
Chin J Plant Ecol ›› 2014, Vol. 38 ›› Issue (7): 665-674.DOI: 10.3724/SP.J.1258.2014.00062 cstr: 32100.14.SP.J.1258.2014.00062
• Research Articles • Previous Articles Next Articles
SHI Qing-Ru1,2, XU Ming-Shan1,2, ZHAO Yan-Tao1,2, ZHOU Liu-Li1,2, ZHANG Qing-Qing1,2, MA Wen-Ji1,2, ZHAO Qi3, YAN En-Rong1,2,*( )
)
Received:2014-01-15
Accepted:2014-05-09
Online:2014-01-15
Published:2014-07-10
Contact:
YAN En-Rong
SHI Qing-Ru, XU Ming-Shan, ZHAO Yan-Tao, ZHOU Liu-Li, ZHANG Qing-Qing, MA Wen-Ji, ZHAO Qi, YAN En-Rong. Testing of corner’s rules across woody plants in Tiantong region, Zhejiang Province: effects of micro-topography[J]. Chin J Plant Ecol, 2014, 38(7): 665-674.
Add to citation manager EndNote|Ris|BibTeX
URL: https://www.plant-ecology.com/EN/10.3724/SP.J.1258.2014.00062
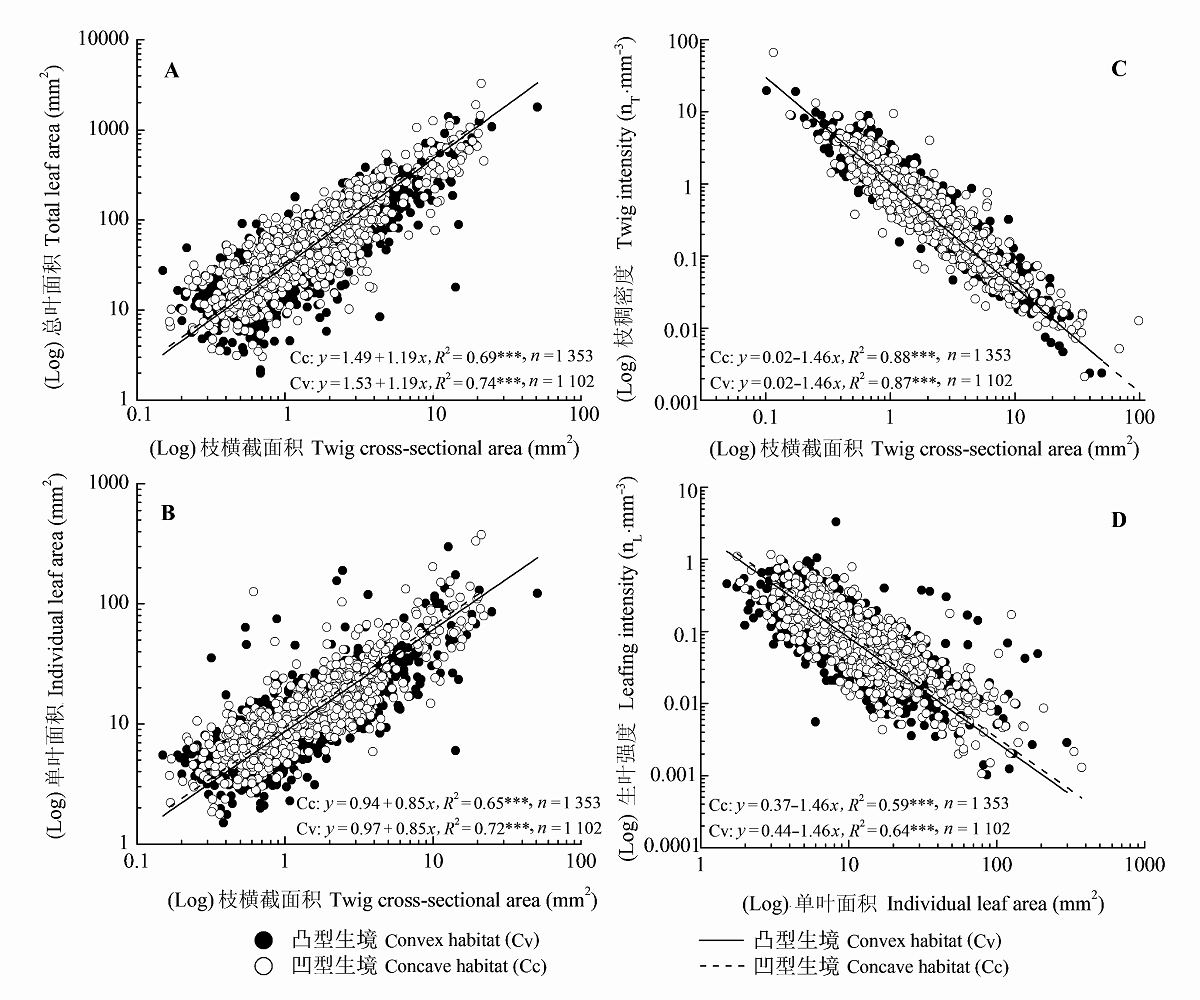
Fig. 1 Regression relationships of twig size-leaf size, twig size-twig intensity, and leaf size-leafing intensity for plants in concave and convex habitats at individual level. ***, p < 0.001.
