



详尽的植物群落调查可以为定量或定性描述群落特征提供依据, 并且为植被类专著编研提供基础性资料(乔鲜果等, 2017)。本研究采用样地调查法对蒙古莸群落的群落结构、群落特征、群系分类及其主要影响因子进行研究, 拟解决蒙古莸群落样方数据缺乏的问题, 并为植被生态学研究及生物多样性保护进一步提供参考依据, 为《中国植物志》的编研提供基础数据和样方资料。
1 材料和方法
1.1 研究区域
研究区域基本涵盖了蒙古莸在我国北方最主要的自然地理分布区域, 从东向西分别包括内蒙古、河北、山西、陕西、宁夏、甘肃等省和自治区(表1)。研究区域内气候属温带大陆性气候, 昼夜温差较大, 日照时间较长, 年平均气温1.03-9.59 ℃, 年降水量67- 475 mm, 气候干燥, 降水主要集中在7、8月份, 区域内地形复杂多样, 包括高原、山地、沟谷、草原等。
表1 蒙古莸天然群落调查样地信息
Table 1
| 编号 No. | 调查地点 Survey site | 纬度 Latitude (° N) | 经度 Longitude (° E) | 海拔 Altitude (m) | 年降水量 Mean annual precipitation (mm) | 年平均气温 Mean annual air temperature (℃) | 调查时间 Survey time |
|---|---|---|---|---|---|---|---|
| 1 | 内蒙古呼和浩特市土默特左旗 Tumd Left Banner, Hohhot, Nei Mongol | 40.77 | 111.22 | 1 123 | 291 | 5.92 | 2018-08-02 |
| 2 | 内蒙古呼和浩特市武川县 Wuchuan County, Hohhot, Nei Mongol | 41.10 | 111.77 | 1 625 | 291 | 2.89 | 2018-08-12 |
| 3 | 内蒙古呼和浩特市清水河县 Qingshuihe County, Hohhot, Nei Mongol | 39.65 | 111.44 | 1 210 | 397 | 6.55 | 2018-08-10 |
| 4 | 内蒙古呼和浩特市和林格尔县 Horinger County, Hohhot, Nei Mongol | 40.41 | 111.93 | 1 299 | 341 | 5.11 | 2018-08-11 |
| 5 | 内蒙古鄂尔多斯市达拉特旗 Dalad Banner, Ordos, Nei Mongol | 39.94 | 109.58 | 1 349 | 325 | 6.37 | 2018-08-03 |
| 6 | 内蒙古鄂尔多斯市鄂托克旗乌兰镇 Ulan Town, Otog Banner, Ordos, Nei Mongol | 39.25 | 108.04 | 1 396 | 288 | 7.02 | 2018-08-03 |
| 7 | 内蒙古鄂尔多斯市鄂托克旗蒙西镇 Mengxi Town, Otog Banner, Ordos, Nei Mongol | 40.08 | 106.92 | 1 196 | 152 | 8.02 | 2018-08-04 |
| 8 | 内蒙古鄂尔多斯市鄂托克前旗 Otog Front Banner, Ordos, Nei Mongol | 38.41 | 107.61 | 1 326 | 304 | 7.75 | 2018-08-07 |
| 9 | 内蒙古鄂尔多斯市乌审旗 Uxin Banner, Ordos, Nei Mongol | 37.72 | 108.55 | 1 321 | 371 | 8.44 | 2018-08-07 |
| 10 | 内蒙古鄂尔多斯市准格尔旗 Jungar Banner, Ordos, Nei Mongol | 39.84 | 110.98 | 1 271 | 382 | 6.32 | 2018-08-10 |
| 11 | 内蒙古乌兰察布市凉城县 Liangcheng County, Ulan Qab, Nei Mongol | 40.68 | 112.22 | 1 336 | 317 | 4.75 | 2018-08-11 |
| 12 | 内蒙古锡林郭勒盟苏尼特右旗 Sonid Right Banner, Xilin Gol, Nei Mongol | 42.57 | 112.43 | 1 269 | 223 | 4.60 | 2018-08-12 |
| 13 | 内蒙古锡林郭勒盟二连浩特市 Erenhot, Xilin Gol, Nei Mongol | 43.58 | 112.02 | 971 | 175 | 4.13 | 2018-08-13 |
| 14 | 内蒙古锡林郭勒盟阿巴嘎旗 Abag Banner, Xilin Gol, Nei Mongol | 43.90 | 115.35 | 1 190 | 246 | 1.83 | 2018-08-13 |
| 15 | 内蒙古锡林郭勒盟东乌珠穆沁旗 Dong Ujimqin Banner, Xilin Gol, Nei Mongol | 45.66 | 116.76 | 1 018 | 247 | 1.03 | 2018-08-14 |
| 16 | 内蒙古锡林郭勒盟锡林浩特市 Xilinhot City, Xilin Gol, Nei Mongol | 43.58 | 116.14 | 1 295 | 302 | 1.72 | 2018-08-15 |
| 17 | 内蒙古锡林郭勒盟正镶白旗 Zhengxiangbai Banner, Xilin Gol, Nei Mongol | 42.26 | 114.97 | 1 421 | 319 | 2.96 | 2018-08-15 |
| 18 | 内蒙古阿拉善盟阿拉善右旗 Alxa Right Banner, Alxa, Nei Mongol | 39.79 | 103.41 | 1 382 | 88 | 8.67 | 2021-08-01 |
| 19 | 内蒙古阿拉善盟阿拉善左旗 Alxa Left Banner, Alxa, Nei Mongol | 38.88 | 105.73 | 1 654 | 202 | 6.95 | 2018-08-05 |
| 20 | 宁夏中卫市沙坡头区 Shapotou District, Zhongwei, Ningxia | 37.66 | 105.37 | 1 332 | 194 | 9.59 | 2018-08-05 |
| 21 | 宁夏银川市贺兰县 Helan County, Yinchuan, Ningxia | 38.69 | 105.98 | 1 329 | 193 | 8.38 | 2018-08-06 |
| 22 | 河北省张家口市康保县 Kangbao County, Zhangjiakou, Hebei | 41.99 | 114.85 | 1 621 | 362 | 2.73 | 2018-08-16 |
| 23 | 陕西省榆林市神木市大柳塔乡 Daliuta Town, Shenmu County, Yulin, Shaanxi | 39.29 | 110.33 | 1 206 | 399 | 7.19 | 2018-08-09 |
| 24 | 陕西省榆林市神木市西沟街 Xigou Street, Shenmu County, Yulin, Shaanxi | 38.81 | 110.48 | 978 | 422 | 8.53 | 2020-08-25 |
| 25 | 陕西省榆林市榆阳区红石桥乡 Hongshiqiao Town, Yuyang District, Yulin, Shaanxi | 38.10 | 109.12 | 1 123 | 366 | 8.80 | 2020-08-18 |
| 26 | 陕西省榆林市榆阳区镇川镇 Zhenchuan Town, Yuyang District, Yulin, Shaanxi | 37.86 | 110.11 | 911 | 446 | 9.43 | 2020-08-20 |
| 27 | 陕西省榆林市横山区 Hengshan District, Yulin, Shaanxi | 38.02 | 109.68 | 1 064 | 425 | 8.98 | 2020-08-18 |
| 28 | 陕西省榆林市靖边县 Jingbian County, Yulin, Shaanxi | 37.50 | 108.98 | 1 556 | 405 | 7.60 | 2020-08-19 |
| 29 | 山西省吕梁市兴县 Xing County, Lüliang, Shanxi | 38.48 | 111.06 | 975 | 475 | 8.38 | 2020-08-22 |
| 30 | 山西省忻州市保德县 Baode County, Xinzhou, Shanxi | 39.01 | 111.10 | 997 | 434 | 8.11 | 2020-08-24 |
| 31 | 山西省忻州市河曲县 Hequ County, Xinzhou, Shanxi | 39.28 | 111.25 | 931 | 405 | 8.00 | 2020-08-24 |
| 32 | 甘肃省兰州市安宁区 Anning District, Lanzhou, Gansu | 36.11 | 103.74 | 1 624 | 345 | 9.24 | 2019-08-22 |
| 33 | 甘肃省白银市景泰县 Jingtai County, Baiyin, Gansu | 37.05 | 104.04 | 1 735 | 209 | 7.45 | 2021-07-30 |
