


种子萌发是植物生长繁殖和群落更新演替的重要环节。在长期的生存进化过程中, 不同种群的植物受到不同环境条件的选择以及基因交流的限制, 产生了遗传性状的分化, 形成了适应各自生境的特征(Bucharova et al., 2017; Hamidzadeh et al., 2021), 因而同一物种不同种群间种子的大小与生理状态都存在较大差异, 进而会影响种子的萌发以及休眠状况。例如, 在对我国甘肃安西荒漠自然保护区的裸果木(Gymnocarpos przewalskii) 3个种群的种子研究中发现, 种子的长宽比、质量、吸水率、萌发势和发芽率等方面的变异系数都较大(李树森等, 2020)。在种子休眠打破方法上, 对全国各省区18个种群的苘麻(Abutilon theophrasti)种子进行温水浸种处理, 发现不同种群发芽率以及萌发时间存在显著差异, 最高和最低分别为89.95%和41.5% (孟帅帅等, 2021)。这些种子的差异大多与地理环境因素相关, 然而关于地理环境因素对不同种群种子萌发与休眠差异影响的研究却很少。
在湿地植被恢复中应用率很高的水毛花(Schoenoplectiella triangulata)是莎草科多年生草本、湿地挺水植物, 具有适应性强、根茎发达的优良特性, 且具极高饲料和园林造景的潜在价值。虽然水毛花应用前景高(Reznicek, 1990), 但其种子的萌发极易受到温度、光照、水分、埋土深度、土壤酸碱度等环境因素以及自身形态特性、生理特性等因素的影响, 出现种子休眠、萌发困难等问题(唐金刚等, 2013)。受环境和遗传因素影响, 其不同地区种群间的种质资源存在较大差异(Bhatt et al., 2020), 严重制约这一重要资源的开发利用。因此了解不同地区的水毛花种群的种子萌发变异性对于科学有效的种质收集以及进行植物群落恢复和保护至关重要; 探究水毛花种子的休眠特性, 提高水毛花种子的萌发效率, 对其在园林中的应用具有重要意义。
目前国内外对水毛花的研究主要集中于药用成分研究以及水质净化作用研究, 如何解除其种子休眠并提高其种子萌发的研究少之又少。本研究以采集自不同地区种群的水毛花为实验材料, 探究不同地区种群的种子萌发机制与休眠机制, 以期发现野生种群建群中存在的问题, 为植物种质资源的科学收集、保存以及在园林中的应用提供理论依据。
1 材料和方法
1.1 实验材料
实验所需的所有种子采自中国科学院庐山植物园鄱阳湖分园的同质园(29.67° N, 116.89° E)中的水毛花植株。2019-2020年从5个不同地区(表1)引种幼苗或植株, 移栽至同质园。引种幼苗或植株时, 在每个地区水毛花大量集中分布区域选择6个样点, 每个样点边缘间隔不小于5 m, 每个样点随机采取6株能够代表该样点水毛花群落整体生长状况的幼苗或植株带回同质园。每个样点采集的6株幼苗或植株均匀种植在装满1/2土壤的塑料水盆(内径86 cm, 高40 cm)内。在2021和2022年8-11月种子成熟后, 将每个样点采集的6株植株所产生的种子采集并混匀, 阴干后装入种子袋中备用。每个样点产生的种子作为一个重复, 每个地区产生的所有种子视为一个种群。2021年产生的种子用于种子形态特征测定, 2022年产生的种子用于种子萌发特性和休眠特性的实验。
表1 水毛花种源地信息
Table 1
| 原生地 Site of collection | 种群编号 Population ID | 位置 Location | 经纬度 Latitude and longitude | 年平均气温 Mean annual air temperature (℃) | 年降水量 Annual precipitation (mm) |
|---|---|---|---|---|---|
| 海南 Hainan | HN1 | 海口昌旺 Changwang, Haikou | 19.80° N, 110.25° E | 25.4 | 1 639 |
| HN2 | 海口羊山 Yangshan, Haikou | 19.94° N, 110.31° E | 25.4 | 1 639 | |
| 贵州 Guizhou | GZ1 | 草海 Caohai | 26.89° N, 104.23° E | 10.5 | 950 |
| GZ2 | 草海 Caohai | 26.99° N, 104.24° E | 10.5 | 950 | |
| 江西 Jiangxi | JX | 上饶余干 Yugan, Shangrao | 28.91° N, 116.49° E | 19.0 | 1 700 |
1.2 种子形态特征
本实验所用种子取自每个种群中5个结种率较高的重复组, 每个重复组随机选取100粒饱满、阴干的成熟种子, 利用天平称量, 取平均值乘以10即得到种子千粒质量。
从每个种群的上述5个重复中各随机选取10粒饱满、阴干的成熟种子, 共计50粒种子, 利用体视解剖镜测量每一粒种子的长、宽、厚(mm), 并拍照记录种子颜色、形状等外形特征。
每个重复组随机选取500粒饱满、阴干的成熟种子, 称量初始质量, 置于蒸馏水中, 25 ℃保存, 每2、4、6、8、24、36、48 h取出, 擦干表面水分, 称量至恒质量。根据以下公式计算种子的吸水率(Ci)和吸水速率(Vi, i+1)。
式中, M0为种子干质量; Mi为种子湿质量; Ri和Ri+1为两个相邻测试时间的平均吸水系数; t为间隔时间(h); 吸水速率的单位为mg·g-1·h-1。
1.3 种子萌发特性
本研究中所有种子萌发实验均采用水培法。
为探究光照对水毛花种子萌发的影响, 从每个种群4个结种率较高的重复组中随机选取成熟、饱满的种子500粒, 共计2 000粒种子用于实验。将5个种群不同重复组的种子分别置于10个表层铺有双层湿润滤纸的培养皿中, 每个培养皿50粒种子。种子进行两种光照处理: 光强为3 000 lx的8 h光照16 h黑暗处理和全黑暗处理。每个光照处理再进行5种变温处理: 15/10 ℃、20/15 ℃、25/20 ℃、30/25 ℃、35/30 ℃。光照下每天记录萌发的种子数量, 黑暗环境3天测1次, 尽量避免光照带来影响。本次萌发实验为期50天, 整个实验期间保持滤纸湿润。
为探究温度对种子萌发的影响, 从每个种群4个结种率较高的重复组中随机选取成熟饱满的种子450粒, 共计1 800粒种子用于实验。将5个种群不同重复组的种子置于9个铺有双层湿润滤纸的培养皿中, 并置于16 h光照、8 h黑暗的培养箱中, 每个培养皿50粒种子。将种子分别进行恒温处理(15、20、25和30 ℃)和变温处理(15/10 ℃、20/15 ℃、25/20 ℃、30/25 ℃和35/30 ℃), 光照时高温, 黑暗时低温。每天记录一次发芽的种子数量。本次萌发实验为期40天, 整个实验期间保持滤纸湿润。