| 组别 Group | 凸型生境 Convex habitat | 凹型生境 Concave habitat | p值 p value |
|---|---|---|---|
| 枝横截面积-总叶面积 Twig cross-sectional area-Total leaf area | 1.49 | 1.53 | 0.000 |
| 枝横截面积-单叶面积 Twig cross-sectional area-Individual leaf area | 0.94 | 0.97 | 0.000 |
| 枝横截面积-枝稠密度 Twig cross-sectional area-Twig intensity | -0.24 | -0.25 | 0.579 |
| 单叶面积-生叶强度 Individual leaf area-Leafing intensity | 0.37 | 0.44 | 0.000 |
Table 1 Results of significance test for differences in intercept in three types of twig-leaf relationships across individual plants in two types of micro-habitat
| 组别 Group | 凸型生境 Convex habitat | 凹型生境 Concave habitat | p值 p value |
|---|---|---|---|
| 枝横截面积-总叶面积 Twig cross-sectional area-Total leaf area | 1.49 | 1.53 | 0.000 |
| 枝横截面积-单叶面积 Twig cross-sectional area-Individual leaf area | 0.94 | 0.97 | 0.000 |
| 枝横截面积-枝稠密度 Twig cross-sectional area-Twig intensity | -0.24 | -0.25 | 0.579 |
| 单叶面积-生叶强度 Individual leaf area-Leafing intensity | 0.37 | 0.44 | 0.000 |
| 组别 Group | 生境类型 Habitat type | 物种数 Species number | 回归系数R2 Regression coefficient | 分组斜率(置信区间) Group slope (CIs) | 共斜率(置信区间) Common slope (CIs) | 截距 Intercept |
|---|---|---|---|---|---|---|
| 枝横截面积-总叶面积 Twig cross-sectional area-total leaf area | 凸型生境 Convex habitat | 65 | 0.76*** | 1.04 (0.92-1.17)a | 1.10 (1.01-1.21)§ | 1.56b |
| 凹型生境 Concave habitat | 45 | 0.81*** | 1.19 (1.04-1.36)a※ | 1.64a | ||
| 枝横截面积-单叶面积 Twig cross-sectional area-individual leaf area | 凸型生境 Convex habitat | 65 | 0.65*** | 0.88 (0.76-1.02)a | 0.91 (0.82-1.01) | 0.97a |
| 凹型生境 Concave habitat | 45 | 0.73*** | 0.94 (0.80-1.10)a | 1.03a | ||
| 枝横截面积-枝稠密度 Twig cross-sectional area-twig intensity | 凸型生境 Convex habitat | 65 | 0.63*** | -1.55 (-1.80- -1.34)a※ | -1.57 (-1.72- -1.43)§ | 0.09a |
| 凹型生境 Concave habitat | 45 | 0.72*** | -1.57 (-1.77- -1.40)a※ | 0.04a | ||
| 单叶面积-生叶强度 Individual leaf area-leafing intensity | 凸型生境 Convex habitat | 65 | 0.66*** | -1.33 (-1.55- -1.14)a※ | -1.28 (-1.43- -1.15)§ | 0.26a |
| 凹型生境 Concave habitat | 45 | 0.85*** | -1.23 (-1.45- -1.05)a※ | 0.33a |
Table 2 Summary of regression parameters based on standardized major axis estimation for scaling relationships of twig size-leaf size, twig size-twig intensity, and leaf size-leafing intensity for plant species in concave and convex habitats
| 组别 Group | 生境类型 Habitat type | 物种数 Species number | 回归系数R2 Regression coefficient | 分组斜率(置信区间) Group slope (CIs) | 共斜率(置信区间) Common slope (CIs) | 截距 Intercept |
|---|---|---|---|---|---|---|