| 34 | 甘肃省武威市凉州区 Liangzhou District, Wuwei, Gansu | 37.69 | 102.56 | 2 532 | 272 | 4.07 | 2021-07-31 |
| 35 | 甘肃省张掖市甘州区 Ganzhou District, Zhangye, Gansu | 39.10 | 100.52 | 1 565 | 190 | 7.49 | 2021-08-04 |
| 36 | 甘肃省张掖市肃南裕固族自治县 Sunan Yugurzu Autonomous County, Zhangye, Gansu | 39.11 | 99.48 | 2 165 | 278 | 1.61 | 2021-08-05 |
| 37 | 甘肃省嘉峪关市文殊镇 Wenshu Town, Jiayuguan, Gansu | 39.69 | 97.95 | 2 016 | 124 | 4.85 | 2021-08-06 |
| 38 | 甘肃省酒泉市瓜州县 Guazhou County, Jiuquan, Gansu | 41.12 | 95.51 | 1 784 | 67 | 6.22 | 2021-08-08 |
| 39 | 甘肃省酒泉市肃北蒙古族自治县鹰嘴山 Yingzui Mountain, Subei Mongolzu Autonomous County, Jiuquan, Gansu | 39.75 | 96.35 | 2 805 | 131 | 1.43 | 2021-08-07 |
| 40 | 甘肃省酒泉市肃北蒙古族自治县马鬃山 Mazong Mountain, Subei Mongolzu Autonomous County, Jiuquan, Gansu | 41.55 | 96.97 | 2 036 | 88 | 4.06 | 2021-08-09 |
1.2 样地及样方调查
野外调查于2018-2021年的6-8月进行, 根据国家标本资源共享平台(
1.3 数据处理
相对盖度= (样方内某种植物的盖度/所有种的盖度之和) × 100%
相对密度= (样方内某种植物的株数/所有种的株数之和) × 100%
相对高度= (样方内某种植物的高度/所有种的高度之和) × 100%
IV = (相对盖度+相对密度+相对高度)/3
以重要值为测度依据, 计算群落的物种丰富度(S)、Shannon-Wiener多样性指数(Hʹ)、Simpson多样性指数(D)及Pielou均匀度指数(J), 公式如下:
式中, pi为样地内物种i的重要值, S是样地内总物种数。
依据WorldClim (
2 结果
2.1 蒙古莸的分布与环境特征
根据40个样地的调查数据和现有文献资料(国家标本资源共享平台(
根据样地环境数据显示, 蒙古莸生长的环境为温带荒漠及温带干旱草原的石质山坡、沙地、干河床、沟谷等地, 主要发育在典型的沙土及沙壤土上。40个样地的平均海拔为1 416 m, 最高海拔位于甘肃省鹰嘴山(2 805 m), 最低海拔位于陕西省镇川镇(911 m); 平均年降水量为287 mm, 最大值为山西省兴县(475 mm), 最小值为甘肃省瓜州县(67 mm), 平均年平均气温为6.08 ℃, 最高值位于宁夏自治区的沙坡头(9.59 ℃), 最低值位于内蒙古乌里雅斯太山(1.03 ℃)。
2.2 蒙古莸群落特征
2.2.1 群落数量特征及结构特征
蒙古莸种群所处群落盖度差异较大。根据40个样地的统计, 以蒙古莸为优势种的群落共31个, 31个蒙古莸群落内灌木层盖度11%-75%, 平均36%; 草本层盖度1%-73%, 平均32%。群落中平均物种丰富度每100 m2有16种, 最少每100 m2有7种, 最多每100 m2有30种。灌木层平均Shannon-Wiener多样性指数为1.02, 草本层为1.94; 灌木层平均Pielou均匀度指数为0.80, 草本层为0.79; 灌木层平均Simpson多样性指数为0.55, 草本层为0.78。群落中蒙古莸平均高度为43.79 cm, 平均株数为每100 m2有24株。
群落的垂直结构可大致分为3层: 第1层由灌木、小灌木等构成, 高度>40 cm, 主要有蒙古莸、锦鸡儿属(Caragana)植物、黄刺玫(Rosa xanthina)等; 第2层由低矮灌木或半灌木及多年生草本构成, 高10-40 cm, 主要有丛生隐子草(Cleistogenes caespitosa)、糙隐子草(Cleistogenes squarrosa)、短花针茅(Stipa breviflora)、黄囊薹草(Carex korshinskyi)、细叶石头花(Gypsophila licentiana)、草木樨状黄耆(Astragalus melilotoides)等; 第3层由半灌木、低矮杂类草构成, 高度<10 cm, 主要有百里香(Thymus mongolicus)、蒺藜(Tribulus terrestris)、牛枝子(Lespedeza potaninii)等。
2.2.2 物种组成
40个样地共记录149种植物, 以蒙古莸为优势种的31个群落中共记录种子植物140种, 蕨类植物1种, 隶属于35科104属, 其中, 单子叶植物有3科16属20种, 双子叶植物有29科86属117种, 裸子植物3种。物种较多的科有禾本科、菊科、苋科、豆科, 其次是唇形科、蔷薇科、蒺藜科、石竹科、毛茛科、夹竹桃科、麻黄科、柽柳科、鼠李科、瑞香科、伞形科、百合科、列当科、旋花科, 其余17科各含1个物种。含种数较多的属有蒿属(Artemisia, 9种)、委陵菜属(Potentilla, 4种)、锦鸡儿属(4种)、黄耆属(Astragalus, 3种)、猪毛菜属(Salsola, 3种)等。
按照存在度等级(宋永昌, 2001)可以将物种划分为5级: 0 < I ≤ 20%、20% < II ≤ 40%、40% < III ≤ 60%、60% < IV ≤ 80%、80% < V ≤ 100% (图1A)。31个蒙古莸群落中, 除蒙古莸外无V级植物和IV级植物; III级物种有7种, 包括黄囊薹草(58.06%)、蒙古韭(Allium mongolicum) (58.06%)、狗尾草(Setaria viridis) (51.61%)、糙隐子草(48.39%)、冷蒿(Artemisia frigida) (45.16%)、阿尔泰狗娃花(Aster altaicus) (45.16%)及兴安胡枝子(Lespedeza davurica) (45.16%), 皆为群落中常见伴生种; II级植物有9种, 包括黑沙蒿(Artemisia ordosica)、灌木铁线莲(Clematis fruticosa)、牛枝子3种灌木或小半灌木及黄花蒿(Artemisia annua)、虫实属(Corispermum)、芨芨草(Neotrinia splendens)等常见草本植物; I级植物最多, 占总物种数的87.94%, 多为群落中的偶见种。
图1
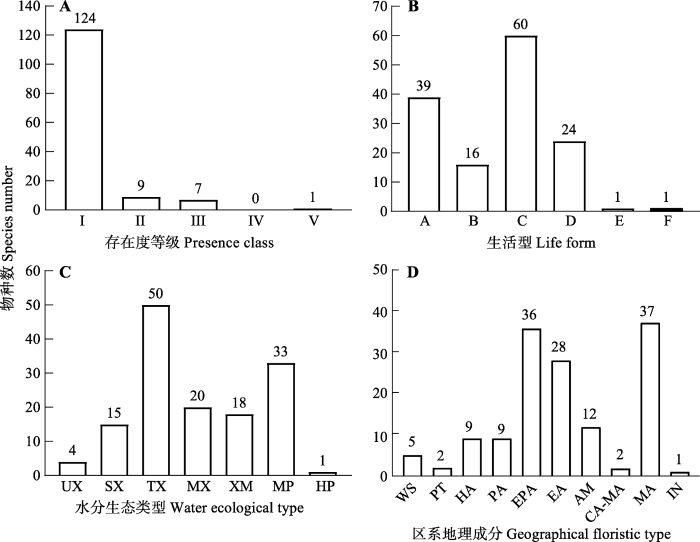
图1
蒙古莸群落的物种存在度等级分布(A)、生活型(B)、水分生态类型(C)和区系地理成分组成(D)。I, (0-20%]; II, (20%-40%]; III, (40%-60%]; IV, (60%-80%]; V, (80%-100%]。A, 灌木和小/半灌木; B, 单子叶多年生草本; C, 双子叶多年生杂类草; D, 一/二年生草本; E, 蕨类植物; F, 小半乔木。HP, 湿生; MP, 中生; MX, 中旱生; SX, 强旱生; TX, 旱生; XM, 旱中生; UX, 超旱生。AM, 古地中海; CA-MA, 中亚-亚洲中部; EA, 东亚; EPA, 东古北极; HA, 泛北极; IN, 外来入侵种; MA, 亚洲中部; PA, 古北极; PT, 泛温带; WS, 世界。
Fig. 1
Presence (A), life forms (B), water ecological types (C) and geographical floristic types (D) of the species in Caryopteris mongholica community. I, (0-20%]; II, (20%-40%]; III, (40%-60%]; IV, (60%-80%]; V, (80%-100%]. A, shrubs and dwarf/semi-shrubs; B, monocotyledonous perennial herbs; C, dicotyledonous perennial herbs; D, annuals and biennials; E, fern; F, dwarf/semi-arbor. HP, hygrophytes; MP, mesophytes; MX, meso-xerophytes; SX, strong xerophytes; TX, typical xerophytes; XM, xero-mesophytes; UX, super-xerophytes. AM, Ancient Mediterranean; CA-MA, Central Asia-Middle Asia; EA, East Asia; EPA, East Palearctic; HA, Holarctic; IN, invasive; MA, Middle Asia; PA, Palearctic; PT, pan-temperate; WS, world spread.