发芽率(GR)和萌发指数(GI)计算公式如下:
式中, N为供试种子粒数, N1为全部正常发芽粒数, Gt为实验发芽开始到结束期间内每日发芽数, Dt为发芽日数。萌发时滞(GD)即从萌发实验开始到第1粒种子发芽所持续的时间(d)。
1.4 种子休眠特性
打破休眠的物理方法采用低温层积法。将采集的种子储藏于以下6种环境中: 室内常温干燥环境(常干), 室内常温水浸(常水), 室内常温湿沙藏(常沙), 4 ℃低温干燥环境(冷干), 4 ℃低温水浸(冷水), 4 ℃低温湿沙藏(冷沙)。其中, 常温与低温做对比, 干燥、水浸和沙藏三者做对比。在6种储藏处理中, 将种子置于培养皿中, 封口膜密封, 减少水分散失, 常水、常沙、冷水和冷沙及时加水; 分别在第2、4、6个月时从6组重复中各取未萌发50粒种子, 共计300粒种子进行萌发实验。每天记录一次发芽的种子数量, 本次萌发实验为期30天。用1.3方法计算GR、GI和GD。
打破休眠的化学方法采用激素处理方法。赤霉素(GA3)和氟啶酮(FL)可显著打破莎草科植物嵩草(Carex myosuroides)、风车草(Cyperus involucratus)、碎米莎草(Cyperus iria)和球穗扁莎(Pycreus flavidus)等植物种子的休眠, 提升其发芽率(李欣勇等, 2021a, 2021b); 唐金刚等(2013)采用0-300 μmol·L-1的GA3溶液浸种对水毛花种子发芽率的提升无显著效果; 而周芝琴等(2013)用激素处理的种子进行萌发实验, 发现激素对种子萌发有显著促进作用。因此, 本研究使用50 μmol·L-1的FL溶液和100、200、300 μmol·L-1的GA3溶液培养种子, 以蒸馏水培养作为对照, 每种处理6个重复。萌发实验为期30天, 每天记录一次萌发的种子数据。用1.3方法计算GR、GI和GD。
1.5 数据分析
所有数据的统计分析均在Excel和SPSS软件上进行, 用Origin 2017和Chiplot软件作图。数据符合方差齐性和正态分布, 采用单因素方差分析对水毛花种子的长、宽、高和不同温度、光照和化学处理下的发芽率、萌发指数和萌发时滞进行差异性分析; 采用广义线性模型分析种子的收集种群、存储方式、存储温度以及存储时间对种子发芽率的影响; 采用Pearson相关系数对植物种子的形态特性、萌发特性与环境因子进行相关性分析; 采用聚类分析对植物种子形态特性以及各温度下萌发状况进行遗传关系远近的分析。统计数据均为平均值±标准误(mean ± SE), 显著度为0.05。
2 结果
2.1 种子形态特征
图1

图1
水毛花江西种群种子形态特征。
Fig. 1
Seed morphological characteristics of Schoenoplectiella triangulata in Jiangxi.
表2 水毛花种子形态和千粒质量的种群间差异 (平均值±标准误)
Table 2
| 种群编号 Population ID | 长 Length (mm) | 宽 Width (mm) | 高 Height (mm) | 千粒质量 Thousand grain mass (g) |
|---|---|---|---|---|
| HN1 | 2.12 ± 0.12a | 1.57 ± 0.11a | 0.81 ± 0.08b | 1.24 ± 0.03a |
| HN2 | 1.94 ± 0.11b | 1.47 ± 0.09b | 0.71 ± 0.07d | 0.85 ± 0.02d |
| GZ1 | 1.89 ± 0.16c | 1.44 ± 0.09c | 0.75 ± 0.06c | 1.04 ± 0.06bc |
| GZ2 | 1.86 ± 0.11c | 1.23 ± 0.14d | 0.63 ± 0.09e | 0.99 ± 0.03c |
| JX | 1.84 ± 0.24c | 1.48 ± 0.10b | 0.86 ± 0.09a | 1.04 ± 0.03b |
HN1, 海口昌旺种群; HN2, 海口羊山种群; GZ1、GZ2, 贵州草海不同种群; JX, 上饶余干种群。同列不同小写字母表示差异显著(p < 0.05)。
HN1, Changwang population in Haikou; HN2, Yangshan population in Haikou; GZ1 and GZ2, different populations in Caohai, Guizhou; JX, Yugan population in Shangrao. Different lowercase letters in the same column indicate significant differences (p < 0.05).
不同地区种群水毛花的吸水率随着时间的延长呈现先快后缓的递增趋势。其中浸种0-2 h吸水速率最快, 为快速吸水期, 由快到慢依次是JX、HN2、HN1、GZ2、GZ1, 后趋于平缓。GZ1、GZ2和HN1在24 h后吸水率增长趋于平缓, 并且GZ1、GZ2和HN1、HN2于48 h后达到吸水饱和。JX水毛花在浸种2-96 h吸水率一直处于缓慢增长, 120 h后饱和, 且其吸水率在5个生境中一直居于最高。JX、GZ1、GZ2、HN1、HN2水毛花最大吸水率分别为25%、14.3%、16.6%、10.9%和8.9% (图2)。
图2

图2
不同地区种群水毛花种子吸水率和吸水速率变化(平均值±标准误)。不同小写字母表示差异显著(p < 0.05)。HN1, 海口昌旺种群; HN2, 海口羊山种群; GZ1、GZ2, 贵州草海不同种群; JX, 上饶余干种群。
Fig. 2
Water uptake percentage and water uptake rate of Schoenoplectiella triangulata seeds from different populations (mean ± SE). Different lowercase letters indicate significant differences (p < 0.05). HN1, Changwang population in Haikou; HN2, Yangshan population in Haikou; GZ1 and GZ2, different populations in Caohai, Guizhou; JX, Yugan population in Shangrao.
