


探索多响应变量与多解释变量之间关系的约束性排序类方法(或称典范分析, canonical analysis)已被广泛用作生态学数据分析的基本框架, 具体包括冗余分析(RDA)、典范对应分析(CCA)和基于距离的冗余分析(db-RDA)等(Legendre & Legendre, 2012)。在群落生态学研究中, 典范分析的主要目标之一是定量给定环境因子(解释变量)对目标生物群落组成(响应变量)的解释程度(即R2), 以揭示物种组成(多样性)的环境响应机制。但除了模型的总体解释率之外, 解释变量的相对重要性也是研究人员普遍感兴趣的问题(Peres-Neto et al., 2006)。解释变量的相对重要性的一种常用判断依据是每个变量对R2的贡献比例(Johnson & LeBreton, 2004)。当所有解释变量彼此完全相互独立(即相关系数为0), 则不同解释因子的单独R2之和就等于总R2, 这种情况被称为R2可加性(Genizi, 1993)。但这种情况在真实场景下很难出现, 由于生态学数据多来自野外观测, 不同指标间的相互关系错综复杂, 存在共线性是普遍现象(Graham, 2003)。以土壤理化数据为例, 土壤有机质含量、总氮含量、pH等指标通常具有相关性, 如将这些变量全部纳入模型中, 就会出现解释变量的共线性。共线性对解释率的影响主要体现在不同因子的解释变差会存在共同解释的部分, 统计上被称为共享R2 (Legendre & Legendre, 2012)。共享R2的比例越大, 说明共线性越强。由于共享R2的存在, 导致不同环境因子的简单R2 (即单个解释变量作为唯一解释变量时模型的R2)加和通常会大于模型的总体R2, 因此, 有必要对共享R2进行分割, 以明确不同解释变量对响应变量的独立贡献。
分割共享R2的难点在于很难从数学上确定共享R2由相关变量中的哪个引起, 因此就很难将共享的R2准确分配给不同的变量。面对这一问题, 生态学家通常采用模型筛选、变量顺序R2与变差分解3种方法进行处理。其中, 模型筛选主要通过减少模型中的共线性变量来降低模型的共线性(Burnhan & Anderson, 2004; Blanchet et al., 2008)。这一方法大多基于一定的准则(如p值、赤池信息准则(AIC)值)得到一个所谓的最优模型, 但这种仅通过统计学手段得到的最优变量集合可能不具生态学意义, 且仍可能存在共线性的问题。另一种替代方案是人为筛选, 依据主成分分析(PCA)、方差膨胀因子(VIF)等方法评估解释变量间的相关关系或共线性强弱, 并综合考虑变量的身份以及其生物学意义, 最终人为做出决策。但这种筛选变量的过程无疑带有主观判断, 很难让人信服。变量顺序R2的方法是将解释变量按一定的顺序加入模型中, 并观察R2的增加情况来判断不同变量的贡献(图1A)。通常情况下, 按照p值由小到大或增加的R2由大到小的顺序进行模型添加, 加入变量前后R2的差值就被称为随机R2, 或条件效应(conditional effect)(Šmilauer & Lepš, 2014)。CANOCO软件与R程序包“adespital”的forward.sel函数均采用了这种方法, 在过去的研究中被广泛使用。这种方法的问题在于解释变量所分配的顺序R2显然取决于变量进入模型的顺序。如果解释变量存在共线性, 共享R2更容易分配给先加入模型的解释变量, 而后加入的解释变量所分配的共享R2变少, 导致先加入模型的解释变量占更大的优势。除非分析者具有极强的先验知识, 这种方法实际上存在一定问题。第三种方法是变差分解(variation partitioning, VP), 也常称作方差分解, 最早出现于20世纪60年代用于多元回归的共性分析(commonality analysis)(Newton & Spurrell, 1967; Mood, 1969), 20世纪90年代被引入至典范分析中(Borcard et al., 1992; Peres-Neto et al., 2006)。VP可以将模型的R2分解为由每个变量(或变量组)所能独自解释的组分(称为独自效应(unique effect), 或边际效应(marginal effect)以及共同解释的组分(称为共同效应(common effect))来理解解释变量之间复杂的关系(图1B)。然而随着纳入的变量数目的增加将导致计算量呈指数型增长, 因此VP分组数量经常受到一定限制, 例如“vegan”包的varpart函数最多只能纳入4组变量。此外, 即使不对分组进行限制, 也依然无法解决共同效应的分割问题。
图1
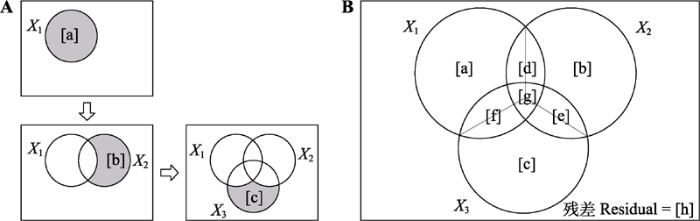
图1
以3组解释变量X1、X2和X3为例, 使用韦恩图概括了响应变量Y的总变差组成。A, 基于变量的顺序R2的方法, 根据分配的顺序, [a]、[b]和[c]分别为X1、X2和X3的条件效应。B, 基于变差分解和层次分割法, [a]、[b]和[c]分别为X1、X2和X3的边际效应, [d]、[e]和[f]分别为X1、X2和X3两两之间的共同效应, [g]为三者之间的共同效应, X1、X2和X3的单独效应分别为[a] + [d]/2 + [f]/2 + [g]/3、[b] + [d]/2 + [e]/2 + [g]/3以及[c] + [e]/2 + [f]/2 + [g]/3。残差[h]代表了未被X1、X2和X3解释的Y的变差部分。
Fig. 1
Venn diagram representing the variation composition of a response matrix Y regressed against three correlated predictors of X1, X2, and X3. A, According to the order of assignment, [a], [b] and [c] are the conditional effect of X1, X2 and X3, respectively. B, Based on variation partitioning and hierarchical partitioning, [a], [b] and [c] are the marginal effect of X1, X2 and X3, respectively; [d], [e] and [f] are the common effect of X1, X2 and X3, respectively; [g] is the common effect among X1, X2 and X3; the individual effect of X1, X2 and X3 can be expressed as [a] + [d]/2 + [f]/2 + [g]/3, [b] + [d]/2 + [e]/2 + [g]/3 and [c] + [e]/2 + [f]/2 + [g]/3. Residual [h] represents the fraction of Y that is not explained by X1, X2 or X3.
