
|
|
||
|
中国森林生态系统土壤呼吸温度敏感性空间变异特征及影响因素
植物生态学报
2020, 44 (6):
687-698.
DOI: 10.17521/cjpe.2019.0300
土壤呼吸的温度敏感性(Q10)是陆地碳循环与气候系统间相互作用的关键参数。尽管已有大量关于不同类型森林Q10季节和年际变化规律的研究, 但是对Q10在区域尺度的空间变异特征及其影响因素仍认识不足, 已有结果缺乏一致结论。该研究通过整合已发表论文, 构建了中国森林生态系统年尺度Q10数据集, 共包含399条记录、5种森林类型(落叶阔叶林(DBF)、落叶针叶林(DNF)、常绿阔叶林(EBF)、常绿针叶林(ENF)、混交林(MF))。分析了不同森林类型Q10的空间变异特征及其与地理、气候和土壤因素的关系。结果显示, 1) Q10介于1.09到6.24之间, 平均值(±标准误差)为2.37 (± 0.04), 且在不同森林类型之间无显著差异; 2)当考虑所有森林类型时, Q10随纬度、海拔、土壤有机碳含量(SOC)和土壤全氮含量(TN)的增加而增大, 随经度、年平均气温(MAT)、平均年降水量(MAP)的增加而减小。气候(MAT、MAP)和土壤(SOC、TN)因素间存在相互作用, 共同解释了33%的Q10空间变异, 其中MAT和SOC是Q10空间变异的主要驱动因素; 3)不同类型森林Q10对气候和土壤因素的响应存在差异。在DNF中Q10随MAP的增加而减小, 而其他类型森林中Q10与MAP无显著相关性; 在EBF、DBF、ENF中Q10随TN的增加而增大, 但Q10对TN的敏感性在EBF中最高, 在ENF中最低。这些结果表明, 尽管Q10有一定的集中分布趋势, 但仍有较大范围的空间变异, 在进行碳收支估算时应注意尺度问题。Q10的主要驱动因素和Q10对环境因素的响应随森林类型而变化, 在气候变化情景下, 不同森林类型间Q10可能发生分异。因此, 未来的碳循环-气候模型还应考虑不同类型森林碳循环关键参数对气候变化的响应差异。 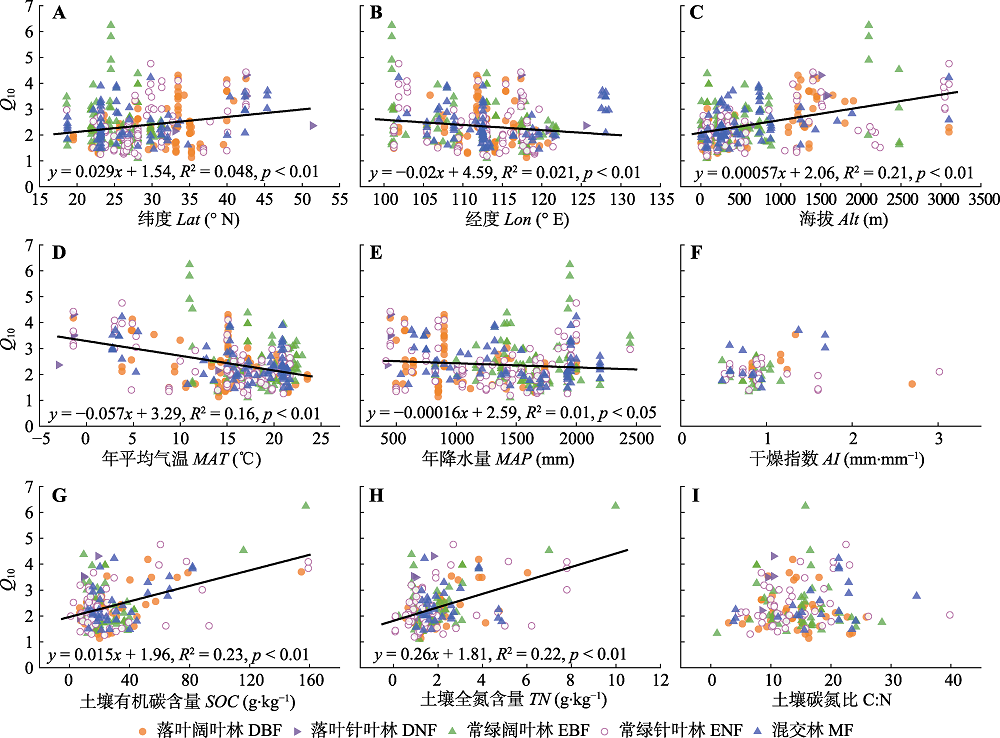 View image in article
图4
土壤呼吸温度敏感性(Q10)与纬度(A)、经度(B)、海拔(C)、年平均气温(D)、年降水量(E)、干燥指数(F)、土壤有机碳含量(G)、土壤全氮含量(H)、土壤碳氮比(I)的关系。
正文中引用本图/表的段落
中国森林生态系统土壤呼吸Q10随纬度(R2 = 0.048, p < 0.01)和海拔(R2 = 0.21, p < 0.01)的增加而增大, 随经度(R2 = 0.021, p < 0.01)的增加而减小(图4A-4C)。协方差分析表明Q10受森林类型和经度交互作用的影响(协方差分析(ANCOVA), Lon, F1,398 = 9.03, p < 0.01; 林分类型(Stand type), F4,398 = 3.21, p < 0.01; Lon × Stand type, F4,398 = 4.78, p < 0.01)。回归分析也表明, 地理因素对Q10的影响在不同森林类型的生态系统中有差异(附录II)。在DBF、ENF、MF中Q10随纬度的增加而增大, 而在DNF和EBF中Q10不随纬度变化; 在DNF、EBF、ENF中Q10随经度的增加而减小, 而在DBF和MF中Q10不随经度变化; 在5种森林类型中Q10均随海拔增加而增大, 但DNF中Q10随海拔增加而增大的幅度高于其他森林类型。
中国森林生态系统Q10随MAT (R2 = 0.16, p < 0.01)和MAP的增加而减小(R2 = 0.01, p < 0.05), 不随AI变化(图4D-4F)。协方差分析表明Q10受森林类型和MAP (ANCOVA, MAP, F1,397 = 4.00, p = 0.04; Stand type, F4,397 = 1.81, p = 0.13; MAP × Stand type, F4,397 = 2.66, p = 0.03), 森林类型和AI (ANCOVA, AI, F1,79 = 1.12, p = 0.29; Stand type, F3,79 = 4.09, p < 0.01; AI × Stand type, F3,79 = 4.05, p = 0.01)交互作用的影响。回归分析也表明, 气候因素对Q10 的影响在不同森林类型的生态系统中有差异(附录II)。在DBF、EBF、ENF、MF中Q10随MAT的增加而减小, 而在DNF中Q10与MAT无显著关系; 在DNF中Q10随MAP的增加而减小, 而其余林型中Q10与MAP无显著关系。在MF中Q10随AI的增加而增大, 其余林型中Q10与AI无显著关系。
