


植物种群空间分布格局反映了种群个体在空间上的分布状态, 对种群间相互关系以及种群与生境之间的关系具有重要的作用(刘振等, 2012)。植物种群分布格局不仅因物种而异, 同一物种在不同生长发育阶段、不同生境条件下也存在明显差异(常伟等, 2016)。一般分为随机、均匀与集群3种类型, 合理的分布格局能够促进森林生态系统功能发挥作用(弓文艳等, 2018)。目前, 多采用样方法、点格局分析法等研究方法进行格局分析。贾炜伟等(2017)通过样方法中的7种聚集度指数得知落叶松(Larix gmelinii)和白桦(Betula platyphylla)更新幼株的聚集强度差异明显, 点格局分析表明急尖长苞冷杉(Abies georgei var. smithii)更新幼株在小尺度上呈聚集分布, 种群聚集强度随树木直径和空间尺度的增加而减小(解传奇等, 2015)。这都表明对植物种群分布格局进行研究有助于了解种群生物学以及生态学特性。
植物种群更新前期是更新个体生命周期中最为脆弱的阶段, 其个体特征与空间分布将影响该种群未来组成、结构和功能(Yang et al., 2015)。新疆野杏(Prunus armeniaca)是野果林重要的组成树种, 对伊犁河谷落叶阔叶林生态系统稳定与当地经济发展具有重要意义。以往有学者对新疆野杏的遗传特性(Li et al., 2020a, 2020b)、表型性状(曹倩等, 2016)、生长发育(尤璐瑶等, 2019), 以及天然更新前期种子的散布(张渊等, 2016)、萌发及影响因素(翟朝阳等, 2019)等方面进行了研究。如今, 受气候变化、病虫危害、过度放牧、旅游开发等多重因素影响, 新疆野杏种群未得到有效保护, 树体受损数量逐年增多, 生长衰退、天然更新障碍等问题突出(方紫妍等, 2019; 刘瑜, 2019; 杨其享等, 2022)。鉴于此, 依据野杏种群在山地多呈片状, 沿沟谷呈带状, 疏林和散生等方式分布的特点, 对分布区域内种群更新幼株的特征和分布状况进行实地调查, 分析其在不同区域、生境(林下、林窗、空地)的变化, 探讨种群更新的空间分布格局与影响因素, 可为保护和人工促进新疆野杏天然更新提供理论依据。
1 材料和方法
1.1 研究区概况
研究区位于伊犁河谷野果林集中分布的霍城县、巩留县、新源县, 地处43.19°-44.43° N, 80.77°-83.43° E之间, 属北温带大陆性半干旱气候, 年平均气温10.4 ℃, 年日照时间2 532-2 722 h, 山区年降水量600 mm, 年积温2 700-3 500 ℃, 无霜期为156天(马闯等, 2018)。该环境下种群间的分布特征存在一定的差异。其中霍城县大西沟、巩留县小莫乎儿沟野杏种群为混交林, 主要分布于中下坡位, 常年受到人类活动干扰, 区别在于大西沟野杏种群为均匀混交, 主沟内次级小沟众多。小莫乎儿沟野杏种群从沟口到沟底由优势种逐渐变为伴生种, 直至沟底为针叶林, 无野杏分布, 沟内野杏主要沿主沟两侧分布。新源县杏花沟野杏种群则为纯林, 海拔跨度较大, 受干扰程度由低海拔到高海拔呈递减趋势。以上3个沟野杏种群的分布状况是其在野果林最为主要的分布方式, 故选择这3个沟为研究区域。研究区内主要伴生树种为新疆野苹果(Malus sieversii)、樱桃李(Prunus cerasifera)、野山楂(Grataegus cuneata)、胡桃(Juglans regia)、欧洲李(Prunus domestica)、稠李(Padus racemosa)等, 灌木有黑果小檗(Berberis atrocarpa)、天山绣线菊(Spiraea tianschanica)、新疆锦鸡儿(Caragana turkestanica)、 新疆忍冬(Lonicera tatarica)等, 草本种类丰富。
1.2 研究方法
1.2.1 样地设置
在研究区内踏查新疆野杏更新幼株的基础上,于各种群的林下设20 m × 20 m (400 m2)样地, 并结合Zhu等(2015)的研究结果, 用林窗平均直径(d)与林窗边缘野杏平均树高(7 m)的比值(d/7)来确定林窗面积(13-600 m2), 设为林窗样地, 超过林窗面积上限(600 m2以上)的设为空地样地。分别在大西沟、杏花沟、小莫乎儿沟野杏林内选择能够反映野杏种群分布特点、林相完整以及林分密度均匀的林分, 先设林下样地, 在距其最近的林窗和空地设立林窗和空地样地, 重复3组, 每个种群设9块样地, 3个种群共27块样地。林下样地编号为A1、A2、A3 (大西沟)、A4、A5、A6 (杏花沟)、A7、A8、A9 (小莫乎儿沟), 林窗为B1、B2、B3 (大西沟)、B4、B5、B6 (杏花沟)、B7、B8、B9 (小莫乎儿沟), 空地为C1、C2、C3 (大西沟)、C4、C5、C6 (杏花沟)、C7、C8、C9 (小莫乎儿沟)。将各样地连续划分为5 m × 5 m方格样方, 林下样地为16个方格样方, 林窗和空地以实际面积所能设的方格样方数量为准(林窗、空地样地为不规则形状)。使用GPS采集林窗、空地边缘木所围成的轨迹, 计算出样地面积, 测定经纬度和海拔; 使用地质罗盘仪测量样地的坡度、坡向, 根据样地所在坡面的海拔范围分为上、中、下坡位。样地基本情况见表1。
表1 新疆野杏种群样地基本情况
Table 1
| 分布区域 Distribution area | 样地 Sample plot | 经度 Longitude (° E) | 纬度 Latitude (° N) | 海拔 Altitude (m) | 坡向 Slope aspect (°) | 坡位 Slope position | 坡度 Slope gradient (°) | 样地面积 Plot area (m2) | 样方(5 m × 5 m)数量 Quadrat (5 m × 5 m) number |
|---|---|---|---|---|---|---|---|---|---|
| DXG | A1 | 80.78 | 44.44 | 1 216.9 | 178.3 | 下 Down | 21.0 | 400.0 | 16 |
| A2 | 80.77 | 44.43 | 1 290.0 | 197.7 | 中 Middle | 35.3 | 400.0 | 16 | |
| A3 | 80.79 | 44.43 | 1 219.2 | 247.3 | 下 Down | 11.7 | 400.0 | 16 | |
| B1 | 80.78 | 44.44 | 1 228.0 | 177.6 | 下 Down | 21.0 | 130.0 | 5 | |
| B2 | 80.77 | 44.43 | 1 323.6 | 203.3 | 中 Middle | 31.7 | 256.2 | 10 | |
| B3 | 80.79 | 44.43 | 1 222.2 | 251.0 | 下 Down | 10.7 | 144.5 | 5 | |
| C1 | 80.78 | 44.44 | 1 242.5 | 180.7 | 中 Middle | 25.7 | 896.2 | 35 | |
| C2 | 80.77 | 44.43 | 1 324.2 | 167.3 | 中 Middle | 25.7 | 1 011.0 | 40 | |
| C3 | 80.79 | 44.43 | 1 240.1 | 225.0 | 中 Middle | 23.7 | 1 058.0 | 42 | |
| XHG | A4 | 83.44 | 43.55 | 1 231.8 | 182.5 | 中 Middle | 23.9 | 400.0 | 16 |
| A5 | 83.43 | 43.55 | 1 340.4 | 136.3 | 中 Middle | 23.9 | 400.0 | 16 | |
| A6 | 83.43 | 43.55 | 1 566.5 | 120.0 | 上 Up | 27.5 | 400.0 | 16 | |