事实上, “共线性解释变量相对重要性的比较”在多元回归领域一直是活跃的主题。20世纪80年代, 由Lindeman等(1980)、Cox Jr (1985)和Kruskal (1987)分别独立地提出了一种“平均排序(averaging over orderings)” (也称LMG指数)的方法, 基于所有模型子集获取解释变量的相对重要性。与Chevan和Sutherland (1991)提出的层次分割(hierarchical partitioning, HP)结果一样, 并推广至广义线性模型(GLM)。R程序包“relaimpo”可用于计算多元回归中解释变量的LMG值, 并提供自举法检验LMG值的显著性(Grömping, 2007)。“hier.part”包基于HP来估计GLM中解释变量的相对重要性(Mac Nally & Walsh, 2004)。最近, Lai等(2022a)将VP和HP建立数学联系, 首次将HP算法引入典范分析并开发了“rdacca.hp”包来获取单个典范分析解释变量的解释率, 同时实现了不限解释变量数量限制的VP。HP强调基于所有可能的模型子集下各解释变量的单独效应(individual effect), 等同于解释变量的边际效应加上与其他解释变量的共同效应的平均分配值(图1B), 为典范分析中评估共线性的解释变量的相对重要性提供了新型定量指标。
为进一步推广该方法在生态学领域的应用, 帮助科学工作者在复杂环境背景下寻找关键因子, 本文旨在: (1)介绍“rdacca.hp”包的主要功能; (2)通过案例分析详解“rdacca.hp”包的使用; (3)结合“rdacca.hp”包自发布以来的相关进展对该程序包在生态学及更多学科领域中的应用和升级进行展望。
1 “rdacca.hp”包的结构
截至目前, “rdacca.hp”包的最新版本为1.0-8 (发布时间是2022年4月5日)。用户可以在R语言中输入“install.packages("rdacca.hp")”直接从CRAN安装“rdacca.hp”包, 或者使用“devtools::install_github ("laijiangshan/rdacca.hp")”在GitHub获取。该程序包封装的函数及核心功能见表1, 各函数具体参数和输出的解释可进一步参阅R文档。
表1 “rdacca.hp”包的函数介绍
Table 1
| 函数 Function | 介绍 Description |
|---|---|
| rdacca.hp | 运行不限制解释变量数量的变差分解和层次分割, 计算每组解释变量对典范分析中解释变异(R2或校正R2)的边际、共同和单独效应Implements both variation partitioning and hierarchical partitioning in canonical analysis without limiting in the number of predictors/matrices of predictors. Output include the marginal, common, as well as individual effect of each (group) variables to total R2 or adjusted R2 |
| permu.hp | 对解释变量的单独效应进行置换检验以评估其显著性 Implements the significance testing for individual contribution from hierarchical partitioning by permutation routine |
| plot.rdaccahp | 基于“rdacca.hp”函数的输出, 绘制柱形图展示解释变量单独效应 Plot a bar plot of the individual contribution of predictors based on the output of rdacca.hp() |
2 “rdacca.hp”包的操作案例——塑造甲螨(Oribatida)群落的重要环境和空间驱动因素分析
> data(mite, package="vegan")
> data(mite.env, package="vegan")
> data(mite.pcnm, package="vegan")
表2 R语言“vegan”包的甲螨多度及环境和空间因子数据集
Table 2
| 数据框 Data frame | 介绍 Description |
|---|---|
| mite | 物种多度矩阵, 包含70个采样点的共计35种甲螨形态种的个体计数 Species abundance matrix that contains the abundance of 35 oribatid mites morphospecies at 70 sampling sites |
| mite.env | 环境因子矩阵, 包含70个采样点的共计5种环境变量: 基质密度、基质含水量、基质类型、灌丛密度和微地形特征 Environmental matrix that contains 5 environmental variables at 70 sampling sites, including substrate density, water content of the substrate, substrate type, shrub density, and microtopography |
| mite.pcnm | 空间因子矩阵, 基于70个采样点的地理坐标, 通过邻体矩阵主坐标分析(PCNM)构建的特征向量, 反映了70个采样点的潜在空间结构 Spatial matrix, which is based on the geographic coordinates of 70 sampling sites and constructed by principal coordinates of neighbour matrices (PCNM) (Borcard & Legendre, 2002), represents the potential spatial structure of 70 sampling points |
2.1 单个解释变量的相对重要性分析
我们首先通过典范分析确定环境异质性对甲螨群落组成的影响。考虑到物种多度矩阵中存在较多0值, 预先进行Hellinger转化以消除极端值的影响并减少高丰度物种的权重(Legendre & Gallagher, 2001)。随后, 基于RDA的典范分析用于拟合物种多度(作为响应变量)与所有5个环境因子(作为解释变量)的多变量回归, 并通过VP和HP为各环境因子分配相应的重要性指标。未对环境因子进行共线性评估或者其他任何方式的筛选, 目的是确保获取所有可能的模型子集, 同时比较了VP和HP方法的差异。
> library(rdacca.hp)
> mite.hel <- decostand(mite, method="hellinger")
> mod <- rdacca.hp(mite.hel, mite.env, method ="RDA", var.part=TRUE, type="adjR2")
> UpSetVP::upset_vp(mod, plot.hp=TRUE, order.part="degree")
