|
|
||||||||||||||||||||||||||||
|
不同生境条件下蕨类和被子植物的气孔形态特征及其对光强变化的响应
植物生态学报
2014, 38 (8):
868-877.
DOI: 10.3724/SP.J.1258.2014.00081
气孔是植物与大气环境进行气体交换的重要通道, 在调控植物碳水平衡方面发挥着重要作用。为探讨生境和植物类型对气孔形态特征的影响以及气孔对光强变化的响应格局在不同植物间和不同生境条件下的变异, 选取开阔生境和林下生境的5种蕨类植物和4种被子植物, 测定了它们的气孔形态特征和气孔导度对光强变化的响应。此外, 还收集了8篇文献中开阔和林下生境的45种蕨类植物和70种被子植物的气孔密度和气孔长度数据, 以增大样本量从而更好地探讨不同生境条件下蕨类和被子植物气孔密度及长度的变异格局, 并通过分析生境和植物类型对气孔形态特征的影响来推测生境和植物类型对气孔响应行为的可能影响。实验结果表明, 与林下植物相比, 开阔环境下的植物气孔密度更大, 气孔长度更小, 气孔对光强降低的响应更敏感; 但植物类型对气孔形态特征的影响以及对气孔响应光强的敏感程度的影响均不显著。对文献数据的分析表明, 生境和植物类型对气孔形态特征均有显著影响。考虑到气孔响应快慢与气孔形态特征密切相关, 与蕨类植物相比, 被子植物小而密的气孔可能为其更快地响应环境变化提供了基础。研究表明生境和植物类型对气孔响应行为均有显著影响。
表2
生境和植物类型对9种植物气孔响应光强变化过程中的各个指标的影响。
正文中引用本图/表的段落
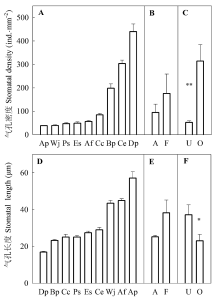
在9种植物中, 来自开阔生境的蕨类植物芒萁(Dicranopteris pedata)气孔密度最大(440.23 ind.·mm-2), 气孔长度最短(16.88 μm); 而来自林下生境的蕨类植物长叶铁角蕨(Asplenium prolongatum)气孔密度最小(38.48 mm-2), 气孔长度最长(57.10 μm); 前者的气孔密度为后者的10倍左右, 而后者的气孔长度约为前者的4倍(图2)。方差分析表明生境对气孔密度和气孔长度均有显著影响, 与林下植物相比, 来自开阔生境的植物气孔密度更大(图2C), 而气孔长度更小(图2F)。此外, 在9种植物中, 植物类型对气孔密度和气孔长度的影响均不显著(图2B、2E)。
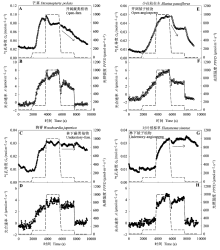
当光照强度增加时(从低光强到高光强), 两种生境的蕨类植物和被子植物的气孔导度和光合速率都逐渐增加(图6); 生境和植物类型对气孔张开速率(绝对速率和相对速率)的影响不显著(p > 0.05, 表2)。而当光照强度降低时(从高光强到低光强), 所有植物的光合速率都迅速下降(图6B、6D、6F和6H), 但只有开阔生境的蕨类和被子植物气孔导度迅速下降(图6A和6E), 而林下生境的蕨类和被子植物气孔导度则下降缓慢(图6C和6G)。生境对气孔关闭速率(绝对速率和相对速率)的影响显著(p < 0.05, 表2), 而植物类型对气孔关闭速率(绝对速率和相对速率)的影响不显著(p > 0.05, 表2)。此外, 生境对气孔响应光强过程中的气孔导度最大值和光合速率最大值均有显著影响(p < 0.05, 表2), 而植物类型对两者的影响不显著(p > 0.05, 表2)。
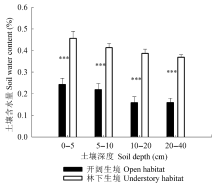
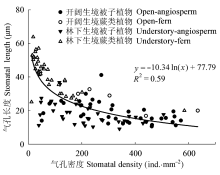
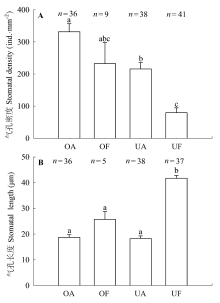
生长在不同环境下的植物可以通过不同的适应机制如改变气孔的形态特征和行为来适应环境, 从而能有效地利用环境资源, 维持较高的生理活动强度(李庆康和马克平, 2002; Franks & Beerling, 2009; Casson & Hetherington, 2010)。针对9个物种的研究表明, 与林下植物相比, 开阔生境下的植物气孔密度更大(图2C), 气孔更小(图2F), 气孔对光强降低的响应也更加敏感(图6)。结合文献中的气孔形态数据, 我们发现生境对气孔密度和长度有显著的影响(图4A、4B)。在开阔环境中, 光照强且波动大, 空气湿度低且蒸散需求高, 小而密的气孔可以使植物获取更大的气孔导度和光合速率, 更快地响应光强和VPD等环境因子的变化, 进而拥有较高的水分利用效率(El-Sharkawy et al., 1985; Tinoco-Ojanguren & Pearcy, 1993; Drake et al., 2013)。本研究里生长在开阔环境下的蕨类植物芒萁就是一个例子; 它的气孔密度较大(440.23 ind.·mm-2), 气孔长度小(16.88 μm), 气孔对光强降低的响应很敏感(图6A), 这些特征使其快速地响应开阔生境中环境条件的变化, 从而能够广泛分布于旷地、林缘等开阔环境中, 并与被子植物共存(Nobel et al., 1984; Zhao et al., 2012),相关研究也发现开阔环境下的蕨类植物能够快速改变气孔导度以响应光照、CO2和VPD等的变化(Nobel et al., 1984; Czerniak, 2013)。另一方面, 林下植物对光强变化的缓慢响应可能是适应林下光斑(sunflecks)的结果。光斑是太阳光移动引起的光照强度突然增加, 占林下光合有效辐射的50%以上(Chazdon, 1988)。当光照强度突然降低, 类似于自然光斑消失, 林下植物的气孔以较慢的速度响应(图6C和6G), 可以减少光合诱导时间, 提高光斑利用效率(Pearcy, 1987; Valladares et al., 1997; Allen & Pearcy, 2000)。由于林下光照强度低, 相对湿度高, 土壤含水量也较高(图1), 植物主要受光而不是水的限制, 所以不灵敏的气孔响应反而有利于增大光的利用效率, 以较低的蒸腾成本获取较大的光合收益(Tinoco- Ojanguren & Pearcy, 1993)。
本文的其它图/表
|