Relationships between xylem structure and embolism vulnerability in six species of drought tolerance trees
LI Rong1, DANG Wei1, CAI Jing1, 3, ZHANG Shuo-Xin1, 3, JIANG Zai-Min2, *,
1College of Forestry, Northwest A&F University, Yangling, Shaanxi 712100, China 2College of Life Sciences, Northwest A&F University, Yangling, Shaanxi 712100, Chinaand 3Qinling National Forest Ecosystem Research Station, Yangling, Shaanxi 712100, China
通讯作者: * 通讯作者Author for correspondence (E-mail: jiangzmz@163.com)
The study of embolism vulnerability to drought has become a hot and key topic under global climate change. The objective of the study was: 1) to identify the relationship between xylem structure and embolism vulnerability; 2) to define the differences in resistance of embolism in xylem structure of each species; and 3) to establish drought tolerance indexes in xylem structure of six species.
Methods
Drought tolerance trees of Robinia pseudoacacia, Hippophae rhamnoides, Ulmus pumila, Corylus heterophylla, Salix matsudana, Acer truncatum were studied. Cochard Cavitron centrifuge was used to establish embolism vulnerability curves and to calculate xylem vulnerability value. Staining and silicone injection techniques were used to to measure xylem structure of drought tolerance trees including vessel diameter, conduit wall span, number of vessels per unit area, contact faction, vessel length and wood density.
Important findings
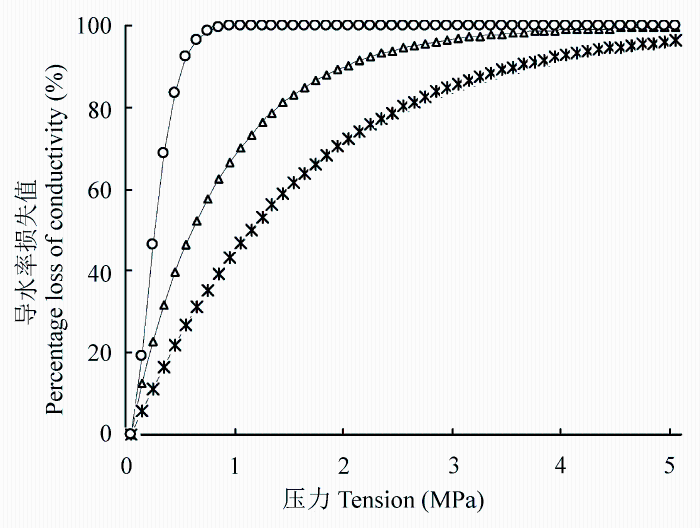
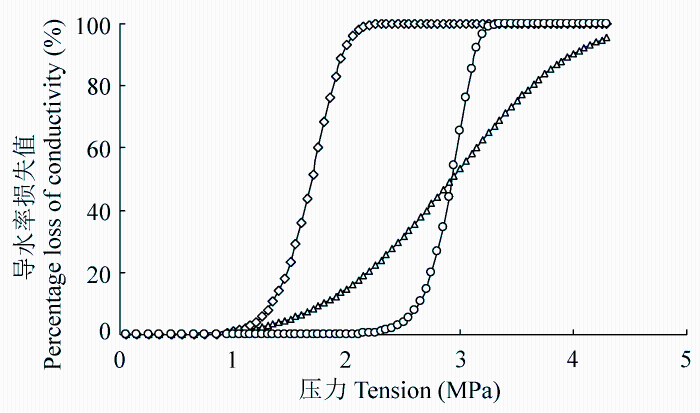
The results showed: 1) xylem embolism vulnerability of the six species ranked as Robinia pseudoacacia > Ulmus pumila > Hippophae rhamnoides > Salix matsudana > Acer truncatum > Corylus heterophylla; 2) the vulnerability curves is “r” shape for Robinia pseudoacacia, Hippophae rhamnoides, Ulmus pumila and is “s” shape for Corylus heterophylla, Salix matsudana, Acer truncatum, respectively; 3) the xylem vulnerability values is significantly different in trees of “r” shape and “s” shape (p < 0.01). Furthermore, linear analysis showed that the different effects between the xylem structure of the species was closely related to the vulnerability in the following order: the maximum effect was from wood density (t = 0.702), the medium effect was from vessel diameter (t = 0.532), and the minimum effect was from vessel length (t = 0.01).
LIRong, DANGWei, CAIJing, ZHANGShuo-Xin, JIANGZai-Min. Relationships between xylem structure and embolism vulnerability in six species of drought tolerance trees[J]. Chinese Journal of Plant Ecology, 2016, 40(3): 255-263 https://doi.org/10.17521/cjpe.2015.0260
干旱是全球性的自然灾害, 近年来因干旱引起的森林枯梢及死亡已严重影响许多国家的森林生态系统(Maherali et al., 2004)。在我国, 干旱限制了干旱半干旱地区植物的生长和生态修复, 生长在干旱半干旱地区的植物在其生命周期内经常遭遇水分的胁迫(Chaves et al., 2002; Rennenberg et al., 2006; Nahm et al., 2007), 由干旱引起的木质部空穴化和栓塞则是制约树木生长发育和生态修复的一个重要因素, 干旱胁迫导致的抗栓塞性在不同树种和不同生态系统表现不同(Delzon et al., 2010), 在全球气候变化的大背景下, 干旱导致的抗栓塞性的研究已越来越重要(Cochard et al., 2013)。木质部栓塞是木本植物在干旱环境下遭受水分胁迫时产生的木质部导管输水功能障碍, 而植物对木质部栓塞的易受性可用“脆弱曲线(VC)”来描述(Tyree & Sperry, 1989), 该曲线是根据木质部导水率损失的百分数(PLC)与相对应水势绘制而成的曲线, 且绝大多数脆弱曲线可用单威布尔累积分布函数(CDF)较好地拟合, 栓塞脆弱性一般用引起枝条50%导水率损失的压力值(P50)来表示。Cochard等(2008)和Awad等(2010)通过对蔷薇科李属(Prunus)的10个树种和杨属(Populus)的2个树种进行栓塞脆弱曲线的测定以及不同干旱条件的处理, 认为抗栓塞性可作为评估植物抵御干旱环境的一个指标。Choat等(2012)和Anderegg等(2013)研究认为抗栓塞性的大小是评估森林生产力和干旱导致的死亡率的一项重要生理指标, 树木栓塞抗性的研究将会有很大发展, 这些研究也将被运用到模型当中, 以估算植物生产力(生理生态模型)以及物种分布(生物地理学模型)。张硕新等(1997)对我国北方5种抗旱树种栓塞脆弱性的研究表明, 耐旱的刺槐(Robinia pseudoacacia)和沙棘(Hipp- ophae rhamnoides)是栓塞脆弱性较大、较易发生栓塞的树种, 在水势下降时木质部导水率急剧下降, 即木质部很快发生栓塞, 而植物栓塞脆弱性与木质部结构密切相关(Wheeler et al., 2005; Sperry et al., 2006; Hacke & Jansen, 2009; Cai & Tyree, 2010; 张海昕等, 2013), 植物栓塞脆弱性与木质部结构关系的研究是了解耐旱树种抗旱机制的一个有效途径。
试验地选取在西北农林科技大学校园和博览园(34.27° N, 108.07° E, 海拔457 m), 样地处于关中平原中部, 属半干旱地区, 暖温带大陆性季风气候, 四季分明, 雨热同季。年平均气温12.9 ℃, 最热月(7月)平均气温25.8 ℃, 最冷月(1月)平均气温-1.1 ℃, 年极端最高气温42 ℃, 年极端最低气温-19.4 ℃, ≥0 ℃积温4903 ℃, 持续天数309天, ≥10 ℃积温4185 ℃, 持续天数206天, 无霜期221天, 年降水量651 mm, 生长季(4-9月份)降水量占全年降水量的79%。
1.2 材料
选取西北农林科技大学校园、博览园内自然状况下生长良好的耐旱树种刺槐、沙棘、榆树(Ulmus pumila)、榛、旱柳(Salix matsudana)、元宝枫为研究对象, 各树种所处立地条件基本一致。于2014年8-10月在清晨在树冠中部北向剪取一年生枝条, 所剪枝条基部直径6-8 mm, 长30-50 cm, 迅速放入装有湿纸的黑色塑料袋中(防止水分散失和外界空气等进入被切开的导管内), 立即带回实验室。
Table 1 The value of tension at 50% loss of conductivity (P50) and characteristic values of xylem structure of the six species (mean ± SE)
因子 Parameter
耐旱树种 Drought tolerance trees
沙棘 Hippophae rhamnoides
刺槐 Robinia pseudoacacia
榆树 Ulmus pumila
元宝枫 Acer truncatum
旱柳 Salix matsudana
榛 Corylus heterophylla
P50 (MPa)
-1.31 ± 0.183Ab
-0.22 ± 0.026Ac
-0.55 ± 0.064Ac
-2.64 ± 0.214Ba
-1.55 ± 0.103Bb
-2.84 ± 0.054Ba
导管直径 Vessel diameter (μm)
23.57 ± 0.579Ac
56.96 ± 1.913Aa
49.48 ± 1.206Aa
31.26 ± 0.405Bb
32.11 ± 0.415Bb
26.81 ± 0.319Bc
导管内径跨度 Conduit wall span (μm)
40.10 ± 1.651Aa
62.20 ± 0.905Ab
58.07 ± 2.414Aa
34.29 ± 0.729Bb
36.19 ± 0.583Bc
27.04 ± 0.749Bb
导管密度 Number of vessels per unit area
216.52 ± 0.006Ab
32.22 ± 2.588Aa
53.64 ± 0.011Aa
143.55 ± 3.874Bb
284.37 ± 0.015Bb
270.11 ± 0.013Bb
导管连接度 Contact fraction
0.03 ± 0.006Aa
0.02 ± 0.002Aa
0.04 ± 0.004Aa
0.13 ± 0.008Bb
0.07 ± 0.007Bb
0.10 ± 0.008Bb
导管长度 Vessel length (cm)
13.35 ± 0.827Ab
20.55 ± 1.913Aa
17.23 ± 0.834Aa
3.08 ± 0.253Bc
4.10 ± 0.397Bc
3.23 ± 0.223Bc
木质部密度 Wood density (g·cm-3)
1.54 ± 0.086Aa
1.17 ± 0.129Aa
1.17 ± 0.062Aa
1.42 ± 0.167Ba
1.46 ± 0.209Ba
0.83 ± 0.039Bb
Different capital letters indicate significant difference (p < 0.01) between trees of “r” and “s” shape and lowercase letters (a or b) indicate significant difference (p < 0.01) of trees by each species.不同大写字母表示脆弱曲线为“r”形的树种与脆弱曲线为“s”形的树种差异显著性(p < 0.01); 不同小写字母表示各树种种间差异显著性(p < 0.01)。
Common trade-offs bet- ween xylem resistance to cavitation and other physiolo- gical traits do not hold among unrelated Populus deltoides × Populus nigra hybrids.