> permu.hp(mite.hel, mite.env, method="RDA", type="adjR2", permutations=999)
RDA显示5个环境因子共解释了物种多度总变差的43.7% (校正R2 = 0.437) (图2)。“rdacca.hp”包成功实现了大于4组解释变量的VP, 然而随着解释变量数量的增加, 从中解读所有边际效应和共同效应来理解变量间的复杂关系非常具有挑战性。相比之下, HP通过继续分解并组合这些组分来分配单独效应, 提供了更便捷的模型解读框架: 5个环境因子的相对重要性排序依次为基质含水量(16.42%)、灌丛密度(10.18%)、微地形特征(8.43%)、基质类型(5.43%)和基质密度(3.2%)。所有环境因子的单独效应均显著, 但必须注意的是, 由于解释变量的相对重要性通常用作探索性框架而不是推理工具, 置换检验的结果仅用作参考(Lai et al., 2022a)。尽管过去流行使用VP的边际效应来估计解释变量的相对重要性, 但片面的信息容易产生误报。例如, 仅比较边际效应时, 灌丛密度(2.88%)则是对甲螨群落组成贡献最小的环境因子。两种方法产生分歧的一个主要原因在于灌丛密度和基质含水量以及微地形特征等其他环境因子间存在较大的共同效应(归因于共线性), 后者忽略这些共同解释的部分导致灌丛密度的贡献被严重低估。此外, 出于解释变量之间的抑制效应或校正R²等原因, 部分共同效应出现负值。观察到基质密度的边际效应(3.63%) >单独效应(3.2%), 意味着仅关注边际效应也会使个别解释变量的重要性被高估。综上, HP在揭示变量的混杂关系中具有优势。
图2
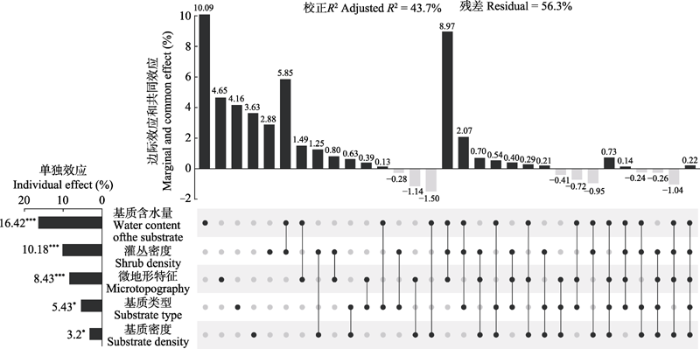
图2
5种环境因子对甲螨物种多度影响的相对重要性。变差分解和层次分割分析结果使用UpSet图来呈现。右侧点阵图中, 每行对应一个环境因子。对于每一列, 孤立黑点表示各环境因子的边际效应, 多点间连线表示这些环境因子间的共同效应, 各组分解释的变差百分比(来自变差分解)展示在上方柱形图中。左侧柱形图为各环境因子的单独效应(来自层次分割), 其值等同于该环境因子的边际效应加上与其他环境因子的共同效应的平均分配值。*, p < 0.05; ***, p < 0.001。
Fig. 2
UpSet matrix layout of variation partitioning and hierarchical partitioning results to show the relative importance of five environmental factors on oribatid mites abundance. In the dot-matrix plot on the right, each row corresponds to an environmental factor. For each column, the isolated black dot represents the marginal effect of each environmental factor, lines connecting multiple dots represent the common effect among these corresponding environmental factors, and the percentage of variation explained by each component (from variation partitioning) are shown in the top column diagram. Column diagram on the left shows individual effect of each environmental factor (from hierarchical partitioning), its value is equal to its marginal effect plus its average shared common effect with other environmental factors. *, p < 0.05; ***, p < 0.001.
2.2 解释变量组的相对重要性分析
“rdacca.hp”包同样提供了用于解释变量组的VP和HP。我们继续将空间因子纳入典范分析, 使用RDA的VP和HP量化空间结构和环境异质性对甲螨群落组成的贡献。
> iv <- list(PCNM=mite.pcnm, ENV=mite.env)
> mod <- rdacca.hp(mite.hel, iv, method="RDA", var.part=TRUE, type="adjR2")
> UpSetVP::upset_vp(mod, plot.hp=TRUE, order.part ="degree")
> permu.hp(mite.hel, iv, method="RDA", type ="adjR2", permutations=999)
结果显示, 空间因子(单独效应27.39%, p < 0.001)和环境因子(单独效应26.41%, p < 0.001)均显著影响甲螨群落组成, 二者共同解释了物种多度总变差的53.8% (校正R2 = 0.538) (图3)。由于相差不足1%, 暗示扩散限制和环境选择压力在塑造当地甲螨群落中的重要性近似相等。值得注意的是两组解释变量存在较高比例的共同效应(34.53%), 可能意味着这些环境因子具有空间结构性, 促使群落形成类似的空间格局, 或者其他空间结构过程(如历史事件或生物相互作用)引发的环境和群落的一致响应。
图3
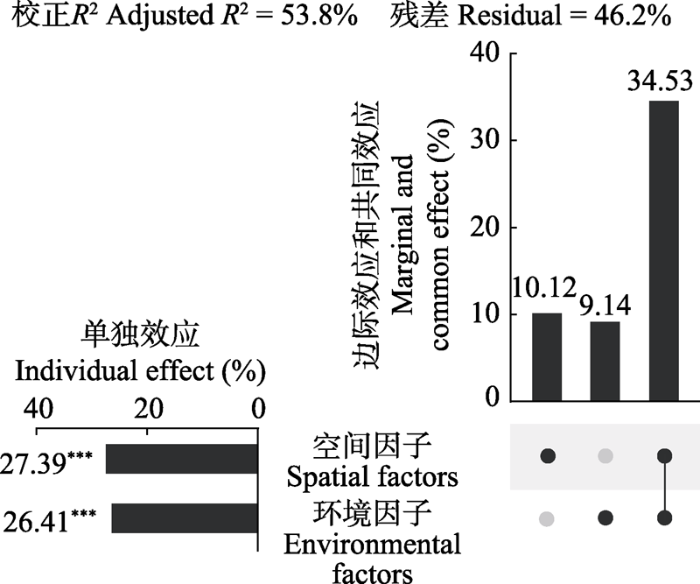
图3
空间和环境因子对甲螨物种多度影响的相对重要性。变差分解和层次分割分析结果使用UpSet图来呈现。右侧点阵图中, 3列分别表示空间因子和环境因子的边际效应以及二者的共同效应(来自变差分解)。左侧柱形图表示二者的单独效应(来自层次分割)。***, p < 0.001。
Fig. 3
UpSet matrix layout of variation partitioning and hierarchical partitioning results to show the relative importance of spatial and environmental factors on oribatid mites abundance. In the dot-matrix plot on the right, three columns represent the marginal effect of spatial factors, the marginal effect of environmental factors, and the common effect between them, respectively (from variation partitioning). Column diagram on the left shows individual effect of spatial and environmental factors (from hierarchical partitioning). ***, p < 0.001.
