


大多数有花植物依赖动物传粉繁育后代, 为了与不同类群的传粉者相适应, 植物的花形成了各种特殊的性状, 这些性状体现在花的颜色、大小、结构等方面。其中, 有一类特殊的花部结构——蜜距(nectar spur), 因为增加了花报酬(花蜜)和生殖结构(雌蕊和雄蕊)之间的距离, 潜在影响了传粉动物的访花效率(Hodges et al., 2004)。通过长度和形状的演变, 蜜距帮助植物适应新的传粉类群, 占据新的传粉生态位, 进而造成生殖隔离, 促进物种形成(Fulton & Hodges, 1999; Shivanna, 2014; Minelli, 2016)。因此蜜距被认为是被子植物进化进程中的一种“关键创新性状” (key innovation trait; Fernández-Mazuecos et al., 2019)。特别是通过对耧斗菜属(Aquilegia)的长期研究, 蜜距已经被认为是植物关键创新性状教科书式的例子(Hodges & Arnold, 1995; Whittall & Hodges, 2007; Puzey et al., 2012), 是研究被子植物平行/趋同进化的理想材料(Box, 2010)。
从形态上看, 蜜距是被子植物的花瓣、萼片等花部器官上的管状或囊状突起(Cullen, 2020)。在大部分有距植物中, 蜜距是一种非分泌性结构, 其作用仅仅是储存花蜜; 花蜜从花的其他地方分泌出来, 汇聚在蜜距中(Box, 2010), 而在一些植物中, 蜜距已经进化成既储存花蜜, 也能分泌花蜜的结构(Stpiczyńska & Matusiewicz, 2001; Box et al., 2008; Bell et al., 2009)。从系统发育方面看, 蜜距不存在于被子植物基部类群中(Box, 2010), 而是衍生于单子叶植物(如兰科、百合科)和真双子叶植物(如毛茛科、车前科和狸藻科)中(Rudall et al., 2003; Endress & Matthews, 2006), 经历了多次独立进化(Hodges, 1997)。在不同科中, 蜜距的发育来源、长度、形状、方向、颜色、弯曲情况和附属物存在差异, 表现出很高的多样性(Hodges & Arnold, 1995; Hodges, 1997), 并且蜜距也是很多植物的分类依据(Koopman & Ayers, 2005), 但有关被子植物蜜距类型的多样性、生态学适应意义和研究现状还缺乏针对性的总结和归纳。系统地总结蜜距植物及其形态特征和传粉/盗蜜者, 将有助于全面了解这一性状在被子植物中的系统发育分布和进化生态学意义。
本文对国内外蜜距相关的数据库和文献进行了梳理, 总结了蜜距的主要来源及其系统分布、蜜距的形态多样性及其生态学适应意义, 统计了不同科植物蜜距长度的差异, 介绍了蜜距植物的主要传粉者和盗蜜者类群, 以及调节蜜距长度的进化机制, 并提出未来需要解决的科学问题。
1 数据来源和分析方法
1.1 数据来源
1.1.1 数据库
中国植物使用“植物智” (
1.1.2 文献
在“谷歌学术” (
1.2 分析方法
从数据库和文献中获得的距长信息中提取可用于统计分析的数据。具体的处理方式是对于单个数据的, 如约6 mm则取值6 mm; 对于范围数据, 如3-4 mm取最大值即4 mm。使用R 4.2.2软件对17个科和不同类型的蜜距分别进行单因素方差分析并计算变异系数(标准差/平均值)。将变异系数与传粉者属数、标准差、平均值、数据量(详细数据见附录III)进行皮尔逊相关分析。
2 蜜距的类型
根据蜜距发育来源的不同, 可将蜜距分为6大类: 花瓣蜜距(petal spur)、花冠蜜距(corolla spur)、萼片蜜距(sepal spur)、花萼蜜距(calyx spur)、花被片蜜距(tepal spur)以及被丝托蜜距(hypanthium spur) (图1)。
图1

图1
被子植物蜜距的多样性。A, 二叶舌唇兰(兰科)。B, 木鱼坪淫羊藿(小檗科)。C, 中甸乌头(毛茛科)。D, 华北耧斗菜(毛茛科)。E, 早开堇菜(堇菜科)。F, 囊距紫堇(罂粟科)。G, 大花花锚(龙胆科)。H, 摩洛哥柳穿鱼(车前科)。I, 黄花狸藻(狸藻科)。J, 凤仙花属(凤仙花科)。K, 油点草(百合科)。L, 旱金莲(旱金莲科)。其中A-F为花瓣蜜距, G-I为花冠蜜距, J为萼片蜜距, K为花被片蜜距, L为被丝托蜜距。白色箭头所指为各种形态的蜜距。拍摄者: 金晓芳。
Fig. 1
Diversity of nectar spurs in angiosperms. A, Platanthera chlorantha (Orchidaceae). B, Epimedium franchetii (Berberidaceae). C, Aconitum piepunense (Ranunculaceae). D, Aquilegia yabeana (Ranunculaceae). E, Viola prionantha (Violaceae). F, Corydalis benecincta (Papaveraceae). G, Halenia elliptica var. grandiflora (Gentianaceae). H, Linaria maroccana (Plantaginaceae). I, Utricularia aurea (Lentibulariaceae). J, Impatiens sp. (Balsaminaceae). K, Tricyrtis macropoda (Liliaceae). L, Tropaeolum majus (Tropaeolaceae). A-F are petal spurs, G-I are corolla spurs, J is sepal spur, K is tepal spur, and L is hypanthium spur. The white arrows indicate the various forms of the nectar spurs. Photoed by JIN Xiao-Fang.
2.1 花被片蜜距
表1 被子植物蜜距的发育来源及其系统分布
Table 1
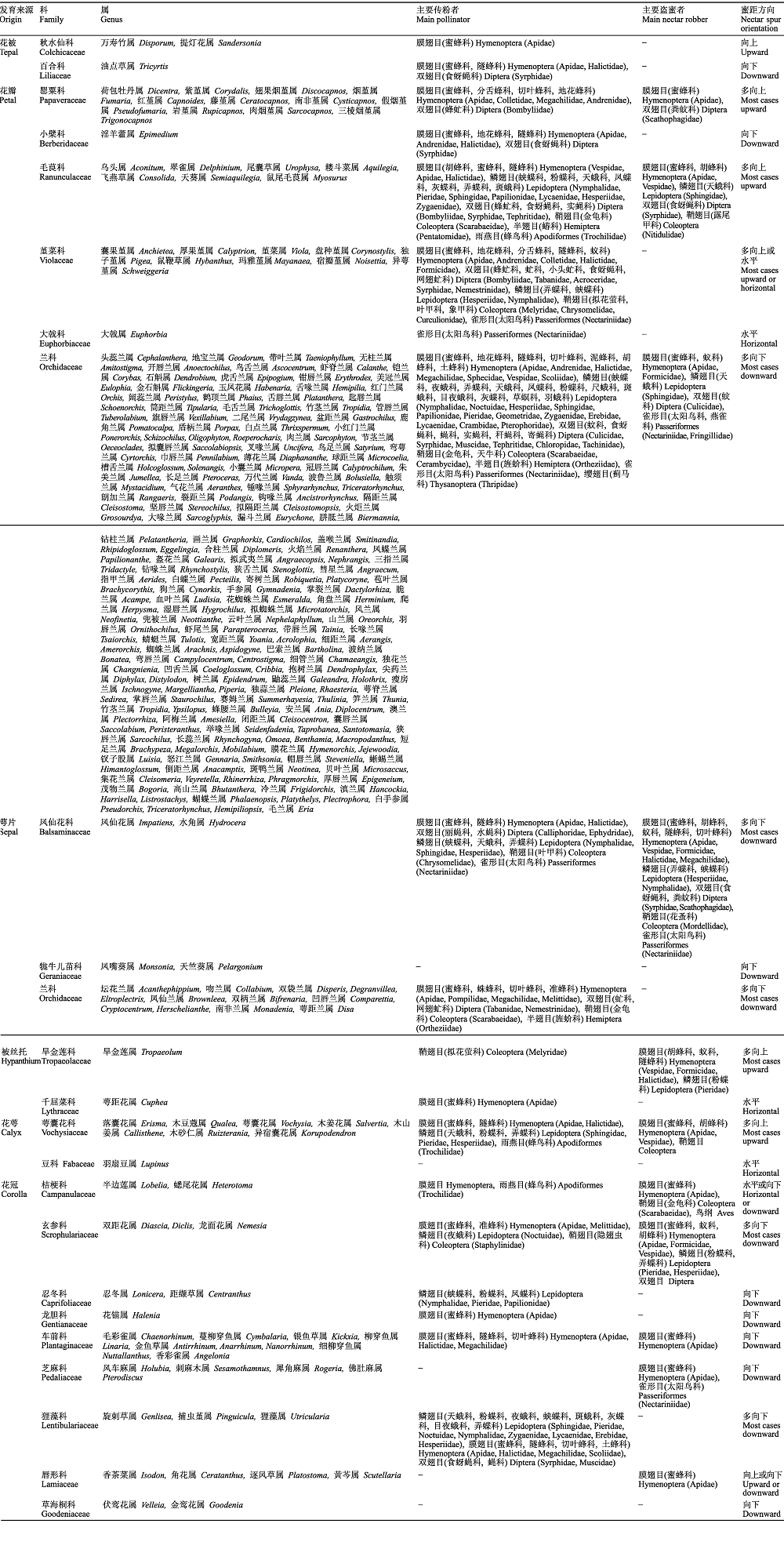 |
-,未调查到传粉数据。
-,no pollination data was investigated.
2.2 花瓣蜜距
花瓣蜜距是指由离瓣花花瓣发育而来的蜜距, 是最常见的蜜距类型。主要存在于罂粟科、小檗科、毛茛科、堇菜科和大戟科的29个属, 以及兰科的183个属(表1)。
毛茛科中, 耧斗菜属(图1D)花瓣下方常向下延长形成蜜距, 极少数为囊状或不明显(Hodges, 1997), 如无距耧斗菜(Aquilegia ecalcarata); 乌头属(Aconitum) (图1C)、翠雀属与飞燕草属有2枚花瓣卷缩形成管状的蜜距(蜜腺位于蜜距末端), 并由1枚萼片包裹(图2A、2B) (Jabbour & Renner, 2012; Antoń & Kamińska, 2015), 乌头属少数种无距, 如空茎乌头(A. apetalum)等; 尾囊草属(Urophysa)的距瓣尾囊草(U. rockii)花瓣有短距, 而尾囊草(U. henryi)花瓣无距(Zhao et al., 2016)。
图2

图2
花瓣卷缩形成的蜜距(翠雀属、乌头属)和被丝托蜜距的结构解剖。A, 中甸乌头的花瓣蜜距。B, 中甸翠雀花的花瓣蜜距(被包裹在萼距里面)。C, 旱金莲的被丝托蜜距。NS, 蜜距; P, 花瓣; S, 萼片。拍摄者: 金晓芳。
Fig. 2
Nectar spur formed by petal rolling (Aconitum, Delphinium) and hypanthium spur. A, Petal spur of A. piepunense. B, Petal spur of D. yuanum (encased in calyx spur). C, Hypanthium spur of Tropaeolum majus. NS, nectar spur; P, petal; S, sepal. Photoed by JIN Xiao-Fang.