不同地区的水毛花种子的种皮颜色随着吸水时间的延长而逐渐加深, 吸水达到饱和2天后种子开始萌发。不同地区的种子在吸水率变化上存在显著差异, 说明不同地区种群种子在休眠与萌发特性上存在较大差异。
2.2 种子萌发特性
不同光照条件下种子萌发结果显示, 光照条件促进水毛花种子萌发, 不同地区种群发芽率对温度的响应有差异; 黑暗条件下各种群水毛花几乎不萌发(表3)。本研究结果表明, 水毛花的种子萌发是需要光照的。
表3 不同温度及光照条件对水毛花种子萌发的影响 (平均值±标准误)
Table 3
| 温度 Temperature (℃) | 光照条件 Light condition | 发芽率 Germination percentage (%) | ||||
|---|---|---|---|---|---|---|
| HN1 | HN2 | JX | GZ1 | GZ2 | ||
| 15/10 | 光照 Light | 8.50 ± 1.50Ba | 9.50 ± 2.50Ca | 0 ± 0Ba | 0 ± 0Ba | 0 ± 0Aa |
| 黑暗 Dark | 0 ± 0Ab | 0 ± 0Ab | 0 ± 0Aa | 0 ± 0Aa | 0 ± 0Aa | |
| 20/15 | 光照 Light | 32.50 ± 5.50Aa | 53.00 ± 8.50Aa | 6.00 ± 5.00Aa | 1.50 ± 0.50Ba | 0 ± 0Aa |
| 黑暗 Dark | 0 ± 0Ab | 1.00 ± 0.50Ab | 0 ± 0Ab | 0 ± 0Aa | 0 ± 0Aa | |
| 25/20 | 光照 Light | 7.75 ± 3.25Ba | 22.00 ± 4.00Ba | 10.0 ± 8.00Aa | 9.00 ± 1.00Aa | 0 ± 0Aa |
| 黑暗 Dark | 0 ± 0Ab | 0 ± 0Ab | 0.50 ± 0.25Ab | 0 ± 0Ab | 0 ± 0Aa | |
| 30/25 | 光照 Light | 3.50 ± 2.50Ca | 3.50 ± 4.50Da | 2.00 ± 1.50Ba | 7.50 ± 4.50Aa | 0 ± 0Aa |
| 黑暗 Dark | 0 ± 0Ab | 0 ± 0Aa | 0.50 ± 0.50Aa | 0.50 ± 0.50Ab | 0 ± 0Aa | |
| 35/30 | 光照 Light | 0 ± 0Ca | 3.50 ± 2.50Da | 0.75 ± 0.50Ba | 1.00 ± 0.50Ba | 0.75 ± 0.50Aa |
| 黑暗 Dark | 0 ± 0Aa | 0 ± 0Aa | 0 ± 0Aa | 0 ± 0Aa | 0 ± 0Aa | |
| 15 | 光照 Light | 0 ± 0Ca | 1.50 ± 1.25Da | 0 ± 0Ba | 0 ± 0Ba | 0 ± 0Aa |
| 黑暗 Dark | 0 ± 0Aa | 0 ± 0Aa | 0 ± 0Aa | 0 ± 0Aa | 0 ± 0Aa | |
| 20 | 光照 Light | 1.75 ± 0.75Ca | 2.75 ± 1.25Da | 0.75 ± 0.50Ba | 0.50 ± 0.50Ba | 0 ± 0Aa |
| 黑暗 Dark | 0 ± 0Aa | 0 ± 0Aa | 0 ± 0Aa | 0 ± 0Aa | 0 ± 0Aa | |
| 25 | 光照 Light | 2.50 ± 1.00Ca | 0.50 ± 1.00Da | 0.50 ± 1.00Ba | 0.25 ± 0.50Ba | 0 ± 0Aa |
| 黑暗 Dark | 0 ± 0Aa | 0 ± 0Aa | 0 ± 0Aa | 0 ± 0Aa | 0 ± 0Aa | |
| 30 | 光照 Light | 0 ± 0Ca | 0 ± 0Da | 0 ± 0Ba | 0 ± 0Ba | 0.25 ± 0.50Aa |
| 黑暗 Dark | 0 ± 0Aa | 0 ± 0Aa | 0 ± 0Aa | 0 ± 0Aa | 0 ± 0Aa | |
不同大写字母表示同一种群在相同光照、不同温度条件下发芽率差异显著; 不同小写字母表示同一种群在相同温度、不同光照条件下差异显著(p < 0.05)。HN1, 海口昌旺种群; HN2, 海口羊山种群; GZ1、GZ2, 贵州草海不同种群; JX, 上饶余干种群。
Different uppercase letters indicate significant differences in germination rates among temperature conditions under same lighting condition of the same population; different lowercase letters indicate significant differences among lighting conditions under same temperature condition of the same population (p < 0.05). HN1, Changwang population in Haikou; HN2, Yangshan population in Haikou; GZ1 and GZ2, different populations in Caohai, Guizhou; JX, Yugan population in Shangrao.
恒温不利于水毛花种子的萌发, 各种群发芽率皆低于5%。而变温有利于水毛花的种子萌发, HN1与HN2升高最多, 其种子萌发最佳变温为20/15 ℃, 发芽率高达32.5%、52%; JX水毛花与GZ1最佳萌发变温为25/20 ℃, 而GZ2在各种恒温与变温条件下发芽率低或均极少萌发(图3A)。且HN1和JX幼苗较其他种群萌发幼苗粗壮。水毛花发芽率和萌发指数呈现先增后减的趋势, JX与GZ1种子在25/20 ℃发芽率最高, HN1与HN2种子在20/15 ℃发芽率最高, 高达6.6%、15.9%, 而GZ2在各温度条件下都几乎无萌发(图3B)。各种群的水毛花种子变温与恒温相比萌发时滞差异不大。萌发时滞随着萌发变温的递增呈现先短后长的递减趋势, 在25/20 ℃达到最短15天, 在35/30 ℃变温时萌发时滞又延长, 其中JX水毛花正好相反, 在35/30 ℃达到萌发时滞最短为5天; 恒温下萌发时滞呈现递减趋势, 即温度越高, 萌发所需时间越短(图3C)。
图3

图3
不同温度条件下水毛花种子萌发动态(平均值±标准误)。不同小写字母表示不同种群间差异显著(p < 0.05)。HN1, 海口昌旺种群; HN2, 海口羊山种群; GZ1、GZ2, 贵州草海不同种群; JX, 上饶余干种群。
Fig. 3
Germination dynamics of Schoenoplectiella triangulate seeds under different temperature conditions (mean ± SE). Different lowercase letters indicate significant differences among different populations (p < 0.05). HN1, Changwang population in Haikou; HN2, Yangshan population in Haikou; GZ1 and GZ2, different populations in Caohai, Guizhou; JX, Yugan population in Shangrao.