中国森林生态系统Q10随SOC (R2 = 0.23, p < 0.01)和TN的增加而增大(R2 = 0.22, p < 0.01), 且随TN增大的趋势强于SOC (TN斜率0.26, SOC斜率0.015), 与C:N无显著关系(图4G-4I)。协方差分析表明Q10受森林类型和TN交互作用的影响(ANCOVA, TN, F1,206 = 62.50, p < 0.01; Stand type, F4,206 = 2.27, p = 0.06; TN × Stand type, F4,206 = 3.33, p = 0.02)。回归分析也表明, 土壤因素对Q10的影响在不同森林类型的生态系统中有差异(附录II)。在DBF、EBF、ENF、MF中Q10随SOC的增加而增大, 在DNF中Q10不随SOC变化。在EBF、DBF、ENF中Q10随TN的增加而增大, 且Q10对TN的敏感性在EBF中最高, 在ENF中最低; 在DNF、MF中Q10不随TN变化。
中国森林Q10随MAT增加而减小(图4D), 表明较寒冷地区的土壤呼吸对温度升高的敏感性高于较温暖地区, 支持了现有的研究结论(Peng et al., 2009; Xu et al., 2015; Li et al., 2020)。研究表明, MAT可以通过直接和间接途径影响Q10。首先, 根呼吸和微生物呼吸均需要酶的参与, 温度升高会导致酶活性的相对增加幅度下降(Peterson et al., 2007; Schipper et al., 2014; Li et al., 2020), 从而降低Q10; 大分子速率理论(MMRT)还表明Q10受酶催化反应热容差(ΔC?p)的强烈影响, ΔC?p随温度升高而降低, 导致Q10随温度升高而降低(Schipper et al., 2014; Li et al., 2020)。其次, 尽管温度升高通过促进土壤微生物快速分解活性有机碳导致Q10随SOC增加而增大(Giardina & Ryan, 2000; Rustad et al., 2001), 但温暖地区较高的分解速率降低了SOC, 因此Q10较低(图5A)。此外, MAT和MAP相互影响, 共同作用于Q10。本研究中MAT通过对Q10的直接和间接影响最终降低了Q10 (图5B)。MAT每增加1 ℃, Q10下降5.7% (图4D), 该值与Peng等(2009)和Li等(2019)的估计值(6%)类似, 高于Xu等(2015)的估计值(3.8%), 造成这种差异的机制尚未得到证实, 可能是不同研究中使用的样本量和生态系统类型的差异引起的。例如Xu等(2015)的研究中包括74个Q10观测值, 本研究中的399个观测值能更全面地量化中国森林生态系统土壤呼吸对温度升高的响应。未来在气候变暖的趋势下, 有必要厘清温度对Q10的直接和间接影响, 以提高碳循环-气候模型的预测能力。
中国森林Q10随MAP增加而减小(图4E), 与现有的研究结果(Peng et al., 2009; Xu et al., 2015)类似, 可能是潮湿地区中过高的土壤水分限制了氧气扩散, 使植物根系和好氧微生物的活动受到抑制, 进而降低了Q10。此外, 有研究发现水分与温度的交互作用会影响Q10, 如在一些干旱和地中海气候的生态系统中温度升高时土壤中的水分降低、土壤呼吸底物扩散受限, 进而导致土壤呼吸速率与温度解耦(Reichstein et al., 2002; Wang et al., 2014), 而本研究中MAP和MAT呈正相关关系(图5A), 但二者对Q10无交互作用(ANCOVA, MAT, F1,394 = 79.98, p < 0.01; MAP, F1,394 = 16.28, p < 0.01; MAT × MAP, F1,394 = 0.62, p = 0.43), 这可能表明只有在极端的温度和水分条件下, 二者才对Q10有交互作用。