Seasonal courses of key parameters of nitrogen, carbon and waterbalance in European beech (Fagus sylvatica L.) grown on four different study sites along a European North-South climate gradient during the 2003 drought.
Estrategias de resistencia al deficit hídrico en Pistacia lentiscus L. y Quercus coccifera L. Implica- ciones en la repoblación forestal. PhD dissertation, University of Alicante, Alicante
Common trade-offs bet- ween xylem resistance to cavitation and other physiolo- gical traits do not hold among unrelated Populus deltoides × Populus nigra hybrids.
1
2010
... 研究表明木质部导管的导水率与导管直径的四次方成正比(Hagen-Poiseuille方程)(Tyree & Zim- mermann, 2002), 即导管直径越大, 导管的导水能力就越强.多数研究认为木质部导管直径与栓塞脆弱性正相关, 即同一树种内大径导管比小径导管更易发生栓塞(Hargrave et al., 1994; Gullo et al., 1995; Wheeler et al., 2005; Hacke et al., 2006; Cai & Tyree, 2010; 张海昕等, 2013), 但也有研究表明栓塞脆弱性与导管直径成反比(Lemoine et al., 2002; Hacke et al., 2009)或不相关(Rosner et al., 2007; Fichot et al., 2010).Cai和Tyree (2010)以杨属植物为例, 按照植物导管直径径级构建的栓塞脆弱曲线表明: 随着导管直径增大, P50值增大(负值), 即导管直径与P50呈正相关关系.Cai等(2010b)的研究表明同一树种内导管直径与导管长度正相关, 即大径级导管其导管长度也越长, 张海昕等(2013)通过对4个杨树无性系的研究得出相同结论.此外, 有研究表明木质部密度和导管内径跨度(Hacke et al., 2001; Jacobsen et al., 2005; Cochard et al., 2008)、导管密度和导管连接度等与栓塞脆弱性具有一定的相关性(Wheeler et al., 2005; Hacke et al., 2006; Cai et al., 2014).安峰和张硕新(2005)对刺槐、元宝枫(Acer truncatum)、榛(Corylus heterophylla)等研究表明, 同一树种小枝和根的栓塞脆弱性主要由木质部结构决定, 小枝易发生栓塞的, 其根也较易发生栓塞, 根和小枝的栓塞脆弱性与植物的抗栓塞性有关.但目前, 通过木质部结构与栓塞脆弱性的关系来说明分布于我国干旱半干旱地区的耐旱树种在木质部结构上的差异尚未见报道, 因此, 本研究通过采用Cochard Cavitron离心机技术(仅需一根枝条且在0.5 h之内即可建立一条完整的栓塞脆弱曲线)(Cochard, 2002; Cochard et al., 2005)建立6个耐旱树种栓塞脆弱曲线, 计算其木质部P50, 找出6个耐旱树种木质部结构间的差异, 以期获得耐旱树种在木质部结构上的抗栓塞性指标, 为干旱、半干旱地区植被恢复工作中选育节水耐旱树种提供新的理论基础. ...
Relations between vulnerability to xylem embolism and xylem conduit dimensions in young trees of Quercus corris.
1
1995
... 研究表明木质部导管的导水率与导管直径的四次方成正比(Hagen-Poiseuille方程)(Tyree & Zim- mermann, 2002), 即导管直径越大, 导管的导水能力就越强.多数研究认为木质部导管直径与栓塞脆弱性正相关, 即同一树种内大径导管比小径导管更易发生栓塞(Hargrave et al., 1994; Gullo et al., 1995; Wheeler et al., 2005; Hacke et al., 2006; Cai & Tyree, 2010; 张海昕等, 2013), 但也有研究表明栓塞脆弱性与导管直径成反比(Lemoine et al., 2002; Hacke et al., 2009)或不相关(Rosner et al., 2007; Fichot et al., 2010).Cai和Tyree (2010)以杨属植物为例, 按照植物导管直径径级构建的栓塞脆弱曲线表明: 随着导管直径增大, P50值增大(负值), 即导管直径与P50呈正相关关系.Cai等(2010b)的研究表明同一树种内导管直径与导管长度正相关, 即大径级导管其导管长度也越长, 张海昕等(2013)通过对4个杨树无性系的研究得出相同结论.此外, 有研究表明木质部密度和导管内径跨度(Hacke et al., 2001; Jacobsen et al., 2005; Cochard et al., 2008)、导管密度和导管连接度等与栓塞脆弱性具有一定的相关性(Wheeler et al., 2005; Hacke et al., 2006; Cai et al., 2014).安峰和张硕新(2005)对刺槐、元宝枫(Acer truncatum)、榛(Corylus heterophylla)等研究表明, 同一树种小枝和根的栓塞脆弱性主要由木质部结构决定, 小枝易发生栓塞的, 其根也较易发生栓塞, 根和小枝的栓塞脆弱性与植物的抗栓塞性有关.但目前, 通过木质部结构与栓塞脆弱性的关系来说明分布于我国干旱半干旱地区的耐旱树种在木质部结构上的差异尚未见报道, 因此, 本研究通过采用Cochard Cavitron离心机技术(仅需一根枝条且在0.5 h之内即可建立一条完整的栓塞脆弱曲线)(Cochard, 2002; Cochard et al., 2005)建立6个耐旱树种栓塞脆弱曲线, 计算其木质部P50, 找出6个耐旱树种木质部结构间的差异, 以期获得耐旱树种在木质部结构上的抗栓塞性指标, 为干旱、半干旱地区植被恢复工作中选育节水耐旱树种提供新的理论基础. ...
Embolism resistance of three boreal conifer species varies with pit structure.
2
2009
... 干旱是全球性的自然灾害, 近年来因干旱引起的森林枯梢及死亡已严重影响许多国家的森林生态系统(Maherali et al., 2004).在我国, 干旱限制了干旱半干旱地区植物的生长和生态修复, 生长在干旱半干旱地区的植物在其生命周期内经常遭遇水分的胁迫(Chaves et al., 2002; Rennenberg et al., 2006; Nahm et al., 2007), 由干旱引起的木质部空穴化和栓塞则是制约树木生长发育和生态修复的一个重要因素, 干旱胁迫导致的抗栓塞性在不同树种和不同生态系统表现不同(Delzon et al., 2010), 在全球气候变化的大背景下, 干旱导致的抗栓塞性的研究已越来越重要(Cochard et al., 2013).木质部栓塞是木本植物在干旱环境下遭受水分胁迫时产生的木质部导管输水功能障碍, 而植物对木质部栓塞的易受性可用“脆弱曲线(VC)”来描述(Tyree & Sperry, 1989), 该曲线是根据木质部导水率损失的百分数(PLC)与相对应水势绘制而成的曲线, 且绝大多数脆弱曲线可用单威布尔累积分布函数(CDF)较好地拟合, 栓塞脆弱性一般用引起枝条50%导水率损失的压力值(P50)来表示.Cochard等(2008)和Awad等(2010)通过对蔷薇科李属(Prunus)的10个树种和杨属(Populus)的2个树种进行栓塞脆弱曲线的测定以及不同干旱条件的处理, 认为抗栓塞性可作为评估植物抵御干旱环境的一个指标.Choat等(2012)和Anderegg等(2013)研究认为抗栓塞性的大小是评估森林生产力和干旱导致的死亡率的一项重要生理指标, 树木栓塞抗性的研究将会有很大发展, 这些研究也将被运用到模型当中, 以估算植物生产力(生理生态模型)以及物种分布(生物地理学模型).张硕新等(1997)对我国北方5种抗旱树种栓塞脆弱性的研究表明, 耐旱的刺槐(Robinia pseudoacacia)和沙棘(Hipp- ophae rhamnoides)是栓塞脆弱性较大、较易发生栓塞的树种, 在水势下降时木质部导水率急剧下降, 即木质部很快发生栓塞, 而植物栓塞脆弱性与木质部结构密切相关(Wheeler et al., 2005; Sperry et al., 2006; Hacke & Jansen, 2009; Cai & Tyree, 2010; 张海昕等, 2013), 植物栓塞脆弱性与木质部结构关系的研究是了解耐旱树种抗旱机制的一个有效途径. ...