2.3 种子休眠特性
对水毛花种子的种群、存储方式、存储时间、存储温度进行广义线性分析(表4)显示, 种子的收集种群、存储方式、存储时间以及存储温度对种子发芽率的双向交互作用均具有显著影响。
表4 基于广义线性分析的种群、存储方式、存储时间、存储温度及其交互作用对水毛花种子萌发的影响
Table 4
| 因子 Factor | Wald’s χ2 | df | p | 因子 Factor | Wald’s χ2 | df | p |
|---|---|---|---|---|---|---|---|
| 种群 Population (P) | 664.030 | 4 | <0.001 | M × Te | 88.044 | 2 | <0.001 |
| 存储方式 Storage manner (M) | 190.189 | 2 | <0.001 | M × Ti | 170.335 | 6 | <0.001 |
| 存储温度 Storage temperature (Te) | 142.543 | 1 | <0.001 | Ti × Te | 58.457 | 3 | <0.001 |
| 存储时间 Storage time (Ti) | 515.916 | 3 | <0.001 | P × M × Te | 107.331 | 8 | <0.001 |
| P × M | 193.495 | 8 | <0.001 | P × M × Ti | 165.427 | 24 | <0.001 |
| P × Te | 139.538 | 4 | <0.001 | P × Ti × Te | 109.271 | 12 | <0.001 |
| P × Ti | 553.715 | 12 | <0.001 | M × Ti × Te | 65.653 | 6 | <0.001 |
2个月的低温层积有助于水毛花种子休眠的打破(图4)。与常干(对照组)相比, 常沙、冷水、冷沙层积方式可以显著打破HN1、HN2和JX 3个种群的水毛花种子休眠, 进而提高种子发芽率, 显著降低萌发时滞和缩短种子萌发周期, 但不同地区种群的种子休眠的最佳打破方式不同; 而低温层积对GZ1和GZ2两个种群水毛花种子休眠的打破无显著效果。
图4

图4
存储条件对水毛花种子的累积发芽率的影响。A, 海口昌旺种群。B, 海口羊山种群。C、D, 贵州草海不同种群。E, 上饶余干种群。
Fig. 4
Effects of storage conditions on the cumulative germination percentage of Schoenoplectiella triangulate seeds. A, Changwang population in Haikou. B, Yangshan population in Haikou. C, D, Different populations in Caohai, Guizhou. E, Yugan population in Shangrao.
存储时间对大部分种群水毛花种子萌发影响显著(图5)。GZ1、GZ2种群随着存储时间增长发芽率降低, 趋于0; 其他种群种子在各种层积方式下存储两个月时, 发芽率最高, 后逐渐降低。存储方式对种子萌发也有显著影响, 其中冷沙、冷水和常沙3种存储方式储藏后发芽率最高, 达50%以上。温度对水毛花种子的萌发也有显著影响, 其中发芽率冷沙小于常沙, 冷水高于常水, 冷干高于常干(图4)。不同地区种群种子的发芽率差异显著, HN1和JX两个种群的水毛花种子在6种存储方式下存储两个月, 其发芽率有显著提高, HN2种群的种子在冷沙、冷水、常沙和常水存储条件下存储两个月发芽率也得到了显著提高; 但GZ1、GZ2种群的种子在6种存储条件下发芽率皆不高。
图5
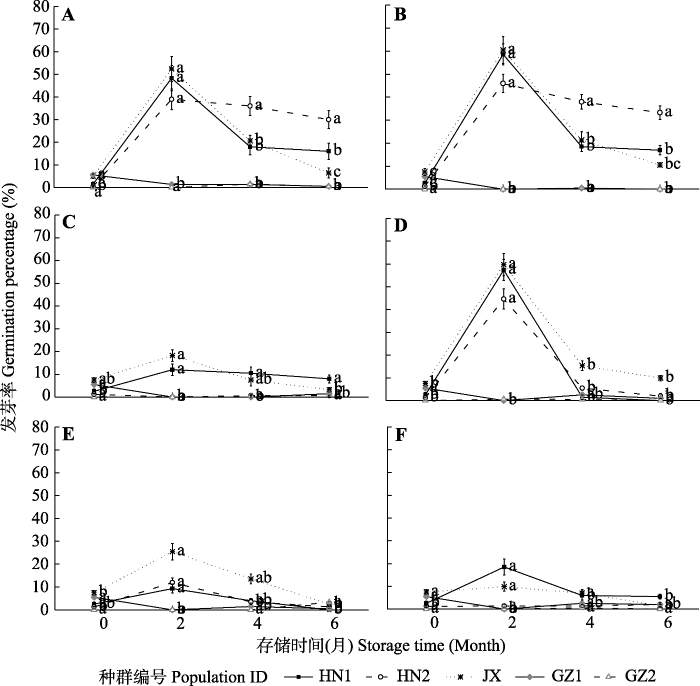
图5
存储时间对水毛花种子发芽率的影响(平均值±标准误)。A, 冷沙。B, 冷水。C, 冷干。D, 常沙。E, 常水。F, 常干。不同小写字母表示不同存储时间间差异显著(p < 0.05)。HN1, 海口昌旺种群; HN2, 海口羊山种群; GZ1、GZ2, 贵州草海不同种群; JX, 上饶余干种群。
Fig. 5
Effects of storage time on the germination percentage of Schoenoplectiella triangulate seeds (mean ± SE). A, Low temperature plus sand reservoir. B, Low temperature plus water reservoir. C, Low temperature plus dry reservoir. D, Normal temperature plus sand reservoir. E, Normal temperature plus water reservoir. F, Normal temperature plus dry reservoir. Different lowercase letters indicate significant differences among different storage time (p < 0.05). HN1, Changwang population in Haikou; HN2, Yangshan population in Haikou; GZ1 and GZ2, different populations in Caohai, Guizhou; JX, Yugan population in Shangrao.