| B4 | 83.44 | 43.54 | 1 205.2 | 151.3 | 下 Down | 33.7 | 192.9 | 7 | |
| B5 | 83.43 | 43.55 | 1 341.4 | 97.6 | 中 Middle | 38.0 | 110.0 | 4 | |
| B6 | 83.43 | 43.55 | 1 563.4 | 89.6 | 上 Up | 41.7 | 143.1 | 5 | |
| C4 | 83.44 | 43.55 | 1 291.0 | 163.7 | 中 Middle | 24.3 | 911.0 | 36 | |
| C5 | 83.43 | 43.55 | 1 459.3 | 191.3 | 上 Up | 38.7 | 975.6 | 39 | |
| C6 | 83.43 | 43.55 | 1 555.8 | 138.3 | 上 Up | 37.0 | 1 065.0 | 42 | |
| XMHE | A7 | 82.73 | 43.19 | 1 360.8 | 232.3 | 中 Middle | 28.0 | 400.0 | 16 |
| A8 | 82.72 | 43.21 | 1 293.9 | 81.3 | 下 Down | 26.7 | 400.0 | 16 | |
| A9 | 82.71 | 43.22 | 1 297.0 | 90.3 | 下 Down | 34.7 | 400.0 | 16 | |
| B7 | 82.73 | 43.19 | 1 360.8 | 234.6 | 中 Middle | 18.3 | 207.5 | 8 | |
| B8 | 82.72 | 43.21 | 1 289.5 | 161.7 | 下 Down | 22.7 | 207.4 | 8 | |
| B9 | 82.72 | 43.22 | 1 315.6 | 59.6 | 下 Down | 35.0 | 179.8 | 7 | |
| C7 | 82.73 | 43.19 | 1 345.8 | 231.3 | 中 Middle | 22.0 | 889.2 | 35 | |
| C8 | 82.72 | 43.21 | 1 326.0 | 92.0 | 下 Down | 32.3 | 946.6 | 37 | |
| C9 | 82.72 | 43.22 | 1 324.4 | 54.3 | 下 Down | 37.3 | 1 115.0 | 44 |
DXG, 大西沟; XHG, 杏花沟; XMHE, 小莫乎儿沟。A1-A9, 林下样地; B1-B9, 林窗样地; C1-C9, 空地样地。
DXG, Daxigou; XHG, Xinghuagou; XMHE, Xiaomohu’ergou. A1-A9, forest understory plots; B1-B9, forest gap plots; C1-C9, forest glade plots.
1.2.2 更新幼株个体特征测定
表2 新疆野杏种群更新等级划分标准
Table 2
| 基本结构 Basic structure | 等级 Grade | ||||
|---|---|---|---|---|---|
| I | II | III | IV | Ⅴ | |
| 基径 Basal diameter (cm) | ≤1 | (1, 2] | (2, 3] | (3, 5] | (5, 10] |
| 高度 Height (cm) | ≤30 | (30, 60] | (60, 100] | (100, 200] | >200 |
| 冠幅 Canopy (cm) | ≤25 | (25, 50] | (50, 100] | (100, 150] | >150 |
1.2.3 空间分布格局
表3 新疆野杏种群各指标计算公式
Table 3
| C | I | m* | PAI | Ca | K | Iδ | ||
|---|---|---|---|---|---|---|---|---|
C, 扩散型指数(方差/平均值); Ca, Cassie指数; I, Mrisita指数; Iδ, 扩散型指数; K, 负二项参数; m*, 平均拥挤度; n, 样地内野杏幼株个体总株数; N, 样地内5 m × 5 m样方总数; PAL, 聚块性指数;
C, diffusing index (variance/mean); Ca, Cassie index; I, Mrisita index; Iδ, diffusing index; K, negative binomial distribution; m*, mean crowding; n, total individual number of seedlings of Prunus armeniaca in sample plot; N, the number of 5 m × 5 m quadrats in the sample plot; PAL, index of patchiness;
表4 新疆野杏种群聚集类型判断标准
Table 4
| 分布类型 Distribution pattern | 格局指数 Pattern index | ||||||
|---|---|---|---|---|---|---|---|
| C | I | m* | PAI | Ca | K | Iδ | |
| 聚集 Cluster | > 1 | > 0 | >$\bar{X}$ | >1 | >0 | >0 | >1 |
| 随机 Random | 1 | 0 | $\bar{X}$ | 1 | 0 | 0 | 1 |
| 均匀 Uniform | <1 | <0 | <$\bar{X}$ | <1 | <0 | <0 | <1 |
指数同
Indexes see
点分布格局: 参考相关点格局研究方法(刘振等, 2012), 以样地内各幼株的坐标点为基础数据, 使用Riply’s K函数对样地内植株间距离进行分析。计算方式如下:
式中, r为空间尺度;
在实际运用中经常将Riply’s K函数变成Riply’s L函数, 转换公式如下:
本研究采用Monte-Carlo随机模拟方法计算上下包迹线, 假设种群为随机分布, 对每个r值计算出
1.3 数据分析
利用SPSS 26软件单因素方差分析法对同一分布区域不同生境、不同分布区域、不同生境野杏幼株个体特征以及不同更新等级的分布数量进行差异显著性分析(p < 0.05), 使用多变量方差分析法解析分布区域及生境类型对幼株个体特征的影响, 通过Excel 2016算出5 m × 5 m样方尺度下更新幼株的分布类型, 并用2D核密度图展现更新幼株的空间分布状况, 使用Programita 2018分析点分布格局, Origin 2021作图。
2 结果和分析
2.1 野杏幼株个体特征与分布
种群内的不同生境野杏更新幼株特征如图1所示。杏花沟林下未发现更新幼株, 更新密度表现为林窗显著大于空地, 其余种群内的不同生境间均无显著差异。大西沟更新幼株的基径、高度和冠幅均表现为空地显著大于林窗和林下。而杏花沟林窗和空地更新幼株的特征均无显著差异。小莫乎儿沟不同生境更新幼株特征的变化无明显规律, 基径表现为空地、林窗显著大于林下, 高度由林窗、空地至林下依次减小, 其中林窗和林下高度呈显著差异, 而不同生境的冠幅无显著差异。各种群内不同生境野杏更新幼株的基径、高度和冠幅多呈由林下、林窗至空地逐渐增大趋势。
图1
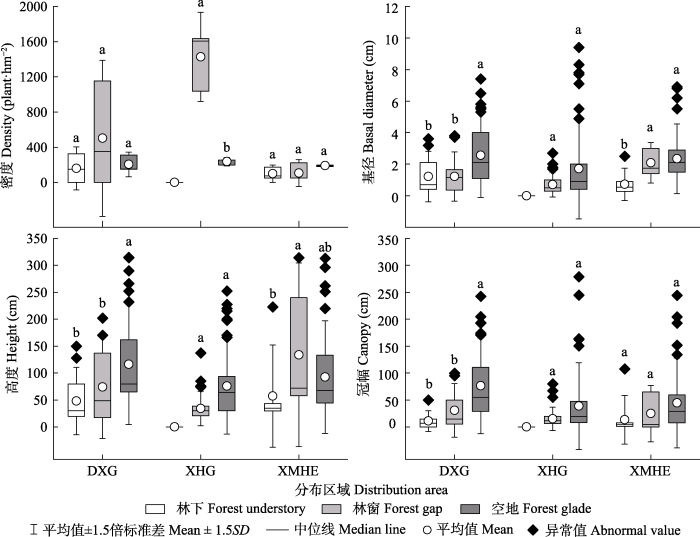
图1
新疆野杏种群内不同生境更新幼株个体特征。
不同小写字母表示同一种群下不同生境更新幼株特征的差异显著(p < 0.05)。DXG, 大西沟; XHG, 杏花沟; XMHE, 小莫乎儿沟。
Fig. 1
Characteristics of regeneration seedlings of Prunus armeniaca populations in Xinjiang in different habitats.