兰科植物有距类群分布广泛, 多由花瓣发育而来, 部分属具明显的长距, 如虾脊兰属(Calanthe)、彗星兰属(Angraecum)等(Whittall & Hodges, 2007; Suetsugu & Fukushima, 2014), 部分属的蜜距呈兜状或囊状, 如盆距兰属(Gastrochilus)、鹿角兰属(Pomatocalpa)等(Watthana, 2007; Nguyen et al., 2022), 还有部分属的蜜距不明显或仅形成小突起, 如地宝兰属(Geodorum)、带叶兰属(Taeniophyllum)等(Chowlu et al., 2012; Teoh, 2022)。
2.3 萼片蜜距
萼片蜜距是指来源于萼片的蜜距, 存在于萼片离生的凤仙花科、牻牛儿苗科的4个属, 以及兰科的12个属(表1)。
大部分兰科蜜距属于花瓣蜜距, 目前仅调查到12个属的蜜距来源于萼片, 分布于兰科系统发育树的几个分支(Hobbhahn et al., 2013; Li et al., 2016)。其中Degranvillea、Eltroplectris、双柄兰属(Bifrenaria)和凹唇兰属(Comparettia)主要分布在南美洲, 而凤仙兰属(Brownleea)分布于非洲(Szlachetko, 1995; Koehler et al., 2002; Pansarin et al., 2015)。值得注意的是分布于非洲的萼距兰属(Disa), 大部分有距植物的蜜腺位于蜜距内, 但D. longicornu的蜜腺位于花瓣上, 花蜜通过延伸的花瓣流入蜜距。
2.4 被丝托蜜距
2.5 花萼蜜距
2.6 花冠蜜距
花冠蜜距是指来源于被子植物合瓣花花冠的蜜距, 包括桔梗科、龙胆科、车前科、玄参科、芝麻科(胡麻科)、狸藻科、唇形科、忍冬科和草海桐科的30个属(表1)。
总体而言, 根据本文调查被子植物约有13个目23个科(参照APG IV系统)具有蜜距, 图3所展示的是本文调查的13个目的蜜距植物所在系统发育树的位置。不同蜜距类型在被子植物系统发育树上反复出现, 表明蜜距在被子植物中经历了多次独立的演化。其中毛茛目和唇形目具有蜜距的科数最多, 分别为3和5个。天门冬目中兰科同时存在花瓣蜜距和萼片蜜距; 桃金娘目同时存在花萼蜜距和被丝托蜜距; 其他目仅存在单一蜜距。另外, 花被片蜜距仅存在于百合目; 花瓣蜜距存在于单子叶植物分支的天门冬目和真双子叶植物分支的毛茛目及金虎尾目; 萼片蜜距存在于单子叶植物分支的天门冬目和超菊类分支的牻牛儿苗目及杜鹃花目; 花萼蜜距均存在于真双子叶植物; 所有花冠蜜距植物均位于真菊类分支(The Angiosperm Phylogeny Group, 2016)。
图3
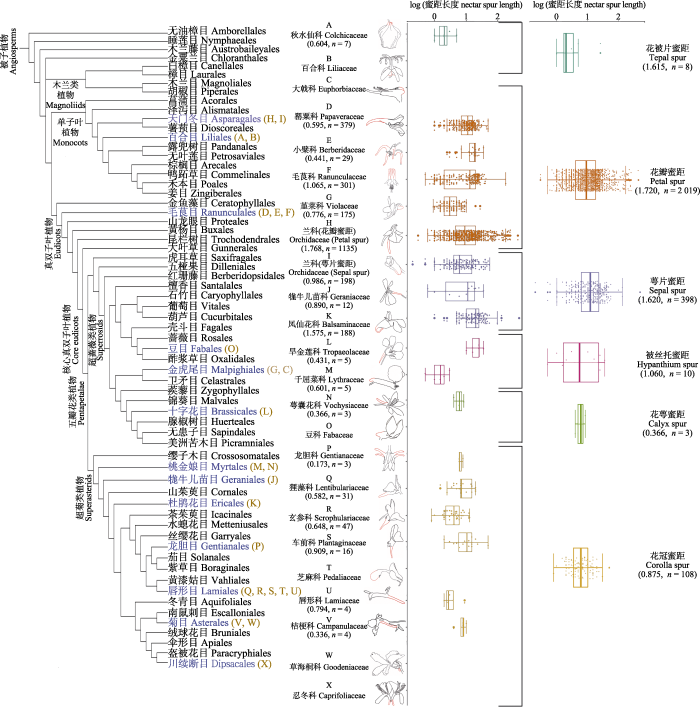
图3
被子植物蜜距在目一级的系统发育分布。系统树根据Angiosperm phylogeny poster (APP)—Flowering plant systematics, 2019绘制(Cole et al., 2019)。线描图红色部分为蜜距所在位置。植物科和蜜距类型所附括号内为距长变异系数和样本量。两列箱型图分别为17个科和6种蜜距类型的蜜距长度log值(平均值±标准差)。
Fig. 3
Distribution of nectar spur types across angiosperm orders. The phylogeny of angiosperm orders is drawn according to the Angiosperm phylogeny poster (APP)—Flowering plant systematics, 2019 (Cole et al., 2019). The red parts of the line diagrams show the location of the nectar spurs. Coefficient of variation of spur length and sample size are enclosed in parentheses for plant families and nectar spur types. The log values (mean ± SD) of spur length for 17 families and 6 types of nectar spur are shown in the two box plots.
3 蜜距的形态多样性
3.1 蜜距的长度
有距植物蜜距长度变化很大, 比如千屈菜科、秋水仙科、龙胆科、桔梗科、萼囊花科、唇形科等的蜜距通常较短, 长度一般不超过10 mm; 而车前科、凤仙花科、堇菜科、兰科、狸藻科、牻牛儿苗科、毛茛科、小檗科、玄参科、罂粟科等的蜜距则有长有短, 差异很大, 其中部分物种蜜距极长, 如凤仙花科的Impatiens simbiniensis和兰科的Angraecum sesquipedale距长分别可达到48 cm和43 cm (Wasserthal, 1997)。
本文提取了2 546种蜜距植物最大距长信息, 排除数据量少于3的科, 剩下17个科2 542种植物进行单因素方差分析。因距长原始数据非正态分布, 因此经log转化后分析, 结果显示不同科之间蜜距长度存在显著性差异(F(17, 2524) = 21.67, p < 0.001) (图3)。其中凤仙花科的平均距长最长, 为23.8 mm; 千屈菜科最短, 为1.6 mm。通过单因素方差分析比较6类型(2 546种)蜜距的距长(经log转化), 结果显示不同类型之间蜜距长度存在显著性差异(F(5, 2540) =8.638, p < 0.001) (图3)。萼片蜜距的平均距长最长, 为17.0 mm; 花被片蜜距最短, 为5.25 mm。
相关分析结果(表2)显示, 距长在“科”水平的变异系数显著与标准差、平均值、数据量和传粉者属数显著正相关, 即科的平均距长越长, 蜜距长度的变异越大; 蜜距的长度变异越大, 传粉者属数越多。
表2 距长变异系数与距长其他特征的相关分析统计结果
Table 2
| 标准差 Standard deviation | 平均值 Average value | 数据量 Amount of data | 传粉者属数 Genus number of pollinators | |||||
|---|---|---|---|---|---|---|---|---|
| r | p | r | p | r | p | r | p | |
| 蜜距类型 Type of nectar spur | 0.781 | 0.067 | 0.563 | 0.245 | 0.542 | 0.266 | 0.413 | 0.416 |
| 科 Family | 0.889 | <0.001 | 0.502 | 0.034 | 0.716 | <0.001 | 0.752 | <0.001 |
| 属 Genus | 0.699 | <0.001 | 0.371 | 0.107 | 0.099 | 0.679 | -0.079 | 0.808 |
皮尔逊相关性(r)和显著性(双尾, p)。
Pearson correlation coefficient (r) and significance (two-tailed, p).
3.2 蜜距尾部弯曲情况
蜜距尾部弯曲也是一个常见特征, 广泛存在于车前科、萼囊花科、凤仙花科、兰科、狸藻科、毛茛科、忍冬科、小檗科、玄参科、罂粟科等多个类群。不同类群植物蜜距的弯曲程度差异很大, 大部分物种的蜜距稍有弯曲, 如柳穿鱼(Linaria vulgaris); 部分物种的蜜距极度弯曲甚至形成环状, 如Impatiens oxyanthera (Wang et al., 2013)。
3.3 蜜距的数量
在有距植物的不同类群中, 蜜距的数量存在差异。绝大多数类群蜜距仅有1个, 少数类群具有多个蜜距, 从2个到6个不等。例如, 兰科铠兰属(Corybas) (Dransfield et al., 1986)和鸟足兰属(Satyrium) (Johnson et al., 2011), 玄参科双距花属(Vogel, 1984), 毛茛科乌头属(Antoń & Kamińska, 2015)和翠雀属(Antoń & Kamińska, 2015), 兰科的双袋兰属(Disperis) (Kurzweil & Manning, 2005)都具有2个蜜距。仅有极少数高度特化的物种具有3个蜜距, 如倒距兰(Anacamptis pyramidalis f. fumeauxiana), 该物种是一种欺骗性、不产生花蜜的兰科物种, 其中一枚蜜距来源于唇瓣, 另外两枚蜜距来源于侧萼片(Kowalkowska et al., 2010, 2012)。具有4个蜜距的植物包括小檗科的淫羊藿属和龙胆科的花锚属(Li et al., 2009)。耧斗菜属具有5枚花瓣, 部分物种每枚花瓣向下延长成蜜距, 共形成5个蜜距(Antoń & Kamińska, 2015)。秋水仙科具有花被片6枚, 部分物种每枚花被片基部形成蜜距, 共形成6个蜜距, 如Disporum cantonniense和Sandersonia aurantiaca (Zou et al., 2001)。
研究表明, 多个蜜距能够诱导传粉者在花上来回运动, 明显增加传粉者访花的时间, 增加花粉的传递效率, 是植物提高繁殖成功率的一种开花策略(Jabbour & Renner, 2012)。
4 蜜距的进化生态学研究概况
4.1 宏观进化
4.1.1 蜜距植物的传粉者和盗蜜者
4.1.1.1 传粉者
有蜜距的被子植物主要依靠长喙的昆虫和鸟类传粉, 其中昆虫和鸟类的喙与蜜距的长度之间的匹配对授粉效率至关重要(Krishna & Keasar, 2018)。这些传粉动物为获得更多的花蜜会将喙尽可能地伸入蜜距中, 其身体会更加充分地接触花粉和柱头, 提高传粉效率(Whittall & Hodges, 2007)。一些膜翅目昆虫具有较长的喙, 使之能够伸入蜜距中吸取花蜜, 是有距植物的重要传粉者(图4A-4F), 比如蜜蜂属(Apis)、熊蜂属(Bombus)、木蜂属(Xylocopa)、胡蜂科等类群(Macior, 1978; Peter & Johnson, 2009; Johnson et al., 2011; Vargas et al., 2017)。鳞翅目昆虫也是有距植物的重要传粉者, 其典型代表为兰科植物(Boberg et al., 2014)。双翅目昆虫常不具备长喙, 难以获取有距植物的花蜜, 很少作为有距植物的主要传粉者(Suetsugu et al., 2015)。然而, 部分双翅目昆虫具有较长的喙, 可以吸食有距植物的花蜜并传粉, 如网翅虻科的Moegistorynchus longirostris可以为兰科植物Disa draconis传粉(Johnson & Steiner, 1997)。 鞘翅目与半翅目昆虫几乎不具备长喙, 虽然可以作为部分短距植物的访花者, 但其传粉效率较低, 因此多数情况下不是有距植物的有效传粉者(Johnson et al., 2007, 2011; Orellana et al., 2008; van der Niet et al., 2010)。表1列出了文献中查阅的部分传粉者,但并不能完全代表所属类群的传粉者全貌, 只能作为参考。
图4

图4
蜜距植物的主要访花者和盗蜜者。A, 南熊蜂访中甸翠雀花。B, 独居蜂访柔毛淫羊藿。C, 条蜂访刻叶紫堇。D, 熊蜂访椭圆叶花锚。E, 熊蜂访顶喙凤仙花。F, 条蜂访摩洛哥柳穿鱼。G, 条蜂盗蜜地锦苗。H, 黄胸木蜂盗蜜凤仙花。拍摄者: 金晓芳。
Fig. 4
Pollinators and nectar robbers of some plants with nectar spurs. A, Bombus secures visiting Delphinium yuanum. B, Solitary bee visiting Epimedium pubescens. C, Anthophoridae bee visiting Corydalis incisa. D, Bombus sp. visiting Halenia elliptica. E, Bombus sp. visiting Impatiens compta. F, Anthophoridae bee visiting Linaria maroccana. G, Anthophoridae bee robbing nectar of Corydalis sheareri. H, Xylocopa appendiculata robbing nectar of Impatiens balsamina. Photoed by JIN Xiao-Fang.