... 研究表明木质部导管的导水率与导管直径的四次方成正比(Hagen-Poiseuille方程)(Tyree & Zim- mermann, 2002), 即导管直径越大, 导管的导水能力就越强.多数研究认为木质部导管直径与栓塞脆弱性正相关, 即同一树种内大径导管比小径导管更易发生栓塞(Hargrave et al., 1994; Gullo et al., 1995; Wheeler et al., 2005; Hacke et al., 2006; Cai & Tyree, 2010; 张海昕等, 2013), 但也有研究表明栓塞脆弱性与导管直径成反比(Lemoine et al., 2002; Hacke et al., 2009)或不相关(Rosner et al., 2007; Fichot et al., 2010).Cai和Tyree (2010)以杨属植物为例, 按照植物导管直径径级构建的栓塞脆弱曲线表明: 随着导管直径增大, P50值增大(负值), 即导管直径与P50呈正相关关系.Cai等(2010b)的研究表明同一树种内导管直径与导管长度正相关, 即大径级导管其导管长度也越长, 张海昕等(2013)通过对4个杨树无性系的研究得出相同结论.此外, 有研究表明木质部密度和导管内径跨度(Hacke et al., 2001; Jacobsen et al., 2005; Cochard et al., 2008)、导管密度和导管连接度等与栓塞脆弱性具有一定的相关性(Wheeler et al., 2005; Hacke et al., 2006; Cai et al., 2014).安峰和张硕新(2005)对刺槐、元宝枫(Acer truncatum)、榛(Corylus heterophylla)等研究表明, 同一树种小枝和根的栓塞脆弱性主要由木质部结构决定, 小枝易发生栓塞的, 其根也较易发生栓塞, 根和小枝的栓塞脆弱性与植物的抗栓塞性有关.但目前, 通过木质部结构与栓塞脆弱性的关系来说明分布于我国干旱半干旱地区的耐旱树种在木质部结构上的差异尚未见报道, 因此, 本研究通过采用Cochard Cavitron离心机技术(仅需一根枝条且在0.5 h之内即可建立一条完整的栓塞脆弱曲线)(Cochard, 2002; Cochard et al., 2005)建立6个耐旱树种栓塞脆弱曲线, 计算其木质部P50, 找出6个耐旱树种木质部结构间的差异, 以期获得耐旱树种在木质部结构上的抗栓塞性指标, 为干旱、半干旱地区植被恢复工作中选育节水耐旱树种提供新的理论基础. ...
Xylem function of arid-land shrubs from California, USA: An ecological and evolutionary analysis.
2009
Water transport in vessel less angiosperms: Conducting efficiency and cavitation safety.
1
2007
... 用硅胶注射法进行导管长度的测定(Wheeler et al., 2005; Hacke et al., 2007; Cai et al., 2010a) : 将清晨采回的枝条(长度30-50 cm)用刀片将切面切平, 然后在0.12 MPa压力下用100 mmol·L-1去气KCl溶液冲洗枝条30 min.硅胶配置: 硅胶是硅胶树脂和固化剂以10:1混合而成(15 g RTV141 part A + 1.5 g RTV141 part B), 0.025 g荧光增白剂Uvitex (使硅胶在紫外光下可见)与2.5 mL氯仿溶解后取15滴与硅胶混合, 搅拌使二者混合均匀后静置1 h以除去硅胶中的气泡.从枝条冲洗端橡胶管注入硅胶(硅胶占橡胶管的2/3即可), 在0.12 MPa压力下持续注射24 h, 之后在室温22 ℃条件下干燥2-3天, 干燥后切片(厚度20 μm), 切片分别从距离注射硅胶端的0.2、0.3、0.5、1.0、1.5、2.0、3.0、4.0 cm直到被注射导管个数占整个扇面导管个数的2%为止, 选取被注射上导管均匀分布的3个扇面获取照片(操作同上).所得照片用Win-CELL 2007图像分析软件统计出单位面积内被硅胶注射的导管数目, 由此可得单位横截面内导管的数量(N =横截面所有被硅胶注射导管的数量/横截面的面积).以截面到注射端的距离x为横坐标, 以lnN为纵坐标进行线性拟合, 得出线性拟合图的斜率λv (Cohen et al., 2003). ...
Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure.
5
2001
... 研究表明木质部导管的导水率与导管直径的四次方成正比(Hagen-Poiseuille方程)(Tyree & Zim- mermann, 2002), 即导管直径越大, 导管的导水能力就越强.多数研究认为木质部导管直径与栓塞脆弱性正相关, 即同一树种内大径导管比小径导管更易发生栓塞(Hargrave et al., 1994; Gullo et al., 1995; Wheeler et al., 2005; Hacke et al., 2006; Cai & Tyree, 2010; 张海昕等, 2013), 但也有研究表明栓塞脆弱性与导管直径成反比(Lemoine et al., 2002; Hacke et al., 2009)或不相关(Rosner et al., 2007; Fichot et al., 2010).Cai和Tyree (2010)以杨属植物为例, 按照植物导管直径径级构建的栓塞脆弱曲线表明: 随着导管直径增大, P50值增大(负值), 即导管直径与P50呈正相关关系.Cai等(2010b)的研究表明同一树种内导管直径与导管长度正相关, 即大径级导管其导管长度也越长, 张海昕等(2013)通过对4个杨树无性系的研究得出相同结论.此外, 有研究表明木质部密度和导管内径跨度(Hacke et al., 2001; Jacobsen et al., 2005; Cochard et al., 2008)、导管密度和导管连接度等与栓塞脆弱性具有一定的相关性(Wheeler et al., 2005; Hacke et al., 2006; Cai et al., 2014).安峰和张硕新(2005)对刺槐、元宝枫(Acer truncatum)、榛(Corylus heterophylla)等研究表明, 同一树种小枝和根的栓塞脆弱性主要由木质部结构决定, 小枝易发生栓塞的, 其根也较易发生栓塞, 根和小枝的栓塞脆弱性与植物的抗栓塞性有关.但目前, 通过木质部结构与栓塞脆弱性的关系来说明分布于我国干旱半干旱地区的耐旱树种在木质部结构上的差异尚未见报道, 因此, 本研究通过采用Cochard Cavitron离心机技术(仅需一根枝条且在0.5 h之内即可建立一条完整的栓塞脆弱曲线)(Cochard, 2002; Cochard et al., 2005)建立6个耐旱树种栓塞脆弱曲线, 计算其木质部P50, 找出6个耐旱树种木质部结构间的差异, 以期获得耐旱树种在木质部结构上的抗栓塞性指标, 为干旱、半干旱地区植被恢复工作中选育节水耐旱树种提供新的理论基础. ...
... 取同样测量导管直径的照片, 用Win-CELL 2007图像分析软件测量每个横切面上相连导管的内径跨度(b)(Hacke et al., 2001).5个枝条的导管内径跨度平均值即为每树种的平均导管内径跨度. ...
... 每个枝条取一定枝段, 测定枝段干质量和鲜体积, 则木质部密度(Dt) (Hacke et al., 2001)为: ...