Different lowercase letters indicate significant differences in the characteristics of seedlings in different habitats in the same population (p < 0.05). DXG, Daxigou; XHG, Xinghuagou; XMHE, Xiaomohu’ergou.
在种群间, 野杏更新幼株的密度均无显著差异, 但纯林状况的杏花沟更新密度相对较大(表5)。大西沟和小莫乎儿沟野杏更新幼株的基径均≥2.0 cm, 且高度大于90.0 cm, 与杏花沟更新幼株的基径1.2 cm、高度56.5 cm呈显著差异。而冠幅表现为大西沟显著大于小莫乎儿沟和杏花沟, 差值分别为15.9 cm、25.6 cm。种群间更新幼株特征的差异, 表明种群更新受到分布区域内生物和非生物条件的影响。
表5 不同种群新疆野杏天然更新幼株特征的差异(平均值±标准差)
Table 5
| 分布区域 Distribution area | 林分密度 Density of forests | 野杏幼株个体特征 Individual characteristics of P. armeniaca seedlings | ||||
|---|---|---|---|---|---|---|
| 密度 Density (plant·hm-2) | 野杏占比 P. armeniaca proportion (%) | 密度 Density (plant·hm-2) | 基径 Basal diameter (cm) | 高度 Height (cm) | 冠幅 Canopy (cm) | |
| DXG | 542 ± 95a | 44.0 | 288 ± 202a | 2.0 ± 0.2a | 93.7 ± 7.1a | 53.3 ± 5.5a |
| XHG | 583 ± 14a | 100.0 | 554 ± 684a | 1.2 ± 0.1b | 56.5 ± 4.4b | 27.7 ± 3.7b |
| XMHE | 283 ± 72b | 55.9 | 132 ± 74a | 2.1 ± 0.2a | 90.2 ± 8.6a | 38.4 ± 6.0b |
DXG, 大西沟; XHG, 杏花沟; XMHE, 小莫乎儿沟。不同小写字母表示种群间的差异显著(p < 0.05)。
DXG, Daxigou; XHG, Xinghuagou; XMHE, Xiaomohu’ergou. Different lowercase letters indicate significant differences between populations (p < 0.05).
在不同生境间, 林窗内野杏幼株更新密度显著大于空地和林下, 分别为678、210和86株·hm-2 (表6)。而空地更新幼株的基径、高度和冠幅均显著大于林下和林窗, 其中基径为2.2 cm, 比林下和林窗分别多出1.2和1.3 cm; 高度93.8 cm, 分别高出林下和林窗42.0、42.5 cm; 冠幅为52.5 cm, 与林下、林窗的差值分别为40.7和32.7 cm。除此之外, 基径、高度和冠幅的最大值也均出现在空地, 表明空地生境对更新幼株的生长最为有利。总体上野杏更新幼株的平均密度、基径、高度和冠幅分别为325株·hm-2、1.7 cm、77.0 cm和38.7 cm, 参照林业天然更新等级标准(LY/T 1812), 高度≥51 cm且更新密度<501株·hm-2时则为更新不良, 该种群符合这一判定结果。
表6 不同生境新疆野杏天然更新幼株特征的差异(平均值±标准差)
Table 6
| 生境 Habitat | 野杏幼株个体特征 Individual characteristics of P. armeniaca seedlings | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 密度 Density (plant·hm-2) | 基径 Basal diameter (cm) | 高度 Height (cm) | 冠幅 Canopy (cm) | |||||||
| 最小 Min | 最大 Max | 平均值 Mean | 最小 Min | 最大 Max | 平均值 Mean | 最小 Min | 最大 Max | 平均值 Mean | ||
| 林下 Forest understory | 86 ± 37b | 0.2 | 3.6 | 1.0 ± 1.0b | 12.0 | 223.0 | 51.8 ± 50.3b | 0 | 107.5 | 11.8 ± 20.8b |
| 林窗 Forest gap | 678 ± 227a | 0.1 | 3.8 | 0.9 ± 0.8b | 9.0 | 314.0 | 51.3 ± 53.2b | 0 | 99.5 | 19.8 ± 23.2b |
| 空地 Forest glade | 210 ± 18b | 0.1 | 9.4 | 2.2 ± 1.9a | 12.0 | 315.0 | 93.8 ± 69.3a | 0 | 279.0 | 52.5 ± 58.2a |
| 总体 Population | 325 ± 89 | 0.1 | 9.4 | 1.7 ± 1.7 | 9.0 | 315.0 | 77.0 ± 66.3 | 0 | 279.0 | 38.7 ± 52.0 |
不同小写字母表示生境间的差异显著(p < 0.05)。
Different lowercase letters indicate significant differences between habitats (p < 0.05).
种群内不同更新等级野杏幼株分布密度如图2所示。不同等级基径的分布密度表现为, 大西沟I-II级均显著大于III-V级。杏花沟I级分布密度显著大于其他等级。而小莫乎儿沟仅有II级和V级存在显著差异。不同等级高度的分布密度表现为: 大西沟I级、III级和IV级均与V级存在显著差异, 并且I级还与II级存在显著差异。杏花沟I级和II级均与后3级存在显著差异。而小莫乎儿沟仅有II级和V级存在显著差异。不同等级冠幅的分布密度表现为: 大西沟I级与其他等级均存在显著差异, 并且II级、III级均与IV级、V级存在显著差异。杏花沟、小莫乎儿沟均仅有I级与其他等级存在显著差异, 其他等级间无显著差异。
图2
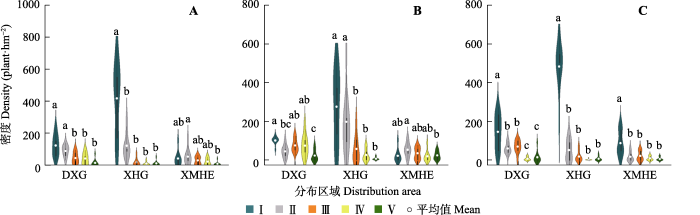
图2
新疆野杏种群内不同更新等级幼株分布密度(平均值±1.5倍标准差)。
A, 基径。B, 高度。C, 冠幅。DXG, 大西沟; XHG, 杏花沟; XMHE, 小莫乎儿沟。不同小写字母表示同一种群下不同更新等级幼株密度的差异显著(p < 0.05)。I-V见
Fig. 2
Distribution density of young plants of Prunus armeniaca with different regeneration grade in the same population in Xinjiang (Mean ± 1.5SD).
A, Basal diameter. B, Height. C, Canopy. DXG, Daxigou; XHG, Xinghuagou; XMHE, Xiaomohu’ergou. Different lowercase letters indicate significant differences in the density of seedlings of different regeneration grades under the same population (p < 0.05). I-V see
同一生境内不同更新等级野杏幼株分布密度如图3所示, 不同等级基径的分布密度表现为: 林下、林窗的I级与II-IV级均存在显著差异, 且林窗的II级与III级、IV级也存在显著差异; 空地I级、II级均与V级呈显著差异。不同等级高度的分布密度表现为: 林下I级与其他等级均存在显著差异, 且II级、III级均与IV级、V级也存在显著差异; 林窗I级和II级均与III-V级存在显著差异; 空地I级和III级与其他等级均存在显著差异, 且这两个等级之间也存在显著差异。不同等级冠幅的分布密度表现为: 林下I级和II级、IV级均存在显著差异; 林窗I级和II级、III级均存在显著差异; 空地I级与其他等级均存在显著差异, 并且II级、III级均与IV级、V级存在显著差异。整体上, 野杏种群更新幼株I-V级基径、高度和冠幅的个体分布密度均呈逐渐下降趋势, 其中I-V级基径分布密度由190株·hm-2降到7株·hm-2, 高度分布密度由122株·hm-2减少至14株·hm-2, 冠幅分布密度从238株·hm-2降至8株·hm-2, 表明野杏种群幼株在生存过程中丢失率较大, 更新受到限制。
图3
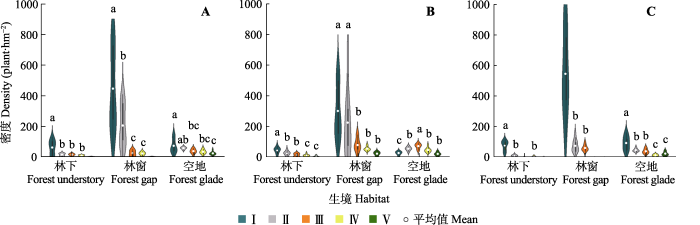
图3
新疆野杏种群生境内不同更新等级幼株分布密度(平均值±1.5倍标准差)。
A, 基径。B, 高度。C, 冠幅。不同小写字母表示同一生境下不同等级更新幼株密度的差异显著(p < 0.05). I-V见
Fig. 3
Distribution density of young plants of Prunus armeniaca with different regeneration grade in the same habitat in Xinjiang (Mean ± 1.5SD).
