


天然林树冠下自然更新起来的树木, 幼苗阶段是其生活史中最脆弱、对生境变化最敏感的阶段(刘帅等, 2016)。幼苗的光合产物分配策略影响各个器官的相对生长速率, 并深刻地影响着幼苗的形态、活力与竞争力(Lacointe, 2000; 平晓燕等, 2010)。在小尺度上, 微生境直接决定了幼苗的存活和生长(Li et al., 2003; Li & Yang, 2004), 基岩、地表碎屑物、腐木等能改变更新幼苗生长的微地貌, 使不同生境的光照强度、温度、风速、土壤水分、积雪厚度等产生较大的差异, 从而影响幼苗的定居(Holtmeier & Broll, 1992; Resler et al., 2013)和生长(Batllori et al., 2009)。Wang等(2022)通过meta分析表明地表凋落物覆盖对森林幼苗的出苗率和成活率的总体影响是不利的, 对根、叶和总生物量的影响为中性, 对茎生物量和幼苗高度的影响为正向。在祁连山高海拔地区, 地表苔藓覆盖的盖度和厚度适中时, 其对青海云杉(Picea crassifolia)幼苗的萌发和生长才会具有积极影响(Wang et al., 2017)。此外, 在瑞士亚高山森林的研究表明林下植被过密时, 幼苗接收的光照不足且与周边植被的竞争加剧, 其光合物质积累可能会受到阻碍(von Arx et al., 2013)。
当林冠逐渐郁闭之后, 较大尺度的地形因子则决定着树木的生长(Li et al., 2003)。地形因子如海拔和坡向的变化可以实现光照、温度、水分、CO2、养分等的重新分配, 导致区域性的生境条件随地形而转变, 进而影响树木的更新定居, 对树木幼苗的生长和生物量积累起着不可忽视的作用(Antonelli et al., 2018; Zhang et al., 2022)。在瑞士阿尔卑斯山脉, 高海拔处的山赤松(Pinus uncinata)幼苗具有比低海拔处更大的叶生物量分配比例, 而其根生物量分配比例随着海拔的升高呈下降趋势(Bernoulli & Körner, 1999)。Li等(2003)在奥地利亚高山森林的研究表明, 随海拔增加, 28年以内的欧洲云杉(Picea abies)的茎生物量呈下降趋势, 而枝和叶生物量则没有明显的变化趋势。Zhang等(2022)综合海拔与坡向因子, 发现在青藏高原东南部的高海拔生境中急尖长苞冷杉(Abies georgei var. smithii)幼苗增加了对地下生物量的投入; 在北坡, 植被因子对幼苗生物量增长的贡献更大, 而在南坡土壤因子对其贡献更多。可见, 不同地区针叶树幼苗器官生物量对生境的响应规律存在地域性差别。
川西亚高山地处青藏高原东缘, 是我国西南天然林区的主体, 也是长江上游流域的重要生态屏障。独特的高山、亚高山地貌使该区域拥有多样化的植被特征, 丰富的天然林资源使其拥有复杂的森林经营历史(张远东等, 2005b)。岷江冷杉(Abies fargesii var. faxoniana)作为川西亚高山森林的优势种, 其幼苗的光合产物分配对增温的响应成为了近年来的研究热点(徐振锋等, 2008; 尹华军等, 2008; 杨兵等, 2010; 庞晓瑜等, 2018)。同时, 多数学者还关注了海拔梯度上岷江冷杉大树根系生物量的分布特征、周转速率、季节动态变化等(胡建利等, 2009; 马姜明等, 2010; 刘顺等, 2018)。尽管已有研究探讨了成熟郁闭林分内林窗与林下不同微生境条件下岷江冷杉幼苗的生物量分配格局的差异(鲜骏仁等, 2007), 但是林内微生境与地形生境各要素之间的相互作用对更新幼苗生长的影响极为复杂, 且大多数研究主要集中在单一指标上, 不同因素对岷江冷杉幼苗生物量分配的相对贡献仍未得到深入揭示。
川西亚高山不同恢复类型的森林, 其植被的物种组成和空间结构差异显著, 导致幼苗生境显著不同。因此, 本研究拟探索两个科学问题: 1)处于红桦(Betula albosinensis)阔叶林、红桦-岷江冷杉针阔混交林和岷江冷杉原始林3种恢复类型林分生境内的岷江冷杉幼苗, 其生物量分配格局及各器官间的异速生长关系是否存在差异? 2)红桦阔叶林和红桦-岷江冷杉针阔混交林2个次生林中制约岷江冷杉幼苗生物量增长的生境因子是否与岷江冷杉原始林不同? 本研究通过多重比较、异速生长分析和冗余分析方法, 揭示不同林分生境内岷江冷杉幼苗的生物量分配格局并着重探讨生境因子对岷江冷杉幼苗生物量积累和分配的制约作用, 以期为川西亚高山森林岷江冷杉更新幼苗的抚育管护以及次生林的结构优化调整提供支撑。
1 材料和方法
1.1 研究区概况及样点选取
研究区域位于川西阿坝藏族羌族自治州理县米亚罗林区(31.40°-31.92° N, 102.58°-103.07° E), 地处青藏高原向四川盆地过渡的高山峡谷区, 海拔2 200-5 500 m, 坡度多在30°以上。该区地处青藏高原气候区, 冬寒夏凉。年平均气温9 ℃左右, 年降水量700-1 000 mm, 年蒸发量1 000-1 900 mm (张远东等, 2019)。该区植被呈明显的垂直成带状态, 亚高山暗针叶林是此地的常见林型, 主要优势树种为岷江冷杉。该区的森林状况和经营历史在整个西南林区具有典型性和代表性(张远东等, 2005a)。目前, 红桦-岷江冷杉针阔混交林及红桦阔叶林主要分布在米亚罗海拔2 800-3 600 m的阴坡、半阴坡, 以及沟谷两旁山坡下部的阴湿生境中; 杜鹃(Rhododendron spp.) -岷江冷杉原始林主要分布在海拔3 700 m以上阴坡和半阴坡的高寒生境中(张远东等, 2005b)。
本研究中的红桦阔叶林与红桦-岷江冷杉针阔混交林均为岷江冷杉原始林被采伐后经自然恢复形成的次生林(表1), 自20世纪70年代大规模采伐后几乎未经历人为干扰, 恢复时间均约50年; 红桦阔叶林与红桦-岷江冷杉针阔混交林的主要区别在于采伐强度不同而导致岷江冷杉保留木的密度不同, 红桦-岷江冷杉林中岷江冷杉保留木密度更大, 恢复速率更快, 与红桦阔叶林所处恢复阶段不同; 目前, 红桦阔叶林中乔木层优势种为红桦, 而红桦-岷江冷杉针阔混交林中优势种为红桦和岷江冷杉。原始林为20世纪采伐时在高海拔林线交错带区域保留的母树林斑块, 几无人为干扰。
表1 川西亚高山样点的林分特征和岷江冷杉幼苗生物量样本量
Table 1
| 林分类型 Forest type | 样点 Sample site | 样点编号 Site code | 背景 Background | 乔木层 Tree layer | 灌木层 Shrub layer | 草本层 Herb layer | 坡向 Aspect slope | 坡度 Slope (°) | 海拔 Altitude (m) | 样本量 Sample size |
|---|---|---|---|---|---|---|---|---|---|---|
| BB | SHG | 1 | 盖度 Coverage (%) | 70 | 35 | 85 | NW | 25-28 | 3 396 | 45 |
| 平均高度 Average height (m) | 13 | 3.5 | 0.4 | |||||||
| BA | 272 | 2 | 盖度 Coverage (%) | 60 | 35 | 70 | N | 25-35 | 3 258 | 72 |
| 平均高度 Average height (m) | 14 | 3.5 | 0.4 | |||||||
| AP | 189 | 3 | 盖度 Coverage (%) | 70 | 20 | 45 | N | 35-37 | 3 995 | 44 |
| 4 | 平均高度 Average height (m) | 12 | 2.0 | 0.3 | 35-40 | 3 793 | 44 | |||
| SJB | 5 | 盖度 Coverage (%) | 47 | 25 | 27 | W | 32-35 | 3 958 | 44 | |
| 6 | 平均高度 Average height (m) | 12 | 2.0 | 0.2 | 27-30 | 3 793 | 45 | |||
| 295 | 7 | 盖度 Coverage (%) | 60 | 50 | 40 | NW | 30-33 | 3 974 | 43 | |
| 8 | 平均高度 Average height (m) | 10 | 4.0 | 0.3 | 30-33 | 3 812 | 45 | |||
| JBG | 9 | 盖度 Coverage (%) | 65 | 25 | 10 | NE | 35-38 | 4 008 | 45 | |
| 10 | 平均高度 Average height (m) | 13 | 5.0 | 0.1 | 30-35 | 3 780 | 45 |
N, 北坡; NE, 东北坡; NW, 西北坡; W, 西坡。JBG, 夹壁沟样点; SHG, 珊瑚沟样点; SJB, 山脚坝样点。AP, 岷江冷杉原始林; BA, 红桦-岷江冷杉针阔混交林; BB, 红桦阔叶林。272、295和189为样点所在林班的编号, 1-10为采样样点编号。
N, north slope aspect; NE, northeast slope aspect; NW, northwest slope aspect; W, west slope aspect. JBG, the Jiabigou site; SHG, the Shanhugou site; SJB, the Shanjiaoba site. AP, Abies fargesii var. faxoniana primary forest; BA, Betula albosinensis - Abies fargesii var. faxoniana needleleaf-broadleaf forest; BB, Betula albosinensis broadleaf forest. 272, 295, and 189 are forest compartment numbers in the Miyaluo forest region, and 1-10 are the sample site codes.