FL处理可以有助于部分种群水毛花种子休眠的解除。与对照组对比, FL处理可打破HN1、JX和GZ1的水毛花种子休眠, 显著提高种子发芽率, 但发芽率低于50%; 但是其对于HN2、GZ2的作用不显著。FL处理后萌发的幼苗呈现白化状态(图6D), 且不久后死亡, 无法移栽土培。
图6

图6
氟啶酮(FL)对水毛花种子萌发的影响(平均值±标准误)。 A, 发芽率。B, 萌发指数。C, 萌发时滞。D, 幼苗形态, 从左到右依次为FL处理JX种子萌发后第2、4、5、6、7、9、11、14天幼苗形态。不同小写字母表示不同处理间差异显著(p < 0.05)。HN1, 海口昌旺种群; HN2, 海口羊山种群; GZ1、GZ2, 贵州草海不同种群; JX, 上饶余干种群。CK, 对照。
Fig. 6
Effects of fluridone (FL) on the germination of Schoenoplectiella triangulate seeds (mean ± SE). A, Germination percentage. B, Germination index. C, Germination time-lag. D, Seedling morphology (from left to right, on day 2, 4, 5, 6, 7, 9, 11 and 14 after JX seeds treated by FL germinated). Different lowercase letters indicate significant differences between different treatments (p < 0.05). HN1, Changwang population in Haikou; HN2, Yangshan population in Haikou; GZ1 and GZ2, different populations in Caohai, Guizhou; JX, Yugan population in Shangrao. CK, control.
GA3处理有助于部分种群水毛花种子休眠的打破。GA3处理可打破HN1和JX的水毛花种子休眠, 显著提高种子发芽率, 提升种子活力; 但是对于HN2、GZ1和GZ2的作用不显著。GA3处理后萌发的幼苗形态比正常水培幼苗细长并且颜色较浅(图7D), 移栽土培时易死亡。
图7

图7
赤霉素(GA3)对水毛花种子萌发的影响(平均值±标准误)。A, 发芽率。B, 萌发指数。C, 萌发时滞。D, 幼苗形态, 从左到右依次为JX种子萌发后第2、4、6、8、10、12天幼苗形态。不同小写字母表示不同处理间差异显著(p < 0.05)。HN1, 海口昌旺种群; HN2, 海口羊山种群; GZ1、GZ2, 贵州草海不同种群; JX, 上饶余干种群。
Fig. 7
Effects of Gibberellins on the germination of Schoenoplectiella triangulate seeds (mean ± SE). A, Germination percentage. B, Germination index. C, Germination time-lag. D, Seedling morphology (from left to right, on day 2, 4, 6, 8, 10 and 12 after JX seeds treated by GA3 germinated). Different lowercase letters indicate significant differences among different treatments (p < 0.05). HN1, Changwang population in Haikou; HN2, Yangshan population in Haikou; GZ1 and GZ2, different populations in Caohai, Guizhou; JX, Yugan population in Shangrao.
3 分析与讨论
3.1 不同地区种群水毛花种子的形态与生理存在差异性
种子的表型可塑性反映了植物的遗传变异及其对环境的适应策略, 在野生植物资源开发和选种育种等方面有较大研究价值(李永霞等, 2022)。种子里胚胎周围的母体组织, 如种皮的差异、种子内的物质含量差异等会影响种子的萌发(Joseph et al., 2010; Hammami et al., 2020)。种子萌发需要启动多重代谢途径, 为种子萌发做准备, 这个过程需要大量能量和中间物质(薛梅真等, 2018)。种皮的厚度以及种皮外的硬质角质会影响种子的透气性, 阻碍内外水分、氧气等物质的交流; 种子内物质(如淀粉、蛋白质、可溶性糖、植物激素和酶等)含量的差异, 也可能在萌发差异中发挥作用(谢坤秀等, 2022)。淀粉、蛋白质、可溶性糖等物质在各种酶的催化下发生代谢反应, 为种子萌发提供能量; 植物激素则可在打破种子休眠上起调节作用(Lando et al., 2020)。本研究结果显示, 不同地区的水毛花种群的种子大小和吸水性存在较大差异, 并且对种子萌发影响显著, 种子的饱满程度与发芽率成正比。