A, Basal diameter. B, Height. C, Canopy. Different lowercase letters indicate significant differences in the density of young plants of different regeneration grades under the same habitat (p < 0.05). I-V see
2.2 分布区域及生境对野杏幼株个体特征的影响
以分布区域和生境为自变量, 新疆野杏幼株个体特征为因变量进行多变量双因素方差分析, 结果如表7所示。在分布区域和生境两个因素中, 除了生境对冠幅主效应不显著外, 其余主效应都极显著。分布区域和生境对高度和冠幅的交互效应极显著, 对基径不显著。且偏Eta平方值的大小可表示主效应或交互效应的大小, 因此, 对更新幼株基径的影响从大到小依次为生境(0.102)、分布区域(0.047)、生境和分布区域交互效应(0.009)。对高度的影响从大到小分别为分布区域(0.090)、生境(0.074)、生境和分布区域交互效应(0.033)。而对冠幅的影响从大到小依次为分布区域(0.122)、生境和分布区域交互效应(0.071)、生境(0.001)。表明不同生境光、水、热条件的异质性对野杏幼株基径产生重要影响, 而高度和冠幅更多是受到分布区域内环境差异以及生物活动强度等因素的影响。
表7 新疆野杏幼株特征的多变量双因素方差分析
Table 7
| 自变量 Argument | 因变量 Dependent variable | 均方 Mean square | F | p | 偏Eta平方 Partial Eta square |
|---|---|---|---|---|---|
| 分布区域 Distribution area | 基径 Basal diameter | 17.098 | 7.449** | 0.001 | 0.047 |
| 高度 Height | 53 578.294 | 14.710** | 0.000 | 0.090 | |
| 冠幅 Canopy | 44 242.587 | 20.837** | 0.000 | 0.122 | |
| 生境 Habitat | 基径 Basal diameter | 38.769 | 16.890** | 0.000 | 0.102 |
| 高度 Height | 43 240.548 | 11.872** | 0.000 | 0.074 | |
| 冠幅 Canopy | 295.686 | 0.139 | 0.870 | 0.001 | |
| 分布区域×生境 Distribution area × habitat | 基径 Basal diameter | 2.023 | 0.881 | 0.451 | 0.009 |
| 高度 Height | 12 459.757 | 3.421** | 0.018 | 0.033 | |
| 冠幅 Canopy | 16 295.174 | 7.674** | 0.000 | 0.071 |
**, p < 0.01.
2.3 5 m × 5 m样方分布格局
由表8可知, 除了未发现更新幼株的5块样地, 其余样地在样方尺度为5 m × 5 m时, 7个聚集指标结果均一致。其中, 大西沟野杏种群6块样地的更新幼株均呈聚集分布(林下A1、A2, 林窗B2, 空地C1、C2、C3), 占6/7, 而B1样地呈均匀分布, A3与B3样地未发现更新幼株。杏花沟野杏种群, 林下样地均未发现更新幼株, 林窗样地(B4、B5、B6)中呈均匀分布, 空地样地(C4、C5和C6)中呈聚集分布, 各占1/2。小莫乎儿沟野杏种群, 更新幼株呈聚集(A7、A8、C7、C8、C9)、均匀(A9、B9)和随机(B7、B8)分布的样地占比分别为5/9、2/9和2/9。
表8 不同生境5 m × 5 m样方尺度下新疆野杏幼株的分布格局
Table 8
| 分布区域 Distribution area | 样地 Sample plot | 格局指数 Pattern index | 分布类型 Distribution pattern | ||||||
|---|---|---|---|---|---|---|---|---|---|
| C | I | m* | PAI | Ca | K | Iδ | |||
| DXG | A1 | 3.318 | 2.318 | 3.130 | 3.853 | 2.853 | 0.351 | 3.897 | C |
| A2 | 2.089 | 1.089 | 1.464 | 3.904 | 2.904 | 0.344 | 4.267 | C | |
| A3 | - | - | - | - | - | - | - | - | |
| B1 | 0.167 | -0.833 | 2.167 | 0.722 | -0.278 | -3.600 | 0.762 | U | |
| B2 | 2.086 | 1.086 | 1.986 | 2.207 | 1.207 | 0.828 | 2.222 | C | |
| B3 | - | - | - | - | - | - | - | - | |
| C1 | 1.603 | 0.603 | 1.403 | 1.754 | 0.754 | 1.327 | 1.759 | C | |
| C2 | 2.556 | 1.556 | 1.931 | 5.148 | 4.148 | 0.241 | 5.333 | C | |
| C3 | 1.787 | 0.787 | 1.168 | 3.065 | 2.065 | 0.484 | 3.150 | C | |
| XHG | A4 | - | - | - | - | - | - | - | - |
| A5 | - | - | - | - | - | - | - | - | |
| A6 | - | - | - | - | - | - | - | - | |
| B4 | 0.983 | -0.017 | 2.841 | 0.994 | -0.006 | -171.429 | 0.995 | U | |