图1
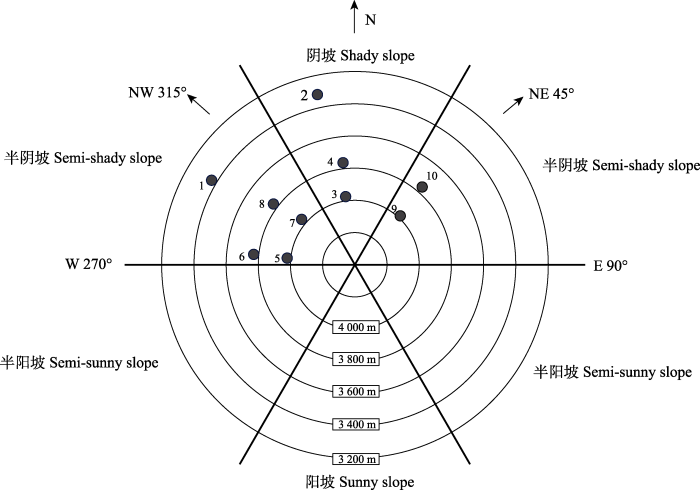
图1
川西亚高山岷江冷杉幼苗样点位置相对关系。E, 东坡; N, 北坡; NE, 东北坡; NW, 西北坡; W, 西坡。1-10为采样样点编号, 详细信息见
Fig. 1
Relative sample site locations of Abies fargesii var. faxoniana seedlings in subalpine area of western Sichuan. E, east slope aspect; N, north slope aspect; NE, northeast slope aspect; NW, northwest slope aspect; W, west slope aspect. 1-10 are sample site codes, detailed information see
1.2 岷江冷杉幼苗生物量的获取和生境调查
为了探究地形和微生境因素对幼苗生物量的影响, 本研究在3个恢复阶段林分样点内, 选取年龄≤20年(查数轮枝法确定)的岷江冷杉幼苗开展其所处生境因子调查。调查的生境因子包括: 微生境因子、基质类型(土壤、倒木、伐桩和岩石)、冠层盖度、草本盖度及高度、灌木盖度及高度、箭竹(Fargesia)盖度及高度、苔藓盖度及厚度和凋落物盖度及厚度(表2); 地形因子包括坡向和海拔(每株幼苗测量)。微生境因子的调查以获取生物量样本的目标苗木为中心拉取小样方进行, 样方大小为2 m × 2 m, 样方数量与幼苗样本数量对应。
表2 不同林型内岷江冷杉幼苗所处微生境的特征
Table 2
| 变量 Variable | 缩写 Abbreviation | 林型 Forest type | 平均值±标准误 Mean ± SE |
|---|---|---|---|
| 郁闭度 Canopy coverage (%) | CC | AP | 46 ± 2c |
| BA | 59 ± 2b | ||
| BB | 72 ± 1a | ||
| 灌木盖度 Shrub coverage (%) | SC | AP | 18 ± 1a |
| BA | 7 ± 2b | ||
| BB | 9 ± 2b | ||
| 灌木高度 Shrub height (m) | SH | AP | 0.43 ± 0.04b |
| BA | 0.88 ± 0.26a | ||
| BB | 0.32 ± 0.05b | ||
| 箭竹盖度 Fargesia coverage (%) | FC | AP | - |
| BA | 29 ± 3a | ||
| BB | 4 ± 1b | ||
| 箭竹高度 Fargesia height (m) | FH | AP | - |
| BA | 1.38 ± 0.16a | ||
| BB | 0.22 ± 0.03b | ||
| 草本盖度 Herbage coverage (%) | HC | AP | 22 ± 1c |
| BA | 34 ± 3a | ||
| BB | 27 ± 4b | ||
| 草本高度 Herbage height (m) | HH | AP | 0.08 ± 0.01b |
| BA | 0.23 ± 0.01a | ||
| BB | 0.17 ± 0.02a | ||
| 苔藓盖度 Moss coverage (%) | MC | AP | 88 ± 1a |
| BA | 44 ± 4b | ||
| BB | 32 ± 5b | ||
| 苔藓厚度 Moss thicknesses (cm) | MT | AP | 6.62 ± 0.19a |
| BA | 2.35 ± 0.16b | ||
| BB | 2.32 ± 0.24b | ||
| 凋落物盖度 Litter coverage (%) | LC | AP | 25 ± 1b |
| BA | 48 ± 2a | ||
| BB | 22 ± 2b | ||
| 凋落物厚度 Litter thicknesses (cm) | LT | AP | 1.26 ± 0.07b |
| BA | 2.73 ± 0.17a | ||
| BB | 2.64 ± 0.29a |
AP, 岷江冷杉原始林; BA, 红桦-岷江冷杉针阔混交林; BB, 红桦阔叶林。不同小写字母代表不同林型间差异显著(p < 0.05)。
AP, Abies fargesii var. faxoniana primary forest; BA, Betula albosinensis - Abies fargesii var. faxoniana needleleaf-broadleaf forest; BB, Betula albosinensis broadleaf forest. Different lowercase letters indicate significant difference among different forest types (p < 0.05).
