



土壤是陆地生态系统最大的碳库, 超过植被和大气的碳库之和, 土壤碳库的微小变化都会对大气CO2浓度和全球气候变化产生巨大的影响, 在调节全球碳循环和气候变化中起着重要的作用(Lal, 2004)。土壤有机碳及其对全球变化的响应是生物地球化学和全球变化领域的研究热点, 受到国内外学者的广泛关注(Jackson et al., 2017; Lajtha et al., 2018; Lu et al., 2021a)。土壤有机碳库的变化主要取决于植物以地上凋落物和根系分泌物与残体的形式向土壤的碳输入过程和微生物分解土壤有机碳产生的碳输出过程。植物向土壤的碳输入过程受到植物生长与生产力、凋落物生产和地下碳分配等因素的影响, 碳输出过程主要受到土壤微生物的调控, 如微生物生物量、碳利用效率(CUE)、酶活性和群落组成等(Cusack et al., 2011a; 李嵘和常瑞英, 2015; Riggs & Hobbie, 2016)。因此, 在外界干扰下, 植物生长和土壤微生物分解活性等与碳输入和输出密切相关过程的变化会对土壤有机碳库的动态变化产生影响。
土壤有机碳的组成成分复杂, 在全球变化等外界因素的影响下, 其变化不仅体现在碳库大小的变化上, 也体现在其组分和分子组成等物理和化学属性的变化上。土壤有机碳库由代表着不同稳定机制及周转速率的多种功能碳库所构成, 一般而言, 可将其区分为活性有机碳库和稳定性有机碳库(Six et al., 2002; von Lützow et al., 2007; Lavallee et al., 2020)。同样地, 土壤有机碳也是由具有不同化学结构的有机碳分子所构成, 它们具有不同的稳定性和降解速度。全球变化要素如氮(N)、磷(P)沉降等可以通过化学反应作用、改变植物地上和地下凋落物的质量以及改变微生物周转和底物利用偏好等来改变土壤有机碳的组分和分子组成(Janssens et al., 2010; Mori et al., 2018)。因此, 了解并剖析土壤有机碳的碳库大小、组分和分子组成的变化对于准确预测土壤有机碳动态具有重要作用。
N和P被认为是限制植物生长和生态系统生产力的两种重要元素(Elser et al., 2007; Vitousek et al., 2010)。自工业革命以来, 人为活动如化石燃料燃烧、农田矿质肥料施用以及大气N、P沉降等极大地增加了陆地生态系统的N、P输入(Galloway et al., 2004; Peñuelas et al., 2012)。据估计, 从19世纪60年代到20世纪90年代人类活动导致的N输入量已从15 Tg·a-1上升到156 Tg·a-1, 预计到2050年将增加到270 Tg·a-1 (Galloway et al., 2004)。与此类似, 人类活动已经使全球P输入量由工业革命前的0.3 Tg·a-1增加到2005-2010年间的14-16 Tg·a-1, 并将持续增加(Peñuelas et al., 2012)。氮磷输入和沉降及其所导致的养分可利用性增加和养分不平衡对区域和全球陆地生态系统碳循环产生了显著的影响, 如改变植被生长与生产力、微生物活性及其对土壤有机碳的分解作用(Feng & Zhu, 2019; Xu et al., 2021a)。氮磷富集所导致的这些植物和微生物的变化会对土壤有机碳动态产生影响, 如改变其碳库大小、组分和分子组成。
因此, 探究氮磷添加对土壤有机碳动态的影响对于理解碳与养分之间的耦合作用以及准确预测陆地生态系统碳循环动态及其与气候变化之间的反馈至关重要。目前, 在全球范围内已经开展了很多关于氮磷添加对土壤有机碳影响的案例研究, 但是不同研究的结果存在很大的差异, 受到诸多因素的影响与调控。为此, 本文以氮磷添加对土壤碳输入和输出过程的影响为切入点, 系统综述了氮磷添加(N添加、P添加和氮磷共同(N + P)添加)对土壤有机碳的碳库大小、组分和分子组成的影响模式与调控机理, 指出了当前研究中存在的不足, 并提出了未来的研究建议。
1 氮磷添加对土壤碳输入和输出过程的影响
1.1 氮磷添加对土壤碳输入过程的影响
植物碳输入是土壤有机碳的主要来源, 对土壤有机碳的形成与积累至关重要, 氮磷添加可直接通过影响地上凋落物和地下根系碳输入过程而调控土壤有机碳动态。研究表明, 氮磷添加对森林、草地和农田等各类陆地生态系统的地上生产力具有显著的促进作用, 且N添加引起的促进作用在北方森林更为强烈, 而P添加导致的促进作用在热带气候区的生态系统(主要为热带森林)中更为强烈(Li et al., 2016; Čapek et al., 2018; Schulte-Uebbing & de Vries, 2018; Hou et al., 2020)。这种N、P添加对陆地生态系统生产力的促进结果可能和植被生产力养分限制的纬度格局有关(Du et al., 2020; 冯继广和朱彪, 2020; Hou et al., 2021), 即高纬度地区主要为N限制, 低纬度地区主要为P限制(Vitousek, 1984; Vitousek et al., 2010; Norby et al., 2016)。
地上凋落物及其分解产生的可溶性有机碳输入是土壤有机碳的一个重要来源途径(Cotrufo et al., 2015; Lu et al., 2021b)。研究表明, N、P添加对不同养分状况下的生态系统的凋落物生产具有不同的影响。一般而言, 在N或P限制的生态系统, 相应的N或P添加能够通过提高植被生产力而促进凋落物生产; 而在N或P不受限制的生态系统, N或P添加并不会对植物生产力和凋落物生产产生显著的影响(Janssens et al., 2010; Feng & Zhu, 2019; Wright, 2019; Xu et al., 2021a)。此外, 研究发现, 土壤可溶性有机碳对N添加的响应和凋落物生产对N添加的响应存在显著的正相关关系, 说明凋落物的输入能够显著地提高土壤可溶性有机碳的含量, 进而提高土壤有机碳含量(图1; Xu et al., 2021a, 2021b)。
图1
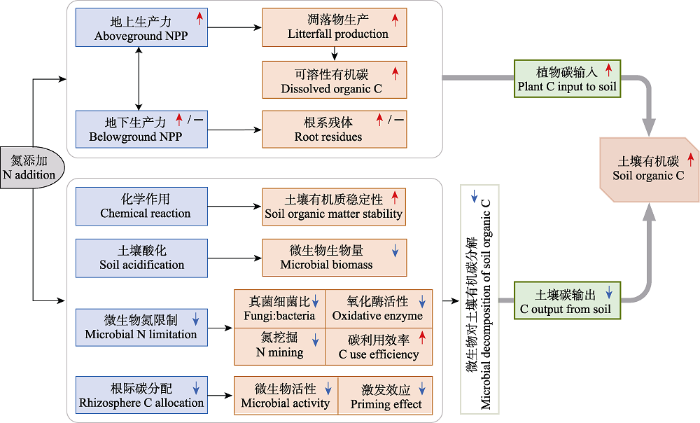
图1
氮添加促进土壤有机碳积累的潜在机制。框内向上箭头表示增加, 向下箭头表示降低, 横线表示没有响应, 这些符号表示不同研究的总体响应情况。
Fig. 1
Potential mechanisms of how nitrogen (N) addition stimulates soil organic carbon (C) accumulation. In the box, upward arrows indicate increase, downward arrows indicate decrease, short horizontal lines indicate no response, and these symbols indicate the overall responses across studies. NPP, net primary productivity.
除地上凋落物外, 根系也是土壤有机碳的重要来源, 而且根系对土壤有机碳形成与积累的贡献远高于地上凋落物(Jackson et al., 2017; Sokol & Bradford, 2019; Villarino et al., 2021)。这主要与3个因素有关: (1)根系能够比地上凋落物向土壤供应更多的碳; (2)根系死亡后可以立即而直接地与土壤矿物、微生物和团聚体相互作用, 具有较高的有机碳形成效率; (3)根系来源碳(如根际沉积物)比凋落物来源碳更能有效地形成稳定的土壤有机碳(Rasse et al., 2005; Crow et al., 2009; Jackson et al., 2017; Sokol et al., 2019; Sokol & Bradford, 2019)。与地上生产力不同, 氮磷添加对不同类型生态系统地下生产力的影响较为复杂, 可表现为促进作用、没有显著影响或抑制作用(Li et al., 2016; Yue et al., 2017; Jiang et al., 2019; Xing et al., 2022a)。氮磷添加的促进作用可能是由于植物生长受到N、P限制, 或者单一养分的添加会加剧植物对另一种养分的需求, 进而促使植物分配更多的光合产物到地下以获取所缺乏的养分(Li et al., 2016; Čapek et al., 2018); 而没有显著影响或抑制作用的原因可能是植物根系的主要功能之一是用于获取养分, 当植物面对养分可利用性提高之后, 不需要分配更多的光合产物到地下部分来获取养分(Peng et al., 2017; Ning et al., 2021)。虽然氮磷添加对生态系统地下生产力的影响不尽相同, 但目前的大尺度整合分析研究大都表明, 氮磷(N、P和N + P)添加对地下生产力总体上表现为促进作用或没有显著影响(图1; Li et al., 2016; Yue et al., 2017; Jiang et al., 2019)。
1.2 氮磷添加对土壤碳输出过程的影响
研究表明, 在一些N限制的生态系统, N添加能够通过提高微生物活性而促进土壤有机碳的分解(Zhang et al., 2014; Zhong et al., 2016), 但总体上N添加倾向于降低土壤有机碳的分解(Janssens et al., 2010; Ramirez et al., 2012; Zhou et al., 2014; Xu et al., 2021a)。具体而言, N添加主要可通过4条途径抑制土壤有机碳的分解(图1): (1)添加的外源N直接和土壤有机质发生缩合反应(condensation reaction)等化学作用, 从而增强土壤有机质的化学稳定性使其难以被微生物分解利用(Fog, 1988; Janssens et al., 2010)。(2) N添加可降低土壤pH, 导致土壤酸化和铝离子的活化而产生毒害(Treseder, 2008; Tian & Niu, 2015; Chen et al., 2020b), 从而抑制微生物生物量及其分解作用(Ye et al., 2018; Zhang et al., 2018)。(3) N添加减缓微生物N限制程度, 这既可直接提高微生物CUE、降低氧化酶活性和微生物对土壤有机质的“氮挖掘” (Manzoni et al., 2012; Ramirez et al., 2012; Spohn et al., 2016; Liu et al., 2018), 也可改变微生物群落结构(即降低真菌细菌比)和功能, 降低对稳定性有机碳的分解, 进而导致异养呼吸降低(Frey et al., 2004; Janssens et al., 2010; Kamble et al., 2013)。(4) N添加提高土壤养分可利用性后, 植物会调整其资源获取策略而减少向地下部分尤其是根际的碳分配, 导致微生物活性下降和激发效应降低, 进而抑制根际微生物呼吸(Janssens et al., 2010; Feng & Zhu, 2021; Ning et al., 2021)。
与N添加可通过土壤酸化等途径抑制微生物对土壤碳的分解输出不同, P添加对土壤有机碳分解的影响较为复杂(Cleveland et al., 2002; Camenzind et al., 2018; Mori et al., 2018; Feng & Zhu, 2019)。研究表明, 在P限制的热带森林中, P添加可通过缓解微生物P限制而促进微生物的生长和活性, 从而提高其对土壤有机碳的分解(Johnston et al., 2019; Hui et al., 2020)。此外, P添加也可通过非生物途径促进土壤有机碳的分解, 其机制是: 与有机化合物相比, 磷酸盐与土壤矿物表面的吸附位点具有更高的亲和力, 加入的无机P会使得被土壤矿物吸附的有机化合物(如可溶性有机碳)发生解吸, 从而使这些碳底物能够被微生物利用分解(Spohn & Schleuss, 2019; Spohn et al., 2022)。也有研究指出, P添加可通过多种途径降低土壤有机碳分解, 如降低植物向地下的碳分配(Wang et al., 2017), 降低微生物“磷挖掘” (Mori et al., 2015), 提高微生物CUE (Manzoni et al., 2012), 以及增强植物N限制及其对N的吸收而抑制微生物分解活性(Feng & Zhu, 2019)。尽管不同研究得到的结果不尽相同, 但综合各类生态系统而言, P添加总体上对微生物活性及其对土壤有机碳的分解表现为无显著影响或促进作用(Camenzind et al., 2018; 范珍珍等, 2018; Feng & Zhu, 2019; Jiang et al., 2021)。
2 氮磷添加对土壤有机碳的影响与潜在机制
2.1 氮磷添加对土壤总有机碳库的影响
目前, 全球或区域尺度的整合分析结果大都表明, 氮磷添加总体上有显著增加土壤有机碳库的趋势, N添加的促进程度为4%-11%, P添加的促进程度为5%左右, N + P添加的促进程度为8% (Janssens et al., 2010; Liu & Greaver, 2010; Yue et al., 2017; Chen et al., 2018b; Xu et al., 2021a)。氮磷添加(尤其是单独的N添加)下, 土壤有机碳库的增加通常被归因于植物向土壤中的碳输入(通常以植被生产力为表征)增加和土壤有机碳分解输出降低两个方面(图1), 而且这两个方面可以耦合发生或单独发生而起作用(Ye et al., 2018; Xu et al., 2021a)。与单独的N或P添加相比, N + P添加总体上对土壤有机碳的促进作用更强, 可能与N + P添加对植物碳输入的促进程度更大有关(Elser et al., 2007; Yue et al., 2017; Jiang et al., 2019)。就N、P添加的交互作用(N × P)而言, 其对土壤总有机碳库的影响可能取决于生态系统中植物和微生物的养分限制状况。有研究发现, N × P在苔原和农田中表现为拮抗作用, 但就不同生态系统来说, N × P通常不显著, 而是更多地表现为加和效应(即N + P添加的效果相当于单独N添加和单独P添加效果的叠加)(Sundqvist et al., 2014; Yue et al., 2017; Luo et al., 2019; Jiang et al., 2021)。就调控因素而言, 研究表明, N添加对土壤有机碳库的促进作用受到实验处理时间和N添加量的调控, 表现为随实验处理时间和N添加量的增加促进作用增强(Xu et al., 2021a; Lu et al., 2022); 而P添加和N + P添加并没有类似的规律, 但案例研究有限, 还存在较大的不确定性。