| 枝横截面积-总叶面积 Twig cross-sectional area-total leaf area | 凸型生境 Convex habitat | 65 | 0.76*** | 1.04 (0.92-1.17)a | 1.10 (1.01-1.21)§ | 1.56b |
| 凹型生境 Concave habitat | 45 | 0.81*** | 1.19 (1.04-1.36)a※ | 1.64a | ||
| 枝横截面积-单叶面积 Twig cross-sectional area-individual leaf area | 凸型生境 Convex habitat | 65 | 0.65*** | 0.88 (0.76-1.02)a | 0.91 (0.82-1.01) | 0.97a |
| 凹型生境 Concave habitat | 45 | 0.73*** | 0.94 (0.80-1.10)a | 1.03a | ||
| 枝横截面积-枝稠密度 Twig cross-sectional area-twig intensity | 凸型生境 Convex habitat | 65 | 0.63*** | -1.55 (-1.80- -1.34)a※ | -1.57 (-1.72- -1.43)§ | 0.09a |
| 凹型生境 Concave habitat | 45 | 0.72*** | -1.57 (-1.77- -1.40)a※ | 0.04a | ||
| 单叶面积-生叶强度 Individual leaf area-leafing intensity | 凸型生境 Convex habitat | 65 | 0.66*** | -1.33 (-1.55- -1.14)a※ | -1.28 (-1.43- -1.15)§ | 0.26a |
| 凹型生境 Concave habitat | 45 | 0.85*** | -1.23 (-1.45- -1.05)a※ | 0.33a |
| 生境类型 Habitat type | 个体水平 Individual level | 物种水平 Species level | |||
|---|---|---|---|---|---|
| 凸型生境 Convex habitat | 凹型生境 Concave habitat | 凸型生境 Convex habitat | 凹型生境 Concave habitat | ||
| 总叶面积 Total leaf area (mm2) | 68.68 ± 3.14b | 96.66 ± 5.26a | 136.90 ± 26.09b | 208.80 ± 60.57a | |
| 单叶面积 Individual leaf area (mm2) | 13.96 ± 0.47b | 18.00 ± 0.70a | 26.86 ± 4.84a | 32.46 ± 7.21a | |
| 枝横截面积 Twig cross-sectional area (mm2) | 1.84 ± 0.07b | 2.25 ± 0.09a | 3.15 ± 0.41a | 3.16 ± 0.59a | |
| 枝稠密度 Twig intensity (No.·mm-3) | 0.09 ± 0.04a | 0.08 ± 0.07a | 0.82 ± 1.49a | 0.82 ± 0.18a | |
| 生叶强度 Leafing intensity (No.·mm-3) | 0.13 ± 0.004a | 0.11 ± 0.004b | 0.07 ± 0.01a | 0.07 ± 0.01a | |
Table 3 Trait values for plants in concave and convex habitats (mean ± SE)
| 生境类型 Habitat type | 个体水平 Individual level | 物种水平 Species level | |||
|---|---|---|---|---|---|
| 凸型生境 Convex habitat | 凹型生境 Concave habitat | 凸型生境 Convex habitat | 凹型生境 Concave habitat | ||
| 总叶面积 Total leaf area (mm2) | 68.68 ± 3.14b | 96.66 ± 5.26a | 136.90 ± 26.09b | 208.80 ± 60.57a | |
| 单叶面积 Individual leaf area (mm2) | 13.96 ± 0.47b | 18.00 ± 0.70a | 26.86 ± 4.84a | 32.46 ± 7.21a | |
| 枝横截面积 Twig cross-sectional area (mm2) | 1.84 ± 0.07b | 2.25 ± 0.09a | 3.15 ± 0.41a | 3.16 ± 0.59a | |
| 枝稠密度 Twig intensity (No.·mm-3) | 0.09 ± 0.04a | 0.08 ± 0.07a | 0.82 ± 1.49a | 0.82 ± 0.18a | |
| 生叶强度 Leafing intensity (No.·mm-3) | 0.13 ± 0.004a | 0.11 ± 0.004b | 0.07 ± 0.01a | 0.07 ± 0.01a | |
| [1] |
Ackerly DD, Donoghue MJ (1988). Leaf size, sapling allometry, and Corner’s rules: phylogeny and correlated evolution in maples ( Acer). The American Naturalist, 152, 767-791.