不同传粉者对蜜距的形态和颜色有不同的偏好, 蜜蜂属往往倾向于访问短而弯曲的、蓝紫色的蜜距, 蜂鸟科更加偏好红色或者黄色的、中等长度的直距, 而天蛾科倾向于访问颜色较淡的、极长的直距(Edwards et al., 2022)。
4.1.1.2 盗蜜者
蜜距可能会提高植物的传粉效率, 但是也会面临被盗蜜的风险, 损失了给正当传粉者提供报酬物的量。所谓盗蜜是指一些昆虫、鸟类等访花者通过绕过花开口并在花基部打孔获取花蜜, 而没有或极少给花提供传粉服务的一种特殊访花行为(Inouye, 1980; 张彦文等, 2006)。盗蜜的发生往往是因为盗蜜者喙太短而无法通过正常访花取食到花蜜或者觅食效率太低, 只能采取打孔的方式更加高效地获取花蜜(Irwin & Brody, 1998)。盗蜜现象在开花植物中很常见, 几乎所有具有长管或者蜜距的开花植物都可能成为潜在的盗蜜对象(Irwin et al., 2010)。目前在59个科214种植物中发现盗蜜现象(Irwin & Maloof, 2002), 然而大部分蜜距植物并未产生防止盗蜜的保护结构, 可能是因为盗蜜对植物的繁育并非完全有害。
有距植物的主要盗蜜者为膜翅目昆虫(图4G、4H), 包括熊蜂属、胡蜂科、蜜蜂属、木蜂属、切叶蜂属(Megachile)、无垫蜂属(Amegilla)、芦蜂属(Ceratina)、蚁类科、回条蜂属(Habropoda)等多个类群, 此外, 盗蜜者还有鳞翅目、双翅目、半翅目、鸟纲等(Irwin & Maloof, 2002; Newman & Thomson, 2005; Chapurlat et al., 2015; Li et al., 2021)。其中, 熊蜂属、无垫蜂属、木蜂属等类群的口器可以破坏花组织, 为初级盗蜜者, 而蜜蜂属、鳞翅目等类群只能通过初级传粉者产生的孔洞盗取花蜜, 为次级盗蜜者(Rust, 1979)。此外, 合法的访花者也可能转变成盗蜜者, 这一方面是因为盗蜜可以高效地获取花蜜, 另一方面盗蜜可以更好地与其他访花者竞争花蜜资源(Irwin et al., 2010)。表1中列出了文献中查阅到的部分盗蜜者, 其中也包含了窃蜜者, 即不破坏花组织, 但是窃取花蜜同时不提供传粉服务的访花者。
4.1.2 距长的进化假说
4.1.2.1 距长和喙长的协同进化假说-达尔文机械模型(Darwin’s Mechanistic Model)
早在1862年, 关于Angraecum sesquipedale如何进化出超长蜜距, 达尔文提出了首个协同进化过程的机械模型, 认为A. sesquipedale的蜜距与某种蛾类的喙在长度上存在军备竞赛(Johnson & Anderson, 2010)。当天蛾科昆虫从蜜距底部吸走花蜜时, 它们把喙的基部贴在花的繁殖器官上, 花粉落在天蛾的身体上被其传播给其他A. sesquipedale。因此, 只有那些蜜距比访花天蛾喙更长的A. sesquipedale个体才能够被传粉, 也只有喙更长的天蛾才能吸到更多花蜜。于是, 兰花进化出更长的蜜距来完成繁殖, 天蛾进化出更长的喙来吸到花蜜, 循环往复。当然, 大多数传粉者和植物不会一直增加距长和喙长, 因为协同进化在早期就被持续变化的选择压力阻止了, 例如过长的喙对天蛾飞行的限制阻止了喙的增长(Johnson & Anderson, 2010)。
植物距长和传粉者喙长的协同进化具有地理镶嵌的规律(Steiner & Whitehead, 1988, 1990; Robertson & Wyatt, 1990; 王晓月等, 2019)。所谓地理镶嵌的协同演化理论(geographic mosaic theory of coevolution, GMC)是指由于时间和空间的分隔, 不同群体中相互作用的物种形成了不同的协同进化(Thompson, 1999)。距长和喙长地理镶嵌的例子很多, 如由短喙的天蛾Deilephila porcellus传粉的二叶舌唇兰(Platanthera bifolia)居群的蜜距较短, 而由长喙的天蛾Sphinx ligustri传粉的居群则具有较长的蜜距(Boberg et al., 2014); 再如相较于依赖膜翅目传粉的变色耧斗菜(Aquilegia caerulea)居群, 更加依赖天蛾传粉的居群具有明显更长的蜜距(Miller, 1981)。
4.1.2.2 传粉者转变模型(Pollinator Shift Model)
协同进化并不是长蜜距进化的唯一假说(Johnson & Anderson, 2010), 另一个合理的解释是, 当植物在进化过程中面临不同传粉者转换时, 蜜距的长度也随之改变, 称之为传粉者转变模型, 由Grant和Grant (1965)及Stebbins (1970)的研究发展而来。在该模型中, 传粉者的转变是由当地传粉者动物群的变化引起的, 这可能由传粉动物生存范围的扩大或物种组成变化导致(Hilton & Harder, 2006)。在传粉者转变模型中, 传粉者的喙长被认为是相对固定的, 这可能是由于受到了体型大小的限制或其他植物蜜距长度的选择, 植物通过单方面的进化(one-sided evolution)适应已存在的传粉者喙长(Whittall & Hodges, 2007)。当植物传播到新的环境中, 蜜距长度就会变化以适应新的传粉者的喙长。理论上, 传粉者转变为喙明显较短的类群的可能性较小, 因为传粉者在无法获得回报时会选择避开花朵(Grant & Temeles, 1992; Hodges, 1995)。因此, 该模型预测, 随着时间的推移, 蜜距通常会变得越来越长。
总之, 该假说与协同进化假说主要区别在于: 蜜距长度进化是在向新的传粉者转变的过程中迅速发生的, 而不是在一个物种与传粉者协同进化的过程中逐渐发生(Whittall & Hodges, 2007)。
4.2 微观进化
植物的蜜距在从无到有, 从短到长, 或从有到无等过程中, 宏观层面表现出传粉者的喙长或传粉者类群发生变化等现象; 同时在微观层面也能发现众多与蜜距演化相关的变化并体现了相应的调节机制。
4.2.1 细胞层面的变化
4.2.2 基因层面的调控
对于蜜距长度调控的分子机制, 首先被考虑的是和细胞伸长或分裂相关的基因。例如, 生长素应答因子家族中的AqARF6和AqARF8基因是耧斗菜属花瓣蜜距细胞伸长和蜜腺成熟特别需要的(Zhang et al., 2020), 这些基因可能在不同距长的耧斗菜属植物间存在某种结构或表达上的差异, 或者可能会受到传粉者选择的影响, 从而导致不同耧斗菜属植物蜜距长度的差异。对耧斗菜属植物的研究也指出转录因子AqIBL1可能抑制蜜距一侧的细胞伸长, 导致蜜距弯曲, 但仍需要进一步的实验验证(Edwards et al., 2022)。另外, 关于细胞分裂相关的基因方面, 有研究表明KNOX基因家族通过影响细胞的分裂和生长从而促进柳穿鱼属植物蜜距形成(Box et al., 2011)。
5 展望
目前, 关于蜜距进化生态学的相关研究主要集中于耧斗菜属和兰科某些类群, 这些类群的距长和不同传粉者喙长有明显的一一对应关系(Wasserthal, 1997; Whittall & Hodges, 2007; Fernández-Mazuecos et al., 2019); 对柳穿鱼属、乌头族、凤仙花科和淫羊藿属也有少量研究(Jabbour & Renner, 2012; Vandelook et al., 2019; Guo et al., 2022; Xie et al., 2022), 而其余大部分蜜距植物的蜜距进化都没有得到很好的研究。并且, 关于距长的军备竞赛假说、传粉者转移假说, 以及地理镶嵌情况在很多植物类群的适用性还未可知。特别是关于蜜距演化的分子机制还有很多属于研究热点(Box et al., 2011; Zhang et al., 2020; Edwards et al., 2022)。为此, 我们从宏观和微观两个方面提出未来值得深入研究的科学问题。
宏观方面: (1)盗蜜行为是否引起蜜距的形态改变? 植物是否产生了某些阻止或减少被盗蜜的机制? (2)蜜距在亲缘关系近的类群间存在着特征的变异和分化, 这些变化的选择压力是否来自传粉类群的差异?
微观方面: (1)蜜距的伸长主要来源于细胞的伸长、分裂, 是否由来自传粉者的刺激导致? 还是完全来自自然选择, 如来自于繁殖成功方面的影响? (2)蜜距的各个特征, 特别是距长变化的分子机制是什么? 相关基因有哪些? (3)来自同一器官, 如花瓣的蜜距是否具有更相似的起源和进化机制? (4)蜜距在被子植物中多次起源, 在不同目、科、属之间, 起源和平行进化的分子机制有何不同?