3 “rdacca.hp”包的相关应用实例
最后, 我们汇总了使用“rdacca.hp”包开展分析的相关研究, 以进一步加深用户对该程序包应用场景的认识。以Google学术为数据源检索关键词“rdacca.hp”和“rdaenvpart” (“rdacca.hp”包的前身, 下文统称为“rdacca.hp”)并根据内容进行甄别, 截至2022年8月31日共计93篇正式发表的论文。“rdacca.hp”包自发布于CRAN以来便得到学术界的高度关注, 被引次数呈现逐年递增趋势, 特别是2022年1月该方法学论文(Lai et al., 2022a)在线发表后被更多国内外学者所熟知(图4)。继续梳理相关研究涉及的学科领域、研究区域或环境类型、典范分析的响应变量和解释变量类别的关键词并进行共词分析, 结果同样显示“rdacca.hp”包正在多学科交叉的数据分析中流行(图5)。其中, 环境科学与生态学、生物学和农林科学是“rdacca.hp”包集中出现的领域, 并涵盖陆地、水域的多种自然系统以及耕地、城市等人工系统, 部分面向微观系统或上升到景观尺度。考虑到“rdacca.hp”包的主要目标是提供定量指标量化响应变量和解释变量的关系, 下文依据这些研究中典范分析的对象和目的, 举例讨论该程序包的当前应用。
图4
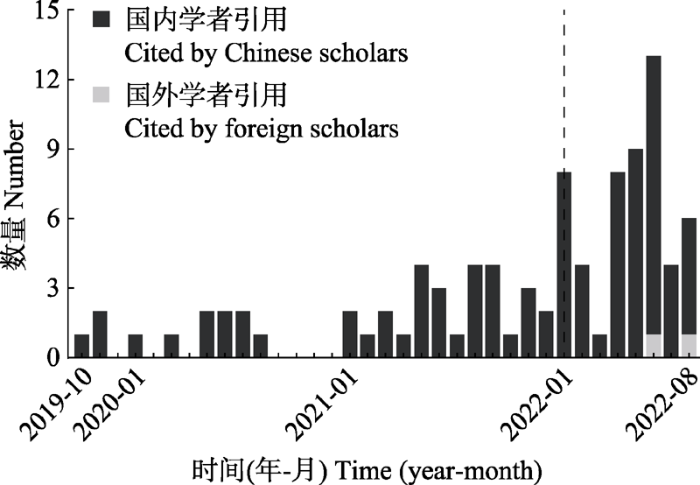
图4
引用“rdacca.hp”包的论文发表时间及数量统计。虚线表示“rdacca.hp”包相关的方法学论文的正式发表时间。
Fig. 4
Statistics of published papers citing “rdacca.hp” package (publication time and number of published paper). Dashed line indicates the publication time of methodological paper related to “rdacca.hp” package.
图5
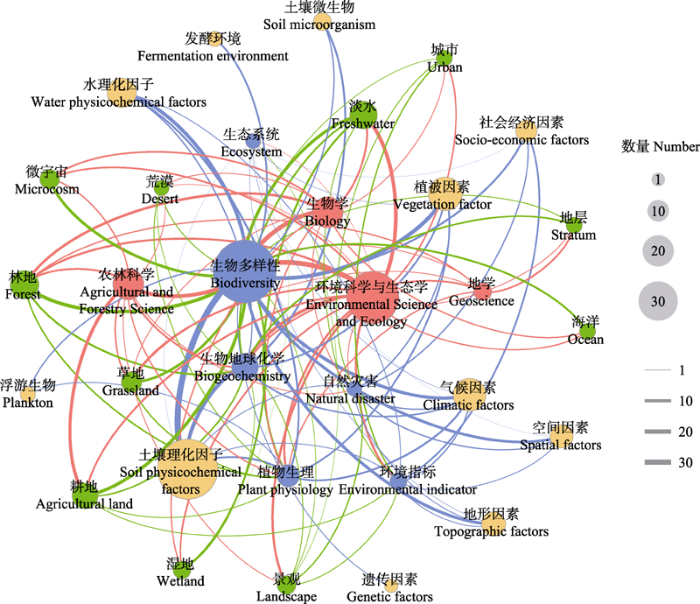
图5
引用“rdacca.hp”包的论文所属的大类学科(红色)、论文中涉及的研究区域或环境类型(绿色)、典范分析的响应变量(蓝色)和解释变量类别(橙色)关键词的共现网络。连线表示关键词的共现关系, 节点大小和连线粗细代表了对应文献的数量。典范分析中解释变量的相对重要性分析均来自“rdacca.hp”包。
Fig. 5
Co-occurrence network of subject (red), study area or environment type (green), response variables (blue) and predictor variables (orange) key words according to published papers citing “rdacca.hp” package. Lines represent the co-occurrence relationship of keywords, node size and line width represent the number of related papers. All of the relative importance of predictors in canonical analysis are from “rdacca.hp” package.
3.1 生物多样性
在所有典范分析的对象中, 生物多样性、群落结构及其与环境、气候、地形或其他生物因子的关系受到最多关注, 陆地生物群落的研究比水生生物群落更广泛(图5, 图6)。揭示生物多样性和群落构建的驱动因素是生态学研究中的核心问题之一, 特别是在当前全球变化背景下, 对于了解生态群落和相关生态系统功能的变化至关重要(Kreft & Jetz, 2007; Hooper et al., 2012; Guo et al., 2018)。保护生态学更注重对相对重要性的分析, 因为明确不同生态因子的重要性对于制定保护策略具有重要价值。尽管不同生物类群对资源需求和环境适应存在异质性响应, 但多项研究的HP结果呈现出一些共性。例如, 地形或气候条件通常比土壤理化因子更主导森林和草原植被特征的变化(Liu et al., 2022b; Xie et al., 2022; Zhao et al., 2022)。淡水湖泊浮游植物群落受养分梯度的强烈影响, 氮是关键的驱动因素(Hu et al., 2021; Zhang et al., 2023)。这些均暗示需要采取适应措施以应对未来气候和营养负荷变化可能带来的不利影响。
图6
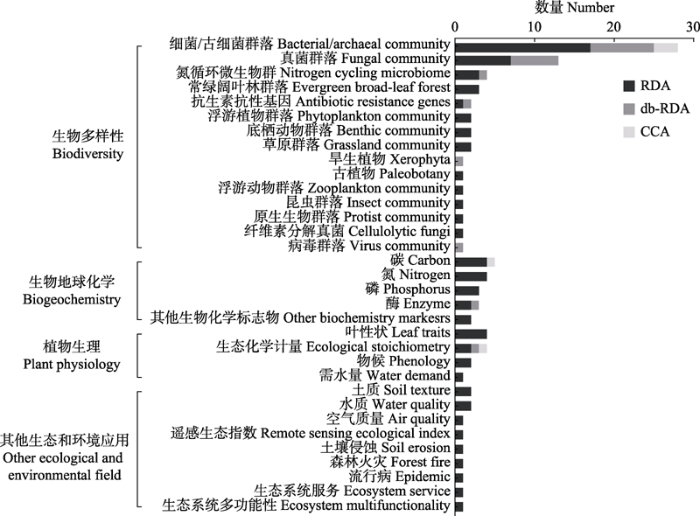
图6
引用“rdacca.hp”包的论文中所涉及典范分析的响应变量类型及数量统计。CCA, 典范对应分析; db-RDA, 基于距离的冗余分析; RDA, 冗余分析。
Fig. 6
Statistics of published papers citing “rdacca.hp” package (type and number of response variables). CCA, canonical correspondence analysis; db-RDA, distance-based redundancy analysis; RDA, redundancy analysis.
