


近几十年来, 化石燃料的燃烧以及化学肥料的大量使用造成全球范围内氮沉降急剧增加。草地作为地球上分布范围最广的陆地生态系统, 具有植物个体小和生活史相对较短的特点, 且比其他生态系统更易受到氮沉降的影响。氮沉降对草地生态系统的影响主要有两个方面: 一方面氮沉降可以缓解生态系统的氮限制, 对植物生长发育产生促进作用, 进而增加群落生物量(Fay et al., 2015; Kotas et al., 2017); 另一方面, 有效氮的增加打破了物种间原有的资源竞争平衡, 导致一些物种被竞争排除, 最终引起群落中物种多样性降低(Harpole & Tilman, 2007; Harpole et al., 2016)。但是不同的添加时间常常导致实验结果不同(Humbert et al., 2016), 且氮添加导致草地生物量与物种多样性变化的机制尚有争论(Isbell et al., 2013)。
当前在全球范围内开展了大量氮添加实验以探究草地生物量的响应。迄今为止, 大部分研究结果都表明氮添加增加草地群落地上生物量, 这一结论在多种类型的草地研究当中都已得到证实, 如荒漠草原(何玉惠等, 2015)、典型草原(Lü et al., 2018)、高寒草原(Li et al., 2015a)、高寒草甸(Ren et al., 2010)、沼泽化草甸(Vivanco et al., 2015)以及人工草地(Fridley, 2003)。然而, 前期的研究只关注于地上生物量, 针对氮添加条件下地下生物量变化所开展的研究较少。事实上, 草地群落的地下生物量远远大于地上生物量, 对群落总生产力变化起着重要作用。主要有两种假说来解释地上和地下生物量对氮添加的响应: 其一, 最优分配假说认为, 氮限制解除后, 植物会将一部分原来分配到地下的生物量重新分配到地上部分的茎和叶, 以获取限制性的光资源, 从而适应加剧的光竞争, 因此氮添加会导致群落地下生物量下降; 其二, 等速生长假说认为, 氮添加后植物不再受氮限制, 光合速率加快且植物个体增大, 因此植物可以将更多的光合产物等比例分配到地上和地下, 最终使得地上与地下生物量共同增加。当前的大部分氮添加实验结果都支持第一种假说(Ram et al., 1991; 辛小娟等, 2014), 但第二种假说在观察实验也获得了一些证据支持(Enquist & Niklas, 2002)。高寒草原的地下生物量对氮添加的响应符合哪种假说仍有待进一步验证。
在不同类型的草地中所开展的研究普遍证实氮添加导致植物群落物种多样性下降(Ren et al., 2010; Yang et al., 2011)。以往的研究都只关注于群落内的物种多样性变化, 对氮添加后群落功能多样性和群落水平功能性状的响应模式关注较少, 而阐明氮添加导致的群落结构和生态系统功能变化需将基于功能性状的研究纳入其中(McGill et al., 2006; Suding & Goldstein, 2008)。首先, 多样性包含不同方面, 不仅指群落内的物种多样性, 也包括功能多样性; 物种多样性与功能多样性之间关系复杂, 不仅仅局限于简单的正相关关系(Niu et al., 2014; Li et al., 2015b)。其次, 群落内物种具有不同的生理、形态和物候特征, 因此对生态系统功能的贡献不同, 而物种多样性在计算过程中将所有物种视为同等贡献, 从而导致各个物种功能性状信息的丢失。再次, 当前解释生态系统功能变化的假说可借助功能性状的途径进行验证。例如, 质量比假说认为, 生态系统功能主要是由群落内优势种的特性所决定(Grime, 1998), 物种的功能性状值与群落内多度加权获得的群落加权性状值是检验该假说的有力指标(García-Palacios et al., 2017); 生态位互补假说认为, 生态系统功能由群落内物种的生态位分化程度所决定(Tilman et al., 1997; Hector, 1998), 功能离散度作为衡量生态位分化的经典指标可以对该假说进行验证(Luo et al., 2021)。此外, 氮添加后群落的变化应该是一个层级响应的过程(Smith et al., 2009): 首先是功能性状的变化, 然后是物种多度的变化, 最终导致物种的迁入与迁出, 即物种多样性变化。因此, 在短期氮添加的条件下群落水平上的功能性状变化可能比物种多样性变化更加敏感, 进而更好地解释群落生物量的变化。
位于天山山脉南麓的巴音布鲁克草原是新疆乃至中亚的生物多样性热点地区, 同时也是新疆最大的畜牧业基地, 在物种保护、水源涵养、水土保持及南疆水资源调配方面具有重要意义。前期的研究表明该地区的氮沉降量已接近当地生态系统功能变化的阈值(李凯辉, 2012), 阐明当地生产力与多样性对氮添加的响应机制已成为迫切需求。因此本研究在当地建立氮添加实验, 通过调查功能性状和群落物种组成变化, 基于性状和物种两种途径, 试图回答以下科学问题: 1)氮添加第一年与第二年群落地上与地下生物量如何变化? 物种多样性、功能多样性与群落水平性状值对短期氮添加如何响应? 2)基于功能多样性和物种多样性的途径对生物量变化的解释能力如何?
