
植物生态学报 ›› 2016, Vol. 40 ›› Issue (3): 255-263.DOI: 10.17521/cjpe.2015.0260
李荣1, 党维1, 蔡靖1,3, 张硕新1,3, 姜在民2,*( )
)
收稿日期:2015-07-08
修回日期:2016-01-17
出版日期:2016-03-10
发布日期:2016-03-25
通讯作者:
姜在民
基金资助:
Rong LI1, Wei DANG1, Jing CAI1,3, Shuo-Xin ZHANG1,3, Zai-Min JIANG2,*()
Received:2015-07-08
Revised:2016-01-17
Online:2016-03-10
Published:2016-03-25
Contact:
Zai-Min JIANG
摘要:
木质部栓塞脆弱性对干旱响应的研究已成为全球气候变化背景下的热点和重要内容。该文以6个耐旱树种刺槐(Robinia pseudoacacia)、沙棘(Hippophae rhamnoides)、榆树(Ulmus pumila)、元宝枫(Acer truncatum)、旱柳(Salix matsudana)、榛(Corylus heterophylla)为研究对象, 采用Cochard Cavitron离心机技术建立木质部栓塞脆弱曲线, 计算木质部栓塞脆弱性, 利用染色法、硅胶注射法等测定木质部导管直径、导管内径跨度、导管连接度、导管密度、导管长度和木质部密度, 探究木质部结构与栓塞脆弱性的关系, 区分6个耐旱树种木质部结构在抗栓塞性上的差异, 以期建立6个耐旱树种在木质部结构方面的抗栓塞性指标。结果表明: 6个耐旱树种木质部栓塞脆弱性大小为刺槐>榆树>沙棘>旱柳>元宝枫>榛, 其中, 刺槐、沙棘和榆树的栓塞脆弱曲线为“r”形, 而元宝枫、旱柳和榛的栓塞脆弱曲线为“s”形, 脆弱曲线为“r”形的树种与脆弱曲线为“s”形的树种栓塞脆弱性差异极显著(p < 0.01)。线性分析表明: 木质部结构影响各树种的栓塞脆弱性, 其中, 木质部密度影响最大(t = 0.702), 导管直径次之(t = 0.532), 导管长度影响最小(t = 0.010)。
李荣, 党维, 蔡靖, 张硕新, 姜在民. 6个耐旱树种木质部结构与栓塞脆弱性的关系. 植物生态学报, 2016, 40(3): 255-263. DOI: 10.17521/cjpe.2015.0260
Rong LI, Wei DANG, Jing CAI, Shuo-Xin ZHANG, Zai-Min JIANG. Relationships between xylem structure and embolism vulnerability in six species of drought tolerance trees. Chinese Journal of Plant Ecology, 2016, 40(3): 255-263. DOI: 10.17521/cjpe.2015.0260
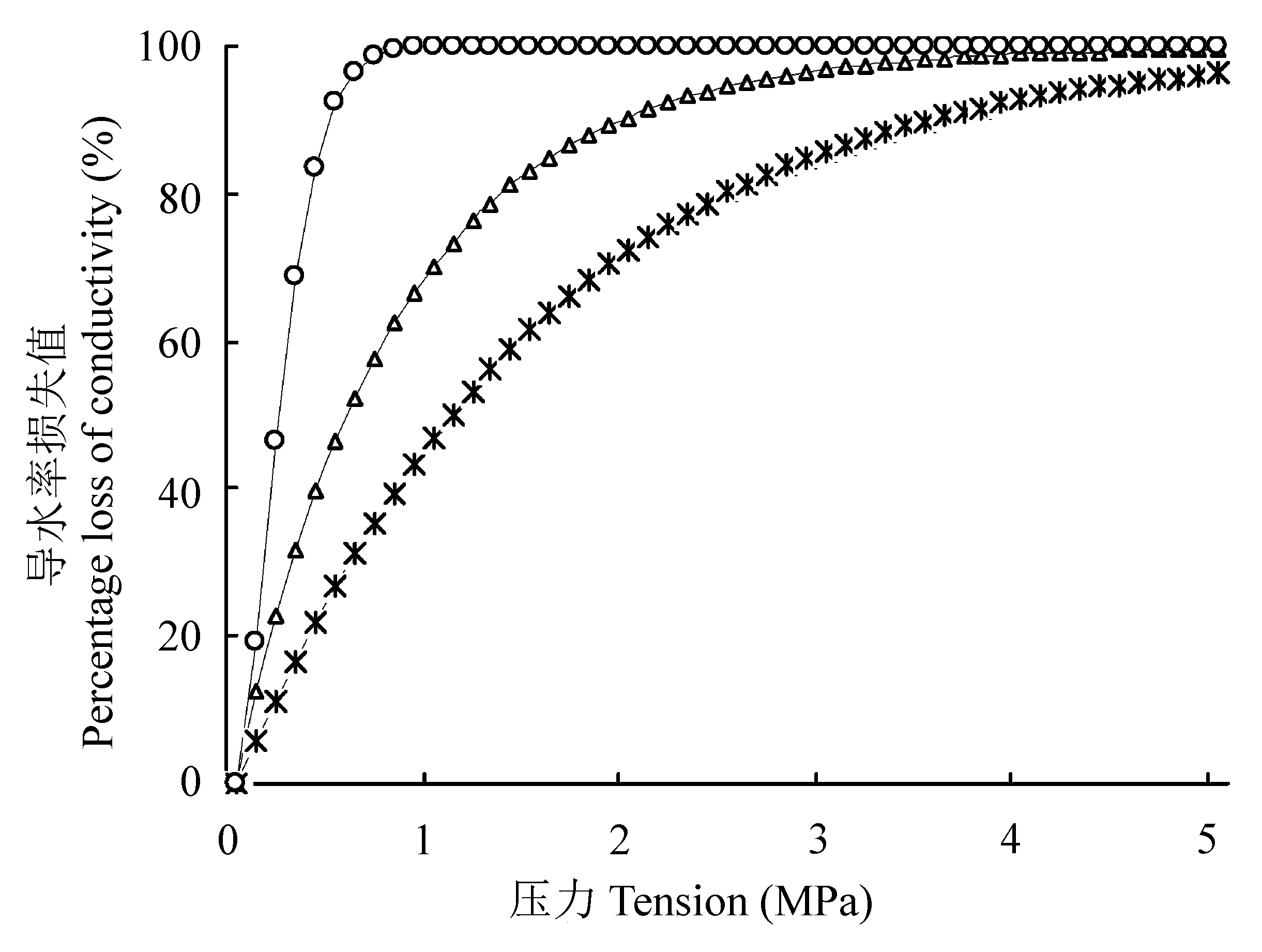
图1 耐旱树种沙棘(*)、刺槐(○)、榆树(△)的栓塞脆弱曲线(“r”形)。
Fig. 1 The vulnerability curves of Hippophae rhamnoides (*), Robinia pseudoacacia (○) and Ulmus pumila (△) (“r” shape).
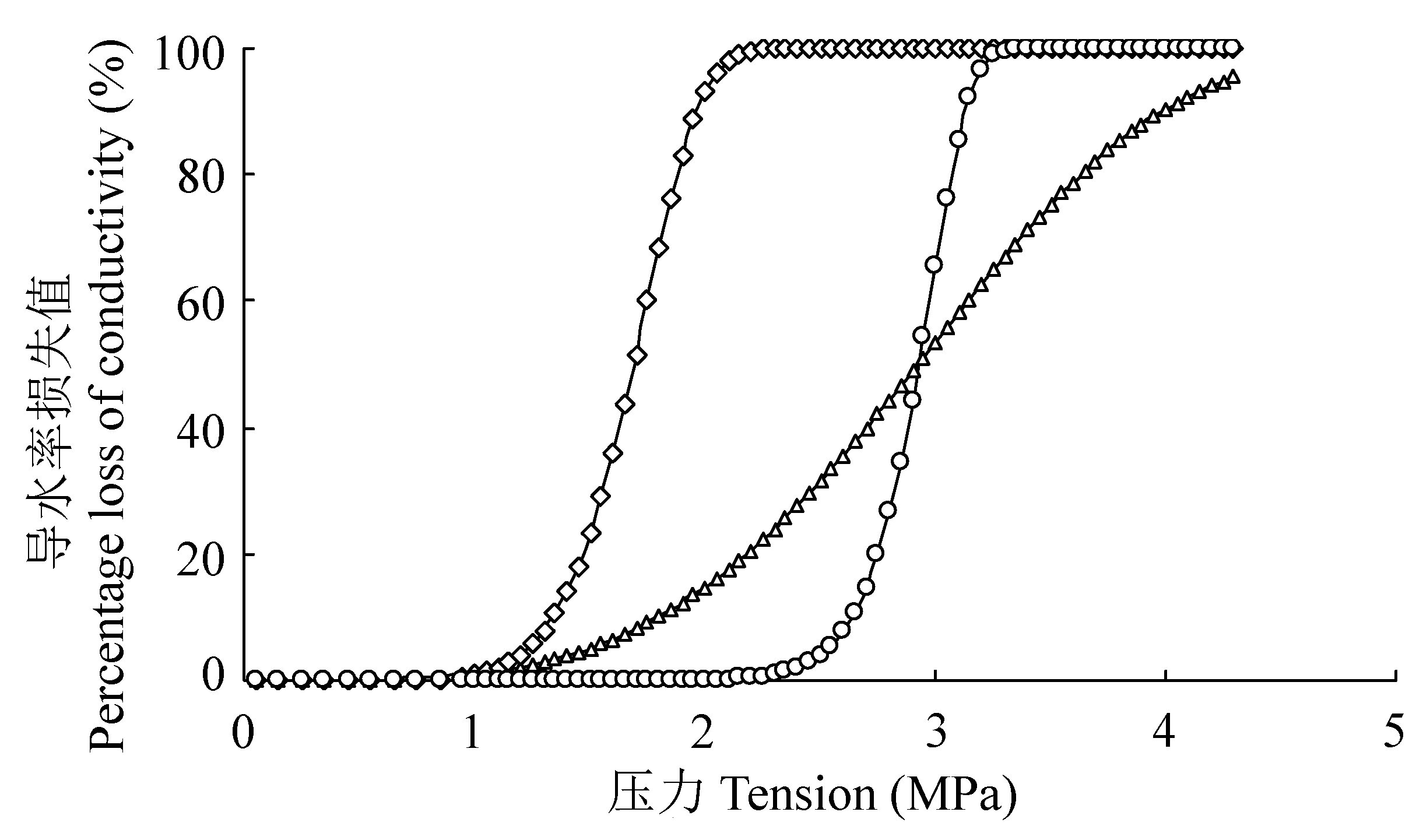
图2 耐旱树种元宝枫(△)、旱柳(◇)、榛(○)的栓塞脆弱曲线(“s”形)。
Fig. 2 The vulnerability curves of Acer truncatum (△), Salix matsudana (◇) and Corylus heterophylla (○) (“s” shape).
| 因子 Parameter | 耐旱树种 Drought tolerance trees | |||||
|---|---|---|---|---|---|---|
| 沙棘 Hippophae rhamnoides | 刺槐 Robinia pseudoacacia | 榆树 Ulmus pumila | 元宝枫 Acer truncatum | 旱柳 Salix matsudana | 榛 Corylus heterophylla | |
| P50 (MPa) | -1.31 ± 0.183Ab | -0.22 ± 0.026Ac | -0.55 ± 0.064Ac | -2.64 ± 0.214Ba | -1.55 ± 0.103Bb | -2.84 ± 0.054Ba |
| 导管直径 Vessel diameter (μm) | 23.57 ± 0.579Ac | 56.96 ± 1.913Aa | 49.48 ± 1.206Aa | 31.26 ± 0.405Bb | 32.11 ± 0.415Bb | 26.81 ± 0.319Bc |