种子萌发极易受外界光照、温度等环境因子的影响。光照促进或抑制种子萌发的现象受种子内的光敏色素调控(Vicente et al., 2020)。种子内的光敏色素是其在成熟过程中或吸水后合成的, 它存在两种形态——远红光吸收态(Pfr)和红光吸收态(Pr)。而种子萌发主要由Pfr含量和Pfr/(Pr + Pfr)比值决定(张敏等, 2012)。沉水植物苦草(Vallisneria natans)的种子可在黑暗条件下萌发, 发芽率随光照强度增加而增高(王瑞等, 2021), 而莎草科挺水植物风车草的种子在黑暗条件下不萌发(李欣勇等, 2021b), 本研究的结果发现水毛花种子在黑暗条件下其发芽率近乎为0, 而在光照条件下其发芽率大幅度提高, 表明光能驱动水毛花种子的萌发。温度是影响种子萌发最重要的因素之一。温度会影响种子内酶的活力, 温度过高或过低都会使种子内生理活动出现异常, 如内源生长调节物质含量变化、糖代谢减慢等, 从而阻碍种子的发育和萌发(罗艳等, 2021)。在适应环境过程中, 植物会形成最佳萌发温度。本研究结果显示不同地区水毛花种群的种子最佳萌发温度差异较大。
本研究结果显示水毛花种子的形态特性、萌发特性在不同地区种群间存在明显差异, 通过对种子的长、宽、高、千粒质量、吸水率以及不同变温条件下发芽率等指标进行聚类分析(图8), 结果显示HN1、HN2相似性最近, GZ1、GZ2相似性最近, 而JX与GZ1、GZ2相似性最近, 也证明了种子的形态特性与萌发特性与采集地的地理环境相关。
图8
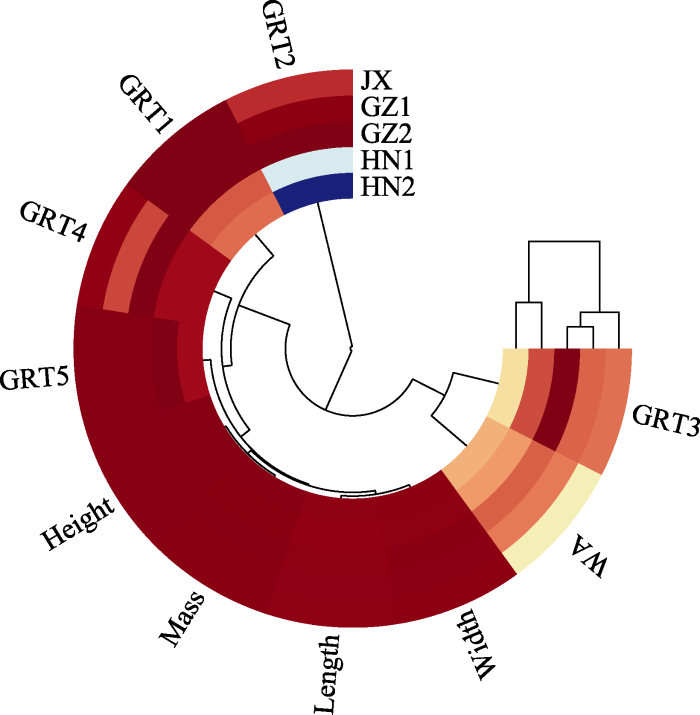
图8
水毛花不同地区种群种子的聚类分析。Length、Width、Height、Mass、WA为种子的长、宽、高、千粒质量、吸水率; GRT1、GRT2、GRT3、GRT4、GRT5依次为5个变温下发芽率。HN1、HN2、GZ1、GZ2、JX同
Fig. 8
Cluster analysis of Schoenoplectiella triangulata seeds from different populations. Length, Width, Height, Mass, and WA are the length, width, height, 1 000-grain mass, and water absorption rate of seeds, respectively; GRT1, GRT2, GRT3, GRT4 and GRT5 are the germination percentages under five variable temperatures, respectively. HN1, HN2, GZ1, GZ2 and JX were the same as those in
不同地区种群之间种子萌发的差异是复杂的,不能简单地用种子植株采集地的温度、降水等单一条件来解释(Zhang et al., 2022)。因此本研究将采集地的环境因素经度、纬度、年平均气温、年降水量, 种子的形态长、宽、高、千粒质量、吸水率以及5个变温下的发芽率进行Pearson相关性分析(图9), 结果证明不同地区水毛花种群的种子长、种子宽均与纬度呈显著负相关关系, 同时各地区的种子发芽率与纬度也呈负相关关系, 并且萌发温度越低, 差异性越显著; 种子的宽、高、吸水率与经度呈正相关关系, 而种子发芽率与经度相关性不显著; 种子的长、宽、高、吸水率与发芽率、采集地的年降水量以及年平均气温呈正相关关系, 但发芽率随萌发变温的升高显著性会降低, 并且萌发温度在30/25 ℃、35/30 ℃时发芽率与年降水量、年平均气温的相关性不显著性; 且种子的质量与采集地相关性不大。不同地区的水毛花种群的种子形态特征、发芽率与采集地信息呈现相关性, 这也与张祖新(2020)的研究结果较为一致。Imbert (2002)认为种子形态差异可能源于种子个体发育的限制, 种子个体发育的限制与头部花轴形态相关(van Mölken et al., 2005; 秦启娟等, 2022), 但对于水毛花的种子形态的发育限制还有待研究。
图9

图9
不同地区种群水毛花种子的形态特性、发芽率与环境因子的相关性分析。GRT1、GRT2、GRT3、GRT4、GRT5依次为5个变温下发芽率; Lat, 纬度; Length、Width、Height、Mass、WA依次为种子的长、宽、高、千粒质量、吸水率; Long, 经度; MAAT, 年平均气温; MAP, 年降水量。*, p < 0.05; **, p < 0.01。
Fig. 9
Correlation analyses among morphological characteristics, germination percentage and environmental factors of Schoenoplectiella triangulata seeds from different populations. GRT1, GRT2, GRT3, GRT4, GRT5, the germination percentages under five variable temperatures, respectively; Lat, latitude; Length, Width, Height, Mass, and WA are the length, width, height, 1 000-grain mass, and water absorption rate of seeds, respectively; Long, longitude; MAAT, mean annual air temperature; MAP, mean annual precipitation. *, p < 0.05; **, p < 0.01.