2.2.3 生活型及水分生态类型
31个群落中的141种植物可分划分为小半乔木、灌木和小/半灌木、单子叶多年生草本、双子叶多年生杂类草、一/二年生草本植物及蕨类植物6类生活型(图1B)。其中, 双子叶多年生杂类草最多, 主要由菊科、豆科、唇形科、蔷薇科及苋科植物组成, 占总种数的42.55%; 其次是灌木和小/半灌木, 主要有蒙古莸、黑沙蒿、锦鸡儿属植物、胡枝子属植物等, 占总种数的27.66%; 一/二年生草本占总种数的17.02%, 以黄花蒿、狗尾草、蒺藜等植物为主; 单子叶多年生草本占总种数的11.35%, 以隐子草属、黄囊薹草及短花针茅等植物为主; 小半乔木为梭梭(Haloxylon ammodendron); 蕨类植物仅卷柏(Selaginella tamariscina)。
通过查阅相关资料(赵一之, 2012)将31个群落中141种植物细化为超旱生植物、强旱生植物、典型旱生植物、中旱生植物、旱中生植物、中生植物和湿生植物共7种水分生态类型(图1C)。典型旱生植物最多, 占35.46%, 且多数是群落的优势种和常见种, 如蒙古莸、锦鸡儿属植物、隐子草属植物、黑沙蒿、百里香等; 其次是典型中生植物, 占23.40%, 主要有狗尾草、苦苣菜(Sonchus oleraceus)、黄花蒿等; 中旱生和旱中生植物分别占14.18%和12.77%, 主要有委陵菜属植物、阿尔泰狗娃花、羊草(Leymus chinensis)等; 强旱生植物占10.64%, 基本为灌木和小半灌木, 主要有沙冬青(Ammopiptanthus mongolicus)、四合木(Tetraena mongolica)、猫头刺(Oxytropis aciphylla)等; 超旱生植物有4种, 占2.84%, 为红砂(Reaumuria soongarica)、珍珠猪毛菜(Salsola passerina)、松叶猪毛菜(Salsola laricifolia)和合头藜(Sympegma regelii); 湿生植物1种, 为芦苇(Phragmites australis)。
2.2.4 物种区系地理成分
其中, 亚洲中部分布种最多, 占26.24%, 主要有黑沙蒿、柠条(Caragana korshinskii)、糙隐子草等; 其次为东古北极(亚洲温带)成分, 有蒙古莸、狭叶锦鸡儿(Caragana stenophylla)、委陵菜属植物、百里香、羊草、阿尔泰狗娃花等常见植物, 占25.53%; 东亚成分有绣线菊(Spiraea salicifolia)、黄刺玫、光药大黄花(Cymbaria mongolica)、丛生隐子草等, 占19.86%; 古地中海分布种占8.51%, 有芨芨草、驼绒藜(Krascheninnikovia ceratoides)等; 古北极分布种和泛北极分布种各占6.38%, 古北极成分包括猪毛蒿(Artemisia scoparia)和白羊草(Bothriochloa ischaemum)等, 泛北极成分包括冷蒿、黄花蒿等; 世界分布种为狗尾草及菟丝子等, 占3.55%; 中亚-亚洲中部分布种包括北芸香(Haplophyllum dauricum)和小叶锦鸡儿(Caragana microphylla) 2种, 占1.42%; 泛温带分布种有蒺藜和藜(Chenopodium album) 2种, 占1.42%; 外来入侵种仅续随子(Euphorbia lathyris) 1种。
2.3 蒙古莸群落分类
表2 蒙古莸群落分类
Table 2
| 群丛组 Association group | 群丛 Association | 样地编号 Plot number |
|---|---|---|
| 蒙古莸-草本 Caryopteris mongholica - herb | 蒙古莸-黄囊薹草 Caryopteris mongholica - Carex korshinskyi | P1, P3, P5, P19, P22, P23 |
| 蒙古莸-糙隐子草 Caryopteris mongholica - Cleistogenes squarrosa | P7, P15, P16, P20 | |
| 蒙古莸-短花针茅 Caryopteris mongholica - Stipa breviflora | P17 | |
| 蒙古莸-丛生隐子草 Caryopteris mongholica - Cleistogenes caespitosa | P11, P14 | |
| 蒙古莸-白莲蒿 Caryopteris mongholica - Artemisia stechmanniana | P2 | |
| 蒙古莸-冷蒿 Caryopteris mongholica - Artemisia frigida | P4, P8 | |
| 蒙古莸-细叶石头花 Caryopteris mongholica - Gypsophila licentiana | P6,P24 | |
| 蒙古莸-蒺藜 Caryopteris mongholica - Tribulus terrestris | P31 | |
| 蒙古莸+灌木-草本 Caryopteris mongholica + shrub - herb | 蒙古莸+醉鱼草-狗尾草 Caryopteris mongholica + Buddleja lindleyana - Setaria viridis | P21 |
| 蒙古莸+黑沙蒿-糙隐子草 Caryopteris mongholica + Artemisia ordosica - Cleistogenes squarrosa | P25 | |
| 蒙古莸+黑沙蒿-兴安胡枝子 Caryopteris mongholica + Artemisia ordosica - Lespedeza daurica | P27 | |
| 蒙古莸+黑沙蒿-白羊草 Caryopteris mongholica + Artemisia ordosica - Bothriochloa ischaemum | P29 | |
| 蒙古莸+黑沙蒿-狗尾草 Caryopteris mongholica + Artemisia ordosica - Setaria viridis | P30 | |
| 蒙古莸+灌木铁线莲-白羊草 Caryopteris mongholica + Clematis fruticosa - Bothriochloa ischaemum | P26 | |
| 蒙古莸+灌木铁线莲-菊蒿 Caryopteris mongholica + Clematis fruticosa - Tanacetum vulgare | P32 | |
| 蒙古莸+灌木 Caryopteris mongholica + shrub | 蒙古莸+霸王 Caryopteris mongholica + Zygophyllum xanthoxylon | P18, P36 |
| 蒙古莸+狭叶锦鸡儿 Caryopteris mongholica + Caragana stenophylla | P33 | |
| 蒙古莸+荒漠锦鸡儿 Caryopteris mongholica + Caragana roborovskyi | P34 | |
| 蒙古莸+灌木亚菊 Caryopteris mongholica + Ajania fruticulosa | P39 |