虽然典范分析方法最初是为解决植物群落问题而开发, 但“rdacca.hp”包更多被应用于微生物群落分析(图6)。事实上, 很多植物生态学研究方法后来均在微生物生态学中得到验证, 这得益于近年来分子测序技术的发展推动微生物生态学研究成为热点。与天然群落相比, 与农林牧有关的土壤微生物更受青睐。在包括农田、人工林、牧场等在内的人工生态系统中, β多样性和HP分析强调pH相较其他理化属性对土壤细菌群落的影响更直接(Mei et al., 2021; Xiao et al., 2021; Xu et al., 2021; Li et al., 2022), 而土壤真菌群落对有机碳组分的响应更敏感(Xu et al., 2021; Wang et al., 2022a, 2022d)。理解土地利用或管理制度如何影响土壤性质以及微生物功能, 有助于指导有益的农林牧实践并提升生态效益。同时, 植物-微生物的相互作用也被广泛关注。以根系相关微生物最具代表性, 植物种类和生理条件的重要性通常大于非生物因子(Wang et al., 2022b, 2022c, 2022d)。地上植被与地下微生物群落构成维持生态功能的系统实体, 有益的生物协同作用促进养分转化并提升生产力, 为可持续生态提供了更多机会。此外, 在环境修复、发酵生产等工程领域也偶见使用“rdacca.hp”包的方法分析微生物群落构建的驱动因素, 由于相关文献数量尚少暂不讨论。
3.2 生物地球化学
作为生物与环境综合作用的结果, 生物地球化学过程塑造地质和生物历史。除了群落生态学应用, 典范分析也常用做环境中碳氮磷化学或生物产物及其驱动因素的解释框架(图5, 图6)。生物多样性的变化是否会导致生态系统功能和相关的生物地球化学过程的改变同样是当代生态学中的关键问题(Isbell et al., 2018; van der Plas, 2019)。尽管当前案例尚少, 但仍可根据HP得知林分结构显著影响土壤微生物生物量、物种组成、酶活性以及化学计量(Xu et al., 2021; Yan et al., 2022a), 改善森林结构和植物多样性有助于减少土壤养分限制和多功能性的维持。类似地, 河湖交错带的植被恢复通过重新分配有机质、氮和生物有效磷组分, 缓解水体富营养化增强流域系统的生态功能(Yan et al., 2022b)。仅考虑非生物因子时, 有机质组分及氮库变动是影响土壤磷有效性、氮矿化速率以及温室气体排放的重要环境参数(Chen et al., 2021; Kong et al., 2022; Liu et al., 2022a)。由于不可能将这些地球化学变化单独归因于物理化学条件, 微生物的活动无疑是联系这些地球化学变化的潜在桥梁。此外, 基于HP的分析还展现了特定生物化学标志物在气候或环境建模中的应用价值, 例如甘油二烷基甘油四醚表现出与温度、降水和土壤pH的强相关关系(Duan et al., 2022a, 2022b), 对于更好地理解生物地球化学循环和气候反馈具有重要意义。
3.3 植物生理
从群落、种群至个体生态学层面, 以植物为代表的生理学或表型相关研究也是一大侧重点(图5, 图6)。这些典范分析案例中的一个主要部分是了解植物性状变异与气候、环境因子等的相关性。其中, 植物叶片作为光合作用的主要器官, 在生态系统中起着决定植物生存和生产力的关键作用, 其形态结构和营养状态成为研究重点。陆生和水生植物与环境因子的HP均强调了环境过滤在塑造叶片性状方面的重要性(Gao et al., 2021; Ren et al., 2021)。考虑到不同植物类群的叶片性状具有独特的进化和生态价值, 叶片性状和环境的关系对于解释和预测叶片营养模式以及识别生态系统营养限制非常重要。淡水的不同理化因子在调节浮游植物、潜水植物以及沉水植物的生态化学计量中的贡献差异巨大(Rao et al., 2021; Zhang et al., 2021, 2023), 充分体现出3种水生植物的生态位高度分化。另一主要部分是物候学, 量化植物物候及其与气候的相互作用对于应对气候变化和物候变化至关重要(Cleland et al., 2007; Richardson et al., 2013)。HP结果显示, 气候是控制植物形态、物候和生理学变化的主要非生物因子(Zhai et al., 2021; Cheng et al., 2022)。利用功能性状评估沙漠植物开花物候的变异性, 为理解气候变化与植物繁殖策略之间的关系提供了新的视角(Wang et al., 2020)。性状特征还可用于诠释生物多样性。在一个中营养亚热带高原湖泊中, 通过将形态学鉴定和分子生物学的遗传标记分型相结合来揭示浮游植物生态策略, 为制定水华防控措施提供了依据(Hu et al., 2021)。
3.4 其他生态和环境应用
典范分析的其他方面应用还包括对环境指标、自然灾害或生态系统的建模(图5, 图6)。类似于生物地球化学的分析, 环境指标的响应变量亦是一组物理化学参数, 因受人类活动影响强烈而常用做评价环境质量的指标, 如水透明度、空气PM2.5指数等(Tao et al., 2021; Gao et al., 2022; Ke et al., 2022)。了解这些指标变动的原因有助于制定相应的污染控制措施来降低环境风险。面向自然灾害或生态系统的研究大多上升到景观尺度, 综合调查了区域的自然或人文属性。典范分析对象包括黄土高原的细沟侵蚀、西南森林的火灾时空格局、三峡流域的生态系统服务、黄河流域的土地利用及其与当地的地理地貌、气候、区域环境、生物特征或社会经济驱动因素的联系等(Xiong et al., 2020; Yang et al., 2020; Gou et al., 2021; Cui et al., 2022)。由于因素复杂, 在景观尺度上整合多种生态系统服务仍存在一定挑战。随着学科发展, 景观配置控制的生态系统服务对景观多样性的影响和反馈可能是未来扩大研究的领域。
4 总结和展望
“rdacca.hp”包的HP算法在评估典范分析中共线性的解释变量的相对重要性中表现出优越性, 目前正广泛用于生态学、环境科学和其他相关学科问题的数据分析。本文可进一步加深国内学者对这一方法及其应用的认识。尽管“rdacca.hp”包对输入的解释变量的数量并无限制, 但进一步纳入置换检验评估HP中单独效应的显著性时, 解释变量数量和置换次数的增加使计算过程仍然复杂且耗时(Lai et al., 2022a)。因此, 未来有待继续优化算法或开发多线程模式以提高效率。此外, 截至当前应用“rdacca.hp”包进行数据分析的公开发表的研究中绝大多数为国人的成果, 国外暂且仅两篇引用(图4), 未来也有待进一步推广至国际应用。此外, 考虑到“rdacca.hp”包无法处理混合效应模型的相对重要性信息提取, 基于同一思路的“glmm.hp”包已经发布至CRAN (
参考文献
Forward selection of explanatory variables
DOI:10.1890/07-0986.1
PMID:18831183
[本文引用: 1]

This paper proposes a new way of using forward selection of explanatory variables in regression or canonical redundancy analysis. The classical forward selection method presents two problems: a highly inflated Type I error and an overestimation of the amount of explained variance. Correcting these problems will greatly improve the performance of this very useful method in ecological modeling. To prevent the first problem, we propose a two-step procedure. First, a global test using all explanatory variables is carried out. If, and only if, the global test is significant, one can proceed with forward selection. To prevent overestimation of the explained variance, the forward selection has to be carried out with two stopping criteria: (1) the usual alpha significance level and (2) the adjusted coefficient of multiple determination (Ra(2)) calculated using all explanatory variables. When forward selection identifies a variable that brings one or the other criterion over the fixed threshold, that variable is rejected, and the procedure is stopped. This improved method is validated by simulations involving univariate and multivariate response data. An ecological example is presented using data from the Bryce Canyon National Park, Utah, U.S.A.