植物碳输入的增加和微生物分解输出的降低是氮磷添加促进土壤有机碳积累的两个潜在机制, 但它们的主导作用在N添加和P添加下存在差异。基于热带森林实验和数据整合分析, 最近的一项研究提出了土壤碳吸存假说, 该假说认为: 在“氮限制”的生态系统(如北方森林)中, N添加促进植被净初级生产力, 地上凋落物输入增加和土壤碳排放降低导致土壤碳吸存增加; 而在“氮富集”的生态系统(如热带森林)中, 长期N添加对净初级生产力无显著影响, 即植物向土壤中的碳输入没有增加, 土壤碳吸存增加的主要驱动因素是土壤碳排放和可溶性有机碳淋溶降低(Lu et al., 2021b)。此外, 也有研究指出, N添加下土壤有机碳的增加主要与微生物对有机碳的分解输出减弱有关, 而与植物碳输入的增加无密切关系(Crowther et al., 2019; Lu et al., 2022)。综合不同研究而言, N添加下土壤有机碳积累的主要驱动力可能是微生物对土壤有机碳分解输出的降低(Crowther et al., 2019; Lu et al., 2021b, 2022)。鉴于P添加对微生物分解总体表现为无显著影响或促进作用, P添加对土壤有机碳的促进作用可能主要与植物碳输入的增加有关(Yue et al., 2017; Feng & Zhu, 2019; Hou et al., 2020)。由于N添加和P添加促进土壤有机碳增加的主要原因不同, N + P添加对土壤有机碳的促进作用可能是微生物分解输出降低和植物碳输入增加共同作用的结果。然而, P添加和N + P添加对土壤有机碳的影响仍缺乏较为系统和定量化的研究, 仍需要进一步探讨。
虽然氮磷(N、P和N + P)添加提高土壤有机碳库的现象在全球尺度上普遍存在, 但是氮磷添加对土壤有机碳的负效应也经常在实验中被观测到(Waldrop et al., 2004; Keeler et al., 2009; Crowther et al., 2019; Huang et al., 2019; Luo et al., 2020)。这种负效应可能与以下原因有关: (1)在N和/或P限制的生态系统, 氮磷添加提高了微生物活性(如生物量和酶活性增加)、降低了微生物CUE, 促进了微生物对土壤碳的分解输出, 而且对碳分解输出的促进作用强于对碳输入的促进作用(Waldrop et al., 2004; Keeler et al., 2009; Luo et al., 2019)。(2)氮磷添加提高了植物地上部分的碳输入, 但没有改变或降低了植物地下部分生产力, 而植物地下部分碳输入对土壤有机碳的形成与积累更为重要(Jackson et al., 2017; Sokol & Bradford, 2019; Villarino et al., 2021)。换言之, 地上碳输入增加对土壤有机碳的促进作用不足以弥补地下碳输入减少和微生物分解作用所带来的土壤有机碳降低(Song et al., 2013; Keller et al., 2022)。(3)加入对植物生长非限制性的养分会加剧微生物与植物之间对另一种养分的需求与竞争(Čapek et al., 2018); 一方面会导致植物生产力和其向土壤碳输入的降低, 另一方面也会导致微生物从土壤有机质中挖掘养分而促进土壤碳的分解输出。(4)不平衡的或过量的氮磷添加超过了植物和微生物对养分的需求, 导致养分失衡或产生毒害(Harpole et al., 2011), 对植物生长和微生物活性产生抑制, 降低植物碳输入、限制微生物对植物来源碳转化成稳定的土壤有机碳, 进而导致土壤有机碳净损失。
2.2 氮磷添加对土壤有机碳组分的影响
2.2.1 土壤有机碳分组方法
粒径分组是指根据粒径大小将土壤有机碳分为不同的组分, 一般划分为颗粒态有机碳(POC, >53 μm)和矿物结合态有机碳(MAOC, <53 μm)两个组分(von Lützow et al., 2007; Chen et al., 2020b, 2021)。如果将土壤有机碳分为大团聚体有机碳(>250 μm)、微团聚体有机碳(53-250 μm)和黏粉粒有机碳(<53 μm, 即MAOC) 3个组分, 该分组方法也可以称为团聚体分组(Six et al., 2000; Huang et al., 2019)。密度分组是指根据土壤颗粒在一定密度(通常为1.60-1.85 g·cm-3)溶液中的沉降情况, 将土壤有机碳分为轻组分有机碳和重组分有机碳两部分, 轻组分有机碳可进一步分为游离态的轻组分有机碳和包裹态的轻组分有机碳(von Lützow et al., 2007; Cusack et al., 2011b; Ye et al., 2018; Lavallee et al., 2020)。尽管两种分组方法的操作不同, 但它们所得到的碳组分的性质具有一定的相似性(表1; Six et al., 2002; von Lützow et al., 2007; Lavallee et al., 2020)。一般认为, 轻组分有机碳和POC类似, 主要来源于新输入的、分解不完全的凋落物和根系残体, 由于缺少矿物或物理化学保护较容易被微生物分解利用, 被认为是活性碳组分; 重组分有机碳和MAOC类似, 由于与矿物结合受到保护而比较稳定, 难以被微生物分解利用, 被认为是稳定性碳组分(Six et al., 2002; von Lützow et al., 2007; Lavallee et al., 2020)。此外, 游离态的轻组分有机碳和大团聚体有机碳类似, 包裹态的轻组分有机碳和微团聚体有机碳类似(von Lützow et al., 2007; Cusack et al., 2011a, 2011b)。目前, 采用粒径分组、密度分组或者二者结合起来的方法将土壤有机碳分为POC (>53 μm或<1.60-1.85 g·cm-3)和MAOC (<53 μm或>1.60- 1.85 g·cm-3)被认为是一种行之有效的有机碳组分区分方法,有助于更好地理解土壤有机碳的形成和稳定以及预测土壤有机碳对环境变化的响应(Lavallee et al., 2020; Rocci et al., 2021; Feng et al., 2022)。
表1 土壤有机碳的分组方法及其组分特性
Table 1
| 粒径分组 Size fractionation | 密度分组 Density fractionation | 特性 Characteristic | 文献 Reference |
|---|---|---|---|
| 大团聚体有机碳 Macroaggregate OC (>250 μm) | 游离态的轻组分有机碳 Free light-fraction OC | 来源于新输入的植物残体, 最容易被微生物分解, 周转速率最快 Originated from newly-inputted plant residues, most easily to be decomposed by microbes, and with the fastest turnover rate | Six et al., 2002; Marín-Spiotta et al., 2008 |
| 微团聚体有机碳 Microaggregate OC (53-250 μm) | 包裹态的轻组分有机碳 Occluded light-fraction OC | 来源于半分解的植物残体, 但在团聚体内部受到了物理保护, 较易分解, 周转速率较快 Originated from partly-decomposed plant residues, but physically protected within aggregates, easier to be decomposed, and with a faster turnover rate | Trumbore, 1993; von Lützow et al., 2007 |
| 黏粉粒/矿物结合态有机碳 Silt-clay/mineral-associated OC (<53 μm) | 重组分有机碳 Heavy-fraction OC | 与土壤矿物紧密结合, 较难被分解, 周转速率较慢 Closely bound to soil minerals, difficult to be decomposed, and with a slow turnover rate | John et al., 2005; Lavallee et al., 2020 |
括号内指的是土壤粒径大小。
The notes in parenthesis indicate the particle-size of soil fractions. OC, organic carbon.
2.2.2 氮磷添加对土壤有机碳组分的影响
氮磷添加对土壤有机碳不同组分的影响比较复杂, 在不同研究中存在很大的差别。研究发现, 氮磷(N、P和N + P)添加促进了土壤活性碳组分(POC, 指碳库大小), 主要原因是氮磷添加促进了植物碳输入、抑制了微生物活性及其对该碳组分的分解(Ye et al., 2018; Chen et al., 2020b; Yuan et al., 2020)。与活性碳组分不同, 这些研究表明, N添加降低了稳定性碳组分(MAOC), 主要原因是N添加所导致的土壤酸化降低了微生物生物量碳和钙结合态有机碳(Ye et al., 2018; Chen et al., 2020b); 而P添加和N + P添加没有显著改变稳定性碳组分(Yuan et al., 2020)。然而, 也有一些研究发现, N添加促进了高山苔原和热带森林土壤中的稳定性碳组分(Neff et al., 2002; Cusack et al., 2011b)。这可能是因为, N添加能够促进植物残体来源的有机质与矿质N进行缩合反应而使其进入矿物结合态的土壤组分中, 或者通过微生物对植物残体的转化而将其以微生物残体的形式进入稳定性碳组分中(Moran et al., 2005; Cusack et al., 2011b; Mori et al., 2018)。与N和土壤有机质的缩合反应不同, 有研究发现, P添加和N + P添加降低了稳定性碳组分(Liu et al., 2013; Luo et al., 2020), 这可能与添加的P使矿物吸附的可溶性有机碳解吸附有关(Spohn et al., 2022)。也有研究发现, 活性碳组分在单独的N或P添加下降低, 而在N + P添加下增加, 即N × P表现为拮抗作用; 这可能是单一养分添加加剧了养分不平衡, 促使微生物对活性碳组分的分解, 而N + P添加促进了植物碳输入(Luo et al., 2019, 2020)。然而, 也有很多研究表明, N + P添加对土壤碳组分的影响与单独的N或P添加类似, 即N和P之间不存在交互作用(Liu et al., 2013; Huang et al., 2019; Luo et al., 2020; Yuan et al., 2020)。
综合以往的研究(Averill & Waring, 2018; Ye et al., 2018; Chen et al., 2020b, 2021), 氮磷添加主要通过提高土壤N、P可利用性和/或导致土壤酸化对植物残体与根际沉积物输入、土壤微生物生物量与残体积累以及金属离子的淋失(如Ca2+)或释放(如Fe3+和Al3+)产生影响, 进而对土壤有机碳组分产生影响(图2)。然而, N添加和P添加对土壤有机碳组分影响的途径可能不同。N添加可通过导致土壤酸化和增加土壤N可利用性而对碳组分产生影响, 而P添加应该是更多地通过增加土壤P可利用性而对碳组分产生影响(图2), 这是因为P添加对土壤pH的影响一般比较小, 不易导致土壤酸化(Feng & Zhu, 2019)。
图2
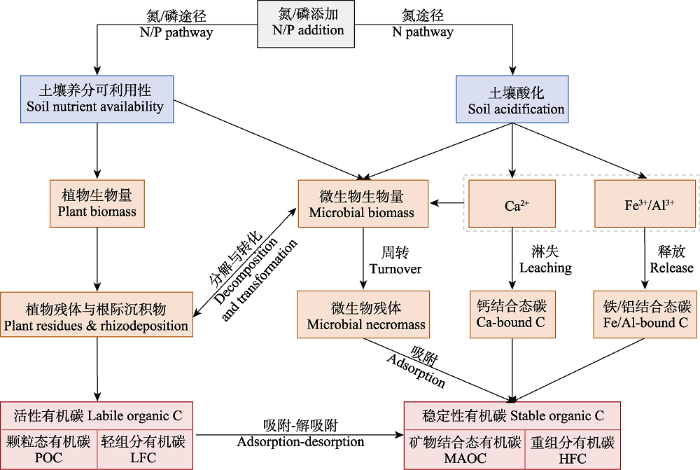
图2
氮磷添加对土壤有机碳组分影响的潜在机制。颗粒态有机碳和矿物结合态有机碳为粒径分组得到的组分, 轻组分有机碳和重组分有机碳为密度分组得到的组分, 它们的性质分别类似。
Fig. 2
Potential mechanisms of how nitrogen (N) and/or phosphorus (P) addition affect the fractions of soil organic carbon (C). Particulate organic C (POC) and mineral-associated organic C (MAOC) are the fractions separated by size fractionation, light-fraction organic C (LFC) and heavy-fraction organic C (HFC) are the fractions separated by density fractionation, and the characteristics of POC vs. LFC and MAOC vs. HFC are similar, respectively.