| B5 | 0.370 | -0.630 | 3.870 | 0.860 | -0.140 | -7.147 | 0.889 | U | |
| B6 | 0.391 | -0.609 | 3.991 | 0.868 | -0.132 | -7.557 | 0.889 | U | |
| C4 | 2.686 | 1.686 | 2.186 | 4.371 | 3.371 | 0.297 | 4.471 | C | |
| C5 | 2.093 | 1.093 | 1.734 | 2.705 | 1.705 | 0.587 | 2.730 | C | |
| C6 | 2.035 | 1.035 | 1.678 | 2.610 | 1.610 | 0.621 | 2.632 | C | |
| XMHE | A7 | 1.578 | 0.578 | 0.765 | 4.082 | 3.082 | 0.325 | 5.333 | C |
| A8 | 2.429 | 1.429 | 1.866 | 4.265 | 3.265 | 0.306 | 4.571 | C | |
| A9 | 0.933 | -0.067 | 0.058 | 0.467 | -0.533 | -1.875 | 0.000 | U | |
| B7 | 1.000 | - | 0.125 | 1.000 | - | - | - | R | |
| B8 | 1.000 | - | 0.125 | 1.000 | - | - | - | R | |
| B9 | 0.500 | -0.500 | 0.071 | 0.125 | -0.875 | -1.143 | 1.167 | U | |
| C7 | 1.862 | 0.862 | 1.347 | 2.774 | 1.774 | 0.564 | 2.831 | C | |
| C8 | 1.644 | 0.644 | 1.103 | 2.401 | 1.401 | 0.714 | 2.449 | C | |
| C9 | 1.442 | 0.442 | 0.942 | 1.884 | 0.884 | 1.132 | 1.905 | C | |
DXG, 大西沟; XHG, 杏花沟; XMHE, 小莫乎儿沟。C, 聚集分布; R, 随机分布; U, 均匀分布。其他指数同
DXG, Daxigou; XHG, Xinghuagou; XMHE, Xiaomohu’ergou. C, cluster distribution; R, random distribution; U, uniform distribution. Other indexes see
有更新幼株的5块林下样地, 聚集和均匀分布样地分别占4/5和1/5; 有更新幼株的8块林窗样地,聚集、均匀和随机分布样地分别占1/8、5/8和2/8; 9块空地样地中均为聚集分布。即聚集、均匀和随机分布样地的比率分别为63.6%、27.3%和9.1%。因此, 在5 m × 5 m样方法中, 新疆野杏种群天然更新幼株呈聚集分布。
2.4 点分布格局
在不同分布区选择具有代表性的样地作为点格局分布研究样地, 其中大西沟选择A1、B1、C1, 杏花沟为A4、B4、C4, 小莫乎儿沟因林窗出现单株随机分布状况, 故选择A8、B9、C8样地。由图4、图5可知新疆野杏更新幼株空间分布尺度, 大西沟林下天然更新幼株在1-10 m尺度内均为聚集分布, 当尺度为7 m时聚集强度达到最大(5.4); 林窗中幼株在1-2 m尺度为聚集分布, 2-5 m尺度为随机分布, 1 m尺度时聚集强度最大, 为4.6; 空地中幼株在1-7 m尺度内为聚集分布, 8-15 m尺度为随机分布, 在5 m尺度时聚集强度达到最大(5.0)。杏花沟林下未发现野杏更新幼株个体; 林窗的更新幼株在1-2 m尺度内为聚集分布, 3-7 m尺度时为随机分布, 尺度等于1 m时聚集强度为2.2; 空地中更新幼株在1-15 m尺度内均为聚集分布, 尺度在8 m时聚集强度最大, 为3.6。小莫乎儿沟林下野杏天然更新幼株, 在1-9 m尺度内为聚集分布, 10 m尺度为随机分布, 当尺度为5 m时聚集强度达到最大值(5.9); 林窗中幼株在尺度为1 m时呈聚集分布, 2-6 m时呈随机分布, 其中, 尺度等于1 m时聚集强度为7.0; 空地中更新幼株在1-13 m尺度内均为聚集分布, 14-15 m尺度为随机分布, 6 m尺度时聚集强度达到最大(4.5)。新疆野杏种群天然更新幼株在林下、空地的聚集强度随尺度变大而先增后减, 最高聚集强度在5-8 m尺度。其中林下、空地的聚集规模分别为91.6-109.3 m2和40.7-78.5 m2, 当尺度为8 m时, 分布格局由聚集趋向随机; 林窗中聚集强度随尺度增加而减小, 在1 m时聚集强度达到最大, 即聚集规模为15.2- 153.9 m2, 而当尺度≥2 m后, 呈随机分布。新疆野杏更新幼株总体分布格局随尺度增加由聚集分布趋向随机分布。
图4
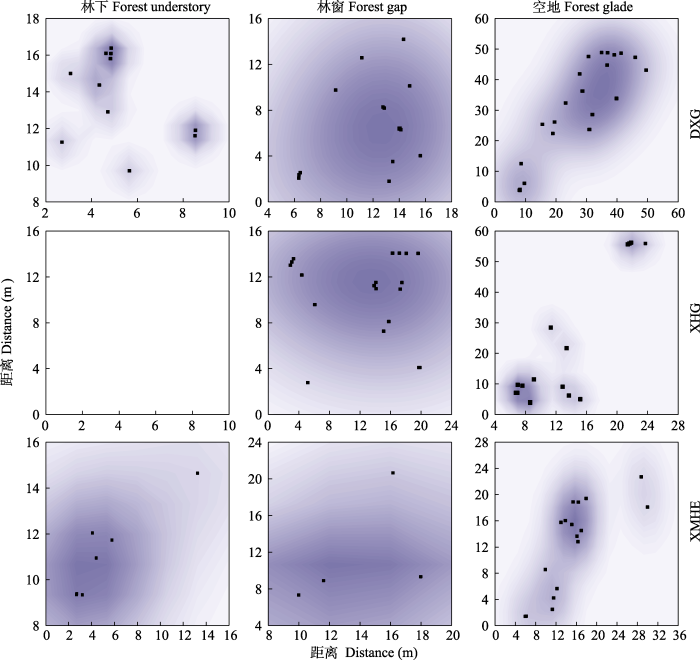
图4
新疆野杏种群内不同生境幼株的空间分布点。
DXG, 大西沟; XHG, 杏花沟; XMHE, 小莫乎儿沟。
Fig. 4
Spatial distribution point of young plants of Prunus armeniaca with different habitats in the populations in Xinjiang.
DXG, Daxigou; XHG, Xinghuagou; XMHE, Xiaomohu’ergou.
图5
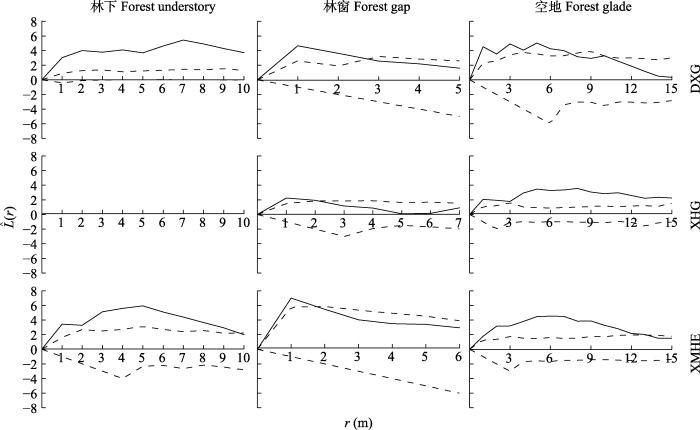
图5
新疆野杏种群内不同生境幼株空间分布格局。
图中曲线为
Fig. 5
Space distribution pattern of Prunus armeniaca young plants with different habitats in the population in Xinjiang.