All-scale spatial analysis of ecological data by means of principal coordinates of neighbour matrices
DOI:10.1016/S0304-3800(01)00501-4 URL [本文引用: 1]
Partialling out the spatial component of ecological variation
DOI:10.2307/1940179 URL [本文引用: 2]
Differential response of soil CO2, CH4, and N2O emissions to edaphic properties and microbial attributes following afforestation in central China
DOI:10.1111/gcb.v27.21 URL [本文引用: 1]
Precipitation dominates the relative contributions of climate factors to grasslands spring phenology on the Tibetan Plateau
DOI:10.3390/rs14030517 [本文引用: 1]
Hierarchical partitioning
Shifting plant phenology in response to global change
DOI:10.1016/j.tree.2007.04.003 URL [本文引用: 1]
A new measure of attributable risk for public health applications
DOI:10.1287/mnsc.31.7.800
URL
[本文引用: 1]
A persistent conceptual problem in the theory of risk accounting and risk analysis methodology is that of allocating or attributing a risk among the joint causes or “factors” producing it. When multiple factors interact, the especially difficult problem of “risk externalities” arises. Drawing on related work in the mathematical theory of games that seeks solutions to similar philosophical and conceptual problems, this paper proposes a method for solving the risk attribution problem. The proposed solution is justified on the grounds that it is the unique risk attribution function satisfying three reasonable principles, or “axioms,” of risk accounting. A mathematical framework for formulating risk attribution problems, and several possible applications, are also discussed.
Vegetation restoration restricts rill development on dump slopes in coalfields
DOI:10.1016/j.scitotenv.2022.153203 [本文引用: 1]
The impact of precipitation on the distributions of branched tetraethers in alkaline soils
DOI:10.1016/j.orggeochem.2022.104410 [本文引用: 1]
General Holocene warming trend in arid Central Asia indicated by soil isoprenoid tetraethers
DOI:10.1016/j.gloplacha.2022.103879 [本文引用: 1]
UV/sodium percarbonate for bisphenol A treatment in water: impact of water quality parameters on the formation of reactive radicals
DOI:10.1016/j.watres.2022.118457 [本文引用: 1]
Rapid adaptive responses of rosette-type macrophyte Vallisneria natans juveniles to varying water depths: the role of leaf trait plasticity
DOI:10.1002/ece3.v11.20 URL [本文引用: 1]
Decomposition of R2 in multiple regression with correlated regressors
Identifying and analyzing ecosystem service bundles and their socioecological drivers in the Three Gorges Reservoir Area
DOI:10.1016/j.jclepro.2021.127208 [本文引用: 1]
Confronting multicollinearity in ecological multiple regression
DOI:10.1890/02-3114 URL [本文引用: 1]
Estimators of relative importance in linear regression based on variance decomposition
DOI:10.1198/000313007X188252 URL [本文引用: 1]
Climate warming leads to divergent succession of grassland microbial communities
DOI:10.1038/s41558-018-0254-2 [本文引用: 1]
A global synthesis reveals biodiversity loss as a major driver of ecosystem change
DOI:10.1038/nature11118 [本文引用: 1]
Environmental factors associated with cyanobacterial assemblages in a mesotrophic subtropical plateau lake: a focus on bloom toxicity
DOI:10.1016/j.scitotenv.2021.146052 [本文引用: 2]
Quantifying effects of biodiversity on ecosystem functioning across times and places
DOI:10.1111/ele.12928
PMID:29493062
[本文引用: 1]
Biodiversity loss decreases ecosystem functioning at the local scales at which species interact, but it remains unclear how biodiversity loss affects ecosystem functioning at the larger scales of space and time that are most relevant to biodiversity conservation and policy. Theory predicts that additional insurance effects of biodiversity on ecosystem functioning could emerge across time and space if species respond asynchronously to environmental variation and if species become increasingly dominant when and where they are most productive. Even if only a few dominant species maintain ecosystem functioning within a particular time and place, ecosystem functioning may be enhanced by many different species across many times and places (β-diversity). Here, we develop and apply a new approach to estimate these previously unquantified insurance effects of biodiversity on ecosystem functioning that arise due to species turnover across times and places. In a long-term (18-year) grassland plant diversity experiment, we find that total insurance effects are positive in sign and substantial in magnitude, amounting to 19% of the net biodiversity effect, mostly due to temporal insurance effects. Species loss can therefore reduce ecosystem functioning both locally and by eliminating species that would otherwise enhance ecosystem functioning across temporally fluctuating and spatially heterogeneous environments.© 2018 John Wiley & Sons Ltd/CNRS.