1 材料和方法
1.1 区域概况
本研究的研究地点位于天山山脉南麓的巴音布鲁克草原, 实验样地选在中国科学院巴音布鲁克草原生态系统研究站(42.52°-42.53° N, 83.41°-83.43° E)。研究地点海拔约2 470 m, 年降水量265.7 mm, 年平均气温-4.8 ℃, 年蒸发量1 022.9-1 247.5 mm, 年日照时间2 466-2 616 h, 全年积雪日达150-180 天, 无绝对无霜期, 属于典型的高寒气候(李凯辉, 2012)。植被类型为高寒草原, 土壤为栗钙土, 优势物种为羊茅(Festuca ovina)、苔草(Koeleria cristata)、草地早熟禾(Poa pratensis)等。
1.2 实验设计
2018年4月, 选择生境均匀的典型高寒草地建立实验样地, 使用围栏进行围封, 避免马、牛、羊等大型家畜的放牧, 但是小型野生哺乳动物(如鼠兔)可以出入。氮添加实验于2018年5月开始, 构建包含对照与氮添加两种处理的实验样地, 每块样地大小为5 m × 5 m, 均匀分成两个半区, 其中一个半区进行样方调查, 另一个半区进行植物与土壤采样。样地间用1 m缓冲带隔开。样地分布采用完全随机设计, 每个处理共有6个重复。选用尿素(CO(NH2)2) 进行氮添加, 每m2添加21.4 g尿素以保证纯氮添加量为10 g·m-2。每年5月中旬将预先称好的颗粒状肥料一次性均匀撒在样方内。为了保证养分添加效果, 养分添加选择在阴雨天进行。
1.3 功能性状测量
表1 天山高寒草地氮添加2年后群落内常见物种相对多度变化(平均值±标准误)
Table 1
| 种名 Species | PFG | RAC | RAN |
|---|---|---|---|
| 冰草 Agropyron cristatum | G | 15.28 ± 0.05 | 31.64 ± 0.01 |
| 草地早熟禾 Poa pratensis | G | 13.51 ± 0.02 | 17.30 ± 0.03 |
| 苔草 Koeleria cristata | G | 15.77 ± 0.02 | 2.02 ± 0.01 |
| 细果薹草 Carex stenocarpa | G | 0.54 ± 0.01 | 0.01 |
| 羊茅 Festuca ovina | G | 0.20 | 32.35 ± 0.01 |
| 紫花针茅 Stipa purpurea | G | 7.45 ± 0.01 | 6.28 ± 0.02 |
| 二裂委陵菜 Potentilla bifurca | F | 5.15 ± 0.01 | 3.27 ± 0.01 |
| 莓叶委陵菜 Potentilla fragarioides | F | 4.23 ± 0.01 | 0.98 |
| 多裂委陵菜 Potentilla multifida | F | 0.05 | 0.01 |
| 天山蒲公英 Taraxacum tianschanicum | F | 0.04 ± 0.01 | 0.21 ± 0.01 |
| 防风 Saposhnikovia divaricata | F | 0.48 ± 0.01 | 0.25 ± 0.01 |
| 斜茎黄耆 Astragalus adsurgens | L | 29.05 ± 0.02 | 4.72 ± 0.01 |
| 小花棘豆 Oxytropis glabra | L | 7.77 ± 0.02 | 0.46 |
PFG, 植物功能群; RAC, 对照中相对多度; RAN, 氮添加处理中相对多度。F, 杂类草; G, 禾草类; L, 豆科。
PFG, plant functional group; RAC, relative abundance in control plots; RAN, relative abundance in nitrogen addition plots. F, forbs; G, graminoids; L, legume.
于2018与2019年的8月初(即氮添加第一年与第二年), 在每块样地中对每个物种分别随机选择3株完整植株剪下后带回实验室, 使用直尺测量植株高度; 从每个植株上选取一片健康成熟叶片使用扫描仪测定叶面积, 烘干后使用电子天平测定叶片干质量, 精确至0.000 1 g。同期, 在样地中采集足量常见种的叶片, 在室内烘箱70 ℃烘干, 研磨过筛进行叶片碳、氮、磷含量测定。叶片碳含量测定使用重铬酸钾外加热法, 叶片氮含量测定使用半微量凯氏定氮法, 叶片磷含量测定使用钼锑抗比色法(鲍士旦, 2012)。种子采集于每年的8月初至9月中旬进行, 在种子充分成熟并自然脱落时采集, 采集时挑选植株将繁殖部分整体剪下装入信封。在室内风干(温度约15 ℃)后, 对种子进行剥离、筛选并将种子的附属结构去除(如菊科的冠毛和禾本科的芒)仅保留胚、胚乳和种皮。每个物种在将种子混合后再随机选取100粒种子称质量, 重复3次并精确至0.000 1 g。本研究中种子质量均为百粒质量。
1.4 群落物种组成与生物量测定
每年8月中旬, 使用样方法对群落物种组成进行调查, 样方大小为0.5 m × 0.5 m。通过目测法估计样方总盖度和物种分盖度, 统计样方中物种个体数(禾草类每个无性分株记为一株), 随后按物种剪取其地上部分装入信封, 在温度为70 ℃的烘箱进行烘干后称质量, 将每个物种的生物量相加获得群落地上生物量。最后, 根据当地植物根系普遍的聚集分布情况(柳妍妍等, 2013), 在每个样方内使用土钻钻取3个20 cm深度的土壤样品, 混装至土壤袋中, 使用50目的筛子将植物根系与土壤分离, 土壤用于理化性质测量, 植物根系用水充分洗净, 然后在70 ℃烘干后称质量并换算为地下生物量。
