown. The thickness of the arrows reflects the magnitude of the standardized regression coefficients. R2 denotes the proportion of variance explained. *, p < 0.05; **, p < 0.01; ***, p < 0.001.
3 讨论
3.1 林分结构对群落生物量稳定性的影响大于超产效应
尽管超产效应的作用并不显著, 但这并不代表物种多样性不重要, 我们的结果表明, 一方面, 物种多样性通过在统计上降低μ多于降低σ, 从而对群落生物量稳定性产生直接负向影响, 在草地生态系统中也有类似的研究结果(Roscher et al., 2011); 另一方面, 物种多样性通过增加物种异步性对群落生物量稳定性产生间接正向影响, 这与大部分研究(Craven et al., 2018; Ouyang et al., 2021)一致, 但Xu等(2021)的meta分析指出, 种群平均稳定性可能随物种多样性的增加而下降, 在这种情况下增加物种异步性必须克服种群稳定性下降的问题, 以在更多样化的群落中维持良好的生态系统稳定性。并且物种多样性在一定程度上也会增加结构多样性。
3.2 物种异步性对群落生物量稳定性及其两个部分均有显著影响
物种异步性与μ和σ均呈显著正相关关系, 并且主要通过增加μ来对群落生物量稳定性产生正向影响, 这个机制可能是由于不同耐阴树种组成的群落可能具有更强的快速响应局部干扰和维持生物量稳定性的能力(Pretzsch, 2014; del Río et al., 2017), 同时物种异步性可以通过减少种间竞争和生境异质性来响应不断变化的环境条件(Loreau & de Mazancourt, 2013), 从而在一定范围内促进树木生物量的增加, 最近有研究表明大树(即林分中生物量或胸径占比超过1%的树木)在解释地上生物量存量、增长和损失方面始终比植物多样性和功能性状组成更重要(Yuan et al., 2021)。生物量大的树木通常具有更强的光捕获能力, 而优化光捕获的能力有助于物种动态的异步性(Ali et al., 2016; Dӑnescu et al., 2016)。也有研究表明物种异步性与物种间的时间互补性有关, 可以通过降低σ产生稳定影响(Loreau & de Mazancourt, 2013)。但由于森林生态系统中树木对环境变化响应较慢, 物种异步性的作用有时候并不显著(Yu et al., 2020)。还有研究使用年度数据来表明树种异步性是群落稳定性一个强大的驱动因子(Schnabel et al., 2019), 树种异步性在短时间范围内保障了稳定性(Aussenac et al., 2019), 证明物种异步性对稳定性的这些不同效果可归因于调查的时间尺度和森林类型(Yu et al., 2020)。此外, 尽管人们认为物种异步性对于稳定生态系统特性很重要, 但其在群落中普遍存在的经验证据是模棱两可的(Valencia et al., 2020), 基于这些结果, 我们建议未来的研究应测试物种异步性对不同天然森林类型和不同时间尺度稳定性的影响。
3.3 优势物种稳定性是直接影响群落生物量稳定性的最主要因素
虽然物种异步性对群落生物量稳定性具有显著影响, 但优势物种稳定性的影响相对更大, 是直接影响群落生物量稳定性的最主要因素。这与前人的研究结果(Hillebrand et al., 2008; Yang et al., 2011)一致, 当生物量在物种间均匀分布时, 组合效应对于高物种均匀度的群落尤其重要, 但当群落被几个优势物种所支配时, 这些优势种的时间变化很可能是群落生物量稳定性更重要的促进因素, 因为均匀度通常比物种丰富度等其他生态系统属性对环境变化的响应更快, 这可能导致生态系统功能更快速的响应(Chapin III et al., 2000)。此外, 响应环境条件变化的种群稳定性的变化可能转化为群落水平稳定性的变化, 特别是当群落少数物种占极大优势时(Wayne Polley et al., 2007; Xu et al., 2015)。
由于树木的寿命和生物量产生所需的时间较长, 森林生态系统的物种组成的变化相对缓慢, 这削弱了物种异步性作为稳定性促进因素的重要性(Jucker et al., 2014)。优势物种通常是森林群落中最主要、最常见和最多的物种, 可能通过对其他植物物种的出现、大小、分布或丰度施加强烈影响而决定森林结构、多样性和生物量(Hillebrand et al., 2008; Rodrigues et al., 2019), 因为在质量比效应的假设下, 优势物种可能会减少物种多样性对生物量的影响(Hillebrand et al., 2008; Chiang et al., 2016; Ali & Yan, 2017), 即优势物种直接通过促进生物量来影响群落生物量稳定性。优势物种通常包含林分中更高比例的大树, 通常也具有较大的叶面积指数, 使它们能够通过光合作用拦截更多的光并固存更多的碳(Michaletz et al., 2014), 且净生物量的精细空间变化很大程度上取决于大型成熟树木死亡导致的生物量损失(Yuan et al., 2019)。此外, 优势物种对气候变化和土壤条件的反应可能会影响天然林中其他剩余物种的分布、丰度和优势度(Figueiredo et al., 2018; Ali et al., 2020)。
3.4 影响和局限性
值得注意的是, 森林生态系统稳定性或群落生物量稳定性大多指生产力时间稳定性, 但由于有限的森林资源调查数据, 我们研究了地上生物量的时间稳定性, 而不是生产力随时间的变化。具体而言, 计算森林生产力和地上生物量的时间稳定性都需要较长的时间重复才能尽量减小误差, 且森林生产力稳定性的计算至少需要4次重复的森林调查数据。目前, 地上生物量也经常被看作重要森林功能(Conti & Díaz, 2013), 因此是天然森林生产力的理想代表(Lohbeck et al., 2015; Yuan et al., 2018)。另外, 天然森林的地上生物量和生产力在所有空间尺度上都是正相关的(Chisholm et al., 2013)。然而, 进一步的研究可能需要利用森林生产力的时间稳定性来阐明和确认这些复杂的路径关系。尽管如此, 这项研究展示了超产效应、胸径结构、物种异步性和优势物种稳定性对天然林地上生物量时间稳定性的影响, 证明了质量比效应在温带针阔混交林中的重要作用。