History and use of relative importance indices in organizational research
DOI:10.1177/1094428104266510
URL
[本文引用: 1]
The search for a meaningful index of the relative importance of predictors in multiple regression has been going on for years. This type of index is often desired when the explanatory aspects of regression analysis are of interest. The authors define relative importance as the proportionate contribution each predictor makes to R2, considering both the unique contribution of each predictor by itself and its incremental contribution when combined with the other predictors. The purposes of this article are to introduce the concept of relative importance to an audience of researchers in organizational behavior and industrial/organizational psychology and to update previous reviews of relative importance indices. To this end, the authors briefly review the history of research on predictor importance in multiple regression and evaluate alternative measures of relative importance. Dominance analysis and relative weights appear to be the most successful measures of relative importance currently available. The authors conclude by discussing how importance indices can be used in organizational research.
Three-dimensional building morphology impacts on PM2.5 distribution in urban landscape settings in Zhejiang, China
DOI:10.1016/j.scitotenv.2022.154094 [本文引用: 1]
Effects of vegetation presence on soil net N mineralization are independent of landscape position and vegetation type in an eroding watershed
DOI:10.1016/j.agee.2021.107743 [本文引用: 1]
Global patterns and determinants of vascular plant diversity
DOI:10.1073/pnas.0608361104
PMID:17379667
[本文引用: 1]
Plants, with an estimated 300,000 species, provide crucial primary production and ecosystem structure. To date, our quantitative understanding of diversity gradients of megadiverse clades such as plants has been hampered by the paucity of distribution data. Here, we investigate the global-scale species-richness pattern of vascular plants and examine its environmental and potential historical determinants. Across 1,032 geographic regions worldwide, potential evapotranspiration, the number of wet days per year, and measurements of topographical and habitat heterogeneity emerge as core predictors of species richness. After accounting for environmental effects, the residual differences across the major floristic kingdoms are minor, with the exception of the uniquely diverse Cape Region, highlighting the important role of historical contingencies. Notably, the South African Cape region contains more than twice as many species as expected by the global environmental model, confirming its uniquely evolved flora. A combined multipredictor model explains approximately 70% of the global variation in species richness and fully accounts for the enigmatic latitudinal gradient in species richness. The models illustrate the geographic interplay of different environmental predictors of species richness. Our findings highlight that different hypotheses about the causes of diversity gradients are not mutually exclusive, but likely act synergistically with water-energy dynamics playing a dominant role. The presented geostatistical approach is likely to prove instrumental for identifying richness patterns of the many other taxa without single-species distribution data that still escape our understanding.
Relative importance by averaging over orderings
Generalizing hierarchical and variation partitioning in multiple regression and canonical analyses using the rdacca.hp R package
DOI:10.1111/mee3.v13.4 URL [本文引用: 4]
glmm.hp: an R package for computing individual effect of predictors in generalized linear mixed models
DOI:10.1093/jpe/rtac096
URL
[本文引用: 1]
Generalized linear mixed models (GLMMs) have been widely used in contemporary ecology studies. However, determination of the relative importance of collinear predictors (i.e. fixed effects) to response variables is one of the challenges in GLMMs. Here, we developed a novel R package, glmm.hp, to decompose marginal R2 explained by fixed effects in GLMMs. The algorithm of glmm.hp is based on the recently proposed approach ‘average shared variance’ i.e. used for multivariate analysis. We explained the principle and demonstrated the use of this package by simulated dataset. The output of glmm.hp shows individual marginal R2s that can be used to evaluate the relative importance of predictors, which sums up to the overall marginal R2. Overall, we believe the glmm.hp package will be helpful in the interpretation of GLMM outcomes.
Ecologically meaningful transformations for ordination of species data
DOI:10.1007/s004420100716
PMID:28547606
[本文引用: 1]
This paper examines how to obtain species biplots in unconstrained or constrained ordination without resorting to the Euclidean distance [used in principal-component analysis (PCA) and redundancy analysis (RDA)] or the chi-square distance [preserved in correspondence analysis (CA) and canonical correspondence analysis (CCA)] which are not always appropriate for the analysis of community composition data. To achieve this goal, transformations are proposed for species data tables. They allow ecologists to use ordination methods such as PCA and RDA, which are Euclidean-based, for the analysis of community data, while circumventing the problems associated with the Euclidean distance, and avoiding CA and CCA which present problems of their own in some cases. This allows the use of the original (transformed) species data in RDA carried out to test for relationships with explanatory variables (i.e. environmental variables, or factors of a multifactorial analysis-of-variance model); ecologists can then draw biplots displaying the relationships of the species to the explanatory variables. Another application allows the use of species data in other methods of multivariate data analysis which optimize a least-squares loss function; an example is K-means partitioning.
Liming mitigates the spread of antibiotic resistance genes in an acid black soil
DOI:10.1016/j.scitotenv.2022.152971 [本文引用: 1]
Introduction to Bivariate and Multivariate Analysis
Nitrogen addition promotes soil phosphorus availability in the subalpine forest of eastern Tibetan Plateau
DOI:10.1007/s11368-021-03064-0 [本文引用: 1]
Classification and distribution of evergreen broad-leaved forests in Jiangxi, East China
DOI:10.1093/jpe/rtac059 [本文引用: 1]
Hierarchical partitioning public-domain software
DOI:10.1023/B:BIOC.0000009515.11717.0b URL [本文引用: 1]
Effects of 40 years applications of inorganic and organic fertilization on soil bacterial community in a maize agroecosystem in northeast China
DOI:10.1016/j.eja.2021.126332 [本文引用: 1]
Macro-analysis of the American educational system
DOI:10.1287/opre.17.5.770
URL
[本文引用: 1]
This paper presents a rudimentary model of the public school system as an input-output process. The inputs are students' own abilities and attitudes, parental support, peer support, quality of the school system, community support, and society's posture with respect to education. Outputs are various categories of academic achievement as well as social competence, responsibility, self confidence, creativeness, ethics, and ambition. All these factors must be measured by index numbers or simple indicators. The model is a set of regression equations relating outputs to inputs. Some implementation of the model has been made possible by means of data gathered in the U.S. Office of Education's Equality-of-Educational-Opportunity Survey.
Examples of the use of elements for clarifying regression analyses
Variation partitioning of species data matrices: estimation and comparison of fractions
DOI:10.1890/0012-9658(2006)87[2614:vposdm]2.0.co;2
PMID:17089669
[本文引用: 2]
Establishing relationships between species distributions and environmental characteristics is a major goal in the search for forces driving species distributions. Canonical ordinations such as redundancy analysis and canonical correspondence analysis are invaluable tools for modeling communities through environmental predictors. They provide the means for conducting direct explanatory analysis in which the association among species can be studied according to their common and unique relationships with the environmental variables and other sets of predictors of interest, such as spatial variables. Variation partitioning can then be used to test and determine the likelihood of these sets of predictors in explaining patterns in community structure. Although variation partitioning in canonical analysis is routinely used in ecological analysis, no effort has been reported in the literature to consider appropriate estimators so that comparisons between fractions or, eventually, between different canonical models are meaningful. In this paper, we show that variation partitioning as currently applied in canonical analysis is biased. We present appropriate unbiased estimators. In addition, we outline a statistical test to compare fractions in canonical analysis. The question addressed by the test is whether two fractions of variation are significantly different from each other. Such assessment provides an important step toward attaining an understanding of the factors patterning community structure. The test is shown to have correct Type I. error rates and good power for both redundancy analysis and canonical correspondence analysis.