3.2 不同地区水毛花种群种子的休眠类型与休眠打破方法的差异
种子休眠是由环境和遗传决定的复杂行为, 是植物为了适应环境变化所产生的应对策略(王因花等, 2022)。由Nikolaeva所建立并由美国著名种子生态学家Baskin和Baskin所修订的休眠分类系统将植物种子休眠分为物理休眠(PY)、生理休眠(PD)、形态休眠(MD)、形态生理休眠(MPD)以及复合休眠(PY+PD) 5种类型(Baskin & Baskin, 2004, 2014)。本研究中不同地区的水毛花种群的种子在休眠类型上无显著差异, 但在打破休眠方式上存在细微差异。水毛花种子可以在不破坏种皮情况下吸收一定水分, 因此水毛花的种子不存在物理休眠。种子的种胚随存储时间增长无显著发育, 因此水毛花的种子不存在形态休眠。HN1、HN2、JX、GZ1种群的水毛花种子可以通过变温条件、低温层积、FL、GA3处理打破休眠, 从而使种子发芽率显著提升, 因此存在PD。GZ2种群的水毛花种子在各种处理下都几乎不萌发, 可能与种子形态或生理限制有关。
种子的萌发是经多种酶催化后发生一系列物质转化和能量传递(Kucera et al., 2005)而产生的, 因此温度对其的影响极大。本研究结果也显示, 变温条件下水毛花种子发芽率显著高于恒温, 是由于变温可以加速酶的催化作用, 并且促进种壳软化, 有利于胚突破种皮, 从而促进种子萌发(李欣勇等, 2021a)。低温层积是通过温度大幅度骤变来降低种子内部脱落酸等抑制种子萌发的物质含量, 进而打破生理休眠促进萌发。而本研究中的低温水藏、低温沙藏条件一方面降低了脱落酸等物质的含量, 另一方面软化种皮, 加速物质流通, 两者共同促进种子萌发。低温沙藏、低温水藏、常温沙藏3种层积方式确实显著提升水毛花发芽率, 与刘贵华等(2005)的研究结果基本一致, 但是低温沙藏、低温水藏、常温沙藏存储方式不适合种子的长期储存, 这3种存储方式下, 种子活力降低速度快, 并且遇到合适温度极易萌发。
激素可以通过调节种子内部萌发抑制物质和萌发促进物质的含量比例, 增加赤霉素等萌发促进物质的含量, 降低脱落酸等萌发抑制物质的含量, 进而解除休眠, 促进萌发(Lando et al., 2020)。常用的化学激素包括赤霉素、细胞分裂激素、萘乙酸、乙烯利、氟啶酮、褪黑激素等等。化学激素氟啶酮可破除因存在脱落酸而抑制种子萌发的休眠, 通过阻碍脱落酸合成、促进赤霉素合成解除休眠, 可以打破碎米莎草、球穗扁莎、风车草等莎草科植物种子休眠(李欣勇等, 2020, 2021a, 2021b), 显著促进萌发; 而化学激素赤霉素可使莎草科植物萤蔺(Schoenoplectiella juncoides)的种子内存在的萌发抑制物钝化或失效进而促进萌发, 唐金刚等(2013)采用0-300 μmol·L-1的GA3溶液浸种对水毛花的种子发芽率的提升无显著效果, 其原因可能是浓度过低。因此本研究采用50 μmol·L-1的FL溶液和0- 300 μmol·L-1的GA3溶液培养种子, 结果显示50 μmol·L-1的FL可打破HN1、JX、GZ1的种子休眠, 提升其发芽率, 但FL处理后幼苗为白化苗; 300 μmol·L-1的GA3可打破HN1、JX的种子休眠, 显著提升其发芽率, 但无法提升HN2、GZ1、GZ2水毛花的种子发芽率, 幼苗细弱且绿色较浅。其原因可能是FL和GA3溶液在调动种子萌发相关激素的同时, 损伤了叶绿素, 破坏了叶绿素的合成途径, 具体反应机制还有待研究(Hamadina & Hamadina, 2018; 罗忍忍等, 2022)。不同地区种群休眠的打破方法上存在一定差异, 与种子的形态特性或母本植物采集地环境相关。
4 结论
综上, 湿地挺水植物水毛花为萌发喜光性植物, 种子存在生理休眠, 不同地区种群间的种子形态特性、萌发特性以及休眠打破上存在显著差异。在形态特性上, 各种群的水毛花种子大小依次为: HN1 > JX > GZ1 > GZ2 > HN2, 其中JX种子吸水率最高, 为25%; 在萌发特性上, 不同地区的水毛花种群的种子萌发的最佳温度差异显著, HN1、HN2最佳萌发温度为20/15 ℃, JX、GZ1最佳萌发温度为25/20 ℃, GZ2在各温度条件下基本无萌发; 在休眠打破方法上, 低温沙藏、低温水藏、常温沙藏3种层积方式可打破水毛花种子休眠, 显著提升其发芽率; FL可打破HN1、JX、GZ1的种子休眠, GA3处理则可打破HN1、JX的种子休眠, 显著提升其发芽率; HN1、JX、HN2、GZ1种群存在PD, GZ2种群种子可能存在形态或生理限制。而不同地区的水毛花种群的种子形态特性、萌发特性以及休眠特性与采集地的环境息息相关。种子的长、宽以及发芽率与纬度呈负相关关系, 且设置的萌发温度越低, 这种差异性越显著; 种子的长、宽、高、吸水率、发芽率以及采集地的年降水量与年平均气温呈正相关关系, 但随所设置萌发温度的升高, 发芽率显著性降低, 并且萌发温度在30/25 ℃、35/30 ℃时发芽率与年降水量、年平均气温的相关性不显著; 种子的质量与采集地无相关性。
致谢
感谢中铁十局集团第五工程有限公司技术开发项目(2019-003)资助。
参考文献
A classification system for seed dormancy
Inter-population variabilities in seed mass and germination of Panicum turgidum and Pennisetum divisum on the desert of Kuwait
Genetic differentiation and regional adaptation among seed origins used for grassland restoration: lessons from a multispecies transplant experiment
Rapid adaptation of seed germination requirements of the threatened Mediterranean species Malcolmia littorea (Brassicaceae) and implications for its reintroduction
Residual fluridone in humid tropical soils: carryover effects on germination and seedling growth of maize (Zea mays L.)