本研究中MAP每增加100 mm, Q10下降1.6% (图4E), 该值低于现有的研究(8%, Peng et al., 2009; 7%, Xu et al., 2015; 7%, Li et al., 2019), 可能与样本量的差异有关, 本研究在较潮湿地区的观测值(大于1 900 mm: 91个)远多于前人研究(11个, Peng et al., 2009; 6个, Xu et al., 2015), Li等(2019)的研究为室内培养实验, 与野外观测条件差异较大。此外, Li等(2020)最近的整合研究表明, 在全球尺度上Q10与MAP无显著关系, 因此MAP与Q10的负相关关系可能很难扩展到较大的尺度。全球气候变暖伴随着降水时空格局的改变(Knapp et al., 2015), 因此需要在进一步的研究中明确水分对Q10空间变异的作用机制, 在碳循环-气候模型中考虑温度和降水的相互作用。
Q10随SOC和TN的增加而增大, 与现有的研究结果一致(Zheng et al., 2009; Song et al., 2014)。土壤有机碳是土壤微生物呼吸的重要底物(Bond-Lamberty et al., 2018), 在其他环境因素不变的条件下, SOC较高的生态系统其土壤微生物呼吸速率随温度升高的幅度也较大, 因此Q10更高。由于土壤中碳氮存在紧密的耦合关系, 二者是协同积累的(G?rden?s et al., 2011), 因此SOC和TN呈正相关关系(图5A)。此外, 尽管有研究发现Q10与底物质量(C:N)呈负相关关系(Fierer et al., 2006; Liu et al., 2017; Wang et al., 2018a), 但本研究中Q10与C:N无显著关系(图4I), 这表明Q10的空间变异可能不受底物质量的影响。一些室内培养实验也表明有机质分解对温度的响应与底物质量无关(Fang et al., 2005; Li et al., 2019), 还有研究发现土壤基质对有机质的保护作用对Q10的影响远超过底物质量(Moinet et al., 2018), Li等(2019)的研究进一步强调了土壤保护性因子(如黏粒含量)对Q10空间变异的重要性。
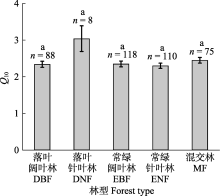
DNF的Q10在所有森林类型中最高(图3), 这与现有的研究结果一致(Peng et al., 2009; Song et al., 2014)。本研究的DNF主要为东北地区的落叶松(Larix gmelinii)林和华北地区的华北落叶松(Larix principis-rupprechtii)林, 少数为华东地区的水杉(Metasequoia glyptostroboides)林。低温是东北和华北地区DNF中根系和微生物酶活性的主要限制因素, 因此DNF中土壤呼吸对温度的变化最敏感, 这也可以由Q10与MAT的负相关关系来检验(图4D)。Q10对环境因素的响应随森林类型变化(附录II)。DNF中Q10与MAT和SOC无显著关系, 但随MAP增加而减小; 其余森林类型中Q10均随MAT增加而减小, 随SOC增加而增大, 但与MAP无显著关系, 这些结果可能表明DNF的根系和微生物群落对MAT、MAP和SOC的敏感性与其他森林类型显著不同, 也可能是DNF的观测数据(6个)较少, 不足以准确体现Q10对环境因素的响应, 未来仍需要开展更多的野外观测以明确DNF中Q10对环境因素的响应机制。此外, 森林类型与TN存在交互作用。尽管在DNF和MF中Q10与TN无显著关系, 但在DBF、EBF、ENF中Q10均随TN增加而增大, 且阔叶林(EBF、DBF)中Q10对TN的敏感性强于针叶林(ENF), 这可能是阔叶林与针叶林的根系或微生物群落在利用TN时采取了不同的策略。本研究中阔叶林的TN (2.87 g·kg-1)高于针叶林(1.84 g·kg-1), 以往的研究也发现, 相比于针叶林, 阔叶林的凋落物和枯死根系的归还量高且更易于分解, 因此其养分循环和分解速度更快(Reich et al., 2005; Olsson et al., 2012), 这种差异可能导致阔叶林的土壤微生物群落在利用TN时采用资源获取型策略, 而针叶林则为保守型策略, 但具体机制仍需要大量实验验证。不同森林类型对环境因素的差异性响应预示着未来在气候变化加剧的背景下, 不同森林类型间Q10的差异可能会增大, 这将影响碳循环-气候模型的预测精度, 因此需要予以考虑。
本文的其它图/表
|


{kind=link}