4 结论
超产效应对温带针阔混交林群落稳定性的影响并不显著, 更多的是物种丰富度通过其他机制产生间接影响, 而林分结构的影响显著大于超产效应。尽管物种异步性和优势树种稳定性均对温带针阔混交林群落生物量稳定性有显著影响, 但优势物种稳定性是直接影响群落生物量稳定性的主要因素, 我们强调优势树种而不是物种异步性或超产效应在维持森林生态系统稳定性中的重要作用, 为森林生态系统维稳措施提供了一定的理论基础。此外, 各种机制与群落生物量稳定性间的关系是不同且复杂的, 其复杂性很可能源于各机制和稳定性的不同定义、时间空间尺度、森林类型、森林演替阶段以及生态系统受到干扰的背景等, 未来我们需要在不同研究背景下探索更准确和更全面的机制。
附录I 温带针阔混交林各树种生物量计算公式
Supplement I Biomass calculation formula of each tree species in a temperate coniferous and broadleaved mixed forest
参考文献
Impacts of climatic and edaphic factors on the diversity, structure and biomass of species-poor and structurally-complex forests
DOI:10.1016/j.scitotenv.2019.135719 URL [本文引用: 1]
Functional identity of overstorey tree height and understorey conservative traits drive aboveground biomass in a subtropical forest
DOI:10.1016/j.ecolind.2017.07.054 URL [本文引用: 1]
Stand structural diversity rather than species diversity enhances aboveground carbon storage in secondary subtropical forests in Eastern China
DOI:10.5194/bg-13-4627-2016 URL [本文引用: 1]
Intraspecific variability in growth response to environmental fluctuations modulates the stabilizing effect of species diversity on forest growth
DOI:10.1111/1365-2745.12728 URL [本文引用: 1]
Interactions among trees: a key element in the stabilising effect of species diversity on forest growth
DOI:10.1111/1365-2435.13257
[本文引用: 2]

There is mounting evidence that species diversity increases the temporal stability of forest growth. This stabilising effect of diversity has mainly been attributed to species differences in their response to fluctuating environmental conditions. Interactions among individuals could also contribute to the stabilising effect of diversity by increasing the mean and reducing the variance of tree growth, however, this has never been directly demonstrated. We used tree-ring width chronologies from temperate and boreal mixed stands of Eastern Canada to identify the role of interactions among individuals in the stabilising effect of diversity on forest growth. Using neighbourhood competition index and a mixed model, we compared the effect of interspecific and intraspecific interactions on the mean and the variance of tree growth. We found that interspecific interactions are less detrimental to tree growth than intraspecific interactions. We also found that interspecific interactions buffer tree response to drought and thereby reduce the variance of tree growth. Our results indicate diversity may increase the mean and reduce the variance of tree growth through interactions among individuals. Thus, we demonstrate interactions among individuals play a role in the stabilising effect of diversity on forest growth, and in doing so, we bring to light other mechanisms of the insurance hypothesis.