虽然N添加对土壤有机碳组分的影响在不同的案例研究中存在差异, 但总体而言, N添加对活性碳组分(游离态的轻组分有机碳、大团聚体有机碳和POC)和稳定性碳组分(重组分有机碳和MAOC)都表现为促进作用(Chen et al., 2018a; Lu et al., 2021a; Rocci et al., 2021)。具体来说, N添加对活性碳组分的促进程度(15%-18%)强于对稳定性碳组分的促进程度(3%-9%); 因此, N添加提高了活性碳组分在土壤总有机碳中的比例, 降低了稳定性碳组分的比例(Chen et al., 2018a; Lu et al., 2021a; Rocci et al., 2021)。N添加对活性碳组分的影响受到土壤pH和植物碳输入的调控, N添加促进活性碳组分的机理是: 促进碳输入和/或通过土壤酸化抑制微生物活性(如生物量)及其对活性碳组分的分解作用(Averill & Waring, 2018; Chen et al., 2018a; Ye et al., 2018; Lu et al., 2021a)。然而, N添加对稳定性碳组分的影响还存在一定的争议。一方面, N添加可能会通过抑制氧化酶活性而降低微生物对稳定性有机碳的分解而提高稳定性碳组分(Janssens et al., 2010; Chen et al., 2018b)。另一方面, N添加也可能会通过抑制微生物生长与残体积累及其对土壤有机碳的贡献而使稳定性碳组分减少(Averill & Waring, 2018; Chen et al., 2018a, 2020b; Ye et al., 2018)。因此, N添加对土壤稳定性碳组分的影响可能取决于两个过程在实验持续时间上的平衡, N添加在短期内即可对微生物的分解活性产生影响, 但需要较长的时间才能对微生物残体积累及其对土壤有机碳的贡献产生作用(Ye et al., 2018; Ma et al., 2021)。因此, 未来还需要研究长期N添加对土壤稳定性碳组分的影响与机制。
2.3 氮磷添加对土壤有机碳分子组成的影响
2.3.1 土壤有机碳分子组成测定方法
在以往的研究中, 固态13C核磁共振波谱技术(13C-NMR)和傅里叶变换红外光谱分析常被用于研究土壤有机碳的分子组成, 这两种方法都是基于所测定的含碳官能团来表征土壤有机碳的分子组成(Baldock et al., 1992; Demyan et al., 2012; Hou et al., 2019)。相比而言, 13C-NMR因其对土壤有机碳分子组成的解析更加贴近真实状态而被广泛应用(叶成龙等, 2018; 李娜等, 2019; Yuan et al., 2020)。按照13C-NMR测定结果, 一般将土壤有机碳分为4个含碳功能团(Baldock et al., 1992), 分别为烷基碳(alkyl C)、烷氧碳(O-alkyl C)、芳香碳(aromatic C)和羰基碳(carbonyl C), 不同含碳功能团的性质如表2所示。烷基碳和芳香碳较难被微生物分解利用, 是难以降解的较稳定的有机碳成分; 烷氧碳和羧基碳较易被微生物分解利用, 是不稳定的活性有机碳成分。通常采用各含碳功能团含量的百分比表征土壤有机碳的分子组成, 采用芳香度(芳香碳/烷氧碳)、脂化度(烷基碳/烷氧碳)和总指数(芳香度与脂化度之和) 3个指数来表征土壤有机碳的总体分子组成和难降解性, 指数越大表明土壤有机碳越难被降解(Baldock et al., 1997; Wagai et al., 2013; 叶成龙等, 2018)。
表2 固态13C核磁共振波谱技术测定的土壤有机碳功能团及其主要形式与特性
Table 2
| 碳功能团 Functional group | 化学位移 Chemical shift (δ) | 碳的主要形式 Dominant forms of carbon | 特性 Characteristic |
|---|---|---|---|
| 烷基碳 Alkyl C | 0-45 | 主要为脂肪族化合物等, 来自于植物角质、蜡质、木栓质 Mainly aliphatic compounds, originating from plant cutin, waxes, suberin | 较稳定, 不易被分解, 为难分解碳 Relatively stable, not easy to be decomposed, and categorized as the recalcitrant C |
| 烷氧碳 O-alkyl C | 45-110 | 主要为碳水化合物, 如纤维素、半纤维素等 Mainly carbohydrates, such as cellulose, hemicellulose, etc. | 容易被分解, 为易分解碳 Easy to be decomposed, and categorized as the easily- decomposed C |
| 芳香碳 Aromatic C | 110-165 | 主要为单宁、木质素等 Mainly tannin, lignin, etc. | 难以被分解, 为难分解碳 Difficult to be decomposed, and categorized as the recalcitrant C |
| 羰基碳 Carbonyl C | 165-210 | 大多为脂肪酸、氨基酸、酰胺、酯、酮醛类物质 Mostly fatty acids, amino acids, amide, esters, ketones and aldehydes | 容易被分解, 为易分解碳 Easy to be decomposed, and categorized as the easily- decomposed C |
根据Baldock等(
Generated according to Baldock et al. (
2.3.2 氮磷添加对土壤有机碳分子组成的影响
养分添加对土壤有机碳分子组成的影响复杂多样, 并没有一致性的规律。研究表明, N添加对土壤有机碳分子组成的影响受到土壤N有效性、N添加量和N形态等多种因素的调节。在土壤N有效性方面, 有研究发现, 在N有效性较高的热带低地森林中, N添加对烷基碳占比无影响, 但降低了烷氧碳占比; 而在N有效性较低的热带山地森林中, N添加提高了烷氧碳占比, 但降低了烷基碳占比(Cusack et al., 2011a)。在N添加量方面, 有研究表明, 土壤有机碳分子组成随N添加量的增加呈现非线性响应(Li et al., 2019; Man et al., 2021)。一项在温带草原的研究表明, 不同剂量的N添加均提高了烷基碳占比, 但对芳香碳占比存在不同的影响, 即随N添加量的增加芳香碳占比呈现先降低后增加的趋势(Li et al., 2019)。与此相反, 一项在农田生态系统的研究发现, 烷基碳占比随N添加量的增加呈现先降低后升高的趋势, 而芳香碳占比呈现先升高后降低的趋势(Man et al., 2021)。在N形态方面, 有研究发现, 硝态氮和铵态氮添加对土壤有机碳分子组成具有相异性的影响; 具体而言, 铵态氮添加降低了北方森林土壤轻组分中的烷基碳占比, 但硝态氮添加对其无显著影响; 与此相反, 硝态氮添加降低了芳香度, 但铵态氮添加对其没有显著影响(Cheng et al., 2017)。与N添加相比, P添加、N + P添加和N × P交互作用对土壤有机碳分子组成影响的研究还很少(Guo et al., 2017; Yuan et al., 2020; Li et al., 2021)。一项在高寒草甸的研究发现, N + P添加改变了土壤有机碳的分子组成, 提高了烷基碳占比、降低了烷氧碳占比, 使得土壤有机碳的难降解性增强(Guo et al., 2017)。然而, 在高寒草甸的另一项研究发现, 为期10年的P添加和N + P添加没有改变各类含碳功能团的占比, 也没有改变脂化度和芳香度等表征有机碳可降解性的指数, 而且N × P不显著(Yuan et al., 2020)。与此类似, 为期35年的P添加和N + P添加也没有改变农田土壤有机碳的分子组成(Li et al., 2021)。在这些研究中, P添加对土壤有机碳的分子组成没有显著影响, 这可能与P添加没有改变土壤微生物群落结构有关。虽然在不同的研究中, 氮磷添加对各类含碳功能团占比的影响不尽相同, 但总体上影响比较小, 这可能与土壤有机碳的分子组成相对稳定, 需要在长期养分添加下才能发生较大的变化有关(Wang et al., 2019; Lu et al., 2021b; van den Enden et al., 2021)。
在氮磷添加调控土壤有机碳分子组成的机制方面, 有研究指出土壤有机碳分子组成与微生物群落结构存在一定的关联(Cusack et al., 2011a; Xing et al., 2022b), 这表明氮磷添加可通过改变微生物群落结构而对土壤有机碳的分子组成产生影响。两项在高寒草甸的研究表明, 10年的N添加和15年的N + P添加显著降低了土壤烷氧碳占比, 其占比的降低可能是由于N添加改变了微生物群落结构、使其由真菌向细菌转变, 进而使微生物对包含烷氧碳在内的不稳定碳底物的分解利用增加(Guo et al., 2017; Yuan et al., 2020)。此外, 基于土壤密度分组的一项研究发现, 土壤轻组分中的烷氧碳占比与革兰氏阳性细菌生物量呈负相关关系, 而烷基碳占比与真菌相对丰度呈负相关关系; 同时, N添加提高了真菌丰度但降低了烷氧碳占比(Cusack et al., 2011a)。这些研究表明, 氮磷添加可通过改变微生物群落结构及其对土壤碳底物的利用偏好而改变土壤有机碳的分子组成, 但目前这方面的研究案例十分有限, 限制了我们对养分添加如何调控土壤有机碳分子组成的深层认识与理解, 未来仍需要进一步探讨。
3 总结和展望
土壤有机碳库的变化取决于碳输入和输出两个过程之间的动态变化, 而这两个过程都受到土壤氮磷可利用性的调节, 因此, 外源氮磷的输入会通过改变碳输入和输出过程而对土壤有机碳的碳库大小、组分和分子组成等产生影响。整合目前的研究结果, N添加和P添加整体上对土壤总有机碳表现为促进作用(Yue et al., 2017; Chen et al., 2018b; Feng & Zhu, 2019; Xu et al., 2021a), 但二者的促进机制可能存在差异。N添加对土壤有机碳的促进作用是由于N添加增加了碳输入并同时降低了碳输出(图1; Lu et al., 2011; Zhang et al., 2018; Xu et al., 2021a), 而P添加对土壤有机碳的促进作用可能主要是由于P添加促进了碳输入(Yue et al., 2017; Feng & Zhu, 2019)。在土壤有机碳组分方面, N添加对活性和稳定性有机碳组分均表现为促进作用, 但对活性有机碳组分的促进作用更大; 但P添加对土壤有机碳组分的影响尚不清楚。此外, N添加和P添加对土壤有机碳分子组成的影响并没有一致性的结果, 而且相关的研究案例也很少, 氮磷添加调控土壤有机碳分子组成的机制还有待进一步探讨。
总体而言, 以往的研究为我们认识养分富集如何调控土壤有机碳的碳库大小、组分和分子组成提供了思路与借鉴, 有利于准确预测未来氮磷沉降背景下土壤有机碳的动态变化。然而, 目前关于氮磷添加如何影响土壤有机碳仍然存在一些尚不清楚的方面, 需要在未来的研究中进行加强。
(1)加强P添加对土壤有机碳影响的研究。与N添加相比, P添加对不同类型生态系统土壤有机碳的影响无论是在其碳库大小还是在其组分和分子组成上都十分欠缺, 导致我们对相关的机制认知非常有限。热带森林的净初级生产力占全球净初级生产力的38%, 总碳库占到全球陆地生态系统碳库的40%, 在全球陆地碳循环中具有重要作用(Pan et al., 2011; Townsend et al., 2011)。热带森林作为P限制的生态系统, 在全球变化背景下备受关注, 但P添加对其土壤有机碳的影响与机制也尚不清楚。一方面, P添加可通过缓解植物P限制而促进热带森林的树木生长和生产力, 进而促进植物对土壤的碳输入(Wright et al., 2018; Wright, 2019)。另一方面, P添加也可通过缓解微生物P限制而促进其生长和对土壤有机碳的分解, 进而使土壤碳输出增加(Cleveland et al., 2002; Camenzind et al., 2018; Hui et al., 2020)。然而, 长期P添加能否持续促进微生物活性, 以及植物的碳输入和微生物的碳分解输出两个过程在调控热带森林土壤有机碳动态中的相对重要性尚不清楚, 使得P添加对土壤有机碳的影响仍存在较大的不确定性。因此, 未来需要加强P添加对不同类型生态系统(尤其是热带森林)土壤有机碳影响的研究, 并在已有的P添加实验平台上持续开展研究并进行系统测定, 这将有利于揭示P富集对土壤有机碳的碳库大小、组分和分子组成影响的潜在机制。
(2)加强氮磷添加下植物和微生物在调控土壤有机碳及其组分变化中的作用和相对贡献的研究。根据以往的研究, 植物来源的活性碳组分可主要通过两种途径转化为稳定性碳组分: 一是土壤矿物的吸附和团聚体的物理保护等非生物途径, 二是微生物对植物来源碳的“体内周转”与转化的生物途径(Lehmann & Kleber, 2015; Liang et al., 2017)。微生物对植物来源碳的转化及其周转而形成的残体是稳定性碳组分的重要来源(Cotrufo et al., 2013; Chen et al., 2020a; Wang et al., 2021), 而微生物残体与活体微生物生物量密切相关(Yang et al., 2020; Wang et al., 2021)。大部分研究表明, N添加总体上对土壤微生物生物量具有抑制趋势(Xiao et al., 2018; Zhang et al., 2018; Xu et al., 2021a); 因此, 从这一角度而言, N添加可能会通过降低微生物对植物来源碳的“体内周转”而减少稳定性碳的形成与积累。然而, 目前的研究表明, N添加促进了土壤稳定性碳组分(Chen et al., 2018b; Rocci et al., 2021), 这表明土壤物理化学过程参与的非生物途径在N添加调控土壤稳定性碳组分中可能比微生物参与的生物途径更加重要。土壤微生物不仅可以作为贡献者调控土壤有机碳库中微生物来源碳的动态, 也可作为分解者调控植物来源碳的动态(Liang et al., 2017)。然而, 目前的研究主要是从植物碳输入和微生物对土壤碳的分解输出两个过程探讨了N、P添加对土壤有机碳动态的影响, 但并未对他们的相对贡献进行量化。因此, 未来的研究可结合生物标志物和碳同位素标记等方法和技术来定量碳输入和输出过程对土壤有机碳及其组分的调控作用, 以及植物和微生物来源碳对土壤有机碳及其组分的贡献。