The curve in the figure is the
3 讨论
3.1 分布区域及生境对新疆野杏幼株个体特征的影响
本研究结果表明不同分布区域野杏更新幼株个体特征差异显著, 以往有研究表明栎类(Quercus spp.)在不同林型中更新密度和生长状况存在显著差异(黄朗等, 2019)。一方面, 种群分布环境非生物因素的异质性是导致更新状况出现差异的因素之一(Dey et al., 2019; 翟朝阳等, 2019)。大西沟整体坡向朝正南, 充足的光照和温度利于更新幼株生长(马莉薇等, 2010); 同样为中下坡位的小莫乎儿沟, 幼株生长状况与大西沟相似, 但成年母树株数较少, 造成林分结构差异, 可能是导致更新密度低的原因; 而杏花沟海拔高, 坡度大, 更新幼株缺乏适宜的温度和水分条件, 生长受到抑制。立地条件对更新幼株的存活有显著影响(Pardos et al., 2005), 本研究结果与此相符。另一方面, 种群分布环境的生物因素也会引起更新状况出现差异(李宁等, 2011), 3个种群均受到不同程度的生物干扰, 包括旅游、放牧以及农垦等。实地调查发现, 大西沟、小莫乎儿沟常 年有人类、牧群活动, 强干扰状态下草本层植物低, 区别在于大西沟次级小沟众多, 种群分布范围广, 并且沟内较多灌丛起到保护更新幼株的作用。小莫乎儿沟野杏沿主沟两侧分布, 沟内面积狭小, 且沟口有村庄, 平时生物活动强度较大。而杏花沟因坡陡且高, 人类活动主要在下坡位, 造成野杏幼株主要分布于中上坡位, 同时夏季牧群转场, 草本植物高度增长明显, 增加了植物之间的相互竞争(胡雪凡等, 2021), 虽然更新密度高, 但生长也明显受到抑制, 这也是造成3个野杏种群更新幼株个体特征存在差异的重要因素。整体上野杏天然更新幼株个体特征受研究区域内生物和非生物因素影响显著, 在今后的研究中还应该关注如何度量各因素对更新的影响程度。
不同生境条件可使幼株的更新密度存在显著差异(贺丹妮等, 2020), 对植物天然更新具有重要的影响。新疆野杏以实生繁殖进行天然更新, 种子具有非深度生理休眠特性(张渊等, 2016), 即在不同生境中均可萌发定居, 但生境条件的异质性会导致不同生境更新幼株的特征存在差异(何中声等, 2012), 这已被以往樟子松(Pinus sylvestris var. mongolica)更新幼苗的研究所证实(王锋和卢琦, 2019)。本研究中, 林窗生境更新密度显著高于空地和林下, 除了更新密度, 野杏幼株在空地生境中个体特征均显著优于林窗和林下, 其原因可能是林下至空地中心光照、温度、湿度显著的梯度性所致(Gagnon et al., 2003; 刘少冲等, 2012)。野杏具有喜光、耐旱的特性, 更新幼株可利用空地所提供的较大空间尺度, 充分接收气候资源, 降低个体之间的竞争作用, 从而促进生长(朱凯月等, 2017)。说明稀疏林分或无上层林木遮挡更利于野杏幼株更新。
3.2 野杏幼株的空间分布格局
种群在一定空间或不同发育阶段均会形成特有的分布格局, 是不断适应环境条件和自然选择的结果(兰国玉和雷瑞德, 2003)。以往有研究表明新疆野核桃种群在25 m2时整体呈聚集分布(张维等, 2016), 锐齿栎(Quercus aliena var. acuteserrata)从幼树到大树由聚集到随机甚至均匀分布(常伟等, 2016), 黄枝润楠(Machilus versicolora)林更新幼株呈聚集分布(吴联杯等, 2019)。本研究中, 新疆野杏更新幼株在5 m × 5 m尺度下主要呈聚集分布, 这与多数以往种群更新幼株分布结果研究相似, 而不同生境更新幼株的聚集强度有所不同, 其原因是野杏生物学与生态特性、野生生境条件、取样尺度和样地面积的综合作用所致(兰国玉和雷瑞德, 2003)。虽然林下生境更易聚集大量种源, 但树冠下充足的土壤水分带来的正效应并不足以弥补太阳辐射减弱和土壤温度降低带来的负效应(张盈和李君, 2018), 使得更新幼株只有在光照、温度适宜的小尺度下才能顺利更新(Calviño-Cancela, 2007), 从而造成聚集效果明显。通常林窗形成后改善了光照的利用率, 物种丰富度相对林下明显增加, 本研究中林窗生境的草本和灌木可直接截获野杏扩散的种源, 同时还可以改善土壤养分(Gómez-Aparicio, 2009)、水分(Li et al., 2007)条件, 形成利于种子萌发的适宜生境, 使得更新幼株分布数量显著高于其他生境, 呈均匀分布, 但其对更新幼株后期的生长起抑制作用。空地多为土层较薄的脊坡与陡坡, 植被覆盖度低, 草本生物量较林下和林窗低, 易受动物活动干扰, 建植的野杏幼株个体常分布于散生灌木丛中, 或者以灌木丛为中心聚集分布, 有效减少了动物啃食带来的机械损伤, 灌木为其建造了适宜生境(Peláez et al., 2019; Guignabert et al., 2020)。在本研究尺度范围内, 新疆野杏更新幼株点格局总体呈聚集分布, 不同生境下均随尺度增大由聚集趋向随机分布, 这与急尖长苞冷杉(Shen et al., 2016)、栓皮栎(Quercus variabilis) (樊登星和余新晓, 2016)等树种天然更新格局相似。其原因与野杏种子扩散机制(张维等, 2016)、生境条件(张中惠等, 2021)有关。如野杏成熟种子颗粒饱满(质量大), 自由落种时被截获数量由林下、林窗至空地依次递减, 同时坡度、网状牧道、植被覆盖度也会影响种子扩散距离。而动物对种子的二次搬运也是影响其分布格局形成的重要因素, 在果实成熟期, 除了大型动物的食用搬运外, 小型动物常将果实运至灌木丛中及其周边, 食用果肉后留下成堆种子, 这也是空地生境稀疏植被条件下野杏更新幼株依靠灌木分布的重要因素。野杏天然更新以聚集为主的格局分布, 是其适应自然选择、实现更新建植的策略。其能否实现天然更新, 与母树分布生境及萌发条件、土壤因子、水分保持、光照等环境条件密切相关, 同时, 与被搬运的种子能获得适宜的更新生境有关。如灌丛的微气候和小生境适宜其更新建植, 比单株孤立个体有更好的抵御能力(宋萍等, 2005)。
3.3 野杏幼株个体特征与分布格局的关系
4 结论
通过调查发现, 新疆野杏更新幼株的生长对不同区域、不同生境作出了不同程度的响应, 其中, 大西沟和小莫乎儿沟均表现为更新幼株个体不足, 杏花沟缺乏更新后期幼株。
在不同生境中, 林下虽然是种子萌芽的适宜生境, 但是其环境条件最大程度上抑制了更新幼株的数量, 只有在小尺度上个别植株能实现更新; 林窗利于扩散后种子的萌发定植, 但幼株生长显著受阻; 空地显著促进更新植株的生长发育, 但阻碍更新前期幼株生存, 是限制种群在此生境更新的主要原因。
综合分析表明, 该种群天然更新在一定程度上受到抑制, 更新幼株主要呈聚集分布则是更新受阻下依靠群体抵抗限制的表现。现有更新与建植, 是其对生境变化、生存胁迫以及放牧干扰等因素作出的自然选择, 是种群可以延续发展的策略。在此背景下, 应加强干预保护, 以促进新疆野杏种群更新。
参考文献
Population structure, age and growth rates of conifer species and their relation to environmental variables at Malam Jabba, Swat District, Pakistan
DOI:10.1007/s11676-018-0820-5 [本文引用: 1]
A study on the structure and dynamics of Cyclobalanopsis glauca population at hills around lake in Hangzhou
杭州西湖山区青冈种群结构和动态的研究
Seed and microsite limitations of recruitment and the impacts of post-dispersal seed predation at the within population level
DOI:10.1007/s11258-006-9223-3 URL [本文引用: 1]
Morphological diversity of branches and leaves in Armeniaca vulgaris at Daxigou of Huocheng
霍城县大西沟野杏枝叶形态多样性
Spatial pattern of secondary Quercus aliena var. acuteserrata forests in the Qinling Mountains
秦岭林区锐齿栎次生林种群空间分布格局
Barriers to natural regeneration in temperate forests across the USA
DOI:10.1007/s11056-018-09694-6
[本文引用: 1]

For millennia, natural disturbance regimes, including anthropogenic fire and hunting practices, have led to forest regeneration patterns that created a diversity of forest lands across the USA. But dramatic changes in climates, invasive species, and human population, and land use have created novel disturbance regimes that are causing challenges to securing desired natural regeneration. Climate is an ever-present background disturbance and determinant of species distribution. Changes in certain other factors such as large herbivore populations, wildfire, and pests modify forest composition and structure, and are common barriers to natural regeneration of desired species. Changes in long-standing disturbance regimes have led to the homogenization of forest landscape composition and structure. Today, forests have low regeneration potential and are low in resilience. They have reduced productivity and are prone to widespread health issues including severe forest mortality. In addition to epidemics of native invasive species due to climate change and availability of habitat at landscape scales, the continued introduction and spread of non-native pests and diseases are causing large-scale forest mortality. These ecological changes have cascading ecological consequences such as increases in severe wildfire, which pose new barriers to natural regeneration. Equally challenging are the barriers to natural regeneration that arise from social, political and economic factors. To address many of these issues requires active management that links all critical stages in the regeneration niche necessary for achieving desired regeneration to sustain forest development and production in a socially acceptable manner and economically viable market system.