... 一是因为刺槐和榆树导管直径较旱柳、元宝枫和榛的导管直径大, 这与前人的具有大径导管植物比小径导管植物更易栓塞观点相同(Wheeler et al., 2005; Hacke et al., 2006; Markesteijn et al., 2011; Cai et al., 2010b; 张海昕等, 2013), 刺槐和榆树之所以容易栓塞是气种假说(air-seeding hypothesis)机理中的导管壁间纹孔膜的表面积所决定的(Wheeler et al., 2005), 纹孔膜在木质部水分传导过程中充当重要的角色, 其上有大量的微孔, 是水流从一个导管(管胞)流向另一个导管(管胞)的入口(Choat et al., 2007; Loepfe et al., 2007), 纹孔面积假说(pit area hypothesis)又是气种假说的必然结果, 按照气种假说, P50值由导管上所有纹孔面积中具有最大孔洞的纹孔决定, 因此, 导管直径越大, 导管上纹孔所占面积越大, 纹孔膜上拥有“易漏”的微孔的几率就越高, 植物栓塞脆弱性就大.导管直径、导管壁上纹孔面积与P50的相关性很接近(R2介于0.72-0.75), 它们在水分运输的有效性与安全性间具有权衡性(Wheeler et al., 2005, Cai et al., 2010b).二是因为沙棘、刺槐和榆树较旱柳、元宝枫和榛的导管内径跨度大, 导管内径跨度越大, 在负水势条件下导管腔越易垮塌, 植物越易栓塞, 这与Hargrave等(1994)的研究结果相同.三是因为沙棘、刺槐和榆树的木质部导管密度小、导管连接度低, 而旱柳、元宝枫和榛则恰好相反, 在低水势条件下, 水分可通过相邻导管或相邻导管壁纹孔进行水分运输, 而沙棘、刺槐和榆树通过相邻导管或纹孔运输水分几率显著小于旱柳、元宝枫和榛, 因此, 沙棘、刺槐和榆树较其他三树种导管易发生栓塞, 这支持Hacke等(2006) 的研究结果.四是因为刺槐和榆树木质部密度小于旱柳、元宝枫和榛, 木质部密度越小, 植物栓塞脆弱性越大, 这也与前人研究结果(Hacke et al., 2001; Jacobsen et al., 2005)相同.此外, 沙棘、刺槐和榆树与旱柳、元宝枫和榛相比, 前者较后者的导管直径大、导管长度长, 植物运输水分速度快, 从而较易发生栓塞. ...
The relevance of xylem network structure for plant hydraulic efficiency and safety.
1
2007
... 一是因为刺槐和榆树导管直径较旱柳、元宝枫和榛的导管直径大, 这与前人的具有大径导管植物比小径导管植物更易栓塞观点相同(Wheeler et al., 2005; Hacke et al., 2006; Markesteijn et al., 2011; Cai et al., 2010b; 张海昕等, 2013), 刺槐和榆树之所以容易栓塞是气种假说(air-seeding hypothesis)机理中的导管壁间纹孔膜的表面积所决定的(Wheeler et al., 2005), 纹孔膜在木质部水分传导过程中充当重要的角色, 其上有大量的微孔, 是水流从一个导管(管胞)流向另一个导管(管胞)的入口(Choat et al., 2007; Loepfe et al., 2007), 纹孔面积假说(pit area hypothesis)又是气种假说的必然结果, 按照气种假说, P50值由导管上所有纹孔面积中具有最大孔洞的纹孔决定, 因此, 导管直径越大, 导管上纹孔所占面积越大, 纹孔膜上拥有“易漏”的微孔的几率就越高, 植物栓塞脆弱性就大.导管直径、导管壁上纹孔面积与P50的相关性很接近(R2介于0.72-0.75), 它们在水分运输的有效性与安全性间具有权衡性(Wheeler et al., 2005, Cai et al., 2010b).二是因为沙棘、刺槐和榆树较旱柳、元宝枫和榛的导管内径跨度大, 导管内径跨度越大, 在负水势条件下导管腔越易垮塌, 植物越易栓塞, 这与Hargrave等(1994)的研究结果相同.三是因为沙棘、刺槐和榆树的木质部导管密度小、导管连接度低, 而旱柳、元宝枫和榛则恰好相反, 在低水势条件下, 水分可通过相邻导管或相邻导管壁纹孔进行水分运输, 而沙棘、刺槐和榆树通过相邻导管或纹孔运输水分几率显著小于旱柳、元宝枫和榛, 因此, 沙棘、刺槐和榆树较其他三树种导管易发生栓塞, 这支持Hacke等(2006) 的研究结果.四是因为刺槐和榆树木质部密度小于旱柳、元宝枫和榛, 木质部密度越小, 植物栓塞脆弱性越大, 这也与前人研究结果(Hacke et al., 2001; Jacobsen et al., 2005)相同.此外, 沙棘、刺槐和榆树与旱柳、元宝枫和榛相比, 前者较后者的导管直径大、导管长度长, 植物运输水分速度快, 从而较易发生栓塞. ...
Adaptive variation in the vulnerability of woody plants to xylem cavitation.
1
2004
... 干旱是全球性的自然灾害, 近年来因干旱引起的森林枯梢及死亡已严重影响许多国家的森林生态系统(Maherali et al., 2004).在我国, 干旱限制了干旱半干旱地区植物的生长和生态修复, 生长在干旱半干旱地区的植物在其生命周期内经常遭遇水分的胁迫(Chaves et al., 2002; Rennenberg et al., 2006; Nahm et al., 2007), 由干旱引起的木质部空穴化和栓塞则是制约树木生长发育和生态修复的一个重要因素, 干旱胁迫导致的抗栓塞性在不同树种和不同生态系统表现不同(Delzon et al., 2010), 在全球气候变化的大背景下, 干旱导致的抗栓塞性的研究已越来越重要(Cochard et al., 2013).木质部栓塞是木本植物在干旱环境下遭受水分胁迫时产生的木质部导管输水功能障碍, 而植物对木质部栓塞的易受性可用“脆弱曲线(VC)”来描述(Tyree & Sperry, 1989), 该曲线是根据木质部导水率损失的百分数(PLC)与相对应水势绘制而成的曲线, 且绝大多数脆弱曲线可用单威布尔累积分布函数(CDF)较好地拟合, 栓塞脆弱性一般用引起枝条50%导水率损失的压力值(P50)来表示.Cochard等(2008)和Awad等(2010)通过对蔷薇科李属(Prunus)的10个树种和杨属(Populus)的2个树种进行栓塞脆弱曲线的测定以及不同干旱条件的处理, 认为抗栓塞性可作为评估植物抵御干旱环境的一个指标.Choat等(2012)和Anderegg等(2013)研究认为抗栓塞性的大小是评估森林生产力和干旱导致的死亡率的一项重要生理指标, 树木栓塞抗性的研究将会有很大发展, 这些研究也将被运用到模型当中, 以估算植物生产力(生理生态模型)以及物种分布(生物地理学模型).张硕新等(1997)对我国北方5种抗旱树种栓塞脆弱性的研究表明, 耐旱的刺槐(Robinia pseudoacacia)和沙棘(Hipp- ophae rhamnoides)是栓塞脆弱性较大、较易发生栓塞的树种, 在水势下降时木质部导水率急剧下降, 即木质部很快发生栓塞, 而植物栓塞脆弱性与木质部结构密切相关(Wheeler et al., 2005; Sperry et al., 2006; Hacke & Jansen, 2009; Cai & Tyree, 2010; 张海昕等, 2013), 植物栓塞脆弱性与木质部结构关系的研究是了解耐旱树种抗旱机制的一个有效途径. ...
Ecological differentiation in xylem cavitation resistance is associated with stem and leaf structural traits.
1
2011
... 一是因为刺槐和榆树导管直径较旱柳、元宝枫和榛的导管直径大, 这与前人的具有大径导管植物比小径导管植物更易栓塞观点相同(Wheeler et al., 2005; Hacke et al., 2006; Markesteijn et al., 2011; Cai et al., 2010b; 张海昕等, 2013), 刺槐和榆树之所以容易栓塞是气种假说(air-seeding hypothesis)机理中的导管壁间纹孔膜的表面积所决定的(Wheeler et al., 2005), 纹孔膜在木质部水分传导过程中充当重要的角色, 其上有大量的微孔, 是水流从一个导管(管胞)流向另一个导管(管胞)的入口(Choat et al., 2007; Loepfe et al., 2007), 纹孔面积假说(pit area hypothesis)又是气种假说的必然结果, 按照气种假说, P50值由导管上所有纹孔面积中具有最大孔洞的纹孔决定, 因此, 导管直径越大, 导管上纹孔所占面积越大, 纹孔膜上拥有“易漏”的微孔的几率就越高, 植物栓塞脆弱性就大.导管直径、导管壁上纹孔面积与P50的相关性很接近(R2介于0.72-0.75), 它们在水分运输的有效性与安全性间具有权衡性(Wheeler et al., 2005, Cai et al., 2010b).二是因为沙棘、刺槐和榆树较旱柳、元宝枫和榛的导管内径跨度大, 导管内径跨度越大, 在负水势条件下导管腔越易垮塌, 植物越易栓塞, 这与Hargrave等(1994)的研究结果相同.三是因为沙棘、刺槐和榆树的木质部导管密度小、导管连接度低, 而旱柳、元宝枫和榛则恰好相反, 在低水势条件下, 水分可通过相邻导管或相邻导管壁纹孔进行水分运输, 而沙棘、刺槐和榆树通过相邻导管或纹孔运输水分几率显著小于旱柳、元宝枫和榛, 因此, 沙棘、刺槐和榆树较其他三树种导管易发生栓塞, 这支持Hacke等(2006) 的研究结果.四是因为刺槐和榆树木质部密度小于旱柳、元宝枫和榛, 木质部密度越小, 植物栓塞脆弱性越大, 这也与前人研究结果(Hacke et al., 2001; Jacobsen et al., 2005)相同.此外, 沙棘、刺槐和榆树与旱柳、元宝枫和榛相比, 前者较后者的导管直径大、导管长度长, 植物运输水分速度快, 从而较易发生栓塞. ...