(3)加强长期氮磷添加及其交互作用对土壤有机碳影响的研究。目前, 在全球范围内开展的氮磷添加对土壤有机碳影响的研究中, 一半以上(55%)的实验其处理持续时间不超过5年, 仅有12%的实验其处理持续时间超过20年(Feng & Zhu, 2019; Xu et al., 2021a)。研究表明, 氮磷添加对土壤总有机碳的促进强度随实验持续时间延长而增加(Lu et al., 2021a; Xu et al., 2021a)。然而, 土壤的固碳能力是有限的, 会发生饱和(Jackson et al., 2017), 这意味着在长期氮磷添加下土壤有机碳可能不会持续增加。因此, 未来的研究需要关注土壤的碳饱和能力, 并结合模型进行模拟预测, 这有助于理解土壤有机碳在较长时间尺度上的变化趋势。与土壤总有机碳类似, 在短期内, 氮磷添加通常对土壤有机碳的稳定性组分和分子组成没有显著的影响(Cheng et al., 2017; Yuan et al., 2020; Xu et al., 2021a)。然而, 有机碳组分和分子组成对土壤有机碳能够长期稳定存储至关重要, 而且往往需要在较长的时间后才能观测到其变化。此外, 在全球变化背景下, N沉降和P沉降是相伴发生的, 而且其沉降速率的不同会导致养分之间的不平衡(Peñuelas et al., 2013; Zhu et al., 2016), 会对植物碳输入和微生物分解作用产生影响, 并进而影响土壤有机碳的碳库大小、组分和分子组成。然而, 以往的研究主要关注单一养分添加(尤其是N添加)对土壤有机碳的影响, 忽视了N + P添加及其交互作用的影响。研究表明, N + P添加对土壤总有机碳的促进作用强于单独的N或P添加(Yue et al., 2017), 这表明当前的单独的N或P添加实验结果可能总体上会低估未来氮磷沉降背景下土壤有机碳的增加幅度。因此, 未来需要利用已有的氮磷添加实验平台并增加N × P双因子实验的开展, 加强长期的N添加、P添加、N + P添加及N × P对土壤有机碳的碳库大小、组分和分子组成影响的研究, 这将有利于提高对未来氮磷沉降背景下土壤有机碳动态的准确预测。
(4)加强氮磷添加对深层土壤有机碳影响的研究。目前, 大部分的研究主要关注氮磷添加对表层土壤有机碳的影响, 而忽视了深层土壤(Rocci et al., 2021; Xu et al., 2021a)。研究表明, 氮磷添加对深层(20 cm以下)土壤有机碳大小、组分和分子组成的影响与表层(主要为0-20 cm)土壤在强度与方向上并不一致(Yu et al., 2020; Rocci et al., 2021; Xu et al., 2021a)。这一结果启示我们, 忽视深层土壤将会高估或低估氮磷添加对土壤有机碳的影响, 进而使得氮磷沉降背景下对土壤有机碳动态的预测具有不确定性。因此, 未来需要加强对深层土壤有机碳的关注, 系统研究氮磷添加对其碳库大小、组分和分子组成的影响。
参考文献
Nitrogen limitation of decomposition and decay: How can it occur?
DOI:10.1111/gcb.13980
PMID:29121419
[本文引用: 3]

The availability of nitrogen (N) is a critical control on the cycling and storage of soil carbon (C). Yet, there are conflicting conceptual models to explain how N availability influences the decomposition of organic matter by soil microbial communities. Several lines of evidence suggest that N availability limits decomposition; the earliest stages of leaf litter decay are associated with a net import of N from the soil environment, and both observations and models show that high N organic matter decomposes more rapidly. In direct contrast to these findings, experimental additions of inorganic N to soils broadly show a suppression of microbial activity, which is inconsistent with N limitation of decomposition. Resolving this apparent contradiction is critical to representing nutrient dynamics in predictive ecosystem models under a multitude of global change factors that alter soil N availability. Here, we propose a new conceptual framework, the Carbon, Acidity, and Mineral Protection hypothesis, to understand the effects of N availability on soil C cycling and storage and explore the predictions of this framework with a mathematical model. Our model simulations demonstrate that N addition can have opposing effects on separate soil C pools (particulate and mineral-protected carbon) because they are differentially affected by microbial biomass growth. Moreover, changes in N availability are frequently linked to shifts in soil pH or osmotic stress, which can independently affect microbial biomass dynamics and mask N stimulation of microbial activity. Thus, the net effect of N addition on soil C is dependent upon interactions among microbial physiology, soil mineralogy, and soil acidity. We believe that our synthesis provides a broadly applicable conceptual framework to understand and predict the effect of changes in soil N availability on ecosystem C cycling under global change.© 2017 John Wiley & Sons Ltd.
Cycling and composition of organic matter in terrestrial and marine ecosystems
DOI:10.1016/j.marchem.2004.06.016 URL [本文引用: 2]
Assessing the extent of decomposition of natural organic materials using solid-state 13C NMR spectroscopy
Aspects of the chemical structure of soil organic materials as revealed by solid-state 13C NMR spectroscopy
DOI:10.1007/BF02402261 URL [本文引用: 4]
Nutrient limitation of soil microbial processes in tropical forests
DOI:10.1002/ecm.1279 URL [本文引用: 3]
A plant-microbe interaction framework explaining nutrient effects on primary production
Different responses of soil organic carbon fractions to additions of nitrogen
DOI:10.1111/ejss.12716 URL [本文引用: 4]
A keystone microbial enzyme for nitrogen control of soil carbon storage
DOI:10.1126/sciadv.aaq1689 URL [本文引用: 4]
Dynamics of microbial residues control the responses of mineral- associated soil organic carbon to N addition in two temperate forests
DOI:10.1016/j.scitotenv.2020.141318 URL [本文引用: 1]
Nitrogen addition has contrasting effects on particulate and mineral- associated soil organic carbon in a subtropical forest
DOI:10.1016/j.soilbio.2020.107708 URL [本文引用: 6]
Global annual soil respiration in relation to climate, soil properties and vegetation characteristics: summary of available data
Particulate organic carbon is more vulnerable to nitrogen addition than mineral-associated organic carbon in soil of an alpine meadow
DOI:10.1007/s11104-019-04279-4 URL [本文引用: 3]
Contrasting effects of NH4+ and NO3- amendments on amount and chemical characteristics of different density organic matter fractions in a boreal forest soil
DOI:10.1016/j.geoderma.2017.01.023 URL [本文引用: 2]
Phosphorus limitation of microbial processes in moist tropical forests: evidence from short-term laboratory incubations and field studies
DOI:10.1007/s10021-002-0202-9 URL [本文引用: 2]
Soil carbon storage informed by particulate and mineral- associated organic matter
DOI:10.1038/s41561-019-0484-6 URL [本文引用: 1]
Formation of soil organic matter via biochemical and physical pathways of litter mass loss
DOI:10.1038/NGEO2520
[本文引用: 1]
Cotrufo, M. Francesca; Soong, Jennifer L.; Horton, Andrew J.; Campbell, Eleanor E.; Haddix, Michelle L.; Wall, Diana H.; Parton, Andwilliam J. Colorado State Univ, Nat Resources Ecol Lab, Ft Collins, CO 80523 USA. Cotrufo, M. Francesca Colorado State Univ, Dept Soil & Crop Sci, Ft Collins, CO 80523 USA. Wall, Diana H. Colorado State Univ, Dept Biol, Ft Collins, CO 80523 USA.
The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter?
DOI:10.1111/gcb.12113
PMID:23504877
[本文引用: 1]
The decomposition and transformation of above- and below-ground plant detritus (litter) is the main process by which soil organic matter (SOM) is formed. Yet, research on litter decay and SOM formation has been largely uncoupled, failing to provide an effective nexus between these two fundamental processes for carbon (C) and nitrogen (N) cycling and storage. We present the current understanding of the importance of microbial substrate use efficiency and C and N allocation in controlling the proportion of plant-derived C and N that is incorporated into SOM, and of soil matrix interactions in controlling SOM stabilization. We synthesize this understanding into the Microbial Efficiency-Matrix Stabilization (MEMS) framework. This framework leads to the hypothesis that labile plant constituents are the dominant source of microbial products, relative to input rates, because they are utilized more efficiently by microbes. These microbial products of decomposition would thus become the main precursors of stable SOM by promoting aggregation and through strong chemical bonding to the mineral soil matrix.© 2012 Blackwell Publishing Ltd.