致谢
感谢中国科学院武汉植物园的叶忠铭和魏能在数据收集和文章构思中给予的帮助。
附录I 有蜜距植物名录及其文献来源
Supplement I Catalogue of plants with nectar spur and its documentary sources
附录II 蜜距植物的传粉者和盗蜜者类群
Supplement II Pollinators and nectar robbers of plants with nectar spur
附录III 距长的变异系数及相关性计算数据
Supplement III Calculation data for coefficients of variation and correlation of nectar spur length
参考文献
The inheritance of flower doubleness and nectary spur in Pelargonium × hortorum Bailey
DOI:10.1007/BF00022407 URL [本文引用: 1]
Comparative floral spur anatomy and nectar secretion in four representatives of Ranunculaceae
DOI:10.1007/s00709-015-0794-5
PMID:25772682
[本文引用: 4]

Nectaries are common in Ranunculaceae. These secretory structures, however, have not been studied in detail despite their importance in plant-animal interactions, and data relating to the structure of nectary spurs, which are so characteristic of several genera of this family, remain scarce. In order to redress this imbalance, we sought, in the present paper, to analyze the anatomical and ultrastructural organization of the nectary spurs of four representatives of Ranunculaceae, i.e., Aconitum lycoctonum L., Aquilegia vulgaris L., Consolida regalis Gray, and Delphinium elatum L. Nectary spurs were examined using light, fluorescence, scanning electron, and transmission electron microscopy. The floral nectaries of A. lycoctonum and A. vulgaris are situated at the apices of the spurs, whereas in C. regalis and D. elatum, the nectary is located along the floor surface of the spurs. Nectar in C. regalis and D. elatum is exuded through micro-channels in the cuticle, whereas in A. lycoctonum and A. vulgaris, it is released by means of cell wall disruption, indicating that the method of nectar secretion here is holocrine. Structurally, the nectary of all four investigated species is quite similar, and its cells are typical of nectar-producing cells described in the literature. It is proposed that in A. lycoctonum and A. vulgaris, disruption of the cell wall and the release of the entire cell contents into the spur cavity contribute to the composition of the nectar that the latter contains, enriching it with cytoplasmic components. We conclude that the manner of nectar exudation may vary considerably between closely related plant species, regardless of their geographical origin and phylogeny.
Four species from western Mexico new to Lobelia (Campanulaceae: Lobelioideae)
DOI:10.2307/2807317 URL [本文引用: 2]
Systematics of Heterotoma (Campanulaceae) and the evolution of nectar spurs in the New World Lobelioideae
DOI:10.2307/2419185 URL [本文引用: 1]
Phylogenetic context, generic affinities and evolutionary origin of the enigmatic Balkan orchid Gymnadenia frivaldii Hampe ex Griseb
DOI:10.2307/25065532
URL
[本文引用: 1]
Although originally ascribed to the genus Gymnadenia R. Br. (Orchidinae: Orchidaceae), the Balkan endemic orchid G. frivaldii Hampe ex Griseb. has since been more frequently assigned to Pseudorchis Séguier (syn. Leucorchis E. Mey., Bicchia Parl.). Molecular phylogenetic analysis using the ITS region of rDNA reveals a large disparity between the two genera and demonstrates that frivaldii is embedded well within Gymnadenia s.s. Macromorphological and SEM studies further elucidate the floral and vegetative similarities between G. frivaldii and Pseudorchis, notably the heterochronically reduced gynostemium and small, short‐spurred labellum; these similarities represent convergent evolutionary transitions, whereas other characters such as contrasting stigma and tuber morphologies provide stronger phylogenetic signals. The sequence‐based phylogeny suggests that G. frivaldii represents one of three cases of independent paedomorphic floral reduction inferred in the genus; simplification has been more severe than in G. odoratissima but less severe than in the closely related Gymnadenia subgenus Nigritella. Alternatively, an effectively instantaneous evolutionary origin through hybridisation with a (most likely diploid) species of subgenus Nigritella remains possible. Reports of rare hybridisation between G. frivaldii and members of subgenus Nigritella are acceptably well documented, whereas reports of hybridisation with several other more phylogenetically distant orchid species (including the often sympatric Pseudorchis albida) are considered less secure.
Is spur length of Platanthera species in the British Isles adaptively optimized or an evolutionary red herring?
Comparative micromorphology of nectariferous and nectarless labellar spurs in selected clades of subtribe Orchidinae (Orchidaceae)
DOI:10.1111/boj.2009.160.issue-4 URL [本文引用: 1]
A new species of Viola (Violaceae) from south-west Turkey
DOI:10.1111/j.1095-8339.2004.00294.x URL [本文引用: 1]
Pollinator shifts and the evolution of spur length in the moth-pollinated orchid Platanthera bifolia
DOI:10.1093/aob/mct217
PMID:24169591
[本文引用: 2]
Plant-pollinator interactions are thought to have shaped much of floral evolution. Yet the relative importance of pollinator shifts and coevolutionary interactions for among-population variation in floral traits in animal-pollinated species is poorly known. This study examined the adaptive significance of spur length in the moth-pollinated orchid Platanthera bifolia.Geographical variation in the length of the floral spur of P. bifolia was documented in relation to variation in the pollinator fauna across Scandinavia, and a reciprocal translocation experiment was conducted in south-east Sweden between a long-spurred woodland population and a short-spurred grassland population.Spur length and pollinator fauna varied among regions and habitats, and spur length was positively correlated with the proboscis length of local pollinators. In the reciprocal translocation experiment, long-spurred woodland plants had higher pollination success than short-spurred grassland plants at the woodland site, while no significant difference was observed at the grassland site.The results are consistent with the hypothesis that optimal floral phenotype varies with the morphology of the local pollinators, and that the evolution of spur length in P. bifolia has been largely driven by pollinator shifts.
Plant- pollinator specialization: origin and measurement of curvature
DOI:10.1086/717677 URL [本文引用: 1]
Floral ontogenetic evidence of repeated speciation via paedomorphosis in subtribe Orchidinae (Orchidaceae)
DOI:10.1111/boj.2008.157.issue-3 URL [本文引用: 2]
Characterization of Linaria KNOX genes suggests a role in petal-spur development
DOI:10.1111/tpj.2011.68.issue-4 URL [本文引用: 3]
Notes on African Pedaliaceae
DOI:10.2307/4115530 URL [本文引用: 1]
The response of floral traits associated with pollinator attraction to environmental changes expected under anthropogenic climate change in high-altitude habitats
DOI:10.1086/705591 URL [本文引用: 1]
Appendicular origin and structure of the spur of Vochysiaceae flowers
DOI:10.1590/0102-33062017abb0117 URL [本文引用: 1]
Spatial variation in pollinator-mediated selection on phenology, floral display and spur length in the orchid Gymnadenia conopsea
DOI:10.1111/nph.13555
PMID:26183369
[本文引用: 1]
Spatial variation in plant-pollinator interactions may cause variation in pollinator-mediated selection on floral traits, but to establish this link conclusively experimental studies are needed. We quantified pollinator-mediated selection on flowering phenology and morphology in four populations of the fragrant orchid Gymnadenia conopsea, and compared selection mediated by diurnal and nocturnal pollinators in two of the populations. Variation in pollinator-mediated selection explained most of the among-population variation in the strength of directional and correlational selection. Pollinators mediated correlational selection on pairs of display traits, and on one display trait and spur length, a trait affecting pollination efficiency. Only nocturnal pollinators selected for longer spurs, and mediated stronger selection on the number of flowers compared with diurnal pollinators in one population. The two types of pollinators caused correlational selection on different pairs of traits and selected for different combinations of spur length and number of flowers. The results demonstrate that spatial variation in interactions with pollinators may result in differences in directional and correlational selection on floral traits in a plant with a semi-generalized pollination system, and suggest that differences in the relative importance of diurnal and nocturnal pollinators can cause variation in selection. © 2015 The Authors. New Phytologist © 2015 New Phytologist Trust.
Two new additions to the orchid flora of Manipur, India
DOI:10.2307/4120346 URL [本文引用: 1]
Angiosperm phylogeny poster (APP)—Flowering plant systematics, 2019
Evolution of nectar spur length in a clade of Linaria reflects changes in cell division rather than in cell expansion
DOI:10.1093/aob/mcx213
PMID:29370374
[本文引用: 2]
Nectar spurs (tubular outgrowths of a floral organ which contain, or give the appearance of containing, nectar) are hypothesized to be a 'key innovation' which can lead to rapid speciation within a lineage, because they are involved in pollinator specificity. Despite the ecological importance of nectar spurs, relatively little is known about their development. We used a comparative approach to investigate variation in nectar spur length in a clade of eight Iberian toadflaxes.Spur growth was measured at the macroscopic level over time in all eight species, and growth rate and growth duration compared. Evolution of growth rate was reconstructed across the phylogeny. Within the clade we then focused on Linaria becerrae and Linaria clementei, a pair of sister species which have extremely long and short spurs, respectively. Characterization at a micromorphological level was performed across a range of key developmental stages to determine whether the difference in spur length is due to differential cell expansion or cell division.We detected a significant difference in the evolved growth rates, while developmental timing of both the initiation and the end of spur growth remained similar. Cell number is three times higher in the long spurred L. becerrae compared with L. clementei, whereas cell length is only 1.3 times greater. In addition, overall anisotropy of mature cells is not significantly different between the two species.We found that changes in cell number and therefore in cell division largely explain evolution of spur length. This contrasts with previous studies in Aquilegia which have found that variation in nectar spur length is due to directed cell expansion (anisotropy) over variable time frames. Our study adds to knowledge about nectar spur development in a comparative context and indicates that different systems may have evolved nectar spurs using disparate mechanisms.
Petal, sepal, or tepal? B-genes and monocot flowers
DOI:S1360-1385(16)30189-3
PMID:27894712
[本文引用: 1]
In petaloid monocots expansion of B-gene expression into whorl 1 of the flower results in two whorls of petaloid organs (tepals), as opposed to sepals in whorl 1 of typical eudicot flowers. Recently, new gene-silencing technologies have provided the first functional data to support this, in the genus Tricyrtis (Liliaceae).Copyright © 2016 Elsevier Ltd. All rights reserved.
A monograph of the genus Disporum
DOI:10.1111/j.1095-8339.1838.tb00201.x URL [本文引用: 1]
A synopsis of Corybas (Orchidaceae) in west Malesia and Asia
DOI:10.2307/4103118 URL [本文引用: 1]
Complex developmental and transcriptional dynamics underlie pollinator-driven evolutionary transitions in nectar spur morphology in Aquilegia (columbine)
DOI:10.1002/ajb2.16046
PMID:35971626
[本文引用: 4]
Determining the developmental programs underlying morphological variation is key to elucidating the evolutionary processes that generated the stunning biodiversity of the angiosperms. Here, we characterize developmental and transcriptional dynamics of the elaborate petal nectar spur of Aquilegia (columbine) in species with contrasting pollination syndromes and spur morphologies.We collected petal epidermal cell number and length data across four Aquilegia species, two with short, curved nectar spurs of the bee-pollination syndrome, and two with long, straight spurs of the hummingbird syndrome. We also performed RNA-seq on A. brevistyla (bee) and A. canadensis (hummingbird) distal and proximal spur compartments at multiple developmental stages. Finally, we intersected these datasets with a previous QTL mapping study on spur length and shape to identify new candidate loci.The differential growth between the proximal and distal surfaces of curved spurs is primarily driven by differential cell division. However, independent transitions to straight spurs in the hummingbird syndrome have evolved by increasing differential cell elongation between spur surfaces. The RNA-seq data reveal these tissues to be transcriptionally distinct and point to auxin signaling as being involved with the differential cell elongation responsible for the evolution of straight spurs. We identify several promising candidate genes for future study.Our study, taken together with previous work in Aquilegia, reveals the complexity of the developmental mechanisms underlying trait variation in this system. The framework we established here will lead to exciting future work examining candidate genes and processes involved in the rapid radiation of the genus. This article is protected by copyright. All rights reserved.This article is protected by copyright. All rights reserved.