生境调查后, 对岷江冷杉幼苗植株进行生物量的全收获。收获时用锄头整株挖取, 去除根系大部分泥土, 装入塑料袋做好标记带回, 置于阴凉处。采样当天在实验室将幼苗根部泥土及其他杂质除去后, 将幼苗从基部土痕处切开, 分为地上部分和地下部分, 地上部分又分为叶和茎。将各部分样品编号并装入信封袋在通风干燥箱中以70 ℃烘24 h至恒质量后, 在电子天平上分别测量各部分干质量。共获取幼苗生物量样本472份(表1)。
1.3 数据处理与统计分析
1.3.1 岷江冷杉幼苗的生物量分配格局
用根生物量分配比例(RMF)、茎生物量分配比例(SMF)和叶生物量分配比例(LMF)分别表示光合产物向根、茎和叶的分配量占总生物量的比例(Poorter & Nagel, 2000); 用平均年生物量增量(AMI), 即生物量总量/年龄表示幼苗平均每年的光合产物积累量, 反映不同恢复阶段林分中幼苗生物量的分配格局。
采用Kolmogorov-Smirnov检验RMF、SMF、LMF和AMI数据是否符合正态分布(α = 0.05)。若不符合正态分布, 则通过Kruskal-Wallis非参数检验比较不同恢复阶段林分间幼苗生物量分配格局的差异,并对样本进行多重比较(α = 0.05)。对所有生物量数据进行以10为底的对数转换, 使其满足标准化正态分布, 再使用回归分析探究生物量随年龄变化的趋势, 并探索这些关系在不同生境中的差异。统计分析使用SPSS 26.0软件完成。
1.3.2 岷江冷杉幼苗生物量的异速生长
为了定量描述个体发育(植物生长)过程中器官生物量分配(资源分配)的速率在不同恢复阶段生境的差异, 采用标准主轴(SMA)回归进行器官-器官生物量的异速生长分析。异速生长关系表示为
化常数,
1.3.3 各生境因子对岷江冷杉幼苗生物量积累的作用分析
首先, 采用冗余分析(RDA)量化所有生境因子对岷江冷杉幼苗生物量积累和分配的影响。RDA分析包含两个数据矩阵(连续型变量均进行了Z-score标准化): 物种矩阵(RMF、SMF、LMF和AMI数据)和生境矩阵(地形因子与微生境因子数据), 利用R 4.3.1中的“vegan”包完成分析。进一步使用Lai等(2022)提出的“rdacca.hp”包提取众多生境因子中单个生境变量对生物量积累和分配的相对贡献率。其次, 分林型采用逐步回归筛选出众多环境因子中最显著的生境因子进行多元线性回归分析, 探索不同林型内生境因子对生物量分配或积累的作用差异。由于岷江冷杉原始林样点包含不同海拔、坡向共8个样点, 考虑样点的影响对岷江冷杉原始林生物量数据采用线性混合模型(LMM)完成回归分析, 将样点编号作为随机效应, 所有生境因子作为固定效应, 通过R 4.3.1中的“lme4”包实现。红桦阔叶林和红桦-岷江冷杉针阔混交林内数据则采用一般线性模型(GLM)完成分析(生境因子仅包含微生境)。
2 结果和分析
2.1 不同恢复阶段林型中岷江冷杉幼苗的生物量分配格局
在红桦阔叶林中岷江冷杉幼苗的AMI (0.82 g·a-1)显著高于红桦-岷江冷杉针阔混交林(0.39 g·a-1)与岷江冷杉原始林(0.40 g·a-1, 图2A)。在红桦阔叶林中, 幼苗各器官生物量的分配比例呈现RMF < SMF < LMF的趋势; 红桦-岷江冷杉针阔混交林中, 幼苗的RMF显著低于茎和叶, SMF与LMF之间没有显著差异; 岷江冷杉原始林中, 幼苗各器官生物量的分配比例表现为LMF < RMF < SMF (图2B)。在3种林型之间, 幼苗的RMF表现为岷江冷杉原始林(30.3%) >红桦-岷江冷杉针阔混交林(25.9%) >红桦阔叶林(22.3%, 图2B)。岷江冷杉原始林中幼苗的SMF (43.3%)显著大于红桦-岷江冷杉针阔混交林(36.7%)和红桦阔叶林(35.5%), 而LMF (26.4%)则显著低于另2种林型(图2B)。
图2
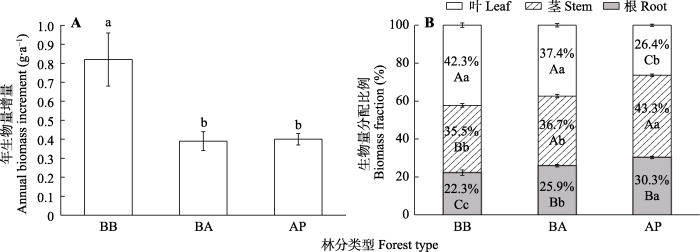
图2
不同林型岷江冷杉幼苗生物量年增量(A)和器官生物量比例(B) (平均值±标准误)。不同大写字母表示不同器官间差异显著(p < 0.05), 不同小写字母表示不同林型间差异显著(p < 0.05)。AP, 岷江冷杉原始林; BA, 红桦-岷江冷杉针阔混交林; BB, 红桦阔叶林。
Fig. 2
Annual biomass increment (A) and organ biomass fraction (B) of Abies fargesii var. faxoniana seedlings in different forest types (mean ± SE). Different uppercase letters indicate significant differences between organs (p < 0.05), different lowercase letters indicate significant differences between the forest types (p < 0.05). AP, Abies fargesii var. faxoniana primary forest; BA, Betula albosinensis - Abies fargesii var. faxoniana needleleaf-broadleaf forest; BB, Betula albosinensis broadleaf forest.
随着幼苗年龄的增加, 岷江冷杉幼苗总生物量增长速率始终表现为红桦阔叶林>红桦-岷江冷杉针阔混交林>岷江冷杉原始林(图3)。
图3
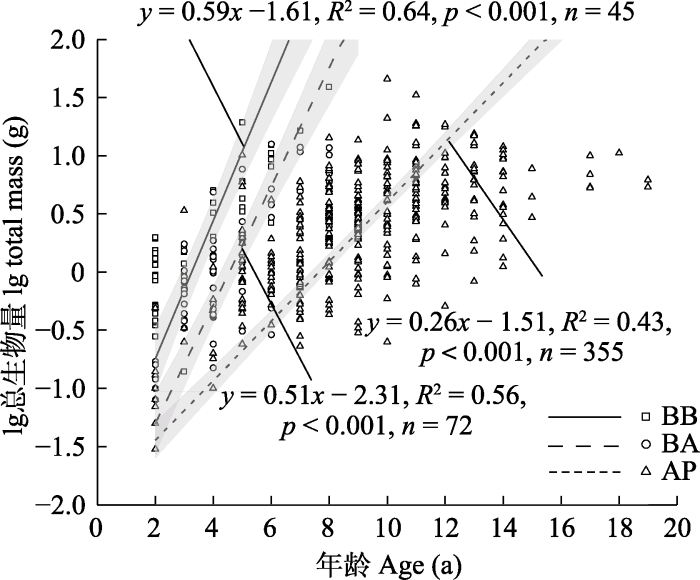
图3
不同林型岷江冷杉幼苗的总生物量与年龄的关系。所有回归模型均在α = 0.001水平上显著。AP, 岷江冷杉原始林; BA, 红桦-岷江冷杉针阔混交林; BB, 红桦阔叶林。
Fig. 3
Relationships between total seedling biomass and seedling age of Abies fargesii var. faxoniana in different forest types. All regression models were significant at the α = 0.001 level. AP, Abies fargesii var. faxoniana primary forest; BA, Betula albosinensis - Abies fargesii var. faxoniana needleleaf-broadleaf forest; BB, Betula albosinensis broadleaf forest.
2.2 不同恢复阶段林型中岷江冷杉幼苗器官间的异速生长关系
在红桦阔叶林中, 幼苗叶-根、茎-根和叶-茎之间的异速生长指数(即回归斜率α)与1.0均无显著差异(图4, p > 0.05), 各器官生物量间均表现为等速生长。在红桦-岷江冷杉针阔混交林和岷江冷杉原始林中, 幼苗茎-根之间的异速生长指数显著大于1.0 (p < 0.001), 而叶-茎之间的异速生长指数则显著小于1.0 (p < 0.001), 这说明在红桦-岷江冷杉针阔混交林和岷江冷杉原始林中茎生物量的增长速率显著大于根, 叶生物量的增长速率显著小于茎; 但两林型中的幼苗叶-根之间的异速生长指数与1.0差异不显著(p > 0.05), 两者呈现等速生长关系。
图4
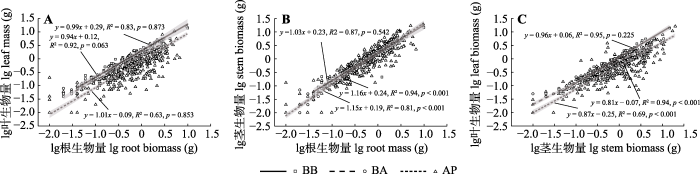
图4
不同林型岷江冷杉幼苗各器官间(A, 叶-根; B, 茎-根; C, 叶-茎)生物量的异速生长方程。所有回归模型均在α = 0.001水平上显著。p表示斜率与1.0的差异显著性。AP, 岷江冷杉原始林; BA, 红桦-岷江冷杉针阔混交林; BB, 红桦阔叶林。
Fig. 4
Allometric relationships for different dimensions (A, leaf-root; B, stem-root; C, leaf-stem) of Abies fargesii var. faxoniana seedlings in different forest types. All regression models were significant at the α = 0.001 level. p indicates a significant difference in slope with 1.0. AP, Abies fargesii var. faxoniana primary forest; BA, Betula albosinensis - Abies fargesii var. faxoniana needleleaf-broadleaf forest; BB, Betula albosinensis broadleaf forest.