| 导管内径跨度 Conduit wall span (μm) | 40.10 ± 1.651Aa | 62.20 ± 0.905Ab | 58.07 ± 2.414Aa | 34.29 ± 0.729Bb | 36.19 ± 0.583Bc | 27.04 ± 0.749Bb |
| 导管密度 Number of vessels per unit area | 216.52 ± 0.006Ab | 32.22 ± 2.588Aa | 53.64 ± 0.011Aa | 143.55 ± 3.874Bb | 284.37 ± 0.015Bb | 270.11 ± 0.013Bb |
| 导管连接度 Contact fraction | 0.03 ± 0.006Aa | 0.02 ± 0.002Aa | 0.04 ± 0.004Aa | 0.13 ± 0.008Bb | 0.07 ± 0.007Bb | 0.10 ± 0.008Bb |
| 导管长度 Vessel length (cm) | 13.35 ± 0.827Ab | 20.55 ± 1.913Aa | 17.23 ± 0.834Aa | 3.08 ± 0.253Bc | 4.10 ± 0.397Bc | 3.23 ± 0.223Bc |
| 木质部密度 Wood density (g·cm-3) | 1.54 ± 0.086Aa | 1.17 ± 0.129Aa | 1.17 ± 0.062Aa | 1.42 ± 0.167Ba | 1.46 ± 0.209Ba | 0.83 ± 0.039Bb |
表1 6个耐旱树种50%导水率损失的压力值(P50)及木质部结构特征值(平均值±标准误差)
Table 1 The value of tension at 50% loss of conductivity (P50) and characteristic values of xylem structure of the six species (mean ± SE)
| 因子 Parameter | 耐旱树种 Drought tolerance trees | |||||
|---|---|---|---|---|---|---|
| 沙棘 Hippophae rhamnoides | 刺槐 Robinia pseudoacacia | 榆树 Ulmus pumila | 元宝枫 Acer truncatum | 旱柳 Salix matsudana | 榛 Corylus heterophylla | |
| P50 (MPa) | -1.31 ± 0.183Ab | -0.22 ± 0.026Ac | -0.55 ± 0.064Ac | -2.64 ± 0.214Ba | -1.55 ± 0.103Bb | -2.84 ± 0.054Ba |
| 导管直径 Vessel diameter (μm) | 23.57 ± 0.579Ac | 56.96 ± 1.913Aa | 49.48 ± 1.206Aa | 31.26 ± 0.405Bb | 32.11 ± 0.415Bb | 26.81 ± 0.319Bc |
| 导管内径跨度 Conduit wall span (μm) | 40.10 ± 1.651Aa | 62.20 ± 0.905Ab | 58.07 ± 2.414Aa | 34.29 ± 0.729Bb | 36.19 ± 0.583Bc | 27.04 ± 0.749Bb |
| 导管密度 Number of vessels per unit area | 216.52 ± 0.006Ab | 32.22 ± 2.588Aa | 53.64 ± 0.011Aa | 143.55 ± 3.874Bb | 284.37 ± 0.015Bb | 270.11 ± 0.013Bb |
| 导管连接度 Contact fraction | 0.03 ± 0.006Aa | 0.02 ± 0.002Aa | 0.04 ± 0.004Aa | 0.13 ± 0.008Bb | 0.07 ± 0.007Bb | 0.10 ± 0.008Bb |
| 导管长度 Vessel length (cm) | 13.35 ± 0.827Ab | 20.55 ± 1.913Aa | 17.23 ± 0.834Aa | 3.08 ± 0.253Bc | 4.10 ± 0.397Bc | 3.23 ± 0.223Bc |
| 木质部密度 Wood density (g·cm-3) | 1.54 ± 0.086Aa | 1.17 ± 0.129Aa | 1.17 ± 0.062Aa | 1.42 ± 0.167Ba | 1.46 ± 0.209Ba | 0.83 ± 0.039Bb |
| 因子 Parameters | 系数 Coefficient | t |
|---|---|---|