Seasonal courses of key parameters of nitrogen, carbon and waterbalance in European beech (Fagus sylvatica L.) grown on four different study sites along a European North-South climate gradient during the 2003 drought.
1
2007
... 干旱是全球性的自然灾害, 近年来因干旱引起的森林枯梢及死亡已严重影响许多国家的森林生态系统(Maherali et al., 2004).在我国, 干旱限制了干旱半干旱地区植物的生长和生态修复, 生长在干旱半干旱地区的植物在其生命周期内经常遭遇水分的胁迫(Chaves et al., 2002; Rennenberg et al., 2006; Nahm et al., 2007), 由干旱引起的木质部空穴化和栓塞则是制约树木生长发育和生态修复的一个重要因素, 干旱胁迫导致的抗栓塞性在不同树种和不同生态系统表现不同(Delzon et al., 2010), 在全球气候变化的大背景下, 干旱导致的抗栓塞性的研究已越来越重要(Cochard et al., 2013).木质部栓塞是木本植物在干旱环境下遭受水分胁迫时产生的木质部导管输水功能障碍, 而植物对木质部栓塞的易受性可用“脆弱曲线(VC)”来描述(Tyree & Sperry, 1989), 该曲线是根据木质部导水率损失的百分数(PLC)与相对应水势绘制而成的曲线, 且绝大多数脆弱曲线可用单威布尔累积分布函数(CDF)较好地拟合, 栓塞脆弱性一般用引起枝条50%导水率损失的压力值(P50)来表示.Cochard等(2008)和Awad等(2010)通过对蔷薇科李属(Prunus)的10个树种和杨属(Populus)的2个树种进行栓塞脆弱曲线的测定以及不同干旱条件的处理, 认为抗栓塞性可作为评估植物抵御干旱环境的一个指标.Choat等(2012)和Anderegg等(2013)研究认为抗栓塞性的大小是评估森林生产力和干旱导致的死亡率的一项重要生理指标, 树木栓塞抗性的研究将会有很大发展, 这些研究也将被运用到模型当中, 以估算植物生产力(生理生态模型)以及物种分布(生物地理学模型).张硕新等(1997)对我国北方5种抗旱树种栓塞脆弱性的研究表明, 耐旱的刺槐(Robinia pseudoacacia)和沙棘(Hipp- ophae rhamnoides)是栓塞脆弱性较大、较易发生栓塞的树种, 在水势下降时木质部导水率急剧下降, 即木质部很快发生栓塞, 而植物栓塞脆弱性与木质部结构密切相关(Wheeler et al., 2005; Sperry et al., 2006; Hacke & Jansen, 2009; Cai & Tyree, 2010; 张海昕等, 2013), 植物栓塞脆弱性与木质部结构关系的研究是了解耐旱树种抗旱机制的一个有效途径. ...
Physiological responses of forest trees to heat and drought.
1
2006
... 干旱是全球性的自然灾害, 近年来因干旱引起的森林枯梢及死亡已严重影响许多国家的森林生态系统(Maherali et al., 2004).在我国, 干旱限制了干旱半干旱地区植物的生长和生态修复, 生长在干旱半干旱地区的植物在其生命周期内经常遭遇水分的胁迫(Chaves et al., 2002; Rennenberg et al., 2006; Nahm et al., 2007), 由干旱引起的木质部空穴化和栓塞则是制约树木生长发育和生态修复的一个重要因素, 干旱胁迫导致的抗栓塞性在不同树种和不同生态系统表现不同(Delzon et al., 2010), 在全球气候变化的大背景下, 干旱导致的抗栓塞性的研究已越来越重要(Cochard et al., 2013).木质部栓塞是木本植物在干旱环境下遭受水分胁迫时产生的木质部导管输水功能障碍, 而植物对木质部栓塞的易受性可用“脆弱曲线(VC)”来描述(Tyree & Sperry, 1989), 该曲线是根据木质部导水率损失的百分数(PLC)与相对应水势绘制而成的曲线, 且绝大多数脆弱曲线可用单威布尔累积分布函数(CDF)较好地拟合, 栓塞脆弱性一般用引起枝条50%导水率损失的压力值(P50)来表示.Cochard等(2008)和Awad等(2010)通过对蔷薇科李属(Prunus)的10个树种和杨属(Populus)的2个树种进行栓塞脆弱曲线的测定以及不同干旱条件的处理, 认为抗栓塞性可作为评估植物抵御干旱环境的一个指标.Choat等(2012)和Anderegg等(2013)研究认为抗栓塞性的大小是评估森林生产力和干旱导致的死亡率的一项重要生理指标, 树木栓塞抗性的研究将会有很大发展, 这些研究也将被运用到模型当中, 以估算植物生产力(生理生态模型)以及物种分布(生物地理学模型).张硕新等(1997)对我国北方5种抗旱树种栓塞脆弱性的研究表明, 耐旱的刺槐(Robinia pseudoacacia)和沙棘(Hipp- ophae rhamnoides)是栓塞脆弱性较大、较易发生栓塞的树种, 在水势下降时木质部导水率急剧下降, 即木质部很快发生栓塞, 而植物栓塞脆弱性与木质部结构密切相关(Wheeler et al., 2005; Sperry et al., 2006; Hacke & Jansen, 2009; Cai & Tyree, 2010; 张海昕等, 2013), 植物栓塞脆弱性与木质部结构关系的研究是了解耐旱树种抗旱机制的一个有效途径. ...
Hydraulic and mechanical properties of young Norway spruce clones related to growth and wood structure.
1
2007
... 研究表明木质部导管的导水率与导管直径的四次方成正比(Hagen-Poiseuille方程)(Tyree & Zim- mermann, 2002), 即导管直径越大, 导管的导水能力就越强.多数研究认为木质部导管直径与栓塞脆弱性正相关, 即同一树种内大径导管比小径导管更易发生栓塞(Hargrave et al., 1994; Gullo et al., 1995; Wheeler et al., 2005; Hacke et al., 2006; Cai & Tyree, 2010; 张海昕等, 2013), 但也有研究表明栓塞脆弱性与导管直径成反比(Lemoine et al., 2002; Hacke et al., 2009)或不相关(Rosner et al., 2007; Fichot et al., 2010).Cai和Tyree (2010)以杨属植物为例, 按照植物导管直径径级构建的栓塞脆弱曲线表明: 随着导管直径增大, P50值增大(负值), 即导管直径与P50呈正相关关系.Cai等(2010b)的研究表明同一树种内导管直径与导管长度正相关, 即大径级导管其导管长度也越长, 张海昕等(2013)通过对4个杨树无性系的研究得出相同结论.此外, 有研究表明木质部密度和导管内径跨度(Hacke et al., 2001; Jacobsen et al., 2005; Cochard et al., 2008)、导管密度和导管连接度等与栓塞脆弱性具有一定的相关性(Wheeler et al., 2005; Hacke et al., 2006; Cai et al., 2014).安峰和张硕新(2005)对刺槐、元宝枫(Acer truncatum)、榛(Corylus heterophylla)等研究表明, 同一树种小枝和根的栓塞脆弱性主要由木质部结构决定, 小枝易发生栓塞的, 其根也较易发生栓塞, 根和小枝的栓塞脆弱性与植物的抗栓塞性有关.但目前, 通过木质部结构与栓塞脆弱性的关系来说明分布于我国干旱半干旱地区的耐旱树种在木质部结构上的差异尚未见报道, 因此, 本研究通过采用Cochard Cavitron离心机技术(仅需一根枝条且在0.5 h之内即可建立一条完整的栓塞脆弱曲线)(Cochard, 2002; Cochard et al., 2005)建立6个耐旱树种栓塞脆弱曲线, 计算其木质部P50, 找出6个耐旱树种木质部结构间的差异, 以期获得耐旱树种在木质部结构上的抗栓塞性指标, 为干旱、半干旱地区植被恢复工作中选育节水耐旱树种提供新的理论基础. ...
Vulnerability curves by centrifugation: Is there an open vessel artefact, and are ‘r’ shaped curves necessarily invalid?
2012
Size and function in conifer tracheids and angiosperm vessels.