DOI URL PMID |
| [2] | Bond WJ, Midgley J (1988). Allometry and sexual differences in leaf size. The American Naturalist, 131, 901-910. |
| [3] | Bragg JG, Westoby M (2002). Leaf size and foraging for light in a sclerophyll woodland. Functional Ecology, 16, 633-639. |
| [4] | Brouat C, Gibernau M, Amsellem L, McKey D (1998). Corner’s rules revisited: ontogenetic and interspecific patterns in leaf-stem allometry. New Phytologist, 139, 459-470. |
| [5] | Callaway RM, Delucia EH, Schlesinger WH (1994). Biomass allocation of montane and desert ponderosa pine: an analog for response to climate change. Ecology, 75, 1474-1481. |
| [6] | Corner EJH (1949). The durian theory or the origin of the modern tree. Annals of Botany, 13, 367-414. |
| [7] | Enquist BJ, West GB, Charnov EL, Brown JH (1999). Allometric scaling of production and life-history variation in vascular plants. Nature, 401, 907-911. |
| [8] | Falster DS, Warton DI, Wright IJ (2006). User’s Guide to SMATR: Standardised Major Axis Tests & Routines Version 2.0. http://www.bio.mq.edu.au/ecology/SMATR/. Cited 2014-01-13. |
| [9] | Falster DS, Westoby M (2003). Leaf size and angle vary widely across species: What consequences for light interception? New Phytologist, 158, 509-525. |
| [10] | Gartner BL (1991). Stem hydraulic properties of vines vs. shrubs of western poison oak, Toxicodendron diversilobum. Oecologia, 87, 180-189. |
| [11] | Givnish TJ (1987). Comparative studies of leaf form: assessing the relative roles of selective pressures and phylogenetic constraints. New Phytologist, 106(Suppl.), 131-160. |
| [12] | Harvey PH, Pagel MD (1991). The Comparative Method in Evolutionary Biology. Oxford University Press, Oxford, UK. |
| [13] | Huston MA (1994). Biological Diversity: the Coexistence of Species on Changing Landscape. Cambridge University Press, Cambridge, UK. |
| [14] | Jones RH, Sharitz RR, Dixon PM, Segal DS, Schneider RL (1994). Woody plant regeneration in four floodplain forests. Ecological Monographs, 64, 345-367. |
| [15] | Kleiman D, Aarssen LW (2007). The leaf size/number trade-off in trees. Journal of Ecology, 95, 376-382. |
| [16] | Li CY, Berninger F, Koskela J, Sonninen E (2000). Drought responses of Eucalyptus microtheca provenances depend on seasonality of rainfall in their place of origin. Australian Journal of Plant Physiology, 27, 231-238. |
| [17] | Li H, Reynolds JF (1995). On definition and quantification of heterogeneity. Oikos, 73, 280-283. |
| [18] | Liao JX, Chen J, Jiang MX, Huang HD (2012). Leaf traits and persistence of relict and endangered tree species in a rare plant community. Functional Plant Biology, 39, 512-518. |
| [19] | Liu ZG, Cai YL, Li K (2008). Studies on the leaf size-twig size spectrum of subtropical evergreen board-leaved woody species. Journal of Plant Ecology (Chinese Version), 32, 363-369. (in Chinese with English abstract) |
| [刘志国, 蔡永立, 李恺 (2008). 亚热带常绿阔叶林植物叶-小枝的异速生长. 植物生态学报, 32, 363-369.] | |
| [20] |
McCulloh KA, Sperry JS (2005). Patterns in hydraulic architecture and their implications for transport efficiency. Tree Physiology, 25, 257-267.