1.5 数据分析
物种的相对多度为样方中每个物种地上生物量除以样方中所有物种的地上生物量总和(Zhou et al., 2016)。使用群落均值加权性状值(CWM traits)获得对照与氮添加处理中群落水平上的功能性状变化。计算公式如下: CWM = ∑Mi × Traiti, 式中Mi代表特定样方内第i个物种的生物量, Traiti是特定样方内第i个物种的功能性状值。使用物种丰富度、香农-维纳指数、辛普森指数和均匀度指数来表征物种多样性; 本研究选用4种指标来代表功能多样性: 功能丰富度、功能离散度、功能均匀度和Rao指数, 其中功能丰富度表示群落内物种所占据的功能空间的规模; 功能均匀度表示具有不同性状的物种在功能空间里分布的均匀程度; 功能离散度表示群落内不同物种在功能空间内分布的分散程度; Rao指数则包含了功能丰富度和功能离散度两方面的信息, 取决于群落内物种所占据的功能空间范围和物种之间的相似度(Mason et al., 2005)。
获得上述指标后, 首先使用非参数检验(Wilcoxon test)比较氮添加处理与对照中的各个指标之间的差异是否显著。因为地上生物量、地下生物量和总生物量对氮添加的响应模式相同, 所以在后续分析中只使用总生物量。然后对总生物量和物种多样性、功能多样性及群落均值加权性状值分别进行一元线性回归。随后使用上述指标构建多元回归模型, 并通过逐步回归筛选出最优指标组合, 并对模型的方差膨胀因子(VIF)进行检验。最后挑选一元线性回归中显著的指标对其标准化后构建多元回归模型, 并对各个指标的独立效应进行分解, 进而比较各个指标对总生物量的解释能力。
所有数据均在R语言中进行分析, 物种多样性的各类指标使用“vegan”包, 群落均值加权性状值与功能多样性的各个指标使用“FD”包, 多元线性模型筛选及检验用到了“MASS”、“car”及“hier.part”包, 绘图使用“ggplot2”包。
2 结果和分析
2.1 短期氮添加对群落物种相对多度的影响
两年的氮添加显著增加了冰草(Agropyron cristatum)与羊茅这两种禾草的相对多度, 分别由对照中的15.28%和0.20%增加到氮添加处理中的31.64%和32.35%。同时, 氮添加导致两种豆科植物(斜茎黄耆(Astragalus adsurgens)和小花棘豆(Oxytropis glabra))的相对多度分别从对照中的29.05%和7.77%下降到4.72%和0.46% (表1)。氮添加2年之后当地群落由禾本科(52.21%)与豆科(36.82%)共同占优势, 转变为禾本科植物占据绝对优势(89.59%)。
2.2 群落生物量、物种多样性、功能多样性及群落水平功能性状对短期氮添加的响应
氮添加第一年与第二年, 地上地下生物量、物种多样性及功能多样性各指标的变化趋势基本一致。氮添加第一年, 相较于对照, 氮添加处理中地上、地下与总生物量分别增加60.25%、21.75%和25.91%, 但是均未达到显著水平; 氮添加第二年, 群落地上、地下和总生物量均显著增加(p < 0.05), 增加幅度分别为64.49%、21.66%和31.56% (图1)。表征群落物种多样性的4个指标在氮添加第一年与第二年均有降低趋势, 但没有显著差异(图2)。氮添加导致群落Rao指数和功能离散度降低, 在氮添加第一年未达到显著水平, 在氮添加第二年达到显著水平, 但是氮添加对功能丰富度和功能均匀度没有显著影响(图3)。氮添加第一年与第二年群落水平功能性状的变化趋势一致, 即群落水平上的植物高度和叶片碳含量显著增加, 比叶面积、种子质量与叶片磷含量均显著降低, 而叶片氮含量没有显著差异(图4)。在功能群水平上, 禾草类的植株高度(11.76 cm)高于杂类草(8.38 cm)而其比叶面积(125 cm2·g-1)则低于杂类草(143 cm2·g-1)和豆科(169 cm2·g-1), 而豆科植物种子质量(0.253 g)和叶片氮含量(45.93 mg·g-1)大于禾草类(种子质量0.078 g, 叶片氮含量13.56 mg·g-1)和杂类草(种子质量0.04 g, 叶片氮含量25.51 mg·g-1)。
图1
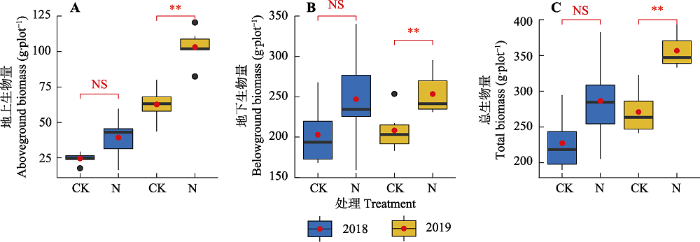
图1
氮添加对天山高寒草地群落生物量的影响(平均值±标准误)。CK, 对照; N, 氮添加。**, p < 0.01; NS, p > 0.05。
Fig. 1
Effects of nitrogen addition on community biomass in Tianshan alpine grassland (mean ± SE). CK, control; N, nitrogen addition. **, p < 0.01; NS, p > 0.05.
图2
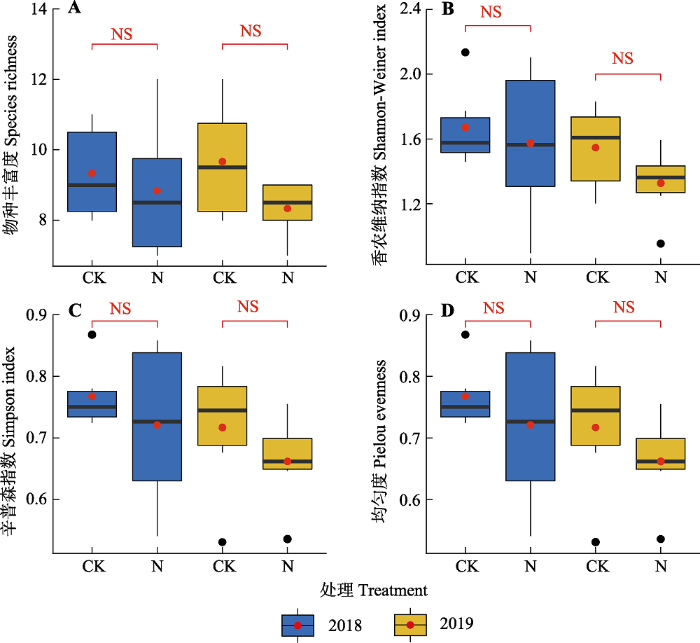
图2
氮添加对天山高寒草地群落物种多样性的影响(平均值±标准误)。CK, 对照; N, 氮添加。NS, p > 0.05。
Fig. 2
Effects of nitrogen addition on species diversity in Tianshan alpine grassland (mean ± SE). CK, control; N, nitrogen addition. NS, p > 0.05.