1
2006
... 干旱是全球性的自然灾害, 近年来因干旱引起的森林枯梢及死亡已严重影响许多国家的森林生态系统(Maherali et al., 2004).在我国, 干旱限制了干旱半干旱地区植物的生长和生态修复, 生长在干旱半干旱地区的植物在其生命周期内经常遭遇水分的胁迫(Chaves et al., 2002; Rennenberg et al., 2006; Nahm et al., 2007), 由干旱引起的木质部空穴化和栓塞则是制约树木生长发育和生态修复的一个重要因素, 干旱胁迫导致的抗栓塞性在不同树种和不同生态系统表现不同(Delzon et al., 2010), 在全球气候变化的大背景下, 干旱导致的抗栓塞性的研究已越来越重要(Cochard et al., 2013).木质部栓塞是木本植物在干旱环境下遭受水分胁迫时产生的木质部导管输水功能障碍, 而植物对木质部栓塞的易受性可用“脆弱曲线(VC)”来描述(Tyree & Sperry, 1989), 该曲线是根据木质部导水率损失的百分数(PLC)与相对应水势绘制而成的曲线, 且绝大多数脆弱曲线可用单威布尔累积分布函数(CDF)较好地拟合, 栓塞脆弱性一般用引起枝条50%导水率损失的压力值(P50)来表示.Cochard等(2008)和Awad等(2010)通过对蔷薇科李属(Prunus)的10个树种和杨属(Populus)的2个树种进行栓塞脆弱曲线的测定以及不同干旱条件的处理, 认为抗栓塞性可作为评估植物抵御干旱环境的一个指标.Choat等(2012)和Anderegg等(2013)研究认为抗栓塞性的大小是评估森林生产力和干旱导致的死亡率的一项重要生理指标, 树木栓塞抗性的研究将会有很大发展, 这些研究也将被运用到模型当中, 以估算植物生产力(生理生态模型)以及物种分布(生物地理学模型).张硕新等(1997)对我国北方5种抗旱树种栓塞脆弱性的研究表明, 耐旱的刺槐(Robinia pseudoacacia)和沙棘(Hipp- ophae rhamnoides)是栓塞脆弱性较大、较易发生栓塞的树种, 在水势下降时木质部导水率急剧下降, 即木质部很快发生栓塞, 而植物栓塞脆弱性与木质部结构密切相关(Wheeler et al., 2005; Sperry et al., 2006; Hacke & Jansen, 2009; Cai & Tyree, 2010; 张海昕等, 2013), 植物栓塞脆弱性与木质部结构关系的研究是了解耐旱树种抗旱机制的一个有效途径. ...
Vulnerability of xylem to cavitation and embolism.
1
1989
... 干旱是全球性的自然灾害, 近年来因干旱引起的森林枯梢及死亡已严重影响许多国家的森林生态系统(Maherali et al., 2004).在我国, 干旱限制了干旱半干旱地区植物的生长和生态修复, 生长在干旱半干旱地区的植物在其生命周期内经常遭遇水分的胁迫(Chaves et al., 2002; Rennenberg et al., 2006; Nahm et al., 2007), 由干旱引起的木质部空穴化和栓塞则是制约树木生长发育和生态修复的一个重要因素, 干旱胁迫导致的抗栓塞性在不同树种和不同生态系统表现不同(Delzon et al., 2010), 在全球气候变化的大背景下, 干旱导致的抗栓塞性的研究已越来越重要(Cochard et al., 2013).木质部栓塞是木本植物在干旱环境下遭受水分胁迫时产生的木质部导管输水功能障碍, 而植物对木质部栓塞的易受性可用“脆弱曲线(VC)”来描述(Tyree & Sperry, 1989), 该曲线是根据木质部导水率损失的百分数(PLC)与相对应水势绘制而成的曲线, 且绝大多数脆弱曲线可用单威布尔累积分布函数(CDF)较好地拟合, 栓塞脆弱性一般用引起枝条50%导水率损失的压力值(P50)来表示.Cochard等(2008)和Awad等(2010)通过对蔷薇科李属(Prunus)的10个树种和杨属(Populus)的2个树种进行栓塞脆弱曲线的测定以及不同干旱条件的处理, 认为抗栓塞性可作为评估植物抵御干旱环境的一个指标.Choat等(2012)和Anderegg等(2013)研究认为抗栓塞性的大小是评估森林生产力和干旱导致的死亡率的一项重要生理指标, 树木栓塞抗性的研究将会有很大发展, 这些研究也将被运用到模型当中, 以估算植物生产力(生理生态模型)以及物种分布(生物地理学模型).张硕新等(1997)对我国北方5种抗旱树种栓塞脆弱性的研究表明, 耐旱的刺槐(Robinia pseudoacacia)和沙棘(Hipp- ophae rhamnoides)是栓塞脆弱性较大、较易发生栓塞的树种, 在水势下降时木质部导水率急剧下降, 即木质部很快发生栓塞, 而植物栓塞脆弱性与木质部结构密切相关(Wheeler et al., 2005; Sperry et al., 2006; Hacke & Jansen, 2009; Cai & Tyree, 2010; 张海昕等, 2013), 植物栓塞脆弱性与木质部结构关系的研究是了解耐旱树种抗旱机制的一个有效途径. ...
Xylem Structure and the Ascent of Sap.
2002
Estrategias de resistencia al deficit hídrico en Pistacia lentiscus L. y Quercus coccifera L. Implica- ciones en la repoblación forestal. PhD dissertation, University of Alicante, Alicante
2002
Cavitation, stomatal conductance and leaf dieback in seedlings of two co-occurring Mediterranean shrubs during an intense drought.
2003
Inter-vessel pitting and cavitation in woody Rosaceae and other vesselled plants: A basis for a safety versus efficiency trade-off in xylem transport.
8
2005
... 干旱是全球性的自然灾害, 近年来因干旱引起的森林枯梢及死亡已严重影响许多国家的森林生态系统(Maherali et al., 2004).在我国, 干旱限制了干旱半干旱地区植物的生长和生态修复, 生长在干旱半干旱地区的植物在其生命周期内经常遭遇水分的胁迫(Chaves et al., 2002; Rennenberg et al., 2006; Nahm et al., 2007), 由干旱引起的木质部空穴化和栓塞则是制约树木生长发育和生态修复的一个重要因素, 干旱胁迫导致的抗栓塞性在不同树种和不同生态系统表现不同(Delzon et al., 2010), 在全球气候变化的大背景下, 干旱导致的抗栓塞性的研究已越来越重要(Cochard et al., 2013).木质部栓塞是木本植物在干旱环境下遭受水分胁迫时产生的木质部导管输水功能障碍, 而植物对木质部栓塞的易受性可用“脆弱曲线(VC)”来描述(Tyree & Sperry, 1989), 该曲线是根据木质部导水率损失的百分数(PLC)与相对应水势绘制而成的曲线, 且绝大多数脆弱曲线可用单威布尔累积分布函数(CDF)较好地拟合, 栓塞脆弱性一般用引起枝条50%导水率损失的压力值(P50)来表示.Cochard等(2008)和Awad等(2010)通过对蔷薇科李属(Prunus)的10个树种和杨属(Populus)的2个树种进行栓塞脆弱曲线的测定以及不同干旱条件的处理, 认为抗栓塞性可作为评估植物抵御干旱环境的一个指标.Choat等(2012)和Anderegg等(2013)研究认为抗栓塞性的大小是评估森林生产力和干旱导致的死亡率的一项重要生理指标, 树木栓塞抗性的研究将会有很大发展, 这些研究也将被运用到模型当中, 以估算植物生产力(生理生态模型)以及物种分布(生物地理学模型).张硕新等(1997)对我国北方5种抗旱树种栓塞脆弱性的研究表明, 耐旱的刺槐(Robinia pseudoacacia)和沙棘(Hipp- ophae rhamnoides)是栓塞脆弱性较大、较易发生栓塞的树种, 在水势下降时木质部导水率急剧下降, 即木质部很快发生栓塞, 而植物栓塞脆弱性与木质部结构密切相关(Wheeler et al., 2005; Sperry et al., 2006; Hacke & Jansen, 2009; Cai & Tyree, 2010; 张海昕等, 2013), 植物栓塞脆弱性与木质部结构关系的研究是了解耐旱树种抗旱机制的一个有效途径. ...
... 研究表明木质部导管的导水率与导管直径的四次方成正比(Hagen-Poiseuille方程)(Tyree & Zim- mermann, 2002), 即导管直径越大, 导管的导水能力就越强.多数研究认为木质部导管直径与栓塞脆弱性正相关, 即同一树种内大径导管比小径导管更易发生栓塞(Hargrave et al., 1994; Gullo et al., 1995; Wheeler et al., 2005; Hacke et al., 2006; Cai & Tyree, 2010; 张海昕等, 2013), 但也有研究表明栓塞脆弱性与导管直径成反比(Lemoine et al., 2002; Hacke et al., 2009)或不相关(Rosner et al., 2007; Fichot et al., 2010).Cai和Tyree (2010)以杨属植物为例, 按照植物导管直径径级构建的栓塞脆弱曲线表明: 随着导管直径增大, P50值增大(负值), 即导管直径与P50呈正相关关系.Cai等(2010b)的研究表明同一树种内导管直径与导管长度正相关, 即大径级导管其导管长度也越长, 张海昕等(2013)通过对4个杨树无性系的研究得出相同结论.此外, 有研究表明木质部密度和导管内径跨度(Hacke et al., 2001; Jacobsen et al., 2005; Cochard et al., 2008)、导管密度和导管连接度等与栓塞脆弱性具有一定的相关性(Wheeler et al., 2005; Hacke et al., 2006; Cai et al., 2014).安峰和张硕新(2005)对刺槐、元宝枫(Acer truncatum)、榛(Corylus heterophylla)等研究表明, 同一树种小枝和根的栓塞脆弱性主要由木质部结构决定, 小枝易发生栓塞的, 其根也较易发生栓塞, 根和小枝的栓塞脆弱性与植物的抗栓塞性有关.但目前, 通过木质部结构与栓塞脆弱性的关系来说明分布于我国干旱半干旱地区的耐旱树种在木质部结构上的差异尚未见报道, 因此, 本研究通过采用Cochard Cavitron离心机技术(仅需一根枝条且在0.5 h之内即可建立一条完整的栓塞脆弱曲线)(Cochard, 2002; Cochard et al., 2005)建立6个耐旱树种栓塞脆弱曲线, 计算其木质部P50, 找出6个耐旱树种木质部结构间的差异, 以期获得耐旱树种在木质部结构上的抗栓塞性指标, 为干旱、半干旱地区植被恢复工作中选育节水耐旱树种提供新的理论基础. ...