Do pollinators determine hybridization patterns in sympatric Satyrium (Orchidaceae) species?
DOI:10.1007/BF00985575 URL [本文引用: 1]
Elaborate petals and staminodes in eudicots: diversity, function, and evolution
Macroevolutionary dynamics of nectar spurs, a key evolutionary innovation
DOI:10.1111/nph.15654
PMID:30570752
[本文引用: 5]
Floral nectar spurs are widely considered a key innovation promoting diversification in angiosperms by means of pollinator shifts. We investigated the macroevolutionary dynamics of nectar spurs in the tribe Antirrhineae (Plantaginaceae), which contains 29 genera and 300-400 species (70-80% spurred). The effect of nectar spurs on diversification was tested, with special focus on Linaria, the genus with the highest number of species. We generated the most comprehensive phylogeny of Antirrhineae to date and reconstructed the evolution of nectar spurs. Diversification rate heterogeneity was investigated using trait-dependent and trait-independent methods, and accounting for taxonomic uncertainty. The association between changes in spur length and speciation was examined within Linaria using model testing and ancestral state reconstructions. We inferred four independent acquisitions of nectar spurs. Diversification analyses revealed that nectar spurs are loosely associated with increased diversification rates. Detected rate shifts were delayed by 5-15 Myr with respect to the acquisition of the trait. Active evolution of spur length, fitting a speciational model, was inferred in Linaria, which is consistent with a scenario of pollinator shifts driving diversification. Nectar spurs played a role in diversification of the Antirrhineae, but diversification dynamics can only be fully explained by the complex interaction of multiple biotic and abiotic factors.© 2018 The Authors. New Phytologist © 2018 New Phytologist Trust.
Floral isolation between Aquilegia formosa and Aquilegia pubescens
New species of Cuphea section Melvilla (Lythraceae) and an annotated key to the section
DOI:10.2307/2807021 URL [本文引用: 1]
Foraging ability of rufous hummingbirds on hummingbird flowers and hawkmoth flowers
Plastid genome data provide new insights into the phylogeny and evolution of the genus Epimedium
DOI:10.1016/j.jare.2021.06.020 URL [本文引用: 2]
The diversification of Halenia (Gentianaceae): ecological opportunity versus key innovation
The plant genus Halenia (Gentianaceae) consists of herbs growing in temperate and tropical alpine habitats and most species possess flowers in which nectar is produced in spurs. This probably helps reward only specialized long-tongued pollinators, and a narrow pollinator/flower relationship is thought to accelerate diversification rates (a key innovation). To test the pattern of diversification of Halenia against the unspurred sister group we reconstructed phylogenetic relationships among 22 species plus outgroups using nuclear ITS and chloroplast rpl16 intron sequence data. We show that Halenia originated in East Asia and migrated via North America into Central America. From there, it colonized South America three times independently, probably within the last million years. Significant changes in diversification rates were found during the evolution of Halenia using a sister group method, a likelihood method, and a diversity-through-time plot. In contrast to other studies, we could not observe a direct speciation rate effect of the evolution of nectar spurs in comparison with the unspurred sister group of Halenia. Rather, increases in diversification occurred following the colonization of Central and South America by spurred progenitor taxa. This later switch in diversification may have resulted from the availability of new geographical and ecological opportunities, or from the availability of more and different pollinators in these regions. Following the latter hypothesis, the nectar spurs were a preadaption and functioned as a key innovation only in this new biotic environment. After an initial rapid increase, a reduction in diversification rate was observed in Central America, probably illustrating density dependence of speciation rates. Finally, we found preliminary evidence for the key innovation hypothesis in geologically young spurred and unspurred lineages of Halenia in South America.
The evolution of floral nectaries in Disa (Orchidaceae: Disinae): recapitulation or diversifying innovation?
DOI:10.1093/aob/mct197
PMID:23997231
[本文引用: 1]
The Orchidaceae have a history of recurring convergent evolution in floral function as nectar production has evolved repeatedly from an ancestral nectarless state. However, orchids exhibit considerable diversity in nectary type, position and morphology, indicating that this convergence arose from alternative adaptive solutions. Using the genus Disa, this study asks whether repeated evolution of floral nectaries involved recapitulation of the same nectary type or diversifying innovation. Epidermis morphology of closely related nectar-producing and nectarless species is also compared in order to identify histological changes that accompanied the gain or loss of nectar production.The micromorphology of nectaries and positionally equivalent tissues in nectarless species was examined with light and scanning electron microscopy. This information was subjected to phylogenetic analyses to reconstruct nectary evolution and compare characteristics of nectar-producing and nectarless species.Two nectary types evolved in Disa. Nectar exudation by modified stomata in floral spurs evolved twice, whereas exudation by a secretory epidermis evolved six times in different perianth segments. The spur epidermis of nectarless species exhibited considerable micromorphological variation, including strongly textured surfaces and non-secreting stomata in some species. Epidermis morphology of nectar-producing species did not differ consistently from that of rewardless species at the magnifications used in this study, suggesting that transitions from rewardlessness to nectar production are not necessarily accompanied by visible morphological changes but only require sub-cellular modification.Independent nectary evolution in Disa involved both repeated recapitulation of secretory epidermis, which is present in the sister genus Brownleea, and innovation of stomatal nectaries. These contrasting nectary types and positional diversity within types imply weak genetic, developmental or physiological constraints in ancestral, nectarless Disa. Such functional convergence generated by morphologically diverse solutions probably also underlies the extensive diversity of nectary types and positions in the Orchidaceae.
The influence of nectar production on hawkmoth behavior, self pollination, and seed production in Mirabilis multiflora (Nyctaginaceae)
DOI:10.1002/ajb2.1995.82.issue-2 URL [本文引用: 2]
Floral nectar spurs and diversification
DOI:10.1086/297508 URL [本文引用: 3]
Spurring plant diversification: Are floral nectar spurs a key innovation?
Verne Grant and evolutionary studies of Aquilegia
DOI:10.1046/j.1469-8137.2003.00950.x
URL
[本文引用: 1]
One of Verne Grant's lasting contributions to plant evolutionary biology has been the recognition that differences between plants in floral characters can have a dramatic impact on both pollinator visitation and pollen transfer and thus affect reproductive isolation between nascent plant species (collectively, floral isolation). Here we review some of the concepts and findings from Grant's work on floral isolation, particularly with respect to the genus Aquilegia (Ranunculaceae). It has now been over 50 yr since Grant first published on the role of floral isolation on reproductive isolation and speciation in Aquilegia and we compare and contrast his findings with our own work on this genus. We find that the data largely support Grant's findings and that Aquilegia will continue to offer great opportunities to learn about the processes of adaptation and speciation.
Two new species of Viola (Violaceae) from the Intermountain West, USA
DOI:10.2307/2806928 URL [本文引用: 1]
A geographical perspective on the relationship between Impatiens spur lengths and bill lengths of sunbirds in Afrotropical Mountains
DOI:10.1002/ece3.v11.7 URL [本文引用: 1]
The terminology of floral larceny
DOI:10.2307/1936841 URL [本文引用: 1]
Nectar robbing in Ipomopsis aggregata: effects on pollinator behavior and plant fitness
DOI:10.1007/s004420050617 URL [本文引用: 1]
Nectar robbing: ecological and evolutionary perspectives
DOI:10.1146/ecolsys.2010.41.issue-1 URL [本文引用: 2]
Variation in nectar robbing over time, space, and species
DOI:10.1007/s00442-002-1060-z
PMID:28466170
[本文引用: 2]
Spatiotemporal variation in the interactions among plants and animals is widespread; yet our conceptual and empirical understanding of this variation is limited to a few types of visitors, mainly herbivores, pollinators, seed predators, and seed dispersers. Despite the ubiquity of nectar robbing and the strength of its effects on plant fitness, we know relatively little about the magnitude and intensity of spatial and temporal variation in interactions among plants and nectar robbers. The purpose of the present study was to quantify spatial and temporal variation in the interactions between a nectar-robbing bumblebee and its host plants. In the Rocky Mountains of Colorado, USA, over 7 years, and multiple locations, we measured levels of nectar robbing by the bumblebee Bombus occidentalis and its interactions with four different host plants, Delphinium nuttallianum (Ranunculaceae), Ipomopsis aggregata (Polemoniaceae), Corydalis caseana (Fumariaceae), and Linaria vulgaris (Scrophulariaceae). Significant variation was found in the robbing rates experienced by different species. Within species, there was variation in robbing rates on an annual basis, on a seasonal basis, among different sites, and within sites. This variation may have important consequences with respect to the population dynamics of host plants as well as selection on floral and flowering traits. Electronic supplementary material is available if you access this article at http://dx.doi.org/10.1007/s00442-002-1060-z. On that page (frame on the left side), a link takes you directly to the supplementary materials.
Spurs in a spur: perianth evolution in the Delphinieae (Ranunculaceae)
DOI:10.1086/667613 URL [本文引用: 3]
Convergent evolution of sunbird pollination systems of Impatiens species in tropical Africa and hummingbird systems of the New World
DOI:10.1111/bij.2015.115.issue-1 URL [本文引用: 1]
Floral development of Hydrocera and Impatiens reveals evolutionary trends in the most early diverged lineages of the Balsaminaceae
DOI:10.1093/aob/mcs065
PMID:22499855
[本文引用: 1]
Balsaminaceae consist of two genera, the monospecific Hydrocera and its species-rich sister Impatiens. Although both genera are seemingly rather similar in overall appearance, they differ in ecology, distribution range, habitat preference and morphology. Because morphological support for the current molecular phylogenetic hypothesis of Impatiens is low, a developmental study is necessary in order to obtain better insights into the evolutionary history of the family. Therefore, the floral development of H. triflora and I. omeiana was investigated, representing the most early-diverged lineage of Impatiens, and the observations were compared with the literature.Flowers at all developmental stages were examined using scanning electron microscopy and light microscopy.In Hydrocera, two whorls of five free perianth primordia develop into a less zygomorphic perianth compared with its sister genus. The androecial cap originates from five individual stamen primordia. Post-genital fusion of the upper parts of the filaments result in a filament ring below the anthers. The anthers fuse forming connivent anther-like units. The gynoecium of Hydrocera is pentamerous; it is largely synascidiate in early development. Only then is a symplicate zone formed resulting in style and stigmas. In I. omeiana, the perianth is formed as in Hydrocera. Five individual stamen primordia develop into five stamens, of which the UPPER PART OF THE FILAMENTS CONVERGE WITH EACH OTHER. THE GYNOECIUM OF I. OMEIANA IS TETRAMEROUS; IT APPEARS ANNULAR IN EARLY DEVELOPMENT.Comparison of the present results with developmental data from the literature confirms the perianth morphocline hypothesis in which a congenital fusion of the parts of the perianth results in a shift from pentasepalous to trisepalous flowers. In addition, the development of the androecial cap and the gynoecium follows several distinct ontogenetic sequences within the family.