图3
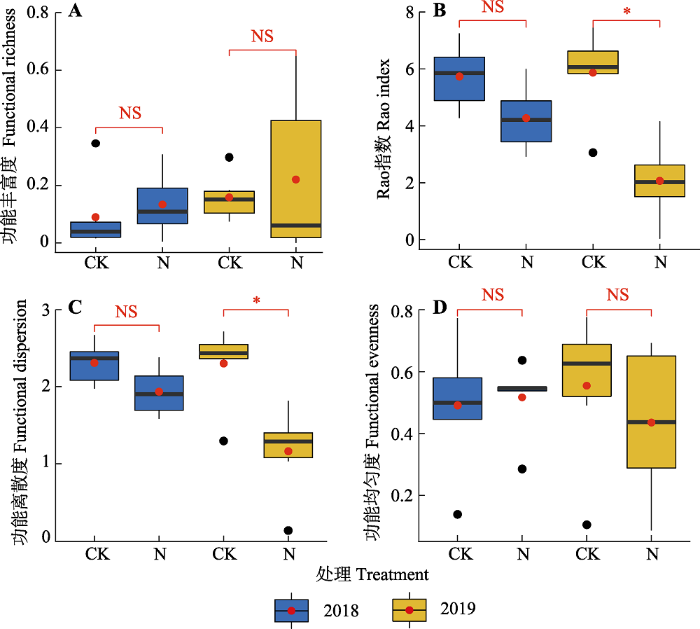
图3
氮添加对天山高寒草地群落功能多样性的影响(平均值±标准误)。CK, 对照; N, 氮添加。*, p < 0.05; NS, p > 0.05。
Fig. 3
Effects of nitrogen addition on functional diversity in Tianshan alpine grassland (mean ± SE). CK, control; N, nitrogen addition. *, p < 0.05; NS, p > 0.05.
图4
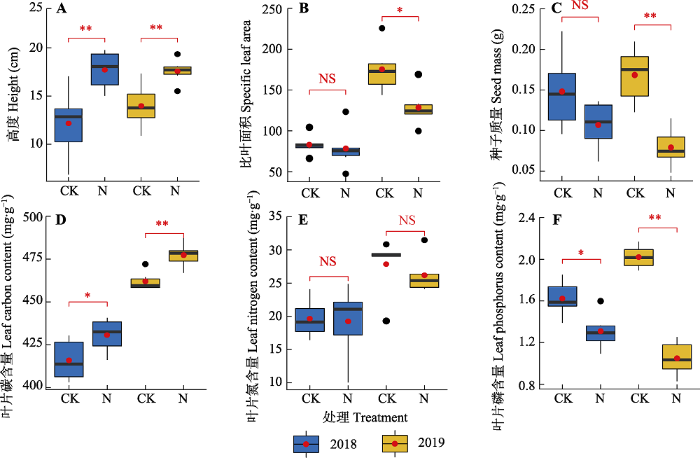
图4
氮添加对天山高寒草地群落水平功能性状群落均值加权性状值的影响(平均值±标准误)。CK, 对照; N, 氮添加。*, p < 0.05; **, p < 0.01; NS, p > 0.05。
Fig. 4
Effects of nitrogen addition on community level functional traits and community weighted mean in Tianshan alpine grassland (mean ± SE). CK, control; N, nitrogen addition. *, p < 0.05; **, p < 0.01; NS, p > 0.05。
2.3 群落生物量与物种多样性、功能多样性和群落功能性状之间的关系
群落生物量与物种多样性之间不存在显著的线性回归关系。氮添加第一年群落生物量与功能多样性之间不存在显著回归关系, 氮添加第二年群落生物量随着Rao指数(斜率(slope) = -17.65, p < 0.05)和功能离散度(slope = -51.13, p < 0.05)的增加而下降(图5)。群落生物量与群落水平功能性状之间的关系较为复杂, 氮添加第二年群落生物量随着高度(slope = 12.71, p < 0.05)和叶片碳含量(slope = 3.50, p < 0.05)的增加而增加; 而群落生物量随着比叶面积(slope = -1.19, p < 0.05)、种子质量(slope = -789.32, p < 0.05)和叶片磷含量(slope = -85.95, p < 0.001)的增加而下降; 氮添加第一年群落生物量随着叶片氮含量增加而下降(slope = -7.81, p < 0.05), 与其他功能性状之间均未出现显著关系(图6)。
图5
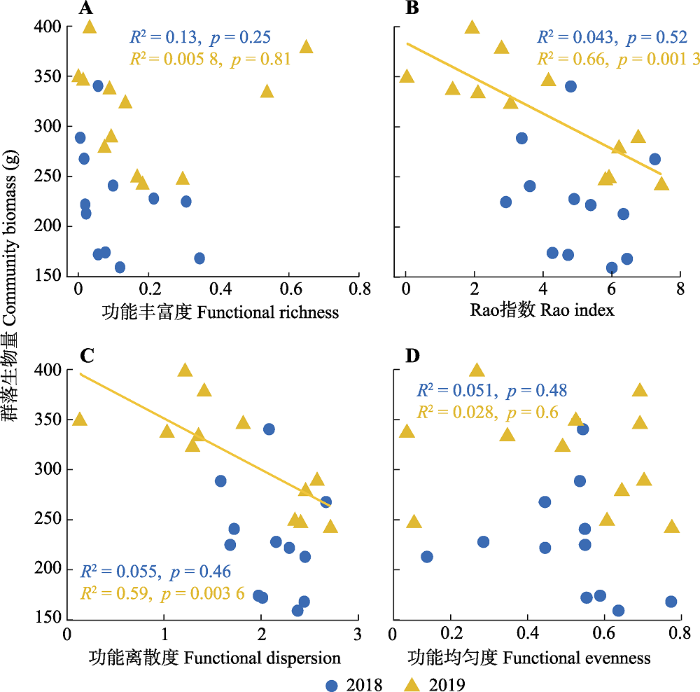
图5
天山高寒草地群落总生物量与功能多样性之间的线性回归关系。
Fig. 5
Linear regression relationships between total community biomass and functional diversity in Tianshan alpine grassland.
图6
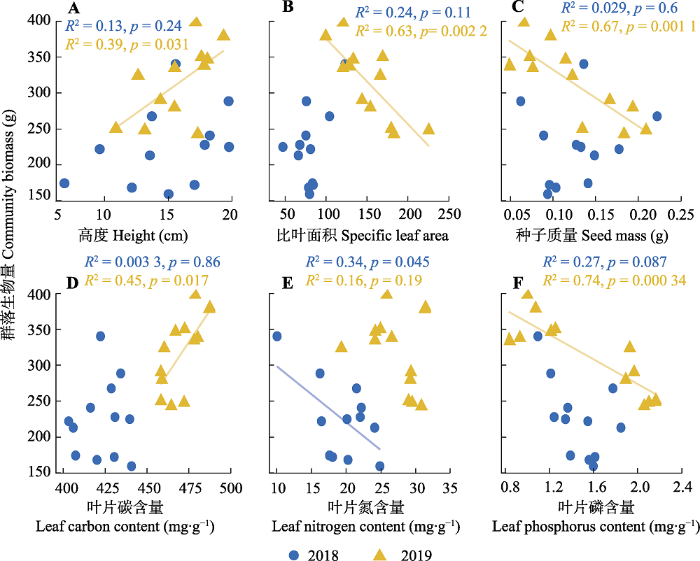
图6
天山高寒草地群落总生物量与功能性状之间的线性回归关系。
Fig. 6
Linear regression relationships between total community biomass and functional traits in Tianshan alpine grassland.