Redroot pigweed (Amaranthus retroflexus L.) and lamb’s quarters (Chenopodium album L.) populations exhibit a high degree of morphological and biochemical diversity
Geographical variation in seed germination and biochemical response of milk thistle (Silybum marianum) ecotypes exposed to osmotic and salinity stresses
Ecological consequences and ontogeny of seed heteromorphism
Imbibition duration, seed treatment, seed mass and population influence germination of annatto (Bixa orellana L.) seeds
Plant hormone interactions during seed dormancy release and germination
The physiological relationship between abscisic acid and gibberellinduring seed germination of Trichocline catharinensis (Asteraceae) is associated with polyamine and antioxidant enzymes
Analysis on seed characteristics of different populations of naked fruit trees
裸果木不同种群种子特性分析
Seed dormancy and germination characteristics of Cyperus iria
碎米莎草种子休眠与萌发特性研究
Seed dormancy and germination characteristics of Pycreus globosus
球穗扁莎种子的休眠与萌发特性
Study on dormancy and germination characteristics of Cyperus alternifolius seeds
风车草种子休眠及萌发特性研究
Seed phenotype and germination characteristics of different populations of Cupressus gigantea
不同种群巨柏种子表型及萌发特性
Effects of storage conditions and duration on seed germination of six wetland perennials
储藏条件和时间对六种多年生湿地植物种子萌发的影响
Spatiotemporal heterogeneity in precipitation patterns explain population-level germination strategies in an edaphic specialist
Effects of plant growth regulators on physiological characteristics and related gene expression in melon seedlings under cold stress
植物生长调节剂对冷胁迫下甜瓜幼苗生理特性及相关基因表达的影响
Effects of temperature on pollen viability and germination of cherry tomato
温度对樱桃番茄花粉活力及萌发率的影响
Seed germination of different populations of Abutilon theophrasti
苘麻不同种群的种子萌发特性
Dormancy and germination characteristics of the dimorphic seeds of Polygonum aviculare
扁蓄二型性种子休眠及萌发特性
Evolution in sedges (Carex, Cyperaceae)
Germination characteristics of Cakile edentula (Brassicaceae) seeds from two different climate zones
Study on 5 species of cyperceae aquatic plant seed germination in Caohai Lake, Weining County, Guizhou Province
威宁草海莎草科5种挺水植物种子萌发研究
Only seed size matters for germination in different populations of the dimorphic Tragopogon pratensis subsp
Effect of light, temperature, and salinity and drought stresses on seed germination of Hypericum ericoides, a wild plant with ornamental potential
Factors on seed germination, tuber sprout and plant growth of Vallisneria species in China
中国苦草属(Vallisneria)植物萌发与生长的影响因素
Study on germination characteristics and dormancy breaking methods of Tilia amurensis seeds
紫椴种子萌发特性及休眠解除方法研究
DOI:10.11924/j.issn.1000-6850.casb2021-1015
[本文引用: 1]

为探明紫椴种子休眠原因并找到休眠解除方法,以紫椴种子为材料,对其透性、萌发特性及休眠解除方法进行了研究。种子透性试验结果表明,紫椴种子经浓硫酸处理后透性增加,吸水率显著提高。种子萌发特性试验显示,紫椴完整种子萌发率为0,去掉种皮后,保留完整胚乳的种子萌发率为4.0%,切除部分胚乳(根尖未露出)的种子萌发率为23.3%,剥除胚乳帽露出根尖的种子萌发率为84.7%,离体胚萌发率为100%,此结果表明紫椴种子存在休眠,但其种胚并不存在休眠,其休眠是由胚外因素导致,胚乳(尤其是胚乳帽)是造成紫椴种子休眠的主要原因。种子休眠解除方法试验结果表明,浓H<sub>2</sub>SO<sub>4</sub>处理15 min+500 mg/L GA<sub>3</sub>溶液浸泡12 h+低温层积30天,可使紫椴种子发芽率达92.0%,可作为快速解除紫椴种子休眠的有效方法。综上,紫椴种子的种皮具有一定的透性障碍,但胚乳帽是造成其休眠的主要原因,使用浓H<sub>2</sub>SO<sub>4</sub>处理+GA<sub>3</sub>浸泡+低温层积的综合处理方法可在30天内解除紫椴种子的休眠。
Studies on morphological and germination characteristics of Zostera marina seed from different populations in Shandong Peninsula
山东半岛不同种群鳗草种子形态学特征及萌发特性研究
Research on soluble sugar metabolic regulation during the initial period of seed germination in maize
玉米种子萌发初期可溶性糖代谢途径的调控研究
Review on influence mechanisms of light in seed germination
DOI:10.3724/SP.J.1258.2012.00899
[本文引用: 1]
<p>Seed germination is a key process in the natural regeneration of plant populations and is mainly controlled by favorable temperature, water or light conditions. Light plays a critical role in determining the ability and rate of germination of photoblastic seeds. Furthermore, the regulation of seed germination by light involves not only</br>complicated physiological processes but also rigorous signal transductions and gene expression pathways. We summarized available data on the relationships between seed germination and other factors (i.e., light attributes, coupling of light and water/heat and phytochromes in seeds). To assess the effects of light on seed germination, we also comprehensively summarized the physiological reaction and light signal transduction mechanisms regulated by phytochromes in seed germination.</p>
光对种子萌发的影响机理研究进展
DOI:10.3724/SP.J.1258.2012.00899
[本文引用: 1]
种子萌发是植物成功实现天然更新的关键环节, 需要适宜的温度、水分或光照条件。对于需光性种子, 光照是决定其萌发与否或萌发率高低的主要因素。光对植物种子萌发的影响不仅是一个复杂的生理过程, 也是受到调控的信号传递和基因表达过程。该文系统总结了影响种子萌发的光照属性、光与水/热耦合作用和种子的光属性(光敏色素)与种子萌发的关系, 明确了光调控种子萌发的生态意义; 重点综述了种子内光敏色素调控种子萌发的生理反应模式和光敏色素的光信号转导途径。试图为全面评估光对种子萌发的影响和将来开展更深入的研究提供参考。
Effect of temperature and water potential on the germination of seeds from three different populations of Bidens pilosa as a potential Cd hyperaccumulator
DOI:10.1186/s12870-022-03876-3
PMID:36224525
[本文引用: 1]
Bidens pilosa L., an annual herb, has recently been shown to be a potential Cd-hyperaccumulating plant. The germination characteristics of B. pilosa have been documented, while the difference among populations remains unclear. Understanding variability in seed germination among populations is crucial for determining which populations to use for soil remediation programs.Present study was conducted to compare the requirements of temperature and water potential for germination of B. pilosa cypselae (the central type, hereafter seeds) from three populations using the thermal time, hydrotime, and hydrothermal time models. Seeds of three populations were incubated at seven constant temperatures (8, 12, 15, 20, 25, 30, and 35 °C) and at each of four water potentials (0, -0.3, -0.6, and -0.9 MPa). The results showed that germination percentage and rate of B. pilosa seeds were significantly by population, temperature, water potential and their interaction except for the interaction of population and water potential. Seeds from Danzhou population displayed a higher base temperature (T) for germination than those from Guilin and Baoshan population, however the ceiling temperature (T) had no consistent level among the populations but varied according to the water potential. In addition, the median base water potential [ψ] for germination of seeds from Danzhou population was higher than that for seeds from Baoshan and Guilin population at low temperatures (< 25 °C), which was opposite at high temperatures (≥ 25 °C).Seed germination requirements of B. pilosa on temperature and water differed significantly among populations. Differences in seed germination among populations may be complicated, which could not be simply explained by the temperature and rainfall conditions where the seeds were produced as previously reported. The results suggested that programme management should consider variation in seed germination traits when select which population could be applied to what kind of target remediation sites.© 2022. The Author(s).
Seed dormancy and germination characteristics of four Cyperaceae species
莎草科4种植物种子休眠与萌发特性的研究

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}