URL PMID |
| [21] | Milla R (2009). The leafing intensity premium hypothesis tested across clades, growth forms and altitudes. Journal of Ecology, 97, 972-983. |
| [22] | Moles AT, Westoby M (2000). Do small leaves expand faster than large leaves, and do shorter expansion times reduce herbivore damage? Oikos, 90, 517-524. |
| [23] | Niklas KJ (1992). Plant Biomechanics: an Engineering Approach to Plant Form and Function. University of Chicago Press, Chicago. |
| [24] | Niklas KJ (1994). Plant Allometry: the Scaling of Form and Process. The University of Chicago Press, Chicago. |
| [25] | Normand F, Bissery C, Damour G, Lauri PÉ (2008). Hydraulic and mechanical stem properties affect leaf-stem allometry in mango cultivars. New Phytologist, 178, 590-602. |
| [26] | Olson ME, Aguirre-Hernández R, Rosell JA (2009). Universal foliage-stem scaling across environments and species in dicot trees: plasticity, biomechanics and Corner’s rules. Ecology Letters, 12, 210-219. |
| [27] | Pickup M, Westoby M, Basden A (2005). Dry mass costs of deploying leaf area in relation to leaf size. Functional Ecology, 19, 88-97. |
| [28] | Pitman EJG (1939). A note on normal correlation. Biometrika, 31, 9-12. |
| [29] | Preston KA, Ackerly DD (2003). Hydraulic architecture and the evolution of shoot allometry in contrasting climates. American Journal of Botany, 90, 1502-1512. |
| [30] | Reich PB, Oleksyn J (2004). Global patterns of plant leaf N and P in relation to temperature and latitude. Proceedings of the National Academy of Sciences of the United States of America, 101, 11001-11006. |
| [31] | Reynolds HL, Hungate BA, Chapin III FS, D’Antonio CM (1997). Soil heterogeneity and plant competition in an annual grassland. Ecology, 78, 2076-2090. |
| [32] | Silvertown J (2004). Plant coexistence and the niche. Trends in Ecology & Evolution, 19, 605-611. |
| [33] | Song YC, Wang XR (1995). Vegetation and Flora of Tiantong National Forest Park Zhejiang Province. Shanghai Scientific and Technical Document Publishing House, Shanghai. (in Chinese with English abstract) |
| [宋永昌, 王祥荣 (1995). 浙江天童国家森林公园的植被和区系. 上海科学技术文献出版社, 上海.] | |
| [34] | Sprugel DG, Hinckley TM, Schaap W (1991). The theory and practice of branch autonomy. Annual Review of Ecology and Systematics, 22, 309-334. |
| [35] |
Sun SC, Jin DM, Shi PL (2006). The leaf size-twig size spectrum of temperate woody species along an altitudinal gradient: an invariant allometric scaling relationship. Annals of Botany, 97, 97-107.
URL PMID |
| [36] | Tilman D (1994). Competition and biodiversity in spatially structured habitats. Ecology, 75, 2-16. |
| [37] | Vandermeer JH (1972). Niche theory. Annual Review of Ecology and Systematics, 3, 107-132. |
| [38] | Villar-Salvador P, Castro-Díez P, Pérez-Rontomé C, Montserrat-Martí G (1997). Stem xylem features in three Quercus (Fagaceae) species along a climatic gradient in NE Spain. Trees, 12, 90-96. |
| [39] | Warton DI, Weber NC (2002). Common slope tests for bivariate errors-in-variables models. Biometrical Journal, 44, 161-174. |
| [40] | Warton DI, Wright IJ, Falster DS, Westoby M (2006). Bivariate line-fitting methods for allometry. Biological Reviews, 81, 259-291. |
| [41] |
West GB, Brown JH, Enquist BJ (1997). A general model for the origin of allometric scaling laws in biology. Science, 276, 122-126.
URL PMID |
| [42] | West GB, Brown JH, Enquist BJ (1999). A general model for the structure and allometry of plant vascular systems. Nature, 400, 664-667. |
| [43] | Westoby M, Falster DS, Moles AT, Vesk PA, Wright IJ (2002). Plant ecological strategies: some leading dimensions of variation between species. Annual Review of Ecology and Systematics, 33, 125-159. |
| [44] |
Westoby M, Wright IJ (2003). The leaf size-twig size spectrum and its relationship to other important spectra of variation among species. Oecologia, 135, 621-628.
URL PMID |
| [45] | White PS (1983a). allometry and its implications for the adaptive architecture of trees. Bulletin of the Torrey Botanical Club, 110, 203-212. |
| [46] | White PS (1983b). Evidence that temperate East North American evergreen woody plants follow Corner’s rules. New Phytologist, 95, 139-145. |
| [47] | Whitman T, Aarssen LW (2010). The leaf size/number trade-off in herbaceous angiosperms. Journal of Plant Ecology, 3, 49-58. |
| [48] | Xu Y, Yang XD, Xie YM, Xu YL, Chang SX, Yan ER (2012). Twig size-number trade-off among woody plants in Tiantong region, Zhejiang Province of China. Chinese Journal of Plant Ecology, 36, 1268-1276. (in Chinese with English abstract) |
| [许月, 杨晓东, 谢一鸣, 徐艺露, Chang SX, 阎恩荣 (2012). 浙江天童木本植物小枝的“大小-数量”权衡. 植物生态学报, 36, 1268-1276.] | |
| [49] | Yan ER, Milla R, Aarssen LW, Wang XH (2012). Functional relationships of leafing intensity to plant height, growth form and leaf habit. Acta Oecologica, 41, 20-29. |
| [50] |
Yan ER, Wang XH, Chang SX, He FL (2013). Scaling relationships among twig size, leaf size and leafing intensity in a successional series of subtropical forests. Tree Physiology, 33, 609-617.