2.3 生境因子对岷江冷杉幼苗生物量积累和分配的影响
2.3.1 生境因子对岷江冷杉幼苗生物量影响的综合分析
图5
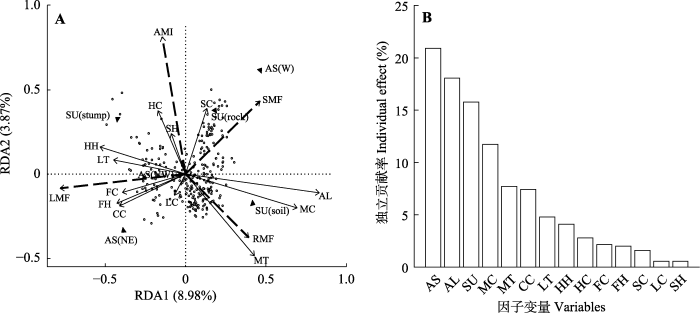
图5
岷江冷杉幼苗生物量与各生境因子的冗余分析(RDA) (A)和单个因子独立贡献率(B)。该模型在α = 0.001水平上显著。AL, 海拔; AMI, 平均年生物量增量; AS, 坡向; CC, 冠层盖度; FC, 箭竹盖度; FH, 箭竹高度; HC, 草本盖度; HH, 草本高度; LC, 凋落物盖度; LMF, 叶生物量分配比例; LT, 凋落物厚度; MC, 苔藓盖度; MT, 苔藓厚度; RMF, 根生物量分配比例; SC, 灌木盖度; SH, 灌木高度; SMF, 茎生物量分配比例; SU, 基质类型。log, 倒木; rock, 岩石; soil, 土壤; stump, 树桩。N, 北坡; NE, 东北坡; NW, 西北坡; W, 西坡。
Fig. 5
Redundancy analysis (RDA) diagram of all factors and biomass of Abies fargesii var. faxoniana seedlings (A) and the relative importance of individual factors (B). The model was significant at the α = 0.001 level. AL, altitude; AMI, annual biomass increment; AS, aspect slope; CC, canopy coverage; FC, Fargesia coverage; FH, Fargesia height; HC, herbage overage; HH, herbage height; LC, litter coverage; LMF, leaf biomass fraction; LT, litter thickness; MC, moss coverage; MT, moss thickness; RMF, root biomass fraction; SC, shrub coverage; SH, shrub height; SMF, stem biomass fraction; SU, substrate type. N, north slope aspect; NE, northeast slope aspect; NW, northwest slope aspect; W, west slope aspect.
14个因子的独立贡献率中, 坡向(20.9%)对生物量积累和分配的贡献率大于海拔(18.1%), 坡向、海拔和苗床基质类型3个因子的贡献率总计达54.7%,是影响生物量的主要影响因子(图5B)。微生境因子中, 基质类型(15.8%)、苔藓盖度(11.7%)及厚度(7.7%)和冠层盖度(7.4%)是对岷江冷杉幼苗生物量积累和分配累积贡献度前四的因子。
2.3.2 不同恢复阶段林型中生境因子对生物量积累和分配的影响
在海拔跨度约200 m的岷江冷杉原始林中, 海拔对生物量的影响变得不显著; 此时, 坡向仍然是影响显著的因子之一(表3, p < 0.05)。其中, 西北坡对岷江冷杉幼苗RMF具有显著的正效应, 西坡相对于其他坡向来说对幼苗的SMF具有显著的正效应(p < 0.001), 西北坡和西坡相对于其他坡向而言对LMF具有显著的负效应(p < 0.05)。总体来看, 西坡对岷江冷杉幼苗AMI比其他坡向具有显著的正效应 (p = 0.009)。在红桦-岷江冷杉针阔混交林中, 苔藓盖度对岷江冷杉幼苗的RMF具有显著正效应(p = 0.002), 而苔藓厚度对其具有显著的负效应(p = 0.044)。在红桦阔叶林中, 草本高度对岷江冷杉幼苗的RMF具有显著的正效应(p = 0.018); 草本盖度对LMF具有显著的负效应(p < 0.05); 冠层盖度和草本高度对AMI具有显著的负效应(p < 0.05), 而箭竹盖度对其具有显著的正效应(表3, p = 0.002)。
表3 不同林分中岷江冷杉幼苗生物量与各生境因子的关系
Table 3
| 林分类型 Forest type | 响应变量 Response variable | 解释变量 Explanatory variable | 参数 Estimate | 标准误 SE | t | p |
|---|---|---|---|---|---|---|
| 岷江冷杉原始林 Abies fargesii var. faxoniana primary forest | RMF | AS (NW) | 0.44 | 0.15 | 2.93 | 0.004** |
| SU (soil) | 0.72 | 0.23 | 3.06 | 0.002** | ||
| SH | -0.17 | 0.08 | -2.06 | 0.041* | ||
| HC | -0.14 | 0.07 | -2.01 | 0.045* | ||
| MT | 0.25 | 0.08 | 3.22 | 0.001** | ||
| SMF | AS (W) | 0.51 | 0.14 | 3.52 | <0.001*** | |
| LMF | AS (NW) | -0.47 | 0.17 | -2.68 | 0.030* | |
| AS (W) | -0.58 | 0.18 | -3.12 | 0.013* | ||
| SU (rock) | -0.90 | 0.35 | -2.54 | 0.012* | ||
| SU (soil) | -0.64 | 0.23 | -2.80 | 0.005** | ||
| MT | -0.19 | 0.07 | -2.75 | 0.008** | ||
| AMI | AS (W) | 0.67 | 0.20 | 3.33 | 0.009** | |
| SH | 0.20 | 0.08 | 2.51 | 0.012* | ||
| 红桦-岷江冷杉针阔混交林 Betula. albosinensis - Abies fargesii var. faxoniana needleleaf-broadleaf forest | RMF | MC | 0.49 | 0.15 | 3.27 | 0.002** |
| MT | -0.65 | 0.31 | -2.06 | 0.044* | ||
| SMF | ns | |||||
| LMF | ns | |||||
| AMI | ns | |||||
| 红桦阔叶林 Betula albosinensis broadleaf forest | RMF | HH | 0.34 | 0.14 | 2.48 | 0.018* |
| SMF | ns | |||||
| LMF | HC | -0.35 | 0.15 | -2.24 | 0.031* | |
| AMI | CC | -0.35 | 0.15 | -2.40 | 0.021* | |
| FC | 1.59 | 0.49 | 3.24 | 0.002** | ||
| HH | -0.29 | 0.12 | -2.41 | 0.020* | ||
所有模型在α = 0.05水平上显著; ns, p > 0.05; *, p < 0.05; **, p < 0.01; ***, p < 0.001。AMI, 平均年生物量增量; AS, 坡向; FC, 箭竹盖度; CC, 冠层盖度; HC, 草本盖度; HH, 草本高度; LMF, 叶生物量分配比例; MC, 苔藓盖度; MT, 苔藓厚度; RMF, 根生物量分配比例; SH, 灌木高度; SMF, 茎生物量分配比例; SU, 基质类型。log, 倒木; rock, 岩石; soil, 土壤; stump, 树桩。N, 北坡; NE, 东北坡; NW, 西北坡; W, 西坡。
All models were significant at the α = 0.05 level; ns, p > 0.05; *, p < 0.05; **, p < 0.01; ***, p < 0.001. AMI, annual biomass increment; AS, aspect slope; CC, canopy coverage; FC, Fargesia coverage; HC, herbage overage; HH, herbage height; LMF, leaf biomass fraction; MC, moss coverage; MT, moss thickness; RMF, root biomass fraction; SH, shrub height; SMF, stem biomass fraction; SU, substrate type. N, north slope aspect; NE, northeast slope aspect; NW, northwest slope aspect; W, west slope aspect.