Sources of plant-derived carbon and stability of organic matter in soil: implications for global change
DOI:10.1111/j.1365-2486.2009.01850.x URL [本文引用: 1]
Sensitivity of global soil carbon stocks to combined nutrient enrichment
DOI:10.1111/ele.13258
PMID:30884085
[本文引用: 3]
Soil stores approximately twice as much carbon as the atmosphere and fluctuations in the size of the soil carbon pool directly influence climate conditions. We used the Nutrient Network global change experiment to examine how anthropogenic nutrient enrichment might influence grassland soil carbon storage at a global scale. In isolation, enrichment of nitrogen and phosphorous had minimal impacts on soil carbon storage. However, when these nutrients were added in combination with potassium and micronutrients, soil carbon stocks changed considerably, with an average increase of 0.04 KgCm year (standard deviation 0.18 KgCm year ). These effects did not correlate with changes in primary productivity, suggesting that soil carbon decomposition may have been restricted. Although nutrient enrichment caused soil carbon gains most dry, sandy regions, considerable absolute losses of soil carbon may occur in high-latitude regions that store the majority of the world's soil carbon. These mechanistic insights into the sensitivity of grassland carbon stocks to nutrient enrichment can facilitate biochemical modelling efforts to project carbon cycling under future climate scenarios.© 2019 John Wiley & Sons Ltd/CNRS.
Changes in microbial community characteristics and soil organic matter with nitrogen additions in two tropical forests
DOI:10.1890/10-0459.1 URL [本文引用: 5]
Effects of nitrogen additions on above- and belowground carbon dynamics in two tropical forests
DOI:10.1007/s10533-010-9496-4 URL [本文引用: 4]
Use of specific peaks obtained by diffuse reflectance Fourier transform mid-infrared spectroscopy to study the composition of organic matter in a Haplic Chernozem
DOI:10.1111/j.1365-2389.2011.01420.x URL [本文引用: 1]
Global patterns of terrestrial nitrogen and phosphorus limitation
DOI:10.1038/s41561-019-0530-4 URL [本文引用: 1]
Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater marine and terrestrial ecosystems
DOI:10.1111/j.1461-0248.2007.01113.x URL [本文引用: 2]
Effect of nitrogen and phosphorus addition on soil enzyme activities: a meta-analysis
DOI:10.13287/j.1001-9332.201804.024
[本文引用: 1]
We conducted a meta-analysis to analyze the effects of nitrogen (N) and combined N and phosphorus (N+P) addition on soil enzyme activities, which being involved in soil carbon (C), N, and P cycles as well as oxidative processes. Nitrogen addition significantly increased the activities of soil C acquisition enzymes, N acquisition enzymes, and P acquisition enzymes by 6.9%, 5.6% and 10.7%, respectively, while the enhancement was much stronger under N+P treatment with 13.4%, 37.4% and 13.3%, respectively. In contrast, both N addition and N+P addition decreased the activities of oxidative enzymes by 6.1% and 0.4%, respectively. The effect sizes of N and N+P addition on soil enzyme activities varied with ecosystem types, N fertilizer types, N addition rates, and the duration of fertilization experiments. Our results suggested that the changes in soil microbes and their enzymes under increasing N deposition and P addition would have profound impacts on soil biogeochemical processes and functions.
整合分析氮磷添加对土壤酶活性的影响
DOI:10.13287/j.1001-9332.201804.024
[本文引用: 1]
本文通过整合分析(Meta-analysis)的方法分析了氮、磷添加对土壤碳、氮和磷素循环水解酶以及土壤氧化酶活性的影响.结果表明: 氮添加显著增加了碳、氮和磷循环水解酶的活性,增幅分别为6.9%、5.6%和10.7%;与氮添加相比,在氮磷同时添加下,3类土壤酶的活性增加更为显著,增幅分别达13.4%、37.4%和13.3%.然而,对于土壤氧化酶,氮以及氮磷的添加都使其活性降低,分别降低了6.1%和0.4%.不同生态系统类型、氮肥类型、施肥速率和施肥试验时间都对土壤酶活性具有影响.在全球大气氮沉降与磷添加逐渐增加的背景下,土壤微生物活性和酶的变化将会对土壤生物地球化学循环过程和土壤生态系统功能产生重要影响.
Global patterns and associated drivers of priming effect in response to nutrient addition
DOI:10.1016/j.soilbio.2020.108118 URL [本文引用: 1]
Changes in plant inputs alter soil carbon and microbial communities in forest ecosystems
DOI:10.1111/gcb.16107 URL [本文引用: 1]
A global meta-analysis of soil respiration and its components in response to phosphorus addition
DOI:10.1016/j.soilbio.2019.04.008 URL [本文引用: 10]
A review on the effects of nitrogen and phosphorus addition on tree growth and productivity in forest ecosystems
DOI:10.17521/cjpe.2019.0176 URL [本文引用: 1]
氮磷添加对树木生长和森林生产力影响的研究进展
DOI:10.17521/cjpe.2019.0176
[本文引用: 1]
人为活动所导致的氮、磷输入和大气氮、磷沉降使生态系统中的氮、磷可利用性大幅提高, 对陆地生态系统的碳循环过程产生了显著影响。森林树木生长和生产力在全球碳循环中发挥着重要作用, 它决定着陆地碳固存的大小和方向。目前, 在全球范围内开展了很多氮、磷添加调控树木生长和生产力的野外控制实验, 但是研究结果并不一致, 受到多种生物、环境和实验处理条件等因素的影响。该文从野外氮添加和磷添加实验的文献数量、实验数量及其全球空间分布三个方面概述了氮、磷添加对森林树木生长和生产力影响的研究现状, 并总结了氮、磷添加实验中森林树木生长和生产力的评估方法, 包括相对生长速率和绝对增长量。基于相关的研究结果, 阐述了氮、磷添加影响树木生长和生产力的调控因素及其潜在影响机制, 包括气候、树木径级与林龄、植物功能性状(共生菌根类型、树木固氮属性和保守性与获得性性状)、植物和微生物相互作用关系、区域养分沉降速率和实验处理条件等。最后, 基于当前的研究进行了系统总结, 并指出今后需要加强的几个方面的研究, 以期为后续研究提供参考: 树木生长响应氮磷添加的生理学机制, 树木各部分生长对氮、磷添加响应的权衡与分配, 植物功能性状在调节与预测树木生长响应氮、磷添加中的作用, 树木之间的竞争关系如何调控氮、磷添加对树木生长的影响, 以及开展长期的和联网的氮、磷添加对树木生长和生产力影响的野外控制实验。
The effect of added nitrogen on the rate of decomposition of organic matter
DOI:10.1111/j.1469-185X.1988.tb00725.x URL [本文引用: 1]
Chronic nitrogen enrichment affects the structure and function of the soil microbial community in temperate hardwood and pine forests
DOI:10.1016/j.foreco.2004.03.018 URL [本文引用: 1]
Nitrogen cycles: past present and future
DOI:10.1007/s10533-004-0370-0 URL [本文引用: 2]
Long-term nitrogen & phosphorus additions reduce soil microbial respiration but increase its temperature sensitivity in a Tibetan alpine meadow
DOI:10.1016/j.soilbio.2017.05.024 URL [本文引用: 5]
Nutrient co-limitation of primary producer communities
DOI:10.1111/j.1461-0248.2011.01651.x
PMID:21749598
[本文引用: 1]
Synergistic interactions between multiple limiting resources are common, highlighting the importance of co-limitation as a constraint on primary production. Our concept of resource limitation has shifted over the past two decades from an earlier paradigm of single-resource limitation towards concepts of co-limitation by multiple resources, which are predicted by various theories. Herein, we summarise multiple-resource limitation responses in plant communities using a dataset of 641 studies that applied factorial addition of nitrogen (N) and phosphorus (P) in freshwater, marine and terrestrial systems. We found that more than half of the studies displayed some type of synergistic response to N and P addition. We found support for strict definitions of co-limitation in 28% of the studies: i.e. community biomass responded to only combined N and P addition, or to both N and P when added separately. Our results highlight the importance of interactions between N and P in regulating primary producer community biomass and point to the need for future studies that address the multiple mechanisms that could lead to different types of co-limitation.© 2011 Blackwell Publishing Ltd/CNRS.
Global meta-analysis shows pervasive phosphorus limitation of aboveground plant production in natural terrestrial ecosystems
DOI:10.1038/s41467-020-14492-w
PMID:32005808
[本文引用: 2]
Phosphorus (P) limitation of aboveground plant production is usually assumed to occur in tropical regions but rarely elsewhere. Here we report that such P limitation is more widespread and much stronger than previously estimated. In our global meta-analysis, almost half (46.2%) of 652 P-addition field experiments reveal a significant P limitation on aboveground plant production. Globally, P additions increase aboveground plant production by 34.9% in natural terrestrial ecosystems, which is 7.0-15.9% higher than previously suggested. In croplands, by contrast, P additions increase aboveground plant production by only 13.9%, probably because of historical fertilizations. The magnitude of P limitation also differs among climate zones and regions, and is driven by climate, ecosystem properties, and fertilization regimes. In addition to confirming that P limitation is widespread in tropical regions, our study demonstrates that P limitation often occurs in other regions. This suggests that previous studies have underestimated the importance of altered P supply on aboveground plant production in natural terrestrial ecosystems.
Latitudinal patterns of terrestrial phosphorus limitation over the globe
DOI:10.1111/ele.13761 URL [本文引用: 1]
Changes in soil organic matter stability with depth in two alpine ecosystems on the Tibetan Plateau
DOI:10.1016/j.geoderma.2019.05.034 URL [本文引用: 2]
Differential mechanisms drive changes in soil C pools under N and P enrichment in a subalpine spruce plantation
DOI:10.1016/j.geoderma.2019.01.027 URL [本文引用: 3]
Phosphorus rather than nitrogen enhances CO2 emissions in tropical forest soils: evidence from a laboratory incubation study
DOI:10.1111/ejss.12885 URL [本文引用: 2]
The sensitivity of soil respiration to soil temperature, moisture, and carbon supply at the global scale
DOI:10.1111/gcb.13489
PMID:27594213
[本文引用: 1]
Soil respiration (Rs) is a major pathway by which fixed carbon in the biosphere is returned to the atmosphere, yet there are limits to our ability to predict respiration rates using environmental drivers at the global scale. While temperature, moisture, carbon supply, and other site characteristics are known to regulate soil respiration rates at plot scales within certain biomes, quantitative frameworks for evaluating the relative importance of these factors across different biomes and at the global scale require tests of the relationships between field estimates and global climatic data. This study evaluates the factors driving Rs at the global scale by linking global datasets of soil moisture, soil temperature, primary productivity, and soil carbon estimates with observations of annual Rs from the Global Soil Respiration Database (SRDB). We find that calibrating models with parabolic soil moisture functions can improve predictive power over similar models with asymptotic functions of mean annual precipitation. Soil temperature is comparable with previously reported air temperature observations used in predicting Rs and is the dominant driver of Rs in global models; however, within certain biomes soil moisture and soil carbon emerge as dominant predictors of Rs. We identify regions where typical temperature-driven responses are further mediated by soil moisture, precipitation, and carbon supply and regions in which environmental controls on high Rs values are difficult to ascertain due to limited field data. Because soil moisture integrates temperature and precipitation dynamics, it can more directly constrain the heterotrophic component of Rs, but global-scale models tend to smooth its spatial heterogeneity by aggregating factors that increase moisture variability within and across biomes. We compare statistical and mechanistic models that provide independent estimates of global Rs ranging from 83 to 108 Pg yr, but also highlight regions of uncertainty where more observations are required or environmental controls are hard to constrain.© 2016 John Wiley & Sons Ltd.
The ecology of soil carbon: pools, vulnerabilities, and biotic and abiotic controls
DOI:10.1146/annurev-ecolsys-112414-054234 URL [本文引用: 5]
Reduction of forest soil respiration in response to nitrogen deposition
DOI:10.1038/ngeo844 URL [本文引用: 8]
Antagonistic and additive interactions dominate the responses of belowground carbon-cycling processes to nitrogen and phosphorus additions
DOI:10.1016/j.soilbio.2021.108216 URL [本文引用: 2]
Interactive effects of nitrogen and phosphorus additions on plant growth vary with ecosystem type
DOI:10.1007/s11104-019-04119-5
[本文引用: 3]
Aims Co-limitation of ecosystem productivity by nitrogen (N) and phosphorus (P) is gaining increasing recognition, but how co-limitation through N and P interactions differs among different terrestrial ecosystems remains unclear. Methods We performed a meta-analysis of 133 independent studies conducted in four natural terrestrial ecosystems to examine the interactive effects of N and P additions on ten plant growth-related variables. Results Adding N and P individually or in combination significantly increased aboveground biomass (AGB), and the interactions were uniformly synergistic for AGB, and additive for belowground biomass (BGB), but variable for other eight growth-related variables among four different ecosystems. The interaction was synergistic for leaf P and soil NO3-N only in tropical forests, and antagonistic for soil available P (AP) in tropical forests, leaf N in grasslands, root P in wetlands, and leaf P and soil NH4-N in tundra. The interaction for leaf N: P ratios was additive only in tropical forests, and synergistic in the other three ecosystems. Conclusions Our results highlighted the interactions of N and P additions can promote uptake of both nutrients by plants, and plants tend to maintain the optimal nutrient balance for growth and reproduction through regulating biomass production and tissue nutrient concentrations.