Consequences of changing biodiversity
DOI:10.1038/35012241 URL [本文引用: 1]
Embracing scale-dependence to achieve a deeper understanding of biodiversity and its change across communities
DOI:10.1111/ele.13151 URL [本文引用: 1]
Tree species identity surpasses richness in affecting soil microbial richness and community composition in subtropical forests
DOI:10.1016/j.soilbio.2018.12.008 URL [本文引用: 1]
Functional composition drives ecosystem function through multiple mechanisms in a broadleaved subtropical forest
DOI:10.1007/s00442-016-3717-z URL [本文引用: 1]
Scale-dependent relationships between tree species richness and ecosystem function in forests
DOI:10.1111/1365-2745.12132 URL [本文引用: 1]
Plant functional diversity and carbon storage-An empirical test in semi-arid forest ecosystems
DOI:10.1111/1365-2745.12012 URL [本文引用: 1]
Multiple facets of biodiversity drive the diversity-stability relationship
Structural diversity promotes productivity of mixed, uneven-aged forests in southwestern Germany
DOI:10.1007/s00442-016-3623-4
PMID:27059713
[本文引用: 1]
Forest diversity-productivity relationships have been intensively investigated in recent decades. However, few studies have considered the interplay between species and structural diversity in driving productivity. We analyzed these factors using data from 52 permanent plots in southwestern Germany with more than 53,000 repeated tree measurements. We used basal area increment as a proxy for productivity and hypothesized that: (1) structural diversity would increase tree and stand productivity, (2) diversity-productivity relationships would be weaker for species diversity than for structural diversity, and (3) species diversity would also indirectly impact stand productivity via changes in size structure. We measured diversity using distance-independent indices. We fitted separate linear mixed-effects models for fir, spruce and beech at the tree level, whereas at the stand level we pooled all available data. We tested our third hypothesis using structural equation modeling. Structural and species diversity acted as direct and independent drivers of stand productivity, with structural diversity being a slightly better predictor. Structural diversity, but not species diversity, had a significant, albeit asymmetric, effect on tree productivity. The functioning of structurally diverse, mixed forests is influenced by both structural and species diversity. These sources of trait diversity contribute to increased vertical stratification and crown plasticity, which in turn diminish competitive interferences and lead to more densely packed canopies per unit area. Our research highlights the positive effects of species diversity and structural diversity on forest productivity and ecosystem dynamics.
Species interactions increase the temporal stability of community productivity in Pinus sylvestris - Fagus sylvatica mixtures across Europe
DOI:10.1111/1365-2745.12727 URL [本文引用: 1]
Biodiversity effects in the wild are common and as strong as key drivers of productivity
DOI:10.1038/nature23886 URL [本文引用: 1]
Carbon-focused conservation may fail to protect the most biodiverse tropical forests
DOI:10.1038/s41558-018-0225-7
[本文引用: 1]
As one of Earth's most carbon-dense regions, tropical forests are central to climate change mitigation efforts. Their unparalleled species richness also makes them vital for safeguarding biodiversity. However, because research has not been conducted at management-relevant scales and has often not accounted for forest disturbance, the biodiversity implications of carbon conservation strategies remain poorly understood. We investigated tropical carbon-biodiversity relationships and trade-offs along a forest-disturbance gradient, using detailed and extensive carbon and biodiversity datasets. Biodiversity was positively associated with carbon in secondary and highly disturbed primary forests. Positive carbon-biodiversity relationships dissipated at around 100 MgC ha(-1), meaning that in less disturbed forests more carbon did not equal more biodiversity. Simulated carbon conservation schemes therefore failed to protect many species in the most species-rich forests. These biodiversity shortfalls were sensitive to opportunity costs and could be decreased for small carbon penalties. To ensure that the most ecologically valuable forests are protected, biodiversity needs to be incorporated into carbon conservation planning.