Spatial point pattern analysis of Quercus variabilis and Pinus tabulaeformis populations in a mountainous area of Beijing
北京山区栓皮栎林优势种群点格局分析
Effects of human disturbance on plant diversity of wild fruit forests in western Tianshan Mountain
人为干扰对西天山野果林群落结构和物种多样性的影响
Dynamics of artificial regeneration in gaps within a longleaf pine flatwoods ecosystem
DOI:10.1016/S0378-1127(01)00808-8 URL [本文引用: 1]
The role of plant interactions in the restoration of degraded ecosystems: a meta-analysis across life-forms and eco-systems
DOI:10.1111/jec.2009.97.issue-6 URL [本文引用: 1]
Spatial structure of natural mixed coniferous-broadleaf secondary forest in Dahuofang Reservoir area, eastern Liaoning
To understand the merit of spatial structure of coniferous-broadleaf secondary forests in Dahuofang Reservoir area, eastern Liaoning, we measured four structure parameters, i.e., angular scale, neighborhood comparison, mixing degree and stand spatial structure index, to analyze the characteristics of stand spatial structure in upper slope and lower slope. The results showed that the horizontal distribution of the forest in the upper slope was randomly distributed, while that in the lower slope was characterized by a slight clustering distribution. In terms of individual plant size in the two positions, a subdominant state was found in the lower slope, while moderate neutral state was found in upper slope. The mixing degree was 0.29 and 0.56 for the upper and lower slope respectively, with the latter one being better. The spatial structure index was 54.81 and 68.04 for the upper and lower slope, respectively, so the integral structure in the lower slope was better. Our results suggested that the forest spatial structure in the lower slope was better than that in the upper slope, which provided a basis for managing the forest structure in this area.
辽东大伙房水库库区天然针阔次生林林分空间结构
为了解辽东大伙房水库库区天然针阔次生林内林分空间结构的优劣,本研究以角尺度、大小比数、混交度和林分空间结构指数这4个结构参数作为切入点,对天然针阔次生林上坡位和下坡位的林分空间结构特征进行了调查。结果表明:上坡位的林木水平分布格局表现为随机分布,下坡位表现为轻微的聚集分布,从两种坡位中林木的大小分化程度来看,下坡位偏亚优势状态,上坡位偏中庸状态;上坡位混交度为0.29,下坡位混交度为0.56,上坡位林分呈弱度混交,下坡位为中度混交,下坡位混交程度较上坡位好;下坡位林分的林分空间结构指数为68.04,上坡位为54.81,林分整体结构较上坡位更稳定。通过4种林分空间结构参数的分析,得出天然针阔次生林下坡位的林分空间结构较上坡位稳定,本研究结果可为该区林分空间结构调整提供一定的依据。
Complex biotic interactions mediated by shrubs, revisiting the stress-gradient hypothesis and consequences for tree seedling survival
DOI:10.1111/jpe.v57.7 URL [本文引用: 1]
Effects of gap size on seedling natural regeneration in artificial Pinus tabulaeformis plantation
油松人工林林窗对幼苗天然更新的影响
Density and spatial distribution of seedlings and saplings in different gap sizes of a spruce-fir mixed stand in Changbai Mountains, China
长白山云冷杉针阔混交林不同林隙下幼苗幼树密度及空间分布
DOI:10.13287/j.1001-9332.202006.004
[本文引用: 1]
2019年8月在云冷杉针阔混交林样地(0.36 hm<sup>2</sup>),对48个林隙及幼苗(0.2<更新高度RH<1 m)、幼树(RH≥1 m,胸径DBH<5 cm)进行调查,分析林隙大小(<20 m<sup>2</sup>,小;20~50 m<sup>2</sup>,中;50~120 m<sup>2</sup>,大;>120 m<sup>2</sup>,特大)对林隙内红松、鱼鳞云杉及冷杉幼苗幼树密度和生长指标(高、基径)的短期影响,并采用核密度估计法分析其空间分布规律。结果表明: 云冷杉更新的密度通常随林隙增大而降低,仅对幼树影响显著,小林隙下云冷杉幼树密度分别为0.34和1.74株·m<sup>-2</sup>,红松密度不受林隙大小的影响。林隙大小对冷杉幼苗幼树生长指标的影响最大,对红松影响最小,平均最大值多出现在大林隙。红松和云杉幼树的基径和树高最大值均分布在小、中、大林隙东北部,在特大林隙中转移至冠空隙西北部。小林隙有助于幼苗的建立和萌发,可通过择伐冷杉创造小林隙,随后扩大林隙面积(>50 m<sup>2</sup>)促进幼树生长,需要持续监测来确定林隙大小对森林更新的长期影响。
Studies on the seeds dispersal and seedlings regeneration in gaps and understory of Castanopsis kawakamii natural forest
格氏栲天然林林窗和林下种子散布及幼苗更新研究
Establishment and evaluation of tree competition index based on intersection and crowding
基于交角和密集度的竞争指数构建及评价
natural secondary forests in Hunan Province, China
湖南栎类天然次生林幼树更新特征及影响因子
Spatial distribution pattern of seedlings and saplings of three forest types by natural regeneration in Daxing’an Mountains Xinlin Forestry Bureau, China
大兴安岭新林林业局3种林分类型天然更新幼苗幼树的空间分布格局
DOI:10.13287/j.1001-9332.201709.014
[本文引用: 1]
基于大兴安岭新林林业局落叶松林、白桦林和混交林3种主要林分类型16块样地的调查数据,依照幼苗幼树的株高划分为3个等级: Ⅰ级, 株高≤60 cm; Ⅱ级, 60 cm<株高≤200 cm; Ⅲ级, 株高>200 cm且胸径<5 cm.在4种取样尺度下,采用负二项指数等7种聚集度指数,判定幼苗幼树的分布格局及格局强度,分析格局变化和格局规模.结果表明: 除落叶松林的落叶松幼苗幼树Ⅱ级最多,且随等级升高幼苗幼树密度先升高后降低外,白桦林和混交林的落叶松以及3种林分的白桦幼苗幼树Ⅲ级最多,而Ⅰ级最少,随等级升高,幼苗幼树密度均升高.落叶松幼苗幼树在3种林分4种取样尺度下均呈聚集分布.白桦幼苗幼树在白桦林中除在10 m×20 m尺度为随机分布外,均呈聚集分布.除落叶松幼苗幼树在混交林中5 m×10 m尺度时聚集强度最大外,落叶松在其他林分以及白桦在3种林分均在5 m×5 m尺度下聚集强度最大.在落叶松林,落叶松和白桦幼苗幼树的聚集强度随等级增大而减小,其中白桦由聚集分布变为随机或均匀分布;在白桦林和混交林中,落叶松和白桦幼苗幼树的聚集强度随等级升高不断增大,由均匀分布变为聚集分布.在3种林分类型中,不同更新等级落叶松和白桦幼苗幼树种群的格局规模均为25 m<sup>2</sup>.落叶松和白桦幼苗幼树种群格局强度在不同林分类型、不同更新等级中差异明显.