蒙古莸-草本群丛组由蒙古莸和根茎禾草、丛生禾草及杂类草组成, 共包括8个群丛类型。蒙古莸-黄囊薹草群丛分布最为广泛, 主要见于内蒙古的土默特左旗、清水河县、达拉特旗、阿拉善左旗及河北康保县、陕西大柳塔乡的草原带石质山坡, 地表多砾石。丛生禾草的主要优势种为糙隐子草、短花针茅和丛生隐子草。蒙古莸-糙隐子草群丛主要分布于内蒙古鄂托克旗蒙西镇、锡林郭勒盟东部, 宁夏沙坡头区域的山地草原。蒙古莸-短花针茅群丛位于内蒙古阿巴嘎旗的荒漠化草原, 分布于栗钙土或棕钙土上, 是偏干旱的类型。蒙古莸-丛生隐子草群丛位于草原带的山坡灌丛。杂类草优势种包括白莲蒿(Artemisia stechmanniana)、冷蒿、细叶石头花和蒺藜, 蒙古莸-白莲蒿群丛分布于内蒙古的武川县; 蒙古莸-冷蒿群丛位于和林格尔县和鄂托克前旗; 蒙古莸-细叶石头花群丛分布于内蒙古鄂尔多斯市乌兰镇和陕西神木市; 蒙古莸-蒺藜群丛位于山西河曲县, 多位于高原地带中部, 群丛周围坡度较缓, 受到放牧及人类活动的干扰。
蒙古莸+灌木-草本群丛组包括7个群丛。以蒙古莸+黑沙蒿-草本群丛最为常见, 主要位于陕西榆林市、山西兴县和保德县, 分布于毛乌素沙地腹部及其边缘地区的沙地和覆沙土壤上; 蒙古莸+醉鱼草(Buddleja lindleyana)-狗尾草群丛位于宁夏贺兰山景区内, 人为干扰较严重; 蒙古莸+灌木铁线莲-草本群丛位于陕西镇川镇及甘肃兰州市, 灌木铁线莲为旱生小灌木, 广泛分布于草原和草原化荒漠带的石质山坡。
蒙古莸+灌木群丛组包括4个群丛类型, 主要分布于河西走廊及附近区域的山地荒漠或荒漠带的石质山坡。蒙古莸+霸王(Zygophyllum xanthoxylon)群丛位于内蒙古阿拉善右旗和甘肃省张掖市; 蒙古莸+狭叶锦鸡儿群(Caragana roborovskyi)丛和蒙古莸+荒漠锦鸡儿群丛分别位于白银市和武威市; 蒙古莸+灌木亚菊(Ajania fruticulosa)群丛位于甘肃酒泉市。该群丛类型具有一定程度的荒漠化, 旱生/强旱生灌木为群落中的优势种, 常见草本物种为戈壁针茅(Stipa tianschanica var. gobica)。
2.4 蒙古莸群落物种多样性与地理及气候因子的关系
根据物种多样性指数与地理和气候因子的相关性分析结果, 31个蒙古莸群落灌木层及草本层的物种多样性与地理及气候因子有一定的相关性, 包括纬度、经度、降水和太阳辐射量等因素(表3)。
表3 蒙古莸群落的物种多样性指数与经纬度及气候因子相关性
Table 3
| 指标 Index | 生活型 Life form | 相关系数 Correlation coefficient | |||||
|---|---|---|---|---|---|---|---|
| 纬度 Latitude | 经度 Longitude | 海拔 Altitude | 年降水量 Mean annual precipitation | 太阳辐射量 Solar radiation | 年平均气温 Mean annual air temperature | ||
| 物种丰富度 Species richness | 灌木 Shrub | -0.288 | -0.434* | 0.181 | -0.564** | 0.315 | 0.310 |
| 草本 Herb | 0.426* | 0.641** | -0.337 | 0.418* | -0.414* | -0.248 | |
| Shannon-Wiener多样性指数 Shannon-Wiener diversity index | 灌木 Shrub | -0.368* | -0.465** | 0.218 | -0.413* | 0.245 | 0.327 |
| 草本 Herb | 0.310 | 0.587** | -0.281 | 0.532** | -0.406* | -0.233 | |
| Simpson多样性指数 Simpson diversity index | 灌木 Shrub | -0.401* | -0.465** | 0.218 | -0.261 | 0.179 | 0.322 |
| 草本 Herb | 0.207 | 0.470** | -0.175 | 0.525** | -0.373* | -0.239 | |
| Pielou均匀度指数 Pielou evenness index | 灌木 Shrub | -0.193 | -0.170 | 0.024 | 0.092 | -0.026 | 0.184 |
| 草本 Herb | 0.061 | 0.238 | 0.016 | 0.453** | -0.292 | -0.261 | |
*, p < 0.05; **, p < 0.01.
进一步将地理因子、气候因子及物种多样性指数进行RDA分析, 结果显示, 灌木、草本层物种多样性指数与地理及气候因子在前两轴的解释率分别为51.42%和3.73%, 第一轴占可解释变异的90.57%, 其相关性主要由第一排序轴决定。图2的箭头在第一轴的长度显示出环境因子和各项物种多样性指数的相关性大小: 经度>年降水量>太阳辐射量>纬度>海拔>年平均气温, 年降水量和经度对群落中各项多样性指数影响最大。由表3和图2可知, 年降水量与灌木层的物种丰富度、Shannon-Wiener多样性指数显著负相关, 与草本层的多项物种多样性指数显著正相关。经度主要与草本层物种丰富度、Shannon-Wiener多样性指数及Simpson多样性指数极显著正相关, 与灌木层多项物种多样性指数显著负相关。其次影响较大的为纬度和太阳辐射量, 纬度和灌木层Shannon-Wiener多样性指数、Simpson多样性指数显著负相关; 太阳辐射量和草本层物种丰富度、Shannon-Wiener多样性指数及Simpson多样性指数显著负相关, 但与灌木层的物种多样性指数无显著相关性。同时, 海拔和年平均气温对群落中各项物种多样性指数无显著影响。
图2
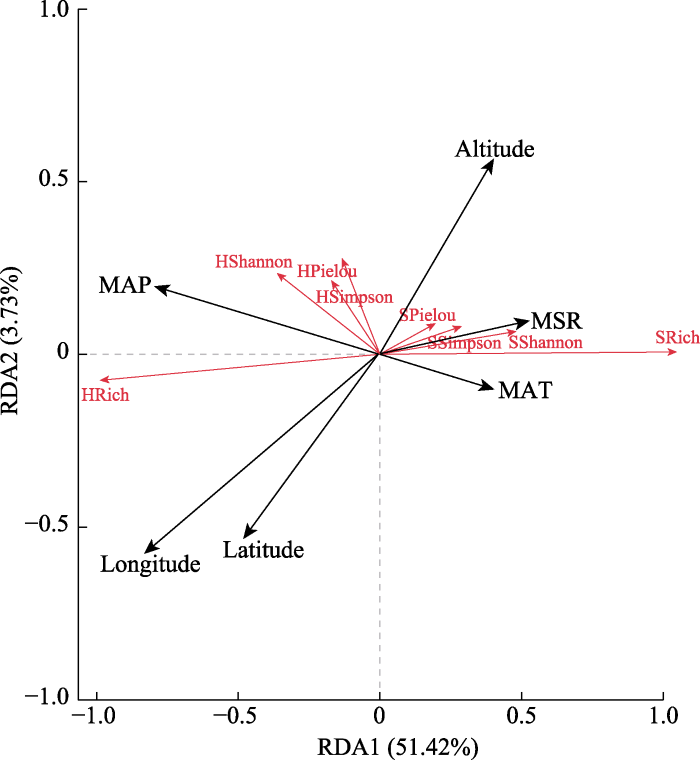
图2
蒙古莸群落物种多样性指数与环境因子的冗余分析(RDA)排序。HPielou, 草本层Pielou均匀度指数; HRich, 草本层物种丰富度; HShannon, 草本层Shannon-Wiener多样性指数; HSimpson, 草本层Simpson多样性指数; SPielou, 灌木层Pielou均匀度指数; SRich, 灌木层物种丰富度; SShannon, 灌木层Shannon-Wiener多样性指数; SSimpson, 灌木层Simpson多样性指数。Altitude, 海拔; Latitude, 纬度; Longtitude, 经度; MAP, 年降水量; MAT, 年平均气温; MSR, 太阳辐射量。
Fig. 2
Redundancy analysis (RDA) ordinations between species diversity indices of Caryopteris mongholica communities and environmental factors. HPielou, Pielou evenness index of herb; HRich, species richness of herb layer; HShannon, Shannon-Wiener diversity index of herb layer; HSimpson, Simpson diversity index of herb layer; SPielou, Pielou evenness index of shrub layer; SRich, species richness of shrub layer; SShannon, Shannon-Wiener diversity index of shrub layer; SSimpson, Simpson diversity index of shrub layer. MAP, mean annual precipitation; MAT, mean annual air temperature; MSR, solar radiation.