| 木质部密度 Wood density (g·cm-3) | 0.326 | 0.702 |
| 导管直径 Vessel diameter (μm) | 0.037 | 0.532 |
| 导管内径跨度 Conduit wall span (μm) | -0.071 | 0.306 |
| 导管密度 Number of vessels per unit area | -0.007 | 0.265 |
| 导管连接度 Contact fraction | -19.762 | 0.188 |
| 导管长度 Vessel length (cm) | -0.281 | 0.010 |
表2 6个耐旱树种木质部结构特征值对50%导水率损失的压力值(P50)的影响程度
Table 2 The significance in characteristic values of xylem structure to value of tension at 50% loss of conductivity (P50)
| 因子 Parameters | 系数 Coefficient | t |
|---|---|---|
| 木质部密度 Wood density (g·cm-3) | 0.326 | 0.702 |
| 导管直径 Vessel diameter (μm) | 0.037 | 0.532 |
| 导管内径跨度 Conduit wall span (μm) | -0.071 | 0.306 |
| 导管密度 Number of vessels per unit area | -0.007 | 0.265 |
| 导管连接度 Contact fraction | -19.762 | 0.188 |
| 导管长度 Vessel length (cm) | -0.281 | 0.010 |
| [1] | An F, Zhang SX (2005). Studies of roots and shoots vulner- ability to xylem embolism in seven woody plants. Acta Ecologica Sinica, 25, 1928-1933. (in Chinese with Eng- lish abstract)[安锋, 张硕新 (2005). 七种木本植物根和小枝木质部栓塞的脆弱性. 生态学报, 25, 1928-1933.] |
| [2] | Anderegg WRL, Plavcov L, Anderegg LDL, Hacke UG, Berry JA, Field CB (2013). Drought’s legacy: Multi-year hydraulic deterioration underlies widespread aspen forest die-off and portends increased future risk.Global Change Biology, 19, 1188-1196. |
| [3] | Awad H, Barigah T, Badel E, Cochard H, Herbette S (2010). Poplar vulnerability to xylem cavitation acclimates to drier soil conditions.Physiologia Plantarum, 139, 280-288. |
| [4] | Cai J, Hacke U, Zhang S, Tyree MT (2010a). What happens when stems are embolized in a centrifuge? Testing the cavitron theory.Physiologia Plantarum, 140, 311-320. |
| [5] | Cai J, Li S, Zhang HX, Zhang SX, Tyree MT (2014). Recalcitrant vulnerability curves: Methods of analysis and the concept of fibre bridges for enhanced cavitation resistance.Plant, Cell & Environment, 37, 35-44. |
| [6] | Cai J, Tyree MT (2010). The impact of vessel size on vulnerability curves: Data and models for within-species variability in saplings of aspen, Populus tremuloides Michx.Plant, Cell & Environment, 33, 1059-1069. |