3 讨论
3.1 岷江冷杉幼苗适应不同恢复阶段林型生境的生物量积累和分配
本研究中, 高海拔岷江冷杉原始林中岷江冷杉幼苗的根和茎生物量占比显著高于低海拔次生林, 而次生林中幼苗的叶生物量占比则显著高于岷江冷杉原始林(图2B), 这与Bernoulli和Körner (1999)的高海拔山赤松的茎枝生物量占总生物量的比例增加的研究结果一致。高海拔区域的寒冷土壤养分供应少、流动性差, 树木采取增加根比例的生长策略来促进高寒地区低营养土壤的养分吸收(Reich et al., 2014)。另外, 高海拔树木幼苗变矮变粗, 发出更多分枝, 来抵御外部强风、积雪和霜冻环境(Li et al., 2003)。幼苗向茎和根增大生物量投入以适应高寒生境, 这种调整可以减缓低温对生物量积累的抑制(Köerner, 2012)和冻害对新叶造成的机械损伤(邵佳怡等, 2019)。此外, 岷江冷杉原始林中岷江冷杉幼苗的总生物量积累慢于红桦-岷江冷杉针阔混交林和红桦阔叶林(图3), 这可能是两种次生林所处海拔较低, 水热条件优越, 幼苗的生长季较高海拔原始林更长所导致(Coll & Ameztegui, 2019)。
在本研究中, 两种次生林中岷江冷杉幼苗的生物量分配比例相似, 均表现为叶>茎>根。但是, 红桦阔叶林中幼苗的平均年生物量增量和根生物量占比显著高于红桦-岷江冷杉针阔混交林(图2)。红桦-岷江冷杉针阔混交林中箭竹盖度(29.1%)与高度(1.38 m)均显著高于红桦阔叶林。已有研究表明, 箭竹通过地下茎的无性繁殖蔓延扩张, 并通过其密集交错的秆枝和根系与岷江冷杉幼苗争夺光照、水分、养分等资源(Taylor & Qin, 1988; Taylor et al., 2006; 马姜明等, 2009)。因此, 这种抑制可能是针阔混交林中幼苗生物量增长速率较慢的主要原因。此外, 红桦-岷江冷杉针阔混交林中幼苗所处生境的草本盖度(33.6%)也显著高于红桦阔叶林(27.4%), 过密的草本植物也可能会导致岷江冷杉幼苗生长受到抑制(庞晓瑜等, 2018)。
3.2 地形和微生境因子对岷江冷杉幼苗生物量影响的相对贡献
地形条件是影响高山森林树种生物量积累和分配的重要因素。其中, 高海拔促进了亚高山针叶树幼苗根茎生物量的积累已被广泛揭示(Reich et al., 2014; Coll & Ameztegui, 2019), 而坡向对其生物量变化的影响研究相对较少。坡向可以对水热进行二次分配, 改变凋落物分解和土壤养分循环速率, 从而间接影响树木生物量的分配(Hicks Jr & Frank Jr, 1984; Thakur et al., 2019)。Zhang等(2022)在藏东南色季拉山的研究发现, 海拔和坡向的单一作用对急尖长苞冷杉幼苗的生物量影响不显著, 但其与年龄的交互作用显著, 意味着坡向和海拔均对生物量有较强的间接效应。本研究发现, 随着海拔跨度减小至200 m时, 坡向的影响依然存在(表3; 图5)。西坡生境中幼苗平均年生物量积累最高, 这可能是西坡相对于其他坡向生境更偏阳, 幼苗能够获取的光照更多。光照不仅直接关系到幼苗自身的光合与蒸腾, 还会影响环境中的湿度和温度, 进而促进或限制幼苗的碳积累(邵佳怡等, 2019)。
在幼苗微地形或者微生境尺度上, 腐烂树桩、倒木、岩石等覆盖在地表, 使林地出现复杂而独特的微地形, 这些独特的微地形地表在许多特性上与森林地面不同, 例如含水量(Simon et al., 2011)、温度(Beatty & Sholes, 1988)、微生物群落组成(Šamonil et al., 2020)、凋落物组成和数量(Simon et al., 2011)。在没有或只有较少凋落物的地方, 裸露的基质(木材或土壤)可以暴露出来或覆盖上苔藓, 这也会影响幼苗的定居和生长(Wang et al., 2017)。本研究发现, 基质类型(15.8%)、苔藓盖度及厚度(19.5%)和冠层盖度(7.4%)是多个微生境因子中对岷江冷杉幼苗生物量的影响贡献度前四的因子(图5B)。Inman-Narahari等(2014)在热带山地雨林中的研究发现, 基质类型(35%-37%)和光照(11%-12%)是仅次于地形(51%-53%)对木本植物幼苗相对生长速率贡献较大的因子。基质类型之中, 岩石和树桩相比其他类型更有利于岷江冷杉幼苗年均生物量的增长(图5A), 这可能是因为岩石和树桩往往是森林地表的凸起生境。相比在倒木基质上, 幼苗定居在土壤上不利于其生长和存活, 因为土壤中抑制针叶树幼苗存活的病原菌感染更为严重, 且土壤上的幼苗更易被下层植被遮蔽, 尤其是在混交林中, 土壤上的幼苗可能会被较厚的阔叶凋落物掩埋(Narukawa et al., 2003; Narukawa & Yamamoto, 2003; Mori et al., 2004)。树桩基质的特点与倒木基质类似, 随着木材的腐烂, 营养和水分条件变得与土壤相似, 增加了幼苗被下层植被和阔叶凋落物遮蔽和掩埋以及感染致病真菌的可能性(Mori et al., 2004)。川西亚高山的类似研究发现, 幼苗在地表植被丰富、竞争激烈的林内经常偏好林地凸起生境, 以减少箭竹等林下灌木对其的影响(Taylor et al., 2006; 马姜明等, 2009)。在红桦-岷江冷杉混交林中, 虽然苔藓盖度(约44%)促进根生物量的积累, 但苔藓厚度(约2 cm)对其产生负效应(表3)。苔藓对幼苗建立的积极作用一般表现为其对种子的庇护作用(Jeschke & Kiehl, 2008)、对土壤水分的保持(Parker et al., 1997; Zamfir, 2000)和土壤温度的缓和作用(Zamfir, 2000)。然而, 地表苔藓层过厚反而会阻碍幼苗根系与土壤的接触, 不利于根系获取水和养分(Freestone, 2006)。Wang等(2017)发现生长在祁连山高海拔地区的青海云杉在苔藓厚度4-5 cm、盖度约50%时, 其更新幼苗的数量最多。对岷江冷杉幼苗, 苔藓盖度适中时促进其根系生物量的积累, 混交林中苔藓厚度过薄(2.35 cm), 可能不足以为幼苗根系生长起到缓冲土壤温度等的作用。
4 结论
红桦阔叶林中岷江冷杉幼苗的生物量积累速率(生长速率)最高, 随着林型向红桦-岷江冷杉针阔混交林正向演替过程中趋于降低。高海拔林型中岷江冷杉幼苗的茎生物量占比最高(43.7%), 低海拔次生林中幼苗的叶生物量占比最高(42.3%和37.4%)。在红桦阔叶林中, 各器官生物量间均表现为等速生长, 而在红桦-岷江冷杉针阔混交林和岷江冷杉原始林中, 茎生物量的增长速率显著大于根, 叶生物量的增长速率显著小于茎。相比于东北、北和西北坡, 西坡生境中的岷江冷杉幼苗年均生物量的积累最高。坡向(20.9%)对幼苗生物量积累和分配贡献率大于海拔(18.1%), 基质类型(15.8%)、苔藓盖度(11.7%)及厚度(7.7%)和冠层盖度(7.4%)是微生境因子中对岷江冷杉幼苗生物量累积贡献度前四的因子。针对岷江冷杉幼苗所在林分的地形和微生境特点, 可以在疏伐、抚育等森林经营活动中保留伐桩、倒木以创建幼苗更新偏好的地表凸起类基质生境, 以促进幼苗的定居和生长。可对冠层盖度较高的幼苗生境地段开展箭竹清除、透光抚育等措施, 促进幼苗的生长。
参考文献
Geological and climatic influences on mountain biodiversity
DOI:10.1038/s41561-018-0236-z
[本文引用: 1]

Mountains are key features of the Earth's surface and host a substantial proportion of the world's species. However, the links between the evolution and distribution of biodiversity and the formation of mountains remain poorly understood. Here, we integrate multiple datasets to assess the relationships between species richness in mountains, geology and climate at global and regional scales. Specifically, we analyse how erosion, relief, soil and climate relate to the geographical distribution of terrestrial tetrapods, which include amphibians, birds and mammals. We find that centres of species richness correlate with areas of high temperatures, annual rainfall and topographic relief, supporting previous studies. We unveil additional links between mountain-building processes and biodiversity: species richness correlates with erosion rates and heterogeneity of soil types, with a varying response across continents. These additional links are prominent but under-explored, and probably relate to the interplay between surface uplift, climate change and atmospheric circulation through time. They are also influenced by the location and orientation of mountain ranges in relation to air circulation patterns, and how species diversification, dispersal and refugia respond to climate change. A better understanding of biosphere-lithosphere interactions is needed to understand the patterns and evolution of mountain biodiversity across space and time.