3 讨论
本研究中, 31个蒙古莸群落中维管植物科的数目占全国维管植物的10.00%, 属数占3.30%, 而种数仅占0.44% (金孝峰等, 2022), 物种数量偏少是由于蒙古莸种群主要分布于干旱和半干旱区, 区域内降水量较少, 生态环境较为恶劣。
黄囊薹草、冷蒿、狗尾草等为群落中常见的伴生种, 黄囊薹草、冷蒿多生于荒漠草原带及荒漠带, 而狗尾草的分布则较为广泛。群落中双子叶多年生杂类草占42.55%, 多年生草本在水分条件适宜的环境中具备更高的水分利用效率和更强的适应能力(王姣月等, 2020); 广幅旱生(包括强旱生、超旱生、典型旱生、中旱生)植物占63.12%, 反映了研究区域内干旱生态环境的严酷。物种的区系地理成分中, 东古北极(亚洲温带)成分居多, 与李登武(2008)对黄土高原区域种子植物区系的研究结果大致相同, 说明研究区域内种子植物地理成分符合该区所处地理位置为温带的特征, 广布的温带性质物种更能适应蒙古莸种群分布区的干旱、半干旱的环境, 是植物长期适应进化的结果。
对蒙古莸群落进行数量分类研究有助于了解其在空间、时间上的变化。TWINSPAN数量分类是以二歧式分割法同时进行样方和物种分类, 以此来划分植物群落类型, 其根据“指示种”将群落与物种组成依次划分为各个等级的生态类群, 具有较好的分类效果(刘博等, 2019)。本研究中, 蒙古莸既可成为群落中的优势种, 也能够成为狭叶锦鸡儿、灌木亚菊等旱生灌木群落的亚优势种或者伴生种。物种在群落中的功能或者角色受到生境地生物和非生物因素的综合影响。利用TWINSPAN将研究区内的31个蒙古莸群落划分3个群丛组, 19个群丛类型。其中蒙古莸-多年生草本群丛组包含了19个样地, 占据了蒙古莸群落样地数的61.29%, 其中草本植物多为多年生旱生禾草, 如黄囊薹草、糙隐子草、短花针茅、丛生隐子草等, 多为荒漠化草原、沙质草原的常见物种。蒙古莸+灌木-草本群丛组主要位于陕西、山西等地, 黑沙蒿、狗尾草是群落中常见的优势种, 此区域受水分胁迫较小, 高艳等(2016)的研究结果也表明, 狗尾草适宜在轻度水分胁迫条件下生长。蒙古莸+灌木群丛组共包含5个样地, 5个区域平均年降水量195 mm, 受到严重的干旱影响, 因此群落中常见的灌木植物多为超旱生或强旱生灌木, 灌木植物能以其较深的根系来固定土壤, 从而维持群落环境的稳定性(司建华等, 2011), 这与干旱、半干旱区域日照充足、降水较少、蒸发强烈、降水蒸发比例严重失衡的自然环境极其相关。
附录
附录 蒙古莸植物群落样方数据集
Supplement A plot-based dataset of Caryopteris mongholica communities
参考文献
Effects of origin climate on light response characteristics of Caryopteris mongholica
种源地气候对蒙古莸光响应特性的影响
Methods and protocols for plant community inventory
DOI:10.3724/SP.J.1003.2009.09253
[本文引用: 1]

A plant community is an assemblage of plant populations that live in certain area, and interact with and adapt to one another in the context of long-term environmental changes. Plant communities maintain global ecosystem functions, and provide food and habitats for animals and other organisms. Plant communities also provide primary resources for human survival and development, and are therefore indispensable to human societies. China is among the countries with the most diverse plant communities in the world. However, no systematic national inventory has been conducted for Chinese plant communities. This fact obstructs exploitation and protection of China’s plant resources, and also hampers the development of the fields of Chinese ecology and geography. There is an urgent need to survey Chinese plant communities using consis-tent methods and protocols. In this paper, we review major concepts in plant community ecology, and pro-pose a framework for developing plant community inventories based on recent progress in community ecol-ogy and our own experience with long-term field surveys. Our framework provides protocols for site selec-tion and plot design, items to be measured in a plot, and measurements of functional traits of dominant spe-cies. We also review protocols for field surveys of large, long-term plots. The protocols proposed in this pa-per are expected to be a base for standardizing methodology for inventory of Chinese plant communities.
植物群落清查的主要内容、方法和技术规范
DOI:10.3724/SP.J.1003.2009.09253
[本文引用: 1]
植物群落是不同植物在长期环境变化中相互作用、相互适应而形成的组合。它提供着人类赖以生存的主要物质资源, 维系着地球生态系统的健康和功能, 也为各种动物和其他生物提供食物来源和栖息地, 是人类生存和发展不可或缺的物质基础, 具有不可替代的作用。我国植物群落类型多样, 在世界上首屈一指, 但我国至今尚没有一次全面和系统的植物群落清查, 不仅影响了人们对我国植物资源的了解、利用和保护, 也不利于我国生态学、环境科学和地理学等相关学科的发展。采用统一的方法体系和技术规范开展我国植物群落的清查工作势在必行, 并具有紧迫性。本文基于作者长期的野外工作实践和国内外的群落调查方法, 首先简要定义了与植物群落清查有关的重要概念, 在此基础上, 论述了调查样地的设置原则和体系、群落清查的技术指标和方法、主要优势种生态属性的测定方法和规范, 并介绍了大样地调查的主要步骤。通过本文的介绍、归纳和总结, 试图为制定我国植物群落清查的技术规范提供基础材料和技术储备。
Distribution, community characteristics and classification of Thymus mongolicus steppe in China
DOI:10.17521/cjpe.2018.0047 URL [本文引用: 1]
中国百里香草原的分布、群落特征和分类
DOI:10.17521/cjpe.2018.0047
[本文引用: 1]
百里香(Thymus mongolicus)草原是以唇形科小半灌木百里香为建群种的草原类型。该文以编写《中国植被志》为契机, 结合2015-2017年生长季野外调查数据和文献资料, 对中国百里香草原的分布、生态特征、群落特征和分类等进行了总结。结果表明: (1)百里香草原主要分布于辽河上游西拉木伦河流域的黄土丘陵、河北西北部坝上高原、阴山山脉两翼低山丘陵、鄂尔多斯高原东部和黄土高原北部, 常分布在水蚀风蚀强烈的黄土丘陵和砾石质化基质的坡地。(2)根据91个样地记录统计, 百里香草原组成物种较丰富, 有种子植物167种, 隶属于34科101属, 以菊科、豆科和禾本科为主; 其中有8种半灌木和小半灌木, 112种多年生杂类草; 水分生态类型以典型旱生植物(58种)和中旱生植物(45种)为主; 区系地理成分以东古北极成分(70种)和东亚成分(46种)为主。(3)根据生活型和优势度, 百里香草原可以划分为百里香、半灌木/小半灌木群丛组, 百里香群丛组, 百里香、丛生禾草群丛组, 百里香、根茎禾草群丛组, 百里香、薹草群丛组, 百里香、杂类草群丛组, 共计6个群丛组28个群丛。
Interspecific competition of six succession series species with different soil moisture treatments in loess hilly region
土壤水分对黄土丘陵区演替序列种种间竞争的影响
An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV
DOI:10.1111/boj.2016.181.issue-1 URL [本文引用: 1]
A revised scheme of vegetation classification system of China
DOI:10.17521/cjpe.2019.0271
[本文引用: 2]
The principles and scheme of vegetation classification system of China (China-VCS) were first proposed by Hou in China’s Vegetation in 1960. An improved hierarchy framework and criteria of China-VCS were published in Vegetation of China in 1980. China-VCS was revised in Vegetation of China and Its Geographic Pattern — Illustration of the Vegetation Map of the People’s Republic of China (1:1 000 000) in 2007 and in Flora and Vegetation Geography of China in 2014. Song proposed a new scheme in the text book Vegetation Ecology in 2017. To facilitate Chinese vegetation study and the compilation of the book series on the vegetation of China (Vegegraphy of China), this paper provides a revised scheme of China-VCS. The scheme adopts the principles, units and hierarchy structure in the Vegetation of China that was considered as a milestone in the development of Chinese vegetation science. Based on the phytocoenological-ecological principles, plant community features and its ecological conditions are considered as the key criteria for vegetation classification. There are three levels including eight units in the hierarchy framework of China-VCS: upper level (Vegetation Formation Group, Vegetation Formation, and Vegetation Subformation), middle level (Alliance Group, Alliance, Suballiance), and lower level (Association Group, Association). Nine Vegetation Formation Groups are defined as Forest, Shrubland, Herbaceous Vegetation (Grassland), Desert, Alpine Tundra and Sparse Vegetation, Swamp and Aquatic Vegetation (Wetlands), Agricultural Vegetation, Urban Vegetation, and Non-vegetated Area. Forty-eight Vegetation Formations (30 for natural and semi-natural vegetation, 12 for agricultural vegetation, 5 for urban vegetation, and one for Non-vegetated Area) are defined, including 81 Vegetation Subformations from 23 natural and semi-natural vegetation formations. These vegetation formation groups and 30 natural and semi-natural vegetation formations were described briefly. Some issues on China-VCS, especially on the framework, criterion identifying mixed-forest, and the role of soil in vegetation classification, are discussed.