Beyond climate control on species range: the importance of soil data to predict distribution of Amazonian plant species
DOI:10.1111/jbi.13104 URL [本文引用: 1]
Linking forest growth with stand structure: tree size inequality, tree growth or resource partitioning and the asymmetry of competition
DOI:10.1016/j.foreco.2019.05.053
[本文引用: 2]
Stand structure can strongly influence forest growth and other processes, such as the water balance, carbon partitioning, nutrient cycling and light dynamics. However, individual structural variables can be positively or negatively correlated with growth. This is the case for variables such as size inequality and those that describe resource partitioning, such as the degree of symmetric/asymmetric competition and growth dominance. Several contrasting growth-structure correlations are reviewed and linked to forest processes by considering the different types of tree interactions they are associated with. Contrasting growth-structure correlations appear to converge when they are examined using a simple framework where stand growth is a function of three variables as opposed to any one of the variables alone; stand density, size distributions and tree size-growth relationships. The size distributions quantify how the stand density is distributed between the different sizes while the size growth relationships quantify how growth is partitioned between different sizes. Size inequality may not often be a useful explanatory variable and instead it appears to sometimes correlate with growth because it can be correlated with other variables that influence growth. The spatial and temporal dynamics of the effects of structure on growth have received little attention and a long-term growth and yield data set from central Europe was used to examine how the effects of structure can change along climatic gradients. The simple framework of three variables could be used to separate the effects of structure and functioning when comparing mixed and monospecific forests, as well as to design silvicultural interventions or to determine whether past management interventions have achieved their goals. The implications for selecting which structural variables to use and when scaling up to the stand level, are also discussed.
Tree-species interactions increase light absorption and growth in Chinese subtropical mixed-species plantations
DOI:10.1007/s00442-019-04495-w
PMID:31463782
[本文引用: 1]
Light-related interactions can increase productivity in tree-species mixtures compared with monocultures due to higher stand-level absorption of photosynthetically active radiation (APAR) or light-use efficiency (LUE). However, the effects of different light-related interactions, and their relative importance, have rarely been quantified. Here, measurements of vertical leaf-area distributions, tree sizes, and stand density were combined with a tree-level light model (Maestra) to examine how crown architecture and vertical or horizontal canopy structure influence the APAR of 16 monocultures and eight different two-species mixtures with 16 different species in a Chinese subtropical tree diversity experiment. A higher proportion of crown leaf area occurred in the upper crowns of species with higher specific leaf areas. Tree-level APAR depended largely on tree leaf area and also, but to a lesser extent, on relative height (i.e., tree dominance) and leaf-area index (LAI). Stand-level APAR depended on LAI and canopy volume, but not on the vertical stratification or canopy leaf-area density. The mixing effects, in terms of relative differences between mixtures and monocultures, on stand-level APAR were correlated with the mixing effects on basal area growth, indicating that light-related interactions may have been responsible for part of the mixing effects on basal area growth. While species identity influences the vertical distributions of leaf area within tree crowns, this can have a relatively small effect on tree and stand APAR compared with the size and vertical positioning of the crowns, or the LAI and canopy volume.