逐步回归结果表明, 对群落生物量解释最优的指标组合为功能离散度+种子质量+叶片碳含量+叶片磷含量的组合, 该组合可以解释高达69.75%的群落生物量变化。对VIF进行检验的结果表明二者膨胀系数(VIF = 1)模型符合标准(VIF < 10)。挑选一元回归中显著的变量标准化后拟合多元回归模型并对每个变量的解释能力进行比较, 结果表明解释能力最好的指标为叶片碳含量(26.13%), 解释能力最低的指标为叶片氮含量(4.88%), 功能离散度和Rao指数分别可以解释14.5%和12.96%。
3 讨论
3.1 短期氮添加同时增加天山高寒草地的地上和地下生物量
研究结果表明, 为期两年的氮添加导致高寒草地群落地上、地下及总生物量均显著增加(图1)。与前期的研究结果(Ren et al., 2010)一致, 地上生物量的增加主要是禾草类生物量的增加所致。在本研究中, 冰草和羊茅两种禾草的生物量增加尤为显著, 二者在群落中的生物量占比分别从对照中的15.28%和0.20%增加到氮添加处理中的31.64%和32.35% (表1), 这可能与两种植物高度较高且可以无性繁殖有关, 使得它们能够对氮添加产生快速的响应。最优资源分配假说认为在养分充足的环境中(如氮添加), 植物会将更多的资源分配到地上部分以获得充足的光资源, 从而导致地上生物量增加而地下生物量减少, 本研究的结果并不支持该假说。在本研究中地上与地下生物量同时增加, 但是地上生物量的增加比例(64.5%)要高于地下生物量的增加比例(21.7%), 因此也不能严格支持等速生长假说。本研究中地下生物量的增加可能有两方面原因: 一是解除氮限制以后植物获得更多的光合产物进而向地下部分分配; 二是氮添加处理中须根系的禾本科植物快速增加占据了原来土壤中的空缺。
3.2 短期氮添加降低天山高寒草地的功能多样性
研究结果表明短期氮添加未导致功能丰富度和功能均匀度发生变化, 但是显著降低功能离散度和Rao指数(图3)。这说明短期氮添加并未导致群落功能空间规模变化, 功能性状值的范围在对照与短期氮添加处理中没有太大变化; 不同物种在功能空间内分布的均匀程度也几乎没有变化。然而, 氮添加后功能离散度显著降低, 表明优势种在功能空间中的分布呈现聚集趋势, 这可能是由于氮添加群落中物种的功能性状都朝着相同方向变化, 如增加高度以适应加剧的光竞争, 增加叶片氮含量以提高光合效率(图4)。功能离散度与群落内物种的生态位占据与资源利用程度具有密切联系, 氮添加导致功能离散度下降, 表明氮添加导致群落内优势种朝着功能空间中心靠近, 对资源的竞争加剧。这可能是由两方面的原因造成的: 首先, 生态位维度-多样性假说指出土壤中的限制性因子驱动的生态位分化是维系群落内物种共存的重要机制, 施加氮肥后氮不再成为限制性因子, 因此导致生态位维度减少和物种丧失(Harpole & Tilman, 2007); 其次, 氮添加后群落内的物种之间的竞争由对地下土壤养分的竞争转为对地上光资源的竞争, 导致对光资源和地上空间资源的竞争加剧(Hautier et al., 2009)。
本研究中为期两年的氮添加对物种多样性未产生显著影响(图2), 这可能是因为实验处理年限相对较短, 群落结构尚未达到稳定状态。该结果也表明氮添加在不引起物种多样性变化的情况下可以导致功能多样性变化, 这和以前的研究结果有着很大的不同。如Li等(2015b)在高寒草甸为期4年的氮磷添加实验表明, 养分添加导致物种丰富度和香农-维纳多样性指数下降, 但是功能丰富度与Rao指数保持不变, 养分添加后群落内物种的生态位分化和性状冗余物种的保险作用对该结果有着重要的贡献。而Niu等(2014)的研究表明施肥第二年和第三年后物种丰富度与香农-维纳多样性指数下降而功能丰富度与Rao指数增加, 因为群落内存活物种的生态位分化加剧, 抵消了物种丧失的影响。同样, 前期在青藏高原的研究表明, 为期5年的氮添加导致物种丰富度和功能丰富度同时下降, 因为物种的丧失同时导致功能性状从群落中消失(Zhou et al., 2017)。本研究与上述研究的不同之处在于上述研究均在高寒草甸群落中开展, 当地群落的物种数目(每m2五六十种)和稀有种数目更多, 因此性状冗余物种更多, 而且青藏高原高寒草甸群落植物平均高度更高, 氮添加以后可以很快地在群落上层形成郁闭空间, 强烈的光竞争导致稀有种迅速丧失。然而, 本研究的研究地点位于天山高寒草原, 植株高度较低, 稀有种较少, 导致物种丧失可能需要更长时间。综上, 我们认为氮添加以后群落功能多样性的变化取决于土壤养分与光照环境变化所导致的性状分化过程与物种丧失所导致的性状丢失过程之间的权衡, 而权衡的结果则与氮添加的时间和群落组成的初始状态有关。
3.3 短期氮添加改变天山高寒草地群落水平功能性状
本研究中5种功能性状在群落水平上对氮添加有着显著的响应, 其中高度与叶片碳含量在氮添加处理中显著增加, 而比叶面积、种子质量和叶片磷含量则随着氮添加而降低。由群落水平功能性状的计算公式可知, 群落水平上功能性状的变化受到功能性状与物种多度两方面的影响。与前期研究结果(Li et al., 2015b; Zhou et al., 2018)相一致, 本研究中氮添加导致群落水平上高度显著增加, 这主要有两方面的原因: 一是氮添加后群落内物种的资源竞争由地下的养分竞争转变为地上的光竞争, 因此群落内物种增加高度以获得更多光资源; 二是氮添加后高度更高的禾草类植物的相对多度增加。本研究中群落水平上比叶面积和叶片碳含量变化主要是由于禾草类植物大量增加, 与杂类草相比, 禾草类植物具有更加致密紧实的叶片, 比叶面积较小而叶片碳含量高, 因此禾草类植物的大量增加引起群落水平上比叶面积的降低和叶片碳含量的增加。与此同时, 群落水平上种子质量的下降主要是由于豆科植物相对多度降低所致, 因为在我们的研究地点豆科植物有着远大于杂类草和禾草类植物的种子质量。叶片磷含量的降低主要是由于稀释效应导致, 即氮添加后群落内物种的叶面积普遍增大而叶片磷含量变化不大, 因此单位面积上的磷含量下降。与其他研究不同的是, 本研究中群落水平上氮添加处理中叶片氮含量没有增加, 这可能是因为豆科植物具有较高的叶片氮含量, 氮添加后豆科植物的多度降低抵消了群落内其他物种叶片氮含量增加的影响。
本研究结果表明, 群落水平上的功能性状对氮添加的响应较为敏感, 结合功能多样性与物种多样性的变化模式, 可以认为氮添加后群落的变化是一个层级响应的过程: 群落内的物种首先通过调节功能性状适应环境的变化, 具有不同性状的物种的表现与适应能力不同, 因此导致群落内物种相对多度的改变, 而竞争排除与物种丧失过程则相对滞后。
3.4 功能多样性与群落水平功能性状能更好解释群落生物量变化