| [7] | Cai J, Zhang SX, Tyree MT (2010b). A computational algorithm addressing how vessel length might depend on vessel diameter.Plant, Cell & Environment, 33, 1234-1238. |
| [8] | Chaves MM, Pereira JS, Maroco J, Rodriggues ML, Ricardo CPP, Osorio ML (2002). How plants cope with water stress in the field? Photosynthesis and growth.Annals of Botany, 89, 907-916. |
| [9] | Choat B, Cobb AR, Jansen S (2007). Structure and function of bordered pits: New discoveries and impacts on whole-plant hydraulic function.New Phytologist, 177, 608-626. |
| [10] | Choat B, Jansen S, Brodribb TJ, Cochard H, Delzon S, Bhaskar R, Bucci SJ, Feild TS, Gleason SM, Hacke UG (2012). Global convergence in the vulnerability of forests to drought. Nature, 491, 752-755. |
| [11] | Cochard H (2002). A technique for measuring xylem hydraulic conductance under high negative pressures.Plant, Cell & Environment, 25, 815-819. |
| [12] | Cochard H, Barigah S, Kleinhentz M, Eshel A (2008). Is xylem cavitation resistance a relevant criterion for screening drought resistance among Prunus species?Plant Physiology, 165, 976-982. |
| [13] | Cochard H, Bréda N, Granier A, Aussenac G (1992). Vulnerability to air embolism of three European oak species (Quercus petraea (Matt) Liebl, Q. pubescens Willd, Q. robur L).Annals of Forest Science, 49, 225-233. |
| [14] | Cochard H, Damour G, Bodet C, Tharwat I, Poirier M, Améglio T (2005). Evaluation of a new centrifuge technique for rapid generation of xylem vulnerability curves.Physiologia Plantarum, 124, 410-418. |
| [15] | Cochard H, Lens F, Tixier A, Sperry JS, Jansen S, Herbette S (2013). Embolism resistance as a key mechanism to understand adaptive plant strategies.Plant Biology, 16, 287-292. |
| [16] | Cohen S, Bennink J, Tyree M (2003). Air method measure- ments of apple vessel length distributions with improved apparatus and theory.Journal of Experimental Botany, 54, 1889-1897. |
| [17] | Delzon S, Douthe C, Sala A, Cochard H (2010). Mechanism of water-stress induced cavitation in conifers: Bordered pit structure and function support the hypothesis of seal capillary-seeding.Plant, Cell & Environment, 33, 2101-2111. |