DOI URL PMID |
| [51] |
Yang DM, Li GY, Sun SC (2008). The generality of leaf size versus number trade-off in temperate woody species. Annals of Botany, 102, 623-629.
DOI URL PMID |
| [52] | Yang DM, Li GY, Sun SC (2009). The effects of leaf size, leaf habit, and leaf form on leaf/stem relationships in plant twigs of temperate woody species. Journal of Vegetation Science, 20, 359-366. |
| [53] | Zhang N, Wang XH, Zheng ZM, Ma ZP, Yang QS, Fang XF, Xie YB (2012). Spatial heterogeneity of soil properties and its relationships with terrain factors in broadleaved forest in Tiantong of Zhejiang Province, East China. Chinese Journal of Applied Ecology, 23, 2361-2369. (in Chinese with English abstract) |
| [张娜, 王希华, 郑泽梅, 马遵平, 杨庆松, 方晓峰, 谢玉彬 (2012). 浙江天童常绿阔叶林土壤的空间异质性及其与地形的关系. 应用生态学报, 23, 2361-2369.] |
| [1] | CHEN Wen-Yi, WANG Zhi-Yong, ZHOU Meng-Yan, MA Wen-Jun, WANG Jun-Hui, LUO Zhi-Bin, ZHOU Jing. Biomass allocation and allometric growth model of young Catalpa bungei [J]. Chin J Plant Ecol, 2025, 49(2): 356-366. |
| [2] | TAO Qiong, MIAO Ning, YUE Xi-Ming, LUO Jian-Qiong, XUE Pan-Pan, WANG Hui. Influencing factors of biomass accumulation and allocation of Abies fargesii var. faxoniana seedlings in the subalpine region of western Sichuan, China [J]. Chin J Plant Ecol, 2024, 48(11): 1459-1470. |
| [3] | Nan DONG, Ming-Ming TANG, Wen-Qian CUI, Meng-Yao YUE, Jie LIU, Yu-Jie HUANG. Growth of chestnut and tea seedlings under different root partitioning patterns [J]. Chin J Plant Ecol, 2022, 46(1): 62-73. |
| [4] | XING Lei, DUAN Na, LI Qing-He, LIU Cheng-Gong, LI Hui-Qing, SUN Gao-Jie. Variation in biomass allocation of Nitraria tangutorum during different phenological phases [J]. Chin J Plant Ecol, 2020, 44(7): 763-771. |
| [5] | CHEN Guo-Peng, YANG Ke-Tong, WANG Li, WANG Fei, CAO Xiu-Wen, CHEN Lin-Sheng. Allometric relations for biomass partitioning of seven alpine Rhododendron species in south of Gansu [J]. Chin J Plant Ecol, 2020, 44(10): 1040-1049. |
| [6] | ZHANG Jian-Hong, SHI Qing-Ru, XU Ming-Shan, ZHAO Yan-Tao, ZHONG Qiang, ZHANG Fu-Jie, YAN En-Rong. Testing of Corner’s rules across woody plants in Tiantong region, Zhejiang Province: effects of individual density [J]. Chin J Plant Ecol, 2014, 38(7): 655-664. |
| [7] | WANG Yi-Kun, JIN Ai-Wu, ZHU Qiang-Gen, QIU Yong-Hua, JI Xin-Liang, ZHANG Si-Hai. Effects of fertilization on the relations of diameter at breast height between different-aged ramets of Phyllostachys edulis population [J]. Chin J Plant Ecol, 2014, 38(3): 289-297. |
| Viewed | ||||||
|
Full text |
|
|||||
|
Abstract |
|
|||||
Copyright © 2026 Chinese Journal of Plant Ecology
Tel: 010-62836134, 62836138, E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
![]()