中国植被分类系统修订方案
DOI:10.17521/cjpe.2019.0271
[本文引用: 2]
为了推动《中国植被志》研编工作, 该文回顾了中国植被分类系统的发展过程和主要阶段性成果, 提出了作为《中国植被志》研编技术框架组成部分的中国植被分类系统修订方案, 对各植被型组及各植被型进行了简单定义和描述, 并针对中国植被分类系统若干问题, 特别就中国植被分类系统总体框架、混交林的界定以及土壤在植被分类中的重要性等问题进行了讨论。1960年侯学煜在《中国的植被》中首次提出了中国植被分类的原则和系统, 1980年出版的《中国植被》制定了分类等级和划分依据等更加完善的系统, 之后《中国植被及其地理格局——中华人民共和国1:1 000 000植被图说明书》和《中国植物区系与植被地理》以及很多省区的植被专著对该系统进行过修订。2017年宋永昌在《植被生态学》中提出了一个分类等级单位调整的方案。本次提出的中国植被分类系统修订方案基本沿用《中国植被》的植被分类原则、分类单位及系统, 采用“植物群落学-生态学”分类原则, 主要以植物群落特征及其与环境的关系作为分类依据, 包含三级主要分类单位, 即植被型(高级单位)、群系(中级单位)和群丛(低级单位); 在三个主要分类单位之上分别增加辅助单位植被型组、群系组和群丛组, 在植被型和群系之下主要根据群落的生态差异和实际需要可再增加植被亚型或亚群系。修订方案包含了森林、灌丛、草本植被(草地)、荒漠、高山冻原与稀疏植被、沼泽与水生植被(湿地)、农业植被、城市植被和无植被地段9个植被型组, 划分为48个植被型(含30个自然植被型、12个农业植被型、5个城市植被型和无植被地段)。自然植被中有23个植被型进一步划分出了81个植被亚型。
Geographical provenance variation of leaf anatomical structure of Caryopteris mongholica and its significance in response to environmental changes
DOI:10.17521/cjpe.2019.0117 URL [本文引用: 1]
蒙古莸叶片解剖结构的地理种源变异及其对环境变化响应的意义
DOI:10.17521/cjpe.2019.0117
[本文引用: 1]
长期受到生长环境影响而形成的遗传变异对植物生长发育有着显著的影响。叶片是植物对环境变化最敏感的器官, 了解叶片解剖结构在不同环境中产生的适应性变异是探索植物对环境适应的基础。同质园试验是研究遗传与环境因素对植物生长代谢等影响的一种有效方法, 该研究利用同质园试验排除了环境梯度的影响, 通过常规石蜡切片、多重比较、相关性分析、一般线性模型分析等方法, 对7个不同种源地的蒙古莸(Caryopteris mongholica)叶片解剖结构及其影响因素进行了定量比较。结果表明, 7个种源地的蒙古莸叶片均为等面叶, 无海绵组织分化, 其上表皮细胞较下表皮细胞厚, 上栅栏组织较下栅栏组织厚; 叶片各解剖结构参数间存在显著的自相关性, 不同种源叶片解剖结构存在显著差异: 随种源地年平均气温升高, 叶厚度、栅栏组织厚度呈增大趋势, 其中, 最西南部的阿左旗种源蒙古莸叶片的上下栅栏组织、叶厚度及叶片结构紧密度值均最大, 表现出明显的抗旱特征。种源地经纬度、气温、降水等对解剖结构指标有显著的影响, 其解释程度为34.09%-81.43%。同质园试验说明, 种源地气候差异驱动的遗传变异是引起不同种源叶片解剖结构差异的重要因素。
Species cataloging of the seed plants in Zhejiang, East China
浙江种子植物物种编目
The importance of biotic vs. abiotic drivers of local plant community composition along regional bioclimatic gradients
Study on rapid propagation technology of tissue culture of Caryopteris mongolica Bunge
蒙古莸组培快繁技术研究
A study on the character of ecology and physiology of aromatic plant——Caryopteris mongolica Bunge
芳香植物——蒙古莸的生态生理特性及其栽培技术
蒙古莸是半荒漠沙区的珍贵芳香植物之一, 不仅具有药用价值, 而且还可提取芳香油。本文从生态、生物学、水分生理等几个方面, 论证了它对沙区环境有较强的适应性, 适宜在人工固定、半固定的沙丘上生长, 可作为一个新的固沙植物种在生产中应用。
Distribution of 45S rDNA sequence on chromosomes of five species in Artemisia
45S rDNA在蒿属5种植物染色体中的分布
以小麦45S rDNA为探针,通过荧光原位杂交(FISH)技术,研究了45S rDNA在蒿属(Artemisia)5个物种染色体中的分布情况。结果发现,在根尖体细胞有丝分裂中期,中亚苦蒿(A. absinthum Linn.)有1对染色体出现杂交信号;大籽蒿(A. sieversiana Ehrhart ex Willd.)和黄花蒿(A. annua Linn.)各有两对染色体上出现信号;欧亚艾蒿(A. abrotanum Linn.)和牡蒿(A. japonica Thumb.)各有4对染色体出现杂交信号。信号位点除了分布在随体上,主要分布在非随体染色体的端部,而在欧亚艾蒿中发现1对信号位点分布在近端部,而且信号间的强弱也不尽相同。基于FISH技术的核型分析,不仅能够提供稳定、可靠的细胞学标记,而且进一步丰富了染色体分析的基础资料。
Quantitative classification and sequencing of communities in pyrogenic succession of Kanas taiga
喀纳斯泰加林火成演替群落数量分类与排序
DOI:10.16258/j.cnki.1674-5906.2019.10.005
[本文引用: 1]
可下载PDF全文。
The complete chloroplast genome of Caryopteris mongholica and phylogenetic implications in Lamiaceae
DOI:10.1007/s12686-017-0802-5 [本文引用: 1]
Distribution, community characteristics and classification of Stipa tianschanica var. klemenzii steppe in China
DOI:10.17521/cjpe.2016.0053 URL [本文引用: 1]
中国石生针茅草原的分布、群落特征和分类
DOI:10.17521/cjpe.2016.0053
[本文引用: 1]
石生针茅(Stipa tianschanica var. klemenzii)草原是中国荒漠草原最具代表性的植被类型。该文利用2010-2016年生长季的野外调查数据并结合已有文献资料, 对中国石生针茅草原的生态地理分布、群落学特征以及类型划分进行了全面系统的分析。结果表明: 中国石生针茅草原主要分布在内蒙古锡林郭勒高原西部和乌兰察布高原, 常分布在棕钙土基质的高平原上; 分布区气候干旱, 群落高度、盖度、生物量和物种丰富度较低; 根据80个样地的数据统计, 石生针茅草原共记录到种子植物165种, 隶属于29科85属, 其中以偶见种(存在度<20%)居多, 占87%, 常见种和恒有种较少, 存在度大于50%的物种有石生针茅、银灰旋花(Convolvulus ammannii)、无芒隐子草(Cleistogenes songorica)、细叶韭(Allium tenuissimum); 物种生活型以地面芽植物和一年生植物居多, 分别占55%和20%; 水分生态类型以典型旱生植物居多, 占47%, 其次为强旱生植物和中旱生植物; 区系地理成分以亚洲中部和东古北极成分为多, 分别占37%和26%; 根据生活型和物种的优势度, 石生针茅群系可划分为6个群丛组、29个群丛。
Study on the relationship between vegetation community characteristics and soil moisture in Badain Jaran Desert
巴丹吉林沙漠植被群落特征与土壤水分关系研究
Changes of non-structural carbohydrates in Caryopteris mongolica seedlings during the process of drought- induced mortality
蒙古莸幼苗干旱致死过程中非结构性碳水化合物的变化
DOI:10.13287/j.1001-9332.201908.005
[本文引用: 1]
以一年生蒙古莸幼苗为对象,设置适宜水分、慢速干旱致死和快速干旱致死3个处理,研究不同干旱强度致死下蒙古莸幼苗各器官中非结构性碳水化合物(NSC,包括可溶性糖和淀粉)的含量变化及其分配规律.结果表明: 慢速干旱致死胁迫下各器官可溶性糖含量与适宜水分组无显著差异.随时间的推移,茎可溶性糖含量先增加后减少,淀粉和NSC含量增加;粗根可溶性糖含量减少,淀粉和NSC含量增加;叶可溶性糖含量增加,淀粉和NSC含量减少.致死时(80 d),叶、茎、粗根和细根的NSC含量分别为6.2%、7.8%、8.3%和7.4%.快速干旱致死胁迫下,各器官可溶性糖含量均高于适宜水分处理组,而淀粉和NSC含量均低于适宜水分组.随时间的推移,根可溶性糖含量下降,淀粉和NSC含量上升;茎可溶性糖、淀粉和NSC含量均上升;叶可溶性糖含量上升,淀粉和NSC含量下降.致死时(30 d),叶、茎、粗根和细根的NSC含量分别为5.9%、6.6%、8.9%和7.7%.应对不同的干旱致死情况,蒙古莸幼苗各器官间非结构性碳水化合物呈现出不同的动态变化.在慢速干旱致死胁迫下,NSC优先为维持各器官生理代谢活动提供能量;而在快速干旱致死下,NSC主要以可溶性糖形式维持植物代谢,调节渗透势,促进吸水,应对急剧的干旱胁迫.