Climate mediates the biodiversity-ecosystem stability relationship globally
DOI:10.1073/pnas.1800425115
PMID:30061405
[本文引用: 1]
The insurance hypothesis, stating that biodiversity can increase ecosystem stability, has received wide research and political attention. Recent experiments suggest that climate change can impact how plant diversity influences ecosystem stability, but most evidence of the biodiversity-stability relationship obtained to date comes from local studies performed under a limited set of climatic conditions. Here, we investigate how climate mediates the relationships between plant (taxonomical and functional) diversity and ecosystem stability across the globe. To do so, we coupled 14 years of temporal remote sensing measurements of plant biomass with field surveys of diversity in 123 dryland ecosystems from all continents except Antarctica. Across a wide range of climatic and soil conditions, plant species pools, and locations, we were able to explain 73% of variation in ecosystem stability, measured as the ratio of the temporal mean biomass to the SD. The positive role of plant diversity on ecosystem stability was as important as that of climatic and soil factors. However, we also found a strong climate dependency of the biodiversity-ecosystem stability relationship across our global aridity gradient. Our findings suggest that the diversity of leaf traits may drive ecosystem stability at low aridity levels, whereas species richness may have a greater stabilizing role under the most arid conditions evaluated. Our study highlights that to minimize variations in the temporal delivery of ecosystem services related to plant biomass, functional and taxonomic plant diversity should be particularly promoted under low and high aridity conditions, respectively.
Integrative modelling reveals mechanisms linking productivity and plant species richness
DOI:10.1038/nature16524 URL [本文引用: 1]
Benefits of plant diversity to ecosystems: immediate, filter and founder effects
DOI:10.1046/j.1365-2745.1998.00306.x URL [本文引用: 2]
Eutrophication weakens stabilizing effects of diversity in natural grasslands
DOI:10.1038/nature13014 URL [本文引用: 1]
Allometric biomass equations for 12 tree species in coniferous and broadleaved mixed forests, Northeastern China
DOI:10.1371/journal.pone.0186226 URL [本文引用: 1]
General stabilizing effects of plant diversity on grassland productivity through population asynchrony and overyielding
Insurance effects of biodiversity can stabilize the functioning of multispecies ecosystems against environmental variability when differential species' responses lead to asynchronous population dynamics. When responses are not perfectly positively correlated, declines in some populations are compensated by increases in others, smoothing variability in ecosystem productivity. This variance reduction effect of biodiversity is analogous to the risk-spreading benefits of diverse investment portfolios in financial markets. We use data from the BIODEPTH network of grassland biodiversity experiments to perform a general test for stabilizing effects of plant diversity on the temporal variability of individual species, functional groups, and aggregate communities. We tested three potential mechanisms: reduction of temporal variability through population asynchrony; enhancement of long-term average performance through positive selection effects; and increases in the temporal mean due to overyielding. Our results support a stabilizing effect of diversity on the temporal variability of grassland aboveground annual net primary production through two mechanisms. Two-species communities with greater population asynchrony were more stable in their average production over time due to compensatory fluctuations. Overyielding also stabilized productivity by increasing levels of average biomass production relative to temporal variability. However, there was no evidence for a performance-enhancing effect on the temporal mean through positive selection effects. In combination with previous work, our results suggest that stabilizing effects of diversity on community productivity through population asynchrony and overyielding appear to be general in grassland ecosystems.
Consequences of dominance: a review of evenness effects on local and regional ecosystem processes
The composition of communities is strongly altered by anthropogenic manipulations of biogeochemical cycles, abiotic conditions, and trophic structure in all major ecosystems. Whereas the effects of species loss on ecosystem processes have received broad attention, the consequences of altered species dominance for emergent properties of communities and ecosystems are poorly investigated. Here we propose a framework guiding our understanding of how dominance affects species interactions within communities, processes within ecosystems, and dynamics on regional scales. Dominance (or the complementary term, evenness) reflects the distribution of traits in a community, which in turn affects the strength and sign of both intraspecifc and interspecific interactions. Consequently, dominance also mediates the effect of such interactions on species coexistence. We review the evidence for the fact that dominance directly affects ecosystem functions such as process rates via species identity (the dominant trait) and evenness (the frequency distribution of traits), and indirectly alters the relationship between process rates and species richness. Dominance also influences the temporal and spatial variability of aggregate community properties and compositional stability (invasibility). Finally, we propose that dominance affects regional species coexistence by altering metacommunity dynamics. Local dominance leads to high beta diversity, and rare species can persist because of source-sink dynamics, but anthropogenically induced environmental changes result in regional dominance and low beta diversity, reducing regional coexistence. Given the rapid anthropogenic alterations of dominance in many ecosystems and the strong implications of these changes, dominance should be considered explicitly in the analysis of consequences of altered biodiversity.