| [18] | Enquist BJ, West GB, Charnov EL, Brown JH (1999). Allometric scaling of production and life-history variation in vascular plants.Nature, 401, 907-911. |
| [19] | Fichot R, Barigah TS, Chamaillard S, lethiec D, Laurans F, Cochard H, Brignolas F (2010). Common trade-offs bet- ween xylem resistance to cavitation and other physiolo- gical traits do not hold among unrelated Populus deltoides × Populus nigra hybrids.Plant, Cell & Environment, 33, 1553-1568. |
| [20] | Gullo MA, Salleo S, Piaceri EC, Rosso R (1995). Relations between vulnerability to xylem embolism and xylem conduit dimensions in young trees of Quercus corris.Plant, Cell & Environment, 18, 661-669. |
| [21] | Hacke UG, Jansen S (2009). Embolism resistance of three boreal conifer species varies with pit structure.New Phytologist, 182, 675-686. |
| [22] | Hacke UG, Jacobsen AL, Pratt RB (2009). Xylem function of arid-land shrubs from California, USA: An ecological and evolutionary analysis.Plant, Cell & Environment, 32, 1324-1333. |
| [23] | Hacke UG, Sperry JS, Field TS (2007). Water transport in vessel less angiosperms: Conducting efficiency and cavitation safety.International Journal of Plant Sciences, 168, 1113-1126. |
| [24] | Hacke UG, Sperry JS, Pockman WT, Davis SD, McCulloh KA (2001). Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure.Oecologia, 126, 457-461. |
| [25] | Hacke UG, Sperry JS, Wheeler JK, Castro L (2006). Scaling of angiosperm xylem structure with safety and efficiency.Tree Physiology, 26, 689-701. |
| [26] | Han RL, Li LX, Liang ZS, Xue ZD, Du ZK (2002). The research of Hippophae rhamnoides membrane lipid peroxide under drought stress protection system.Journal of Northwest Forestry College, 17, 1-5. (in Chinese with English abstract)[韩蕊莲, 李丽霞, 梁宗锁, 薛智德, 杜正科 (2002). 干旱胁迫下沙棘膜脂过氧化保护体系研究. 西北林学院学报, 17, 1-5.] |
| [27] | Hargrave KR, Kolb KJ, Ewers FW, Davis SD (1994). Conduit diameter and drought-induced embolism in Salvia mellifera (Labiatae).New Phytologist, 126, 695-705. |
| [28] | Jacobsen AL, Ewers FW, Pratt RB, Paddock WA, Davis SD (2005). Do xylem fibers affect vessel cavitation resistance?Plant Physiology, 139, 546-556. |