... )、导管密度和导管连接度等与栓塞脆弱性具有一定的相关性(Wheeler et al., 2005; Hacke et al., 2006; Cai et al., 2014).安峰和张硕新(2005)对刺槐、元宝枫(Acer truncatum)、榛(Corylus heterophylla)等研究表明, 同一树种小枝和根的栓塞脆弱性主要由木质部结构决定, 小枝易发生栓塞的, 其根也较易发生栓塞, 根和小枝的栓塞脆弱性与植物的抗栓塞性有关.但目前, 通过木质部结构与栓塞脆弱性的关系来说明分布于我国干旱半干旱地区的耐旱树种在木质部结构上的差异尚未见报道, 因此, 本研究通过采用Cochard Cavitron离心机技术(仅需一根枝条且在0.5 h之内即可建立一条完整的栓塞脆弱曲线)(Cochard, 2002; Cochard et al., 2005)建立6个耐旱树种栓塞脆弱曲线, 计算其木质部P50, 找出6个耐旱树种木质部结构间的差异, 以期获得耐旱树种在木质部结构上的抗栓塞性指标, 为干旱、半干旱地区植被恢复工作中选育节水耐旱树种提供新的理论基础. ...
... 取同样测量导管直径的照片, 用Win-CELL 2007图像分析软件测量每个扇面中导管与导管之间相连长度占该扇面中所有导管周长的比例, 即导管连接度(Cf) (Wheeler et al., 2005).5个枝条的导管连接度平均值即为每树种的平均导管连接度. ...
... 用硅胶注射法进行导管长度的测定(Wheeler et al., 2005; Hacke et al., 2007; Cai et al., 2010a) : 将清晨采回的枝条(长度30-50 cm)用刀片将切面切平, 然后在0.12 MPa压力下用100 mmol·L-1去气KCl溶液冲洗枝条30 min.硅胶配置: 硅胶是硅胶树脂和固化剂以10:1混合而成(15 g RTV141 part A + 1.5 g RTV141 part B), 0.025 g荧光增白剂Uvitex (使硅胶在紫外光下可见)与2.5 mL氯仿溶解后取15滴与硅胶混合, 搅拌使二者混合均匀后静置1 h以除去硅胶中的气泡.从枝条冲洗端橡胶管注入硅胶(硅胶占橡胶管的2/3即可), 在0.12 MPa压力下持续注射24 h, 之后在室温22 ℃条件下干燥2-3天, 干燥后切片(厚度20 μm), 切片分别从距离注射硅胶端的0.2、0.3、0.5、1.0、1.5、2.0、3.0、4.0 cm直到被注射导管个数占整个扇面导管个数的2%为止, 选取被注射上导管均匀分布的3个扇面获取照片(操作同上).所得照片用Win-CELL 2007图像分析软件统计出单位面积内被硅胶注射的导管数目, 由此可得单位横截面内导管的数量(N =横截面所有被硅胶注射导管的数量/横截面的面积).以截面到注射端的距离x为横坐标, 以lnN为纵坐标进行线性拟合, 得出线性拟合图的斜率λv (Cohen et al., 2003). ...
... 一是因为刺槐和榆树导管直径较旱柳、元宝枫和榛的导管直径大, 这与前人的具有大径导管植物比小径导管植物更易栓塞观点相同(Wheeler et al., 2005; Hacke et al., 2006; Markesteijn et al., 2011; Cai et al., 2010b; 张海昕等, 2013), 刺槐和榆树之所以容易栓塞是气种假说(air-seeding hypothesis)机理中的导管壁间纹孔膜的表面积所决定的(Wheeler et al., 2005), 纹孔膜在木质部水分传导过程中充当重要的角色, 其上有大量的微孔, 是水流从一个导管(管胞)流向另一个导管(管胞)的入口(Choat et al., 2007; Loepfe et al., 2007), 纹孔面积假说(pit area hypothesis)又是气种假说的必然结果, 按照气种假说, P50值由导管上所有纹孔面积中具有最大孔洞的纹孔决定, 因此, 导管直径越大, 导管上纹孔所占面积越大, 纹孔膜上拥有“易漏”的微孔的几率就越高, 植物栓塞脆弱性就大.导管直径、导管壁上纹孔面积与P50的相关性很接近(R2介于0.72-0.75), 它们在水分运输的有效性与安全性间具有权衡性(Wheeler et al., 2005, Cai et al., 2010b).二是因为沙棘、刺槐和榆树较旱柳、元宝枫和榛的导管内径跨度大, 导管内径跨度越大, 在负水势条件下导管腔越易垮塌, 植物越易栓塞, 这与Hargrave等(1994)的研究结果相同.三是因为沙棘、刺槐和榆树的木质部导管密度小、导管连接度低, 而旱柳、元宝枫和榛则恰好相反, 在低水势条件下, 水分可通过相邻导管或相邻导管壁纹孔进行水分运输, 而沙棘、刺槐和榆树通过相邻导管或纹孔运输水分几率显著小于旱柳、元宝枫和榛, 因此, 沙棘、刺槐和榆树较其他三树种导管易发生栓塞, 这支持Hacke等(2006) 的研究结果.四是因为刺槐和榆树木质部密度小于旱柳、元宝枫和榛, 木质部密度越小, 植物栓塞脆弱性越大, 这也与前人研究结果(Hacke et al., 2001; Jacobsen et al., 2005)相同.此外, 沙棘、刺槐和榆树与旱柳、元宝枫和榛相比, 前者较后者的导管直径大、导管长度长, 植物运输水分速度快, 从而较易发生栓塞. ...
... ), 刺槐和榆树之所以容易栓塞是气种假说(air-seeding hypothesis)机理中的导管壁间纹孔膜的表面积所决定的(Wheeler et al., 2005), 纹孔膜在木质部水分传导过程中充当重要的角色, 其上有大量的微孔, 是水流从一个导管(管胞)流向另一个导管(管胞)的入口(Choat et al., 2007; Loepfe et al., 2007), 纹孔面积假说(pit area hypothesis)又是气种假说的必然结果, 按照气种假说, P50值由导管上所有纹孔面积中具有最大孔洞的纹孔决定, 因此, 导管直径越大, 导管上纹孔所占面积越大, 纹孔膜上拥有“易漏”的微孔的几率就越高, 植物栓塞脆弱性就大.导管直径、导管壁上纹孔面积与P50的相关性很接近(R2介于0.72-0.75), 它们在水分运输的有效性与安全性间具有权衡性(Wheeler et al., 2005, Cai et al., 2010b).二是因为沙棘、刺槐和榆树较旱柳、元宝枫和榛的导管内径跨度大, 导管内径跨度越大, 在负水势条件下导管腔越易垮塌, 植物越易栓塞, 这与Hargrave等(1994)的研究结果相同.三是因为沙棘、刺槐和榆树的木质部导管密度小、导管连接度低, 而旱柳、元宝枫和榛则恰好相反, 在低水势条件下, 水分可通过相邻导管或相邻导管壁纹孔进行水分运输, 而沙棘、刺槐和榆树通过相邻导管或纹孔运输水分几率显著小于旱柳、元宝枫和榛, 因此, 沙棘、刺槐和榆树较其他三树种导管易发生栓塞, 这支持Hacke等(2006) 的研究结果.四是因为刺槐和榆树木质部密度小于旱柳、元宝枫和榛, 木质部密度越小, 植物栓塞脆弱性越大, 这也与前人研究结果(Hacke et al., 2001; Jacobsen et al., 2005)相同.此外, 沙棘、刺槐和榆树与旱柳、元宝枫和榛相比, 前者较后者的导管直径大、导管长度长, 植物运输水分速度快, 从而较易发生栓塞. ...