Population structure and dynamics of Torreya fargesii Franch., a plant endemic to China
中国特有植物巴山榧树的种群结构与动态
Size-class structure and distribution pattern of Emmenopterys henryi in different habitats
不同生境香果树种群的径级结构与分布格局
Brief introduction of spatial methods to distribution patterns of population
植物种群空间分布格局研究方法概述
Effect of plant species on shrub fertile island at an oasis-desert ecotone in the South Junggar Basin, China
DOI:10.1016/j.jaridenv.2007.03.015 URL [本文引用: 1]
Recruitment limitation of plant population, from seed production to sapling establishment
植物种群更新限制——从种子生产到幼树建成
Determination of genome size and chromosome ploidy of selected taxa from Prunus armeniaca by flow cytometry
DOI:10.1016/j.scienta.2019.108987 [本文引用: 1]
Genetic diversity, population structure, and relationships of apricot (Prunus) based on restriction site-associated DNA sequencing
DOI:10.1038/s41438-020-0284-6 [本文引用: 1]
Dynamic changes in soil temperature water content nutrition and microorganisms of different size gaps in the mixed broadleaved Korean pine forest
阔叶红松林不同大小林隙土壤温度、水分、养分及微生物动态变化
Identification of pathogenic fungi causing leaf spot of Malus sieversii and Impatiens brachycentra growing in the wild fruit forest of Tianshan Mountain
天山野果林塞威士苹果和短距凤仙花叶斑病病原真菌的鉴定
The spatial point pattern of Ulmus pumila population in two habitats in the Otindag sandy land
浑善达克沙地2种生境下榆树种群空间点格局
Path analysis between diameter at breast height, height and crown width of Cunninghamia lanceolata in different age
不同林龄杉木胸径树高与冠幅的通径分析
Effects of spatial structure on DBH increment of natural spruce-fir forest
云冷杉天然林林分空间结构对胸径生长量的影响
Age composition and dynamic characteristics of the main populations of endangered Malus sieversii
新疆野苹果(Malus sieversii)种群年龄结构及其动态特征
Growth characteristics and influencing factors of Quercus variabilis seedlings on the north slope of Qinling Mountains
秦岭北坡不同生境栓皮栎实生苗生长及其影响因素
Ecophysiology of natural regeneration of forest stands in Spain
DOI:10.5424/srf/2005143-00939 [本文引用: 1]
Nurse plant size and biotic stress determine quantity and quality of plant facilitation in Oak savannas
DOI:10.1016/j.foreco.2019.02.010 URL [本文引用: 1]
Spatial pattern analysis and associations of different growth stages of populations of Abies georgei var. smithii in Southeast Tibet, China
DOI:10.1007/s11629-016-3849-y URL [本文引用: 1]
Population structure and its dynamics of rare and endangered plant Alsophila spinulosa
珍稀濒危植物桫椤种群结构与动态研究
采用空间序列代替时间变化的方法,对7个地段中的桫椤种群大小结构进行分析,运用C、K、m<sup>*</sup>、m<sup>*</sup>/m和Iδ5种聚集度指标测定不同地段内桫椤种群的空间分布格局及其动态,并根据植株个体点位图,考察桫椤种群在不同取样尺度上的空间分布格局.结果表明,不同地段中的桫椤种群结构存在增长型、稳定型、成熟型和衰退型4种类型;不同地段中的桫椤种群空间分布格局有所差异,表现为集群分布或随机分布;在桫椤种群生长过程中,分布格局从集群型向随机型转变;种群扩散型指数Iδ随取样尺度的增大而减小.
A spatial-explicit seedling recruitment model for scattered individual trees of Pinus sylvestris var
沙地樟子松散生单木的天然更新幼苗空间分布模型
Spatial structure pattern of regeneration seedlings from over- logged forest in Jingouling forest farm
金沟岭林场过伐林更新幼苗空间结构分析
Basic characteristics and distribution type of regeneration saplings and seedling under Machilus versicolora stand
黄枝润楠林下更新幼树幼苗基本特征及分布类型
Spatial point pattern analysis of Abies georgei var. smithii in forest of Sygera Mountains in southeast Tibet, China
西藏色季拉山急尖长苞冷杉种群点格局分析
基于4 hm<sup>2</sup>色季拉山长期监测样地调查资料,采用成对相关函数点格局方法,分析西藏色季拉山冷杉林群落优势种急尖长苞冷杉种群活立木不同生长阶段和不同大小死亡木的空间格局及它们之间的空间关联性.结果表明: 急尖长苞冷杉种群活立木径级分布连续,呈倒“J”型,为增长型种群.小树在小尺度(0~7 m)上呈显著聚集分布,随着龄级和研究尺度的增加,冷杉种群聚集强度逐渐减弱,最终趋向于均匀或随机分布格局.小树与大树、中树与大树在小尺度(0~35和0~30 m)上呈负关联,在大尺度(45~100和80~100 m)上呈正关联,且个体间龄级相差越大,其关联性越强.冷杉死亡所释放的空间不足以影响大树对小树的距离制约,大径级死亡木与小树在小尺度(0~34和 5~27 m)上具有负关联性,在大尺度(49~100和73~100 m)上具有正关联性.自疏作用和Janzen-Connell假说中的距离制约效应可能是色季拉山急尖长苞冷杉种群空间格局形成的重要原因.
Population structure characteristics and health evaluation of Armeniaca vulgaris Lam
新疆野杏种群结构特征与健康评价
Completing the life history of Castanopsis fargesii: changes in the seed dispersal, seedling and sapling recruitment patterns
DOI:10.1007/s10342-015-0916-9 URL [本文引用: 1]
Dynamics and correlation analysis of growth and development of root system and above-ground part of Armeniaca vulgaris Lam. in Xinyuan wild fruit forest
新源野果林野杏根系与地上部生长发育动态及相关性分析
Effects of microtopography on germination layer soil factors in Armeniaca vulgaris Lam. in Daxigou
微地形对大西沟新疆野杏萌发层土壤因子的影响
Distribution patterns of Juglans cathayensis populations at different slope aspects in Tianshan valley in Xinjiang, China
新疆天山峡谷不同坡向野核桃种群分布格局
DOI:10.13287/j.1001-9332.201610.004
[本文引用: 3]
在天山峡谷野核桃分布的4条沟谷的阳坡和阴坡设置8个4000 m<sup>2</sup>样地,应用相邻格子法对野核桃每木调查,在不同尺度(面积)上采用方差/均值比率法和偏离指数(C<sub>x</sub>)、负二项参数(K)、丛生指数(I)、平均拥挤度(m<sup>*</sup>)、聚块性指数(PAI)、聚集指数(C<sub>a</sub>)等聚集强度指数对野核桃种群分布格局进行分析.结果表明: 随尺度增大,野核桃种群由集群分布向随机分布过渡;在25 m<sup>2</sup>尺度上,种群整体呈集群分布,但不同样地种群聚集强度存在较大差异,西沟阴坡种群聚集程度最大,南沟阴坡种群聚集程度最小;在50~400 m<sup>2</sup>尺度上,种群整体呈随机分布.在25 m<sup>2</sup>尺度上,幼树、小树和壮年树均呈集群分布,壮年树聚集强度最大,小树聚集强度最小;当取样尺度达到200~400 m<sup>2</sup>时,幼树、小树、壮年树和大树均呈随机分布.野核桃种群格局规模为25 m<sup>2</sup>,格局强度为2.49~9.38 m<sup>2</sup>;种群组分中小树格局强度最大,壮年树格局强度最小.取样尺度与种群斑块大小接近,采用适合的测定方法并应用多指数综合判定,一般可获得可靠的结论.
Spatial patterns of soil seed bank and seedlings in the Gurbantunggut desert, the nurse effect of shrubberies
古尔班通古特沙漠土壤种子库和幼苗的空间格局——灌丛的“保护效应”
Study on seed storage and germination characteristics of Armeniaca vulgaris Lam
新疆野杏种子贮藏方式和萌发特性的研究
Population structure and spatial distribution pattern of Quercus wutaishanica in Liupan Mountains
六盘山辽东栎林种群结构和空间分布格局
On the size of forest gaps: can their lower and upper limits be objectively defined?
DOI:10.1016/j.agrformet.2015.06.015 URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}