大叶藻对氮磷营养盐的吸收动力学特征
李诗奇, 张彦浩, 李政, 张沛东
植物生态学报
2020, 44 ( 7):
772-781.
DOI: 10.17521/cjpe.2019.0335
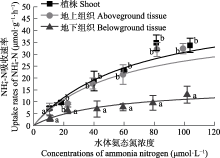
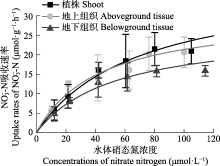
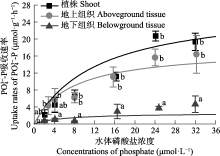
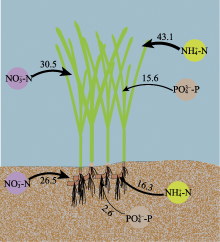
营养盐是影响海草生长的关键因子, 目前有关海草不同组织对不同形式氮和磷的吸收特征尚不明确。该研究通过利用海草地上和地下组织分隔培养装置, 设置不同的氨态氮、硝态氮和磷酸盐浓度, 探究了大叶藻(Zostera marina)植株及其地上和地下组织对氮磷营养盐的吸收动力学特征。结果显示: (1)大叶藻对氮磷的吸收符合饱和吸收动力学特征, 吸收速率和水体氮磷浓度可用米式方程描述; (2)大叶藻植株对NH4+-N的最大吸收速率(Vmax, 52 μmol·g-1·h-1)显著高于其对NO3--N的Vmax (39 μmol·g-1·h-1); (3)大叶藻地上组织和地下组织均可吸收氮磷, 但地上组织对氨态氮、硝态氮、磷酸盐的Vmax分别为43.1、30.5和15.6 μmol·g-1·h-1, 为地下组织的2.6、1.2和6倍。结果表明, 大叶藻对氨态氮的吸收能力高于硝态氮, 且对氮磷的吸收可能主要依赖地上组织(叶片)。结果为查明大叶藻对氮磷的吸收利用机制及评估大叶藻的海洋生态效应提供了理论依据。
分隔装置
Separation apparatus | 未分隔组
No-separating group | 分隔且地上组织添加组
Separation group with N, P addition in the leaf compartment | 分隔且地下组织添加组
Separation group with N, P addition in the root- rhizome compartment | 玻璃水槽
Glass aquarium | 设置的营养盐浓度
Designed nutrient concentration | 设置的营养盐浓度
Designed nutrient concentration | 天然海水营养盐浓度
Nutrient concentration of natural seawater | 聚乙烯瓶
Polyethylene bottle | 设置的营养盐浓度
Designed nutrient concentration | 天然海水营养盐浓度
Nutrient concentration of natural seawater | 设置的营养盐浓度
Designed nutrient concentration |
View table in article
表1
大叶藻分隔培养不同实验装置水体营养盐浓度设置方案
正文中引用本图/表的段落
van Katwijk等(1997)研究发现, 水体氨态氮浓度≥125 μmol·L-1时对大叶藻产生毒性; Udy和Dennison (1997)的研究表明, 水体磷酸盐浓度≥ 30 μmol·L-1时对海草生长具有显著促进作用。根据这些已有报道以及预实验结果, 设置氨态氮(NH4Cl)和硝态氮(KNO3)浓度分别为10、20、40、60、80和100 μmol·L-1, 磷酸盐(KH2PO4)浓度为2、4、8、16、24和32 μmol·L-1。各营养盐水平分别设置3个实验组: 未分隔组(UG)、分隔且地上组织添加组(SGA)和分隔且地下组织添加组(SGB), 分别用于评价大叶藻植株、地上组织和地下组织对氨态氮、硝态氮和磷酸盐的吸收动力学特征。不同实验组水体营养盐浓度设置方案见表1。其中天然海水氨态氮浓度为(3.04 ±0.14) μmol·L-1, 硝态氮浓度为(2.41 ± 0.18) μmol·L-1, 磷酸盐浓度为(0.25 ± 0.02) μmol·L-1。各实验组设置12个重复。
图中数字表示最大吸收速率(Vmax, μmol·g-1·h-1)。