本研究中表征物种多样性的4个指标与群落生物量之间均不存在显著的回归关系, 表明物种多样性对群落生物量变化的解释能力非常有限。这可能是由于群落生物量与物种多样性变化不同步所导致, 在本研究中地上、地下和总生物量在氮添加后均显著增加, 而表征物种多样性的几个指标则均未发生显著变化。与此相反, 表征功能多样性的4个指标中有2个指标(功能离散度和Rao指数)可以很好地解释群落生物量变化, 同时6种功能性状均可以很好地解释群落生物量变化。多元回归和逐步筛选的结果也表明功能离散度、种子质量、叶片碳含量和叶片磷含量是解释生物量变化的最优组合指标, 该模型可以解释69.75%的生物量变化。上述结果表明功能性状在解释短期氮添加导致的生物量变化中起到了重要作用, 忽略物种之间性状差异的物种多样性会导致信息的丢失, 无法有效地解释短期氮添加群落的生物量变化。在本研究中, 生物量与5种群落水平功能性状之间的关系在氮添加第一年并不显著, 在氮添加第二年则变得显著(图6), 这说明氮添加对两者关系的影响具有累积效应, 随着氮添加时间的增加, 生物量与群落水平功能性状的线性回归关系均变得显著(图1, 图4)。
本研究中群落生物量与功能离散度之间呈负的线性回归关系, 说明生态位互补假说并未得到支持。质量比假说认为群落生物量主要由优势种的特征所决定, 群落均值加权性状值可以作为验证该假说的有力指标(Häger & Avalos, 2017)。本研究中生物量与群落均值加权性状值之间普遍存在着线性回归关系, 因此很好地支持了质量比假说。综上所述, 我们的研究结果符合质量比假说, 说明在短期氮添加群落中优势种的功能性状驱动了生物量的增加。
综上, 本研究表明天山高寒草地中, 功能多样性与群落水平功能性状对短期氮添加响应迅速, 而物种多样性对短期氮添加的响应不敏感; 物种多样性变化与群落生物量变化并不是同步发生, 生物量变化快于物种多样性变化, 氮添加导致群落地上和地下生物量同时增加, 但是地上生物量的增加比例高于地下生物量; 在短期氮添加群落中, 物种多样性对生物量变化的解释能力非常有限, 而基于功能性状的途径可以很好地解释群落的生物量变化。优势植物类群在生物量变化中起着决定作用。
致谢
感谢中国科学院巴音布鲁克草原生态系统研究站在野外工作中提供便利。
参考文献
Global allocation rules for patterns of biomass partitioning in seed plants
A general allometric model has been derived to predict intraspecific and interspecific scaling relationships among seed plant leaf, stem, and root biomass. Analysis of a large compendium of standing organ biomass sampled across a broad sampling of taxa inhabiting diverse ecological habitats supports the relations predicted by the model and defines the boundary conditions for above- and below-ground biomass partitioning. These canonical biomass relations are insensitive to phyletic affiliation (conifers versus angiosperms) and variation in averaged local environmental conditions. The model thus identifies and defines the limits that have guided the diversification of seed plant biomass allocation strategies.
Grassland productivity limited by multiple nutrients
DOI:10.1038/nplants.2015.80
PMID:27250253
[本文引用: 1]

Terrestrial ecosystem productivity is widely accepted to be nutrient limited(1). Although nitrogen (N) is deemed a key determinant of aboveground net primary production (ANPP)(2,3), the prevalence of co-limitation by N and phosphorus (P) is increasingly recognized(4-8). However, the extent to which terrestrial productivity is co-limited by nutrients other than N and P has remained unclear. Here, we report results from a standardized factorial nutrient addition experiment, in which we added N, P and potassium (K) combined with a selection of micronutrients (K+μ), alone or in concert, to 42 grassland sites spanning five continents, and monitored ANPP. Nutrient availability limited productivity at 31 of the 42 grassland sites. And pairwise combinations of N, P, and K+μ co-limited ANPP at 29 of the sites. Nitrogen limitation peaked in cool, high latitude sites. Our findings highlight the importance of less studied nutrients, such as K and micronutrients, for grassland productivity, and point to significant variations in the type and degree of nutrient limitation. We suggest that multiple-nutrient constraints must be considered when assessing the ecosystem-scale consequences of nutrient enrichment.