Impacts of species richness on productivity in a large-scale subtropical forest experiment
DOI:10.1126/science.aat6405 URL [本文引用: 1]
Linking the influence and dependence of people on biodiversity across scales
DOI:10.1038/nature22899 URL [本文引用: 1]
Taxonomic and functional diversity change is scale dependent
DOI:10.1038/s41467-018-04889-z
PMID:29967400
[本文引用: 1]
Estimates of recent biodiversity change remain inconsistent, debated, and infrequently assessed for their functional implications. Here, we report that spatial scale and type of biodiversity measurement influence evidence of temporal biodiversity change. We show a pervasive scale dependence of temporal trends in taxonomic (TD) and functional (FD) diversity for an similar to 50-year record of avian assemblages from North American Breeding Bird Survey and a record of global extinctions. Average TD and FD increased at all but the global scale. Change in TD exceeded change in FD toward large scales, signaling functional resilience. Assemblage temporal dissimilarity and turnover (replacement of species or functions) declined, while nestedness (tendency of assemblages to be subsets of one another) increased with scale. Patterns of FD change varied strongly among diet and foraging guilds. We suggest that monitoring, policy, and conservation require a scale-explicit framework to account for the pervasive effect that scale has on perceived biodiversity change.
Stabilizing effects of diversity on aboveground wood production in forest ecosystems: linking patterns and processes
DOI:10.1111/ele.12382 URL [本文引用: 2]
Biomass is the main driver of changes in ecosystem process rates during tropical forest succession
DOI:10.1890/14-0472.1 URL [本文引用: 1]
Species synchrony and its drivers: neutral and nonneutral community dynamics in fluctuating environments
DOI:10.1086/589746 URL [本文引用: 1]
Biodiversity and ecosystem stability: a synthesis of underlying mechanisms
DOI:10.1111/ele.12073 URL [本文引用: 3]
Convergence of terrestrial plant production across global climate gradients
DOI:10.1038/nature13470 URL [本文引用: 1]
Temperate forest health in an era of emerging megadisturbance
DOI:10.1126/science.aaa9933 URL [本文引用: 1]
Temporal stability in forest productivity increases with tree diversity due to asynchrony in species dynamics
DOI:10.1111/ele.12357 URL [本文引用: 1]
Stability in subtropical forests: the role of tree species diversity, stand structure, environmental and socio- economic conditions
DOI:10.1111/geb.13235 URL [本文引用: 1]
Canopy space filling and tree crown morphology in mixed-species stands compared with monocultures
DOI:10.1016/j.foreco.2014.04.027 URL [本文引用: 1]
Fine-scale topography shape richness, community composition, stem and biomass hyperdominant species in Brazilian Atlantic forest
DOI:10.1016/j.ecolind.2019.02.033
[本文引用: 1]
Topography is an important driver that determines diversity patterns and ecosystem functioning in tropical forests. However, there are few studies analyzing contrasting topographical conditions on the relative importance of species to ecosystem functioning, mainly on those who have a greater contribution (i.e., hyperdominant species). We aimed to evaluate whether contrasting topographical conditions determine changes in tree species richness, community composition, and the number of stem and biomass hyperdominant species in a Brazilian Atlantic remnant forest fragment. We selected two areas on distinct hillsides with contrasting topographic conditions, at the biological reserve of the Federal University of Vicosa, Minas Gerais state, southeastern Brazil. Each area (100x100 m) was sub-divided into 100 plots of 10x10 m. From each plot, all trees having diameter at breast height (DBH) >= 10 cm were identified to the species level and tagged for measurement. We measured three topographic variables (slope, elevation, and convexity) in each plot, based on the assumption that these variables may affect tree species diversity, species composition, and ecosystem function (aboveground biomass). The AGB of individual stems was calculated in all plots. We performed a multivariate regression tree for estimating topographical heterogeneity in each area. We found that species richness differed significantly between areas. Species richness in the Northeast area (the more topographically heterogeneous one) was 48% higher than that in the Southeast area, which is less topographically heterogeneous. The tree species composition varied considerably between areas, with similar AGB patterns being registered among plots. The number of stem hyperdominants varied significantly between areas. In the Southeast area, only two species out of the 85 recorded (2.38%) accounted for 50% of the number of stems hyperdominants, while in the Northeast area 10 species (7.94%) accounted for 50% of stems hyperdominants. Our results showed that high topographic heterogeneity induces high species richness and that the number of stem and biomass hyperdominant species increase along with richness on a local scale. Based on our results, we presume that biomass hyperdominance can also strongly influence forest ecosystem functioning on a local scale.