Seedling recruitment, survival and facilitation in alpine Pinus uncinata tree line ecotones. Implications and potential responses to climate warming
Leaf litter effect on plant species composition of deciduous forest treefall pits
Dry matter allocation in treeline trees
Elevation modulates the phenotypic responses to light of four co-occurring Pyrenean forest tree species
Facilitation drives local abundance and regional distribution of a rare plant in a harsh environment
The importance of facilitation to local community dynamics is becoming increasingly recognized. However, the predictability of positive interactions in stressful environments, the balance of competition and facilitation along environmental gradients, and the scaling of local positive interactions to regional distributions are aspects of facilitation that remain unresolved. I explored these questions in a habitat specialist, Delphinium uliginosum, and a moss, Didymodon tophaceus, both found in small serpentine wetlands. I tested three hypotheses: (1) moss facilitates germination, growth, and/or fecundity of D. uliginosum; (2) facilitation is stronger at the harsher ends of gradients in soil moisture, toxicity, and/or biomass; and (3) facilitation is reflected in positive associations at the levels of local abundance and regional occurrence. Although considerable competitive interactions occurred in later life stages, moss strongly facilitated D. uliginosum seedling emergence. There was no evidence that this facilitative effect weakened, or switched to competition, in benign environments. D. uliginosum was more locally abundant and more frequently present, across a large portion of its range, with than without moss, indicating a net facilitative effect in the face of competitive influences. Facilitated recruitment, possibly by seed retention, was found to be an important control on abundance and distribution in this rare species.
Relationship of aspect to soil nutrients, species importance and biomass in a forested watershed in West Virginia
The influence of tree islands and microtopography on pedoecological conditions in the forest-alpine tundra ecotone on Niwot Ridge, Colorado front range, U.S.A.
Characteristics of biomass and carbon stock of fir and birch fine roots in subalpine forest of western Sichuan, China
川西亚高山冷杉和白桦细根生物量与碳储量特征
Trade-offs in seedling growth and survival within and across tropical forest microhabitats
DOI:10.1002/ece3.1196
PMID:25614790
[本文引用: 1]
For niche differences to maintain coexistence of sympatric species, each species must grow and/or survive better than each of the others in at least one set of conditions (i.e., performance trade-offs). However, the extent of niche differentiation in tropical forests remains highly debated. We present the first test of performance trade-offs for wild seedlings in a tropical forest. We measured seedling relative growth rate (RGR) and survival of four common native woody species across 18 light, substrate, and topography microhabitats over 2.5 years within Hawaiian montane wet forest, an ideal location due to its low species diversity and strong species habitat associations. All six species pairs exhibited significant performance trade-offs across microhabitats and for RGR versus survival within microhabitats. We also found some evidence of performance equivalence, with species pairs having similar performance in 26% of comparisons across microhabitats. Across species, survival under low light was generally positively associated with RGR under high light. When averaged over all species, topography (slope, aspect, and elevation) explained most of the variation in RGR attributable to microhabitat variables (51-53%) followed by substrate type (35-37%) and light (11-12%). However, the relative effects of microhabitat differed among species and RGR metric (i.e., RGR for height, biomass, or leaf area). These findings indicate that performance trade-offs among species during regeneration are common in low-diversity tropical forest, although other mechanisms may better explain the coexistence of species with small performance differences.
Effects of a dense moss layer on germination and establishment of vascular plants in newly created calcareous grasslands
Treelines will be understood once the functional difference between a tree and a shrub is.
Carbon allocation among tree organs: a review of basic processes and representation in functional-structural tree models
Generalizing hierarchical and variation partitioning in multiple regression and canonical analyses using the rdacca.hp R package
Effects of microsite on growth of Pinus cembra in the subalpine zone of the Austrian Alps
Growth responses of Picea abies and Larix decidua to elevation in subalpine areas of Tyrol, Austria
Effects of biotic neighbors and habitat heterogeneity on tree seedling survival in a secondary mixed conifer and broad-leaved forest in Changbai Mountain
DOI:10.17521/cjpe.2015.0366
[本文引用: 1]
Aims Our study aimed to understand the effects and the relative importance of biotic neighborhood and habitat heterogeneity for tree seedling survival in a secondary mixed conifer and broad-leaved forest in Changbai Mountain, north-eastern China. Methods The generalized mixed linear model was used to examine the relative effects of biotic neighborhood and habitat heterogeneity on seedling survival over two years. Important findings Our results showed that both biotic neighborhood and habitat heterogeneity had significant effects on the seedling survival at community level. The local environment suitable for the adult growth was also suitable for seedling survival. The soil moisture and soil available nitrogen exhibited significant positive effects on seedling survival. On the other hand, seedling density had significant negative effects on seedling survival due to the individual competition. Particularly, we found significant negative density-dependent effects on seedling survival which was caused by conspecific adult and seedling neighbors. As expected, with the increasing of seedling survival age, the habitat heterogeneity became more important on seedling survival. These results suggest that both local biotic neighborhood and habitat heterogeneity drive seedling survival in this temperate forest, and their relative importance varies with different seedling age classes and species traits.