Diversity effects on production in different light and fertility environments: an experiment with communities of annual plants
DOI:10.1046/j.1365-2745.2003.00775.x URL [本文引用: 1]
Contrasting mass-ratio vs. niche complementarity effects on litter C and N loss during decomposition along a regional climatic gradient
DOI:10.1111/1365-2745.12730 URL [本文引用: 1]
Benefits of plant diversity to ecosystems: immediate, filter and founder effects
DOI:10.1046/j.1365-2745.1998.00306.x URL [本文引用: 1]
Do functional diversity and trait dominance determine carbon storage in an altered tropical landscape
DOI:10.1007/s00442-017-3880-x
PMID:28516322
[本文引用: 1]
Altered landscapes play a major role in biodiversity conservation and carbon (C) storage in the tropics. There is increasing evidence that C storage potential is controlled by tree functional diversity, but underlying mechanisms are debated. We analyzed the effects of trait dominance (mass-ratio hypothesis), species diversity, and trait variation (species complementarity) on C storage in the soils and vegetation of 20 agroforestry systems (AFS) and seven forested sites in Costa Rica. AFS consisted of organic and conventional coffee farms and pastures with trees. We used the community weighted mean (CWM) to measure trait dominance, and functional divergence (FDvar) to evaluate trait variation of wood densities (WD) and maximum heights (H ) of woody plants at each site. Species richness, the number of woody plants per hectare, and slope of the terrain were also considered as independent variables. Soil organic carbon (SOC) increased with higher CWM and with higher variability of H ([Formula: see text]) across land-use types. Aboveground carbon (AGC) was controlled by the number of woody plants per hectare and by species richness. Our results suggest that dominant traits as well as species complementarity play an important role in determining C storage. Diverse, multilayered AFS which incorporate trees with high WD, combined with the conservation of remnant forests, can maximize C storage in the soils and vegetation of altered tropical landscapes.
Grassland species loss resulting from reduced niche dimension
DOI:10.1038/nature05684 URL [本文引用: 2]
Addition of multiple limiting resources reduces grassland diversity
DOI:10.1038/nature19324 URL [本文引用: 1]
Competition for light causes plant biodiversity loss after eutrophication
DOI:10.1126/science.1169640
PMID:19407202
[本文引用: 1]
Human activities have increased the availability of nutrients in terrestrial and aquatic ecosystems. In grasslands, this eutrophication causes loss of plant species diversity, but the mechanism of this loss has been difficult to determine. Using experimental grassland plant communities, we found that addition of light to the grassland understory prevented the loss of biodiversity caused by eutrophication. There was no detectable role for competition for soil resources in diversity loss. Thus, competition for light is a major mechanism of plant diversity loss after eutrophication and explains the particular threat of eutrophication to plant diversity. Our conclusions have implications for grassland management and conservation policy and underscore the need to control nutrient enrichment if plant diversity is to be preserved.
Effect of nitrogen addition on species diversity and plant productivity of herbaceous plants in desert grassland of the Loess Plateau
氮素添加对黄土高原荒漠草原草本植物物种多样性和生产力的影响
The effect of diversity on productivity: detecting the role of species complementarity
DOI:10.2307/3546380 URL [本文引用: 1]
Impacts of nitrogen addition on plant biodiversity in mountain grasslands depend on dose, application duration and climate: a systematic review
DOI:10.1111/gcb.12986 URL [本文引用: 1]
Nutrient enrichment, biodiversity loss, and consequent declines in ecosystem productivity
DOI:10.1073/pnas.1310880110
PMID:23818582
[本文引用: 1]
Anthropogenic drivers of environmental change often have multiple effects, including changes in biodiversity, species composition, and ecosystem functioning. It remains unknown whether such shifts in biodiversity and species composition may, themselves, be major contributors to the total, long-term impacts of anthropogenic drivers on ecosystem functioning. Moreover, although numerous experiments have shown that random losses of species impact the functioning of ecosystems, human-caused losses of biodiversity are rarely random. Here we use results from long-term grassland field experiments to test for direct effects of chronic nutrient enrichment on ecosystem productivity, and for indirect effects of enrichment on productivity mediated by resultant species losses. We found that ecosystem productivity decreased through time most in plots that lost the most species. Chronic nitrogen addition also led to the nonrandom loss of initially dominant native perennial C4 grasses. This loss of dominant plant species was associated with twice as great a loss of productivity per lost species than occurred with random species loss in a nearby biodiversity experiment. Thus, although chronic nitrogen enrichment initially increased productivity, it also led to loss of plant species, including initially dominant species, which then caused substantial diminishing returns from nitrogen fertilization. In contrast, elevated CO2 did not decrease grassland plant diversity, and it consistently promoted productivity over time. Our results support the hypothesis that the long-term impacts of anthropogenic drivers of environmental change on ecosystem functioning can strongly depend on how such drivers gradually decrease biodiversity and restructure communities.