Bird pollination in South African species of Satyrium (Orchidaceae)
DOI:10.1007/BF00985239 URL [本文引用: 2]
Coevolution between food-rewarding flowers and their pollinators
Specialization for pollination by beetles and wasps: the role of lollipop hairs and fragrance in Satyrium microrrhynchum (Orchidaceae)
DOI:10.3732/ajb.94.1.47
PMID:21642207
[本文引用: 1]
Exposed nectar presentation is a key trait in flowers specialized for pollination by short-tongued insects. We investigated the pollination of Satyrium microrrhynchum, a rare South African orchid in which nectar is secreted as droplets on long floral hairs ("lollipop hairs") at the mouth of a shallow labellum. Our observations indicate that this orchid is pollinated specifically by two insect species: a cetoniid beetle (Atrichelaphinus tigrina) and a pompilid wasp (Hemipepsis hilaris). Both insects have short mouthparts and remove nectar from the hairs with sweeping motions of their mouthparts. Pollinaria become attached to the upper surface of their heads while they feed on the nectar. Beetles damage the hairs while feeding, which may explain the positive relationship between hair damage and pollination success in plants of S. microrrhynchum from populations where beetles were common. The orchid has cryptic green-yellow flowers with spectral reflectance similar to that of its leaves. The fragrance from plants in three populations, analyzed using gas chromatography coupled to mass spectrometry, was dominated by various terpenoids; linalool was the most abundant. Plants in different populations emitted similar compounds, but eugenol and derivatives of this compound were found in only one of the three populations. In an electrophysiological study (gas chromatography coupled to electroantennography), using antennae of A. tigrina, clear signals were elicited by some of the floral scent compounds.
Diverse pollination systems of the twin-spurred orchid genus Satyrium in African grasslands
DOI:10.1007/s00606-010-0411-1 URL [本文引用: 3]
Long-tongued fly pollination and evolution of floral spur length in the Disa draconis complex (Orchidaceae)
DOI:10.1111/j.1558-5646.1997.tb02387.x
PMID:28568792
[本文引用: 2]
Field studies in South Africa showed that floral spur length in the Disa draconis complex (Orchidaceae) varies enormously between populations in the southern mountains (means = 32-38 mm), lowland sandplain (mean = 48 mm), and northern mountains (means = 57-72 mm). We tested the hypothesis that divergence in spur length has resulted from selection exerted through pollinator proboscis length. Short-spurred plants in several southern mountain populations, as well as long-spurred plants in one northern mountain population, were pollinated by a horsefly, Philoliche rostrata (Tabanidae), with a proboscis length that varied from 22 to 35 mm among sites. Long-spurred plants on the sandplain were pollinated by the tanglewing fly, Moegistorynchus longirostris (Nemestrinidae), which has a very long proboscis (mean = 57 mm). Selection apparently favors long spurs in sandplain plants, as artificial shortening of spurs resulted in a significant decline in pollen receipt and fruit set, although pollinaria removal was not significantly affected. Fruit set in the study populations was limited by pollen availability, which further suggests that selection on spur length occurs mainly through the female component of reproductive success.© 1997 The Society for the Study of Evolution.
Unrecorded and introduced taxon in Korea: Cymbalaria muralis P. Gaetrn. (Scrophulariaceae)
DOI:10.11110/kjpt.2009.39.2.120 URL [本文引用: 1]
Phylogeny of the Bifrenaria (Orchidaceae) complex based on morphology and sequence data from nuclear rDNA internal transcribed spacers (ITS) and chloroplast trnL-trnF region
DOI:10.1086/342035 URL [本文引用: 1]
Nectar spur evolution in the Mexican lobelias (Campanulaceae: Lobelioideae)
DOI:10.3732/ajb.92.3.558
URL
[本文引用: 2]
Phylogenetic studies are often hampered by the independent evolution of characters that may potentially obscure relationships. The adaptive significance of the nectar spur and its evolution within the Mexican lobeliads (Campanulaceae) is considered here. The taxonomic delimitations of Heterotoma from the Mexican species within the genera Lobelia and Calcaratolobelia were tested. Independent molecular data were gathered to determine whether the Mexican spurred lobeliads should be treated as distinct genera. The internal transcribed spacer (ITS) region from 18–26S nuclear rDNA and chloroplast DNA from the 3′ trnK intron were sequenced from 14 representative species. Our data suggest that Heterotoma, as originally conceived, is a good evolutionary unit within Lobelia and that the presence of a nectar spur is an important morphological character that can be used in defining phylogenetic position. This study also suggests that morphological changes associated with hummingbird pollination have evolved more than once in the Mexican lobeliads, from small blue‐flowered, insect‐pollinated relatives.
Comparative anatomy of the lip spur and additional lateral sepal spurs in a three-spurred form (f. fumeauxiana) of Anacamptis pyramidalis
Studies on the ultrastructure of a three-spurred fumeauxiana form of Anacamptis pyramidalis
DOI:10.1007/s00606-012-0611-y URL [本文引用: 1]
Morphological complexity as a floral signal: from perception by insect pollinators to co-evolutionary implications
A synopsis of the genus Disperis Sw. (Orchidaceae)
Do flowers removed of either nectar or pollen attract fewer bumblebee pollinators? An experimental test in Impatiens oxyanthera
Pollen and nectar are the primary rewards offered by flowers to pollinators. In floral visitors of some plant species, pollen thieves and nectar robbers cause the reduction in pollen grain number and nectar volume, respectively. However, it remains unclear whether the absence of either of the two rewards in a given flower reduces its attraction to nectar- and pollen-collecting pollinators. We hypothesized that flowers removed of either nectar or pollen would attract fewer pollinators. We studied protandrous Impatiens oxyanthera, whose flowers provide bumblebee pollinators with both nectar and pollen in the male phase. We conducted floral reward manipulation experiments to explore how the removal of either nectar or pollen from flowers influences pollinator behaviour by comparing their visitation rates and visit duration. Compared with the control flowers, the flowers removed of pollen attracted significantly more bumblebee pollinators per 30 min, but the flowers removed of nectar or those removed of both pollen and nectar attracted significantly fewer bumblebee pollinators per 30 min. Moreover, the visit duration of bumblebee pollinators to control flowers or flowers removed of pollen was longer than that to flowers removed of nectar or those removed of both pollen and nectar. Our investigations indicated that compared with control flowers, the flowers removed of nectar attracted fewer bumblebee pollinators, supporting our hypothesis. However, our other hypothesis that pollen removal would reduce pollinator visits was not supported by our results. Instead, compared with control flowers, the flowers that contained only nectar attracted more bumblebee pollinators. Nectar seems to be the main reward, and bumblebee pollinators mainly used the absence of pollen as a visual signal to locate I. oxyanthera flowers with a potentially higher amount of nectar.
A molecular phylogeny of Chinese orchids
DOI:10.1111/jse.v54.4 URL [本文引用: 1]
Effect of floral morphology on fruit set in Epimedium sagittatum (Berberidaceae)
DOI:10.1007/s00606-009-0145-0 URL [本文引用: 1]
Floral developmental evidence for the systematic relationships of Tropaeolum (Tropaeolaceae)
DOI:10.1006/anbo.2001.1525 URL [本文引用: 1]
Flower nectar trichome structure of carnivorous plants from the genus butterworts Pinguicula L. (Lentibulariaceae)
DOI:10.1007/s00709-019-01433-8 [本文引用: 1]
The pollination ecology of Dicentra cucullaria
DOI:10.1002/ajb2.1970.57.issue-1 URL [本文引用: 1]
The pollination ecology of Delphinium tricorne (Ranunculaceae)
DOI:10.1002/ajb2.1975.62.issue-10 URL [本文引用: 1]
Pollination interactions in sympatric Dicentra species
DOI:10.1002/ajb2.1978.65.issue-1 URL [本文引用: 1]
The relationship between cell division and elongation during development of the nectar- yielding petal spur in Centranthus ruber (Valerianaceae)
DOI:10.1093/aob/mcu261 URL [本文引用: 1]
A revision of Fumariaceae (Fumarioideae) in southern Africa, including naturalized taxa
DOI:10.4102/abc.v39i1.229
URL
[本文引用: 1]
The native and naturalized species of Fumariaceae in southern Africa are reviewed, and keys and full descriptions are provided. All relevant regional synonyms are given and the indigenous species are illustrated. Three native genera with four species are recognized. The previously taxonomically unplaced genus Cysticapnos Mill. (3 spp.) is included with two other monotypic native South African genera, Discocapnos Cham. Schltdl. and Trigonocapnos Schltr., in an enlarged circumscription of subtribe Discocapninae, which is morphologically defined by tendrilliferous leaves and mostly sessile racemes with the lowermost flower basal. Two subspecies are recognized in Cysticapnos vesicaria (E.Mey. ex Bemh.) Lidén, subsp. vesicaria with fewer, smaller flowers and subsp. namaquensis J.C.Manning Goldblatt for plants from Namaqualand with more numerous, often larger flowers. C. parviflora Lidén appears to be nothing more than a dwarf-fruited form of C. vesicaria, in which heterocarpy has long been known. C. pruinosa (E.Mey. ex Bemh.) Lidén is recorded for the first time to be a short-lived perennial and not an annual, thus unique in the tribe Fumarieae. Discocapnos mundii var. dregei Harv. from the southern Cape is treated as subsp. dregei (Harv.) J.C.Manning Goldblatt on account of its slightly smaller fruits and significant geographical disjunction from subsp. mundii. Three introduced species are included, Fumaria capreolata L., F. muralis Sond. ex W.D.Koch subsp. muralis and F. parviflora Lam. var. parviflora, although only the last two appear to be truly naturalized.