Interspecific association of dominant herb species in ridge vegetation community in the loess area
黄土区土坎植被群落草本优势种的种间关联性研究
Potential geographical distribution and the dominant climatic factors of three common Artemisia species in China
中国三种常见蒿属植物潜在地理分布及其主导气候因子
Community structure and species diversity of desert plants in the wind-sand area of Yabulai
阿拉善雅布赖风沙区荒漠植物群落结构和物种多样性研究
Distribution and species diversity patterns of Vitex negundo var. heterophylla shrublands in north China
DOI:10.17521/cjpe.2018.0162
[本文引用: 1]
Aims To explore the factors controlling the spatial distribution, as well as the species diversity, of the Vitex negundo var. heterophylla shrublands in North China.Methods We investigated the species composition and environments for 370 shrubland plots of Vitex negundo var. heterophylla shrublands in North China. We calculated biodiversity indices (Shannon-Wiener index, Simpson index and Pielou’s evenness index) and then applied multivariate regression trees (MRT), analysis of variance (ANOVA), Pearson correlation analysis to disentangle distribution of the Vitex negundo var. heterophylla community and patterns of species diversity along both horizontal and elevational gradients.Important findings The results showed that Vitex negundo var. heterophylla shrublands distributed in a large area stretching for more than 10° of both latitude and longitude, together with an elevational range of more than 1 000 m (117 m to 1 248 m). The Shannon-Wiener index, Simpson index and Pielou’s evenness index were higher in the herbaceous layer than those in the shrub layers. The species diversity of shrub layer increased with latitude but decreased with elevation. In contrast, that of the herbaceous layer decreased with latitude but increased with elevation. The Shannon-Wiener indexes of both layers were significantly correlated with environmental factors. The results of Pearson correlation analysis indicated that the Shannon-Wiener indexes of shrub layer had a significantly negative relationship with its temperature and slope position, and a significantly positive relationship with its precipitation. However, that of herb layer showed opposite trends.
华北地区荆条灌丛分布及物种多样性空间分异规律
DOI:10.17521/cjpe.2018.0162
[本文引用: 1]
生境差异是影响植物群落及其物种多样性分布格局的重要因素。该研究基于370个荆条(Vitex negundo var. heterophylla)灌丛的野外调查样方, 结合多元回归树(MRT)、显著性分析、Pearson相关分析等技术手段, 分析了华北地区荆条灌丛及其物种多样性的空间分布格局。结果表明: 荆条灌丛的经纬度均跨越10°以上, 可分为高、中、低纬度分布区; 分布海拔则从117 m至1 248 m跨越了1 000 m以上, 也可分为高、中、低海拔分布区。所有分布区中荆条灌丛草本层的Shannon-Wiener指数、Simpson指数和均匀度指数均高于灌木层。灌木层物种多样性随着纬度的降低而降低, 随着海拔的升高而降低; 草本层相反, 物种多样性随着纬度降低而升高, 随着海拔升高而升高。灌木层和草本层的物种丰富度(Shannon-Wiener指数)与水、热、地形等环境因子显著相关, 其中灌木层物种多样性随着气温、坡位升高而显著减少, 随着降水的增加而显著增加;草本层则相反。
Preliminary study on the Compositae flora in the Loess Plateau
黄土高原菊科植物区系初步研究
Contents and protocols for the classification and description of Vegetation Formations, Alliances and Associations of vegetation of China
DOI:10.17521/cjpe.2019.0272
[本文引用: 2]
Vegegraphy, a compound word of prefix “vege-” of “vegetation” and suffix of “-graphy” (description), is a series of monographs that describe species composition, structures, functions, distribution and environmental settings of a set of plant communities and/or their combinations. The Vegegraphy of China, which will be composed of 48 volumes and about 110 issues, is the first version to describe the medium-level units (Alliance Group, Alliance) and the lower-level units (Association Group, Association), and summarizes the higher-level units (Vegetation Formation Group, Vegetation Formation, Vegetation Subformation) in the Chinese Vegetation Classification System. The description of the higher-level units is highly generalized, mainly based on the data and evidences from the classification and description of the medium- and the lower-level classification units of the Chinese vegetation classification system, focusing on the geographic distribution, natural environment, community physiognomy, vegetation type and diversity, dominant species, values for biodiversity conservation, and the status of current vegetation resources at the scales of both worldwide and China. It is an important revision and expansion of the relevant content of Vegetation of China (1980) and will be the most authoritative and accurate description of the basic characteristics of Chinese vegetation. Alliance Group in each issue is briefly described on geographical distribution, natural environment, vegetation types, significance and conservation etc. Alliance as a key medium- level unit is to be described on several aspects, i.e., geographical distribution, natural environment, ecological characteristics, vegetation composition, vegetation structure, vegetation types and characteristics, biological characteristics of dominant species, biomass and productivity, vegetation dynamics and succession, and significance and conservation. As a core content of the above mentioned aspects, vegetation classification and description are conducted under the guidance of the “80 scheme” of Vegetation of China, using vegetation survey data as credentials to quantitatively differentiate vegetation types accordingly. Specifically, a vegetation classification scheme is determined based on supraterraneous stratification, and a set of diagnostic species while vegetation descriptions focus on physiognomy, community structure, species composition, including quantitative features of dominant species, companion species, constant species and accidental species, as well as their environmental preferences. In this study, we first put forward a code of vegetation nomenclature for the medium-level units (Alliance Group, Alliance) and the lower-level units (Association Group, Association) in the Chinese Vegetation Classification System. According to this vegetation nomenclature, the name of a vegetation type is composed of the name of a diagnostic species (may also be a dominant species) of each layer of the vegetation type and the name of the higher-level units (Vegetation Formation or Vegetation Formation Group) to which it belongs, which guarantees both scientific soundness and practical sense of the vegetation nomenclature. Contents and protocols for the research and editing of “Vegegraphy of China”, including vegetation survey and sample collection, literature compilation, climate, soil and topography data collection and compilation, vegetation classification, vegetation nomenclature, vegetation description outlines, are elaborated or exemplified in detail.