Linking above- and belowground responses to 16 years of fertilization, mowing, and removal of the dominant species in a temperate grassland
DOI:10.1007/s10021-016-0031-x URL [本文引用: 1]
Response of alpine grassland to elevated nitrogen deposition and water supply in China
DOI:10.1007/s00442-014-3122-4 URL [本文引用: 1]
Plant functional diversity can be independent of species diversity: observations based on the impact of 4-yrs of nitrogen and phosphorus additions in an alpine meadow
DOI:10.1371/journal.pone.0136040 URL [本文引用: 3]
Vertical differentiation of plant species diversity and biomass in alpine grassland in the middle section of Tianshan Mountains southern slope, Xinjiang of Northwest China
天山南坡中段高寒草地物种多样性与生物量的垂直分异特征
Testing nitrogen and water co-limitation of primary productivity in a temperate steppe
DOI:10.1007/s11104-018-3791-6 URL [本文引用: 1]
Plant traits and soil fertility mediate productivity losses under extreme drought in C3 grasslands
DOI:10.1002/ecy.3465 [本文引用: 1]
Functional richness, functional evenness and functional divergence: the primary components of functional diversity
DOI:10.1111/j.0030-1299.2005.13886.x URL [本文引用: 1]
Rebuilding community ecology from functional traits
DOI:10.1016/j.tree.2006.02.002 URL [本文引用: 1]
Fertilization decreases species diversity but increases functional diversity: a three-year experiment in a Tibetan alpine meadow
DOI:10.1016/j.agee.2013.07.015 URL [本文引用: 2]
Effect of fertilizer on plant biomass distribution and net accumulation rate in an alpine meadow in central Himalaya, India
DOI:10.2307/4002312 URL [本文引用: 1]
Effects of resource additions on species richness and ANPP in an alpine meadow community
DOI:10.1093/jpe/rtp034 URL [本文引用: 3]
A framework for assessing ecosystem dynamics in response to chronic resource alterations induced by global change
In contrast to pulses in resource availability following disturbance events, many of the most pressing global changes, such as elevated atmospheric carbon dioxide concentrations and nitrogen deposition, lead to chronic and often cumulative alterations in available resources. Therefore, predicting ecological responses to these chronic resource alterations will require the modification of existing disturbance-based frameworks. Here, we present a conceptual framework for assessing the nature and pace of ecological change under chronic resource alterations. The "hierarchical-response framework" (HRF) links well-documented, ecological mechanisms of change to provide a theoretical basis for testing hypotheses to explain the dynamics and differential sensitivity of ecosystems to chronic resource alterations. The HRF is based on a temporal hierarchy of mechanisms and responses beginning with individual (physiological/metabolic) responses, followed by species reordering within communities, and finally species loss and immigration. Each mechanism is hypothesized to differ in the magnitude and rate of its effects on ecosystem structure and function, with this variation depending on ecosystem attributes, such as longevity of dominant species, rates of biogeochemical cycling, levels of biodiversity, and trophic complexity. Overall, the HRF predicts nonlinear changes in ecosystem dynamics, with the expectation that interactions with natural disturbances and other global-change drivers will further alter the nature and pace of change. The HRF is explicitly comparative to better understand differential sensitivities of ecosystems, and it can be used to guide the design of coordinated, cross-site experiments to enable more robust forecasts of contemporary and future ecosystem dynamics.
Testing the Holy Grail framework: using functional traits to predict ecosystem change
DOI:10.1111/j.1469-8137.2008.02650.x PMID:19138225 [本文引用: 1]
Plant diversity and ecosystem productivity: theoretical considerations
Ecosystem processes are thought to depend on both the number and identity of the species present in an ecosystem, but mathematical theory predicting this has been lacking. Here we present three simple models of interspecific competitive interactions in communities containing various numbers of randomly chosen species. All three models predict that, on average, productivity increases asymptotically with the original biodiversity of a community. The two models that address plant nutrient competition also predict that ecosystem nutrient retention increases with biodiversity and that the effects of biodiversity on productivity and nutrient retention increase with interspecific differences in resource requirements. All three models show that both species identity and biodiversity simultaneously influence ecosystem functioning, but their relative importance varies greatly among the models. This theory reinforces recent experimental results and shows that effects of biodiversity on ecosystem functioning are predicted by well-known ecological processes.
Nonlinear responses in salt marsh functioning to increased nitrogen addition
Salt marshes provide storm protection to shorelines, sequester carbon (C), and mitigate coastal eutrophication. These valuable coastal ecosystems are confronted with increasing nitrogen (N) inputs from anthropogenic sources, such as agricultural runoff, wastewater, and atmospheric deposition. To inform predictions of salt marsh functioning and sustainability in the future, we characterized the response of a variety of plant, microbial, and sediment responses to a seven-level gradient of N addition in three Californian salt marshes after 7 and 14 months of N addition. The marshes showed variable responses to the experimental N gradient that can be grouped as neutral (root biomass, sediment respiration, potential carbon mineralization, and potential net nitrification), linear (increasing methane flux, decreasing potential net N mineralization, and increasing sediment inorganic N), and nonlinear (saturating aboveground plant biomass and leaf N content, and exponentially increasing sediment inorganic and organic N). The three salt marshes showed quantitative differences in most ecosystem properties and processes rates; however, the form of the response curves to N addition were generally consistent across the three marshes, indicating that the responses observed may be applicable to other marshes in the region. Only for sediment properties (inorganic and organic N pool) did the shape of the response differ significantly between marshes. Overall, the study suggests salt marshes are limited in their ability to sequester C and N with future increases in N, even without further losses in marsh area.
A leaf-height-seed (LHS) plant ecology strategy scheme
DOI:10.1023/A:1004327224729 URL [本文引用: 1]
The worldwide leaf economics spectrum
DOI:10.1038/nature02403 URL [本文引用: 1]
Effects of N, P addition on above/below ground biomass allocation in a subalpine meadow
氮、磷添加对亚高山草甸地上/地下生物量分配的影响
Plant community responses to nitrogen addition and increased precipitation: the importance of water availability and species traits
DOI:10.1111/j.1365-2486.2011.02423.x URL [本文引用: 1]
Shift in community functional composition following nitrogen fertilization in an alpine meadow through intraspecific trait variation and community composition change
DOI:10.1007/s11104-018-3771-x URL [本文引用: 1]
Different categories of biodiversity explain productivity variation after fertilization in a Tibetan alpine meadow community
DOI:10.1002/ece3.2723
PMID:28515882
[本文引用: 1]
The relationship between productivity and biodiversity has long been an important issue in ecological research. However, in recent decades, most ecologists have primarily focused on species diversity while paying little attention to functional diversity and phylogenetic diversity (PD), especially in alpine meadow communities following fertilization. In this study, a fertilization experiment involving the addition of nitrogen, phosphorus, and a mixture of both was implemented in an alpine meadow on the Tibetan Plateau. Species diversity, functional diversity, and PD were measured, and the responses of these parameters to the variation in productivity were analyzed. We found that the productivity of alpine plant communities was colimited by N and P, with N being the principal and P being the secondary limiting nutrient. Our results supported the prediction of both the mass ratio hypothesis and niche complementarity hypothesis in fertilized communities, but these hypotheses were not mutually exclusive. The combination of different aspects of biodiversity not only provides a crucial tool to explain the variation in productivity and to understand the underlying mechanisms but also plays an important role in predicting the variation in productivity of alpine meadow communities, which are sensitive to nutrient enrichment in the context of global change.
The effects of fertilization on the trait-Abundance relationships in a Tibetan alpine meadow community
DOI:10.1093/jpe/rtv043 URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}