Storage of organic carbon in aggregate and density fractions of silty soils under different types of land use
DOI:10.1016/j.geoderma.2004.12.013 URL [本文引用: 1]
Phosphate addition increases tropical forest soil respiration primarily by deconstraining microbial population growth
DOI:10.1016/j.soilbio.2018.11.026 URL [本文引用: 1]
Bacterial growth and growth-limiting nutrients following chronic nitrogen additions to a hardwood forest soil
DOI:10.1016/j.soilbio.2012.12.017 URL [本文引用: 1]
Effects of long- term nitrogen addition on microbial enzyme activity in eight forested and grassland sites: implications for litter and soil organic matter decomposition
DOI:10.1007/s10021-008-9199-z URL [本文引用: 2]
Soil carbon stocks in temperate grasslands differ strongly across sites but are insensitive to decade-long fertilization
DOI:10.1111/gcb.15988 URL [本文引用: 1]
The macromolecular organic composition of plant and microbial residues as inputs to soil organic matter
DOI:10.1016/S0038-0717(01)00158-4 URL [本文引用: 2]
The detrital input and removal treatment (DIRT) network: insights into soil carbon stabilization
Soil carbon sequestration impacts on global climate change and food security
DOI:10.1126/science.1097396
PMID:15192216
[本文引用: 1]
The carbon sink capacity of the world's agricultural and degraded soils is 50 to 66% of the historic carbon loss of 42 to 78 gigatons of carbon. The rate of soil organic carbon sequestration with adoption of recommended technologies depends on soil texture and structure, rainfall, temperature, farming system, and soil management. Strategies to increase the soil carbon pool include soil restoration and woodland regeneration, no-till farming, cover crops, nutrient management, manuring and sludge application, improved grazing, water conservation and harvesting, efficient irrigation, agroforestry practices, and growing energy crops on spare lands. An increase of 1 ton of soil carbon pool of degraded cropland soils may increase crop yield by 20 to 40 kilograms per hectare (kg/ha) for wheat, 10 to 20 kg/ha for maize, and 0.5 to 1 kg/ha for cowpeas. As well as enhancing food security, carbon sequestration has the potential to offset fossil fuel emissions by 0.4 to 1.2 gigatons of carbon per year, or 5 to 15% of the global fossil-fuel emissions.
Conceptualizing soil organic matter into particulate and mineral-associated forms to address global change in the 21st century
DOI:10.1111/gcb.14859
PMID:31587451
[本文引用: 6]
Managing soil organic matter (SOM) stocks to address global change challenges requires well-substantiated knowledge of SOM behavior that can be clearly communicated between scientists, management practitioners, and policy makers. However, SOM is incredibly complex and requires separation into multiple components with contrasting behavior in order to study and predict its dynamics. Numerous diverse SOM separation schemes are currently used, making cross-study comparisons difficult and hindering broad-scale generalizations. Here, we recommend separating SOM into particulate (POM) and mineral-associated (MAOM) forms, two SOM components that are fundamentally different in terms of their formation, persistence, and functioning. We provide evidence of their highly contrasting physical and chemical properties, mean residence times in soil, and responses to land use change, plant litter inputs, warming, CO enrichment, and N fertilization. Conceptualizing SOM into POM versus MAOM is a feasible, well-supported, and useful framework that will allow scientists to move beyond studies of bulk SOM, but also use a consistent separation scheme across studies. Ultimately, we propose the POM versus MAOM framework as the best way forward to understand and predict broad-scale SOM dynamics in the context of global change challenges and provide necessary recommendations to managers and policy makers.© 2019 John Wiley & Sons Ltd.
The contentious nature of soil organic matter
DOI:10.1038/nature16069 URL [本文引用: 1]
Different traits from the paddy soil and upland soil regulate bacterial community and molecular composition under long-term fertilization regimes
DOI:10.1016/j.apsoil.2021.103982 URL [本文引用: 2]
Advancement in research on application of 13C NMR techniques to exploration of chemical structure of soil organic matter
应用13C核磁共振技术研究土壤有机质化学结构进展
Effects of external nitrogen additions on soil organic carbon dynamics and the mechanism
DOI:10.17521/cjpe.2015.0098
[本文引用: 1]
What would be the impact of external nitrogen additions on soil carbon, an issue still under debating, as reported experimental results were either positive, negtive or neutral. Several factors may be related to these seemingly controversial results: differences in ecosystem types and soil properties, soil carbon detection methods, soil depths, and contents of soil labile and recalcitrant carbon that affect the responses to nitrogen additions, all could cause discrepancies and variations in carbon sequestration. The several processes that contribute to enhance soil organic carbon storage include increasing litter input, decresing soil carbon output, particularly, by supressed decomposition of recalcitrant carbon, promoting soil humifiction and formation of recalcitrant carbon storage. However, there are still many uncertainties associated with these issues. To improve our understanding, the research about carbon in deep soil layers, dissolved organic carbon leaching and accumulation, and the effect of labile and recalcitrant soil C ratios on N addition responses, should be further investigated in the future studies.
土壤有机碳对外源氮添加的响应及其机制
DOI:10.17521/cjpe.2015.0098
[本文引用: 1]
土壤有机碳库是陆地生态系统碳库的重要组成, 在全球碳循环中发挥着重要的作用。受元素化学计量平衡调控作用, 氮输入的增加将会对土壤有机碳库产生重要影响。然而, 目前关于陆地生态系统碳库对氮添加的响应主要集中在植被碳库, 对土壤碳库研究较少, 且研究结论争议较大, 尤其对其响应机制缺少系统梳理。该文作者通过对已有文献进行梳理, 认为生态系统类型、土壤碳变化的检测方法、土壤深度, 以及土壤稳定性碳和易变碳含量的差异可能是造成当前研究土壤碳汇增量(每克氮输入所增加的碳)差异的重要原因。氮添加条件下土壤有机碳的积累机制可能包括3个方面: 1)氮添加增加了凋落物输入, 促进了碳积累; 2)氮添加减少土壤碳输出, 尤其是抑制了稳定性碳的分解; 3)促进土壤腐殖质及稳定性碳的形成。此外, 该文结合当前研究中存在的不足, 提出今后需加强对深层土壤碳、土壤可溶性有机碳的淋溶及吸附, 以及不同土壤碳组分对氮添加的响应研究, 并通过改进检测方法减少氮添加条件下碳储量的测量误差。
Soil microbial community composition closely associates with specific enzyme activities and soil carbon chemistry in a long-term nitrogen fertilized grassland
DOI:10.1016/j.scitotenv.2018.11.031 URL
Aggravated phosphorus limitation on biomass production under increasing nitrogen loading: a meta-analysis
DOI:10.1111/gcb.13125
PMID:26463578
[本文引用: 4]
Nitrogen (N) and phosphorus (P), either individually or in combination, have been demonstrated to limit biomass production in terrestrial ecosystems. Field studies have been extensively synthesized to assess global patterns of N impacts on terrestrial ecosystem processes. However, to our knowledge, no synthesis has been done so far to reveal global patterns of P impacts on terrestrial ecosystems, especially under different nitrogen (N) levels. Here, we conducted a meta-analysis of impacts of P addition, either alone or with N addition, on aboveground (AGB) and belowground biomass production (BGB), plant and soil P concentrations, and N : P ratio in terrestrial ecosystems. Overall, our meta-analysis quantitatively confirmed existing notions: (i) colimitation of N and P on biomass production and (ii) more P limitation in tropical forest than other ecosystems. More importantly, our analysis revealed new findings: (i) P limitation on biomass production was aggravated by N enrichment and (ii) plant P concentration was a better indicator of P limitation than soil P availability. Specifically, P addition increased AGB and BGB by 34% and 13%, respectively. The effect size of P addition on biomass production was larger in tropical forest than grassland, wetland, and tundra and varied with P fertilizer forms, P addition rates, or experimental durations. The P-induced increase in biomass production and plant P concentration was larger under elevated than ambient N. Our findings suggest that the global limitation of P on biomass production will become severer under increasing N fertilizer and deposition in the future. © 2015 John Wiley & Sons Ltd.
The importance of anabolism in microbial control over soil carbon storage
DOI:10.1038/nmicrobiol.2017.105
PMID:28741607
[本文引用: 2]
Studies of the decomposition, transformation and stabilization of soil organic matter (SOM) have dramatically increased in recent years owing to growing interest in studying the global carbon (C) cycle as it pertains to climate change. While it is readily accepted that the magnitude of the organic C reservoir in soils depends upon microbial involvement, as soil C dynamics are ultimately the consequence of microbial growth and activity, it remains largely unknown how these microorganism-mediated processes lead to soil C stabilization. Here, we define two pathways-ex vivo modification and in vivo turnover-which jointly explain soil C dynamics driven by microbial catabolism and/or anabolism. Accordingly, we use the conceptual framework of the soil 'microbial carbon pump' (MCP) to demonstrate how microorganisms are an active player in soil C storage. The MCP couples microbial production of a set of organic compounds to their further stabilization, which we define as the entombing effect. This integration captures the cumulative long-term legacy of microbial assimilation on SOM formation, with mechanisms (whether via physical protection or a lack of activation energy due to chemical composition) that ultimately enable the entombment of microbial-derived C in soils. We propose a need for increased efforts and seek to inspire new studies that utilize the soil MCP as a conceptual guideline for improving mechanistic understandings of the contributions of soil C dynamics to the responses of the terrestrial C cycle under global change.
Interactive effects of nitrogen and phosphorus on soil microbial communities in a tropical forest
DOI:10.1371/journal.pone.0061188 URL [本文引用: 2]
A global perspective on belowground carbon dynamics under nitrogen enrichment
DOI:10.1111/j.1461-0248.2010.01482.x
PMID:20482580
[本文引用: 1]
Nitrogen (N) effects on ecosystem carbon (C) budgets are critical to understand as C sequestration is considered as a mechanism to offset anthropogenic CO(2) emissions. Interactions between aboveground C and N cycling are more clearly characterized than belowground processes. Through synthesizing data from multiple terrestrial ecosystems, we quantified the responses of belowground C cycling under N addition. We found that N addition increased litter input from aboveground (+20%) but not from fine root. N addition inhibited microbial activity as indicated by a reduction in microbial respiration (-8%) and microbial biomass carbon (-20%). Although soil respiration was not altered by N addition, dissolved organic carbon concentration was increased by 18%, suggesting C leaching loss may increase. N addition increased the C content of the organic layer (+17%) but not the mineral soil layer. Overall, our meta-analysis indicates that N addition will increase short term belowground C storage by increasing C content of organic layer. However, it is difficult to predict the response of long term C sequestration since there is no significant change in mineral soil C content.