| [29] | Lemoine D, Cochard H, Granier A (2002). Within crown variation in hydraulic architecture in beech (Fagus sylvatica L.): Evidence for a stomatal control of xylem embolism.Annals of Forest Science, 59, 19-27. |
| [30] | Li DQ, Liang YM, Huang J, Jiang J (2003). Study on characteristic of anatomy in Hippophae rhamnoides subsp.sinensis. Journal of Northwest Forestry College, 23, 1224-1230. (in Chinese with English abstract)[李代琼, 梁一民, 黄瑾, 姜峻 (2003). 沙棘的形态解剖学特性研究. 西北植物学报, 23, 1224-1230.] |
| [31] | Loepfe L, Martinez-Vilalta J, Pinol J, Mencuccini M (2007). The relevance of xylem network structure for plant hydraulic efficiency and safety.Theoretical Biology, 247, 788-803. |
| [32] | Maherali H, Pockman WT, Jackson RB (2004). Adaptive variation in the vulnerability of woody plants to xylem cavitation.Ecology, 85, 2184-2199. |
| [33] | Markesteijn L, Poorter L, Paz H, Sack L, Bongers F (2011). Ecological differentiation in xylem cavitation resistance is associated with stem and leaf structural traits.Plant, Cell & Environment, 34, 137-148. |
| [34] | Nahm M, Matzarakis A, Rennenberg H, Geβler A (2007). Seasonal courses of key parameters of nitrogen, carbon and waterbalance in European beech (Fagus sylvatica L.) grown on four different study sites along a European North-South climate gradient during the 2003 drought.Trees, 21, 79-92. |
| [35] | Rennenberg H, Loreto F, Polle A, Brilli F, Fares S, Beniwal RS (2006). Physiological responses of forest trees to heat and drought.Plant Biology, 8, 556-571. |
| [36] | Rosner S, Klein A, Müller U (2007). Hydraulic and mechanical properties of young Norway spruce clones related to growth and wood structure.Tree Physiology, 27, 1165-1178. |
| [37] | Sperry JS, Christman MA, Torres-ruiz JM, Taneda H, Smith DD (2012). Vulnerability curves by centrifugation: Is there an open vessel artefact, and are ‘r’ shaped curves necessarily invalid?Plant, Cell & Environment, 35, 601-610. |
| [38] | Sperry JS, Hacke UG, Pittermann J (2006). Size and function in conifer tracheids and angiosperm vessels.Botany, 93, 1490-1500. |
| [39] | Tyree MT, Sperry JS (1989). Vulnerability of xylem to cavitation and embolism. Annual Review of Plant Biology, 40, 19-36. |