《中国植被志》研编内容与规范
DOI:10.17521/cjpe.2019.0272
[本文引用: 2]
植被志是基于植被(或植物群落)调查资料, 全面记叙植被的外貌、物种组成、结构和功能, 以及地理分布和生境条件等特征, 并对同类植被进行归纳和总结的志书。《中国植被志》是第一部对中国植被进行全面记述的志书, 预计完成约48卷110册。在借鉴《中国植被》(1980)(简称“80方案”)植被分类基本原则的基础上,《中国植被志》将对中国植被分类系统的高级分类单位(植被型组、植被型和植被亚型)进行归纳和总结, 对中级和低级分类单位(群系组、群系、亚群系, 群丛组、群丛)进行详细描述。植被高级分类单位的描述具有概括性质, 是在中国植被分类系统中级和低级分类单位描述的基础上, 对其在全球和中国境内的地理分布、自然环境、群落外貌、植被类型及多样性、优势种或共优势种、生物多样性保育价值以及资源现状等进行概述, 并对“80方案”相关内容进行修订和拓展, 将提供对中国植被基本特征客观、准确的记述。在植被中级和低级分类单位中, 群系组描述的内容包括地理分布、自然环境、群落外貌、植被类型以及价值与保育等内容; 群系描述的主要内容包括地理分布、自然环境、生态特征、物种组成、群落结构、群丛组和群丛的分类与描述、优势种的生物学特性、生物量与生产力、植被动态与演替以及价值与保育等方面。作为植被志研编的核心内容, 群丛组和群丛的分类与描述主要基于植被调查资料, 采用数量分类方法, 根据群落结构和物种组成的差异划分出不同的植被类型, 并对其基本特征进行定量描述和归纳。其中, 群落的层片结构、特定植被分类单元的特征种或特征种组的筛选与甄别是植被类型划分的关键环节; 而群落外貌, 群落结构, 物种组成, 各类物种的生长习性、生境的偏适性等是群丛组和群丛描述与归纳的重点内容。该文提出了中国植被中级和低级分类单位的命名方案, 其特点在于植被类型的科学名称中同时体现了植被分类单元特征种或优势种的名称及其所属的高级植被分类单位(植被型组或植被型)的名称, 兼顾了植被名称的规范性与实用性。《中国植被志》的研编工作由文献整编、群落调查、数据分析与整理、文本撰写等环节组成。该文对植被样方的调查与收集, 文献收集与整编, 气候、土壤、地形等相关数据的来源及其整理方法, 植被分类方法, 植被命名, 植被分类单元描述的内容, 植被志章节编写大纲、体裁及撰写等多个规范进行了详细的阐述或示例。
Spatial-temporal patterns of vegetation water use efficiency in the Mu Us Desert
DOI:10.7522/j.issn.1000-694X.2020.00005
[本文引用: 1]
Water use efficiency (WUE) is a key proxy for linking carbon and water exchange processes in terrestrial ecosystems. Understanding the spatial-temporal variations in WUE helps to evaluate the adaptability of vegetation to local environmental conditions. This study estimated the vegetation WUE in the Mu Us Desert during 2001-2016 using remote-sensing data and meteorological observations. Carbon sequestration (i.e., net primary productivity, NPP) was estimated based on the Carnegie-Ames-Stanford Approach (CASA) model, while water consumption (i.e., evapotranspiration, ET) was obtained from the MODIS product. The spatial-temporal patterns of WUE and their responses to meteorological factors were investigated by trend analysis and correlation analysis, respectively. The results showed that: (1) the mean annual WUE across the Mu Us Desert was 0.62 gC·mm-1·m-2 with a trend of decrease from the east to the west, and among different vegetation types, the values of WUE ranked in the order of cultivated vegetation>shrub>meadow>steppe>desert; (2) WUE increased at the rate of 0.009 gC·mm-1·m-2·yr-1 during the study period, especially significantly in eastern, and the increase in WUE mainly attributed to the rapid rise in NPP; and (3) the relationships between WUE and meteorological factors followed a sequence of solar radiation, precipitation, wind speed, and air temperature from strong to weak, and there were relatively insignificant relationships between WUE and meteorological factors in the western part of the Mu Us Desert. The study highlight that the revegetation programs implemented in the Mu Us Desert have achieved good outputs that both NPP and WUE have increased since 2001, which contributed to the improvement of local ecological environment; however, in the central and western part of the Mu Us Desert with poor condition of water and heat, the mismatch between revegetation magnitude and water should be paid more attention.
毛乌素沙地植被水分利用效率的时空格局
DOI:10.7522/j.issn.1000-694X.2020.00005
[本文引用: 1]
水分利用效率(WUE)是衡量生态系统碳水循环耦合程度的重要指标,明晰其动态变化规律有助于判断区域植被生态建设是否与当地的自然条件相适应。利用基于CASA(Carnegie-Ames-Stanford Approach)模型得到的植被净初级生产力(NPP)数据和MODIS系列产品中的蒸散发(ET)数据,估算了2001—2016年毛乌素沙地植被的WUE,并探讨了其时空演变特征及与气象因子的关系。结果显示:(1)WUE的多年平均值为0.62 gC·mm<sup>-1</sup>·m<sup>-2</sup>,空间上自东向西递减,在不同植被类型间表现为栽培植物>灌丛>草甸>草原>荒漠;(2)WUE以0.009 gC·mm<sup>-1</sup>·m<sup>-2</sup>·a<sup>-1</sup>的速率上升,东部地区变化尤为显著,NPP的快速增长是WUE呈增加趋势的主要原因;(3)WUE与各气象因子的相关程度由大至小依次为太阳辐射、降水量、风速和气温,分布在西部地区的荒漠植被WUE受气象因子影响最小。经过一系列生态建设工程的治理,毛乌素沙地大部分地区自2001年以来NPP和WUE均呈增加趋势,即生态系统在变“绿”的同时还实现了水分的高效利用,有利于当地生态环境的健康发展;但在水热条件较差的中西部地区,新建设的植被生长缓慢且导致蒸散发产生强烈变化,使WUE呈减少趋势,暴露出植被建设规模与水资源承载能力不匹配的问题,需引起格外重视。
Establishment and optimization of tissue culture and rapid propagation system in Caryopteris mongholica
蒙古莸(Caryopteris mongholica)组培快繁体系的建立和优化
Study on the biological and ecological characteristics and economic value of the ecological shrub of Caryopteris mongolica
生态灌木蒙古莸的生物生态学特性及其经济价值评价
Multiscale spatial patterns of species diversity and biomass together with their correlations along geographical gradients in subalpine meadows
Composition of seed plant species and flora features in different areas of the Kumtag Desert of China
库姆塔格沙漠不同区域种子植物物种组成与区系特征研究
Studies on the chemical constituents of Caryopteris mongholica
蒙古莸的化学成分研究
Distribution, community characteristics and classification of Stipa tianschanica var. gobica steppe in China
DOI:10.17521/cjpe.2017.0314 URL [本文引用: 1]
中国戈壁针茅草原的分布、群落特征和分类
DOI:10.17521/cjpe.2017.0314
[本文引用: 1]
戈壁针茅(Stipa tianschanica var. gobica)草原主要分布于荒漠草原区的石质丘陵或石质山坡, 向东可以分布在典型草原区的石质丘陵或山地, 向西也可以分布在荒漠区的石质山坡上。该研究调查了中国戈壁针茅草原的主要植被类型, 通过对115个样地的样方数据分析, 量化描述了这一草原类型的主要植物群落特征。结果表明, 中国戈壁针茅草原共有维管植物272种, 隶属于38科127属; 其中裸子植物3种, 隶属于1科1属; 被子植物269种, 隶属于37科126属, 物种数大于等于10种的科为: 菊科、禾本科、豆科、蔷薇科、百合科、藜科、石竹科; 戈壁针茅草原植物区系的生活型谱以地面芽植物为主, 共178种植物, 占总种数的65.44%; 水分生态类型以旱生植物为主, 共120种, 占总种数的44.12%; 植物区系地理成分上以东古北极分布种和亚洲中部分布种(含其变型)最多, 分别为82种和80种, 占总种数的30.14%和29.41%; 盖度等级划分上以0.1%-1.0%的盖度最多, 共174种, 占总种数的63.97%; 恒有度等级划分上以I级(0-20%)的物种数最多, 共258种植物, 占所有植物种数的94.85%。基于群落调查数据, 按照群落-外貌分类原则, 将戈壁针茅群系划分为8个群丛组106个群丛。

{kind=link}
{kind=link}
{kind=link}
{kind=link}