... 介于0.72-0.75), 它们在水分运输的有效性与安全性间具有权衡性(Wheeler et al., 2005, Cai et al., 2010b).二是因为沙棘、刺槐和榆树较旱柳、元宝枫和榛的导管内径跨度大, 导管内径跨度越大, 在负水势条件下导管腔越易垮塌, 植物越易栓塞, 这与Hargrave等(1994)的研究结果相同.三是因为沙棘、刺槐和榆树的木质部导管密度小、导管连接度低, 而旱柳、元宝枫和榛则恰好相反, 在低水势条件下, 水分可通过相邻导管或相邻导管壁纹孔进行水分运输, 而沙棘、刺槐和榆树通过相邻导管或纹孔运输水分几率显著小于旱柳、元宝枫和榛, 因此, 沙棘、刺槐和榆树较其他三树种导管易发生栓塞, 这支持Hacke等(2006) 的研究结果.四是因为刺槐和榆树木质部密度小于旱柳、元宝枫和榛, 木质部密度越小, 植物栓塞脆弱性越大, 这也与前人研究结果(Hacke et al., 2001; Jacobsen et al., 2005)相同.此外, 沙棘、刺槐和榆树与旱柳、元宝枫和榛相比, 前者较后者的导管直径大、导管长度长, 植物运输水分速度快, 从而较易发生栓塞. ...
4个杨树无性系木质部导管结构与栓塞脆弱性的关系
3
2013
... 干旱是全球性的自然灾害, 近年来因干旱引起的森林枯梢及死亡已严重影响许多国家的森林生态系统(Maherali et al., 2004).在我国, 干旱限制了干旱半干旱地区植物的生长和生态修复, 生长在干旱半干旱地区的植物在其生命周期内经常遭遇水分的胁迫(Chaves et al., 2002; Rennenberg et al., 2006; Nahm et al., 2007), 由干旱引起的木质部空穴化和栓塞则是制约树木生长发育和生态修复的一个重要因素, 干旱胁迫导致的抗栓塞性在不同树种和不同生态系统表现不同(Delzon et al., 2010), 在全球气候变化的大背景下, 干旱导致的抗栓塞性的研究已越来越重要(Cochard et al., 2013).木质部栓塞是木本植物在干旱环境下遭受水分胁迫时产生的木质部导管输水功能障碍, 而植物对木质部栓塞的易受性可用“脆弱曲线(VC)”来描述(Tyree & Sperry, 1989), 该曲线是根据木质部导水率损失的百分数(PLC)与相对应水势绘制而成的曲线, 且绝大多数脆弱曲线可用单威布尔累积分布函数(CDF)较好地拟合, 栓塞脆弱性一般用引起枝条50%导水率损失的压力值(P50)来表示.Cochard等(2008)和Awad等(2010)通过对蔷薇科李属(Prunus)的10个树种和杨属(Populus)的2个树种进行栓塞脆弱曲线的测定以及不同干旱条件的处理, 认为抗栓塞性可作为评估植物抵御干旱环境的一个指标.Choat等(2012)和Anderegg等(2013)研究认为抗栓塞性的大小是评估森林生产力和干旱导致的死亡率的一项重要生理指标, 树木栓塞抗性的研究将会有很大发展, 这些研究也将被运用到模型当中, 以估算植物生产力(生理生态模型)以及物种分布(生物地理学模型).张硕新等(1997)对我国北方5种抗旱树种栓塞脆弱性的研究表明, 耐旱的刺槐(Robinia pseudoacacia)和沙棘(Hipp- ophae rhamnoides)是栓塞脆弱性较大、较易发生栓塞的树种, 在水势下降时木质部导水率急剧下降, 即木质部很快发生栓塞, 而植物栓塞脆弱性与木质部结构密切相关(Wheeler et al., 2005; Sperry et al., 2006; Hacke & Jansen, 2009; Cai & Tyree, 2010; 张海昕等, 2013), 植物栓塞脆弱性与木质部结构关系的研究是了解耐旱树种抗旱机制的一个有效途径. ...
... 研究表明木质部导管的导水率与导管直径的四次方成正比(Hagen-Poiseuille方程)(Tyree & Zim- mermann, 2002), 即导管直径越大, 导管的导水能力就越强.多数研究认为木质部导管直径与栓塞脆弱性正相关, 即同一树种内大径导管比小径导管更易发生栓塞(Hargrave et al., 1994; Gullo et al., 1995; Wheeler et al., 2005; Hacke et al., 2006; Cai & Tyree, 2010; 张海昕等, 2013), 但也有研究表明栓塞脆弱性与导管直径成反比(Lemoine et al., 2002; Hacke et al., 2009)或不相关(Rosner et al., 2007; Fichot et al., 2010).Cai和Tyree (2010)以杨属植物为例, 按照植物导管直径径级构建的栓塞脆弱曲线表明: 随着导管直径增大, P50值增大(负值), 即导管直径与P50呈正相关关系.Cai等(2010b)的研究表明同一树种内导管直径与导管长度正相关, 即大径级导管其导管长度也越长, 张海昕等(2013)通过对4个杨树无性系的研究得出相同结论.此外, 有研究表明木质部密度和导管内径跨度(Hacke et al., 2001; Jacobsen et al., 2005; Cochard et al., 2008)、导管密度和导管连接度等与栓塞脆弱性具有一定的相关性(Wheeler et al., 2005; Hacke et al., 2006; Cai et al., 2014).安峰和张硕新(2005)对刺槐、元宝枫(Acer truncatum)、榛(Corylus heterophylla)等研究表明, 同一树种小枝和根的栓塞脆弱性主要由木质部结构决定, 小枝易发生栓塞的, 其根也较易发生栓塞, 根和小枝的栓塞脆弱性与植物的抗栓塞性有关.但目前, 通过木质部结构与栓塞脆弱性的关系来说明分布于我国干旱半干旱地区的耐旱树种在木质部结构上的差异尚未见报道, 因此, 本研究通过采用Cochard Cavitron离心机技术(仅需一根枝条且在0.5 h之内即可建立一条完整的栓塞脆弱曲线)(Cochard, 2002; Cochard et al., 2005)建立6个耐旱树种栓塞脆弱曲线, 计算其木质部P50, 找出6个耐旱树种木质部结构间的差异, 以期获得耐旱树种在木质部结构上的抗栓塞性指标, 为干旱、半干旱地区植被恢复工作中选育节水耐旱树种提供新的理论基础. ...
... 一是因为刺槐和榆树导管直径较旱柳、元宝枫和榛的导管直径大, 这与前人的具有大径导管植物比小径导管植物更易栓塞观点相同(Wheeler et al., 2005; Hacke et al., 2006; Markesteijn et al., 2011; Cai et al., 2010b; 张海昕等, 2013), 刺槐和榆树之所以容易栓塞是气种假说(air-seeding hypothesis)机理中的导管壁间纹孔膜的表面积所决定的(Wheeler et al., 2005), 纹孔膜在木质部水分传导过程中充当重要的角色, 其上有大量的微孔, 是水流从一个导管(管胞)流向另一个导管(管胞)的入口(Choat et al., 2007; Loepfe et al., 2007), 纹孔面积假说(pit area hypothesis)又是气种假说的必然结果, 按照气种假说, P50值由导管上所有纹孔面积中具有最大孔洞的纹孔决定, 因此, 导管直径越大, 导管上纹孔所占面积越大, 纹孔膜上拥有“易漏”的微孔的几率就越高, 植物栓塞脆弱性就大.导管直径、导管壁上纹孔面积与P50的相关性很接近(R2介于0.72-0.75), 它们在水分运输的有效性与安全性间具有权衡性(Wheeler et al., 2005, Cai et al., 2010b).二是因为沙棘、刺槐和榆树较旱柳、元宝枫和榛的导管内径跨度大, 导管内径跨度越大, 在负水势条件下导管腔越易垮塌, 植物越易栓塞, 这与Hargrave等(1994)的研究结果相同.三是因为沙棘、刺槐和榆树的木质部导管密度小、导管连接度低, 而旱柳、元宝枫和榛则恰好相反, 在低水势条件下, 水分可通过相邻导管或相邻导管壁纹孔进行水分运输, 而沙棘、刺槐和榆树通过相邻导管或纹孔运输水分几率显著小于旱柳、元宝枫和榛, 因此, 沙棘、刺槐和榆树较其他三树种导管易发生栓塞, 这支持Hacke等(2006) 的研究结果.四是因为刺槐和榆树木质部密度小于旱柳、元宝枫和榛, 木质部密度越小, 植物栓塞脆弱性越大, 这也与前人研究结果(Hacke et al., 2001; Jacobsen et al., 2005)相同.此外, 沙棘、刺槐和榆树与旱柳、元宝枫和榛相比, 前者较后者的导管直径大、导管长度长, 植物运输水分速度快, 从而较易发生栓塞. ...

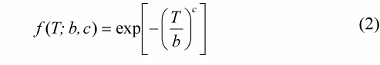
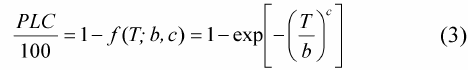


{kind=link}
{kind=link}
{kind=link}
{kind=link}