Identifying population and community-level mechanisms of diversity-stability relationships in experimental grasslands
DOI:10.1111/j.1365-2745.2011.01875.x URL [本文引用: 1]
Dominant species, rather than diversity, regulates temporal stability of plant communities
DOI:10.1007/s00442-011-1916-1 URL [本文引用: 1]
Drivers of productivity and its temporal stability in a tropical tree diversity experiment
DOI:10.1111/gcb.14792
PMID:31486578
[本文引用: 3]
There is increasing evidence that mixed-species forests can provide multiple ecosystem services at a higher level than their monospecific counterparts. However, most studies concerning tree diversity and ecosystem functioning relationships use data from forest inventories (under noncontrolled conditions) or from very young plantation experiments. Here, we investigated temporal dynamics of diversity-productivity relationships and diversity-stability relationships in the oldest tropical tree diversity experiment. Sardinilla was established in Panama in 2001, with 22 plots that form a gradient in native tree species richness of one-, two-, three- and five-species communities. Using annual data describing tree diameters and heights, we calculated basal area increment as the proxy of tree productivity. We combined tree neighbourhood- and community-level analyses and tested the effects of both species diversity and structural diversity on productivity and its temporal stability. General patterns were consistent across both scales indicating that tree-tree interactions in neighbourhoods drive observed diversity effects. From 2006 to 2016, mean overyielding (higher productivity in mixtures than in monocultures) was 25%-30% in two- and three-species mixtures and 50% in five-species stands. Tree neighbourhood diversity enhanced community productivity but the effect of species diversity was stronger and increased over time, whereas the effect of structural diversity declined. Temporal stability of community productivity increased with species diversity via two principle mechanisms: asynchronous responses of species to environmental variability and overyielding. Overyielding in mixtures was highest during a strong El Niño-related drought. Overall, positive diversity-productivity and diversity-stability relationships predominated, with the highest productivity and stability at the highest levels of diversity. These results provide new insights into mixing effects in diverse, tropical plantations and highlight the importance of analyses of temporal dynamics for our understanding of the complex relationships between diversity, productivity and stability. Under climate change, mixed-species forests may provide both high levels and high stability of production.© 2019 The Authors. Global Change Biology published by John Wiley & Sons Ltd.
Increasing stand structural heterogeneity reduces productivity in Brazilian Eucalyptus monoclonal stands
DOI:10.1016/j.foreco.2016.04.035 URL [本文引用: 2]
Understanding diversity- stability relationships: towards a unified model of portfolio effects
DOI:10.1111/ele.12019
PMID:23095077
[本文引用: 1]
A major ecosystem effect of biodiversity is to stabilise assemblages that perform particular functions. However, diversity-stability relationships (DSRs) are analysed using a variety of different population and community properties, most of which are adopted from theory that makes several restrictive assumptions that are unlikely to be reflected in nature. Here, we construct a simple synthesis and generalisation of previous theory for the DSR. We show that community stability is a product of two quantities: the synchrony of population fluctuations, and an average species-level population stability that is weighted by relative abundance. Weighted average population stability can be decomposed to consider effects of the mean-variance scaling of abundance, changes in mean abundance with diversity and differences in species' mean abundance in monoculture. Our framework makes explicit how unevenness in the abundances of species in real communities influences the DSR, which occurs both through effects on community synchrony, and effects on weighted average population variability. This theory provides a more robust framework for analysing the results of empirical studies of the DSR, and facilitates the integration of findings from real and model communities.© 2012 Blackwell Publishing Ltd/CNRS.