Stoichiometric and physiological mechanisms that link hub traits of submerged macrophytes with ecosystem structure and functioning
DOI:10.1016/j.watres.2021.117392 [本文引用: 1]
Environmental filtering rather than phylogeny determines plant leaf size in three floristically distinctive plateaus
DOI:10.1016/j.ecolind.2021.108049 [本文引用: 1]
Climate change, phenology, and phenological control of vegetation feedbacks to the climate system
DOI:10.1016/j.agrformet.2012.09.012 URL [本文引用: 1]
Songhua River Basin’s improving water quality since 2005 based on Landsat observation of water clarity
DOI:10.1016/j.envres.2021.111299 [本文引用: 1]
Biodiversity and ecosystem functioning in naturally assembled communities
DOI:10.1111/brv.12499
PMID:30724447
[本文引用: 1]
Approximately 25 years ago, ecologists became increasingly interested in the question of whether ongoing biodiversity loss matters for the functioning of ecosystems. As such, a new ecological subfield on Biodiversity and Ecosystem Functioning (BEF) was born. This subfield was initially dominated by theoretical studies and by experiments in which biodiversity was manipulated, and responses of ecosystem functions such as biomass production, decomposition rates, carbon sequestration, trophic interactions and pollination were assessed. More recently, an increasing number of studies have investigated BEF relationships in non-manipulated ecosystems, but reviews synthesizing our knowledge on the importance of real-world biodiversity are still largely missing. I performed a systematic review in order to assess how biodiversity drives ecosystem functioning in both terrestrial and aquatic, naturally assembled communities, and on how important biodiversity is compared to other factors, including other aspects of community composition and abiotic conditions. The outcomes of 258 published studies, which reported 726 BEF relationships, revealed that in many cases, biodiversity promotes average biomass production and its temporal stability, and pollination success. For decomposition rates and ecosystem multifunctionality, positive effects of biodiversity outnumbered negative effects, but neutral relationships were even more common. Similarly, negative effects of prey biodiversity on pathogen and herbivore damage outnumbered positive effects, but were less common than neutral relationships. Finally, there was no evidence that biodiversity is related to soil carbon storage. Most BEF studies focused on the effects of taxonomic diversity, however, metrics of functional diversity were generally stronger predictors of ecosystem functioning. Furthermore, in most studies, abiotic factors and functional composition (e.g. the presence of a certain functional group) were stronger drivers of ecosystem functioning than biodiversity per se. While experiments suggest that positive biodiversity effects become stronger at larger spatial scales, in naturally assembled communities this idea is too poorly studied to draw general conclusions. In summary, a high biodiversity in naturally assembled communities positively drives various ecosystem functions. At the same time, the strength and direction of these effects vary highly among studies, and factors other than biodiversity can be even more important in driving ecosystem functioning. Thus, to promote those ecosystem functions that underpin human well-being, conservation should not only promote biodiversity per se, but also the abiotic conditions favouring species with suitable trait combinations.© 2019 Cambridge Philosophical Society.
Grazing greatly reduces the temporal stability of soil cellulolytic fungal community in a steppe on the Tibetan Plateau
DOI:10.1016/j.jes.2021.09.023 URL [本文引用: 1]
N-induced root exudates mediate the rhizosphere fungal assembly and affect species coexistence
DOI:10.1016/j.scitotenv.2021.150148 [本文引用: 1]
Different bacterial co-occurrence patterns and community assembly between rhizosphere and bulk soils under N addition in the plant-soil system
Soil bacterial and fungal communities respond differently to Bombax ceiba (Malvaceae) during reproductive stages of rice in a traditional agroforestry system
DOI:10.1007/s11104-022-05542-x [本文引用: 2]
Flowering phenology shifts in response to functional traits, growth form, and phylogeny of woody species in a desert area
DOI:10.3389/fpls.2020.00536 [本文引用: 1]
Variation in abundance, diversity, and composition of nirK and nirS containing denitrifying bacterial communities in a red paddy soil as affected by combined organic-chemical fertilization
DOI:10.1016/j.apsoil.2021.104001 [本文引用: 1]
Mismatch between specific and genetic diversity in an evergreen broadleaf forest in southeast china: a study case of 10.24 ha forest dynamics plot of Huangshan
DOI:10.3389/fpls.2021.706006 [本文引用: 1]
Fire from policy, human interventions, or biophysical factors? Temporal-spatial patterns of forest fire in southwestern China
DOI:10.1016/j.foreco.2020.118381 [本文引用: 1]
Abundant fungal and rare bacterial taxa jointly reveal soil nutrient cycling and multifunctionality in uneven-aged mixed plantations
DOI:10.1016/j.ecolind.2021.107932 [本文引用: 3]
Planted forests intensified soil microbial metabolic nitrogen and phosphorus limitation on the Loess Plateau, China
DOI:10.1016/j.catena.2021.105982 [本文引用: 1]
Response of spatio-temporal changes in sediment phosphorus fractions to vegetation restoration in the degraded river-lake ecotone
DOI:10.1016/j.envpol.2022.119650 [本文引用: 1]
Determinations of environmental factors on interactive soil properties across different land-use types on the Loess Plateau, China
DOI:10.1016/j.scitotenv.2020.140270 [本文引用: 1]
The powdery mildew disease of rubber (Oidium heveae) is jointly controlled by the winter temperature and host phenology
DOI:10.1007/s00484-021-02125-w [本文引用: 1]
Topsoil nutrients drive leaf carbon and nitrogen concentrations of a desert phreatophyte in habitats with different shallow groundwater depths
DOI:10.3390/w13213093 [本文引用: 1]
Total nitrogen and community turnover determine phosphorus use efficiency of phytoplankton along nutrient gradients in plateau lakes
DOI:10.1016/j.jes.2022.02.005 URL [本文引用: 2]
Soil development mediates precipitation control on plant productivity and diversity in alpine grasslands
DOI:10.1016/j.geoderma.2022.115721 [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}