Km, 半饱和常数; Vmax, 最大吸收速率. ... Interaction of leaves and roots of Ruppia maritima in the uptake of phosphate, ammonia and nitrate 1 1984 ... 尽管已有研究表明, 海草既可以通过根系从沉积物中吸收氮磷, 也可以通过叶片从海水中获取, 但多数海草种类的叶片是吸收氮磷的主要部位(Lee & Dunton, 1999; Gras et al., 2003; Nielsen et al., 2006).如Pedersen等(1997)研究表明, 南极根枝草(Amphibolis antarctrica)的叶片对NH4+的吸收速率是其根茎的5-38倍; Hemminga等(1994)研究表明, 大叶藻叶片在氮吸收过程中发挥主要作用, 68%-92%的氮吸收量来源于叶片; 龟裂泰来草叶片对PO43-P的最大吸收速率是地下组织的3.3倍(Gras et al., 2003); 川蔓草(Ruppia maritima)叶片对NH4+-N的最大吸收速率是地下组织的5倍(Thursby & Harlin, 1984).本研究也发现, 尽管大叶藻的地上组织和地下组织均可吸收氮磷营养盐, 但地上组织的Vmax值更高, 因此大叶藻可能更依赖叶片吸收氮磷, 其对氮磷的吸收模式如图5所示.海草叶片对氮磷营养盐的吸收率较高, 与其谷氨酰胺合成酶和硝酸还原酶活力显著高于地下组织密切相关, 叶片在酶的催化下可以高效吸收并充分利用氮磷营养盐(Kraemer et al., 1997; Alexandre et al., 2004). ... 几种作物NO3 吸收动力学参数测定方法初探 1 2001 ... 在米氏方程中, Vmax表示底物饱和时的吸收速率, 越大则吸收的内在潜力越大, Km表示当吸收速率为最大吸收速率一半时的底物浓度, Km值越小, 对底物亲和力越大(刘静雯和董双林, 2001; 田霄鸿等, 2001).研究表明, 大叶藻植株对NH4+-N、NO3--N和PO43--P表现出饱和吸收动力学的特征, 其对营养盐的吸收速率和水体氮磷浓度可用米式方程描述, 这与之前对有些藻类和海草吸收动力学特征的研究结果(Lavery & McComb, 1991; Hanisak & Harlin, 2010)相符合.Rubio等(2018)研究发现, 大洋波喜荡草(Posidonia oceanica)对NO3--N和PO43--P的吸收亦符合饱和吸收动力学, 但其Km值显著低于大叶藻.这可能反映了不同海草种类栖息环境氮磷营养盐条件的差异以及对NO3--N和PO43--P的转运系统不同, 大洋波喜荡草在低氮、低磷条件下, 主要通过Na+依赖式转运系统吸收NO3--N和PO43--P, 而大叶藻则主要采用H+依赖式转运系统(Lara et al., 1993; Bethoux et al., 2005; Rubio et al., 2018). ... 几种作物NO3 吸收动力学参数测定方法初探 1 2001 ... 在米氏方程中, Vmax表示底物饱和时的吸收速率, 越大则吸收的内在潜力越大, Km表示当吸收速率为最大吸收速率一半时的底物浓度, Km值越小, 对底物亲和力越大(刘静雯和董双林, 2001; 田霄鸿等, 2001).研究表明, 大叶藻植株对NH4+-N、NO3--N和PO43--P表现出饱和吸收动力学的特征, 其对营养盐的吸收速率和水体氮磷浓度可用米式方程描述, 这与之前对有些藻类和海草吸收动力学特征的研究结果(Lavery & McComb, 1991; Hanisak & Harlin, 2010)相符合.Rubio等(2018)研究发现, 大洋波喜荡草(Posidonia oceanica)对NO3--N和PO43--P的吸收亦符合饱和吸收动力学, 但其Km值显著低于大叶藻.这可能反映了不同海草种类栖息环境氮磷营养盐条件的差异以及对NO3--N和PO43--P的转运系统不同, 大洋波喜荡草在低氮、低磷条件下, 主要通过Na+依赖式转运系统吸收NO3--N和PO43--P, 而大叶藻则主要采用H+依赖式转运系统(Lara et al., 1993; Bethoux et al., 2005; Rubio et al., 2018). ... Review of nitrogen and phosphorus metabolism in seagrasses 1 2000 ... 