Microbial carbon use efficiency and priming effect regulate soil carbon storage under nitrogen deposition by slowing soil organic matter decomposition
DOI:10.1016/j.geoderma.2018.07.008 URL [本文引用: 1]
Minor stimulation of soil carbon storage by nitrogen addition: a meta-analysis
DOI:10.1016/j.agee.2010.12.010 URL [本文引用: 1]
Decrease in soil pH has greater effects than increase in above-ground carbon inputs on soil organic carbon in terrestrial ecosystems of China under nitrogen enrichment
DOI:10.1111/1365-2664.14091 URL [本文引用: 3]
Nitrogen addition stimulates soil aggregation and enhances carbon storage in terrestrial ecosystems of China: a meta-analysis
DOI:10.1111/gcb.15604 URL [本文引用: 5]
Nitrogen deposition accelerates soil carbon sequestration in tropical forests
DOI:10.1073/pnas.2020790118 [本文引用: 4]
Nitrogen and phosphorus enrichment accelerates soil organic carbon loss in alpine grassland on the Qinghai-Tibetan Plateau
DOI:10.1016/j.scitotenv.2018.09.038 URL [本文引用: 3]
Nutrient addition reduces carbon sequestration in a Tibetan grassland soil: disentangling microbial and physical controls
DOI:10.1016/j.soilbio.2020.107764 URL [本文引用: 4]
Effects of nitrogen addition on microbial residues and their contribution to soil organic carbon in Chinaʼs forests from tropical to boreal zone
DOI:10.1016/j.envpol.2020.115941 URL [本文引用: 1]
Altered soil organic matter composition and degradation after a decade of nitrogen fertilization in a temperate agroecosystem
DOI:10.1016/j.agee.2021.107305 URL [本文引用: 2]
Environmental and stoichiometric controls on microbial carbon-use efficiency in soils
DOI:10.1111/j.1469-8137.2012.04225.x
PMID:22924405
[本文引用: 2]
Carbon (C) metabolism is at the core of ecosystem function. Decomposers play a critical role in this metabolism as they drive soil C cycle by mineralizing organic matter to CO(2). Their growth depends on the carbon-use efficiency (CUE), defined as the ratio of growth over C uptake. By definition, high CUE promotes growth and possibly C stabilization in soils, while low CUE favors respiration. Despite the importance of this variable, flexibility in CUE for terrestrial decomposers is still poorly characterized and is not represented in most biogeochemical models. Here, we synthesize the theoretical and empirical basis of changes in CUE across aquatic and terrestrial ecosystems, highlighting common patterns and hypothesizing changes in CUE under future climates. Both theoretical considerations and empirical evidence from aquatic organisms indicate that CUE decreases as temperature increases and nutrient availability decreases. More limited evidence shows a similar sensitivity of CUE to temperature and nutrient availability in terrestrial decomposers. Increasing CUE with improved nutrient availability might explain observed declines in respiration from fertilized stands, while decreased CUE with increasing temperature and plant C : N ratios might decrease soil C storage. Current biogeochemical models could be improved by accounting for these CUE responses along environmental and stoichiometric gradients.© 2012 The Authors. New Phytologist © 2012 New Phytologist Trust.
Chemical and mineral control of soil carbon turnover in abandoned tropical pastures
DOI:10.1016/j.geoderma.2007.10.001 URL [本文引用: 1]
Role of mineral-nitrogen in residue decomposition and stable soil organic matter formation
DOI:10.2136/sssaj2004.0301 URL [本文引用: 1]
Phosphorus addition reduced microbial respiration during the decomposition of Acacia mangium litter in South Sumatra, Indonesia
DOI:10.3759/tropics.24.113 URL [本文引用: 1]
Reconsidering the phosphorus limitation of soil microbial activity in tropical forests
DOI:10.1111/1365-2435.13043 URL [本文引用: 3]
Variable effects of nitrogen additions on the stability and turnover of soil carbon
DOI:10.1038/nature01136 URL [本文引用: 1]
Carbon limitation overrides acidification in mediating soil microbial activity to nitrogen enrichment in a temperate grassland
DOI:10.1111/gcb.15819
PMID:34343388
[本文引用: 2]
Higher ecosystem nitrogen (N) inputs resulting from human activities often suppress soil microbial biomass and respiration, thereby altering biogeochemical cycling. Soil acidification and carbon (C) limitation may drive these microbial responses, yet their relative importance remains elusive, which limits our understanding of the longer-term effects of increasing N inputs. In a field experiment with continuous N addition at seven different rates from 0 to 50 g N m yr over six years in a temperate grassland of Inner Mongolia, China, we examined the responses of soil microbial biomass and respiration to changes in soil acidity and C availability by adding lime and/or glucose to soil samples. Soil microbial biomass and respiration did only weakly respond to increasing soil pH, but increased strongly in response to higher C availability with increasing N addition rates. Soil net N immobilization increased in response to glucose addition and soil microbial biomass increased at higher rates than microbial respiration along the gradient of previous N addition rates, both suggesting increasingly reinforced microbial C limitation with increasing N addition. Our results provide clear evidence for strong N-induced microbial C limitation, but only little support for soil acidity effects within the initial pH range of 4.73 to 7.86 covered by our study. Field data support this conclusion by showing reduced plant C allocation belowground in response to N addition, resulting in soil microbial C starvation over the long term. In conclusion, soil microbial biomass and respiration under N addition were strongly dependent on C availability, most likely originating from plant belowground C inputs, and was much less affected by changes in soil pH. Our data help clarify a long-standing debate about how increasing N input rates affect soil microbial biomass and respiration, and improve the mechanistic understanding of the linkages between ecosystem N enrichment and C cycling.This article is protected by copyright. All rights reserved.
Model-data synthesis for the next generation of forest free-air CO2 enrichment (FACE) experiments
DOI:10.1111/nph.13593 URL [本文引用: 1]
A large and persistent carbon sink in the worldʼs forests
DOI:10.1126/science.1201609 URL [本文引用: 1]
Global patterns of root dynamics under nitrogen enrichment
DOI:10.1111/geb.12508 URL [本文引用: 1]
Human-induced nitrogen-phosphorus imbalances alter natural and managed ecosystems across the globe
DOI:10.1038/ncomms3934
PMID:24343268
[本文引用: 1]
The availability of carbon from rising atmospheric carbon dioxide levels and of nitrogen from various human-induced inputs to ecosystems is continuously increasing; however, these increases are not paralleled by a similar increase in phosphorus inputs. The inexorable change in the stoichiometry of carbon and nitrogen relative to phosphorus has no equivalent in Earth's history. Here we report the profound and yet uncertain consequences of the human imprint on the phosphorus cycle and nitrogen: phosphorus stoichiometry for the structure, functioning and diversity of terrestrial and aquatic organisms and ecosystems. A mass balance approach is used to show that limited phosphorus and nitrogen availability are likely to jointly reduce future carbon storage by natural ecosystems during this century. Further, if phosphorus fertilizers cannot be made increasingly accessible, the crop yields projections of the Millennium Ecosystem Assessment imply an increase of the nutrient deficit in developing regions.
The human-induced imbalance between C, N and P in earthʼs life system
DOI:10.1111/j.1365-2486.2011.02568.x URL [本文引用: 2]
Consistent effects of nitrogen amendments on soil microbial communities and processes across biomes
DOI:10.1111/j.1365-2486.2012.02639.x URL [本文引用: 2]
Is soil carbon mostly root carbon? Mechanisms for a specific stabilisation
DOI:10.1007/s11104-004-0907-y URL [本文引用: 1]
Mechanisms driving the soil organic matter decomposition response to nitrogen enrichment in grassland soils
DOI:10.1016/j.soilbio.2016.04.023 URL [本文引用: 1]
Soil organic carbon response to global environmental change depends on its distribution between mineral-associated and particulate organic matter: a meta-analysis
DOI:10.1016/j.scitotenv.2021.148569 URL [本文引用: 6]
Global-scale impacts of nitrogen deposition on tree carbon sequestration in tropical, temperate, and boreal forests: a meta-analysis
Stabilization mechanisms of soil organic matter: implications for C- saturation of soils
DOI:10.1023/A:1016125726789 URL [本文引用: 4]
Soil macroaggregate turnover and microaggregate formation: a mechanism for C sequestration under no-tillage agriculture
DOI:10.1016/S0038-0717(00)00179-6 URL [本文引用: 2]
Microbial formation of stable soil carbon is more efficient from belowground than aboveground input
DOI:10.1038/s41561-018-0258-6
[本文引用: 3]
The relative contributions of aboveground versus belowground plant carbon inputs to the stable soil organic carbon pool are the subject of much debate-with direct implications for how the carbon cycle is modelled and managed. The belowground rhizosphere pathway (that is, carbon exiting the living root) is theorized to form stable soil carbon more efficiently than the aboveground pathway. However, while several mechanisms have been invoked to explain this efficiency, few have been empirically tested or quantified. Here, we use soil microcosms with standardized carbon inputs to investigate three posited mechanisms that differentiate aboveground from belowground input pathways of dissolved organic carbon-through the microbial biomass-to the mineral-stabilized soil organic carbon pool: (1) the physical distance travelled, (2) the microbial abundance in the region in which a carbon compound enters (that is, rhizosphere versus bulk soil) and (3) the frequency and volume of carbon delivery (that is, infrequent 'pulse' versus frequent 'drip'). We demonstrate that through the microbial formation pathway, belowground inputs form mineral-stabilized soil carbon more efficiently than aboveground inputs, partly due to the greater efficiency of formation by the rhizosphere microbial community relative to the bulk soil community. However, we show that because the bulk soil has greater capacity to form mineral-stabilized soil carbon due to its greater overall volume, the relative contributions of aboveground versus belowground carbon inputs depend strongly on the ratio of rhizosphere to bulk soil.
Evidence for the primacy of living root inputs, not root or shoot litter, in forming soil organic carbon
DOI:10.1111/nph.15361
PMID:30067293
[本文引用: 1]
Soil organic carbon (SOC) is primarily formed from plant inputs, but the relative carbon (C) contributions from living root inputs (i.e. rhizodeposits) vs litter inputs (i.e. root + shoot litter) are poorly understood. Recent theory suggests that living root inputs exert a disproportionate influence on SOC formation, but few field studies have explicitly tested this by separately tracking living root vs litter inputs as they move through the soil food web and into distinct SOC pools. We used a manipulative field experiment with an annual C grass in a forest understory to differentially track its living root vs litter inputs into the soil and to assess net SOC formation over multiple years. We show that living root inputs are 2-13 times more efficient than litter inputs in forming both slow-cycling, mineral-associated SOC as well as fast-cycling, particulate organic C. Furthermore, we demonstrate that living root inputs are more efficiently anabolized by the soil microbial community en route to the mineral-associated SOC pool (dubbed 'the in vivo microbial turnover pathway'). Overall, our findings provide support for the primacy of living root inputs in forming SOC. However, we also highlight the possibility of nonadditive effects of living root and litter inputs, which may deplete SOC pools despite greater SOC formation rates.© 2018 The Authors. New Phytologist © 2018 New Phytologist Trust.
Effect of nitrogen addition on soil organic carbon in freshwater marsh of Northeast China
DOI:10.1007/s12665-013-2252-z URL [本文引用: 1]
Sorption and desorption of organic matter in soils as affected by phosphate
DOI:10.1016/j.geoderma.2021.115377 URL [本文引用: 2]
Soil microbial carbon use efficiency and biomass turnover in a long-term fertilization experiment in a temperate grassland
DOI:10.1016/j.soilbio.2016.03.008 URL [本文引用: 1]
Addition of inorganic phosphorus to soil leads to desorption of organic compounds and thus to increased soil respiration
DOI:10.1016/j.soilbio.2018.12.018 URL [本文引用: 1]
Plant and microbial responses to nitrogen and phosphorus addition across an elevational gradient in subarctic tundra
Temperature and nutrients are major limiting factors in subarctic tundra. Experimental manipulation of nutrient availability along elevational gradients (and thus temperature) can improve our understanding of ecological responses to climate change. However, no study to date has explored impacts of nutrient addition along a tundra elevational gradient, or across contrasting vegetation types along any elevational gradient. We set up a full factorial nitrogen (N) and phosphorus (P) fertilization experiment in each of two vegetation types (heath and meadow) at 500 m, 800 m, and 1000 m elevation in northern Swedish tundra. We predicted that plant and microbial communities in heath or at lower elevations would be more responsive to N addition while communities in meadow or at higher elevations would be more responsive to P addition, and that fertilizer effects would vary more with elevation for the heath than for the meadow. Although our results provided little support for these predictions, the relationship between nutrient limitation and elevation differed between vegetation types. Most plant and microbial properties were responsive to N and/or P fertilization, but responses often varied with elevation and/or vegetation type. For instance, vegetation density significantly increased with N + P fertilization relative to the other fertilizer treatments, and this increase was greatest at the lowest elevation for the heath but at the highest elevation for the meadow. Arbuscular mycorrhizae decreased with P fertilization at 500 m for the meadow, but with all fertilizer treatments in both vegetation types at 800 m. Fungal to bacterial ratios were enhanced by N+ P fertilization for the two highest elevations in the meadow only. Additionally, microbial responses to fertilization were primarily direct rather than indirect via plant responses, pointing to a decoupled response of plant and microbial communities to nutrient addition and elevation. Because our study shows how two community types differ in their responses to fertilization and elevation, and because the temperature range across this gradient is approximately 3 degrees C, our study is informative about how nutrient limitation in tundra may be influenced by temperature shifts that are comparable to those expected under climate change during this century.