Synchrony matters more than species richness in plant community stability at a global scale
DOI:10.1073/pnas.1920405117
PMID:32900958
[本文引用: 1]
The stability of ecological communities is critical for the stable provisioning of ecosystem services, such as food and forage production, carbon sequestration, and soil fertility. Greater biodiversity is expected to enhance stability across years by decreasing synchrony among species, but the drivers of stability in nature remain poorly resolved. Our analysis of time series from 79 datasets across the world showed that stability was associated more strongly with the degree of synchrony among dominant species than with species richness. The relatively weak influence of species richness is consistent with theory predicting that the effect of richness on stability weakens when synchrony is higher than expected under random fluctuations, which was the case in most communities. Land management, nutrient addition, and climate change treatments had relatively weak and varying effects on stability, modifying how species richness, synchrony, and stability interact. Our results demonstrate the prevalence of biotic drivers on ecosystem stability, with the potential for environmental drivers to alter the intricate relationship among richness, synchrony, and stability.
Relationships between tree biomass productivity and local species diversity
DOI:10.1002/ecs2.1562 [本文引用: 2]
Biodiversity and ecosystem stability across scales in metacommunities
DOI:10.1111/ele.12582 URL [本文引用: 1]
Variability in community productivity-Mediating effects of vegetation attributes
DOI:10.1111/1365-2435.13080 URL [本文引用: 2]
Dominant species constrain effects of species diversity on temporal variability in biomass production of tallgrass prairie
DOI:10.1111/j.2007.0030-1299.16080.x URL [本文引用: 1]
The meaning and measurement of size hierarchies in plant populations
DOI:10.1007/BF00379630
PMID:28311058
[本文引用: 1]
The term "size hierarchy" has been used frequently by plant population biologists but it has not been defined. Positive skewness of the size distribution, which has been used to evaluate size hierarchies, is inappropriate. We suggest that size hierarchy is equivalent to size inequality. Methods developed by economists to evaluate inequalities in wealth and income, the Lorenz curve and Gini Coefficient, provide a useful quantification of inequality and allow us to compare populations. A measure of inequality such as the Gini Coefficient will usually be more appropriate than a measure of skewness for addressing questions concerning plant population structure.
Consistently positive effect of species diversity on ecosystem, but not population, temporal stability
DOI:10.1111/ele.13777 URL [本文引用: 1]
Environmental changes drive the temporal stability of semi-arid natural grasslands through altering species asynchrony
DOI:10.1111/1365-2745.12441 URL [本文引用: 2]
The effects of long-term fertilization on the temporal stability of alpine meadow communities
DOI:10.1007/s11104-011-0784-0 URL [本文引用: 1]
Species dominance rather than species asynchrony determines the temporal stability of productivity in four subtropical forests along 30 years of restoration
DOI:10.1016/j.foreco.2019.117687 URL [本文引用: 2]
Multiple abiotic and biotic pathways shape biomass demographic processes in temperate forests
DOI:10.1002/ecy.2650 URL [本文引用: 1]
Few large trees, rather than plant diversity and composition, drive the above-ground biomass stock and dynamics of temperate forests in northeast China
DOI:10.1016/j.foreco.2020.118698 URL [本文引用: 1]
Abiotic and biotic determinants of coarse woody productivity in temperate mixed forests
DOI:10.1016/j.scitotenv.2018.02.125 URL [本文引用: 2]
Species- habitat associations in a northern temperate forest in China
Individual size inequality links forest diversity and above-ground biomass
DOI:10.1111/1365-2745.12425 URL [本文引用: 1]
Nitrogen addition does not reduce the role of spatial asynchrony in stabilising grassland communities
DOI:10.1111/ele.13212 URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}