长白山次生针阔混交林乔木幼苗存活的影响因素分析
DOI:10.17521/cjpe.2015.0366
[本文引用: 1]
基于长白山次生针阔混交林样地, 以520个1 m × 1 m幼苗样方中胸径小于1 cm的乔木幼苗为研究对象, 选取2013年和2014年的幼苗调查数据, 运用广义线性混合模型(GLMM)分析了生物邻体和生境异质性对幼苗存活的影响, 探讨了次生针阔混交林幼苗存活影响因素及物种共存机制。结果表明: (1)适宜大树生长的局域生境同样也适宜幼苗的存活, 幼苗存活率与土壤含水量和有效氮等土壤养分显著正相关。(2)幼苗个体之间存在明显的竞争, 较多的幼苗邻体显著降低幼苗的存活率。同种大树邻体和同种幼苗邻体与幼苗存活显著负相关, 表明存在负密度制约效应。(3)随着幼苗年龄的增加, 生境异质性对幼苗存活的影响逐渐增大。该研究证实了密度制约效应和生境异质性对幼苗存活有着重要影响, 其相对重要性随着幼苗年龄级、功能群以及物种种类而变化。
Fine root biomass, productivity and turnover of Abies faxoniana primary forest in subalpine region of western Sichuan, China
川西亚高山岷江冷杉原始林细根生物量、生产力和周转
原始林在全球碳收支中具有重要的意义,其细根在地下碳循环过程中发挥着重要作用。本研究采用连续土钻法对川西亚高山岷江冷杉原始林0~30 cm土层细根(≤2 mm)生物量及其季节动态进行了测定,并采用决策矩阵法对细根生产量和周转速率进行了估算。结果表明:岷江冷杉原始林0~30 cm土层活细根生物量和年生产量分别为286.89 g·m<sup>-2</sup>和168.94 g·m<sup>-2</sup>·a<sup>-1</sup>,平均细根周转速率为0.56 a<sup>-1</sup>;细根生物量、生产量和死亡量在生长期内具有明显的动态特征;活细根生物量和生产量总体呈现单峰曲线特征,以9月最大;死细根生物量和死细根/活细根生物量在生长期内总体呈“U型”变化趋势,而死亡量总体呈增加的趋势;土层深度是影响细根动态的重要因素,活细根生物量、死细根生物量、生产量和周转速率随着土层深度的增加呈现下降的变化趋势,而死细根/活细根生物量略有增加。
Root biomass in the restoration process of subalpine dark coniferous forests in western Sichuan, China
川西亚高山暗针叶林恢复过程中根系生物量研究
Natural regeneration of Abies faxoniana along restoration gradients of subalpine dark coniferous forest in western Sichuan, China
DOI:10.3773/j.issn.1005-264x.2009.04.003
[本文引用: 2]
<EM>Aims</EM> Subalpine dark coniferous forests in western Sichuan, China include natural old-growth forests and successional forests that regenerated mainly following logging over the past 50 years. Both the bamboo and moss forest types are comprised of three different restoration stages: broad-leaved forest 20–40 years old, conifer/broad-leaved mixed forest 50 years old and old-growth dark coniferous forest 160–200 years old. Our objectives were to understand how subalpine dark coniferous forests naturally regenerated following disturbance and determine key factors influencing natural regeneration. <BR><EM>Methods</EM> We investigated densities of seedlings, saplings and small trees of the climax dominant tree species <EM>Abies faxoniana</EM> in 50 quadrats along a gradient of the different successional stages in both the bamboo and moss forest types located at 3 100–3 600 m elevation on north or northwest aspects. <BR><EM>Important findings</EM> The densities of seedlings, saplings and small trees of <EM>A. faxoniana </EM>were generally relatively low in both the bamboo and moss forest types, but higher than that in moss forest types. Key factors enhancing natural regeneration of <EM>A. faxoniana</EM> in the bamboo forest types were density of seed-bearing trees and the stock of fallen wood. Negative factors were coverage of bamboo and thickness of moss. In the moss forest types, the coverage of shrubs and thickness of moss enhanced the occurrence of seedlings of <EM>A. faxoniana</EM> but limited the development of saplings and small trees. <BR>
川西亚高山暗针叶林恢复过程中岷江冷杉天然更新状况及其影响因子
DOI:10.3773/j.issn.1005-264x.2009.04.003
[本文引用: 2]
在川西亚高山米亚罗林区海拔3 100~3 600 m 阴坡、半阴坡, 以立地条件基本一致的箭竹和藓类林型不同恢复阶段 (20~40 a生的箭竹阔叶林、50 a生的箭竹针阔混交林、160~200 a生的箭竹原始暗针叶老龄林; 20~40 a生的藓类阔叶林、50 a生的藓类针阔混交林、160~200 a生的藓类原始暗针叶老龄林) 的群落为研究对象, 共设置了50个样方 (20 m×20 m ), 采用空间代时间的方法分析了岷江冷杉(Abies faxoniana)的天然更新状况, 并采用通径分析法对其影响因子进行分析。结果表明: 箭竹和藓类两种森林类型岷江冷杉幼苗、幼树和小树的密度偏低。对于箭竹林型不同恢复阶段, 岷江冷杉幼苗密度<幼树密度<小树密度; 对于藓类林型不同恢复阶段, 藓类阔叶林幼树密度大于幼苗和小树密度, 藓类针阔混交林小树密度大于幼苗和幼树密度, 而藓类原始暗针叶老龄林幼苗密度大于幼树和小树密度。藓类林型岷江冷杉天然更新状况好于箭竹林型。对箭竹林型而言, 影响岷江冷杉天然更新的关键因子为母树密度、倒木蓄积量、箭竹盖度和苔藓层厚度, 其中母树密度和倒木蓄积量对岷江冷杉天然更新起着促进作用, 箭竹盖度和苔藓层厚度对岷江冷杉天然更新起着阻碍作用; 对于藓类林型而言, 影响岷江冷杉天然更新的关键因子为灌木盖度和苔藓层厚度。灌木和苔藓有利于幼苗的发生, 但不利于幼苗向幼树、小树的过渡。
Substrate-associated seedling recruitment and establishment of major conifer species in an old-growth subalpine forest in central Japan
State of fallen logs and the occurrence of conifer seedlings and saplings in boreal and subalpine old-growth forests in Japan
Development of conifer seedlings roots on soil and fallen logs in boreal and subalpine coniferous forests of Japan
Effects of simulated warming and functional group removal on survival and growth of Abies faxoniana seedlings
模拟增温和功能群去除对岷江冷杉幼苗存活和生长的影响
DOI:10.13287/j.1001-9332.201803.010
[本文引用: 2]
通过模拟增温和功能群去除试验,研究不同功能群的草本植物在岷江冷杉幼苗更新定植过程的作用,分析其在温度升高条件下生理指标的变化.结果表明: 增温提高了岷江冷杉幼苗的存活率和植物体内非结构性碳水化合物含量,但抑制了幼苗的生长和根冠比.岷江冷杉幼苗的非结构性碳水化合物含量,尤其是可溶性糖含量与存活率呈显著正相关.不增温处理中,草本植物降低了幼苗的存活率,促进幼苗的株高生长和地上生物量积累,禾本科植物和杂类草抑制了幼苗根系生长和地下生物量的积累;增温处理中,杂类草的存在提高了幼苗的存活率,莎草科则显著抑制了幼苗根系生长和地下生物量的积累,禾本科植物和杂类草抑制了幼苗的高生长和地上生物量积累.模拟增温提高了林线树种岷江冷杉幼苗的存活率,但使其面临来自草本植物更剧烈的竞争,抑制其生长.
The role of hair-cap mosses (Polytrichum spp.) in natural regeneration of white spruce (Picea glauca (Moench) Voss).