A global analysis of soil acidification caused by nitrogen addition
DOI:10.1088/1748-9326/10/2/024019 URL [本文引用: 1]
Multi-element regulation of the tropical forest carbon cycle
DOI:10.1890/100047 URL [本文引用: 1]
Nitrogen additions and microbial biomass: a meta-analysis of ecosystem studies
DOI:10.1111/j.1461-0248.2008.01230.x
PMID:18673384
[本文引用: 1]
Nitrogen (N) enrichment is an element of global change that could influence the growth and abundance of many organisms. In this meta-analysis, I synthesized responses of microbial biomass to N additions in 82 published field studies. I hypothesized that the biomass of fungi, bacteria or the microbial community as a whole would be altered under N additions. I also predicted that changes in biomass would parallel changes in soil CO2 emissions. Microbial biomass declined 15% on average under N fertilization, but fungi and bacteria were not significantly altered in studies that examined each group separately. Moreover, declines in abundance of microbes and fungi were more evident in studies of longer durations and with higher total amounts of N added. In addition, responses of microbial biomass to N fertilization were significantly correlated with responses of soil CO2 emissions. There were no significant effects of biomes, fertilizer types, ambient N deposition rates or methods of measuring biomass. Altogether, these results suggest that N enrichment could reduce microbial biomass in many ecosystems, with corresponding declines in soil CO2 emissions.
Comparison of carbon dynamics in tropical and temperate soils using radiocarbon measurements
DOI:10.1029/93GB00468 URL [本文引用: 1]
Biogeochemical evolution of soil organic matter composition after a decade of warming and nitrogen addition
DOI:10.1007/s10533-021-00837-0 URL [本文引用: 1]
Plant rhizodeposition: a key factor for soil organic matter formation in stable fractions
DOI:10.1126/sciadv.abd3176 URL [本文引用: 2]
Litterfall, nutrient cycling, and nutrient limitation in tropical forests
DOI:10.2307/1939481 URL [本文引用: 1]
Terrestrial phosphorus limitation: mechanisms, implications, and nitrogen-phosphorus interactions
DOI:10.1890/08-0127.1 URL [本文引用: 2]
SOM fractionation methods: relevance to functional pools and to stabilization mechanisms
DOI:10.1016/j.soilbio.2007.03.007 URL [本文引用: 9]
Linking temperature sensitivity of soil organic matter decomposition to its molecular structure, accessibility, and microbial physiology
DOI:10.1111/gcb.12112
PMID:23504889
[本文引用: 1]
Temperature sensitivity of soil organic matter (SOM) decomposition may have a significant impact on global warming. Enzyme-kinetic hypothesis suggests that decomposition of low-quality substrate (recalcitrant molecular structure) requires higher activation energy and thus has greater temperature sensitivity than that of high-quality, labile substrate. Supporting evidence, however, relies largely on indirect indices of substrate quality. Furthermore, the enzyme-substrate reactions that drive decomposition may be regulated by microbial physiology and/or constrained by protective effects of soil mineral matrix. We thus tested the kinetic hypothesis by directly assessing the carbon molecular structure of low-density fraction (LF) which represents readily accessible, mineral-free SOM pool. Using five mineral soil samples of contrasting SOM concentrations, we conducted 30-days incubations (15, 25, and 35 °C) to measure microbial respiration and quantified easily soluble C as well as microbial biomass C pools before and after the incubations. Carbon structure of LFs (<1.6 and 1.6-1.8 g cm(-3) ) and bulk soil was measured by solid-state (13) C-NMR. Decomposition Q10 was significantly correlated with the abundance of aromatic plus alkyl-C relative to O-alkyl-C groups in LFs but not in bulk soil fraction or with the indirect C quality indices based on microbial respiration or biomass. The warming did not significantly change the concentration of biomass C or the three types of soluble C despite two- to three-fold increase in respiration. Thus, enhanced microbial maintenance respiration (reduced C-use efficiency) especially in the soils rich in recalcitrant LF might lead to the apparent equilibrium between SOM solubilization and microbial C uptake. Our results showed physical fractionation coupled with direct assessment of molecular structure as an effective approach and supported the enzyme-kinetic interpretation of widely observed C quality-temperature relationship for short-term decomposition. Factors controlling long-term decomposition Q10 are more complex due to protective effect of mineral matrix and thus remain as a central question.© 2012 Blackwell Publishing Ltd.
Nitrogen deposition modifies soil carbon storage through changes in microbial enzymatic activity
DOI:10.1890/03-5120 URL [本文引用: 2]
Microbial necromass as the source of soil organic carbon in global ecosystems
DOI:10.1016/j.soilbio.2021.108422 URL [本文引用: 2]
Long-term nitrogen addition suppresses microbial degradation, enhances soil carbon storage, and alters the molecular composition of soil organic matter
DOI:10.1007/s10533-018-00535-4 URL [本文引用: 1]
N and P fertilization reduced soil autotrophic and heterotrophic respiration in a young Cunninghamia lanceolata forest
DOI:10.1016/j.agrformet.2016.08.007 URL [本文引用: 1]
Plant responses to nutrient addition experiments conducted in tropical forests
DOI:10.1002/ecm.1382 [本文引用: 2]
Plant responses to fertilization experiments in lowland, species-rich, tropical forests
DOI:10.1002/ecy.2193
PMID:29460277
[本文引用: 1]
We present a meta-analysis of plant responses to fertilization experiments conducted in lowland, species-rich, tropical forests. We also update a key result and present the first species-level analyses of tree growth rates for a 15-yr factorial nitrogen (N), phosphorus (P), and potassium (K) experiment conducted in central Panama. The update concerns community-level tree growth rates, which responded significantly to the addition of N and K together after 10 yr of fertilization but not after 15 yr. Our experimental soils are infertile for the region, and species whose regional distributions are strongly associated with low soil P availability dominate the local tree flora. Under these circumstances, we expect muted responses to fertilization, and we predicted species associated with low-P soils would respond most slowly. The data did not support this prediction, species-level tree growth responses to P addition were unrelated to species-level soil P associations. The meta-analysis demonstrated that nutrient limitation is widespread in lowland tropical forests and evaluated two directional hypotheses concerning plant responses to N addition and to P addition. The meta-analysis supported the hypothesis that tree (or biomass) growth rate responses to fertilization are weaker in old growth forests and stronger in secondary forests, where rapid biomass accumulation provides a nutrient sink. The meta-analysis found no support for the long-standing hypothesis that plant responses are stronger for P addition and weaker for N addition. We do not advocate discarding the latter hypothesis. There are only 14 fertilization experiments from lowland, species-rich, tropical forests, 13 of the 14 experiments added nutrients for five or fewer years, and responses vary widely among experiments. Potential fertilization responses should be muted when the species present are well adapted to nutrient-poor soils, as is the case in our experiment, and when pest pressure increases with fertilization, as it does in our experiment. The statistical power and especially the duration of fertilization experiments conducted in old growth, tropical forests might be insufficient to detect the slow, modest growth responses that are to be expected.© 2018 by the Ecological Society of America.
A meta-analysis of soil extracellular enzyme activities in response to global change
DOI:10.1016/j.soilbio.2018.05.001 URL [本文引用: 1]
Nonlinear responses of ecosystem carbon fluxes to nitrogen deposition in an old-growth boreal forest
DOI:10.1111/ele.13906 URL [本文引用: 1]
Disentangling the effects of nitrogen availability and soil acidification on microbial taxa and soil carbon dynamics in natural grasslands
DOI:10.1016/j.soilbio.2021.108495 URL [本文引用: 1]
Long-term, amplified responses of soil organic carbon to nitrogen addition worldwide
DOI:10.1111/gcb.15489 URL [本文引用: 15]
Aboveground litter inputs determine carbon storage across soil profiles: a meta-analysis
DOI:10.1007/s11104-021-04881-5 URL [本文引用: 1]
Decline in the contribution of microbial residues to soil organic carbon along a subtropical elevation gradient
DOI:10.1016/j.scitotenv.2020.141583 URL [本文引用: 1]
Reconciling multiple impacts of nitrogen enrichment on soil carbon: plant microbial and geochemical controls
DOI:10.1111/ele.13083 URL [本文引用: 9]
Effects of nitrogen additions on soil microbial respiration and its temperature sensitivity in a Tibetan alpine meadow
氮添加对高寒草甸土壤微生物呼吸及其温度敏感性的影响
Divergent responses of soil organic carbon accumulation to 14 years of nitrogen addition in two typical subtropical forests
DOI:10.1016/j.scitotenv.2019.136104 URL [本文引用: 1]
Sensitivity of soil carbon dynamics to nitrogen and phosphorus enrichment in an alpine meadow
DOI:10.1016/j.soilbio.2020.107984 URL [本文引用: 8]
Influence of multiple global change drivers on terrestrial carbon storage: additive effects are common
DOI:10.1111/ele.12767
PMID:28370812
[本文引用: 9]
The interactive effects of multiple global change drivers on terrestrial carbon (C) storage remain poorly understood. Here, we synthesise data from 633 published studies to show how the interactive effects of multiple drivers are generally additive (i.e. not differing from the sum of their individual effects) rather than synergistic or antagonistic. We further show that (1) elevated CO, warming, N addition, P addition and increased rainfall, all exerted positive individual effects on plant C pools at both single-plant and plant-community levels; (2) plant C pool responses to individual or combined effects of multiple drivers are seldom scale-dependent (i.e. not differing from single-plant to plant-community levels) and (3) soil and microbial biomass C pools are significantly less sensitive than plant C pools to individual or combined effects. We provide a quantitative basis for integrating additive effects of multiple global change drivers into future assessments of the C storage ability of terrestrial ecosystems.© 2017 John Wiley & Sons Ltd/CNRS.
Effects of simulated nitrogen deposition on soil respiration components and their temperature sensitivities in a semiarid grassland
DOI:10.1016/j.soilbio.2014.04.013 URL [本文引用: 1]
Soil organic carbon fractionation methods and their applications in farmland ecosystem research: a review
Soil organic carbon is of heterogeneity in components. The active components are sensitive to agricultural management, while the inert components play an important role in carbon fixation. Soil organic carbon fractionation mainly includes physical, chemical, and biological fractionations. Physical fractionation is to separate the organic carbon into active and inert components based on the density, particle size, and its spatial distribution; chemical fractionation is to separate the organic carbon into various components based on the solubility, hydrolizability, and chemical reactivity of organic carbon in a variety of extracting agents. In chemical fractionation, the dissolved organic carbon is bio-available, including organic acids, phenols, and carbohydrates, and the acid-hydrolyzed organic carbon can be divided into active and inert organic carbons. Simulated enzymatic oxidation by using KMnO4 can separate organic carbon into active and non-active carbon. Biological fractionation can differentiate microbial biomass carbon and potential mineralizable carbon. Under different farmland management practices, the chemical composition and pool capacity of soil organic carbon fractions will have different variations, giving different effects on soil quality. To identify the qualitative or quantitative relationships between soil organic carbon components and carbon deposition, we should strengthen the standardization study of various fractionation methods, explore the integrated application of different fractionation methods, and sum up the most appropriate organic carbon fractionation method or the appropriate combined fractionation methods for different farmland management practices.
土壤有机碳分组方法及其在农田生态系统研究中的应用
Global negative effects of nitrogen deposition on soil microbes
DOI:10.1038/s41396-018-0096-y URL [本文引用: 3]
Effects of climate change on molecular structure and stability of soil carbon pool: a general review
气候变化对土壤有机碳库分子结构特征与稳定性影响研究进展
The effects of nitrogen enrichment on soil CO2 fluxes depending on temperature and soil properties
DOI:10.1111/geb.12430 URL [本文引用: 1]
Different responses of soil respiration and its components to nitrogen addition among biomes: a meta- analysis
DOI:10.1111/gcb.12490 URL [本文引用: 1]
Imbalanced atmospheric nitrogen and phosphorus depositions in China: implications for nutrient limitation

{kind=link}
{kind=link}
{kind=link}
{kind=link}