海草床生态系统较脆弱, 受全球变化和人类活动影响, 目前已是地球生物圈退化速率最快的生态系统, 且退化速率仍在逐年增加(Waycott et al., 2009; 郑凤英等, 2013; 郭美玉等, 2017; Perry et al., 2020).然而在海草床修复过程中, 对促进海草快速生长的关键因子及其生态幅的科学认知还远远不够, 限制了高效修复方法的建立.光照、温度和营养盐是影响海草生长的三大外界因素(Lee et al., 2007).Dennison等(1993)在对澳大利亚、古巴、美国等17个国家或地区的15种海草种类进行研究后分别得出了不同地区不同海草种类的最小需光量(%); Lee等(2007)综合分析已有文献报道, 提出大叶藻(Zostera marina)生长的适宜温度范围为15-20 ℃; 尽管20世纪70年代就有学者尝试探究海草的营养生理生态学理论, 但至今仍未得出海草的适宜营养盐浓度范围(Orth, 1973; van Katwijk et al., 1999; Hemminga & Duarte, 2000; Touchette & Burkholder, 2000; Romero et al., 2006; Kaldy, 2014; Pereda-Briones et al., 2018). ... Effects of inorganic n availability on algal photosynthesis and carbon metabolism 1 1991 ... 陆生高等植物主要以硝态氮和氨态氮作为无机氮素的来源, 但更多的是吸收硝态氮, 只有在某些还原性较强的土壤或者水淹地, 其主要的氮源才可能是氨态氮(McCarthy et al., 2004).本研究结果发现, 大叶藻优先吸收氨态氮作为氮源, 与大多数海草种类相同(Brun et al., 2002; Guo et al., 2007; Alexandre et al., 2015), 这是因为海草营沉水生活, 海草体内空间不足以积聚硝态氮, 因此当硝态氮被海草吸收后, 其将转化为氨态氮的形式进行储存或者存于液泡中, 该过程需要消耗大量的能量和磷, 从而增加磷限制风险(Turpin, 1991; Burkholder et al., 1992; Alexandre et al., 2004; Burkholder et al., 2007). ... Growth and physiological responses of three seagrass species to elevated sediment nutrients in Moreton Bay, Australia 1 1997 ... van Katwijk等(1997)研究发现, 水体氨态氮浓度≥125 μmol·L-1时对大叶藻产生毒性; Udy和Dennison (1997)的研究表明, 水体磷酸盐浓度≥ 30 μmol·L-1时对海草生长具有显著促进作用.根据这些已有报道以及预实验结果, 设置氨态氮(NH4Cl)和硝态氮(KNO3)浓度分别为10、20、40、60、80和100 μmol·L-1, 磷酸盐(KH2PO4)浓度为2、4、8、16、24和32 μmol·L-1.各营养盐水平分别设置3个实验组: 未分隔组(UG)、分隔且地上组织添加组(SGA)和分隔且地下组织添加组(SGB), 分别用于评价大叶藻植株、地上组织和地下组织对氨态氮、硝态氮和磷酸盐的吸收动力学特征.不同实验组水体营养盐浓度设置方案见表1.其中天然海水氨态氮浓度为(3.04 ±0.14) μmol·L-1, 硝态氮浓度为(2.41 ± 0.18) μmol·L-1, 磷酸盐浓度为(0.25 ± 0.02) μmol·L-1.各实验组设置12个重复. ... Seagrass meadows support global fisheries production 1 2019 ... 海草是一类海洋高等单子叶开花植物, 具有极高的初级生产力, 在我国主要分布于温带的山东、辽宁、河北, 亚热带、热带的广东、广西和海南等沿岸海域(李文涛和张秀梅, 2009; Short et al., 2011; 郑凤英等, 2013).