The pollination ecology of Aquilegia elegantula and A. caerulea (Ranunculaceae) in Colorado
DOI:10.1002/ajb2.1978.65.issue-4 URL [本文引用: 1]
Hawkmoths and the geographic patterns of floral variation in Aquilegia caerulea
DOI:10.2307/2408246 URL [本文引用: 1]
The pollination ecology of Aquilegia micrantha (Ranunculaceae) in Colorado
DOI:10.2307/3671384 URL [本文引用: 1]
Species diversity vs. morphological disparity in the light of evolutionary developmental biology
DOI:10.1093/aob/mcv134
PMID:26346718
[本文引用: 1]
Two indicators of a clade's success are its diversity (number of included species) and its disparity (extent of morphospace occupied by its members). Many large genera show high diversity with low disparity, while others such as Euphorbia and Drosophila are highly diverse but also exhibit high disparity. The largest genera are often characterized by key innovations that often, but not necessarily, coincide with their diagnostic apomorphies. In terms of their contribution to speciation, apomorphies are either permissive (e.g. flightlessness) or generative (e.g. nectariferous spurs).Except for Drosophila, virtually no genus among those with the highest diversity or disparity includes species currently studied as model species in developmental genetics or evolutionary developmental biology (evo-devo). An evo-devo approach is, however, potentially important to understand how diversity and disparity could rapidly increase in the largest genera currently accepted by taxonomists. The most promising directions for future research and a set of key questions to be addressed are presented in this review.From an evo-devo perspective, the evolution of clades with high diversity and/or disparity can be addressed from three main perspectives: (1) evolvability, in terms of release from previous constraints and of the presence of genetic or developmental conditions favouring multiple parallel occurrences of a given evolutionary transition and its reversal; (2) phenotypic plasticity as a facilitator of speciation; and (3) modularity, heterochrony and a coupling between the complexity of the life cycle and the evolution of diversity and disparity in a clade. This simple preliminary analysis suggests a set of topics that deserve priority for scrutiny, including the possible role of saltational evolution in the origination of high diversity and/or disparity, the predictability of morphological evolution following release from a former constraint, and the extent and the possible causes of a positive correlation between diversity and disparity and the complexity of the life cycle.© The Author 2015. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Effects of nectar robbing on nectar dynamics and bumblebee foraging strategies in Linaria vulgaris (Scrophulariaceae)
DOI:10.1111/oik.2005.110.issue-2 URL [本文引用: 1]
Gastrochilus pankajkumarii, (Aeridinae, Epidendroideae, Orchidaceae), a new lithophytic orchid from southern Vietnam
Effects of local abundance on pollination and reproduction in the narrow endemic endangered species Delphinium bolosii (Ranunculaceae)
Floral features, pollination biology, and breeding system of Comparettia coccinea (Orchidaceae: Oncidiinae)
Classification and species of Platostoma and its relationship with Haumaniastrum (Labiatae)
DOI:10.2307/4110385 URL [本文引用: 2]
Pollination by flower chafer beetles in Eulophia ensata and Eulophia welwitschii (Orchidaceae)
DOI:10.1016/j.sajb.2009.07.008 URL [本文引用: 1]
Nectar trichome structure of aquatic bladderworts from the section Utricularia (Lentibulariaceae) with observation of flower visitors and pollinators
DOI:10.1007/s00709-018-1216-2 [本文引用: 1]
Evolution of spur-length diversity in Aquilegia petals is achieved solely through cell-shape anisotropy
DOI:10.1098/rspb.2011.1873
URL
[本文引用: 2]
\n The role of petal spurs and specialized pollinator interactions has been studied since Darwin.\n Aquilegia\n petal spurs exhibit striking size and shape diversity, correlated with specialized pollinators ranging from bees to hawkmoths in a textbook example of adaptive radiation. Despite the evolutionary significance of spur length, remarkably little is known about\n Aquilegia\n spur morphogenesis and its evolution. Using experimental measurements, both at tissue and cellular levels, combined with numerical modelling, we have investigated the relative roles of cell divisions and cell shape in determining the morphology of the\n Aquilegia\n petal spur. Contrary to decades-old hypotheses implicating a discrete meristematic zone as the driver of spur growth, we find that\n Aquilegia\n petal spurs develop via anisotropic cell expansion. Furthermore, changes in cell anisotropy account for 99 per cent of the spur-length variation in the genus, suggesting that the true evolutionary innovation underlying the rapid radiation of\n Aquilegia\n was the mechanism of tuning cell shape.\n
Generation of shape complexity through tissue conflict resolution
Out-of-plane tissue deformations are key morphogenetic events during plant and animal development that generate 3D shapes, such as flowers or limbs. However, the mechanisms by which spatiotemporal patterns of gene expression modify cellular behaviours to generate such deformations remain to be established. We use the Snapdragon flower as a model system to address this problem. Combining cellular analysis with tissue-level modelling, we show that an orthogonal pattern of growth orientations plays a key role in generating out-of-plane deformations. This growth pattern is most likely oriented by a polarity field, highlighted by PIN1 protein localisation, and is modulated by dorsoventral gene activity. The orthogonal growth pattern interacts with other patterns of differential growth to create tissue conflicts that shape the flower. Similar shape changes can be generated by contraction as well as growth, suggesting tissue conflict resolution provides a flexible morphogenetic mechanism for generating shape diversity in plants and animals.
Evidence for pollination ecotypes in the yellow-fringed orchid, Platanthera ciliaris
DOI:10.1111/j.1558-5646.1990.tb04283.x
PMID:28568207
[本文引用: 1]
Platanthera ciliaris is a butterfly-pollinated, terrestrial orchid with a loose terminal raceme of 10-50 orange flowers, characterized by a long nectariferous spur. In the southeastern United States, P. ciliaris occurs in the Appalachian mountains and coastal-plain physiographic provinces, but it is found rarely in the intervening Piedmont. In 1983 and 1984, detailed observations of two populations within these disjunct areas revealed that the butterfly species that serve as the primary pollinators differ sharply. In the mountains, Papilio troilus (spicebush swallowtail) was the most frequent and effective visitor, whereas in the coastal plain, P. palamedes (palamedes swallowtail) was the predominant pollinator. Proboscis lengths of P. troilus (mean = 23.3 mm) were significantly shorter than those of P. palamedes (mean = 28.7 mm). Floral characters, most notably spur length, also differed significantly between mountain (mean = 23.8 mm) and coastal-plain (mean = 25.6 mm) plants. In both 1983 and 1984, levels of pollinator service, as assessed by rates of removal and insertion of pollinia, were higher in the mountains (0.81 and 0.86) than in the coastal plain (0.63 and 0.67). In addition, fruit-set was significantly greater in mountain (83.9% in 1983, 86.5% in 1984) than in coastal-plain (63.8% in 1983, 65.5% in 1984) populations. We hypothesize that selection pressure exerted through pollinator proboscis lengths has resulted in pollination ecotypes of P. ciliaris. The short-spurred mountain plants appear to be in equilibrium with their short-tongued butterfly pollinators, receiving high levels of effective pollination and achieving high levels of fruit-set. Coastal-plain plants produce flowers with longer spurs which, nevertheless, are shorter than optimal for insuring that very long-tongued butterflies make contact with their pollinia. Thus, effective pollinator service and fruit-set are reduced. Correlation analyses in 1984 showed a positive relationship between spur length and fruit-set only in the coastal plain. The presence of long-tongued nectar thieves in the coastal plain may also contribute to lower effective pollinator service and fruit-set. Results of reciprocal-transplant studies attempting to determine the genetic basis of floral characters, including spur length, were inconclusive but suggest that differences between mountain and coastal-plain plants are not due solely to phenotypic plasticity.© 1990 The Society for the Study of Evolution.
Evolution of floral nectaries in Iridaceae
DOI:10.2307/3298546 URL [本文引用: 1]
Pollination of Impatiens capensis: pollinators and nectar robbers
Adaptive radiation of reproductive characteristics in angiosperms, I: Pollination mechanisms
DOI:10.1146/ecolsys.1970.1.issue-1 URL [本文引用: 1]
Chromosome numbers and relationships in tribe Hemimerideae (Scrophulariaceae)
DOI:10.2307/2419563 URL [本文引用: 1]
The association between oil-producing flowers and oil-collecting bees in the Drakensberg of southern Africa. Monogr
Pollinator adaptation to oil-secreting flowers—Rediviva and Diascia
DOI:10.1111/j.1558-5646.1990.tb03857.x PMID:28564320 [本文引用: 2]
Anatomy and ultrastructure of spur nectary of Gymnadenia conopsea (L.) Orchidaceae
DOI:10.5586/asbp.2001.034
URL
[本文引用: 1]
The anatomy and the ultrastructure of spur nectaries of <em>Cymnadenia conopsea</em> at different developmental stages were investigated. The secretory epidermis surrounded the inside of spur and formed many unicellular papillae, which significantly enlarged the secretory surface. At the activity stage the epidermal cells contained characteristic plastids with well developed intraplastidal membrane system and numerous osmiophillic globules. The contact of plastids and endoplasmic reticulum indicates a possibility of the involvement of these structures in the secretory processes. The cell wall and the cuticle did not form a barrier for the secreted nectar and no pores or cracks were visible in the cuticle covering secretory papillae.
Five new species of Platostoma (Lamiaceae) from North-Eastern Thailand
Bee pollination of the endangered orchid Calanthe discolor through a generalized food-deceptive system
DOI:10.1007/s00606-013-0894-7 URL [本文引用: 1]
Pollination system and the effect of inflorescence size on fruit set in the deceptive orchid Cephalanthera falcata
DOI:10.1007/s10265-015-0716-9
PMID:25801274
[本文引用: 1]
Larger inflorescences in reward-producing plants can benefit plants by increasing both pollinator attraction and the duration of visits by individual pollinators. However, ultimately, inflorescence size is determined by the balance between the benefits of large inflorescences and the increased cost of geitonogamy. At present, little is known about the relationship between inflorescence size and fecundity in deceptive plants. Given that pollinators are likely to leave inflorescences lacking rewards quickly, it seems unlikely that longer pollinator visits and the risk of geitonogamy would be strong selective pressures in these species, which indicates that pollinator attraction might be the most important factor influencing their inflorescence size. Here we examined the pollination ecology of the deceptive orchid Cephalanthera falcata in order to clarify the effects of inflorescence size on the fruit set of this non-rewarding species. Field observations of the floral visitors showed that C. falcata is pollinated by the andrenid bee Andrena aburana, whilst pollination experiments demonstrated that this orchid species is neither autogamous nor apogamous, but is strongly pollinator dependent. Three consecutive years of field observations revealed that fruit set was positively correlated with the number of flowers per inflorescence. These results provide strong evidence that the nectarless orchid C. falcata benefits from producing larger inflorescences that attract a greater number of innate pollinators. Large inflorescences may have a greater positive effect on fruit set in deceptive plants because a growing number of studies suggest that fruit set in reward-producing plants is usually unaffected by display size.
Deep flowers for long tongues
DOI:10.1016/S0169-5347(98)01359-7 URL [本文引用: 1]
Eltroplectris brachycentron Szlachetko (Orchidaceae), a new orchid species from Bolivia
DOI:10.2307/3391968 URL [本文引用: 1]
An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV
DOI:10.1111/boj.2016.181.issue-1 URL [本文引用: 1]
Specific hypotheses on the geographic mosaic of coevolution
DOI:10.1086/303208 URL [本文引用: 1]
The relationship between nectar spur curvature in jewelweed (Impatiens capensis) and pollen removal by hummingbird pollinators
DOI:10.1139/b03-014
URL
[本文引用: 1]
Floral nectar spurs are hypothesized to have had a major role in the evolution of floral diversity and plant-pollinator coadaptation. We examined variation in the degree of nectar spur curvature in two species of jewelweed (Impatiens capensis and Impatiens pallida) pollinated by different sets of pollinators. To distinguish between adaptive and nonadaptive explanations for between-species differences in curvature, we determined the relationship between spur curvature and pollen removal, which is one estimate of male reproductive success. Spur curvature exhibited considerable variation both within and among three populations, with spur angles ranging from 0° to 297°. A greenhouse experiment determined that spur curvature of I. capensis flowers has a broad-sense heritability of 0.636. Laboratory experiments indicated that flowers having recurved spurs deposit significantly more pollen grains on hummingbird visitors than flowers having perpendicular spurs, apparently as a result of greater contact between the androecium of curve-spurred flowers and the upper bill of hummingbirds. We also found a significant relationship between spur curvature and flower length, suggesting a developmental link between the two traits. We discuss the degree of spur curvature in bird-pollinated I. capensis as a function of both adaptive evolution and developmental constraint.Key words: pollination, nectar spurs, hummingbirds, Impatiens capensis, Impatiens pallida, pollen export, floral morphology, floral polymorphisms.