| [40] | Tyree MT, Zimmermann MH (2002). Xylem Structure and the Ascent of Sap. Springer, Berlin. 45-56. |
| [41] | Vilagrosa A (2002). Estrategias de resistencia al deficit hídrico en Pistacia lentiscus L. y Quercus coccifera L. Implica- ciones en la repoblación forestal. PhD dissertation, University of Alicante, Alicante, Spain. |
| [42] | Vilagrosa A, Bellot J, Vallejo VR, Gil-Pelegrín E (2003). Cavitation, stomatal conductance and leaf dieback in seedlings of two co-occurring Mediterranean shrubs during an intense drought.Experimental Botany, 54, 2015-2024. |
| [43] | West NE (1989). Intermountain deserts, shrub steppes, and wood-lands. In: Barbour MG, Billings WD eds. North American Terrestrial Vegetation. Cambridge University Press, Cambridge, UK. 209-230. |
| [44] | Wheeler JK, Sperry JS, Hacke UG, Hoang N (2005). Inter-vessel pitting and cavitation in woody Rosaceae and other vesselled plants: A basis for a safety versus efficiency trade-off in xylem transport.Plant, Cell & Environment, 28, 800-812. |
| [45] | Zhang HX, Li S, Zhang SX, Xiong XY, Cai J (2013). Relationships between xylem structure and embolism vulnerability in four Populus clones.Scientia Silvae Sinicae, 49(5), 54-61. (in Chinese with English abstract)[张海昕, 李珊, 张硕新, 熊晓燕, 蔡靖 (2013). 4个杨树无性系木质部导管结构与栓塞脆弱性的关系. 林业科学, 49(5), 54-61.] |
| [46] | Zhang SX, Shen WJ, Zhang YY, Zhou XX (1997). The research on the xylem embolism vulnerability of several woody plants.Journal of Northwest Forestry College, 12(2), 1-6. (in Chinese with English abstract)[张硕新, 申卫军, 张远迎, 周新霞 (1997). 几个抗旱树种木质部栓塞脆弱性的研究. 西北林学院学报, 12(2), 1-6.] |
| [1] | 李伟斌, 张红霞, 张玉书, 陈妮娜. 昼夜不对称增温对长白山阔叶红松林碳汇能力的影响[J]. 植物生态学报, 2023, 47(9): 1225-1233. |
| [2] | 王嘉仪, 王襄平, 徐程扬, 夏新莉, 谢宗强, 冯飞, 樊大勇. 北京市行道树绒毛梣的水力结构对城市不透水表面比例的响应[J]. 植物生态学报, 2023, 47(7): 998-1009. |
| [3] | 程向芬, 马晋, 赵涵, 姜在民, 蔡靖. 木本植物水力学结构之导管长度研究进展[J]. 植物生态学报, 2018, 42(6): 609-618. |
| [4] | 安瑞, 孟凤, 尹鹏先, 杜光源. 刺槐木质部栓塞脆弱性检测的方法比较[J]. 植物生态学报, 2018, 42(11): 1113-1119. |
| [5] | 李荣, 姜在民, 张硕新, 蔡靖. 木本植物木质部栓塞脆弱性研究新进展[J]. 植物生态学报, 2015, 39(8): 838-848. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19