海草床具有重要的生态功能和极高的生态服务价值(Unsworth et al., 2019).Short等(2011)估算发现, 海草床的生态服务价值高达34 000 $·hm-2·a-1, 超过大多数陆地生境及海洋 生境. ... Potential uptake of dissolved organic matter by seagrasses and macroalgae 1 2011 ... 氮磷是水生植物最主要的限制性营养盐, 学者普遍使用米氏方程来表示稳态时水生植物对营养盐的吸收特征(Duarte, 1990; Geider et al., 1997; Thormann & Bayley, 1997; 刘静雯等, 2001; Güsewell et al., 2003; Pietro et al., 2006; Rubio et al., 2018).研究表明, 大多数海草种类都优先吸收氨态氮(Vonk et al., 2008; van Engeland et al., 2011), 但其地上组织和地下组织对营养盐的吸收能力存在差异(Gras et al., 2003; Nielsen et al., 2006).因此, 查明海草对营养盐的吸收动力学特征, 得出不同组织对不同形式氮和磷的吸收参数, 对明确海草最适营养盐生态幅具有重要科学意义. ... Effects of salinity and nutrient load and their interaction on Zostera marina 1 1999 ... 海草床生态系统较脆弱, 受全球变化和人类活动影响, 目前已是地球生物圈退化速率最快的生态系统, 且退化速率仍在逐年增加(Waycott et al., 2009; 郑凤英等, 2013; 郭美玉等, 2017; Perry et al., 2020).然而在海草床修复过程中, 对促进海草快速生长的关键因子及其生态幅的科学认知还远远不够, 限制了高效修复方法的建立.光照、温度和营养盐是影响海草生长的三大外界因素(Lee et al., 2007).Dennison等(1993)在对澳大利亚、古巴、美国等17个国家或地区的15种海草种类进行研究后分别得出了不同地区不同海草种类的最小需光量(%); Lee等(2007)综合分析已有文献报道, 提出大叶藻(Zostera marina)生长的适宜温度范围为15-20 ℃; 尽管20世纪70年代就有学者尝试探究海草的营养生理生态学理论, 但至今仍未得出海草的适宜营养盐浓度范围(Orth, 1973; van Katwijk et al., 1999; Hemminga & Duarte, 2000; Touchette & Burkholder, 2000; Romero et al., 2006; Kaldy, 2014; Pereda-Briones et al., 2018). ... Ammonium toxicity in eelgrass Zostera marina 2 1997 ... van Katwijk等(1997)研究发现, 水体氨态氮浓度≥125 μmol·L-1时对大叶藻产生毒性; Udy和Dennison (1997)的研究表明, 水体磷酸盐浓度≥ 30 μmol·L-1时对海草生长具有显著促进作用.根据这些已有报道以及预实验结果, 设置氨态氮(NH4Cl)和硝态氮(KNO3)浓度分别为10、20、40、60、80和100 μmol·L-1, 磷酸盐(KH2PO4)浓度为2、4、8、16、24和32 μmol·L-1.各营养盐水平分别设置3个实验组: 未分隔组(UG)、分隔且地上组织添加组(SGA)和分隔且地下组织添加组(SGB), 分别用于评价大叶藻植株、地上组织和地下组织对氨态氮、硝态氮和磷酸盐的吸收动力学特征.不同实验组水体营养盐浓度设置方案见表1.其中天然海水氨态氮浓度为(3.04 ±0.14) μmol·L-1, 硝态氮浓度为(2.41 ± 0.18) μmol·L-1, 磷酸盐浓度为(0.25 ± 0.02) μmol·L-1.各实验组设置12个重复. ...
本文的其它图/表
|