Pollinators, floral morphology and scent chemistry in the southern African orchid genus Schizochilus
DOI:10.1016/j.sajb.2010.07.004 URL [本文引用: 1]
Nectar traits differ between pollination syndromes in Balsaminaceae
DOI:10.1093/aob/mcz072
PMID:31120478
[本文引用: 1]
The attractiveness of nectar rewards depends both on the quantity of nectar produced and on its chemical composition. It is known that nectar quantity and chemical composition can differ in plant species depending on the main pollinator associated with the species. The main aims of this study were to test formally whether nectar traits are adapted to pollination syndromes in the speciose Balsaminaceae and, if so, whether a combination of nectar traits mirrors pollination syndromes.Comparative methods based on Ornstein-Uhlenbeck models were used to test whether nectar volume, nectar sucrose proportion, sugar and amino acid concentration and amino acid composition had evolved as a function of pollination syndromes in 57 species of Balsaminaceae. Cluster analysis and ordination were performed to derive clusters of species resembling each other in nectar composition.Evolutionary models for nectar volume and nectar sucrose proportion performed best when including information on pollination syndrome, while including such information improve model fit neither for sugar and amino acid concentration nor for amino acid composition. A significant relationship emerged between pollination syndrome and the combined nectar traits.Our results show that nectar volume and nectar sucrose proportion evolve rapidly towards optimal values associated with different pollination syndromes. The detection of a signal indicating that nectar traits in combination are to a certain extent able to predict pollination syndromes in Balsaminaceae suggests that a holistic approach including the whole set of nectar traits helps us to better understand evolution of nectar composition in response to pollinators.© The Author(s) 2019. Published by Oxford University Press on behalf of the Annals of Botany Company.
Flower specialisation: the occluded corolla of snapdragons (Antirrhinum) exhibits two pollinator niches of large long-tongued bees
DOI:10.1111/plb.12588
PMID:28590517
[本文引用: 1]
Flower specialisation of angiosperms includes the occluded corollas of snapdragons (Antirrhinum and some relatives), which have been postulated to be one of the most efficient structures to physical limit access to pollinators. The Iberian Peninsula harbours the highest number of species (18 Iberian of the 20 species of Antirrhinum) that potentially share similar pollinator fauna. Crossing experiments with 18 Iberian species from this study and literature revealed a general pattern of self-incompatibility (SI) - failure in this SI system has been also observed in a few plants - which indicates the need for pollinator agents in Antirrhinum pollination. Field surveys in natural conditions (304 h) found flower visitation (>85%) almost exclusively by 11 species of bee (Anthophora fulvitarsis, Anthophora plumipes, Anthidium sticticum, Apis mellifera, Bombus hortorum, Bombus pascuorum, Bombus ruderatus, Bombus terrestris, Chalicodoma lefebvrei, Chalicodoma pyrenaica and Xylocopa violacea). This result covering the majority of Antirrhinum species suggests that large bees of the two long-tongued bee families (Megachilidae, Apidae) are the major pollinators of Antirrhinum. A bipartite modularity analysis revealed two pollinator systems of long-tongued bees: (i) the long-studied system of bumblebees (Bombus spp.) associated with nine primarily northern species of Antirrhinum; and (ii) a newly proposed pollinator system involving other large bees associated with seven species primarily distributed in southern Mediterranean areas.© 2017 German Botanical Society and The Royal Botanical Society of the Netherlands.
The Diascia flower and its bee—An oil-based symbiosis in southern Africa
DOI:10.1111/plb.1984.33.issue-4 URL [本文引用: 1]
Pollinators and nectar robbers cause directional selection for large spur circle in Impatiens oxyanthera (Balsaminaceae)
DOI:10.1007/s00606-013-0794-x URL [本文引用: 2]
Geographic mosaic theory of coevolution in plant and pollinator interaction: process, evidence, prospect
植物与传粉者地理镶嵌的协同演化: 过程、证据与展望
Comparative pollen and dye transfer by pollinators of Delphinium nelsonii
DOI:10.2307/2389458 URL [本文引用: 1]
The pollinators of the Malagasy star orchids Angraecum sesquipedale, A. sororium and A. compactum and the evolution of extremely long spurs by pollinator shift
DOI:10.1111/plb.1997.110.issue-5 URL [本文引用: 2]
The genus Pomatocalpa (Orchidaceae) a taxonomic monograph
DOI:10.3100/1043-4534(2007)11[207:TGPOAT]2.0.CO;2 URL [本文引用: 1]
Structure of trichomatous nectaries in flowers of Lonicera kamtschatica (Sevast.) Pojark
DOI:10.5586/aa.2008.002
URL
[本文引用: 1]
The structure of the floral nectaries of <i>Lonicera kamtschatica</i> was examined using light microscopy, scanning electron microscopy and transmission electron microscopy. Nectariferous tissues are located in the lower portion of the corolla tube. It was found that the secretory tissue of the nectary was composed of two layers of epidermal formations: short papillae and about 3x longer unicellular trichomes. They cover the adaxial surface of a small spur. Nectar secretion takes place through the apical portion of the trichomes and papillae. The cell wall of the upper part of the trichome has protuberances participating in nectar transfer to the subcuticular space which reaches large dimensions. The lateral walls of the trichomes are saturated with cutin. The papillae have much thicker walls than the trichomes. In the papillae, there are no wall protuberances. Less secretion accumulates in the subcuticular cavities of the papillae than in the trichomes.
Pollinator shifts drive increasingly long nectar spurs in columbine flowers
DOI:10.1038/nature05857 [本文引用: 8]
Are the spurs more complex than other petal types in Epimedium? Evidence from development, micromorphology, and nectary structure
Equipped for migrations across high latitude regions? Reduced spur length and outcrossing rate in a biennial Halenia elliptica (Gentianaceae) with mixed mating system along a latitude gradient
Selection on spur shape in Impatiens capensis
DOI:10.1007/s00442-008-1014-1 URL [本文引用: 1]
Molecular phylogeny of the mainly Mediterranean genera Chaenorhinum, Kickxia and Nanorrhinum (Plantaginaceae, tribe Antirrhineae), with focus on taxa in the Flora Iranica region
DOI:10.1111/njb.2016.v34.i4 URL [本文引用: 1]
A role for the Auxin Response Factors ARF6 and ARF8 homologs in petal spur elongation and nectary maturation in Aquilegia
DOI:10.1111/nph.16633
PMID:32356309
[本文引用: 2]
The petal spur of the basal eudicot Aquilegia is a key innovation associated with the adaptive radiation of the genus. Previous studies have shown that diversification of Aquilegia spur length can be predominantly attributed to variation in cell elongation. However, the genetic pathways that control the development of petal spurs are still being investigated. Here, we focus on a pair of closely related homologs of the AUXIN RESPONSE FACTOR family, AqARF6 and AqARF8, to explore their roles in Aquileiga coerulea petal spur development. Expression analyses of the two genes show that they are broadly expressed in vegetative and floral organs, but have relatively higher expression in petal spurs, particularly at later stages. Knockdown of the two AqARF6 and AqARF8 transcripts using virus-induced gene silencing resulted in largely petal-specific defects, including a significant reduction in spur length due to a decrease in cell elongation. These spurs also exhibited an absence of nectar production, which was correlated with downregulation of STYLISH homologs that have previously been shown to control nectary development. This study provides the first evidence of ARF6/8 homolog-mediated petal development outside the core eudicots. The genes appear to be specifically required for cell elongation and nectary maturation in the Aquilegia petal spur.© 2020 The Authors. New Phytologist © 2020 New Phytologist Trust.
The effects of nectar robbing on plant reproduction and evolution
DOI:10.17521/cjpe.2006.0091 URL [本文引用: 1]
盗蜜行为在植物繁殖生态学中的意义
DOI:10.17521/cjpe.2006.0091
[本文引用: 1]
在动植物的相互关系中,盗蜜行为被认为是一种不同于普通传粉者的非正常访花行为。动物之所以要采取这种特殊的觅食策略,有假说认为是由访花者的口器和植物的花部形态不匹配造成的,也有认为是盗蜜行为提高了觅食效率从而使盗蜜者受益。在盗蜜现象中, 盗蜜者和宿主植物之间的关系是复杂的。盗蜜对宿主植物的影响尤其是对其繁殖适合度的影响归纳起来有正面、负面以及中性3类。与此同时,盗蜜者的种类, 性别及其掠食行为差异不仅与生境因素密切相关,而且会对宿主植物的繁殖成功产生直接或间接的影响。另外,盗蜜者的存在无疑对其它正常传粉者的访花行为也产生一定的影响,从而间接地影响宿主植物的繁殖成功, 而植物在花部形态上也出现了对盗蜜现象的适应性进化。作者认为, 盗蜜是短嘴蜂对长管型花最有效的一种掠食策略, 它不仅增加了盗蜜者对资源的利用能力, 而且由于盗蜜对宿主植物繁殖成功的不同的影响使其具有调节盗蜜者和宿主之间种群动态的作用, 两者的彼此适应是一种协同进化的结果。
Floral organogenesis in Urophysa rockii, a rediscovered endangered and rare species of Ranunculaceae
DOI:10.1139/cjb-2015-0232
URL
[本文引用: 1]
Urophysa is an Asian endemic genus in the Ranunculaceae, but data on floral organogenesis, which would be a useful complement to molecular data in clarifying the relationship with closely related taxa (Aquilegia and Semiaquilegia) in Ranunculaceae, are completely lacking. We used scanning electron microscopy and light microscopy to study the floral development of Urophysa rockii Ulbrich, a recently rediscovered species in this genus. The sepals are initiated spirally, whereas other organs are nonsimultaneously whorled; the floral phyllotaxis is whorled. Primordia of the sepals are lunular and truncate, but those of the petals and stamens are hemispherical and rounded. After sepal initiation, there is a delay in development, but the initiation of petals and stamens is continuous. The developmental sequence of the microspores in the stamens is centrifugal, although the stamens are initiated centripetally. The early developmental stages of the staminodes are similar to those of the stamens, although much smaller, so they may be phylogenetically homologous organs. The carpel primordia are lunular in shape and plicate. The mature ovule is anatropous and bitegmic. Urophysa shows similar floral development features as Aquilegia and Semiaquilegia, although with some differences, which supports the relationship inferred by DNA sequence data.
Anatomical and morphological studies of seed development in Sandersonia aurantiaca (Hook.)
DOI:10.1016/S0254-6299(15)31118-2 URL [本文引用: 2]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}