Advances in the study of photosynthate allocation and its controls
植物光合产物分配及其影响因子研究进展
DOI:10.3773/j.issn.1005-264x.2010.01.013
[本文引用: 1]
植物光合产物分配受环境因子和生物因子的共同影响。为增进对植物对全球变化响应的理解, 从植物个体水平与群落/生态系统水平综述了植物光合产物分配的影响因子与影响机理的最新研究进展。植物个体在光照增强及受水分和养分胁迫时, 会将光合产物更多地分配到根系; CO<sub>2</sub>浓度升高对植物光合产物分配的影响受土壤氮素的制约, 植物受氮素胁迫时, CO<sub>2</sub>浓度升高会促进光合产物更多地分配到根系; 反之, 对植物光合产物分配没有影响。植物群落/生态系统的光合产物分配对环境因子的响应不敏感; 光合产物向根系的分配比例随其生长阶段逐渐降低。功能平衡假说、源汇关系假说和相关生长关系假说分别从环境因子、个体发育和环境因子与个体发育协同作用方面阐述了植物光合产物分配的影响机理。在此基础上,指出了未来拟重点加强的研究方向: 1)生态系统尺度的光合产物向呼吸部分的分配研究; 2)地下净初级生产力(belowground net primary productivity, BNPP)研究; 3)温室和野外条件下及幼苗和成熟林光合产物分配对环境因子响应的比较研究; 4)生态系统尺度的多因子控制试验; 5)整合环境因子和个体发育对植物光合产物分配格局的影响研究。
The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: a quantitative review
Temperature drives global patterns in forest biomass distribution in leaves, stems, and roots
DOI:10.1073/pnas.1216053111
PMID:25225412
[本文引用: 2]
Whether the fraction of total forest biomass distributed in roots, stems, or leaves varies systematically across geographic gradients remains unknown despite its importance for understanding forest ecology and modeling global carbon cycles. It has been hypothesized that plants should maintain proportionally more biomass in the organ that acquires the most limiting resource. Accordingly, we hypothesize greater biomass distribution in roots and less in stems and foliage in increasingly arid climates and in colder environments at high latitudes. Such a strategy would increase uptake of soil water in dry conditions and of soil nutrients in cold soils, where they are at low supply and are less mobile. We use a large global biomass dataset (>6,200 forests from 61 countries, across a 40 °C gradient in mean annual temperature) to address these questions. Climate metrics involving temperature were better predictors of biomass partitioning than those involving moisture availability, because, surprisingly, fractional distribution of biomass to roots or foliage was unrelated to aridity. In contrast, in increasingly cold climates, the proportion of total forest biomass in roots was greater and in foliage was smaller for both angiosperm and gymnosperm forests. These findings support hypotheses about adaptive strategies of forest trees to temperature and provide biogeographically explicit relationships to improve ecosystem and earth system models. They also will allow, for the first time to our knowledge, representations of root carbon pools that consider biogeographic differences, which are useful for quantifying whole-ecosystem carbon stocks and cycles and for assessing the impact of climate change on forest carbon dynamics.
Topographic shelter and conifer establishment and mortality in an alpine environment, Glacier National Park, Montana
Convergence, divergence or chaos? Consequences of tree trunk decay for pedogenesis and the soil microbiome in a temperate natural forest
Tree seedling distribution, regeneration mechanism and response to climate change in alpine treeline ecotone
高山林线生态交错区木本植物幼苗分布特征、更新机制及其对气候变化的响应
DOI:10.13287/j.1001-9332.201908.007
[本文引用: 2]
木本植物幼苗是高山林线生态交错区的重要组成部分,其更新对气候变化背景下树线的移动至关重要.本研究通过对近几十年来全球范围内林线生态交错区的木本植物幼苗分布特征、更新机制及其对气候变化响应的研究总结得出:林线生态交错区木本植物幼苗的空间分布类型主要为渐变型和聚集型,且不同分布类型对树线动态的指示意义各异.在全球尺度上,其分布的海拔高限通常与生长季长度、均温和物种特性等有关,而在区域尺度上则多受降水影响.在幼苗更新初期,种源在很大程度上决定了种子的萌发及分布位置,之后微环境的促进作用为幼苗的定植提供庇护,提高其存活率,而在更新后期多种生物和非生物因素及其相互作用则非常关键.气候变暖促使林线生态交错区气温升高、降水充沛,有利于幼苗生长,使其向高海拔区域扩张而成为树线上移的先兆,但部分物种受遗传特性或适应策略影响,仅表现为密度增加,使树线保持相对稳定.未来应借助树轮、<sup>14</sup>C等精确定年技术,通过长期的野外定位观测和室内模拟,加强多时空尺度下林线幼苗的空间分布特征和更新机制研究,分析不同类型林线内木本植物幼苗的适应策略,预测气候变化背景下的树线动态,为山地生态系统恢复及保护提供科学依据.
The influence of windthrow microsites on tree regeneration and establishment in an old growth mountain forest
Regeneration patterns in old-growth Abies-Betula forests in the Wolong Natural Reserve, Sichuan, China
Regeneration patterns and tree species coexistence in old-growth Abies-Picea forests in southwestern China
Assessment of biomass and net primary productivity of a dry tropical forest using geospatial technology
DOI:10.1007/s11676-018-0607-8
[本文引用: 1]
This study quantifies biomass, aboveground and belowground net productivity, along with additional environmental factors over a 2-3year period in Barnawapara Sanctuary of Chhattisgarh, India through satellite remote-sensing and GIS techniques. Ten sampling quadrates 20x20, 5x5 and 1x1m were randomly laid for overstorey (OS), understorey (US) and ground vegetation (GS), respectively. Girth of trees was measured at breast height and collar diameters of shrubs and herbs at 0.1m height. Biomass was estimated using allometric regression equations and herb biomass by harvesting. Net primary productivity (NPP) was determined by summing biomass increment and litter crop values. Aspect and slope influenced the vegetation types, biomass and NPP in different forests. Standing biomass and NPP varied from 18.6 to 101.5Mgha(-1) and 5.3 to 12.7Mgha(-1) a(-1), respectively, in different forest types. The highest biomass was found in dense mixed forest, while net production recoded in Teak forests. Both were lowest in degraded mixed forests of different forest types. OS, US and GS contributed 90.4, 8.7 and 0.7%, respectively, for the total mean standing biomass in different forests. This study developed spectral models for the estimation of biomass and NPP using Normalized Difference Vegetation Index and other vegetation indices. The study demonstrated the potential of geospatial tools for estimation of biomass and net productivity of dry tropical forest ecosystem.
Microclimate in forests with varying leaf area index and soil moisture: potential implications for seedling establishment in a changing climate
Effects of environmental variables on seedling-sapling distribution of Qinghai spruce (Picea crassifolia) along altitudinal gradients
Meta-analysis of effects of forest litter on seedling establishment
Smatr 3—An R package for estimation and inference about allometric lines
Effects of forest canopy gap on Abies faxoniana seedling’s biomass and its allocation in subalpine coniferous forest of west Sichuan
林窗对川西亚高山岷江冷杉幼苗生物量及其分配格局的影响
Responses of phenology and growth of Betula utilis and Abies faxoniana in subalpine timberline ecotone to simulated global warming, western Sichuan, China
川西亚高山林线交错带糙皮桦和岷江冷杉幼苗物候与生长对模拟增温的响应
DOI:10.3773/j.issn.1005-264x.2008.05.011
[本文引用: 1]
采用开顶式生长室(Open-top chamber, OTC)模拟增温对植被影响的研究方法, 研究了川西亚高山林线交错带糙皮桦(Betula utilis) 和岷江冷杉(Abies faxoniana)幼苗物候及生长特性对模拟增温的响应。结果表明, 温度升高使岷江冷杉幼苗芽开放时间显著提前(15.2 d); 糙 皮桦春季芽物候期变化不显著, 而落叶时间明显推迟(19.7 d), 叶寿命延长(22.8 d)。与对照(CK)相比, OTC内糙皮桦叶面积和岷江冷杉叶片长度及两者侧枝生长速率都显著加快。模拟增温对两物种基径相对生长速率都表现为正效应, 增温对两物种枝叶特性及分布格局表现为不同程度 的正效应、负效应或无影响。不同功能型两物种对模拟增温响应方式存在一定程度差异。
Effect of long-term warming on growth and biomass allocation of Abies faxoniana seedlings
长期模拟增温对岷江冷杉幼苗生长与生物量分配的影响
Warming effects on growth and physiology of seedlings of Betula albosinensis and Abies faxoniana under two contrasting light conditions in subalpine coniferous forest of western Sichuan, China
增温对川西亚高山针叶林内不同光环境下红桦和岷江冷杉幼苗生长和生理的影响
Effects of bryophytes and lichens on seedling emergence of alvar plants: evidence from greenhouse experiments
Influence of topography, soil properties and plant community on the biomass of Abies georgei var. smithii seedlings in Southeast Tibet
Litter composition and its dynamic in five main forest types in subalpine areas of west Sichuan, China
川西亚高山五种主要森林类型凋落物组成及动态
The influence factors of sub-alpine forest restoration in Miyaluo, west Sichuan
川西米亚罗林区森林恢复的影响因子分析

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}